Область техники.
Изобретение относится к области иммунологии, инфекционных заболеваний и исследований СПИДа. Более конкретно, настоящее изобретение относится к новому способу определения пациентов со СПИДом или другим образом подавленным иммунитетом при высоком риске развития криптококкоза.
Предшествующий уровень техники.
Одним из отличительных особенностей синдрома приобретенного иммунодефицита (СПИД) является тот факт, что большая часть смертельных случаев не связана с инфицированием самим вирусом иммунодефицита человека (HIV). На практике большинство смертельных случаев вызывается различными оппортунистическими простейшими, грибками, бактериями и вирусами, которые почти не оказывают влияния на здоровых индивидуумов. Наиболее обычные оппортунистические инфекции включают Pneumocystis carinii, Toxoplasma gondii, Mycobacterium spp. Human Cytomegalovirus и дрожжи Cryptococcus neoformans.
C. neoformans представляет собой дрожжи, которые повсеместно встречаются в нашем окружении. По-видимому, многие индивидуумы инфицированы этими видами дрожжей, но их иммунная система предотвращает развитие заболевания. Потеря иммунной функции, с большой вероятностью, приводит к распространению дрожжей из их первоначально латентного положения, возможно из легких, в кровь и, в конечном счете, в оболочки мозга.
В лечении пациентов со СПИДом наблюдается прогресс. Длительность их жизни почти удвоилась, улучшилось также качество жизни пациентов со СПИДом. Основным усовершенствованием в лечении пациентов со СПИДом было профилактическое применение антибиотиков для предотвращения пневмоцистной пневмонии. Такой подход, при котором всем пациентам со СПИДом проводят пневмоцистозную профилактику, является эффективным, так как у большинства (70%) пациентов со СПИДом развивается пневмоциститная пневмония.
Профилактика криптококкоза у пациентов со СПИДом имеет большие возможности, но подход должен отличаться от подхода, который используют при лечении против Pneumocystis carinii. Криптококкоз ожидается у 5-15% пациентов со СПИДом. Если лишь у 5-15% пациентов развивается криптококкоз, универсальное профилактическое лечение пациентов со СПИДом против криптококкоза нереально. Вместо этого, профилактика пациентов со СПИДом будет зависеть от разработки способов идентификации пациентов со СПИДом, наиболее подверженных риску развития криптококкоза.
Ранее не существовало эффективных средств для определения пациентов со СПИДом или вызванным другими причинами иммунодефицитом, подверженных высокому риску заболевания криптококкозом. Настоящее изобретение восполняет эту давно существующую необходимость в этой области.
Раскрытие изобретения.
В одном из вариантов настоящего изобретения предложен способ определения индивидуумов, подверженных высокому риску развития криптококкоза, который включает стадию определения уровней антител к глюкуроноксиломанну в сыворотке указанных индивидуумов.
В другом варианте настоящего изобретения предложен способ, обеспечивающий присоединение полисахаридаглюкуроноксиломанна к твердой фазе, пригодной для определения связывания антитела с антигеном, который включает стадии активации полисахарида при pH около 8,5 с использованием бензохинона; присоединения активированного полисахарида к гидрофобному лиганду при pH около 7,5 и связывание модифицированного GXM (глюкуроноксиломанна) с указанной твердой фазой при pH около 7,0.
В еще одном варианте настоящего изобретения предложен способ определения антител к глюкуроноксиломанну в сыворотке, включающий стадии инкубирования микротитровальной пластины, ячейки которой покрыты антигеном, в течение 90 минут при 25oC с сывороткой, разбавленной в смеси PBS-Tween; удаления сыворотки пациента из ячеек и промывки указанных ячеек смесью PBS Tween для удаления несвязанных антител; инкубирования промытых ячеек в течение 30 минут при 25oC с разбавленным антителом, коньюгированным с энзимом и специфичным к IgG человека; удаления антител, коньюгированных с энзимом из ячеек и удаления несвязанных коньюгированных с энзимом антител за счет промывки ячеек смесью PBS-Tween; добавления субстрата, соответствующего энзим-коньюгированному антителу и инкубирования в течение 30 минут при 25oC; добавления прекращающего реакцию раствора и определения оптической плотности энзимного продукта.
В другом варианте настоящего изобретения предложен набор для определения присутствия антител к глюкуроноксиломанну в сыворотке индивидуумов с высокой степенью риска развития криптококкоза, который включает твердую фазу; гидрофобный лиганд, который обеспечивает гидрофобное связывание глюкуроноксиломанна с твердой фазой; энзимную индикаторную систему и субстрат, соответствующий связанному с энзимом антителу.
В еще одном варианте настоящего изобретения предложен способ лечения индивидуумов с высокой степенью риска развития криптококкоза, включающий стадии: определение уровней антител к глюкуроноксиломанну в сыворотке указанных индивидуумов и введение антиCryptococcis neoformans агента.
Другие и дополнительные аспекты, особенности и преимущества настоящего изобретения будут очевидны из последующего описания предложенных предпочтительных вариантов изобретения, приведенных с целью раскрытия изобретения.
Краткое описание рисунков
С тем чтобы предмет изобретения, в котором вышеперечисленные особенности, преимущества и цели изобретения, а также те, которые станут очевидны впоследствии, были разъяснены более подробно, более конкретные описания изобретения, кратко изложенного ранее, можно проиллюстрировать со ссылками на некоторые его варианты, которые проиллюстрированы в прилагаемых рисунках. Эти рисунки составляют часть описания. Однако, следует отметить, что прилагаемые рисунки иллюстрируют предпочтительные варианты изобретения и поэтому не должны рассматриваться как ограничивающие объем изобретения.
Фиг. 1(a-f) представляет влияние композиции кроющего буфера на связывание 1251-меченного тираминированного глюкуроноксиломанна на (T-GXM) с Immulon 1 пластинами. Данные представлены как количество связанного T-GXM после добавления различных количеств T-GXM (рисунки слева) или как процент добавляемого T-GXM, который связан в ячейках (рисунки справа). T-GXM разбавляют в 0,05 М карбонатном буфере (верхние рисунки) фосфатным буфером (центральные рисунки) или цитратным буфером (нижние рисунки). Буферы используют либо в неизменном виде (треугольники), либо дополненные 0,1М NaCl (квадраты), либо дополненные 0,95 М NaCl (кружки). Представленные результаты являются средними из четырех повторяющихся ячеек.
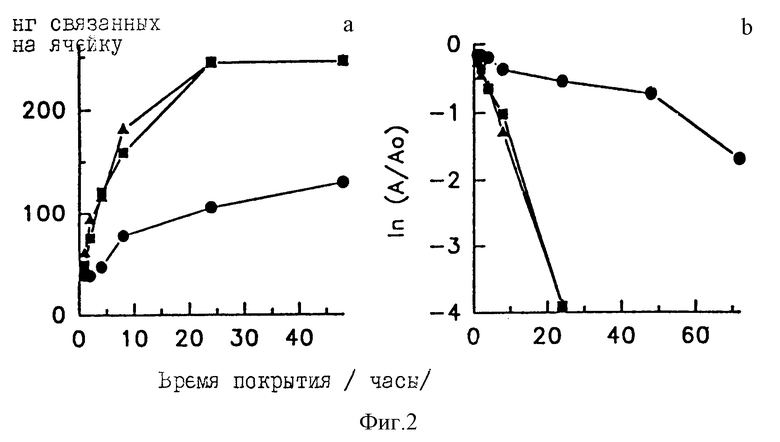
Фиг. 2(a, b) представляет влияние времени инкубирования и температуры на связывание T-GXM с пластинами Immulon 1. Кинетические кривые связывания построены как количество связей для каждого временного интервала (рисунки слева) и как графики псевдопервого порядка, где A0 представляет ожидаемый максимум количества связанных T-GXM, а A представляет возможную связывающую способность для каждого временного интервала. Ячейки инкубируют в течение различных промежутков времени при 4oC (кружки), 20oC (квадраты) или 37oC (треугольники) со 100 мкл 10 мкг/мл радиомеченого тираминированного GXM в фосфатном кроющем буфере, содержащем 0,95 М NaCl. Представленные результаты являются средними из трех повторяющихся ячеек.
