Настоящее изобретение относится к антителам к рецептору инсулинподобного фактора роста I (IGF-IR), способам их получения, фармацевтическим композициям, содержащим указанные антитела, и их применению.
Рецептор инсулинподобного фактора роста I (IGF-IR, КФ 2.7.112, антиген CD 221) относится к семейству трансмембранных протеин-тирозин-киназ (LeRoith D. и др., Endocrin. Rev. 16, 1995, cc.143-163; и Adams Т.Е. и др., Cell. Mol. Life Sci. 57, 2000, cc.1050-1093). IGF-IR связывается с IGF-I с высокой аффинностью и инициирует физиологический ответ на этот лиганд in vivo. IGF-IR связывается также с IGF-II, однако с несколько более низкой аффинностью. Сверхэкспрессия IGF-IR стимулирует неопластическую трансформацию клеток, и имеются данные о том, что IGF-IR участвует в трансформации клеток в злокачественные клетки и поэтому является пригодной мишенью при разработке терапевтических средств лечения рака (Adams Т.Е. и др., Cell. Mol. Life Sci. 57, 2000, cc.1050-1093).
Антитела к IGF-IR хорошо известны в данной области, и изучено их противоопухолевое действие in vitro и in vivo (Benini S. и др., Clin. Cancer Res. 7, 2001, cc.1790-1797; Scotlandi K. и др., Cancer Gene Ther. 9, 2002, cc.296-307; Scotlandi K. и др., Int. J. Cancer 101, 2002, cc.11-16; Brunetti А. и др., Biochem. Biophys. Res. Commun. 165, 1989, cc.212-218; Prigent S.A. и др., J. Biol. Chem. 265, 1990, cc.9970-9977; Li S.L. и др., Cancer Immunol. Immunother. 49, 2000, cc.243-252; Pessino А. и др., Biochem. Biophys. Res. Commun. 162, 1989, cc.1236-1243; Surinya K.H. и др., J. Biol. Chem. 277, 2002, cc.16718-16725; Soos M.A. и др., J. Biol. Chem., 267, 1992, cc.12955-12963; Soos M.A. и др., Proc. Natl. Acad. Sci. USA 86, 1989, cc.5217-5221; O'Brien R.M. и др., ЕМВО J. 6, 1987, cc.4003-4010; Taylor R. и др., Biochem. J. 242, 1987, cc.123-129; Soos M.A. и др., Biochem. J. 235, 1986, cc.199-208; Li S.L. и др., Biochem. Biophys. Res. Commun. 196, 1993, cc.92-98; Delafontaine Р. и др., J. Mol. Cell. Cardiol. 26, 1994, cc.1659-1673; Kull F.C. Jr. и др. J. Biol. Chem. 258, 1983, cc.6561-6566; Morgan D.O. и Roth R.A., Biochemistry 25, 1986, cc.1364-1371; Forsayeth J.R. и др., Proc. Natl. Acad. Sci. USA 84, 1987, cc.3448-3451; Schaefer E.M. и др., J. Biol. Chem. 265, 1990, cc.13248-13253; Gustafson T.A. и Rutter W.J., J. Biol. Chem. 265, 1990, cc.18663-18667; Hoyne P.A. и др., FEBS Lett. 469, 2000, cc.57-60; Tulloch P.A. и др., J. Struct. Biol. 125, 1999, cc.11-18; Rohlik Q.T. и др., Biochem. Biophys. Res. Comm. 149, 1987, cc.276-281; и Kalebic T. и др., Cancer Res. 54, 1994, cc.5531-5534; Adams Т.Е. и др., Cell. Mol. Life Sci. 57, 2000, cc.1050-1093; Dricu А. и др., Glycobiology 9, 1999, cc.571-579; Kanter-Lewensohn L. и др., Melanoma Res. 8, 1998, cc.389-397; Li S.L. и др., Cancer Immunol. Immunother. 49, 2000, cc.243-252). Антитела к IGF-IR описаны также в многочисленных других публикациях, например, у Arteaga C.L. и др., Breast Cancer Res. Treatment 22, 1992, cc.101-106; и Hailey J. и др., Mol. Cancer Ther. 1, 2002, cc.1349-1353.
В частности, моноклональное антитело к IGF-IR, имеющее название αIR3, нашло широкое применение при исследовании опосредуемых IGF-IR процессов и опосредуемых IGF-I заболеваний, таких как рак. Альфа-IR-3 описано у Kull F.C., J. Biol. Chem. 258, 1983, cc.6561-6566. С тех пор опубликовано около ста работ, посвященных исследованию и терапевтическому применению αIR3, в которых рассматривается его противоопухолевое действие индивидуально и в сочетании с цитостатическими агентами, такими как доксорубицин и винкристин. αIR3 представляет собой мышиное моноклональное антитело, в отношении которого известно, что оно ингибирует связывание IGF-I с рецептором IGF, но не ингибирует связывание IGF-II с IGF-IR. В высоких концентрациях αIR3 стимулирует пролиферацию опухолевых клеток и фосфорилирование IGF-IR (Bergmann U. и др., Cancer Res, 55, 1995, cc.2007-2011; Kato H. и др., J. Biol. Chem. 268, 1993, cc.2655-2661). Существуют другие антитела (например, 1Н7, см. Li S.L. и др., Cancer Immunol. Immunother. 49, 2000, cc.243-252), которые ингибируют связывание IGF-II с IGF-IR в большей степени, чем связывание IGF-I. Обзор уровня техники, касающийся антител и их свойств и характеристик, приведен у Adams Т.Е. и др., Cell. Mol. Life Sci. 57, 2000, cc.1050-1093.
Большинство антител, известных из существующего уровня техники, имеют происхождение из организма мыши. Как хорошо известно, из существующего уровня техники, такие антитела нельзя применять в терапии человека без дополнительных изменений, таких как химеризация или гуманизация. Вследствие указанных недостатков в качестве терапевтических агентов для лечения людей наиболее предпочтительно применяют человеческие антитела. Человеческие антитела хорошо известны из существующего уровня техники (van Dijk М.А. и van de Winkel J.G., Curr. Opin. Chem. Biol. 5, 2001, cc.368-374). С использованием этой технологии можно получать человеческие антитела к широкому разнообразию мишеней. Примеры человеческих антител к IGF-IR представлены в WO 02/053596, WO 2004071529, WO 2005016967, WO 2006008639, US 20050249730, US 20050084906, WO 2005058967, WO 2006013472, US 20030165502, WO 2005082415, WO 2005016970, WO 03106621, WO 04083248, WO 2003100008, WO 2004087756, WO 2005005635 и WO 2005094376.
Однако все еще сохраняется потребность в антителах к IGF-IR, обладающих выраженными благоприятными действиями для пациентов, нуждающихся в противоопухолевой терапии. Проще говоря, благоприятное действие для пациента означает уменьшение роста опухоли и существенное увеличение времени до прогрессирования заболевания, обусловленное лечением с применением противоонкогенного средства.
У Routier F.H. и др., Glycoconjugate J. 14, 1997, cc.201-207, описана схема гликозилирования гуманизированного антитела в виде IgG1, экспрессируемого в клетках линии CHO-DUKX. Для этого антитела молярное соотношение остатков фукозы и маннозы (Fuc:Man) составляет 0,8:3,0, что соответствует степени гликозилирования, составляющей 80%. У Niwa R. и др., J. Immunol. Methods 306, 2005, cc.151-160, описаны антитела к CD20 в виде IgG1 и IgG3, которые рекомбинантно продуцируются в клетках линии СНО DG44, имеющие степень фукозилирования 90% и 91% соответственно. У Mimura Y. и др., J. Immunol. Methods 247, 2001, cc.205-216, описано, что бутират повышает производство человеческого химерного IgG в клетках линии СНО-K1, сохраняя при этом функцию и профиль гликозилирования. Олигосахаридные профили свидетельствуют о значительном содержании нефукозилированных гликановых структур. У Raju T.S., BioProcess International 1, 2003, cc.44-53, описано влияние вариации гликозилирования, обусловленной экспрессионными системами, на биологическую активность и номенклатуру терапевтических иммуноглобулинов. У Ма S. и др., Anal. Chem. 71, 1999, cc.5185-5192 описан анализ углеводного состава ритуксимаба. Ритуксимаб имеет степень фукозилирования 9-10% (Niwa R. и др., J. Immunol. Methods 306, 2005, cc.151-160). У Fujii S., J. Biol. Chem 265, 1990, cc.6009-6018 указано, что бычий IgG содержит примерно 11% нефукозилированного IgG. У Mizuochi Т., J. Immunol. 129, 1982, cc.2016-2020, приведены данные о том, что человеческий IgG является примерно на 14% нефукозилированным. У Bergwerff A.A., Glycoconjugate J. 12, 1995, cc.318-330 указано, что антитела, продуцируемые в организме мышей линии SP2/0, содержат в больших количествах олигосахариды N-гликолилнейраминовой кислоты (NGNA). У Nahrgang S. и др., In: Animal Cell Technology: Products from Cells, Cells as Products, под ред. Bernard А. и др., изд-во Kluwer Academic Publishers, Dordrecht, NL, 1999, cc.259-261, указано, что при экспрессии IgG1 в клетках СНО после кратковременной трансфекции обнаружено в целом незначительное гликозилирование. У Lund J. и др., Mol. Immunol. 30, 1993, cc.741-748 описано рекомбинантное получение мышиного-человеческого антитела в клетках мышиной трансфектомы. Антитело в виде IgG1 является нефукозилированным примерно на 13%. У Patel Т.Р. и др., Biochem. J. 285, 1992, cc.839-845, описано гликозилирование антител, полученных из клеток гибридомы и мышиных асцитов. У Niwa R. и др., J. Immunol. Methods 306, 2005, cc.151-160, указано, что антитело в виде IgGI к CD20, полученное методом рекомбинации в клетках СНО DG44, имеет фукозилирование, составляющее 91%, а у Mori К. и др., Biotech. Bioeng. 88, 2004, cc.901-908, указано, что фукозилирование составляет 94%. У Davies J. и др., Biotechnol. Bioeng. 74, 2001, cc.288-294, указано, что экспрессия антител с измененными гликоформами приводит к повышению антитело-обусловленной клеточнозависимой цитотоксичности (ADCC). У Sheeley D.M. и др., Anal. Biochem. 247, 1997, cc.102-110, приведено сравнение гликозилирования антител, полученных в различных экспрессионных системах. У Shields R.L. и др., J. Biol. Chem. 277, 2002, cc.26733-26740, указано, что отсутствие фукозы на Fc-фрагменте человеческого IgG1 повышает связывание FcγRIII и повышает ADCC. Антитело Her2, которое имеет степень гликозилирования, составляющую примерно 90%, обладает также значительной ADCC. У Zhu L. и др., Nature Biotechnol. 23, 2005, cc.1159-1169, описано получение человеческих антител в куриных яйцах.
Краткое изложение сущности изобретения
Изобретение относится к антителу, обладающему способностью к связыванию с IGF-IR, которое представляет собой человеческое антитело IgG1- или IgG3-типа и является гликозилированным сахарной цепью на остатке Asn297, где антитело отличается тем, что количество остатков фукозы в сахарной цепи составляет по меньшей мере 98% («полностью фукозилированное антитело», предпочтительные версии приведены ниже) и, кроме того, количество остатков NGNA составляет 1% или менее и/или количество остатков N-концевой альфа-1,3-галактозы составляет 1% или менее.
В контексте изобретения понятие «количество» означает количество сахарных остатков в сахарной цепи, присоединенной к Asn297, взятое по отношению к сумме G0, G1, G2 (без маннозы (4 и 5)), принимаемой за 100%, и его рассчитывают, как описано в примере 3.
Согласно изобретению можно создавать антитела, связывающиеся с IGF-IR, которые имеют степень фукозилирования, составляющее даже 99,4% или более, 99,5% или более или 99,9% или более.
Предпочтительно количество NGNA составляет 0,5% или менее, более предпочтительно 0,1% или менее, и даже представляет собой количество, не обнаруживаемое с помощью ЖХ-МС (жидкостная хроматография/масс-спектрометрия).
Предпочтительно количество N-концевой альфа-1,3-галактозы составляет 0,5% или менее, более предпочтительно 0,1% менее, и даже представляет собой количество, не обнаруживаемое с помощью ЖХ-МС.
Сахарная цепь предпочтительно обладает характеристиками N-связанных гликанов, присоединенных к Asn297 антитела, которое связывается с IGF-IR, рекомбинантно экспрессируемого в клетке СНО (клетка яичника китайского хомячка).
