Настоящее изобретение относится к новому фактору роста/дифференциации TGF- β -семейства и кодирующим его ДНК-последовательностям.
TGF- β -семейство факторов роста, к которому относятся родственные BMP, TGF и ингибину протеины /Poberts и Sporn, Handbook of Experimental Pharmacology, 95 (1990), 419-472/, особенно уместно для широкой области медицинских методов лечения и применений. Эти факторы пригодны в способах, которые относятся к заживлению ран и регенерации тканей. Далее, некоторые члены TGF- β -семейства индуцируют рост тканей, в особенности рост костей, и поэтому играют центральную роль при индуцировании развития хрящей и костей.
Wozney /Progress in Growth Fact or Research, 1, (1989), 267-280 и Vale и др. / Handbook of Experimental Pharmacology, 95 (1990), 211-248/ описывают различные факторы роста, как, например, таковые, которые являются родственными группе BMP (костно-морфогенетические протеины) и группе ингибина. Члены этих групп обладают значительным структурным сходством. Предшественник протеина состоит из аминоконцевой сигнальной последовательности, пропептид- и карбоксиконцевой последовательности примерно из 110 аминокислот, которая отщепляется от предшественника и представляет собой зрелый протеин. Далее, ее члены определяются путем гомологии аминокислотных последовательностей. Зрелый протеин содержит высококонсервативные последовательности, в особенности семь цистеиновых остатков, которые консервативны у членов семейства. TGF- β -образные протеины представляют собой многофункциональные, гормонально активные факторы роста. Они обладают также родственными биологическими активностями, как, например, хемотактическое притяжение клеток, способствование дифференциации клеток и индуцирующие ткани способности, как, например, индуцирующие хрящи и кости способности. В патенте США N 5013649 раскрыты ДНК-последовательности, которые кодируют остеоиндуктивные протеины, обозначаемые как BMP-2, а из патентных заявок США N 179101 и N 170197 известны BMP-протеины: BMP-1 и BMP-3. Далее, многие типы клеток в состоянии синтезировать TGF- β -образные протеины, и практически все клетки содержат TGF- β -рецепторы.
В целом, эти протеины различаются по своей структуре, что приводит к значительным вариациям в их точной биологической функции. Далее, они находятся в широкой области различных видов тканей и стадий развития. Следовательно, они могут иметь различия в отношении их точной функции, например, необходимой клеточной физиологической среды (окружения), продолжительности их жизни, их местоположения, их потребности во вспомогательных факторах и их устойчивости к деструкции. Итак, хотя и описываются многочисленные протеины, которые обладают индуктивным в отношении тканей, в особенности остеоиндуктивным, потенциалом, их естественные задачи в организме и - еще важнее - их медицинскую применимость еще нужно детально исследовать. С большой вероятностью предполагается наличие еще неизвестных членов TGF- β -семейства, которые важны для остеогенеза или для дифференциации/индукции других видов тканей. Большая трудность при выделении этих новых TGF- β -образных протеинов, однако, состоит в том, что их функции еще недостаточно точно могут быть описаны для разработки биологического анализа с целью их распознавания. С другой стороны, ожидаемая гомология нуклеотидных последовательностей для известных членов семейства слишком незначительна, чтобы сделать возможным скрининг с помощью классических методов гибридизации нуклеиновых кислот. Однако дальнейшее выделение и характеристика новых TGF- β -образных протеинов необходимы, чтобы получить другие протеины дифференцировки и индукции, которые удовлетворяют всем желательным медицинским требованиям. Эти факторы могут найти применение в медицине при лечении (заживлении) повреждений и при лечении дегенеративных заболеваний костей и/или других видов тканей, как, например, почки или печень.
В патентной заявке PCT / EP 93 / 00350 указана нуклеотидная и аминокислотная последовательности для TGF- β -протеина MP-52, причем указана соответствующая зрелому пептиду последовательность и большая часть соответствующей пропептиду MP-52 последовательности. Полная последовательность пропептида MP-52 не раскрыта.
Положенная в основу настоящего изобретения задача состоит в том, чтобы получить ДНК-последовательности, которые кодируют новые члены семейства TGF- β -протеинов с митогенным и/или дифференцирующе-индуктивным, например остеоиндуктивным, потенциалом. В особенности задача настоящего изобретения состоит в том, чтобы получить полную ДНК- и аминокислотную последовательность TGF-протеина MP-52.
Эта задача решается благодаря молекуле ДНК, которая кодирует протеин TGF- β -семейства и включает:
(а) кодирующую зрелый протеин часть (участок) и в случае необходимости другие функциональные части указанной в SEQ ID NO 1 нуклеотидной последовательности;
(б) соответствующую последовательность из п. (а) в рамках дегенерации генетического кода нуклеотидную последовательность;
(в) аллельное производное соответствующей последовательностям из п.п. (а) и (б) нуклеотидной последовательности; или
(г) гибридизирующуюся с одной из последовательностей (а), (б) или (в) последовательность;
при предположении, что молекула ДНК согласно п. (г) содержит по меньшей мере одну часть, кодирующую зрелый протеин TGF- β -семейства.
Другие варианты осуществления настоящего изобретения относятся к предмету п. п. 2-10 формулы изобретения. Дальнейшие признаки и преимущества изобретения следуют из описания предпочтительных вариантов осуществления и чертежей. Ниже кратко описываются протоколы (схемы) последовательностей и чертежи.
В SEQ ID NO 1 представлена полная нуклеотидная последовательность ДНК, кодирующей TGF- β -протеин MP-52. ATG - Стартовый кодон начинается с нуклеотида 640. "Старт" зрелого протеина начинается после нуклеотида 1782.
В SEQ ID NO 2 представлена полная аминокислотная последовательность TGF- β -протеина MP-52, которая производится от указанной в SEQ ID NO 1 нуклеотидной последовательности. Последовательности SEQ ID NO1 и SEQ ID NO2 приведены в графической части.
На фиг. 1 представлено сравнение аминокислотной последовательности MP-52 с некоторыми членами семейства BMP-протеинов с началом на первом из семи сохранившихся цистеиновых остатков. Знак * обозначает, что аминокислота одинакова во всех сравниваемых протеинах; знак + обозначает, что аминокислота совпадает по меньшей мере в одном из протеинов по сравнению с MP-52.
На фиг. 2 представлены нуклеотидные последовательности олигонуклеотидных праймеров, которые применяются в настоящем изобретении, и сравнение этих последовательностей с известными членами TGF- β -семейства. M обозначает A или C; S обозначает C или G; R обозначает A или G; и K обозначает G или T, "2a" обозначает последовательность праймера OD; "2b" обозначает последовательность праймера OID.
Настоящее изобретение охватывает по меньшей мере кодирующую зрелый протеин часть и в случае необходимости функциональную часть указанной в SEQ ID NO 1 нуклеотидной последовательности, а также последовательности, которые соответствуют этой последовательности в рамках вырожденности генетического кода, и аллельные производные таких последовательностей. Далее, настоящее изобретение охватывает также последовательности, гибридизирующиеся с такого рода последовательностями, при предположении, что такая молекула ДНК содержит часть, кодирующую зрелый протеин TGF- β -семейства.
Понятие "функциональная часть" в смысле настоящего изобретения обозначает часть протеина, которая в состоянии действовать, как, например, часть сигнального пептида, пропептида, соответственно, зрелого протеина, т.е. выполнять по меньшей мере одну из биологических функций естественных частей протеина MP-52.
Кодирующая зрелую часть протеина область простирается от нуклеотидов 1783-2142 указанной в SEQ ID NO 1 последовательности. В случае необходимости, молекула ДНК может еще содержать функциональные части указанной в SEQ ID NO 1 последовательности, а именно кодирующие сигнальную и/или пропептидную часть нуклеотидные последовательности. Особенно предпочтительно молекула ДНК содержит последовательность сигнальной и пропептидной части и часть зрелого протеина, т. е. нуклеотиды 640-2142 указанной в SEQ ID NO 1 последовательности. С другой стороны, молекула ДНК, наряду с кодирующей зрелый протеин частью, также может содержать еще функциональные сигнальные и/или пропептидные части других протеинов, в особенности других протеинов TGF- β -семейства, например вышеуказанных BMP-протеинов. Соответствующие нуклеотидные последовательности можно представить из вышеуказанных ссылок, на раскрытие которых здесь ссылаются.
