Область техники
Настоящее изобретение относится к О-фосфосерин-экспортирующему белку и к способу получения О-фосфосерина, цистеина и производных цистеина с его применением.
Предшествующий уровень техники
L-цистеин, аминокислота, которая играет важную роль в метаболизме серы у всех живых организмов, используется не только в синтезе биологических белков, таких как кератин волос и т.д., глутатион, биотин, метионин, и других серосодержащих метаболитов, но также как предшественник в биосинтезе коэнзима А.
Способы получения L-цистеина с использованием микроорганизмов, известные в данной области техники, включают: 1) способ биологического превращения D,L-2-амино-2-тиазолин-4-карбоновой кислоты (D,L-ATC) в L-цистеин с использованием микроорганизмов, 2) способ получения L-цистеина путем прямой ферментации с использованием Е. coli (ЕР 0885962 В; Wada М and Takagi Н, Appl. Microbiol. Biochem., 73:48-54, 2006) и 3) способ получения О-фосфосерина (ниже "OPS") посредством ферментации с использованием микроорганизмов и последующего превращения О-фосфосерина в L-цистеин посредством взаимодействия О-фосфосерина с сульфидом при каталитическом действии О-фосфосеринсульфгидрилазы (ниже "OPSS") (европейский патент 2444481).
В частности, чтобы получить цистеин посредством способа 3) с высоким выходом, OPS, являющийся предшественником, должен быть получен в избыточном количестве.
В таких обстоятельствах авторы настоящего изобретения приложили значительные усилия для определения подходящего экспортирующего фактора, который может беспрепятственно экспортировать О-фосфосерин, полученный в OPS-продуцирующем штамме, за пределы клеток и увеличивать продуцирование OPS. В результате они обнаружили новый белок, экспортирующий OPS, тем самым осуществив настоящее изобретение.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Техническая задача
Авторы настоящего изобретения приложили значительные усилия для определения подходящих экспортирующих факторов, которые могут беспрепятственно экспортировать О-фосфосерин за пределы клеток и увеличивать продуцирование OPS, и в результате они обнаружили новый OPS-экспортирующий белок, тем самым осуществив настоящее изобретение.
Решение технической задачи
Одна задача настоящего изобретения заключается в предложении рекомбинантного микроорганизма для получения О-фосфосерина, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью.
Другая задача настоящего изобретения заключается в предложении способа получения О-фосфосерина с использованием рекомбинантного микроорганизма, продуцирующего О-фосфосерин, по настоящему изобретению.
Еще одна задача по настоящему изобретению заключается в предложении способа получения цистеина или его производного с использованием рекомбинантного микроорганизма, продуцирующего О-фосфосерин, по настоящему изобретению.
Полезные эффекты
Если О-фосфосерин получают с использованием рекомбинантного микроорганизма, продуцирующего О-фосфосерин, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью, это может привести к высокопродуктивному продуцированию О-фосфосерина по сравнению с использованием существующего немодифицированного штамма.
Подробное описание предпочтительных воплощений
Настоящее изобретение будет описано подробно. При этом, каждое описание и воплощение, представленное здесь, может быть применено к другим описаниям и воплощениям соответственно. То есть все комбинации различных элементов, раскрытых в данном документе, входят в объем настоящего изобретения. Кроме того, объем настоящего раскрытия не ограничен конкретным описанием, представленным ниже.
В одном аспекте настоящего изобретения для выполнения вышеприведенных задач в настоящем изобретении предложен рекомбинантный микроорганизм, продуцирующий О-фосфосерина, в котором активность белка mdtH повышена по сравнению с его эндогенной активностью.
При использовании здесь термин "О-фосфосерин" (в дальнейшем "OPS") относится к эфиру фосфорной кислоты серина, который является составляющим компонентом многих белков. В частности, OPS является предшественником L-цистеина и может быть превращен в цистеин посредством взаимодействия с сульфидом при каталитическом действии OPS-сульфгидрилазы (ниже "OPSS"), но без ограничения этим (европейский патент 2444481).
В частности, рекомбинантный микроорганизм по настоящему изобретению может иметь повышенную О-фосфосерин-экспортирующую активность по сравнению с эндогенной активностью. Белок mdtH по настоящему изобретению может иметь активность экспорта О-фосфосерина за пределы клеток и такая активность была идентифицирована в настоящем изобретении впервые.
В одном примере белок mdtH по настоящему изобретению может быть мембранным белком и может быть получен из Escherichia coli.
Кроме того, белок mdtH по настоящему изобретению может быть транспортером, принадлежащим к суперсемейству главных фасилитаторов (MFS).
В частности, белок mdtH по настоящему изобретению может быть белком, включающим аминокислотную последовательность SEQ ID NO: 1, или аминокислотную последовательность, имеющую по меньшей мере 95% идентичность с SEQ ID NO: 1, при этом имеющим О-фосфосерин-экспортирующую активность. Например, белок, включающий аминокислотную последовательность, имеющую по меньшей мере 95% идентичность с аминокислотной последовательностью SEQ ID NO: 1, может включать любой белок, который имеет OPS экспрессирующую активность, идентичную или соответствующую белку mdtH, включающему аминокислотную последовательность SEQ ID NO: 1 без ограничений.
Более конкретно, аминокислотная последовательность, имеющая по меньшей мере 95% идентичность с аминокислотной последовательностью SEQ ID NO: 1, может быть последовательностью, имеющей идентичность, составляющую 96% или более, 97% или более, 98% или более, 99% или более, 99,5% или более или 99,75% или более, и 100% или менее с аминокислотной последовательностью SEQ ID NO: 1.
Кроме того, белок mdtH по настоящему изобретению может включать варианты белка, у которых часть последовательности делетирована, модифицирована, заменена, консервативно заменена или добавлена к аминокислотной последовательности SEQ ID NO: 1 или к аминокислотной последовательности, имеющей по меньшей мере 95% идентичность с аминокислотной последовательностью SEQ ID NO: 1, при условии, что они имеют О-фосфосерин-экспортирующую активность и могут по существу входить в объем настоящего изобретения.
Кроме того, специалистам в данной области очевидно, что белок mdtH по настоящему изобретению может включать бессмысленную последовательность, добавленную выше или ниже аминокислотной последовательности SEQ ID NO: 1 или аминокислотной последовательности, имеющей по меньшей мере 95% идентичность с аминокислотной последовательностью SEQ ID NO: 1, но при этом имеющей О-фосфосерин-экспортирующую активность, природную мутацию, или ее молчащую мутацию.
При использовании здесь термин "гомология" или "идентичность" относится к степени сходства двух данных аминокислотных последовательностей или нуклеотидных последовательностей и может быть выражена в процентах. Термины "гомология" и "идентичность" часто могут использоваться взаимозаменяемо друг с другом.
Гомология или идентичность последовательности консервативного полинуклеотида или полипептида может быть определена стандартными алгоритмами выравнивания с использованием штрафа за пробел по умолчанию, установленного используемой программой. По существу, гомологичные или идентичные последовательности, как правило, гибридизуются со всеми или частью последовательностей при умеренных или жестких условиях. Очевидно, что также включена гибридизация с полинуклеотидами, содержащими общий кодон или вырожденные кодоны в гибридизирующихся полинуклеотидах.
Обладают ли последовательности двух полинуклеотидов или полипептидов гомологией, сходством или идентичностью можно определить, например, посредством известного компьютерного алгоритма, такого как программа "FASTA" (Pearson et at., (1988) Proc. Natl. Acad. Sci. USA 85:2444) с использованием параметров по умолчанию. В качестве альтернативы, это может быть определено посредством алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), который выполняют с использованием программы Needleman пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16:276-277) (предпочтительно, версия 5.0.0 или более поздняя) (пакет программы GCG (Devereux, J. et al, Nucleic Acids Research 12:387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S.F. et al., JMOLEC BIOL 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994, and CARILLO et al. (1988) SIAM J Applied Math 48:1073). Например, гомология, сходство или идентичность может быть определена с использованием BLAST или ClustalW Национального центра биотехнологической информации (NCBI).
Гомология, сходство или идентичность полинуклеотидов или полипептидов может быть, например, определена путем сравнения информации о последовательности с использованием, например, компьютерной программы GAP, такой как Needleman et al. (1970), J Mol Biol. 48:443, описанной в работе Smith and Waterman, Adv. Appl. Math (1981) 2:482. Таким образом, программа GAP определяет гомологию, сходство или идентичность как значение, полученное делением числа сходных выровненных символов (то есть нуклеотидов или аминокислот) на общее количество символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать (1) унарную матрицу сравнения (содержащую значение 1 для идентичностей и 0 для не-идентичностей) и взвешенную матрицу сравнения из Gribskov et al. (1986), Nucl. Acids Res. 14:6745, как описано в Schwartz and Dayhoff, eds., Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, pp. 353-358 (1979) (или EDNAFULL матрицу замены (EMBOSS версия NCBI NUC4.4)); (2) штраф 3.0 за каждый пробел и дополнительный штраф 0.10 за каждый символ в каждом пробеле (или штраф за открытие пробела 10 и штраф за продление пробела 0.5); и (3) отсутствие штрафа за концевые пробелы.
При использовании здесь термин "рекомбинантный микроорганизм для продуцирования (9-фосфосерина (OPS)" может относиться к микроорганизму, который обладает природной слабой OPS-продуцирующей способностью, или к предложенному микроорганизму с OPS-продуцирующей способностью посредством естественной или искусственной генетической модификации родительского штамма, который не обладает OPS-продуцирующей способностью. В настоящем изобретении термин "рекомбинантный микроорганизм, продуцирующий OPS" может использоваться взаимозаменяемо с "микроорганизмом, имеющим OPS-продуцирующую способность".
Применительно к целям настоящего изобретения, в случае OPS-продуцирующего микроорганизма, количество продуцированного OPS может быть увеличено по сравнению с микроорганизмом дикого типа или микроорганизмом до модификации, так как увеличена активность белка mdtH. Это важно в том смысле, что способность к продуцированию OPS может быть увеличена путем усиления активности белка mdtH по настоящему изобретению, в то время как микроорганизмы дикого типа или микроорганизмы до модификации не могут продуцировать OPS или могут продуцировать только следовые количества, даже когда они способны продуцировать OPS.
При использовании здесь термин "улучшение" полипептидной активности означает, что активность полипептида увеличена по сравнению с его эндогенной активностью. Улучшение может использоваться взаимозаменяемо с такими терминами, как активация, повышение, сверхэкспрессия, увеличение и т.д. В частности, активация, улучшение, повышение регуляции, сверхэкспрессия и увеличение могут включать в себя оба случая - когда демонстрируется изначально отсутствующая активность или активность повышается по сравнению с эндогенной активностью или активностью до модификации. "Эндогенная активность" относится к активности конкретного полипептида, которой исходно обладал родительский штамм до трансформации или немодифицированный микроорганизм, когда признак изменяется в результате генетической модификации, вызванной естественными или искусственными факторами и может использоваться взаимозаменяемо с "активностью до модификации". "Улучшение", "активация", "сверхэкспрессия" или "увеличение" активности полипептида по сравнению с эндогенной активностью означает, что активность и/или концентрация (уровень экспрессии) полипептида увеличивается по сравнению активностью конкретного полипептида, которой исходно обладал родительским штамм до трансформации или немодифицированный микроорганизм.
Улучшение может быть достигнуто путем введения чужеродного полипептида или посредством повышения активности и/или концентрации (уровня экспрессии) эндогенного полипептида. Усиление активности полипептида может быть подтверждено увеличением уровня активности полипептида, уровня экспрессии или количества продукта, выделенного из полипептида.
Усиление активности полипептида может осуществляться различными способами, хорошо известными в данной области, и не ограничено при условии, что оно может усилить активность целевого полипептида по сравнению с активностью микроорганизма до модификации. В частности, можно использовать генетическую инженерию и/или белковую инженерию, хорошо известную специалистам в данной области, которая является обычным способом молекулярной биологии, но способ не ограничивается этим (e.g., Sitnicka et al. Functional Analysis of Genes. Advances in Cell Biology. 2010, Vol.2. 1-16; Sambrook et al. Molecular Cloning 2012, etc.).
В частности, усиление полипептида по настоящему изобретению может быть достигнуто посредством:
1) увеличения числа внутриклеточных копий полинуклеотида, кодирующего полипептид;
2) модификации регуляторной области экспрессии гена, кодирующего полипептид, на хромосоме (например, индукция модификации регуляторной области экспрессии, замена последовательности на последовательность, имеющую более сильную активность, или вставка последовательности, имеющей более сильную активность);
3) модификации нуклеотидной последовательности, кодирующей кодон инициации или 5'-UTR транскрипта гена, кодирующего полипептид;
4) модификации аминокислотной последовательности полипептида, приводящая к усилению активности полипептида;
5) модификации полинуклеотидной последовательности, кодирующей полипептид, приводящая к усилению активности полипептида (например модификация полинуклеотидной последовательности гена полипептида для кодирования полипептида, который модифицирован с повышением активности этого полипептида);
6) введения чужеродного полипептида, демонстрирующего указанную активность полипептида, или чужеродного полинуклеотида, кодирующего его;
7) оптимизации кодона полинуклеотида, кодирующего полипептид;
8) анализа третичной структуры полипептида и, посредством этого, выбора и модификации экспонированного сайта, или его химической модификации; или
9) комбинации двух или более методов, выбранных из вышеуказанных 1-8, но без конкретного ограничения ими.