Предпочтительно клетка СНО представляет собой клетку СНО, имеющую делецию (например, DG44) или функциональную инактивацию обоих аллелей DHFR или делецию одного аллеля DHFR и функциональную инактивацию второго аллеля DHFR (например, DXB11).
Предпочтительно антитело представляет собой моноклональное антитело. Предпочтительно антитело представляет собой химерное, гуманизированное или человеческое антитело.
Изобретение предпочтительно относится к полностью фукозилированному антителу, обладающему способностью связываться с IGF-IR и ингибировать связывание IGF-I и IGF-II с IGF-IR, где антитело отличается тем, что оно обладает одним или несколькими свойствами, выбранными из группы, включающей:
а) соотношение значений IC50, характеризующих ингибирование связывания IGF-I с IGF-IR и ингибирование связывания IGF-II с IGF-IR, составляет от 1:3 до 3:1;
б) в концентрации 5нМ ингибирует по меньшей мере на 80%, предпочтительно по меньшей мере на 90% фосфорилирование IGF-IR по данным клеточного анализа фосфорилирования с использованием клеток линии НТ29 в среде, содержащей 0,5% инактивированной тепловой обработкой фетальной телячьей сыворотки (FCS), по сравнению с фосфорилированием по данным анализа без антитела;
в) не обладает IGF-IR-стимулирующей активностью (отсутствие передачи сигнала, отсутствие IGF-1-подобной активности), измеряемой по РKВ-фосфорилированию при использовании концентрации 10 мкМ в клеточном анализе фосфорилирования с использованием клеток линии 3Т3, имеющих 400000-600000 молекул IGF-IR на клетку, в среде, содержащей 0,5% инактивированной тепловой обработкой фетальной телячьей сыворотки (FCS) по сравнению с фосфорилированием в анализе без антитела;
г) осуществляет понижающую регуляцию на 50% или более IGF-IR, экспрессируемых на опухолевых клетках (например, линии НТ29) через 24 ч после добавления антитела к клеткам.
Антитела, предлагаемые в изобретении, обладают благоприятным действием для пациента, нуждающегося в противоопухолевой терапии, и обеспечивают снижение роста опухоли и значительное увеличение времени до прогрессирования заболевания. Антитела, предлагаемые в изобретении, обладают новыми и имеющими признаки изобретения свойствами, оказывают благоприятное действие на пациента, страдающего заболеванием, ассоциированным с нарушением регуляции IGF, прежде всего заболеванием, связанным с опухолью. Антитела, предлагаемые в изобретении, отличаются указанными выше свойствами.
При создании изобретения неожиданно было установлено, что антитело, предлагаемое в изобретении («полностью фукозилированное антитело»), не вызывает ADCC (антитело-обусловленная клеточнозависимая цитотоксичность) (ее величина отклоняется не более чем на 3×С.К.О. (стандартное отклонение) от используемого в качестве эталона стандартного антитела (референс-антитело) (антитело к гемоцианину лимфы улитки, т.е. антитело к KLH)) как продемонстрировано в ADCC-анализе, описанном в примере).
Предпочтительно антитело, которое обладает способностью специфически связываться с IGF-IR, ингибировать связывание IGF-1 и IGF-II с IGF-IR в указанном выше соотношении, представляет собой антитело IgG1-изотипа и оно не активирует передачу сигнала IGF-IR даже в клетках, сверхэкспрессирующих IGF-IR, при концентрациях, в 200 раз превышающих его значение IC50.
Антитела, связывающиеся с IGF-IR, которые не обладают «IGF-1-подобной активностью» и характеризуются «полным фукозилированием», имеют выраженное преимущество при применении в качестве терапевтического средства.
Предпочтительно антитела, предлагаемые в изобретении, в концентрации 5нМ полностью ингибируют трансдукцию опосредуемого IGF-I сигнала IGF-IR в опухолевых клетках.
Ниже перечислены кодируемые нуклеиновыми кислотами полипептиды, которые обладают способностью к сборке с соответствующей другой цепью антитела с образованием антитела, предлагаемого в изобретении:
а) тяжелая цепь антитела, содержащая в качестве CDR CDR1 (ак 31-35), CDR2 (ак 50-66) и CDR3 (ак 99-107) последовательности, представленной в SEQ ID NO:1 или 3;
б) легкая цепь антитела, содержащая в качестве CDR CDR1 (ак 24-34), CDR2 (ак 50-56) и CDR3 (ак 89-98) последовательности, представленной в SEQ ID NO:2 или 4.
Предпочтительно антитело представляет собой моноклональное антитело и, кроме того, химерное антитело (содержащее человеческую константную цепь), гуманизированное антитело и наиболее предпочтительно человеческое антитело.
Антитело предпочтительно конкурирует с антителом 18 за связывание с человеческим IGF-IR (КФ 2.7.1.112, SwissProt P08069):
Кроме того, антитело предпочтительно характеризуется аффинностью к связыванию, составляющей 10-8 М (KD) или менее, предпочтительно от примерно 10-9 до 10-13 М.
Предпочтительно антитело не обладает обнаруживаемой зависимостью способности ингибировать связывание инсулина с рецептором инсулина от концентрации.
Предпочтительно антитело представляет собой антитело IgG1-типа.
Антитело, предлагаемое в изобретении, значительно увеличивает время до прогрессирования заболевания на моделях с использованием соответствующих ксенотрансплантатов опухолей по сравнению с животными, обработанными наполнителем, и замедляет рост опухоли. Антитело ингибирует связывание IGF-I и IGF-II с IGF-IR in vitro и in vivo, предпочтительно в примерно одинаковой степени как касательно IGF-I, так и IGF-II.
Предпочтительно антитела, предлагаемые в изобретении, содержат в качестве гипервариабельных участков (CDR) следующие последовательности:
а) тяжелую цепь антитела, содержащую в качестве CDR CDR1 (ак 31-35), CDR2 (ак 50-66) и CDR3 (ак 99-107) последовательности, представленной в SEQ ID NO:1 или 3;
б) легкую цепь антитела, содержащую в качестве CDR CDR1 (ак 24-34), CDR2 (ак 50-56) и CDR3 (ак 89-98) последовательности, представленной в SEQ ID NO:2 или 4.
Предпочтительные вариабельные области и CDR, прежде всего CDR3 тяжелой цепи антител, предлагаемых в изобретении, получают из клона 18 <IGF-1R> HUMAB (антитело 18) и клона 22 <IGF-1R> HUMAB (антитело 22), депонированных в Немецкой коллекции микроорганизмов и клеточных культур ГмбН (DSMZ), Германия.
Указанные антитела подробно описаны в WO 2005/005635. Другие предпочтительные вариабельные области и CDR, прежде всего CDR3 тяжелой цепи антител, предлагаемых в изобретении, получают из клона 1а <IGF-1R> HuMab (антитело 1А, At 1A или Ak 1A), клона 23 <IGF-1R> HuMab (антитело 23) и клона 8 <IGF-1R> HuMab (антитело 8), депонированных в Немецкой коллекции микроорганизмов и клеточных культур ГмбН (DSMZ), Германия:
Указанные антитела подробно описаны в WO 2005/087756.
Изобретение относится также к способам рекомбинантного получения таких антител.
Изобретение относится также к способам лечения рака, заключающимся в том, что пациенту, у которого диагностирован рак (и который, следовательно, нуждается в противоопухолевой терапии), вводят в эффективном количестве антагонистическое антитело к IGF-IR, предлагаемое в изобретении. Антитело можно вводить индивидуально в виде фармацевтической композиции или в альтернативном варианте в сочетании с цитотоксическим лечением, таким как радиотерапия, или с цитотоксическим средством или его пролекарством.
Кроме того, изобретение относится к применению антитела, предлагаемого в изобретении, для лечения рака и для приготовления фармацевтической композиции, предлагаемой в изобретении. Изобретение относится также к способу приготовления фармацевтической композиции, предлагаемой в изобретении.
Кроме того, изобретение относится к фармацевтической композиции, содержащей антитело, предлагаемое в изобретении, в фармацевтически эффективном количестве необязательно в сочетании с буфером и/или адъювантом, который можно применять в фармацевтических целях для приготовления лекарственных форм.
В изобретении предложены также фармацевтические композиции, содержащие такие антитела в фармацевтически приемлемом носителе. В одном из вариантов осуществления изобретения фармацевтическую композицию можно включать в состав изделия или набора.
Кроме того, изобретение относится к способу получения рекомбинантного человеческого антитела, предлагаемого в изобретении, отличающемуся тем, что осуществляют экспрессию нуклеиновой кислоты, кодирующей антитело, которое обладает способностью связываться с IGF-IR, в клетке-хозяине, представляющей собой СНО-клетку, в результате чего происходит полное фукозилирование антитела, и выделяют антитело из клетки. Изобретение относится также к антителу, которое можно получать таким методом рекомбинации.
Краткое описание чертежа
На чертеже показана столбиковая диаграмма, иллюстрирующая ADCC-активность (или ее отсутствие) антител, предлагаемых в изобретении, и контроля и антител, используемых для сравнения.
Подробное описание изобретения
Понятие «антитело» относится к различным формам антител, включая (но, не ограничиваясь только ими) полные антитела, фрагменты антител, человеческие антитела, гуманизированные антитела и созданные с помощью генной инженерии антитела, если они сохраняют отличительные признаки, предлагаемые в изобретении.
Понятие «фрагменты антитела» относится к части полноразмерного антитела, как правило, по меньшей мере к его антигенсвязывающему центру или вариабельной области. Примерами фрагментов антител являются двойные антитела, молекулы одноцепочечных антител, иммунотоксины и мультиспецифические антитела, образованные из фрагментов антител.
Понятия «моноклональное антитело» или «композиция моноклонального антитела» в контексте настоящего описания относятся к препарату, содержащему молекулы антитела, которые имеют одинаковый состав аминокислот. Соответственно понятие «человеческое моноклональное антитело» относится к антителам, обладающим одинаковой специфичностью к связыванию, которые имеют вариабельную и константную области, полученные из последовательностей иммуноглобулина человеческой зародышевой линии.
Понятие «химерное антитело» относится к моноклональному антителу, содержащему вариабельную область, т.е. связывающую область, из одного источника или видов и по меньшей мере часть константной области из другого источника или других видов, которое, как правило, получают с помощью методов рекомбинантной ДНК. Особенно предпочтительными являются химерные антитела, содержащие мышиную вариабельную область и человеческую константную область. Такие мышиные/человеческие химерные антитела представляют собой продукт экспрессированных генов иммуноглобулина, которые содержат ДНК-сегменты, кодирующие вариабельные области мышиного иммуноглобулина, и ДНК-сегменты, кодирующие константные области человеческого иммуноглобулина. Другие формы «химерных антител», подпадающие под объем настоящего изобретения, представляют собой такие формы, в которых класс или подкласс модифицирован или заменен относительно исходного антитела. Такие «химерные» антитела обозначают также как «антитела переключенного класса». Методы получения химерных антител включают общепринятые методы, основанные на применении рекомбинантной ДНК и генной трансфекции, которые в настоящее время хорошо известны в данной области (см., например, Morrison S.L. и др., Proc. Natl. Acad Sci. USA 81, 1984, cc.6851-6855; US 5202238 и 5204244).
Понятие «гуманизированное антитело» относится к антителам, в которых каркасный участок или «гипервариабельные участки» (CDR) модифицированы таким образом, что они содержат CDR иммуноглобулина другой специфичности по сравнению с родительским иммуноглобулином. В предпочтительном варианте осуществления изобретения для получения «гуманизированного антитела» мышиный CDR трансплантируют в каркасный участок человеческого антитела (см., например, Riechmann L. и др., Nature 332, 1988, cc.323-327; и Neuberger M.S. и др., Nature 314, 1985, cc.268-270). Наиболее предпочтительные CDR соответствуют репрезентативным последовательностям, которые распознают антигены, указанные выше для химерных и бифункциональных антител.
