I. Предпосылки создания изобретения.
Настоящее изобретение относится к общей области биосинтеза серина и родственных серину соединений, в частности триптофана, а также к продуктам и способам, применяемым в таком биосинтезе.
Серин является первичным промежуточным соединением в биосинтезе самых различных клеточных метаболитов, включая такие экономически важные соединения, как холин, глицин, цистеин и триптофан. Кроме того, серин является единственным источником углерода и отвечает за 60 - 75% потребления клеткой C1-звеньев в ходе образования 5,10-метилентетрагидрофолата из тетрагидрофолата. Такие C1-звенья используют в самых различных схемах биосинтеза, в том числе синтезе метионина, монофосфата инозина, других пуринов и некоторых пиримидинов (например, тимидина и гидроксиметилцитидина).
Схема биосинтеза серина, приведенная на фиг.1, как правило, реализуется в самых разнообразных тканях и микроорганизмах. Первая стадия по этой схеме состоит в превращении 3-фосфо-D-глицериновой кислоты (ФГК) под действием фермента 3-фосфоглицератдегидрогеназы (ФГД) в 3-фосфогидроксипировиноградную кислоту (ФПК). Ген, кодирующий ФГД, был клонирован и секвенирован, была также установлена аминокислотная последовательность субъединицы ФГД. Tobey и Grant, J. Biol. Chem. 261, 12179-12183 (1980).
В прокариотах (в частности, бактериях) и микроорганизмах, например, дрожжах, но не в высших эукариотах активность ФГД дикого типа ингибируется содержащимся в клетках серином. Изучена кинетика подобного ингибирования, которое, как сообщается, протекает аллостерическим путем. Tobey и Grant, J. Biol. Chem. 261, 12179-12183 (1986); Dubrow и Pizer, J. Biol. Chem. 252, 1527-1551 (1977); Mc Kitrick и Pizer, J. Bacteriol 141, 235-245 (1980).
Tosa и Pizer (J. Bacteriol, 106, 972-982 (1971)) исследовали действие обычно токсичного аналога серина (L-серин-гидроксамата) на штамм E.coli. Отбор на среде для выращивания, содержащей этот аналог, привел к получению устойчивых к серину мутантов. Некоторые мутанты, как показано, обладают модификацией в ферменте, не имеющем отношения к ФГД, а именно серил-тРНК-синтетазе. Сырой экстракт одного из мутантов показал ФГД активность со сниженной чувствительностью к серину (см. J. Bacteriol, 106, 972-982 (1971), фиг.5, таблица 6, а также стр. 973, нижний левый столбец, стр. 977, нижний левый столбец).
II. Краткое изложение сущности изобретения.
Одним из своих аспектов изобретение направлено на ДНК, кодирующую 3-фосфоглицератдегидрогеназу (ФГД) с пониженной чувствительностью к ингибированию серином по сравнению с ФГД дикого типа, т.е. на ДНК, кодирующую ФГД, обладающую хоть какой-то полезной для биосинтеза ферментативной активностью и сохраняющей эту активность при более высоком содержании серина в отличие от (немодифицированной) ФГД дикого типа.
