Область техники, к которой относится изобретение
Изобретение относится к генным конструкциям, которые пригодны в качестве компонентов вакцин против вируса гепатита С в протоколах генетической иммунизации, к способам защиты индивидуумов от заражения вирусом гепатита С, к способам лечения индивидуумов, страдающих от заражения вирусом гепатита С, к рекомбинантным химерным генным конструкциям, которые полезны в качестве компонентов вакцин против вируса гепатита В и/или вируса гепатита С в протоколах генетической иммунизации, к способам защиты индивидуумов от заражения вирусом гепатита В и/или вирусом гепатита С и к способам лечения индивидуумов, страдающих от заражения вирусом гепатита В и/или вирусом гепатита С. Настоящая заявка родственна заявке на патент США, зарегистрированной 5 октября 1994, регистрационный номер 08/318248, и заявке на патент США, зарегистрированной 6 июня 1995, регистрационный номер 08/467859, которые обе включены в настоящее изобретение в качестве ссылок.
Предпосылки создания изобретения
Вирус гепатита С (HCV), основной этиологический фактор приобретаемого при трансфузии не-А и не-В гепатита, ответственен приблизительно за 150000 новых случаев острого вирусного гепатита в США ежегодно. См. Greenberger, N. J. , 1983, "New Approaches for Hepatitis C", Contemporary Internal Medicine Feb:64. Примерно половина этих случаев заражения переходит в хронические инфекционные заболевания, которые могут ассоциироваться с циррозом и/или печеночно-клеточным раком (Alter, et al., Science, 1992, 258, 135-140; и Alter, et al. , New Eng. J. Med., 1992, 327, 1899-1905). Кроме того, HCV-инфекция является независимым фактором риска для развития печеночно-клеточного рака, как показывает широкое распространение антител против HCV (Colombo, et al., Lancet, 1989, ii, 1006-1008; Saito, et al., Proc. Natl. Acad. Sci. USA, 1990, 87, 6547-6549; Simonetti, et al., An. Int. Med., 1992, 116, 97-102; и Tsukuma, et al., New Eng. J. Med., 1993, 328, 1797-1801).
HCV представляет собой оболочечный, плюс-нитевой РНК-вирус, длиной приблизительно в 9500 нуклеотидов, который в последнее время относят к отдельному роду в семействе Flavivirus (Heinz, F.X., Аrсh. Virol. (Suppl.), 1992, 4, 163-171). Различные иэоляты показывают большое разнообразие нуклеотидных последовательностей, приводящее к подразделению геномов HCV по крайней мере на восемь генотипов (Simmonds, et al., J. Gen. Virol., 1993, 74, 2391-2399). Во всех генотипах вирусный геном содержит большую открытую рамку считывания (ORF), которая кодирует белок-предшественник из 3010-3033 аминокислот приблизительно в 330 кД (Choo, et al., Proc. Natl. Acad. Sci. USA, 1991, 88, 2451-2455; Inchauspe, et al., Proc. Natl. Acad. Sci. USA, 1991, 88, 10292-10296; Kato, et al., Proc. Natl. Acad. Sci. USA, 1990, 87, 9524-9528; Okamoto, et al., J. Gen. Virol., 1991, 72, 2697-2704; и Takamizava, et al., J. Gen. Virol., 1991, 65, 1105-1113).
Отдельные полипептиды HCV продуцируются протеолитическим процессингом полипептида-предшественника с образованием корового белка (С), белков оболочки (El, E2) и неструктурных белков (NS2-NS5) (Bartenschlager, et al., J. Gen. Virol., 1993, 67, 3835-3844; Grakoui, et al., J. Gen. Virol., 1993, 67, 2832-2843; и Selby, et al., J. Gen. Virol., 1993, 74, 1103-1113). Такой протеолиз катализируется сочетанием как клеточных, так и вирусных кодированных протеаз.
Помимо транслируемой области, геном HCV содержит как 5'-нетранслируемую область (5'-UTR), так и 3'-нетранслируемую область (3'-UTR). 5'-UTR из 324-341 нуклеотидов представляет наиболее консервативную последовательность среди всех изолятов, о которых приводятся данные (Han, et al., Proc. Natl. Acad. Scl. USA, 1991, 88, 1711-1715; и Bukh, et al., Proc. Natl. Acad. Sci. USA, 1992, 89, 4942-4946). Постулировано, что эта 5'-UTR содержит важные регуляторные элементы для репликации и/или трансляции РНК HCV. 5'-UTR также содержит несколько небольших открытых рамок считывания (ORF), но в настоящее время нет оснований полагать, что эти ORF-последовательности действительно являются транслируемыми.
Коровый ген HCV может представлять собой важную мишень для противовирусного подхода, основанного на нуклеиновых кислотах. Полагают, что первые 191 аминокислот полипротеинового предшественника HCV представляют собой вирусный нуклеокапсидный белок. Этот белок состоит из основного РНК-связывающего аминоконцевого домена и высокогидрофобной области карбоксильного конца (Bukh, et al. , Proc. Natl. Acad. Sci. USA, 1994, 91, 8239-8243; и Santolini et al., J. Virol., 1994, 68, 3631-3641). Зрелый коровый белок в 21 кД отщепляется от полипротеинового предшественника клеточной сигнальной пептидазой, и есть основания предполагать, что нуклеокапсидный белок HCV устойчиво ассоциируется с цитоплазматической поверхностью мембраны эндоплазматического ретикулума (Hijikata, et al., Proc. Natl. Acad. Sci. USA, 1991, 88, 5547-5551; и Santolini, et al. , J. Virol., 1994, 68, 3631-3641). В противоположность гликопротеинам оболочки, которые включают гипервариабельную область в аминоконцевой области Е2 (Weiner, et al., Virol., 1991, 180, 842-848; и Weiner, et al. , Proc. Natl. Acad. Sci. USA, 1992, 89, 3468-3472), коровый белок является весьма консервативным среди различных генотипов HCV и генерирует иммунный ответ организма-хозяина (Bukh, et al., Proc. Natl. Acad. Sci. USA, 1994, 91, 8239-8243; и Houghton, et al., Hepatology, 1991, 14, 381-388). Предыдущие исследования показали, что большинство инфицированных HCV индивидуумов вырабатывают антитела к коровому белку HCV в начале заражения. (Chiba, et al., Proc. Natl. Acad. Sci. USA, 1991, 88, 4641-4645; Hosein, et al., Proc. Natl. Acad. Sci. USA, 1991, 88, 3647-3651; Hsu, et al., Hepatology, 1993, 17, 763-771; Katayama, et al., Hepatology, 1992, 15, 391-394; Nasoff, et al., Proc. Natl. Acad. Sci. USA, 1991, 88, 5462-5466; и Okamoto, et al., Hepatology, 1992, 15, 180-186). Более того, нуклеокапсидный белок представляется важной мишенью для клеточного иммунного ответа против HCV (Botarelli, et al., Gastroenterol., 1993, 104, 580-587; Koziel, et al., J. Virol., 1993, 67, 7522-7532; и Shirai, et al., J. Virol. , 1993, 68, 3334-3342). Наконец, последние наблюдения позволяют предположить, что коровый белок HCV может также иметь некоторые регуляторные функции (Shin, et al., J. Virol., 1993, 67, 5823-5832). Мутация вирусного генома с высокой частотой происходит, вероятно, во время репликации генома вируса и через иммунную селекцию. Это явление может быть связано с установлением персистентной вирусной инфекции и последующим хроническим характером болезни (Weiner, et al., Proc. Natl. Acad. Sci. USA, 1992, 89, 3468-3472; Kato, et al., Biochem.Biophys. Res. Comm., 1992, 189, 119-127; и Alter, et al., New Eng. J. Med., 1992, 327, 1899-1905).
Клеточные иммунные события, вовлеченные в повреждение печени и вирусный клиренс во время HCV-инфекции, установлены только частично. При попытке изучить потенциальную патогенную роль инфильтрирующих печень лимфоцитов у пациентов с хронической HCV-инфекцией, Koziel, et al. изучили ответ цитотоксических Т-лимфоцитов (ЦТЛ) таких клеток и продемонстрировали НLA класса I-ограниченный СD8+ЦТЛ-ответ, направленный против как структурных, так и неструктурных областей HCV-полипептидов (Koziel, et аl., J. Virol., 1993, 67, 7522-7532; и Koziel, et al., J. Immunol., 1992, 149, 3339-3344). Другие исследователи также отметили наличие ЦТЛ в популяциях мононуклеарных клеток периферической крови, которые во время хронической HCV-инфекции распознают эпитопы на коровом и других родственных вирусных белках (Kita, et аl., Hepatol., 1993, 18, 1039-1044; и Cerny, et al., Intl. Symp. Viral. Hepatitis Liver Dis., 1993, 83 (реферат)).
