Введение
[0001] По настоящей заявке испрашивается приоритет заявки США с серийным номером 16/790,277, поданной 13 февраля 2020 г., содержание которой полностью включено в настоящее описание посредством ссылки.
[0002] Настоящее изобретение было сделано при поддержке правительства по контрактам №№ AI 064745 и AI 116441, заключенным Национальными институтами здравоохранения. Правительство имеет определенные права на изобретение.
Уровень техники
[0003] Лимфатический филяриоз, вызываемый филяриозными нематодами Wuchereha bancrofti, Brugia malayi и Brugia timori, поражает более 120 миллионов человек во всем мире (WHO (1992) World Health Organ. Tech. Rep.Ser. 821:1-71). Программа массового применения лекарственных средств Всемирной организацией здравоохранения значительно снижает уровень заболеваемости лимфатическим филяриозом во многих частях мира (Hotez (2009) Clin. Pharmacol. Ther. 85(6):659-64). Тем не менее, в нескольких эндемичных регионах сообщалось об отсутствии эффективности массового применения лекарственных средств в основном из-за нарушения схем терапии (Babu & (2008) Trans. R. Soc. Trop.Med. Hyg. 102(12):1207-13; El-Setouhy, et al. (2007) Am. J. Trop. Med. Hyg. 77 (6):1069-73). Кроме того, сообщалось о лекарственной резистентности, по меньшей мере, к одному из лекарственных средств в массовой комбинации лекарственных средств (Horton (2009) Ann. Trop.Med. Parasitol. 103(1):S33-40; Schwab, et al. (2007)). Parasitology 134(Pt 7): 1025-40). Поскольку для эффективной борьбы требуется ежегодное массовое применение лекарственных средств, возникает тревожная проблема при отборе резистентных к лекарственным средствам паразитов. Таким образом, существует неотложная необходимость в комплексном подходе к борьбе с этой переносимой комарами инфекцией.
[0004] Как и в случае с лимфатическим филяриозом, лечение дирофиляриоза (дирофиляриоза сердца) у псовых и кошачьих включало использование макролидных агентов, таких как ивермектин, оксим милбемицина, моксидектин и селамектин, которые предотвращают развитие личинок в течение первых 2 месяцев после заражения. Однако для эффективности эти средства необходимо вводить ежемесячно, и они могут быть очень дорогими для владельца домашнего животного.
[0005] Вакцинация является одной из стратегий борьбы с этими инфекциями, и несколько субъединичных вакцинных антигенов-кандидатов были протестированы на лабораторных животных с переменными результатами (Bottazzi, et al. (2006) Expert Rev. Vaccines 5(2):189-98; Chenthamarakshan, et al. (1995) Parasite Immunol. 17(6):277-85, Dissanayake, et al. (1995), Am. J. Trop.Med. Hyg. 53(3):289-94, Li, et al. (1993) J. Immunol. 150(5):1881-5; Maizels, et al., (2001) Int. J. Parasitol. 31(9):889-98; Thirugnanam, et al. (2007), Exp.Parastiol. 116(4):483-91; Veera path ran, et al. (2009) PLoS Negl. Trop.Dis. 3(6):e457). Лимфатический филяриоз представляет собой многоклеточный организм со сложным жизненным циклом и продуцирует большое количество модулирующих молекул-хозяев. Таким образом, борьба с этой инфекцией с помощью вакцины с одним антигеном может быть затруднена. Путем скрининга библиотеки экспрессии кДНК фагового дисплея паразита В. malayi с сывороткой от иммунных индивидуумов было идентифицировано несколько потенциальных вакцин-кандидатов (Gnanasekar, et al. (2004) Infect. Immun. 72(8):4707-15). Однако различная степень защиты была достигнута с каждым из вакцинных антигенов-кандидатов при введении в виде ДНК, белка или прайм-буст вакцины (Veerapathran, et al. (2009) выше).
Сущность изобретения
[0006] Настоящее изобретение представляет собой поливалентную иммуногенную композицию, состоящую из двух или нескольких антигенов Dirofilaria immitis. В некоторых вариантах осуществления, антигены основаны на белке, ДНК или их комбинации. В других вариантах осуществления, антигены включают многокопийный личиночный транскрипт (ALT), малый белок теплового шока (HSP) 12.6, тиоредоксинпероксидазу 2 (ТХР2) или, необязательно, тетраспанин (TSP). В некоторых аспектах, антигены включают антиген ALT, имеющий аминокислотную последовательность SEQ ID NO: 98 или SEQ ID NO: 99; антиген HSP12.6, имеющий аминокислотную последовательность SEQ ID NO: 100 или SEQ ID NO: 101; и/или антиген ТХР2, имеющий аминокислотную последовательность SEQ ID NO: 83 или SEQ ID NO: 101. В некоторых аспектах, антиген ALT имеет аминокислотную последовательность SEQ ID NO: 93; антиген HSP12.6 имеет аминокислотную последовательность SEQ ID NO: 91; и антиген ТХР2 имеет аминокислотную последовательность SEQ ID NO: 95. В некоторых аспектах, антиген ALT имеет аминокислотную последовательность SEQ ID NO: 121 или SEQ ID NO: 122; антиген HSP 12.6 имеет аминокислотную последовательность SEQ ID NO: 81 или SEQ ID NO: 123; антиген TSP имеет аминокислотную последовательность SEQ ID NO: 82; и антиген ТХР2 имеет аминокислотную последовательность SEQ ID NO: 83 или SEQ ID NO: 124. В некоторых аспектах, антигены присоединены ковалентно. Настоящее изобретение также предлагает рекомбинантный вектор, содержащий нуклеиновые кислоты, кодирующие поливалентную иммуногенную композицию, рекомбинантную клетку-хозяина, содержащую рекомбинантный вектор, и включение адъюванта в поливалентную иммуногенную композицию. Также предложены способы индукции защитного иммунного ответа у субъекта и иммунизации животного против филяриоза или дирофиляриоза. В некоторых вариантах осуществления этих способов, поливалентную иммуногенную композицию вводят с адъювантом, например, в одной или нескольких дополнительных дозах путем подкожной или внутримышечной инъекции.
Краткое описание чертежей
[0007] На фиг.1 показан титр анти-BmHSP и анти-BmALT2 IgG антител в сыворотке вакцинированных мышей. 6-недельных мышей balb/c иммунизируют с использованием подхода примирования/стимулирования одновалентной иммуногенной композицией (примирование Bmhsp и стимулирование rBmHSP или примирование Bmalt2 и стимулирование rBmALT2) и поливалентной иммуногенной композицией (примирование Bmhsp/Bmalt2 и стимулирование rBmHSP и rBmALT2). Титр IgG антител измеряют в сыворотке с помощью непрямого ELISA. Предложенные данные представляют собой титр антител через 2 недели после последнего стимулирования. Результаты показывают, что как двухвалентные, так и поливалентные иммуногенные композиции индуцируют значительные IgG антитела против каждого из входящих в их состав антигенов. Результаты также показывают, что антигены в одновалентных и поливалентных препаратах действуют синергетически, усиливая иммунный ответ.N=5. Статистически значимо **р 0,001, *р<0,05. Представленные значения являются средними ±CO.
[0008] На фиг.2А-2 В показано количество клеток, секретирующих IL-4 (фиг.2А) и IFN-((фиг.2 В), в селезенке мышей, вакцинированных одновалентной (BmHSP или BmALT2) или поливалентной иммуногенной композицией. Анализ ELISPOT проводят после стимуляции клеток с rBmHSP или rBmALT (1 мкг/мл). Одноклеточные препараты клеток селезенки стимулируют соответствующими антигенами в течение 48 часов и подсчитывают клетки, образующие «ореол». Результаты показывают, что как одновалентные, так и поливалентные иммуногенные композиции стимулируют клетки, секретирующие IL-4. Поливалентная вакцинация индуцирует большее количество клеток, продуцирующих IL-4, чем контрольные. Количество клеток, продуцирующих IFN-γ, является сравнительно низким. Эти данные дополнительно подтверждают, что BmHSP и BmALT2 синергически усиливают иммунный ответ у вакцинированных животных после поливалентной вакцинации. N=5. Результаты выражены как среднее число единиц, образующих «ореол», на 3×106 клеток ±СО.
[0009] На ФИГ. 3 показана степень защиты, обеспечиваемой поливалентной иммуногенной композицией в мышиной модели. Мышей линии Balb/c иммунизируют гибридной ДНК HAT (HSP/ALT2/TSP), рекомбинантным белком HAT или их комбинацией с использованием подхода примирования/стимулирования. Гибридную ДНК HAT используют для примирования. Через две недели после примирования, мышей стимулируют гибридным белком HAT. Другую группу мышей иммунизируют гибридной ДНК HAT или гибридным белком HAT. Контрольные группы мышей получают только пустой вектор или квасцовый адъювант.Через две недели после последней иммунизации, мышей заражают 20 инфекционными личинками Brugia malayi, помещая их в камеру с микропорами в брюшной полости иммунизированных мышей. Через 48 часов измеряют гибель личинок для определения успеха вакцинации.
[0010] На ФИГ. 4 показана индуцированная поливалентной иммуногенной композицией защита от инфекции Brugia malayi у макак. Всех животных (вакцинированных и контрольных) заражают 130-180 L3 Brugia malayi через месяц после последней иммунизации. На 5, 10, 15 и 18 неделе после заражения, у каждой макаки отбирают 10 мл крови между 18:00 и 22:00 часами, проводят скрининг на наличие микрофилярий с использованием модифицированного метода Нотта и анализируют с помощью ПЦР на повторы Hha-1 повтор. Отсутствие инфекции у животных, отрицательных по микрофилярий (Mf), дополнительно подтверждают с помощью ELISA на SXP-1 (диагностический антиген В. malayi). Результаты показывают, что rBmHAXT+AL019 (квасцы плюс стабильная эмульсия глюкопиранозила-липидного адъюванта) является более иммуногенным составом композиции, чем другие протестированные составы (n=10 на группу). Тест хи-квадрат и точный критерий Фишера используют для сравнения пропорций между группами.
[0011] На ФИГ. 5 показаны результаты анализа антителозависимой клеточно-опосредованной цитотоксичности (ADCC). Приблизительно 10 личинок Brugia malayi инкубируют в течение 72 часов при 37°С с 2×105 мононуклеарных клеток периферической крови (РВМС) и 50 мкл образцов сыворотки от каждой макаки. Гибель личинок в каждой лунке контролируют под световым микроскопом. Каждая точка данных указывает долю гибели личинок с использованием образца сыворотки от одного животного. «+» указывает среднюю долю ларвицидного действия для этой группы, n=10 макак в группе. *Р<0,005 по сравнению с группой AL019 (квасцы плюс стабильная эмульсия глюкопиранозила-липидного адъюванта). Статистический анализ проводят с помощью теста Крускала-Уоллиса с последующей поправкой Бонферрони для нескольких тестов. rBmHAXT, рекомбинантный В. malayi HSP/ALT-2/TPX-2/TSPLEL.
[0012] На ФИГ. 6 показаны результаты анализа ADCC для уничтожения чувствительных к лекарственным средствам и резистентных к лекарственным средствам Dirofilaria immitis у собак. Приблизительно 8-10 личинок D. immitis инкубируют в течение 96 часов при 37°С с 0,5 миллиона РВМС и 100 мкл образцов сыворотки от каждой собаки. Гибель личинок в каждой лунке контролируют под световым микроскопом. Каждая точка данных указывает долю гибели личинок с использованием образца сыворотки от одного животного. rBmHAXT, рекомбинантный В. malayi HSP/ALT-2/TPX-2/TSPLEL.
Подробное описание изобретения
[0013] В настоящее время разработана поливалентная иммуногенная композиция против филяриоза. Комбинации антигенов, таких как многокопийный личиночный транскрипт (ALT2), тетраспанин (TSP), малый белок теплового шока (HSP) 12.6, белок, подобный гомологу аллергена яда осы (VAL-1), глутатион-S-трансфераза (GST) и тиоредоксинпероксидаза 2 (ТРХ-2) и их фрагменты были протестированы на экспериментальных животных (т.е., мышах, песчанках, мастомисах, макаках и собаках) и, как было показано, обеспечивают>80% защиту от заражения филяриозными нематодами, такими как Brugia malayi и Dirofilaria immitis. Соответственно, в настоящем изобретении представлены композиции на основе белков и ДНК, состоящие из антигенов филяриозной нематоды, или нуклеиновых кислот, кодирующих их, и применение иммуногенных композиций для профилактики или борьбы с филяриозом у людей и животных, в частности, псовых и кошачьих. В дополнение к вакцинации, настоящее изобретение также предлагает тесты и наборы для обнаружения присутствия филяриозной нематоды.
[0014] Для целей настоящего изобретения, мультивалентная или поливалентная иммуногенная композиция относится к иммуногенной композиции или вакцине, приготовленной из нескольких антигенов. В соответствии с некоторыми вариантами осуществления, антиген представляет собой молекулу нуклеиновой кислоты, которая упоминается в настоящем документе как антиген «на основе ДНК». В соответствии с другими вариантами осуществления, антиген представляет собой белок или полипептид, который упоминается в настоящем документе как антиген «на основе белка». Поливалентная иммуногенная композиция по изобретению может состоять из двух, трех, четырех, пяти, шести или до десяти антигенов или их фрагментов в различных комбинациях пермутаций. В конкретных вариантах осуществления поливалентная иммуногенная композиция состоит из двух, трех или четырех антигенов. В некоторых вариантах осуществления поливалентная иммуногенная композиция состоит исключительно из белковых антигенов. В других вариантах осуществления поливалентная иммуногенная композиция состоит исключительно из антигенов на основе ДНК. В еще других вариантах осуществления поливалентная иммуногенная композиция состоит из смеси антигенов на основе белка и на основе ДНК.
[0015] Антигены по настоящему изобретению могут быть получены или экспрессированы из одной молекулы нуклеиновой кислоты, содержащей, например, участки внутренней посадки рибосомы между антигенами. Кроме того, антигены поливалентной иммуногенной композиции по настоящему изобретению могут быть ковалентно присоединены с образованием гибридной или химерной молекулы или слитого белка, где антигены непосредственно примыкают друг к другу (например, слияние внутри рамки с или без короткого спейсера). Альтернативно, антигены по настоящему изобретению могут быть предложены в виде смеси отдельных антигенов. Более того, предполагается, что настоящая иммуногенная композиция может состоять из гибридной молекулы, содержащей, например, два антигена в смеси с третьим нековалентно присоединенным антигеном. В качестве иллюстрации, поливалентная иммуногенная композиция по изобретению может состоять из химерного белка TSP-HSP в смеси с молекулой нуклеиновой кислоты, кодирующей ALT2.
[0016] В одном варианте осуществления антигены поливалентной иммуногенной композиции представляют собой разные белки одного вида филяриозной нематоды. В качестве примера этого варианта осуществления поливалентная иммуногенная композиция состоит из антигенов ALT2, HSP и TSP и/или ТРХ2 или GST, выделенных из одного или нескольких штаммов В. malayi или D. immitis. В другом варианте осуществления антигены являются одинаковыми, но из разных видов филяриозных нематод. В качестве примера этого варианта осуществления поливалентная иммуногенная композиция состоит из антигена ALT2, выделенного из W. bancrofti, В. malayi, В. timori и D. immitis. В еще одном варианте осуществления поливалентная иммуногенная композиция состоит из комбинации различных антигенов различных видов филяриозных нематод. В качестве иллюстрации поливалентная иммуногенная композиция может состоять из антигена ALT2, выделенного из W. bancrofti, О. volvulus и L. 1оа, и антигена HSP, выделенного из В. malayi и D. immitis.
[0017] Для получения иммуногенной композиции на основе поливалентной ДНК или поливалентной рекомбинантной ДНК последовательность ДНК представляющего интерес гена (также используемого взаимозаменяемо как молекула ДНК) не обязательно должна содержать полную длину ДНК, кодирующей соответствующий белок. Аналогичным образом, при приготовлении иммуногенных композиций на основе слитых белков или поливалентных рекомбинантных белков последовательность белка не обязательно должна содержать полноразмерный белок. В большинстве случаев для иммунизации достаточно фрагмента белка или гена, кодирующего область эпитопа. ДНК/белковая последовательность области эпитопа может быть найдена путем секвенирования соответствующей части гена из различных штаммов или видов и их сравнения. Основными антигенными детерминантами, вероятно, будут те, которые демонстрируют наибольшую гетерологию. Кроме того, эти области, вероятно, открыто лежат в конформационной структуре белков. Один или несколько таких фрагментов белков или генов, кодирующих антигенные детерминанты, могут быть получены химическим синтезом или технологией рекомбинантной ДНК. Эти фрагменты белков или генов, при желании, могут быть соединены между собой или соединены с другими белками или молекулами ДНК, соответственно.
[0018] Как описано в настоящем документе, антигены ALT2, TSP, VAL-1, GST и HSP были идентифицированы как обеспечивающие защиту от заражения личинками филярий. Соответственно, в конкретных вариантах осуществления, настоящая иммуногенная композиция включает белковые антигены ALT2, TSP, VAL-1, ТРХ2, GST и/или HSP и/или молекулы нуклеиновой кислоты, кодирующие ALT2, TSP, VAL-1, ТРХ2, GST и/или или белок HSP, или его фрагменты. Последовательности белков и нуклеиновых кислот для этих антигенов доступны под номерами доступа GENBANK и/или в виде последовательностей, перечисленных в таблице 1.
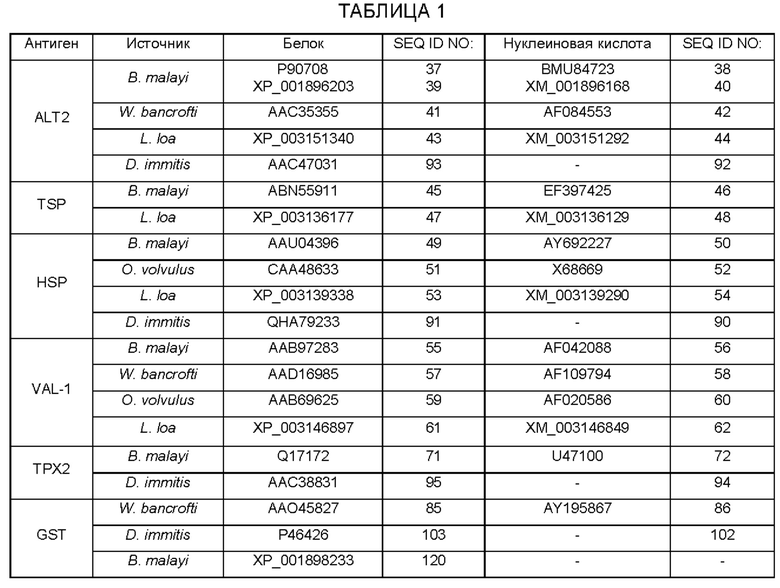
[0019] Кроме того, нуклеотидная последовательность, кодирующая TSP О. volvulus, может быть найдена под №доступа в GENBANK JN861043. Белковые антигены и молекулы нуклеиновых кислот по изобретению могут быть использованы в виде полноразмерных молекул или менее чем полноразмерных молекул. В этом отношении настоящее изобретение дополнительно включает использование фрагментов указанных выше белковых антигенов и молекул нуклеиновых кислот.Фрагменты определяются в настоящем документе как фрагменты из 20, 30, 40, 50, 60, 70, 80, 90,100, 150 или 200 аминокислотных частей полноразмерных белковых антигенов (например, перечисленных в таблице 1) или 60, 90, 120, 150, 180, 210, 240, 270, 300, 350 или 600 нуклеотидных частей полноразмерных молекул нуклеиновых кислот (например, перечисленных в таблице 1). Примеры белковых фрагментов включают домен большой внеклеточной петли (LEL) TSP (см., например, домен LEL В. malayi TSP SEQ ID NO: 63 или SEQ ID NO: 77) и N-концевую делецию HSP 12.6 (cHSP; см., например, В. malayi фрагмент HSP SEQ ID NO: 64), а также молекулы нуклеиновой кислоты, кодирующие его (см. SEQ ID NO: 65 и SEQ ID NO: 66, соответственно). Типовой слитый белок, содержащий белковые последовательности ALT2, HSP и TSP, представлен в SEQ ID NO: 70. Типовой слитый белок, содержащий белковые последовательности ALT2, HSP и ТРХ2, представлен в SEQ ID NO: 73 и SEQ ID NO: 97. Типовой гибридный белок, содержащий белковые последовательности ALT2, HSP, TSP и ТРХ2, представлен в SEQ ID NO: 74.
[0020] В конкретных вариантах осуществления белок или фрагменты белка по настоящему изобретению имеют одну или несколько антигенных последовательностей для индукции иммунного ответа у животного. В некоторых вариантах осуществления белок ALT2 по изобретению представляет собой белок ALT2 В, malayi или его фрагмент, содержащий или состоящий из последовательности VSESDEEFDDSAADDTDDSEAGGGSEGGDEYVT (SEQ ID NO: 78) и/или EFVETDGKKKECSSHEACYDQREPQ (SEQ ID NO: 79) или белок ALT2 D. immitis или фрагмент, содержащий или состоящий из последовательности ASESQEETVSFEESDEDYEDDSE (SEQ ID NO: 98) и/или
FVESDGKMKHCKTHEACYDQREPQ (SEQ ID NO: 99), который, на основе способа прогнозирования линейного эпитопа BepiPpred (Larsen, et al. (2006) Immunome Res, 2:2), являются спрогнозированными В-клеточными эпитопами. В других вариантах осуществления белок HSP по изобретению представляет собой белок HSP В, malayi или его фрагмент, содержащий или состоящий из последовательности WSAEQWDWPLQH (SEQ ID NO: 80) и/или KLPSDVDTKTL (SEQ ID NO: 81), или белок HSP D. immitis или фрагмент, содержащий или состоящий из последовательности NWSADQWDWPLQHNDDVVKVTNTNDK (SEQ ID NO: 100) и/или KLPSDVDTKTL (SEQ ID NO: 81), которые являются спрогнозированными В-клеточными эпитопами. В дополнительных вариантах осуществления белок TSP по изобретению представляет собой белок TSP В. malayi или его фрагмент, содержащий или состоящий из последовательности KTGESEDEMQ (SEQ ID NO: 82), которая представляет собой спрогнозированный В-клеточный эпитоп.В еще одном варианте осуществления белок ТРХ2 по изобретению представляет собой белок ТРХ2 В, malayi или его фрагмент, содержащий или состоящий из последовательности FIGQPAPNFKT (SEQ ID NO: 83) и/или GEVCPANWHPGSETIKPGVKESKA (SEQ ID NO: 84) или белок или фрагмент ТРХ2 D. immitis, содержащий или состоящий из последовательностей FIGQPAPNFKT (SEQ ID NO: 83) и/или GEVCPANWQPGSEAIKPGVKESKA (SEQ ID NO: 101), которые являются спрогнозированными В-клеточными эпитопами.
[0021] В некоторых вариантах осуществления фрагменты белка ALT2 по настоящему изобретению содержат или состоят из аминокислотных последовательностей X1X2ESDEX3X4X5DX6 (SEQ ID NO: 121), где, независимо, X1 представляет собой V или F, Х2 представляет собой S или Е, Х3 представляет собой Е или D, Х4 представляет собой F или Y, X5 представляет собой D или Е и Х6 представляет собой S или D; или FVEX1DGKX2KX3CX4X5HEACYDQREPQ (SEQ ID NO: 122), где, независимо, Xi представляет собой S или Т; Х2 представляет собой М или K; Х3 представляет собой Е или Н, Х4 представляет собой S или K, и X5 представляет собой S или Т. В других вариантах осуществления фрагменты белка HSP по настоящему изобретению содержат или состоят из аминокислотных последовательностей WSAX1QWDWPLQH (SEQ ID NO: 123), где, независимо, X1 представляет собой Glu или Asp; или KLPSDVDTKTL (SEQ ID NO: 81). В дополнительных вариантах осуществления фрагменты белка ТРХ2 по настоящему изобретению содержат или состоят из аминокислотных последовательностей FIGQPAPNFKT (SEQ ID NO: 83); или GEVCPANWX1PGSEX2IKPGVKESKA (SEQ ID NO: 124), где, независимо, X1 представляет собой Н или Q, и Х2 представляет собой Т или А.
[0022] Что касается некоторых вариантов осуществления изобретения, поливалентная иммуногенная композиция по изобретению включает другие известные антигены филяриозных нематод. Примеры других подходящих антигенов включают, но не ограничены ими, глутатионпероксидазу (см. Cookson, et al. (1992) Proc. Natl. Acad. Sci. USA 89:5837-5841; Maizels, et al. (1983) Parasitology 87:249-263; Maizels, et al. (1983) Clin. Exp.Immunol. 51:269-277); рекомбинантный антиген (BmR1; см. Noordin, et al. (2004) Filaha J. 3:10); аминоацил-тРНК синтетазу II класса (см. Kron, et al. (1995) FEBS Lett. 374:122-4); родственный белок теплового шока 70 (hsc70) (см. Selkirk, et al. (1989) J. Immunol. 143:299-308); парамиозин (см. Li, et al. (1991) Mol. Biochem. Parasitol. 49:315-23); тропомиозин (Hartmann, et al. (2006) Vaccine 24(17):3581-90); хитиназу (Adam, et al. (1996) J. Biol. Chem. 271 (3):1441-7); многокопийный личиночный транскрипт (ALT)-1 (Gregory, et al. (2000) Infect. Immun. 68(7):4174-9); иммунодоминантный гиподермальный антиген SPX1 (Bradley, et al. (1993) Exp.Parasitoi. 77(4):414-424). В некоторых вариантах осуществления антиген получают из филяриозной нематоды, выбранной из группы W. bancrofti, В. malayi, О. volvulus, L. loa, D. immitis и В. timori. В некоторых вариантах осуществления антиген представляет собой тропомиозин В. malayi или Dirofilaria, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 104 и SEQ ID NO: 105, соответственно, или его фрагмент; хитиназу В. malayi или Dirofilaria, имеющую аминокислотную последовательность, указанную в SEQ ID NO: 106 и SEQ ID NO: 107, соответственно, или ее фрагмент; ALT-1 В. malayi или Dirofilaria, имеющий аминокислотную последовательность, указанную в SEQ ID NO: 108 и SEQ ID NO: 109, соответственно, или ее фрагмент; SPX1 В. malayi или Dirofilaria, имеющий аминокислотную последовательность, указанную в SEQ ID NO: 110 и SEQ ID NO: 111, соответственно, или ее фрагмент; белок, подобный антигену 5 аллергена яда В. malayi или D. immitis, имеющий аминокислотную последовательность, указанную в SEQ ID NO: 112 и SEQ ID NO: 113, соответственно, или его фрагмент; белок фактора ингибирования миграции макрофагов (MIF)-1 В. malayi или D. immitis, имеющий аминокислотную последовательность, указанную в SEQ ID NO: 114 и SEQ ID NO: 115, соответственно, или его фрагмент; белок MIF-2 В. malayi или Dirofilaria, имеющий аминокислотную последовательность, указанную в SEQ ID NO: 116 и SEQ ID NO: 117, соответственно, или его фрагмент; или цистатиновый белок В. malayi или Dirofilaria, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 118 и SEQ ID NO: 119, соответственно, или его фрагмент.
[0023] Согласно настоящему изобретению антигены слитого белка и иммуногенной композиции выделяют из филяриозной нематоды. В этом отношении выделенная молекула нуклеиновой кислоты или белка представляет собой молекулу нуклеиновой кислоты или белка, которые были удалены из своей естественной среды (т.е. которые были подвергнуты манипуляциям человека). Таким образом, «выделенный» не отражает степень, до которой была очищена молекула нуклеиновой кислоты или белка. В конкретных вариантах осуществления антигены очищают (например, очищают до более чем 95% гомогенности). Выделенная и, необязательно, очищенная молекула нуклеиновой кислоты или белка по настоящему изобретению может быть получена из его природного источника или продуцирована с использованием технологии рекомбинантной ДНК (например, амплификации или клонирования с помощью полимеразной цепной реакции (ПЦР)) или химического синтеза. Выделенные молекулы нуклеиновой кислоты и белка могут также включать, например, природные аллельные варианты или изомеры, которые вызывают иммунный ответ у хозяина.
[0024] Один вариант осуществления настоящего изобретения включает рекомбинантный вектор, который включает, по меньшей мере, одну выделенную молекулу нуклеиновой кислоты по настоящему изобретению, встроенную в вектор, способный доставлять молекулу нуклеиновой кислоты в клетку-хозяина. Такой вектор содержит гетерологичные последовательности нуклеиновых кислот, которые представляют собой последовательности нуклеиновых кислот, которые в природе не встречаются рядом с молекулами нуклеиновых кислот по настоящему изобретению, и которые, предпочтительно, получают от видов, отличных от видов, от которых получена(ы) молекула(ы) нуклеиновой кислоты. Рекомбинантные векторы можно использовать для клонирования, секвенирования и/или других манипуляций с молекулами нуклеиновых кислот по настоящему изобретению.
[0025] Настоящее изобретение также включает вектор экспрессии, который включает молекулу нуклеиновой кислоты по настоящему изобретению в рекомбинантном векторе, способном экспрессировать молекулу нуклеиновой кислоты при трансформации в клетку-хозяина. Предпочтительно, вектор экспрессии также способен к репликации в клетке-хозяине. Векторы экспрессии могут быть либо прокариотическими, либо эукариотическими, и обычно представляют собой вирусы или плазмиды. Векторы экспрессии по настоящему изобретению включают любые векторы, которые функционируют (т.е. направляют экспрессию генов) в рекомбинантных клетках по настоящему изобретению, в том числе в клетках бактерий, грибов, паразитов, насекомых, других животных и растений. Предпочтительные векторы экспрессии по настоящему изобретению могут направлять экспрессию генов в клетках бактерий, дрожжей, червей или других паразитов, насекомых и млекопитающих.
[0026] В частности, векторы экспрессии по настоящему изобретению содержат регуляторные последовательности, такие как регуляторные последовательности транскрипции, регуляторные последовательности трансляции, точки начала репликации и другие регуляторные последовательности, которые совместимы с рекомбинантной клеткой и контролируют экспрессию молекул нуклеиновых кислот по настоящему изобретению. В частности, рекомбинантные молекулы по настоящему изобретению включают регуляторные последовательности транскрипции. Регуляторные последовательности транскрипции представляют собой последовательности, которые контролируют инициацию, удлинение и терминацию транскрипции. Особенно важными регуляторными последовательностями транскрипции являются такие, которые контролируют инициацию транскрипции, такие как промоторные, энхансерные, операторные и репрессорные последовательности. Подходящие регуляторные последовательности транскрипции включают любую регуляторную последовательность транскрипции, которая может функционировать, по меньшей мере, в одной из рекомбинантных клеток по настоящему изобретению. Множество таких контрольных последовательностей транскрипции известно специалистам в данной области техники. Предпочтительные регуляторные последовательности транскрипции включают последовательности, функционирующие в клетках бактерий, дрожжей, червей или других эндопаразитов, или клетках насекомых и млекопитающих, такие как, но не ограничиваясь ими, tac, lac, trp, trc, oxy-pro, omp/lpp, rrnB, бактериофаг лямбда (например, лямбда pL и лямбда pR и гибриды, которые включают такие промоторы), бактериофаг Т7, T7lac, бактериофаг Т3, бактериофаг SP6, бактериофаг SP01, металлотионеин, фактор альфа-спаривания, алкогольоксидаза Pichia, субгеномный промотор альфавируса, ген резистентности к антибиотику, бакуловирус, вирус насекомых Heliothis zea, вирус коровьей оспы, герпесвирус, поксвирус енота, другие поксвирусы, аденовирус, цитомегаловирус (например, немедленно-ранний промотор), вирус 40 обезьян, ретровирус, актин, ретровирусный длинный терминальный повтор, вирус саркомы Рауса, тепловой шок, регуляторные последовательности транскрипции фосфата и нитрата, а также другие последовательности, способные контролировать экспрессию генов в прокариотических или эукариотических клетках. Дополнительные подходящие регуляторные последовательности транскрипции включают тканеспецифические промоторы и энхансеры, а также промоторы, индуцируемые лимфокинами (например, промоторы, индуцируемые интерферонами или интерлейкинами). Регуляторные последовательности транскрипции по настоящему изобретению могут также включать встречающиеся в природе регуляторные последовательности транскрипции, естественным образом связанные с паразитическими червями, такие как регуляторные последовательности транскрипции В. malayi или D. immitis,
[0027] Рекомбинантные молекулы по настоящему изобретению могут также содержать (а) секреторные сигналы (т.е. последовательности нуклеиновых кислот сигнального сегмента), чтобы сделать возможным секретирование экспрессированного белка по настоящему изобретению из клетки, которая продуцирует белок, и/или (b) слитые последовательности, которые приводят к экспрессии молекул нуклеиновых кислот по настоящему изобретению в виде слитых белков. Примеры подходящих сигнальных сегментов включают любой сигнальный сегмент, способный направлять секрецию белка по настоящему изобретению. Предпочтительные сигнальные сегменты включают, но не ограничены ими, сигнальные сегменты тканевого активатора плазминогена (t-PA), интерферона, интерлейкина, гормона роста, гистосовместимости и гликопротеина вирусной оболочки. Кроме того, молекула нуклеиновой кислоты по настоящему изобретению может быть присоединена к слитому сегменту, который направляет кодируемый белок в протеосому, такому как сегмент слияния убиквитина. Эукариотические рекомбинантные молекулы могут также включать промежуточные и/или нетранслируемые последовательности, окружающие и/или находящиеся внутри последовательностей нуклеиновых кислот молекул нуклеиновых кислот по настоящему изобретению.
[0028] Другой вариант осуществления настоящего изобретения включает рекомбинантную клетку-хозяина, содержащую одну или несколько рекомбинантных молекул по настоящему изобретению. Трансформация молекулы нуклеиновой кислоты в клетку может быть осуществлена любым способом, с помощью которого молекула нуклеиновой кислоты может быть встроена в клетку. Методы трансформации включают, но не ограничены ими, трансфекцию, электропорацию, микроинъекцию, липофекцию, адсорбцию и слияние протопластов. Рекомбинантная клетка может оставаться одноклеточной или может вырасти в ткань, орган или многоклеточный организм. Молекулы трансформированных нуклеиновых кислот по настоящему изобретению могут оставаться внехромосомными, или могут интегрироваться в один или несколько сайтов внутри хромосомы трансформированной (т.е. рекомбинантной) клетки таким образом, что сохраняется их способность к экспрессии.
[0029] Клетки-хозяева, подходящие для трансформации, включают любые клетки, которые можно трансформировать молекулой нуклеиновой кислоты по настоящему изобретению. Клетки-хозяева могут быть либо нетрансформированными клетками, либо клетками, которые уже трансформированы с, по меньшей мере, одной молекулой нуклеиновой кислоты (например, молекулами нуклеиновой кислоты, кодирующими один или несколько белков по настоящему изобретению и/или другие белки, пригодные для получения поливалентных иммуногенных композиций). Клетки-хозяева по настоящему изобретению либо могут быть способны к эндогенному (т.е. естественным образом) продуцированию белков по настоящему изобретению, либо могут быть способны к продуцированию таких белков после трансформации, по меньшей мере, одной молекулой нуклеиновой кислоты по настоящему изобретению. Клетки-хозяева по настоящему изобретению могут быть любой клеткой, способной продуцировать, по меньшей мере, один белок по настоящему изобретению, и включают клетки бактерий, грибов (включая дрожжи), паразитов (включая червей, простейших и эктопаразитов), других насекомых, других животных и растений. Предпочтительные клетки-хозяева включают клетки бактерий, микобактерий, дрожжей, червей, насекомых и млекопитающих. Более предпочтительные клетки-хозяева включают Salmonella, Escherichia, Bacillus, Listeria, Saccharomyces, Spodoptera, Mycobacteria, Trichoplusia, клетки BHK (почек детеныша хомячка), клетки MDCK (линию клеток почек собак Madin-Darby), клетки CRFK (линию клеток почек кошек Crandell), клетки CV-1 (линию клеток почек африканских обезьян, используемую, например, для культивирования вируса оспы енота), клетки COS (например, COS-7) и клетки Vero. Особенно предпочтительными клетками-хозяевами являются Escherichia coli, включая производные Е. coil K-12; Salmonella typhi; Salmonella typhimurium; Spodoptera frugiperda; Trichoplusia ni; клетки BHK; клетки MDCK; клетки CRFK; клетки CV-1; клетки COS; клетки Vero; и неканцерогенные миобласты мыши G8 (например, АТСС CRL 1246). Дополнительные подходящие клетки-хозяева млекопитающих включают другие клеточные линии почек, другие клеточные линии фибробластов (например, клеточные линии фибробластов человека, мыши или куриного эмбриона), клеточные линии миеломы, клетки яичников китайского хомячка, клетки мыши NIH/3T3, клетки LMTK31 и/или или клетки HeLa. В одном варианте осуществления, белки могут быть экспрессированы в виде гетерологичных белков в клеточных линиях миеломы с использованием промоторов иммуноглобулина.
[0030] Рекомбинантную клетку предпочтительно продуцируют путем трансформации клетки-хозяина с одной или несколькими рекомбинантными молекулами, каждая из которых содержит одну или несколько молекул нуклеиновой кислоты по настоящему изобретению и одну или несколько регуляторных последовательностей транскрипции, примеры которых описаны в настоящем документе.
[0031] Технологии рекомбинантной ДНК можно использовать для улучшения экспрессии трансформированных молекул нуклеиновых кислот, манипулируя, например, числом копий молекул нуклеиновых кислот в клетке-хозяине, эффективностью, с которой транскрибируются эти молекулы нуклеиновых кислот, эффективностью, с которой полученные транскрипты транслируются, и эффективностью пост-трансляционных модификаций. Рекомбинантные методы, применимые для увеличения экспрессии молекул нуклеиновых кислот по настоящему изобретению, включают, но не ограничены ими, оперативное связывание молекул нуклеиновых кислот с плазмидами с большим числом копий, интеграцию молекул нуклеиновых кислот в одну или несколько хромосом клетки-хозяина, добавление последовательностей стабильности вектора в плазмиды, замены или модификации регуляторных сигналов транскрипции (например, промоторов, операторов, энхансеров), замены или модификации регуляторных сигналов трансляции (например, сайтов связывания рибосом, последовательностей Шайна-Дальгарно), модификации молекул нуклеиновой кислоты по настоящему изобретению, чтобы они соответствовали использованию кодонов в клетке-хозяине, делеции последовательностей, которые дестабилизируют транскрипты, и использование регуляторных сигналов, которые временно отделяют рост рекомбинантных клеток от продуцирования рекомбинантных ферментов во время ферментации. Активность экспрессированного рекомбинантного белка по настоящему изобретению можно улучшить путем фрагментации, модификации или дериватизации молекул нуклеиновой кислоты, кодирующих такой белок. Более того, хотя не кодон-оптимизированные последовательности могут быть использованы для экспрессии слитых белков в клетках-хозяевах, таких как Е, coli (см. таблицу 1), в вариантах осуществления, относящихся к ДНК вакцинам, молекула нуклеиновой кислоты может быть кодон-оптимизирована для облегчения экспрессии в клетки млекопитающих. В этом отношении, кодон-оптимизированные последовательности для BmALT2, N-концевого удаленного HSP 12.6 (cHSP) В. malayi и домена LEL В. malayi Тетраспанина представлены в SEQ ID NO: 67, SEQ ID NO: 68 и SEQ ID NO: 69, соответственно. Кроме того, для облегчения экспрессии одного или нескольких рекомбинантных белков в рекомбинантной клетке-хозяине можно манипулировать последовательностью белка. В качестве иллюстрации, было обнаружено, что вставка глицинового остатка после N-концевого метионинового остатка белка ALT2 В. malayi улучшает экспрессию этого белка в Е. coli.
[0032] Выделенные антигены на основе белков по настоящему изобретению могут быть получены различными способами, включая продуцирование и восстановление природных белков, продуцирование и восстановление рекомбинантных белков и химический синтез белков. В одном варианте осуществления, выделенный белок по настоящему изобретению продуцируют путем культивирования клетки, способной экспрессировать белок, в условиях, эффективных для продуцирования белка, и восстановления белка. Предпочтительной клеткой для культивирования является рекомбинантная клетка по настоящему изобретению. Эффективные условия культивирования включают, но не ограничены ими, эффективную среду, биореактор, температуру, рН и кислородные условия, которые обеспечивают продуцирование белка. Эффективная среда относится к любой среде, в которой культивируют клетку для продуцирования белка по настоящему изобретению. Такая среда обычно включает водную среду, содержащую источники усвояемого углерода, азота и фосфата, и соответствующие соли, минералы, металлы и другие питательные вещества, такие как витамины. Клетки по настоящему изобретению можно культивировать в обычных ферментационных биореакторах, встряхиваемых колбах, пробирках, микротитровальных чашках и чашках Петри. Культивирование можно проводить при температуре, рН и содержании кислорода, подходящих для рекомбинантной клетки. Такие условия культивирования находятся в компетенции специалиста в данной области техники.
[0033] В зависимости от вектора и системы-хозяина, используемых для получения, полученные белки по настоящему изобретению могут либо оставаться в рекомбинантной клетке; секретироваться в ферментационную среду; секретироваться в пространство между двумя клеточными мембранами, такое как периплазматическое пространство Е. coli; либо удерживаться на внешней поверхности клетки или вирусной мембраны.
[0034] Восстановление белков по изобретению может включать сбор всей ферментационной среды, содержащей белок, и не требует дополнительных стадий разделения или очистки. Белки по настоящему изобретению могут быть очищены с использованием различных стандартных методов очистки белков, таких как, но не ограничиваясь ими, аффинная хроматография, ионообменная хроматография, фильтрация, электрофорез, хроматография гидрофобного взаимодействия, гель-фильтрационная хроматография, хроматография с обращенной фазой, хроматография с конканавалином А, хроматофокусирование и дифференциальная солюбилизация. Белки по настоящему изобретению предпочтительно извлекают в по существу чистой форме, что позволяет эффективно использовать белок в качестве терапевтической композиции. Терапевтическая композиция для животных, например, не должна проявлять существенной токсичности и, предпочтительно, должна быть способна стимулировать продуцирование антител у леченного животного.
[0035] Одним из вариантов осуществления настоящего изобретения является иммуногенная композиция или вакцина, которая, при эффективном введении животному способна защитить это животное от филяриоза или дирофиляриоза, вызываемого филяриозной нематодой, такой как нематода Dirofilaria, В некоторых вариантах осуществления изобретение предлагает способ лечения или защиты животного от заболевания, вызванного филяриозной нематодой. В других вариантах осуществления изобретение предлагает способ лечения или защиты животного, например, собаки или кошки, от дирофиляриоза (дирофиляриоза сердца). Иммуногенные композиции включают защитные молекулы, такие как выделенный антигенный белок по настоящему изобретению, выделенная молекула нуклеиновой кислоты по настоящему изобретению и их гибриды и смеси. Используемая в настоящем документе поливалентная иммуногенная композиция по изобретению индуцирует защитный иммунный ответ при эффективном введении животному, такому как человек, кошка или собака, тем самым вылечивая, облегчая и/или предотвращая заболевание, вызванное филяриозной или дирофиляриозной нематодой, включая, но не ограничиваясь ими, W. bancrofti, В, malayi, О. volvulus, L, loa, D, immitis, Mansonella streptocerca, Dracunculus medinensis, M. perstans, M, ozzardi и/или В, timoh. Иммуногенную композицию по настоящему изобретению можно вводить любому животному, восприимчивому к такой терапии, предпочтительно, млекопитающим, и более предпочтительно, людям, домашним животным, таким как собаки и кошки, и животным, потребляемым в пищу, и/или животным зоопарка.