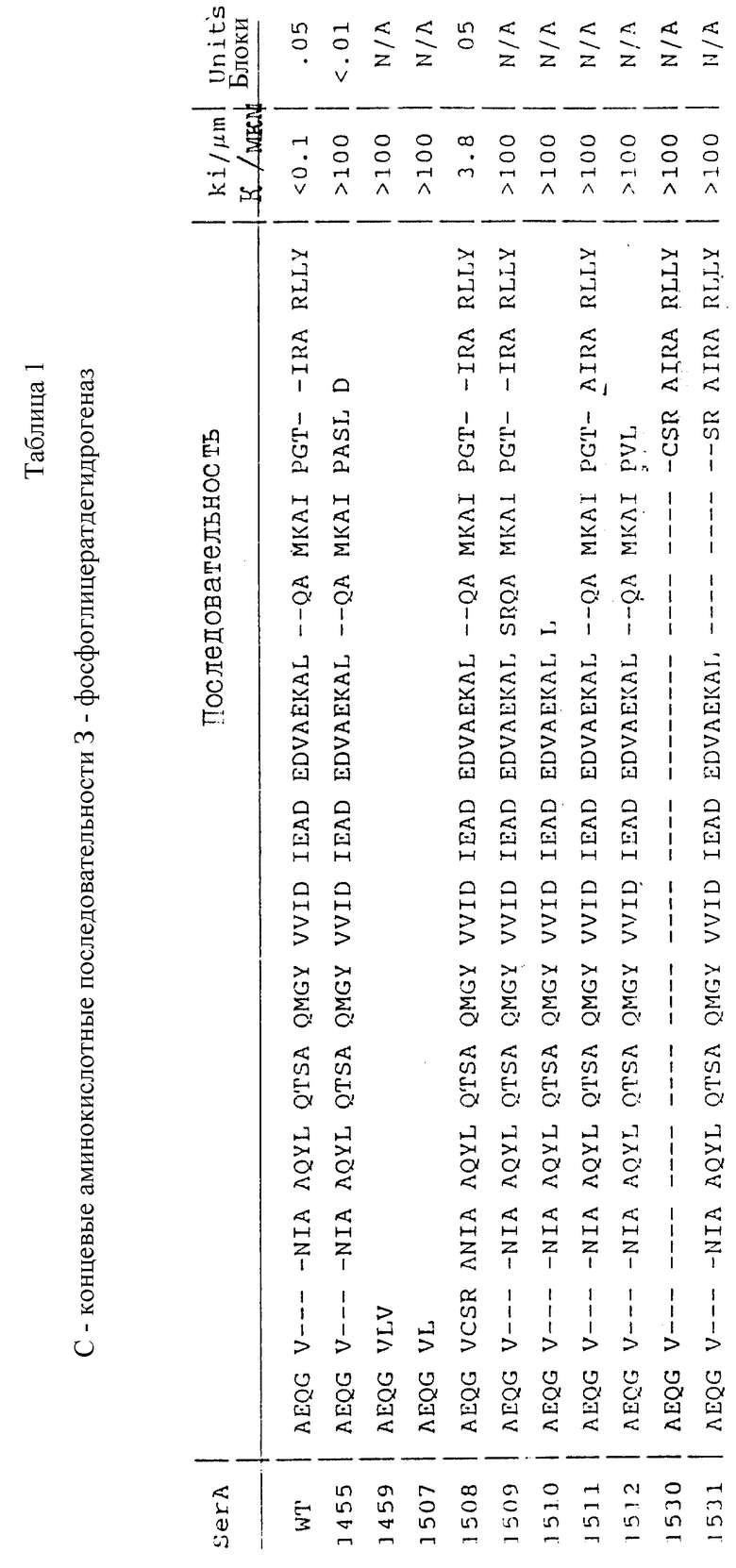
В рекомендуемых воплощениях изобретения ФГД дикого типа представлена микробной ФГД или дрожжевой ФГД. Рекомендуется также созданная генной инженерией ДНК, кодирующая ФГД с изменениями в 25% C-конца ФГД дикого типа, предпочтительно в 52 аминокислотах C-конца. К примеру, созданная генной инженерией ДНК способна кодировать ФГД с делецией части или всего C-конца. Рекомендуется также, чтобы созданная генной инженерией ДНК кодировала ФГД, имеющую вставку (например, между VAL363 и ASN364 или между ALA392 и CLN394) помимо вышеуказанной делеции или в виде независимого изменения.
Изобретение также направлено на: a) ФГД с аминокислотной последовательностью вышеуказанной созданной генной инженерией ДНК; b) экспрессирующие вектора, состоящие из созданной генной инженерией ДНК и регуляторной ДНК, расположенной и ориентированной для экспрессии рекомбинантной ДНК в хозяйской системе экспрессии; с) клетки, содержащие такие экспрессирующие векторы, и d) способы получения серина или происходящих из серина соединений культивированием подобных клеток. Что касается вышеприведенного пункта c), то рекомендуются клетки с делецией SerA дикого типа.
И еще в одном своем воплощении изобретение направлено на клеточную инженерию (например, включает рекомбинантные генетические конструкции) с целью получения ФГД-кодирующего мРНК транскрипта с измененным 3'-концом, причем такой транскрипт транслируется клеткой с образованием ФГД, обладающей пониженной чувствительностью к ингибированию серином по сравнению с ФГД дикого типа.