Более конкретно:
1) способ увеличения внутриклеточного количества копий полинуклеотида, кодирующего полипептид, может быть выполнен путем введения вектора, который функционально связан с полинуклеотидом, кодирующим полипептид, и способен к репликации и функционированию независимо от клетки-хозяина, в клетку-хозяина. В качестве альтернативы, способ может быть достигнут посредством введения одной копии или двух копий полинуклеотидов, кодирующих полипептид, в хромосому клетки-хозяина. Введение в хромосому может быть выполнено посредством введения вектора, который способен вставлять полинуклеотид в хромосому клетки-хозяина, в клетку-хозяина, но без ограничения этим. Вектор является таким, как описано выше.
2) Способ замены регуляторной области экспрессии (или регуляторной последовательности экспрессии) гена, кодирующего полипептид на хромосоме, последовательностью, имеющей сильную активность, может быть выполнен, например, путем модификации последовательности посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации для дальнейшего повышения активности регуляторной области экспрессии или заменой последовательности последовательностью, имеющей сильную активность. Регуляторная область экспрессии может включать, но не ограничивается этим, промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосом, и последовательность, регулирующую завершение транскрипции и трансляции и т.д. В одном примере способ может включать замену исходного промотора сильным промотором, но без ограничения этим.
Примеры известного сильного промотора могут включать промоторы CJ1 - CJ7 (US 7662943 В2), промотор lac, промотор trp, промотор trc, промотор tac, lambda phage промотор PR, промотор PL, промотор tet, промотор gapA, промотор SPL7, промотор SPL13 (sm3) (US 10584338 В2), промотор O2 (US 10273491 В2), промотор tkt, промотор усе А, и т.д., но сильный промотор не ограничен этим.
3) Способ модификации нуклеотидной последовательности, кодирующей кодон инициации или 5'-UTR транскрипта гена, кодирующего полипептид, может быть выполнен, например, посредством замены нуклеотидной последовательности нуклеотидной последовательностью, кодирующей другой кодон инициации, имеющей более высокий уровень экспрессии полипептида по сравнению с эндогенным кодоном инициации, но не ограничивается этим.
4) и 5) Способы модификации аминокислотной последовательности или полинуклеотидной последовательности могут быть выполнены посредством индукции модификации последовательности в результате делеции, вставки, неконсервативной или консервативной замены в аминокислотной последовательности полипептида или полинуклеотидной последовательности, кодирующей полипептид, или их комбинация для усиления активности полипептида, или посредством замены последовательности аминокислотной последовательностью или полинуклеотидной последовательностью, модифицированной так, чтобы иметь более сильную активность, или аминокислотной последовательностью или полинуклеотидной последовательностью, модифицированной для усиления активности, но без ограничения этим. В частности, замена может быть выполнена путем вставки полинуклеотида в хромосому посредством гомологичной рекомбинации, но без ограничения этим. Вектор, используемый здесь, может включать селективный маркер для подтверждения вставки в хромосому. Селективный маркер является таким, как описано выше.
Способ 6) введения чужеродного полинуклеотида, демонстрирующего активность полипептида, может быть выполнен путем введения в клетку-хозяина чужеродного полинуклеотида, кодирующего полипептид, который проявляет такую же/аналогичную активность с полипептидом. Чужеродный полинуклеотид может использоваться без ограничения, независимо от его происхождения или последовательности, если он проявляет такую же/аналогичную активность с полипептидом. Введение может быть выполнено специалистами в данной области посредством выбора подходящего, известного в данной области способа трансформации, а экспрессия введенного полинуклеотида в клетку-хозяин позволяет продуцировать полипептид, тем самым увеличивая его активность.
Способ 7) оптимизации кодона полинуклеотида, кодирующего полипептид, может быть выполнен посредством оптимизации кодона эндогенного полинуклеотида для увеличения транскрипции или трансляции внутри клетки-хозяина или посредством оптимизации его кодонов, благодаря чему можно достичь оптимизированной транскрипции и трансляции чужеродного полинуклеотида в клетке-хозяине.
Способ 8) анализа третичной структуры полипептида и посредством этого выбора и модификации экспонируемого сайта или его химической модификации может быть выполнен, например, посредством сравнения информации о последовательности полипептида, анализируемой с помощью базы данных, в которой хранится информация о последовательности известных белков, для определения кандидатов эталонов белка в соответствии со степенью сходства последовательностей, и, таким образом, подтверждение структуры на основе этой информации, и посредством этого выбора и трансформации или модификации сайта, который следует модифицировать или химически модифицировать.
Такое улучшение активности полипептида может означать, что активность или концентрация соответствующего полипептида увеличивается по сравнению с активностью или концентрацией полипептида, экспрессируемого диким типом или микроорганизмом до модификации, или что количество продукта, полученного от полипептида, увеличивается, без ограничения этим.
При использовании здесь термин "вектор" может включать конструкцию ДНК, содержащую нуклеотидную последовательность полинуклеотида, кодирующего целевой полипептид, функционально связанную с участками регуляции экспрессии (или с последовательностью регуляции экспрессии), подходящими для экспрессии целевого полипептида в подходящем хозяине. Участки регуляции экспрессии могут включать промотор, способный инициировать транскрипцию, любую последовательность оператора для регуляции транскрипции, последовательность, кодирующую подходящий сайт связывания рибосомы на мРНК и последовательность для регулирования окончания транскрипции и трансляции. Будучи трансформированным в подходящую клетку-хозяина, вектор может реплицироваться или функционировать независимо от генома хозяина или может интегрироваться в геном.
Вектор, используемый в настоящем изобретении, конкретно не ограничен и можно использовать любой известный в данной области вектор. Примеры обычных векторов могут включать природную или рекомбинантную плазмиду, космиду, вирус и бактериофаг. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A и Charon21A. В качестве плазмидного вектора можно использовать тип pDZ, тип pBR, тип pUC, тип pBluescriptII, тип pGEM, тип pTZ, тип pCL и тип рЕТ. В частности, можно использовать векторы pDZ, pDC, pDCM2, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118 и pCC1BAC.
Например, полинуклеотид, кодирующий целевой полипептид, может быть встроен в хромосому с использованием вектора для хромосомной вставки. Встраивание полинуклеотида в хромосому может быть выполнено с помощью любого способа, известного в данной области, например гомологичной рекомбинации, без ограничения ею. Полинуклеотид может дополнительно включать селективный маркер для подтверждения хромосомной вставки. Селективный маркер используют для отбора клеток, которые трансформированы вектором, то есть для подтверждения вставки нужной молекулы нуклеиновой кислоты и примеры селективных маркеров могут включать маркеры, обеспечивающие селектируемые фенотипы, такие как устойчивость к лекарственному средству, потребность в питательных веществах, устойчивость к цитотоксическим агентам или экспрессия поверхностного полипептида. Только клетки, экспрессирующие селективный маркер, способны выживать или проявлять разные фенотипы в среде, обработанной селективным агентом, и, таким образом могут быть отобраны трансформированные клетки.
При использовании здесь термин "трансформация" относится к процессу введения вектора, включающего полинуклеотид, кодирующий целевой полипептид, в клетку-хозяина или микроорганизм таким образом, что полипептид, кодируемый полинуклеотидом, экспрессируется в клетке-хозяине. Трансформированный полинуклеотид может быть либо в форме, встроенной в хромосому клетки-хозяина, либо в форме, расположенной вне хромосомы, при условии что белок экспрессируется в клетке-хозяине. Кроме того, полинуклеотид включает ДНК и/или РНК, кодирующую целевой полипептид. Полинуклеотид может быть введен в клетку-хозяина в любой форме, при условии, что этот полинуклеотид введен в клетку-хозяина и в ней экспрессируется полипептид. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генную конструкцию, включающую все основные элементы, необходимые для самовоспроизведения. Экспрессионная кассета, как правило, может включать промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, сайт связывания рибосомы и сигнал терминации трансляции. Экспрессионная кассета может находиться в форме самовоспроизводимого вектора экспрессии. Также, полинуклеотид может быть введен в клетку-хозяина в своей первоначальной форме и функционально связан с последовательностью, необходимой для экспрессии в клетке-хозяине, без ограничения этим.
Кроме того, при использовании здесь термин "функционально связанный" относится к функциональной связи между последовательностью промотора, которая позволяет осуществлять инициацию и опосредование транскрипции полинуклеотида, кодирующего целевой вариант по настоящему изобретению, и полинуклеотидной последовательностью.
В частности, рекомбинантный микроорганизм по настоящему изобретению может быть микроорганизмом с введенным геном/полинуклеотидом, кодирующим белок mdtH; микроорганизмом, в котором увеличено число внутриклеточных копий полинуклеотида, кодирующего белок mdtH; микроорганизмом, трансформированным вектором, содержащим полинуклеотид, кодирующий белок mdtH; микроорганизмом, в котором последовательность регуляции экспрессии гена в хромосоме, кодирующей белок mdtH, заменена последовательностью с более сильной активностью; или микроорганизмом, у которого промотор, кодирующий белок mdtH, в хромосоме заменен промотором с сильной активностью.
Более конкретно, ген, кодирующий белок mdtH, может быть геном mdtH. Кроме того, полинуклеотид, кодирующий белок mdtH может быть полинуклеотидом, включающим последовательность нуклеотидов SEQ ID NO: 2. Последовательности SEQ ID NO: 1 и SEQ ID NO: 2 могут быть получены из известной базы данных GenBank NCBI.
Белок/полипептид, включающий аминокислотную последовательность с определенным номером последовательности или аминокислотную последовательность, описанную при помощи конкретного номера последовательности, может быть белком/полипептидом, имеющим аминокислотную последовательность с конкретным номером последовательности или аминокислотную последовательность, описанную при помощи конкретного номера последовательности, или может быть белком/полипептидом, состоящим или по существу состоящим из аминокислотной последовательности с конкретным номером последовательности или аминокислотной последовательности, описанной при помощи конкретного номера последовательности. Кроме того, полинуклеотид, включающий нуклеотидную последовательность с конкретным номером последовательности или нуклеотидную последовательность, описанную при помощи конкретного номера последовательности, может быть полинуклеотидом, имеющим нуклеотидную последовательность с конкретным номером последовательности или нуклеотидной последовательностью, описанной при помощи конкретного номера последовательности, или может быть полинуклеотидом, состоящим или по существу состоящим из нуклеотидной последовательности с конкретным номером последовательности или нуклеотидной последовательности, описанной при помощи конкретного номера последовательности.
Микроорганизм по настоящему изобретению не ограничен его типом при условии, что он может продуцировать OPS, и может быть любым прокариотическим или эукариотическим микроорганизмом, в частности прокариотическим микроорганизмом. Примеры прокариотического микроорганизма могут включать штаммы микроорганизмов, принадлежащие к роду Escherichia, роду Erwinia, роду Serratia, роду Providencia, роду Corynebacterium и роду Brevibacterium, в частности микроорганизма, принадлежащего к роду Escherichia, и более конкретно Escherichia coli, но не ограничиваются ими. В частности, в случае микроорганизма, принадлежащего к роду Escherichia или Corynebacterium, может продуцироваться OPS и L-серин (Ahmed Zahoor, Computational and structural biotechnology journal, Vol. 3, 2012 October; Wendisch, V.F. et al., Curr Opin Microbiol. 2006 Jun; 9(3):268-74; Peters-Wendisch, P. et al., Appl Environ Microbiol 2005 Nov; 7 1(ll): 7 139-44.).
Рекомбинантный микроорганизм по настоящему изобретению может быть таким, в котором активность фосфосеринфосфатазы (SerB) дополнительно ослаблена по сравнению с его эндогенной активностью.