В контексте настоящего описания подразумевается, что понятие «человеческое антитело» относится к антителам, имеющим вариабельную и константную области, полученные из последовательностей иммуноглобулина человеческой зародышевой линии. Вариабельную тяжелую цепь предпочтительно получают из последовательности зародышевой линии DP-50 (GenBank L06618), а вариабельную легкую цепь предпочтительно получают из последовательности зародышевой линии L6 (GenBank X01668), или вариабельную тяжелую цепь предпочтительно получают из DP-61 (GenBank М99682), а вариабельную легкую цепь предпочтительно получают из последовательности зародышевой линии L15 (GenBank K01323). Константные области антитела представляют собой константные области человеческого иммуноглобулина IgG1-типа. Такие области могут быть аллотипическими, они описаны, например, у Johnson G. и Wu Т.Т., Nucleic Acids Res. 28, 2000, cc.214-218, и в указанных в этой публикации ссылках.
Понятие «рекомбинантное человеческое антитело» относится к антителам, имеющим вариабельную и константную области, которые получены из последовательностей иммуноглобулина человеческой зародышевой линии и находятся в преобразованной форме. Рекомбинантные человеческие антитела, предлагаемые в изобретении, были подвергнуты in vivo соматической гипермутации. Так, аминокислотные последовательности VH- и VL-областей рекомбинантных антител представляют собой последовательности, которые, хотя и выведены из последовательностей VH- и VL-областей человеческой зародышевой линии и являются родственными им, могут не встречаться in vivo в спектре зародышевой линии человеческого антитела.
В контексте настоящего описания понятие «связывание» относится к связыванию антитела с IGF-IR, которое характеризуется аффинностью, составляющей примерно от 10-13 до 10-8 М (KD), предпочтительно примерно от 10-13 до 10-9 М.
В контексте настоящего описания подразумевается, что понятие «молекула нуклеиновой кислоты» включает молекулы ДНК и молекулы РНК. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, но предпочтительно она представляет собой двухцепочечную ДНК.
Человеческие константные области IgG1- или IgG3-типа подробно описаны у Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-е изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD., 1991, и у Bruggemann M. и др., J. Exp. Med. 166, 1987, cc.1351-1361; Love T.W. и др., Methods Enzymol. 178, 1989, cc.515-527. Их примерами являются последовательности, представленные в SEQ ID NO:5-8. Другие пригодные и предпочтительные константные области представляют собой константные области антител, которые можно получать из линий клеток гибридомы, депонированных в DSMZ для целей настоящего изобретения.
Константные области IgG1- или IgG3-типа гликозилированы на остатке Asn297. В контексте настоящего описания «Asn 297» означает аминокислоту аспарагин, локализованную примерно в положении 297 в Fc-фрагмента; вследствие небольших вариаций последовательностей антител Asn297 может находиться также на несколько аминокислот (как правило, не более чем на ±3 аминокислоты) выше или ниже. Например, в одном из антител, предлагаемых в изобретении, «Asn297» находится в положении 298 аминокислотной последовательности.
Гликозилирование человеческого IgG1 или IgG3 имеет место на Asn297 в виде гликозилирования, осуществляемого с помощью сложного биантенного олигосахарида с коровым фукозилированием, на концах которого располагаются вплоть до 2 остатков Gal. Эти структуры обозначены как гликановые остатки G0, G1 (α1,6 или α1,3) или G2, в зависимости от количества концевых остатков Gal (Raju T.S., BioProcess Int. 1, 2003, cc.44-53). СНО-тип гликозилирования Fc-фрагментов антитела описан, например, у Routier F.H., Glycoconjugate J. 14, 1997, сс.201-207.
Понятие «вариабельная область» (вариабельная область легкой цепи (VL), вариабельная область тяжелой цепи (VH)) в контексте настоящего описания относится к каждой из пары легких и тяжелых цепей, которые непосредственно принимают участие в связывании антитела с антигеном. Домены вариабельных областей легких и тяжелых цепей имеют одинаковую общую структуру, и каждый домен содержит 4 каркасных участка (FR), последовательности которых являются высококонсервативными, связанных тремя «гипервариабельными участками» (или определяющими комплементарность областями, CDR). Каркасные участки адаптированы к β-складчатой конформации, а CDR-участки могут формировать петли, соединяющие β-складчатую структуру. CDR в каждой цепи поддерживаются в виде своей трехмерной структуры с помощью каркасных участков и формируют с CDR-участками другой цепи антигенсвязывающий центр. СОР3-участки тяжелой и легкой цепи антитела играют наиболее важную роль в специфичности связывания/аффинности антител, предлагаемых в изобретении, и вследствие этого являются дополнительным объектом изобретения.
Понятия «гипервариабельный участок» или «антигенсвязывающий центр антитела» в контексте настоящего описания относятся к аминокислотным остаткам антитела, которые ответственны за связывание с антигеном. Гипервариабельный участок содержит аминокислотные остатки из «определяющих комплементарность областей» или «CDR». «Каркасные» или «FR»-участки представляют собой участки вариабельной области, остатки которых отличны от остатков гипервариабельного участка, как они определены в настоящем описании. Таким образом, легкие и тяжелые цепи антитела содержат в направлении от N- к С-концу домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. CDR3 тяжелой цепи представляет собой участок, который обеспечивает главным образом связывание с антигеном. К CDR- и FR-участкам относят участки, которые определяют согласно стандартному определению Кэбота и др. (Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991) и/или остатки из «гипервариабельной петли».
В контексте настоящего описания понятие «связывание с IGF-IR» означает связывание антитела с IGF-IR, обнаруживаемое в анализе in vitro, предпочтительно в анализе связывания, в котором антитело связывают с поверхностью и связывание IGF-IR измеряют с помощью резонанса поверхностного плазмона (SPR). Наличие связывания означает, что аффинность к связыванию (KD) составляет 10-8 М или менее, предпочтительно от 10-13 до 10-9 М.
Связывание с IGF-IR можно исследовать с помощью BIAcore-анализа (фирма Pharmacia Biosensor AB, Уппсала, Швеция). Аффинность к связыванию определяют в понятиях ka (константа скорости ассоциации антитела при формировании комплекса антитело/антиген), kd (константа диссоциации) и KD (kd/ka). Антитела, предлагаемые в изобретении, характеризуются значением KD, составляющим 10-10 M или менее.
Антитела, предлагаемые в изобретении, ингибируют также связывание IGF-I и IGF-II с IGF-IR. Ингибирование оценивают по величине IC50 в анализе связывания IGF-I/IGF-II с IGF-IR на опухолевых клетках. Такой анализ описан в примере 7. В таком анализе измеряют количество несущего радиоактивную метку IGF-I или IGF-II или их IGF-IR-связывающих фрагментов, связанных с IGF-IR, находящимся на поверхности указанных опухолевых клеток (например, линии НТ29), в отсутствие антитела и в присутствии возрастающих концентраций антитела. Значения IC50 для антител, предлагаемых в изобретении, характеризующие ингибирование связывания IGF-I и IGF-II с IGF-IR, составляют не более 2 нМ и отношение значений IC50, характеризующих ингибирование связывания IGF-I/IGF-II с IGF-IR, составляет примерно от 1:3 до 3:1. Значения IC50 определяют как средние или медианные значения по результатам по меньшей мере трех независимых измерений. Отдельные значения IC50 могут выпадать из указанного диапазона.
В контексте настоящего описания понятие «ингибирование связывания IGF-I и IGF-II с IGF-IR» относится к ингибированию связывания меченного с помощью I125 IGF-I или IGF-II с IGF-IR, присутствующим на поверхности опухолевых клеток линии НТ29 (АТСС НТВ-38), в анализе in vitro. Наличие ингибирования означает, что значение IC50 составляет 2 нМ или менее.
Понятие «клетки, экспрессирующие IGF-IR» относится к таким клеткам, в которых происходит сверхэкспрессия рецептора IGF-I с уровнем, составляющим по меньшей мере примерно 20000 рецепторов/клетку. Такими клетками являются, например, линии опухолевых клеток, такие как NCI H322M или НТ29, или линия клеток (например, 3Т3 АТСС CRL1658), которая сверхэкспрессирует IGF-IR после трансфекции экспрессионным вектором, обеспечивающим экспрессию IGF-IR. Количество рецепторов на клетку оценивают согласно методу, описанному у Lammers, R. и др., EMBO J. 8, 1989, cc.1369-1375.
Понятие «ингибирование фосфорилирования IGF-IR» относится к ингибированию, которое выявляют с помощью клеточного анализа фосфорилирования с использованием клеток линии 3Т3, экспрессирующих по 400000-600000 молекул IGF-IR на клетку в среде, содержащей 0,5% инактивированной тепловой обработкой фетальной телячьей сыворотки (FCS), при сравнении с результатами такого анализа без присутствия антитела. Фосфорилирование обнаруживают методом Вестерн-блоттинга с использованием антитела, специфического в отношении белков с фосфорилированным остатком тирозина. Такой анализ описан в примере 11. Тепловую инактивацию FCS осуществляют путем кратковременного нагрева до 56°C с целью инактивации системы комплемента.
Понятие «ингибирование фосфорилирования протеинкиназы В (РKВ)» относится к ингибированию, которое выявляют с помощью клеточного анализа фосфорилирования с использованием клеток линии 3Т3, экспрессирующих по 400000-600000 молекул IGF-IR на клетку в среде, содержащей 0,5% инактивированной тепловой обработкой фетальной телячьей сыворотки (FCS), при сравнении с результатами такого анализа без присутствия антитела. Фосфорилирование обнаруживают методом Вестерн-блоттинга с использованием антитела, специфического в отношении РKВ, фосфорилированной на остатке серина в положении 473 РКВ (Akt 1, Swiss Prot Асе. No. P31749). Такой анализ описан в примере 11.
Понятие «антитело-обусловленная клеточнозависимая цитотоксичность (ADCC)» относится к лизису человеческих опухолевых клеток-мишеней с помощью антитела, предлагаемого в изобретении, в присутствии эффекторных клеток. ADCC предпочтительно измеряют путем обработки препарата клеток, которые экспрессируют IGF-IR, антителом, предлагаемым в изобретении, в присутствии эффекторных клеток, таких как свежевыделенные РВМС (мононуклеарные клетки периферической крови) или очищенные эффекторные клетки из лейкоцитных пленок, такие как моноциты или NK-клетки (естественные клетки-киллеры).
Понятие «полное ингибирование опосредуемой IGF-I трансдукции сигнала» относится к ингибированию опосредуемого IGF-I фосфорилирования IGF-IR. В таком анализе клетки, экспрессирующие IGF, предпочтительно клетки линии Н322М, стимулируют IGF-I и обрабатывают антителом, предлагаемым в изобретении (можно применять антитело в концентрации 5нМ или более). Затем осуществляют ДСН-ПААГ и измеряют фосфорилирование IGF-IR методом Вестерн-блоттинга с использованием антитела, специфического в отношении фосфорилированного остатка тирозина. Считается, что имеет место полное ингибирование трансдукции сигнала, если на Вестерн-блоте визуально нельзя обнаружить полосы, соответствующей фосфорилированному IGF-IR.
Антитела, предлагаемые в изобретении, предпочтительно обладают способностью связываться с тем же самым эпитопом IGF-IR, что и антитело 18, или имеет место ингибирование их связывания с IGF-IR вследствие стерической помехи, обусловленной связыванием с антителом 18. Ингибирование связывания можно обнаруживать с помощью SPR-анализа с использованием иммобилизованного антитела 18 и IGF-IR в концентрации 20-50 нМ и антитела, подлежащего обнаружению, в концентрации 100 нМ. Уменьшение сигнала на 50% или более означает, что антитело конкурирует с антителом 18. Такой анализ можно осуществлять аналогичным образом с использованием антитела 22 в качестве иммобилизованного антитела.
Понятие «эпитоп» обозначает белковую детерминанту, обладающую способностью специфически связываться с антителом. Эпитопы, как правило, состоят из химически активных расположенных на поверхности групп молекул, таких как боковые цепи аминокислот или сахаров, и, как правило, имеют специфические характеристики трехмерной структуры, а также специфические характеристики зарядов. Конформационные и неконформационные эпитопы различаются тем, что связывание с эпитопами первого, но не второго типа, нарушается в присутствии денатурирующих растворителей.
Антитела, предлагаемые в изобретении, ингибируют фосфорилирование тирозина IGF-IR и предпочтительно также фосфорилирование тирозина РKB в близкой степени.
Антитела, предлагаемые в изобретении, предпочтительно осуществляют понижающую регуляцию уровня белка IGF-IR в опухолевых клетках и в опухолях, например в ксенотрансплантированных опухолях.
Антитела, предлагаемые в изобретении, предпочтительно ингибируют трехмерное разрастание опухолевых клеток в анализе образования колоний, а также пролиферацию клеток, экспрессирующих IGF-IR (например, клеток линии NIH 3Т3).