Изобретением обеспечивается разрегулирование важной для биосинтеза точки регулирования с усилением тем самым образования многочисленных соединений в нисходящем направлении от этой точки, в том числе, в частности, серина и происходящих из серина соединений, например, триптофана. Другие происходящие из серина клеточные метаболиты (т.е., для которых в их биосинтезе серин является первичным промежуточным соединением) включают: холин, глицин, цистеин и зависимые от C1-источника соединения, например, метионин, монофосфат инозина, пурины и некоторые пиримидины (напр.: тимидин и гидроксиметилцитозин).
III. Описание рекомендуемых воплощений изобретения.
A. Фиг. 1 - 3.
Фиг. 1 отражает стадии биосинтеза L-серина из глюкозы.
На фиг. 2 приведены последовательность E.coli SerA гена согласно Tobey и Grant (см. выше) и аминокислотная последовательность, выведенная на основе гена.
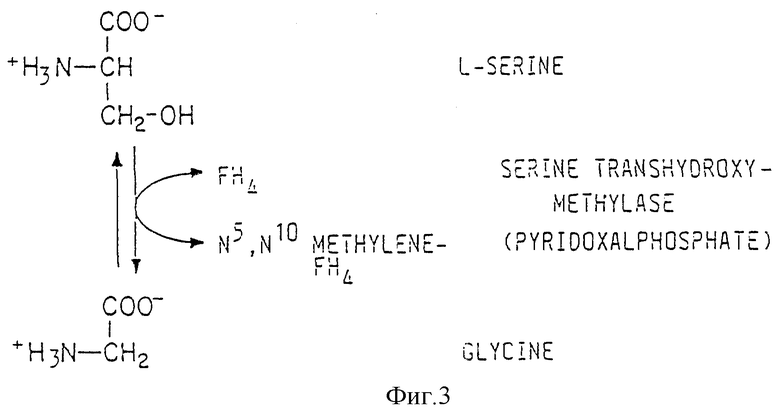
Фиг. 3 отражает биопревращение L-серина и тетрагидрофолата в глицин и N5,N10-метилентетрагидрофолат.
B. Создание не чувствительной к серину ФГД.
1. Создание генной инженерией конструкции.
Рекомендуемые воплощения изобретения направлены на биосинтез серина и родственных серину соединений, например, вышеперечисленных соединений, происходящих из серина в результате биосинтеза. Первая стадия в биосинтезе таких соединений, как показано ниже, состоит в создании не чувствительной к серину ФГД.
Нами обнаружено, что в ФГД существует специфичный связанный с серином обратной связью домен, и что такой домен может быть изменен с понижением чувствительности к серину, но с сохранением уровня полезной активности ФГД. На фиг. 2 приведена одна из конкретных ФГД генетических и аминокислотных последовательностей, которая может быть использована в качестве ссылки при последующем обсуждении. Последовательность на фиг. 2 включает 410 аминокислот (включая начальный Mel, отщепленный в зрелом белке). Домен ФГД, способный снизить чувствительность к серину без нарушения ФГД активности, находится в пределах 25% C-конца молекулы, наиболее предпочтительно в пределах 50 C-концевых остатков.
Примеры модификаций ФГД, охватываемых объемом изобретения, включают делеции некоторых или всех 42 C-концевых аминокислот, а также вставки или замены в пределах этой области, понижающие чувствительность к серину при сохранении полезного действия ФГД. К примеру, вставка аминокислотных остатков между Val 363 и Asn 364 приводит к повышению значения Ki для ФГД по сравнению с диким типом при сохранении активности ФГД.
Более заметное повышение значения Ki происходит при делеции некоторых или всех C-концевых аминокислотных остатков. Например, делеция C-концевых остатков GTIRARLLY и замена на ASLD приводит к возрастанию величины Ki на несколько порядков с одновременным сохранением уровня полезной активности ФГД. Примеры других вставок в объеме изобретения включают вставки между Ala392 и Gln394.
Другие полезные модификации включают делеции из C-конца помимо вставок и модификаций, приведенных выше.