SerB по настоящему изобретению обладает активностью превращения OPS в L-серин, и, следовательно, микроорганизм, модифицированный так, что активность SerB ослаблена, обладает свойством накапливать OPS в себе, и поэтому является полезным для продуцирования OPS. SerB по настоящему изобретению может быть белком, включающим аминокислотную последовательность, описанную SEQ ID NO: 3, но не ограничивается ею. Кроме того, SerB может включать аминокислотную последовательность, имеющую идентичность 80% или более, конкретно 90% или более, более конкретно 95% или более, или еще более конкретно 99% или более с аминокислотной последовательностью, описанной SEQ ID NO: 3, при условии, что он демонстрирует SerB активность, но не ограничивается этим. Кроме того, полинуклеотид, кодирующий SerB, может иметь нуклеотидную последовательность, кодирующую аминокислоту, описанную SEQ ID NO: 3. Различные модификации могут быть сделаны в кодирующем участке полинуклеотида в пределах, не изменяющих аминокислотную последовательность белка, из-за вырожденности кодона или с учетом кодона, предпочтительного для организма, в котором белок должен быть экспрессирован. Полинуклеотид, кодирующий SerB по настоящему изобретению, может, например, включать нуклеотидную последовательность SEQ ID NO: 4, или нуклеотидную последовательность, имеющую идентичность, составляющую 80% или более, конкретно 90% или более, более конкретно 95% или более, или еще более конкретно 99% или более с нуклеотидной последовательностью SEQ ID NO: 4, но не ограничивается этим.
При использовании здесь термин "ослабление" полипептида предполагает понятие, включающее как снижение, так и отсутствие активности по сравнению с изначально присущей ему активностью. Ослабление может быть использовано взаимозаменяемо с инактивацией, дефицитом, понижением, уменьшением, снижением, истощением или тому подобным.
Ослабление может также включать: случай, когда активность самого полипептида снижена или ликвидирована по сравнению с активностью самого полипептида, которой обладал исходный микроорганизм, вследствие мутации или т.п. полинуклеотида, кодирующего полипептид; случай, где активность и/или концентрация (уровень экспрессии) всех полипептидов в клетке ниже по сравнению с нативным штаммом вследствие ингибирования экспрессии гена полинуклеотида, кодирующего полипептид, или ингибирования трансляции в полипептид; случай, когда экспрессия полинуклеотида не осуществляется; и/или случай, когда полипептид не обладает активностью, несмотря на экспрессию полинуклеотида. "Эндогенная активность" относится к активности конкретного полипептида, которой исходно обладает родительский штамм до модификации, или дикий тип, или немодифицированный микроорганизм, когда трансформация происходит из-за генетической вариации, вызванной природными или искусственными факторами. Этот термин может использоваться взаимозаменяемо с термином "активность до модификации". "Ослабление", "инактивация", "недостаток", "уменьшение", "негативная регуляция", "снижение" или "истощение" активности полипептида по сравнению с эндогенной активностью означает, что активность полипептида снижена по сравнению с активностью конкретного полипептида, которой исходно обладает родительский штамм до модификации или немодифицированный микроорганизм.
Ослабление активности полипептида может быть достигнуто любым способом, известным в данной области техники, но не ограничивается этим, и может быть достигнуто путем применения различных способов, хорошо известных в данной области техники (e.g., Nakashima N et al., Bacterial cellular engineering by genome editing and gene silencing. Int J Mol Sci. 2014; 15(2):2773-2793, Sambrook et al., Molecular Cloning 2012, etc.).
В частности, ослабление полипептида по настоящему изобретению может представлять:
1) делецию части или всего гена, кодирующего полипептид;
2) модификацию области контроля экспрессии (или последовательности контроля экспрессии) для уменьшения экспрессии гена, кодирующего полипептид;
3) модификацию аминокислотной последовательности, составляющей полипептид, чтобы устранить или ослабить активность полипептида (например делецию/замену/добавление по меньшей мере одной аминокислоты в аминокислотной последовательности).
4) модификацию последовательности генов, кодирующей полинуклеотид, чтобы устранить или ослабить активность полипептида (например удаление/замещение/добавление по меньшей мере одного нуклеотида на нуклеотидную последовательность гена полипептида, чтобы кодировать полипептид, модифицированный для устранения или ослабления активности полипептида);
5) модификацию нуклеотидной последовательности, кодирующей кодон инициации или 5'-UTR область транскрипта гена, кодирующего полипептид;
6) введение антисмыслового олигонуклеотида (например антисмысловой РНК), комплементарно связывающегося с транскриптом гена, кодирующего полипептид;
7) добавление последовательности, комплементарной последовательности Шайна-Далгарно гена, кодирующего полипептид выше последовательности Шайна-Далгарно, чтобы сформировать вторичную структуру, которая делает прикрепление рибосом невозможным;
8) добавление промотора обратной транскрипции к 3'-концу открытой рамки считывания (ORF) последовательности гена, кодирующего полипептид (инженерия обратной транскрипции, RTE); или
9) комбинация двух или более, выбранных из пунктов (1)-(8), без особого ограничения ими.
Например, они описаны следующим образом.
Делеция части или всего гена, кодирующего полипептид в пункте (1) может представлять собой исключение всего полинуклеотида, кодирующего эндогенный целевой белок в хромосоме, замену полинуклеотида с делецией некоторых нуклеотидов или замену маркерным геном.
Модификация области, контролирующей экспрессию (или последовательности, контролирующей экспрессию) в пункте (2) может представлять собой мутацию области, контролирующей экспрессию (или последовательности, контролирующей экспрессию) посредством удаления, вставки, неконсервативной или консервативной замены или их комбинации, или замену последовательностью, имеющей более слабую активность. Область контроля экспрессии включает промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, и последовательность контроля за терминацией транскрипции и трансляции, но не ограничена этим.
Модификация нуклеотидной последовательности, кодирующей кодон инициации или 5'-UTR-область транскрипта гена, кодирующего полипептид в пункте (3), может заключаться, например, в замещении нуклеотидной последовательностью, кодирующей, вместо эндогенного кодона инициации, другой кодон инициации, имеющий более низкий уровень экспрессии полипептида, но не ограничивается этим.
Модификация аминокислотной последовательности или полинуклеотидной последовательности в пунктах (4) и (5) может представлять собой модификацию последовательности посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации в аминокислотной последовательности полипептида или в полинуклеотидной последовательности, кодирующей этот полипептид, чтобы ослабить активность полипептида, или в замене аминокислотной последовательности или полинуклеотидной последовательности модифицированной, чтобы иметь более слабую активность, или аминокислотной последовательностью или полинуклеотидной последовательностью, модифицированной так, чтобы не иметь активности, но не ограничивается этим. Например, экспрессия гена может быть ингибирована или ослаблена путем введения мутации в полинуклеотидную последовательность для создания терминирующего кодона.
Введение антисмыслового олигонуклеотида (например антисмысловой РНК), комплементарно связывающегося с транскриптом гена, кодирующего полипептид в пункте (6) может быть, например, упомянут в литературе (Weintraub, Н. et al., Antisense-RNA as a Genetics, Vol. 1(1) 1986).
Добавление последовательности, которая комплементарна последовательности Шайна-Далгарно гена, кодирующего полипептид, выше последовательности Шайна-Далгарно так, чтобы создать вторичную структуру, которая делает невозможным присоединение рибосомы в пункте (7), может сделать невозможной трансляцию мРНК или может уменьшить ее уровень.
Добавление промотора обратной транскрипции к 3'-концу открытой рамки считывания (ORF) последовательности гена, кодирующего полипептид (инженерия обратной транскрипции, RTE) в пункте (8) может сделать антисмысловой нуклеотид комплементарным транскрипту гена, кодирующего полипептид, тем самым ослабляя активность полипептида.
Модификация части или всего полинуклеотида в микроорганизме по настоящему изобретению может быть вызвана: (а) редактированием генома с использованием гомологичной рекомбинации или инженерной нуклеазы (например CRISPR-Cas9) с использованием вектора для встраивания в хромосому в микроорганизме и/или (б) обработкой светом, таким как ультрафиолетовый свет и радиация, и/или химическим агентами, но без ограничения этим. Способ модификации части или всего гена может включать способ с использованием технологии рекомбинантной ДНК. Например, нуклеотидная последовательность или вектор, содержащий(ая) нуклеотидную последовательность, гомологичную целевому гену, может быть введен(а) в микроорганизм, чтобы вызвать гомологичную рекомбинацию, что приводит к удалению части или всего гена. Нуклеотидная последовательность или вектор, которые должны быть введены, могут включать селективный маркер, но не ограничены этим.
Кроме того, рекомбинантный микроорганизм по настоящему изобретения может быть таким, в котором активность любой из фосфоглицератдегидрогеназы (SerA), фосфосеринаминотрансферазы (SerC) или их комбинации дополнительно повышена по сравнению с его эндогенной активностью.
SerA по настоящему изобретению представляет собой белок, обладающий активностью превращения 3-фосфоглицерата в 3-фосфо-гидроксипируват. SerC по настоящему изобретению представляет собой белок, имеющий активность превращения 3-фосфо-гидроксипирувата в OPS. Соответственно, любой микроорганизм с повышенной активностью SerA и/или SerC может быть эффективно использован в качестве OPS-продуцирующего микроорганизма.
SerA по настоящему изобретению может быть белком, включающим аминокислотную последовательность SEQ ID NO: 5 или 6. Аминокислотная последовательность SEQ ID NO: 5 представляет собой последовательность дикого типа SerA, а аминокислотная последовательность SEQ ID NO: 6 представляет собой последовательность варианта SerA, где осуществляется ингибирование по типу обратной связи по серину. Кроме того, SerA может включать аминокислотную последовательность, имеющую идентичность 80% или более, конкретно 90% или более, более конкретно 95% или более, или еще более конкретно 99% или более с аминокислотной последовательностью, описанной SEQ ID NO: 5 или 6, при условии, что он демонстрирует активность дикого типа SerA или активность варианта SerA, у которого осуществляется ингибирование по типу обратной связи по серину, но не ограничивается этим. Варианты SerA, у которых осуществляется ингибирование по типу обратной связи по серину, относятся к таким белкам, у которых модификация введена в SerA-кодирующий ген посредством вставки, замены и т.д., тем самым поддерживается активность от ингибирования по типу обратной связи серином или глицином, или обеспечивается их повышенная активность, и варианты SerA, у которых осуществляется ингибирование по типу обратной связи по серину, уже хорошо известны (Grant, G.A. et al., J. Biol. Chem., 39:5357-5361, 1999; Grant, G.A. et al., Biochem., 39:7316-7319, 2000; Grant, G.A. et al., J. Biol. Chem., 276:17844-17850, 2001; Peters-Wendisch, P. et al., Appl. Microbiol. Biotechnol, 60:37-441, 2002; EP 0943687 B).
Кроме того, полинуклеотид, кодирующий SerA дикого типа или вариант SerA, у которого осуществляется ингибирование по типу обратной связи по серину, могут включать нуклеотидную последовательность, кодирующую любую аминокислотную последовательность, описанную SEQ ID NO: 5 или 6, но не ограничиваются этим. Полинуклеотидная последовательность, кодирующая SerA дикого типа или вариант SerA, в котором осуществляется ингибирование по типу обратной связи по серину, могут претерпевать различные модификации в кодирующей области при условии, что не меняется аминокислотная последовательность полипептида, из-за вырожденности кодона или с учетом кодонов, предпочтительных в организме, в котором должен экспрессироваться полипептид. Полинуклеотид, кодирующий SerA дикого типа или вариант SerA, в котором осуществляется ингибирование по типу обратной связи по серину, может быть, например, полинуклеотидом, включающим нуклеотидную последовательность SEQ ID NO: 7 или 8, или может быть полинуклеотидом, включающим нуклеотидную последовательность, имеющую гомологию 80% или более, конкретно 90% или более, более конкретно 95% или более, или даже более конкретно 99% или более с нуклеотидной последовательностью SEQ ID NO: 7 или 8, но не ограничивается этим.
SerC может быть, например, белком, включающим аминокислотную последовательность, описанную SEQ ID NO: 9, но не ограничивается этим. Кроме того, SerC может включать аминокислотную последовательность, имеющую идентичность, составляющую 80% или более, конкретно 90% или более, более конкретно 95% или более, или даже более конкретно 99% или более с аминокислотной последовательностью, описанной SEQ ID NO: 9, при условии, что он демонстрирует активность SerC, но не ограничивается этим.
Кроме того, полинуклеотид, кодирующий SerC, может включать нуклеотидную последовательность, кодирующую аминокислотную последовательность, описанную SEQ ID NO: 9. Полинуклеотид, кодирующий SerC, может претерпевать различные модификации в кодирующей области при условии, что не меняется аминокислотная последовательность полипептида, из-за вырожденности кодона или принимая во внимание кодоны, предпочтительные в организме, в котором должен экспрессироваться полипептид. Полинуклеотид, кодирующий SerC, может включать, например, нуклеотидную последовательность SEQ ID NO: 10, или нуклеотидную последовательность, имеющую гомологию 80% или более, в частности 90% или более, более конкретно 95% или более, или даже более конкретно 99% или более с аминокислотной последовательностью SEQ ID NO: 10, но не ограничивается этим.
При использовании здесь термин "повышение по сравнению с его эндогенной активностью" и способ повышения являются такими же, как описано выше.
