ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к рекомбинантному микроорганизму для продукции метионина и к способу продукции метионина путем культивирования рекомбинантного микроорганизма в подходящей культуральной среде, содержащей источник углерода и источник серы. Данный микроорганизм модифицирован таким образом, что возрастает выход из источника метионина/углерода путем ослабления активности кобаламин-независимой метионинсинтазы. В частности, в данном рекомбинантном микроорганизме подвергается делеции ген metE.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Серосодержащие соединения, такие как цистеин, гомоцистеин, метионин или S-аденозилметионин являются критически важными для клеточного метаболизма и производятся промышленным путем для применения в качестве пищевых или кормовых добавок и фармацевтических средств. В частности, метионин, незаменимая аминокислота, которая не может синтезироваться животными, играет важную роль во многих функциях организма. Помимо его роли в биосинтезе белка, метионин участвует в трансметилировании и в регуляции биодоступности селена и цинка. Метионин также непосредственно применяется в качестве средства для лечения расстройств, подобных аллергии и ревматическому полиартриту. Тем не менее, большую часть произведенного метионина добавляют в корм животных.
С уменьшением применения белков животного происхождения в результате BSE (губкообразная энцефалопатия крупного рогатого скота) и птичьего гриппа возросла потребность в чистом метионине. Обычно D, L-метионин производится химически из акролеина, метилмеркаптана и гидроцианида. Однако данная рацемическая смесь не работает так же хорошо, как чистый L-метионин (Saunderson, 1985). Кроме того, хотя чистый L-метионин и может быть получен из рацемического метионина, например, посредством обработки N-ацетил-D,L-метионина ацилазой, это кардинально увеличивает себестоимость. Соответственно, возрастающая потребность в чистом L-метионине в сочетании с заботой об окружающей среде делают микробную продукцию метионина привлекательной перспективой.
Оптимизация получения химического соединения из микроорганизма типично включает сверхэкспрессию белков, участвующих в пути биосинтеза, ослабление активности белков, участвующих в репрессии пути биосинтеза, или ослабление активности белков, участвующих в продукции нежелательных побочных продуктов. Все эти подходы для оптимизации получения L-метионина в микроорганизмах были описаны ранее (см., например, патенты или патентные заявки US 7790424, US 7611873, WO 2002/010209, WO 2005/059093 и WO 2006/008097); однако, для промышленного производства L-метионина из микроорганизмов требуются дальнейшие улучшения.
В Escherichia coli и в других микроорганизмах, подобных Corynebacterium glutamicum, конечную стадию в биосинтезе метионина de novo катализируют два отличных фермента (Foster et al., 1961; Gonzalez et al., 1992). Кобаламин-зависимая метионинсинтаза (MetH, EC 2.1.1.13) кодируется геном metH и содержит простетическую группу, которая требуется для активности. Кобаламин-независимая метионинсинтаза (MetE, ЕС 2.1.1.14) кодируется геном metE и не имеет известной потребности в простетической группе, происходящей из витамина.
Многочисленные патентные заявки относятся к сверхпродукции ферментов MetH и MetE для усиления последней стадии биосинтеза метионина, как, например:
- в WO 2007/012078 и WO 2007/135188 от BASF описаны генетические изменения, приводящие к сверхэкспрессии генов metE и/или metH.
- в WO 2009/144270 от EVONIK описан способ продукции метионина с использованием микроорганизма, который демонстрирует повышенное количество и/или активность системы реактивации коб(I)аламин-зависимой MetH.
Удивительным образом и неожиданно авторы изобретения обнаружили, что уменьшение количества и/или активности кобаламин-независимой метионинсинтазы (MetE) приводит к улучшенной продукции метионина. Впервые предложено, что потеря активности одного из ферментов, принадлежащих к пути биосинтеза метионина, является полезной для продукции метионина.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Изобретение относится к рекомбинантному микроорганизму, оптимизированному для продукции метионина, в котором ослаблена активность кобаламин-независимой метионинсинтазы MetE. Предпочтительно ген metE, кодирующий фермент MetE, подвергнут делеции или мутации. Данный рекомбинантный микроорганизм также может содержать другие генетические модификации, такие как:
- повышенная экспрессия по меньшей мере одного из следующих генов: ptsG, pyc, pntAB, cysP, cysU, cysW, cysA, cysM, cysJ, cysI, cysH, gcvT, gcvH, gcvP, Ipd, serA, serB, serC, cysE, metF, metH, thrA, аллель metA (metA*), кодирующий фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи, thrA или аллель thrA (thrA*), кодирующий фермент с пониженным ингибированием треонином по механизму обратной связи и/или
- ослабленная экспрессия одного из следующих генов: metJ, pykA, pykF, purU, ybdL или yncA.
В конкретном воплощении настоящее изобретение относится к микроорганизму, в котором: а) ген metE подвергнут делеции, б) усилена экспрессия генов metA*, metH, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE, thrA* и pyc и в) ослаблена экспрессия генов metJ, pykA, pykF, purU и yncA.
Данное изобретение также относится к способу получения метионина или производных метионина в ферментативном процессе, включающем следующие стадии: а) культивирование рекомбинантного микроорганизма согласно изобретению в подходящей культуральной среде, включающей ферментируемый источник углерода, содержащий глюкозу, и источник серы, и б) выделение метионина или производных метионина из культуральной среды.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Перед подробным описанием настоящего изобретения следует понимать, что данное изобретение не ограничивается конкретными приведенными в качестве примеров способами и, естественно, может изменяться. Также следует понимать, что использованная здесь терминология предназначена лишь для цели описания конкретных воплощений изобретения и не подразумевает того, что она является ограничивающей, и что она будет ограничиваться лишь приложенной формулой изобретения.
Все публикации, патенты и патентные заявки, процитированные здесь выше или ниже, являются тем самым включенными посредством ссылки во всей их полноте. Однако упомянутые здесь публикации процитированы с целью описания и раскрытия протоколов, реактивов и векторов, которые приведены в данных публикациях, и которые возможно могут использоваться в связи с изобретением. Ничто здесь не следует истолковывать как допущение того, что изобретение не дает право противопоставлять такое раскрытие с предыдущим изобретением.
Кроме того, в воплощении настоящего изобретения на практике, если не указано иное, используются традиционные методики микробиологии и молекулярной биологии в пределах квалификации в данной области. Такие методики хорошо известны специалисту и полностью объяснены в литературе. См., например, Prescott et al. (1999) и Sambrook et al. (1989) (2001).
Следует отметить, что формы в единственном числе, в том виде, в котором они используются здесь и в приложенной формуле изобретения, включают ссылку на множественное число, если контекст явно не диктует иное. Таким образом, например, ссылка на «микроорганизм» включает множество таких микроорганизмов, и ссылка на «эндогенный ген» представляет собой ссылку на один или более чем один эндогенный ген и так далее. Если не определено иначе, все использованные здесь технические и научные термины имеют такое значение, которое обычно понятно обычному специалисту в области, к которой принадлежит изобретение. Несмотря на то, что в воплощении на практике или тестировании настоящего изобретения можно использовать любые материалы и методы, аналогичные или эквивалентные описанным здесь материалам и методам, сейчас описаны предпочтительные материалы и методы.
В формуле изобретения, которая следует далее, и в последующем описании изобретения, за исключением тех случаев, когда контекст требует иного из-за прямой формулировки или необходимого подтекста, слово «содержать» или «включать» или вариации, такие как «содержит», «содержащий», «включающий», «включенный», «включает», используются во включительном смысле, т.е. для определения присутствия утверждаемых характеристик, но не для исключения присутствия или добавления дополнительных характеристик в разных воплощениях данного изобретения.
Определения
Термин «метионин» обозначает незаменимую серосодержащую аминокислоту с химической формулой HO2CCH(NH2)CH2CH2SCH3 и номером CAS 59-51-8 или 63-68-3 для специфического L-изомера.
Фраза «производные метионина» относится к молекулам-аналогам метионина, которые имеют такой же химический остов, но отличаются от метионина по меньшей мере одной химической группой. В данном изобретении предпочтительными производными метионина являются N-ацетилметионин (NAM), S-аденозилметионин (SAM) и гидрокси-метионин.
Термин «микроорганизм» в том виде, как он здесь используется, относится к бактерии, дрожжам или грибку, которые не являются искусственно модифицированными. Предпочтительно микроорганизм выбран из Enterobacteriaceae, Bacillaceae, Streptomycetaceae и Corynebacteriaceae. Более предпочтительно микроорганизм представляет собой вид Escherichia, Klebsiella, Pantoea, Salmonella или Corynebacterium. Даже более предпочтительно микроорганизм представляет собой либо вид Escherichia coli, либо Corynebacterium glutamicum.
Термин «рекомбинантный микроорганизм» или «генетически модифицированный микроорганизм» в том виде, как он здесь используется, относится к бактерии, дрожжам или грибку, который отсутствует в природе и является генетически отличным от его эквивалента, обнаруженного в природе. Это означает, что он модифицирован либо путем введения, либо делеции, либо модификации генетических элементов. Он также может быть трансформирован путем принуждения к развитию и эволюции новых метаболических путей путем объединения прямого мутагенеза и эволюции под специфичным давлением отбора (см., например, WO 2004/076659).
Микроорганизм может быть модифицирован для экспрессии экзогенных генов, если данные гены вводятся в данный микроорганизм со всеми элементами, обеспечивающими их экспрессию в микроорганизме-хозяине. Модификация или «трансформация» микроорганизмов экзогенной ДНК представляет собой рутинную задачу для специалистов в данной области.
Микроорганизм может быть модифицирован для модуляции уровня экспрессии эндогенного гена.