Гены, кодирующие вышеохарактеризованную не чувствительную к серину ФГД, могут быть сконструированы методами генной инженерии, включающими изменение 3'-конца кодирующей области, которая кодирует C-концевые аминокислоты, с последующим трансформированием штамма-хозяина посредником с целью экспрессии измененного ФГД фермента.
Нижеприведенными методами осуществлен поиск ферментов с измененными сродством к серину (Ki) и ФГД активностью (см. ниже).
2. Поиск созданных генной инженерией конструкций.
В поиске генетических конструкций, полученных вышеприведенными способами, использованы следующие методы анализа ФГД активности и чувствительности к серину.
Хотя и не имеющей для изобретения особого значения, анализ ФГД активности тем не менее необходим для установления степени чувствительности к серину измененного фермента. Как хорошо известно специалистам, ферментативная активность является функцией общего числа молекул фермента и каталитической активности каждой молекулы. Так, при сравнении каталитической активности связанных обратной связью вариантов ФГД необходимо предпринять меры для адекватного регулирования относительного числа молекул ФГД в образцах, для которых сравнивается относительная каталитическая активность. Существует целый ряд подходов, позволяющих это осуществить. Однако, поскольку трудно определить однозначно уровень генной экспрессии в клетках, трансформированных усеченными SerA генами (вследствие их пониженной выживаемости), наиболее приемлемый подход к сравнению ФГД активности, создаваемой различными конструктами и диким типом, состоит в интеграции на хромосомном уровне измененного SerA гена, содержащего стандартные регуляторные элементы, в единственную копию с последующим сбором трансформантов и определением относительной каталитической активности в сравнении с ФГД из клеток дикого типа.
Может быть использован любой метод, пригодный для определения ФГД активности. ФГД активность может быть определена измерением как прямой, так и обратной реакции по методике Mc Kitrick Iohn C. и Lewis I. Pizer, J. Bacteriol 141, 235-245 (1980).
Ферментативный анализ, приведенный выше, пригоден для определения чувствительности к серину любого ФГД фермента, включая ферменты с химически модифицированным C-концом. Анализ проводят в присутствии различных концентраций серина. Каталитическую активность в присутствии серина сравнивают с каталитической активностью в отсутствие серина, после чего рассчитывают величину Ki.
В большинстве случаев рекомендуется понижать чувствительность к серину без значительного изменения ФГД каталитической активности. Могут быть использованы, конструкции с C-концевыми аминокислотными последовательностями 3-фосфоглицератдигидрогеназ, перечисленные в таблице 1.
Другие конструкции с модифицированными 3'-концами также охватываются объемом настоящего изобретения, поскольку не составляет труда получить испытуемую конструкцию и трансформировать клетку способом изобретения и провести анализ ингибирования серином ФГД активности.
Любой вектор, применение которого ведет к экспрессии ФГД белка с отсутствием чувствительности к ингибированию серином, приемлем для настоящего изобретения. В целом, однако, при отсутствии чувствительности к серину следует избегать высокого уровня экспрессии не обладающего обратной связью ФГД, поскольку образующиеся большие цитоплазмические количества серина или происходящих из серина метаболитов могут оказаться токсичными для клетки. Таким образом, любая конструкция, кодирующая ингибированный по типу обратной связи ФГД с нормальной каталитической активностью и уровнем экспрессии, аналогичным уровню нативного гена, в результате трансформации скорее всего покажет высокий уровень экспрессии ФГД с понижением клеточной выживаемости. Токсичность большого количества образующегося серина может на деле привести к появлению мутантов с пониженной экспрессией ФГД. Таким образом, хотя в некоторых вариантах трансформация с применением многокопийных плазмид может оказаться полезной в первоначальном поиске, тем не менее рекомендуется интегрировать SerA конструкты в единственных копиях в геном на хромосомном уровне. Кроме того, хромосомная интеграция, как показано ниже, облегчает определение активности делетированной по типу обратной связи ФГД. Таким образом, в большинстве вариантов, когда ожидается или желательна высокая каталитическая активность, рекомендуется применять векторы, пригодные для однокопийной хромосомной интеграции. Специалистам известны многие такие векторы и стратегия их применения. См. например, Backman, заявка на патент США N 07/091837, поданная 1 сентября 1987 г., которая вводится здесь в качестве ссылки. Могут быть получены векторы и конструкты, позволяющие успешно трансформировать и экспрессировать фермент в соответствующем хозяине с целью получения целевого продукта. Способы осуществления этого хорошо известны специалистам и в настоящем изобретении решающей роли не играют. Помимо измененной ФГД кодирующей ДНК экспрессионный вектор должен содержать различные другие элементы, охарактеризованные ниже.
