Изобретение относится к способу трансформации, регенерации и селекции растений семейства бобовых рода Cyamopsis, в частности трансформации гуара (Cyamopsis Tetraqonoloba), опосредованной Agrobacterium, к генетически модифицированным растениям, полученным данным способом, также как и к применению веществ, таких как сулбактам, ингибитора β-лактамазы, способствующего трансформации гуара и других растений.
Известный уровень техники
Трансформация бобовых
Семейство Fabaceae (Lequminosae) является наиболее важным семейством двудольных растений в мире. Из-за его громадного экономического значения предпринимались многочисленные усилия для улучшения его сельскохозяйственных показателей с помощью генной инженерии.
Трансформация, опосредованная Agrobacterium, является обычно используемым способом для переноса генов в растения. Виды растений, которые к настоящему времени подверглись успешной трансформации с помощью Agrobacterium, представлены исключительно двудольными (в противоположность однодольным), однако не все двудольные растения легко трансформировать.
Одни семейства растений, например Solanaceae, особенно хорошо приспособлены для генного переноса, опосредованного Agrobacterium, тогда как другие, такие как Fabaceae, поддаются этому переносу с трудом.
Трансгенные растения сои (Glycine max) пытались получить разными путями. В качестве источников эксплантантов использовали листья и протопласты, однако при данном способе в трансформированных растениях регенерацию не получали. Семядоли сои, инокулированные Agrobacterium tumefaciens, давали трансгенные растения, но в этом случае только один из множества проверенных генотипов оказался успешно трансформированным (Hinchee et al., Bio/Technology 6: 915, 1988). WO 94/02620 описывает способ получения трансгенных растений сои, используя гипокотили или семядольные узлы и ряд операций, специально предназначенных для трансформации сои, в том числе особые температурные условия, pH и концентрации Aqrobacterium.
Однако использовать семядоли в качестве эксплантантов для трансформации бобовых, в целом, не представляется возможным и в большинстве случаев используют другие источники эксплантантов. Например, для трансформации гороха (Pisum sativum) в качестве эксплантантов использовали эксплантанты из проростковых культур и эпикотили сеянцев и, полученный таким образом трансгенный каллус, спустя 6 месяцев регенерировали в растения (Puonti-Kaerlas et al. Plant Cell Rep. 8:321, 1989).
Для трансформации клевера ползучего (Trifolium repens) кончики его проростков инокулировали почвенной бактерией и получали трансгенные растения (Voisey et al. Plant Cell Rep. 13:309, 1994).
Предпринимали попытки трансформировать ряд других бобовых. Например, семядольные узлы и гипокотили Phaseolus vulgaris, инкубированные с Agrobacterium tumefaciens, превращали в трансгенный каллус, но не в трансгенные растения (Mc-Clean et al. Plant Cell, Tissue & Orq. Cult. 24:131, 1991). Такие же результаты получили и для представителей рода Vigna (Garcia et al. Plant Sciens 48-49, 1986). Несмотря на предпринятые значительные усилия, не было сообщений о получении трансгенных растений земляного ореха (Arachis hypogaea).
Авторы настоящего изобретения предприняли неудачную попытку трансформировать гуар, используя методику инокуляции семядолей сои, описанную Hinchee et al. Наряду с результатами, полученными другими исследователями, это свидетельствует, что выбор способа трансформации бобовых является полностью эмпирическим и что в данном случае невозможно установить общие научно обоснованные принципы. Поэтому методика трансформации и источник получения эксплантанта для трансформации бобового растения, которую разработали в соответствии со специфическими особенностями рода, вида и даже генотипа, ненадежны.
Сообщения о многочисленных попытках получить трансгенные растения различных бобовых ясно показывают, что осуществить трансформацию бобовых очень трудно, даже ученым, специалистам в данной области. Дополнительным подтверждением этому является тот факт, что из приблизительно 100 видов бобовых, представляющих коммерческий интерес, трансформированы менее 5 видов. Поэтому успешная трансформация прежде не трансформированных родов или видов бобовых является нестандартным достижением.
Гуар
Гуар (Cyamopsis tetraqonoloba) является бобовым растением, представляющим значительный коммерческий интерес из-за высокого содержания в его семенах галактоманнана. Гуаровый галактоманнан известен также как гуаровая камедь и используется для пищевых и непищевых целей в качестве вещества, повышающего вязкость.
Галактоманнан найден в эндосперме, который составляет около 35% сухого веса семян или 80-90% чистого галактоманнана. Большие эндоспермы являются необычным признаком у представителей Fabaceae, в семенах которых эндосперм или отсутствует, или находится в рудиментарном состоянии; запасные питательные вещества для прорастания у бобовых чаще всего отложены в увеличенных семядолях.