Термин «эндогенный ген» означает то, что ген присутствовал в микроорганизме до какой-либо генетической модификации в штамме дикого типа. Эндогенные гены могут подвергаться сверхэкспрессии путем введения гетерологичных последовательностей помимо или для того, чтобы замещать эндогенные регуляторные элементы, или путем введения одной или более чем одной дополнительной копии гена в хромосому или плазмиду. Эндогенные гены также могут быть модифицированы для модуляции их экспрессии и/или активности. Например, в кодирующую последовательность могут быть введены мутации для модификации генного продукта, или могут быть введены гетерологичные последовательности помимо или для замещения эндогенных регуляторных элементов. Модуляция эндогенного гена может приводить к повышающей регуляции и/или усилению активности генного продукта, или, в качестве альтернативы, к понижающей регуляции и/или уменьшению активности эндогенного генного продукта.
Другим способом модуляции их экспрессии является замена эндогенного промотора гена (например, промотора дикого типа) на более сильный или более слабый промотор для повышающей или понижающей регуляции экспрессии эндогенного гена. Данные промоторы могут быть гомологичными или гетерологичными. Выбор подходящих промоторов находится целиком в пределах способностей специалиста в данной области.
Термин «экзогенный ген» означает то, что ген был введен в микроорганизм способами, хорошо известными специалисту в данной области, в то время как данный ген не встречается в природе в данном микроорганизме. Экзогенные гены могут быть интегрированы в хромосому хозяина, или экспрессироваться внехромосомно плазмидами или векторами. В данной области хорошо известен целый ряд плазмид, которые отличаются в отношении их репликатора и числа их копий в клетке. Данные гены могут быть гетерологичными или гомологичными.
Термин «гетерологичный ген» означает то, что ген происходит из вида микроорганизма, отличного от микроорганизма-реципиента, который его экспрессирует. Он относится к гену, который не встречается в данном микроорганизме в природе.
В настоящей заявке ссылка на все гены дается с использованием их обычных названий в Е.coli. Их нуклеотидные последовательности доступны на веб-сайтах http://www.ncbi.nlm.nih.gov/gene или http://www.ebi.ac.uk/embl/.
С использованием ссылок, приведенных в Genbank для известных генов, специалисты в данной области могут определять эквивалентные гены в других организмах, бактериальных штаммах, дрожжах, грибах, млекопитающих, растениях и т.д. Эта рутинная работа преимущественно осуществляется с использованием консенсусных последовательностей, которые могут быть определены путем проведения выравниваний последовательностей с генами, происходящими из других микроорганизмов, и разработки вырожденных зондов для клонирования соответствующего зонда в другом организме. Эти традиционные методы молекулярной биологии хорошо известны специалистам в данной области и заявлены, например, в Sambrook et al, (1989) и (2001).
Термины «улучшенная продукция метионина», «улучшать продукцию метионина» и их грамматические эквиваленты в том виде, как они здесь используются, относятся к повышенному выходу метионина/источник углерода (отношение грамм/моль метионина, продуцированного на грамм/моль потребленного источника углерода, которое может быть выражено в процентах). Способы определения количества потребленного источника углерода и продуцированного метионина хорошо известны специалистам в данной области. Выход выше у рекомбинантного микроорганизма по сравнению с соответствующим немодифицированным микроорганизмом.
Фраза «микроорганизм, оптимизированный для ферментативной продукции метионина» относится к микроорганизмам, эволюционировавшим и/или генетически модифицированным для осуществления улучшенной продукции метионина по сравнению с эндогенной продукцией соответствующих микроорганизмов дикого типа. Такие микроорганизмы, «оптимизированные» для продукции метионина, хорошо известны в данной области и были раскрыты, в частности, в патентных заявках WO 2005/111202, WO 2007/077041 и WO 2009/043803.
Согласно данному изобретению термины «ферментативная продукция», «культура» или «ферментация» используются для обозначения роста бактерий. Этот рост обычно проводится в ферментерах с подходящей культуральной средой, адаптированной для используемого микроорганизма и содержащей по меньшей мере один простой источник углерода и, при необходимости, косубстраты.
Фраза «подходящая культуральная среда» обозначает среду (например, стерильные жидкие среды), содержащую незаменимые или полезные питательные вещества для поддержания и/или роста клетки, такие как источники углерода или углеродные субстраты, источники азота, например, пептон, дрожжевые экстракты, мясные экстракты, экстракты солода, мочевина, сульфат аммония, хлорид аммония, нитрат аммония и фосфат аммония; источники фосфора, например, монокалия фосфат или дикалия фосфат; микроэлементы (например, соли металлов), например, соли магния, соли кобальта и/или соли марганца, а также факторы роста, такие как аминокислоты и витамины.
Термин «углеродный источник» или «углеродный субстрат», или «источник углерода» согласно настоящему изобретению обозначает любой источник углерода, который может использоваться специалистами в данной области для поддержки нормального роста микроорганизма, включая моносахариды (такие как глюкоза, галактоза, ксилоза, фруктоза или лактоза), олигосахариды, дисахариды (такие как сахароза, целлобиоза или мальтоза), меласса, крахмал или его производные, гемицеллюлозы и их комбинации. Особенно предпочтительным простым источником углерода является глюкоза. Другим предпочтительным простым источником углерода является сахароза. Источник углерода может происходить из возобновляемого сырья. Возобновляемое сырье определяется как сырье, требующееся для определенных промышленных процессов, которое может быть регенерировано в пределах короткого промежутка времени и в достаточном количестве для обеспечения его превращения в желательный продукт. Растительная биомасса, обработанная или необработанная, является интересным возобновляемым источником углерода.
Термин «источник серы» согласно данному изобретению относится к сульфату, тиосульфату, гидросульфиду, дитионату, дитиониту, сульфиту, метилмеркаптану, диметилсульфиду и другим сульфидам с метильной группой или к комбинации разных источников. Более предпочтительно, источник серы в культуральной среде представляет собой сульфат или тиосульфат, или их смесь.
Термин «источник азота» соответствует либо соли аммония, либо газообразному аммиаку. Источник азота поставляется в форме аммония или аммиака.
Термины «ослабление» или с «ослабленной экспрессией» в данном контексте означают то, что экспрессия гена или фермента уменьшается или подавляется по сравнению с немодифицированным микроорганизмом. Уменьшение или подавление экспрессии фермента получают путем ослабления экспрессии гена, кодирующего указанный фермент.
Ослабление экспрессии генов может достигаться средствами и способами, известными специалисту в данной области. В общем, ослабление экспрессии гена может достигаться посредством:
- мутирования кодирующей области или промоторной области, или
- удаления всей или части промоторной области, необходимой для экспрессии гена, или
- удаления кодирующей области гена путем гомологичной рекомбинации или
- вставки в кодирующую область или в промоторную область внешнего элемента или
- проведения экспрессии гена под контролем слабого промотора.
Специалист в данной области знает целый ряд промоторов, которые демонстрируют разную силу, и какой промотор использовать для слабой генетической экспрессии.
Термин «активность» фермента используется взаимозаменяемо с термином «функция» и обозначает, в контексте изобретения, реакцию, которая катализируется ферментом. Специалист в данной области знает как измерять ферментативную активность указанного фермента. В частности, относительно измерения активности белка MetE см. пример 5.
Термины «ослабленная активность» или «пониженная активность» фермента означают либо пониженную удельную каталитическую активность белка, полученную мутацией в аминокислотной последовательности, и/или пониженные концентрации белка в клетке, полученные мутацией нуклеотидной последовательности или делецией кодирующей области гена.
Термины «повышенная активность» или «увеличенная активность» фермента обозначают или увеличенную удельную каталитическую активность фермента и/или повышенное количество/доступность фермента в клетке, полученные, например, сверхэкспрессией гена, кодирующего фермент.
Термины «увеличенная экспрессия», «повышенная экспрессия» или «сверхэкспрессия» и их грамматические эквиваленты используются в тексте взаимозаменяемо и имеют аналогичное значение. Данные термины означают то, что экспрессия гена или фермента увеличивается по сравнению с немодифицированным микроорганизмом. Увеличение экспрессии фермента получают путем увеличения экспрессии гена, кодирующего указанный фермент.
Для увеличения экспрессии гена специалист в данной области знает разные методики:
- Увеличение числа копий гена в микроорганизме. Данный ген кодируется хромосомно или внехромосомно. При локализации гена на хромосоме, на хромосому можно вводить несколько копий гена способами рекомбинации, известными эксперту в данной области (включая замену гена). При внехромосомной локализации гена его можно проводить разными типами плазмид, которые отличаются в отношении их репликатора и, таким образом, их числа копий в клетке. Данные плазмиды присутствуют в микроорганизме в 1-5 копиях или примерно в 20 копиях или в вплоть до 500 копиях, в зависимости от природы плазмиды: плазмиды с низким числом копий с ограниченной репликацией (pSC101, RK2), плазмиды с низким числом копий (pACYC, pRSF1010) или плазмиды с большим числом копий (pSK bluescript II).
- С использованием промотора, индуцирующего высокий уровень экспрессии гена. Специалист в данной области знает, какие промоторы являются самыми удобными, например, широко используются промоторы Ptrc, Ptac, Plac или промотор лямбда cl. Данные промоторы могут быть «индуцибельными» посредством конкретного соединения или конкретными внешними условиями, подобными температуре или свету. Данные промоторы могут быть гомологичными или гетерологичными.
- Путем ослабления активности или экспрессии репрессора транскрипции специфического или неспецифического гена.
- С использованием элементов, стабилизирующих соответствующую матричную РНК (Carrier and Keasling, 1998), или элементов, стабилизирующих белок (например, GST (глутатион-S-трансфераза) метки, GE Healthcare).