Далее настоящее изобретение включает также молекулу ДНК, такую, как указанная выше, которая дополнительно между нуклеотидами 1270 и 1271 указанной в SEQ ID NO 1 последовательности содержит некодирующую интрононую последовательность. Эта интрононая последовательность содержится в депонированной в DSM плазмиде SKL 52 (H3) MP 12, которая имеет геномную последовательность нуклеиновых кислот MP-52.
Изобретение также охватывает кодированную фагом λ 15. 1 кДНК-последовательность MP-52-протеина. Эта последовательность начинается с нуклеотида 321 указанной в SEQ ID NO 1 последовательности.
Хотя аллельные, вырожденные и гибридизирующие последовательности, которые входят в настоящее изобретение, имеют структурные различия на основании незначительных изменений в нуклеотидной и/или аминокислотной последовательности, кодированные такого рода последовательностями протеины обладают еще по существу такими же полезными свойствами, которые делают возможным их использование в принципе для таких же применений в медицине.
Согласно настоящему изобретению, обозначение "гибридизация" представляет собой обычные условия гибридизации, предпочтительно условия с концентрацией соли 6 х SSC при 62-66oC, с последующей, осуществляемой в течение 1 часа, промывкой с помощью 0,6 х SSC, 0,1% SDS, при 62-66oC. Особенно предпочтительно обозначение "гибридизация" относится к строгим условиям гибридизации с концентрацией соли 4 х SSC при 62-66oC, с последующей, осуществляемой в течение 1 часа, промывкой с помощью 0,1 х SSC, 0,1% SDS, при 62-66oC.
Предпочтительными вариантами осуществления настоящего изобретения являются ДНК-последовательности, как указанные выше, которые получают от позвоночных животных, предпочтительно млекопитающих, как, например, свинья, коровы и грызуны, как, например, крысы или мыши, и в особенности от приматов, как, например, люди.
Особенно предпочтительной формой осуществления настоящего изобретения является указанная в SEQ ID NO 1 и обозначаемая как MP-52 последовательность. Транскрипты MP-52 получаются из эмбриональной ткани и кодируют протеин, который проявляет значительную гомологию аминокислот по отношению к зрелой части BMP-образных протеинов (см. фиг. 1). Протеиновые последовательности BMP 2 (=BMP 2A) и BMP 4 (=BMP 2B) описаны Wozney и др., Science, 242 (1988), 1528-1534. Соответствующие последовательности BMP 5, BMP 6 и BMP 7 описаны Celeste и др., Proc. Natl. Acad. Sci. USA, 87, (1990), 9843-9847. Некоторые типичные гомологии последовательностей, которые специфичны для известных BMP-последовательностей, находятся также в пропептидной части MP-52, в то время как другие части предшественника MP-52 имеют значительные различия с BMP-предшественниками.
Следующим предметом настоящего изобретения является вектор, который содержит по меньшей мере одну копию предлагаемой согласно изобретению молекулы ДНК. В такого рода векторе предлагаемая согласно изобретению ДНК-последовательность предпочтительно оперативно связана с последовательностью, контролирующей экспрессию. Такие векторы пригодны для получения TGF- β -образных протеинов в стабильно- или временно-трансформированных клетках. Различные системы животных, растений, грибов и бактерий можно применять для трансформации и последующего культивирования. Предлагаемые согласно изобретению векторы предпочтительно содержат необходимые для репликации в клетке-хозяине последовательности и автономно реплицируемы. Далее, предпочтительно применение векторов, которые содержат селектируемые маркерные гены, благодаря чему может быть доказана трансформация клетки-хозяина.
Другим предметом изобретения является клетка-хозяин, которая трансформирована с помощью предлагаемой согласно изобретению ДНК или предлагаемого согласно изобретению вектора. Примеры пригодных клеток-хозяев включают различные эукариотные и прокариотные клетки, как, например, E. coli, клетки насекомых, клетки растений, клетки млекопитающих и грибы, как, например, дрожжи.
Следующим предметом изобретения является протеин TGF- β -семейства, который кодируется ДНК-последовательностью по п. 1 формулы изобретения. Предлагаемый согласно изобретению протеин предпочтительно содержит указанную в SEQ ID NO 2 аминокислотную последовательность или, в случае необходимости, ее функциональные части и обладает биологическими свойствами, как, например, индуктивные для ткани, в особенности остеоиндуктивные, и/или митогенные способности, которые могут быть пригодны для терапевтического применения. Вышеуказанные признаки протеина могут изменяться в зависимости от образования гомодимеров или гетеродимеров. Такие структуры могут оказаться также пригодными для клинических применений.
Биологические свойства предлагаемых согласно изобретению протеинов, в особенности митогенный и остеоиндуктивный потенциал, можно определять, например, путем испытания согласно Seyedin и др., PNAS, 82 (1985), 2267-2271; или Sampath и Reddi, PNAS, 78 (1981), 7599-7603.
Другим предметом настоящего изобретения является способ получения протеина TGF- β -семейства, который отличается тем, что культивируют трансформированную с помощью предлагаемой согласно изобретению ДНК или предлагаемого согласно изобретению вектора клетку-хозяина и TGF- β -протеин получают из клетки и/или надосадочной жидкости после культивирования. Такой способ включает культивирование трансформированной клетки-хозяина в пригодной питательной среде и очистку полученного TGF- β -образного протеина. Таким образом способ позволяет получить достаточное количество желательного протеина для использования при лечении в медицине или при тех применениях, где употребляют методы с использованием культуры клеток, при которых необходимы факторы роста. Клеткой-хозяином может быть бактерия, как, например, Bacillus или E. coli; гриб, как правило, дрожжи; клетка растения, как, например, табак, картофель или Arabidopsis; или клетка животного, в особенности линия клеток позвоночного животного, как, например, MO-, COS- или CHO-линии клеток; или линия клеток насекомых.
Еще одним следующим предметом настоящего изобретения является приготовление фармацевтических композиций, которые содержат фармацевтически эффективное количество предлагаемого согласно изобретению TGF- β -протеина в качестве биологически активного вещества. В случае необходимости такая композиция включает фармацевтически приемлемый носитель, вспомогательное вещество, разбавитель или наполнитель. Такая фармацевтическая композиция может применяться при заживлении ран и регенерации тканей, а также при лечении повреждений костей, хрящей, соединительных тканей, кожи, слизистой оболочки, эпителия или зубов и в случае зубных имплантатов, либо индивидуально, либо в комбинации с другими биологически активными веществами, например, с другими протеинами TGF- β -семейства или факторами роста, как, например, EGF (эпидермальный фактор роста) или PDGF (происходящий от тромбоцитов фактор роста). Далее, такую фармацевтическую композицию можно применять для предупреждения заболевания, как, например, для предупреждения остеопороза и артроза.
Другим возможным клиническим применением предлагаемого согласно изобретению TGF- β -образного протеина является использование в качестве супрессора иммунной реакции для избежания отторжения трансплантатов органов или использование в связи с антиогенезом. Предлагаемую согласно изобретению фармацевтическую композицию можно также применять для профилактики или в косметической хирургии. Далее, применение композиции не ограничивается людьми, а ее можно применять также в случае животных, в особенности домашних животных.
Наконец, дальнейшим предметом изобретения является антитело, которое может специфически связываться с предлагаемыми согласно изобретению протеинами, или такого рода фрагмент антитела (например, Fab или Fab1). Способы получения такого специфического антитела или фрагмента антитела относятся к общему специальному знанию среднего специалиста. Предпочтительно такое антитело представляет собой моноклональное антитело. Такие антитела или фрагменты антител могут быть пригодны также для диагностических методов.
Далее, изобретение должно быть более наглядным благодаря следующему примеру.
Пример 1
Выделение MP-52
1.1. Полную РНК выделяют из человеческой эмбриональной ткани (в возрасте 8-9 недель) по методу Chirgwin и др., Biochemistry, 18 (1979), 5294-5299. Поли (A+)-РНК отделяют от полной РНК путем олиго (dT) - хроматографии согласно инструкциям изготовителя (Stratagene Poly (A) Quick-колонки).