[0036] В одном варианте осуществления поливалентная иммуногенная композиция по настоящему изобретению при введении хозяину может вырабатывать антитела, способные убивать паразитов в векторе, в котором развивается филяриозная нематода, например, в комарах, когда они кормят хозяина.
[0037] Для защиты животного от заболевания, вызванного филяриозной нематодой, иммуногенную композицию по настоящему изобретению вводят животному эффективным образом, так что композиция способна защитить это животное от заболевания, вызываемого филяриозной нематодой. Композиции по настоящему изобретению можно вводить животным до заражения для профилактики инфекции (т.е. в качестве профилактической вакцины) и/или можно вводить животным после заражения для лечения заболевания, вызванного филяриозной нематодой (т.е. в качестве терапевтической вакцины).
[0038] Композиции по настоящему изобретению могут быть составлены в эксципиенте, который может переносить животное, подлежащее лечению. Примеры таких эксципиентов включают воду, физиологический раствор, раствор Рингера, раствор декстрозы, раствор Хенка и другие водные физиологически сбалансированные растворы соли. Также можно использовать не водные носители, такие как нелетучие масла, кунжутное масло, этилолеат или триглицериды. Другие полезные составы включают суспензии, содержащие агенты, повышающие вязкость, такие как карбоксиметилцеллюлоза натрия, сорбит или декстран. Эксципиенты также могут содержать незначительные количества добавок, таких как вещества, повышающие изотоничность и химическую стабильность. Примеры буферов включают фосфатный буфер, бикарбонатный буфер и Трис буфер, тогда как примеры консервантов включают тимеросал, м- или о-крезол, формалин и бензиловый спирт.Стандартные составы могут представлять собой либо жидкие инъекционные препараты, либо твердые вещества, которые можно вводить в подходящую жидкость в виде суспензии или раствора для инъекций. Таким образом, в не жидком составе, эксципиент может содержать декстрозу, сывороточный альбумин человека, консерванты и т.д., к которым перед введением можно добавить стерильную воду или солевой раствор.
[0039] В одном варианте осуществления настоящего изобретения, иммуногенная композиция может включать адъювант.«Адъювант», как определено в настоящем документе, представляет собой вещество, которое служит для усиления иммуногенности иммуногенной композиции по изобретению. Иммунный адъювант может усиливать иммунный ответ на антиген, который является слабо иммуногенным при отдельном введении, например, не индуцируя или индуцируя слабые титры антител или клеточно-опосредованный иммунный ответ, повышать титры антител к антигену и/или снижать дозу антигена, эффективного для достижения иммунного ответа у индивидуума. Таким образом, адъюванты часто вводят для усиления иммунного ответа, и они хорошо известны специалистам в данной области техники.
[0040] Подходящие адъюванты для повышения эффективности иммуногенной композиции включают, но не ограничены ими:
(1) соли алюминия (квасцы), такие как гидроксид алюминия, фосфат алюминия, сульфат алюминия и т.д.;
(2) соли на основе кальция;
(3) диоксид кремния;
(4) составы эмульсии масло-в-воде (с или без других специфических иммуностимулирующих агентов, таких как мурамиловые пептиды (определенные ниже) или компоненты клеточной стенки бактерий), такие как, например,
(a) MF59 (WO 90/14837), содержащая 5% сквалена, 0,5% полисорбата 80 и 0,5% сорбитантриолеата (необязательно содержащий различные количества мурамилтрипептидфосфатидилэтаноламина), приготовленный в виде субмикронных частиц с использованием микрофлюидизатора, такого как микрофлюидизатор Model 110Y (Microfluidics, Newton, MA),
(b) SAF, содержащая 10% сквалена, 0,4% полисорбата 80, 5% плюроник-блокированного полимера L121 и thr-MDP, либо микрофлюидизированная в субмикронную эмульсию, либо вортексированная для получения эмульсии с большим размером частиц,
(c) адъювантная система Ribi((RAS) (Corixa, Hamilton, МТ), содержащая 2% сквалена, 0,2% полисорбата 80 и один или несколько компонентов бактериальной клеточной стенки из группы, состоящей из 3-О-деацилированного монофосфорилипида A (MPL™), описанного в US 4,912,094, димиколата трегалозы (TDM) и скелета клеточной стенки (CWS), предпочтительно, MPL+CWS (Detox™); и
(d) ISA монтанид;
(5) могут быть использованы сапониновые адъюванты, такие как продаваемые под торговыми названиями QUIL-A(или QS-21 STIMULON((Antigenics, Framingham, MA) (см., например, US 5,057,540), или частицы, полученные из них, такие как ISCOM (иммуностимулирующие комплексы, образованные комбинацией холестерина, сапонина, фосфолипида и амфипатических белков) и Iscomatrix((имеющий по существу ту же структуру, что и ISCOM, но без белка);
(6) бактериальные компоненты (например, эндотоксины, в частности, суперантигены, экзотоксины и компоненты клеточной стенки) и липополисахариды, синтетические аналоги липида А, такие как аминоалкилглюкозаминфосфатные соединения (AGP), или их производные или аналоги, которые доступны от Corixa, и описаны в US 6,113,918; один из таких AGP представляет собой 2-[(R)-3-тетрадеканоилокситетрадеканоиламино]этил 2-дезокси-4-O-фосфоно-3-O-[(R)-3-тетрадеканоилокситетрадеканоил]-2-[(R)-3-тетрадеканоилокситетрадеканоиламино]-b-D-глюкопиранозид, также известный как 529 (ранее известный как RC529), который составляют в виде водной формы или в виде стабильной эмульсии;
(7) синтетические полинуклеотиды, такие как олигонуклеотиды, содержащие мотив(ы) CpG (US 6,207,646);
(8) цитокины и хемокины (например, гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), гранулоцитарный колониестимулирующий фактор (G-CSF), макрофагальный колониестимулирующий фактор (M-CSF), колониестимулирующий фактор (CSF), эритропоэтин (ЕРО), интерлейкин 2 (IL-2), IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-10, IL-12, IL-15, IL-18, интерферон гамма, фактор I, индуцирующий интерферон гамма (IGIF), трансформирующий фактор роста бета, RANTES (регулируемый при активации, экспрессируемый и, предположительно, секретируемый нормальными Т-клетками), воспалительные белки макрофагов (например, MIP-1 альфа и MIP-1 бета), фактор некроза опухоли (TNF), костимулирующие молекулы В7-1 и В7-2 и фактор, инициирующий удлинение Leishmania (LEIF));
(9) комплемент, такой кактример компонента комплемента C3d;
(10) агонисты toll-подобных рецепторов, например, агонисты TLR4, такие как глюкопиранозил липидный адъювант (GLA);
(11) белки сыворотки, например, трансферрин;
(12) белки вирусной оболочки, например, белок капсида VP6 ротавируса; и
(13) блок-сополимерные адъюванты, например, адъювант Хантера TITERMAX((VAXCEL, Inc. Norcross, GA).
[0041] Мура мил овые пептиды включают, но не ограничены ими, N-ацетилмурамил-L-треонил-D-изоглутамин (thr-MDP), N-ацетилнормурамил-L-аланин-2-(1'-2'дипальмитоил-sn-глицеро-3-гидроксифосфорилокси)этиламин (МТР-РЕ) и т.д.
[0042] Белковые адъюванты по настоящему изобретению могут быть доставлены в форме самих белков или молекул нуклеиновых кислот, кодирующих такие белки, с использованием способов, описанных в настоящем документе.
[0043] В некоторых вариантах осуществления, адъювант включает соль алюминия. Адъювант на основе соли алюминия может быть вакциной, осажденной квасцами, или вакциной, адсорбированной квасцами. Адъюванты на основе соли алюминия хорошо известны в данной области техники и описаны, например, в Harlow & Lane ((1988) Antibodies: A Laboratory Manual, Cold Spring Harbour Laboratory) и Nicklas ((1992) Res. Immunol. 143:489-493). Соль алюминия включает, но не ограничена ими, гидратированный оксид алюминия, гидрат оксида алюминия, тригидрат оксида алюминия (АТН), гидрат алюминия, тригидрат алюминия, гидроксид алюминия (III), сульфат гидроксифосфата алюминия, адъювант фосфата алюминия (АРА), аморфный оксид алюминия, тригидрат оксида алюминия или тригидроксиалюминий.
[0044] АРА представляет собой водную суспензию гидроксифосфата алюминия. АРА производят путем смешивания хлорида алюминия и фосфата натрия в объемном соотношении 1:1 для осаждения гидроксифосфата алюминия. После процесса смешивания, материал измельчают с помощью смесителя с высоким сдвигом для достижения монодисперсного распределения размеров частиц. Затем продукт диафильтруют против солевого раствора и стерилизуют паром.
[0045] В некоторых вариантах осуществления, коммерчески доступный Al(ОН)3 (например, гель гидроксида алюминия, продаваемый под торговой маркой Alhydrogel®) используется для адсорбции белков в соотношении 50-200 мкг белка/мг гидроксида алюминия. В другом варианте осуществления, адсорбция белка зависит от pl (изоэлектрического рН) белка и рН среды. Белок с более низким pl адсорбируется на положительно заряженном ионе алюминия сильнее, чем белок с более высоким pl. Соли алюминия могут образовывать депо антигена, который медленно высвобождается в течение 2-3 недель, участвующее в неспецифической активации макрофагов и активации комплемента, и/или стимулировать врожденный иммунный механизм (возможно, посредством стимуляции мочевой кислоты). См., например, Lambrecht, et al. (2009) Curr, Opin. Immunol. 21:23.
[0046] В некоторых вариантах осуществления, адъювант представляет собой смесь 2, 3 или более вышеуказанных адъювантов, например, SBAS2 (эмульсия масло-в-воде, также содержащая 3-деацилированный монофосфориллипид А и QS-21); или квасцы в комбинации с GLA (AL019).
[0047] Поливалентная иммуногенная композиция по изобретению может быть составлена в виде флаконов с одной дозой, флаконов с несколькими дозами или в виде предварительно заполненных стеклянных или пластиковых шприцев.
[0048] В одном варианте осуществления, поливалентные иммуногенные композиции по настоящему изобретению вводят перорально и, таким образом, готовят в форме, подходящей для перорального введения, т.е. в виде твердого или жидкого препарата. Твердые пероральные составы включают таблетки, капсулы, пилюли, гранулы, пеллеты и т.п. Жидкие пероральные составы включают растворы, суспензии, дисперсии, эмульсии, масла и подобные.
[0049] Фармацевтически приемлемыми носителями для жидких составов являются водные или не водные растворы, суспензии, эмульсии или масла. Примерами не водных растворителей являются пропиленгликоль, полиэтиленгликоль и органические сложные эфиры для инъекций, такие как этилолеат.Водные носители включают воду, спиртовые/водные растворы, эмульсии или суспензии, включая солевой раствор и буферные среды. Примерами масел являются масла животного, растительного или синтетического происхождения, например, арахисовое масло, соевое масло, оливковое масло, подсолнечное масло, рыбий жир, другой жир морских животных или липид из молока или яиц.
[0050] Фармацевтическая композиция может быть изотонической, гипотонической или гипертонической. Однако часто предпочтительно, чтобы композиция для инфузии или инъекции была по существу изотонической при ее введении. Следовательно, хранение композиции, предпочтительно, может быть изотоническим или гипертоническим. Если композиция является гипертонической для хранения, ее можно развести до изотонического раствора перед введением.
[0051] Изотонический агент может быть ионным изотоническим агентом, таким как соль, или неионным изотоническим агентом, таким как углевод. Примеры ионных изотонических агентов включают, но не ограничены ими, NaCl, CaCl2, KCl и MgCl2. Примеры неионных изотонических агентов включают, но не ограничены ими, маннит, сорбит и глицерин.
[0052] Также предпочтительно, чтобы, по меньшей мере, одна фармацевтически приемлемая добавка представляла собой буфер. Для некоторых целей, например, когда композиция предназначена для инфузии или инъекции, часто желательно, чтобы композиция включала буфер, способный буферировать раствор до рН в диапазоне от 4 до 10, например, от 5 до 9, например, от 6 до 8.
[0053] Буфер может быть, например, выбран из Tris, ацетатного, глутаматного, лактатного, малеатного, тартратного, фосфатного, цитратного, карбонатного, глицинатного, гистидинового, глицинового, сукцинатного и триэтаноламинового буфера. Буфер может быть выбран из USP-совместимых буферов для парентерального применения, в частности, когда состав предназначен для парентерального применения. Например, буфер может быть выбран из группы одноосновных кислот, таких как уксусная, бензойная, глюконовая, глицериновая и молочная; двухосновных кислот, таких как аконитовая, адипиновая, аскорбиновая, карбоновая, глутаминовая, яблочная, янтарная и винная, многоосновных кислот, таких как лимонная и фосфорная; и оснований, таких как аммиак, диэтаноламин, глицин, триэтаноламин и Tris.
[0054] Парентеральные носители (для подкожной, внутривенной, внутриартериальной или внутримышечной инъекции) включают раствор хлорида натрия, декстрозу Рингера, декстрозу и хлорид натрия, лактированный раствор Рингера и нелетучие масла. Внутривенные носители включают вещества для восполнения жидкости и питательных веществ, вещества для восполнения электролитов, такие как средства на основе декстрозы Рингера, и подобные. Примерами являются стерильные жидкости, такие как вода и масла, с или без добавления поверхностно-активного вещества и других фармацевтически приемлемых адъювантов. Как правило, предпочтительными жидкими носителями являются вода, солевой раствор, водная декстроза и родственные растворы сахаров, гликоли, такие как пропиленгликоли или полиэтиленгликоль, полисорбат 80 (PS-80), полисорбат 20 (PS-20) и полоксамер 188 (Р188), в частности, для растворов для инъекций. Примерами масел являются масла животного, растительного или синтетического происхождения, например, арахисовое масло, соевое масло, оливковое масло, подсолнечное масло, рыбий жир, жир других морских обитателей или липид из молока или яиц.
[0055] Составы по изобретению могут также содержать поверхностно-активное вещество. Предпочтительные поверхностно-активные вещества включают, но не ограничены ими, поверхностно-активные вещества на основе сложных эфиров полиоксиэтиленсорбитана, особенно PS-20 и PS-80; сополимеры этиленоксида (ЕО), пропиленоксида (РО) и/или бутиленоксида (ВО), продаваемые под торговой маркой DOWFAX™, такие как линейные блок-сополимеры ЕО/РО; октоксинолы, которые могут различаться по количеству повторяющихся этокси(окси-1,2-этанд пильных) групп, где особый интерес представляет октоксинол-9 (Тритон Х-100 или трет-октилфеноксиполиэтоксиэтанол); (октилфенокси)полиэтоксиэтанол (IGEPAL CA-630/NP-40); фосфолипиды, такие как фосфатидилхолин (лецитин); этоксилаты нонилфенола, такие как серия Tergitol(NP; жирные эфиры полиоксиэтилена, полученные из лаурилового, цетилового, стеарилового и олеилового спиртов (известные как поверхностно-активные вещества Brij), такие как монолауриловый эфир триэтиленгликоля (Brij 30); и сложные эфиры сорбитана, такие как сорбитантриолеат и сорбитанмонолаурат.Предпочтительным поверхностно-активным веществом для включения в эмульсию является PS-80.
[0056] Можно использовать смеси поверхностно-активных веществ. Комбинация сложного эфира полиоксиэтиленсорбитана, такого как моноолеат полиоксиэтиленсорбитана (PS-80), и октоксинола, такого как трет-октилфеноксиполиэтоксиэтанол, также является подходящей. Другая полезная комбинация включает лаурет 9 плюс сложный эфир полиоксиэтиленсорбитана и/или октоксинол.
[0057] Полоксамер также можно использовать в композициях по изобретению. Полоксамер представляет собой неионный триблоксополимер, состоящий из центральной гидрофобной цепи полиоксипропилена (поли(пропиленоксида)), фланкированной двумя гидрофильными цепями полиоксиэтилена (поли(этиленоксида)). Полоксамеры также известны под торговой маркой Pluronic®. Поскольку длину полимерных блоков можно регулировать, существует множество различных полоксамеров, которые имеют немного отличающиеся свойства. Для общего термина «полоксамер» эти сополимеры обычно обозначаются буквой «Р» (для полоксамера), за которой следуют три цифры, первые две цифры ×100 дают приблизительную молекулярную массу полиоксипропиленового ядра, и последняя цифра ×10 дает долю полиоксиэтилена (например, Р407=полоксамер с молекулярной массой полиоксипропилена 4000 г/моль и содержанием полиоксиэтилена 70%). Для торговой марки Pluronic®, кодирование этих сополимеров начинается с буквы, обозначающей их физическую форму при комнатной температуре (L=жидкость, Р=паста, F=хлопья (твердое вещество)), за которой следуют две или три цифры. Первая цифра (две цифры в трехзначном числе) в числовом обозначении, умноженная на 300, указывает примерную молекулярную массу гидрофоба; и последняя цифра ×10 указывает долю полиоксиэтилена (например, L61 представляет собой Pluronic(с молекулярной массой полиоксипропилена 1800 г/моль и содержанием полиоксиэтилена 10%). См. US 3,740,421.
[0058] Предпочтительно, полоксамер обычно имеет молекулярную массу в диапазоне от 1100 до 17400 Да, от 7500 до 15000 Да или от 7500 до 10000 Да. Полоксамер может быть выбран из полоксамера 188 или полоксамера 407. Конечная концентрация полоксамера в составах составляет от 0,001% до 5% масса/объем или от 0,025% до 1% масса/объем. В некоторых аспектах, полиол представляет собой пропиленгликоль и имеет конечную концентрацию от 1% до 20% масса/объем. В некоторых аспектах, полиол представляет собой полиэтиленгликоль 400 и имеет конечную концентрацию от 1% до 20% масса/объем.
[0059] Подходящими полиолами для составов по изобретению являются полимерные полиолы, в частности, диолы простых полиэфиров, включая, но не ограничиваясь ими, пропиленгликоль и полиэтиленгликоль, монометиловые эфиры полиэтиленгликоля. Пропиленгликоль доступен в диапазоне молекулярной массы мономера от примерно 425 до примерно 2700. Полиэтиленгликоль и монометиловый эфир полиэтиленгликоля также доступны в диапазоне молекулярной массы в диапазоне от примерно 200 до примерно 35000, включая, но не ограничиваясь ими, PEG200, PEG300, PEG400, PEG1000, PEG ММЕ 550, PEG ММЕ 600, PEG ММЕ 2000, PEG ММЕ 3350 и PEG ММЕ 4000. Предпочтительным полиэтиленгликолем является полиэтиленгликоль 400. Конечная концентрация полиола в композициях по изобретению может составлять от 1% до 20% масса/объем или от 6% до 20% масса/объем.
[0060] Состав может также содержать рН-забуференный солевой раствор. Буфер может быть, например, выбран из группы, состоящей из Tris, ацетата, глутамата, лактата, малеата, тартрата, фосфата, цитрата, карбоната, глицината, гистидина, глицина, сукцината, HEPES (4-(2-гидроксиэтил)-1-пиперазинэтансульфоновой кислоты), MOPS (3-(N-морфолино)пропансульфоновой кислоты), MES (2-(N-морфолино)этансульфоновой кислоты) и триэтаноламинового буфера. Буфер способен буферировать раствор до рН в диапазоне от 4 до 10, от 5,2 до 7,5 или от 5,8 до 7,0. В некоторых аспектах изобретения, буфер выбран из группы, включающей фосфат, сукцинат, гистидин, сольват, MOPS, HEPES, ацетат или цитрат.Кроме того, буфер может быть выбран, например, из USP-совместимых буферов для парентерального применения, в частности, когда фармацевтическая композиция предназначена для парентерального применения. Концентрации буфера будут варьироваться от 1 мМ до 100 мМ. Концентрации буфера будут варьироваться от 10 мМ до 80 мМ. Концентрации буфера будут варьироваться от 1 мМ до 50 мМ или от 5 мМ до 50 мМ.
[0061] В то время как солевой раствор (т.е. раствор, содержащий NaCl) предпочтителен, другие соли, подходящие для составления, включают, но не ограничены ими, CaCl2, KCl и MgCl2 и их комбинации. Вместо соли можно использовать неионные изотонические агенты, включая, но не ограничиваясь ими, сахарозу, трегалозу, маннит, сорбит и глицерин. Подходящие диапазоны солей включают, но не ограничены ими, от 25 мМ до 500 мМ или от 40 мМ до 170 мМ. В одном аспекте, солевой раствор представляет собой NaCl, необязательно присутствующий в концентрации от 20 мМ до 170 мМ.
[0062] В некоторых аспектах композицию по изобретению вводят субъекту одним или несколькими способами, известными специалисту в данной области техники, например, парентерально, трансмукозально, трансдермально, внутримышечно, внутривенно, внутрикожно, интраназально, подкожно, внутрибрюшинно и в соответствующем составе. В одном варианте осуществления композицию по настоящему изобретению вводят посредством эпидермальной инъекции, внутримышечной инъекции, внутривенной, внутриартериальной, подкожной инъекции или инъекции через слизистую оболочку дыхательных путей жидкого препарата. Жидкие составы для инъекций включают растворы и подобные.
[0063] Один из вариантов осуществления настоящего изобретения представляет собой препарат с контролируемым высвобождением, который способен медленно высвобождать композицию по настоящему изобретению у животного. Используемый в настоящем документе состав с контролируемым высвобождением включает композицию по настоящему изобретению в носителе с контролируемым высвобождением. Подходящие носители с контролируемым высвобождением включают, но не ограничены ими, биосовместимые полимеры, другие полимерные матрицы, капсулы, микрокапсулы, микрочастицы, болюсные препараты, осмотические насосы, диффузионные устройства, липосомы, липосферы и системы трансдермальной доставки. Другие составы с контролируемым высвобождением по настоящему изобретению включают жидкости, которые при введении животному образуют твердое вещество или гель in situ. Предпочтительные составы с контролируемым высвобождением являются биоразлагаемыми (т.е. биоразрушаемыми).
[0064] Предпочтительный состав с контролируемым высвобождением способен высвобождать иммуногенную композицию по настоящему изобретению в кровь лечимого животного с постоянной скоростью, достаточной для достижения терапевтических уровней дозы композиции для защиты животного от заболевания, вызываемого филяриозной нематодой. Например, иммуногенную композицию можно вводить с помощью внутривенной инфузии, трансдермального пластыря, липосом или других способов введения. В другом варианте осуществления, используются полимерные материалы, например, в виде микросфер или имплантата. Иммуногенная композиция предпочтительно высвобождается в течение периода времени от примерно 1 до примерно 12 месяцев. Состав с контролируемым высвобождением по настоящему изобретению способен оказывать лечебное действие, предпочтительно, в течение, по меньшей мере, примерно 1 месяца, более предпочтительно, в течение, по меньшей мере, примерно 3 месяцев, еще более предпочтительно, в течение, по меньшей мере, примерно 6 месяцев, еще более предпочтительно, в течение, по меньшей мере, примерно 9 месяцев, и еще более предпочтительно, в течение, по меньшей мере, примерно 12 месяцев.
[0065] Иммуногенные композиции или вакцины по настоящему изобретению можно вводить животным до заражения для профилактики инфекции, и/или можно вводить животным после заражения для лечения заболевания, вызванного филяриозной нематодой. Например, в качестве иммунотерапевтических агентов можно использовать белки, нуклеиновые кислоты и их смеси. Приемлемые протоколы эффективного введения композиций включают размер индивидуальной дозы, количество доз, частоту введения дозы и способ введения. Определение таких протоколов может быть выполнено специалистами в данной области техники. Подходящая разовая доза представляет собой дозу, которая способна защитить животное от заболевания при введении один или несколько раз в течение подходящего периода времени. Например, предпочтительная разовая доза вакцины на основе белка составляет от примерно 1 микрограмма (пг) до примерно 10 миллиграммов (мг) вакцины на основе белка на килограмм массы тела животного. Стимулирующие вакцинации можно вводить от примерно 2 недель до нескольких лет после первоначального введения. Стимулирующие введения предпочтительно вводят, когда иммунный ответ животного становится недостаточным для защиты животного от болезни. Предпочтительной схемой введения является такая, при которой от примерно 10 мкгдо примерно 1 мг вакцины на кг массы тела животного вводят от примерно одного до примерно двух раз в течение периода времени от примерно 2 недель до примерно 12 месяцев. Способы введения могут включать, но не ограничены ими, подкожный, внутрикожный, внутривенный, интраназальный, пероральный, трансдермальный и внутримышечный пути.
[0066] Если иммуногенная композиция включает молекулу нуклеиновой кислоты, иммуногенную композицию можно вводить животному таким образом, чтобы обеспечить экспрессию этой молекулы нуклеиновой кислоты в защитный белок у животного. Молекулы нуклеиновой кислоты могут быть доставлены животному различными способами, включая, но не ограничиваясь ими, введение голой (т.е. не упакованной в вирусную оболочку или клеточную мембрану) нуклеиновой кислоты в качестве генетической вакцины (например, в виде голой ДНК молекулы, как описано, например, в Wolff, et al., (1990) Science 247:1465-1468); или введение молекулы нуклеиновой кислоты, упакованной в виде рекомбинантной вирусной вакцины или рекомбинантной клеточной вакцины (т.е. молекула нуклеиновой кислоты доставляется с помощью вирусного или клеточного носителя).
[0067]Генетическая (т.е. голая нуклеиновая кислота) вакцина по настоящему изобретению включает молекулу нуклеиновой кислоты по настоящему изобретению и, предпочтительно, включает рекомбинантную молекулу по настоящему изобретению, которая, предпочтительно, способна к репликации или иным образом, к амплификации. Генетическая вакцина по настоящему изобретению может включать одну или несколько молекул нуклеиновой кислоты по настоящему изобретению в форме, например, дицистронной рекомбинантной молекулы. Предпочтительные генетические вакцины включают, по меньшей мере, часть вирусного генома (т.е. вирусный вектор). Предпочтительные вирусные векторы включают векторы на основе альфавирусов, поксвирусов, аденовирусов, герпесвирусов, пикорнавирусов и ретровирусов, где особенно предпочтительными являются векторы на основе альфавирусов (таких как синдбис или лесной вирус Семлики), видоспецифические герпесвирусы и поксвирусы. Можно использовать любую подходящую регуляторную последовательность транскрипции, в том числе описанную как подходящую для продуцирования белка. Особенно предпочтительные регуляторные последовательности транскрипции, включают немедленно-ранний цитомегаловирус (предпочтительно, в сочетании с интроном-А), длинный концевой повтор вируса саркомы Рауса и тканеспецифические регуляторные последовательности транскрипции, а также регуляторные последовательности транскрипции, эндогенные для вирусных векторов, если используются вирусные векторы. Включение «сильного» сигнала полиаденилирования также является предпочтительным.
[0068] Генетические вакцины по настоящему изобретению можно вводить различными способами, включая внутримышечный, подкожный, внутрикожный, чрескожный, интраназальный и пероральный пути введения. Кроме того, предполагается, что вакцина может быть доставлена с помощью генной пушки, кожного пластыря, электропорации или доставки на основе нанотехнологий. В этом отношении вакцины на основе ДНК и белка можно вводить одновременно. Предпочтительная однократная доза генетической вакцины находится в диапазоне от примерно 1 нанограмма (нг) до примерно 600 мкг, в зависимости от пути введения и/или способа доставки, что может быть определено специалистами в данной области техники. Подходящие способы доставки включают, например, инъекцию, капли, аэрозоль и/или местную доставку. Генетические вакцины по настоящему изобретению могут содержаться в водном эксципиенте (например, солевом растворе с фосфатным буфером) отдельно или в носителе (например, носителях на основе липидов).
[0069] Рекомбинантная вирусная вакцина по настоящему изобретению включает рекомбинантную молекулу по настоящему изобретению, которая упакована в вирусную оболочку, и которая может экспрессироваться в организме животного после введения. Предпочтительно, рекомбинантная молекула является дефицитной по упаковке или репликации и/или кодирует ослабленный вирус. Можно использовать ряд рекомбинантных вирусов, включая, но не ограничиваясь ими, вирусы, основанные на альфавирусах, поксвирусах, аденовирусах, герпесвирусах, пикорнавирусах и ретровирусах. Предпочтительными рекомбинантными вирусными вакцинами являются вакцины на основе альфавирусов (таких как вирус Синдбис), поксвирусов енота, видоспецифичных герпесвирусов и видоспецифичных поксвирусов. Примеры способов получения и применения альфавирусных рекомбинантных вирусных вакцин описаны в публикации РСТ №WO 94/17813.
[0070] При введении животному рекомбинантная вирусная вакцина по настоящему изобретению заражает клетки иммунизированного животного и направляет продуцирование защитного белка, способного защитить животное от филяриоза, вызываемого филяриозными нематодами. В качестве иллюстрации разовая доза рекомбинантной вирусной вакцины по настоящему изобретению может составлять от примерно 1×104 до примерно 1×108 вирусных бляшкообразующих единиц (pfu) на килограмм массы тела животного. Протоколы введения аналогичны протоколам, описанным в настоящем документе для белковых вакцин, с подкожным, внутримышечным, интраназальным и пероральным способами введения.
[0071] Рекомбинантная клеточная вакцина по настоящему изобретению включает рекомбинантные клетки по настоящему изобретению, которые экспрессируют белок по настоящему изобретению. Предпочтительные рекомбинантные клетки для этого варианта осуществления включают Salmonella, Е, coll, Listeria, Mycobacterium, S. frugiperda, дрожжи (включая Saccharomyces cerevisiae и Pichia pastoris), BHK, CV-1, миобласт G8, COS (например, COS-7), рекомбинантные клетки Vero, MDCKn CRFK. Рекомбинантные клеточные вакцины по настоящему изобретению можно вводить различными способами, но их преимущество заключается в том, что их можно вводить перорально, предпочтительно, в дозах от примерно 108 до примерно 1012 клеток на килограмм массы тела. Протоколы введения аналогичны протоколам, описанным в настоящем документе для вакцин на основе белка. Рекомбинантные клеточные вакцины могут включать целые клетки, клетки, лишенные клеточных стенок, или клеточные лизаты.
[0072] В некоторых вариантах осуществления композиции по изобретению, все антигены присутствуют в композиции в одинаковом количестве. В дополнительных вариантах осуществления, антигены присутствуют в композиции в различных количествах (т.е., по меньшей мере, один антиген присутствует в количестве, которое отличается от количества одного или нескольких других антигенов композиции).
[0073] Оптимальные количества компонентов для конкретной иммуногенной композиции могут быть установлены стандартными исследованиями, включающими наблюдение соответствующих иммунных ответов у субъектов. Например, в другом варианте осуществления, дозу для вакцинации человека определяют путем экстраполяции данных исследований на животных на данные о людях. В другом варианте осуществления дозировку определяют эмпирически.
[0074] Как известно в данной области техники, существуют три группы филяриозных нематод, классифицируемых в соответствии с занимаемой ими нишей в организме: лимфатический филяриоз, подкожный филяриоз и филяриоз серозных полостей. Лимфатический филяриоз вызывается червями W, bancrofti, В, malayi и В, timori. Эти черви заселяют лимфатическую систему, включая лимфатические узлы, и вызывают лихорадку, лимфаденит (отек лимфатических узлов), лимфангит (воспаление лимфатических сосудов в ответ на инфекцию) и лимфедему (слоновость). Подкожный филяриоз может быть вызван Loa loa (африканский глазной червь), Mansonella stretocerca, О. volvulus, Dracunculus medinensis или Dirofilaria immitis. Многие из этих червей занимают подкожный слой кожи, в жировом слое, и проявляются кожной сыпью, уртикарными папулами и артритом, а также пятнами гипер- и гипопигментации. Onchocerca volvulus проявляется в глазах, вызывая «речную слепоту». Взрослые Dirofilaria immitis обитают в легочных артериях и являются возбудителями дирофиляриоза сердца. Филяриоз серозной полости вызывается червями М. perstans и М, ozzardi, заселяющими серозную полость живота. Филяриоз серозной полости проявляется симптомами, сходными с симптомами подкожного филяриоза, помимо болей в животе, поскольку эти черви также обитают в глубоких тканях.
[0075] У собак, инфицированных Brugia malayi, развиваются клиническая лимфедема, увеличение мошонки, конъюнктивит и лимфагит, сходные с лимфатическим филяриозом человека; однако патология протекает не так тяжело, как у человека. Поскольку собаки переносят инфекцию в природе, люди могут заразиться Brugia malayi от собак. Таким образом, зоонозные инфекции распространены в эндемичных районах, где собаки и кошки переносят инфекцию в природе и передают инфекцию человеку. Собаки и кошки также могут быть инфицированы Brugia malayi в лабораторных условиях. Таким образом, иммуногенная композиция, разработанная против лимфатического филяриоза у собак, также важна для блокирования передачи заболевания человеку.
[0076] Эффективность поливалентной иммуногенной композиции по настоящему изобретению для защиты животного от филяриоза или дирофиляриоза, вызываемого филяриозными нематодами, можно тестировать различными способами, включая, но не ограничиваясь ими, обнаружение защитных антител (с использованием, например, белков настоящего изобретения), обнаружение клеточного иммунитета у леченного животного и/или контрольное заражение леченного животного филяриозной нематодой для определения резистентности обработанного животного к заболеванию и отсутствия одного или нескольких признаков заболевания. Исследования с заражением могут включать имплантацию камер, содержащих личинок филяриозных нематод, животному, получающему лечение, и/или прямое введение личинок животному, получающему лечение. В одном варианте осуществления терапевтические композиции можно тестировать на животных моделях, таких как мыши, песчанки (Meriones unguiculatus), мастомисы (например, Mastomys natalensis) и/или собаки. Такие способы известны специалистам в данной области техники.
[0077] Для обнаружения присутствия/количества антител против филяриозных нематод, например, защитных или нейтрализующих антител, полученных в результате вакцинации животного, настоящее изобретение также предлагает способ и набор для оценки эффективности, а также для выявления предшествующего воздействия белков филярий и/или заражения филяриозной нематодой. В соответствии с таким способом один или несколько антигенных белков/эпитопов контактируют с биологическим образцом животного, и связывание между антигенными белками/эпитопами и антителами в биологическом образце определяют количественно или качественно, как описано в настоящем документе, где присутствие и/или количество антител к антигенным белкам/эпитопам указывает на эффективность вакцины, а также на предшествующее воздействие белков филярий или существующее заражение филяриозной нематодой. В некоторых вариантах осуществления, в способе и наборе используется формат на основе матрицы, в котором печатаются серийные разведения одного или нескольких антигенов или эпитопов. В некоторых вариантах осуществления один или несколько белков филяриозной нематоды присутствуют на одной или нескольких твердых поверхностях или частицах. В других вариантах осуществления один или несколько белков филяриозной нематоды находятся в матрице, так что присутствие множественных антител можно оценить в одном анализе благодаря возможности мультиплексирования подхода на основе матрицы. В этом отношении, матрица может содержать один или несколько белков ALT2, TSP, VAL-1, ТРХ2, GST или HSP или их эпитопов. В других вариантах осуществления матрица содержит, по меньшей мере, каждый из белков, используемых в поливалентной иммуногенной композиции. Например, для анализа защитных или нейтрализующих антител против поливалентной иммуногенной композиции, содержащей HSP, ALT2 и TSP, матрица должна содержать HSP, ALT2 и TSP или их слитый белок.
[0078] Для тестирования на наличие филяриозной нематоды, настоящее изобретение также предлагает способ и набор для обнаружения филяриозной нематоды. Способ анализа обычно включает стадии контакта in vitro биологического образца с одним или несколькими агентами, связывающими белки филяриозных нематод, выбранными из группы, состоящей из ALT2, TSP, VAL-1, ТРХ2, GST и HSP или их фрагментов. Затем обнаруживают связанные связывающие агенты. Связанные связывающие агенты могут быть обнаружены с использованием автоматического обнаружения связывания, такого как считыватель изображений анализа ELISA, и если связанный связывающий агент обнаружен, данные, указывающие на обнаружение связанного связывающего агента, могут быть переданы, например, на дисплей компьютера или распечатаны на бумаге. Обнаружение белка филяриозной нематоды указывает на то, что образец или субъект, от которого был получен образец, болен филяриозом. Таким образом, обнаружение позволяет выбрать варианты лечения субъекта. Таким образом, в одном варианте осуществления, при обнаружении одного или нескольких из ALT2, TSP, VAL-1, ТРХ2, GST и HSP пациенту будет назначено лечение, подходящее для филяриоза, включая, но не ограничиваясь ими, лечение диэтилкарбамазином, мебендазолом, флубендазолом, альбендазолом, ивермектином или их комбинацией.
[0079] Биологический образец представляет собой любой материал, подлежащий тестированию на наличие или количество представляющего интерес белка (например, антитела или антигена/эпитопа). Образец может представлять собой жидкий образец, предпочтительно, образец жидкости. Примеры образцов жидкости, которые можно тестировать в соответствии с настоящим изобретением, включают жидкости организма, включая кровь, сыворотку, плазму, слюну, мочу, глазную жидкость, сперму и спинномозговую жидкость. Вязкие жидкие, полутвердые или твердые образцы (например, ткани человека, ткани комаров или мух) можно использовать для создания жидких растворов, элюатов, суспензий или экстрактов, которые могут быть образцами. В некоторых вариантах осуществления биологический образец является неразведенным. В других вариантах осуществления образец разводят или концентрируют в зависимости от применения обнаружения.
[0080] В некоторых вариантах осуществления можно концентрировать белки в образце, используя твердую поверхность, покрытую моноклональным антителом, для захвата белка. Восстановленные захваченные белки затем могут быть проанализированы с использованием любого подходящего способа, описанного в настоящем документе. Твердая поверхность может представлять собой, например, микроносители, такие как магнитные микроносители, полистироловые микроносители или золотые микроносители, или быть в формате матрицы или микроматрицы с использованием стеклянного, пластикового или кремниевого чипа. Такой захват белка также может быть частью канала в микрожидкостном устройстве.
[0081] Связывающие агенты, используемые в настоящем изобретении, включают антитело, фрагмент антитела или производное антитела (например, аптамер), которое специфически связывается с родственным белком филяриозной нематоды. Специфическое связывание между двумя объектами обычно относится к аффинности, по меньшей мере, 106, 107, 108, 109 или 1010 М-1. Аффинность больше 108 М-1 необходима для достижения специфического связывания.
[0082] Когда связывающим агентом является антитело, антитело может быть получено естественным путем (т.е. иммунизацией) или частично или полностью синтетическим путем. Антитела могут быть моноклональными или поликлональными, и включают коммерчески доступные антитела. Антитело может быть представителем любого класса иммуноглобулинов, включая любой из классов человека: IgG, IgM, IgA, IgD и IgE. Биспецифические и химерные антитела также входят в объем настоящего изобретения. Однако желательны производные IgG класса. Кроме того, антитело может происходить от человека, мыши, крысы, козы, овцы, кролика, курицы, верблюда или осла, или от других видов, которые можно использовать для получения нативных или человеческих антител (т.е. рекомбинантных бактерий, бакуловирусов или растений).
[0083] Например, продуцируемые в природе моноклональные антитела могут быть созданы с использованием классических способов или способов клонирования и слияния клеток, в которых В-клетки захватывают и амплифицируют нуклеиновые кислоты, кодирующие специфическое антитело (см., например, US 20060051348). В таких способах набор белков или отдельный белок (например, пептид или полипептид) можно использовать для начальной иммунизации, и в контексте продуцирования антител он упоминается в настоящем документе как антиген. Исследуемый антиген обычно вводят (например, внутрибрюшинной инъекцией) мышам дикого типа или инбредным мышам (например, BALB/c) или крысам, кроликам, курам, овцам, козам или другим видам животных, которые могут продуцировать нативные или человеческие антитела. Антиген можно вводить отдельно или в смеси с адъювантом. После стимуляции животного, например, два или несколько раз, селезенку или большой лимфатический узел, такой как подколенный у крысы, удаляют, и спленоциты или лимфоциты выделяют и сливают с клетками миеломы с использованием хорошо известных процессов, например, см. Kohler & Milstein ((1975) Nature 256:495-497) или Harlow & Lane (Antibodies: A Laboratory Manual (Cold Spring Harbor Laboratory, New York (1988)). Полученные гибридные клетки затем клонируют обычным способом, например, с использованием лимитирующего разведения, и полученные клоны, которые продуцируют желаемые моноклональные антитела, культивируют (см. Stewart (2001) Monoclonal Antibody Production. In: Basic Methods in Antibody Production and Characterization, Howard and Bethell (eds.), CRC Press, Boca Raton, FL, стр. 51-67).
[0084] Альтернативно, антитела могут быть получены способом фагового дисплея. Способы продуцирования антител фаговым дисплеем известны в данной области техники, например, см. Huse, et al. ((1989) Science 246(4935): 1275-81). Выбор антител основан на аффинности связывания с представляющим интерес белком или белками.
[0085] Фрагмент антитела охватывает, по меньшей мере, значительную часть специфической связывающей способности полноразмерного антитела. Примеры фрагментов антител включают, но не ограничены ими, Fab, Fab', F(ab')2, scFv, Fv, dsFv, диатело, Fd фрагменты или микротела. Фрагмент антитела может содержать несколько цепей, связанных друг с другом, например, дисульфидными связями. Фрагмент также может необязательно представлять собой мультимолекулярный комплекс. Функциональный фрагмент антитела обычно включает, по меньшей мере, примерно 50 аминокислотных остатков, и более типично, включает, по меньшей мере, примерно 200 аминокислотных остатков. Фрагмент антитела может быть получен любым способом. Например, фрагмент антитела можно получить ферментативно или химически путем фрагментации интактного антитела, или его можно получить рекомбинантным путем из гена, кодирующего частичную последовательность антитела. Альтернативно, фрагмент антитела может быть полностью или частично получен синтетическим путем.
[0086] Пептидные аптамеры, которые специфически связываются с белком, обычно рационально конструируют или подвергают скринингу в библиотеке аптамеров (например, предоставленной Aptanomics SA, Lyon, France). В целом, пептидные аптамеры представляют собой синтетические молекулы распознавания, дизайн которых основан на структуре антител. Пептидные аптамеры состоят из вариабельной пептидной петли, прикрепленной на обоих концах к белковому каркасу. Это двойное структурное ограничение значительно увеличивает аффинность связывания пептидного аптамера до уровней, сравнимых с таковыми у антитела (наномолярный диапазон).
[0087] Рекомбинантное продуцирование связывающих агентов настоящего изобретения может быть достигнуто с использованием обычных методов молекулярной биологии и коммерчески доступных систем экспрессии. Кроме того, связывающие агенты могут быть получены с использованием твердофазных технологий (см., например, Merrifield (1963) J. Am. Chem. Soc. 85:2149-2154; Seeberger (2003) Chem. Commun. (Camb) (10):1115-21). Синтез белка может быть выполнен с использованием ручных методов или с помощью автоматизации. Автоматизированный синтез может быть осуществлен, например, с использованием пептидного синтезатора Applied Biosystems 431A (Perkin Elmer, Boston, MA). Различные фрагменты связывающего агента могут быть химически синтезированы по отдельности и объединены с использованием химических способов для получения полноразмерной молекулы.
[0088] Более того, подходы комбинаторной химии могут быть использованы для получения связывающих агентов (см., например, Lenssen, et al. (2002) Chembiochem. 3(9):852-8; Khersonsky, et al. (2003) Curr. Top.Med. Chem. 3(6):617-43; Anthony-Cahill & Magliery (2002) Curr. Pharm. Biotechnol. 3(4):299-315).
[0089] Связывающие агенты, описанные в настоящем документе, могут быть помечены. В некоторых вариантах осуществления связывающий агент представляет собой антитело, меченое путем ковалентного связывания антитела с прямой или косвенной меткой. Прямая метка может быть обнаружена как объект, который в своем естественном состоянии виден либо невооруженным глазом, либо с помощью оптического фильтра и/или приложенной стимуляции, например ультрафиолетового света, для стимулирования флуоресценции. Примеры цветных меток, которые можно использовать, включают частицы золя металлов, частицы золя золота, частицы золя красителя, частицы окрашенного латекса или красители, инкапсулированные в липосомы. Другие прямые метки включают радионуклиды и флуоресцентные или люминесцентные группы.