Антитела, предлагаемые в изобретении, предпочтительно не ингибируют связывание инсулина с рецептором инсулина в анализе конкурентного связывания на клетках линии 3Т3, сверхэкспрессирующих рецептор инсулина, в концентрации 200 нмолей/л.
Антитела, предлагаемые в изобретении, получают методом рекомбинации в СНО-клетках, где происходит полное фукозилирование антитела. Для осуществления экспрессии белка нуклеиновые кислоты, кодирующие легкую и тяжелую цепи или их фрагменты, встраивают в экспрессионные векторы с помощью стандартных методов. Экспрессию осуществляют в таких СНО-клетках и антитело выделяют из клеток (из супернатанта или клеток после лизиса).
Пригодные СНО-клетки-хозяева можно получать с помощью метода, заключающегося в том, что культивируют СНО-клетки, трансфектированные нуклеиновой кислотой, которая кодирует антитело, предлагаемое в изобретении, в условиях давления отбора с использованием DHFR, отбирают отдельные клоны, размножают клоны и отбирают клон, продуцирующий антитело, имеющее схему гликозилирования, предлагаемую в изобретении. Предпочтительно культивирование осуществляют в течение по меньшей мере двух, предпочтительно по меньшей мере трех недель. СНО-клетка предпочтительно представляет собой клетку линии DG44.
Понятие «СНО-клетка» относится к различным формам клеток яичника китайского хомячка (СНО), основанных на двух неактивных, предпочтительно уделенных путем делеции аллелях dhfr (дефицит дигидрофолатредуктазы (dhfr-)). Такие dhfr--клетки и методы их получения описаны, например, у Urlaub G. И др., Cell 33, 1983, cc.405-412; Chasm L. и др., Som. Cell Molec. Genet. 12, 1986, cc.555-556; Kolkekar A.S. и др., Biochemistry 36, 1997, cc.10901-10909. Предпочтительно клетка представляет собой клетку линии DG44. Такие dhfr--клетки СНО можно получать с использованием гамма-излучения для удаления всего локуса dhfr. В клетках дикого типа, не имеющих мутацию, dhfr является ферментом, необходимым для синтеза de novo глицина, пуринов и тимидилата. Это позволяет использовать ген dhfr, кодируемый плазмидами, в качестве доминантного селектируемого маркера и гена-амплификатора для экспрессии белков в dhfr--клеточных линиях (клеточных линиях с дефицитом dhfr). dhfr--мутация в клетках линии DG44 является стабильной и необратимой. СНО-клетки, успешно котрансфектированные экспрессионным(и) вектором(ами), несущими антитело человеческого IgG1- или IgG3-типа и ген DHFR, должны иметь фенотип dhfr+ и их можно легко отбирать путем выращивания колоний на средах, не содержащих тимидин и гипоксантин и необязательно содержащих метотрексат (МТХ) для амплификации.
Клетки линии DG44 хорошо известны из существующего уровня техники, и их можно покупать в виде поступающих на рынок линий клеток, например, от фирмы Invitrogen Corp. (США). Клетки линии DG44 могут расти в прикрепленном состоянии, в суспензии и/или в бессывороточной среде. В контексте настоящего описания понятия «клетка», «клеточная линия» и «клеточная культура» используются взаимозаменяемо, и все такие обозначения применительно к dhfr--клеточным линиям СНО (с делецией двух аллелей dhfr) включают потомство. Так, понятия «трансформанты» и «трансформированные клетки» относятся к первичной рассматриваемой клетке и полученным из нее культурам безотносительно к количеству пересевов. Следует понимать также, что все потомство может не быть полностью идентичным по составу ДНК из-за преднамеренных или случайных мутаций. Под объем изобретения подпадают варианты потомства, имеющие такие же особенности схемы гликозилирования, которые выбраны путем скрининга исходных трансформированных клеток.
Предпочтительно dhfr--клеточную линию СНО подвергают коамплификации по меньшей мере с DHFR, в качестве единственного селектируемого маркерного гена. Например, осуществляют совместную трансфекцию клеток-реципиентов СНО экспрессионным вектором для экспрессии в клетках млекопитающих, содержащим селектируемый(е) маркер(ы) и ген антитела. Образовавшиеся колонии можно подвергать селекции, колонии, обладающие ожидаемым фенотипом, обладают способностью экспрессировать антитело. Дополнительные серектируемые маркеры не являются иди могут не являться доминантными. Примерами дополнительных селектируемых маркеров, применяемых для котрансфекции, являются аденозиндезаминаза (Kaufman R.J. и др., Proc. Natl. Acad. Sci. USA 83, 1986, cc.3136-3140), аспарагинсинтетаза (Cartier M. и др., Mol. Cell Biol. 7, 1987, cc.1623-1628), ген trpB E.coli и ген hisD Salmonella (Hartman S.C. и Mulligan R.C., Proc. Natl. Acad. Sci. USA 85, 1988, cc.8047-8051), мышиная рибонуклеотидредуктаза М2 (Thelander M. и Thelander L., EMBO J. 8, 1989, cc.2475-2479), человеческий ген, обусловливающий устойчивость к многим лекарственным средствам (Kane S.E. и др., Gene 84, 1989, с.439-446), глутаминсинтетаза (Bebbington C.R. и др., DNA Cloning, том III, под ред. D.M. Glover, изд-во IRL Press, 1987, cc.163-188), ксантин-гуанин-фосфорибозилтрансфераза (gpt) (Mulligan R.C. и Berg, P., Science 209, 1980, cc.1422-1427), гигромицин В (Santerre R.F. и др., Gene 30, 1984, cc.147-156), ген неомицина (Southern P.J. и Berg P., J. Mol. Appl. Genet. 1, 1982, cc.327-341). Селектируемые маркеры могут также представлять собой основу, с помощью которой можно осуществлять амплификацию генов, кодирующих антитело. При осуществлении котрансфекции линии клеток СНО векторные ДНК часто интегрируют в хромосому клетки в один и тот же локус. Таким образом, использование только одного из селектируемых маркеров в качестве основы для амплификации, как правило, приводит к параллельному увеличению количества копий обоих генов. Одним из конкретных селектируемых маркеров, применяемых таким путем, является dhfr, который позволяет осуществлять требуемую амплификацию при использовании возрастающих концентраций МТХ. Вторым предпочтительным селектируемым маркером является GS, который позволяет осуществлять амплификацию при добавлении метионинсульфоксимина (MSX).
Селектируемые маркеры, конечно, находятся под контролем регуляторных элементов ДНК, что обеспечивает их экспрессию. В случае использования dhfr в качестве селектируемого маркера предпочтительно применяют регуляторные элементы из вирусных источников, из таких, как ДНК опухолевых вирусов. Особенно предпочтительно применять главный поздний промотор SV40 или аденовируса. В этом случае особенно предпочтительно удалять энхансерный элемент из промотора, тем самым эффективно «повреждая» его. Такая модификация позволяет повышать уровни амплификации гена при каждой выбранной концентрации метотрексата, что в ином случае достигается при использовании сильного промотора. В случае применения неомицина в качестве селектируемого маркера, примером пригодного промотора является мышиный промотор металлотионеина.
Общие методы рекомбинантного получения антител хорошо известны из существующего уровня техники, и они описаны, например, в обзорных статьях Makrides S.C., Protein Expr. Purif. 17, 1999, cc.183-202; Geisse S. и др., Protein Expr. Purif. 8, 1996, cc.271-282; Kaufman R.J., Mol. Biotechnol. 16, 2000, cc.151-161; Werner R.G., Drug Res. 48, 1998, cc.870-880.
Антитела могут присутствовать в целых клетках, в супернатанте, клеточном лизате или могут находиться в частично очищенной или практически чистой форме. Очистку для удаления других клеточных компонентов или других примесей, например, других клеточных нуклеиновых кислот или белков, осуществляют с помощью стандартных методов, включая обработку щелочью/ДСН, CsCl-бэндинг, хроматографию на колонках, электрофорез в агарозном геле и другие методы, хорошо известные в данной области (см. в Current Protocols in Molecular Biology, под ред. Ausubel F. и др., изд-во Greene Publishing and Wiley Interscience, New York, 1987).
Контролирующие последовательности, которые можно применять для прокариотических организмов, представляют собой, например, промотор, необязательно операторную последовательность и сайт связывания рибосом. Известно, что для эукариотических клеток применяют промоторы, энхансеры и сигналы полиаденилирования.
Нуклеиновая кислота «функционально связана», когда она находится в функциональной связи с другой нуклеотидной последовательностью. Например, ДНК предпоследовательности или лидерной секреторной последовательности функционально связана с ДНК полипептида, если при ее экспрессии образуется предбелок, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он оказывает воздействие на транскрипцию последовательности; или сайт связывания рибосом функционально связан с кодирующей последовательностью, если он расположен так, что облегчает трансляцию. Как правило, понятие «функционально связана» означает, что последовательности ДНК, будучи связаны, являются смежными, а в случае лидерной секреторной последовательности, являются смежными и находятся в рамке считывания. Однако не требуется, чтобы энхансеры были смежными. Связывание осуществляют путем лигирования в соответствующих сайтах рестрикции. Если такие сайты не существуют, то в соответствии с принятой практикой применяют синтетические олигонуклеотидные адаптеры или линкеры.
Моноклональные антитела можно отделять от культуральной среды гибридомы с помощью общепринятых методов очисти иммуноглобулинов, таких, например, как, хроматография на протеин А-сефарозе, хроматография на гидроксилапатите, гель-электрофорез, диализ или аффинная хроматография. ДНК и РНК, которые кодируют моноклональные антитела, можно легко выделять из гибридомы и секвенировать с помощью общепринятых методов. Клетки гибридом могут служить источником таких ДНК и РНК, После идентификации и выделения ДНК можно встраивать в экспрессионные векторы, которыми затем трансфектируют СНО-клетки, которые в противном случае не могут продуцировать белок иммуноглобулина, для синтеза рекомбинантных моноклональных антител в клетках-хозяевах.
Изобретение относится также к иммуноконъюгатам, содержащим антитело, предлагаемое в изобретении, конъюгированное с цитотоксическим агентом, таким как химиотерапевтический агент, токсин (например, обладающий ферментативной активностью токсин бактериального, грибного, растительного или животного происхождения или его фрагменты), радиоактивный изотоп (т.е. радиоконъюгат) или пролекарство цитотоксического агента. Агенты, которые можно применять для получения таких иммуноконъюгатов, описаны выше. К обладающим ферментативной активностью токсинам и их фрагментам, которые можно применять, относятся цепь А токсина дифтерии, несвязывающиеся активные фрагменты токсина дифтерии, цепь А эндотоксина (из Pseudomonas aeruginosa), цепь А рицина, цепь А абрина, цепь А модеццина, альфа-сарцин, белки Aleuritesfordii, белки диантина, белки Phytolaca americana (PAPI, PAPII, и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены.
Конъюгаты антитела и цитотоксического агента получают с использованием разнообразных бифункциональных агентов для сшивания белков, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат·HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидопроизводные (такие как бис(пара-азидобензоил)гександиамин), производные бис-диазония (такие как бис(пара-диазонийбензоил)этилендиатнин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные фторсодержащие соединения (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получать согласно методу, описанному у Vitetta E.S. и др., Science 238, 1987, cc.1098-1104). Меченная с помощью С14 1-изотиоцианатбензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) является примером хелатирующего агента, применяемого для конъюгации радионуклеотида с антителом (см. WO 94/11026).
Следующим объектом настоящего изобретения является композиция, например, фармацевтическая композиция, которая содержит антитело, предлагаемое в настоящем изобретении, включенное в состав препаративной формы в сочетании с фармацевтически приемлемым носителем.
Фармацевтические композиции, предлагаемые в изобретении, можно применять также в совместной терапии, т.е. в сочетании с другими средствами. Например, для совместной терапии можно применять композицию, предлагаемую в настоящем изобретении, в сочетании по меньшей мере с одним противоопухолевым средством, таким как химиотерапевтический агент, цитотоксический агент или пролекарство, или другой общепринятой терапией.
«Химиотерапевтическое средство» представляет собой химическое соединение, которое можно применять при лечении рака. Примерами химиотерапевтических средств являются адриамицин, доксорубицин, 5-фторурацил, цитозина арабиносид («Ara-С»), циклофосфамид, тиотепа, таксотер (доксетаксел), бусульфан, гемцитабин, цитоксин, таксол, метотрексат, цисплатин, мелфалан, винбластин, блеомицин, этопосид, ифосфамид, митомицин С, митоксантрон, винкристин, винорелбин, карбоплатин, тенипосид, дауномицин, карминомицин, аминоптерин, дактиномицин, митомицины, эсперамицины (см. US 4675187), мелфалан и другие родственные производные горчичного газа.