Botarelli, et al., (Botarelli, et al., Gastroenterol.,1993, 104, 580-587) и Ferrari, et al., (Ferrari et al., Hepatol., 1994, 19, 286-295) обнаружили HLA класса II-ограниченный CD4+ опосредуемый Т-клетками пролиферативный ответ на некоторые рекомбинантные белки, полученные из различных областей HCV, у пациентов с хронической HCV-инфекцией. Примечательно, что существовала корреляция между ответами Т-клеток на коровый белок HCV и клинически доброкачественным течением болезни печени так же, как и последующим уничтожением вируса. Однако подобные исследования показали, что пролиферативный ответ на коровый белок HCV не предсказывает доброкачественное клиническое течение относительно серьезности болезни печени (Schupper, et al., Hepatol. , 1993, 18, 1055-1060). Таким образом, важно прояснить связь между активным клеточным иммунитетом и клиническим течением вирусной инфекционной болезни в отношении типа повреждения печени и клинической реакции HCV-инфекции на интерфероновую (IFN) терапию. В этом отношении проводились исследования, включающие ответы мононуклеарных клеток периферической крови (РВМС) на рекомбинантный GST-HCV коровый слитый белок, которые включали оценку способности таких клеток продуцировать IFN-γ; и проводились корреляции с различными клиническими результатами заражения HCV. Обнаружено, что мононуклеарные клетки 24 из 46 пациентов (52%) с хронической болезнью печени отвечали на коровый белок; бессимптомные носители HCV показали низкую степень ответа (15%, р<0,05). Что более существенно, пациенты, которые получили лечение IFN-α и пришли к клинической и вирусологической эмиссии, имели более высокую степень реакции (75%, р<0,05) к коровому белку HCV по сравнению с пациентами с постоянным гепатитом, лечение которых не удалось (31%). Из 25 пациентов, мононуклеарные клетки которых реагировали на коровый белок HCV, 18 имели заметную реакцию на один или несколько пептидов; 12 пациентов реагировали на пептидную смесь, содержащую гидрофильные последовательности. Аминокислотная последовательность корового пептида 140-160 распознавалась 9 пациентами. Интересно, что 7 из 8 пациентов, несущих HLA DR4 и гаплотипы w53, узнавали пептидную последовательность 140-160. Таким образом, оказалось, что ответ мононуклеарных клеток ограничен HLA DR, и отвечающие клетки идентифицированы как CD4+ Т-клетки. Это исследование показывает присутствие иммунодоминантных Т-клеточных эпитопов в коровом белке HCV в ассоциации с фенотипами HLA DR у пациентов с заболеванием печени, связанным с HCV.
Вирус гепатита В (HBV) является основным человеческим патогеном, для которого не существует эффективного лечения. Установлено, что более 300 миллионов человек в мире хронически инфицированы этим вирусом. Воздействие HBV может привести к острому или хроническому гепатиту, циррозу печени и развитию печеночно-клеточного рака. Клинические последствия этой серьезной инфекционной болезни являются предметом особого отношения в разработках всего мира, особенно там, где HBV-инфекция является одной из ведущих причин смертности, а также является основной причиной острых или хронических болезней печени в Соединенных Штатах, как и в Европе. HBV является эталонным членом семейства hepadnavirus, группы близкородственных вирусов (Ganem, et al. , Annu. Rev. Blochem., 1987, 56, 651-693), которая включает, среди прочих, вирус гепатита В утки (DHBV) и вирус гепатита лесного североамериканского сурка (WHV). Экспериментально инфицированные утки или лесные американские сурки точно воспроизводили многие признаки болезни человека, такой как острая или хроническая инфекционная болезнь, и, в случае хронически инфицированных лесных американских сурков развитие печеночно-клеточного рака (Shodel, et al. , "The Biology of Avian Hepatitis В viruses", в Molecular Biology of the Hepatitis В Virus, 1991, Vol. 3, CRC Press, Boca Raton, FL, pp. 53-80; Korba, et al., J. Virol., 1989, 63, 1360-1370; и Коrbа, et al., Hepatology, 1989, 9, 461-470). Хотя вирусные репликативные промежуточные формы обнаружены в других тканях, печень является органом-мишенью, и гепатоцитное повреждение ассоциируется с персистентной вирусной инфекцией. Представляется, что сам по себе HBV не является цитопатическим вирусом. Поэтому вероятно, что иммунный ответ организма-хозяина, продуцированный против вирусных эпитопов, продуцирует повреждение печени, и стратегия лечения, разработанная для уменьшения геномной репликации вируса в печени, может иметь положительное клиническое действие.
Геном HBV кодирует 4 открытые рамки считывания (ORF), которые включают 1) 5-ген, кодирующий белок оболочки, с 2 полипептидами pre-Sl и pre-S2 внутри рамки; 2) полимеразную ORF, кодирующую белок обратную транскриптазу, которая ответственна за обратную транскрипцию предгеномной РНК в 3,6 кб в ДНК; 3) коровый ген, кодирующий белок, который собран для завершения вирусного нуклеокапсида; и 4) НВх ORF, которая кодирует белок неизвестной функции. Ген ро1 заключает в себе 80% генома и перекрывает три другие ORF. Коровый ген предваряется внутрирамочной последовательностью, которая кодирует сигнальный пептид, и последующее протеолитическое расщепление приводит к антигенно отличному белку, названному HBeAg. Обнаружено, что НВх-белок не является существенным для вирусного жизненного цикла in vitro (Blum, et al., J. Virol. , 1992, 66, 123-127), но выясняется, что он является необходимым для установления продуктивной инфекции in vivo (Chen, et al., J. Virol., 1993, 67, 1218-1226). НВх может функционировать как транскрипционный трансактиватор на множестве клеточных и вирусных генов, и предполагается, что он может вносить вклад в развитие НСС (Schek, et аl., "The Hepadnaviral X Protein" в Molecular Biology of the Hepatitis В Virus, 1991, Vol. 3, CRC Press, Boca Raton, FL, pp. 181-192).
Прямая инъекция ДНК животным является перспективным способом доставки специфических антигенов в целях иммунизации (Barry, et al., Bio Techniques, 1994, 16, 616-619; Davis, et al., Hum. Mol. Genet., 1993, 11, 1847-1851; Tang, et al., Nature, 1992, 356, 152-154; Wang, et al., J. Virol., 1993, 67, 3338-3344; и Wolff, et al., Science, 1990, 247, 1465-1468). Такой подход с успехом применяют для создания защитного иммунитета против вируса гриппа у мышей и кур, против бычьего вируса 1 герпеса у мышей и крупного рогатого скота и против вируса бешенства у мышей (Сох, et al., J. Virol., 1993, 67, 5664-5667; Fynan, et al., DNA and Cell Biol., 1993, 12, 785-789; Ulmer, et al. , Science, 1993, 259, 1745-1749; и Xiang, et al., Virol., 1994, 199, 132-140). В большинстве случаев с контролем инфекции ассоциируются сильное, но высоковариабельное антитело и цитотоксические Т-клеточные ответы. В действительности, возможность генерации ЦТЛ с долго сохраняющейся памятью без использования печеночного переносчика делает такой подход особенно привлекательным по сравнению со способами, включающими вакцины с убитыми вирусами и генерирования реакции ЦТЛ, которая не только защищает против острой инфекции, но также может быть благоприятной при уничтожении персистентной вирусной инфекции (Wolff, et al., Science, 1990, 247, 1465-1468; Wolff, et al., Hum. Mol. Genet., 1992, 1, 363-369; Manthorpe, et al., Human Gene Therapy, 1993, 4, 419-431; Ulmer, et al., Science, 1993, 259, 1745-1749; Yankauckas, et al. , DNA and Cell Biol., 1993, 12, 777-783; Montgomery, et al., DNA and Cell Biol., 1993, 12, 777-783; Fynan, et al., DNA and Cell Blol., 1993, 12, 785-789; Wang, et al., Proc. Natl. Acad. Sci. USA, 1993, 90, 4156-4160; Wang, et al. , DNA and Cell Biol., 1993, 12, 799-805; Xiang, et al., Virol., 1994, 199, 132-140; и Davis, et al., Hum. Mol. Genet., 1993, 11, 1847-1851), а заражение HCV и HBV являются заболеваниями человека, имеющими значение во всем мире.
В настоящее время не существует универсального высокоэффективного способа лечения хронической HBV- и/или HCV-инфекции. Разработка вакцинной стратегии против HBV и/или HCV осложнена не только значительной гетерогенностью изолятов HBV и HCV, но также смесью гетерогенных геномов в пределах изолята (Martell, et al. , J. Virol., 1992, 66, 3225). Кроме того, вирус содержит высоковариабельную область оболочки.
Вакцинация и иммунизация обращаются, главным образом, к введению невирулентного фактора, против которого иммунная система индивидуума может инициировать иммунный ответ, который затем будет полезен для защиты против заражения патогеном. Иммунная система идентифицирует вторгающиеся "чужеродные" композиции и факторы, в первую очередь, путем опознавания белков и других больших молекул, которые, по природе, отсутствуют у индивидуума. Чужеродный белок представляет мишень, против которой осуществляется иммунный ответ.
В заявке на международный патент PCT/US90/01348 дается информация о последовательностях клонов генома HCV, аминокислотных последовательностях белков вируса HCV и способах приготовления и применения таких композиций, включая вакцины против HCV, содержащие белки HCV и пептиды, полученные из них.
В каждой из заявок на патент США, зарегистрированных 26 января 1993, регистрационный номер 08/008342, 11 марта 1993, регистрационный номер 08/029336, 21 сентября 1993, регистрационный номер 08/125012, в заявке на международный патент, зарегистрированной 26 января 1994, регистрационный номер PCT/US94/00899, и в заявке на патент США, зарегистрированной 1 апреля 1994, регистрационный номер 08/221579, содержится описание протоколов генетической иммунизации. В каждой из них описываются вакцины против HCV.