Кроме того, рекомбинантный микроорганизм по настоящему изобретению может быть микроорганизмом, у которого его способность вводить OPS в клетку или разлагать OPS дополнительно ослаблена.
Что касается содержимого OPS-продуцирующего микроорганизма, то описания изобретений в европейском патенте 2444481 или патентной публикации США 2012-0190081 могут быть использованы в качестве ссылок для настоящего изобретения в дополнение к описанным выше.
В другом аспекте настоящее изобретение предложен способ продуцирования О-фосфосерина, включающий культивирование в среде рекомбинантного микроорганизма, продуцирующего О-фосфосерин, у которого активность белка mdtH в среде повышена по сравнению с его эндогенной активностью.
Белок mdtH, эндогенная активность, улучшение, О-фосфосерин и микроорганизм являются такими, как описано выше.
При использовании здесь термин "культивирование" означает, что микроорганизм выращивают в соответствующим образом контролируемых условиях окружающей среды. Процесс культивирования по настоящему изобретению может быть выполнен в подходящей культуральной среде и при условиях культивирования, известных в данной области. Такой процесс культивирования может быть легко скорректирован для использования специалистами в данной области в соответствии с выбранным штаммом. В частности, культивирование может быть периодическим культивированием, непрерывным культивированием и культивированием с подпиткой, но без ограничения этим.
При культивировании рекомбинантного микроорганизма, у которого активность SerB ослаблена по сравнению с его эндогенной активностью, среда может дополнительно содержать глицин или серии, поскольку индуцируются потребность микроорганизма в серине. Глицин может быть представлен в форме очищенного глицина, глицин-содержащего дрожжевого экстракта или триптона. Концентрация глицина, содержащегося в среде, обычно составляет от 0,1 г/л до 10 г/л и, в частности, от 0,5 г/л до 3 г/л. Кроме того, серии может быть предоставлен в виде очищенного серина, серин-содержащего дрожжевого экстракта или триптона. Концентрация серина, содержащегося в среде, обычно составляет от 0,1 г/л до 5 г/л и, в частности, от 0,1 г/л до 1 г/л.
Примеры источника углерода, содержащегося в среде, могут включать сахариды и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза; масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол; и органические кислоты, такие как уксусная кислота. Эти источники углерода могут быть использованы по отдельности или в комбинации, но не ограничены этим.
Примеры источника азота, содержащегося в среде, могут включать в себя органические источники азота, такие как пептон, дрожжевой экстракт, мясной соус, солодовый экстракт, кукурузный экстракт и мука из фасоли; и неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота могут быть использованы по отдельности или в комбинации, но не ограничены этим.
Примеры источника фосфора, содержащегося в среде, могут включать дигидрофосфат калия, дикалия гидрофосфат, и соответствующие натрий-содержащие соли, но не ограничены этим.
Кроме того, культуральная среда может включать соли металлов, такие как сульфат магния или сульфат железа, и может также содержать аминокислоты, витамины и соответствующие предшественники. Такие культуральные среды или предшественники могут быть добавлены к культуре в случае периодической культуры или непрерывной культуры, но не ограничена этим.
рН культуры может быть скорректирован путем добавления соединений, таких как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота при культивировании подходящим образом. Кроме того, образование пузырьков при культивировании может быть предупреждено с использованием пеногасителя, такого как сложный полигликолевый эфир жирных кислот. Кроме того, газообразный кислород или газ, содержащий кислород, может быть введен в культуру, чтобы поддерживать аэробные условия культивирования; или анаэробные или микроаэробные условия могут сохранены без введения газа или путем введения азота, водорода или углекислого газа. Температура культуры может быть в диапазоне от 25°С до 40°С и, в частности, от 30°С до 35°С. Культивирование может продолжаться, пока не будет получено необходимое вещество, и может, в частности, выполняться в течение от 10 часов до 100 часов, но не ограничено этими иллюстративными примерами.
Способ продуцирования О-фосфосерина по настоящему изобретению может, дополнительно включать получение микроорганизма по настоящему изобретению, получение культуральной среды для культивирования штамма или любую их комбинацию (независимо от порядка, в любом порядке), например, до стадии культивирования.
Способ получения О-фосфосерина по настоящему изобретению может дополнительно включать извлечение О-фосфосерина из культуральной среды после культивирования (культуральной среды, в которой выполняли культивирование) или из микроорганизма по настоящему изобретению. Стадия извлечения также может быть включена после стадии культивирования.
Извлечение может заключаться в сборе необходимого О-фосфосерина с использованием способа культивирования по настоящему изобретению, например подходящего способа, известного в данной области, такого как периодическое, непрерывное культивирование или культивирование с подпиткой. Например, могут быть использованы центрифугирование, фильтрация, обработка агентом, осаждающим белки (высаливание), экстракция, ультразвуковая дезинтеграция, ультрафильтрация, диализ, различные хроматографические методы, такие как молекулярно-ситовая хроматография (гель-проникающая), адсорбционная хроматография, ионообменная хроматография и аффинная хроматография, высокоэффективная жидкостная хроматография (ВЭЖХ), и любая их комбинация, и необходимый О-фосфосерин может быть выделен из культуральной среды или микроорганизма с использованием подходящего способа, хорошо известного в данной области.
Кроме того, способ получения О-фосфосерина по настоящему изобретению может дополнительно включать очистку О-фосфосерина. Стадия очистки может быть выполнена с использованием подходящего способа, хорошо известного в данной области. В одном воплощении, когда способ получения О-фосфосерина по настоящему изобретению включает стадии и выделения и очистки, эти стадии выделения и очистки могут быть выполнены непрерывно или с перерывами, независимо от порядка, или могут быть выполнены одновременно или в виде интегрированной стадии, без ограничения этим.
В способе по настоящему изобретению вариант, полинуклеотид, вектор, микроорганизм и тому подобное являются такими, как описано выше.
В еще одном аспекте настоящего изобретения предложен способ получения цистеина или его производного, включающий:
а) получение О-фосфосерина или среды, содержащей О-фосфосерин, путем культивирования в среде рекомбинантного микроорганизма, продуцирующего О-фосфосерин, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью; и
б) взаимодействие О-фосфосерина или среды, содержащей О-фосфосерин, полученных на стадии (а) с сульфидом в присутствии О-фосфосеринсульфгидрилаз ы (OPSS) или содержащего ее микроорганизма.
При использовании здесь "включать/включающий" конкретный белок в отношении микроорганизма означает состояние, в котором конкретный интересующий белок введен в микроорганизм или экспрессируется в микроорганизме.
Белок mdtH, эндогенная активность, усиление, О-фосфосерин и микроорганизм являются такими, как описано выше.
При использовании здесь термин "производное" относится к аналогичным соединениям, полученным посредством химической модификации части любого соединения. Этот термин обычно относится к соединениям, в которых атом водорода или определенная группа атомов заменены другим атомом или группой атомов.
При использовании здесь термин "производное цистеина" относится к соединениям, в которых атом водорода или конкретная группа атомов цистеина заменена другим атомом или другой группой атомов. Например, производные цистеина могут иметь форму, в которой атом азота аминогруппы (-NH2) или атом серы тиольной группы (-SH) в цистеине имеет другой атом или группу атомов, присоединенную к нему, и примеры производных цистеина могут включать NAC (N-ацетилцистеин), SCMC (S-карбоксиметилцистеин), Boc-Cys(Me)-OH, (R)-S-(2-амино-2-карбоксиэтил)-L-гомоцистеин, (R)-2-амино-3-сульфо-пропионовую кислоту, D-2-амино-4-(этилтио)масляную кислоту, 3-сульфино-L-аланин, Fmoc-Cys(Boc-метил)-OH, селено-L-цистин, S-(2-тиазолил)-L-цистеин, S-(2-тиенил)-L-цистеин, S-(4-толил)-L-цистеин и т.д., но не ограничиваются ими.
При условии, что цистеин производится в соответствии с способом по настоящему изобретению, цистеин может быть превращен в различные производные цистеина, и превращение в производные цистеина может быть легко выполнено посредством способов, хорошо известных в данной области.
В частности, способ получения производных цистеина может дополнительно включать превращение цистеина, полученного на стадии (б) в производное цистеина. Например, цистеин может быть синтезирован в виде N-ацетилцистеина (NAC) посредством взаимодействия с ацетилирующим агентом, или он может быть синтезирован в виде S-карбоксиметилцистеина (SCMC) посредством взаимодействия с галогензамещенной уксусной кислотой в щелочных условиях, но способ этим не ограничивается.
Производные цистеина используют, главным образом, в качестве фармацевтических веществ для противокашлевых средств, средств, облегчающих кашель, и терапевтических агентов для бронхита, бронхиальной астмы, ларингофарингита и т.д., но не ограничиваются этим.
При использовании здесь термин "O-фосфосеринсульфгидрилаза (OPSS)" относится к ферменту, который катализирует реакцию, посредством которой OPS превращается в цистеин путем передачи тиольной группы (группа SH) на OPS. Фермент мог быть впервые найден в Aeropyrum pernix, Mycobacterium tuberculosis, Mycobacterium smegmatis and Trichomonas vaginalis (Mino, K. and Ishikawa, K., FEBS Letters, 551:133-138, 2003; Bums, K.E. et al.,J.Am. Chem. Soc., 127:11602-11603, 2005). Кроме того, OPSS может включать не только белки OPSS дикого типа, но также варианты белков, которые включают делецию, замену или добавление в части последовательности в полинуклеотидной последовательности, кодирующей OPSS, которые демонстрируют активность, равную или превышающую биологическую активность белков OPSS дикого типа, и может также включать все белки OPSS, раскрытые в европейском патенте 2444481 и патенте US 9127324, и варианты этих белков.
Сульфид может быть любым сульфидом, представленным не только в твердой форме, обычно используемой в данной области, но и в жидкой или газообразной форме из-за разницы в рН, давлении и растворимости, и, таким образом, может быть превращен в тиольную (SH) группу в форме сульфида (S2) или тиосульфата (S2O32-). В частности, сульфид может включать Na2S, NaSH, H2S, (NH4)2S и Na2S2O3, которые могут обеспечивать тиольную группу для OPS, но не ограничен ими. В данном взаимодействии одна тиольная группа предоставляется одной реакционной группе OPS с получением одного цистеина или его производного. В этом взаимодействии сульфид, в частности, добавляют в количестве от 0,1 до 3 молярных эквивалентов, и, в частности, от 1 до 2 молярных эквивалентов на основе молярной концентрации OPS, но без ограничения этим.
Кроме того, способ по настоящему изобретению может дополнительно включать стадию извлечения цистеина, полученного на вышеуказанной стадии взаимодействия. В частности, необходимый цистеин может быть выделен путем разделения и очистки реакционного раствора с использованием подходящей реакции, известной в данной области.
В еще одном аспекте настоящего изобретения предложено применение рекомбинантного микроорганизма, продуцирующего О-фосфосерин, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью, для получения О-фосфосерина, цистеина или производных цистеина.
В еще одном аспекте настоящего изобретения предложено применение для экспорта О-фосфосерина белка mdtH из микроорганизма.
Белок mdtH, эндогенная активность, улучшение, О-фосфосерин, цистеин, производные цистеина и микроорганизм являются таким, как описано выше.
Способ осуществления изобретения
Ниже настоящее изобретение будет описано более подробно с помощью Примеров. Однако специалистам в области, к которой принадлежит данное изобретение, понятно, что эти Примеры представлены только для иллюстративных целей и что не предполагается ограничивать объем изобретения этими Примерами.
Пример 1: Идентификация мембранных белков mdtH
Для того, чтобы идентифицировать мембранные белки Е. coli, вовлеченные в экспорт OPS, выполняли скрининговый анализ с использованием библиотеки геномной ДНК Escherichia coli W3110 (АТСС27325), штамма Е. coli дикого типа.
Чтобы установить условия, в которых рост Е. coli ингибировался OPS, был подготовлен референсный штамм для продуцирования OPS. Референсный штамм для скрининга представлял собой OPS-продуцирующий штамм, мутированный так, чтобы ослабить активность эндогенной фосфосеринфосфатазы (serB) в W3110, и был назван СА07-0012 (KCCM11212P, европейский патент №2444481; публикация патентной заявки США 2012-0190081).
СА07-0012 культивировали в среде, содержащей OPS, чтобы установить оптимальные условия скрининга, демонтрирующие ингибирование роста. Плазмиду геномной библиотеки W3110 трансформировали в СА07-0012 посредством электропорации (van der Rest et al. 1999) и отбирали колонии, у которых наблюдалось ингибирование роста в условиях среды с добавлением избытка OPS. Из отобранных колоний были получены плазмиды, и нуклеотидные последовательности анализировали с помощью методов секвенирования. Таким образом, были идентифицированы два типа мембранных белков Е. coli, вовлеченных в ингибирование роста в условиях с добавкой избытка OPS.