В контексте описания понятие «цитотоксическое средство» относится к субстанции, которая ингибирует или препятствует функции клеток и/или вызывает деструкцию клеток. Следует понимать, что понятие включает радиоактивные изотопы, химиотерапевтические средства и токсины, такие как обладающие ферментативной активностью токсины бактериального, грибного, растительного или животного происхождения.
В контексте настоящего описания понятие «пролекарство» относится к предшественнику или производному обладающей фармацевтической активностью субстанции, которое обладает меньшей цитотоксичностью в отношении опухолевых клеток, чем родительское лекарственное средство, и обладает способностью активироваться или превращаться под действием ферментов в более активную родительскую форму (см., например, Wilman D.E., Biochemical Society Transactions 14, 1986, cc.375-382, и Stella V.J. и др., «Prodrugs: A Chemical Approach to Targeted Drug Delivery», в: Directed Drug Delivery, под ред. Borchardt R.T., изд-во Humana Press, Clifton, New Jersey, 1985, cc.247-267). К пролекарствам, предлагаемым в настоящем изобретении, относятся (но не ограничиваясь только ими) фосфатсодержащие пролекарства, тиофосфатсодержащие пролекарства, сульфатсодержащие пролекарства, пептидсодержащие пролекарства, пролекарства с модифицированными D-аминокислотами, гликозилированные пролекарства, содержащие β-лактамовое кольцо пролекарства, пролекарства, содержащие необязательно замещенный феноксиацетамид или пролекарства, содержащие необязательно замещенный фенилацетамид, 5-фторцитозин- и другие 5-фторуридинсодержащие пролекарства, которые могут превращаться в обладающие большей цитотоксической активностью свободные лекарственные средства. Примерами цитотоксических лекарственных средств, которые можно дериватизировать с получением формы в виде пролекарства для применения согласно настоящему изобретению, являются (но, не ограничиваясь только ими), указанные выше химиотерапевтические средства.
В контексте настоящего описания понятие «фармацевтически приемлемый носитель» включает любой и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, придающие изотоничность и замедляющие абсорбцию агенты и т.п., которые являются физиологически совместимыми. Предпочтительно носитель можно применять для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии).
В контексте настоящего описания понятие «фармацевтически приемлемая соль» относится к соли, которая сохраняет требуемую биологическую активность антитела и не оказывает никаких нежелательных токсикологических действий (см., например, Berge S.M. и др., J. Pharm. Sci. 66, 1977, cc.1-19). Такие соли подпадают под объем изобретения. Примерами таких солей являются кислотно-аддитивные соли и соли присоединения оснований. К кислотно-аддитивным солям относятся соли, образованные с нетоксичными неорганическими кислотами, такие как гидрохлориды.
Композицию, предлагаемую в настоящем изобретении, можно вводить с помощью различных методов, известных в данной области. Как должно быть очевидно специалисту в данной области, путь и/или форма введения может варьироваться в зависимости от требуемых результатов.
Для введения соединения, предлагаемого в изобретении, с помощью определенных путей введения может оказаться необходимым наносить на соединение покрытие из материала, препятствующего его инактивации, или осуществлять введение соединения совместно с таким материалом. Например, соединение можно вводить индивидууму в соответствующем носителе, например, в липосомах или в разбавителе. К фармацевтически приемлемым разбавителям относятся физиологический раствор и водные забуферивающие растворы.
К фармацевтически приемлемым носителям относятся стерильные водные растворы или дисперсии и стерильные порошки для приготовления стерильных инъекционных растворов или дисперсий непосредственно перед введением. Применение таких сред и агентов для обладающих фармацевтической активностью субстанций известно в данной области.
В контексте настоящего описания понятия «парентеральное введение» и «введение парентеральным путем» означают пути введения, отличные от энтерального и местного введения, как правило, они относятся к введению путем инъекции и включают (но, не ограничиваясь только ими) внутривенную, внутримышечную, внутриартериальную, подоболочечную, внутриглазную, внутрисердечную, внутрикожную, внутрибрюшинную, транстрахеальную, подкожную, субкутикулярную, внутрисуставную, субкапсулярную, субарахноидальную, интраспинальную, эпидуральную и интрастернальную инъекцию и инфузию.
Указанные композиции могут содержать также адъюванты, такие как консерванты, смачивающие агенты, эмульгаторы и диспергирующие агенты. Отсутствие микроорганизмов можно обеспечивать как с помощью процедур стерилизации (см. выше), так и путем включения различных антибактериальных и противогрибковых средств, таких, например, как парабен, хлорбутанол, фенол, сорбиновая кислота и т.п. Может оказаться целесообразным включать в композиции агенты для придания изотоничности, такие как сахара, хлорид натрия и т.п. Кроме того, можно пролонгировать абсорбцию инъекционной фармацевтической формы путем включения веществ, которые замедляют абсорбцию, таких как моностеарат алюминия и желатин.
Вне зависимости от выбранного пути введения соединения, предлагаемые в настоящем изобретении, которые можно применять в пригодной гидратированной форме, и/или фармацевтические композиции, предлагаемые в настоящем изобретении, приготавливают в виде фармацевтически приемлемых форм лекарственного средства с помощью общепринятых методов, известных специалистам в данной области.
Фактические уровни доз действующих веществ в фармацевтических композициях, предлагаемых в настоящем изобретении, можно варьировать для получения количества действующего вещества, которое является эффективным для достижения требуемого терапевтического ответа у конкретного пациента при использовании конкретной композиции и пути введения, но не является токсичным для пациента. Выбранный уровень доз должен зависеть от различных фармакокинетических факторов, включая активность конкретных применяемых композиций, предлагаемых в настоящем изобретении, путь введения, время введения, скорость экскреции конкретного применяемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или материалы, которые используют в сочетании с конкретными применяемыми композициями, возраст, пол, вес, состояние, общее состояние здоровья и предшествующую историю болезни пациента, подлежащего лечению, и другие подобные факторы, хорошо известные в области медицины.
Композиция должна быть стерильной и текучей в той степени, чтобы ее можно было вводить с помощью шприца. Помимо воды предпочтительным носителем является изотонический забуференный физиологический раствор.
Соответствующую текучесть можно поддерживать, например, путем использования покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ. Во многих случаях предпочтительно включать в композицию агенты для придания изотоничности, например, сахара, многоатомные спирты, такие как маннит или сорбит, и хлорид натрия.
Предпочтительно полностью фукозилированное антитело, предлагаемое в изобретении, следует применять для лечения немелкоклеточного рака легкого (NSCLC) в сочетании с эрлотинибом (Tarceva®), для лечения рака молочной железы - в сочетании с Herceptin® (трастузумаб) и опухолей поджелудочной железы - предпочтительно в сочетании с гемцитабином (Gemzar®).
Ниже приведены примеры, чертеж и перечень последовательностей для облегчения понимания настоящего изобретения, истинный объем которого изложен в прилагаемой формуле изобретения. Следует иметь ввиду, что в описанные процедуры можно вносить модификации без отклонения от сущности изобретения.
Примеры
Линии клеток
Родительская линия клеток, которую использовали для создания линии клеток, экспрессирующих рекомбинантный IgG, представляла собой линию клеток яичника китайского хомячка (СНО) CHO-DG44 (Flintoff W.F. и др., Somat. Cell Genet. 2, 1976, cc.245-261; Flintoff и др., Mol. Cell. Biol. 2, 1982, cc.275-285; Urlaub G. и др., Cell 33, 1983, cc.405-412; Urlaub G. и др., Somat. Cell Mol. Genet. 12, 1986, cc.555-566). В клетках линии CHO-DG44 отсутствуют оба эндогенных локуса фермента дигидрофолатредуктазы (DHFR).
Клетки линии CHO-DG44 выращивали в среде MEM alpha Minus (фирма Gibco, №22561), дополненной 10% диализированной FCS (фирма Gibco, №26400-044) и 2 ммолями/л L-глутамина, 100 мкМ гипоксантином, 16 мкМ тимидином (НТ-добавка).
Плазмиды
Экспрессионная система содержала промотор CMV, и она описана в таблице 1. В качестве антитела использовали антитело к IGF-1R (WO 2005005635; АK18 или АK22).
Пример 1
Трансфекция и отбор
Трансфекцию экспрессионной плазмидой осуществляли с помощью препарата Fugene (фирма Roche Diagnostics GmbH). Через один день после трансфекции клетки линии DG44 подвергали давлению отбора в среде, представляющей собой среду MEM alpha Minus, дополненную 10% диализированной PCS и 2 ммолями/л L-глутамина и 20 нМ метотрексатом (МТХ). После выдерживания в течение 3 недель в условиях давления отбора с планшета отбирали отдельные клоны и размножали.
Собирали супернатанты и присутствие антитела выявляли с помощью специфического в отношении человеческого IgG ELISA. Затем субклоны размножали и анализировали в отношении продуцирования специфического антитела.
Клоны адаптировали к росту в суспензионной культуре и бессывороточной среде HyQ SFM4 CHO-Utility (HyClone № SH30516), дополненной 20 нМ МТХ. Параллельно определяли профиль гликозилирования. Отбирали субклоны, которые характеризовались степенью дефукозилирования, составляющей 2,0% или менее (по отношению к общему молярному количеству олигосахаридов).
Пример 2
Культивирование и очистка
Клетки (3×105) выращивали во встряхиваемых колбах вместимостью 125 мл (фирма Coming), содержащих по 30 мл среды, при 37°C, 5% СО2, 100 об./мин. Плотность клеток измеряли с помощью счетчика типа CASY Counter и супернатант отбирали для определения концентрации антитела с помощью аффинной хроматографии на протеине А. Примерно по 20 мл каждого супернатанта очищали для дальнейшей биохимической характеризации с помощью хроматографии на протеине А (уравновешивание ЗФР, отмывка 25 мМ натрий-цитратным буфером, рН 5,2, элюирование 100 мМ натрий-цитратным буфером, рН 2,8, CIP с использованием 10 мМ NaOH).
Пример 3
Анализ гликоструктуры антитела
Материал, представляющий собой очищенное антитело, анализировали с использованием пептидной карты, полученной с помощью жидкостной хроматографии/масс-спектрометрии (ЖХ-МС). Образцы восстанавливали (0,4М Трис/HCl, 8М гуанидин/HCl, рН 8,5, ДТТ (3 мг/мл), карбоксиконцы метилировали (йодуксусная кислота) и расщепляли трипсином. Пептидную-гликопептидную смесь разделяли с помощью ОФ-ЖХВР и анализировали в он-лайновом режиме с помощью масс-спектрометрии с ионизацией электроспреем. Интегрировали m/z-спектры содержащего гликоструктуру пептида, результаты представлены в таблице 2.
Клон 5 линии клеток СНО (hu MAb<IGF-1R>B1-4E10_9-16) был депонирован в соответствии с Будапештским договором о международном признании депонирования микроорганизмов для целей процедуры патентования в Немецкой коллекции микроорганизмов и клеточных культур ГмбН (DSMZ)), Германия, 21 июня 2006 г. под регистрационным номером DSM АСС 2795.
Среды, применяемые для культивирования различных клонов, получали от фирмы Hyclone (HyQ SFM4 CHO-Utility применяли для клонов 4-6) или фирмы Sigma (C-8862 применяли для клонов 1-3 и 7).
Анализ полученной с помощью ЖХ-МС пептидной карты осуществляли путем интегрирования специфических ионных хроматограмм всех заряженных состояний для всех гликопептидов.
Разветвленные GlcNac, NGNA и структуры с высоким содержанием маннозы определяли таким же образом.
Разветвленные GlcNac и NGNA не были обнаружены. Следовательно, количество NGNA составляет 0,5% или менее, а также 0,1% или менее. Количество разветвленных GlcNac также составляет 0,5% или менее, и 0,1% или менее.
Пример расчета степени гликозилирования представлен в таблице 3 (таблица 2а: клон 3, таблица 3б: клон 5; пептид, содержащий asn298, обозначен как Н27).