Сообщений о генетической трансформации какого-либо вида бобовых, с большим содержанием галактоманнана в эндосперме, не было.
Сулбактам
Недостаток, присущий генному переносу, опосредованному Agrobacterium, заключается в том, что бактерии продолжают расти после трансформации. Для предотвращения чрезмерного роста растительного материала необходимо удалять бактерии, обычно с помощью добавления антибиотика пенициллинового ряда (β-лактамы), такие как карбенициллин, цефотаксим и др.
Вещества пенициллинового ряда выбраны по той причине, что они в принципе не токсичны для растительных тканей. Однако на практике эти соединения часто оказывают на эксплантанты значительный токсический эффект. Одна из возможных причин фитотоксичности, кроме возможного прямого токсического действия, состоит в том, что антибиотики подвержены постепенному разрушению во время длительной инкубации в присутствии бактерий и растительных тканей. Примером нежелательной деградации продукта является обычно используемый антибиотик карбенициллин, который может разрушаться до фенилуксусной кислоты. Фенилуксусная кислота обладает ауксинподобными свойствами и поэтому улучшает рост каллюса в эксплантанте, что в свою очередь снижает регенерацию.
Поэтому было бы очень полезно научиться использовать небольшие количества антибиотиков и/или антибиотиков, которые не обладают нежелательными побочными эффектами. Во время своей работы по трансформации гуара авторы настоящего изобретения обнаружили, что β-лактомазный ингибитор сулбактам резко понижает концентрации веществ пенициллинового ряда, в результате чего повышается эффективность трансформации и значительно снижаются расходы на ее осуществление.
Этот новый подход к контролю роста почвенной бактерии в целом пригоден для трансформации растений, поскольку он относится к бактериям, которые необходимо всегда удалять во время трансформации и поэтому не ограничен каким-либо отдельным растительным видом.
Краткое описание изобретения
В соответствии с поставленной задачей настоящее изобретение относится к генетически модифицированному растению, или его части, из рода Cyamopsis, отличающееся тем, что растение или часть растения содержит в своем геноме не менее одной последовательности рекомбинантной ДНК.
В соответствии с другой целью настоящее изобретение относится к способу получения генетически модифицированного растения, или его части, из рода Cyamopsis, включающего в себя операции встраивания последовательности рекомбинантной ДНК, по крайней мере, в одну клетку или протопласт и образования генетически модифицированных эксплантантов, используя, как минимум, одну селективную или ростовую среду для проростков, включающую в себя хотя бы одно соединение, выбранное из ингибитора ауксина, ингибитора лактамазы и ингибитора этилена, так чтобы получить генетически модифицированное растение, или часть растения, содержащее в своем геноме не менее одной последовательности рекомбинантной ДНК.
Другим аспектом настоящего изобретения является способ получения генетически модифицированного растения, в котором используется, как минимум, одна среда для селекции или роста клеток, протопластов, каллюса или частей растения, включающая в себя не менее одного вещества, которое ингибирует рост бактерий или которое повышает действие ингибитора бактериального роста без какой-либо существенной регуляции роста растения или токсического воздействия на растение.
В соответствии с еще одной целью настоящее изобретение относится к химерным растениям, наделенным способностью производить трансгенные семена, а полученный путем прививки генетически модифицированный проросток выращивали ин витро в не ин витро выращенном растении.
Подробное описание изобретения
Термины "генетически модифицированное растение" и "трансгенное растение" в контексте настоящей заявки относятся к общепринятому пониманию значений этих терминов в технике, т.е. растения, которые были изменены таким образом, что их геном включает в себя не менее одной последовательности рекомбинантной ДНК. "Последовательностью рекомбинантной ДНК" обычно называют ту последовательность, которая может быть экспрессирована или которая влияет на экспрессию гена в растении, а также последовательность, которая может служить в качестве маркера, необязательно, которая экспрессирована или влияющая на экспрессию гена. Последовательности, которые экспрессированы или которые влияют на экспрессию гена, часто оказываются чужеродными генами для растения, о котором идет речь, в его природной форме, но могут также представлены, например, слегка измененной формой природного гена или, к примеру, промоторной или регуляторной последовательностью, которая изменяет экспрессию природного гена. Способ настоящего изобретения по получению генетически модифицированных растений следует отнести к генетической трансформации в целом, и не ограничивается включением каких-либо отдельных категорий последовательностей ДНК.
Термин "части растения" относится, в общем, к любой части растения, например ткани или органу, которые не представляют целое растение, в том числе недифференцированный каллюс, также как и дифференцированные части растения, такие как проростки, листья, корни, плоды, семена и т.д.