В результате был идентифицирован ген, кодирующий мембранный белок Е. coli (SEQ ID NO: 1), как переносчик mdtH, принадлежащий к суперсемейству главных фасилитаторов (MFS).
Пример 2: Получение векторов сверхэкспрессии mdtH
Когда mdtH, который вовлечен в осуществление ингибирования роста, вызванного OPS, был усилен в каждом из OPS-продуцирующих штаммов, для каждого гена получали векторы сверхэкспрессии для подтверждения, была ли улучшена OPS-экспортирующая способность. Кроме того, при усилении yhhS, который является экспортером фосфосерина, в OPS-продуцирующих штаммах, было подтверждено увеличение концентрации OPS (WO 2016/024771 A1) и, соответственно, это может быть использовано в качестве положительного контроля. Кроме того, мембранный белок yfaV MFS транспортер из E. coli, принадлежащий у суперсемейству главных фасилитаторов (MFS), был также оценен таким же образом, как mdtH. Фрагмент ДНК, кодирующий mdtH, был получен посредством ПЦР с использованием геномной ДНК W3110 в качестве матрицы (SEQ ID NO: 2). Последовательности праймеров, используемые для получения векторов сверхэкспрессии для гена каждого мембранного белка, показаны в Таблице 1 ниже.

Для того чтобы получить pCL_Ptrc-yhhS, использовали pCL_Ptrc-gfp (WO 2016/024771 A1) в качестве матрицы, и ПЦР выполняли с использованием SEQ ID NO: 11 и SEQ ID NO: 12 с получением фрагментов Ptrc DNA. Фрагменты yhhS ДНК получали посредством ПЦР с использованием SEQ ID NO: 13 и SEQ ID NO: 14 на основе W3110 в качестве матрицы. Амплифицированные фрагменты подвергали IST с вектором pCL1920, обработанным рестрикционными ферментами XbaI и HindIII, с получением посредством этого pCL_Ptrc-yhhS. IST (Gibson, D.G. et al., NATURE METHODS, Vol. 6 No. 5, MAY 2009, NEBuilder HiFi DNA Assembly Master Mix) относится к способу клонирования с использованием способа сборки Гибсона и ниже все время упоминается как IST.
Для получения pCL_Ptrc-mdtH, также использовали pCL_Ptrc-gfp в качестве матрицы, и фрагменты Ptrc DNA были получены с использованием SEQ ID NOS: 15 и 16. Фрагменты ДНК mdtH были получены с использованием SEQ ID NO: 17 и SEQ ID NO: 18 на основе W3110 в качестве матрицы, и pCL_Ptrc-mdtH был получен посредством IST таким же образом, как pCL_Ptrc-yhhS.
pCL_Ptrc-yfaV был также клонирован таким же образом, как при получении двух плазмид, и фрагменты Ptrc DNA были получены с использованием pCL_Ptrc-gfp в качестве матрицы, и фрагменты ДНК yfaV были получены с использованием W3110 в качестве матрицы. ПЦР была выполнена с использованием SEQ ID NO: 19 и SEQ ID NO: 20 для Ptrc и SEQ ID NO: 21 и SEQ ID NO: 22 для yfaV В качестве праймеров.
Пример 3: Получение mdtH MFS транспортер-усиленных штаммов и оценка их способности продуцировать OPS
3-1. Получение mdtH MFS транспортер-усиленных штаммов с использованием СА07-0012 и оценка их способности продуцировать OPS
Были получены штаммы, где три типа плазмид, полученных в Примере 2, были введены в OPS-продуцирующий штамм СА07-0012 и оценивали их способность продуцировать OPS.
Каждый из штаммов высевали на твердую среду LB и затем культивировали в инкубаторе при 33°С в течение ночи. Штаммы, культивируемые на твердой LB среде в течение ночи, инокулировали в 25 мл титрованной среды, показанной в Таблице 2 ниже и затем культивировали в инкубаторе при 33°С со скоростью 200 об/мин в течение 48 часов. Их OPS-продуцирующая способность показана в Таблице 3.
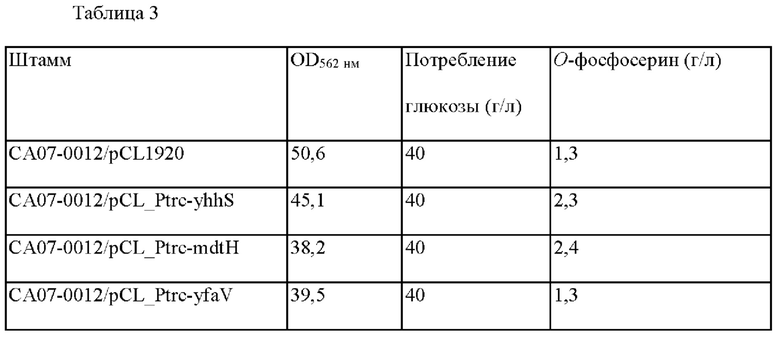
Как видно из Таблицы 3, среди случаев, в которых ген мембранного белка Е. coli был дополнительно введен в происходящий от Е. coli штамм СА07-0012, производство OPS было увеличено у штаммов с yhhS и mdtH, усиленными по сравнению с СА07-0012. В частности, было подтверждено, что концентрация OPS увеличилась приблизительно в 2-раза в mdtH-усиленном штамме по настоящему изобретению. Напротив, в случае yfaV-усиленного штамма, который представляет собой группу сравнения, продуцирование OPS не увеличивалось.
Соответственно, CA07-0012/pCL_Ptrc-mdtH был назван СА07-0354. Штамм СА07-0354 был депонирован в Корейском Центре Культур Микроорганизмов (KCCM), согласно Будапештскому договору, 28 августа 2020 года с регистрационным номером KCCM12781P.
3-2. Получение mdtH MFS транспортер-усиленных штаммов с использованием serA- и serC-усиленных штаммов и оценка их способности продуцировать OPS
СА07-0022 (KCCM11103P, патент US 9689009), OPS-продуцирующий штамм с повышенной способностью продуцировать OPS посредством повышения активности serA (3-фосфоглицератдегидрогеназы) и serC (3-фосфосеринаминотрансферазы), которые представляют собой пути биосинтеза OPS, использовали для определения влияния гена мембранного белка Е. coli, и векторы сверхэкспрессии для гена мембранного белка получали в комбинации с serA и serC. Последовательности используемых праймеров, являются такими, как показано в Таблице 4 ниже.
Во-первых, получали pCL_Ptrc-serA*C для того, чтобы приготовить плазмиды для отрицательного контроля, в которые были введены serA и serC. Фрагменты ДНК Ptrc были получены с использованием SEQ ID NO: 23 и SEQ ID NO: 24 на основе pCL_Ptrc-gfp в качестве матрицы. ПЦР выполняли с использованием SEQ ID NO: 25 и SEQ ID NO: 26 на основе рС_Prmf-serA*C (WO 2016/024771 A1) в качестве матрицы с получением фрагментов ДНК serA*C. Амплифицированные фрагменты подвергали IST с вектором pCL1920, обработанным рестриктазами XbaI и HindIII, с получением pCL_Ptrc-serA*C.
Для получения pCL_Ptrc-ser А* С_Ptrc-yhhS, ПЦР выполняли с использованием SEQ ID NO: 27 и SEQ ID NO: 28 на основе pCL_Ptrc-yhhS, который был приготовлен выше, в качестве матрицы. На основе этого были получены фрагменты ДНК Ptrc-yhhS. Амплифицированные фрагменты были подвергнуты IST с помощью вектора pCL_Ptrc-serA*C, обработанного HindIII, с получением pCL_Ptrc-serA* С_Ptrc-yhhS.
pCL_Ptrc-serA*C_Ptrc-mdtH и pCL_Ptrc-serA*C_Ptrc-yfaV были получены также, как в случае pCL_Ptrc-serA*C_Ptrc-yhhS. ДНК фрагменты Ptrc-mdtH были получены на основе pCL_Ptrc-mdtH, используемой в качестве матрицы, с использованием SEQ ID NO: 29 и SEQ ID NO: 30, и ДНК фрагменты Ptrc-yfaV были получены на основе pCL_Ptrc-yfaV в качестве матрицы с использованием SEQ ID NO: 31 и SEQ ID NO: 32.
Способность продуцировать OPS была подтверждена путем введения полученных плазмид в СА07-0022, OPS-продуцирующий штамм, и результаты показаны в Таблице 5 ниже.
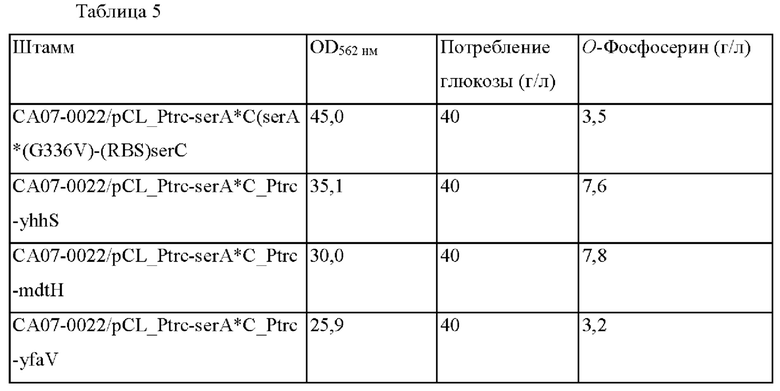
Как видно из представленной выше Таблицы 5, среди штаммов, у которых ген мембранного белка Е. coli был дополнительно введен в CA07-0022/pCL_Ptrc-serA*C, который имел улучшенную способность продуцировать OPS по сравнению со штаммом СА07-0012, полученным из Е. coli, еще раз было подтверждено, что продуцирование OPS увеличено в штамме с усиленным yhhS, который служит положительным контролем, по сравнению с контролем. В частности, в случае mdtH-усиленного штамма по настоящему изобретению, концентрация OPS была повышенной, аналогично результатам, показанным в Таблице 3. Напротив, в случае yfaV-усиленной группы, которая является группой сравнения, продуцирование OPS было уменьшено по сравнению с контрольной группой.
3-3. Получение mdtH транспортер-усиленного штамма в соответствии с силой хромосомного промотора и оценка его способности продуцировать OPS
Для того, чтобы определить будет ли экспортирующая способность улучшена в случае замены промотора mdtH на более сильный промотор в хромосоме, были получены штаммы, в которых аутологичный промотор был заменен на промотор trc, и была оценена их способность продуцировать OPS. Способ введения промотора trc в хромосому Е. coli был выполнен с помощью следующего часто используемого способа. Для вставки в хромосому использовали вектор pSKH130, имеющий белок PI (ген pir)-зависимый R6K репликон, в который введен ген sacB (левансахараза). Кроме того, вектор содержит ген устойчивости к канамицину, который используют в качестве селективного маркера для получения штаммов. После получения необходимого штамма с использованием R6K и канамицина в первом кроссинговере с использованием вектора, антибиотики были удалены из среды, содержащей сахарозу, для получения штаммов. Для того чтобы заменить промотор mdtH промотором trc в СА07-0022, получали вектор pSKH130_Ptrc-mdtH и использовали последовательности праймера, как показано в Таблице 6 ниже.
Для получения pSKH130_Ptrc-mdtH, были получены фрагменты ДНК mdtHUP путем выполнения ПЦР на основе W3110 с использованием SEQ ID NO: 33 и SEQ ID NO: 34. Кроме того, фрагменты ДНК Ptrc-mdtH были получены посредством выполнения ПЦР на основании pCL_Ptrc-mdtH, полученного выше с использованием SEQ ID NO: 35 и SEQ ID NO: 36. Амплифицированные фрагменты были подвергнуты IST с вектором pSKH130, обработанным рестриктазой EcoRV, с получением pSKH_mdtHUP_Ptrc-mdtH. Полученные таким образом плазмиды были трансформированы в штамм СА7-0022 посредством электропорации. После селекции штамма со вставкой в хромосомы в твердой среде LB с добавкой канамицина посредством рекомбинации (кроссинговера), область плазмиды вырезали из хромосомы посредством вторичной рекомбинации (замены) в среде, содержащей сахарозу. Штамм, в котором была завершена вторичная рекомбинация, был получен с использованием праймеров SEQ ID NO: 37 и SEQ ID NO: 38 (CA07-0022::Ptrc-mdtH).
Чтобы определить эффект полученных OPS-продуцирующих штаммов, способность продуцировать OPS показана в таблице 7 ниже.

В результате, как видно из таблицы 7, когда экспрессия мембранного белка E.coli была повышена путем усиления промотора, было подтверждено, что концентрация OPS была увеличена по сравнению с контрольной группой.