площадь: площадь пика;
H27_GO-H27_G4: гликопептид Н27 (содержащий Asn298) с фукозилированным углеводом биантенного сложного типа, содержащим х-концевую галактозу (например, G4 с 4 единицами галактозы);
относительное количество без Fuc: процентное содержание Fuc по отношению ко всем G0, G1, G2 без маннозной (4 и 5) гликоструктуры (высокое содержание маннозы);
H27_G1_1NGNA-H27_G3_2NGNA: гликопептид Н27 (содержащий Asn298) с фукозилированным углеводом биантенного сложного типа, содержащим х-концевые галактозные звенья (например, G2 с 2 звеньями галактозы), который имеет один-два остатка N-гликолилнейраминовой кислоты;
относительное количество без Fuc: процентное содержание Fuc по отношению ко всем G0, G1, G2 без маннозной (4 и 5) гликоструктуры (с высоким содержанием маннозы).
Пример 4
Оценка эффекторных функций, опосредуемых антителами к IGF-IR HuMAb
Для оценки способности созданных антител HuMAb вызывать иммунные эффекторные механизмы проводили исследования антитело-зависимой клеточно-опосредованной цитотоксичности (ADCC).
Для изучения воздействия антител на ADCC, клетки рака предстательной железы линии DU145 (НТВ081 АТСС; 1×106 в 2-4 мл среды RPMI-FM), экспрессирующие IGF-IR, метили с использованием 1 мкл раствора бис(ацетоксиметил)-2,2':6'2"-терпиридин-6,6"-дикарбоксилата (BATDA) в течение 25 мин при 37°C в инкубаторе для клеток. Клетки отмывали четыре раза с помощью 10 мл среды RPMI-FM и центрифугировали в течение 10 мин при 200×g с торможением. После этого концентрацию клеток доводили до 1×105 клеток/мл. Клетки высевали из расчета 5000 клеток/лунку в круглодонный планшет в объеме, соответствующем 50 мкл. Антитела HuMAb добавляли в конечной концентрации 25-0,1 нг/мл в 50 мкл среды для культуры клеток. Затем добавляли 50 мкл эффекторных клеток, таких как РВМС, свежевыделенных из цельной крови, или очищенных эффекторных клеток из лейкоцитных пленок, в соотношении Е:Т, составляющем 25:1. Сразу после этого планшеты центрифугировали в течение 1 мин при 200×g с торможением и инкубировали в течение 2 ч при 37°C. После инкубации клетки центрифугировали в течение 10 мин при 200×g и 20 мкл супернатанта переносили в титрационный микропланшет типа Optiplate 96-F. Добавляли 200 мкл раствора европия (при комнатной температуре) и смесь инкубировали в течение 15 мин на шейкере. Испускаемую флуоресценцию измеряли с помощью флуориметра с временным разрешением с использованием протокола EU-TDA фирмы Perkin Elmer.
Степень лизиса клеток в результате ADCC выражали в виде % от максимального высвобождения TDA из клеток-мишеней, подвергшихся лизису в результате действия поверхностно-активного вещества, скорректированного на спонтанное высвобождение TDA из соответствующих клеток-мишеней. В качестве используемого для сравнения стандарта антитела, «не обладающего ADCC», использовали (моноклональное) антитело к KLH (гемоцианин лимфы улитки) такого же IgG-типа или смесь IgG, полученную примерно от 35000 доноров («Redimune»). В качестве положительного контроля использовали антитело к IGF-IR с уменьшенным на 75% содержанием фукозы. Антитело, предлагаемое в изобретении, вызывало высвобождение TDA, которое находилось в пределах 3×С.К.О. относительно высвобождения TDA, вызываемого стандартным антителом (фиг.1).
Пример 5
Определение аффинности антител к IGF-IR к IGF-IR
Для оценки аффинности антитела к человеческому FCγ (из кролика) связывали с поверхностью чипа с целью презентации антитела для IGF-IR. Добавляли внеклеточный домен IGF-IR в различных концентрациях в растворе. Ассоциацию оценивали, осуществляя инъекцию IGF-IR в течение 3 мин; диссоциацию оценивали, осуществляя отмывку поверхности чипа буфером в течение 5 мин. Результаты оценки аффинности антител 18 и 22 представлены в таблице 4.
Пример 6
Ингибирование связывания IGF-I и IGF-II с опухолевыми клетками, экспрессирующими IGF-IR
Для оценки способности антитела, предлагаемого в изобретении, блокировать связывание лигандов IGF-I и IGF-II с рецептором IGF-I (IGF-IR) осуществляли эксперименты по конкурентному связыванию с использованием радиоактивномеченых пептидов-лигандов.
Человеческие опухолевые клетки (НТ29, NCI H322M, 0,5-1×105/мл) высевали в среду RPMI 1640 (фирма РАА, каталожный номер Е15-039), дополненную 2 мМ L-глутамином, 1 × заменимыми аминокислотами (фирма Gibco, каталожный номер 11140-035), 1 мМ пируватом натрия (фирма Gibco, каталожный номер 11360-039) и 10% инактивированной тепловой обработкой FCS (фирма РАА, каталожный номер А15-771). Для осуществления эксперимента инокулировали шесть колб формата Т175 20 мл клеток в соответствующей среде и культивировали в течение двух дней при 37°C и 5% CO2 до получения конфлюэнтных монослоев клеток.
Для сбора индивидуальных клеток добавляли по 2 мл 1 хтрипсин/ЭДТК (фирма Gibco, каталожный номер 25300-054) на Т175-колбу и наблюдали за отделением клеток с помощью микроскопа типа Zeiss Axiovert25. Клетки собирали и добавляли описанную выше среду, содержащую 10% FCS, до получения конечного объема 50 мл. Клетки повторно выделяли путем центрифугирования в течение 10 мин при 1000 об/мин (фирма Heraeus sepatech, центрифуга типа Omnifuge 2.0 RS) и ресуспендировали в 50 мл буфера для связывания (120 мМ NaCl, 5 мМ KСl, 1,2 мМ MgSO4, 1 мМ ЭДТК, 10 мМ D(+)глюкоза, 15 мМ NaAc, 100 мМ Hepes, pH 7,6, 1% БСА). Клетки подсчитывали, повторно выделяли путем центрифугирования и доводили их концентрацию до 1×106 клеток/мл с помощью буфера для связывания.
Меченные с помощью I125 пептиды IGF-I и IGF-II (фирма Amersham, ~2000 Ки/ммоль, каталожные номера IM172 и IM238), солюбилизованные в 0,1% СН3СООН, разводили в буфере для связывания до достижения конечной активности, составляющей 4×105 импульсов /(мин × мл). 75 мкл антитела в определенных концентрациях вместе с 25 мкл предварительно разведенного меченного с помощью I пептида IGF-I или IGF-II добавляли к 200 мкл суспензии клеток и инкубировали в течение 3,5 ч при 4°C. Клетки повторно выделяли путем центрифугирования в течение 5 мин при 2000 об/мин (фирма Eppendorf, центрифуга типа 5415С) и удаляли супернатант. После двукратной отмывки в 1 мл буфера для связывания клетки ресуспендировали в 1 мл буфера для связывания и переносили в сцинтилляционные флаконы. Количество радиоактивного пептида, связанного с рецепторами клеточной поверхности, измеряли с помощью сцинтилляционного счетчика.
Средние значения IC50 для антитела 18 составляли 0,3 нМ. Не было выявлено обнаруживаемого ингибирования связывания IGF-II.
Пример 7
Анализ конкурентного связывания антитела с IGF-IR
Для эпитопного картирования моноклональных антител к IGF-IR был выбран формат, аналогичный формату для оценки аффинности (пример 5), но IGF-IR предварительно инкубировали по меньшей мере в течение 0,5 ч при КТ с антителом в растворе. Эту смесь инъецировали и выявляли связывание с IGF-IR (или его ингибирование). Этот анализ позволяет оценивать реципрокную ингибирующую активность моноклональных антител в отношении связывания с IGF-IR. Было установлено, что антитела, предлагаемые в изобретении, конкурируют за связывание с IGF-IR с антителом αIR3, которое, как известно, связывается с ак 217-274 (Gustafson Т.А. и Rutter, W.J., J. Biol. Chem. 265, 1990, cc.18663-18667).
Пример 8
Ингибирование опосредуемого IGF-I фосфорилирования IGF-IR и Akt/РKВ
Для оценки способности антитела, предлагаемого в изобретении, ингибировать активацию и фосфорилирование рецептора IGF-I (IGF-IR) осуществляли эксперименты по изучению конкурентного связывания с использованием пептида IGF-I и последующий анализ методом Вестерн-блоттинга с использованием антител, обладающих специфичностью в отношении фосфорилированного тирозина.
Человеческие опухолевые клетки (НТ29, NCI H322M, 5×104/мл) высевали в среду RPMI 1640 (фирма РАА, каталожный номер Е15-039), дополненную 2 мМ L-глутамином, 1 × заменимыми аминокислотами (фирма Gibco, каталожный номер 11140-035), 1 мМ пируватом натрия (фирма Gibco, каталожный номер 11360-039) и 0,5% инактивированной тепловой обработкой FCS (фирма РАА, каталожный номер А 15-771). Для оценки величин IC50 для каждого эксперимента осуществляли инокуляцию 12-луночных планшетов с использованием 1 мл клеток в соответствующей среде и культивировали в течение 2 дней при 37°C и 5% CO2.
После культивирования в течение 48 ч в среде с низким содержанием сыворотки среду осторожно удаляли и заменяли антителом, разбавленным соответствующей средой до различных концентраций. После инкубации в течение 5 мин при 37°C и 5% CO2 добавляли пептид IGF-I в конечной концентрации 2нМ и клетки еще раз инкубировали в течение 10 мин в указанных выше условиях. Среду осторожно удаляли путем аспирации и в каждую лунку добавляли по 100 мкл холодного буфера для лизиса (50 мМ Hepes, pH 7,2, 150 мМ NaCl, 1 мМ ЭГТК, 10% глицерина, 1% Triton®-X100. 100 мМ NaF, 10 мМ Na4P2O7, ингибитор протеаз Complete®). Клетки отделяли с помощью скребка для клеток (фирма Corning, каталожный номер 3010) и содержимое лунок переносили в реакционные пробирки Эппендорфа. Фрагменты клеток удаляли центрифугированием в течение 10 мин при 13000 об/мин и 4°C и половину супернатанта добавляли к 2× буферу Laemmli для образца в соотношении 1:1 (об./об.). Для иммунопреципитации IGF-IR оставшийся супернатант клеточных лизатов подвергали осветляющему центрифугированию (10 мин при 13000 об/мин и 4°C) непосредственно перед добавлением 1 мкл поликлонального антитела к IGF-IRβ (С-20, фирма Santa Cruz Biotechnologies) или мышиного моноклонального антитела (IgG1), которое распознает эпитоп в положениях 440-586 аминокислотной последовательности внеклеточного домена (α-цепь) человеческого рецептора IGF типа 1 (МАт 24-55, фирма GroPep). После инкубации в течение 2 ч при 4°C во вращающейся реакционной пробирке Эппендорфа добавляли 25 мкл гранул протеин G Sepharose® (фирма Amersham Biosciences, каталожный номер 17-0618-01), после чего осуществляли еще одну стадию инкубации в течение 1 ч при 4°C. Гранулы со связанными комплексами антитело-белок выделяли центрифугированием (1 мин при 2000 об/мин и 4°С) и отмывали трижды буфером для отмывки (буфер для лизиса, содержащий Triton®-X100 в концентрации, составляющей только 0,1%). После кипячения гранул в буфере Laemmli для образца клеточные белки выделяли с помощью ДСН-ПААГ и переносили на нитроцеллюлозную мембрану (PROTRAN® ВА 85, фирма Schleicher&Schuell) с помощью полусухого Вестерн-блоттинга.
Для определения статуса фосфорилирования иммуноочищенного IGF-IR использовали специфическое в отношении фосфотирозина антитело (фирма Upstate, клон 4G10, каталожный номер 05-321). Для обнаружения фосфорилированной Akt/PKB использовали антитело, обладающее специфичностью в отношении фосфорилированного Ser473 (фирма Cell Signalling, каталожный номер 9271).
Было установлено, что антитело 18 обладает способностью ингибировать опосредуемое IGF-I фосфорилирование IGF-IR и РKВ, которая характеризуется значением IC50, составляющим 0,5 нМ.
Пример 9
Индукция опосредуемой антителом понижающей регуляции IGF-IR in-vitro
Для оценки воздействий антитела, предлагаемого в изобретении, на количество рецептора IGF-I (IGF-IR) в опухолевых клетках проводили эксперименты по оценке их зависимости от времени и последующий анализ методом Вестерн-блоттинга с использованием специфических в отношении IGF-IR антител.