Как указано выше, настоящее изобретение относится, в частности, к генетически модифицированным растениям рода Cyamopsis и, прежде всего, к растениям вида C.tetragonoloba (гуар). Генетически модифицированные растения рода Cyamopsis можно получить по вышеупомянутому способу, в котором первой операцией является интродукция последовательности рекомбинантной ДНК хотя бы в одну клетку или протопласт. Интродукция рекомбинантной ДНК может быть достигнута с помощью способа, обычно используемого для получения геноинженерных растений, включая перенос, опосредованный Agrobacterium, например, посредством Ti-плазмиды A.tumefaciens, или Ri-плазмиды A. rhizogenes в качестве вектора, также как и в результате, например, микроинъекции, электропорации или бомбардировкой частиц. Предпочтительным методом (описанный ниже в примерах) является генный перенос, опосредованный Agrobacterium. Как объяснено ниже, хорошие результаты были получены путем трансформации семядолей растения Agrobacterium tumefaciens, даже если использованные семядоли происходили из семян, которые прорастали на протяжении относительного долгого периода времени, такого как 11-12 дней.
После интродукции желаемой рекомбинантной ДНК в избранный растительный материал (например, ткань, клетки или протопласты) получали генетически модифицированные эксплантанты, используя, как минимум, одну элективную или питательную среду для выращивания проростков, содержащую по меньшей мере, одно соединение, выбранное из ингибитора ауксина, ингибитора β-лактамазы и ингибитора этилена, поскольку было обнаружено, что присутствие одного или больше из этих соединений в селективной и/или питательной среде приводит к уменьшению каллюса и увеличению частоты регенерированных и трансформированных проростков. Предпочтительно, чтобы селективная среда включала в себя, как минимум, ингибитор ауксина и ингибитор β-лактамазы, а питательная среда хотя бы ингибитор β-лактамазы. Более предпочтительно, чтобы селективная среда включала в себя ингибитор ауксина, ингибитор β-лактамазы и ингибитор этилена, а питательная среда включала в себя ингибитор ауксина и ингибитор этилена.
Ингибиторы (ингибитор ауксина, ингибитор β-лактамазы и ингибитор этилена) могут действовать, удаляя или уменьшая количество соответствующих соединений (т.е. в результате ингибирования биосинтеза соединений или деградации соединений) или в результате подавления деятельности этих соединений. Не замыкаясь в рамках какой-либо одной теории, очевидно, что действие ингибиторов, по крайней мере частично, связано с антиауксиновым действие или с ингибированием "ауксин-подобных" эффектов, поскольку присутствие ауксина ведет к повышению роста каллюса и, следовательно, к снижению частоты регенерации проростков и выделению трансформированных проростков. Это справедливо для тех соединений, которые непосредственно функционируют как ингибиторы ауксина. Что касается ингибитора β-лактамазы, то он описывается в примере, в котором одним из предполагаемых действий сулбактама является элиминация продукта деградации - фенилуксусной кислоты (из антибиотика карбенициллина), которая обладает нежелательным ауксин-подобным свойством, приводящее к повышению роста каллюса. Также полагают, что ингибитор этилена, кроме его прямого действия на этилен, который, по-видимому, служит для предотвращения преждевременного физиологического старения в развивающихся проростках, обладает также полезным действием, поскольку этилен, как известно, влияет на восприимчивость к ауксину. Использование ингибитора ауксина и ингибитора этилена приемлемо для любого типа использованного переноса гена, например переноса опосредованного бактериями, например Agrobacterium, наряду с другими способами, такими как микроинъекция, электропорация и бомбардировка частицами, тогда как использование ингибитора β-лактамазы особенно подходит для методики, использующей перенос гена посредством Agrobacterium или другие штаммы бактерий, продуцирующие β-лактамазу.
Предпочтительным ингибитором ауксина является 2-(п-хлорофенокси)-2-метилпропионовая кислота (PCIB), которую можно использовать в элективной среде и факультативно также в питательной среде для выращивания проростков, в концентрациях около 0,01-10 мг/л, обычно около 0,05-5 мг/л, например около 0,1-2 мг/л. Другими ингибиторами ауксина, которые можно использовать, являются например, 2,3,5-трииодбензойная кислота (TIBA), N-нафтилф таламиновая кислота и 7-хлороиндолуксусная кислота.
Предпочтительным ингибитором β-лактамазы является сулбактам (продает Pfizer под коммерческим наименованием Betamaze). Сулбактам можно использовать в элективной среде или питательной среде для выращивания проростков в концентрации около 10-1000 мг/л, обычно около 20-500 мг/л, например около 50-200 мг/л.
Предпочтительным ингибитором этилена является тиосульфат серебра, который обычно используют в селективной или питательной среде для прорастания в концентрации вплоть до 50 мкМ, обычно около 0,1-10 мкМ, например около 0,5-5 мкМ. Другими ингибиторами этилена, которые можно использовать, являются, например, аминоэтоксивинилглицин (AVG), кобальт и норборнеол.