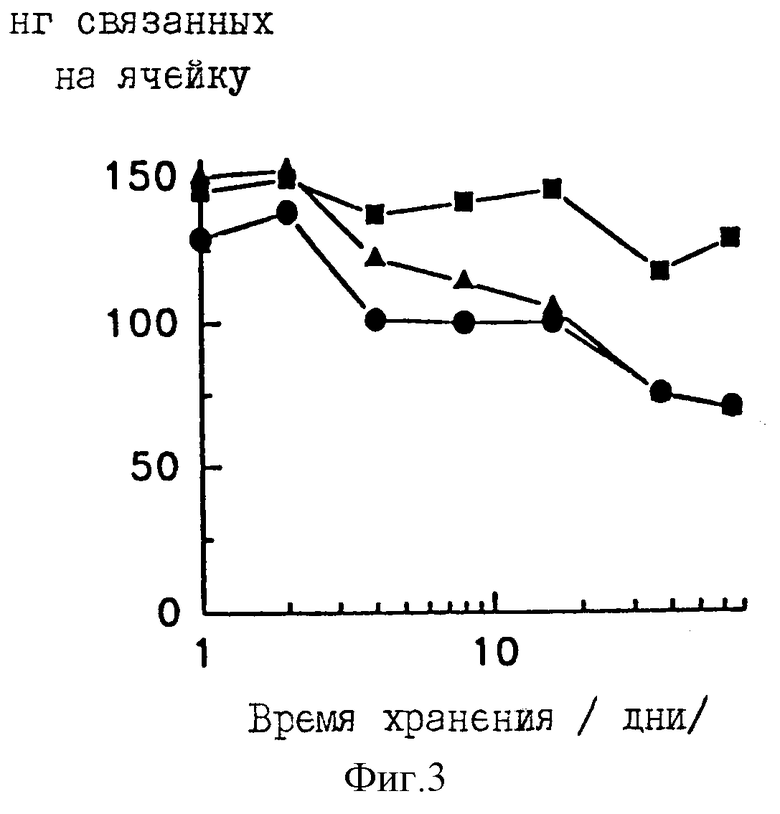
Фиг. 3 представляет влияние времени хранения и условий хранения на сохранность T-GXM на пластинах Immulon 1. Ячейки покрывают T-GXM и (a) промывают PBS-Tween и хранят сухими (кружки), (b) промывают PBS-Tween с последующей промывкой дистиллированной водой и хранят сухими (квадраты) или (c) промывают PBS-Tween и хранят с покрывающим слоем PBS-Tween (треугольники). После указанного промежутка времени все ячейки промывают PBS-Tween и определяют количество связанного T-GXM. Результаты представлены как среднее для четырех повторяющихся ячеек.
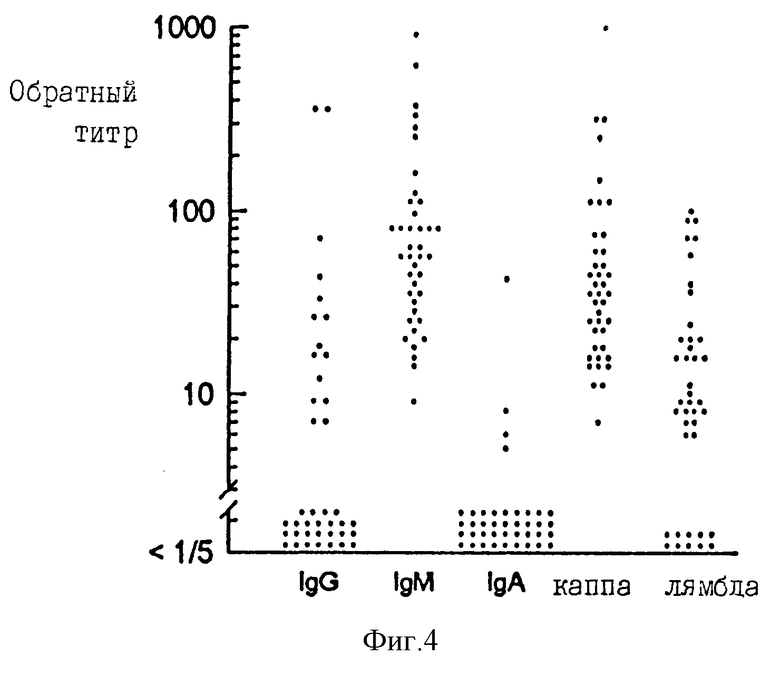
Фиг. 4 представляет АнтиGXM антитела в сыворотке от 40 здоровых взрослых доноров. Каждая точка представляет отдельный образец сыворотки, проанализированный на антитела с IgG, IgM или IgA тяжелыми цепями, или каппа, или лямбда легкими цепями.
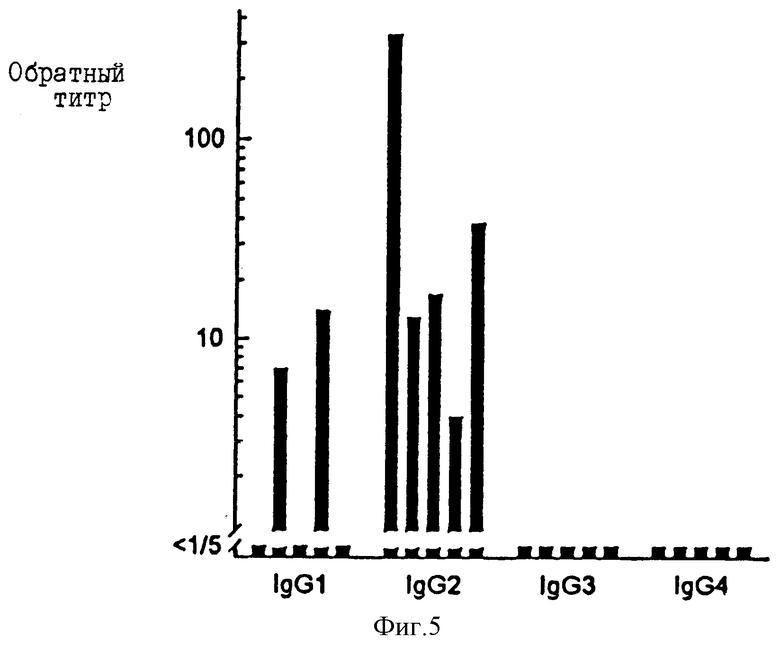
Фиг. 5 представляет анализ сыворотки здоровых людей на антиGXM антитела IgG1, IgG2, IgG3 и IgG4 подклассов. Выбранная для анализа сыворотка имеет титры антиGXM IgG антител, более или равные 1/50.
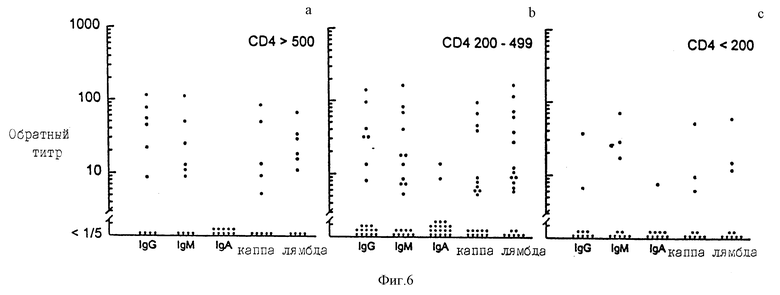
Фиг. 6(a, b, c) представляет нахождение и характеристики антиGXM антител в сыворотке, полученной на различных стадиях развития от HIV инфицирования до СПИДа. Сыворотку отбирали у 10 пациентов, которые были HIV позитивны, с СД4 > 500/мкл (рисунки слева), 20 пациентов с СД4 200-499/мкл (рисунки в центре) и 10 пациентов с СД4 < 200/мкл (рисунки справа).