1.2. Для обратной реакции транскрипции 1-2,5 мкг поли (A+)-РНК в течение 5 минут нагревают при 65oC и быстро охлаждают льдом. Реакционная смесь содержит 27 ед. РНК - Guard (Pharmacia), 2,5 мкг олиго (dT) 12-18 (Pharmacia), 5 х буфер (250 ммоль/л ТРИС/HCl, pH 8,5; 50 ммоль/л MgCl2; 50 ммоль/л ДТТ (дитиотреитола); 5 ммоль/л любого дНТФ (дезоксинуклеозидтрифосфата); 600 ммоль/л KCl) и 20 ед. AMV обратной транскриптазы (Boehringer Mannheim) на 1 мкг поли (A+)-РНК. Реакционную смесь (25 мкл) инкубируют в течение 2 часов при 42oC.
1.3. Указанные на фиг. 2 дезоксинуклеотидные праймеры OD и OID получают на автоматическом ДНК-синтезаторе (Biosearch). Очистку осуществляют путем денатурирующего электрофореза на полиакриламидном геле и выделения основной полосы из геля путем изотахофореза. Путем сравнения последовательностей нуклеиновых кислот известных членов TGF- β -семейства и выбора областей с самым высоким сохранением проектируют олигонуклеотиды. Сравнение этих областей представлено на фиг. 2. Для облегчения клонирования оба нуклеотида содержат сайты рестрикции EcoRI и OD содержит дополнительно сайт рестрикции NcoI на своем 5'-конце.
1.4. В случае полимеразной цепной реакции (PCR-реакции) применяют 20 нг соответствующей поли (A+)-РНК кДНК из исходного материала. Реакцию осуществляют в объеме 50 мкл, и он содержит 1 х PCR-буфер (16,6 ммоль/л (NH4)2SO4; 67 ммоль/л ТРИС/HCl, pH 8,8; 2 ммоль/л MgCl2; 6,7 мкмоль/л ЭДТК, 10 ммоль/л β -меркапто-этанола; 170 мкг/мл альбумина сыворотки крупного рогатого скота (Gibco); 200 мкмоль/л любого дНТФ (Pharmacia); 30 пкмоль каждого олигонуклеотида (OD и OID) и 1,5 ед. Taq-полимеразы (AmpliTaq, Perkin Elmer Cetus). Реакционную смесь покрывают парафином и осуществляют 40 циклов полимеразной цепной реакции. Продукты полимеразной цепной (PCR) реакции очищают путем экстракции с помощью фенола с хлороформом и концентрируют путем осаждения этанолом.
1.5. Продукт полимеразной цепной реакции расщепляют с помощью рестрикционных ферментов SphI (Pharmacia) и ALWNI (Biolabs) соответственно инструкциям изготовителя.
1.6. Продукты рестрикционного расщепления фракционируют путем электрофореза на агарозном геле. После окрашивания с помощью этидиумбромида нерасщепленные продукты амплификации вырезают из геля и выделяют путем экстракции фенолом. Полученную ДНК затем очищают путем двукратной экстракции фенолом с хлороформом.
1.7. После осаждения этанолом одну четвертую или одну пятую часть выделенной ДНК реамплифицируют, причем используют такие же условия, как и для первичной амплификации, кроме того, что число циклов уменьшается до 13. Продукты реамплификации (повторной амплификации) очищают, разрезают с помощью таких же ферментов, как указанные выше, и неразрезанные продукты, как пояснено выше для продуктов амплификации, выделяют из агарозных гелей. Стадию реамплификации (повторной амплификации) повторяют дважды.
1.8. После последнего выделения из геля продукты амплификации расщепляют благодаря 4 ед. EcoRI (Pharmacia) при рекомендуемых изготовителем условиях. Одну четвертую часть смеси после рестрикции лигируют с расщепленным с помощью EcoRI вектором pBLuescriptII SK+ (Stratagene). После лигирования 24 клона анализируют далее путем секвенирования. Расщепленный с помощью ALWNI и SpHI образец дает новую последовательность, которая обозначается как MP-52. Другие клоны содержат преобладающие BMP 6-последовательности и один клон содержит BMP 7-последовательность. Клон на 3'-конце дополняют кДНК согласно подробно описанному Frohmann методу (Амплификации, опубликовано Perkin-Elmer Corp. , Issue 5 (1990), с. 11-15). Такую же эмбриональную мРНК, которая была применена для выделения первого фрагмента MP-52, подвергают обратной транскрипции как описано выше. Амплификацию осуществляют при применении адаптерного праймера (AGAATTCGCATGCCATGGTCGACG) и внутреннего праймера (CTTGAGTACGAGGCTTTCCACTG) MP-52-последовательности. Продукты амплификации реамплифицируют (повторно амплифицируют) при применении перекрывающего адаптерного праймера (ATTCGCATGCCATGGTCGACGAAG) и перекрывающего внутреннего праймера (GGAGCCCACGAATCATGCAGTCA) MP-52-последовательности. Продукты реамплификации, после рестрикционного расщепления с помощью NcoI, клонируют в расщепленном таким же образом векторе [pUC 19 (Pharmacia N 27-4951-01) с одним измененным составным (multiplen) участком клонирования, который содержит единственный сайт рестрикции NcoI], и секвенируют. Клоны характеризуют путем соединения внахлестку их последовательности с 3'-концом известной MP-52-последовательности. Один из них используют в качестве зонда для скрининга человеческого геномного банка генов (Stratagene N 946203) согласно описанному подробно Ausubel и др. методу (Current Protocolsin Molecular Biology, опубликовано Greene Publishing Associates und Wiley-Intersience /1989/). Из 8•105 λ -фагов выделяют фаг ( λ 2.7.4), который содержит вставку примерно из 20 кб и депонирован в DSM под номером 7387. Этот клон, наряду с выделенной из мРНК путем описанных методов амплификации последовательностью, содержит другие, несущие информацию последовательности /Sequenzinformationen/ на 5'-конце.
Для анализа путем секвенирования HindIII-фрагмент длиной примерно 7,5 кб субклонируют в разрезанном таким же образом векторе (Bluescript SK, Stratоgene N 212206). Эта, обозначаемая как SKL 52 (H3) MP12, плазмида также хранится в DSM под номером 7353. Представленная в SEQ ID NO 1 несущая информацию последовательность происходит от фага λ 2.7.4. ATG в положении 640 представляет собой первый ATG в рамке считывания (в положении 403 появляется стоп-кодон). На основании указанной последовательности нужно предполагать, что при этом речь идет о стартовом кодоне для трансляции.
Геномная ДНК содержит интрон длиной примерно 2 кб между парами оснований 1270 и 1271 SEQ ID NO 1. Последовательность интрона не показана. Правильность места сплайсинга подтверждается путем секвенирования продукта амплификации, который происходит от кДНК, содержащей эту область. Эти несущие информацию последовательности получают с помощью слегка модифицированного метода, который подробно описан Frohman (Амплификации, опубликовано Perkin-Elmer-Corporation, Issue 5, (1990), с. 11-15). Такую же эмбриональную РНК, которую использовали для изолирования 3'-конца MP-52, подвергают обратной транскрипции при применении внутреннего, ориентированного в 5'-направлении, праймера MP-52-последовательности (ACAGCAGGTGGGTGGTGTGGACT). Поли-A-конец (хвост) присоединяют к 5'-концу первой кДНК-нити при применении концевой трансферазы. Осуществляют двухстадийную амплификацию, сначала путем применения состоящего из олиго-dT и адаптерной последовательности праймера (AGAATTCGCATGCCATGGTCGACGAAGC /T 16/) и, во-вторых, при применении адаптерного праймера (AGAATTCGCATGCCATGGTCGACG) и внутреннего праймера (CCAGCAGCCCATCCTTCTCC) МP-52-последовательности. Продукты амплификации реамплифицируют (повторно амплифицируют) при применении такого же адаптерного праймера и перекрывающего (соединяющегося внахлестку) внутреннего праймера (TCCAGGGCACTAATGTCAAACACG) MP-52-последовательности. Затем продукты реамплификации повторно амплифицируют при применении перекрывающего адаптерного праймера (ATTCGCATGCCATGGTCGACGAAG) и перекрывающего внутреннего праймера (ACTAATGTCAAACACGTACCTCTG) MP-52-последовательности. Продукты повторной амплификации с помощью гладких концов клонируют в векторе (Bluescript, SK, Stratagene N 212206), который расщеплен с помощью EcoRV. Клоны характеризуют путем перекрывания их последовательности с помощью ДНК фага λ 2.7.4.