Кроме соединений, описанных выше, обнаружено также, что некоторые другие соединения обладают полезным действием при их использовании в элективной и/или питательной среде для выращивания проростков. Например, обнаружено, что при добавлении к селективной среде соли никеля частота трансформации увеличивается. Поэтому элективная среда предпочтительно содержит соль никеля, например NiCb2 • 6H2O, например, в концентрации около 0,1-10 мг/л, например 0,5-5 мг/л. Обнаружено, что 6-бензиламинопурин (ВАР) также обладает полезным действием. Поэтому элективная среда содержит ВАР, например, в концентрации около 0,1-10 мг/л, предпочтительно 1-5 мг/л, для питательной среды выращивания проростков приемлемы те же концентрации.
Наличие канамицина, например, в виде сульфата канамицина, в концентрации около 50-300 мг/л, обычно около 100-200 мг/л, например около 130-160 мг/л, в элективной среде может также оказывать положительное действие, когда инсерционная последовательность ДНК включает ген резистентности к канамицину. Было также установлено, что хорошие результаты получают в том случае, когда эксплантанты, обычно те из которых получали проростки, переносили во вторую элективную среду с пониженной концентрацией канамицина по сравнению с первой элективной средой предпочтительно не более чем 125 мг/л, обычно 20-100 мг/л, предпочтительно 30-70 мг/л. Сходным образом и другие аминогликозидные антибиотики, такие как гигромицин, неомицин, стрептомицин и гентамицин можно применять в элективной среде вместе с инсерционной последовательностью ДНК, включающей в себя нужный ген устойчивости к антибиотику. Можно также использовать и другие селективные агенты, например гербициды или позитивные селективные агенты, такие как манноза или ксилоза.
После селекции и сбора регенерированных проростков наличие генетически трансформированных проростков можно определить различными способами. Один из них (кроме использования антибиотика вместе с инсерционным геном резистентности к антибиотику, как описано выше) осуществляют с помощью репортерного гена β-глюкуронидазы, а его использование описано ниже и в WO 93/05163.
Получаемые трансгенные проростки Cyamopsis можно затем регенерировать в целые растения известными способами, т. е. либо в результате образования корней непосредственно у проростков или с помощью прививки трансгенных проростков на устоявшиеся укорененные растения. Последний способ, прививка трансгенных проростков на стебли устоявшихся растений (которые сами по себе могут быть или не быть трансгенными), оказался наиболее приемлемым, давая химерные растения, способные продуцировать трансгенные семена. Кроме того, обнаружили, что лучшие результаты получали, когда трансгенные проростки прививали например, 7-28-дневную рассаду, обычно на 12-21-дневную.
Как указано выше, другой целью настоящего изобретения является способ получения генетически модифицированного растения, для которого используют, как минимум, одну среду для селекции или роста клеток, протопластов, каллюса или частей растения, включающую в себя, как минимум, одно вещество, которое ингибирует рост бактерий или которое увеличивает эффект ингибитора бактериального роста без какого-либо существенного регулирования роста растения или токсического действия, например ингибитора β-лактамазы. Например, ингибитор β-лактамазы, такой как сулбактам, способствует не только трансформации и селекции растений гуара, он также влияет на перенос гена, опосредованного Agrobacterium (или в присутствии других бактериальных штаммов, продуцирующих β-лактамазу), в любое растение с целью исключения нежелательного роста бактерий после трансформации. Значительная практическая и экономическая выгода этого подхода заключается в том, что количество антибиотиков пенициллинового ряда, используемых в селективной и питательной среде для прорастания можно существенно снизить, например до уровня около 10% от того, которое необходимо в отсутствие ингибитора β-лактамазы. Когда ингибитором β-лактамазы является сулбактам, его используют в вышеуказанном количестве.
Далее изобретение иллюстрируют нижеприведенные примеры, не ограничивающие его объем.
Примеры
Общая методика для трансформации гуара
Рассада
Семена гуара стерилизовали в растворе хлорита натрия, содержащего 2,5% свободного хлора, pH 7,0, и две капли Твина-80 на 100 мл раствора. Семена (около 10 г на 100 мл) перемешивали в течение 25 мин, промывали 5 раз стерильной водой и высушивали на фильтровальной бумаге в течение ночи.
Семена высевали на среду прорастания и помещали в темноту при 25oC на 4 дня. Затем проращивание продолжали в течение 7 дней в режиме 12 ч/12 ч день/ночь. Использование обогащенной среды для прорастания приводило к получению гуаровой рассады высокого качества.
Среда для прорастания:
4,43 г/л MSMO (Siqma М6899)
20 г/л сахарозы
8,0 г/л агара
pH 5,8 (доводили с помощью КОН)
Суспензия Agrobacterium tumefaciens
Инкубирование суспензии Agrobacterium осуществляли в течение 17-18 ч в LB-среде. Ацетосирингон не добавляли к бактериальной культуре.