Подробное описание изобретения.
В контексте настоящего изобретения термин "недостаточный иммунитет" означает такую потерю у пациентов функций T лимфоцитов, что пациенты отличаются повышенной подверженностью диссеминированному криптококкозу.
Cryptococcus neoformans представляет собой оппортунистические дрожжи, которые окружены полисахаридной капсулой. Основным структурным элементом капсулы является полисахарид, носящий название глюкуроноксиломанн (GXM), который состоит из маннозы, ксилозы, глюкуроновой кислоты и O-ацетильных групп. Антикапсулярные и/или антиGXM антитела обладают различными биологическими активностями, включая комплемент фиксации, опсонизации для фагоцитов за счет мышиных резидентных перитоничных макрофагов и нейтрофилов человека, повышением антикриптококкозной активности нативных киллерных клеток человека и ин виво очищение от криптококков за счет пассивной иммунизации.
Негативно заряженные полисахариды, такие как криптококковые GXM, плохо прилипают к полистирольным пластинам, которые обычно используют в твердофазном иммуноанализе. В известных ранее подходах для облегчения связывания полисахаридов с полистиролом и обеспечения возможности изменения антикапсулярных антител использовали: (а) пластины с предварительным покрытием поли-L-лизином, который образует множественные электростатические связи с негативно заряженными полисахаридами; (б) ковалентное связывание полисахаридов с поли-L-лизином, который, в свою очередь, связывается с полистирольной пластиной; (в) ковалентное связывание полисахаридов с тирамином, который в значительной степени повышает способность прилипания; (г) комплексообразование негативно заряженных полисахаридов с позитивно заряженными метилированными протеинами; (д) ковалентное связывание полисахаридов с закрепленными на твердой фазе протеинами или связывание полисахаридов с протеинами с последующим помещением их на полистирольные пластины; (е) биотинилирование полисахаридов с последующим связыванием с авидином покрытыми пластинами и (ж) непосредственное пассивное связывание полисахаридов под покрытием, что облегчает иммобилизацию.
Ковалентное связывание полисахаридов с реагентами, которые облегчают прикрепление, обычно включает активацию полисахарида для связи с аминосодержащими соединениями.
Процедуры активации включают (а) обработку полисахаридов цианогенбромидом для введения реакционноспособных групп, чаще всего имидокарбонатов и сложных эфиров цианата, по полисахаридным гидроксильным группам; (б) лимитированное периодатное окисление полисахаридов для создания альдегидных групп, которые можно было бы конденсировать с подходящими нуклеофилами; и (в) обработку полисахаридов хлорангидридом циануроновой кислоты.
Активация за счет бензохинона имеет существенные преимущества по сравнению с альтернативными процедурами, такими как активация цианогенбромидом, так как эту активацию можно проводить при pH ≤ 8,5. Криптококковый полисахарид снабжен O-ацетильными группами, которые легко гидролизуются при более щелочных pH, которые необходимы для активации цианогенбромидом, или при щелочных pH, создаваемых боргидридом натрия, который обычно используют, когда окисленный периодатом полисахарид связывают с аминогруппами.
Настоящее изобретение включает связывание тирамина с бензохинон-активированным полисахаридом, и определяет условия, которые благоприятствуют связыванию тирамининованного GXM (T-GXM) с полистирольными или поливинилхлоридными микротитровальными пластинами.
Настоящее изобретение иллюстрирует: (а) распределение частоты и иммуноглобулинового класса антиGXM антител среди группы здоровых взрослых добровольцев; (б) определение подкласса IgG антител, обнаруженных в сыворотке здоровых добровольцев; и (в) определение того, что развитие от HIV инфицирования до СПИДа сопровождается количественными и качественными эффектами на продуцирование антиGXM антител. В настоящем изобретении показано, что некоторые здоровые субъекты имеют высокие уровни антиGXM IgG и практически все субъекты имеют легко определяемые антиGXM IgM. Антикапсулярный IgG является преимущественно IgG2 субклассом, причем IgG1 антитела встречаются в сыворотке некоторых индивидуумов. И, наконец, развитие HIV инфекции в СПИД сопровождается селективной потерей антиGXM антител IgM класса; однако, не наблюдается совсем потерь (или незначительные потери) антиGXM антител IgG класса.
В настоящем изобретении предложен способ определения индивидуумов, у которых высок риск развития криптококкоза, который включает стадию определения уровней антител к глюкуроноксиломанну в сыворотке указанных индивидуумов. Далее предложен способ лечения индивидуумов с высоким риском развития криптококкоза, включающий стадии: определения уровней антител к глюкуроноксиломанну в сыворотке указанных индивидуумов и введение им антиCryptococcis neoformans агента.
Понятно, что способы настоящего изобретения, хотя и применимы к любому индивидууму, наиболее желательно использовать для индивидуумов с недостаточным иммунитетом и поэтому с высоким риском развития криптококкоза. Вообще говоря, способы настоящего изобретения можно осуществить для любого человека с дефицитом T-лимфоцитов или с подавленной T-лимфоцитной функцией. Одним из классов индивидуумов с недостаточным иммунитетом являются индивидуумы, инфицированные вирусом иммунодефицита человека. Дополнительную группу индивидуумов с недостаточным иммунитетом составляют пациенты с патофизиологическими состояниями, выбранными из группы, состоящей из синдрома приобретенного иммунодефицита, применения супрафизиологических доз адреналокортикостероидов, саркоидоза и лимфом.
Обычно в способах настоящего изобретения определяют уровни антител к глюкуроноксиломанну в сыворотке выбранных индивидуумов. Предпочтительно, определять антитела IgG.
В настоящем изобретении предложен также набор для определения наличия антител к глюкуроноксиломанну в сыворотке индивидуумов с недостаточным иммунитетом, содержащий твердую фазу, гидрофобный лиганд, который обеспечивает гидрофобное связывание GXM с твердой фазой, энзимную индикаторную систему и субстрат, подходящий для связанного с энзимом антитела.
При использовании набора, предлагаемого в настоящем изобретении, криптококковый глюкуроноксиломанн иммобилизуют на твердой фазе так, чтобы GXM был прочно связан с твердой фазой без модификации, блокирования или деградации эпитопов распознаваемых антикриптококковым антителом. Обычно твердой фазой может быть любая фаза, которая связывает GXM. Предпочтительно, чтобы твердой фазой были полистирольные или поливинилхлоридные микротитровальные пластины; альтернативными твердыми фазами служат нитроцеллюлоза или другие формы бумаги, магнитные шарики или пластиковые погружаемые стержни.
Предпочтительным способом иммобилизации является связывание GXM с гидрофобным лигандом, который обеспечивает гидрофобное связывание GXM с твердой фазой. Гидрофобный лиганд не должен распознаваться ни одним из антител, которые находятся в сыворотке человека. Предпочтительным лигандом является тирамин. Связывание GXM с гидрофобным лигандом должно быть ковалентным и осуществлено таким образом, который не позволяет происходить гидролизу чувствительных к щелочи O-ацетильных боковых цепей. Оптимальные концентрации антигена, оптимальные pH для покрытия микротитровальных пластин и оптимальные температуры для покрытий станут очевидны специалистам после выяснения преимуществ в описании настоящего изобретения.