Пример 4: Сравнение экспорта 3-фосфоглицерата
OPS представляет собой вещество, содержащее фосфат, и имеет химическую структуру, аналогичную 3-фосфоглицерату (ниже называемому 3PG). Соответственно, предполагалось, что экспортер OPS может высвобождать 3PG, и 3PG-продуцирующую способность измеряли в штаммах с повышенным экспортером OPS. 3PG измеряли с использованием высокоэффективной жидкостной хроматографии (ВЭЖХ) и 3PG-продуцирующая способность показана в Таблице 8.
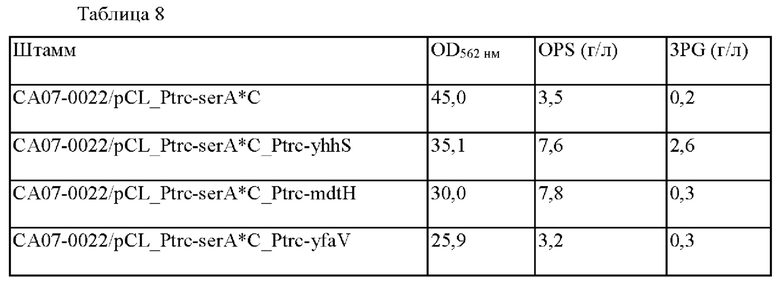
Как можно видеть из Таблицы 8, 3PG накапливался в yhhS-усиленном штамме, но 3PG не был повышен в штамме, усиленном mdtH мембранным белком. То есть было подтверждено, что штамм, в который был введен mdtH, имел OPS-экспортирующую способность, и что экспортирующая способность, специфичная в отношении OPS, была также увеличена.
Специалистам в данной области понятно, что настоящее изобретение может быть воплощено в других конкретных формах без отклонения его сущности или существенных характеристик. Описанные воплощения должны рассматриваться во всех отношениях только как иллюстративные, а не ограничивающие. Таким образом, объем настоящего изобретение определяется прилагаемой формулой изобретения, а не вышеизложенным описанием. Все изменения, которые подпадают под значение и диапазон эквивалентности пунктов формулы изобретения, должны быть включены в объем настоящего изобретения.
Депозитарный номер
Депозитарий: Корейский центр культур микроорганизмов (Международный орган по депонированию)
Регистрационный номер: KCCM12781P
Дата депонирования: 20200828
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> НОВЫЙ O-ФОСФОСЕРИН-ЭКСПОРТИРУЮЩИЙ БЕЛОК И МЕТОДЫ ПОЛУЧЕНИЯ
O-ФОСФОСЕРИНА, ЦИСТЕИНА И ПРОИЗВОДНЫХ ЦИСТЕИНА С ЕГО ПРИМЕНЕНИЕМ
<130> OPA21268
<150> KR 10-2020-0115569
<151> 2020-09-09
<160> 38
<170> KoPatentIn 3.0
<210> 1
<211> 402
<212> PRT
<213> Unknown
<220>
<223> mdtH
<400> 1
Met Ser Arg Val Ser Gln Ala Arg Asn Leu Gly Lys Tyr Phe Leu Leu
1 5 10 15
Ile Asp Asn Met Leu Val Val Leu Gly Phe Phe Val Val Phe Pro Leu
20 25 30
Ile Ser Ile Arg Phe Val Asp Gln Met Gly Trp Ala Ala Val Met Val
35 40 45
Gly Ile Ala Leu Gly Leu Arg Gln Phe Ile Gln Gln Gly Leu Gly Ile
50 55 60
Phe Gly Gly Ala Ile Ala Asp Arg Phe Gly Ala Lys Pro Met Ile Val
65 70 75 80
Thr Gly Met Leu Met Arg Ala Ala Gly Phe Ala Thr Met Gly Ile Ala
85 90 95
His Glu Pro Trp Leu Leu Trp Phe Ser Cys Leu Leu Ser Gly Leu Gly
100 105 110
Gly Thr Leu Phe Asp Pro Pro Arg Ser Ala Leu Val Val Lys Leu Ile
115 120 125
Arg Pro Gln Gln Arg Gly Arg Phe Phe Ser Leu Leu Met Met Gln Asp
130 135 140
Ser Ala Gly Ala Val Ile Gly Ala Leu Leu Gly Ser Trp Leu Leu Gln
145 150 155 160
Tyr Asp Phe Arg Leu Val Cys Ala Thr Gly Ala Val Leu Phe Val Leu
165 170 175
Cys Ala Ala Phe Asn Ala Trp Leu Leu Pro Ala Trp Lys Leu Ser Thr
180 185 190
Val Arg Thr Pro Val Arg Glu Gly Met Thr Arg Val Met Arg Asp Lys
195 200 205
Arg Phe Val Thr Tyr Val Leu Thr Leu Ala Gly Tyr Tyr Met Leu Ala
210 215 220
Val Gln Val Met Leu Met Leu Pro Ile Met Val Asn Asp Val Ala Gly
225 230 235 240
Ala Pro Ser Ala Val Lys Trp Met Tyr Ala Ile Glu Ala Cys Leu Ser
245 250 255
Leu Thr Leu Leu Tyr Pro Ile Ala Arg Trp Ser Glu Lys His Phe Arg
260 265 270
Leu Glu His Arg Leu Met Ala Gly Leu Leu Ile Met Ser Leu Ser Met
275 280 285
Met Pro Val Gly Met Val Ser Gly Leu Gln Gln Leu Phe Thr Leu Ile
290 295 300
Cys Leu Phe Tyr Ile Gly Ser Ile Ile Ala Glu Pro Ala Arg Glu Thr
305 310 315 320
Leu Ser Ala Ser Leu Ala Asp Ala Arg Ala Arg Gly Ser Tyr Met Gly
325 330 335
Phe Ser Arg Leu Gly Leu Ala Ile Gly Gly Ala Ile Gly Tyr Ile Gly
340 345 350
Gly Gly Trp Leu Phe Asp Leu Gly Lys Ser Ala His Gln Pro Glu Leu
355 360 365
Pro Trp Met Met Leu Gly Ile Ile Gly Ile Phe Thr Phe Leu Ala Leu
370 375 380
Gly Trp Gln Phe Ser Gln Lys Arg Ala Ala Arg Arg Leu Leu Glu Arg
385 390 395 400
Asp Ala
<210> 2
<211> 1209
<212> DNA
<213> Unknown
<220>
<223> mdtH
<400> 2
atgtcccgcg tgtcgcaggc gaggaacctg ggtaaatatt tcctgctcat cgataatatg 60
ctggtcgtgc tggggttctt tgttgtcttc ccgctgatct ctatccgctt cgttgatcaa 120
atgggctggg ccgccgtcat ggtcggtatt gctctcggtc tacgccaatt tattcagcaa 180
ggtctgggta ttttcggcgg tgcaattgcc gaccgctttg gtgccaaacc gatgattgtt 240
accggtatgc tgatgcgcgc cgccggattc gccacaatgg gtatcgccca cgaaccgtgg 300
ctattgtggt tttcatgcct gctctcggga ctcggtggca cgttgtttga tccgccgcgt 360
tcggcgctgg tggtgaaatt aatccgtcca cagcagcgtg gtcgtttttt ctcgctgttg 420
atgatgcagg acagtgccgg tgcggtcatt ggcgcattgt tggggagctg gctgttgcaa 480
tacgactttc gcctggtctg cgccacaggg gcagttctat ttgtgctatg tgcggcgttc 540
aatgcgtggt tgttaccagc atggaaactc tccaccgtac gcacgcccgt tcgcgaaggc 600
atgacccgcg tgatgcgtga caagcgtttt gtcacctatg ttctgacgct ggcgggttac 660
tacatgctgg ctgtacaagt gatgctgatg ctgccaatta tggtcaacga cgtggctggc 720
gcgccctctg ccgttaaatg gatgtatgcc attgaagcgt gtctgtcgtt aacgttgctc 780
taccctatcg cccgctggag tgaaaagcat tttcgtctgg aacaccggtt gatggctggg 840
ctgttgataa tgtcattaag catgatgccg gtgggcatgg tcagcggcct gcaacaactt 900
ttcaccctga tttgtctgtt ttatatcggg tcgatcattg ccgagcctgc gcgtgaaacc 960
ttaagtgctt cgctggcgga cgcaagagct cgcggcagct atatggggtt tagccgtctg 1020
ggtctggcga ttggcggcgc tattggttat atcggtggcg gctggctgtt tgacctgggc 1080
aaatcggcgc accagccaga gcttccgtgg atgatgctgg gcattattgg catcttcact 1140
ttccttgcgc tgggttggca gtttagccag aaacgcgccg cgcgtcgttt gcttgaacgc 1200
gacgcctga 1209
<210> 3
<211> 322
<212> PRT
<213> Unknown
<220>
<223> serB
<400> 3
Met Pro Asn Ile Thr Trp Cys Asp Leu Pro Glu Asp Val Ser Leu Trp
1 5 10 15
Pro Gly Leu Pro Leu Ser Leu Ser Gly Asp Glu Val Met Pro Leu Asp
20 25 30
Tyr His Ala Gly Arg Ser Gly Trp Leu Leu Tyr Gly Arg Gly Leu Asp
35 40 45
Lys Gln Arg Leu Thr Gln Tyr Gln Ser Lys Leu Gly Ala Ala Met Val
50 55 60
Ile Val Ala Ala Trp Cys Val Glu Asp Tyr Gln Val Ile Arg Leu Ala
65 70 75 80
Gly Ser Leu Thr Ala Arg Ala Thr Arg Leu Ala His Glu Ala Gln Leu
85 90 95
Asp Val Ala Pro Leu Gly Lys Ile Pro His Leu Arg Thr Pro Gly Leu
100 105 110
Leu Val Met Asp Met Asp Ser Thr Ala Ile Gln Ile Glu Cys Ile Asp
115 120 125
Glu Ile Ala Lys Leu Ala Gly Thr Gly Glu Met Val Ala Glu Val Thr
130 135 140
Glu Arg Ala Met Arg Gly Glu Leu Asp Phe Thr Ala Ser Leu Arg Ser
145 150 155 160
Arg Val Ala Thr Leu Lys Gly Ala Asp Ala Asn Ile Leu Gln Gln Val
165 170 175
Arg Glu Asn Leu Pro Leu Met Pro Gly Leu Thr Gln Leu Val Leu Lys
180 185 190
Leu Glu Thr Leu Gly Trp Lys Val Ala Ile Ala Ser Gly Gly Phe Thr
195 200 205
Phe Phe Ala Glu Tyr Leu Arg Asp Lys Leu Arg Leu Thr Ala Val Val
210 215 220
Ala Asn Glu Leu Glu Ile Met Asp Gly Lys Phe Thr Gly Asn Val Ile
225 230 235 240
Gly Asp Ile Val Asp Ala Gln Tyr Lys Ala Lys Thr Leu Thr Arg Leu
245 250 255
Ala Gln Glu Tyr Glu Ile Pro Leu Ala Gln Thr Val Ala Ile Gly Asp
260 265 270
Gly Ala Asn Asp Leu Pro Met Ile Lys Ala Ala Gly Leu Gly Ile Ala
275 280 285
Tyr His Ala Lys Pro Lys Val Asn Glu Lys Ala Glu Val Thr Ile Arg
290 295 300
His Ala Asp Leu Met Gly Val Phe Cys Ile Leu Ser Gly Ser Leu Asn
305 310 315 320
Gln Lys
<210> 4
<211> 969
<212> DNA
<213> Unknown
<220>
<223> serB
<400> 4
atgcctaaca ttacctggtg cgacctgcct gaagatgtct ctttatggcc gggtctgcct 60
ctttcattaa gtggtgatga agtgatgcca ctggattacc acgcaggtcg tagcggctgg 120
ctgctgtatg gtcgtgggct ggataaacaa cgtctgaccc aataccagag caaactgggt 180
gcggcgatgg tgattgttgc cgcctggtgc gtggaagatt atcaggtgat tcgtctggca 240
ggttcactca ccgcacgggc tacacgcctg gcccacgaag cgcagctgga tgtcgccccg 300
ctggggaaaa tcccgcacct gcgcacgccg ggtttgctgg tgatggatat ggactccacc 360
gccatccaga ttgaatgtat tgatgaaatt gccaaactgg ccggaacggg cgagatggtg 420
gcggaagtaa ccgaacgggc gatgcgcggc gaactcgatt ttaccgccag cctgcgcagc 480
cgtgtggcga cgctgaaagg cgctgacgcc aatattctgc aacaggtgcg tgaaaatctg 540
ccgctgatgc caggcttaac gcaactggtg ctcaagctgg aaacgctggg ctggaaagtg 600
gcgattgcct ccggcggctt tactttcttt gctgaatacc tgcgcgacaa gctgcgcctg 660
accgccgtgg tagccaatga actggagatc atggacggta aatttaccgg caatgtgatc 720
ggcgacatcg tagacgcgca gtacaaagcg aaaactctga ctcgcctcgc gcaggagtat 780
gaaatcccgc tggcgcagac cgtggcgatt ggcgatggag ccaatgacct gccgatgatc 840
aaagcggcag ggctggggat tgcctaccat gccaagccaa aagtgaatga aaaggcggaa 900
gtcaccatcc gtcacgctga cctgatgggg gtattctgca tcctctcagg cagcctgaat 960
cagaagtaa 969
<210> 5
<211> 410
<212> PRT
<213> Unknown
<220>
<223> SerA дикого типа
<400> 5
Met Ala Lys Val Ser Leu Glu Lys Asp Lys Ile Lys Phe Leu Leu Val
1 5 10 15
Glu Gly Val His Gln Lys Ala Leu Glu Ser Leu Arg Ala Ala Gly Tyr
20 25 30
Thr Asn Ile Glu Phe His Lys Gly Ala Leu Asp Asp Glu Gln Leu Lys
35 40 45
Glu Ser Ile Arg Asp Ala His Phe Ile Gly Leu Arg Ser Arg Thr His
50 55 60
Leu Thr Glu Asp Val Ile Asn Ala Ala Glu Lys Leu Val Ala Ile Gly
65 70 75 80
Cys Phe Cys Ile Gly Thr Asn Gln Val Asp Leu Asp Ala Ala Ala Lys
85 90 95
Arg Gly Ile Pro Val Phe Asn Ala Pro Phe Ser Asn Thr Arg Ser Val
100 105 110
Ala Glu Leu Val Ile Gly Glu Leu Leu Leu Leu Leu Arg Gly Val Pro