LB-среда:
10 г/л бактотриптона
10 г/л NaCl
5,0 г/л дрожжевого экстракта
pH 7,4 (доводили с помощью NaOH)
Трансформация и сокультивирование
Суспензию бактерий разбавляли в большинстве случаев до OD (оптическая плотность) 0,1 (660 нм) LB-средой, но хорошие результаты получали также при OD около 1.
Семядоли с приблизительно 2 мм подсемядольным коленом вырезали из 12-дневных стеблей. Затем семядоли отрывали, используя щипчики для создания раневой поверхности, и помещали в суспензию Agrobacterium на 30 мин.
Предпочтительной методикой для проведения сокультивирования был так называемый метод сэндвича, где эксплантанты помещали на фильтровальную бумагу, которую поочередно помещали на среду сокультивирования.
Фильтровальную бумагу, насыщенную в жидкой среде сокультивирования, также помещали сверху на эксплантанты с целью предотвращения высыхания эксплантантов.
Среда сокультивирования:
0,43 г/л смеси основных солей MS (Siqma М5524)
20 г/л сахарозы
100 мг/л миоинозитола
0,1 мг/л тиамина • HCl
0,5 мг/л пиридоксина • HCl
0,5 мг/л никотиновой кислоты
1,0 мкМ тиосульфата серебра
8,0 г/л агара
pH 5,1
Сокультивирование проводили в течение 3 дней при 25oC ив режиме 12 ч/12 ч день/ночь. Затем сокультуру эксплантантов отмывали 1/10 MS-среды, к которой добавляли 100 мг/л карбенициллина, 100 мг/л цефотаксима и 1000 мг/л лизозима, перемешивая 2-3 раза по 45 мин при 100 об/мин.
Селекция
Эксплантанты переносили на селективную среду и инкубировали, как указано выше (25oC, 12 ч днем/12 ч ночью).
Селективная среда
3,2 г/л Гамборг B5 (Siqma G5893)
20 г/л сахарозы
1,0 мг/л бензиламинопурина
0,05 мг/л гибберелловой кислоты (GA3)
1,0 мкМ тиосульфата серебра
1,0 мг/л NiCl2 • 6H2O
0,5 мг/л 2-(п-хлорофенокси)-2-метилпропионовой кислоты (PCIB)
50 мг/л цефотаксима
50 мг/л карбенициллина
100 мг/л сулбактама (Betamaze)
145 мг/л сульфата канамицина
pH 5,7
Сбор трансгенных проростков
Первый сбор
Спустя 4 недели собирали проростки, выросшие более 3 мм, и переносили их в среду для проростков. Через 10-14 дней у проростков проверяли активность репортерного гена β-глюкуронидазы (GUS), смотри ниже. GUS-позитивные проростки переносили в свежую среду для проростков, а GUS -негативные проростки отбрасывали.
Среда для проростков:
3,2 г/л Гамборга B5 (Siqma G 5893)
20 г/л сахарозы
0,1 мг/л бензиламинопурина
1,0 мкМ тиосульфата серебра
0,1 мг/л гибберелловой кислоты (GA3)
100 мг/л цефотаксима
100 мг/л сулбактама (Betamaze)
8,0 г/л агара
pH 5,7
Второй сбор
После первого сбора эксплантанты (от которых собрали урожай проростков) переносили во вторую селективную среду с пониженной концентрацией канамицина (50 мг/л). После следующих 4 недель проростки собирали и подвергали GUS-тестированию. Позитивные проростки переносили в свежую среду, а негативные отбрасывали.
Анализ трансгенных проростков
Верхушки молодых листьев срезали и переносили в лунки планшета, содержащих 200 мкл Х-глюкуронатного раствора. После инкубирования в течение 16 ч при 35oC верхушки листьев обесцвечивали в 96% этаноле, а уровень голубой окраски определяли под микроскопом.
Х-глюкуронатный раствор (50 мл):
0,2 М Na2HPO4 - 15,5 мл
0,2 М NaH2PO4 - 9,5 мл
H2O - 19,5 мл
0,1 М K3(Fe(CN)6) - 0,25 мл
0,1 М K4(Fe(CN)63H2O - 0,25 мл
0,1 М Na2 ЭДТА - 5,0 мл
Х-глук (циклогексиламмоний 5-бром-4-хлор-3-индолил)-β-D-глюкуронат - 50 мг
Прививка
Укоренение трансгенных проростков осуществляли прививкой. Проростки, образующие здоровую зелень и длиной 0,5-1,0 см, отбирали для прививки на 1,5-2-месячные растения гуара, выросшие в режиме 14 ч день/10 ч ночь при 32oC/25oC. Перед прививкой все листья, за исключением двух самых верхних листьев, удаляли, а трансгенные проростки прививали непосредственно в вертикальные разрезы на узлах стебля. Привитые растения помещали во влажную камеру на 5-6 дней.