[0090] Косвенные метки, такие как ферменты, также могут быть использованы по изобретению. Известно использование в качестве меток различных ферментов, таких как, например, щелочная фосфатаза, пероксидаза хрена, лизоцим, глюкозо-6-фосфатдегидрогеназа, лактатдегидрогеназа и уреаза. Подробное обсуждение ферментов в иммуноанализах см. в Engvall (1980) Methods of Enzymology 70:419-439.
[0091] Описанные в настоящем документе белки (т.е. антитела или антигены/эпитопы) могут быть присоединены к поверхности. Примеры полезных поверхностей, на которые может быть прикреплен белок для диагностических целей, включают нитроцеллюлозу, PVDF, полистирол, нейлон или другой подходящий пластик. Поверхность или подложка также может быть пористой подложкой (см., например, US 7,939,342).
[0092] Кроме того, белки по изобретению могут быть присоединены к частице или микроносителю. Например, антитела к белкам филяриозной нематоды или сами белки филяриозной нематоды могут быть конъюгированы с суперпарамагнитными микрочастицами, например, как это используется в мультиплексных анализах на основе LUMINEX.
[0093] Белки филяриозных нематод по настоящему изобретению могут быть выделены и/или очищены или продуцированы синтетическим путем или с использованием технологии рекомбинантных нуклеиновых кислот.Очистка может быть частичной или существенной. Что касается белковых фрагментов филяриозной нематоды, термин «фрагмент» относится к белку, имеющему более короткую аминокислотную последовательность, чем у описанных в настоящем документе белков. Предпочтительно, такие фрагменты имеют длину, по меньшей мере, 5 последовательных аминокислот или до 35 аминокислот.В некоторых вариантах осуществления, фрагмент белка включает, по меньшей мере, один эпитоп.«Эпитоп» представляет собой характеристику молекулы, такую как первичная, вторичная и/или третичная пептидная структура и/или заряд, которая образует сайт, распознаваемый иммуноглобулином, Т-клеточным рецептором или молекулой HLA. Альтернативно, эпитоп можно определить как набор аминокислотных остатков, которые участвуют в распознавании определенным иммуноглобулином, или, в контексте Т-клеток, те остатки, которые необходимы для распознавания белками Т-клеточных рецепторов и/или рецепторами главного комплекса гистосовместимости (МНС).
[0094] В некоторых вариантах осуществления фрагмент белка по изобретению представляет собой фрагмент ALT2, содержащий или состоящий из эпитопа SEQ ID NO: 121, в частности, эпитопов SEQ ID NO: 78 или SEQ ID NO: 98. В других вариантах осуществления фрагмент белка по изобретению представляет собой фрагмент ALT2, содержащий или состоящий из эпитопа SEQ ID NO: 122, в частности, SEQ ID NO: 79 или SEQ ID NO: 99. В дополнительных вариантах осуществления фрагмент белка по изобретению представляет собой фрагмент HSP, содержащий или состоящий из эпитопа SEQ ID NO: 81 или SEQ ID NO: 123, в частности, SEQ ID NO: 80 или SEQ ID NO: 100. В некоторых вариантах осуществления фрагмент белка по изобретению представляет собой фрагмент TSP, содержащий или состоящий из эпитопа SEQ ID NO: 82. В других вариантах осуществления фрагмент белка по изобретению представляет собой фрагмент ТРХ2, содержащий или состоящий из эпитопа SEQ ID NO: 83 или SEQ ID NO: 124, в частности, SEQ ID NO: 84 или SEQ ID NO: 101.
[0095] Фрагменты по изобретению могут быть выделены, очищены или иным образом приготовлены/получены человеческими или не человеческими способами. Например, эпитопы могут быть получены путем выделения фрагмента белка филяриозной нематоды из бактериальной культуры, или их можно синтезировать в соответствии со стандартными протоколами в данной области техники. Синтетические эпитопы также могут быть получены из аминокислотных миметиков, таких как D-изомеры встречающихся в природе L-аминокислот или аминокислот неприродного происхождения, таких как циклогексилаланин.
[0096] В некоторых вариантах осуществления белок или фрагмент белка филяриозной нематоды конъюгируют или сливают с высокомолекулярным белковым носителем для облегчения продуцирования антител. В некоторых вариантах осуществления высокомолекулярный белок представляет собой бычий сывороточный альбумин, тиреоглобулин, овальбумин, фибриноген или гемоцианин лимфы улитки. Особенно предпочтительным носителем является гемоцианин лимфы улитки.
[0097] Можно использовать любой подходящий способ иммуноанализа, включая коммерчески доступные, для определения уровня, по меньшей мере, одного из специфических белков филяриозной нематоды, фрагментов белка или защитных/нейтрализующих антител по изобретению. Подробное обсуждение известных методов иммуноанализа в настоящем документе не требуется, поскольку они известны специалистам в данной области техники. Типовые подходящие методы иммуноанализа включают твердофазные иммуноферментные анализы (ELISA), радиоиммуноанализы (РИА), анализы конкурентного связывания, гомогенные анализы, гетерогенные анализы и т.д. Обзор различных известных методов иммуноанализа, например, содержится в Methods in Enzymology (1980) 70:30-70 и 166-198.
[0098] В некоторых вариантах осуществления способ или анализ иммуноанализа включает метод двойного антитела для измерения уровня белков или фрагментов белка филяриозной нематоды в биологическом образце. В соответствии с этим методом одно из антител является «захватывающим» антителом, и другое является «обнаруживающим» антителом. Захватывающее антитело иммобилизуют на твердой подложке, которая может быть любого из различных типов, известных в данной области техники, таких как, например, лунки титровального микропланшета, микроносители, пробирки и пористые материалы, такие как нейлон, стекловолокно и другие полимерные материалы. В этом способе твердую фазу составляет твердая подложка, например, лунки титровального микропланшета, покрытые захватывающим антителом, предпочтительно моноклональным, индуцированным против конкретного представляющего интерес белка. Биологический образец, который может быть разведен или нет, обычно, по меньшей мере 1, 2, 3, 4, 5, 10 или более стандартов и контролей, добавляют к отдельным твердым носителям и инкубируют.Когда представляющий интерес белок присутствует в образце, он захватывается иммобилизованным антителом, специфичным для рассматриваемого белка. После инкубации и промывки к твердой подложке добавляют обнаруживающее антитело, например, поликлональное антитело кролика против маркерного белка. Обнаруживающее антитело связывается с белком, связанным с захватывающим антителом, образуя сэндвич-структуру. После инкубации и промывания к твердой подложке добавляют анти-IgG антитело, например, поликлональное антитело козы против IgG кролика, меченное ферментом, таким как пероксидаза хрена (HRP). После инкубации и промывки к твердой подложке добавляют субстрат для фермента с последующей инкубацией и добавлением раствора кислоты для остановки ферментативной реакции.
[0099] Степень ферментативной активности иммобилизованного фермента определяют путем измерения оптической плотности окисленного ферментативного продукта на твердой подложке при соответствующей длине волны, например, 450 нм для HRP. Абсорбция при длине волны пропорциональна количеству представляющего интерес белка в образце. Набор стандартов маркерного белка используют для получения стандартной кривой зависимости абсорбции от концентрации белка филяриозной нематоды. Этот способ удобен, потому что результаты теста могут быть получены через 45-50 минут, и способ является и чувствительным в представляющем интерес диапазоне концентраций для каждого белка филяриозной нематоды, и обладает высокой специфичностью.
[0100] Стандарты могут быть положительными образцами, содержащими различные концентрации обнаруживаемого белка, чтобы гарантировать, что реагенты и условия работают должным образом для каждого анализа. Стандарты также обычно включают отрицательный контроль, например, для обнаружения примесей. В некоторых аспектах вариантов осуществления изобретения, положительные контроли могут быть титрованы до различных концентраций, включая необнаруживаемые количества и четко обнаруживаемые количества, и в некоторых аспектах, также включают образец, который показывает сигнал на пороговом уровне обнаружения в биологическом образце.
[0101] Способ по изобретению может быть осуществлен в различных форматах устройств для анализа, включая описанные в патентах US 6,426,050, US 5,910,287, US 6,229,603 и US 6,232,114, выданных Aurora Biosciences Corporation. Устройства для анализа, используемые в соответствии с изобретением, могут быть приспособлены для получения количественного или качественного (присутствует/отсутствует) результата. В некоторых вариантах осуществления, способ включает использование формата титровального микропланшета или микрожидкостного устройства. Анализы также можно проводить в автоматических анализаторах иммуноанализа, которые известны в данной области техники, и которые могут выполнять анализы на ряде различных образцов. Эти автоматические анализаторы включают типы непрерывного/произвольного доступа. Примеры таких систем описаны в US 5,207,987, US 5,518,688, US 6,448,089 и US 6,814,933. Различные автоматические анализаторы, которые коммерчески доступны, включают анализаторы OPUS(и OPUS MAGNUM®.
[0102] Другой формат анализа, который можно использовать в соответствии с изобретением, представляет собой ручной экспресс-тест, который можно проводить в месте оказания медицинской помощи в любом месте. Как правило, такие устройства для анализа в месте оказания медицинской помощи будут давать результат, который является либо «положительным», т.е. показывающим присутствие белка, либо «отрицательным», показывающим, что белок отсутствует. Как правило, контроль, показывающий, что реагенты работали, в целом входит в состав такой системы для места оказания медицинской помощи. Системы, анализы и устройства для места оказания медицинской помощи хорошо описаны для других целей, таких как обнаружение беременности (см., например, патент US 7,569,397 и US 7,959,875). Соответственно, изобретение также предлагает устройства, такие как тест-полоски для места оказания медицинской помощи и микрожидкостные устройства для проведения анализов in vitro по настоящему изобретению.
[0103] Следует также понимать, что устройства для анализа, используемые по изобретению, могут быть предназначены для проведения одного единственного анализа для конкретного белка или для проведения множества анализов, из одного объема жидкости организма, для соответствующего количества различных белков филяриозной нематоды или антител к ним. В некоторых вариантах осуществления, аналитическое устройство последнего типа представляет собой такое устройство, которое может обеспечить полуколичественный результат для белка филяриозной нематоды или антител, измеренных в соответствии с изобретением, т.е. одного или нескольких из ALT2, TSP, VAL-1, ТРХ2, GST и HSP или их антител. Эти устройства, как правило, адаптированы для предоставления отчетливой визуально обнаруживаемой цветной полосы в месте, где находится конкретный представляющий интерес белок, когда концентрация белка выше порогового уровня. Для дополнительного подробного обсуждения типов анализов, которые можно использовать по изобретению, а также различных форматов анализов и автоматических анализаторов см., например, US 5,747,274. Обнаружение белка филяриозной нематоды может быть дополнительно выполнено с использованием мультиплексных технологий.
[0104] В других вариантах осуществления анализы или иммуноанализы по изобретению включают микроносители, покрытые связывающим агентом против белка филяриозной нематоды или его фрагмента, или антитела. Обычно используются микроносители из полистирола, которые можно маркировать для установления уникальной идентичности. Обнаружение проводят с помощью проточной цитометрии. В данной области техники известны другие типы иммуноанализов на основе микроносителей, например, иммуноанализ с лазерными микроносителями и родственные анализы с магнитными микроносителями (см., например, Fritzler, et al. (2009) Expert Opinion on Medical Diagnostics 3:81 -89).
[0105] Способы по изобретению могут быть автоматизированы с использованием робототехники и систем, управляемых компьютером. Биологический образец может быть введен в систему, такую как микрожидкостное устройство, полностью управляемое роботизированной станцией от ввода образца до вывода результата. Стадия отображения результата также может быть автоматизирована и подключена к той же системе или к удаленной системе. Таким образом, анализ образца может быть выполнен в одном месте, и анализ результатов в другом месте, где единственным соединением является, например, подключение к Интернету, где анализ впоследствии отображается в формате, подходящем для чтения как медицинским работником, так и пациентом.
[0106] В некоторых вариантах осуществления присутствие любого одного или любой комбинации защитных/нейтрализующих антител, описанных в настоящем документе, идентифицирует субъекта как иммунизированного поливалентной иммуногенной композицией против филяриозной нематоды. Таким образом, в зависимости от титра антител, субъект может получать или не получать дополнительные стимулирующие вакцинации.
[0107] В некоторых вариантах осуществления присутствие любого одного или любой комбинации белков филяриозных нематод, описанных в настоящем документе, идентифицирует субъекта как инфицированного филяриозной нематодой. Таким образом, у субъекта диагностируют филяриоз, и, в некоторых вариантах осуществления настоящего изобретения, его лечат диэтилкарбамазином, мебендазолом, флубендазолом, альбендазолом, ивермектином или их комбинацией. В одном варианте осуществления диагноз может быть поставлен, если в образце субъекта обнаружено присутствие любого из белков филяриозной нематоды. В другом варианте осуществления лечение назначается или проводится, если, по меньшей мере, два белка филяриозных нематод положительно идентифицируются в биологическом образце.
[0108] Наборы, предлагаемые в соответствии с настоящим изобретением, включают один или несколько связывающих агентов, например, антител или фрагментов антител, или белки филяриозной нематоды, и, необязательно, устройство с твердой поверхностью. В некоторых вариантах осуществления твердая поверхность представляет собой микроноситель, предметное стекло, аналитический планшет (например, многолуночный планшет) или устройство, основанное на принципе растекания жидкости в радиальном направлении, с которым связываются связывающие агенты/белки. В некоторых вариантах осуществления набор дополнительно включает один или несколько стандартов или контролей.
[0109] В некоторых вариантах осуществления, изобретение предлагает матрицу на основе микропланшетов для мультиплексных иммуноанализов. В соответствии с некоторыми вариантами осуществления, каждая лунка может содержать одно антитело против, по меньшей мере, одного из перечисленных белков филяриозной нематоды. В других вариантах осуществления каждая лунка содержит матрицу антител против, по меньшей мере, двух или нескольких из перечисленных белков филяриозных нематод. В некоторых вариантах осуществления каждая лунка планшета содержит антитело к двум, трем, четырем или пяти из следующих белков: ALT2, TSP, VAL-1, ТРХ2, GST и HSP. В конкретных вариантах осуществления каждая лунка планшета содержит антитело к каждому из ALT2, TSP, VAL-1, ТРХ2, GST и HSP.
[0110] В других вариантах осуществления каждая лунка содержит матрицу, по меньшей мере, двух или нескольких белков филяриозных нематод по настоящему изобретению. В некоторых вариантах осуществления каждая лунка планшета содержит два, три, четыре или пять следующих белков: ALT2, TSP, VAL-1, ТРХ2, GST и HSP. В конкретных вариантах осуществления каждая лунка планшета содержит ALT2, TSP, VAL-1, ТРХ2, GST и HSP.
[0111] В других вариантах осуществления изобретение предлагает простые в использовании диагностические тест-полоски для мест оказания медицинской помощи, аналогичные полоскам для определения беременности, где полоска содержит, по меньшей мере, одно антитело против, по меньшей мере, одного из перечисленных белков филяриозной нематоды. В альтернативных вариантах осуществления изобретение предлагает простые в использовании диагностические тест-полоски для мест оказания медицинской помощи, где полоска содержит, по меньшей мере, один из указанных белков филяриозной нематоды.
[0112] Тест-полоска может включать положительный и отрицательный контроль, чтобы показать пользователю, что реагенты работают правильно, и/или что образец был правильно добавлен на полоску. Полоски могут поставляться с или без упаковки, с или без дополнительных реагентов. Диагностические тест-полоски для иммунохроматорафических анализов, такие как тест-полоски, описанные в настоящем документе, могут быть сконструированы, как описано в данной области техники, см., например, US 2010/0196200; US 2010/0129935; US 2009/0253119; и US 2009/0111171. Подходящие материалы для тест-полосок включают, но не ограничены ими, материалы, полученные из целлюлозы, такие как фильтровальная бумага, хроматографическая бумага, нитроцеллюлоза и ацетат целлюлозы, а также материалы, изготовленные из стекловолокна, нейлона, дакрона, ПВХ, полиакриламида, поперечно-связанного декстрана, агарозы, полиакрилата, керамических материалов и подобных. Материал или материалы тест-полоски необязательно могут быть обработаны для изменения их характеристик капиллярного потока или характеристик нанесенного образца. Например, область нанесения образца тест-полоски может быть обработана буферами для корректировки рН или удельного веса нанесенного образца, чтобы обеспечить оптимальные условия тестирования.
[0113] Изобретение более подробно описано с помощью следующих неограничивающих примеров.
Пример 1: Вакцина против малого белка теплового шока
[0114] Паразиты. В. malayi L3 получают из Ресурсного центра исследований реагентов против филяриоза NIAID/NIH (FR3) в Университете Джорджии, Athens, GA.
[0115] Образцы сыворотки человека. Примерно 10 мл образцов крови собирают у следующих клинических групп субъектов (1) эндемичные нормальные (EN) субъекты, которые представляют собой индивидуумов, бессимптомных или с отсутствием микрофиляриемии; (2) бессимптомные субъекты с микрофиляриемией (Mf), которые имеют циркулирующие микрофилярии в крови и которые идентифицированы при микроскопическом исследовании их ночных мазков крови; (3) пациенты с хронической патологией (CP) включают субъектов, у которых наблюдается лимфатический отек и другие хронические клинические симптомы филяриоза, и (4) неэндемичные нормальные субъекты (NEN), которые живут в не эндемичных областях и не имеют циркулирующих паразитов или антител и не демонстрируют признаки какого-либо филяриоза. Из этих образцов крови отделяют сыворотки и хранят при -80°С до использования.
[0116] Экспрессия и очистка рекомбинантного белка теплового шока В. malayi. Для получения рекомбинантного малого белка теплового шока В. malayi 12.6 (rBmHSP) полноразмерную последовательность гена клонируют в pRSET-A (с N-концевой гексагистидиновой меткой) и трансформируют в BL21(DE3), содержащий pLysS (Invitrogen, Carlsbad, СА), чтобы свести к минимуму токсичность из-за белка. Когда абсорбция культур достигает значения OD 0,6, к культурам добавляют 1 мМ IPTG (изопропилтио^-галактопиранозида) и инкубируют в течение дополнительных 3 часов для индукции экспрессии генов. После лизиса клеток общие белки разделяют в 15% SDS-PAGE для подтверждения экспрессии his-меченного белка. Затем меченный гистидином рекомбинантный белок очищают с использованием аффинной колоночной хроматографии с иммобилизованным металлическим кобальтом (Clontech, Mountain View, CA) в соответствии с рекомендациями производителя. Затем рекомбинантный белок разделяют в 15% SDS-PAGE и окрашивают бриллиантовым синим COOMASSIE R250. Единственную полосу получают после очистки колонки.
[0117] Трехмерная модель BmHSP, Трехмерную модель белка BmHSP строят путем моделирования гомологии. Поиски гомологии последовательности BLAST проводят для идентификации матричных белков в базе данных PDB. Альфа-кристаллин А человека, недавно кристаллизованный белок, показывает значительную идентичность последовательности, и поэтому его выбирают в качестве матрицы для моделирования BmHSP. Построение модели выполняют с использованием MODELLER 9v6 (Sali & Blundell (1993) J. Mol. Biol. 234:779-815). Полученную 3-D структуру затем подтверждают с использованием программы PROCHECK (Laskowski, et al. (1993) J, Appl, Cryst. 26:283-29). Лучшая модель, спрогнозированная PROCHECK, имеет оценку -0,46, и ее выбирают для дальнейшего моделирования и создания 3-D структуры с использованием программы Rasmol.
[0118] Анализ структуры BmHSP. Вторичную структуру и сайт взаимодействия белок-белок BmHSP прогнозируют в PDBsum и на сервере электронной почты Predict Protein в Европейской лаборатории молекулярной биологии в Гейдельберге (Roos, et al. (1995) Parasitol, Today 11:148-150). Сканирование мотивов проводят с помощью анализа паттернов PROSITE для идентификации функциональных мотивов в BmHSP. Эпитопы В-клеток, Т-клеток и CTL в последовательностях BmHSP прогнозируют с использованием базы данных по иммунным эпитопам и ресурса анализа (IEDB).
[0119] Филогенетический анализ BmHSP. Аминокислотные последовательности BmHSP сравнивают с членами других семейств малых белков теплового шока из разных организмов. Анализируют следующие последовательности. Номера доступа указаны в скобках. Aconthocheilonema vitae (САА48631); Archaeoglobus fulgidus (028308); Artibeus jamaicensis (P02482); Aspergillus fumigatus (Q4WV00); Arabidopsis thaliana (081822); Artemia persimilis (DQ310578); Azotobacter vinelandii (P96193); Brugia pahangi (CAA61152), Brugia malayi (AAU04396); Buchnera aphidicola (P57640); Bombyx mori (AF315318_1); Bradyrhizobium japonicum (P70918); Caenorhabditis elegans (Q7JP52); Coccidioides immitis (Q1E6R4); Carica papaya (Q69BI7); Caenorhabditis remanei (AAZ42349); Dictyostelium discoideum (Q54I91); Escherichia coli (ibpA; P0C054); Escherichia coli (ibpB; P0C058); Homo sapiens (P02489); Haemonchus contortus (AAN05752); Lygodactylus picturatus (Q6EWI0); Onchocercara volvulus (CAA48633), Ostertagia ostergi (CAG25499); Macaca muiatta (P02488); Mycobacterium tuberculosis (P0A5B7); Mus musculus (AAA37861); Nippostrongylus brasiliensis (BAI81970); Plasmodium falciparum (Q8IB02); Rattus (CAA42910); Saccharomyces cerevisiae (P15992); Solanum lycopersicum (082545); Streptococcus thermophilus (P80485); Trichinella spiralis (ABJ55914); Trypanosoma brucei (Q57V53); Toxoplasma gondii (Q6DUA8). Домен альфа-кристалл и на из всех последовательностей sHSP выравнивают с использованием алгоритма ClustalW, и набор данных используют для построения филогенетического дерева с помощью программного обеспечения PHYLIP. Деревья создают с использованием метода объединения соседних пар с расстоянием между аминокислотами с поправкой Пуассона.
[0120] Анализ белков теплового шока. Одной из типовых характеристик белков теплового шока является то, что они могут связываться с клеточными белками и защищать их от теплового повреждения. Когда белки подвергаются тепловому повреждению, они агрегируют (тепловая агрегация). Белки теплового шока предотвращают эту агрегацию. Чтобы определить, может ли BmHSP предотвращать тепловую агрегацию, используют анализ тепловой агрегации цитрулинсинтазы (CS) (Sigma, St. Louis, МО). CS выбирают потому, что этот белок очень чувствителен к тепловой денатурации. Используют утвержденный способ (Gnanasekar, et al. (2009) Biochem. Biophys. Res, Commun, 386:333-337). Коротко, 1 мкМ CS подвергают воздействию 45°C в присутствии или в отсутствие BmHSP (2 мкМ), суспендированного в 50 мМ буфера на основе фосфата натрия с рН 7,4, содержащего 100 мМ NaCl. В качестве контроля используют BSA. CS инкубируют с BmHSP в молярном соотношении (1:2) в течение различных временных интервалов от 0 до 40 минут.Тепловую денатурацию (агрегацию) отслеживают спектрофотометрически при 360 нм.
[0121] In vitro анализ связывания пептидов на активность белков теплового шока. Другой характеристикой белков теплового шока является то, что они могут связываться с различными белками. Чтобы определить, обладает ли BmHSP также этой функцией, CS и другой белок, люциферазу, химически денатурируют 6М гидрохлоридом гуанидина в соответствии с известными способами (Gnanasekar, et al. (2009) выше). Затем нативные и химически денатурированные белки наносят на 96-луночные планшеты в течение ночи при 4°С. После промывания PBS, лунки блокируют 3% ВБАпри комнатной температуре. После дополнительной промывки, лунки инкубируют с his-меченным rBmHSP в течение 1 часа при 37°С. После повторной промывки PBS, добавляют оптимально разведенный конъюгат анти-Мэ-меченного HRP и инкубируют при 37°С в течение 1 часа. После окончательной промывки цвет проявляют с помощью ABTS [2,2'-азинобис(3-этилбензотиазолин-6-сульфоновой кислоты)], и измеряют OD при 405 нм.
[0122] Уровни анти-BmHSP антитела е сыворотке человека, В общей сложности 20 образцов сыворотки, принадлежащих к различным клиническим группам, таким как Mf, CP, EN и NEN, анализируют на наличие и титр анти-BmHSP IgG антител с использованием косвенного ELISA (Cheirmaraj, et al. (1992) J, Trop.Med. Hyg. 95:47-51). Коротко, лунки 96-луночного титровального микропланшета покрывают rBmHSP (1 мкг/мл) в карбонатном буфере, рН 9,6, в течение ночи при 4°С и блокируют 3% BSA в течение 1 часа при 37°С. В лунки добавляют образцы сыворотки, и планшеты инкубируют в течение ночи при 4°С. После промывания лунок, добавляют HRP-меченное антитело мыши против IgG человека (1:5000) и инкубируют еще в течение 1 часа при 37°С. Цвет проявляют с использованием субстрата ABTS. Абсорбцию измеряют при 405 нм в микропланшетном ридере (BIO-RAD, Hercules, СА). Изотип анти-BmHSP IgG антител в сыворотке субъектов также определяют с помощью изотип-специфического ELISA. Биотинилированные моноклональные антитела мыши против IgG1, IgG2, IgG3 и IgG4 человека используют в качестве вторичных антител, и цвет проявляют конъюгатом авидин-HRP (Sigma, St. Louis, МО) в качестве вторичных антител.
[0123] Клонирование кодон-оптимизированного BmHSP в вектор pVAX для ДНК вакцины. Кодон-оптимизированные гены Bmhsp клонируют в эукариотический вектор экспрессии pVAX (Invitrogen) с использованием вставка-специфичных праймеров (прямой праймер, 5'-CGC GGA ТСС ATG GAA GAG AAG GTG-3((SEQ ID NO: 1), содержащий сайт BamHI, и обратный праймер, 5'-CCG GAA ТТС ТСА СТТ GTC GTT GGT G-3((SEQ ID NO: 2), содержащий сайт Eco RI). Параметры ПЦР следующие: 94°С денатурация 30 секунд, 50°С отжиг праймеров в течение 30 секунд, 72°С удлинение праймера в течение 30 секунд за 30 циклов; и окончательное удлинение в течение 5 минут проводят при 72°С. Вставку ДНК секвенируют для обеспечения аутентичности клонированной нуклеотидной последовательности на обеих цепях. Плазмиды сохраняют и размножают в клетках Е. coli TOP10F'. Затем плазмиды очищают с использованием набора для экстракции плазмид без эндотоксина (Qiagen, Hilden, Germany). ДНК анализируют с помощью электрофореза в агарозном геле и количественно определяют с помощью спектрофотометрии (OD 260/280, соотношение>1,8).
[0124] Иммунизация мышей. В этих экспериментах используют шестинедельных самцов мышей Balb/c, приобретенных в Charles River Laboratories. Гуманное использование животных в этом исследовании и протокол одобрены комитетом IACUC Медицинского колледжа Университета Иллинойса в Рокфорде. Каждая группа состоит из пяти (5) мышей, и всех мышей иммунизируют внутрибрюшинно с использованием трех различных схем иммунизации. Мышей группы А иммунизируют по схеме примирования/стимулирования. Мышей примируют дважды с двухнедельными интервалами 100 мкг свободной от эндотоксина кодон-оптимизированной pVAX Bmhsp ДНК, суспендированной в объеме 50 мкл. После примирования все мыши получают две стимулирующие дозы по 15 мкг белка rBmHSP (по 50 мкл каждая), суспендированного в квасцах, с интервалом в две недели. Мышей группы В иммунизируют только белком rBmHSP. Эти мыши получают четыре дозы по 15 мкг белка rBmHSP, суспендированного в квасцах, с двухнедельными интервалами. Мышей группы С иммунизируют только ДНК. Эти мыши получают четыре дозы по 100 мкг pVAX Bmhsp ДНК, вводимых с двухнедельными интервалами. Животные группы D получают 100 мкг контрольного вектора pVAX и адъюванта с тем же интервалом и остаются в качестве отрицательных контролей. Образцы крови собирают у каждой мыши перед иммунизацией и через месяц после последней стимулирующей дозы. После отделения сыворотки определяют титр циркулирующих анти-BmHSP IgG антител и соответствующие изотипы. Сыворотки, показавшие высокий титр анти-BmHSP антител, используют в описанном в настоящем документе анализе антителозависимой клеточной цитотоксичности (ADCC).
[0125] Уровни анти-BmHSP антител в сыворотке мышей. Уровни анти-BmHSP IgG антител в сыворотке иммунизированных и контрольных групп мышей определяют с помощью косвенного ELISA (Veerapathran, et al. (2009) PLoS Negl. Trop.Dis. 3:e457). Уровни анти-BmHSP IgG1, IgG2a, IgG2b и IgG3 антител также определяют с использованием набора ELISA (ThermoFisher Scientific, Rockford, IL) для изотипирования антител мыши. Цвет проявляют с помощью хромогенного субстрата ABTS (2,2'-азинобис(3-этилбензотиазолин-6-сульфоновой кислоты) и измеряют оптическую плотность при 405 нм в ридере ELISA (BIO-RAD).
[0126] Истощение анти-BmHSP антител из сыворотки человека и мыши. Объединенные сыворотки субъектов EN и иммунизированных мышей обедняют анти-BmHSP антителами путем инкубации объединенных сывороток с кобальтовой смолой IMAC, сопряженной с his-меченным rBmHSP в соответствии с утвержденными способами (Veerapathran, et al. (2009) выше). Коротко, 1 мг his-меченного rBmHSP связывают с 2 мл объемным слоем смолы IMAC в течение 2 часов при 37°С. После однократной промывки смолы 10 мл PBS (рН 8), добавляют 200 мкл объединенной сыворотки и инкубируют в течение ночи при 4°С. После инкубации, смесь смол центрифугируют в течение 2 минут при 750 об/мин и собирают супернатант. Истощение анти-BmHSP антител в супернатанте подтверждают с помощью ELISA, как описано в настоящем документе.
[0127] Объединенные сыворотки субъектов EN и объединенные сыворотки иммунизированных мышей обедняют анти-BmHSP IgG1, анти-BmHSP IgG2a, анти-BmHSP IgG2b, анти-BmHSP IgG3 и анти-BmHSP IgG4 антителами с использованием смолы NHS (N-гидроксисукцинимидил) (Thermo fisher scientific). Коротко, 1 мкг соответствующих моноклональных антител связывают с колонкой со смолой NHS. После двукратной промывки смолы PBS (рН 8) через колонку пропускают 100 мкл сыворотки. Поток собирают в виде обедненной антителами сыворотки. Истощение специфического изотипа антитела подтверждают с помощью изотип-специфического ELISA, как описано в настоящем документе. После трехкратной промывки колонки PBS (рН 7,4) связанные антитела элюируют из смолы буфером Глицин-HCl (рН 2,7), и рН доводят до 7,4 с 1 М Tris буфера (рН 8). Восстановленный элюат содержит специфическое антитело, что снова подтверждают с помощью ELISA. Сыворотку, обедненную антителами, также восстанавливают с элюированными антителами. Аликвоту обедненной сыворотки восстанавливают элюированными антителами до ее исходной концентрации с использованием значений, определенных в более раннем ELISA на чистых образцах сыворотки. Сыворотки, обедненные антителами, элюированные антитела и восстановленные образцы сыворотки затем используют в анализе ADCC.
[0128] Анализ антителозависимой клеточной цитотоксичности (ADCC). Анализ ADCC in vitro проводят в соответствии с известными способами (Chandrasekhar, et al. (1985) Parasite Immunol. 7:633-641). Коротко, десять (10) L3 В. malayi инкубируют с 2×105 клетками брюшины (РЕС), собранными от нормальных мышей, 50 мкл объединенных образцов сыворотки мыши и 50 мкл среды RPM11640 в 96-луночном культуральном планшете (Thermo Fisher Scientific). Через 48 часов инкубации при 37°С и 5% СО2 жизнеспособность личинок определяют при 400Х увеличении с использованием светового микроскопа. Личинки, которые являются прозрачными и поврежденными, считаются мертвыми. Кроме того, мертвые личинки также имеют прилипшие к ним скопления клеток и являются более прозрачными, чем живые. Личинки, которые являются активными, свернутыми в кольцо и полупрозрачными, считаются живыми. ADCC оценивают как долю гибели личинок, рассчитываемую по формуле:
Количество мертвых личинок (Общее количество личинок (100.
[0129] Анализ ADCC также проводят с объединенными образцами сыворотки человека, как описано в настоящем документе, за исключением того, что образцы сыворотки человека инкубируют с 2×105 РВМС, собранными у здоровых субъектов, и 6-12 В. malayi L3 в течение 48 часов при 37°С и 5% СО2. Жизнеспособность и гибель личинок определяют, как описано выше.
[0130] Исследования защиты на мышах. Вакцинный потенциал BmHSP оценивают на модели контрольного заражения на мышах. Мышей иммунизируют, как описано выше, с использованием примирования/стимулирования, только ДНК или только белка. Группы вектора и квасцов служат в качестве отрицательных контролей. Иммунизированных и контрольных животных заражают с использованием способа камеры с микропорами, известного в данной области техники (Abraham, et al. (1986) Immunology 57:165-169). Коротко, камеры с микропорами собирают с использованием 14×2 мм (акриловых) колец PLEXIGLASS (Millipore Corporations, Bedford, MA) и 5,0 мкм поликарбонатных мембран NUCLEOPORE (Millipore Corporations). Мембраны прикрепляют к кольцам PLEXIGLASS с помощью цианакрилового адгезива и стоматологического цемента. Камеры погружают на ночь при 37°С в стерильную среду RPMI, содержащую гентамицин и антимикотический раствор. Перед экспериментами с заражением, 20 живых инфекционных L3, суспендированных в среде RPMI 1640 с добавлением 15% инактивированной теплом фетальной телячьей сыворотки (FCS), вводят в камеры с микропорами, и отверстие герметично закрывают стоматологическим цементом. Затем камеру с микропорами, содержащую L3, хирургическим путем имплантируют в брюшную полость каждой мыши под анестезией. При хирургических вмешательствах соблюдаются асептические условия. Через 48 часов после имплантации, животных умерщвляют и извлекают камеры из брюшной полости. Содержимое каждой камеры опорожняют, и личинок исследуют под микроскопом на прилипание клеток и гибель личинок. Мертвых и живых личинок идентифицируют, как описано выше в ADCC. Долю защиты показывают как количество мертвых паразитов общее количество извлеченных паразитов х 100.
[0131] Анализ пролиферации спленоцитов. У всех мышей из вышеуказанного эксперимента собирают селезенки и готовят одноклеточную суспензию клеток селезенки. Приблизительно 2×105 клеток/лунку, суспендированных в полной среде RPMI 1640 с добавлением 10% инактивированной теплом FCS, инкубируют при 37°С и 5% СО2 в течение 72 часов с rBmHSP (1 мкг/мл), СопА (1 мкг/мл) или только со средой. После инкубации определяют пролиферацию клеток с использованием набора для подсчета клеток (CCK-8), приобретенного у Dojindo Molecular Technologies, Inc. (Gaithersburg, MD). Индекс стимуляции пролиферации клеток селезенки рассчитывают по формуле: Абсорбция стимулированных клетокАбсорбция не стимулированных клеток.
[0132] Цитокиновый анализ. Клетки селезенки иммунизированных и контрольных мышей культивируют при 37°С и 5% СО2 в течение 72 часов с rBmHSP (1 мкг/мл), СопА (1 мкг/мл) или только со средой, как описано выше. Через 72 часа культуральные супернатанты и осадок клеток собирают отдельно для анализа цитокинов. Для измерения мРНК цитокинов осадок клеток суспендируют в реагенте TRIZOL (фенол, гуанидиний и тиоцианат) (GIBCO-BRL, Life technologies, Carlsbad, СА) и экстрагируют общую РНК в соответствии с инструкциями производителя. После промывки этанолом осадок РНК растворяют в воде, не содержащей РНКазу (Sigma) и обрабатывают ДНКазой I перед определением общей концентрации РНК с использованием спектрофотометра Бекмана при 260 нм. Обратную транскрипцию общей РНК выполняют с использованием набора для синтеза первой цепи кДНК (SABiosciences, Frederick, MD) в соответствии с рекомендациями производителя. Относительную количественную оценку экспрессии представляющих интерес генов измеряют на приборе Applied BioSystems 7300 для ПЦР в реальном времени (Applied BioSystems, Foster City, СА). ПЦР амплификации проводят с использованием смеси LIGHTCYCLER-DNA SYBR Green (цианиновый краситель) (SAbiosciences). Реакцию проводят с использованием следующих условий ПЦР: стадия активации 15 минут при 95°С в течение одного цикла, стадия денатурации 15 секунд при 95°С, отжиг праймеров в течение 20 секунд при 50°С и стадия удлинения в течение 15 секунд при 72°С. ДНК амплифицируют в течение 50 циклов. Отслеживают флуоресцентный ДНК-связывающий краситель SYBR Green (цианиновый краситель). Набор массивов данных РВ-ПЦР создают и анализируют с использованием веб-системы анализа данных SABiosciences.
[0133] Затем культуральные супернатанты собирают из культур спленоцитов через 72 часа после инкубации с rBmHSP (1 мкг/мл), ConA (1 мкг/мл) или только со средой. Уровни секретируемых белков IL-2, IL-4, IFN-(и IL-10 в культуральных супернатантах определяют с использованием набора для сэндвич-ELISA, приобретенного у ThermoFisher Scientific. Концентрацию каждого цитокина определяют по стандартной кривой, построенной с использованием рекомбинантного IL-2, IL-4, IFN-(или IL-10 мыши.
[0134] Статистический анализ. Статистический анализ проводят с использованием программного обеспечения XL STAT v.7.5.2 (Kovach Computing Services, Anglesey, UK). Статистическую значимость между сопоставимыми группами оценивают с помощью соответствующих непараметрических тестов с уровнем значимости, установленным на уровне р<0,05.
[0135] Экспрессия рекомбинантного BmHSP12.6 (Вт HSP). BmHSP клонируют в векторе pRSET А и экспрессируют в виде меченного гистидином (his-меченного) слитого белка в Е. coli BL21 (DE3)PLysS. Затем рекомбинантный белок BmHSP очищают с использованием колонки IMAC. Было обнаружено, что молекулярная масса очищенного рекомбинантного слитого белка с his-меткой составляет примерно 18 кДа. Очищенный на колонке рекомбинантный белок проявляется в виде одной полосы в SDS-PAGE.
[0136] Спрогнозированная трехмерная структура BmHSP. Аминокислотные последовательности альфа-кристаллической А-цепи человека на 42% сходны с BmHSP. Поскольку кристаллическая структура альфа-кристаллической А-цепи человека уже доступна, ее используют в качестве шаблона для моделирования предполагаемой структуры BmHSP с использованием программы Modeller 9v6. Анализ PROCHECK используют для выбора наилучшей модели, которая показывает оценку -0,41 по сравнению с шаблоном (Laskowski, et al. (1993) J. Appl. Cryst. 26:283-29). Анализ графика Рамачандрана также выполняют для последовательности BmHSP. Эти анализы показывают, что 92% остатков находятся в наиболее благоприятной области без пространственных затруднений. В дополнительной разрешенной области обнаруживают примерно 6,7% остатков. Модели, которые показали более 90% остатков в наиболее предпочтительных областях, прогнозируют как наиболее идеальные трехмерные модели, как спрогнозировано графиком Рамачандрана (Balazs, et al. (2001) Protein Eng. Des, Sci, 14:875-880). Анализ прогнозирования вторичной структуры также выполняют для белка BmHSP с использованием сервера PDBsum в EMBL. Этот анализ показал, что каждый альфа-кристаллический домен мономера BmHSP имеет ядро иммуноглобулина, состоящее из семи β-цепей, расположенных в двух антипараллельных листах. Прогнозирование вторичной структуры BmHSP показало два листа, четыре бета шпильки, одну бета выпуклость, семь нитей, две спирали, семь бета витков и один гамма виток в структуре BmHSP.
[0137] Предыдущие исследования показали, что BmHSP связывается с а цепью рецептора I IL-10 человека (Gnanasekar, et al. (2008) Mol, Biochem, Parasit, 159:98-103). Для идентификации сайта связывания рецептора IL-10 на BmHSP проводят прогностический анализ взаимодействий белок-белок (Ofran & Rost (2007) Bioinformatics 23:е13-е16). Результаты прогностического анализа показали, что N-концевой фрагмент BmHSP (аминокислоты от Met1 до Asn26) имеет область сильного взаимодействия белок-белок. Дальнейший анализ последовательности этой области показал, что аминокислотные последовательности от Val5 до Glu42 имеют значительную идентичность последовательности с областью связывания IL-10R человека IL-10 человека. Эти открытия подтверждают, что N-концевая область BmHSP может быть вовлечена в связывание BmHSP с а цепью рецептора I IL-10 человека.
[0138] Мотив и филогенетический анализ BmHSP, Анализ мотивов, проведенный в PROSITE, показал несколько предполагаемых сайтов посттрансляционной модификации, таких как сайты N-гликозилирования (остатки с 11 по 14 и с 98 по 101), сайты фосфорилирования протеинкиназы-с (остатки с 83 по 85 и со 100 по 102), сайты фосфорилирования казеинкиназы II (остатки с 68 по 71 и с 88 по 91) и сайты N-миристилирования (остатки с 40 по 45) на BmHSP. Подобные мотивы также наблюдают в IL-10 человека, что дополнительно указывает на то, что BmHSP может имитировать функцию IL-10 человека (Gnansekar, et al. (2008) выше). Картирование эпитопов на BmHSP выявило наличие областей эпитопов В-клеток, Т-клеток и CTL, что указывает на то, что BmHSP потенциально является высокоиммуногенным белком (таблица 2).
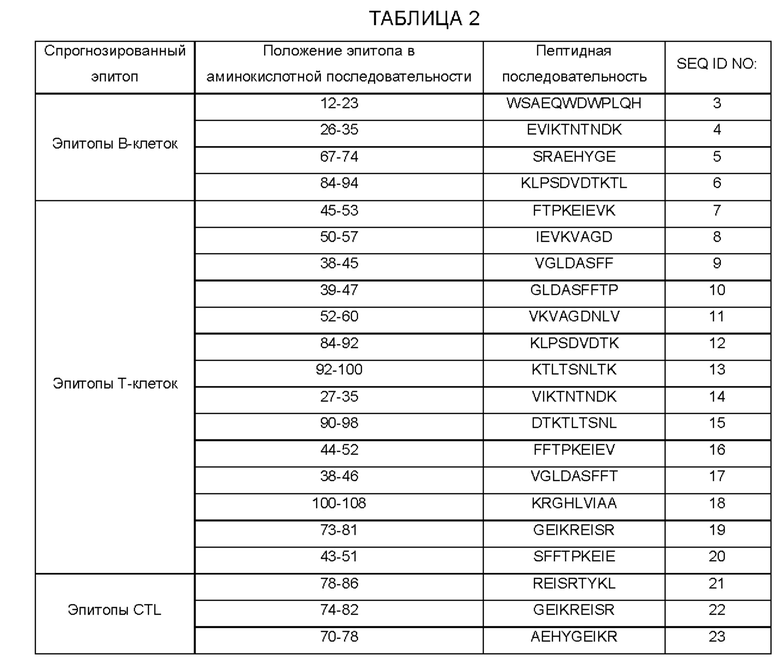
[0139] Филогенетический анализ, проведенный с использованием типовых последовательностей sHSP из разных групп организмов, показал, что BmHSP, HSP С.elegans и HSP С.remani образуют монофилетическую группу, отделенную от других групп организмов.