Термин «кодирующий» относится к процессу, посредством которого полинуклеотид, посредством механизмов транскрипции и трансляции, продуцирует аминокислотную последовательность. Ген(ны), кодирующий(щие) фермент(ты) может быть экзогенным или эндогенным.
Термины «чувствительность по механизму обратной связи» или «ингибирование по механизму обратной связи» относятся к клеточному механизму контроля, в котором один или несколько ферментов, которые катализируют продукцию определенного вещества в клетке, ингибируются или становятся менее активными при накоплении данного вещества до определенного уровня. Так что термины «пониженная чувствительность по механизму обратной связи» или «ослабленное ингибирование по механизму обратной связи» означают то, что активность такого механизма уменьшается или подавляется по сравнению с немодифицированным микроорганизмом. Специалист в данной области знает как модифицировать фермент для получения данного результата. Такие модификации были описаны в патентной заявке WO 2005/111202 или в патенте US 7611873.
Данное изобретение относится к рекомбинантному микроорганизму, оптимизированному для ферментативной продукции метионина, в котором ослаблена активность кобаламин-независимой метионинсинтазы MetE.
Специалист в данной области знает много средств и способов ослабления ферментативной активности, подобных мутации белка, мутации гена или ослаблению экспрессии гена. Мутация белка может достигаться заменой специфических аминокислот, присутствующих в каталитическом сайте фермента, или введением дополнительных аминокислот, или делецией определенных аминокислот.
В первом аспекте изобретения ослабляют экспрессию гена metE, кодирующего кобаламин-независимую метионинсинтазу MetE. Нуклеотидная последовательность гена metE Е.coli показана в SEQ ID NO 20.
Ослабление экспрессии гена может достигаться введением в ген чужеродной ДНК для его инактивации или экспрессированием гена под контролем слабого промотора или индуцибельного промотора. Специалист в данной области знает широкий спектр промоторов, демонстрирующих разную силу экспрессии и/или разные параметры индукции, или как модифицировать промотор для уменьшения его силы экспрессии путем проведения модификации промотора дикого типа, например, в его консенсусной последовательности, сайте связывания рибосомы или инициирующем кодоне. Таким образом, специалист в данной области способен к выбору промотора, который приводит к ослаблению экспрессии metE.
В предпочтительном воплощении изобретения подвергают делеции по меньшей мере часть гена metE. Предпочтительно эта подвергнувшаяся делеции часть составляет по меньшей мере 10% кодирующей последовательности, более предпочтительно по меньшей мере 20%, 30%, 40% или 50% кодирующей последовательности. Более предпочтительно по меньшей мере 80% кодирующей последовательности подвергается делеции. В конкретном воплощении изобретения осуществляется делеция всего гена metE. Специалист в данной области знает много методик для удаления частей гена, таких как гомологичная рекомбинация.
Во втором аспекте изобретения ген metE подвергают мутации для того, чтобы он кодировал модифицированный белок, демонстрирующий ослабленную активность. В предпочтительном воплощении изобретения мутация гена metE приводит к трансляции усеченного белка MetE, который является неактивным. Более предпочтительно мутация представляет собой делецию части из 13 пар оснований (п.о.): от 417-го до 429-го основания гена Е.coli, нуклеотидная последовательность которого показана в SEQ ID NO 20, что приводит к мутации в виде сдвига рамки считывания. Впоследствии трансляция белка сокращается (посредством сдвига рамки считывания вводится терминирующий кодон) и приводит к усеченному белку из 152 аминокислот, как показано в SEQ ID NO 22, вместо 753 аминокислот в последовательности дикого типа, как показано в SEQ ID NO 21. Частью данного изобретения также является любая эквивалентная мутация, обеспечивающая введение терминирующего кодона в ген metE из любого вида микроорганизмов.
Оптимизация пути биосинтеза метионина
Рекомбинантный микроорганизм согласно изобретению модифицируется для улучшения продукции метионина. Гены, участвующие в продукции метионина, хорошо известны в данной области и включают гены, участвующие в специфическом пути биосинтеза метионина, а также гены, участвующие в путях, дающих предшественники, и гены, участвующие в путях, потребляющих метионин.
Для эффективной продукции метионина требуется оптимизация специфического пути биосинтеза метионина и нескольких путей, дающих предшественники. Метионин-продуцирующие штаммы уже были описаны, в частности, в патентных заявках WO 2005/111202, WO 2007/077041 и WO 2009/043803. Данные заявки включены в эту заявку в качестве ссылки.
В конкретном воплощении изобретения рекомбинантный микроорганизм модифицирован, как описано ниже: увеличена экспрессия по меньшей мере одного из следующих генов: ptsG, pyc, pntAB, cysP, cysU, cysW, cysA, cysM, cysJ, cysI, cysH, gcvT, gcvH, gcvP, Ipd, serA, serB, serC, cysE, metF, metH, metA, аллель metA, кодирующий фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи (metA*), thrA и аллель thrA, кодирующий фермент с пониженной чувствительностью к треонину по механизму обратной связи (thrA*).
- ptsG кодирует фермент PTS IICBGlc, как описано в патентной заявке ЕР 11305829,
- pyc кодирует пируваткарбоксилазу, как описано в патентной заявке ЕР 11305829. В предпочтительном воплощении ген pyc является гетерологичным и выбран из генов pyc из Rhizobium etli, Bacillus subtilis, Lactococcus lactis, Pseudomonas fluorescens или видов Corynebacterium,
- pntAB кодирует субъединицы мембраносвязанной трансгидрогеназы, такой как описанная в патентной заявке WO 2012/055798,
- cysP кодирует периплазматический сульфатсвязывающий белок, как описано в WO 2007/077041 и в WO 2009/043803,
- cysU кодирует компонент ABC транспортера сульфата, как описано в WO 2007/077041 и в WO 2009/043803,
- cysW кодирует мембраносвязанный белок, транспортирующий сульфат, как описано в WO 2007/077041 и в WO 2009/043803,
- cysA кодирует сульфатпермеазу, как описано в WO 2007/077041 и в WO 2009/043803,
- cysM кодирует О-ацетилсеринсульфгидралазу, как описано в WO 2007/077041 и в WO 2009/043803,
- cysI и cysJ кодируют, соответственно, альфа и бета субъединицы сульфитредуктазы, как описано в WO 2007/077041 и в WO 2009/043803. Предпочтительно cysI и cysJ сверхэкспрессируются совместно,
- cysH кодирует аденилилсульфатредуктазу, как описано WO 2007/077041 и в WO 2009/043803.
Усиленный С1 метаболизм также представляет собой модификацию, которая приводит к улучшенной продукции метионина. Он относится к увеличению активности по меньшей мере одного фермента, участвующего в С1 метаболизме, выбранного из GcvTHP, Lpd, MetF или MetH. В предпочтительном воплощении изобретения одноуглеродный метаболизм усиливается путем увеличения экспрессии и/или активности по меньшей мере одного из следующих:
- gcvT, gcvH, gcvP и lpd, кодирующих комплекс расщепления глицина, как описано в патентной заявке WO 2007/077041. Комплекс расщепления глицина (GCV) представляет собой мультиферментный комплекс, который катализирует окисление глицина, давая диоксид углерода, аммоний, метилен-THF (тетрагидрофолат) и восстановленный пиридиновый нуклеотид. Комплекс GCV состоит из четырех белковых компонентов: указанного Р-белка глициндегидрогеназы (GcvP), указанного Н-белка - липоил-GcvH-белка (GcvH), указанного Т-белка - аминометилтрансферазы (GcvT) и указанного L-белка - дигидролипоамиддегидрогеназы (GcvL или Lpd). Р-белок катализирует пиридоксальфосфат-зависимое высвобождение CO2 из глицина, оставляющее метиламинную группировку. Метиламинная группировка переносится на группу липоевой кислоты Н-белка, который связан с Р-белком до декарбоксилирования глицина. Т-белок катализирует высвобождение NH3 из метиламинной группы и переносит остающуюся С1 единицу на THF, образуя метилен-THF. L-белок затем окисляет компонент в виде липоевой кислоты Н-белка и переносит электроны на NAD+, образуя NADH;
- MetF, кодирующей метилентетрагидрофолатредуктазу, как описано в патентной заявке WO 2007/077041;
- MetH (В12-зависимая гомоцистеин-N5-метилтетрагидрофолаттрансметилаза), кодирующей метилтрансферазы.
Сверхэкспрессия по меньшей мере одного из следующих генов, участвующих в биосинтезе серина, также уменьшает продукцию побочного продукта - изолейцина:
- serA, который кодирует фосфоглицератдегидрогеназу, как описано в WO 2007/077041 и в WO 2009/043803,
- serB, который кодирует фосфосеринфосфатазу, как описано в WO 2007/077041 и в WO 2009/043803,
- serC, который кодирует фосфосеринаминотрансферазу, как описано в WO 2007/077041 и в WO 2009/043803.
Уже было показано, что сверхэкспрессия следующих генов улучшает продукцию метионина:
- cysE кодирует серинацилтрансферазу; его сверхэкспрессия обеспечивает усиление продукции метионина, как описано в WO 2007/077041;
- metA кодирует гомосеринсукцинилтрансферазу. Аллель MetA* кодирует фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи. Предпочтительно используется аллель MetA*, описанный в патентной заявке WO 2005/111202;
- thrA кодирует аспартокиназу/гомосериндегидрогеназу; аллель thrA* кодирует фермент с пониженным ингибированием треонином по механизму обратной связи, как описано в WO 2005/111202.
В конкретном воплощении изобретения гены могут находиться под контролем индуцибельного промотора. В предпочтительном воплощении изобретения по меньшей мере один из данных генов находится под контролем промотора, индуцируемого температурой. Предпочтительно экспрессия по меньшей мере одного из генов: thrA, cysE, metA прямо или опосредованно находится под контролем индуцибельного промотора. В предпочтительном воплощении изобретения экспрессия гена thrA находится под прямым контролем индуцибельного промотора, и экспрессия гена cysE подвергается полярному влиянию экспрессии гена thrA. В другом предпочтительном воплощении изобретения экспрессия гена thrA находится под прямым контролем индуцибельного промотора, и экспрессия генов cysE и metA подвергается полярному влиянию индуцибельной экспрессии гена thrA.
В наиболее предпочтительном воплощении промотор, индуцируемый температурой, принадлежит к семейству промоторов PR. Продуцирующий метионин штамм имеет гены, находящиеся под контролем индуцибельных промоторов, как описано в патентной заявке WO 2011/073122.
В другом конкретном воплощении изобретения микроорганизм был дополнительно модифицирован, и была ослаблена экспрессия по меньшей мере одного из следующих генов: metJ, pykA, pykF, purU, ybdL или yncA.
- Ген metJ кодирует репрессорный белок MetJ (GenBank 1790373), ответственный за понижающую регуляцию метионинового регулона, как было предложено в патентной заявке JP 2000/157267,