Во-первых, присутствующие на векторе кодирующие последовательности должны сопровождаться соответствующими регуляторными элементами, необходимыми для поддержания соответствующего уровня экспрессии кодирующих последовательностей, в том числе промоторы, места связывания рибосом и терминирующие последовательности. В большинстве случаев нативные SerA регуляторные последовательности будут рекомендуемым источником каталитически активной части молекулы, хотя общепризнанно, что могут быть использованы многие другие регуляторные последовательности, как известные, так и те, которые еще предстоит открыть.
Во-вторых, рекомендуется, чтобы на векторе присутствовали также последовательности, кодирующие селективные маркеры, и/или гены-репортеры вместе с соответствующими регуляторными элементами. Экспрессия таких селективных маркеров оказывается полезной при идентификации трансформантов. Соответствующие селективные маркерные гены включают гены, кодирующие ампициллин, тетрациклин и хлорамфеникол.
В-третьих, желательность точки начала репликации на плазмидном векторе по большей степени зависит от желательности поддержания генов в хромосомах или вне хромосом. Для специалистов очевидна та стратегия, за счет которой отсутствие источника репликации может быть использовано для промотирования интеграции в хромосомы. См., например, патент США 4743546 (Backman и др.), который вводится здесь в качестве ссылки.
После того, как конструкция экспрессионного вектора завершена, подходящая клетка может быть трансформирована вектором, содержащим транскрипционный элемент, кодирующий не чувствительный к серину ФГД белок. В большинстве случаев полезно использовать клетки, для которых известно ингибирование эндогенного ФГД белка серином и в которых эндогенный SerA ген удален и заменен измененным геном изобретения. Подобные клеточные системы применимы для сверхпродуцирования родственных серину метаболитов. К клеткам, заведомо содержащим чувствительные к серину белки, относятся прокариоты и дрожжи.
Нижеследующие примеры иллюстрируют, но не ограничивают изобретение.
Пример 1.