[0140] BmHSP является белком теплового шока. Большинство белков теплового шока, о которых сообщалось на сегодняшний день, обладают функцией шаперона. Чтобы определить, обладает ли BmHSP аналогичной функцией шаперона проводят реакцию тепловой агрегации с использованием модельного субстрата, цитруллинсинтазы (CS). Инкубация CS при 42°С приводит к разворачиванию белка и последующей агрегации в течение 10 минут. Добавление BmHSP к белку CS (в молярном соотношении 1:2) перед термической обработкой значительно (Р<0,01) ингибирует термическую агрегацию белка CS. Контрольный белок, не являющийся белком теплового шока, BSA, не влияет на индуцированную теплом агрегацию белка CS.
[0141] Другая функция белков теплового шока заключается в том, что они могут специфически связываться с денатурированными белками. Чтобы определить, может ли BmHSP специфически связываться с денатурированными белками, rBmHSP инкубируют с субстратами нативной и денатурированной CS или нативной и денатурированной люциферазы. Эти исследования показывают, что rBmHSP предпочтительно связывается с субстратами денатурированных белков по сравнению с нативным или контрольным белком. Таким образом, эти данные подтверждают, что BmHSP может действовать как молекулярный белок теплового шока, потенциально защищая клеточные белки паразита от повреждающего действия хозяина.
[0142] Реакция антитела у человека. Представленные в настоящем документе результаты показывают, что BmHSP имеет несколько Т-клеточных и В-клеточных эпитопов. Поэтому оценивают, имеют ли инфицированные филяриозом индивидуумы антитела к BmHSP. Соответственно, измеряют титр анти-BmHSP IgG-антител в сыворотке субъектов с EN, CP, Mf и NEN. Результаты показали, что субъекты EN имеют самые высокие уровни анти-BmHSP антител (р<0,001). Последующий изотипический анализ IgG антител показывает, что по сравнению с инфицированными группами (Mf и CP) индивидуумов, сыворотки от субъектов EN имеют высокие титры анти-BmHSP IgG1 и IgG3 антител. Носители Mf имеют только значительные уровни анти-BmHSP IgG2 антител в своей сыворотке. Точно так же, индивидуумы CP имеют только значительные уровни анти-BmHSP IgG4 антител в своей сыворотке. Уровни анти-BmHSP IgG1 и IgG3 являются очень низкими в сыворотке этих индивидуумов Mf и СР. Анти-BmHSP антитела не обнаруживаются в сыворотке пациентов NEN.
[0143] Результаты анализа ADCC. Поскольку антитела к BmHSP присутствуют во всех инфицированных группах индивидуумов (Mf и CP) и у субъектов EN, определяют, являются ли эти антитела функциональными. Используя анализ антителозависимой клеточной цитотоксичности, проверяют, обладают ли анти-BmHSPI 2.6 IgG антитела какой-либо защитной функцией против В, malayi. Эти исследования показывают, что объединенные сыворотки EN способствуют прикреплению РВМС к L3 и вызывают значительную (77,37%) гибель L3 В, malayi in vitro (таблица 3), тогда как объединенные сыворотки от Mf и CP не участвуют в функции ADCC. Эти данные показывают, что сыворотки EN обладают антипаразитарной активностью. Чтобы определить, связана ли эта функция с антителами, проводят исследования обеднения антителами. Обеднение сыворотки EN анти-BmHSP антителами приводит к значительному снижению (21,42%) гибели личинок (таблица 3), подтверждая, что анти-BmHSP антитела в сыворотке субъектов EN, но не субъектов Mf или CP, участвуют в уничтожении личинок.
[0144] Дальнейшие исследования обеднения показывают, что противопаразитарное действие анти-BmHSP антител связано с изотипом IgG1 антител. Истощение IgG1 антител из сыворотки EN значительно (40%) ингибирует функцию ADCC (таблица 4). Восстановление сыворотки EN, обедненной анти-BmHSP антителами, с элюированными анти-BmHSP IgG1 антителами восстанавливает функцию ADCC (таблица 3). Таким образом, эти данные показывают, что анти-BmHSP IgG1 антитела имеют решающее значение для функции ADCC.
[0145] Ответы антител у мышей. У мышей, иммунизированных rBmHSP, вырабатываются значительные уровни анти-BmHSP IgG антител. В частности, схема примирования/стимулирования индуцирует значительно более высокий титр IgG антител по сравнению с группой только ДНК вакцины (р<0,05). Однако белковая вакцина rBmHSP индуцирует самый высокий титр IgG антител. Анализ изотипа анти-BmHSP IgG антител показывает, что в сыворотке вакцинированных животных присутствуют преимущественно анти-BmHSP IgG1, IgG2a и IgG2b антитела. Анализ ADCC также проводят с сывороткой мышей. Эти исследования показали, что сыворотка мышей, вакцинированных BmHSP, способствует прилипанию клеток перитонеального экссудата к L3 и участвует в функции ADCC (83,02% уничтожения личинок) по сравнению с контрольной сывороткой (13%) (р<0,002) (таблица 5).
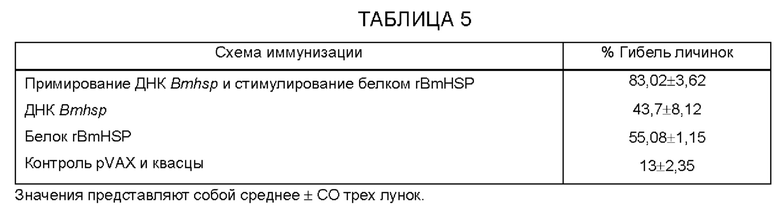
[0146] Подобно сывороткам человека, сыворотки вакцинированных мышей обедняют отдельными изотипами IgG антител для определения изотипа анти-BmHSP антител, которые участвуют в функции ADCC. Результаты этих исследований показывают, что, аналогично тому, что наблюдается с сыворотками EN, анти-BmHSP IgG1 антитела также вовлечены в ADCC-опосредованное уничтожение L3y мышей (таблица 6).

[0147] Вакцинный потенциал BmHSP у мышей. Вакцинный потенциал BmHSP оценивают на мышах Balb/c с использованием способа камеры с микропорами. Результаты показывают, что мыши, иммунизированные с использованием схемы примирования/стимулирования и белковой вакцины BmHSP, демонстрируют почти 72% и 58% смертности, соответственно, L3, имплантированных в брюшную полость иммунизированных мышей (таблица 7). В то время как камеры, имплантированные в контрольные группы животных, показывают только 7% гибели паразита, разница между защитой контрольной группы мышей и вакцинированных мышей является достоверной (Р<0,001). С другой стороны, мыши, иммунизированные только ДНК вакциной, индуцируют защиту только на 31%. Таким образом, схема примирования/стимулирования оказалась высокоэффективной в предоставлении индуцированной вакциной защиты от контрольного заражения по сравнению с протоколами иммунизации только ДНК или только белком.

[0148] Иммунный ответ у мышей, вакцинированных BmHSP, Для определения клеточного иммунного ответа на BmHSP у вакцинированных мышей, клетки селезенки, собранные у вакцинированных и контрольных мышей, культивируют в присутствии белка rBmHSP и оценивают их пролиферативный ответ и цитокиновый профиль. Пролиферативный ответ клеток селезенки у животных, иммунизированных по схеме примирования/стимулирования, значительно (Р>0,05) выше (индекс стимуляции 3,35±0,176) по сравнению с группой, вакцинированной только белком rBmHSP (индекс стимуляции 2,22±0,018) или группой, вакцинированной только ДНК Bmftsp (индекс стимуляции 3,53±0,102). Клетки селезенки контрольной группы животных не пролиферируют в ответ на rBmHSP (индекс стимуляции 0,98±0,013) и аналогичны контрольным клеткам, содержащим только среду. Поскольку клетки селезенки вакцинированных животных значительно пролиферируют, чтобы вспомнить ответ на rBmHSP, измеряют уровни цитокинов в культура л ьных супернатантах. Эти результаты показывают, что IFN-(является преобладающим цитокином, секретируемым клетками селезенки вакцинированных животных через 72 часа после стимуляции rBmHSP. ПЦР в реальном времени матрицы генов цитокинов проводят мРНК, собранной из клеток селезенки, стимулированных rBmHSP. Эти результаты показывают, что гены цитокинов Th1 (IFN-γ, CD-28, IL-12, IL-2) и Th2 (IL-4, IL-5, IL-1 R) значительно увеличены у вакцинированных животных.
Пример 2: rBmALT2+rBmHSP поливалентная иммуногенная композиция
[0149] Паразит.Brugia malayi L3 получают из Ресурсного центра исследований реагентов против филяриоза NIAID/NIH (FR3) в Университете Джорджии, Athens, GA.
[0150] Конструирование одновалентных и поливалентных ДНК вакцин. Одновалентная ДНК вакцина состоит из Bmhsp или Bmalt2 в векторе pVAX1. Для получения одновалентной вакцины, кодон-оптимизированные гены Bmhsp или BmALT2 клонируют в эукариотический вектор экспрессии pVAX1 (Invitrogen, Carlsbad, СА) с использованием вставка-специфичных праймеров (Gnanasekar, et al. (2004) выше). Поливалентная иммуногенная композиция состоит из генов Bmhsp и Bmalt2 в одном векторе pVAX1. Кодон-оптимизированный ген Bmhsp сначала клонируют в вектор pVAX1 без стоп-кодона в обратном праймере (5'-CCG GAA ТТС ТСА СТТ GTC GTT GGT G-3'; SEQ ID NO: 24), но содержит сайт Pstl. Затем в этот клон встраивают кодон-оптимизированный ген Bmalt2 с использованием специфичных для гена праймеров (Gnanasekar, et al. (2004) выше). Параметры ПЦР для всех трех конструкций: денатурация при 94°С в течение 30 секунд, отжиг праймеров при 50°С в течение 30 секунд, удлинение праймеров при 72°С в течение 30 секунд, 30 циклов; окончательное удлинение в течение 5 минут проводят при 72°С. Наконец, вставку ДНК секвенируют, чтобы гарантировать подлинность клонированной нуклеотидной последовательности на обеих цепях. Плазмиды сохраняют и размножают в клетках Е. coli TOP10F'. Плазмиды очищают с использованием набора для экстракции плазмид без эндотоксина (Qiagen, Valencia, СА). ДНК анализируют электрофорезом в агарозном геле и количественно определяют на спектрофотометре (OD 260/280, соотношение>1,8).
[0151] Экспрессия и очистка рекомбинантных белков. Все гены клонируют в вектор pRSET-A (с N-концевой гексагистидиновой меткой) для продуцирования рекомбинантных белков. Конструкции Bmhsp и Bmalt2 трансформируют в BL21 (DE3), содержащий хозяина pLysS Е. coli (Invitrogen), чтобы свести к минимуму токсичность, обусловленную белком. Когда абсорбция культур достигает значения OD 0,6, к культурам добавляют 1 мМ IPTG (изопропилтио-d-галактопиранозид) и инкубируют в течение дополнительных 3 часов, чтобы индуцировать экспрессию гена. После лизиса клеток общие белки разделяют в 15% и 12% SDS-PAGE для подтверждения экспрессии рекомбинантных белков BmHSP (rBmHSP) и rBmALT2 с his-меткой. Затем рекомбинантные белки очищают с использованием аффинной хроматографии на колонке с иммобилизованным металлическим кобальтом (Clontech, Mountain View, СА) в соответствии с рекомендациями производителя. Затем рекомбинантные белки разделяют в SDS-PAGE и окрашивают бриллиантовым синим COOMASSIE R250 и серебряным красителем. Эти исследования показывают, что после очистки колонки получают одну полосу. Эндотоксины, если таковые имеются в рекомбинантных препаратах, удаляют путем пропускания рекомбинантных белков через аффинные колонки с полимиксином В (Thermo Fisher Scientific, Rockford, IL), и уровни эндотоксина в конечных препаратах определяют с использованием набора Е-ТОХАТЕ (Sigma, St Louis, МО) в соответствии с инструкциями производителя. Уровни эндотоксина ниже пределов обнаружения в этих препаратах рекомбинантного белка.
[0152] Иммунизация мышей. В этих экспериментах используют шестинедельных самцов мышей Balb/c, приобретенных в Charles River Laboratories. Гуманное использование животных в этом исследовании и протокол одобрены комитетом IACUC Медицинского колледжа Университета Иллинойса в Рокфорде. Мышей делят на четыре (4) группы по пять (5) животных в каждой. Всех мышей иммунизируют подкожно, используя схему вакцинации с примированием ДНК и стимулированием белком. Все экспериментальные группы мышей примируют двумя инъекциями кодон-оптимизированной ДНК, не содержащей эндотоксинов, в объеме 50 мкл и стимулируют двумя дозами рекомбинантных белков, суспендированных в квасцах (по 50 мкл каждая), которые вводят с интервалом в две недели.
[0153] Мышей группы А примируют 100 мкг pVAXBmhsp и стимулируют 15 мкг rBmHSP; мышей группы В примируют 100 мкг pVAXBmalt2 и стимулируют 15 мкг rBmALT2; мышей группы С примируют 100 мкг ДНК pVAXBmhsp/Bmalt2 и стимулируют 15 мкг rBmHSP и 15 мкг rBmALT2. Мыши группы D получают 100 мкг вектора pVAX1 плюс 50 мкл квасцов и служат контролем. Образцы крови собирают у каждой мыши перед иммунизацией и через месяц после последней стимулирующей дозы. Сыворотки отделяют и хранят при -80°С.
[0154] Оценка ответов антител у мышей. Уровни анти-BmHSP и анти-BmALT2 антител измеряют в сыворотке иммунизированных и контрольных групп мышей с использованием косвенного ELISA согласно утвержденным способам (Veerapathran, et al. (2009) выше; Gnanasekar, et al. (2004) выше). Коротко, лунки 96-луночных титровальных микропланшетов покрывают rBmHSP, rBmALT2 или rBmHSP (1 мкг/мл) в карбонатном буфере (рН 9,6) в течение ночи при 4°С. После промывки лунок не связанные сайты блокируют 3% BSA в течение 1 часа при 37°С. Затем в лунки добавляют разведенные образцы сыворотки и инкубируют далее в течение ночи при 4°С. После промывки лунок добавляют меченное HRP антитело кролика против IgG мыши (1:5000) и инкубируют еще в течение 1 часа при 37°С. Цвет проявляют с использованием субстрата ABTS (2,2'-азинобис(3-этилбензотиазолин-6-сульфоновой кислоты)). Абсорбцию измеряют при 405 нм в микропланшетном ридере (BIO-RAD, Hercules, СА).
[0155] Исследования защиты на мышах. Вакцинный потенциал одновалентных и поливалентных иммуногенных композиций затем оценивают на модели мышей. Мышей иммунизируют, как описано выше, с использованием подхода примирования/стимулирования. Группа "Вектор плюс квасцы" служит отрицательным контролем. Иммунизированных и контрольных животных заражают с использованием способа камеры с микропорами, известного в данной области техники (Abraham, et al. (1989) Am, J, Trop, Med. Hyg. 40(6):598-604). Коротко, камеры с микропорами собирают с использованием 14×2 мм (акриловых) колец PLEXIGLASS (Millipore Corporations, Bedford, MA) и 5,0 мкм поликарбонатных мембран NUCLEOPORE (Millipore Corporations), которые прикрепляют к (акриловым) кольцам PLEXIGLASS с помощью цианакрилового адгезива и стоматологического цемента. Камеры погружают на ночь при 37°С в стерильную среду RPMI, содержащую гентамицин и антимикотический раствор. Перед экспериментами с заражением 20 живых инфекционных L3, суспендированных в среде RPMI 1640 с добавлением 15% инактивированной теплом фетальной телячьей сыворотки (FCS), вводят в камеры с микропорами, и отверстие герметично закрывают стоматологическим цементом. Затем камеру с микропорами, содержащую L3, хирургическим путем имплантируют в брюшную полость каждой мыши под анестезией. При хирургических вмешательствах соблюдаются асептические условия. Через 48 часов после имплантации, животных умерщвляют и извлекают камеры из брюшной полости. Содержимое каждой камеры опорожняют, и личинок исследуют под микроскопом на прилипание клеток и гибель личинок. Жизнеспособность личинок определяют микроскопически при 100×. Долю защиты показывают как количество мертвых паразитов общее количество извлеченных паразитов ×100.
[0156] Анализ цитокинов у мышей. Долю rBmHSP и rBmALT2-специфичных клеток, секретирующих интерферон-у (IFN-γ) и интерлейкин-4 (IL-4), определяют в селезенке контрольных и вакцинированных мышей с использованием анализа ELISPOT. Коротко, фильтровальные планшеты MILLIPORE MULTISCREEN HTS покрывают моноклональными антителами крысы против IFN-(мыши или моноклональными антителами крысы против IL-4 мыши (BD Pharmigen, San Diego, СА) в концентрации 10 мкг/мл в PBS буфере. После промывания планшетов неспецифические участки блокируют путем инкубации лунок в полной RPMI с 10% фетальной телячьей сывороткой в течение одного часа при комнатной температуре. Затем в каждую лунку добавляют приблизительно 3×106 клеток селезенки, суспендированных в полной среде RPMI1640 с добавлением 10% инактивированной теплом FBS. Клетки стимулируют rBmHSP или rBmALT2 (1 мкг/мл). Не стимулированные клетки служат контролем. Через сорок восемь часов после инкубации при 37°С во влажном 5% СО2 планшеты промывают и дополнительно инкубируют в течение 1 часа при комнатной температуре с 2 мкг биотинилированного антитела крысы против IFN-(мыши или биотинилированного антитела крысы против IL-4 мыши (BD Pharmigen). После промывания планшетов в каждую лунку добавляют конъюгированную со стрептавидином пероксидазу хрена (Thermo Fisher Scientific) (1:800) и инкубируют при комнатной температуре в течение одного часа. Планшеты промывают и окрашивают с использованием субстрата DAB (Thermo Fisher Scientific). Общее количество «ореолов» подсчитывают под стереомикроскопом.
[0157] Статистический анализ. Статистический анализ проводят с использованием программного обеспечения XL STAT v.7.5.2 (Kovach Computing Services, Anglesey, UK). Статистическую значимость между сопоставимыми группами оценивают с помощью соответствующих непараметрических тестов с уровнем значимости, установленным на уровне р<0,05.
[0158] Ответы антител у мышей. Сначала определяют, может ли поливалентная иммуногенная композиция образовывать значительные антитела против каждого из антигенных компонентов. Предыдущие исследования показали, что у мышей, аналогичным образом вакцинированных антигенами В. malayi, образуются значительные защитные IgG антитела хозяина (Veerapathran, et al. (2009) выше). Таким образом анализируют титры антител IgG. Результаты этого анализа показывают, что одновалентная иммунизация Bmhsp+rBmHSP и Bmalt2+rBmALT2 вызывает значительные (р<0,005) титры анти-BmHSP и анти-ВьАДТ2 IgG антител (ФИГ. 1). Поливалентная иммуногенная композиция также образует значительные титры IgG антител. После поливалентной иммуногенной композиции, мыши в равной степени продуцируют IgG антитела как против BmHSP, так и против BmALT2, что свидетельствует о том, что антигены не взаимодействуют и не конкурируют за доминирование. Интересным открытием является то, что поливалентная иммуногенная композиция образует в 1,5-1,75 раза более высокие (р<0,005) титры IgG антител по сравнению с одновалентной вакциной (ФИГ. 1). Эти данные указывают на то, что два антигена в поливалентной композиции могут действовать синергически, усиливая индуцированный вакциной ответ антител против каждого антигена у вакцинированных мышей. Открытия также показали, что комбинирование этих двух антигенов в составе вакцины имеет большое преимущество. Учитывая сильный ответ IgG антител, индуцированный после вакцинации, также возможно, что концентрация составляющих антигенов в поливалентном препарате может быть снижена.
[0159] Поливалентная иммуногенная композиция вызывает значительную защиту у мышей. Приведенные в настоящем документе результаты показывают, что после вакцинации одновалентными и поливалентными препаратами иммуногенной композиции образуются значительные IgG антитела. Чтобы проверить, являются ли иммунные ответы, возникшие после вакцинации, защитными, вакцинированных животных заражают живыми инфекционными личинками третьей стадии (L3) В. malayi. Поскольку у этих животных паразиты не достигают зрелости, лучшее извлечение червей достигается, если паразиты имплантируются животным хирургическим путем. Стандартный способ заражения через камеру с микропорами (Abraham, et al. (1989) выше). Эти исследования показывают, что можно достичь почти 61% защиты у мышей, иммунизированных одновалентной вакциной (таблица 8). Это является очень значимым (р<0,001) по сравнению с отрицательным контролем. Это открытие также показывает, что rBmHSP и rBmALT2 можно использовать в вакцинах для лимфатического филяриоза. Эксперименты с заражением мышей, иммунизированных поливалентной иммуногенной композицией, показывают, что может быть достигнута значительно (р<0,005) более высокая защита по сравнению с одновалентной вакцинацией (таблица 8). Эти открытия также четко коррелируют с более высоким титром IgG антител у этих животных и подтверждают приведенный выше вывод о том, что rBmALT2 и rBmHSP могут синергетически усиливать защитные иммунные ответы у вакцинированных животных при введении по схеме примирования/стимулирования (таблица 8).
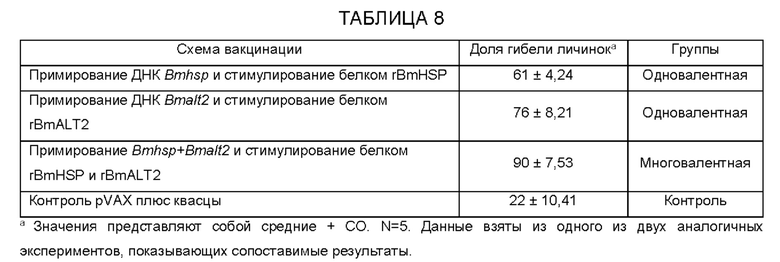
[0160] Чтобы дополнительно продемонстрировать эффективность, мышей иммунизируют различными комбинациями примирования/стимулирования. Как показано на ФИГ. 3, 100% защита может быть достигнута у мышей после иммунизации гибридным белком HAT или после иммунизации примированием/стимулированием гибридной ДНК HAT и гибридным белком HAT.
[0161] Цитокиноеые ответы. Иммунологические характеристики защитных ответов у вакцинированных мышей определяют путем оценки секретируемых цитокинов клеток селезенки в ответ на вакцинные антигены. Когда клетки селезенки стимулируют rBmHSP или rBmALT, наблюдают значительную антигенспецифическую пролиферацию клеток селезенки, что свидетельствует о сильном клеточном ответе на антигены. Для идентификации цитокинового профиля этих антиген-отвечающих клеток, клетки, секретирующие IFN-(и IL-4, подсчитывают с использованием анализа ELISPOT. Результаты этих исследований показывают, что клетки селезенки мышей, вакцинированных поливалентной иммуногенной композицией, преимущественно секретируют IL-4 (ФИГ. 2А). Количество клеток, секретирующих IFN-γ, очень низкое (ФИГ. 2 В). В целом, эти результаты показывают, что индуцированная вакциной защита в значительной степени опосредована ответами типа Th2.
Пример 3: BmVal-1+Bm ALT2 поливалентная иммуногенная композиция
[0162] Сыворотка. Образцы сыворотки, используемые в этом исследовании, берут из архивных образцов, хранящихся в Институте медицинских наук имени Махатмы Ганди, Севаграм, Индия. Эти образцы были собраны в рамках эпидемиологических исследований в районе Вардха, эндемичном по лимфатическому филяриозу, и его окрестностях.
[0163] Для этого исследования нет доступных демографических данных, за исключением того, что образцы сыворотки классифицированы как микрофиляремийные (MF), хронические патологии (CP) или эндемические нормальные (EN) на основании обнаружения циркулирующих паразитов, антигенов паразитов или оценки клинических симптомов лимфатического филяриоза. Циркулирующие микрофилярий обнаруживают в крови субъектов известными способами (Haslbeck, et al. (2005) Nat Struct Mol. Biol. 12:842-846; Yoo, et al. (2005) Biotechnol. Lett. 27: 443-448). Наличие циркулирующего антигена обнаруживают с помощью набора Од4СЗ и иммуноферментного анализа (ELISA) на основе WbSXP. Субъектов без циркулирующего антигена или микрофилярий классифицируют как EN, тогда как субъектов с циркулирующими микрофиляриями и/или циркулирующим антигеном, обнаруженными ELISA, считают MF. Субъектов, демонстрирующих лимфедему и другие видимые клинические симптомы филяриоза собирают в группу СР. Контрольные не эндемичные нормальные (NEN) сыворотки собирают в клинике Университета Иллинойса в Рокфорде, Иллинойс.
[0164] Паразиты. Brugia malayi L3 получены из Ресурсного центра исследований реагентов против филяриоза NIAID/NIH (FR3) в Университете Джорджии, Athens, GA.
[0165] Конструирование одновалентных и поливалентных ДНК-вакцин. Для приготовления одновалентной вакцины, кодон-оптимизированные гены Bmval-1 или Bmalt-2 клонируют в эукариотический вектор экспрессии pVAX1 (Invitrogen, Carlsbad, СА) с использованием вставка-специфичных праймеров (Yoo, et al. (2005), выше; Huang, et al. (2005) Immunol. Lett. 101:71-80). Для получения поливалентной иммуногенной композиции, кодон-оптимизированный ген Bmval-1 сначала клонируют в вектор pVAX1 без стоп-кодона с использованием уже опубликованных последовательностей праймеров с сайтом Pstl. Затем в этот клон встраивают кодон -оптимизированный ген Bmalt-2 с использованием ген-специфических праймеров. Параметры ПЦР для всех конструкций: денатурация при 94°С в течение 30 секунд, отжиг праймеров при 50°С в течение 30 секунд, удлинение праймеров при 72°С в течение 30 секунд, 30 циклов; и окончательное удлинение в течение 5 минут проводят при 72°С. ДНК-вставку секвенируют для обеспечения аутентичности клонированной нуклеотидной последовательности на обеих цепях. Плазмиды сохраняют и размножают в клетках Е. coli TOP10F'. Плазмиды очищают с использованием набора для экстракции плазмид без эндотоксина (Qiagen, Valencia, СА). ДНК анализируют с помощью электрофореза в агарозном геле и количественно определяют на спектрофотометре (OD 260/280, соотношение>1,8).
[0166] Экспрессия и очистка рекомбинантных белков. Рекомбинантные BmVAL-1 и rBmALT2 экспрессируют в векторе pRSET-A и очищают с использованием аффинной хроматографии на колонке с иммобилизованным металлическим кобальтом в соответствии с опубликованными способами (Norimine, et al. (2004) Infect. Immun. 72:1096-1106; Shinnick, et al. (1988) Infect. Immun. 56:446-451). Эндотоксин в рекомбинантных препаратах удаляют пропусканием рекомбинантных белков через аффинные колонки с полимиксином В (Thermo Fisher Scientific, Rockford, IL), и уровни эндотоксина в конечных препаратах определяют с использованием набора Е-ТОХАТЕ (Sigma, St Louis, МО) в соответствии с инструкциями производителя. Уровни эндотоксина в конечных препаратах (0,005 ЕЭ/мл) ниже пределов обнаружения в этих препаратах рекомбинантных белков.
[0167] Иммуноре активность сыворотки человека. Чтобы определить, несут ли образцы человеческой сыворотки антитела против BmVAL-1 или BmALT2, проводят ELISA (Haslbeck, et al. (2005) выше; Yoo, et al. (2005) выше). Для изотип-специфического ELISA в качестве вторичных антител используют конъюгированные с щелочной фосфатазой антитела козы к IgG1 человека, IgG2 человека, IgG3 человека и IgG4 человека (Sigma).
[0168] Протокол иммунизации мышей и песчанок. В этих экспериментах используют шестинедельных самцов мышей Balb/c и беспородных самцов монгольских песчанок весом 35-40 г, приобретенных в Charles River Laboratories (Wilmington, MA). С животными обращаются в соответствии с рекомендациями Руководства по уходу и использованию лабораторных животных. Используют две разные модели на животных, поскольку паразит В. malayi не созревает во взрослых особях у мышей, поэтому на мышиной модели можно оценить индуцированную вакциной защиту от стадий L3. Кроме того, у мышей можно измерить важные иммунологические параметры. Наоборот, паразит В. malayi развивается в зрелых взрослых червей в песчанках. Таким образом, индуцированная вакциной защита может быть оценена против внедрения взрослых червей у песчанок.
[0169] Проводят три серии экспериментов: (1) одновалентная вакцинация BmVAL-1, (2) одновалентная вакцинация BmALT2 и (3) поливалентная вакцинация mVAL-1/BmALT2. Каждая экспериментальная группа состоит из четырех групп (а) примирование ДНК плюс стимулирование ДНК (гомологичное), (b) примирование белком плюс стимулирование белком (гомологичное), (с) примирование ДНК плюс стимулирование белком (гетерологичное) и контроль pVAX плюс квасцы. Каждая группа включает по десять (10) животных. Всех животных иммунизируют подкожно кодон-оптимизированной ДНК (100 мкг) в объеме 50 мкл или рекомбинантным белком (150 мкг) плюс квасцы в объеме 50 мкл. Контрольная группа получает 100 мкг пустого вектора pVAX1 или 50 мкл квасцов. Образцы крови отбирают через частые промежутки времени, сыворотку отделяют и хранят при -80°С. Протокол, используемый для иммунизации мышей и животных, следующий. У животных предварительно берут кровь и дают первую дозу на 0 день. Вторую дозу вводят на 14 день, а затем берут кровь. Третью и четвертую дозы вводят на 28 и 42 день, соответственно, после чего у животных берут кровь. Мышей заражают на 56 день, и защиту определяют на 58 день. Мышей заражают на 60 день, и защиту определяют на 155 день.
[0170] Исследования защиты на мышах. Исследования с заражением проводят на мышах путем хирургической имплантации двадцати живых инфекционных L3 В. malayi в брюшную полость в камере с микропорами (Veerapathran, et al. (2009) выше; Abraham, et al. (1988) выше). При хирургических вмешательствах соблюдают асептические условия. Через сорок восемь часов после имплантации камеры извлекают из брюшной полости и определяют жизнеспособность личинок под световым микроскопом. Долю защиты выражают как количество мертвых паразитов+общее количество извлеченных паразитов (100.
[0171] Пролиферация спленоцитов и анализы цитокинов. Одноклеточную суспензию клеток селезенки (0,5×106 клеток на лунку, суспендированную в 200 мкл среды) готовят от каждой мыши и культивируют в трех лунках либо с (1) 1 мкг/мл rBmVAL-1, (2) 1 мкг/мл rBmALT2, (3) 1 мкг/мл rBmVAL-1+BmALT2, (4) неспецифическим рекомбинантным белком (1 мкг/мл G-связывающего белка Schistosoma mansoni) или (5) оставляют не стимулированными в среде. Все клетки инкубируют в течение 3 дней при 37°С с 5% СО2. Через 3 дня в каждую лунку добавляют 3Н-тимидин (0,5 мкКи на лунку, Amersham Biosciences) и дополнительно инкубируют.Клетки собирают через 16 часов и измеряют поглощение 3Н-тимидина в жидкостном сцинтилляционном счетчике и выражают как индекс стимуляции (SI)=(импульсы в минуту для стимулированных культур, импульсы в минуту для не стимулированных культур). Супернатанты клеточных культур, собранные из культур селезенки, анализируют на IFN-γ, IL-4, IL-5 и IL-10 с использованием набора ELISA, приобретенного у eBioscience Inc. (San Diego, CA).
[0172] BmVAL-1 и BmALT2 специфические IgG антитела в сыворотке иммунизированных мышей. Титр анти-BmVAL-l - и анти-BmALT2-специфических антител определяют в сыворотке иммунизированных мышей с помощью ELISA (Vee га path ran, et al. (2009) выше; Gnanasekar, et al. (2004) Infect, immun. 72:4707-15). Предиммунные сыворотки служат контролем. Конъюгированное с HRP антитело козы против IgG мыши используют в качестве вторичного антитела (Thermo Fisher Scientific) для анализов на мышах. В качестве подложки используют OPD (Sigma) и измеряют оптическую плотность (OD) при 405 нм.
[0173] Анти-BmVAL-1- и анти-BmALT2-специфические IgG1, IgG2a, IgG2b, IgG3 и IgG4 антитела определяют в сыворотке мыши с использованием набора для изотипирования мышиных антител, приобретенного у Thermo Fisher Scientific. Все ELISA выполняют в соответствии с рекомендациями производителя и считывают оптическую плотность при 405 нм. В качестве вторичных антител используют HRP-меченное антитело козы анти-IgG изотипа, и цвет проявляют с использованием субстрата OPD.
[0174] Исследования проблем у песчанок. Песчанок заражают 100 L3 В. malayi и внедрение червей определяют на 95 день после заражения в соответствии с утвержденными способами (Weil, et al. (1992) выше). Песчанки являются допустимыми хозяевами для В. malayi, и черви созревают во взрослых самцов и самок примерно за 75 дней. Наличие половозрелых червей в контрольной группе песчанок подтверждают выявлением микрофилярий в их крови на 80 день после заражения. Долю снижения внедрения червей рассчитывают по формуле: среднее количество червей, извлеченных из контрольных червей - среднее количество червей, извлеченных из вакцинированных животных / среднее количество червей, извлеченных из контрольных животных (100.
[0175] Статистический анализ. Статистический анализ проводят с использованием программы SIGMASTAT (Jandel Scientific, San Rafel, California) и программного обеспечения STATVIEW(SAS Institute, Сагу, NC). Критерий знаковых рангов Уилкоксона используют для сравнения парных данных; сравнение между группами проводят с помощью U-критерия Манна-Уитни. Значение р<0,05 считают статистически значимым.
[0176] Индивидуумы EN несут высокий титр антител против BmVAL-1 и BmALT2. Значительные анти-BmVAL-1 и анти-BmALT2 IgG антитела присутствуют в сыворотке субъектов EN по сравнению с субъектами MF (р<0,01) и субъектами CP (р<0,005). Субъекты NEN не имеют IgG антител ни к одному из антигенов. Последующий анализ антител изотипа IgG в сыворотке субъектов EN показал, что анти-BmVAL-1 и анти-BmALT2 антитела преимущественно имеют изотип IgG1 и IgG3.
[0177] Высокий титр ответов антител в сыворотке иммунизированных мышей. Показано, что мыши, вакцинированные антигенами В, malayi, образуют значительные защитные IgG антитела хозяина. Поэтому определяют титры IgG антител в сыворотке иммунизированных мышей. Одновалентная иммунизация BmVal-1 и одновалентная иммунизация BmAlt2 создает значительные (р<0,005) титры анти-BmVAL-l и анти-BmALT2 IgG антител в сыворотке мышей. По сравнению с контролями, группа, иммунизированная примированием/стимулированием дает максимальный титр антител, за ней следуют группы, иммунизированные белком, и группы, иммунизированные ДНК. Иммунизация поливалентным иммуногенным составом (BmVAL-1+BmALT2) также создает значительные титры IgG антител как против rBmVAL-1, так и против rBmALT2, и титры являются сопоставимыми, что указывает на то, что антигены не мешают друг другу и не конкурируют за доминирование. Интересным открытием является то, что поливалентная иммуногенная вакцина создает значительно более высокий (р<0,001) титр IgG антител у мышей по сравнению с любой из одновалентных вакцин. Эти данные показывают, что два антигена в поливалентной композиции синергетически усиливают индуцированный вакциной гуморальный ответ.
[0178] В целом белковая вакцинация вызывает более высокий титр IgG антител по сравнению с ДНК вакцинами, что указывает на высокую иммуногенность белковых вакцин. Другое наблюдение заключается в том, что гетерологичный подход примирования/стимулирования дает более высокую сероконверсию, чем гомологичный подход примирования/стимулирования. Таким образом, оказалось, что в целом гетерологичный подход примирования/стимулирования стимулирует самый высокий титр антител.
[0179] Анализ субпопуляции IgG антител показал, что вакцинация BmVAL-1 индуцирует, в первую очередь, IgG1 и IgG2a изотип антител, тогда как вакцинация BmALT2 индуцирует IgG1, IgG2a и IgG3 изотип антигенспецифических ответов антител. Ответы антигенспецифического IgG4 антитела не очевидны. Подход примирование/стимулирование значительно усиливает ответы IgG изотипа. После схемы поливалентной вакцинации, в сыворотке мыши присутствуют IgG1, IgG2a и IgG3 субпопуляции антигенспецифических антител.
[0180] Антиген-специфические ответы в селезенке мышей. Клетки селезенки иммунизированных мышей, стимулированных либо rBmVAL-1, либо rBmALT2, значительно пролиферируют (SI 10,8±1,1 и SI 14,6±1,2, соответственно) по сравнению с контрольной средой (SI 2,1±0,9). Клетки селезенки мышей, иммунизированных поливалентной конструкцией, отвечают как на rBmVAL-1 (SI 18,9±2,6), так и на rBmALT2 (SI 23,5±3,1), что указывает на то, что сильный ответный клеточный ответ получен как на BmVAL-1, так и на BmALT2 после вакцинации поливалентной конструкцией.
[0181] Анализ цитокинов из супернатантов пролиферирующей культуры. Для идентификации цитокинового профиля антиген-отвечающих клеток собирают культуральный супернатант клеток селезенки мыши, стимулированных соответствующим антигеном (rBmVAL-1 или rBmALT2), и измеряют уровень IFN-γ, IL-4, IL-5 и IL-10. Эти результаты показывают, что клетки селезенки секретируют значительные уровни IL-5 и IFN-(в ответ на rBmVAL-1. Клетки селезенки, стимулированные rBmALT2, преимущественно секретируют IL-4 и IL-5.
[0182] Поливалентная иммуногенная композиция индуцирует значительную защиту у мышей и песчанок. Приведенные в настоящем документе результаты показывают, что после вакцинации одновалентными и поливалентными иммуногенными препаратами образуются значительные IgG антитела. Чтобы проверить, являются ли иммунные ответы, вызванные после вакцинации, защитными, вакцинированных животных заражают живыми инфекционными личинками третьей стадии (L3) В. malayi. Поскольку паразиты не достигают зрелости у мышей, используют стандартный способ заражения с использованием камеры с микропорами (Gnanasekar, et al. (2004) выше). Эти исследования показывают, что после иммунизации одновалентной вакциной у мышей достигнута защита от 39% до 74% (таблица 9).
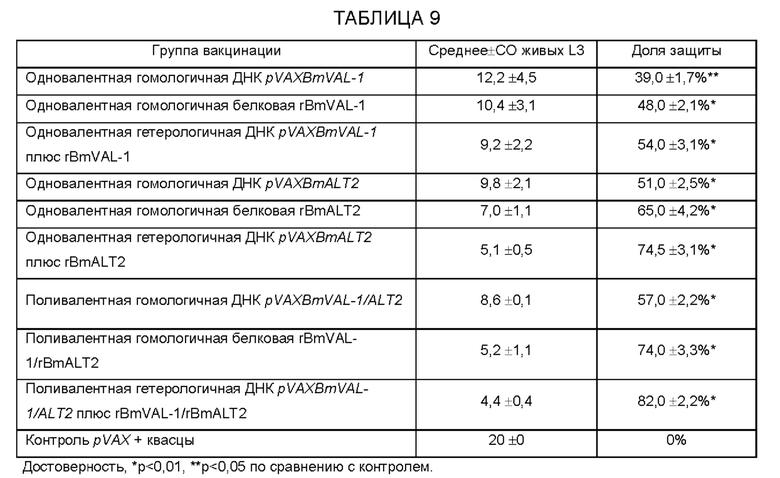
[0183] Белковая вакцинация дает лучшие результаты, чем вакцинация ДНК. Схема примирования/стимулирования дает наилучшие результаты в целом. Вакцинация BmALT2 дает более высокую долю защиты по сравнению с BmVAL-1. Точно так же, схема поливалентной вакцинации дает защиту от 57% до 82% по сравнению со схемой одновалентной вакцинации. Эти данные показывают, что BmVAL-1 и BmALT2 синергически усиливают защитные иммунные ответы у вакцинированных животных при введении в виде поливалентной иммуногенной композиции.
[0184] Анализ толстого мазка крови, полученного от контрольной группы песчанок на 80 день после заражения, показывает, что все пять песчанок являются положительными на микрофилярии, тогда как в периферической крови вакцинированных песчанок микрофилярии не обнаруживаются. Через пятнадцать (15) дней животных умерщвляют и подсчитывают количество самцов и самок червей в брюшной, тазовой и плевральной полостях, и сравнивают результаты контрольной и вакцинированной групп (таблица 10). Результаты вакцинации песчанок также подтверждают, что поливалентная схема примирования/стимулирования обеспечивает наивысший уровень защиты. Из поливалентно вакцинированных животных самок червей извлечено не было.

Пример 4: Поливалентная иммуногенная композиция BmHSP+Bm ALT2+BmTSP
[0185] Паразиты. Brugia malayi L3 получают из Ресурсного центра исследований реагентов против филяриоза NIAID/NIH (FR3) в Университете Джорджии, Athens, GA.
[0186] Конструирование ДНК вакцины pVAX Bmhsp+Bmalt2+Bmtsp.Кодон-оптимизированную последовательность ДНК, кодирующую Bmhsp, амплифицируют с прямым праймером 5'-CGC GGA ТСС АСС GTG АТС CAT TGT CG-3((SEQ ID NO: 25), содержащим сайт рестрикции BamHI, и обратным праймером 5'-ААС TGC AGC TGT ТТТ CCA ТТТ ССА ТТС-3((SEQ ID NO: 26), содержащим сайт рестрикции Pstl без стоп-кодона и клонируют в вектор pVAX. Полученную плазмиду обозначают как pVAX Bmhsp*. Кодон-оптимизированный ген Bmalt2 амплифицируют с прямым праймером 5'-ААС TGC AGA TGG GTA АСА AGC ТСС ТСА TCG-3((SEQ ID NO: 27) и обратным праймером без стоп-кодона 5'-CGC GAA ТТС GGC GCA CTG CCA АСС TGC-3((SEQ ID NO: 28). Подчеркнутые последовательности указывают сайты рестрикции Pstl и EcoRI в прямом и обратном праймерах, соответственно. Амплифицированную ДНК вставку Bmalt2 затем субклонируют в плазмиду pVAX Bmhsp* на сайтах рестрикции Pstl и EcoRI, в результате чего получают плазмиду pVAX Bmhsp+Bmalt2*. Для клонирования конечного продукта плазмиды pVAXBmhsp+Bmalt2+Bmtsp последовательность гена, кодирующего только ECL домен Bmtsp, амплифицируют с прямым праймером 5'-CGC GAA ТТС АСС ATG GTC CTG GAG-3((SEQ ID NO: 29), содержащим сайт рестрикции EcoRI, и обратный праймер со стоп-кодоном 5'-GCT СТА GAT CAG ТСС TTC TGG СТА G-3((SEQ ID NO: 30), содержащий сайт рестрикции XbaI, и клонируют в плазмиду pVAX Bmhsp+Bmalt2*. Двухвалентные конструкции HSP+TSP, TSP+ALT и HSP+ALT также конструируют с соответствующими праймерами.