В заявке на патент США, зарегистрированной 5 октября 1994, регистрационный номер 08/318248, которая включена в настоящее изобретение в качестве ссылки, описываются генетические конструкции, содержащие нуклеотидные последовательности, кодирующие коровый белок HCV, которые пригодны в качестве вакцины. Вакцина на основе корового белка HCV экспрессирует высокий уровень корового антигена in vitro и индуцирует сильный иммунный ответ in vivo.
Сохраняется потребность в вакцинах, пригодных для защиты индивидуумов от заражения вирусом гепатита В и/или вирусом гепатита С. Сохраняется и потребность в способах защиты индивидуумов от заражения вирусом гепатита В и/или вирусом гепатита С.
Краткое изложение сущности изобретения
Настоящее изобретение относится к молекулам рекомбинантной нуклеиновой кислоты, содержащим нуклеотидную кодирующую последовательность, которая кодирует слитый белок. Слитый белок содержит S-генный продукт вируса гепатита В, соединенный с аминокислотами 1-69 корового белка вируса гепатита С.
Настоящее изобретение относится к фармацевтической композиции, содержащей молекулу рекомбинантной нуклеиновой кислоты, которая содержит нуклеотидную кодирующую последовательность, которая кодирует слитый белок. Слитый белок содержит S-генный продукт вируса гепатита В, соединенный с аминокислотами 1-69 корового белка вируса гепатита С. Нуклеотидная кодирующая последовательность, которая кодирует слитый белок, операбельно связана с регуляторными элементами, функциональными в человеческих клетках. Фармацевтическая композиция также содержит фармацевтически приемлемый носитель или разбавитель.
Настоящее изобретение относится к способам иммунизации индивидуума, восприимчивого к или инфицированного вирусом гепатита С и/или вирусом гепатита В, которые включают стадию введения такому индивидууму фармацевтической композиции, содержащей молекулу рекомбинантной нуклеиновой кислоты, которая содержит нуклеотидную кодирующую последовательность, которая кодирует слитый белок. Слитый белок содержит S-генный продукт вируса гепатита В, соединенный с аминокислотами 1-69 корового белка вируса гепатита С. Нуклеотидная кодирующая последовательность, которая кодирует слитый белок, операбельно связана с регуляторными элементами, функциональными в человеческих клетках. Фармацевтическая композиция также содержит фармацевтически приемлемый носитель или разбавитель. Индивидуум получает количество, эффективное для индуцирования защитного или лечебного иммунного ответа против гепатита В и/или гепатита С.
Настоящее изобретение относится к молекулам нуклеиновой кислоты, содержащим неполный вирусный геном гепатита С, включающий послед. 14.
Настоящее изобретение относится к молекулам нуклеиновой кислоты, содержащим неполный вирусный геном гепатита С, включающий нуклеотидную последовательность, которая кодирует коровый белок HCV, и по крайней мере последние 9 нуклеотидов 5'-нетранслируемой области.
Настоящее изобретение относится к фармацевтической композиции, которая содержит молекулу нуклеиновой кислоты, имеющую неполный вирусный геном гепатита С и по крайней мере кодирующие последовательности послед. 14, операбельно соединенные с регуляторными элементами, функциональными в человеческих клетках.
Настоящее изобретение относится к фармацевтической композиции, которая содержит молекулу нуклеиновой кислоты, имеющую неполный вирусный геном гепатита С, включающий нуклеотидную последовательность, которая кодирует коровый белок HCV, и по крайней мере последние 9 нуклеотидов 5'-нетранслируемой области, операбельно связанные с регуляторными элементами, функциональными в человеческих клетках.
Настоящее изобретение относится к способу иммунизации индивидуума, восприимчивого к или инфицированного вирусом гепатита С, включающему стадию введения индивидууму количества плазмиды pHCV2-1 или плазмиды pHCV4-2, эффективного для индуцирования защитного или лечебного иммунного ответа против гепатита С.
Краткое описание чертежей
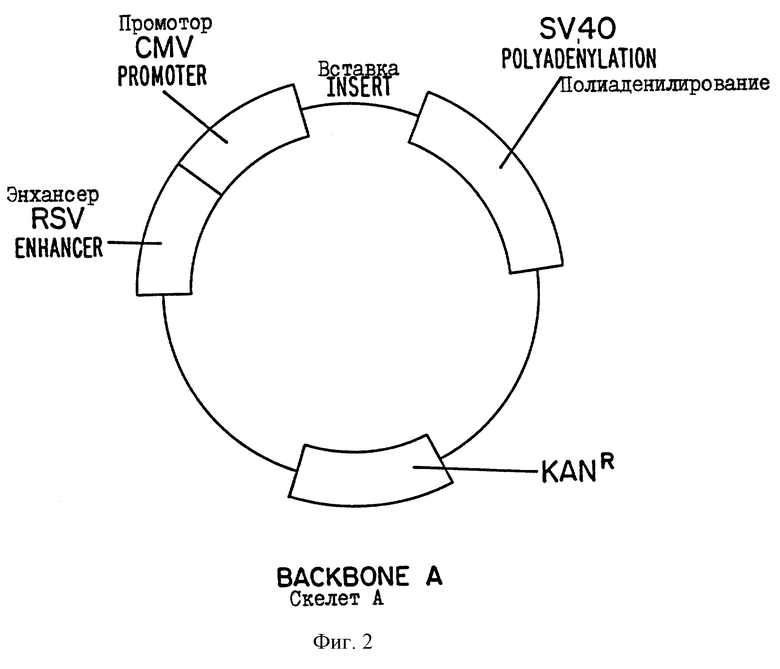
На фиг.1 дается схематическое изображение некоторых конструкций, содержащих коровый ген HCV, S-ген HBV, pre S1 ген HBV и pre S2 ген HBV.
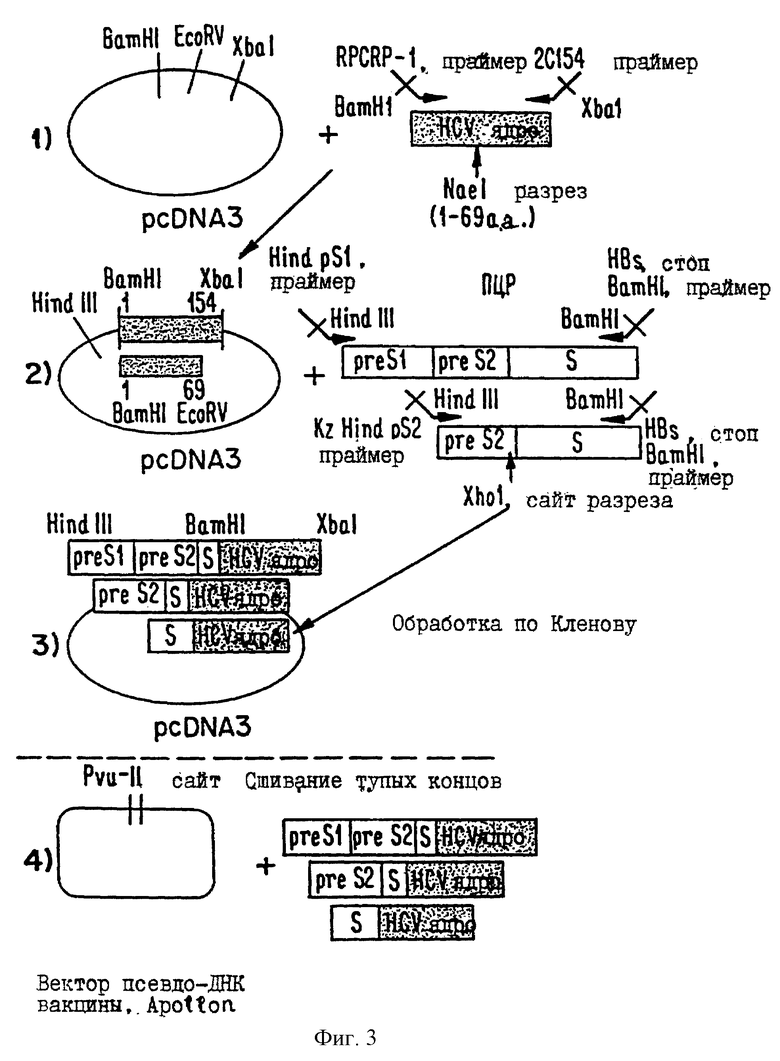
На фиг. 2 приводится диаграмма экспрессионного вектора, в который могут быть вставлены кодирующие последовательности, кодирующие химерный слитый белок HCV/HBV, или в который вставляют кодирующую последовательность HCV, чтобы получить плазмиды pHCV2-1 и pHCV4-2.
На фиг.3 изображаются способы построения гибридных конструкций HBV/HCV.