Конструирование SerA генных аллелей, кодирующих устойчивые по типу обратной связи 3-фосфоглицератдегидрогеназы.
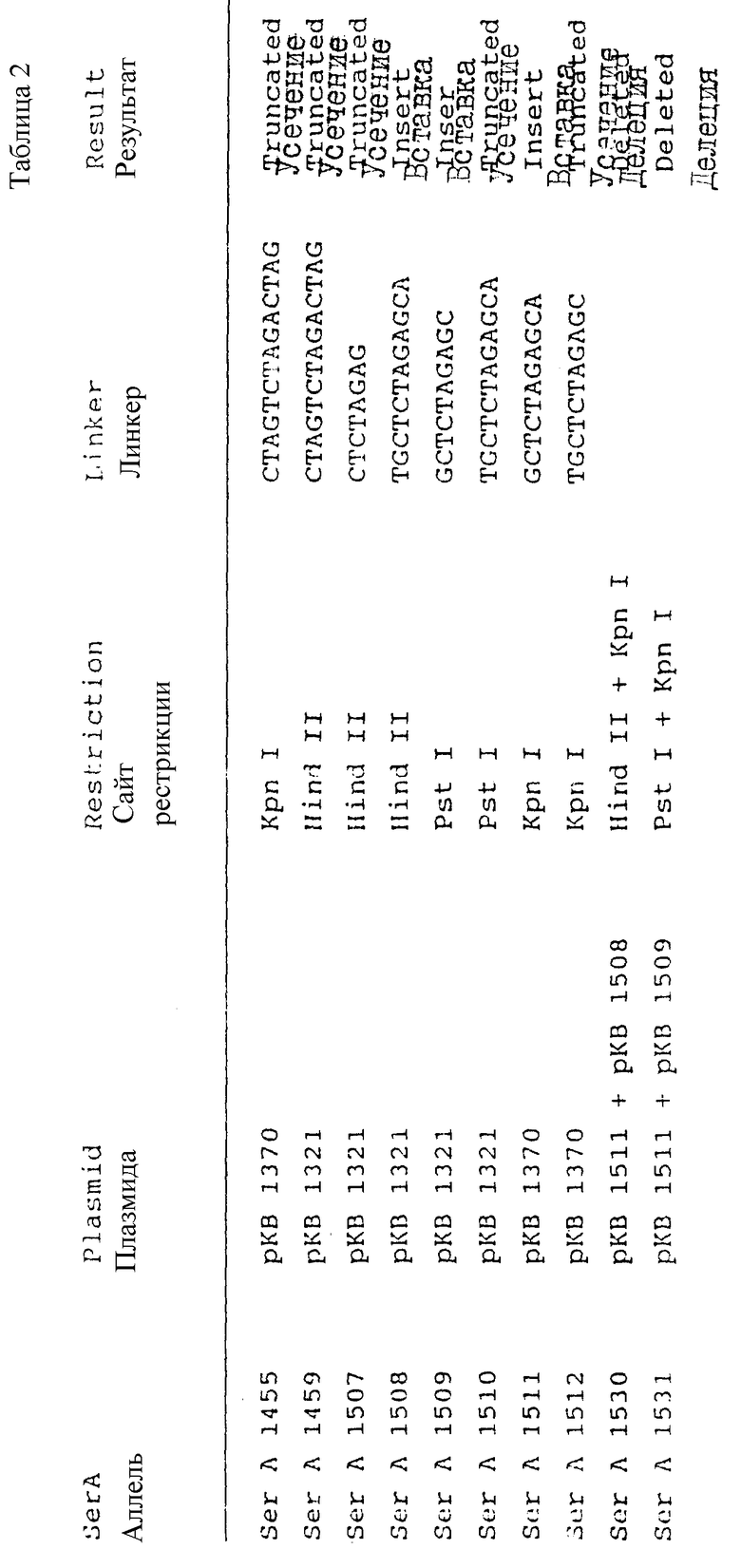
E.coli K12 SerA выделен в виде фрагмента в 6,4 т.п.о. из продукта частичного гидролиза в присутствии Sau3A, клонированного в BcII сайт плазмиды pTR264. См. Backmann и др. , заявка на патент США N 07/285128, подана 16.12.88 и Roberts и др. , Gene 12, 123 (1980). Данная плазмида названа pKB1302. Sal I-Sph I фрагмент в 3 п.т.о. из pKB1302 ДНК, содержащий SerA ген, клонируют в pUC19 с образованием pKB1321. pKB1370 образуют клонированием Hind III - Sal I фрагмента в 3 т.п.о., содержащего SerA ген, в pBR322.
Аллели SerA, кодирующие устойчивые по типу обратной связи 3-фосфоглицератдегидрогеназы, образуют вставкой Xba I линкеров в сайты рестрикции в 3'-области SerA гена. Частичный гидролиз плазмиды pKB1321 в присутствии Hid II приводит к образованию тупых концов в положении 1793, где вставка линкеров дает: a) pKB1507, кодирующую усеченную 3-фосфоглицератдегидрогеназу; b) pKB1507, кодирующую усеченную 3-фосфоглицератдегидрогеназу, и c) pKB1508, которая кодирует 3-фосфоглицератдегидрогеназу со вставкой четырех аминокислотных остатков.
Гидролиз pKB1321 в присутствии Pst I дает 3'-концевую избыточность одной нити ДНК в положении 1888. Тупые концы образуют действием фрагмента Кленова ДНК-полимеразы 1. Линкеры лигируют с тупыми концами фрагментов с образованием плазмиды pKB1509, кодирующей 3-фосфоглицератдегидрогеназу со вставкой из двух аминокислот, и pKB1510, кодирующей усеченную 3-фосфоглицератдегидрогеназу. В продукте гидролиза pKB1370 в присутствии Kpn I образуют тупые концы действием фрагмента Кленова ДНК-полимеразы 1 и после вставки линкеров получают плазмиды, кодирующие усеченные 3-фосфоглицератдегидрогеназы (pKB1455 и pKB1512) или 3-фосфоглицератдегидрогеназу со вставкой из двух аминокислотных остатков (pKB1511). Делеционные плазмиды pKB1530 и pKB1531 образованы вставкой BamHI - XbaI фрагмента в 0,8 т.п.о. из pKB1508 или BamHI - XbaI фрагмента в 0,9 т. п.о. из pKB1509 соответственно в BamHI - XbaI фрагмент в 5,8 т.п.о. плазмиды pKB1511.