[0187] Конструирование pRSETA Bmhsp+Bmalt2+Bmtsp, поливалентного слитого белка. Слитый белок Bmhsp+Bmalt2+Bmtsp конструируют так же, как описано выше. Последовательности праймеров HSP, ALT2 и TSP следующие. Bmhsp, прямой праймер, 5'-CGG GAT CCA TGG AAG AAA AGG TAG TG-3((SEQ ID NO: 31), содержащий BamHI, и обратный праймер, 5'-ССС TCG AGT GCT TTC ТТТ TTG GCA GC-3((SEQ ID NO: 32), содержащий XhoI. Bmalt2, прямой праймер, 5'-ССС TCG AG A TGA ATA ААС ТТТ ТАА TAG САТ-3((SEQ ID NO: 33), содержащий XhoI, или 5'-ААС TGC AGA TGG GTA АСА AGC ТСС ТСА TCG-3((SEQ ID NO: 27), и обратный праймер, 5'-GGG ТАС CCG CGC ATT GCC ААС СС-3((SEQ ID NO: 34), содержащий KpnI. Bmtsp, прямой праймер, 5'-GGG GTA ССС CGG САА GGA ТСА АТТ ТАА АА-3((SEQ ID NO: 35), содержащий Крп\, и обратный праймер, 5'-CGGAAT ТСТ САА ТСТ ТТТ TGA GAT GAA Т-3((SEQ ID NO: 36), содержащий EcoRI, используют для амплификации Bmtsp фрагмента SEQ ID NO: 77. Праймеры также конструируют для амплификации фрагмента большой внеклеточной петли (LEL) тетраспанина SEQ ID NO: 63. Двухвалентные конструкции (НА, НТ и ТА) также клонируют по отдельности в вектор pRSETA.
[0188] Иммунизация животных. Шестинедельных мышей Balb/C иммунизируют 100 мкг ДНК внутрикожно (в/к) в качестве ДНК вакцины или 15 мкг рекомбинантного белка подкожно (п/к) в качестве белковой вакцины или двумя дозами ДНК и двумя дозами белка в качестве примирующей/стимулирующей вакцины. Мышей случайным образом делят на 15 групп по 5 мышей в группе. Животных из групп 1-3 иммунизируют HSP+ALT2 (НА). Группы 4-6 иммунизируют HSP+TSP (НТ), и 7-9 иммунизируют TSP+ALT2 (ТА). Мышей из групп 10-12 иммунизируют поливалентной иммуногенной композицией HSP+ALT2+TSP (HAT). Контрольная группа животных получает вектор pVAX и/или квасцы (научно-исследовательский институт инфекционных болезней (IDRI)). Этот эксперимент повторяют дважды со всеми группами.
[0189] Анализ ответа антител у иммунизированных животных. Уровни IgG антител в сыворотке иммунизированных и контрольных групп животных против всех трех белков определяют с помощью косвенного ELISA (Anandharaman, et al. (2009), выше). Коротко, лунки 96-луночного титровального микропланшета покрывают рекомбинантными белками (rHSP, rALT2 или rTSP; 1 мкг/мл) в карбонатном буфере, рН 9,6, в течение ночи при 4°С и блокируют 3% BSA в течение 1 часа при 37°С. В лунки добавляют образцы сыворотки, и планшеты инкубируют в течение ночи при 4°С. После промывания добавляют HRP-меченное антитело мыши против IgG человека (1:5000) и инкубируют еще в течение 1 часа при 37°С. Цвет проявляют с помощью субстрата OPD (о-фенилендиамина) (Sigma Aldrich, USA). Абсорбцию измеряют при 450 нм в микропланшетном ридере (BIO-RAD, Hercules, СА).
[0190] Иммуноблот анализ также проводят с сывороткой иммунизированных мышей. Образцы сыворотки мышей, иммунизированных рекомбинантными белками (rHAT, rALT2, rTSP или rBmHAT), используют для иммуноблоттинга. Цвет блота проявляют с помощью диаминобензидинового (DAB) субстрата.
[0191] Анализ ADCC, Для оценки защитной эффективности комбинаций антигенов проводят ADCC in vitro с сывороткой мышей, иммунизированных двухвалентными и трехвалентными вакцинными конструкциями. Анализ ADCC in vitro проводят в соответствии с известными способами (Chandrasekhar, et al. (1990) выше). Коротко, клетки перитонеального экссудата (РЕС) собирают у нормальных мышей Balb/c путем промывания брюшной полости стерильной средой RPMI 1640. Клетки промывают и суспендируют в среде RPMI 1640 с добавлением 10% фетальной телячьей сыворотки (FCS). Десять L3 В. malayi добавляют к 2×105 клеток перитонеального экссудата (РЕС)/лунка в 96-луночных культу рал ьных планшетах (Thermo Fisher Scientifics, USA), 50 мкл сыворотки иммунизированных мышей и 50 мкл среды RPMI 1640 добавляют в лунки в трех повторах и инкубируют в течение 48 часов в 5% СО2 при 37°С. Жизнеспособность личинок определяют микроскопически через 48 часов инкубации. Личинки, которые являются прозрачными, поврежденными и с прилипшими к ним скоплениями клеток, считают погибшими. ADCC оценивают как долю гибели личинок, рассчитываемую по формуле: Количество погибших личинок (Общее количество личинок ×100.
[0192] Обеднение образцов сыворотки IgG антителами. Сыворотку мышей, иммунизированных поливалентной иммуногенной композицией, обедняют рекомбинантными антигенспецифическими IgG антителами с использованием кобальтовой смолы IMAC, сопряженной с his-меченными рекомбинантными антигенами (Anandharaman, et al. (2009) выше). Коротко, 1 мг his-меченного рекомбинантного белка (rHSP) сопрягают с 2 мл слоя смолы IMAC в течение 2 часов при 37°С. Кобальтовую колонку промывают десятью объемами слоя PBS (рН 8) и инкубируют в течение ночи при 4°С с 200 мкл объединенной сыворотки мышей, иммунизированных поливалентной иммуногенной композицией. Супернатант, содержащий обедненную сыворотку, собирают центрифугированием. Обедненную анти-HSP сыворотку инкубируют в течение ночи при 4°С в колонке, сопряженной с rALT2. Супернатант, содержащий обедненную анти-HSP и анти-ALT2 сыворотку, собирают и инкубируют в колонке, сопряженной с rTSP. Обедненную анти-HSP, анти-ALT2 и анти-TSP сыворотку собирают и используют.Обеднение IgG антителами против специфических антигенов подтверждают с помощью ELISA, как описано выше. Затем обедненную антителами сыворотку используют в анализе ADCC.
[0193] Анализ цитотоксичности in situ против личинок L3 у иммунизированных мышей (метод камеры с микропорами). Защитную эффективность вакцинации анализируют путем заражения иммунизированных животных инфекционными L3 с использованием способа камеры с микропорами (Abraham, et al. (1989) выше). Камеры с микропорами собирают с использованием 14×2 мм (акриловых) колец PLEXIGLASS и поликарбонатных мембран NUCLEOPORE 5,0 мкм (Millipore Corporations, Bedford, MA). Через 48 часов после имплантации животных умерщвляют и извлекают камеры из брюшной полости. Содержимое каждой камеры исследуют под микроскопом на предмет прилипания клеток и гибели инфекционных L3. Паразит считается погибшим, если он не является подвижным и прозрачным, и на поверхности имеется несколько прилипших клеток. Долю защиты рассчитывают по формуле: количество мертвых паразитов (количество извлеченных паразитов (100. Этот эксперимент повторяют дважды с пятью животными в каждой группе.
[0194] Пролиферация спленоцитов. Вакцинированных и контрольных мышей умерщвляют на 60 день и асептически удаляют селезенки. Суспензии одиночных клеток готовят в среде RPMI 1640 с добавлением 10% термоинактивированной FCS, пропуская через NYLON (алифатический полиамид) сито (BD Biosciences, Bedford, USA). После определения жизнеспособности клеток с помощью исключения красителя трипанового синего, приблизительно 2×106 клеток на лунку в трех повторах высевают в 96-луночные культуральные планшеты (ThermoFisher, USA). Спленоциты стимулируют 1 мкг/100 мкл/лунку рекомбинантных белков (rHSP, rALT2 или rTSP) или СопА или только средой (не стимулированной) в течение 72 часов при 37°С в атмосфере 5% СО2. Пролиферацию клеток определяют с использованием набора для подсчета клеток (CCK-8), приобретенного у Dojindo Molecular Technologies, Inc. (Gaithersburg, MD). Индекс стимуляции пролиферации клеток селезенки рассчитывают по формуле: Абсорбция стимулированных клеток ÷ Абсорбция не стимулированных клеток. Все культуры берут в трех повторах, и результаты выражают как средний И.С. ± СОС.
[0195] ПЦР в реальном времени (РВ-ПЦР), Уровни цитокинов в мРНК осадка клеток селезенки анализируют с помощью ПЦР в реальном времени. Клетки селезенки вакцинированных мышей и мышей контрольной группы культивируют, как указано выше, в концентрации 2×106 клеток/100 мкл/лунку в 96-луночных планшетах и стимулируют рекомбинантными антигенами (1 мкг/мл). Через 72 часа клетки центрифугируют (1000 об/мин в течение 5 минут), и из клеточных осадков экстрагируют общую РНК с использованием реагента TRIZOL (фенол, гуанидиний и тиоцианат) (Invitrogen) в соответствии с описанием производителя. После экстракции РНК, кДНК первой цепи синтезируют с помощью набора RT2 First Strand Kit (SuperArray Bioscience Corporation, Frederick, MD). Анализ матрицы ПЦР проводят в соответствии с протоколом производителя с помощью смеси RT2 Real-Time ТМ SYBR Green (цианиновый краситель) PCR Master Mix. Аликвоты этой смеси добавляют в 96-луночный планшет, где каждая лунка содержит предварительно распределенные наборы ген-специфических праймеров. Относительную количественную оценку представляющих интерес генов, которые экспрессируются, измеряют в приборе для ПЦР в реальном времени Applied BioSystem 7300 (Applied BioSystems, Foster City, СА). Параметры циклов следующие: 95°C в течение 10 минут для активации ДНК-полимеразы HOTSTART, затем 40 циклов денатурации при 95°С в течение 15 секунд и удлинение праймера при 60°С в течение 1 минуты. Набор массивов данных РВ-ПЦР создают и анализируют с использованием веб-системы анализа данных SABiosciences. Результаты выражают в виде кратности изменения иммунизированных мышей по сравнению с контрольными мышами путем нормализации экспрессии генов «домашнего хозяйства».
[0196] Цитокиновый анализ, Супернатанты культуры клеток спленоцитов собирают через 72 часа инкубации, стимулированной рекомбинантными антигенами (1 мкг/мл) или только средой. Уровни секретируемых цитокинов IL-4 и IFN-(в культуральных супернатантах определяют с использованием набора для сэндвич ELISA, приобретенного у Thermo Scientifics, USA. Все концентрации получают из стандартных кривых и данных, выраженных в пг/мл.
[0197] Конструирование плазмиды сНАТ и экспрессия белков слияния. Поскольку N-концевая область HSP участвует в связывании IL-10, эту область удаляют и получают рекомбинантный белок сНАТ в виде His-меченного белка с молекулярной массой 37 кДа.
[0198] Конструирование рекомбинантных плазмид и экспрессия белков слияния. Полноразмерные гены hsp, alt2 и tsp В. malayi стадии L3 конструируют с ожидаемым размером (850 п. о.). Эти фрагменты затем были направленно клонируют в векторы экспрессии pVAX1 и pRSETA с указанными сайтами разрезания ферментами рестрикции. Результаты анализа последовательности ДНК подтверждают направление вставки гена. rBmHAT экспрессируют в виде 45 кДа His-меченного слитого белка, который очищают и анализируют в SDS-PAGE. Результаты показывают, что слитый белок является чистым без каких-либо загрязняющих белков. Наличие антител ко всем трем антигенам подтверждают иммуноблот-анализом.
[0199] Титр антител в сыворотке иммунизированных мышей. Средний пиковый титр антител в образцах сыворотки мышей, иммунизированных примированием/стимулированием или белковой вакциной, значительно выше (р<0,001) по сравнению с группой ДНК. Сыворотки, собранные у животных, иммунизированных rBmHAT, показывают максимальный титр 30000 против антигена rALT2, в то время как титр антител против антигена rHSP или rTSP находится в диапазоне 18000-20000. Точно так же мыши, иммунизированные двухвалентной вакциной, показывают максимальный титр 30000 против антигена ALT2, в то время как анти-HSP и анти-TSP антитела находятся в диапазоне 8000-15000.
[0200] Антителозависимая клеточно-опосредованная цитотоксичность. Опосредованную антителами адгезию и цитотоксичность иммунных клеток к L3 личинкам В. malayi наблюдают через 48 часов инкубации паразитов с сывороткой и нормальными иммунными клетками. ADCC показывает максимальную цитотоксичность приблизительно 90% (р<0,001) в сыворотке мышей, иммунизированных вакцинными конструкциями rBmHAT или rWbHA (таблица 11). Двухвалентные вакцинные конструкции rWbHTn rWbTA также дают лучшую защиту 82% и 87%, соответственно, что является значимым по сравнению с одновалентно вакцинированными и контрольными животными (р<0,001). Для оценки защиты, опосредованной антителами, создаваемыми против антигенов HSP, ALT и TSP, иммунизированную сыворотку обедняют антителами IgG и используют в ADCC. Обедненные антитела показывают только 6% защиту против L3.
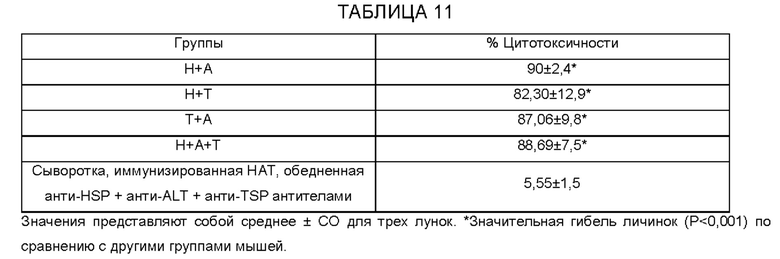
[0201] In situ исследование защиты. Через две недели после последней иммунизации способность вакцин-кандидатов убивать филяриозных паразитов у иммунизированных животных оценивают с помощью исследований камеры с микропорами in situ. Данные объединяют для двух аналогичных экспериментов и представляют как среднее значение ±СОС. Анализ доли снижения количества червей по сравнению с контролем показывает, что поливалентная иммуногенная композиция (HAT) обеспечивает максимальную защиту 100% и 94% для белка и примирующей/стимулирующей вакцины, что является очень значительной защитой (таблица 12) (Р<0,0001) по сравнению с контрольными группами (5%). Интересно, что доля снижения червей двухвалентными вакцинами НА, ТА и НТ составляет 90%, 80% и 82%, соответственно, что также значительно выше по сравнению с контролем. Во всей группе двухвалентной вакцины вакцинация примированием/стимулированием является большей защитой по сравнению с вакцинацией ДНК и белком.
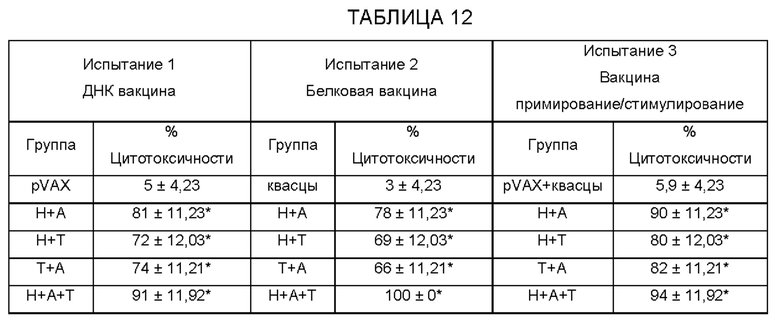

[0202] Пролиферация спленоцитое. Клетки селезенки, выделенные от вакцинированных и контрольных животных, стимулируют in vitro индивидуально с помощью rHSP, rALT2 или rTSP для анализа пролиферации протеин-специфических Т-клеток у вакцинированных животных. Мыши, иммунизированные по схеме примирования/стимулирования всеми комбинациями вакцин и HAT в качестве белковой вакцины, дают самую высокую защиту. Следовательно, спленоциты собирают только у этих животных, проанализированных на иммунный ответ.Спленоциты от двухвалентных и трехвалентных вакцинированных животных, стимулированных соответствующими рекомбинантными белками, показывают достаточно высокую (Р<0,001) пролиферацию (средний И.С.=4,25-5,8) по сравнению с одновалентными и не стимулированными контролями. Индекс пролиферации клеток селезенки, иммунизированных одновалентной конструкцией, показывает значительную пролиферацию. Стимуляция клеток сопоставима с положительными контролями.
[0203] Матрица ОТ-ПЦР. Для определения клеточных иммунных ответов на поливалентные конструкции у вакцинированных мышей клетки селезенки, собранные у вакцинированных и контрольных мышей, культивируют в присутствии соответствующих рекомбинантных белков и оценивают их пролиферативный ответ и цитокиновый профиль. Поскольку клетки селезенки вакцинированных животных значительно пролиферируют на вторичный ответ, измеряют уровни мРНК цитокинов. РВ-ПЦР матрицы генов цитокинов проводят на мРНК, собранной из клеток селезенки, стимулированных рекомбинантными белками. Эти результаты показывают, что гены цитокинов Th1 (IFN-γ, IL-2) и Th2 (IL-4) значительно увеличены у вакцинированных животных.
[0204] Уровни цитокинов. После определения наличия экспрессии цитокинов IFN-(и IL-4 в мРНК, выделенной из вакцинированных клеток селезенки, исследуют секрецию тех же цитокинов в супернатанте. Данные нормализуют с помощью не стимулированного контроля. Интересно, что профили цитокинов, наблюдаемые в супернатанте, показывают значительно более высокие уровни IFN-у, указывая на Th1-смещенный иммунный ответ.Эти результаты показывают, что рекомбинантные белки стимулируют продуцирование IFN-(и индуцируют Th1-опосредованный защитный ответ.
Пример 5: Анализ сНАТ-вакцины в различных адъювантных составах
[0205] Получение сНАТ. Предыдущие исследования показывают, что N-концевая последовательность BmHSP12.6 может связываться с рецептором IL-10 человека и запускать опосредованные IL-10 ответы (Gnanasekar, et al. (2008) MoL Biochem. ParasitoL 159(2):98-103). Поскольку IL-10 является иммунодепрессантом, последовательности связывания рецептора IL-10 удаляют из HSP. Усеченную последовательность называют cHSP. Затем cHSP используют для замены гена HSP и белка HSP в поливалентной гибридной вакцине HAT. Таким образом, полученная новая вакцина получила название сНАТ.
[0206] Исследования защиты с использованием вакцины сНАТ-слитый белок на мышах. Мышей иммунизируют четырьмя дозами слитого белка сНАТ с двухнедельными интервалами. Через месяц после последней иммунизации способность вакцин-кандидатов убивать филяриозных паразитов оценивают с помощью исследований в камере с микропорами in situ. Результаты показывают, что когда мышей иммунизируют слитым белком сНАТ с квасцами в качестве адъюванта, вакцина обеспечивает 81% защиту (таблица 13) (Р<0,0001) по сравнению с контрольными группами (2%), которые получают только фосфатно-солевой буфер (PBS) и квасцы. Затем тестируют различные адъюванты, чтобы увидеть, улучшит ли замена адъюванта защитную способность сНАТ. Тестируют два дополнительных адъюванта: квасцы, содержащие агонист TLR4 (приобретены в Научно-исследовательском институте инфекционных заболеваний, Seattle, Washington) и ALHYDROGEL (приобретены в Sigma, St. Louis, МО). сНАТ без адъювантов оставляют в качестве контроля. Результаты этих исследований (таблица 13) показывают, что 78% защита достигается при использовании квасцов плюс агонист TLR4, и сНАТ, введенная в адъюванте ALHYDROGEL, обеспечивает 70% защиту. Интересным открытием в этих исследованиях является то, что сНАТ без какого-либо адъюванта также дает 72% защиту, что указывает на то, что вакцина на основе слитого белка сНАТ может вводиться без какого-либо адъюванта и при этом обеспечивать значительную защиту.
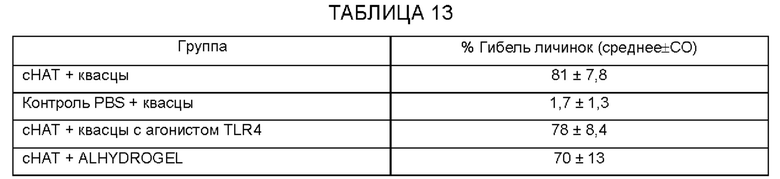
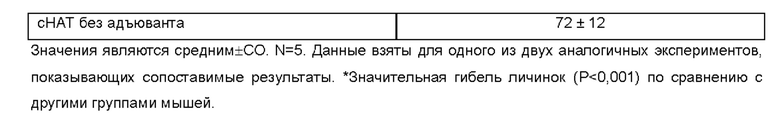
Пример 6: Гомологи HSP, ALT2 и TSP
[0207] Гомологи вакцинных антигенов HSP, ALT2 и тетраспанина присутствуют в О, volvulus и L. loa. Сравнение нуклеотидной последовательности HSP, ALT2 и тетраспанина из О. volvulus и L. loa показывает, что существует значительная гомология последовательностей (>90%) между белками всех филяриозных паразитов. Эти данные показывают, что вакцина на основе слитого белка сНАТ, разработанная в примере 5, может быть использована в качестве вакцины против О. volvulus и L. loa.
[0208] Например, тетраспанин О. volvulus клонируют из библиотеки кДНК L3 О. volvulus и получают рекомбинантные белки. Образец сыворотки мышей, вакцинированных сНАТ вакциной, которая дает 81% защиту по таблице 13, используют для зондирования рекомбинантного тетраспанина О. volvulus после разделения белка в 12% геле SDS-PAGE. В качестве положительного контроля используют тетраспанин В, malayi. Результаты показывают, что образец сыворотки в значительной степени реагирует с тетраспанином О. volvulus, что указывает на то, что вакцина сНАТ, разработанная в примере 5, может быть использована в качестве вакцины против О. volvulus.
Пример 7: Поливалентная иммуногенная композиция против лимфатического филяриоза на модели макаки-резуса
[0209] Паразиты. Инфекционные личинки третьей стадии (L3) В, malayi получают из Ресурсного центра исследований реагентов против филяриоза NIAID/NIH (Университет Джорджии, Athens, GA).
[0210] Поливалентный гибридный белок rBmHAT, Поливалентный слитый белок rBmHAT, экспрессированный в Escherichia coli BL21 (pLysS), очищают и удаляют эндотоксин с помощью высокопроизводительной колонки со смолой для удаления эндотоксинов Pierce (Thermo Fisher Scientific, Rockford, IL), как описано в настоящем документе.
[0211] Иммунизация rBmHAT, Каждая из пяти макак получает по 200 мкг вакцины rBmHAT, смешанной со 100 мкг квасцов AL007 (IDRI, Seattle, WA) в условиях ABSL-2. Пять (5) макак, получивших квасцы (AL007), остаются в качестве только контроля. Каждое животное анестезируют кетамином/ксилазином, и вакцину вводят внутримышечно в каждое бедро (одно место инъекции на бедро на вакцинацию). Животных иммунизируют с 4-недельными интервалами в дни 0, 28 и 56. Внутримышечный путь обычно используют для клинических испытаний вакцины, и, следовательно, туже процедуру применяют для макак. Места инъекций ежедневно контролируют на наличие признаков лихорадки, любых побочных реакций (покраснение, отек и т.д.) в течение 7 дней после иммунизации.
[0212] Контрольное заражение В. malayi L3. На 84 день, через месяц после последней дозы вакцины, макак анестезируют кетамином HCl и подкожно заражают 400-500 В. malayi L3. Для облегчения получения относительно большого количества L3 (500 L3/животное), необходимого для заражения 10 иммунизированных макак, животных внутри каждой группы делят на 2 подгруппы. Подгруппы тестируют с интервалом в одну неделю. Перед заражением В. malayi L3 подсчитывают и исследуют на жизнеспособность под микроскопом. Для заражения используют только жизнеспособных паразитов.
[0213] Отслеживание каждого животного после контрольного заражения. Всех животных ежедневно отслеживают на наличие клинических признаков после заражения. Наблюдения за поведением проводят аналогичным образом в течение всего периода после заражения. Клиническое отслеживание включает химический анализ сыворотки, гематологию, общий анализ крови (СВС) (IDEXX) и анализ проточной цитометрии CD4+/CD8+Т-клеток. Массу тела, состояние тела, лимфатический отек и измерения лимфатических узлов также регистрируют каждый раз, когда животное подвергается седации для процедур (таких как иммунизации, контрольное заражение и забор крови).
[0214] Сбор образцов. Собирают образцы крови и мононуклеарные клетки периферической крови (РВМС). Цельную кровь собирают в пробирки BD VACUTAINER SST в соответствии с инструкциями производителя. Гепаринизированную кровь (1 мл) собирают из бедренной вены каждого животного во время периода иммунизации и из подкожной вены во время периода контрольного заражения. Смещение места забора крови проводят для устранения любого потенциального влияние на измерения паховых лимфатических узлов или оценку отека. Образцы крови получают в несколько моментов времени в течение всего периода наблюдения.
[0215] Выделение РВМС. Осадок крови после отделения плазмы разводят в фосфатно-солевом буфере (PBS; 1:2) и подвергают центрифугированию в градиенте плотности в течение 30 минут при 2200 об/мин с использованием 90% раствора для разделения HISTOPAQUE (Sigma, St. Louis, МО). Непрозрачную поверхность раздела, содержащую мононуклеарные клетки, собирают, трижды промывают в PBS путем центрифугирования при 800 об/мин. РВМС подсчитывают с использованием способа исключения красителя трипанового синего и ресуспендируют в среде RPMI 1640, содержащей 10% FBS (100 ЕД/мл пенициллина/стрептомицина и 2 мМ L-глутамина). РВМС, собранные перед контрольным заражением, анализируют на пролиферацию Т-клеток и секрецию IFN-γ. РВМС, собранные после экспериментов с контрольным заражением, тестируют на пролиферацию Т-клеток и анализы ELISPOT. Анализ пролиферации проводят с РВМС, выделенными в день взятия крови. РВМС, суспендированные в среде RPMI с 10% FBS, используют для анализа ADCC и для анализа цитокинов.
[0216] Пролиферация Т-клеток и проточная цитометрия. Анализ на основе сукцинимидилового эфира карбоксифлуоресцеина диацетата (CFSE) используют для оценки антигенспецифической пролиферации в популяции Т-клеток (Parish, et al. (2009) Curr. Protoc. Immunol. Chapter 4: Unit 49). Исходный раствор 5 мМ CFSE (Invitrogen, Grand Island, NY) готовят в соответствии с инструкциями производителя. РВМС, собранные через четыре недели после последней иммунизации, осторожно ресуспендируют в концентрации 107 клеток/мл в 5 мкМ CFSE и инкубируют в темноте при 37°С в течение 15 минут.Клетки центрифугируют и промывают RPMI, содержащей 10% FBS (100 ЕД/мл пенициллина/стрептомицина и 2 мМ L-глутамина), и инкубируют в течение дополнительных 30 минут при 37°С. Затем клетки промывают, ресуспендируют в RPMI, содержащем FBS, высевают в 24-луночный планшет в количестве 2×106 клеток/мл на лунку и инкубируют в течение ночи при 37°С. Среду (~500 мкл) удаляют на следующий день, и клетки стимулируют 1 мкг/мл rBmHAT. Отрицательным контролем служат образцы, инкубированные только со средой RPMI. В качестве положительного контроля для каждого животного клетки стимулируют фитогемагглютинином (РНА). Клетки культивируют и собирают после 5 дней стимуляции. После стадии промывки PBS/0,2% FBS клетки окрашивают на поверхности коктейлем антител CD3-APC-Cy, CD4-PE и CD8-PerCP и инкубируют в течение 20 минут при комнатной температуре. После дополнительной стадии промывки PBS/0,2% FBS клетки собирают на проточном цитометре BD FACS CANTO II (BD, San Jose, CA) и анализируют с помощью программного обеспечения BD FACS DIVA v6.1.2. Получают, по меньше мере, 50000 событий при гейтировании живых лимфоцитов.
[0217] Подсчет клеток, биохимический анализ сыворотки и общий анализ крови (СВС). Общий анализ крови, биохимический анализ сыворотки и количество эозинофилов анализируют с использованием коммерческих автоматических анализаторов гематологии и биохимического анализа сыворотки IDEXX. Образцы, собранные до начала исследования, служат нормальным исходным эталоном для каждого животного.
[0218] Измерение секретируемых уровней IFN-γ. РВМС (1x106 клеток) стимулируют in vitro 1 мкг/мл rBmHAT в течение 5 дней при 37°С. После стимуляции супернатанты собирают и анализируют на секретируемые уровни IFN-(с использованием набора ELISA (Mabtech АВ, Ashburn, VA) в соответствии с инструкциями производителя.
[0219] Анализ ELISPOT. Анализ ELISPOT проводят для определения клеток, секретирующих антигенспецифический IFN-(и IL-10, в РВМС вакцинированных и контрольных макак. Набор ELISPOT для обезьян, приобретенный у U-Cytech biosciences (Yalelaan, The Netherlands), используют для определения единиц, образующих «ореол» в соответствии с инструкциями производителя. РВМС, собранные через 20 недель после контрольного заражения, высевают в 96-луночные планшеты с концентрацией 1×106 клеток/мл и стимулируют 100 нг/лунку растворимого антигена взрослых В. malayi (BmA) в течение 24 часов при 37°С и 5% СО2. Лунки планшетов ELISPOT покрывают 100 мкл/лунку захватывающими антителами (анти-IL-10 или анти-IFN-γ), разведенными в стерильном буфере для покрытия, и инкубируют в течение ночи при 4°С. Планшеты промывают 2 раза стерильным буфером для покрытия. После блокирования планшетов 200 мкл/лунку блокирующего буфера в течение 1 часа при комнатной температуре, РВМС, которые уже стимулированы антигенами BmA или только средой (отрицательный контроль), добавляют в лунки планшетов ELISPOT по 100 мкл/лунку и инкубируют в течение 24 часов при 37°С и 5% СО2. Все клетки удаляют с планшетов, и мембрану промывают 3 раза стерильным PBS. После промывки в каждую лунку добавляют по 100 мкл обнаруживающих антител и инкубируют при комнатной температуре в течение 2 часов. После промывки планшета 4 раза промывочным буфером добавляют реагент авидин-HRP (100 мкл/лунку) и инкубируют в течение 45 минут при комнатной температуре. После окончательной промывки PBS добавляют свежеприготовленный раствор субстрата 3-амино-9-этилкарбазола (АЕС) (100 мкл/лунку) и наблюдают за развитием «ореолов» при комнатной температуре в течение 10-60 минут.Реакцию субстрата останавливают, промывая лунки 3 раза сверхчистой водой по 200 мкл/лунку. Планшеты сушат на воздухе. «Ореолы» подсчитывают с помощью стереомикроскопа. Планшеты хранят в темноте до считывания. Антигенспецифические ответы определяют путем вычитания количества «ореолов» в лунках с отрицательным контролем из лунок, содержащих антигены. Результаты представлены как среднее значение «ореолов», полученных из трех лунок.
[0220] Анализ титров антител в сыворотке у макак. Уровни IgG, IgG1, IgG2, IgG3, IgA и IgE антител против rBmHSP, rBmALT2, rBmTSP или rBmHAT определяют в сыворотке (собранной через один месяц после последней дозы вакцины) каждой макаки-резуса с использованием косвенного ELISA, как описано в настоящем документе. Коротко, лунки 96-луночных титровальных микропланшетов для ELISA покрывают 100 нг/лунку антигенов (rBmHSP, rBmALT2, rBmTSP или rBmHAT) в 0,05 М карбонатно-бикарбонатном буфере, рН 9,6. Лунки блокируют 3% BSA в 0,05% PBS-TWEEN 20 (PBS-T) и добавляют по 100 мкл образцов сыворотки (разведенных в диапазоне 1:100-1:50000 в PBS-T) от каждой макаки в каждую лунку. Антитела козы против IgG обезьян, конъюгированные с пероксидазой (Rockland Immunochemicals, Gilbertsville, РА), используют в качестве вторичных антител для определения титра IgG антител. Цвет проявляют с использованием подложки OPD, и абсорбцию считывают при 492 нм в ридере ELISA (BioRad, Hercules, СА). Для определения уровней изотипических антител, биотинилированных антител против IgG1 (1:2000), IgG2 (1:200), IgG3 (1:2000), IgA (1:2000) и IgE (1:1000) обезьяны (NHP Reagent Resources, Boston, MA) используют в качестве вторичных антител. После промывки планшетов добавляют оптимально разведенную конъюгированную со стрептавидином пероксидазу хрена (HRP) и дополнительно инкубируют в течение 60 минут при комнатной температуре, и проявляют цвет.
[0221] Анализ ADCC. РВМС готовят из гепаринизированной цельной крови от наивного здорового животного, как описано выше. Коротко, десять В. malayi L3 (суспендированных в 50 мкл среды RPMI 1640, содержащей 10% FBS) инкубируют с 2×105 РВМС (в 50 мкл RPMI 1640) и 50 мкл сыворотки от каждого животного (собранной через месяц после последней дозы вакцины) в 96-луночном круглодонном планшете для тканевых культур. Для каждого образца сыворотки выполняют пять повторов. Контрольные лунки содержат В, malayi L3, инкубированные в среде, только с сывороткой или только с клетками. Планшеты инкубируют при 37°С с 5% СО2 в течение 48 часов. После инкубации, В. malayi L3 исследуют под микроскопом через 24 и 48 часов для определения жизнеспособности личинок. Мертвых L3 определяют как имеющих прозрачный или прямой вид без движений в течение дополнительного периода наблюдения 8 часов при 37°С. Живые личинки являются активными, свернутыми и подвижными. Долю гибели личинок выражают как отношение количества погибших L3 к общему количеству извлеченных в течение экспериментального периода, умноженное на 100. Рассчитывают среднюю гибель личинок в 5 лунках и выражают как долю защиты для каждого животного.
[0222] Тест Нотта для определения микрофиляремии (Mf) у макак. Присутствие Mf в крови макак определяют с использованием методики Нотта, как описано ранее (Liu, et al. (1989) J. Trop.Med. Hyg. 92:93-96). Периферическую кровь макак еженедельно проверяют на наличие Mf, начиная с 5 по 20 неделю после контрольного заражения. Коротко, цельную кровь смешивают с 9 мл 2% раствора формалина (приготовленного в PBS) в 15 мл конической центрифужной пробирке. Пробирки осторожно встряхивают в течение 2 мин при комнатной температуре и центрифугируют при 1500 об/мин в течение 5 мин. Затем супернатант тщательно декантируют, полностью перевернув пробирку вверх дном, чтобы удалить всю жидкость. После этого, к осадку добавляют 5 мл буфера для лизиса АСК (Quality Biologicals, Gaithersburg, MD) и встряхивают пробирку. Затем в пробирки добавляют две-три капли раствора метиленового синего (Fisher Scientific, Hannover Park, IL), осторожно перемешивают и наносят на пять предметных стекол. Образцам дают высохнуть и считывают их под микроскопом с использованием объектива с увеличением 40Х. Сравнение количества Mf в крови, взятой из подкожной и бедренной вен, показывает сходные результаты.
[0223] Обнаружение Mf в периферической крови методом ПЦР. Анализы на основе ПЦР более чувствительны при обнаружении присутствия Mf в образцах крови (Mishra, et al. (2005) Acta Trop.93:233-7; Tao, et al. (2006) J. Clin. Microbiol. 44:3887-93). Поэтому анализ на основе ПЦР также используют для подтверждения присутствия Mf в образцах крови всех макак через 20 недель после контрольного заражения. Образцы цельной крови центрифугируют при 10000 об/мин в течение 5 минут, и надосадочную жидкость, содержащую сыворотку, хранят при -20°С. ДНК выделяют из осадка с использованием набора DNEASY Blood & Tissue Kit (Qiagen, Valencia, CA) в соответствии с инструкциями производителя. Праймеры синтезируют в Integrated DNA Technologies Inc. (Coralville, IA) для тандемных повторов HhaI. Последовательности праймеров для тандемных повторов HnaI представляют собой: прямой 5'-GCG CAT AAA TTC АТС AGC-3((SEQ ID NO: 75) и обратный 5'-GCG САА ААС TTA ATT АСА AAA GC-3((SEQ ID NO: 76). Параметры ПЦР включают начальную денатурацию при 94°С в течение 5 минут, затем 40 циклов по 1 минуте при 94°С, 1 минуте при 56°С, 1 минуте при 72°С и окончательное удлинение в течение 10 минут при 72°С. После реакции ПЦР 10 мкл каждого продукта ПЦР анализируют на 1% агарозном геле.
[0224] Анализ пролиферации РВМС, РВМС, собранные через 10 недель после заражения, культивируют в 96-луночных планшетах для тканевых культур в концентрации 1×106 клеток/лунку в среде RPMI 1640 с добавлением 10% FCS. Клетки стимулируют либо антигеном rBmHAT (1 мг/мл), либо конканавалином А (1 мг/мл), либо только средой (не стимулированные) в трех лунках. РВМС стимулируют в трех лунках, и планшеты инкубируют при 37°С в 5% СО2. Через 72 часа измеряют пролиферацию клеток с использованием набора для подсчета клеток (CCK-8) (Dojindo Molecular Technologies, Inc., Gaithersburg, MD). Индекс стимуляции пролиферации РВМС рассчитывают по формуле: абсорбция стимулированных клеток/абсорбция не стимулированных клеток.
[0225] Статистический анализ. Данные представлены как среднее ± стандартная ошибка. Односторонний анализ ANOVA (Крускала-Уоллиса) выполняют для определения титра антител и пролиферации Т-клеток с использованием программного обеспечения GraphPad Prism. Т-критерий Стьюдента проводят для исследований защиты. Значение вероятности (Р) ≤0,001 считается статистически значимым.
[0226] Вакцинация rBmHAT не вызывает побочных реакций у макак. Места инъекций тщательно контролируют на наличие признаков каких-либо побочных реакций (покраснение, отек и т.д.) в течение 7 дней после иммунизации. Нежелательных реакций ни у вакцинированных, ни у контрольных животных не наблюдается. Клиническое отслеживание не выявляет резкой потери массы тела (>10% от исходной массы), изменений пищевых привычек или каких-либо других поведенческих изменений. Измерения температуры, полученные ежедневно после иммунизации, не показывают каких-либо существенных изменений. Измерения температуры также проводят через регулярные промежутки времени с использованием имплантированных транспондеров. Значительных колебаний температуры тела у вакцинированных и контрольных животных не наблюдается.
[0227] Лимфатические узлы на левой и правой ногах всех животных контролируют еженедельно, начиная примерно за 2 недели до заражения (для установления исходного уровня) и в течение всего периода контрольного заражения. Лимфатические узлы измеряют штангенциркулем и наблюдают за отеком. Измерения показывают общее увеличение среднего размера паховых лимфатических узлов на обеих ногах в течение 5-8 недель после контрольного заражения во всех группах. По сравнению с исходным уровнем (14,5 мм) размер лимфатических узлов у контрольных животных составляет 22±1 мм, и в группе rBmHAT составляет 26,2±1 мм. После этого периода размеры лимфатических узлов уменьшаются почти до уровня до контрольного заражения у всех макак.
[0228] Контрольное заражение В. malayi L3 не влияет на температуру тела макак. Анализы химических и гематологических показателей сыворотки (СВС) показали, что все они находятся в пределах нормы для всех типов клеток, за исключением небольшого увеличения количества эозинофилов после контрольного заражения L3 у инфицированных животных.
[0229] Все три антигена в поливалентной иммуногенной конструкции являются иммуногенными у макак. Анализ титра IgG антител у вакцинированных макак показывает, что все макаки развивают высокие титры (1:40000) IgG антител после третьей иммунизации против rBmHAT. Затем анализируют титр антител против каждого из трех составляющих антигенов в конструкции вакцины. У всех макак вырабатываются высокие титры IgG антител против rBmHSP12.6 (1:16000), rBmALT2 (1:24000) и rBmTSP-LEL (1:16000). Имеются небольшие индивидуальные различия в титре антител между вакцинированными макаками. На сравнительной основе макаки №5242, №5258 и №5259 демонстрируют самый высокий титр IgG антител против составляющих антигенов (за исключением анти-rBmHSPI 2.6 антител у макаки №5258 и анти-rBmTSP антител у макаки №5259). У макак №4996 и 5254 развиваются только низкие титры антител к rBm ALT2 и rBmTSP (таблица 14).

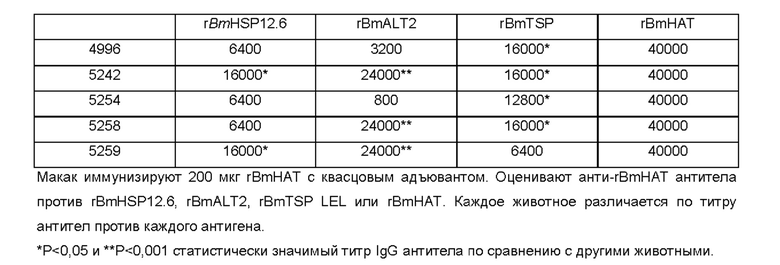
[0230] Изотипический анализ показал, что почти все антитела имеют IgG1 изотип против всех четырех протестированных антигенов (rBmHSP, rBmALT2, rBmTSP и rBmHAT). Уровни IgG2, IgG3, IgA и IgE существенно не отличаются от фоновых значений.
[0231] Клетки, отвечающие на rBmHAT, присутствуют в РВМС иммунизированных макак-резусов. Для определения антигенспецифических пролиферативных ответов, РВМС собирают через четыре недели после последней вакцинации. Пролиферацию клеток определяют после стимуляции CFSE-меченых РВМС с белками rBmHAT в течение 5 дней и подсчета меченных клеток в проточном цитометре. Эти результаты показывают, что частота пролиферации антиген-отвечающих клеток у иммунизированных животных в 3 раза выше (индекс стимуляции 6,1±0,86) по сравнению с контрольными животными (индекс стимуляции 2,2±1,42). Как и ожидалось, РВМС всех животных демонстрируют сильные пролиферативные ответы (индекс стимуляции 87,4±0) при стимуляции пан-Т митогеном, РНА. РВМС, культивированные в контрольной среде, имеют только низкий уровень пролиферации после 5-дневной инкубации. Значение частоты пролиферации для каждого образца получают путем вычитания контрольного значения только для среды.
[0232] Частоту CFSE-меченых CD3+, CD4+и CD8+РВМС, пролиферирующих в ответ на стимуляцию антигеном, определяют с помощью проточной цитометрии. Эти исследования показывают увеличение пролиферации антиген-отвечающих Т-клеток у всех иммунизированных макак по сравнению с контрольными макаками. Анализ субпопуляции показывает, что у иммунизированных животных приблизительно 12,7% антиген-отвечающих Т-клеток являются клетками CD4+, и 7,9% Т-клеток являются CD8+субпопуляциями. Фоновая пролиферация в присутствии антигена rBmHAT в РВМС контрольных животных составляет 1,4% для клеток CD4+и 2,3% для клеток CD8+.
[0233] Антиген-отвечающие клетки в РВМС иммунизированных обезьян секретируют IFN-γ. Антиген-отвечающие клетки в селезенке мышей и песчанок, иммунизированных rBmHAT, преимущественно секретируют высокие уровни IFN-γ. Таким образом определяют, проявляют ли макаки аналогичный ответ после иммунизации, но до контрольного заражения. Эти исследования показывают, что все РВМС от трех иммунизированных макак (№5242, №5258 и №5259) секретируют значительные количества IFN-(при стимуляции антигеном rBmHAT (таблица 15). Культуральные супернатанты РВМС макаки №4996 и №5254 имеют только фоновые уровни IFN-γ, сходные с уровнями РВМС контрольной макаки.