Набор настоящего изобретения включает также энзимную индикаторную систему, такую как антитела, специфичные для тяжелых цепей IgG человека. Предпочтительно, чтобы в качестве антител были использованы меченые пероксидазой хрена козьи или овечьи антитела. Другие энзимные индикаторные системы также хорошо известны специалистам. Обычно в набор включен субстрат, подходящий для связанных с энзимом антител. Представительным примером такого субстрата может служить 3,3',5,5'-тетраметилбензидин и перекись водорода (TMB Microwell Peroxide Substrate System). Специалистам также должны быть очевидны возможные альтернативы.
Настоящее изобретение предлагает далее способ определения антител к глюкуроноксиломанну в сыворотке, включающий стадии: инкубирование микротитровальной пластины с ячейками, покрытыми антигеном, в течение 90 минут при 25oC с сывороткой, разбавленной в смеси PBS-Tween; удаление сыворотки пациентов из ячеек и промывание указанных ячеек смесью PBS-Tween для удаления несвязанных антител; инкубирование промытых ячеек в течение 30 минут при 25oC с разбавленными коньюгированными с энзимом антителами, специфичными для IgG человека; удаление коньюгированных с энзимом антител из ячеек за счет промывания смесью PBS-Tween; добавление субстрата, соответствующего коньюгированным с энзимом антителам, и инкубирование в течение 30 минут при 25oC; добавление останавливающего реакцию раствора и определение оптической плотности энзимного продукта.
В качестве контроля за неспецифическим связыванием аликвоту сыворотки человека предварительно инкубируют с 4 мкг/мл GXM перед инкубированием с антигеном покрытыми ячейками. Антигеном нейтрализованную сыворотку пациентов инкубируют затем с антигеном, иммобилизованным на твердой фазе описанным ранее способом.
Приводимые далее примеры служат лишь для иллюстрации различных вариантов изобретения и никоим образом не ограничивают его объем.
Пример 1
Криптококковый глюкуроноксиломанн
Криптококковый GXM выделяют с помощью модифицированной процедуры описанной Cherniak et. al. Infect. Immun. 59:59 (1991). Короче, C. neoformans серотипа A штамм 24064 (Американская коллекция типовых культур, Rockville, MD) выращивают в жидкой синтетической среде во вращающемся шейкере при 100 об/мин в течение 4 дней при 30oC. Дрожжевые клетки убивают, добавляя формальдегид до окончательной концентрации 1%. Добавляют кристаллы ацетата натрия и ледяную уксусную кислоту до конечной концентрации 10% и 1% соответственно, и дрожжевые клетки удаляют центрифугированием при 10000 g. Полисахарид осаждают из надосадочной жидкости, добавляя 2,5 объема 95% этанола, и осадок собирают центрифугированием. Полисахаридный осадок растворяют до концентрации примерно 5 мг/мл в 0,2М NaCl и полученный раствор осветляют за счет центрифугирования. Добавляют гексадецилтриметиламмонийбромид (CTAB) 3 мг CTAB на мг полисахарида и при перемешивании медленно добавляют два объема 0,05% CTAB. Комплекс GXM-CTAB выделяют центрифугированием и снова растворяют до примерно 5 мг/мл в 0,2М NaCl. Полисахарид осаждают этанолом и CTAB, как было указано ранее. И, наконец, полисахарид осаждают этанолом, диализуют против деионизированной воды и лиофилизируют.
Пример 2
Присоединение тирамина к GXM
GXM ковалентно связывают с тирамином, используя модификации процедуры активации бензохиноном, описанные Kozel and Hermerath J. Immunol. Meth. 107: 53 /1988/ и Infect. Immun. 43:879 (1984). GXM растворяют в деионизированной воде (10 мг/мл) и смешивают с равным объемом 0,2 М бикарбоната натрия (pH 8,5). Добавляют бензохинон (0,25 объема, 250 мМ в абсолютном этаноле) до получения конечной концентрации 50 мМ бензохинона в 20% этаноле и инкубируют с GXM в течение 60 минут при комнатной температуре. pH не должно превышать 8,5 или время инкубирования не должно быть более 60 минут, так как O-ацетильные группы на полисахариде могут быть потеряны за счет щелочного гидролиза. Кристаллы ацетата натрия (10% вес/объем) и ледяную уксусную кислоту (1% объем/объем) добавляют и GXM осаждают, добавляя два объема 95% этанола. Осадок собирают центрифугированием, растворяют в ацетатном буфере (10% кристаллы ацетата натрия и 1% ледяная уксусная кислота) при концентрации примерно 2 мг GXM/мл и осаждают, добавляя два объема этанола. Осадок собирают центрифугированием и растворяют до концентрации примерно 4 мг/GXM/мл в 0,1М натрийфосфате (pH 7,5), содержащем 0,5М NaCl. GXM диализуют против того же буфера и против 0,1М натрийфосфата (pH 7,5).
Активированный бензохиноном GXM связывают с тирамином за счет смешивания активированного полисахарида (примерно 2 мг/мл) с равным объемом тирамингидрохлорида (1 мг/мл в 0,1 М натрийфосфата pH 7,5) и полученную смесь инкубируют в течение ночи при комнатной температуре. Как указано ранее, добавляют кристаллы ацетата натрия и ледяную уксусную кислоту и обработанный тирамином GXM осаждают, добавляя два объема 95% этанола. Осадок собирают центрифугированием, растворяют до концентрации примерно 4 мг/мл в 0,1 М натрийфосфате (pH 7,5), содержащем 0,5 М NaCl, и диализуют последовательно против 0,1 М натрий фосфата (pH 7,5), содержащего 0,5 М NaCl и 0,1М натрийфосфата (pH 7,5). Концентрацию GXM определяют в анализе с фенол-серной кислотой, и связанный с тирамином GXM разбавляют до 2 мг/мл, стерилизуют фильтрованием и хранят при 4oC.
Степень тираминирования GXM определяют, измеряя оптическую плотность на 280 нм, используя тирамингидрохлорид в качестве стандарта и активированных бензохиноном полисахарид в качестве слепого опыта. Примерно 30 молей тирамина связывается с каждым молем GXM в предположении, что молекулярный вес криптококкового GXM составляет 500000.
Пример 3
Анализ связывания T-GXM со стирольными пластинами
Тираминированные GXM метят 125I с помощью Iodogen /Pierce Chemical Co., Rockford, I11/ процедуры. Радиомеченые GXM выделяют от свободного иода за счет фильтрации через Sephadex G-25. Обычно 0,4 мг GXM метят до специфической активности 3,0 • 106 имп. мин/мкг. 125I-меченый GXM смешивают с немеченым GXM до специфической активности 2,5 • 105 имп. мин/мкг GXM.
Условия инкубирования, используемые для связывания
T-GXM с пластиной варьируются в соответствии с экспериментальным протоколом. После стадии нанесения покрытия пластины промывают шесть раз смесью PBS-Tween (8 г NaCl, 0,2 г KH2PO4, 2,9 г Na2HPO4Σ12H2O, 0,2 г KCl и 0,5 мл Tween 20 на литр деионизированной H2O). Ячейки разделяют и количество связанного GXM определяют с помощью счетчика гамма-излучения. Статистическое сравнение количества связанного GXM в различных условиях определяют с помощью т-теста, используя p<0,05 в качестве критерия статистической значимости.