Подробное описание изобретения
Настоящее изобретение относится к композициям и способам, которыми иммунизируют индивидуум для профилактики и/или лечения от HBV- и/или HCV-инфекции. Для иммунизации против HBV- и/или HCV-инфекции индивидууму вводят молекулы рекомбинантной нуклеиновой кислоты, содержащие нуклеотидную кодирующую последовательность, которая кодирует слитый белок, который содержит S-генный продукт HBV, соединенный с аминокислотами 1-69 корового белка HCV. Слитый белок, кодированный генной конструкцией, экспрессируется клетками индивидуума и служит в качестве иммуногенной мишени, против которой индуцируются анти-HBV и/или анти-HCV иммунные ответы. Для иммунизации против заражения HCV индивидууму вводят генетический материал, содержащий неполный геном вируса гепатита С, но который кодирует коровый белок HCV. Белок, кодированный генной конструкцией, экспрессируется клетками индивидуума и служит в качестве иммуногенной мишени, против которой индуцируется анти-HCV иммунный ответ. Получающиеся в результате иммунные ответы являются вполне фундаментальными; кроме гуморального иммунного ответа выявляются обе ветви клеточного иммунного ответа. Способы настоящего изобретения пригодны для создания профилактического и лечебного иммунитета. Таким образом, способ иммунизации включает как способы защиты индивидуума от заражения HBV и/или HCV, так и способы лечения индивидуума, страдающего от заражения HBV и/или HCV.
Показано, что многие белки, известные ранее для индуцирования гуморального и клеточного иммунного ответа после иммунизации ДНК, являются либо нативными белками клеточной поверхности, либо секретируемыми белками, такими как, например, NP вируса гриппа, HBsAg и гликопротеины вируса бешенства. Хотя вакцина на основе коровой ДНК HCV экспрессирует коровый антиген на высоком уровне in vitro и индуцирует иммунный ответ in vivo, вероятно, что коровый белок HCV остается фиксированным в цитоплазме и не экспрессируется на поверхности клетки или не выделяется из клетки. Следовательно, иммунная реакция может быть увеличена посредством обеспечения экспрессии белка на клеточной поверхности и/или секреции белка. Соответственно, в одном из предпочтительных вариантов осуществления изобретения нуклеотидная кодирующая последовательность, кодирующая коровый белок HCV, соединяется с областями нуклеотидной кодирующей последовательности, кодирующей поверхностный антиген HBV, как показано на фиг.1. Таким образом, слитый химерный белок HBV/HBC может индуцировать иммунный ответ к HBV и/или HCV.
Используемое здесь выражение "неполный геном вируса гепатита С" означает молекулу нуклеиновой кислоты, которая не содержит полной кодирующей последовательности для полипротеина HCV, который кодируется вирусным геномом HCV. Включение неполного генома HCV в клетку не привносит достаточной генетической информации для продуцирования инфекционного вируса.
Используемый здесь термин "коровый белок HCV" означает белок, относящийся к усеченным коровым белкам HCV, таким как белки, имеющие аминокислотную последовательность, представленную послед. 1 и послед. 2, которая содержит 154 аминокислоты, или коровый белок HCV полной длины, имеющий аминокислотную последовательность, представляемую послед. 14 и послед. 15, которая является аминокислотной последовательностью корового белка HCV специфического изолята HCV. Полноразмерный коровый белок HCV обычно имеет 191 аминокислоту, но для целей настоящего изобретения, относящихся к усеченному коровому белку HCV, С-концевые 37 аминокислот (аминокислоты 155-191) удаляют, чтобы усилить способность к секреции слитого белка. Постулировано, что двадцать С-концевых аминокислот полноразмерного корового белка HCV содержат сайты связывания с эндоплазматическим ретикулумом, что приводит, в результате, к удержанию корового белка внутри клеточной цитоплазмы. Кроме того, термин "коровый белок HCV" относится к соответствующим коровым белкам HCV, полноразмерным или усеченным, из дополнительных изолятов HCV, которые могут изменяться. Специалисты в этой области техники могут легко идентифицировать коровый белок HCV из дополнительных изолятов. Нуклеотидные и аминокислотные последовательности корового белка HCV описываются в заявке на патент США, зарегистрированный 5 октября 1994, регистрационный номер 08/318248, которая включена в настоящее изобретение в качестве ссылки. Следует иметь в виду, что нуклеотидные замещения в кодоне могут быть приемлемыми, когда кодирована та же аминокислота. Кроме того, следует также иметь в виду, что нуклеотидные изменения могут быть приемлемыми, когда при этом в результате нуклеотидного замещения получается консервативная аминокислота.
Замечено, что коровый белок HCV существует в двух различных формах в форме закрепленного (заякоренного) корового белка и в форме корового белка вириона. Во время трансляции HCV полипептид-предшественник транслируется и затем процессируется клеточными протеазами и вирусными протеазами, образуя индивидуальные вирусные полипептиды. Полагают, что клеточная сигнальная пептидаза расщепляется в сайте, который отделяет часть полипротеина, которая становится коровым белком, отчасти, которая становится белком оболочки, освобождая, посредством этого, закрепленный коровый белок от белка оболочки. Закрепленный коровый белок, который ассоциирован с эндоплазматическим ретикулумом, затем процессируется до зрелого вирионного корового белка. Закрепленный коровый белок отличается от корового белка вириона тем, что он содержит 18 дополнительных аминокислот на С-конце. В действительности, на вирусных частицах закрепленный белок не обнаружен.
Используемый здесь термин "генная конструкция" относится к молекуле рекомбинантной нуклеиновой кислоты, содержащей нуклеотидную кодирующую последовательность, которая кодирует слитый белок, который содержит по крайней мере 69 N-концевых аминокислот корового белка HCV и полный S-генный продукт HCV, или полноразмерный коровый белок HCV, а также сигналы инициации и терминации, операбельно соединенные с регуляторными элементами, включающими промотор и сигнал полиаденилирования, способные регулировать экспрессию в клетках вакцинированного индивидуума. В некоторых предпочтительных вариантах осуществления изобретения генная конструкция также содержит энхансер, последовательность Козака (GCCGCCATG, послед. 13) и, по крайней мере, фрагмент 5'-UTR HCV.
Используемый здесь термин "генетическая вакцина" относится к фармацевтическому препарату, который содержит генную конструкцию. Генетические вакцины включают фармацевтические препараты, пригодные для индукции профилактического и/или терапевтического иммунного ответа к HCV и/или HBV.
В соответствии с настоящим изобретением, генные конструкции вводят в клетки индивидуума, где они экспрессируются, продуцируя, таким образом, слитый белок корового белка HCV и поверхностного антигена HBV или полноразмерный коровый белок HCV. Регуляторные элементы генных конструкций изобретения способны управлять экспрессией в человеческих клетках. Регуляторные элементы включают промотор и сигнал полиаденилирования. Кроме того, в генную конструкцию также могут включаться другие элементы, такие как энхансер и последовательность Козака.
При захвате клеткой генная конструкция настоящего изобретения может оставаться в клетке в качестве функционирующей внехромосомной молекулы, или она может объединиться с хромосомной ДНК клетки. ДНК может быть введена в клетки, где она остается как отдельный генетический материал в форме плазмиды. С другой стороны, в клетку может быть введена линейная ДНК, которая может объединяться с хромосомой. При введении ДНК в клетку могут быть добавлены реагенты, которые промотируют интеграцию ДНК с хромосомой. В молекулу ДНК также могут быть включены ДНК-последовательности, которые пригодны для промотирования интеграции. С другой стороны, в клетку можно ввести РНК. Также предполагается создать генную конструкцию в виде линейной минихромосомы, включающей центромеру, теломеры и источник репликации.
В одном из предпочтительных вариантов осуществления изобретения генная конструкция содержит молекулы рекомбинантной нуклеиновой кислоты, содержащие нуклеотидную кодирующую последовательность, которая кодирует слитый белок, содержащий области S-генного продукта HBV, соединенного с коровым белком HCV, предпочтительно с усеченным коровым белком HCV.
В некоторых предпочтительных вариантах осуществления изобретения молекула рекомбинантной нуклеиновой кислоты содержит нуклеотидную кодирующую последовательность, которая кодирует слитый белок, содержащий полный S-генный продукт HBV, соединенный с аминокислотами 1-69 корового белка HCV. Нуклеотиды, кодирующие С-концевые аминокислоты S-генного продукта HBV, соединяются с нуклеотидами, кодирующими N-концевые аминокислоты усеченного корового белка HCV.
В некоторых предпочтительных вариантах осуществления изобретения молекула рекомбинантной нуклеиновой кислоты содержит нуклеотидную кодирующую последовательность, которая кодирует слитый белок, содержащий полный S-генный продукт HBV, соединенный с аминокислотами (1-70)-(1-154) усеченного корового белка HCV. Усеченный коровый белок HCV может содержать аминокислоты 1-70, 1-71, 1-72, 1-73, 1-74, ..., 1-150, 1-151, 1-152, 1-153 или 1-154. Использованные здесь обозначения" (1-70)-(1-154)" или "1-70, 1-71, 1-72, 1-73, 1-74, . .., 1-150, 1-151, 1-152, 1-153 или 1-154" применяются как взаимозаменяемые для описания каждой из аминокислотных последовательностей фрагментов усеченного корового белка HCV, которые включают аминокислоты 1-69 плюс дополнительные С-концевые остатки до 154 остатка. Нуклеотиды, кодирующие С-концевые аминокислоты S-генного продукта HBV, соединены с нуклеотидами, кодирующими N-концевые аминокислоты усеченного корового белка HCV.