115 120 125
Glu Ala Asn Ala Lys Ala His Arg Gly Val Trp Asn Lys Leu Ala Ala
130 135 140
Gly Ser Phe Glu Ala Arg Gly Lys Lys Leu Gly Ile Ile Gly Tyr Gly
145 150 155 160
His Ile Gly Thr Gln Leu Gly Ile Leu Ala Glu Ser Leu Gly Met Tyr
165 170 175
Val Tyr Phe Tyr Asp Ile Glu Asn Lys Leu Pro Leu Gly Asn Ala Thr
180 185 190
Gln Val Gln His Leu Ser Asp Leu Leu Asn Met Ser Asp Val Val Ser
195 200 205
Leu His Val Pro Glu Asn Pro Ser Thr Lys Asn Met Met Gly Ala Lys
210 215 220
Glu Ile Ser Leu Met Lys Pro Gly Ser Leu Leu Ile Asn Ala Ser Arg
225 230 235 240
Gly Thr Val Val Asp Ile Pro Ala Leu Cys Asp Ala Leu Ala Ser Lys
245 250 255
His Leu Ala Gly Ala Ala Ile Asp Val Phe Pro Thr Glu Pro Ala Thr
260 265 270
Asn Ser Asp Pro Phe Thr Ser Pro Leu Cys Glu Phe Asp Asn Val Leu
275 280 285
Leu Thr Pro His Ile Gly Gly Ser Thr Gln Glu Ala Gln Glu Asn Ile
290 295 300
Gly Leu Glu Val Ala Gly Lys Leu Ile Lys Tyr Ser Asp Asn Gly Ser
305 310 315 320
Thr Leu Ser Ala Val Asn Phe Pro Glu Val Ser Leu Pro Leu His Gly
325 330 335
Gly Arg Arg Leu Met His Ile His Glu Asn Arg Pro Gly Val Leu Thr
340 345 350
Ala Leu Asn Lys Ile Phe Ala Glu Gln Gly Val Asn Ile Ala Ala Gln
355 360 365
Tyr Leu Gln Thr Ser Ala Gln Met Gly Tyr Val Val Ile Asp Ile Glu
370 375 380
Ala Asp Glu Asp Val Ala Glu Lys Ala Leu Gln Ala Met Lys Ala Ile
385 390 395 400
Pro Gly Thr Ile Arg Ala Arg Leu Leu Tyr
405 410
<210> 6
<211> 410
<212> PRT
<213> Artificial Sequence
<220>
<223> SerA мутантного типа
<400> 6
Met Ala Lys Val Ser Leu Glu Lys Asp Lys Ile Lys Phe Leu Leu Val
1 5 10 15
Glu Gly Val His Gln Lys Ala Leu Glu Ser Leu Arg Ala Ala Gly Tyr
20 25 30
Thr Asn Ile Glu Phe His Lys Gly Ala Leu Asp Asp Glu Gln Leu Lys
35 40 45
Glu Ser Ile Arg Asp Ala His Phe Ile Gly Leu Arg Ser Arg Thr His
50 55 60
Leu Thr Glu Asp Val Ile Asn Ala Ala Glu Lys Leu Val Ala Ile Gly
65 70 75 80
Cys Phe Cys Ile Gly Thr Asn Gln Val Asp Leu Asp Ala Ala Ala Lys
85 90 95
Arg Gly Ile Pro Val Phe Asn Ala Pro Phe Ser Asn Thr Arg Ser Val
100 105 110
Ala Glu Leu Val Ile Gly Glu Leu Leu Leu Leu Leu Arg Gly Val Pro
115 120 125
Glu Ala Asn Ala Lys Ala His Arg Gly Val Trp Asn Lys Leu Ala Ala
130 135 140
Gly Ser Phe Glu Ala Arg Gly Lys Lys Leu Gly Ile Ile Gly Tyr Gly
145 150 155 160
His Ile Gly Thr Gln Leu Gly Ile Leu Ala Glu Ser Leu Gly Met Tyr
165 170 175
Val Tyr Phe Tyr Asp Ile Glu Asn Lys Leu Pro Leu Gly Asn Ala Thr
180 185 190
Gln Val Gln His Leu Ser Asp Leu Leu Asn Met Ser Asp Val Val Ser
195 200 205
Leu His Val Pro Glu Asn Pro Ser Thr Lys Asn Met Met Gly Ala Lys
210 215 220
Glu Ile Ser Leu Met Lys Pro Gly Ser Leu Leu Ile Asn Ala Ser Arg
225 230 235 240
Gly Thr Val Val Asp Ile Pro Ala Leu Cys Asp Ala Leu Ala Ser Lys
245 250 255
His Leu Ala Gly Ala Ala Ile Asp Val Phe Pro Thr Glu Pro Ala Thr
260 265 270
Asn Ser Asp Pro Phe Thr Ser Pro Leu Cys Glu Phe Asp Asn Val Leu
275 280 285
Leu Thr Pro His Ile Gly Gly Ser Thr Gln Glu Ala Gln Glu Asn Ile
290 295 300
Gly Leu Glu Val Ala Gly Lys Leu Ile Lys Tyr Ser Asp Asn Gly Ser
305 310 315 320
Thr Leu Ser Ala Val Asn Phe Pro Glu Val Ser Leu Pro Leu His Val
325 330 335
Gly Arg Arg Leu Met His Ile His Glu Asn Arg Pro Gly Val Leu Thr
340 345 350
Ala Leu Asn Lys Ile Phe Ala Glu Gln Gly Val Asn Ile Ala Ala Gln
355 360 365
Tyr Leu Gln Thr Ser Ala Gln Met Gly Tyr Val Val Ile Asp Ile Glu
370 375 380
Ala Asp Glu Asp Val Ala Glu Lys Ala Leu Gln Ala Met Lys Ala Ile
385 390 395 400
Pro Gly Thr Ile Arg Ala Arg Leu Leu Tyr
405 410
<210> 7
<211> 1233
<212> DNA
<213> Unknown
<220>
<223> SerA дикого типа
<400> 7
atggcaaagg tatcgctgga gaaagacaag attaagtttc tgctggtaga aggcgtgcac 60
caaaaggcgc tggaaagcct tcgtgcagct ggttacacca acatcgaatt tcacaaaggc 120
gcgctggatg atgaacaatt aaaagaatcc atccgcgatg cccacttcat cggcctgcga 180
tcccgtaccc atctgactga agacgtgatc aacgccgcag aaaaactggt cgctattggc 240
tgtttctgta tcggaacaaa ccaggttgat ctggatgcgg cggcaaagcg cgggatcccg 300
gtatttaacg caccgttctc aaatacgcgc tctgttgcgg agctggtgat tggcgaactg 360
ctgctgctat tgcgcggcgt gccggaagcc aatgctaaag cgcaccgtgg cgtgtggaac 420
aaactggcgg cgggttcttt tgaagcgcgc ggcaaaaagc tgggtatcat cggctacggt 480
catattggta cgcaattggg cattctggct gaatcgctgg gaatgtatgt ttacttttat 540
gatattgaaa ataaactgcc gctgggcaac gccactcagg tacagcatct ttctgacctg 600
ctgaatatga gcgatgtggt gagtctgcat gtaccagaga atccgtccac caaaaatatg 660
atgggcgcga aagaaatttc actaatgaag cccggctcgc tgctgattaa tgcttcgcgc 720
ggtactgtgg tggatattcc ggcgctgtgt gatgcgctgg cgagcaaaca tctggcgggg 780
gcggcaatcg acgtattccc gacggaaccg gcgaccaata gcgatccatt tacctctccg 840
ctgtgtgaat tcgacaacgt ccttctgacg ccacacattg gcggttcgac tcaggaagcg 900
caggagaata tcggcctgga agttgcgggt aaattgatca agtattctga caatggctca 960
acgctctctg cggtgaactt cccggaagtc tcgctgccac tgcacggtgg gcgtcgtctg 1020
atgcacatcc acgaaaaccg tccgggcgtg ctaactgcgc tgaacaaaat cttcgccgag 1080
cagggcgtca acatcgccgc gcaatatctg caaacttccg cccagatggg ttatgtggtt 1140
attgatattg aagccgacga agacgttgcc gaaaaagcgc tgcaggcaat gaaagctatt 1200
ccgggtacca ttcgcgcccg tctgctgtac taa 1233
<210> 8
<211> 1233
<212> DNA
<213> Artificial Sequence
<220>
<223> SerA мутантного типа
<400> 8
atggcaaagg tatcgctgga gaaagacaag attaagtttc tgctggtaga aggcgtgcac 60
caaaaggcgc tggaaagcct tcgtgcagct ggttacacca acatcgaatt tcacaaaggc 120
gcgctggatg atgaacaatt aaaagaatcc atccgcgatg cccacttcat cggcctgcga 180
tcccgtaccc atctgactga agacgtgatc aacgccgcag aaaaactggt cgctattggc 240
tgtttctgta tcggaacaaa ccaggttgat ctggatgcgg cggcaaagcg cgggatcccg 300
gtatttaacg caccgttctc aaatacgcgc tctgttgcgg agctggtgat tggcgaactg 360
ctgctgctat tgcgcggcgt gccggaagcc aatgctaaag cgcaccgtgg cgtgtggaac 420
aaactggcgg cgggttcttt tgaagcgcgc ggcaaaaagc tgggtatcat cggctacggt 480
catattggta cgcaattggg cattctggct gaatcgctgg gaatgtatgt ttacttttat 540
gatattgaaa ataaactgcc gctgggcaac gccactcagg tacagcatct ttctgacctg 600
ctgaatatga gcgatgtggt gagtctgcat gtaccagaga atccgtccac caaaaatatg 660
atgggcgcga aagaaatttc actaatgaag cccggctcgc tgctgattaa tgcttcgcgc 720
ggtactgtgg tggatattcc ggcgctgtgt gatgcgctgg cgagcaaaca tctggcgggg 780
gcggcaatcg acgtattccc gacggaaccg gcgaccaata gcgatccatt tacctctccg 840
ctgtgtgaat tcgacaacgt ccttctgacg ccacacattg gcggttcgac tcaggaagcg 900
caggagaata tcggcctgga agttgcgggt aaattgatca agtattctga caatggctca 960
acgctctctg cggtgaactt cccggaagtc tcgctgccac tgcacgttgg gcgtcgtctg 1020
atgcacatcc acgaaaaccg tccgggcgtg ctaactgcgc tgaacaaaat cttcgccgag 1080
cagggcgtca acatcgccgc gcaatatctg caaacttccg cccagatggg ttatgtggtt 1140
attgatattg aagccgacga agacgttgcc gaaaaagcgc tgcaggcaat gaaagctatt 1200
ccgggtacca ttcgcgcccg tctgctgtac taa 1233
<210> 9
<211> 362
<212> PRT
<213> Unknown
<220>
<223> SerC
<400> 9
Met Ala Gln Ile Phe Asn Phe Ser Ser Gly Pro Ala Met Leu Pro Ala
1 5 10 15
Glu Val Leu Lys Gln Ala Gln Gln Glu Leu Arg Asp Trp Asn Gly Leu
20 25 30
Gly Thr Ser Val Met Glu Val Ser His Arg Gly Lys Glu Phe Ile Gln
35 40 45
Val Ala Glu Glu Ala Glu Lys Asp Phe Arg Asp Leu Leu Asn Val Pro
50 55 60
Ser Asn Tyr Lys Val Leu Phe Cys His Gly Gly Gly Arg Gly Gln Phe
65 70 75 80
Ala Ala Val Pro Leu Asn Ile Leu Gly Asp Lys Thr Thr Ala Asp Tyr
85 90 95
Val Asp Ala Gly Tyr Trp Ala Ala Ser Ala Ile Lys Glu Ala Lys Lys
100 105 110
Tyr Cys Thr Pro Asn Val Phe Asp Ala Lys Val Thr Val Asp Gly Leu
115 120 125
Arg Ala Val Lys Pro Met Arg Glu Trp Gln Leu Ser Asp Asn Ala Ala
130 135 140
Tyr Met His Tyr Cys Pro Asn Glu Thr Ile Asp Gly Ile Ala Ile Asp
145 150 155 160
Glu Thr Pro Asp Phe Gly Ala Asp Val Val Val Ala Ala Asp Phe Ser
165 170 175
Ser Thr Ile Leu Ser Arg Pro Ile Asp Val Ser Arg Tyr Gly Val Ile
180 185 190
Tyr Ala Gly Ala Gln Lys Asn Ile Gly Pro Ala Gly Leu Thr Ile Val
195 200 205
Ile Val Arg Glu Asp Leu Leu Gly Lys Ala Asn Ile Ala Cys Pro Ser
210 215 220
Ile Leu Asp Tyr Ser Ile Leu Asn Asp Asn Gly Ser Met Phe Asn Thr
225 230 235 240
Pro Pro Thr Phe Ala Trp Tyr Leu Ser Gly Leu Val Phe Lys Trp Leu
245 250 255
Lys Ala Asn Gly Gly Val Ala Glu Met Asp Lys Ile Asn Gln Gln Lys
260 265 270
Ala Glu Leu Leu Tyr Gly Val Ile Asp Asn Ser Asp Phe Tyr Arg Asn
275 280 285
Asp Val Ala Lys Ala Asn Arg Ser Arg Met Asn Val Pro Phe Gln Leu
290 295 300
Ala Asp Ser Ala Leu Asp Lys Leu Phe Leu Glu Glu Ser Phe Ala Ala
305 310 315 320
Gly Leu His Ala Leu Lys Gly His Arg Val Val Gly Gly Met Arg Ala
325 330 335
Ser Ile Tyr Asn Ala Met Pro Leu Glu Gly Val Lys Ala Leu Thr Asp
340 345 350
Phe Met Val Glu Phe Glu Arg Arg His Gly
355 360
<210> 10
<211> 1089
<212> DNA
<213> Unknown
<220>
<223> SerC
<400> 10
atggctcaaa tcttcaattt tagttctggt ccggcaatgc taccggcaga ggtgcttaaa 60
caggctcaac aggaactgcg cgactggaac ggtcttggta cgtcggtgat ggaagtgagt 120
caccgtggca aagagttcat tcaggttgca gaggaagccg agaaggattt tcgcgatctt 180
cttaatgtcc cctccaacta caaggtatta ttctgccatg gcggtggtcg cggtcagttt 240
gctgcggtac cgctgaatat tctcggtgat aaaaccaccg cagattatgt tgatgccggt 300