Пример 4
Оптимальные условия для иммобилизации T-GXM
Чтобы проиллюстрировать важную роль pH и ионных сил для связывания T-GXM с пластинами Imnulon 1, T-GXM приготавливают в концентрациях в интервале от 625 до 20000 нг/мл в (а) 0,05М карбонатном буфере, pH 9,6; (б) 0,05М натрийфосфатном буфере, pH 7,0 с 10 мМ ЕДТА; или (в) 0,05М цитратном буфере, pH 4,5. В качестве кроющих буферов используют: (а) неизмененный, (б) с добавлением NaCl до 0,1М или (в) с добавлением NaCl до 0,95 М. Пластины инкубируют с T-GXM в течение ночи при комнатной температуре. Ячейки промывают для удаления несвязанного T-GXM и определяют количество адсорбированного T-GXM. Данные рассчитывают как нанограммы связанного T-GXM на ячейку и как процент добавленного GXM, который оказался связанным с ячейкой.
Фиг. 1 представляет, что значительные количества GXM оказываются связанными при введении либо в карбонатный, либо в фосфатный буферы. Максимальное связывание в карбонатном и фосфатном буферах составляет примерно 100-150 нг/ячейку. В случае цитратного буфера наблюдается значительное уменьшение связывания, причем максимум наблюдаемого связывания составляет примерно 50 нг/ячейку. Введение NaCl до 0,1 М существенно повышает связывание, которое наблюдается для фосфатного буфера, по сравнению со связыванием для того же буфера, но без добавления NaCl. Аналогично, значительно большее связывание наблюдается для фосфатного буфера с 0,95 М NaCl по сравнению с буфером, в который добавлено 0,1 М NaCl. Повышение ионной силы не оказывает существенного усиливающего эффекта на связывание в карбонатном или цитратном буферах.
Пример 5
Временные и температурные требования
Влияние времени покрытия и температуры также были исследованы. Ячейки на Immulon 1 пластинах инкубируют со 100 мМКл 10 мкг/мл радиомеченого T-GXM в фосфатном кроющем буфере, содержащем 0,95 М NaCl. Пластины инкубируют в течение 1, 2, 4, 8, 24 или 48 часов при 4oC, 20oC или 37oC. Пластины промывают и определяют количество связанного T-GXM.
Фиг. 2 демонстрирует, что не существует значительных различий в уровнях связывания при 20oC или 37oC для любого времени, которое было исследовано. Инкубирование пластин в течение 48 часов при 20oC или 37oC не приводит к заметному дополнительному связыванию по сравнению с тем, которое наблюдается спустя 24 часа. Инкубирование пластин при 4oC обеспечивает значительно меньшее связывание, нежели то, которое наблюдается, если пластины покрывают при 20oC или 37oC для всех периодов времени инкубирования, равных или более 2 часов. На этом основании затем используют покрытия пластин T-GXM в течение 24 часов при 20oC.
График 1 (A/A0) от времени (фиг. 2), где A представляет вакантную связывающую способность в любой момент времени, а A0 представляет полную связывающую способность (определяемую здесь как количество связанного T-GXM при инкубировании в течение 48 часов при 20oC или 37oC) оказывается линейным для пластин, покрытых при 0oC, 20oC или 37oC. Это дает возможность предположить, что связывание происходит с константной скоростью первого порядка, характерной для каждой из температур. Связывание оказывается гораздо более медленным при 4oC (константа скорости псевдо первого порядка ≈ 3,2 • 10-4), нежели при 20oC (константа скорости псевдопервого порядка ≈ 2,7 • 10-3) или 37oC (константа скорости псевдопервого порядка ≈ 2,6 • 10-3).
Пример 6
Стабильность T-GXM иммобилизации на полистирольных пластинах
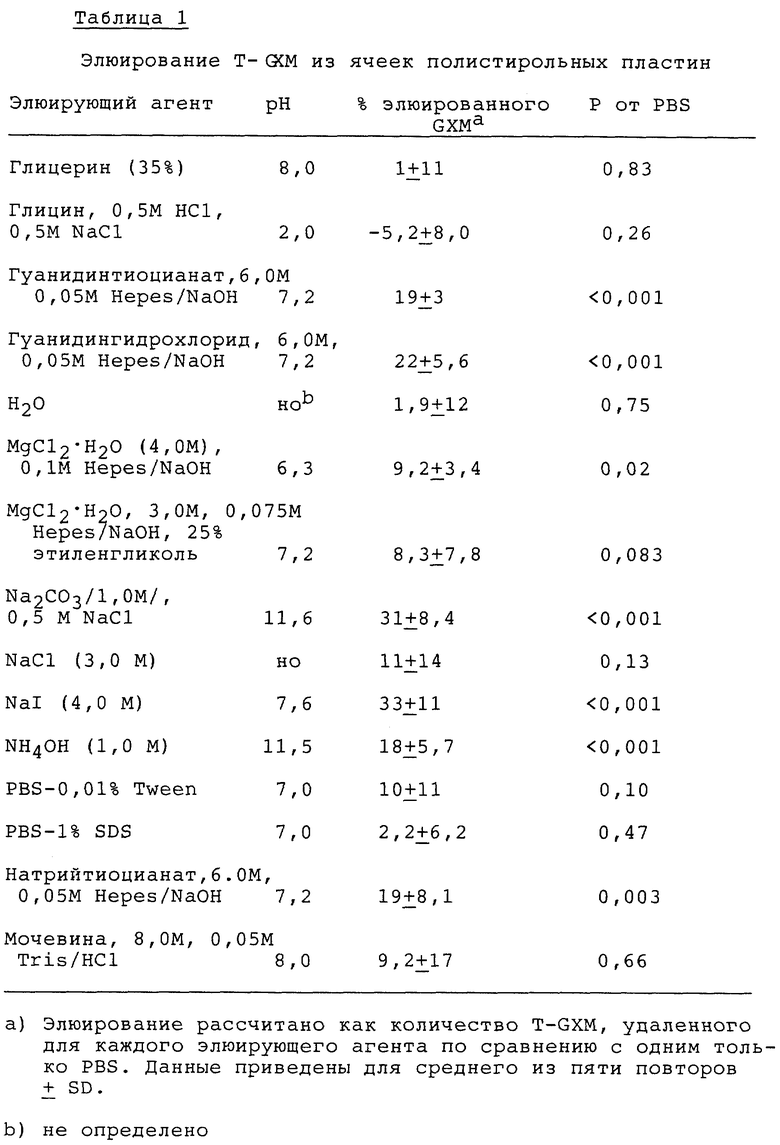
В попытках оценить стабильность связывания T-GXM с пластинами Immulon 1 были исследованы различные элюирующие и диссоциирующие агенты. Ячейки покрывают в течение 24 часов при комнатной температуре 100 мкл 10 мкг/мл радиомеченого T-GXM в фосфатном кроющем буфере, содержащем 0,95М NaCl. Ячейки промывают шесть раз смесью PBS-Tween и в каждую ячейку добавляют 100 мкл элюирующего агента и инкубируют в течение 1 часа при 20oC. Ячейки промывают шесть раз смесью PBS-Tween, и определяют количество связанного тираминированного GXM. Полученные результаты представляют как процент удаленных тираминированных GXM в отношении ячеек, обработанных PBS.
Таблица 1 демонстрирует, что связанный тираминированный GXM значительно устойчив к элюированию большинством диссоциирующих агентов. Значительное элюирование наблюдается для случая, когда ячейки были обработаны щелочными буферами или различными солями. Однако, количество T-GXM, элюированного из пластин, никогда не превышает 35% по сравнению с величинами, наблюдаемыми для пластин, инкубируемых с PBS.