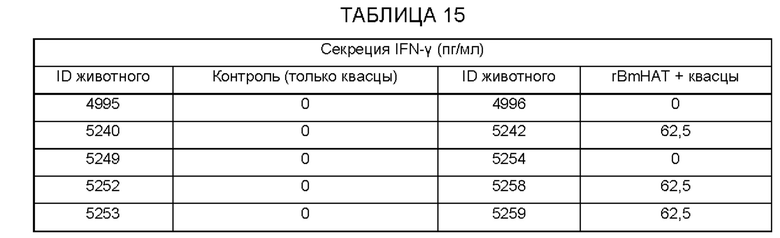
[0234] Анти-rBmHAT антитела в сыворотке иммунизированных макак могут участвовать в уничтожении В. malayi L3. Для определения защитной способности анти-rBmHAT антител в сыворотке иммунизированных макак проводят анализ ADCC in vitro. Результаты показывают, что РВМС вакцинированных макак способны участвовать в уничтожении 35% В, malayi L3 (таблица 16). Когда оценивают сыворотки отдельных макак, максимальный потенциал уничтожения в ADCC составляет 45% в сыворотке макаки №5258. Сыворотки макак №5242 и №5259 также показывают значительный потенциал уничтожения с 38% и 35% уничтожения, соответственно. Сыворотки макак №4996 и №5259 имеют наименьшее свойство ADCC с 25% и 31% уничтожения, соответственно. Когда в этих анализах используют сыворотку контрольных макак, гибели личинок не происходит.
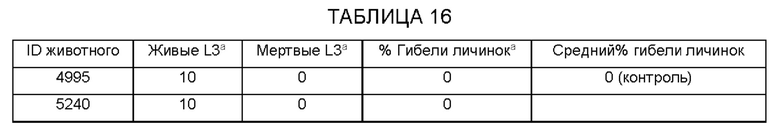
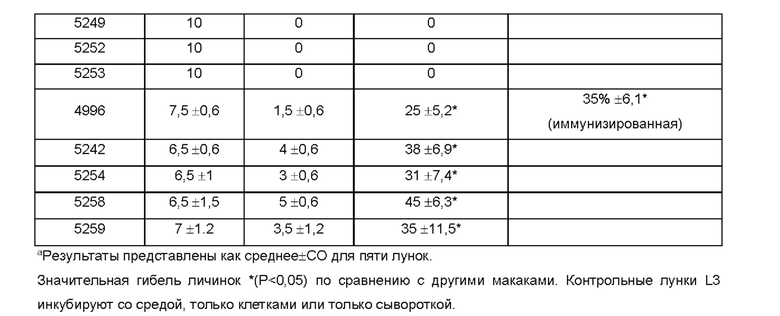
[0235] Иммунизация rBmHAT обеспечивает частичную защиту макак. Через месяц после последней вакцинации всех 10 обезьян контрольно заражают 500 В, malayi L3 и проверяют на появление Mf в периферическом кровообращении. Для обнаружения Mf используют тест Нотта и ПЦР анализ. Тест Нотта проводят еженедельно с 5 недели после контрольного заражения до тех пор, пока животные не становятся положительными. В этих исследованиях зараженные макаки становятся положительными на Mf, начиная с 10 недели после контрольного заражения. В течение 11-20 недель после заражения три контрольных макаки дают положительный результат на Mf. К сожалению, оставшиеся две контрольные макаки остаются отрицательными до конца исследования. В вакцинированной группе три макаки (№5242, №5254 и №5259) остаются отрицательными на протяжении всего исследования. Однако две вакцинированные макаки (№4996 и №5258) являются положительными на Mf. Для дальнейшего подтверждения инфекции проводят ПЦР анализ, где используют антигенспецифические праймеры Hha1 для амплификации на присутствие Mf-специфической ДНК в крови инфицированных обезьян. ПЦР анализ подтвердил инфицирование макак №5249 и №4996. Другие три положительных животных, идентифицированных методом Нотта, являются отрицательными по ПЦР.
[0236] Клетки, отвечающие на rBmHAT, присутствуют в РВМС иммунизированных макак-резусов после заражения. РВМС, собранные через 10 недель после контрольного заражения, стимулируют с rBmHAT для определения антигенспецифического Т-клеточного ответа. РВМС трех животных №5242 (И.С.- 0,928±0,01), №5258 (И.С.- 1,091±0,16) и №5256 (И.С.- 1,0181±0,13) из вакцинированной группы, которые являются отрицательными по Mf, показывают значительную пролиферацию при стимуляции rBmHAT. В то время как у двух вакцинированных животных #4996 (И.С.- 0,258±0,12) и #5254 (И.С.- 0,379±0,03), положительных по Mf, не наблюдается значительной пролиферации при стимуляции rBmHAT. Ни у одного из контрольных животных №4995 (И.С.- 0,280±0,03), 5240 (И.С.- 0,415±0,09), 5249 (И.С.- 0,300±0,26), 5252 (И.С.- 0,507±0,03) или 5253 (И.С.- 0,507±0,03) не наблюдается значительной пролиферации. И.С.РВМС, стимулированных с конканавалином, находится в диапазоне 2,0-3,8.
[0237] Количество эозинофилов является высоким у инфицированных макак, демонстрируя Mf. Микрофиляремические индивидуумы демонстрируют высокое количество эозинофилов в крови (Pearlman, et al. (1993) Exp.Parasitol. 76:200-8; Pearlman, et al. (1993) J. Immunol. 151:4857-64). Подобный результат наблюдается и у макак-резусов. Абсолютное количество эозинофилов определяют за 13, 9 и 5 недель до контрольного заражения, в день контрольного заражения и через 1, 5, 10 и 14 недель после заражения. Результаты показывают увеличение частоты числа эозинофилов в периферической крови микрофиляремийных макак примерно через 10 недель после контрольного заражения. У одной макаки (№5259), которая является отрицательной по Mf, также обнаружена некоторая эозинофилия. Количество эозинофилов в 10 раз выше у контрольных макак с микрофиляриями в периферической крови.
[0238] Высокий титр антигенспецифических IgG антител и повышенная антигенспецифическая секреция IFN-(из РВМС коррелируют с защитой у иммунизированных макак. Поскольку у двух макак в иммунизированной группе обнаружено наличие инфекции после контрольного заражения, иммунные ответы, вызванные вакциной, сравнивают у двух инфицированных макак с аналогичными ответами у трех не инфицированных макак в иммунизированной группе. Сравнивают значения до и после контрольного заражения. Значения перед контрольным заражением устраняют любое смещение из-за контрольного заражения паразитами. Сравнительные иммунологические значения представлены в Таблице 17.
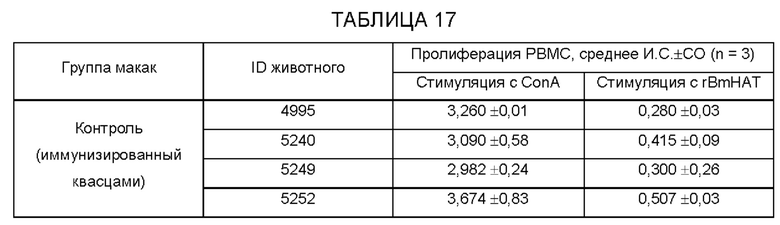
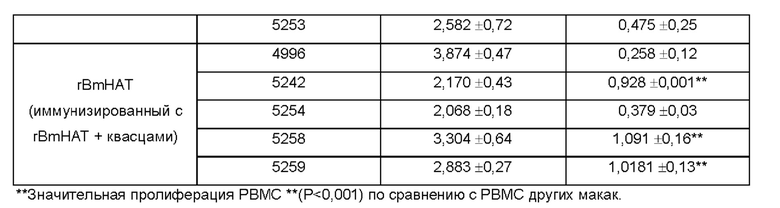
[0239] Результаты показывают, что титр IgG антител значительно выше у трех иммунизированных макак, у которых не развилась инфекция после контрольного заражения. Точно так же, РВМС от тех же трех макак секретируют более высокие уровни IFN-(при стимуляции антигеном rBmHAT. РВМС от двух иммунизированных макак, у которых развилась инфекция после заражения, неспособны секретировать аналогичные уровни IFN-(в ответ на стимуляцию rBmHAT. Анализ ELISPOT проводят с использованием РВМС вакцинированных и контрольных макак. Результаты показывают, что у всех инфицированных макак наблюдается значительное увеличение числа клеток, секретирующих антигенспецифический IL-10, по сравнению с клетками, секретирующими IFN-γ. При сравнении соотношений IFN-(и клеток, секретирующих IL-10, в РВМС иммунизированных макак наблюдается значительное увеличение количества клеток, секретирующих IL-10, у двух вакцинированных макак, у которых обнаружена инфекция (таблица 18). Эти результаты свидетельствуют о четкой корреляции между типом вызванного иммунного ответа и неспособностью установить инфекцию у вакцинированных макак.
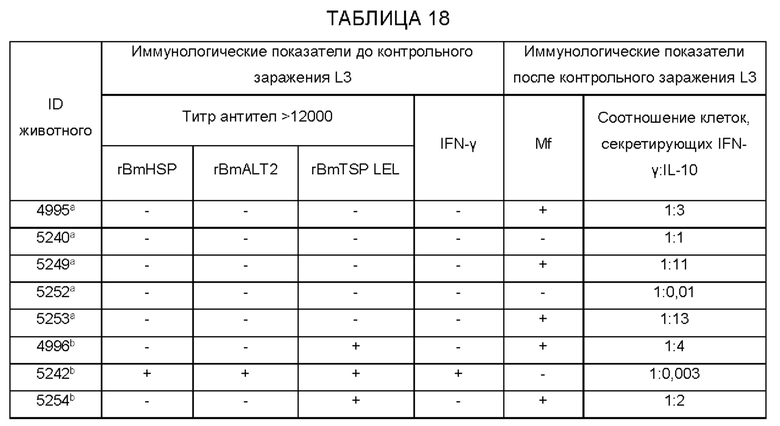
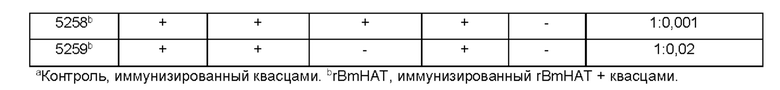
Пример 8: Сравнение валентности
[0240] Испытания с одновалентной, двухвалентной и трехвалетной вакцинацией рекомбинантным белком теплового шока 12,6 (rHSP12.6), обильным личиночным транскриптом-2 (rALT-2) и белками большой внеклеточной петли тетраспанина (rTSP-LEL). Рекомбинантные белки получают, как описано в настоящем документе. Двухвалентные иммуногенные композиции и поливалентные иммуногенные композиции (SEQ ID NO: 70) получают в виде слитых белков. Мышей (N=5) иммунизируют подкожно с использованием схемы белковой примирующей/стимулирующей вакцины. Иммунизированных и контрольных животных заражают живыми инфекционными личинками третьей стадии (L3) В, malayi с использованием способа камеры с микропорами. Через 48 часов после имплантации животных умерщвляют и извлекают камеры из брюшной полости. Содержимое каждой камеры опорожняют, и личинки исследуют под микроскопом при 100Х увеличении для оценки гибели личинок. Результаты этого анализа представлены в таблице 19.
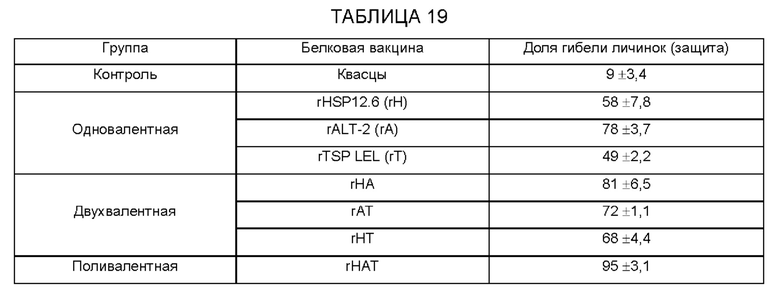
[0241] Результаты показывают, что поливалентная иммуногенная композиция синергетически усиливает защитные иммунные реакции у вакцинированных животных по сравнению с одновалентными и двухвалентными композициями.
[0242] B. malayi не созревает во взрослую форму у мышей. Однако индуцированная вакциной защита от внедрения взрослых червей может быть определена у песчанок. Поэтому одновалентные, двухвалентные и трехвалентные вакцины оценивают у песчанок. Животных (N=10) иммунизируют подкожно с рекомбинантными белками. Животных повторно заражают 100 В. malayi L3, и внедрение червей определяют на 95 день после заражения. Значения доли защиты рассчитывают как долю снижения внедрения червей по сравнению с контрольными песчанками. Результаты этого анализа представлены в таблице 20.
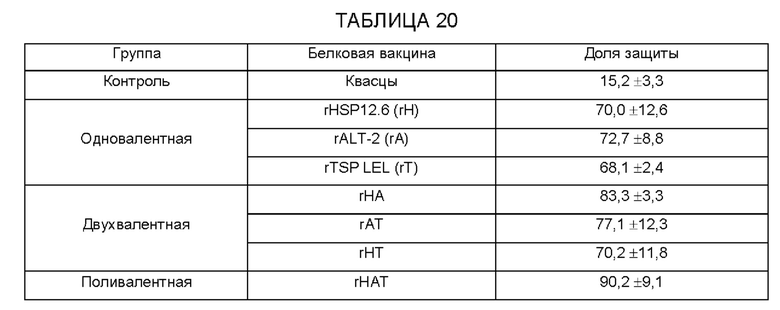
[0243] Результаты показывают, что поливалентная иммуногенная композиция синергетически усиливает защитные иммунные ответы у вакцинированных животных по сравнению с одновалентными и двухвалентными вакцинами.
Пример 9: Сравнение вакцин
[0244] Одновалентные, двухвалентные и поливалентные иммуногенные композиции по данному описанию сравнивают на мышах, песчанках и мастомисах. Животных иммунизируют, как описано, повторно заражают В, malayi L3 и определяли внедрение червей. Результаты этих анализов представлены в таблице 21. Следует отметить, что иммунизация rBmHAX дает 98% защиту у мышей и 97% защиту у песчанок. Эти результаты показывают, что как гвтНАТ, так и гвтНАХ являются отличными кандидатами на вакцины против лимфатического филяриоза.
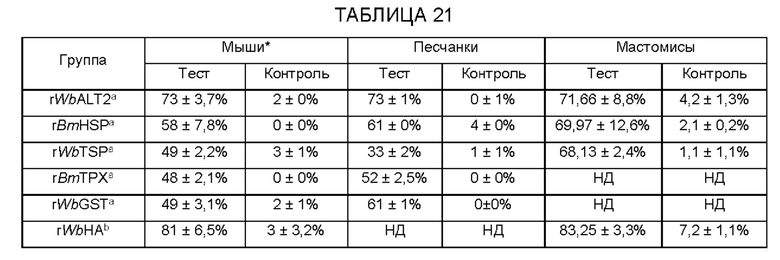
Пример 10: Вакцинный антиген на основе тетравалентного слитого белка (rBmHAXT) против лимфатического филяриоза в модели на мышах
[0245] Клонирование, экспрессия и очистка рекомбинантного белка rBmHAXT. Компания GenScript (Piscataway, NJ) предоставила последовательности bmhsp12.6 (№ доступа GENBNAK AY692227.1), bmalt-2 (№ доступа GENBNAK JF795950.1), bmtpx-2 (№ доступа GENBNAK AF319997.1) и bmtsp. (№ доступа GENBNAK JF795955.1) в векторе pUC57. Гены амплифицируют с использованием прямой 5'-CGGGATCCA TGG AAG AAA AGG TAG TG-3((SEQ ID NO: 31) и обратной 5'-CCC GAA TTC TTA ATG ТТТ CTC AAA ATA TGC TTT-3((SEQ ID NO: 89) с сайтами рестрикции для BamHI и EcoRI. ПЦР-амплифицированные продукты клонируют в вектор экспрессии pRSETA, трансформируют в компетентные клетки BL21 (DE3) Escherichia coli для экспрессии рекомбинантных белков с 6Х гистидиновой меткой. Рекомбинантные слитые белки очищают с использованием аффинной Ni+-заряженной агарозной хроматографической колонки с иммобилизованным металлом, продаваемой под торговым названием SEPHAROSE((GE Healthcare Life Sciences, Pittsburg, PA), и элюируют 300 мМ имидазолом. Эндотоксин в окончательном препарате очищенного белка удаляют с использованием колонки для удаления эндотоксинов (Thermo Fisher Scientific, Rockford, IL). Экспрессию и чистоту рекомбинантных белков подтверждают в 12% SDS-PAGE геле и вестерн-блоттингом с использованием анти-НПэ-антител (Qiagen, Valencia, СА). Концентрацию белка определяют с использованием реактива Бредфорда (Thermo Fisher Scientific).
[0246] Адъюванты, Три различных адъювантных состава используют с рекомбинантным ВтНАХТ. Квасцы (AL007) и квасцы плюс GLA, синтетический агонист TLR4 (AL019), приобретают в Научно-исследовательском институте инфекционных заболеваний, Сиэтл, WA, и маннозилированный хитозан (МСА) подарен компанией Pacific GeneTech, Hong Kong.
[0247] Животные и паразиты. В этих экспериментах используют шести-восьмимесячных мышей Balb/c, приобретенных у Taconic biosciences (Hudson, NY). Использование животных в этом исследовании одобрено комитетом по уходу за животными Иллинойского университета в Рокфорде в соответствии с рекомендациями Национального института здравоохранения по уходу и использованию лабораторных животных. Инфекционных личинок стадии (L3) В. malayi получают из Ресурсного центра исследований реагентов NIAID/NIH (Университет Джорджии, Athens, GA).
[0248] Иммунизация мышей Balb/c. Для иммунизации мышей случайным образом делят на семь групп по пять мышей в группе: (1) rBmHAXT + AL007, вводимый п/к, (2) rBmHAXT + AL019, вводимый п/к, (3) rBmHAXT + МСА (первая доза п/к и стимулирующие дозы вводимые перорально), (4) контроль AL007, вводимый п/к, (5) контроль AL019, вводимый п/к, (6) контроль МСА (первая доза п/к и стимулирующие дозы, вводимые перорально), (7) rBmHAXT + контроль МСА (все дозы вводимые перорально). Каждая мышь получает три дозы по 15 мкг rBmHAXT и 15 мкг соответствующей композиции адъюванта с интервалом в 15 дней.
[0249] Сбор сыворотки, перитонеальной жидкости и селезенки. Образцы крови собирают из поднижнечелюстной вены каждой мыши в 0 день (до иммунизации), и затем через две недели после каждой иммунизации и выдерживают при комнатной температуре в течение 1 часа для образования сгустка. Сыворотку отделяют, и аликвоты хранят замороженными при -80°С для дальнейшего использования. Полость брюшины промывают 500 мкл стерильного солевого раствора, и жидкость собирают у каждой мыши и обрабатывают. Затем у каждого животного собирают селезенку, трижды промывают полной средой RPMI-1640 с добавлением 10% FBS и 1Х раствора антибиотика/микотика (Sigma, St. Louis, МО).
[0250] Титр IgG антител в сыворотке и перитонеальной жидкости. Титры rBmHAXT-специфических IgG антител в образцах сыворотки и перитонеальных жидкостях оценивают с помощью косвенного ELISA. Лунки покрывают 1 мкг/мл rBmHAXT в течение ночи при 4°С. После промывания и блокировки планшетов, добавляют разведенные (1:100, 1:1000, 1:5000, 1:10000, 1:20000 и 1:40000) образцы сыворотки или перитонеальной жидкости и инкубируют в течение 1 часа при комнатной температуре. HRP-конъюгированные антитела курицы против IgG мыши (Thermo Fisher Scientific) используют в качестве вторичных антител, и цвет проявляют с использованием 1-стадийного субстрата Ultra TMB-ELISA (Thermo Fisher Scientific). Реакцию останавливают, используя 0,16 М H2SO4 и определяют оптическую плотность при 450 нм в ридере ELISA BioTek Synergy 2.
[0251] Уровни изотипов антигенспецифических антител в сыворотке и перитонеальной жидкости. Уровни гвтНАХТ-специфических изотипов антител (IgG1, IgG2a, IgG2b, IgG3, IgE, IgM) определяют в образцах сыворотки крови и перитонеальной жидкости с помощью косвенного ELISA. В качестве вторичных антител используют соответствующие изотип-специфические биотинилированные антитела козы против мыши (Sigma) и стрептавидин-HRP (1:20000). Цвет проявляют с помощью 1-стадийного Ultra-TMB. Реакцию останавливают, используя 0,16 М h2so4, и определяют оптическую плотность при 450 нм в ридере ELISA BioTek Synergy 2.
[0252] Исследования контрольного заражения. Для определения защиты, вызванной вакциной, используют способ заражения в камере с микропорами, как описано ранее (Dakshinamoorthy, et al. (2013) Vaccine 31 (12):1616-22). Коротко, 20 L3 В. malayi помещают в камеру с микропорами и хирургическим путем имплантируют в брюшину каждой мыши. Через семьдесят два часа после имплантации камеры с микропорами извлекают. Содержимое каждой камеры опорожняют, личинок подсчитывают и исследуют под микроскопом на прилипание клеток и гибель личинок. Личинок, которые являются прозрачными, прямыми и не двигаются, считают мертвыми. Личинок, которые являются активными, свернутыми в кольцо и полупрозрачными, считают живыми. Долю защиты выражают как количество мертвых паразитов/общее количество извлеченных паразитов, × 100.
[0253] Уровни секретируемых цитокинов в культуральном супернатанте спленоцитов. Готовят одноклеточную суспензию клеток селезенки и стимулируют 1 мкг/мл rBmHAXT или ConA. Не стимулированные клетки селезенки сохраняют в качестве отрицательного контроля для анализа. После 72-часовой инкубации собирают супернатанты культур и определяют уровни IL-2, IL-4, IL-6, IFNy, TNFa, IL-10 и IL-17A с помощью набора для матрицы цитокиновых микроносителей (BD Bio Sciences, San Jose, CA).
[0254] Анализ субпопуляций Т-клеток с помощью проточного цитометра. Клетки селезенки из вышеуказанных культур затем промывают и метят флуоресцентно меченными анти-мышиными CD3 (АРС), CD4 (РЕ) и CD8 (РЕ/суа7) и определяют долю популяции каждого типа клеток в проточном цитометре. Коротко, клетки инкубируют с блокатором рецептора Fcyll в буфере для окрашивания (2% FBS + 0,1% азид натрия) в течение 30 минут при 4°С с последующей промывкой в буфере для окрашивания. К клеткам добавляют все три антитела с флуоресцентной меткой и инкубируют в течение 1 часа при 4°С в темноте. После промывки буфером для окрашивания клетки фиксируют в 4% параформальдегиде и анализируют на проточном цитометре BD FACSCalibur((BD Biosciences).
[0255] Другой набор клеток из вышеуказанного эксперимента окрашивают CD3 (АРС), и в СР3-гейтированной популяции, CD62L (РЕ/Суа7) и CCR7 (РЕ) положительные Т-клетки идентифицируют как Т-клетки центральной памяти. Популяцию клеток также окрашивают на внутриклеточный IFN-((FITC) для определения доли IFN-(положительных Т-клеток центральной памяти.
[0256] Статистический анализ. Представленные данные представляют собой среднее значение ± стандартное отклонение (СО). Статистическая значимость средних различий между различными группами образцов анализируют с использованием непараметрического критерия Крускала-Уоллиса с последующей поправкой Бонферрони для нескольких тестов с использованием программного обеспечения SPSS (v24.0, IBM, NY). Уровень значимости определяют как Р<0,05.
[0257] Титр rBmHAXT-специфических IgG антител. Получают и экспрессируют рекомбинантный белок BmHAXT. На геле SDS-PAGE молекулярная масса rBmHAXT составляет приблизительно 60 кДа и проявляется в виде одной полосы. Уровни эндотоксина в конечных очищенных препаратах составляют <3 ЕЭ/0,1 мг белка. Титр гвтНАХТ-специфических IgG антител является высоким (1:20000) в сыворотках группы rBmHAXT + AL007 и группы rBmHAXT + AL019 (р<0,05). Однако титр меньше (1:10000) в группе rBmHAXT + МСА. В группе rBmHAXT + МСА, где все дозы вводят перорально, имеется очень низкий титр антигенспецифических антител, почти такой же, как в контрольных группах AL007, AL019 и адъюванта МСА. Аналогично, при анализе перитонеальной жидкости наблюдается высокий титр антигенспецифических IgG антител у животных, вакцинированных rBmHAXT + AL019 и rBmHAXT + AL007, по сравнению с группой rBmHAXT + МСА. Титр IgG антител в перитонеальной жидкости меньше по сравнению с соответствующими образцами сывороток тех же животных.
[0258] Изотипы антитела в сыворотке и перитонеальной жидкости. Для определения типа гуморального иммунного ответа, созданного против rBmHAXT, в образцах сыворотки и перитонеальной жидкости, определяют изотипы IgG1, IgG2a, IgG2b, IgG3, IgE, IgM и IgA антител. Результаты показывают, что IgG1 является преобладающим изотипом антител во всех вакцинированных группах (р=0,0001), за исключением группы rBmHAXT + МСА (все пероральные дозы), которая аналогична контролю. Титр IgG2a и IgG2b антител также значительно выше у всех вакцинированных животных (р<0,05) по сравнению с контролем, за исключением группы, вакцинированной rBmHAXT+пероральным МСА. Титр IgG3, IgE, IgM и IgA антител у вакцинированных животных не демонстрирует существенных изменений по сравнению с контролем. Примечательно, что высокие титры IgG1 антител присутствуют в перитонеальных жидкостях животных, вакцинированных rBmHAXT + AL007 и rBmHAXT + AL019 (р=0,0001), по сравнению с подкожной группой rBmHAXT + МСА и контролем. Титр IgG2a и IgG2b антител в перитонеальной жидкости всех вакцинированных животных значительно не отличается от контроля.
[0259] Вакцинация rBmHAXT + AL019 обеспечивает максимальную защиту. Индуцированную вакциной защиту определяют с использованием контрольного заражения способом камеры с микропорами. Результаты показывают, что максимальная защита наблюдается у животных, вакцинированных rBmHAXT + AL019 (88,05±3,9%; р=0,0001), затем rBmHAXT + AL007 (79,47±2,6%; р=0,0001) и rBmHAXT + MCA (78,67±5,47%; р=0,0001). Обнаружено несколько клеток, прикрепленных к поверхности погибших личинок. Эти результаты показывают, что AL019 может быть лучшим адъювантом для rBmHAXT по сравнению с AL007 (р=0,0037) и МСА (р=0,02). Вакцинация rBmHAXT + пероральный MCA дает только 17,97±5,75%, что аналогично контрольным группам садъювантом (AL007, 20,55%; AL019, 24,19% и МСА, 15,742%).
[0260] Клетки селезенки животных, вакцинированных rBmHAXT, секретируют и Th1, и Th2 цитокины. Уровень цитокинов в супернатантах культур клеток селезенки определяют с использованием матрицы цитокиновых микроносителей. Результаты показывают, что секретируемые уровни цитокинов Th1 (IFN-γ, IL-2, IL-6, IL-17A) и Th2 (IL-4 и IL-10) значительно (р<0,05) повышены в супернатантах культур клеток селезенки от животных, вакцинированных rBmHAXT + AL019 и rBmHAXT + AL007, по сравнению с соответствующими контрольными животными с адъювантом. Супернатанты культуры селезенки от животных, вакцинированных rBmHAXT + MCA, имеют значительно более высокие уровни IFN-γ (р=0,01) и IL-6 (р=0,0001) по сравнению с соответствующим адъювантным контролем. Однако не выявлено существенной разницы в уровнях других измеренных цитокинов. Уровни цитокинов в супернатантах культур из группы rBmHAXT + пероральный МСА аналогичны контрольным адъювантам.
[0261] Клетки Тем создают в селезенке животных, вакцинированных rBmHAXT. Клетки селезенки культивируют при 37°С в течение 72 часов, стимулируют 1 мкг/мл белка rBmHAXT. Через 72 часа клетки собирают и окрашивают CD3/CD4/CD8 антителами, и оценивают с помощью проточного цитометра. Наблюдается небольшое, но значительное увеличение популяции клеток CD8+ в группе, леченной rBmHAXT + AL019 (р<0,05), по сравнению с другими группами. Для определения доли клеток Тем в селезенке спленоциты окрашивают CD62L/CCR7 антителами и анализируют в проточном цитометре. Клетки, которые являются дважды положительными по CD62L/CCR7, считают Тем клетками. Результаты показывают, что у животных, леченных rBmHAXT, имеется высокая доля клеток Тем независимо от используемого адъюванта (р<0,001).
[0262] Клетки Тем являются преимущественно IFNγ*. Считается, что клетки Тем, секретирующие IFN-γ, играют основную роль в индуцированной вакциной защите от паразитарных инфекций (Maggioli, et al. (2016) Front. Immunol. 7:421). Поэтому измеряют долю CD62L+ CCR7 TCM клеток, которые экспрессируют внутриклеточный IFN-γ. Результаты показывают, что клетки животных, вакцинированных rBmHAXT + AL019, имеют значительно (р<0,01) высокую долю IFNγ+ TCM клеток по сравнению с группами, вакцинированными rBmHAXT + AL007 и rBmHAXT + МСА.
Пример 11: Профилактическая вакцина против лимфатического филяриоза человека у приматов, отличных от человека
[0263] Приматы, отличные от человека. Сорок самцов или самок здоровых макак-резусов (в возрасте от 3 до 5 лет) приобретают у PrimGen (Hines, IL) и размещают на объекте Bioqual в Rockville, MD. Всех животных перед включением в исследование подвергают скринингу на отсутствие филяриозных инфекций путем анализа крови на наличие микрофилярий Hha-1 с помощью ПЦР (Hoti, et al. (2003) Acta Trop. 88:77-81; Rao, et al. (2006) J. Clin. Microbiol. 44:3887-3893) и сыворотки на наличие антител против rBmSXP-1 (Vasuki, et al. (2003) Acta Trop. 86:109-114; Abdul Rahman, et al. (2007) Filaha J. 6:10) и белки rBmHAXT анализируют с помощью твердофазного иммуноферментного анализа (ELISA). Животные, у которых был положительный результат на любой из белков, не включались в исследование.
[0264] Паразиты. Инфекционные личинки третьей стадии Brugia malayi (L3) получают из Ресурсного центра исследований реагентов NIAID/NIH (Университет Джорджии, Athens, GA).
[0265] Адъюванты. В этом исследовании сравнивают два разных адъюванта. Квасцы (AL007) и квасцы плюс синтетический агонист TLR4 GLA (AL019) приобретают в Научно-исследовательском институте инфекционных заболеваний, Seattle, WA.
[0266] Клонирование и экспрессия поливалентных рекомбинантных белков. Белок rBmHAT экспрессируют в Escherichia coli BL21 (DE3), очищают и анализируют, как описано в настоящем документе. Кодирующую последовательность (CDS) поливалентного слитого белка rBmHAT (состоит из bmhsp 12.6, bmalt-2 и bmtsp) и rBmHAXT (состоит из bmhsp 12.6, bmalt-2, bmtpx2 и bmtsp) синтезируют в GenScript (Piscataway, NJ). Последовательности представлены в векторе pUC51. Оба CDS подвергали ПЦР-амплификации с использованием одних и тех же ген-специфических праймеров (прямой праймер: 5'-CGG GAT CCA TGG AAG AAA AGG TAG TG-3', SEQ ID NO: 31 и обратный праймер: 5'-CGG AAT TCT CAA TCT TTT TGA GAT GAA T-3', SEQ Dl NO: 36) с сайтами рестрикции для BamHI и EcoRI и клонируют в вектор экспрессии pRSETA (Invitrogen, Carlsbad, СА) с 6Х гистидиновой меткой. Лигированные конструкции для bmhat и bmhaxt далее трансформируют в штамм экспрессии Е. coli BL21 (DE3). Экспрессию рекомбинантных белков индуцируют 1 мМ IPTG. Рекомбинантные белки очищают с помощью аффинной колоночной хроматографии на никеле (GE Healthcare Life Sciences, Pittsburg, PA), и чистоту рекомбинантных белков подтверждают в 12% геле SDS-PAGE и вестерн-блоттингом с использованием анти-пента His антител (Qiagen, Velencia, СА). Эндотоксин в конечном препарате удаляют с помощью колонки для удаления эндотоксинов (Thermo Fisher Scientific, Rockford, IL). Конечную концентрацию белков rBmHAT и rBmHAXT определяют с помощью анализа Бредфорда (Qiagen).
[0267] Иммунизация макак. Зона представляет собой двойное слепое испытание вакцины. В общей сложности 40 макак были случайным образом разделены на три лечебные группы и одну контрольную группу по 10 макак в каждой. Все леченные животные получают четыре дозы по 150 мкг вакцинного антигена и 2 мг адъюванта в дни 0, 28, 56 и 84. Группа лечения 1 получает rBmHAT + квасцы, группа лечения 2 получает rBmHAT + AL019 и группа лечения 3 получает rBmHAXT + AL019. Контрольные животные получают только адъювант AL019. Места инъекций ежедневно отслеживают на наличие любых побочных реакций (покраснение, отек и т.д.) в течение до 7 дней после иммунизации. Образцы крови собирают перед каждой иммунизацией и перед контрольным заражением.
[0268] Подсчет клеток, биохимический анализ сыворотки и общий анализ крови (СВС). Общий анализ крови и биохимический анализ сыворотки анализируют с использованием коммерческих автоматических анализаторов гематологии и биохимического анализа сыворотки IDEXX. Образцы, собранные до начала исследования, служат нормальным исходным эталоном для каждого животного.
[0269] Уровни антигенспецифических антител в сыворотке макак. Определяют уровни rBmHAT-, rBmHAXT-, rBmHSP-, rBmALT-2-, rBmTPX- или rBmTSP-специфических общих IgG, IgG1, IgG2, IgG3, IgM и IgE антител в сыворотке каждой макаки-резуса с использованием косвенного ELISA, как описано в другом месте настоящего документа.
[0270] Анализ антителозависимой клеточно-опосредовэнной
цитотоксичности (ADCC). Анализ ADCC выполняют, как описано в другом месте настоящего документа. Приблизительно по десять живых В. malayi L3 инкубируют при 37°С с 5% СО2 в трех лунках вместе с 2×105 РВМС и 50 мкл образцов сыворотки. Через семьдесят два часа после инкубации определяют жизнеспособность В. malayi L3. Долю гибели личинок выражают как отношение количества погибших L3 к общему количеству, извлеченному из каждой лунки, умноженное на 100.
[0271] Супернатант культуры из анализа ADCC также собирают для определения уровня активности миелопероксидазы (МРО) с использованием набора, приобретенного у Biovision (Milpitas, СА), и значения выражают в мЕд/минуту на мл супернатанта культуры.
[0272] Антигенспецифическая пролиферация мононуклеарных клеток периферической крови (РВМС), РВМС (1×107 клеток/лунку в 1 мл) инкубируют при 37°С в темноте в течение 15 минут с 5 мМ сукцинимидилового эфира карбоксифлуоресцеин диацетата (CFSE). После инкубации клетки промывают, инкубируют в течение дополнительных 30 минут при 37°С и высевают по 2×106 клеток/лунку в 1 мл в 24-луночный планшет для тканевых культур. На следующий день удаляют пятьсот мкл среды и в лунки добавляют rBmHAXT (1 мкг в 500 мкл). Клетки, инкубированные только со средой RPMI, служат в качестве отрицательного контроля, и клетки, стимулированные конканавалином А (1 мг/лунку), служат в качестве положительного контроля. Клетки собирают на пятый день после культивирования, метят анти-CD3-АРС антителом, фиксируют в 4% параформальдегиде в течение 10 минут при комнатной температуре, и данные получают на проточном цитометре BD FACSCalibur(и анализируют с использованием программного обеспечения ModFit LT (Verity Software House, Topsham, ME).
[0273] Собирают параллельные культуры клеток, инкубированные в течение 3 дней, фиксируют параформальдегидом и метят комбинациями анти-CD3-АРС, анти-CD4-FITC, анти-CD8-РЕ, анти-CD28-РЕ или анти-CCR7-FITC. Внутриклеточный IFN-γ и IL-4 определяют после фиксации и пермеабилизации. Данные получают на проточном цитометре BD FACSCalibur(и анализируют с использованием Flow Jo v10.1 (FlowJo, LLC, Ashland, OR).
[0274] Секретируемые уровни цитокинов (GM-CSF, IFN-γ, IL-12p70, IL-1β, IL-4, IL-5, IL-6, IL-15, IL-16 и TNF-α) в супернатантах клеточных культур измеряют с помощью основанной на антителах матрицы Rhesus Cytokine Quantibody Array GS1 (RayBiotech, Inc., Norcross, GA) в соответствии с протоколом производителя. Интенсивность сигналов флуоресценции из матриц предметных стекол сканируют, и данные анализируют после вычитания фоновых сигналов и нормализации до положительных контролей.
[0275] Контрольное заражение инфекционными личинками В. malayi (L3), Через месяц после последней иммунизации всем макакам подкожно вводят 130-180 В. malayi L3. Всех личинок исследуют под микроскопом на жизнеспособность, подсчитывают, и только жизнеспособные личинки используют для контрольного заражения. После контрольного заражения, всех макак регулярно отслеживают на наличие любых возможных изменений в панели клинической биохимии (биохимии сыворотки, гематологии, общего анализа крови и количества CD4+/CD8+ Т-клеток), физических параметрах (признаков побочных реакций в месте контрольного заражения, массы тела, температуры тела, состояния тела, лимфедемы и измерения лимфатических узлов) и модели поведения.
[0276] Подтверждение возникновения контрольной инфекции у макак. Практически трудно подсчитать количество взрослых червей, поселившихся в каждой макаке. Однако для подтверждения наличия активной инфекции у макак используют несколько доказательств, включая микроскопию и ПЦР анализ Hha-I. Для микроскопического анализа, на 5, 10, 15 и 18 неделе после заражения, 10 мл крови собирают у каждой макаки между 18-22 часами вечера и подвергают скринингу на наличие микрофилярий с использованием модифицированного метода Нотта. Для ПЦР анализа Hha-I, ДНК, выделенную из 200 мкл образцов крови с использованием набора геномной ДНК крови Gen Elute (Sigma-Aldrich), подвергают ПЦР амплификации на гены тандемных повторов Hha-I, как описано ранее (Hoti, et al. (2001)). Bull. Entomol. Res. 97:87-92) и амплифицированные продукты ПЦР секвенируют для подтверждения последовательности Hha-I.
[0277] Титр анти-rBmSXP-l антител в сыворотке. SmSXP-1 является высокочувствительным и специфичным диагностическим антигеном для инфекций В. malayi. Как правило, однополая инфекция не вызывает какой-либо Mf в периферическом кровообращении. Таким образом, микроскопия и метод ПЦР могут не выявить эти латентные инфекции. Однако наличие активного червя у инфицированных субъектов или животных может быть подтверждено определением титра антител IgG4 против антигена SmSXP-1. Для определения титра этих антител в сыворотке макак используют ELISA. Поскольку не удается получить надежные коммерческие антитела против IgG4 макаки, титр анти-remSXP-1 IgG антител используют в качестве маркера для выявления латентных инфекций.
[0278] Лимфосцинтиграфический анализ. Лимфосцинтиграфический анализ проводят, как описано в другом месте настоящего документа.
[0279] Статистический анализ. Представленные данные представляют собой среднее значение ± стандартное отклонение (СО). Статистическую значимость средних различий между различными группами выборок анализируют с использованием непараметрического критерия Крускала-Уоллиса с последующей поправкой Бонферрони для нескольких тестов с использованием программного обеспечения SPSS (v24.0, IBM, NY). Уровень значимости определяют как Р≤0,05. Для анализа защиты, вызванной вакциной, используют критерий хи-квадрат для сравнения пропорций между группами и точный критерий Фишера, где это уместно. Отношение шансов (OD) рассчитывают для определения различий между группами.
[0280] Иммунизация вакцинами-кандидатами создает высокий титр антигенспецифических IgG антител и их изотипов. Молекулярная масса rBmHAT составляет приблизительно 39 кДа, и для rBmHAXT составляет приблизительно 60 кДа. Оба белка готовят с чистотой более 95% с уровнями эндотоксина <10 ЕЭ/мкг белка. Иммунизация макак очищенными белками создает высокий титр антигенспецифических IgG антител. IgG антитела являются специфичными к каждому компоненту слитого белка. Максимальный титр IgG антител достигается после второй дозы иммунизации (таблица 22), что указывает на то, что двух доз вакцины достаточно для достижения максимального титра антител.
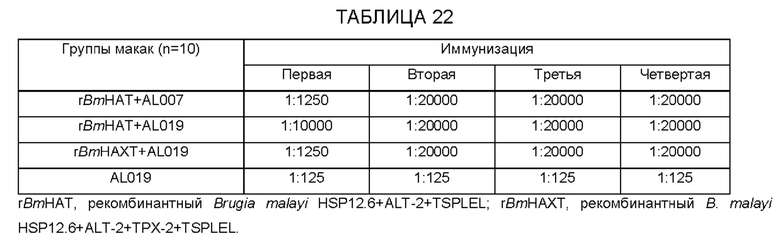
[0281] IgG1 и IgG2 являются наиболее преобладающим изотипом IgG антител в сыворотке всех иммунизированных животных. Уровни IgG3 антител значительно повышены в сыворотке группы, иммунизированной rBmHAXT плюс AL019, по сравнению с контрольными животными. IgM и IgE антитела значительно не отличаются от контроля в сыворотке вакцинированных животных.
[0282] Антиген-отвечающие клетки памяти присутствуют в крови вакцинированных макак. Индекс пролиферации РВМС от вакцинированных животных является высоким в ответ на соответствующие антигены по сравнению с контролем. Клетки из контрольной группы реплицируют только один раз в течение пяти дней в культуре по сравнению с вакцинированной группой, где клетки делятся до восьми поколений. Оценка популяции клеток памяти в делящихся антиген-отвечающих CD4+ и CD8+ Т-клетках показывает, что как CD28+CCR7- Т-клетки эффекторной памяти (ТЕМ), так и CD28+CCR7+ Т-клетки центральной памяти (ТСМ) селективно размножены у макак, иммунизированных rBmHAXT плюс AL019 и rBmHAT плюс AL007. Тем клетки преимущественно являются положительными по внутриклеточному IFN-γ, и Тем клетки преимущественно являются положительными по внутриклеточному IL-4. Анализ супернатантов культур РВМС показывает заметное увеличение секретируемых уровней цитокинов (GM-CSF, IFN-γ, IL-12р70, IL-1β, IL-4, IL-5, IL-6, IL-15, IL-16 и TNF-α) по сравнению с контролями AL019. РВМС от животных, вакцинированных rBmHAXT + AL019, секретируют почти в 10 раз более высокие уровни IFN-(по сравнению с контрольными животными AL019.