Далее, кДНК-банк, полученный из РНК человеческих фибробластов и клонированный в фаге λ qt 10, подвергают скринингу. При этом исследуют 2 х 106 фагов, причем в качестве радиоактивного зонда служит величиной примерно 1 кб фрагмент геномной MP-52-ДНК (2-ой экзон вплоть до сайта рестрикции HindIII в неподвергнутой трансляции 3'-области). Выделяют 17 пятен (зон гемолиза) смеси, которые дополнительно исследуют с помощью PCR при применении праймеров из 5'-3'-области MP-52-последовательности. После этого выбирают и разъединяют 8 стерильных пятен. кДНК выделяют из фага путем частичного расщепления с помощью EcoRI и клонируют в также расщепленном с помощью EcoRI Bluescript-векторе.
Секвенирование одной из результирующих плазмид SK 52 L 15. IMP25 показывает, что самый длинный фаг (15.1) начинается с нуклеотида N 321 SEQ ID K N 1. Далее, путем секвенирования подтверждается место сплайсинга (нуклеотид 1270).
Плазмида SKL 52 (H3) MP 12 хранится под номером 7353 в DSM (немецкая коллекция микроорганизмов и культур клеток, Mascheroder Weg 1b, 38124, Braunschweig) с 10 декабря 1992 г.
Фаг λ 2.7.4. хранится под номером 7387 в DSM с 13 января 1993 г.
Плазмида SK 52 L15. IMP25 хранится под номером 8421 в DSM с 16 июля 1993 г.
Пример 2
Экспрессия MP-52
Для экспрессии MP-52 испытывают различные системы. Применение вирусов оcповакцины в качестве системы экспрессии подробно и как служащее руководством для специалиста описано в Curren Protocols в Molecular Biology (Ausubel и др., Greene Publishing Associates and Wilay-Interscience, Wiley and Sans), в дальнейшем сокращенно называемых CP, в главе 16, часть 16.15 - 16.18. Система основана на том, что чужеродные ДНК при применении определенных векторов путем гомологичной рекомбинации можно интегрировать в геном вируса осповакцины. Для этой цели используемый вектор содержит ТК (тимидинкиназа) ген из генома осповакцины. Для того чтобы сделать возможной селекцию в отношении рекомбинантных вирусов, вектор, далее, содержит E.coli-ксантин-гуанин-фосфорибозил-трансфераза-ген (gpt)/Falkner и др., J. Virol. 62 (1988), 1849-18544/. В этом векторе клонируют кДНК со всей кодирующей областью MP-52. кДНК происходит от плазмиды SK 52L15. IMP25 (DSM, номер 8421), которую для удаления большой части не подвергнутой трансляции 5'-области, однако, сначала подвергают делеции и промежуточно клонируют. Для этого плазмиду SK 52L15. IMP25 линеаризируют с помощью SalI и 5'-конец последовательно подвергают делеции с помощью набора ExoIII/Mung Bean (Stratagene # 200 330) согласно указаниям изготовителя. После рестрикции с помощью BamHI в разном степени подвергшиеся делеции MP-52-кДНК на агарозогеле отделяют от остаточного вектора, изолируют и согласно стандартным методам (Sambrook и др. , Molecular Cloning, 2-е издание, Colg Spring Harbor Laboratory Press, 1989) промежуточно клонируют в подвергнутом рестрикции с помощью EcoRV и BamHI pBLuescriptII SK-векторе (Stratagene #212206) /p.SK52S/. Все рестрикции осуществляют согласно указаниям изготовителя. Дополнительное секвенирование с помощью секвеназы (USB/Amersham #70770) дает, между прочим, клон, который начинается с нуклеотида 576 в SEQ ID NO 1 (на 64 пары оснований удален от стартового кодона). Из него за счет рестрикции с помощью SalI и SacI изолируют кДНК-вставку и клонируют в таким же образом расщепленном векторе для рекомбинации в осповакцине. Результирующую плазмиду (pBPIM52S) хранят в DSM (под номером 9217) с 24 мая 1994 г. и используют для получения рекомбинантных вирусов осповакцины. Для этого до 80% конфлюэнтных 14 3B клеток (HuTk-, ATCC CRL 8303) в чашках для культур диаметром 35 мм инфицируют с помощью вируса осповакцины дикого типа в 2 мл PBS (фосфатно-буферного рассола) в течение 30 минут при комнатной температуре и при встряхивании по случаю (1 вирус на 10 клеток). После отсасывания надосадочной жидкости и добавки 2 мл питательной среды (MEM, Gibco BRL #041-01095) инкубируют в течение 2 часов при 37oC. Среду затем удаляют, и трансформации этих клеток достигают с помощью 100 нг pВPIMP52S, 2 мкг носителя ДНК (телячья вилочковая железа, Boehringer Mannheim # 104175) и 10 мкл липофектина (Gibco BRL # 18292-011) в 1 мл MEM в течение 15 часов при 37oC. После добавки 1 мл MEM с 20% FCS (Gibco BRL # 011-06290) инкубируют в течение следующих 24 часов при 37oC и лизированные клетки затем замораживают. gpt-Селекцию на ксантин-гуанин-фосфорибозил-трансферазу и изоляцию и амплификацию отдельных рекомбинантных вирусов осуществляют по существу как описано в части 16.17 CP, с тем различием, что применяют РК-13 клетки (ATCC CCL 37).
Интеграцию MP-52-кДНК в вирусный геном подтверждают путем анализа при использовании блоттинг-метода по Саузерну и дот-блоттинга (CP, часть 16.18). Рекомбинантный вирус используют для экспрессионных анализов в линии клеток 143 B (HuTk-, ATCC CRL, 8303, человек). Конфлюэнтные клетки инфицируют с помощью соответствующего числу клеток числа вирусов в течение 45 минут при 37oC и затем добавляют соответствующую питательную среду (MEM, Gibco BRL # 041-01095) с 10% FCS и пенициллин/стрептомицин (1:500, Gibco BRL # 043-0514 OH). Спустя 6 часов при 37oC удаляют среду, клетки промывают дважды с помощью, например, HBSS (Gibco BRL # 042-0418 OM) и добавляют питательную среду (например, MEM) без FCS. После продуцирования в течение 20-22 часов собирают надосадочную жидкость культуры клеток. Анализ экспрессии осуществляют путем вестерн-блоттинга по стандартным методам (CP, часть 10.8). Для этого протеины осаждают из 100-500 мкл недосадочной жидкости культуры клеток путем добавки эквивалентного объема ацетона и инкубации по меньшей мере в течение 1 часа на льду и отделяют путем центрифугирования. После повторного суспендирования осадка в заданном буфере (7 М мочевины, 1% SDS, 7 ммоль дигидрофосфата натрия, 0,01% бромфенолового синего и, в случае необходимости, 1% β -меркаптоэтанола) осуществляют разделение на 15%-ном полиакриламидном геле. В качестве маркерных протеинов используют предварительно окрашенный стандарт по молекулярной массе протеина (Gibco BRL # 26041-020). Перенос на PVDF-мембрану (Immobilon # IPVH00010) и блокирование мембраны осуществляют стандартными методами.
Для обнаружения MP-52 на мембране получают поликлональные антитела против MP-52 как в случае кур, так и также в случае кроликов. Для этого зрелую часть MP-52 с 6 гистидинами на N-конце экспрессируют в E.coli и очищают, как, например, описано Hochuli и др. (B 10/Technology, 6, 1321-1325 /1988/). С помощью обоих антител можно обнаруживать специфически экспрессию MP-52, причем димерный MP-52 менее эффективно распознается, чем мономерный. Для вестерн-блоттинга, согласно фиг. 3, применяют куриные антитела, которые специфически очищены путем ПЭГ-преципитации /Thalley и др. B 10/Technology, 8, 934-938 (1990)/ и через связанный с мембраной антиген (зрелый MP-52 с 6 гистидинами) (18, 17; Sambrook и др., Molecular Cloning, 2-е издание, Cold Spring Harbor Laboratory Press, 1989). В качестве второго антитела используют анти-куриный IgG с присоединенной щелочной фосфатозой (Sigma A 9171).