Стабильность иммобилизованного T-GXM определяют в различных условиях хранения. Ячейки полистирольных пластин покрывают на 24 часа при комнатной температуре 100 мкл 10 мкг/мл радиомеченого T-GXM в фосфатном кроющем буфере, содержащем 0,95 М NaCl. Ячейки промывают смесью PBS-Tween и (i) PBS-Tween удаляют, а пластины хранят сухими, (ii) пластины промывают еще два раза дистиллированной водой и хранят сухими или (iii) пластины хранят с нанесенным слоем 100 мкл PBS-Tween. Четыре ячейки удаляют в различные моменты времени, промывают PBS-Tween и определяют количество связанного тираминированного полисахарида. Наблюдается значительная потеря T-GXM для пластин, которые либо хранили сухими после промывки одним PBS-Tween (срок полужизни примерно 46 дней), либо хранили с нанесенным слоем PBS-Tween (срок полужизни примерно 47 дней). Удаление PBS-Tween за счет промывки дистиллированной водой обеспечивает хранение сухих пластин при незначительных потерях T-GXM после 64 дней хранения (срок полужизни оценен как превышающий 1000 дней).
Активирование капсулярных полисахаридов для связывания с тирамином осуществляют, используя CNBr, хлорангидрид циануроновой кислотой и окисление периодатом. Настоящее изобретение демонстрирует, что активация бензохиноном также является эффективным средством для тираминирования полисахаридов для использования в иммунологических анализах. Существует несколько характеристик бензохинона, связанных с процедурой активации. Во-первых, активацию бензохиноном осуществляют при более низких значениях pH (≤ 8,5), нежели те, которые необходимы для альтернативных способов. Это особенно важно для криптококковых полисахаридов, так как O-ацетилирование является важной компонентой антигенного характера GXM. O-ацетильные группы легко можно утратить у полисахарида при pH более 8,5. Во-вторых, активация бензохиноном вводит хромофор в полисахарид, который облегчает контроль за процессом активации.
Количество T-GXM, связывающегося с полистирольными пластинами, сопоставимо со связыванием, наблюдаемым с протеинами. Насыщение сайтов связывания за счет T-GXM, покрытого фосфатным буфером, содержащим 0,95 М NaCl, начинает проявляться при 50-100 нг/см2. Различие в результатах, полученных с протеинами и T-GXM, легко объяснить за счет высокой степени гидратации GXM. Тот факт, что полисахариды являются сильно гидратированными, предсказывает, что для заполнения всех доступных сайтов понадобится меньшее количество полисахарида.
Важным фактором, влияющим на эффективность связывания T-GXM с полистиролом, может оказаться температура. Константа скорости псевдопервого порядка для связывания при 4oC примерно в 8 раз меньше, чем константы для связывания при 20oC или 37oC.
Настоящее изобретение иллюстрирует, что T-GXM связывается с полистиролом достаточно прочно. Немногие элюирующие агенты оказались способными сколько-нибудь существенно удалять T-GXM с пластин. Прочность связывания обеспечивает длительные сроки хранения пластин при незначительных потерях связанного T-GXM. Введение тираминовой группы без сомнения делает молекулы более гидрофобными с сопровождающим усилением притяжения к полистиролу. Однако, при примерно 30 молях тирамина на моль GXM поливалентный характер связывания за счет отдельных GXM молекул, вероятно, является значительным фактором в прочности связывания. В отличие от протеина, в котором многие из гидрофобных фрагментов расположены внутри молекулы, по-видимому, растянутый характер GXM позволяет большинству, если не всем, тираминовым группам связываться с поверхностью полистирола.
Пример 7
Криптококковые клетки и GXM
GXM выделяют, как это указано в примерах 1 и 2. GXM ковалентно связывают с тирамином, как это указано в примере 2.
Бензохиноном активированный GXM связывают с тирамином за счет смешивания активированного полисахарида (примерно 4 мг/мл) с равным объемом тирамингидрохлорида (1 мг/мл в 0,1 М натрийфосфате, pH 7,5) и инкубируют в течение ночи при комнатной температуре. Как указано ранее, добавляют кристаллы ацетата натрия и ледяную уксусную кислоту, и связанный с тирамином GXM осаждают, добавляя два объема 95% этанола. Осадок собирают центрифугированием, растворяют до концентрации примерно 4 мг/мл в 0,1 М натрийфосфате (pH 7,5), содержащем 0,5 М NaCl, и диализуют последовательно против 0,1 М натрийфосфата (pH 7,5), содержащего 0,5 М NaCl, и 0,1 М натрийфосфата (pH 7,5).
Концентрацию GXM определяют в анализе с фенол-серной кислотой по способу Dubois, et. al. Anal. Chem. 28:350-356 (1956), и связанный с тирамином GXM разбавляют до 2 мг/мл, стерилизуют за счет фильтрования и хранят при 4oC.
Пример 8
ELISA анализ для антикриптококковых антител
Ячейки микротитровальных пластин Immulon I (Dynatech Laboratories Inc., Chantilly, Va) покрывают на ночь при 4oC 100 нг тираминированного GXM (T-GXM) в 100 мкл кроющего буфера (0,05 М натрийфосфат, pH 7,4, содержащий 10 мМ ЕДТА). Ячейки трижды промывают блокирующим буфером (0,05 М натрийфосфат, pH 7,4) и блокируют в течение 90 минут 300 мкл 0.05% Tween20 в блокирующем буфере. Пластины, покрытые тираминированным GXM, трижды промывают PBS-Tween (PBS, содержащий 0,05% Tween 20) и инкубируют в течение 90 минут с сериальными двукратными разбавлениями сыворотки человека в PBS-Tween, начиная с разбавления 1:5. В качестве негативного контроля все сыворотки разбавляют в PBS-Tween, содержащем 4 мкг GXM на мл, и инкубируют в течение 30 минут перед добавлением в ячейки, покрытые тираминированным GXM (GXM нейтрализованная сыворотка). После инкубирования с сывороткой или GXM-нейтрализованной сывороткой ячейки промывают трижды PBS-Tween и инкубируют в течение 90 минут при комнатной температуре со вторым антителом, меченным пероксидазой хрена (HRPO).
Вторыми антителами были HRPO-меченые, афинно-очищенные козлиные античеловеческие IgG тяжелые цепи (1/16000; Southern Biotechnology, N 2040-05), HRPO-меченые афинно- очищенные козлиные античеловеческие IgM тяжелые цепи (1/15000; там же кат. N 2020-05), HRPO-меченые афинно-очищенные козлиные античеловеческие IgA тяжелые цепи (1/12000; там же кат. N 2050-05), HRPO-меченые афинно-очищенные козлиные античеловеческие каппа (1/10000; там же кат. N 2060-05), HRPO-меченые, афинно очищенные античеловеческие лямбда (1/5000; там же кат. N 2070-05). Второе антитело используют в восьмикратном разбавлении от антитела, дающего OD450 = 2,5 для ячеек, покрытых очищенным IgG (антиIgG, антикаппа или антилямбда), IgM (антиIgM), или IgA (антиIgA). Пластины промывают трижды PBS-Tween и инкубируют в течение 30 минут со 100 мкл TMB Microwell Peroxidase Substrate Solution (Kirkegaard and Perry Labs, Inc. , Gaithersburg, MD). Реакцию останавливают, добавляя 100 мкл 1 М H3PO4. Считывают поглощение в течение 30 минут на 450 нм с помощью рабочей станции Ceres 900 энзимного иммуносорбентного анализа (Bio-Tek Instruments, Inc., Winooski, VT). Все ELISA анализы осуществляют в дубликате, включая GXM-нейтрализованный контроль. Рассчитывают среднее из дублируемых ячеек и характеристическое OD450 рассчитывают, вычитая RD450, полученное для GXM-нейтрализованной сыворотки из OD450 полученных для случаев, когда сыворотку не инкубировали предварительно с GXM. Полученные данные обрабатывают с помощью Kineticafc EIC Application Software Version 2.12 (Bio-Tek Instrupents Inc.) в титровальном режиме. Наилучшее соответствие с результатами получено для 4-параметрального алгоритма, и порог чувствительности устанавливают на 0,5. В этих условиях разбавление сыворотки, дающее OD450, с поправкой на фон (GXM-нейтрализованный) порядка 0,5 берут за конечную точку. Сыворотку с OD450 менее 0,5 при разбавлении сыворотки 1/5 рассматривают как негативную. Используя мышиные моноклональные IgG1 антитела в качестве стандарта, оказывается, что примерно 500 пг антител дает OD450 порядка 0,5.