В некоторых предпочтительных вариантах осуществления изобретения молекула рекомбинантной нуклеиновой кислоты содержит нуклеотидную кодирующую последовательность, которая кодирует слитый белок, содержащий фрагмент рrе S2-генного продукта HBV, соединенного с полным S-генным продуктом HBV, соединенного с аминокислотами 1-69 усеченного корового белка HCV. Фрагмент рrе S2-генного продукта HBV может включать С-концевую аминокислоту, или 2 С-концевых аминокислоты, или 3 С-концевых аминокислоты, или 4 С-концевых аминокислоты, . . . , или полный рrе S2-генный продукт HBV. Используемое здесь выражение "фрагмент рrе S2-генного продукта HBV может включать С-концевую аминокислоту, или 2 С-концевых аминокислоты, или 3 C-концевых аминокислоты, или 4 C-концевых аминокислоты, ..., или полный рrе S2-генный продукт HBV" предназначено для описания аминокислотных последовательностей, которые включают фрагменты pre S2-генного продукта HBV, начиная с фрагмента, который включает только С-концевой остаток, до фрагментов, которые включают дополнительные остатки непосредственно от N-концевого до С-концевого остатка, т. е. последний С-концевой остаток, последние два С-концевых остатка, последние три С-концевых остатка и так далее до присутствия полного pre S2 белка. Нуклеотиды, кодирующие С-концевые аминокислоты pre S2-генного продукта HBV, соединяются с нуклеотидами, кодирующими N-концевые аминокислоты S-генного продукта HBV. Нуклеотиды, кодирующие С-концевые аминокислоты S-генного продукта HBV, в свою очередь, соединяются с нуклеотидами, кодирующими N-концевые аминокислоты усеченного корового белка HCV.
В некоторых предпочтительных вариантах осуществления изобретения молекула рекомбинантной нуклеиновой кислоты содержит нуклеотидную кодирующую последовательность, которая кодирует слитый белок, содержащий фрагмент pre S2-генного продукта HBV, соединенный с полным S-генным продуктом HBV, соединенным с аминокислотами (1-70)-(1-154) усеченного корового белка HCV. Фрагмент pre S2-генного продукта HBV может включать С-концевую аминокислоту, или 2 С-концевых аминокислоты, или 3 С-концевых аминокислоты, или 4 С-концевых аминокислоты, ..., или полный pre S2-генный продукт HBV. Усеченный коровый белок HCV может содержать аминокислоты 1-70, 1-71, 1-72, 1-73, 1-74, . . ., 1-150, 1-151, 1-152, 1-153 или 1-154. Нуклеотиды, кодирующие С-концевые аминокислоты pre S2-генного продукта HBV, соединяются с нуклеотидами, кодирующими N-концевые аминокислоты S-генного продукта HBV. Нуклеотиды, кодирующие С-концевые аминокислоты S-генного продукта HBV, в свою очередь, соединяются с нуклеотидами, кодирующими N-концевые аминокислоты усеченного корового белка HCV.
В некоторых предпочтительных вариантах осуществления изобретения молекула рекомбинантной нуклеиновой кислоты содержит нуклеотидную кодирующую последовательность, которая кодирует слитый белок, содержащий фрагмент pre S1-генного продукта HBV, соединенного с полным S-генным продуктом HBV, соединенного с аминокислотами 1-69 усеченного корового белка HCV. Фрагмент pre S1-генного продукта HBV может включать С-концевую аминокислоту, или 2 С-концевых аминокислоты, или 3 С-концевых аминокислоты, или 4 С-концевых аминокислоты, . .., или полный pre S1-генный продукт HBV. Используемое здесь выражение "фрагмент pre S1-генного продукта HBV может включать С-концевую аминокислоту, или 2 С-концевых аминокислоты, или 3 С-концевых аминокислоты, или 4 С-концевых аминокислоты, ..., или полный pre S1-генный продукт HBV" предназначено для описания аминокислотных последовательностей, которые включают фрагменты pre S1-генного продукта HBV, начиная с фрагмента, который включает только С-концевой остаток, до фрагментов, которые включают дополнительные остатки непосредственно от N-концевого до С-концевого остатка, т. е. последний С-концевой остаток, последние два С-концевых остатка, последние три С-концевых остатка и так далее до присутствия полного pre S1 белка. Фрагмент pre S2-генного продукта HBV может включать С-концевую аминокислоту, или 2 С-концевых аминокислоты, или 3 С-концевых аминокислоты, или 4 С-концевых аминокислоты, ..., или полный pre S2-генный продукт HBV. Нуклеотиды, кодирующие С-концевые аминокислоты pre S1-генного продукта HBV, соединяются с нуклеотидами, кодирующими N-концевые аминокислоты pre S2-генного продукта HBV. Нуклеотиды, кодирующие С-концевые аминокислоты pre S2-генного продукта HBV, в свою очередь, соединяются с нуклеотидами, кодирующими N-концевые аминокислоты S-генного продукта HBV. Нуклеотиды, кодирующие С-концевые аминокислоты S-генного продукта HBV, в свою очередь, соединяются с нуклеотидами, кодирующими N-концевые аминокислоты усеченного корового белка HCV.
В некоторых предпочтительных вариантах осуществления изобретения молекула рекомбинантной нуклеиновой кислоты содержит нуклеотидную кодирующую последовательность, которая кодирует слитый белок, содержащий фрагмент pre S1-генного продукта HBV, соединенный с фрагментом pre S2-генного продукта HBV, соединенного с S-генным продуктом HBV, соединенным с аминокислотами (1-70)-(1-154) усеченного корового белка HCV. Усеченный коровый белок HCV может содержать аминокислоты 1-70, 1-71, 1-72, 1-73, 1-74, ..., 1-150, 1-151, 1-152, 1-153 или 1-154. Фрагмент pre S1-генного продукта HBV может включать С-концевую аминокислоту или 2 С-концевых аминокислоты, или 3 С-концевых аминокислоты, или 4 С-концевых аминокислоты, ..., или полный pre S1-генный продукт HBV. Фрагмент pre S2-генного продукта HBV может включать С-концевую аминокислоту, или 2 С-концевых аминокислоты, или 3 С-концевых аминокислоты, или 4 С-концевых аминокислоты, ..., или полный pre S2-генный продукт HBV. Нуклеотиды, кодирующие С-концевые аминокислоты pre S1-генного продукта HBV, соединяются с нуклеотидами, кодирующими N-концевые аминокислоты pre S2-генного продукта HBV. Нуклеотиды, кодирующие С-концевые аминокислоты pre S2-генного продукта HBV, в свою очередь, соединяются с нуклеотидами, кодирующими N-концевые аминокислоты S-генного продукта HBV. Нуклеотиды, кодирующие С-концевые аминокислоты S-генного продукта HBV, в свою очередь, соединяются с нуклеотидами, кодирующими N-концевые аминокислоты усеченного корового белка HCV.
Предполагается, что нуклеотидные кодирующие последовательности могут быть соединены одна с другой с помощью или без помощи нуклеотидных спейсеров или линкеров, таким образом, что нуклеотидная кодирующая последовательность в прямом направлении остается в рамке. Таким образом, нуклеотидные кодирующие последовательности могут быть соединены непосредственно, т.е. без каких-либо нуклеотидных спейсеров. С другой стороны, между соседними нуклеотидными последовательностями может быть вставлен спейсер, содержащий от 3 до 60 нуклеотидов. Например, нуклеотидный спейсер может быть вставлен между нуклеотидными кодирующими последовательностями для S-гена HBV и усеченного корового гена HCV, между pre S2-геном HBV и pre S-геном HBV, и pre S1-геном HBV и pre S2-геном HBV. Спейсер или линкер может содержать сайты для рестрикционных эндонуклеаз для целей клонирования.
Также предполагается, что молекула рекомбинантной нуклеиновой кислоты, содержащая нуклеотидную кодирующую последовательность, которая кодирует слитый белок, который может содержать менее полного S-генного продукта HBV, соединенного
с аминокислотами 1-69 усеченного корового белка HCV, по существу, не изменяет эффективности вакцины. Также предполагается, что нуклеотидные замещения могут быть осуществлены в нуклеотидной кодирующей последовательности, не затрагивая аминокислотную последовательность слитого белка. Также предполагается, что замещения консервативных аминокислот могут быть осуществлены в слитом белке без существенного уменьшения иммуногенной активности слитого белка.
В некоторых предпочтительных вариантах осуществления изобретения генная конструкция включает по крайней мере фрагмент 5'-UTR HCV, содержащий последние 9 нуклеотидов 5'-UTR HCV. Используемый здесь термин "последние 9 нуклеотидов 5'-UTR HCV" предназначается для обозначения 9 самых крайних 3'-нуклеотидов 5'-UTR, иными словами для обозначения 9 нуклеотидов 5'-UTR, которые непосредственно предшествуют кодирующей последовательности. Последние 9 нуклеотидов 5'-UTR HCV предпочтительного варианта осуществления изобретения показаны как послед. 3.