Использовали человеческие опухолевые клетки (линия НТ29, 5×104 клеток/мл) в среде RPMI 1640 (фирма РАА, каталожный номер Е15-039), дополненной 2 мМ L-глутамином, 1 × заменимыми аминокислотами (фирма Gibco, каталожный номер 11140-035), 1 мМ пируватом натрия (фирма Gibco, каталожный номер 11360-039) и 10% инактивированной тепловой обработкой FCS (фирма РАА, каталожный номер А15-771). Для каждого периода инкубации осуществляли инокуляцию одного 12-луночного планшета с использованием 1 мл клеток в соответствующей среде для каждого эксперимента и культивировали в течение 24 ч при 37°C и 5% СO2.
Среду осторожно удаляли и заменяли антителом, разведенным в соответствующей среде в различных концентрациях. В двух контрольных лунках среду заменяли либо средой, не содержащей антитело, либо содержащей контрольное антитело (АВ-1, фирма Oncogene, каталожный номер GR11). Клетки инкубировали при 37°C и 5% СO2 и через 15 мин, 24 ч и 48 ч отбирали индивидуальные планшеты для дальнейшего процессинга.
Среду осторожно удаляли путем аспирации и в каждую лунку добавляли по 100 мкл холодного буфера для лизиса (50 мМ Hepes, pH 7,2, 150 мМ NaCl, 1 мМ ЭГТК, 10% глицерина, 1% Triton®-X100, 100 мМ NaF, 10 мМ Na4Р2O7, ингибитор протеаз Complete®). Клетки отделяли с помощью скребка для клеток (фирма Corning, каталожный номер 3010) и содержимое лунок переносили в реакционные пробирки Эппендорфа. Фрагменты клеток удаляли путем центрифугирования в течение 10 мин при 13000 об/мин и 4°C и супернатант добавляли к 2× буферу Laemmli для образца в соотношении 1:1 (об./об.). Клеточные белки разделяли с помощью ДСН-ПААГ и переносили на нитроцеллюлозную мембрану (PROTRAN® ВА 85, фирма Schleicher&Schuell, каталожный номер 10 401196) с помощью полусухого Вестерн-блоттинга.
Для определения уровней белка IGF-IR применяли антитело, специфическое в отношении IGF-IR (C-20, фирма Santa Cruz Biotechnologies, каталожный номер sc-713).
Было установлено, что антитело, предлагаемое в изобретении, индуцирует понижающую регуляцию IGF-IR менее чем через 24 ч после добавления антитела.
Пример 10
Ингибирование связывания инсулина с 3Т3-клетками, экспрессирующими человеческий рецептор инсулина
Для определения того, блокирует ли также антитело, предлагаемое в изобретении, связывание инсулина с рецептором инсулина (IR), проводили эксперименты по конкурентному связыванию с использованием несущего радиоактивную метку пептида-лиганда.
Клетки линии 3Т3 (1×105/мл), рекомбинантно экспрессирующие в больших количествах человеческий IR (>105), высевали в модифицированную по способу Дульбекко среду MEM (DMEM) с высоким содержанием глюкозы (фирма РАА, каталожный номер Е15-009), дополненную 2 мМ L-глутамином (фирма Gibco, каталожный номер 25030-024) и 10% инактивированной тепловой обработкой FCS (фирма РАА, каталожный номер А15-771). Шесть колб формата Т175 инокулировали 20 мл клеток в соответствующей среде для каждого эксперимента и культивировали в течение двух дней при 37°C и 5% СO2 до получения конфлюэнтных монослоев клеток.
Для сбора индивидуальных клеток добавляли по 2 мл 1 × трипсин/ЭДТК (фирма Gibco, каталожный номер 25300-054) на Т175-колбу и отделение клеток выявляли с помощью микроскопа. Клетки собирали и добавляли среду, дополненную, как описано выше 10% FCS, до получения общего объема 50 мл. Клетки повторно выделяли путем центрифугирования в течение 10 мин при 1000 об/мин и ресуспендировали в 50 мл буфера для связывания (120 мМ NaCl, 5 мМ KСl, 1,2 мМ MgSO4, 1 мМ ЭДТК, 10 мМ D(+)глюкоза, 15 мМ NaAc, 100 мМ Hepes, рН 7,6, 1% БСА). Клетки подсчитывали, повторно выделяли путем центрифугирования и доводили с помощью буфера для связывания концентрацию клеток до 1×106 клеток/мл.
Меченный с помощью I125 пептид инсулина (фирма Amersham, каталожный номер IM166, ~2000 Ки/ммоль), солюбилизированный в 0,1% СН3СООН, разводили в буфере для связывания до конечной активности 4×105 импульсов/(мин-мл). Добавляли 75 мкл антитела вместе с 25 мкл предварительно разведенного меченного с помощью I125 пептида инсулина к 200 мкл суспензии клеток (конечная концентрация антитела 200нМ) и инкубировали в течение 3,5 ч при 4°C. Клетки повторно выделяли путем центрифугирования в течение 5 мин при 2000 об/мин и удаляли супернатант. После двукратной отмывки в 1 мл буфера для связывания клетки ресуспендировали в 1 мл буфера для связывания и переносили в сцинтилляционные флаконы. Количество несущего радиоактивную метку пептида, связанного с рецепторами клеточной поверхности, измеряли с помощью сцинтилляционного счетчика.
Результаты продемонстрировали, что антитело, предлагаемое в изобретении, не влияет на связывание лиганда-инсулина с рецептором инсулина.
Пример 11
Фосфорилирование IGF-IR и Akt/PKB в отсутствие стимуляции
Для того чтобы исключить стимулирующую IGF-IR активность антитела, предлагаемого в изобретении, проводили оценку фосфорилирования IGF-IR в отсутствие лиганда IGF-I, но в присутствии антитела, предлагаемого в изобретении и референс-антитела (αIR3, фирма Oncogene, Германия). Этот анализ проводили методом Вестерн-блоттинга с использованием специфических в отношении статуса фосфорилирования антител. Клетки линии 3Т3 (АТСС CRL 1658), трансфектированные IGF-IR (5×104 клеток/мл, Pietrzkowski, Z. и др., Cell Growth Differ. 4, 1992, cc.199-205) высевали в модифицированную по способу Дульбекко среду MEM (DMEM) с высоким содержанием глюкозы (фирма РАА, каталожный номер Е15-009), дополненную 2 мМ L-глутамином (фирма Gibco, каталожный номер 25030-024) и 0,5% инактивированной тепловой обработкой FCS (фирма РАА, каталожный номер А15-771) или человеческие опухолевые клетки (линия НТ29, NCI Н322М, 5×104/мл) в среду RPMI 1640 (фирма РАА, каталожный номер Е15-039), дополненную 2 мМ L-глутамином, 1× заменимыми аминокислотами (фирма Gibco, каталожный номер 11140-035), 1 мМ пируватом натрия (фирма Gibco, каталожный номер 11360-039) и 0,5% инактивированной тепловой обработкой FCS (фирма РАА, каталожный номер А15-771). Для определения величин IC50 12-луночные планшеты инокулировали 1 мл клеток в соответствующей среде для каждого эксперимента и культивировали в течение двух дней при 37°C и 5% CO2.
После культивирования в течение 48 ч в среде с низким содержанием сыворотки среду осторожно удаляли и заменяли антителом в различных концентрациях, разведенным в соответствующей среде. Клетки инкубировали в течение 15 мин в указанных выше условиях. Среду осторожно удаляли путем аспирации и добавляли в каждую лунку по 100 мкл холодного буфера для лизиса (50 мМ Hepes, рН 7,2, 150 мМ NaCl, 1 мМ ЭГТК, 10% глицерина, 1% Triton-Х100, 100 мМ NaF, 10 мМ Na4P2O7, ингибитор протеаз Complete™). Клетки отделяли с помощью скребка для клеток (фирма Corning, каталожный номер 3010) и содержимое лунок переносили в реакционные пробирки Эппендорфа. Фрагменты клеток удаляли путем центрифугирования в течение 10 мин при 13000 об/мин и 4°C (центрифуга Эппендорфа 5415R) и половину супернатанта добавляли к 2× буферу Laemmli для образца в соотношении 1:1 (об./об.). Для иммунопреципитации IGF-IR оставшийся супернатант клеточных лизатов подвергали осветляющему центрифугированию (10 мин при 13000 об/мин и 4°C) непосредственно перед добавлением 1 мкл антитела к IGF-IR (C-20, фирма Santa Cruz Biotechnologies, каталожный номер sc-713, или МАт 24-55, фирма GroPep, каталожный номер MAD1). После инкубации в течение 2 ч при 4°C во вращающейся реакционной пробирке Эппендорфа добавляли 25 мкл гранул протеин G Sepharose® (фирма Amersham Biosciences, каталожный номер 17-0618-01), после чего осуществляли еще одну стадию инкубации в течение 1 ч при 4°C. Гранулы со связанными комплексами антитело-белок выделяли центрифугированием (1 мин при 2000 об/мин и 4°C) и отмывали трижды буфером для отмывки (буфер для лизиса, содержащий только 0,1% Triton-X100). После кипячения гранул в буфере Laemmli для образца клеточные белки выделяли с помощью ДСН-ПААГ и переносили на нитроцеллюлозную мембрану (PROTRAN ВА 85, фирма Schleicher&Schuell) с помощью полусухого Вестерн-блоттинга.
Для определения статуса фосфорилирования иммуноочищенного IGF-IR использовали специфическое в отношении фосфотирозина антитело (фирма Upstate, клон 4G10, каталожный номер 05-321, распознающее белки с фосфорилированным остатком тирозина). Для обнаружения фосфорилированной Akt/PKB использовали антитело к Akt1, обладающее специфичностью в отношении фосфорилированного Ser473 (фирма Cell Signalling, каталожный номер 9271).
Было установлено, что происходила существенная активация киназы Akt/PKB, находящейся ниже в пути передачи сигнала IGF-IR, референс-антителом в концентрациях, превышающих 5нМ, но не происходила активация антителом, предлагаемым в изобретении, в концентрациях вплоть до 10000 нМ.
Пример 12
Индукция понижающей регуляции рецептора на моделях с использованием ксенотрансплантата Н322М
Осуществляли индукцию опухолей в организме бестимусных мышей и однократную обработку антителом, предлагаемым в изобретении, взятым в различных концентрациях. Через 24 ч после обработки опухоли удаляли и гомогенизировали в жидком азоте. Добавляли холодный буфер для лизиса (50 мМ Hepes, рН 7,2, 150 мМ NaCl, 1 мМ ЭГТК, 10% глицерина, 1% Triton-X100, 100 мМ NaF, 1 мМ Na3V04, 10 мМ Na4P2O7, ингибитор протеазы Complete™, 1 мМ фенилметилсульфонил-фторид (PMSF)) в соотношении объем буфера:масса опухоли, равном 3:1, и осторожно смешивали с оттаивающим гомогенатом опухоли. После солюбилизации ткани в течение 15 мин на льду нерастворимые фрагменты удаляли путем центрифугирования в течение 10 мин при 13000 об/мин и 4°C (центрифуга Эппендорфа 5415R). Концентрацию белка в образцах определяли с использованием реагентов Micro BCA™ (фирма Pierce) и добавляли буфер для лизиса до достижения одинаковых концентраций. Часть супернатанта добавляли к 2× буферу Laemmli для образца в соотношении 1:1 (об./об.). Клеточные белки разделяли с помощью ДСН-ПААГ и переносили на нитроцеллюлозную мембрану (PROTRAN ВА 85, фирма Schleicher&Schuell, каталожный номер 10 401196) с помощью полусухого Вестерн-блоттинга. Для обнаружения IGF-IR применяли специфическое в отношении IGF-IR антитело (С-20, фирма Santa Cruz Biotechnologies, каталожный номер sc-713).
После обработки антителом, предлагаемым в изобретении, было выявлено зависящее от концентрации уменьшение уровней IGF-IR, характеризующееся значением ЕС50, составляющим 0,6 мг/кг.