- Гены pykA и pykF кодируют ферменты «пируваткиназы». Ослабление экспрессии по меньшей мере одной или обеих пируваткиназ снижает потребление фосфоенолпирувата (PEP). Повышенная доступность PEP может увеличивать продукцию оксалоацетата, важного предшественника аспартата, который, в свою очередь, является предшественником метионина, как описано в WO 2007/077041 и в WO 2009/043803,
- purU кодирует формилтетрагидрофолатдеформилазу, фермент, который катализирует формил-ТНР-деформилазную реакцию. Ослабление деформилазной активности увеличивает продукцию метил-THF, который требуется для метилирования гомоцистеина. Потеря С1 метаболитов посредством деформилирования приводит к повышенной продукции гомоцистеина, который не может превращаться в метионин. Гомоцистеин затем может быть субстратом фермента цистатионин-гамма-синтазы (MetB), который может катализировать реакцию между О-сукцинилгомосерином и гомоцистеином, приводящую к образованию гомолантионина, как описано в WO 2007/077041 и в WO 2009/043803,
- ybdL кодирует аминотрансферазу, как описано в патентной заявке PCT/FR 2010/052937,
- yncA кодирует N-ацилтрансферазу, как описано в патентной заявке WO 2010/020681.
В более предпочтительном воплощении изобретения ферментативная продукция метионина рекомбинантным микроорганизмом, в котором ослаблена активность кобаламин-независимой метионинсинтазы MetE, осуществляется из глюкозы в качестве главного источника углерода и может достигаться посредством комбинации обсуждавшихся выше модификаций в указанном микроорганизме, например:
- экспрессия гена metJ ослабляется, и экспрессия аллеля metA, кодирующего фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи (MetA*), усиливается;
- экспрессия гена metJ ослабляется; экспрессия аллеля metA, кодирующего фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи (MetA*), усиливается; и экспрессия аллеля thrA, кодирующего фермент с пониженным ингибированием треонином по механизму обратной связи (thrA*), усиливается;
- экспрессия гена metJ ослабляется; экспрессия аллеля metA, кодирующего фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи (MetA*), усиливается; экспрессия аллеля thrA, кодирующего фермент с пониженным ингибированием треонином по механизму обратной связи (thrA*), усиливается; и экспрессия гена cysE усиливается;
- экспрессия гена metJ ослабляется; экспрессия аллеля metA, кодирующего фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи (MetA*), усиливается; экспрессия аллеля thrA, кодирующего фермент с пониженным ингибированием треонином по механизму обратной связи (thrA*), усиливается; экспрессия гена cysE усиливается; и экспрессия генов metF и/или metH усиливается.
В конкретном аспекте изобретения рекомбинантный микроорганизм содержит следующие генетические модификации:
- ген metE подвергнут делеции,
- экспрессия генов metA*, metH, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE, thrA* и pyc усиливается и
- генов metJ, pykA, pykF, purU и yncA - ослабевает.
В конкретном воплощении изобретения микроорганизм происходит из семейства бактерий Enterobacteriaceae или Corynebacteriaceae.
Предпочтительно микроорганизмом является Escherichia coli или Corynebacterium glutamicum.
Культуральные условия
Изобретение также относится к способу получения метионина, включающему следующие стадии:
- культивирование рекомбинантного микроорганизма в подходящей культуральной среде, содержащей ферментируемый источник углерода и источник серы, и
- выделение метионина или его производных из культуральной среды.
Специалисты в данной области способны определять культуральные условия для микроорганизмов согласно изобретению. В частности бактерии осуществляют ферментацию при температуре от 20°С до 55°С, предпочтительно от 25°С до 40°С и более конкретно - при примерно 30°С для С.glutamicum и примерно 37°С для Е.coli.
Для Е.coli культуральная среда может иметь идентичный или аналогичный состав со средой М9 (Anderson, 1946) и средой М63 (Miller, 1992); или такой средой, как определено Schaefer et al., (1999).
Для С.glutamicum культуральная среда может иметь идентичный или аналогичный состав со средой BMCG (Liebl et al., 1989) или такой средой, как описано Riedel et al., (2001).
В некотором воплощении изобретения культуру подвергают ограничению или голоданию в отношении одного или нескольких неорганических субстратов. Оно относится к условиям, при которых рост микроорганизмов управляется количеством поставляемого неорганического химического соединения, которые все еще обеспечивают слабый рост. Такое ограничение роста микроорганизма было описано в патентной заявке WO 2009/043372. В предпочтительном воплощении изобретения культуру подвергают ограничению в отношении количества фосфата.
Действие «выделение метионина или его производных из культуральной среды» обозначает действие по выделению L-метионина и/или одного из его производных, в частности N-ацетилметионина (NAM) и S-аденозилметионина (SAM) и всех других производных, которые могут быть полезными. Способы выделения и очистки полученных соединений хорошо известны специалистам в данной области (см. в частности WO 2005/007862, WO 2005/059155).
Количество продукта в ферментационной среде можно определять с использованием целого ряда способов, известных в данной области, например, высокоэффективной жидкостной хроматографии (ВЭЖХ) или газовой хроматографии (ГХ). Например, количество метионина, полученного в среде, измеряют ВЭЖХ после дериватизации OPA/Fmoc с использованием L-метионина (Fluka, Ref 64319) в качестве стандарта. Количество NAM определяется с использованием рефрактометрической ВЭЖХ с использованием NAM (Sigma, Ref 01310) в качестве стандарта.
ПРИМЕРЫ
Настоящее изобретение, кроме того, определено в следующих примерах. Следует понимать, что данные примеры, в то время как они указывают на предпочтительные воплощения изобретения, приведены лишь ради иллюстрации. Основываясь на приведенном выше раскрытии и данных примерах, специалист в данной области может делать разные изменения изобретения для его адаптации к разным применениям и условиям без модификации существенных способов по изобретению.
В частности, примеры демонстрируют модифицированные штаммы Escherichia coli (Е.coli), но данные модификации можно легко проводить в других микроорганизмах того же самого семейства.
Escherichia coli принадлежит к семейству Enterobacteriaceae, которое содержит членов, которые являются грамотрицательными, палочковидными, неспорообразующими бактериями, типично имеющими длину 1-5 мкм. Большинство членов имеют жгутик, используемый для перемещения, но несколько родов являются неподвижными. Многие члены данного семейства являются нормальным компонентом кишечной флоры, находящимся в кишечнике человека и других животных, тогда как другие встречаются в воде или почве, или представляют собой паразитов на целом ряде разных животных и растений. Е.coli является одним из самых важных модельных организмов, но другие важные члены семейства Enterobacteriaceae включают Klebsiella, в частности Klebsiella terrigena, Klebsiella planticola или Klebsiella oxytoca, и Salmonella.
Кроме того, в нескольких патентных заявках отмечено, что оптимизацию продукции метионина можно легко применять к Е.coli и к Corynebacterium glutamicum без чрезмерного экспериментирования.
ПРИМЕР 1: ПРОТОКОЛЫ
Для конструирования штаммов, продуцирующих метионин, использовали несколько протоколов, описанных в следующих примерах.
Протокол 1: Хромосомные модификации путем гомологичной рекомбинации и отбора рекомбинантов (Datsenko, & Wanner, (2000)).
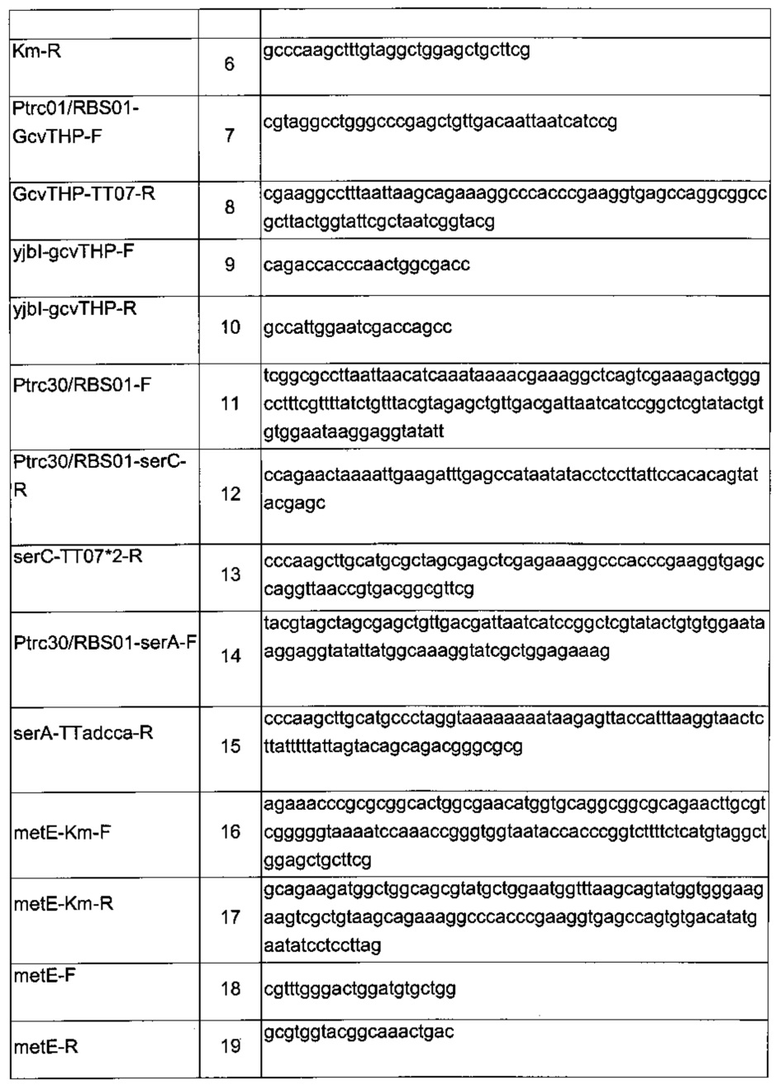
Аллельную замену или вставку гена в определенный хромосомный локус проводили путем гомологичной реомбинации, как описано Datsenko & Wanner (2000). Ген устойчивости к канамицину (Km) - kan, фланкированный сайтами распознавания Flp, амплифицировали ПЦР (полимеразная цепная реакция) с использованием плазмиды pKD4 в качестве матрицы. Образующиеся ПЦР-продукты использовали для трансформации штамма-реципиента Е.coli, содержащего плазмиду pKD46, которая экспрессирует рекомбиназу λ Red (γ, β, экзо). Затем отбирали трансформантов, устойчивых к антибиотику, и хромосомную структуру модифицированного локуса подтверждали ПЦР-анализом с подходящими праймерами, перечисленными в Таблице 3.
Ген устойчивости кап можно вырезать с использованием плазмиды рСР20, которая несет ген, кодирующий Flp рекомбиназу, как описано Datsenko & Wanner (2000). Плазмиду рСР20 вводили в подходящий штамм, и трансформантов распределяли на среде LB, дополненной ампициллином, при 30°С. Для того чтобы экспрессировать ген flp и удалить кассету устойчивости к канамицину, трансформантов культивировали при 37°С. Затем после выделения клоны, чувствительные к антибиотику, подтверждали ПЦР с использованием олигонуклеотидов, перечисленных в Таблице 3.
Протокол 2: трансдукция фага Р1
Хромосомные модификации переносили в данный штамм-реципиент Е.coli посредством трансдукции Р1. Данный протокол включает 2 стадии: (i) получение фагового лизата на штамме-доноре, имеющем устойчивость, ассоциированную с хромосомной модификацией, и (ii) инфекция штамма-реципиента данным фаговым лизатом.
Получение фагового лизата
- Инокулировать 100 мкл выращенной в течение ночи культуры штамма MG1655 с интересующей хромосомной модификацией в 10 мл 50 мкг/мл Km плюс 0,2% глюкоза плюс 5 мМ CaCl2.
- Инкубировать 30 мин при 37°С со встряхиванием.
-Добавить 100 мкл лизата фага Р1, полученного на штамме-доноре MG1655 (приблизительно 1×109 фагов/мл).
- Встряхивать при 37°С в течение 3 часов до полного лизиса клеток.
- Добавить 200 мкл хлороформа и встряхивать на вихревой мешалке.
- Центрифугировать 10 мин при 4500 g для устранения обломков клеток.
- Перенести супернатант в стерильную пробирку.
- Хранить лизат при 4°С.
Трансдукция
- Центрифугировать выращенную в течение ночи культуру штамма-реципиента Е.coli, культивированную в среде LB, 10 мин при 1500 g.
- Суспендировать клеточный осадок в 2,5 мл 10 мМ MgSO4, 5 мМ CaCl2.
- Инфицировать 100 мкл клеток 100 мкл фага Р1 из штамма MG1655 с модификацией на хромосоме (пробирка) и, в качестве контрольных пробирок, 100 мкл клеток без фага Р1 и 100 мкл фага Р1 без клеток.
- Инкубировать 30 мин при 30°С без встряхивания.
- Добавить 100 мкл 1 М цитрата натрия в каждую пробирку и встряхнуть на вихревой мешалке.
- Добавить 1 мл LB.
- Инкубировать 1 час при 37°С со встряхиванием.
- Центрифугировать 3 мин при 7000 об/мин.
- Высадить на чашки с LB плюс 50 мкг/мл Km.
- Инкубировать при 37°С в течение ночи.