Описанный ранее анализ ELISA был модифицирован для определения IgG субкласса антиGXM IgG антител. В этом модифицированном анализе пластины покрывают GXM и инкубируют при разбавлениях сыворотки или в сыворотке, разбавленной PBS-Tween, содержащим GXM, как было раскрыто ранее. Ячейки промывают и инкубируют в течение 90 минут при комнатной температуре с оптимально разбавленными мышиными моноклональными антителами, специфичными для человеческого IgG1 (1/2000; Calbiochem, La Jolla; кат. N 411451), IgG2 (1/8000; там же кат. N 411461), IgG3 (1/40000, там же кат. N 411481) или IgG4 (1/1500, там же N 411492). Затем пластины промывают PBS-Tween, инкубируют со 100 мкл пероксидазой меченого козлиного антимышиного IgG (1/4500, Southern Biotechnology Assoc. , Inc. ; кат. N 1030-05), промывают и инкубируют с субстратом, как было указано ранее. Результаты приводят как разбавления сыворотки с поправкой на фон (GXM-нейтрализованный), который дают OD450 = 0,5.
Пример 9
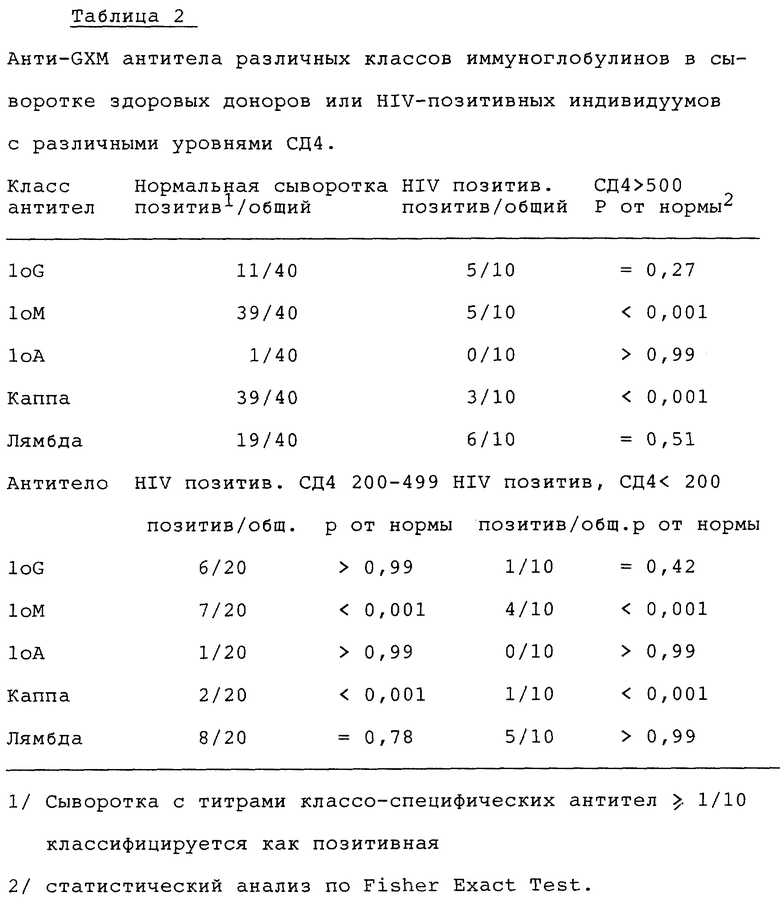
Существование и класс иммуноглобулинов антиGXM антител в сыворотке здоровых доноров
Сыворотку исследуют для определения уровней антител IgG, IgM и IgA классов. Сыворотку получают от 40 здоровых взрослых добровольцев. Эту сыворотку исследуют также для оценки присутствия и уровней антител с каппа или лямбда легкими цепями. Фиг. 4 демонстрирует, что 28% сывороток содержит IgG антитела с титрами ≥ 1/10, 98% имеет IgM антитела с титрами ≥ 1/10 и только в одном образце сыворотки IgA антитела наблюдаются с титрами ≥ 1/10. 98% содержат антитела с каппа цепями с титрами ≥ 1/10 и 45% имеют антитела с лямбда цепями с титрами ≥ 1/10.
Пять образцов сыворотки с IgG антител титрами ≥ 1/50 исследуют далее для определения IgG субкласса антиGXM-IgG. Результаты показывают (фиг. 5), что каждая из этих сывороток содержит детектируемые уровни антиGXM антител IgG2 субкласса. Две из сывороток содержат также детектируемые уровни IgG1 антител. Ни в одной из сывороток не содержатся детектируемые уровни антиGXM антител IgG3 или IgG4 субкласса.
Пример 10
Характеристики антиGXM антител в сыворотке HIV-позитивных доноров
Частоту нахождения и уровни антиGXM антител с различными изотипами тяжелых и легких цепей в сыворотке пациентов исследуют для случаев: (а) HIV- позитивные с СД4 ≥ 500 клеток/мл, (б) HIV-позитивные с СД4 от 200 до 499 клеток/мл и (в) HIV-позитивные с СД4 менее 200 клеток/мл. Позитивные титры определяют как титры антител ≥ 1/10.
На фиг. 6 представлены результаты анализа сыворотки от HIV-позитивных доноров. Сравнение частоты позитивных титров среди здоровых доноров (фиг. 4) и HIV-позитивных доноров (фиг. 6) представлено в таблице 2. Менее часто встречается антиGXM IgG у пациентов с уровнями СД4 ниже 200. Однако, это различие не было значительным различием. АнтиGXM IgM класса встречаются значительно более часто среди здоровых субъектов, нежели среди HIV-позитивных доноров, независимо от СД4 уровней. Аналогично, антиGXM антитела с каппа легкими цепями значительно менее часто встречаются среди HIV-позитивных доноров, нежели среди здоровых субъектов. Возможно, что другие различия можно было бы также заметить, если бы сравнивать реальные уровни антител, а не частоту нахождения субъектов с титрами ≥ 1/10.
Индивидуальные титры различных групп субъектов сравнивали в тесте: Mann-Whitney (rank sum test). За одним исключением значимые различия, определенные в этом тесте, были точно такими же, что и найденные в случае, когда данные оценивали в единицах частоты позитивных титров. Исключение составляет значительное (p ≤ 0,01) снижение (по сравнению со здоровыми субъектами) титров антител с лямбда легкими цепями, обнаруженными в сыворотке HIV-позитивных пациентов с СД4 уровнями ниже 200.
Пример 11
Ретроспективный анализ сыворотки пациентов со СПИДом
В настоящем изобретении предложен высокочувствительный анализ для определения антител реакционноспособных по отношению к основному капсулярному полисахариду Cryptococcus neoformans. Способ настоящего изобретения позволяет определить наличие латентной инфекции, так как эти дрожжевые уровни иммунологических "отпечатков" в форме IgG антител, которые реакционноспособны в отношении полисахаридов. Так, присутствие антител сигнализирует о наличии латентной криптококковой инфекции и поэтому о высоком риске криптококкоза для пациентов со СПИДом.
Анализ антител
Сыворотку оценивают на наличие антител к глюкуроноксиломанну, основному серотипически-специфическому полисахариду C. neoformans. Этот полисахарид является основным капсулярным полисахаридом, который окружает дрожжи. Антитела к GXM оценивают в энзимном иммуносорбентном анализе, в котором GXM иммобилизован на стандартных микротитровальных пластинах. Иммобилизация требует химической модификации GXM таким способом, который сохраняет антигенную структуру полисахарида, при этом повышая гидрофобность, необходимую для прочного прикрепления к пластинам. В этом анализе используют процедуру бензохинонового связывания, которая обеспечивает связывание тирамина с GXM. Полисахарид, модифицированный таким образом, связывается с полистирольными и поливинилхлоридными пластинами с достаточной прочностью.