Обнаружение осуществляют с помощью набора для определения протеина Tropix Western-Light (Serva #WL10 RC), согласно указаниям изготовителя.
Вестерн-блоттинг на фиг. 3 показывает, что только в случае рекомбинантных вирусов, однако не в случае вирусов дикого типа (без интегрированной чужеродной ДНК), появляются специфические для MP-52 полосы. Экспрессия MP-52 приводит к секретированному протеину с проявляющейся в геле при невосстанавливающих условиях молекулярной массой приблизительно 25 кДа. При восстанавливающих условиях протеин с 14-15 кДа проникает в гель. Эти результаты показывают, что MP-52 экспримируется как димерный зрелый протеин. В случае появляющихся при вестерн-блоттинг-анализе слабых полос в области выше 60 кДа речь идет, вероятно, об остатках неразрезанных протеинов-предшественников. Поведение в отношении проникания к тому же подтверждает получаемые из SEQ ID NO 2 теоретические молекулярные массы, сообразно с чем зрелый мономерный MP-52 имеет величину 13.6 кДа.
Экспрессию MP-52 и расщепление протеина-предшественника до зрелого MP-52 можно обнаружить в различных линиях клеток. Испытывали клетки C 127 (ATCC CRL, 1616, мышь), BHK 21 (ATCC CCL, 10, хомяк), MRC-5 (ATCC CCL, 171, человек) и 3TG Swiss albino (ATCC CCL, 96, мышь).
Экспрессия и расщепление до зрелого MP-52 показана также в другой эукариотной системе экспрессии. Для этого кДНК MP-52 (начинающуюся с нуклеотида 576) клонируют в плазмиде экспрессии pSG5 (Stratagene # 216201). Плазмиду pSK52s подвергают рестрикции с помощью ClaI и XbaI и путем обработки T4-полимеразой делают тупыми выступающие концы MP-52-вставки. Клонирование в подвергнутом рестрикции и также с тупыми концами за счет обработки с помощью T4-полимеразы векторе p.SG5 осуществляют стандартными методами. Все ферментативные реакции осуществляют согласно указаниям изготовителя. Правильную ориентацию MP-52-вставки обеспечивают за счет рестрикционного анализа и дополнительного секвенирования с помощью 17-праймера (Stratagene # 300302). Результирующую плазмиду pSG52s (хранится с 17.05.1994 г. в DSM под номером 9204) можно контрасформировать с помощью вектора, который кодирует выбираемый маркер, как, например, ген устойчивости G418, чтобы получить стабильные линии клеток. Для этой цели pSG52s контрансформируют вместе с плазмидой p3616 (хранится в DSM под номером 9203 с 17.05.1994 г.) в L929-клетки (ATCC CCL, 1, мышь) с помощью липофектина (Cibco BRL # 18292-011), согласно указаниям изготовителя. Селекцию с помощью G418 осуществляют согласно известным специалисту методам (CP, часть 9.5) и приходят к линии клеток, которая в вестерн-блоттинге продуцирует обнаруживаемый зрелый MP-52.
Другой вектор экспрессии для MP-52 получают при применении плазмиды pABWN (Niwa и др., Gene, 108 (1991), 193-200; и фиг. 4), который представлен в распоряжение Dr. Miyazaki.
Для этого HindIII-фрагмент из плазмиды p.SK52s, который начинается с нуклеотида 576 в SEQ ID NО 1, изолируют и выступающие концы затупляют путем обработки с помощью фрагмента Кленова. Путем лигирования адаптера в оба конца фрагмента вводят сайт рестрикции NotI. Адаптер:
AGCGGCCGCT
TCGCCGGCGA
Вектор pABWN подвергают рестрикции с помощью XhoI, также обрабатывают фрагментом Кленова и дефосфорилируют с помощью кишечной щелочной фосфатазы теленка (Boehringer Mannheim). Этот фосфорилированный адаптер дополнительно лигируют так, что теперь возможна вставка MP-52-фрагмента после рестрикции с помощью NotI в генерированное NotI место разреза вектора. Результирующий вектор экспрессии ниже обозначается как HindIII-MP-52/pABWN. Все осуществляемые реакции клонирования проводят по стандартным методам (например, CP, часть 3.16). Структура HindIII-MP-52/pABWN-вектора экспрессии подтверждается путем секвенирования и получения карты рестрикции. HindIII-MP-52/pABWN содержит MP-52-последовательность, начинающуюся с нуклеотида 576 и заканчивающуюся нуклеотидом 2278 в SEQ ID NO 1.
HindIII-MP-52/pABWN трансфицируют в L-клетки (мышиные фибробласты), и оттуда получают стабильные трансформанты. Для этого, смотря по обстоятельствам, 4 мкг плазмиды (HindIII-MP-52/pABWN или pABWN) трансфицирую в 5•105 L-клеток, находящихся в чашках для культур диаметром 6 см, при применении 20 мкл реактива Lipofect AMINE (Gibco BRL # 18324-012). Для этой цели раствор A (4 мкг соответствующей плазмиде ДНК в 200 мкл OPTI-MEM I /Gibco BRL # 31985/) осторожно смешивают с раствором B (20 мкл реактива Lipofect AMINE в 200 мкл OPTI-MEM I) и при комнатной температуре в течение 45 минут инкубируют для образования комплекса ДНК-липосомы. Во время этой процедуры клетки промывают один раз с помощью 2 мл OPTI-MEM I. Для каждой трансфекции в сосуд с ДНК-липосомным комплексом вводят 1,6 мл OPTI-MEM I. Раствор осторожно перемешивают и таким образом переслаивают промытые клетки. Клетки инкубируют с разбавленным комплексом в течение 5 часов при 37oC в CO2-содержащем инкубаторе. После инкубации добавляют 2 мл DMEM (Gibco BRL, модифицированная Dulbecco Eagle-среда) с 20% FCS. Спустя 24 часа после трансфекции среду заменяют свежей DMEM с 10% FCS. Спустя 48 часов после начала трансфекции клетки переносят в чашки для культур диаметром 10 см. Спустя 72 часа после начала трансфекции с концентрации 800 мкг/мл начинается селекция G 418. Стабильные клоны появляются спустя 1-2 недели.
5 мл кондиционированной DMEM с или без FCS получают от конфлюэнтных трансформантов, которые выросли в течение 3 дней в чашке для культуры диаметром 10 см. Трансфицированные двумя различными надосадочными жидкостями культур клеток (HindIII-MP-52/pAВWN и pABWN) клетки, а также лизаты клеток исследуют путем вестерн-блоттинга. При этом в кондиционированной среде, а также в лизатах трансфицированных с помощью HindIII-MP-52/pABWN клеток найден зрелый MP-52. Клоны клонируют далее и продуцирующие MP-52 клетки, смотря по обстоятельствам, выбирают согласно вестерн-блоттинг-анализу. Оценки из вестерн-блоттинг-анализов показывают MP-52-продуцирование вплоть до 1 мг/л.
Пример 3
Биологическая активность MP-52
Для того чтобы обнаружить биологическую активность MP-52 и доказать полезность настоящего изобретения для применений в медицине с целью избежания и/или лечения заболеваний костей, осуществляют различные эксперименты ин витро и ин виво.
1. Испытание ин витро
1.1.
Так как усиление синтеза гликозаминогликана (GAG) в хондроцитах после TGF- β -стимуляции описано (Hiraki и др., Biochimica et Biophysica Acta, 969 (1988), 91-99), исследуют, оказывает ли также MP-52 это влияние. При применении надосадочных жидкостей культур клеток (DMEM с 10% FCS), продуцирующих MP-52 трансформантов L-клеток (трансфекция с помощью HindIII-MP-52/pABWN), изучают хондрогенную активность MP-52 в первичных культурах из зародышевых конечностей крыс.