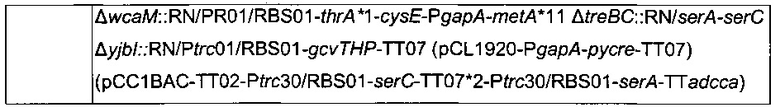


ПРИМЕР 2: Конструкция штамма 5, 


1. Штамм 1
Продуцирующий метионин штамм 1 (генотип в таблице 1) был описан в патентной заявке ЕР 10306164, которая включена в данную заявку посредством ссылки.
2. Конструкция штамма 2
Для увеличения метилен-тетрагидрофолатного пула в клетке сверхпродуцировали комплекс расщепления глицина, кодируемый опероном gcvTHP, путем добавления одной копии данного оперона на хромосому в локусе yibl. Эту дополнительную копию gcvTHP экспрессировали с использованием искусственного индуцибельного промотора trc и оптимизированного сайта связывания рибосомы, получая хромосомную интеграцию 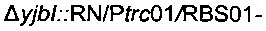

Для удаления гена yibl и замены его областью 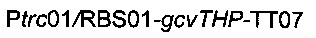 использовали стратегию гомологичной рекомбинации, описанную Datsenko & Wanner (2000). Данная стратегия обеспечивает вставку кассеты устойчивости к хлорамфениколу или канамицину, но также и дополнительной ДНК при удалении большинства из рассматриваемых генов. С этой целью конструировали следующую плазмиду:
использовали стратегию гомологичной рекомбинации, описанную Datsenko & Wanner (2000). Данная стратегия обеспечивает вставку кассеты устойчивости к хлорамфениколу или канамицину, но также и дополнительной ДНК при удалении большинства из рассматриваемых генов. С этой целью конструировали следующую плазмиду: 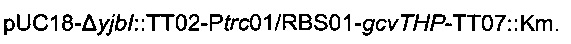
Данная плазмида 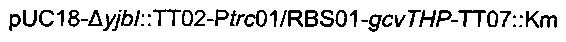 происходит из вектора pUC18 (Norrander et al., 1983) и содержит кассету устойчивости к канамицину, ассоциированную с областью
происходит из вектора pUC18 (Norrander et al., 1983) и содержит кассету устойчивости к канамицину, ассоциированную с областью 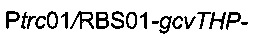
 которые обе клонировали между расположенными выше и ниже областями yibl.
которые обе клонировали между расположенными выше и ниже областями yibl.
Для конструирования 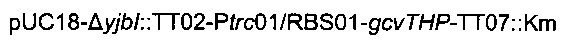 сначала конструировали плазмиду
сначала конструировали плазмиду 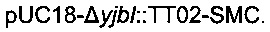 Данная плазмида несет расположенные выше и ниже области yibl, которые разделены терминатором транскрипции (Т1 гена rrnB Е.coli, названный ТТ02) и сайтом множественного клонирования (состоящего из сайтов рестрикции BstZ17I, HindIII, AvrII, ApaI и PacI, названным SMC). Эту последнюю область ПЦР-амплифицировали из геномной ДНК с использованием следующих олигонуклеотидов:
Данная плазмида несет расположенные выше и ниже области yibl, которые разделены терминатором транскрипции (Т1 гена rrnB Е.coli, названный ТТ02) и сайтом множественного клонирования (состоящего из сайтов рестрикции BstZ17I, HindIII, AvrII, ApaI и PacI, названным SMC). Эту последнюю область ПЦР-амплифицировали из геномной ДНК с использованием следующих олигонуклеотидов:
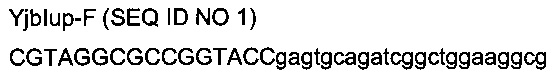
с
- областью (строчные буквы), гомологичной последовательности (4247987-4248009) области yjbl (эталонная последовательность на веб-сайте http://www.ecogene.org/),
- областью (заглавные буквы) рестрикционных сайтов SfoI и KpnI и дополнительных оснований.
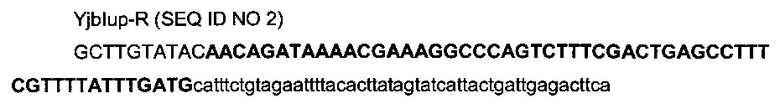
с
- областью (строчные буквы), гомологичной последовательности (4248931-4248980) области yjbl (эталонная последовательность на веб-сайте http://www.ecogene.org/),
- областью (заглавные буквы жирным шрифтом) терминатора транскрипции T1 гена rrnB Е.coli (Orosz et al, 1991),
- областью (заглавные буквы) рестрикционного сайта BstZ17I и частью рестрикционного сайта HindIII сайта множественного клонирования.

с
- областью (строчные буквы), гомологичной последовательности (4250286-4250335) области yjbl (эталонная последовательность на веб-сайте http://www.ecogene.org/),
- областью (заглавные буквы жирным шрифтом) терминатора транскрипции T1 гена rrnB Е.coli (Orosz et al, 1991),
- областью (заглавные буквы) всего сайта множественного клонирования.
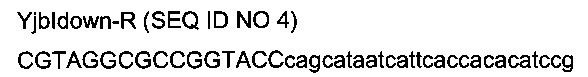
с
- областью (строчные буквы), гомологичной последовательности (4251224-4251249) области yjbl (эталонная последовательность на веб-сайте http://www.ecogene.org/),
- областью (заглавные буквы) рестрикционных сайтов SfoI и KpnI и дополнительных оснований.
Во-первых, фрагменты «вышеYjbI» и «нижеYjbI» ПЦР-амплифицировали из геномной ДНК MG1655 с использованием олигонуклеотидов Yjblup-F / Yjblup-R и Yjbldown-F / Yjbldown-R соответственно. Во-вторых, фрагмент «вышеYjbl-нижеYjbl» амплифицировали из ПЦР-фрагментов «вышеYjbl» и «нижеYjbl» (которые имеют перекрывающуюся область, включающую часть терминатора транскрипции T1 гена rrnB Е.coli и часть сайта множественного клонирования) с использованием олигонуклеотидов Yjblup-F / Yjbldown-R. ПЦР-фрагмент «вышeYjbl-нижeYjbl» вырезали рестрикционным ферментом SfoI и клонировали в сайты с тупыми концами EcoRI / HindIII вектора pUC18, получая плазмиду 
Затем из вектора pKD4 ПЦР-амплифицировали кассету устойчивости к канамицину с использованием следующих олигонуклеотидов:

с
- областью (строчные буквы) для амплификации кассеты устойчивости к канамицину (эталонная последовательность в Datsenko & Wanner, 2000,
- областью (заглавные буквы) рестрикционных сайтов SmaI и BstZ17I и дополнительных оснований.

с
- областью (строчные буквы) для амплификации кассеты устойчивости к канамицину (эталонная последовательность в Datsenko & Wanner, 2000,
- областью (заглавные буквы) рестрикционного сайта HindIII и дополнительных оснований.
ПЦР-фрагмент вырезали рестрикционными ферментами BsfZ17I и HindIII и клонировали в сайты BsfZ17I / HindIII плазмиды  получая плазмиду
получая плазмиду 
Наконец, фрагмент 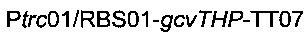 ПЦР-амплифицировали из геномной ДНК штамма 1 с использованием олигонуклеотидов
ПЦР-амплифицировали из геномной ДНК штамма 1 с использованием олигонуклеотидов 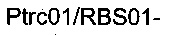
 (описаны ниже). Данный ПЦР-фрагмент вырезали рестрикционными ферментами ApaI и PacI и клонировали в сайты ApaI / PacI плазмиды
(описаны ниже). Данный ПЦР-фрагмент вырезали рестрикционными ферментами ApaI и PacI и клонировали в сайты ApaI / PacI плазмиды 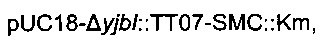 получая плазмиду
получая плазмиду 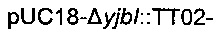
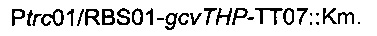
Рекомбинантные плазмиды подтверждали секвенированием ДНК.
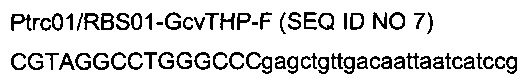
с
- областью (строчные буквы), гомологичной части искусственного индуцибельного промотора trc, локализованного у штамма 1 выше оперона gcvTHP,
- областью (заглавные буквы) рестрикционных сайтов StuI и ApaI и дополнительных оснований.
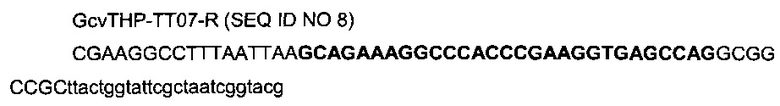
с
- областью (строчные буквы), гомологичной последовательности (3044190-3044214) гена gcvP (эталонная последовательность на веб-сайте http://www.ecogene.org/),
- областью (заглавные буквы жирным шрифтом) последовательности терминатора транскрипции T7te, названного ТТ07 (Harrington et al., 2001),
- областью рестрикционных сайтов PacI, StuI и дополнительных оснований.
Наконец, фрагмент 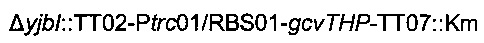 получали разрезанием плазмиды
получали разрезанием плазмиды 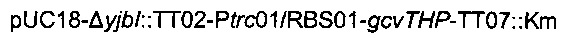 рестрикционным ферментом KpnI и затем вводили посредством электропорации согласно Протоколу 1 в штамм MG1655 metA*11 pKD46. Затем отбирали устойчивых к канамицину трансформантов, и вставку фрагмента
рестрикционным ферментом KpnI и затем вводили посредством электропорации согласно Протоколу 1 в штамм MG1655 metA*11 pKD46. Затем отбирали устойчивых к канамицину трансформантов, и вставку фрагмента 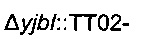
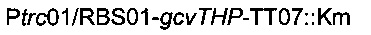 подтверждали ПЦР-анализом с олигонуклеотидами
подтверждали ПЦР-анализом с олигонуклеотидами 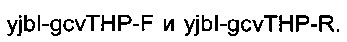 Подтвержденный и отобранный штамм называли
Подтвержденный и отобранный штамм называли 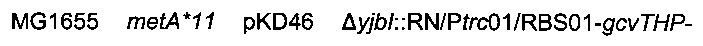
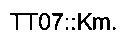