В нижеследующей таблице 2 суммированы данные для различных полученных конструкций.
Для каждой конструкции указан исходный вектор, применяемый сайт рестрикции и последовательность линкерной вставки. Величины Ki для серина приведены в таблице 1, также как и относительные каталитические активности для трех таких конструкций после хромосомной интеграции (см. ниже). N/A указывает на то, что конструкция не подвергалась хромосомной интеграции, вследствие чего уровень активности не был стандартизирован.
3. Химическая модификация.
Для специалиста понятно, что делеции или модификации C-конца в ФГД дикого типа могут быть осуществлены ферментативно или химически, например, действием различных карбоксипептидаз, в том числе карбоксипептидазой Y или йодированием и присутствии лактопероксидазы.
4. Применение антисенсибилизированной мРНК.
С другой стороны имеется возможность снизить чувствительность к серину in vivo созданием ФГД-закодированных транскриптов, усеченных в 3'-конце, путем образования антисенсибилизированной мРНК, включающей нуклеотидные последовательности, комплементарные к частям кодирующих 3'-областей нативных или трансформированных ФГД-кодирующих последовательностей.
C. Получение целевых соединений.
Как показано на фиг. 3, cepин является промежуточным соединением при получении глицина. Серин также является промежуточным соединением при получении N5,N10- метилентетрагидрофолата - общего C1-источника, необходимого для синтеза метионина, пуринов (включая инозин) и некоторых пиримидинов. Таким образом, сверхпродуцирование серина из фосфоглицерата может быть использовано в широком спектре бактериальных систем продуцирования, в том числе систем продуцирования холина, глицина, цистеина, метионина, триптофана и всех пуринов, включая монофосфат инозина.
Нижеследующие характерные примеры иллюстрируют изобретение.
Пример 2.
Получение штамма-хозяина.
Внутренние относительно рожденного в плазмиде SerA гена последовательности заменяют устойчивым к канамицину геном. Полученную плазмиду затем применяют для дезактивации SerA гена в штамме-хозяине с помощью обмена аллелей по следующей методике.
SerA область УМС9 (ATCC 33920) клонируют из хромосомной ДНК, частично гидролизованной в присутствии Sau 3AI, комплементацией PC1523 (arg 161, arg F58, Ser A27, pur A54, thr -25, ton A49, re IAI, spo TI), полученной от Coli генетического Центра хранения, Иельский Университет, Нью-Хавен, коннектикут, фрагмент в 3 т.п.о., несущий SerA ген, субклонируют в pUC19 с образованием плазмиды, названной pKB1321. Из этой плазмиды фрагмент Sa II - Hind III в 3 т. п.о. вновь клонируют в pBR322 с образованием плазмиды pKB1370. Kpn I сайт у 3'-конца SerA гена превращают в BamHI использованием линкера и BamHI фрагмент, внутренний относительно образованного SerA гена, заменяют BamHI фрагментом из pUC-4-K AC (фармация), содержащим Tn903 ген устойчивости к канамицину. Эту новую плазмиду обозначают, как pKB1429. Создают производное pBR322, названное pKB701 (ATCC 39772, см. заявку на патент США N 06/757019, которая вводится здесь в качестве ссылки), в котором MboI и TThIII I, фланкирующие источник репликации, превращают в KpnI сайты. SalI - EcoRI фрагмент, содержащий SerA::KanR из pKB1429, клонируют в pKB701 с образованием pKB1438. Плазмиду pKB1438 гидролизуют в присутствии KpnI с удалением ori области. Больший фрагмент, содержащий область, кодирующую устойчивость к ампициллину, а также SerA::KanR, замыкают в цикл и используют в трансформации УМС9 в присутствии CaCl2. После трансформации хозяйские УМС9 клетки подвергают отбору на ампициллин. В условиях отбора устойчивые к ампициллину клоны развиваются за счет введения кольцевой ДНК путем гомологичной рекомбинации во фланкирующих областях SerA гена. Рост устойчивого к ампициллину изолята при отсутствии ампициллиновой селекции приводит к потере гена устойчивости к ампициллину за счет гомологичной рекомбинации повторяющихся последовательностей SerA гена. Такие штаммы идентифицируют по потере способности продуцировать β-лактамазу использованием АмпСкрин (БРЛ) в соответствии с инструкциями изготовителя. Двукратной штриховой разводкой единственных колоний на среде в присутствии или отсутствие серина выявляют чувствительные к ампициллину клоны, требующие для своего роста на минимальной среде присутствия серина и обладающие также устойчивостью к канамицину. Один из таких изолятов назван KB875.