В некоторых предпочтительных вариантах осуществления изобретения фрагмент 5'-UTR, который включает последние 9 нуклеотидов 5'-UTR HCV, содержит последние 25 нуклеотидов 5'-UTR HCV. В некоторых предпочтительных вариантах осуществления изобретения фрагмент 5'-UTR, который включает последние 9 нуклеотидов 5'-UTR HCV, содержит последние 50 нуклеотидов 5'-UTR HCV. В некоторых предпочтительных вариантах осуществления изобретения фрагмент 5'-UTR, который включает последние 9 нуклеотидов 5'-UTR HCV, содержит последние 75 нуклеотидов 5'-UTR HCV. В некоторых предпочтительных вариантах осуществления изобретения фрагмент 5'-UTR, который включает последние 9 нуклеотидов 5'-UTR HCV, содержит последние 100 нуклеотидов 5'-UTR HCV. В некоторых предпочтительных вариантах осуществления изобретения фрагмент 5'-UTR, который включает последние 9 нуклеотидов 5'-UTR HCV, содержит последние 150 нуклеотидов 5'-UTR HCV. В некоторых предпочтительных вариантах осуществления изобретения фрагмент 5'-UTR, который включает последние 9 нуклеотидов 5'-UTR HCV, содержит последние 200 нуклеотидов 5'-UTR HCV. В некоторых предпочтительных вариантах осуществления изобретения фрагмент 5'-UTR, который включает последние 9 нуклеотидов 5'-UTR HCV, содержит последние 250 нуклеотидов 5'-UTR HCV. В некоторых предпочтительных вариантах осуществления изобретения фрагмент 5'-UTR, который включает последние 9 нуклеотидов 5'-UTR HCV, содержит последние 300 нуклеотидов 5'-UTR HCV.
В некоторых предпочтительных вариантах осуществления изобретения генная конструкция включает полную 5'-UTR HCV. В некоторых предпочтительных вариантах осуществления изобретения генная конструкция включает 9 самых крайних 3'-нуклеотидов 5'-UTR HCV. Полная 5'-UTR HCV предпочтительного варианта осуществления изобретения приводится как послед. 4.
В соответствии с другими аспектами настоящего изобретения генная конструкция включает кодирующие последовательности, которые кодируют полноразмерный коровый белок HCV. В некоторых предпочтительных вариантах осуществления изобретения генная конструкция кодирует "якорную" форму полноразмерного корового белка HCV. Закрепленный коровый белок является предпочтительным в некоторых вариантах осуществления изобретения по следующим причинам. Во-первых, полагают, что желательно правильно компартментализировать коровый белок в эндоплазматический ретикулум, где коровый белок обнаруживается в первую очередь в естественно инфицированных клетках. Если белок компартментализирован неправильно, существует опасность снижения общей экспрессии этого белка вследствие разрушения или отравления клетки. Далее, так как считается, что клеточная протеаза расщепляет закрепленный коровый белок до корового вирионного, оба вида могут присутствовать в клетках, экспрессирующих закрепленную форму. Если между двумя видами существуют различия в антигенности, экспрессией закрепленной формы полноразмерного корового белка потенциально генерируется более широкий иммунный ответ против ядра. Более того, дополнительные 18 аминокислот в закрепленном коровом белке могут содержать либо эпитопы для антител, либо эпитопы для цитотоксических Т-клеток, либо то и другое, экспрессия закрепленной формы обеспечивает присутствие этих областей, которые могут расширить иммунный ответ к коровому белку.
В некоторых предпочтительных вариантах осуществления изобретения генная конструкция кодирует полноразмерный коровый белок, который состоит из аминокислотной последовательности послед. 14 и послед. 15. В некоторых предпочтительных вариантах осуществления изобретения генная конструкция содержит кодирующую последовательность в послед. 14.
В некоторых предпочтительных вариантах осуществления изобретения генная конструкция содержит послед. 14.
В некоторых предпочтительных вариантах осуществления изобретения используемый вектор выбирают среди векторов, описанных на фиг.2. В некоторых предпочтительных вариантах осуществления изобретения в скелет А вставляют нуклеотиды 342-923 послед. 14, чтобы образовать плазмиду pHCV2-l. В некоторых предпочтительных вариантах осуществления изобретения в скелет А вставляют нуклеотидную послед. 14, чтобы образовать плазмиду pHCV4-2.
В плазмидах pHCV2-l и pHCV4-2 кодирующая последовательность, кодирующая полноразмерный коровый белок HCV, находится под регуляторным контролем немедленно-раннего промотора CMV и SV-40-минорного сигнала полиаденилирования. Конструкции могут, необязательно, содержать точку начала репликации SV-40.
Регуляторные элементы, необходимые для генной экспрессии молекулы ДНК, включают промотор, инициирующий кодон, стоп-кодон и сигнал полиаденилирования. Кроме того, для экспрессии гена часто требуются энхансеры. Необходимо, чтобы эти элементы были операбельно соединены с последовательностью, которая кодирует слитый белок или полноразмерный коровый белок HCV, и чтобы регуляторные элементы являлись операбельными в организме индивидуума, которому они вводятся.
Вообще, считается, что инициирующие кодоны и стоп-кодоны являются частью нуклеотидной последовательности, которая кодирует слитый белок или полноразмерный коровый белок HCV.
Используемые промоторы и сигналы полиаденилирования должны быть функциональными в пределах клеток индивидуума. Для максимального продуцирования белка в конструкцию вводят регуляторные последовательности, которые можно выбрать среди последовательностей, которые вполне соответствуют экспрессии гена в клетках. Кроме того, могут быть выбраны кодоны, которые наиболее эффективно транскрибируются в клетке. Специалист в этой области техники может получить ДНК-конструкции, которые являются функциональными в клетках.
Примеры промоторов, пригодных для осуществления настоящего изобретения, особенно при получении генетической вакцины для людей, включают, но не ограничиваются перечисленными: промоторы из вируса 40 обезьян (SV40), промотор вируса опухоли молочной железы мышей (MMTV), вируса иммунодефицита человека (ВИЧ), такой как промотор длинного концевого повтора (LTR) ВИЧ, вируса Молони, ALV, цитомегаловируса (CMV), такой как немедленно-ранний промотор CMV, вируса Эпштейна-Барр (EBV), вируса саркомы Рауса (RSV), а также промоторы из человеческих генов, такие как человеческий актин, человеческий миозин, человеческий гемоглобин, человеческий мышечный креатин и человеческий металлотионеин.
Примеры сигналов полиаденилирования, пригодных для осуществления настоящего изобретения, особенно при получении генетической вакцины для людей, включают, но не ограничиваются перечисленными: сигналы полиаденилирования SV-40 и сигналы полиаденилирования LTR. В частности, используют сигнал полиаденилирования SV-40, который присутствует в плазмиде рСЕР4 (Invitrogen, Сан-Диего, СА), упоминаемый как сигнал полиаденилирования SV-40.
Помимо регуляторных элементов, необходимых для экспрессии гена, в генную конструкцию также могут включаться другие элементы, к таким дополнительным элементам относятся энхансеры. Энхансер можно выбрать из группы, включающей, но не ограничивающейся перечисленными: человеческий актин, человеческий миозин, человеческий гемоглобин, человеческий мышечный креатин и вирусные энхансеры, такие как энхансеры из CMV, RSV и EBV.
Генные конструкции могут снабжаться точкой начала репликации млекопитающего, чтобы сохранить конструкцию вне хромосомы и продуцировать множество копий конструкции в клетке. Плазмиды pCEP4 и pREP4 от Invitrogen (Сан-Диего, СА) содержат точку начала репликации вируса Эпштейна-Барр и кодирующую область ядерного антигена EBNA-1, которая продуцирует высококопийную эписомальную репликацию без интеграции.
В некоторых предпочтительных вариантах осуществления изобретения используемую генную конструкцию выбирают среди векторов, показанных на фиг.2. В некоторых предпочтительных вариантах осуществления изобретения нуклеотидную кодирующую последовательность, кодирующую слитый белок, вставляют в скелет А.
В экспрессирующих векторах настоящего изобретения нуклеотидная кодирующая последовательность, кодирующая слитый белок, находится под регуляторным контролем немедленно-раннего промотора CMV и минорного сигнала полиаденилирования SV-40. Конструкции могут содержать, необязательно, точку начала репликации SV-40.
Способы введения включают, но не ограничиваются перечисленными: внутримышечный, интраперитонеальный, интрадермальный, подкожный, внутривенный, внутриартериальный, внутриглазной и пероральный способ, а также трансдермальное введение или введение с помощью ингаляции или суппозитория. Предпочтительными способами введения являются внутримышечный, интраперитонеальный, интрадермальный и подкожная инъекция. Доставка генных конструкций, которые кодируют химерный белок HBV/HCV или полноразмерный коровый белок HCV, может создать защитные свойства слизистых оболочек у индивидуумов, иммунизированных посредством способа введения, при котором материал присутствует в тканях, связанных с мукозальным иммунитетом. Так, в некоторых примерах генную конструкцию доставляют посредством введения в щечный карман рта индивидуума.
Генные конструкции могут вводиться средствами, к которым относятся, но не ограничиваются перечисленными: традиционные шприцы, устройства для безыгольных инъекций или "бомбардирующие микрочастицами генные пушки" ("microprojectile bombardment gene guns"). С другой стороны, генную вакцину можно различными способами вводить в клетки, которые удалены из индивидуума. Такие способы включают, например, трансфекцию ех vivo, электропорацию, микроинъекцию и бомбардировку микрочастицами. После захвата клетками генной конструкции их реплантируют индивидууму. Предполагается, что индивидууму могут быть имплантированы другие неиммуногенные клетки, в которые включены генные конструкции, даже если вакцинированные клетки первоначально взяты от другого индивидуума.