гомологичная последовательности (4247754-4247774) области yjbl (эталонная последовательность на веб-сайте http://www.ecoqene.org/)
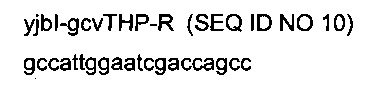
гомологичная последовательности (4251489-4251508) области yjbl (эталонная последовательность на веб-сайте http://www.ecoqene.orq/)
Затем хромосомную модификацию 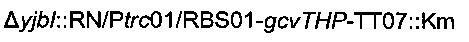 трансдуцировали в штамм 1 согласно Протоколу 2.
трансдуцировали в штамм 1 согласно Протоколу 2.
Отбирали устойчивых к канамицину трансдуктанов, и присутствие хромосомной модификации 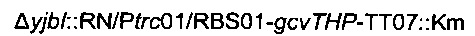 подтверждали посредством ПЦР с праймерами
подтверждали посредством ПЦР с праймерами 
Образовавшийся штамм 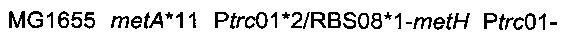

3. Конструирование штамма 3
Для конструирования штамма 3 придающую устойчивость кассету, ассоциированную с хромосомной интеграцией 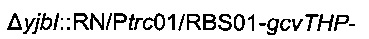
 штамма 2, удаляли согласно Протоколу 1.
штамма 2, удаляли согласно Протоколу 1.
Отбирали чувствительные к канамицину клоны, и отсутствие кассеты, придающей устойчивость к канамицину, подтверждали ПЦР с праймерами yjbl-gcvTHP-F и yjbl-gcvTHP-R.
Образовавшийся штамм 

 назвали штамм 3.
назвали штамм 3.
4. Конструирование штамма 4
Плазмида pCL1920-PgapA-pycre-ТТ07, описанная в патентной заявке PCT/FR2010/052937 (которая включена в данную заявку в виде ссылки), была введена в штамм 2 с образованием следующего штамма 

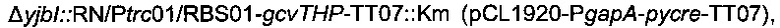 названного штамм 4.
названного штамм 4.
5. Конструирование штамма 5
Для увеличения потока в сериновый путь гены serC и serA сверхэкспрессировали, благодаря искусственным промоторам и оптимизированным сайтам связывания рибосомы и с использованием бактериальной искусственной хромосомы pCC1BAC (Epicentre). С этой целью конструировали следующую плазмиду 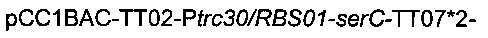
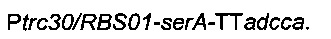
Для конструирования 
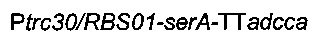 ПЦР-амплифицировали области
ПЦР-амплифицировали области 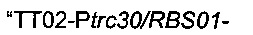
 и
и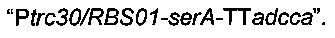
Для области 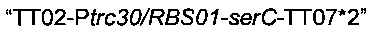 сначала синтезировали мегапраймер, содержащий терминатор транскрипции (Т1 rrnB, аннотированный ТТ02), искусственный промотор (Ptrc30), оптимизированный сайт связывания рибосомы (RBS01) и начало гена serC, посредством короткой ПЦР с использованием олигонуклеотидов Ptrc30/RBS01-F и Ptrc30/RBS01-serC-R (описаны ниже) без добавления матрицы. Во-вторых, фрагмент
сначала синтезировали мегапраймер, содержащий терминатор транскрипции (Т1 rrnB, аннотированный ТТ02), искусственный промотор (Ptrc30), оптимизированный сайт связывания рибосомы (RBS01) и начало гена serC, посредством короткой ПЦР с использованием олигонуклеотидов Ptrc30/RBS01-F и Ptrc30/RBS01-serC-R (описаны ниже) без добавления матрицы. Во-вторых, фрагмент 
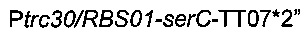 амплифицировали посредством ПЦР с использованием геномной ДНК MG1655 Е.coli в качестве матрицы, синтезированного мегапраймера и олигонуклеотида serC-TT07*-R (описан ниже).
амплифицировали посредством ПЦР с использованием геномной ДНК MG1655 Е.coli в качестве матрицы, синтезированного мегапраймера и олигонуклеотида serC-TT07*-R (описан ниже).
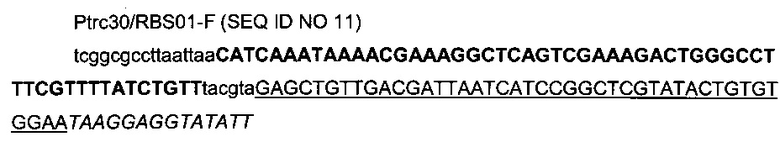
с
- областью (заглавные буквы жирным шрифтом) терминатора транскрипции Т1 гена rrnB Е.coli (Orosz et al., 1991),
- областью (заглавные подчеркнутые буквы), гомологичной искусственному индуцибельному промотору trc,
- областью (заглавные буквы курсивом), гомологичной оптимизированному сайту связывания рибосомы,
- областью (строчные буквы) рестрикционных сайтов NarI, PacI и дополнительных оснований.

с
- областью (заглавные подчеркнутые буквы), гомологичной искусственному индуцибельному промотору trc,
- областью (заглавные буквы курсивом), гомологичной оптимизированному сайту связывания рибосомы,
- областью (строчные буквы), гомологичной последовательности (956876-956904) гена serC (эталонная последовательность на веб-сайте http://www.ecoqene.org/).
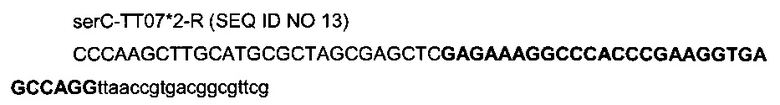
с
- областью (заглавные буквы жирным шрифтом) последовательности терминатора транскрипции T7te (Harrington et al., 2001), которая обладает делецией основания в 29-ом положении (названа ТТ07*2),
- областью (строчные буквы), гомологичной последовательности (957946-957964) гена serC (эталонная последовательность на веб-сайте http://www.ecogene.org/),
- областью (заглавные буквы) рестрикционных сайтов HindIII, SphI, SacI и NheI и дополнительных оснований.
Аналогичным образом фрагмент 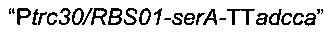 амплифицировали посредством ПЦР с использованием геномной ДНК Е.coli MG1655 в качестве матрицы и олигонуклеотидов Ptrc30/RBS01-serA-F и serA-TTadcca-R (описаны ниже).
амплифицировали посредством ПЦР с использованием геномной ДНК Е.coli MG1655 в качестве матрицы и олигонуклеотидов Ptrc30/RBS01-serA-F и serA-TTadcca-R (описаны ниже).

с
- областью (заглавные подчеркнутые буквы), гомологичной искусственному индуцибельному промотору trc,
- областью (заглавные буквы курсивом), гомологичной оптимизированному сайту связывания рибосомы,
- областью (строчные буквы), гомологичной последовательности (3056408-3056432) гена serA (эталонная последовательность на веб-сайте http://www.ecogene.org/).
- областью (строчные буквы) рестрикционного сайта NheI и дополнительных оснований.
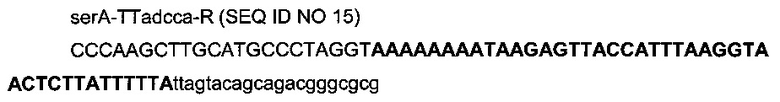
с
- областью (строчные буквы жирным шрифтом) последовательности терминатора транскрипции TTadc (терминатор транскрипции гена adc из Clostridium acetobutylicum, гомологичный от 179847 до 179807 мегаплазмиде pSLO1),
- областью (строчные буквы), гомологичной последовательности (3055200-3055220) гена serA (эталонная последовательность на веб-сайте http://www.ecoqene.org/),
- областью (заглавные буквы) рестрикционных сайтов AvrII, SphI, HindIII и дополнительных оснований.
ПЦР-фрагменты  и
и 
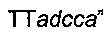 вырезали рестрикционными ферментами NarI / NheI и NheI / SphI соответственно и оба клонировали в сайты NarI / SphI плазмиды рСС1ВАС, получая плазмиду
вырезали рестрикционными ферментами NarI / NheI и NheI / SphI соответственно и оба клонировали в сайты NarI / SphI плазмиды рСС1ВАС, получая плазмиду 
Рекомбинантную плазмиду подтверждали секвенированием ДНК. Наконец, плазмиду 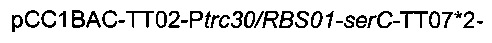
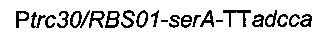 вводили в штамм 4, получая следующий штамм
вводили в штамм 4, получая следующий штамм 
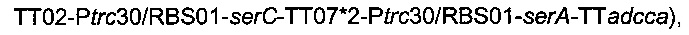 названный штамм 5.
названный штамм 5.
6. Идентификация мутации metE в штамме 5
Путем измерения метионинсинтазной активности (МЕТЕ) штамма 5 авторы изобретения идентифицировали, что ген metE не был функциональным из-за некоторых мутаций, дающих усеченный белок MetE.
Данная мутация представляет собой делецию 13 п.н. (с 417-го по 429-ое основание гена) гена metE, приводящую к мутации, связанной со сдвигом рамки считывания. Следовательно, трансляция белка сокращается (посредством сдвига рамки считывания вводится терминирующий кодон) и приводит к образованию усеченного белка из 152 аминокислот вместо 753.
Здесь приведена последовательность белка MetE дикого типа (SEQ ID NO 21):


Здесь приведена последовательность усеченного белка MetE* (SEQ ID NO 22):
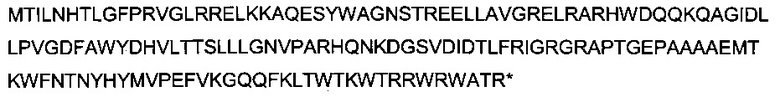
ПРИМЕР 3: конструирование штамма 7, 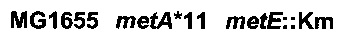

1. Конструирование штамма 6
Для исследования того, могло ли восстановление функционального белка MetE модифицировать продукцию метионина штаммом 5, авторы изобретения заменили усеченный ген metE геном дикого типа с использованием стратегии гомологичной рекомбинации, описанной Datsenko & Wanner (2000).
С этой целью кассету устойчивости к канамицину фланкировали фрагментами, гомологичными области metE, фрагмент «metE::Km» ПЦР-амплифицировали с использованием олигонуклеотидов metE-Km-F и metE-Km-R (описаны ниже). Фрагмент «metE::Km» вводили в штамм MG1655 metA*11 pKD46, который имеет функциональную версию гена metE.