Для этой цели используют 44 конечности крысиных плодов в возрасте 16 дней. После трипсинирования, полученные клетки в F-12-среде (питательная смесь Ham's F-12, Gibco BRL # 21700) с 10% FCS помещают на покрытые коллагеном типа 1 пластины с 24 углублениями в количестве 3•105 клеток и культивируют примерно 2 дня вплоть до конфлюэнции. К 500 мкл питательной среды (F-12-среда с 10% FCS), смотря по обстоятельствам, добавляют 56 мкл кондиционированной среды (KM) трансфектантов за счет HindIII-MP-52/pABWN L-клеток, трансфектантов за счет pABWN-L-клеток или только среду (DMEM с 10% FCS). Через промежуток времени 0, 3, 6 и 9 дней применяют F-12-среду с 10% FCS, а также соответствующие добавки. Все три дня осуществляют перемену среды с соответствующими добавками. Затем следующие 2 дня культуру культивируют в F-12-среде без FCS в присутствии соответствующих добавок (кондиционированные среды, соответственно, контрольная среда) и после этого добавляют 35S-сульфат за 6 часов. Инкорпорированную в полисахариды 35S определяют после проназа-E-переваривания и преципитации, как описано Hiraki и др. (Biochimica et Biophisica Acta, 969 (1988), 91-99).
Как видно из таблицы, 1 надосадочные жидкости культур клеток продуцирующих MP-52 трансфектантов значительно стимулируют синтез GAG по сравнению с чистой питательной средой (DMEM с 10% FCS) или надосадочной жидкостью культуры L-клеток, трансфицированных с помощью pABWN. Это показывает, что MP-52 может стимулировать дифференциацию хондроцитов.
1.2.
Описанный эффект некоторых членов BMP-семейства представляет собой усиление активности щелочной фосфатазы (ALP-активности) в остеобластях. Клональные линии клеток крыс ROВ - C-26 (C-26) причисляют к остеобластам относительно ранней стадии созревания (Yamaguchi и др., Calcif. Tissue Int. 49 (1991), 221-225).
Для остеоиндуктивных протеинов, как, например, BMP-2, описана способность усиливать ALP-активность: Yamaguchi и др., J. Cell. Biol. 113 (1991), 681-687.
Влияние MP-52 на C-26-клетки исследуют следующим образом. C-26-Клетки в количестве 3•104 клеток на углубление высевают на пластину с 24 углублениями и культивируют в среде α -MEM (Gibco BRL) с 10% FCS вплоть до конфлюэнции. На углубление добавляют 56 мкл надосадочной жидкости культуры продуцирующих MP-52 трансфектантов L-клеток (HindIII-MP-52/pABWN), соответственно, надосадочной жидкости трансфектантов L-клеток, полученных с помощью pABWN, или только надосадочной жидкости культуры (DMEM с 10% FCS) L-клеток к 500 мкл среды культуры клеток C-26. Замену среды с соответствующими добавками осуществляют все три дня. ALP-Активность в экстрактах клеток определяют спустя 0, 3, 6, 9 и 12 дней с помощью стандартных способов, базирующихся на п-нитрофенилфосфате в качестве субстрата, как, например, описано Takuwa и др. (Am. J. Physiol. 257 /1989/, E797-E803).
Как следует из таблицы 2, ALF-активность значительно усиливается за счет добавки MP-52 по сравнению с чистой средой DMEM с 10% FCS и средой инфицированных с помощью pABWN L-клеток.
Этот результат показывает, что MP-52 может способствовать не только дифференциации хондроцитов, но и также дифференциации и созреванию остеобластов.
Другая линия клеток остеобластов (MC3T3-EI, мышь), которая, как описано Takuwa и др. (Biochem. Biophys. Res. Com., 174 (1991), 96-101), путем обработки BMP-2 показывает увеличение ALP-активности, после инкубации с кондиционированной средой продуцирующих MP-52 трансфектантов L-клеток (HindIII-MP-52/pABWN) или со средой после продуцирования MP-52 за счет инфекции рекомбинантными вирусами осповакцины не показывает никакого изменения ALP-активности. Это указывает на то, что MP-52 обладает отчасти специфичностью клетки, отклоняющейся от BMP-2. Различные функции, обусловленные разными целевыми участками отдельных членов TGF- β -семейства, могут иметь большую пригодность в медицине.
2. Эксперименты ин виво
2.1.
Наиболее четко высказанная возможность исследования развития костей базируется на эктопическом костеобразовании ин виво. Его можно индуцировать, например, путем имплантации деминерализованной костной матрицы (Urist, Science, 150 (1965), 893-899). Путем комбинации неактивной матрицы с индуцирующими кости протеинами можно индуцировать такой же процесс, который описан, например, Sampath и др. (PNAS (Proc. Natl. Acad. Sci., США), 78 (1981), 7599-7603). Этот процесс костеобразования похож на таковой эмбрионального энхондрального костеобразования и лечения костей во взрослом состоянии. Таким образом, этот метод дает возможность исследовать протеины на их способность к индуктированию костей ин виво.
Для такого эксперимента MP-52-протеин, который получают путем экспрессии в системе осповакцины (см. пример 2), частично очищают и имплантируют.
Для этой цели 143B-клетки (HuTk-, ATCC CRL 8303) выращивают в чашках для культивирования и микролитражных емкостях вплоть до конфлюэнции и, как описано в примере 2 для анализов путем экспрессии, инфицируют с помощью рекомбинантных вирусов, промывают и оставляют аккумулировать примерно в течение 20 часов MP-52 в MEM (Gibco BRL, примерно 1 мл на 106 клеток). В качестве контроля осуществляют такое же приготовление путем инфекции с помощью вирусов дикого типа. Надосадочную жидкость культуры клеток (кондиционированная среда) каждого приготовления (смеси) собирают и центрифугируют (40000•g в течение 30 минут при 4oC). Для удаления вирусов надосадочные жидкости фильтруют через неорганические фильтры (величина пор 0,1 мкм, Whatman, Anotop 25). В ходе охарактеризовывания MP-52 смогли обнаружить, что этот протеин связывается с гепарин-сефарозой. Это поведение используют для частичной очистки. Для этой цели отфильтрованную и отцентрифугированную, кондиционированную среду доводят до конечной концентрации 50 ммоль ТРИС, pH 7,0; 100 ммоль NaCl и 6 моль мочевины и загружают в колонку с гепарином (HiTropTM, Pharmacia # 17-0407-01), которая уравновешена буфером A (50 ммоль ТРИС, pH 7,0, 100 ммоль NaCl и 6 моль мочевины). Нагруженную колонку промывают буфером A и с помощью линейного градиента до 100% буфера B (50 ммоль ТРИС, pH 7,0, 600 ммоль NaCl и 6 моль мочевины) при скорости истечения 0,5 мл/мин элюируют в течение 50 мин (2,5 мл на фракцию). Применение мочевины необязательно. Путем вестерн-блоттинг-анализа (см. пример 2) можно дополнительно проверить, что MP-52 элюируется воспроизводимо в основном в 2 фракциях при концентрации NaCl примерно 250-400 ммоль. Аликвоты этих фракций также, согласно инструкциям изготовителя, проверяют с помощью окрашенных серебром 15%-ных полиакриламидных гелей (Silver Stain-II, Daiichi # SE 140000) и объединяют. Сопоставимые фракции, после очистки от кондиционированной среды после инфекции с помощью вирусов дикого типа, также объединяют после анализа в окрашенных серебром гелях.
Из дальнейших исследований в отношении MP-52 следует, что MP-52 также связывается с гидроксиапатитом. Поэтому в принципе можно достигать дополнительной очистки при использовании колонки с гидроксиапатитом, соответственно заменять колонку с гепарином (например, B10-RAD, Econo-pac HTP). Для дальнейших очисток возможны также другие, известные специалисту методы, как, например, колонки с гель-ситами (Gelsiebsoulen), колонки с ионообменниками, аффинные колонки, колонки с хелатами металлов или колонки, базирующиеся на гидрофобных взаимодействиях.