В соответствии с некоторыми предпочтительными вариантами осуществления настоящего изобретения генную конструкцию вводят индивидууму, используя устройство для безыгольных инъекций. В соответствии с некоторыми предпочтительными вариантами осуществления настоящего изобретения генную конструкцию вводят индивидууму, одновременно интрадермально, подкожно и внутримышечно, используя устройство для безыгольных инъекций. Устройства для безыгольных инъекций хорошо известны и широко доступны. Специалист в этой области техники может, следуя данным здесь указаниям, использовать устройства для безыгольных инъекций для доставки генетического материала к клеткам индивидуума. Устройства для безыгольных инъекций хорошо подходят для доставки генетического материала в любую ткань. Они особенно пригодны для доставки генетического материала в клетки кожи и мышечные клетки. В некоторых предпочтительных вариантах осуществления изобретения устройство для безыгольных инъекций может использоваться для продвижения жидкости, которая содержит молекулы ДНК, к поверхности кожи индивидуума. Жидкость продвигается с достаточной скоростью, такой, что после "соударения" с кожей жидкость пропитывает поверхность кожи, проходит в кожу и мышечную ткань под ней. Таким образом, генетический материал вводится одновременно интрадермально, подкожно и внутримышечно. В некоторых вариантах осуществления изобретения устройство для безыгольных инъекций может применяться для доставки генетического материала к тканям других органов, чтобы ввести молекулу нуклеиновой кислоты в клетки этого органа. Генетические вакцины по настоящему изобретению содержат от 1 нанограмма до 1000 микрограммов ДНК. В некоторых предпочтительных вариантах осуществления изобретения вакцины содержат от 10 нанограммов до 800 микрограммов ДНК. В некоторых предпочтительных вариантах осуществления изобретения вакцины содержат от 0,1 до 500 микрограммов ДНК. В некоторых предпочтительных вариантах осуществления изобретения вакцины содержат от 1 до 350 микрограммов ДНК. В некоторых предпочтительных вариантах осуществления изобретения вакцины содержат от 25 до 250 микрограммов ДНК. В некоторых предпочтительных вариантах осуществления изобретения вакцины содержат около 100 микрограммов ДНК.
Генетические вакцины по настоящему изобретению составляются в соответствии со способом введения, который используют. Специалист в этой области техники может легко составить фармацевтическую композицию, которая содержит генную конструкцию. В некоторых случаях используют изотонический состав. Как правило, добавки для придания изотоничности могут включать хлорид натрия, декстрозу, маннит, сорбит и лактозу. В некоторых случаях предпочтительны такие изотонические растворы, как забуференный фосфатом физиологический раствор. К стабилизаторам относятся желатин и альбумин. В некоторых вариантах осуществления изобретения к составу добавляют сосудосуживающее средство. Фармацевтические препараты по настоящему изобретению относятся к стерильным и свободным от пирогенов.
Генные конструкции изобретения могут быть составлены или могут вводиться в сочетании со средствами, которые увеличивают поглощение и/или экспрессию генной конструкции клетками по сравнению с поглощением и/или экспрессией генной конструкции клетками, которые присутствуют, когда идентичную генетическую вакцину вводят в отсутствие таких средств. Такие средства и протоколы их введения в сочетании с генными конструкциями описаны в заявках на патент США, зарегистрированных 26 января 1993, регистрационный номер 08/008342, 11 марта 1993, регистрационный номер 08/029336, 21 сентября 1993, регистрационный номер 08/125012, заявке на международный патент, зарегистрированной 26 января 1994, регистрационный номер PCT/US94/00899, и в заявке на патент США, зарегистрированной 1 апреля 1994, регистрационный номер 08/221579, которые все включены в настоящее изобретение в качестве ссылок. Примерами таких средств являются СаРО4, декстран DEAE, анионогенные липиды; активные ферменты внеклеточного матрикса; сапонины; лектины; эстрогенные соединения и стероидные гормоны; низшие гидроксилированные алкилы; диметилсульфоксид (ДМСО); мочевина; и сложные эфиры, анилиды, амидины, уретаны бензойной кислоты и их гидрохлориды, такие как относящиеся к группе местных анестетиков. Кроме того, генные конструкции инкапсулируют или вводят в сочетании с липидными и поликатионными комплексами.
ПРИМЕРЫ
Пример 1. Конструирование и построение плазмид экспрессии HBV/HCV
Создают большой S НВ клон, содержащий полные последовательности рrе S1, pre S2 и S (adr-l). Нуклеотидные и аминокислотные последовательности гена pre Sl/pre S2/S изображаются в послед. 5. Коротко, 100 мкл сыворотки, полученной от пациента с хроническим гепатитом, связанным с HBV, инкубируют при 70oС в течение 3 часов в смеси с протеиназой К (100 мкг/мл), 0,5% ДСН, 5 мМ ЭДТК и 10 мМ трис-HCl, рН 8,0. Раствор экстрагируют фенол-хлороформом, и осаждают ДНК этанолом. Проводят реакцию ПЦР в 100 мкл смеси, содержащей 10 мкл образца сывороточной ДНК, 2,5 Е Taq-полимеразы (Perkin-Elmer Cetus, Norwalk, CT), по 50 мкМ каждого дезоксинуклеозид-5'-трифосфата (dNTP) и 0,4 мкМ праймера HBs-3 5'-GGGTCACCATATTCTTGGGAA- 3' (послед. 6) и HBs-1 5'-GCAGCAAAGCCCAAAAGACCC-3' (послед. 7). Реакционная смесь проходит 35 реакционных циклов. Амплифицированную ДНК клонируют в векторе pCRТМ, используя набор для клонирования ТА (Invitrogen). Также создают клон HCV (ТН, генотип II) от пациента с хроническим инфекционным гепатитом С, как описано в работе Wakita, et al. , J. Biol. Chem., 1994, 269, 1405-1410, которая включена в настоящее изобретение в качестве ссылки.
На фиг. 3 изображаются предпочтительные способы, применяемые для получения гибридных конструкций HCV/HBV. Первая стадия требует ПЦР-амплификации корового гена усеченного HCV. Коротко, 10 мкг ТН-ДНК амплифицируют RPCRP-1-праймером 5'-CGGGATCCATGAGCACGAATCCTAAACC-3' (послед. 8) и праймером 2C154XBA-R 5'-TCTCTAGATTACTAGCCATGCGCCAAGGCCCTGG-3' (послед. 9). Продукт ПЦР расщепляют BamHI и XbaI, и лигируют в вектор pcDNA3 (Invitrogen). Последние 20 аминокислот усеченного корового белка HCV прочно связывают с ER-мембраной. Учитывая эту информацию, в отношении секреции слитого белка, удаляют конечные 37 аминокислот полноразмерного корового белка HCV (аминокислоты со 155 по 191). Также получают другую плазмиду, кодирующую коровый ген укороченного или усеченного HCV (коровые аминокислоты HCV с 1 по 69), так как возможно, что слитые белки, содержащие коровые аминокислоты HCV с 1 по 154, будут слишком длинными, чтобы быть секретированными. Чтобы получить укороченную или усеченную HCV-конструкцию, амплифицированную ДНК расщепляют BamHI и NaeI и лигируют в сайт BamHI сайт EcoRI в векторе pcDNA3.
Наконец, осуществляют ПЦР НВs-продукта между праймером HBs-стоп-ВаmНI 5'-CATGGATCCAATGTATACCCAAAGACA-3' (послед. 10) и праймером Hind pSl 5'-AGACACAAGCTTATGGGAGGTTGGTCTTCCAAAC-3' (послед. 11) или праймером Kz Hind pS2 5'-AGACACAAGCTTGCCGCCATGCAGTGGAACT-3' (послед. 12). Амплифицированную ДНК расщепляют BamHI и HindIII, и лигируют в сайт BamHI - сайт HindIII вектора pcDNA3, который включает коровый ген HCV. Ген, кодирующий малый S, продуцируют путем расщепления продукта ПЦР среднего S (pre S2-S) Xho-I с последующей обработкой по Кленову. В последовательность обратного направления слитых конструкций pre-S2-S-HCV в праймер Kz Hind pS2 включают последовательность Козака (GCCGCCATG, послед. 13), и все это добавляют к конструкции preS2-S-HCV, поскольку конструкция preS2-S-HCV без последовательности Козака часто транслируется от кодона ATG гена НВs (малый S), а не от кодона ATG гена pre S2. Как показано на фиг.1, получают шесть гибридных конструкций. Осуществляют полный анализ последовательностей всех конструкций, и не обнаруживают мутаций, индуцированных ПЦР. Наконец, эти шесть HB-HCV вставок лигируют с сайтом Pvu-II вакцинного экспрессионного вектора.
Специалист в этой области техники, имеющий ДНК-последовательности, кодирующие усеченный коровый белок HCV, белок pre S1, белок pre S2 и белок S, может создать праймеры для получения любой из химерных генных конструкций настоящего изобретения. Кроме того, можно осуществить замещения нуклеотидных оснований без нарушения связывания праймеров. Кроме того, праймеры могут быть получены с эндонуклеазными сайтами для целей клонирования и лигирования, что известно специалистам в этой области техники. Таким образом, специалист в этой области техники может получить любую генную конструкцию настоящего изобретения посредством конструирования подходящих праймеров и осуществления ПЦР-амплификации. Продукты ПЦР лигируют в экспрессирующий вектор.
Плазмиды, содержащие нуклеотидную кодирующую последовательность для слитых белков HBV/HCV, описанных выше, содержат, каждая, нуклеотидную кодирующую область для слитого белка, размещенную под транскрипционным контролем промотора CMV и энхансерного элемента RSV.