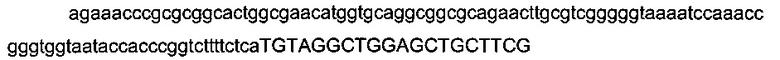
с
- областью (строчные буквы), гомологичной последовательности (4013277-4013376) области metE (эталонная последовательность на веб-сайте http://www.ecogene.org/),
- областью (заглавные буквы) для амплификации кассеты устойчивости к канамицину (эталонная последовательность в Datsenko & Wanner, 2000).
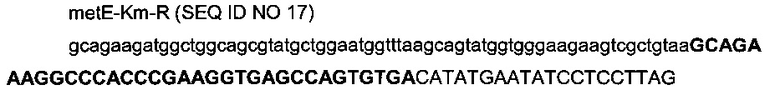
с
- областью (строчные буквы), гомологичной последовательности (4013377-4013442) области metE (эталонная последовательность на веб-сайте http://www.ecogene.org/),
- областью (заглавные буквы жирным шрифтом) последовательности терминатора транскрипции T7te (Harrington et al., 2001),
- областью (заглавные буквы) для амплификации кассеты устойчивости к канамицину (эталонная последовательность в Datsenko & Wanner, 2000).
Отбирали устойчивых к канамицину рекомбинантов, и присутствие кассеты Km ниже гена metE подтверждали ПЦР с олигонуклеотидами metE-F и metE-R (описаны ниже). Подтвержденный и отобранный штамм назвали MG1655 metA*11 pKD46 metE::Km.

гомологичная последовательности (4012495-4012516) области metE (эталонная последовательность на веб-сайте http://www.ecogene.org/)
гомологичная последовательности (4013672-4013691) области metE (эталонная последовательность на веб-сайте http://www.ecogene.org/)
Затем в штамм 3 трансдуцировали хромосомную модификацию metE::Km согласно Протоколу 2.
Отбирали рекомбинантов с устойчивостью к канамицину, и присутствие кассеты Km ниже гена metE подтверждали ПЦР с олигонуклеотидами metE-F и metE-R (описаны ниже). Присутствие гена metE с последовательностью дикого типа подтверждали секвенированием ДНК.
Образовавшийся штамм 

 был назван штамм 6.
был назван штамм 6.
2. Конструирование штамма 7
Плазмиду  (описанную в патентной заявке PCT/FR2010/052937) и плазмиду
(описанную в патентной заявке PCT/FR2010/052937) и плазмиду 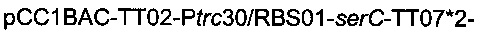
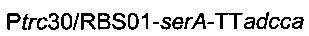 (описанную выше) вводили в штамм 6, получая штамм
(описанную выше) вводили в штамм 6, получая штамм 
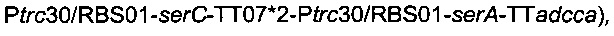 названный штамм 7.
названный штамм 7.
Измерение активности кобаламин-независимой метионинсинтазы (MS, MetE) штамма 6 подтвердило то, что белок MetE является функциональным.
ПРИМЕР 4: ПРОДУКЦИЯ L-МЕТИОНИНА ПОСРЕДСТВОМ ФЕРМЕНТАЦИИ В БИОРЕАКТОРЕ
Штаммы, которые продуцировали существенные количества метионина, затем тестировали при условиях продукции в 0,5 л ферментерах (GX, GPC) с использованием стратегии с подпиткой.
Вкратце, для инокуляции 24-часовой предкультуры в минимальной среде (В1а) использовали 24-часовую культуру, выращенную в 10 мл среды LB с 2,5 г⋅л-1 глюкозы. Данные инкубации проводили в 500 мл колбах с дефлектором, содержащих 40 мл минимальной среды (В1а), на ротационном шейкере (200 об/мин). Первую предкультуру осуществляли при температуре 30°С, вторую - при температуре 34°С.
Третью стадию предкультуры проводили в биореакторах (Sixfors), заполненных 200 мл минимальной среды (B1b), инокулированной до концентрации биомассы 1,2 г⋅л-1 5 мл концентрированной предкультуры. Температуру предкультуры поддерживали на постоянном уровне - 34°С, и рН автоматически подводили до значения 6,8 с использованием 10%-ного раствора NH4OH. Концентрацию растворенного кислорода непрерывно подводили до значения 30% насыщения парциального давления воздуха посредством подачи воздуха и/или встряхивания. После исчерпания глюкозы в загрузочной среде начинали подпитку с исходной скоростью тока 0,7 мл⋅ч-1 и увеличивали ее экспоненциально в течение 24 часов со скоростью увеличения 0,13 ч-1 для того, чтобы получить конечную концентрацию клеток примерно 20 г⋅л-1.
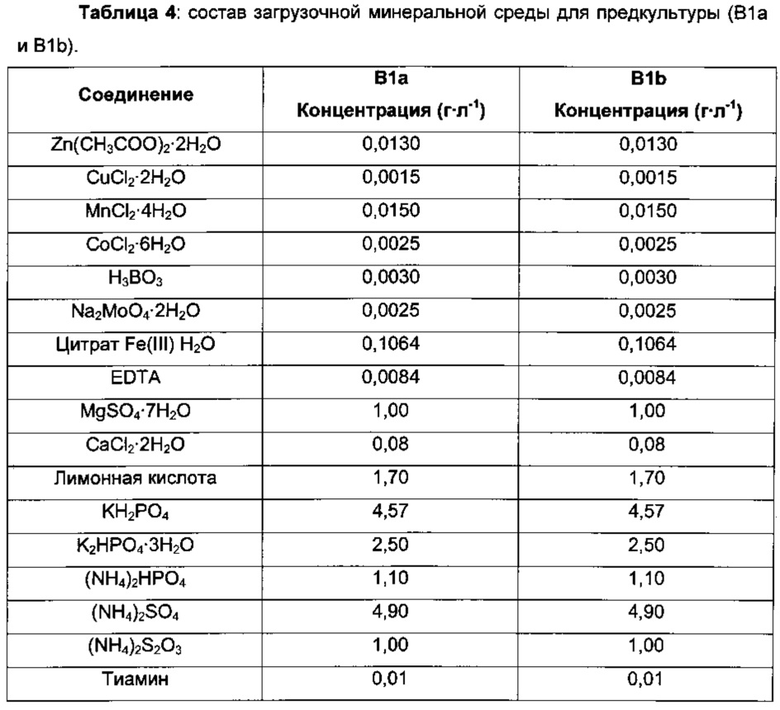

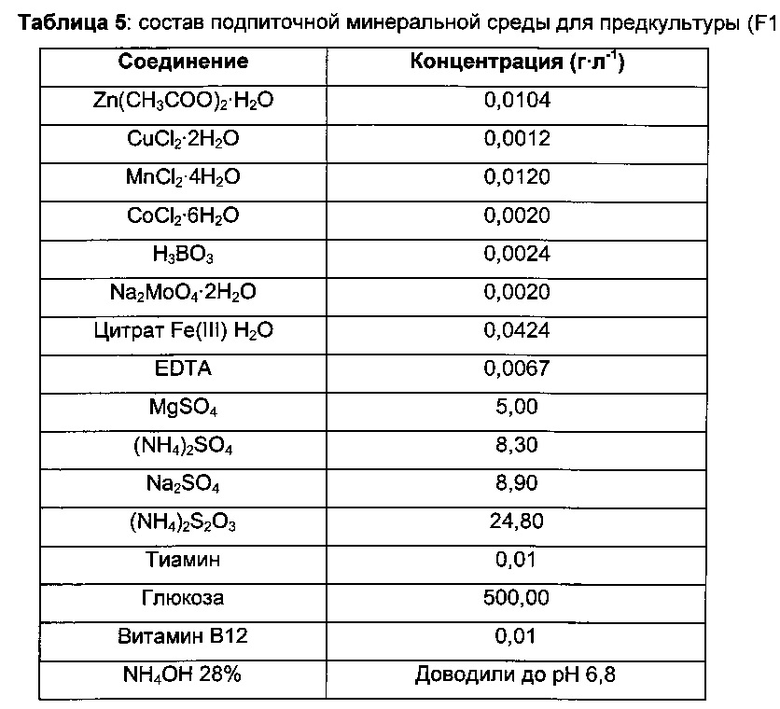
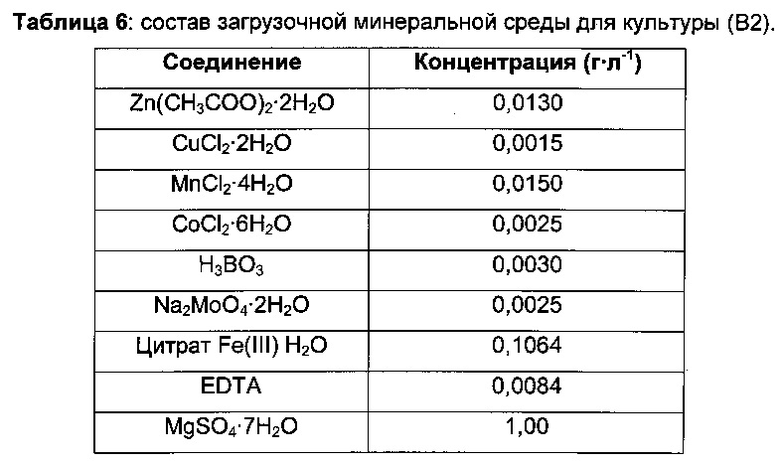
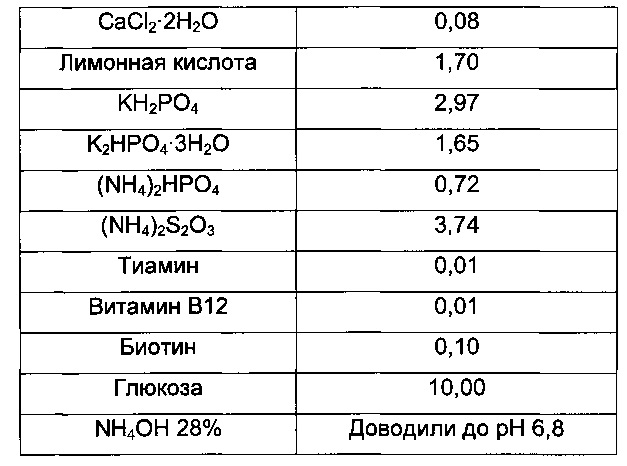
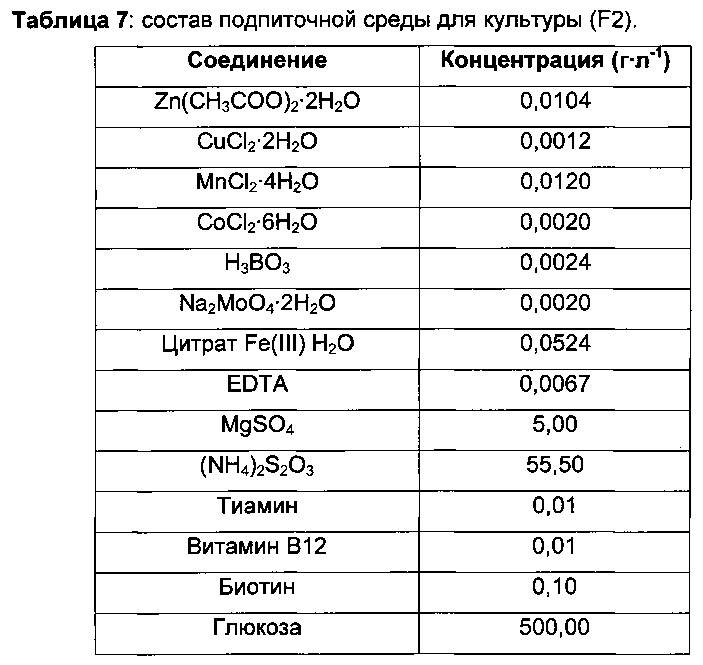
Затем 0,5 л ферментеры GX (GPC) заполняли 220 мл минимальной среды (В2) и инокулировали объемом предкультуры, варьирующим от 20 до 30 мл, до концентрации биомассы 2,1 г⋅л-1.
Температуру культуры поддерживали на постоянном уровне - 37°С, и рН поддерживали в рабочем значении (6,8) путем автоматического добавления растворов NH4OH (10% NH4OH). На протяжении загрузочной фазы устанавливали исходную скорость встряхивания 200 об/мин, и ее увеличивали вплоть до 1000 об/мин во время подпиточной фазы. На протяжении загрузочной фазы устанавливали исходную скорость тока воздуха 0,3 л⋅мин-1, и ее увеличивали вплоть до 0,7 л⋅мин-1 в начале подпиточной фазы. Концентрацию растворенного кислорода поддерживали на уровне от 20 до 40%, предпочтительно 30% от насыщения путем усиления встряхивания.
При достижении клеточной массой концентрации, близкой к 5 г⋅л-1, начинали подпитку с исходной скоростью тока 1,9 мл⋅ч-1. Подпиточный раствор инъецировали с сигмоидным профилем с возрастающей скоростью тока, которая достигала 8,8 мл⋅ч-1 через 26 часов. Точные условия подпитки были рассчитаны посредством уравнения: 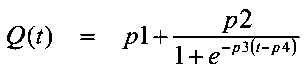
где Q(t) представляет собой скорость тока подпитки в мл⋅ч-1 для загрузочного объема 600 мл с р1 равным 0,66; р2 равным 8,21; р3 равным 0,27, р4 равным 6,50.
После 26 часов подпитки насос с подпиточным раствором останавливали, и культивирование прекращали после полного исчерпания глюкозы.
Внеклеточные аминокислоты количественно измеряли ВЭЖХ (высокоэффективная жидкостная хроматография) после дериватизации OPA/Fmoc, и другие релевантные метаболиты анализировали с использованием ВЭЖХ с рефрактометрической детекцией (органические кислоты и глюкоза) и ГХ-МС (газовая хроматография-масс-спектрометрия) после силирования.