Анализ сыворотки пациентов
Сыворотку получают и подразделяют на две категории: (1) HIV-инфицированные пациенты, у которых развился или (2) не развился криптококкоз после того, как HIV-инфекция развилась до СПИДа. IgG антитела определяют со значительно большей частотой в группе пациентов, у которых развился криптококкоз, нежели в группе пациентов, у которых криптококкоз не развился.
Настоящее изобретение обеспечивает новую информацию относительно антиGXM антител, находящихся в сыворотке, полученной от здоровых взрослых доноров. Частота нахождения антикриптококковых IgM антител в сыворотке составляет около 98% у здоровых доноров, тогда как IgG антитела встречаются с частотой около 28% у здоровых доноров. Настоящее изобретение иллюстрирует резкое снижение нахождения антиGXM антител IgM класса у всех категорий HIV-позитивных пациентов. Аналогичное уменьшение наблюдается для частоты нахождения антител с каппа легкими цепями. Напротив, не наблюдается уменьшения процента HIV-позитивных пациентов, вырабатывающих антитела с лямбда легкими цепями. Значительное снижение нахождения IgG антител не было обнаружено у пациентов с различными стадиями HIV-инфекции. Это указывает на то, что IgG антитела-маркеры для определения риска криптококкоза не утрачивают своего значения, когда иммунная система нарушается при развитии до СПИДа.
Как было обнаружено ранее, снижение реакций на тип 2 T-независимые антигены происходит на относительно ранних стадиях развития HIV-инфекции. Более ранние исследования иммунной реакции на пневмококковые вакцины обнаружили ослабление IgG2, IgM и IgA реакции у HIV позитивных пациентов с персистентной генерализованной лимфаденопатией; однако, не происходит снижения IgG1 реакций. Таким образом, уменьшение количества антиGXM IgG антител может быть связано с селективной утратой IgG2. Это должно было бы согласовываться с нахождением IgG1 в сыворотке от двух из пяти здоровых добровольцев, сыворотку которых подвергали исследованию.
Содержание антигенных стимулов для антиGXM антител в сыворотке здоровых людей неизвестно. Эти антитела не могут быть связаны со стимуляцией перекрестно-реактивных антигенов, которые находятся в окружении. "Природные" антитела, которые реакционно способны в отношении нескольких бактериальных капсул, как считают, связаны с экспонированием перекрестно-реагирующим антигенам непатогенных энтерических и назофарингеальных организмов. Глюкуроновая кислота и O-ацетильные группы являются общими компонентами многих полисахаридов. Так, возможно, что повсеместные антиGXM IgM антитела реагируют с одним из этих компонентов. С другой стороны, ксилоза не является характеристически обнаруживаемой в бактериальных капсулах, хотя она обычно встречается среди растений. IgG антиGXM антитела могут предоставлять поверхность для криптококков либо в качестве природной либо в качестве субклонированной инфекции. Так, антиGXM IgG может быть маркером для латентной криптококковой инфекции и повышать риск криптококкоза у HIV-инфицированных пациентов или пациентов с недостаточным иммунитетом за счет других причин.
Все патенты и публикации, упомянутые здесь, являются указаниями уровней для специалистов, к которым это изобретение имеет отношение. Все патенты и публикации включены по ссылкам.
Специалистам легко понять, что настоящее изобретение хорошо адаптировано для осуществления его целей и конечных результатов и упомянутых преимуществ, а также тех, которые ему присущи. Приведенные примеры наряду со способами, процедурами, обработками, молекулами и конкретными соединениями, описанные здесь, являются представительными для предпочтительных вариантов и приведены в качестве примеров и никоим образом не ограничивают объем изобретения. Все изменения и другие применения, которые очевидны специалистам, также входят в объем изобретения, определенный объемом прилагаемой формулы изобретения.
Изобретение предназначено для скрининга индивидуумов с высоким риском развития криптококкоза. Способ включает стадию определения уровней IgG антител к глюкуроноксиломанну (GXM) в сыворотке указанных индивидуумов. Повышенные уровни антител указывают на наличие воздействия или скрытой инфекции. Титры антител к глюкуроноксиломанну более 1:10 свидетельствуют о высоком риске заражения криптококкозом. Для реализации способа используют набор для определения наличия антител к глюкуроноксиломанну в сыворотке индивидуумов с недостаточным иммунитетом. Набор содержит твердую фазу, гидрофобный лиганд, энзимную индикаторную систему и субстрат. Лиганд обеспечивает гидрофобное связывание GXM с твердой фазой. Изoбретение позволит своевременно принять профилактические меры против заражения криптококкозом. 2 с. и 4 з.п. ф-лы, 2 табл., 14 ил.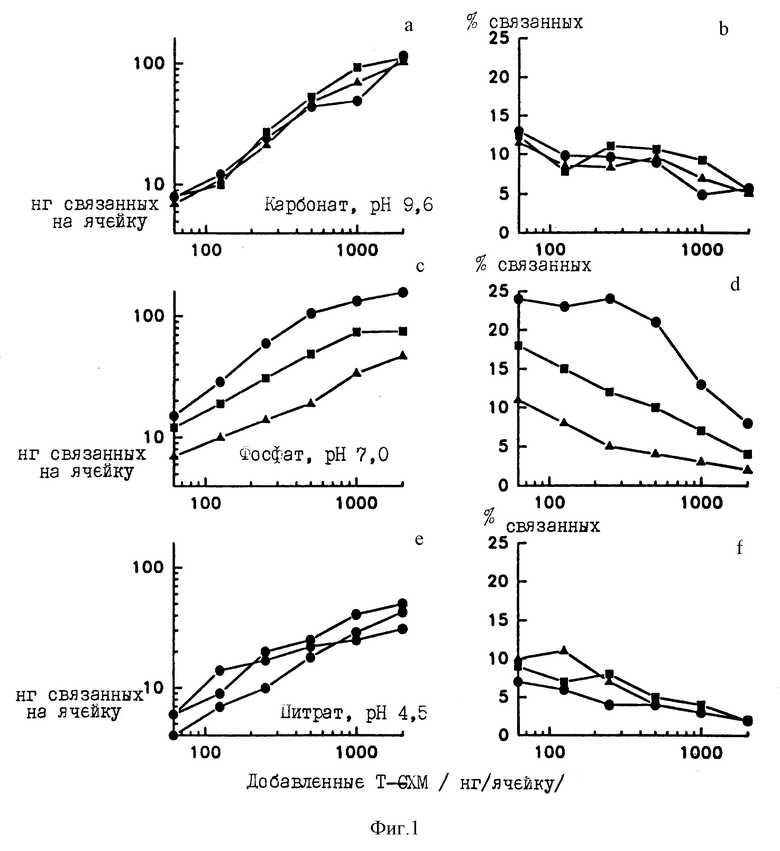
| Todaro - Luck et al | |||
| "Characterization of Cryptococcus neoformans capsular polysaccharride with monoclonal antibodies" // Infect and Immun., 1989, v.57, N 12, p.3882-3887 | |||
| Reiss et al | |||
| "Enzyme immunoassay detection of IgM to galactoxylomannan of Cryprococcus neoformans" // Diagn | |||
| Immunol., 1984, v.2, N 2, p.109-115 | |||
| Dromer et al | |||
| Cryptococcus neoformans antibody levels in patients with AIDS" // Scand | |||
| Journal | |||
| Infect | |||
| Disease, 1988, v.20, N 3, p.283-285. |