[0283] Иммунизация с помощью rBmHAXT обеспечивает максимальную защиту. Через десять недель после заражения В. malayi L3, у семи из 10 контрольных животных (70%) в периферической крови обнаруживают Mf, и они продолжают являться положительными по Mf до завершения эксперимента (18 недель после заражения). Однако только у 3 из 10 макак (30%) в группе rBmHAXT плюс AL019 Mf обнаружен в крови (ФИГ. 4). Эти открытия дополнительно подтверждены ПЦР анализом образцов крови на присутствие В. malayi-специфического Hha-1 и ELISA на IgG антитела против антигена SXP-1. Повторное исследование крови не выявляет каких-либо признаков инфицирования ни у одного из отрицательных животных до завершения эксперимента (через 18 недель), что указывает на то, что иммунизация rBmHAXT плюс AL019 обеспечивает 57,14% защиту после корректировки 30% амикрофиларемических макак в контрольной группе (р=0,073, отношение шансов = 0,18). Пять из 10 животных в группе rBmHAT+AL019 (р=0,649, отношение шансов = 0,42) и 7 из 10 животных в группе rBmHAT+AL007 (р=1,0, отношение шансов = 1,0) являются положительными по Mf. Статистическая корреляция между различными группами показывает, что rBmHAXT+AL019 обеспечивает лучшую защиту, чем другие вакцинированные группы и контроль AL019 (rBmHAXT+AL019 к контролю AL019 - р=0,073; rBmHAXT+AL019 к rBmHAT+AL019 - р=0,649; rBmHAXT+AL019 к rBmHAT+AL007 -р=0,073 и rBmHAT+AL019 к rBmHAT+AL007 - р=0,649). Таким образом, на основании отношения шансов вероятность того, что контрольная группа AL019 будет положительной по Mf, в 2,33 раза выше, чем вероятность того, что группа rBmHAT + AL019 будет положительной по Mf. Вероятность того, что контрольная группа AL019 будет положительной по Mf, в 5,43 раза выше, чем вероятность того, что группа rBmHAXT+AL019 будет положительной по Mf. Вероятность того, что группа rBmHAT+AL019 будет положительной по Mf, в 1,95 раза выше, чем вероятность того, что группа rBmHAXT+AL019 будет положительной по Mf. Эти статистические анализы показывают, что защита, обеспечиваемая иммунизацией rBmHAXT+AL019, значительно лучше, чем иммунизация rBmHAT+AL007 и rBmHAT+AL019.
[0284] Анализ антителозависимой клеточной цитотоксичности (ADCC) также проводят в качестве in vitro суррогата для определения защиты, индуцированной вакциной. Эти анализы также подтверждают, что образцы сыворотки макак, вакцинированных rBmHAXT+AL019, более эффективно уничтожают В. malayi L3 in vitro по сравнению с образцами сыворотки двух других вакцинированных групп (ФИГ. 5). Обнаружено несколько клеток, прикрепленных к мертвым личинкам. Фактически, многие мертвые личинки полностью покрыты клетками в течение 24 часов после инкубации. В этом исследовании морфология клеток не идентифицирована. Однако предыдущие исследования предполагают, что связанные клетки представляют собой преимущественно моноциты. Чтобы определить статус активации этих клеток, измеряют уровни миелопероксидаз (МРО) в супернатантах культур из этих анализов ADCC. Результаты показывают увеличение уровня секретируемых МРО (от 2,08 до 2,78 ммоль/мин/мл) в лунках с сывороткой вакцинированных животных и РВМС по сравнению с лунками с сывороткой контрольных животных (1,99 ммоль/мин/мл), что указывает на активацию клеток, продуцирующих МРО.
[0285] У Mf-отрицательных вакцинированных животных не развивается лимфатическая патология. Взрослые филяриозные паразиты, живущие в лимфатических сосудах, вызывают сильное воспаление, приводящее к лимфедеме и блокированию тока лимфы. Наличие лимфатической блокады в этом исследовании определяют с помощью лимфосцинтиграфии. Лимфоток сравнивают между правой и левой ногой одного и того же животного. Контрольное заражение инфекциями проводят на правой ноге. Результаты показывают, что на 16 неделе после контрольного заражения наблюдается значительное снижение лимфотока в правой ноге всех Mf-положительных животных по сравнению с их левой ногой, что указывает на блокаду лимфы. Лимфоток не показывает каких-либо существенных различий между правой и левой ногой у всех Mf-отрицательных животных в вакцинированной группе. Эти результаты, таким образом, демонстрируют, что лимфатическая патология является минимальной или отсутствует у вакцинированных и контрольно зараженных макак, у которых не обнаруживается Mf в периферической крови.
Пример 12: Профилактическая вакцина против инфекции Dirofilaria immitis у собак
[0286] Протокол иммунизации. Шесть собак делят на две группы по три животных в каждой. Каждому животному первой группы вводят три цикла по 100 мкг вакцины гвтНАХТ плюс 400 мкл адъюванта AL019 (40 мкг агониста TLR4 + 800 мкг квасцов) п/к. Каждое животное второй группы используют в качестве контроля, и оно получает три цикла по 400 мкл только адъюванта AL019 п/к. Инъекции осуществляют в 0, 30 и 60 дни. Кроме того, в 0, 30, 60 и 90 дни, 20 мл крови собирают в пробирки с ЭДТК из подкожной вены каждой собаки перед иммунизацией. Всех животных отслеживают на наличие побочных реакций, включая реакции в месте инъекции, лихорадку, потерю аппетита, аллергию, выпадение волос и потерю веса.
[0287] Уровни антигенспецифических антител в сыворотке собак. Уровни rBmHAXT-специфических общих IgG, IgG1, IgG2, IgG3, IgM и IgE антител определяют в сыворотке каждой собаки с использованием косвенного ELISA, как описано в другом месте настоящего документа.
[0288] Анализ антителозависимой клеточно-опосредованной цитотоксичности (ADCC), Анализ ADCC проводят, как описано в другом месте настоящего документа. Каждую из восьми-десяти живых D. immitis L3 инкубируют при 37°С с 5% СО2 в дублированных лунках вместе с 0,5 миллиона РВМС, выделенных из нормальной крови собаки, 200 мкл среды RPMI 1640 и 100 мкл образцов сыворотки. Планшеты отслеживают под световым микроскопом каждые 24 часа на жизнеспособность D. immitis L3. Личинки, которые являются прозрачными, неподвижными или малоподвижными, считают мертвыми. Долю гибели личинок выражают как отношение количества погибших L3 к общему количеству, извлеченному из каждой лунки, умноженное на 100.
[0289] Иммунизация вакциной rBmHAXT создает высокий титр антигенспецифических IgG антител и их изотипов. Иммунизация собак очищенным rBmHAXT белком создает высокий титр антигенспецифических IgG антител (более 1:20000). Также наблюдается значительное увеличение IgG1, IgG2 и IgA антител в сыворотке крови собак после трех иммунизации по сравнению с контрольной группой, где IgG1 и IgG2 являются наиболее преобладающим изотипом IgG антител. Анализ образцов сыворотки на наличие защитных антител против инфекционных личинок D. immitis показывает, что в сыворотке вакцинированных животных присутствуют значительные уровни защитных антител памяти. Эти защитные антитела пролиферируют в ответ на rBmHAXT и способны убивать как чувствительных к лекарственным средствам (~60%), так и резистентных к лекарственным средствам (~85%) инфекционных личинок после двух-трех иммунизации (ФИГ. 6). Обнаружено несколько клеток, прикрепленных к мертвым личинкам. Кроме того, супернатанты культур клеток селезенки вакцинированных собак показывают повышенные уровни TNF-α и IL-10. Эти данные свидетельствуют о том, что rBmHAXT представляет собой отличную вакцину-кандидат против дирофиляриоза сердца у животных, таких как собаки и кошки.
[0290] Всех собак заражают п/к 50 чувствительными к лекарству инфекционными личинками D. immitis. Через шесть месяцев после контрольного заражения всех животных подвергают эвтаназии для извлечения внедрившихся взрослых червей у каждой собаки. Результаты показывают, что среднее количество червей у животных, вакцинированных rBmHAXT, составляет 7±1,73 по сравнению с контрольной группой AL019, в которой среднее количество червей составляет 29,3±3,51. Это исследование демонстрирует, что вакцинация собак rBmHAXT обеспечивает защиту на 76,11% от заражения чувствительными к лекарственному средству D. immitis. После контрольного заражения в собак, вакцинированных rBmHAXT, внедряется меньше самцов и самок червей.
Пример 13: Сравнение вакцин rBmHAXT и rDiHAX против инфекции Dirofilaria immitis у мышей
[0291] Клонирование, экспрессия и очистка рекомбинантного белка rBmHAXT. Компания GenScript (Piscataway, NJ) предоставила последовательности bmhsp12.6 (№ доступа GENBNAK AY692227.1), bmalt-2 (№ доступа GENBNAK JF795950.1), bmtpx-2 (№ доступа GENBNAK AF319997.1) и bmtsp (№ доступа GENBNAK JF795955.1) в векторе pUC57. Гены амплифицируют с использованием прямой 5'-CGGGATCCA TGG AAG AAA AGG TAG TG-3((SEQ ID NO: 31) и обратной 5'-CCC GAA TTC TTA ATG TTT CTC AAA ATA TGC TTT-3((SEQ ID NO: 89) с сайтами рестрикции для BamHI и EcoRI, ПЦР-амплифицированные продукты клонируют в вектор экспрессии pRSETA, трансформируют в компетентные клетки BL21 (DE3) Е. coli для экспрессии рекомбинантных белков с 6Х гистидиновой меткой. Рекомбинантные слитые белки очищают с использованием аффинной Ni+-заряженной агарозной хроматографической колонки с иммобилизованным металлом, продаваемой под торговым названием SEPHAROSE((GE Healthcare Life Sciences, Pittsburg, PA), и элюируют 300 мМ имидазолом. Эндотоксин в конечном препарате очищенного белка удаляют с использованием колонки для удаления эндотоксинов (Thermo Fisher Scientific, Rockford, IL). Экспрессию и чистоту рекомбинантных белков подтверждают с помощью 12% SDS-PAGE геля и вестерн-блоттинга с использованием анти-Нэ-антител (Qiagen, Valencia, СА). Концентрацию белка определяют с использованием реактива Бредфорда (Thermo Fisher Scientific).
[0292] Клонирование, экспрессия и очистка рекомбинантного белка rDiHAX, Непрерывную нуклеотидную последовательность DiHSP 12.6 (ген = nDi.2.2.2.g00663), DiALT-2 (ген = nDi.2.2.2.g08197) и DiTPX-2 (ген = nDi.2.2.2.g06574) получают от Wormbase ParaSite путем бластинга последовательностей BmHSP 12.6, BmALT-2 и BmTPX-2. Обнаружено, что белки BmHSP 12.6, BmALT-2 и BmTPX-2 имеют 97%, 63% и 96% сходство последовательности с белками DiHSP 12.6, DiALT-2 и DiTPX-2, соответственно. Нуклеотидная и аминокислотная последовательности DiHSP представлены в SEQ ID NO: 90 и SEQ ID NO: 91, соответственно. Нуклеотидная и аминокислотная последовательности DiALT-2 представлены в SEQ ID NO: 92 и SEQ ID NO: 93, соответственно. Нуклеотидная и аминокислотная последовательности DiTPX представлены в SEQ ID NO: 94 и SEQ ID NO: 95, соответственно. Нуклеотидные последовательности DiHSP 12.6, DiALT-2 и DiTPX-2 линейно объединяют, и полученный ген dihax синтезируют с помощью Invitrogen Life Technologies™. Химерный ген (SEQ ID NO: 96) представляют в векторе рЕТ100/D-TOPO® (ThermoFisher Scientific, Rockford, IL), трансформированном в компетентную BL21*DE3 Е. coli и экспрессируют слитый белок rDiHAX (SEQ ID NO: 97). Коротко, ночную посевную культуру инокулируют в 500 мл стерильного бульона LB и позволяют расти в оптимизированных условиях и давлении селекции до тех пор, пока не будет достигнута OD6oo. Затем бактериальные клетки индуцируют 1 мМ IPTG (Research Products International, Mt. Prospect, IL) и оставляют расти еще в течение 4 часов. Затем клетки пеллетируют центрифугированием при 12000 об/мин в течение 30 минут при 4°С. Для очистки белка осадок ресуспендируют в 20 мл Tris-забуференного солевого раствора (TBS), затем к раствору добавляют 150 мкл лизоцима и инкубируют в течение 30 минут на платформе шейкера при комнатной температуре. После инкубации осадок обрабатывают ультразвуком в течение 4 циклов по 1 минуте каждый с 30-секундной паузой между ними. После обработки ультразвуком лизаты центрифугируют при 12000 об/мин в течение 30 минут при 4°С. Супернатант отбрасывают, осадок промывают, затем к осадку добавляют 15 мл 8М мочевины и инкубируют в течение ночи при 4°С при постоянном перемешивании. После инкубации лизат центрифугируют при 12000 об/мин в течение 30 минут при 4°С и супернатант собирают в свежую 50 мл коническую центрифужную пробирку над льдом. Рекомбинантный белокэкспрессируютс N-концевой меткой из шести гистидиновых остатков. Это позволяет очистить белок rDiHAX аффинной хроматографией с использованием иммобилизованного металла (IMAC). В частности, экстрагированный белок инкубируют с 2 мл кобальтовой смолы в течение 30 минут на шейкере при комнатной температуре и упаковывают в 10 мл колонку. После промывки колонки 10 мл TBS, колонку промывают 30 мл 10 мМ имидазола, приготовленного в TBS. Затем связанный белок элюируют 300 мМ имидазола, содержащего 10% глицерина в TBS. Чистоту и молекулярный размер белка rDiHAX оценивают на 14% геле SDS-PAGE. Молекулярная масса рекомбинантного белка составляет приблизительно 60 кДа. После обессоливания эндотоксин из очищенного белка удаляют путем пропускания раствора белка через высокопроизводительную колонку со смолой для удаления эндотоксина (ThermoFisher Scientific). Уровень эндотоксина в образце концентрированного белка анализируют с использованием набора для количественного определения хромогенного эндотоксина Pierce™ LAL. Конечное количество эндотоксина в очищенном препарате rDiHAX составляет 3 ЕЭ на мг белка.
[0293] Дизайн эксперимента. Мышей Balb/c (самцы в возрасте 4-6 недель) объединяют в группы по 10 мышей в каждой группе и иммунизируют подкожно 15 мкг rBmHAXT или rDiHAX антигеном вместе с 10 мкг AL019 (квасцы плюс GLA, синтетический агонист TLR4) в качестве адъюванта. Проводят четыре иммунизации с интервалом в 2 недели. Контрольные животные получают только адъювант AL019. Образцы крови собирают у каждой мыши перед каждой иммунизацией и через 2 недели после последней иммунизации для анализа сывороточных уровней антигенспецифических IgG, IgG1, IgG2a, IgG2b и IgG3. Анализ ADCC проводят путем инкубации 10-15 D. immitis L3 с 50 мкл сыворотки иммунизированных мышей и 1×105 клеток брюшины контрольных мышей. Также проводят контрольный эксперимент, помещая камеру с микропорами, содержащую 15-20 D. immitis L3, в брюшную полость всех мышей. Жизнеспособность личинок определяют через 72 часа после заражения, и клетки селезенки и перитонеальную жидкость/клетки анализируют на предмет иммунологических коррелятов защиты, индуцированной вакциной.
[0294] Иммунизация вакцинами rBmHAXT и rDiHAX создают высокие титры антигенспецифических IgG антител и их изотипов. Иммунизация мышей очищенными белками rBmHAXT и rDiHAX создает высокий титр антигенспецифических IgG антител. В частности, результаты показывают, что у иммунизированных животных развиваются высокие титры (титр 1:10000) IgG1, IgG2a, IgG2b и IgG3 антител против rBmHAXT или rDiHAX по сравнению с контролями (р<0,05).
[0295] Результаты экспериментов ADCC показывают, что образцы сыворотки мышей, иммунизированных rBmHAXT, убивают 93±8,83% личинок, и образцы сыворотки мышей, иммунизированных rDiHAX, убивают 76±5,69% личинок по сравнению с образцами сыворотки из группы AL019, которые вызывают (20±5,93%) гибель личинок. В эксперименте с контрольным заражением гибель личинок составляет 83±4,14% и 71±8,99% для мышей, иммунизированных rBmHAXT и rDiHAX, соответственно, по сравнению с контролем (7,3±2,42%). Примечательно, что наблюдается значительная (р<0,05) перекрестная реактивность антител в образцах сыворотки животных, вакцинированных rBmHAXT и rDiHAX с белками rBmHAXT и rDiHAX, определенная с помощью ELISA и вестерн-блоттинга.
[0296] Анализ клеточного иммунного ответа показывает, что в селезенке вакцинированных животных наблюдается увеличение антигенспецифических CD3+CD62L+CCR7+ Т-клеток памяти по сравнению с контрольными AL019. Супернатанты культур клеток селезенки из обоих групп rBmHAXT и rDiHAX показывают повышенные уровни IL-17A, IL-6, IFN-γ и IL-10.
Пример 14: Внутримышечная инъекция rBmHAXT обеспечивает защиту от инфекции Dirofilaria immitis у собак
[0297] Протокол иммунизации. Шесть собак были делят на две группы по три животных в каждой. Каждое животное первой группы получает три раунда по 100 мкг вакцины rBmHAXT плюс 40 мкг GLA-SE, адсорбированной на квасцах (AL019; лиганд TLR4, GLA в виде эмульсии масло-в-воде) в дни 0, 28 и 56, в/м, в область левого бока. Каждое животное второй группы используется в качестве контроля и получает три раунда только адъюванта на 0, 28 и 56 дни, в/м, в область левого бока. Кроме того, в -1, 0, 28, 56 и 84 дни образцы крови собирают в пробирки с ЭДТК из подкожной вены каждой собаки до иммунизации. Образцы сыворотки анализируют на титр антител (IgG, IgG1, IgG2, IgA, IgM и IgE). Мононуклеарные клетки периферической крови анализируют на индуцированные вакциной клетки памяти и на продуцирование ими цитокинов. Защитные антитела определяют, выполняя анализ ADCC. Всех животных заражают 50 чувствительными к лекарственному средству личинками D, immitis, для определения защиты. Осуществляют отслеживание основных показателей жизнедеятельности и клинико-лабораторных показателей (OAK, общий анализ мочи, функция печени), а также реакцию в месте инъекции (отек, покраснение, боль).
[0298] Уровни антигенспецифических антител в сыворотке собак. Уровни rBmHAXT-специфических общих антител IgG1, IgG2, IgGA, IgM и IgE определяют в сыворотке каждой собаки с использованием косвенного ELISA, как описано в другом месте настоящего документа.
[0299] Анализ антителозависимой клеточно-опосредованной цитотоксичности (ADCC). Анализ ADCC выполняют, как описано в другом месте настоящего документа. Каждую из восьми-десяти живых D. immitis L3 инкубируют при 37°С с 5% СО2 в дублированных лунках вместе с 0,5 миллиона РВМС, выделенными из нормальной крови собаки, 200 мкл среды RPMI 1640 и 100 мкл образцов сыворотки. Планшеты отслеживают под световым микроскопом каждые 24 часа на жизнеспособность D. immitis L3. Личинки, которые являются прозрачными, неподвижными или малоподвижными, считают мертвыми. Долю гибели личинок выражают как отношение количества погибших L3 к общему количеству, извлеченному из каждой лунки, умноженное на 100.
[0300] Иммунизация вакциной rBmHAXT создает высокий титр антигенспецифических антител IgG изотипа. Через двадцать восемь дней после третьей вакцинации у собак, вакцинированных антигеном rBmHAXT, наблюдают значительное увеличение (р<0,01) титров IgG антител (1:10000) по сравнению с контрольной группой, получавшей адъювант. Также наблюдают значительное увеличение IgG1, IgG2 и IgA антител в сыворотке крови собак после трех иммунизации по сравнению с контрольной группой, где IgG1 и IgG2 являются наиболее преобладающим изотипом IgG антител. Хотя изменений в уровнях антител IgE и IgM не наблюдается, этот анализ демонстрирует сбалансированный иммунный ответ Th1/Th2, создаваемый против вакцинных антигенов у вакцинированных животных. ADCC показывает, что образцы сыворотки от животных, вакцинированных rBmHAXT, уничтожили 69±16,82% чувствительных к лекарственному средству личинок (р<0,05) по сравнению с контрольной группой AL019 (20±12,17%). Точно так же, ADCC показал, что образцы сыворотки от животных, вакцинированных rBmHAXT, уничтожили 86±7,50% резистентных к лекарственным средствам личинок (р<0,05) по сравнению с контрольной группой AL019 (38,2±22,80%). У животных, вакцинированных rBmHAXT, нагрузка чувствительными к лекарственному средству червями составляет 7±1,73 (Р=0,0025) по сравнению с контрольной группой AL019 (29,3±3,51). Это представляет собой снижение внедрения червей в вакцинированных животных на 76,11%. Действительно, извлеченные черви после контрольного заражения чувствительным к лекарственным средствам D. immitis L3 показывают значительное снижение числа животных, вакцинированных rBmHAXT, по сравнению с контрольной группой AL019 (6,67 против 29,33). Результаты этого анализа показывают, что после трех доз вакцинации rBmHAXT образуются защитные антитела, и эти антитела являются эффективными для уничтожения как резистентных к лекарственному средству, так и чувствительных к лекарственному средству инфекционных личинок D. immitis in vitro. Кроме того, исследования с контрольным заражением показывают, что вакцинация значительно снижает внедрение червей. Таким образом, вакцина rBmHAXT является безопасной для собак.
--->
Список последовательностей
<110> The Board of Trustees of the University of Illinois
Kalyanasundaram, Ramaswamy
<120> Vaccine and Methods for Detecting and Preventing Filariasis
<130> UIC0030WO.3
<150> US 16/790,277
<151> 2020-02-13
<160> 124
<170> PatentIn version 3.5
<210> 1
<211> 27
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 1
cgcggatcca tggaagagaa ggtggtg 27
<210> 2
<211> 25
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 2
ccggaattct cacttgtcgt tggtg 25
<210> 3
<211> 12
<212> PRT
<213> Brugia malayi
<400> 3
Trp Ser Ala Glu Gln Trp Asp Trp Pro Leu Gln His
1 5 10
<210> 4
<211> 10
<212> PRT
<213> Brugia malayi
<400> 4
Glu Val Ile Lys Thr Asn Thr Asn Asp Lys
1 5 10
<210> 5
<211> 8
<212> PRT
<213> Brugia malayi
<400> 5
Ser Arg Ala Glu His Tyr Gly Glu
1 5
<210> 6
<211> 11
<212> PRT
<213> Brugia malayi
<400> 6
Lys Leu Pro Ser Asp Val Asp Thr Lys Thr Leu
1 5 10
<210> 7
<211> 9
<212> PRT
<213> Brugia malayi
<400> 7
Phe Thr Pro Lys Glu Ile Glu Val Lys
1 5
<210> 8
<211> 8
<212> PRT
<213> Brugia malayi
<400> 8
Ile Glu Val Lys Val Ala Gly Asp
1 5
<210> 9
<211> 8
<212> PRT
<213> Brugia malayi
<400> 9
Val Gly Leu Asp Ala Ser Phe Phe
1 5
<210> 10
<211> 9
<212> PRT
<213> Brugia malayi
<400> 10
Gly Leu Asp Ala Ser Phe Phe Thr Pro
1 5
<210> 11
<211> 9
<212> PRT
<213> Brugia malayi
<400> 11
Val Lys Val Ala Gly Asp Asn Leu Val
1 5
<210> 12
<211> 9
<212> PRT
<213> Brugia malayi
<400> 12
Lys Leu Pro Ser Asp Val Asp Thr Lys
1 5
<210> 13
<211> 9
<212> PRT
<213> Brugia malayi
<400> 13
Lys Thr Leu Thr Ser Asn Leu Thr Lys
1 5
<210> 14
<211> 9
<212> PRT
<213> Brugia malayi
<400> 14
Val Ile Lys Thr Asn Thr Asn Asp Lys
1 5
<210> 15
<211> 9
<212> PRT
<213> Brugia malayi
<400> 15
Asp Thr Lys Thr Leu Thr Ser Asn Leu
1 5
<210> 16
<211> 9
<212> PRT
<213> Brugia malayi
<400> 16
Phe Phe Thr Pro Lys Glu Ile Glu Val
1 5
<210> 17
<211> 9
<212> PRT
<213> Brugia malayi
<400> 17
Val Gly Leu Asp Ala Ser Phe Phe Thr
1 5
<210> 18
<211> 9
<212> PRT
<213> Brugia malayi
<400> 18
Lys Arg Gly His Leu Val Ile Ala Ala
1 5
<210> 19
<211> 9
<212> PRT
<213> Brugia malayi
<400> 19
Gly Glu Ile Lys Arg Glu Ile Ser Arg
1 5
<210> 20
<211> 9
<212> PRT
<213> Brugia malayi
<400> 20
Ser Phe Phe Thr Pro Lys Glu Ile Glu
1 5
<210> 21
<211> 9
<212> PRT
<213> Brugia malayi
<400> 21
Arg Glu Ile Ser Arg Thr Tyr Lys Leu
1 5
<210> 22
<211> 9
<212> PRT
<213> Brugia malayi
<400> 22
Gly Glu Ile Lys Arg Glu Ile Ser Arg
1 5
<210> 23
<211> 9
<212> PRT
<213> Brugia malayi
<400> 23
Ala Glu His Tyr Gly Glu Ile Lys Arg
1 5
<210> 24
<211> 25
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 24
ccggaattct cacttgtcgt tggtg 25
<210> 25
<211> 26
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 25
cgcggatcca ccgtgatcca ttgtcg 26
<210> 26
<211> 27
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 26
aactgcagct gttttccatt tccattc 27
<210> 27
<211> 30
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 27
aactgcagat gggtaacaag ctcctcatcg 30
<210> 28
<211> 27
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 28
cgcgaattcg gcgcactgcc aacctgc 27
<210> 29
<211> 24
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 29
cgcgaattca ccatggtcct ggag 24
<210> 30
<211> 25
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 30
gctctagatc agtccttctg gctag 25
<210> 31
<211> 26
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 31
cgggatccat ggaagaaaag gtagtg 26
<210> 32
<211> 26
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 32
ccctcgagtg ctttcttttt ggcagc 26
<210> 33
<211> 30
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 33
ccctcgagat gaataaactt ttaatagcat 30
<210> 34
<211> 23
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 34
gggtacccgc gcattgccaa ccc 23
<210> 35
<211> 29
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 35
ggggtacccc ggcaaggatc aatttaaaa 29
<210> 36
<211> 28
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 36
cggaattctc aatctttttg agatgaat 28
<210> 37
<211> 129
<212> PRT
<213> Brugia malayi
<400> 37
Met Gly Asn Lys Leu Leu Ile Ala Phe Gly Leu Val Ile Leu Phe Val
1 5 10 15
Thr Leu Pro Cys Val Ser Glu Ser Asp Glu Glu Phe Asp Asp Ser Ala
20 25 30
Ala Asp Asp Thr Asp Asp Ser Glu Ala Gly Gly Gly Ser Glu Gly Gly
35 40 45
Asp Glu Tyr Val Thr Lys Gly Glu Phe Val Glu Thr Asp Gly Lys Lys
50 55 60
Lys Glu Cys Ser Ser His Glu Ala Cys Tyr Asp Gln Arg Glu Pro Gln
65 70 75 80
Ala Trp Cys Arg Leu Ser Glu Asn Gln Ala Trp Thr Asp Arg Gly Cys
85 90 95
Phe Cys Glu Asp Lys Leu His Ser Cys Val Ile Glu Arg Thr Asn Asn
100 105 110
Gly Lys Leu Glu Tyr Ser Tyr Cys Ala Pro Glu Ala Gly Trp Gln Cys
115 120 125
Ala
<210> 38
<211> 572
<212> DNA
<213> Brugia malayi
<400> 38
ggtttattac cccaagtttg agggaggaga aaatgaataa acttttaata gcattcggtt 60
tggtaattct ttttgtgaca ctcccgtgtg tatcagaatc agacgaagag ttcgatgact 120
ccgcagccga tgacaccgac gacagcgagg ccggaggtgg tagtgaagga ggtgatgaat 180
atgtaaccaa aggggaattt gttgaaactg atggcaaaaa gaaagagtgc tcttcgcacg 240
aagcttgcta cgatcaacgt gaaccacaag cgtggtgcag actgagcgag aatcaggcat 300
ggactgacag aggctgcttc tgcgaagata agttgcattc gtgcgtcatc gaaagaacga 360
acaatggtaa attggagtat tcgtactgtg cacctgaagc aggttggcaa tgcgcatagg 420
aatgaatcag tctttgatac taccaacttt ccatccattt cctacacttt tcaaatttaa 480
ttctttctaa caaataacat ttaactttgg ttttaccgtt attttattaa atattggaaa 540
tatgtgcgac aaaaaaaaaa aaaaaaaaaa aa 572
<210> 39
<211> 126
<212> PRT
<213> Brugia malayi
<400> 39
Met Asn Asn Leu Leu Ile Ala Ile Ser Leu Val Ile Leu Leu Val Ile
1 5 10 15
Phe Pro Ser Lys Ser Asp Leu Glu Ile Asp Asp Ser Ser Asp Ser Asp
20 25 30
Tyr Ala His Ser Asn Asp Asp Asp Tyr Asn Glu Glu Glu Asp Glu Tyr
35 40 45
Thr Thr Lys Gly Glu Phe Val Glu Thr Asp Gly Lys Trp Lys Asn Cys
50 55 60
Ser Ser His Ala Asp Cys Tyr Asp Tyr Arg Glu Pro Ile Ala Trp Cys
65 70 75 80
Lys Pro Thr Ala Asn Gln Phe Trp Thr Asp Lys Gly Cys Phe Cys Glu
85 90 95
Asp Met Leu Tyr Ser Cys Val Ile Glu Arg Thr Asn Asn Gly Lys Leu
100 105 110
Glu Tyr Ser Tyr Cys Thr Ser Lys Glu Asn Trp Gln Cys Leu
115 120 125
<210> 40
<211> 381
<212> DNA
<213> Brugia malayi
<400> 40
atgaataatc tattgatagc aatcagttta gtaattcttc ttgtgatatt cccatcaaaa 60
tcagatttag aaattgatga ttcctcagac agtgattatg ctcatagcaa tgacgacgat 120
tacaatgaag aagaagatga atatacgacc aaaggagaat ttgttgaaac tgatggcaaa 180
tggaagaatt gcagttctca tgcagattgc tatgattatc gtgaaccaat agcttggtgc 240
aaacccactg caaatcaatt ttggacagac aaaggttgct tctgtgaaga tatgttgtat 300
tcatgtgtga tcgaaagaac gaacaatggc aaattagaat attcatattg tacatccaaa 360
gaaaactggc aatgtttata a 381
<210> 41
<211> 128
<212> PRT
<213> Wuchereria bancrofti
<400> 41
Met Asn Lys Leu Leu Ile Ala Phe Gly Leu Val Ile Leu Leu Val Thr
1 5 10 15
Leu Pro Cys Ala Ser Glu Ser Asp Glu Glu Phe Asp Asp Gly Ser Asn
20 25 30
Asp Glu Thr Asp Asp Lys Glu Asp Glu Gly Asn Ser Glu Gly Gly Asp
35 40 45
Glu Tyr Val Thr Lys Gly Glu Val Val Glu Thr Asp Gly Lys Lys Lys
50 55 60
Glu Cys Ser Ser His Glu Ala Cys Tyr Asp Gln Arg Glu Pro Gln Ala
65 70 75 80
Trp Cys Arg Pro Asn Glu Asn Gln Ser Trp Thr Asp Lys Gly Cys Phe
85 90 95
Cys Glu Asp Lys Leu His Ser Cys Val Ile Glu Arg Lys Asn Asn Gly
100 105 110
Lys Leu Glu Tyr Ser Tyr Cys Ala Pro Glu Ala Gly Trp Gln Cys Ala
115 120 125
<210> 42
<211> 387
<212> DNA
<213> Wuchereria bancrofti
<400> 42
atgaataaac ttttaatagc attcggttta gtaattcttc ttgtgacact tccgtgtgca 60
tcagaatcag acgaagagtt cgatgacggc tcaaatgatg agaccgacga caaagaggac 120
gaaggtaata gtgaaggagg tgatgaatat gtaaccaaag gagaagttgt tgaaactgat 180
ggcaagaaga aagagtgctc ttcgcacgaa gcttgttacg atcaacgtga accacaagcg 240
tggtgtagac cgaacgagaa tcagtcgtgg actgacaaag gttgcttctg cgaagataag 300
ttgcattcgt gcgtcatcga aagaaagaac aacggtaaat tggagtattc gtattgcgca 360
cctgaagcgg gttggcaatg cgcgtag 387
<210> 43
<211> 83
<212> PRT
<213> Loa loa
<400> 43
Met Ile His Asn Gly Gly Tyr Gln Lys Leu Lys Val Lys Leu Arg Lys
1 5 10 15
Lys Glu Cys Lys Ser His Glu Ala Cys Tyr Asp Gln Arg Glu Pro Gln
20 25 30
Asp Trp Cys Arg Leu Asn Glu Asn Gln Ser Trp Thr Asp Lys Gly Cys
35 40 45
Tyr Cys Asp Asp Lys Leu His Ser Cys Ile Ile Glu Arg Lys Asn Gly
50 55 60
Gly Lys Leu Glu Tyr Ala His Cys Ala Pro Glu Gln Gly Trp Lys Cys
65 70 75 80
Pro Ser Lys
<210> 44
<211> 252
<212> DNA
<213> Loa loa
<400> 44
atgattcata acggtggata ccaaaaatta aaagtcaaat tgaggaagaa agaatgcaag 60
tctcatgaag cttgctacga tcaacgtgaa ccacaagact ggtgcagatt gaacgaaaat 120
caatcgtgga cagacaaagg ttgctactgt gacgataagt tgcattcatg cattattgag 180
cgcaagaacg gtggcaagtt ggaatatgcg cactgtgcgc ctgaacaagg atggaaatgt 240
ccaagcaagt aa 252
<210> 45
<211> 229
<212> PRT
<213> Brugia malayi
<400> 45
Met Val His Gly Cys Gly Asn Arg Thr Leu Lys Phe Leu Phe Phe Thr
1 5 10 15
Ala Asn Leu Leu Thr Cys Ala Phe Gly Ala Leu Leu Phe Gly Ile Ser
20 25 30
Leu Trp Ala Tyr His Asp Glu Asn Phe Leu Gln Lys Leu Glu Lys Val
35 40 45
Gln Glu Ile His Lys Glu Tyr Ser Pro Glu Leu Thr Gln His Gln Ile
50 55 60
Gly Leu Trp Leu Leu Ile Ile Ile Gly Ala Ser Ile Phe Leu Val Gly
65 70 75 80
Phe Leu Gly Cys Cys Gly Ala Ile Cys Glu Ser Thr Lys Leu Leu Ala
85 90 95
Leu Phe Ser Ile Ile Val Leu Ile Leu Ala Ile Leu Glu Val Ala Ser
100 105 110
Ile Val Leu Ile Ile Ala Gly Lys Asp Gln Phe Lys Asn Ala Leu Tyr
115 120 125
Asn Leu Leu Ser Lys Thr Gly Glu Ser Glu Asp Glu Met Gln His Phe
130 135 140
Lys Pro Ile Glu Asp Leu Phe Gln Cys Cys Gly Pro Thr Asn Glu Thr
145 150 155 160
Met Val Arg Tyr Ile Glu Asn Gly Leu Cys Glu Asp Glu Leu Arg Asn
165 170 175
Lys Pro Asn Cys Phe Ala Val Ile Ser Asp His Phe Asp Ser Ser Gln
180 185 190
Lys Asp Ile Ile Lys Ile Ser Val Val Leu Ile Ile Ile Asp Leu Leu
195 200 205
Ala Leu Phe Ser Thr Ser Met Leu Tyr Lys Ala Phe Arg Tyr Gln Thr
210 215 220
Pro Tyr Tyr Tyr Ala
225
<210> 46
<211> 690
<212> DNA
<213> Brugia malayi
<400> 46
atggttcacg gctgtggtaa tcgaacattg aaatttttat ttttcactgc aaatttatta 60
acttgtgcat ttggtgcact tctttttggc atttcattat gggcttatca tgatgaaaat 120
tttttgcaaa aattagaaaa agtacaggaa atacataaag aatattctcc tgaactaaca 180
caacatcaaa taggattgtg gttattaata attattggtg catccatatt tcttgttggt 240
tttctcggtt gttgtggtgc gatatgtgaa agtaccaagc tattggcttt attttctatt 300
attgtattaa ttcttgcaat acttgaagtt gcttcgatag tacttataat tgctggcaag 360
gatcaattta aaaatgcttt atataattta ctatcaaaaa ctggcgaatc agaagatgaa 420
atgcaacatt ttaaacctat cgaagattta ttccaatgtt gtggtccaac aaatgaaaca 480
atggttcgat acatcgagaa tggcttatgt gaggatgaat taagaaataa accgaattgt 540
ttcgcagtaa tatccgatca ttttgattca tctcaaaaag atatcattaa aatttccgtc 600
gttttaataa ttattgattt gcttgcatta ttttctacat ctatgcttta taaagcattt 660
cgctatcaaa caccatacta ttatgcttaa 690
<210> 47
<211> 230
<212> PRT
<213> Loa loa
<400> 47
Met Val His Gly Cys Gly Asn Arg Met Val Lys Phe Leu Phe Phe Thr
1 5 10 15
Ala Asn Leu Leu Ile Cys Leu Phe Gly Ala Leu Ile Phe Gly Phe Ser
20 25 30
Leu Trp Ala Asn Leu Asp Glu Asp Phe Ala Leu Lys Leu Glu Glu Thr
35 40 45
Arg Lys Ile His Lys Glu Asp Phe Asn Val Leu Ala Arg Tyr Gln Ala
50 55 60
Ala Phe Trp Val Leu Val Val Ile Gly Ala Phe Leu Phe Leu Val Gly
65 70 75 80
Phe Leu Gly Cys Cys Gly Ala Met Cys Glu Asn Ile Met Leu Leu Thr
85 90 95
Ala Phe Phe Ile Ile Ile Leu Ile Leu Thr Thr Ile Glu Val Gly Ala
100 105 110
Val Ile Phe Ala Ile Thr Ser Lys Gly Gln Phe Lys Ser Val Leu His
115 120 125
Arg Leu Leu Ser Glu Ala Gly Lys Ala Glu Asp Lys Tyr Leu Pro Asn
130 135 140