Трансгенные растения
Привитые растения последовательно переносили в ростовую камеру с целью дальнейшего роста трансгенных проростков. Условия для роста были, как указано выше. Спустя примерно 2 месяца собирали зрелые стручки с множеством трансгенных семян.
Пример 1
Трансформация различных сортов гуара
Этот пример демонстрирует трансформацию у нескольких сортов гуара с использованием вышеописанного способа.
Сорт гуара - N GUS + проростков
Льюис - 8
Санта Круз - 3
Индийский - 6
Число GUS+(GUS-позитивных, т.е. трансформированных) проростков подсчитывали среди 1000 трансформированных эксплантантов, используя нопалиновый штамм Aqrobacterium С58. Американский сорт Льюис давал наибольшую численность трансгенных проростков, имеющих GUS-ген как маркер успешной трансформации, и этот сорт был одним из использованных в последующих примерах.
Пример 2
Агробактерии
Проверяли способность нескольких разных безжгутиковых штаммов Aqrobacterium tumefaciens трансформировать гуар, и все они оказались пригодны для этой цели. К примеру 500 гуаровых эксплантантов, подвергнутых воздействию одним бактериальным штаммом, испытывали с каждым из четырех различных штаммов (октопиновый штамм LBA 4404, Ditta et al., Proc. Nat. Acad. Sci. 77: 7347, 1980, и три штамма, полученные из С58). Для каждого штамма получили 11-26 регенерированных проростков, из которых 1-3 были GUS-позитивными проростками. Сходным образом другой штамм Agrobacterium (L,L-сукцинаминопиновый штамм ЕНА 101), который использовали для обработки 2500 эксплантантов, привел к получению 25 регенерированных проростков, из которых 8 были GUS-позитивными. Штамм LBA 4404 также использовали для обработки 2000 эксплантантов, давших 67 регенерированных проростков, из которых 17 были GUS-позитивными.
Использованные штаммы A. tumefaciens содержали в области Т-ДНК гены, кодирующие β-глюкуронидазу (для GUS-анализа) и фосфотрансферазу неомицина (для селекции на среде, содержащей канамицин).
Пример 3
Отбор по канамицину
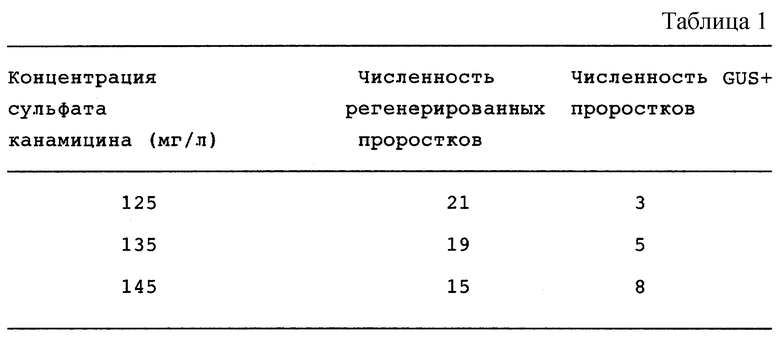
В этом примере оптимальная концентрация сульфата канамицина в элективной среде составляла 145 мг/л, однако эффективную трансформацию осуществили, используя другие концентрации, в пределах 125-145 мг/л. Применение концентрации канамицина 100 мг/л или менее того давало частоту регенерации, близкую к 100%, хотя получили всего только несколько трансгенных проростков (см. табл. 1).
Всего, при каждой обработке штаммом Agrobacterium ЕНА 101, трансформировали 1600 эксплантантов. Около 50% трансформированных проростков обнаружены при втором сборе урожая, где концентрация канамицина была снижена. Когда концентрацию канамицина поддерживали на уровне 125-145 мг/л, при втором сборе урожая найдены только несколько трансгенных проростков.
Пример 4
BAP и NiCl2
Этот пример показывает преимущество добавления 6-бензил-аминопурина (BAP) и NiCl2 к элективной среде (см. табл. 2).
При каждой обработке всего трансформировали 1200 эксплантантов, используя Aqrobacterium штамм ЕНА 101. Добавление ВАР в количестве 5 мг/л увеличивало количество трансформантов, а также общую численность регенерированных проростков примерно вдвое при концентрации ВАР 5 мг/л по сравнению с 1 мг/л.
Добавление 1 мг/л NiCl2 • 6H2O, который обычно отсутствует в используемых средах MS и Гамборг В5, повышало частоту трансформации в 2-4 раза.
Пример 5
Тиосульфат серебра
Этот пример показывает, что тиосульфат серебра (STS) существенно повышает частоту трансформации.