Плазмидный скелет А имеет длину в 3969 пар оснований. Он содержит точку начала репликации PBR для репликации в E.coli. Он также содержит ген устойчивости к канамицину, так что плазмида может быть селектирована в E.coli. Вставки, такие как усеченный или полноразмерный коровый ген HCV, клонируют в полилинкерной области, которая размещает вставку между промотором и сигналом полиаденилирования, операбельно соединенными. Транскрипция клонированных вставок находится под контролем CMV-промоторного и RSV-энхансерного элементов. Сигнал полиаденилирования обеспечен присутствием сигнала SV-40 поли А, расположенного точно в 3' сайта клонирования.
Пример 2. Конструирование и построение экспрессионных пдазмид полноразмерного HCV
Для экспрессии закрепленного корового белка HCV конструируют обе плазмиды pHCV2-1 и pHCV4-2. Закрепленный коровый белок, клеточный предшественник корового вирионного белка, остается ассоциированным с мембраной за счет гидрофобных карбоксильных концов.
Каждая плазмидная конструкция содержит область, кодирующую полноразмерный коровый белок HCV, размещенную под транскрипционным контролем CMV-промотора и RSV-энхансерного элемента. Плазмида pHCV2-1 содержит 9-нуклеотидный фрагмент 5' UTR HCV, который включает 9 концевых 3'-нуклеотидов 5' UTR HCV. Плазмида pHCV4-2 отличается от плазмиды pHCV2-1 тем, что она также содержит полную 5'-UTR HCV.
Специфические для HCV последовательности амплифицируют полимеразной цепной реакцией (ПЦР), используя в качестве матрицы pUC-T7-HCVTH (HCV типа 2), и олигонуклеотиды в качестве праймеров ПЦР содержат последовательности 16, 17 и 18.
Праймеры 5'-ПЦР - послед. 16 и послед. 18 - содержат сайты NatI, а праймер 3'-ПЦР послед. 17 имеет сайт SalI. Три стоп-кодона создают в послед. 17. Послед. 18 и послед. 17 используют в качестве праймеров для генерации продукта ПЦР, который картируется от положения 1 до положения 810 генома HCV, и который содержит полную 5'-нетранслируемую область HCV и коровую кодирующую область.
Использование послед. 16 и послед. 18 в качестве праймеров дает, в результате, продукт ПЦР, картирующийся от положения 333 до положения 810 генома HCV. Этот продукт содержит последние 9 нуклеотидов 5'-нетранслируемой области HCV и полную коровую кодирующую область.
Оба продукта ПЦР расщепляют NatI и SalI (NED), очищают на геле и лигируют в сайты NatI-SalI плазмидного скелета А, показанного на фиг.2, и генерируют pHCV4-2 и pHCV2-l.
Плазмидный скелет А имеет длину в 3969 пар оснований. Он содержит точку начала репликации PBR для репликации в E.coli. Он также содержит ген устойчивости к канамицину, так что плазмида может быть селектирована в E.coll. Вставки, такие как полноразмерный коровый ген HCV, клонируют в полилинкерной области, которая размещает вставку между промотором и сигналом полиаденилирования, операбельно соединенными. Транскрипция клонированных вставок находится под контролем CMV-промоторного и RSV-энхансерного элементов. Сигнал полиаденилирования обеспечен присутствием сигнала SV-40 поли А, расположенного точно в 3' сайта клонирования.
Пример 3. Анализ экспрессии in vitro
Экспрессию полноразмерного корового белка HCV из pHCV2-1 и pHCV4-1 проверяют in vitro в клеточной линии рабдомиосаркомы (RD). Клетки котрансфектируют pHCV2-1 и описанной β-галактозидазной плазмидой pCMVB (Clontech), pHCV4-2 и pCMVB, плазмидным вектором и pCMVB, или псевдотрансфектируют. Все анализы проводят через 48 часов после трансфекции. Эффективность трансфекции определяют, измеряя специфическую активность β-галактозидазы в клеточных лизатах.
Трансфецированные клетки анализируют посредством непрямой иммунофлуоресценции, используя в качестве первичных антител либо антикоровое моноклональное антитело, либо смешанные сыворотки от больных HCV. в качестве вторичных антител используют помеченный ФИТЦ антимышиный IgG (Sigma) и помеченный ФИТЦ античеловеческий IgG (Sigma) соответственно. Иммунное окрашивание заметно в клетках, которые трансфецированы либо pHCV2-l, либо pHCV4-2, но не в клетках, трансфецированных плазмидным вектором или псевдотрансфецированных. Иммунофлуоресценция в фиксированных клетках наблюдается в виде окрашенных пятен, локализованных в цитоплазме, когда в качестве источника первичного антитела используют либо моноклональное антитело, либо смешанные сыворотки пациентов. Оказывается, что нет различий в уровне окрашивания между клетками, трансфецированными pHCV2-l и pHCV4-2. He наблюдается окрашивания в нефиксированных клетках, что указывает, что экспрессированный коровый белок, вероятно, не ассоциирован с поверхностью клеточной мембраны.
Лизаты трансфецированных клеток также анализируют посредством Вестерн-блоттинга и иммунопреципитации. Оказывается, что присутствие или отсутствие 5'-UTR HCV имеет незначительное влияние на экспрессию корового белка, так как pHCV2-l и pHCV4-2 экспрессируют эквивалентные количества корового белка в проверяемых клеточных линиях.
Пример 4. Инокуляция ДНК
Гуморальный иммунитет в группе из шести мышей оценивают после однократной внутримышечной инокуляции 100 мкг pHCV4-2 с добавлением 0,25% бупивакаина. Установлено, что все мыши видоизменяют серологическую специфичность через 21 день после инокуляции ДНК. Гуморальная реакция устойчива в течение нескольких месяцев. В настоящее время исследования продолжаются в направлении оценки действия при многократном дозировании.
Такая вакцина на основе коровой ДНК HCV экспрессирует высокий уровень корового антигена in vitro и индуцирует сильную иммунную реакцию in vivo.








Изобретение относится к биотехнологии и генной терапии и касается вакцины против вирусов гепатита. Сущность изобретения включает молекулу нуклеиновой кислоты, которая содержит неполный геном слитых вирусов гепатита С и гепатита В или неполный геном вируса гепатита С, включающие нуклеотидную последовательность, кодирующую полный коровый белок гепатита С и S-генный белок гепатита В, или неполный вирусный геном гепатита С, включающий нуклеотидную последовательность, кодирующую полный коровый белок гепатита С, операбельно соединенную с регуляторными элементами, функциональными в человеческих клетках. Описываются также фармацевтические композиции, содержащие такие конструкции. Преимущество изобретения заключается в создании генных конструкций, которые могут быть эффективны для защиты против вирусов гепатита В и/или гепатита С. 8 с. и 6 з.п.ф-лы, 3 ил.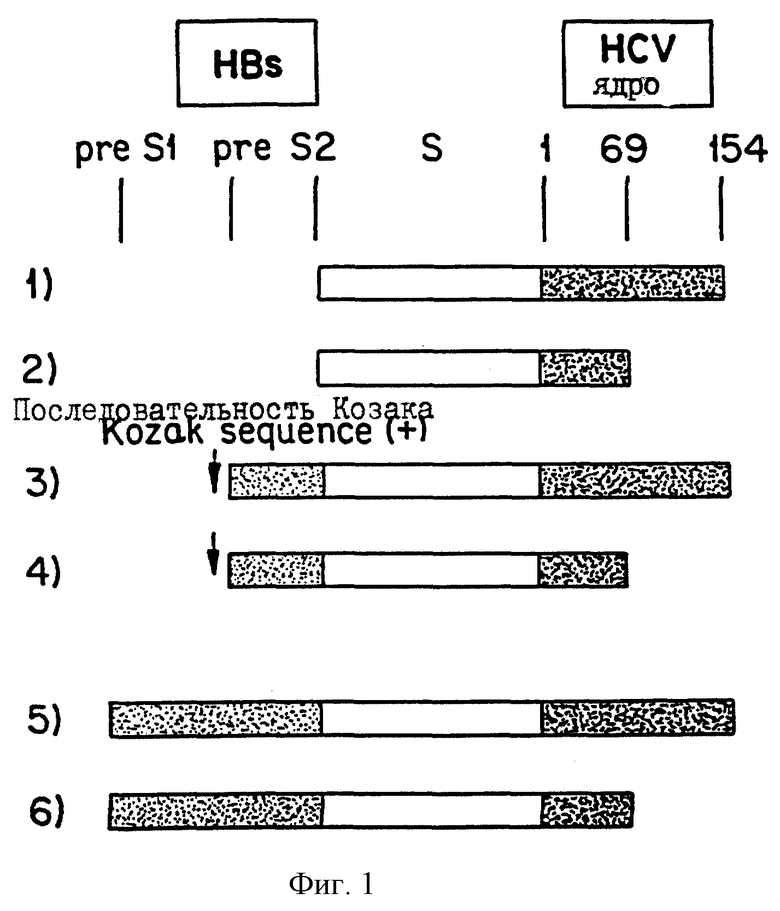
| US 4707542 А, 17.11.1987 | |||
| Способ получения фторидов сульфоновых кислот | 1974 |
|
SU511854A3 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| ЖДАНОВ В.М | |||
| и др | |||
| Вирусные гепатиты | |||
| Пневматический водоподъемный аппарат-двигатель | 1917 |
|
SU1986A1 |