Предварительно очищенный путем хроматографии на гепаринсефарозе MP-52-протеин соответственно, еще соответственно загрязняющие протеины, которые находятся также в инфицированных диким типом надосадочных жидкостях культуры клеток, очищают далее с помощью обращенной фазы ВЭЖХ (высокоэффективной жидкостной хроматографии). Для этой цели C8-колонку (Aquapore RP 300, Applied Biosystems, размер частиц: 7 мкм; величина пор: 300 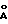 ) уравновешивают с помощью 10% буфера B (буфер A : 0,1% трифторуксусной кислоты; буфер B : 90% ацетонитрила, 0,1% трифторуксусной кислоты). После загрузки колонки объединенными, содержащими MP-52 фракциями гепариновую колонку промывают в большом количестве 10% буфером B. Связанный протеин элюируют с помощью следующих градиентов: 10-50% буфера B в течение 20 минут и 50-100% буфера B в течение 50 минут. Фракции по 500 мкл собирают и анализируют как путем вестерн-блоттинг-анализа, так и также в окрашенных серебром гелях. MP-52-Протеин при выбранных условиях элюируется примерно в области 55-65% ацетонитрила. Фракции с MP-52 объединяют. То же самое осуществляют с соответствующими фракциями из контрольной очистки надосадочной жидкости инфицированных вирусами дикого типа клеток.
) уравновешивают с помощью 10% буфера B (буфер A : 0,1% трифторуксусной кислоты; буфер B : 90% ацетонитрила, 0,1% трифторуксусной кислоты). После загрузки колонки объединенными, содержащими MP-52 фракциями гепариновую колонку промывают в большом количестве 10% буфером B. Связанный протеин элюируют с помощью следующих градиентов: 10-50% буфера B в течение 20 минут и 50-100% буфера B в течение 50 минут. Фракции по 500 мкл собирают и анализируют как путем вестерн-блоттинг-анализа, так и также в окрашенных серебром гелях. MP-52-Протеин при выбранных условиях элюируется примерно в области 55-65% ацетонитрила. Фракции с MP-52 объединяют. То же самое осуществляют с соответствующими фракциями из контрольной очистки надосадочной жидкости инфицированных вирусами дикого типа клеток.
Также частично очищенный MP-52-протеин в определенной путем вестерн-блоттинг-анализа концентрации 50 нг/мл показывает отчетливое повышение ALP-активности на ROB-C-26-клетках после трех стадий инкубации.
Частично очищенный MP-52-протеин, соответственно контрольный протеин из соответственно частично очищенных надосадочных жидкостей культур клеток после инфекции вирусами дикого типа, реконструируют с матрицей и имплантируют крысам, чтобы доказать способность к хряще- и костеобразованию.
В принципе должны быть применимы различные, известные специалисту матричные материалы, т. е. природные (также модифицированные), и синтетически полученные матрицы, предпочтительно, однако, биосовместимые, биологически разрушаемые ин виво пористые материалы. В этих экспериментах применяют костную матрицу крыс, которая по существу приготовлена подобно тому, как описано Sampath и др. (PNAS, 80 (1983), 6591-6595). Крысиные кости (бедро и большеберцовая кость) деминерализуют в течение 24 часов в 0,6 М HCl и затем удаляют еще имеющийся костный мозг. После промывки с помощью воды и обезжиривания в течение 3 часов в смеси хлороформа с метанолом (1:1) кости высушивают на воздухе, замороженными при низкой температуре пульверизуют (распыляют) в мельнице и отделяют путем просеивания частицы размером 400-1000 мкм. После этого матрицу в течение 7 дней при комнатной температуре экстрагируют в 4 М гуанидиний-HCl в присутствии ингибиторов протеазы. После экстенсивных (обильных) промывок водой матрицу лиофилизируют и хранят при 4oC. Таким образом обработанные матрицы не проявляют, ни одна из них, никакой индуктирующей кости активности.
Протеин при применении различных, известных специалисту методов можно комбинировать с проэкстрагированной костной матрицей.
MP-52-Протеин, соответственно контрольный протеин, который очищен как при использовании гепарин-сефарозы, так и также обращенной фазы ВЭЖХ, в растворе ацетонитрила с трифторуксусной кислотой, после элюирования, объединяют в количестве по 25 мг. матрицы на имплантат, хорошо смешивают, замораживают при низких температурах и лиофилизируют.
Для имплантации связанного с матрицей MP-52 используют крыс (Wistar) в возрасте примерно 3 месяца, которых приводят в состояние наркоза путем внутримышечной инъекции наркотизирующего средства (0,2 мл Rompun (Bayer), смешанные с 0,5 мл Ketanest 50 (Parke Davis)) в количестве 0,14 мл на 100 г веса тела. Для имплантатов приготовляют с двух сторон карманы в брюшных мышцах (ниже грудной клетки, начиная примерно на 0,5 см ниже самой нижней реберной дуги). Связанный с матрицей MP-52 (примерно 2-4 мкг согласно определению по вестерн-блоттинг-анализу), а также соответственно связанные с матрицей контрольные протеины увлажняют с помощью 0,9%-ного раствора хлорида натрия (Delta Pharma) и переносят в мышечные карманы. Мышечные карманы, а также необходимые разрезы кожи затем зашивают. Крыс иммуноподавляют с помощью циклоспорина A (Sandimmun).
Спустя 18, соответственно 26 дней имплантаты извлекают из крыс и фиксируют для гистологических исследований. Так как имплантат с MP-52 после 26 дней уже позволяет предполагать макроскопически образование костей, то его помещают (заделывают) в метилметакрилат для приготовления тонких пленок; другие имплантаты заделывают в парафин. Минерализованные хрящевые и костные ткани чернят (делают черными) благодаря способу окраски Von Kossa (Romeis B. , Mikroskopische Technik, изд. Bock P.; Urban и Schwarzenberg; Munchen, Baltimore, Wien /1989/). При окрашивании с помощью трихром-красителей (Trichromfarbung) согласно Masson-Goldner (Romeis B.; Mikroskopisсhe Technik, изд. Bock P.; Urban и Schwarzenberg; Munchen, Baltimore, Wien /1989/) минерализованная костная ткань и коллаген окрашиваются слегка зеленым, остеоид является красным, а цитоплазма имеет красновато-коричневую окраску. К имплантатам из обеих крыс применяют оба способа окрашивания. С помощью обоих способов окрашивания в случае обоих подопытных животных можно доказать отчетливое хряще- и костеобразование в имплантатах, которые содержат MP-52. Соответствующие имплантаты с контрольным протеином не показывают никакого хряще- и костеобразования. Доля предстадий хряща с хондроцитами и хрящевыми областями с начинающимся образованием внеклеточной матрицы и ее минерализация в концентрических кругах в имплантате с MP-52 спустя 18 дней выше, чем в таковом спустя 26 дней. Однако также в имплантате спустя 18 дней уже обнаруживаются зрелые костные ткани с векториальным образованием остеоида, а также отдельные остеоциты в кости. Далее, распознаваемы замкнутые косточки (Ossikel) с начинающимся образованием костного мозга. В случае имплантата спустя 26 дней также еще обнаруживаются хрящевые области с начинающимся образованием матрицы и кальцинозом, доля окрашенной в зеленый цвет минерализованной костной ткани с остеоцитами и остеоидными краями, однако, отчетливо увеличена. Также в этом имплантате обнаруживается образование костного мозга с единичными (отдельными) случаями жировых клеток. Для наглядности на фиг. 5 показано получаемое путем окраски обнаружение (von Kossa) костного материала из всего имплантата спустя 26 дней. На фиг. 6 показан маленький вырез из того же самого имплантата после окрашивания по Masson-Goldner. Видны активные кости с ободком (краем) из кубиодальных остеобластов и в отдельных "вмурованных" остеобластях распознаваемы остеоиды. Далее, видны отдельные остеоциты в минерализованной костной ткани (в подлинном препарате окрашено зеленым). Также обнаруживается образование костного мозга.
Опыт показывает, что рекомбинантно полученный MP-52, один, в комбинации с матрицей, в состоянии индуцировать энхондральное костеобразование.
2.2.
Для подтверждения результатов осуществляют другой тест эктопического костеобразования при применении трансформантов за счет MP-52 L-клеток. Продуцирующие MP-52 (трансфицированные с помощью HindIII-MP-52/pABWN) и непродуцирующие MP-52 (трансфицированные с помощью pABWN) L-клетки (1•106 клеток) вводят путем инъекции с обеих сторон мышц бедра взятым по три самцам голых мышей. Спустя 3 недели всех животных умерщвляют, мышцы бедра отделяют и исследуют их как с помощью рентгеновского излучения с низкой энергией, так и гистопатологически.