Пример 13
Ингибирование роста опухолей линии Н322М
Воздействие антитела 18 in vivo исследовали путем индукции опухолей у бестимусных мышей согласно утвержденным для этой цели методам. Человеческие клетки NSCLC линии Н322М вводили путем совместной подкожной инъекции с матригелем (Matrigel) бестимусным (nu/nu) мышам 6-7-недельного возраста. Для этой цели концентрировали 5×106 клеток линии Н322М в 100 мкл культуральной среды и смешивали с 100 мкл матригеля. 200 мкл этой смеси вводили путем инъекции в правый бок мышей. Объем опухоли рассчитывали на основе измерения диаметров опухоли с помощью штангенциркулей с нониусом дважды в неделю по формуле, впервые опубликованной Geran с соавторами («Protocols for screening chemical agents and natural products against animal tumors and other biological systems». Cancer Chemother. Rep.11.301, 1972), согласно которой объем опухоли [мм3]=(длина × (ширина)). Антитело вводили внутрибрюшинно (i.p.) в дозе 10 мл/кг. Лечение начинали с использованием двойных доз антитела, которые вводили в двойных объемах. Опухоли индуцировали у бестимусных мышей как описано выше. После того, как опухоли вырастали до средней массы, составляющей 160 мг, мышей обрабатывали путем внутрибрюшинного введения шесть раз в неделю по 6, 0,6 и 0,06 мг/кг антитела в виде последующих доз после введения ударных доз, составляющих 12, 1,2 и 0,12 мг/кг соответственно, которые вводили один раз в первый день лечения. Результаты эксперимента свидетельствуют о том, что блокирование IGF-IR-оси с помощью рекомбинантного (rhu) МАт 18 к IGF-IR оказывало противоопухолевое действие в случае введения в качестве индивидуального агента в дозах 6 и 0,6 мг/кг. В отличие от этого доза 0,06 мг/кг не оказывала влияния на рост опухоли.
Кроме того, с использованием этой же модели тестировали антитело 18 в комбинации с гемцитабином. Опухоли индуцировали, как описано выше, и лечение начинали, когда опухоли развивались и вырастали до объема, составлявшего в среднем 170 мм3 для всех групп. Антитело вводили i.p. один раз в неделю в дозах 6 и 0,6 мг/кг и в комбинации с гемцитабином в дозе 62 мг/кг для дозы антитела 0,6 мг/кг. Осуществляли один цикл введения гемцитабина, а именно, каждый третий день, в общей сложности четыре раза. И в этом случае также лечение начинали путем введения двойных доз антитела. Результаты эксперимента показали, что обработка антителом 18 путем его введения один раз в каждые семь дней само по себе ингибирует рост опухоли и повышает эффективность гемцитабина, известного антиметаболического средства.
Пример 14
Ингибирование роста опухолей линии 3Т3
Опухоли индуцировали у бестимусных мышей в основном как описано в примере 15 за исключением того, что применяли мышиные фибробласты 3Т3, сверхэкспрессирующие человеческий IGF-IR. Мышей с развитыми опухолями массой примерно 180 мг обрабатывали путем внутрибрюшинного введения один раз в неделю, которое осуществляли в общей сложности семь раз, по 18, 6 или 0,6 мг/кг антитела 18. И в этом случае также лечение начинали путем введения в качестве ударных доз двойных доз антитела (36, 12 и 1,2 мг/кг). Результаты эксперимента показали, что лечение антителом может приводить к замедлению роста опухоли при введении доз 18 и 6 мг/кг один раз в неделю.
Пример 15
Индукция опосредованной антителом понижающей регуляции IGF-IR in vitro
Для выявления воздействия антитела, предлагаемого в изобретении, на количество рецептора IGF-I (IGF-IR) в опухолевых клетках проводили эксперименты по оценке зависимости от времени и последующий анализ методом Вестерн-блоттинга с использованием специфических в отношении IGF-IR антител.
В опытах применяли человеческие опухолевые клетки (линия НТ29, 5×104 клеток/мл) в среде RPMI 1640 (фирма РАА, каталожный номер Е15-039), дополненной 2 мМ L-глутамином, 1 × заменимыми аминокислотами (фирма Gibco, каталожный номер 11140-035), 1 мМ пируватом натрия (фирма Gibco, каталожный номер 11360-039) и 10% инактивированной тепловой обработкой FCS (фирма РАА, каталожный номер А15-771). Для каждого периода инкубации осуществляли инокуляцию одного 12-луночного планшета с использованием 1 мл клеток в соответствующей среде для каждого эксперимента и культивировали в течение 24 ч при 37°C и 5% CO2.
Среду осторожно удаляли и заменяли антителом, разведенным в соответствующей среде в различных концентрациях. В двух контрольных лунках среду заменяли либо средой, не содержащей антитело, либо средой, содержащей контрольное антитело (АВ-1, фирма Oncogene, каталожный номер GR11). Клетки инкубировали при 37°C и 5% СO2 и через 15 мин, 24 ч и 48 ч отбирали индивидуальные планшеты для дальнейшего процессинга.
Среду осторожно удаляли путем аспирации и в каждую лунку добавляли по 100 мкл холодного буфера для лизиса (50 мМ Hepes, pH 7,2, 150 мМ NaCl, 1 мМ ЭГТК, 10% глицерина, 1% Triton®-X100, 100 мМ NaF, 10 мМ Na4P2O7, ингибитор протеаз Complete®). Клетки отделяли с помощью скребка для клеток (фирма Corning, каталожный номер 3010) и содержимое лунок переносили в реакционные пробирки Эппендорфа. Фрагменты клеток удаляли путем центрифугирования в течение 10 мин при 13000 об/мин и 4°C и супернатант добавляли к 2× буферу Laemmli для образца в соотношении 1:1 (об./об.). Клеточные белки разделяли с помощью ДСН-ПААГ и переносили на нитроцеллюлозную мембрану (PROTRAN® ВА 85, фирма Schleicher&Schuell, каталожный номер 10 401196) с помощью полусухого Вестерн-блоттинга.
Для определения уровней белка IGF-IR применяли антитело, специфическое в отношении IGF-IR (C-20, фирма Santa Cruz Biotechnologies, каталожный номер sc-713).
Было установлено, что антитело, предлагаемое в изобретении, индуцировало понижающую регуляцию IGF-IR на 50% или более через 24 ч после добавления антитела
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ ЧЕТЫРЕХВАЛЕНТНЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2010 |
|
RU2604189C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2011 |
|
RU2573588C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2010 |
|
RU2573914C2 |
| ДВУХВАЛЕНТНЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2008 |
|
RU2547615C2 |
| АНТИТЕЛА ПРОТИВ CSF-1R ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2617966C2 |
| СВЯЗЫВАЮЩИЕ ПРОТЕИНЫ, СПЕЦИФИЧНЫЕ ПО ОТНОШЕНИЮ К ИНСУЛИН-ПОДОБНЫМ ФАКТОРАМ РОСТА, И ИХ ИСПОЛЬЗОВАНИЕ | 2013 |
|
RU2549703C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ АЛЬФА 1 IL-13 И ИХ ПРИМЕНЕНИЕ | 2006 |
|
RU2413736C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CDCP1 | 2010 |
|
RU2571207C2 |
| ПОЛИСПЕЦИФИЧНЫЕ АНТИТЕЛА, ВКЛЮЧАЮЩИЕ АНТИТЕЛА ПОЛНОЙ ДЛИНЫ И ОДНОЦЕПОЧЕЧНЫЕ ФРАГМЕНТЫ FAB | 2010 |
|
RU2598248C2 |
| ДВУХВАЛЕНТНЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2008 |
|
RU2587616C2 |







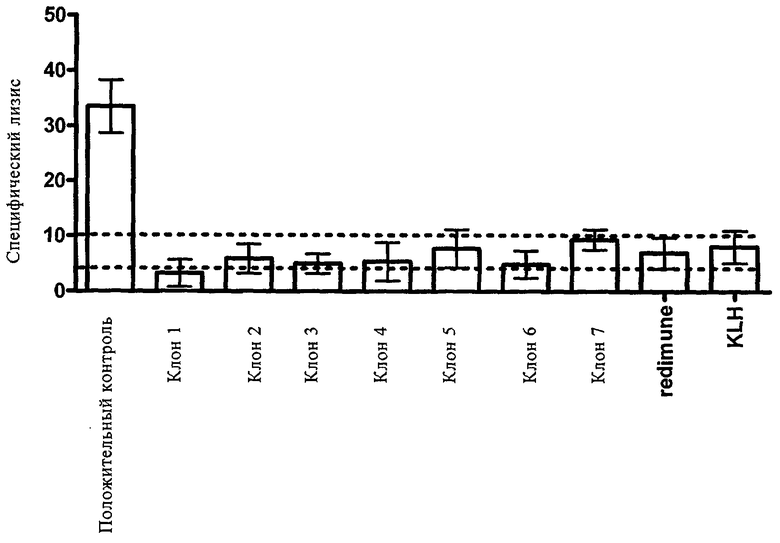
Настоящее изобретение относится к области биотехнологии. Предложена композиция антител, обладающих способностью к связыванию с IGF-IR, которую получают при культивировании клеточной линии CHO DSM ACC 2795, причем количество остатков фукозы в сахарной цепи антител составляет по меньшей мере 99%. Также рассмотрена фармацевтическая композиция для ингибирования роста опухоли и способ получения такой фармацевтической композиция путем смешивания композиции антител по изобретению с фармацевтически приемлемым носителем. Композиция антител обладает пониженной антителозависимой клеточно-опосредованной цитотоксичностью (ADCC) и может найти применение в терапии заболевания, ассоциированных с IGF-IR. 3 н. и 2 з.п. ф-лы, 1 ил., 4 табл., 15 пр.
1. Композиция антител для получения фармацевтической композиции для ингибирования роста опухоли, обладающих способностью к связыванию с IGF-IR, где композиция антител получается при культивировании клеточной линии CHO DSM ACC 2795 и отличается тем, что количество остатков фукозы в сахарной цепи антител составляет по меньшей мере 99% («полностью фукозилированное»), и включает гипервариабельные участки (CDR), имеющие следующие последовательности:
а) тяжелую цепь антитела, содержащую в качестве CDRs CDR1 (ак 31-35), CDR2 (ак 50-66) и CDR (ак 99-107) последовательности, представленной в SEQ ID NO:1;
б) легкую цепь антитела, содержащую в качестве CDRs CDR1 (ак 24-34), CDR2 (ак 50-56) и CDR3 (ак 89-98) последовательности, представленной в SEQ ID NO:2.
2. Композиция антител по п.1, отличающаяся тем, что антитела обладают одним или несколькими свойствами, выбранными из группы, включающей:
а) соотношение значений IC50, характеризующих ингибирование связывания IGF-I с IGF-IR и ингибирование связывания IGF-II с IGF-IIR, составляет от 1:3 до 3:1;
б) в концентрации 5нМ ингибирует по меньшей мере на 80%, предпочтительно по меньшей мере на 90%, фосфорилирование IGF-IR по данным клеточного анализа фосфорилирования с использованием клеток линии НТ29 в среде, содержащей 0,5% инактивированной тепловой обработкой фетальной телячьей сыворотки (FCS), по сравнению с фосфорилированием по данным анализа без антитела;
в) не обладает IGF-IR-стимулирующей активностью (отсутствие передачи сигнала, отсутствие IGF-I-подобной активности), измеряемой по фосфорилированию РКВ, при использовании в концентрации 10мкМ по данным клеточного анализа фосфорилирования с использованием клеток линии 3Т3, имеющих 400000-600000 молекул IGF-IR на клетку в среде, содержащей 0,5% инактивированной тепловой обработкой фетальной телячьей
сыворотки (FCS), по сравнению с фосфорилированием по данным анализа без антитела.
3. Композиция антител по п.1, отличающаяся тем, что аффинность связываний с IGF-IR составляет примерно от 10-13 до 10-9М (KD).
4. Фармацевтическая композиция для ингибирования роста опухоли, содержащая композицию антител по любому из пп.1-3 в фармацевтически эффективном количестве и фармацевтически приемлемые носители.
5. Способ получения фармацевтической композиции, предусматривающий смешивание композиции антител по любому из пп.1-2 в фармацевтически эффективном количестве с фармацевтически приемлемыми носителями.
| SHIELDS R.L | |||
| et al., "Lack of Fucose on Human IgG1 N-Linked Oligosaccharide Improves Binding to Human Fc[gamma]RIII and Antibody-dependent Cellular Toxicity", The Journal of Biological Chemistry (2002), 277(30):26733-26740 | |||
| DAVIES J | |||
| et al., "Expression of GnTIII in a recombinant anti-CD20 CHO production cell line: Expression of antibodies with |