Пример 3.
Хромосомная интеграция измененных SerA последовательностей обменом аллелей.
Аллель SerA1455 вводят в хромосому способом, аналогичным способу введения SerA::KanR, приведенному в примере 2. Вкратце способ состоит в следующем. Фрагмент (SaII - Hind III), несущий SerA1455 аллель, клонируют в pKB 701. Плазмидное начало удаляют гидролизом в присутствии KpnI. Для трансформации с приданием устойчивости к ампициллину и образованием означенного штамма используют циклическую ДНК. После неселективного выращивания использованием Ампскрина и реплики для канамицина выделяют KB904 (SerA1455), который, как показано, является чувствительным к ампициллину и канамицину KB904. Полученные SerA1544 аллели могут быть перенесены в продуцирующие штаммы PI трансдукцией. Miller (1972) "Экспериментальная мол. генетика". Колд Спринг Харбор Пресс, стр. 201-205.
Пример 4.
Хромосомная интеграция измененных SerA последовательностей recD-зависимой заменой генов.
Для введения в хромосому secA1508 аллелей использован иной подход. Штамму KB875 придан recD характер PI трансдукцией из V220 (recD,arg : Tn10, Amundsen и др., Proc. Acad. Sci. США, 82, 5558-5562 (1986))(DSM 6823). Используют ген для существенной третьей субъединицы экзонуклеазы Y для получения JGP101. Плазмиду pKB1506 переводят в линейное состояние и используют для трансформации JGP101 в сериновый прототроф по методике Shevell и др., J. Bacteriol. 170, 3294-3296 (1988) с образованием IC P103. Аллель serA1508 может быть затем введен в продуцирующие штаммы PI трансдукцией. Miller и др. "Экспериментальная мол. генетика". Колд Спринг Харбор Лаб., стр. 201-205 (1972).
Сбор сверхпродуцированных метаболитов.
Для сверхпродуцирования родственных серину метаболитов могут быть созданы клетки, продуцирующие ФГД с пониженной чувствительностью к серину, и выращены в ферменторе в соответствующих условиях, в большинстве случаев на неподвижной фазе. Клетки затем могут быть собраны, лизированы и целевые метаболиты могут быть выделены использованием стандартных биохимических методик. Условия, принципы и ссылки относительно выращивания микробов и сбора специфичных метаболитов можно найти у Gruege и Cruege (1982) (Биотехнология. Справочник промышленной микробиологии), а также Herrman и Somerville (1983) (Аминокислоты. Биосинтез и генетическое регулирование), которые вводятся здесь в качестве ссылок.




Изобретение направлено на ДНК, кодирующую 3-фосфоглицератдегидрогеназу (ФГД) с пониженной чувствительностью к ингибированию серином по сравнению с ФГД дикого типа. ФГД дикого типа представлена микробной или дрожжевой ФГД. Методом генной инженерии также создана ДНК, кодирующая ФГД с изменениями в 25% С-конца ФГД дикого типа, предпочтительно в 52 аминокислотах С-конца. Изобретение обеспечивает получение в чистом виде серина и серин содержащих соединений. 3 с.п. ф-лы, 3 ил., 2 табл.
| EP 91310709 А, 20.11.91. |