Концентрация STS (мкМ) - Численность GUS+проростков
0 - 4
2,5 - 12
5,0 - 8
10,0 - 7
Количество GUS + проростков подсчитывали на 1000 эксплантатов, трансформированных при использовании штамма ЕНА 101. При концентрации 2,5 мкМ тиосульфата серебра получали значительно больше трансформантов, чем при концентрациях 0; 5,0 или 10,0 мкМ тиосульфата серебра.
Повышение частоты трансформации, вызванное STS, связано со значительным улучшением качества проростков. В отсутствие STS трансгенные проростки останавливались в росте и желтели, тогда как присутствие STS поддерживает рост. Поскольку ионы серебра, как известно, подавляют действие этилена, благоприятные эффекты STS могут быть связаны со снижением влияния этилена в сосудах, предотвращая преждевременное физиологическое старение.
Пример 6
PCIB
PCIB (2-(п-хлорфенокси)-2-метилпропионовая кислота) обладает антиауксиновым действием и может подавлять образование каллюса. В отсутствие PCIB во время селекции происходило интенсивное образование каллюса и одновременно ухудшение регенерации.
Добавление в среду 0,1-2 мг/л PCIB существенно снижало количество каллюса, усиливало регенерацию и увеличивало численность трансгенных проростков.
Пример 7
Сулбактам
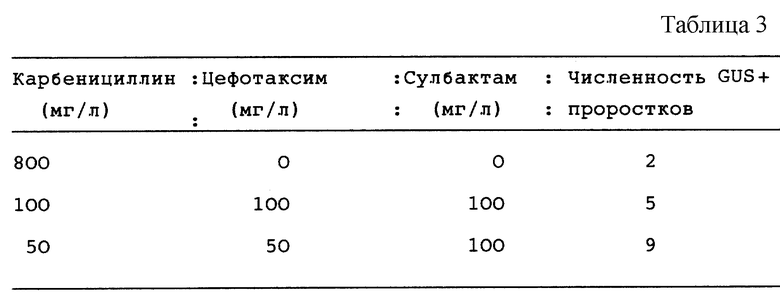
Этот пример показывает, что сулбактам, ингибитор β-лактамазы значительно уменьшает количества карбенициллина и цефотаксима, необходимые для селекции и повышения частоты трансформации (см. табл. 3).
При каждой обработке трансформировали всего 800 эксплантатов, используя штамм ЕНА 101. Все 3 обработки осуществляли при удалении Agrobacteria.
Трансгенные проростки в присутствии карбенициллина, 800 мг/л среды, росли плохо и имели желтоватый оттенок. Эти проростки не восстанавливались после переноса в среду для проростков и многие трансгенные проростки со временем погибали.
Трансгенные проростки, подвергнутые селекции с карбенициллином, 50 или 100 мг/л среды, в присутствии сулбактама, 100 мг/л среды, имели нормальный зеленый вид и могли бы поддерживаться ин витро длительное время.
Пример 8
Тидиазурон
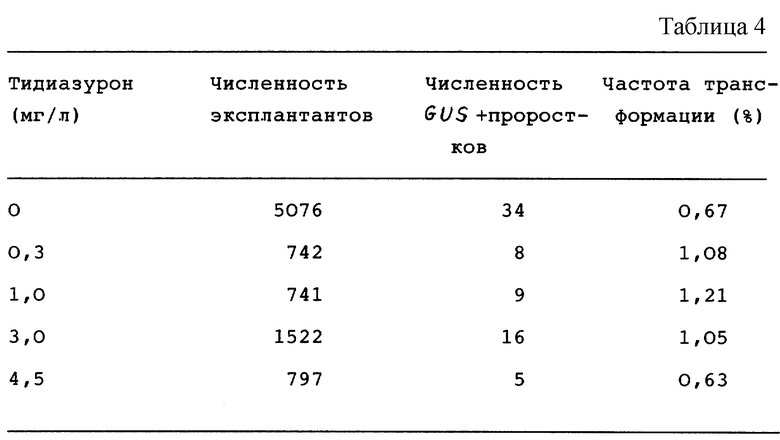
Добавление цитокинина тидиазурона (TDZ) значительно повышало частоту трансформации. Как показано в нижеприведенной таблице, оптимальная концентрация TDZ в элективной среде, которая повышала частоту трансформации в 1,5-1,8 раз, составляла ряд 0,3-3,0 мг/л (см. табл. 4).
Кроме повышения частоты трансформации, тидиазурон также благоприятно влиял на последующее клонирование трансгенных проростков.
Пример 9
Прививка
Укоренение трансгенных проростков осуществляли путем прививки. Хотя прививка на выращенных растениях (1,5-2-месячные) возможна, как показано выше, однако степень успеха повышается после прививки в сеянцы, имеющие возраст 12-21 день.