tactgggcgg caagtgccat taaagaagcg aaaaaatact gcacgcctaa tgtctttgac 360
gccaaagtga ctgttgatgg tctgcgcgcg gttaagccaa tgcgtgaatg gcaactctct 420
gataatgctg cttatatgca ttattgcccg aatgaaacca tcgatggtat cgccatcgac 480
gaaacgccag acttcggcgc agatgtggtg gtcgccgctg acttctcttc aaccattctt 540
tcccgtccga ttgacgtcag ccgttatggt gtaatttacg ctggcgcgca gaaaaatatc 600
ggcccggctg gcctgacaat cgtcatcgtt cgtgaagatt tgctgggcaa agcgaatatc 660
gcgtgtccgt cgattctgga ttattccatc ctcaacgata acggctccat gtttaacacg 720
ccgccgacat ttgcctggta tctatctggt ctggtcttta aatggctgaa agcgaacggc 780
ggtgtagctg aaatggataa aatcaatcag caaaaagcag aactgctata tggggtgatt 840
gataacagcg atttctaccg caatgacgtg gcgaaagcta accgttcgcg gatgaacgtg 900
ccgttccagt tggcggacag tgcgcttgac aaattgttcc ttgaagagtc ttttgctgct 960
ggccttcatg cactgaaagg tcaccgtgtg gtcggcggaa tgcgcgcttc tatttataac 1020
gccatgccgc tggaaggcgt taaagcgctg acagacttca tggttgagtt cgaacgccgt 1080
cacggttaa 1089
<210> 11
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yhhS_Ptrc_F
<400> 11
cggggatcct ctagacgctt gctgcaactc tctca 35
<210> 12
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yhhS_Ptrc_R
<400> 12
tacgggttcg ggcatgatat ctttcctgtg tgaaa 35
<210> 13
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yhhS_F
<400> 13
cacaggaaag atatcatgcc cgaacccgta gccga 35
<210> 14
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yhhS_R
<400> 14
gattacgcca agcttttaag atgatgaggc ggcct 35
<210> 15
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> mdtH_Ptrc_F
<400> 15
cggggatcct ctagacgctt gctgcaactc tctca 35
<210> 16
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> mdtH_Ptrc_R
<400> 16
cgacacgcgg gacatgatat ctttcctgtg tgaaa 35
<210> 17
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> mdtH_F
<400> 17
cacaggaaag atatcatgtc ccgcgtgtcg caggc 35
<210> 18
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> mdtH_R
<400> 18
gattacgcca agctttcagg cgtcgcgttc aagca 35
<210> 19
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yfaV_Ptrc_F
<400> 19
cggggatcct ctagacgctt gctgcaactc tctca 35
<210> 20
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yfaV_Ptrc_R
<400> 20
caaagcggtg ctcatgatat ctttcctgtg tgaaa 35
<210> 21
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yfaV_F
<400> 21
cacaggaaag atatcatgag caccgctttg cttga 35
<210> 22
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yfaV_R
<400> 22
gattacgcca agcttttaat gatgtgccac gtcgg 35
<210> 23
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> serA*,serC_Ptrc_F
<400> 23
cggggatcct ctagaggtac ccgcttgctg caact 35
<210> 24
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> serA*,serC_Ptrc_R
<400> 24
cgataccttt gccatgatat ctttcctgtg tgaaa 35
<210> 25
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> serA*,serC_F
<400> 25
cacaggaaag atatcatggc aaaggtatcg ctgga 35
<210> 26
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> serA*,serC_R
<400> 26
gattacgcca agcttttaac cgtgacggcg ttcga 35
<210> 27
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> serA*,serC,yhhS_Ptrc-yhhS_F
<400> 27
cacggttaaa agcttcgctt gctgcaactc tctca 35
<210> 28
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> serA*,serC,yhhS_Ptrc-yhhS_R
<400> 28
gattacgcca agcttttaag atgatgaggc ggcct 35
<210> 29
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> serA*,serC,mdtH_Ptrc-mdtH_F
<400> 29
cacggttaaa agcttcgctt gctgcaactc tctca 35
<210> 30
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> serA*,serC,mdtH_Ptrc-mdtH_R
<400> 30
gattacgcca agctttcagg cgtcgcgttc aagca 35
<210> 31
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> serA*,serC,mdtH_Ptrc-yfaV_F
<400> 31
cacggttaaa agcttcgctt gctgcaactc tctca 35
<210> 32
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> serA*,serC,mdtH_Ptrc-yfaV_R
<400> 32
gattacgcca agcttttaat gatgtgccac gtcgg 35
<210> 33
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> mdtHUP_F
<400> 33
caggaattcg atatctaatc tctttttcgt ccggg 35
<210> 34
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> mdtHUP_R
<400> 34
gagttgcagc aagcgttccc ctcccgggaa ataaa 35
<210> 35
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Ptrc-mdtH_F
<400> 35
ttcccgggag gggaacgctt gctgcaactc tctca 35
<210> 36
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Ptrc-mdtH_R
<400> 36
gactagcgtg atatccagac caggcgaaag tcgta 35
<210> 37
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Ptrc-mdtH_conf_F
<400> 37
caccgctgcg tttattgt 18
<210> 38
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Ptrc-mdtH_conf_R
<400> 38
aaacgcttgt cacgcatca 19
<---
Изобретение относится к биотехнологии. Предложено применение рекомбинантного микроорганизма, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью, для продуцирования O-фосфосерина. Также предложены способы получения О-фосфосерина, цистеина и производного цистеина с использованием указанного микроорганизма. Также предложены рекомбинантный микроорганизм для продуцирования O-фосфосерина, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью и у которого дополнительно ослаблена активность фосфосеринфосфатазы (SerB) по сравнению с его эндогенной активностью, а также рекомбинантный микроорганизм для продуцирования O-фосфосерина, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью и у которого дополнительно повышена активность любой из фосфоглицератдегидрогеназы (SerA), фосфосеринаминотрансферазы (SerC) или их комбинации по сравнению с его эндогенной активностью. Изобретение обеспечивает высокопродуктивное продуцирование О-фосфосерина. 6 н. и 5 з.п. ф-лы, 8 табл., 4 пр.
1. Применение рекомбинантного микроорганизма, продуцирующего O-фосфосерин, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью, для продуцирования O-фосфосерина.
2. Применение по п.1, где у микроорганизма O-фосфосерин-экспортирующая активность повышена по сравнению с его эндогенной активностью.
3. Применение по п. 1, где белок mdtH содержит аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, имеющую по меньшей мере 95% идентичность с SEQ ID NO: 1, при наличии O-фосфосерин- экспортирующей активности.
4. Рекомбинантный микроорганизм для продуцирования O-фосфосерина, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью и у которого дополнительно ослаблена активность фосфосеринфосфатазы (SerB) по сравнению с его эндогенной активностью.
5. Рекомбинантный микроорганизм для продуцирования O-фосфосерина, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью и у которого дополнительно повышена активность любой из фосфоглицератдегидрогеназы (SerA), фосфосеринаминотрансферазы (SerC) или их комбинации по сравнению с его эндогенной активностью.
6. Применение по п. 1, где рекомбинантный микроорганизм принадлежит к роду Escherichia.
7. Способ получения O-фосфосерина, включающий культивирование в среде рекомбинантного микроорганизма для получения O-фосфосерина, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью.
8. Способ по п. 7, дополнительно включающий извлечение O-фосфосерина из культуральной среды или микроорганизма.
9. Способ получения цистеина, включающий:
а) получение O-фосфосерина или среды, содержащей O-фосфосерин, посредством культивирования в среде рекомбинантного микроорганизма, продуцирующего O-фосфосерин, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью; и
б) взаимодействие O-фосфосерина или среды, содержащей O-фосфосерин, полученных на стадии (a), с сульфидом в присутствии O-фосфосеринсульфгидрилазы (OPSS) или содержащего ее микроорганизма.
10. Способ по п. 9, где сульфид представляет собой по меньшей мере один, выбранный из группы, состоящей из Na2S, NaSH, (NH4)2S, H2S и Na2S2O3.
11. Способ получения производных цистеина, включающий:
а) получение O-фосфосерина или среды, содержащей O-фосфосерин, посредством культивирования в среде рекомбинантного микроорганизма, продуцирующего O-фосфосерин, у которого активность белка mdtH повышена по сравнению с его эндогенной активностью; и
б) взаимодействие O-фосфосерина или среды, содержащей O-фосфосерин, полученных на стадии (a), с сульфидом в присутствии O-фосфосеринсульфгидрилазы (OPSS) или содержащего ее микроорганизма; и
в) превращение цистеина, полученного на стадии б), в производные цистеина.
| ZHANG Y | |||
| ET AL | |||
| Native efflux pumps of Escherichia coli responsible for short and medium chain alcohol | |||
| Biochemical Engineering Journal, Volume 133, 15 May 2018, Pages 149-156, https://doi.org/10.1016/j.bej.2018.02.009 | |||
| МИКРООРГАНИЗМ, ПРОДУЦИРУЮЩИЙ О-ФОСФОСЕРИН, И СПОСОБ ПОЛУЧЕНИЯ О-ФОСФОСЕРИНА ИЛИ L-ЦИСТЕИНА С ИСПОЛЬЗОВАНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2015 |
|
RU2663726C1 |
| WO 2012053794 A2, 26.04.2012 | |||
| Новый вариант эффлюксного белка О-фосфосерина и способ продуцирования О-фосфосерина, цистеина и его производного с использованием этого варианта | 2016 |
|
RU2694589C1 |