Как представлено в таблице 3, анализ путем рентгеновского излучения показывает плотный материал в местах инъекции в мышечную ткань всех продуцирующих MP-52 L-клеток. С помощью гистологических исследований можно установить простое хрящеобразование и кальцинозное хрящеобразование в мышках. Также эти результаты подтверждают, что MP-52 может индуцировать энхондральное костеобразование.
Осуществленные эксперименты подтверждают, что MP-52-протеин стимулирует образование хрящей из недифференцированных мезенхимных клеток, а также дифференциацию и созревание остеобластов. Это приводит к энхондральному костеобразованию, которое равно индукционному каскаду при эмбриональном костеобразовании и излечиванию (срастанию) костей при переломах.
Указанные в опытах условия нужно рассматривать в качестве иллюстрации MP-52-активности, а не как ограничение. Изобретение можно также рассматривать и охарактеризовывать в другой форме.
Для указанных в заявке линий клеток и плазмид в качестве приложения даются материалы, из которых очевидны подтверждающие данные.
Описание чертежей
На фиг. 1 представлено сравнение аминокислотной последовательности MP-52 с некоторыми членами семейства BMP-протеинов с началом на первом из семи сохранившихся цистеиновых остатков. Знак * обозначает, что аминокислота одинакова во всех сравниваемых протеинах; знак + обозначает, что аминокислота совпадает по меньшей мере в одном из протеинов по сравнению с MP-52.
На фиг. 2 представлены нуклеотидные последовательности олигонуклеотидных праймеров, которые применяются в настоящем изобретении, и сравнение этих последовательностей с известными членами TGF- β -семейства. M обозначает A или C; S обозначает C или G; R обозначает A или G; K обозначает G или T, "2a" обозначает последовательность праймера OD, "2b" обозначает последовательность праймера OID.
На фиг. 3 представлен вестерн-блоттинг, показывающий, что только в случае рекомбинантных вирусов, однако, не в случае вирусов дикого типа (без интегрированной чужеродной ДНК), появляются специфические для MP-52 полосы. M обозначает предварительно окрашенный маркер по молекулярной массе протеина с указанными кажущимися молекулярными массами (Cibco BRL # 26041-020), 1 обозначает надосадочную жидкость культуры клеток (100 мкл) после инфекции с помощью рекомбинантных вирусов (со вставленной MP-52-кДНК) при восстанавливающих (1% β- меркаптоэтанола) условиях, 2 обозначает надосадочную жидкость культуры клеток (100 мкл) после инфекции вирусами дикого типа (без вставленной чужеродной ДНК) при восстанавливающих (1% β -меркаптоэтанола) условиях, 3 обозначает надосадочную жидкость культуры клеток (500 мкл) после инфекции с помощью рекомбинантных вирусов (со вставленной MP-52 кДНК) при невосстанавливающих условиях, 4 обозначает надосадочную жидкость культуры клеток (500 мкл) после инфекции вирусами дикого типа (без вставленной чужеродной ДНК) при невосстанавливающих условиях.
На фиг. 4 представлена плазмида pABWN.
На фиг. 5 представлен разрез всего имплантата (после имплантации в течение 26 дней), окрашенный согласно Von Kossa. Минерализованная ткань отчетливо выделяется - черная - по сравнению с окружающей мышечной тканью.
На фиг. 6 представлен вырез из имплантата (после имплантации в течение 26 дней), окрашенный согласно Mass on Goldner. 1 обозначает край из остеобластов (в оригинале розовый), 2 обозначает остеоид (в оригинале красный), 3 обозначает минерализованную костную ткань (в оригинале зеленая) с остеоцитами (в оригинале розовые), 4 обозначает костный мозг (в оригинале от светло-розового до оранжевого).
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЙ ФАКТОР РОСТА/ДИФФЕРЕНЦИАЦИИ TGF-β-СЕМЕЙСТВА И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1994 |
|
RU2246315C2 |
| ДНК (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ БЕЛКА, БЕЛОК, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1993 |
|
RU2208638C2 |
| МАТЕРИАЛЫ, СТИМУЛИРУЮЩИЕ РОСТ ХРЯЩА/КОСТИ, ДЛЯ ПРЕПАРАТА | 1996 |
|
RU2180234C2 |
| ОПТИМИЗИРОВАННЫЙ СПОСОБ ОЧИСТКИ РЕКОМБИНАНТНОГО БЕЛКА ФАКТОРА РОСТА | 2008 |
|
RU2491290C2 |
| БИОРЕЗОРБИРУЕМЫЙ ПЕРЕВЯЗОЧНЫЙ МАТЕРИАЛ ДЛЯ РАН | 2012 |
|
RU2577155C2 |
| БИС-Met-ГИСТОНЫ | 2008 |
|
RU2498997C2 |
| КОМБИНАЦИИ АНТИТЕЛ, ОБЛАДАЮЩИХ СЕЛЕКТИВНОСТЬЮ ПО ОТНОШЕНИЮ К РЕЦЕПТОРУ ЛИГАНДА, ИНДУЦИРУЮЩЕМУ АПОПТОЗ, АССОЦИИРОВАННЫЙ С ФАКТОРОМ НЕКРОЗА ОПУХОЛИ, И ДРУГИХ ТЕРАПЕВТИЧЕСКИХ СРЕДСТВ | 2002 |
|
RU2313368C2 |
| АНТИТЕЛО, ОБЛАДАЮЩЕЕ СЕЛЕКТИВНОСТЬЮ ПО ОТНОШЕНИЮ К РЕЦЕПТОРУ ЛИГАНДА, ИНДУЦИРУЮЩЕМУ АПОПТОЗ, АССОЦИИРОВАННЫЙ С ФАКТОРОМ НЕКРОЗА ОПУХОЛИ, И ЕГО ИСПОЛЬЗОВАНИЕ | 2001 |
|
RU2298013C2 |
| СРЕДСТВА И СПОСОБ ПОЛУЧЕНИЯ ГЕМОПОЭТИЧЕСКОГО БЕЛКА | 1994 |
|
RU2233881C2 |
| ИНДУЦИРУЕМЫЙ ТЕПЛОМ ПРОМОТОР И ЕГО ИСПОЛЬЗОВАНИЕ | 2000 |
|
RU2230786C2 |











Изобретение относится к области молекулярной биологии и касается нового фактора роста/дифференциации ТGЕ-β-семейства и кодирующим его ДНК-последовательностям. Сущность изобретения включает кодирующую зрелый протеин часть (участок) и в случае необходимости другие функциональные части указанной в SEQ ID NO. 1 нуклеотидной последовательности, вектор, содержащий копию молекулы ДНК по п. 1, клетку-хозяин, трансформированную ДНК по п. 1, протеин, экспрессированный данными клетками, способ его получения, фармацевтические композиции, включающие данный протеин и поликлональные антитела к этому протеину. Изобретение позволяет получить ДНК-последовательности, которые кодируют новые члены семейства TGF-β протеинов с остеогенными свойствами. 7 с. и 2 з.п. ф-лы, 6 ил., 3 табл.
(б) нуклеотидную последовательность, соответствующую последовательности из п. (а) в силу вырожденности генетического кода, соответствующая нуклеотидной последовательности, которая кодирует протеин согласно последовательности 1DNO.2 или функциональные части из нее, приведенной в графической части,
(в) нуклеотидную последовательность, гибридизирующуюся с одной из последовательностей из п.п. (а, (б) при условии, что молекула ДНК по п. (в) кодирует, по меньшей мере, полностью часть зрелого протеина TGF-β-семейства, приведенную в графической части.
Приоритет по пунктам:
10.08.93 по пп.1, 4, 6 - 8, 9;
09.06.94 по пп.2 - 4, 5.
| US 5147854 A, 15.09.92 | |||
| US 4929442 A, 29.05.90 | |||
| WALE et al., Handbook of experimental pharmacology, 1990, v.95, pp.211 - 248. |