Рассаду получали, помещая стерилизованные семена гуара в среду прорастания (смотри выше) и затем культивируя их в течение 12-21 дня при 25oC в режиме 13 ч днем/11 ч ночью. Семядоли вырезали и на подсемядольном колене делали вертикальные 0,5-1,0 см надрезы по направлению вниз. Трансгенный проросток помещали в разрез и завязывали коротким куском стерильного волокна. Спустя 5-10 дней волокно удаляли и привитое растеньице переносили в грунт.
Из 49 трансгенных проростков, привитых в такие сеянцы, 42 выжили (89%) и дали плодоносящие растения с нормальным фенотипом.
Пример 10
Саузерн-блот анализ
С целью подтвердить наличие трансгенов и множество генных копий в линиях трансгенного гуара, геномную ДНК экстрагировали из образцов листьев, расщепляли рестриктазой HindIII и подвергали электрофорезу в 0,8% агарозном геле. Саузерн-блоты делали с Hybond N+ (Amersham), а прегибридизацию и гибридизацию проводили при 68oC в буфере, рекомендованном изготовителем. Зонды ДНК представляли собой либо ген PMI, либо ген GUS, меченые путем случайного праймирования, используя набор "Ready to qo" (Pharmacia). 1 • 106 CPM/мл меченого зонда добавляли в гибридизационный буфер.
Саузерн-блот-анализ с использованием гена PMI, в качестве зонда, представлен на фигуре 1. Можно видеть, что каждая колонка, за исключением колонки 1, содержит интенсивную полосу, соответствующую 2,1 т.п.н., которая представляет собой ожидаемый размер фрагмента ДНК, полученный в результате расщепления ДНК с помощью HindIII. Колонка 1 содержит слабую полосу сходного размера. Колонка 8 представляет нетрансгенную линию гуара.
Саузерн-блот-анализ, использующий ген GUS в качестве зонда, представлен на фигуре 2. Можно видеть, что большинство колонок содержат одну интенсивную полосу, указывая тем самым, что эти линии содержат одну копию гена GUS. Расщепленная ДНК, представленная в колонке 7, содержит 4 полосы, означая тем самым, что эта линия содержит 4 копии гена GUS. Колонка 8 представлена нетрансгенной линией гуара.
Фигура 1 показывает результаты анализа по Саузерну трансгенных линий гуара. Образцы из 10 мкг геномной ДНК, полученные от разных трансгенных линий гуара и расщепленные рестриктазой HindIII, подвергали электрофорезу в 0,8% агарозном геле и ДНК блоттировали в Hybond N+ и гибридизовали с зондом, содержащим ген PMI, меченный [32P]dCTP. Маркеры [35S]ДНК (Amersham) использовали в качестве маркера молекулярного веса (МW).
Фигура 2 показывает результаты анализа по Саузерну трансгенных линий гуара. Образцы из 10 мкг геномной ДНК, полученные от разных трансгенных линий гуара, расщепленные рестриктазой HindIII, подвергали электрофорезу в 0,8% агарозном геле и ДНК блоттировали в Hybond N+ и гибридизовали с зондом, содержащим ген GUS, меченный [32P]dCTP. Маркеры [35S] ДНК (Amersham) использовали в качестве маркера молекулярного веса (MW).
Пример 11
Анализ трансгенного потомства
Наследственность и сегрегацию трансгенов анализировали в нескольких не связанных между собой трансформантах гуара. Первичные трансформанты являлись автофертильными. Посев семян осуществляли в количестве 10-20 штук. Наличие гена GUS во втором поколении растений устанавливали с помощью GUS-анализа (смотри выше) (см. табл. 5).
Табл. 5 показывает, что ген GUS устойчиво наследуется и что он расщепляется приблизительно так, как этого бы следовало ожидать для простого доминантного гена. Более того, ген GUS сохраняет свою активность.



Изобретение относится к способу трансформации и регенерации растений из семейства бобовых рода Cyamopsis, в частности трансформации гуара (Cyamopsis tetragonoloba). В результате интродуцирования последовательности рекомбинантной ДНК хотя бы в одну клетку или протопласт получают генетически модифицированные эксплантанты. Для селекции модифицированных клеток и выращивания проростков используют селективные и питательные среды. Применение среды, включающей, по крайней мере, одно соединение, выбранное из ингибитора ауксина, ингибитора β-лактамазы и ингибитора этилена, позволяет уменьшить рост каллюса в эксплантате и обеспечивает повышение частоты трансформации. 20 з. п.ф-лы, 2 ил., 5 табл.
| Способ получения трансгенных растений картофеля | 1990 |
|
SU1782484A1 |
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| Экономайзер | 0 |
|
SU94A1 |