Как можно видеть в таблице 8, приведенной выше, выход продукции метионина значимо возрастал при мутации гена metE. Штамм 5, содержащий мутировавший ген metE, имеет выход, превышающий 4 балла, по сравнению со штаммом 7, который содержит функциональный белок MetE.
Объем ферментера рассчитывали путем суммирования исходного объема реактора, количества растворов, добавленных для регуляции рН и подпитки культуры, и путем вычитания объема, использованного для отбора образцов и потерянного в результате испарения.
Подпиточный объем непрерывно отслеживали путем взвешивания подпиточного маточного раствора. Затем рассчитывали количество инъецированной глюкозы на основе инъецированной массы, плотности раствора и концентрации глюкозы, определенной способом Brix ([Глюкоза]). Выход метионина выражали следующим образом:
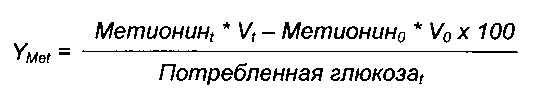
Здесь представлен конечный выход, полученный на протяжении культуры для каждого штамма. Метионин0 и Метионинt представляют собой исходную и конечную концентрацию метионина соответственно, и V0 и Vt представляют собой исходный и конечный объемы.
Потребленную глюкозу рассчитывали следующим образом:

Инъецированная глюкозаt = подпиточный объемt * [Глюкоза]
Потребленная глюкозаt = [Глюкоза]0 * V0 + Инъецированная глюкоза - [Глюкоза]остаточная * Vt, причем [Глюкоза]0, [Глюкоза], [Глюкоза]остаточная представляют собой исходную, подпиточную и остаточную концентрации глюкозы соответственно.
ПРИМЕР 5: ИЗМЕРЕНИЕ АКТИВНОСТИ КОБАЛАМИН-НЕЗАВИСИМОЙ МЕТИОНИНСИНТАЗЫ (MetE)
Для определения активности кобаламин-независимой метионинсинтазы (MS, MetE) in vitro штаммы 7 и 5 Е.coli, несущие ген metE дикого типа или мутировавший соответственно, культивировали в минимальной среде, как описано в примере 3 выше, и собирали в конце логарифмической фазы центрифугированием. Осадки ресуспендировали в холодном 20 мМ калий-фосфатном буфере, рН 7,2, содержащем смесь ингибиторов протеаз с EDTA. Затем клетки разрушали встряхиванием с шариками с использованием системы Precellys (Bertin Technologies; 2 × 10 с при 6500 об/мин), с последующим центрифугированием при 12000 g при 4°С в течение 30 минут. Супернатанты обессоливали и использовали для ферментативных анализов. Концентрации белка определяли с использованием анализа с реактивом Бредфорд (Bradford, 1976).
Для определения активности MS 40 мкг неочищенных клеточных экстрактов инкубировали в течение 15 минут при 37°С с 1 мМ DL-гомоцистеином и 0,25 мМ метил-тетрагидроптероил-триглутаматом в 100 мМ калий-фосфатном буфере, рН 7,2, 5 мМ MgSO4. Метионин, продуцированный ферментом кобаламин-независимой метионинсинтазой, количественно измеряли ГХ-МС после дериватизации трет-бутилдиметилсилилтрифторацетамидом (TBDMSTFA). Аспартат и норлейцин включали в качестве внутренних стандартов.
Результаты по активностям кобаламин-независимой метионинсинтазы представлены в таблице 9 ниже.

Как можно видеть в таблице 9, штамм 5 (ΔmetE) полностью утрачивал его MS активность, тогда как штамм 7 сохранял значительную активность. Эта потеря активности коррелировала со значимым улучшением продукции метионина.
ПРИМЕР 6: ЭФФЕКТ ДЕЛЕЦИИ ГЕНА МЕТЕ НА ПРОДУКЦИЮ L-МЕТИОНИНА
Для оценки эффекта полной делеции гена metE на продукцию L-метионина авторы изобретения подвергали делеции мутировавший ген metE штамма 5. Авторы изобретения ввели чистую делецию гена metE в данный штамм с использованием гомологичной рекомбинации, как описано ранее и с использованием стратегии, предложенной Datsenko & Wanner (2000).
После замены мутировавшего гена metE кассетой устойчивости к канамицину отбирали рекомбинантов, устойчивых к канамицину, и подтверждали секвенированием ДНК.
Один из них культивируется, как описано в примере 4, и продуцированный L-метионин количественно измеряется посредством ВЭЖХ.
Данный штамм с чистой делецией metE продуцирует больше метионина, чем штамм 7, который обладает функциональным белком MetE: делеция metE приводит к увеличению выхода метионина более чем на 15%.













Настоящее изобретение относится к биохимии, в частности к рекомбинантному микроорганизму из семейства бактерий Enterobacteriaceae, например E.coli, оптимизированному для ферментативной продукции метионина. Указанный микроорганизм модифицирован таким образом, что у него ослаблена активность кобаламин-независимой метионинсинтазы MetE и ген metH сверхэкспрессирован по сравнению с немодифицированным микроорганизмом. Изобретение также раскрывает способ ферментативной продукции метионина посредством культивирования указанного рекомбинантного микроорганизма в культуральной среде, содержащей источник углерода и источник серы. Настоящее изобретение позволяет повысить выход метионина. 2 н. и 8 з.п. ф-лы, 9 табл., 6 пр.
1. Рекомбинантный микроорганизм из семейства бактерий Enterobacteriaceae, оптимизированный для ферментативной продукции метионина, где в указанном микроорганизме активность кобаламин-независимой метионинсинтазы MetE отсуствует и ген metH сверхэкспрессирован по сравнению с немодифицированным организмом.
2. Микроорганизм по п. 1, в котором кобаламин-независимая метионинсинтаза MetE кодируется геном metE, в котором по меньшей мере часть гена metE подвергнута делеции или белок MetE кодируется мутировавшим геном metE.
3. Микроорганизм по п. 2, в котором делеция гена metE представляет собой делецию оснований, находящихся между 417-м и 429-м положениями.
4. Микроорганизм по п. 1, в котором увеличена экспрессия по меньшей мере одного из следующих генов: ptsG, рус, pntAB, cysP, cysU, cysW, cysA, cysM, cysJ, cysI, cysH, gcvT, gcvH, gcvP, Ipd, serA, serB, serC, cysE, metF, thrA, аллеля metA, кодирующего фермент с пониженной чувствительностью к S-аденозилметионину и/или метионину по механизму обратной связи (metA*), thrA или аллеля thrA, кодирующего фермент с пониженным ингибированием треонином по механизму обратной связи (thrA*).
5. Микроорганизм по п. 4, в котором по меньшей мере один ген находится под контролем индуцибельного промотора.
6. Микроорганизм по п. 1, в котором ослаблена экспрессия по меньшей мере одного из следующих генов: metJ, pykA, pykF, purU, ybdL или yncA.
7. Микроорганизм по п. 1, в котором:
а) ген metE подвергнут делеции,
б) экспрессия генов metA*, cysPUWAM, cysJIH, gcvTHP, metF, serA, serB, serC, cysE, thrA* и рус усилена; и
в) экспрессия генов metJ, pykA, pykF, purU и yncA ослаблена.
8. Микроорганизм по п. 1, где указанный микроорганизм представляет собой Escherichia coli.
9. Способ ферментативной продукции метионина, включающий следующие стадии:
а) культивирование рекомбинантного микроорганизма по любому из пп. 1-8 в подходящей культуральной среде, содержащей ферментируемый источник углерода и источник серы, и
б) выделение метионина из данной культуральной среды.
10. Способ по п. 9, при котором рост рекомбинантного микроорганизма подвергается действию ограничения или недостаточности в отношении одного или нескольких неорганических субстратов, в частности, фосфата и/или калия в культуральной среде.
| HONDORP ER, et al., Oxidation of cysteine 645 of cobalamin-independent methionine synthase causes a methionine limitation in Escherichia coli., J Bacteriol., 2009, 191(10), pp.3407-3410 | |||
| WO2009144270 A1, 03.12.2009 | |||
| BORJA et al., Engineering Escherichia coli to increase plasmid DNA production in high cell-density cultivations in batch mode, Microbial Cell Factories, 2012, 11, pp.132(1-9) | |||
| RU 2008105480 A, 27.08.2009. |