Leu Lys Pro Ile Glu Asp Leu Phe Tyr Cys Cys Gly Ala Thr Gln Glu
145 150 155 160
Thr Met Asn Arg Tyr Met Glu His Gly Leu Cys Glu Gly Glu Leu Lys
165 170 175
Asn Lys Pro Asp Cys Phe Thr Ala Ile Ser Asp Tyr Leu Glu Ser Thr
180 185 190
Gly Glu Ala Val Val Val Phe Ala Phe Phe Leu Leu Ile Ile Glu Leu
195 200 205
Phe Ala Leu Val Ser Thr Cys Ile Leu Cys Lys Ala Phe Arg Tyr Gly
210 215 220
Arg Pro Tyr Tyr Tyr Ala
225 230
<210> 48
<211> 693
<212> DNA
<213> Loa loa
<400> 48
atggttcacg gttgtggcaa tcgaatggtg aaatttttat ttttcactgc aaatttactg 60
atctgtttat ttggcgcact tatttttggc ttctcattat gggctaattt ggatgaagat 120
tttgcgctaa agctggaaga aacgagaaaa atacataaag aagattttaa tgtgctggca 180
agatatcaag cagcattttg ggtattggtt gttattgggg cattcttgtt tctcgttggt 240
ttcctcggct gttgtggtgc aatgtgtgaa aatatcatgt tgttgactgc atttttcatc 300
ataatactaa ttcttacaac aatcgaagtt ggcgcggtaa tatttgcaat cactagcaag 360
ggccaattta aaagtgtttt acataggcta ctatcagaag ctggtaaagc agaagacaaa 420
tacttgccga atttaaaacc catcgaagat ctgttttatt gctgtggtgc aacgcaagaa 480
acgatgaacc ggtacatgga acatggctta tgtgaaggag aattgaagaa taagccggac 540
tgcttcacgg caatatcgga ttatttggag tcaaccggag aagcggtcgt cgtatttgca 600
ttctttttgc taattatcga gttgtttgct ttagtttcca catgtatact atgtaaagca 660
ttccgttatg gaaggccata ctactatgct taa 693
<210> 49
<211> 113
<212> PRT
<213> Brugia malayi
<400> 49
Met Glu Glu Lys Val Val Glu Leu Thr His Asn Trp Ser Ala Glu Gln
1 5 10 15
Trp Asp Trp Pro Leu Gln His Asn Asp Glu Val Ile Lys Val Thr Asn
20 25 30
Thr Asn Asp Lys Phe Glu Val Gly Leu Asp Ala Ser Phe Phe Thr Pro
35 40 45
Lys Glu Ile Glu Val Lys Val Ala Gly Asp Asn Leu Val Ile His Cys
50 55 60
Arg His Glu Ser Arg Ala Glu His Tyr Gly Glu Ile Lys Arg Glu Ile
65 70 75 80
Ser Arg Thr Tyr Lys Leu Pro Ser Asp Val Asp Thr Lys Thr Leu Thr
85 90 95
Ser Asn Leu Thr Lys Arg Gly His Leu Val Ile Ala Ala Lys Lys Lys
100 105 110
Ala
<210> 50
<211> 342
<212> DNA
<213> Brugia malayi
<400> 50
atggaagaaa aggtagtgga actaacgcac aactggagtg cggagcagtg ggattggccc 60
ttacagcaca acgatgaggt gattaaagta accaacacga atgataagtt tgaagtcggt 120
ttggatgcat cattcttcac gccgaaggaa atcgaggtaa aagttgctgg cgataatctg 180
gtcattcact gcagacatga atcacgtgcc gagcattatg gagaaattaa acgtgaaatt 240
agtcgtacct ataagcttcc gtcggatgta gatacgaaga ctctgacatc aaatttgacc 300
aagcgaggac atttggtcat cgctgccaaa aagaaagcat ga 342
<210> 51
<211> 165
<212> PRT
<213> Onchocerca volvulus
<400> 51
Gln Thr Ser Pro Met Glu Arg Phe Ile Val Asn Leu Leu Asp Ser Thr
1 5 10 15
Phe Asp Asp Arg Ser Ser Arg Pro Leu His Ser Val Ala Pro Tyr Trp
20 25 30
Leu His Gln Pro Glu Leu Asn Glu Cys Asn Ile Gly Asn Ser Leu Gly
35 40 45
Glu Val Ile Asn Glu Lys Asp Lys Phe Ala Val Arg Ala Asp Val Ser
50 55 60
His Phe His Pro Lys Glu Leu Ser Val Ser Val Arg Asp Arg Glu Leu
65 70 75 80
Val Ile Glu Gly His His Glu Glu Arg Thr Asp Pro Ala Gly His Gly
85 90 95
Ser Ile Glu Arg His Phe Ile Arg Lys Tyr Val Leu Pro Glu Glu Val
100 105 110
Gln Pro Asp Thr Ile Glu Ser His Leu Ser Asp Lys Gly Val Leu Thr
115 120 125
Ile Ser Ala Asn Lys Thr Ala Ile Gly Thr Thr Ala Ser Arg Asn Ile
130 135 140
Pro Ile Arg Ala Ser Pro Lys Glu Pro Glu Ala Asn Gln Lys Ser Ala
145 150 155 160
Ile Asn Asp Ala Lys
165
<210> 52
<211> 574
<212> DNA
<213> Onchocerca volvulus
<400> 52
caaacttcac cgatggaacg tttcatagtg aaccttcttg acagtacatt cgatgacaga 60
tcatctcgac cgctccattc tgttgcacca tattggcttc atcaaccgga actgaatgaa 120
tgtaatatcg gaaattctct cggcgaggta ataaatgaga aagacaagtt tgcagttcga 180
gctgatgttt ctcatttcca tccgaaggaa ctttcggtgt cggttcgaga tcgtgaacta 240
gttattgaag gtcaccatga agagcgtacc gatccggcag gacatggtag tattgaacgt 300
cacttcattc gaaaatatgt cttgccagaa gaagtgcagc ctgacacgat cgaatcacat 360
ttgtccgata aaggagtgtt gacaattagc gcaaacaaaa cagctattgg cacgactgca 420
tcacgaaaca ttccaattcg agcatctcca aaagaaccgg aagcaaatca gaaatcggca 480
ataaatgatg caaagtaaaa agaagtagaa attccggata tatttcattc tttattttat 540
acgctttttt tttctttttt tgttcatttt aaag 574
<210> 53
<211> 113
<212> PRT
<213> Loa loa
<400> 53
Met Glu Glu Lys Val Leu Glu Leu Thr His Asn Trp Ser Ala Glu Gln
1 5 10 15
Trp Asp Trp Pro Leu Gln His Asn Asp Asp Val Ile Lys Val Thr Asn
20 25 30
Thr Asn Asp Lys Phe Glu Val Ser Leu Asp Ala Ser Phe Phe Thr Pro
35 40 45
Lys Glu Ile Glu Val Lys Val Ala Gly Asp Asn Leu Val Ile His Cys
50 55 60
Arg His Glu Ser Arg Ala Asp Gln Tyr Gly Glu Ile Lys Arg Glu Ile
65 70 75 80
Ser Arg Thr Tyr Lys Leu Pro Ser Asp Ile Asp Thr Lys Thr Leu Thr
85 90 95
Ser Thr Leu Thr Lys Arg Gly His Leu Val Ile Val Ala Lys Lys Lys
100 105 110
Ala
<210> 54
<211> 342
<212> DNA
<213> Loa loa
<400> 54
atggaagaaa aggtattgga actaacgcat aattggagtg ccgagcagtg ggattggccc 60
ttgcagcata acgatgacgt gattaaagtg accaatacga atgataagtt tgaagtcagt 120
ctggatgctt cattcttcac accgaaggaa atcgaggtga aagttgctgg tgataatctg 180
gtcattcact gcagacatga atcacgtgcc gaccaatacg gtgaaattaa acgtgaaatt 240
agtcgtacct ataagcttcc gtcggatatt gatacgaaga ccctgacatc aactttgacc 300
aaaagaggac atctggtcat cgttgccaaa aagaaggcat ga 342
<210> 55
<211> 220
<212> PRT
<213> Brugia malayi
<400> 55
Met Leu Phe Phe Val Ile Phe Ser Ile Thr Ile Ala Val Val Ala Gly
1 5 10 15
Phe Glu Cys Pro Gly Gly Arg Leu Thr Pro Gln Gln Arg Lys Asp Ile
20 25 30
Val Arg Gln Asn Asn Lys Phe Arg Ser Leu Leu Ile His Gly Lys Leu
35 40 45
Lys Asn Arg Asn Gly Thr Tyr Met Pro Arg Gly Lys Asn Met Leu Leu
50 55 60
Leu Lys Trp Ser Cys Gln Leu Glu Asn Ser Ala Gln Arg Trp Ala Asn
65 70 75 80
Gln Cys Val Phe Gly His Ser Pro Arg Asn Gln Arg Gln Gly Ile Gly
85 90 95
Glu Asn Val Tyr Ala Tyr Trp Ser Ser Glu Ser Val Glu Lys Leu Arg
100 105 110
Asn Thr Ala Gly Thr Glu Ala Gly Lys Ser Trp Trp Ser Glu Leu Pro
115 120 125
Lys Leu Tyr Lys Gln Asn Pro Ser Asn Asn Leu Thr Asp Asp Val Ala
130 135 140
Arg Gln Gly Val Leu His Phe Thr Gln Met Ala Trp Gly Lys Thr His
145 150 155 160
Lys Ile Gly Cys Gly Ile Ala Thr Asn Cys Asp Gly Gly Arg Thr Leu
165 170 175
Ile Ala Ile Cys His Tyr Ser Pro Ala Gly Asn Met Leu Lys Glu Leu
180 185 190
Ile Tyr Glu Leu Gly Glu Pro Cys Lys Thr Asp Ser Asp Cys Asn Thr
195 200 205
Lys Lys Cys Ala Lys Lys Ser Gly Leu Cys Arg Lys
210 215 220
<210> 56
<211> 897
<212> DNA
<213> Brugia malayi
<400> 56
gcacgagata ttttcaaagc taacagtata cattccacca aaaaactcgg caaaatgtta 60
tttttcgtta tattttccat cacgattgct gttgtggcag gttttgaatg tccaggaggt 120
cgactaacac cacagcaacg taaagatatc gttcgtcaga acaataaatt tcgttcatta 180
ctaattcatg gaaaacttaa aaataggaat ggtacatata tgccacgcgg caagaatatg 240
ttgctactga agtggagttg tcagttagaa aattctgcgc aaagatgggc aaatcaatgc 300
gtctttgggc actctccgag aaatcaaaga caaggaatcg gtgaaaatgt ctatgcctac 360
tggtcatcag aaagcgtcga aaagcttaga aataccgctg gtacggaagc cggcaaaagt 420
tggtggtcag aacttccgaa actgtacaaa cagaatcctt cgaacaatct gactgatgat 480
gttgccagac aaggtgtttt acatttcaca cagatggcct ggggtaaaac gcataaaatt 540
ggttgtggta tagcaacaaa ctgcgatggt ggtcgcacac tcatagctat ttgccattat 600
tcacccgctg gaaatatgct aaaagaactg atatatgagc ttggcgaacc atgcaaaacg 660
gatagcgatt gtaacacaaa gaaatgtgcc aaaaaatctg gattgtgtag aaaatgaaag 720
agagtttaaa gtttcacttt ttgcactctt aattttggtt ttatattata aaactgagaa 780
ttattacaaa catatttatt ggctattata agataaaaaa gttccttgaa attcttctca 840
aataaaattt ctctctgata aaagcatgaa acaaaaaaaa aaaaaaaaaa aaaaaaa 897
<210> 57
<211> 220
<212> PRT
<213> Wuchereria bancrofti
<400> 57
Met Leu Leu Phe Val Ile Phe Ser Ala Thr Ile Val Ala Val Ala Ala
1 5 10 15
Phe Glu Cys Pro Gly Gly Gln Leu Thr Pro Gln Gln Arg Lys Asp Ile
20 25 30
Val Arg Gln Asn Asn Lys Phe Arg Ser Leu Leu Ile Arg Gly Lys Leu
35 40 45
Lys Asn Arg Asn Gly Thr Tyr Met Pro Arg Gly Lys Asn Met Leu Gln
50 55 60
Leu Thr Trp Ser Cys Gln Leu Glu Asn Ser Ala Gln Arg Trp Ala Asn
65 70 75 80
Gln Cys Val Phe Gly His Ser Pro Arg Asn Gln Arg Gln Gly Ile Gly
85 90 95
Glu Asn Val Tyr Ala Tyr Trp Ser Ser Ala Ser Val Glu Asn Leu Arg
100 105 110
Lys Thr Ala Gly Thr Glu Ala Gly Lys Ser Trp Trp Ser Glu Leu Pro
115 120 125
Glu Leu Tyr Lys His Asn Pro Ser Asn Asn Leu Thr Asp Asp Val Ser
130 135 140
Arg Gln Gly Val Leu His Phe Thr Gln Met Ala Trp Gly Lys Thr His
145 150 155 160
Lys Ile Gly Cys Gly Ile Ala Thr Asn Cys Asp Gly Gly Arg Thr Leu
165 170 175
Ile Thr Ile Cys His Tyr Ser Pro Ala Gly Asn Ile Leu Lys Asn Leu
180 185 190
Ile Tyr Glu Leu Gly Glu Pro Cys Lys Lys Asp Gly Asp Cys Asn Thr
195 200 205
Lys Lys Cys Ala Lys Lys Ser Gly Leu Cys Arg Lys
210 215 220
<210> 58
<211> 873
<212> DNA
<213> Wuchereria bancrofti
<400> 58
ggaattcggc acgagcgaca tcttctgtta tcaaatatat tttcaaagct aacagtgtac 60
attctgccag aaaatcccgc aaaatgttac ttttcgtcat attttctgca actattgttg 120
ctgtggcggc ttttgaatgt ccaggaggtc aactaacacc gcaacaacgt aaagatatcg 180
ttcgtcagaa caataaattt cgttcattac tgattcgtgg aaaacttaaa aataggaatg 240
gtacatatat gccacgcggc aagaatatgt tgcaactgac atggagttgt cagttagaaa 300
attctgcgca aagatgggca aatcagtgcg tctttggaca ctcaccgaga aatcaaagac 360
aaggaatcgg tgaaaatgtt tacgcttact ggtcgtcagc aagcgttgaa aatcttagaa 420
aaaccgctgg tacggaagcc ggcaaaagtt ggtggtcaga acttccggaa ctgtacaaac 480
acaatccttc gaacaatctg actgacgatg tttccaggca aggtgttttg catttcacac 540
agatggcttg gggtaaaacg cataaaattg gttgtggtat tgctacaaac tgtgatggtg 600
gtcgcacact cataactatt tgccattatt cgcccgctgg aaatatactg aaaaatctga 660
tatatgagct tggcgaaccg tgcaaaaagg atggcgattg taacacaaag aaatgtgcga 720
aaaaatctgg attgtgcaga aagtgaaaga atttgaagag tttcaccttc tcatttctaa 780
ttttggtttt gtatcataaa actgacaatt attacaaaca tatttattgg ctattatggg 840
ataaagaagt ttcttgaaaa aaaaaaaaaa aaa 873
<210> 59
<211> 220
<212> PRT
<213> Onchocerca volvulus
<400> 59
Met Ile Leu Phe Ile Ile Phe Pro Ala Ile Val Val Ala Val Thr Gly
1 5 10 15
Tyr Asn Cys Pro Gly Gly Lys Leu Thr Ala Leu Glu Arg Lys Lys Ile
20 25 30
Val Gly Gln Asn Asn Lys Tyr Arg Ser Asp Leu Ile Asn Gly Lys Leu
35 40 45
Lys Asn Arg Asn Gly Thr Tyr Met Pro Arg Gly Lys Asn Met Leu Glu
50 55 60
Leu Thr Trp Asp Cys Lys Leu Glu Ser Ser Ala Gln Arg Trp Ala Asn
65 70 75 80
Gln Cys Ile Phe Gly His Ser Pro Arg Gln Gln Arg Glu Gly Val Gly
85 90 95
Glu Asn Val Tyr Ala Tyr Trp Ser Ser Val Ser Val Glu Gly Leu Lys
100 105 110
Lys Thr Ala Gly Thr Asp Ala Gly Lys Ser Trp Trp Ser Lys Leu Pro
115 120 125
Lys Leu Tyr Glu Asn Asn Pro Ser Asn Asn Met Thr Trp Lys Val Ala
130 135 140
Gly Gln Gly Val Leu His Phe Thr Gln Met Ala Trp Gly Lys Thr Tyr
145 150 155 160
Lys Ile Gly Cys Gly Val Ala Thr Gln Cys Asp Gly Gly Arg Thr Leu
165 170 175
Ile Val Ile Cys His Tyr Ser Pro Gly Gly Asn Met Val Gly Glu Val
180 185 190
Ile Tyr His Arg Gly Asn Pro Cys Lys Val Asp Lys Asp Cys Tyr Thr
195 200 205
Lys Lys Cys Leu Ser Lys Ser Gly Leu Cys Arg Lys
210 215 220
<210> 60
<211> 883
<212> DNA
<213> Onchocerca volvulus
<400> 60
gcattttcta aagtgacatc ttctgttatt atcaaatata tttctgagat taacagctta 60
tattctatcg gaaaattccg caaaatgata cttttcatca tcttccctgc aattgtcgtc 120
gcggtgacgg gttataattg tccaggaggt aaactaacag cactggaacg taaaaaaatc 180
gttggtcaga ataataaata tcgttcggat ttgattaatg gaaagcttaa gaatagaaat 240
ggtacataca tgccaagagg caagaatatg ttggaactga catgggattg taaattggaa 300
agttcggcac aaaggtgggc caatcagtgt atctttggac attcaccaag acaacaaaga 360
gagggagttg gtgaaaatgt ttatgcgtac tggtcatcag tatcagttga aggtcttaaa 420
aaaactgctg gtacggatgc tgggaaaagc tggtggtcaa agcttccaaa attatatgaa 480
aataaccctt cgaataatat gacttggaaa gttgccggtc aaggtgtttt gcatttcaca 540
caaatggctt ggggtaagac ttataaaatt ggttgcggtg ttgcaacaca atgtgatggt 600
ggtagaacac ttattgttat ttgtcactat tctcctggtg gaaatatggt tggagaggtg 660
atataccacc gaggtaatcc gtgtaaagtc gacaaagatt gctatacgaa aaaatgttta 720
tcaaaatctg gactgtgcag aaaatgaaaa tttttcgctt ttcttcattt aattctcggc 780
tatatatctc ctatattaat ttttcagcaa aaaagctata aagaaatatt cataattaaa 840
taaatatagt aattattatg aaataaaaaa ttaagttctg ctc 883
<210> 61
<211> 220
<212> PRT
<213> Loa loa
<400> 61
Met Leu Cys Ser Phe Met Phe Ala Ala Ile Ile Ile Ala Val Glu Gly
1 5 10 15
Tyr Arg Cys Gln Gly Gly Arg Leu Thr Pro Glu Gln Arg Lys Ala Ile
20 25 30
Val Ile Gln Asn Asn Lys Phe Arg Ser Gln Leu Ile Arg Gly Glu Leu
35 40 45
Lys Asn Lys Ala Gly Glu Phe Met Pro Arg Gly Lys Asn Met Leu Arg
50 55 60
Met Arg Trp Ser Cys Ser Leu Glu Tyr Ser Ala Gln Arg Trp Ala Asp
65 70 75 80
Arg Cys Ile Phe Gly His Ser Pro Arg Asp Gln Arg Asn Asn Ile Gly
85 90 95
Glu Asn Val Tyr Ala Tyr Trp Ser Ser Gly Ser Val Glu Gly His Arg
100 105 110
Lys Thr Ala Gly Thr Asp Ala Gly Lys Asn Trp Trp Ser Glu Leu Pro
115 120 125
Glu Arg Tyr Gly Ser Asn Pro Ser Asn Asn Leu Thr Ala Gln Val Ser
130 135 140
Ser Gln Gly Val Leu His Phe Thr Gln Met Ala Trp Gly Lys Thr Tyr
145 150 155 160
Lys Ile Gly Cys Gly Ile Ala Thr Asn Cys Asp Gly Gly Arg Thr Leu
165 170 175
Met Val Ile Cys His Tyr Ser Pro Ala Gly Asn Met Leu Lys Glu Leu
180 185 190
Ile Tyr Glu Leu Gly Glu Pro Cys Lys Lys Asn Asn Asp Cys Tyr Thr
195 200 205
Lys Lys Cys Ser Val Lys Ser Gly Leu Cys Asn Lys
210 215 220
<210> 62
<211> 663
<212> DNA
<213> Loa loa
<400> 62
atgttatgtt ccttcatgtt tgcagcaatt atcattgctg tggaaggtta tcgatgtcaa 60
ggaggtcgac taacaccgga gcaacgtaaa gcaatcgtta ttcaaaataa taaattccgt 120
tcgcaactga tccgtggcga gcttaagaac aaagctggtg aatttatgcc acgtggcaag 180
aatatgttga gaatgagatg gagttgttcg ctggaatatt ccgcgcagag atgggcagat 240
agatgtatct ttggacactc accaagagat caaagaaaca atatcggtga aaatgtctat 300
gcttactggt catcaggaag cgttgaaggt catagaaaaa ctgctggtac agatgctggt 360
aaaaattggt ggtcagaact tccggaacgg tacggatcta atccttcaaa taatttgact 420
gctcaagttt ccagccaagg tgttttgcat ttcacacaga tggcttgggg taaaacgtac 480
aaaattggct gtggtattgc tacaaactgc gatggtggca ggacactaat ggtcatttgc 540
cattattcgc cagctgggaa tatgttgaaa gagcttatat atgagcttgg cgaaccatgc 600
aagaagaaca atgattgtta taccaagaag tgttcagtca agtctggatt gtgcaacaag 660
tga 663
<210> 63
<211> 89
<212> PRT
<213> Brugia malayi
<400> 63
Met Ile Leu Glu Val Ala Ser Ile Val Leu Ile Ile Ala Gly Lys Asp
1 5 10 15
Gln Phe Lys Asn Ala Leu Tyr Asn Leu Leu Ser Lys Thr Gly Glu Ser
20 25 30
Glu Asp Glu Met Gln His Phe Lys Pro Ile Glu Asp Leu Phe Gln Cys
35 40 45
Cys Gly Pro Thr Asn Glu Thr Met Val Arg Tyr Ile Glu Asn Gly Leu
50 55 60
Cys Glu Asp Glu Leu Arg Asn Lys Pro Asn Cys Phe Ala Val Ile Ser
65 70 75 80
Asp His Phe Asp Ser Ser Gln Lys Asp
85
<210> 64
<211> 53
<212> PRT
<213> Brugia malayi
<400> 64
Val Ile His Cys Arg His Glu Ser Arg Ala Glu His Tyr Gly Glu Ile
1 5 10 15
Lys Arg Glu Ile Ser Arg Thr Tyr Lys Leu Pro Ser Asp Val Asp Thr
20 25 30
Lys Thr Leu Thr Ser Asn Leu Thr Lys Arg Gly His Leu Val Ile Ala
35 40 45
Ala Lys Lys Lys Ala
50
<210> 65
<211> 267
<212> DNA
<213> Brugia malayi
<400> 65
atgatcctgg aggtggcctc catcgtgctg atcatcgccg gcaaggacca gttcaagaac 60
gctctgtaca acctgctgtc taagacaggc gagagcgagg acgagatgca gcacttcaag 120
cctatcgagg acctgttcca gtgctgcggc cctaccaacg agaccatggt gaggtacatc 180
gagaacggac tgtgcgagga cgagctgagg aacaagccta actgtttcgc cgtgatctct 240
gaccacttcg attctagcca gaaggac 267
<210> 66
<211> 162
<212> DNA
<213> Brugia malayi
<400> 66
gtcattcact gcagacatga atcacgtgcc gagcattatg gagaaattaa acgtgaaatt 60
agtcgtacct ataagcttcc gtcggatgta gatacgaaga ctctgacatc aaatttgacc 120
aagcgaggac atttggtcat cgctgccaaa aagaaagcat ga 162
<210> 67
<211> 387
<212> DNA
<213> Brugia malayi
<400> 67
atgggtaaca agctcctcat cgctttcgga ctcgtgatcc tgtttgtcac cctgccttgc 60
gtcagtgaga gcgacgagga gtttgatgac tctgccgctg acgacaccga cgattccgag 120
gccggtggcg gctctgaggg gggcgacgag tacgtgacta agggggagtt tgtggagacc 180
gatggaaaga agaaagagtg ctccagccac gaggcctgtt atgaccagcg ggagcctcag 240
gcttggtgca ggctgtctga gaatcaggcc tggactgata gaggctgttt ctgtgaggac 300
aagctgcatt cttgcgtgat tgagcggact aataatggca agctggaata cagctactgc 360
gctcccgagg caggttggca gtgcgcc 387
<210> 68
<211> 153
<212> DNA
<213> Wuchereria bancrofti
<400> 68
atggtcattc actgcaggca cgagagcaga gccgagcact acggcgagat caaaagagag 60
atcagcagga catacaagct gccctcagac gtggacacta agactctgac tagcaatctg 120
accaagaggg gccacctggt catcgctgcc aag 153
<210> 69
<211> 267
<212> DNA
<213> Wuchereria bancrofti
<400> 69
atgatcctgg aggtggcctc catcgtgctg atcatcgccg gcaaggacca gttcaagaac 60
gctctgtaca acctgctgtc taagacaggc gagagcgagg acgagatgca gcacttcaag 120
cctatcgagg acctgttcca gtgctgcggc cctaccaacg agaccatggt gaggtacatc 180
gagaacggac tgtgcgagga cgagctgagg aacaagccta actgtttcgc cgtgatctct 240
gaccacttcg attctagcca gaaggac 267
<210> 70
<211> 331
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic polypeptide
<400> 70
Met Glu Glu Lys Val Val Glu Leu Thr His Asn Trp Ser Ala Glu Gln
1 5 10 15
Trp Asp Trp Pro Leu Gln His Asn Asp Glu Val Ile Lys Val Thr Asn
20 25 30
Thr Asn Asp Lys Phe Glu Val Gly Leu Asp Ala Ser Phe Phe Thr Pro
35 40 45
Lys Glu Ile Glu Val Lys Val Ala Gly Asp Asn Leu Val Ile His Cys
50 55 60
Arg His Glu Ser Arg Ala Glu His Tyr Gly Glu Ile Lys Arg Glu Ile
65 70 75 80
Ser Arg Thr Tyr Lys Leu Pro Ser Asp Val Asp Thr Lys Thr Leu Thr
85 90 95
Ser Asn Leu Thr Lys Arg Gly His Leu Val Ile Ala Ala Lys Lys Lys
100 105 110
Ala Met Gly Asn Lys Leu Leu Ile Ala Phe Gly Leu Val Ile Leu Phe
115 120 125
Val Thr Leu Pro Cys Val Ser Glu Ser Asp Glu Glu Phe Asp Asp Ser
130 135 140
Ala Ala Asp Asp Thr Asp Asp Ser Glu Ala Gly Gly Gly Ser Glu Gly
145 150 155 160
Gly Asp Glu Tyr Val Thr Lys Gly Glu Phe Val Glu Thr Asp Gly Lys
165 170 175
Lys Lys Glu Cys Ser Ser His Glu Ala Cys Tyr Asp Gln Arg Glu Pro
180 185 190
Gln Ala Trp Cys Arg Leu Ser Glu Asn Gln Ala Trp Thr Asp Arg Gly
195 200 205
Cys Phe Cys Glu Asp Lys Leu His Ser Cys Val Ile Glu Arg Thr Asn
210 215 220
Asn Gly Lys Leu Glu Tyr Ser Tyr Cys Ala Pro Glu Ala Gly Trp Gln
225 230 235 240
Cys Ala Met Ile Leu Glu Val Ala Ser Ile Val Leu Ile Ile Ala Gly
245 250 255
Lys Asp Gln Phe Lys Asn Ala Leu Tyr Asn Leu Leu Ser Lys Thr Gly
260 265 270
Glu Ser Glu Asp Glu Met Gln His Phe Lys Pro Ile Glu Asp Leu Phe
275 280 285
Gln Cys Cys Gly Pro Thr Asn Glu Thr Met Val Arg Tyr Ile Glu Asn
290 295 300
Gly Leu Cys Glu Asp Glu Leu Arg Asn Lys Pro Asn Cys Phe Ala Val
305 310 315 320
Ile Ser Asp His Phe Asp Ser Ser Gln Lys Asp
325 330
<210> 71
<211> 199
<212> PRT
<213> Brugia malayi
<400> 71
Met Thr Leu Ala Gly Ser Lys Ala Phe Ile Gly Gln Pro Ala Pro Asn
1 5 10 15
Phe Lys Thr Thr Ala Val Val Asn Gly Asp Phe Lys Glu Ile Ser Leu
20 25 30
Gly Gln Phe Lys Gly Lys Tyr Val Val Leu Leu Phe Tyr Pro Leu Asp
35 40 45
Phe Thr Phe Val Cys Pro Thr Glu Ile Ile Ala Phe Ser Asp Arg Ile
50 55 60
Ala Glu Phe Lys Gln Leu Asp Val Ala Val Met Ala Cys Ser Thr Asp
65 70 75 80
Ser His Phe Ser His Leu Ala Trp Val Asn Thr Asp Arg Lys Met Gly
85 90 95
Gly Leu Gly Gln Met Asn Ile Pro Ile Leu Ala Tyr Thr Asn His Val
100 105 110
Ile Ser Arg Ala Tyr Gly Val Leu Lys Glu Asp Asp Gly Ile Ala Tyr
115 120 125
Arg Gly Leu Phe Ile Ile Asp Pro Lys Gly Ile Leu Gly Gln Ile Thr
130 135 140
Ile Asn Asp Leu Pro Val Gly Arg Ser Val Asp Glu Thr Leu Arg Leu
145 150 155 160
Ile Gln Ala Phe Gln Phe Val Asp Lys His Gly Glu Val Cys Pro Ala
165 170 175
Asn Trp His Pro Gly Ser Glu Thr Ile Lys Pro Gly Val Lys Glu Ser
180 185 190
Lys Ala Tyr Phe Glu Lys His
195
<210> 72
<211> 713
<212> DNA
<213> Brugia malayi
<400> 72
ggtttaatta cccaagtttg aggaagaaaa gaaaatgaca cttgctggaa gtaaagcatt 60
tattggtcaa ccagcaccta attttaaaac aacagctgtt gtgaacggtg atttcaagga 120
aatttcactt ggtcaattta agggaaaata tgttgttctc ctcttttatc ctcttgattt 180
cacttttgtt tgtccaacgg aaataattgc attttctgat cgtattgcgg aatttaaaca 240
attagatgta gctgttatgg catgttcaac agattcacac ttttcacatc ttgcatgggt 300
aaatactgat cgaaaaatgg gtggacttgg tcaaatgaat ataccaattt tggcttatac 360
aaatcatgta ataagtcgag catatggtgt acttaaagag gatgatggaa ttgcttatcg 420
tggattattc attattgatc caaaaggaat tttggggcaa atcacaatta acgatcttcc 480
agtagggcgt tctgtagacg aaactttacg tttaattcaa gcttttcagt ttgttgacaa 540
gcatggtgaa gtatgtcctg ctaattggca tccaggatct gaaacgatta agcctggtgt 600
gaaagaaagt aaagcatatt ttgagaaaca ttaaaaacca aaaatggttg tacgacatgg 660
aattactcag tggatttttt tgattaaatt atatgctgaa gtaatattaa cca 713
<210> 73
<211> 441
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic polypeptide
<400> 73
Met Glu Glu Lys Val Val Glu Leu Thr His Asn Trp Ser Ala Glu Gln
1 5 10 15
Trp Asp Trp Pro Leu Gln His Asn Asp Glu Val Ile Lys Val Thr Asn
20 25 30
Thr Asn Asp Lys Phe Glu Val Gly Leu Asp Ala Ser Phe Phe Thr Pro
35 40 45
Lys Glu Ile Glu Val Lys Val Ala Gly Asp Asn Leu Val Ile His Cys
50 55 60
Arg His Glu Ser Arg Ala Glu His Tyr Gly Glu Ile Lys Arg Glu Ile
65 70 75 80
Ser Arg Thr Tyr Lys Leu Pro Ser Asp Val Asp Thr Lys Thr Leu Thr
85 90 95
Ser Asn Leu Thr Lys Arg Gly His Leu Val Ile Ala Ala Lys Lys Lys
100 105 110
Ala Met Gly Asn Lys Leu Leu Ile Ala Phe Gly Leu Val Ile Leu Phe
115 120 125
Val Thr Leu Pro Cys Val Ser Glu Ser Asp Glu Glu Phe Asp Asp Ser
130 135 140
Ala Ala Asp Asp Thr Asp Asp Ser Glu Ala Gly Gly Gly Ser Glu Gly
145 150 155 160
Gly Asp Glu Tyr Val Thr Lys Gly Glu Phe Val Glu Thr Asp Gly Lys
165 170 175
Lys Lys Glu Cys Ser Ser His Glu Ala Cys Tyr Asp Gln Arg Glu Pro
180 185 190
Gln Ala Trp Cys Arg Leu Ser Glu Asn Gln Ala Trp Thr Asp Arg Gly
195 200 205
Cys Phe Cys Glu Asp Lys Leu His Ser Cys Val Ile Glu Arg Thr Asn
210 215 220
Asn Gly Lys Leu Glu Tyr Ser Tyr Cys Ala Pro Glu Ala Gly Trp Gln
225 230 235 240
Cys Ala Met Thr Leu Ala Gly Ser Lys Ala Phe Ile Gly Gln Pro Ala
245 250 255
Pro Asn Phe Lys Thr Thr Ala Val Val Asn Gly Asp Phe Lys Glu Ile
260 265 270
Ser Leu Gly Gln Phe Lys Gly Lys Tyr Val Val Leu Leu Phe Tyr Pro
275 280 285
Leu Asp Phe Thr Phe Val Cys Pro Thr Glu Ile Ile Ala Phe Ser Asp
290 295 300
Arg Ile Ala Glu Phe Lys Gln Leu Asp Val Ala Val Met Ala Cys Ser
305 310 315 320
Thr Asp Ser His Phe Ser His Leu Ala Trp Val Asn Thr Asp Arg Lys
325 330 335
Met Gly Gly Leu Gly Gln Met Asn Ile Pro Ile Leu Ala Tyr Thr Asn
340 345 350
His Val Ile Ser Arg Ala Tyr Gly Val Leu Lys Glu Asp Asp Gly Ile
355 360 365
Ala Tyr Arg Gly Leu Phe Ile Ile Asp Pro Lys Gly Ile Leu Gly Gln
370 375 380
Ile Thr Ile Asn Asp Leu Pro Val Gly Arg Ser Val Asp Glu Thr Leu
385 390 395 400
Arg Leu Ile Gln Ala Phe Gln Phe Val Asp Lys His Gly Glu Val Cys
405 410 415
Pro Ala Asn Trp His Pro Gly Ser Glu Thr Ile Lys Pro Gly Val Lys
420 425 430
Glu Ser Lys Ala Tyr Phe Glu Lys His
435 440
<210> 74
<211> 530
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic polypeptide
<400> 74
Met Glu Glu Lys Val Val Glu Leu Thr His Asn Trp Ser Ala Glu Gln
1 5 10 15
Trp Asp Trp Pro Leu Gln His Asn Asp Glu Val Ile Lys Val Thr Asn
20 25 30
Thr Asn Asp Lys Phe Glu Val Gly Leu Asp Ala Ser Phe Phe Thr Pro
35 40 45
Lys Glu Ile Glu Val Lys Val Ala Gly Asp Asn Leu Val Ile His Cys
50 55 60
Arg His Glu Ser Arg Ala Glu His Tyr Gly Glu Ile Lys Arg Glu Ile
65 70 75 80
Ser Arg Thr Tyr Lys Leu Pro Ser Asp Val Asp Thr Lys Thr Leu Thr
85 90 95
Ser Asn Leu Thr Lys Arg Gly His Leu Val Ile Ala Ala Lys Lys Lys
100 105 110
Ala Met Gly Asn Lys Leu Leu Ile Ala Phe Gly Leu Val Ile Leu Phe
115 120 125
Val Thr Leu Pro Cys Val Ser Glu Ser Asp Glu Glu Phe Asp Asp Ser
130 135 140
Ala Ala Asp Asp Thr Asp Asp Ser Glu Ala Gly Gly Gly Ser Glu Gly
145 150 155 160
Gly Asp Glu Tyr Val Thr Lys Gly Glu Phe Val Glu Thr Asp Gly Lys
165 170 175
Lys Lys Glu Cys Ser Ser His Glu Ala Cys Tyr Asp Gln Arg Glu Pro
180 185 190
Gln Ala Trp Cys Arg Leu Ser Glu Asn Gln Ala Trp Thr Asp Arg Gly
195 200 205
Cys Phe Cys Glu Asp Lys Leu His Ser Cys Val Ile Glu Arg Thr Asn
210 215 220
Asn Gly Lys Leu Glu Tyr Ser Tyr Cys Ala Pro Glu Ala Gly Trp Gln
225 230 235 240
Cys Ala Met Thr Leu Ala Gly Ser Lys Ala Phe Ile Gly Gln Pro Ala
245 250 255
Pro Asn Phe Lys Thr Thr Ala Val Val Asn Gly Asp Phe Lys Glu Ile
260 265 270
Ser Leu Gly Gln Phe Lys Gly Lys Tyr Val Val Leu Leu Phe Tyr Pro
275 280 285
Leu Asp Phe Thr Phe Val Cys Pro Thr Glu Ile Ile Ala Phe Ser Asp
290 295 300
Arg Ile Ala Glu Phe Lys Gln Leu Asp Val Ala Val Met Ala Cys Ser
305 310 315 320
Thr Asp Ser His Phe Ser His Leu Ala Trp Val Asn Thr Asp Arg Lys
325 330 335
Met Gly Gly Leu Gly Gln Met Asn Ile Pro Ile Leu Ala Tyr Thr Asn
340 345 350
His Val Ile Ser Arg Ala Tyr Gly Val Leu Lys Glu Asp Asp Gly Ile
355 360 365
Ala Tyr Arg Gly Leu Phe Ile Ile Asp Pro Lys Gly Ile Leu Gly Gln
370 375 380
Ile Thr Ile Asn Asp Leu Pro Val Gly Arg Ser Val Asp Glu Thr Leu
385 390 395 400
Arg Leu Ile Gln Ala Phe Gln Phe Val Asp Lys His Gly Glu Val Cys
405 410 415
Pro Ala Asn Trp His Pro Gly Ser Glu Thr Ile Lys Pro Gly Val Lys
420 425 430
Glu Ser Lys Ala Tyr Phe Glu Lys His Met Ile Leu Glu Val Ala Ser
435 440 445
Ile Val Leu Ile Ile Ala Gly Lys Asp Gln Phe Lys Asn Ala Leu Tyr
450 455 460
Asn Leu Leu Ser Lys Thr Gly Glu Ser Glu Asp Glu Met Gln His Phe
465 470 475 480
Lys Pro Ile Glu Asp Leu Phe Gln Cys Cys Gly Pro Thr Asn Glu Thr
485 490 495
Met Val Arg Tyr Ile Glu Asn Gly Leu Cys Glu Asp Glu Leu Arg Asn
500 505 510
Lys Pro Asn Cys Phe Ala Val Ile Ser Asp His Phe Asp Ser Ser Gln
515 520 525
Lys Asp
530
<210> 75
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 75
gcgcataaat tcatcagc 18
<210> 76
<211> 23
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 76
gcgcaaaact taattacaaa agc 23
<210> 77
<211> 76
<212> PRT
<213> Brugia malayi
<400> 77
Gly Lys Asp Gln Phe Lys Asn Ala Leu Tyr Asn Leu Leu Ser Lys Thr
1 5 10 15
Gly Glu Ser Glu Asp Glu Met Gln His Phe Lys Pro Ile Glu Asp Leu
20 25 30
Phe Gln Cys Cys Gly Pro Thr Asn Glu Thr Met Val Arg Tyr Ile Glu
35 40 45
Asn Gly Leu Cys Glu Asp Glu Leu Arg Asn Lys Pro Asn Cys Phe Ala
50 55 60
Val Ile Ser Asp His Phe Asp Ser Ser Gln Lys Asp
65 70 75
<210> 78
<211> 33
<212> PRT
<213> Brugia malayi
<400> 78
Val Ser Glu Ser Asp Glu Glu Phe Asp Asp Ser Ala Ala Asp Asp Thr
1 5 10 15
Asp Asp Ser Glu Ala Gly Gly Gly Ser Glu Gly Gly Asp Glu Tyr Val
20 25 30
Thr
<210> 79
<211> 25
<212> PRT
<213> Brugia malayi
<400> 79
Glu Phe Val Glu Thr Asp Gly Lys Lys Lys Glu Cys Ser Ser His Glu
1 5 10 15
Ala Cys Tyr Asp Gln Arg Glu Pro Gln
20 25
<210> 80
<211> 12
<212> PRT
<213> Brugia malayi
<400> 80
Trp Ser Ala Glu Gln Trp Asp Trp Pro Leu Gln His
1 5 10
<210> 81
<211> 11
<212> PRT
<213> Brugia malayi
<400> 81
Lys Leu Pro Ser Asp Val Asp Thr Lys Thr Leu
1 5 10
<210> 82
<211> 10
<212> PRT
<213> Brugia malayi
<400> 82
Lys Thr Gly Glu Ser Glu Asp Glu Met Gln
1 5 10
<210> 83
<211> 11
<212> PRT
<213> Brugia malayi
<400> 83
Phe Ile Gly Gln Pro Ala Pro Asn Phe Lys Thr
1 5 10
<210> 84
<211> 24
<212> PRT
<213> Brugia malayi
<400> 84
Gly Glu Val Cys Pro Ala Asn Trp His Pro Gly Ser Glu Thr Ile Lys
1 5 10 15
Pro Gly Val Lys Glu Ser Lys Ala
20
<210> 85
<211> 208
<212> PRT
<213> Wuchereria bancrofti
<400> 85
Met Ser Tyr Lys Leu Thr Tyr Phe Pro Ile Arg Gly Leu Ala Glu Pro
1 5 10 15
Ile Arg Leu Val Leu Val Asp Gln Gly Ile Lys Phe Thr Asp Asp Arg
20 25 30
Ile Asn Ala Ser Asp Trp Pro Ser Met Lys Ser His Phe His Phe Gly
35 40 45
Gln Leu Pro Cys Leu Tyr Asp Gly Asp His Gln Ile Val Gln Ser Gly
50 55 60
Ala Ile Leu Arg His Leu Ala Arg Lys His Asn Leu Asn Gly Gly Asn
65 70 75 80
Glu Leu Glu Thr Thr His Ile Asp Met Phe Cys Glu Gly Ile Arg Asp
85 90 95
Leu His Thr Lys Tyr Ala Lys Met Ile Tyr Gln Ala Tyr Asp Thr Glu
100 105 110
Lys Asp Ser Tyr Ile Lys Asp Ile Leu Pro Val Glu Leu Ala Lys Phe
115 120 125
Glu Lys Leu Leu Ala Thr Arg Asp Asp Gly Lys Asn Phe Ile Leu Gly
130 135 140
Glu Lys Ile Ser Tyr Val Asp Phe Val Leu Phe Glu Glu Leu Asp Ile
145 150 155 160
His Gln Ile Leu Asp Pro His Cys Leu Asp Lys Phe Pro Leu Leu Lys
165 170 175
Ala Tyr His Gln Arg Met Glu Asp Arg Pro Gly Leu Lys Glu Tyr Cys
180 185 190
Lys Gln Arg Asn Arg Ala Lys Ile Pro Val Asn Gly Asn Gly Lys Gln
195 200 205
<210> 86
<211> 627
<212> DNA
<213> Wuchereria bancrofti
<400> 86
atgagttata aactgacgta ttttcctatc cgaggcttgg ctgaaccaat tcgtttagta 60
ctggttgatc aaggtataaa gtttactgat gatcgtatca atgcaagtga ttggccttct 120
atgaaatcac actttcattt tggacaacta ccttgtcttt acgatggtga tcatcaaatt 180
gtgcagtccg gtgctattct gagacatctt gctcgcaaac ataatttaaa tggtggaaat 240
gaactggaaa cgacacatat tgatatgttc tgtgaaggca tccgtgatct acataccaaa 300
tatgcaaaaa tgatttacca ggcttatgac acagaaaagg attcctacat aaaggatata 360
ttgccagtag agctggcgaa atttgaaaag ttgcttgcta cacgagacga tggaaaaaat 420
ttcattctag gagaaaagat ttcatacgta gatttcgtcc ttttcgaaga acttgacatt 480
catcagatcc tggatccgca ctgcttggat aaatttccac tcttaaaagc ttaccatcaa 540
cgaatggaag atagacctgg acttaaggag tactgcaagc agcgtaatag agcaaaaata 600
ccagtgaatg gaaatggaaa acagtga 627
<210> 87
<211> 16
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 87
attttcggcc gtaggc 16
<210> 88
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 88
aggtggccat tgacatgat 19
<210> 89
<211> 33
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 89
cccgaattct taatgtttct caaaatatgc ttt 33
<210> 90
<211> 342
<212> DNA
<213> Dirofilaria immitis
<400> 90
atggaagaga aagtagtgga actaacgcat aattggagtg ccgatcagtg ggattggcct 60
ttacagcata atgatgatgt ggttaaagtg accaatacga atgataagtt tgaagtaggt 120
ttggacgcat cattctttac gccgaaggaa atcgaggtga aggtttgtgg cgataatttg 180
gttattcact gtagacatga aacacgcact gatcaatatg gtgaaattaa acgtgaaatt 240
agtcgcactt ataaactccc atcggatgta gatacaaaga cattaacatc aaatttgact 300
aaacgaggac atctggttat tgctgccaaa aagaaggcat aa 342
<210> 91
<211> 113
<212> PRT
<213> Dirofilaria immitis
<400> 91
Met Glu Glu Lys Val Val Glu Leu Thr His Asn Trp Ser Ala Asp Gln
1 5 10 15
Trp Asp Trp Pro Leu Gln His Asn Asp Asp Val Val Lys Val Thr Asn
20 25 30
Thr Asn Asp Lys Phe Glu Val Gly Leu Asp Ala Ser Phe Phe Thr Pro
35 40 45
Lys Glu Ile Glu Val Lys Val Cys Gly Asp Asn Leu Val Ile His Cys
50 55 60
Arg His Glu Thr Arg Thr Asp Gln Tyr Gly Glu Ile Lys Arg Glu Ile
65 70 75 80
Ser Arg Thr Tyr Lys Leu Pro Ser Asp Val Asp Thr Lys Thr Leu Thr
85 90 95
Ser Asn Leu Thr Lys Arg Gly His Leu Val Ile Ala Ala Lys Lys Lys
100 105 110
Ala
<210> 92
<211> 453
<212> DNA
<213> Dirofilaria immitis
<400> 92
atgaacaaac ttttcatagt tcttggctta gcgcttcttt ttgttgcatt accttccgca 60
tcagaatcac aagaagagac tgtatctttt gaagaaagcg acgaagatta tgaagacgat 120
agtgaagatc aaacaaaaga agaggaacat tcaaaagagg aagatcattc agaagaacac 180
gacgatcatt cagctgaaga cgataaattt gtaactaaag gaaaatttgt tgaaagtgac 240
ggcaagatga agcattgcaa aacccatgaa gcttgctatg atcaacgtga accacaatcg 300
tggtgcatat taaaaccgca tcagtcatgg acacaaagag gttgtttctg cgaatcaaaa 360
aaacatgcat gcgttatcga acgaaaaagt ggcgacaaat tggaatattc gtattgctca 420
ccccgaaaaa actggcagtg ttcatacgat taa 453
<210> 93
<211> 150
<212> PRT
<213> Dirofilaria immitis
<400> 93
Met Asn Lys Leu Phe Ile Val Leu Gly Leu Ala Leu Leu Phe Val Ala
1 5 10 15
Leu Pro Ser Ala Ser Glu Ser Gln Glu Glu Thr Val Ser Phe Glu Glu
20 25 30
Ser Asp Glu Asp Tyr Glu Asp Asp Ser Glu Asp Gln Thr Lys Glu Glu
35 40 45
Glu His Ser Lys Glu Glu Asp His Ser Glu Glu His Asp Asp His Ser
50 55 60
Ala Glu Asp Asp Lys Phe Val Thr Lys Gly Lys Phe Val Glu Ser Asp
65 70 75 80
Gly Lys Met Lys His Cys Lys Thr His Glu Ala Cys Tyr Asp Gln Arg
85 90 95
Glu Pro Gln Ser Trp Cys Ile Leu Lys Pro His Gln Ser Trp Thr Gln
100 105 110
Arg Gly Cys Phe Cys Glu Ser Lys Lys His Ala Cys Val Ile Glu Arg
115 120 125
Lys Ser Gly Asp Lys Leu Glu Tyr Ser Tyr Cys Ser Pro Arg Lys Asn
130 135 140
Trp Gln Cys Ser Tyr Asp
145 150
<210> 94
<211> 600
<212> DNA
<213> Dirofilaria immitis
<400> 94
atgactcttg ctggaagcaa agcattcatt ggtcaaccgg cccctaattt caaaacaaca 60
gcggttgtga atggcgattt caaggaaatt tcactttgtc agttcaaagg aaaatatgtg 120
gtcctcttct tttatcctct cgatttcact ttcgtttgcc caacagagat aattgctttt 180
tctgatcgta ttgcggagtt caaaaaatta gatgtagctg ttatggcatg ctcaactgat 240
tcacattttt cacaccttgc atggataaat actgaccgaa aaatgggtgg cctcggtcag 300
atgaatatac caattcttgc tgataccaat catacaatta gtagggcata tggcgtgctc 360
aaggaagatg atggcattgc ttaccgtgga ttattcatca ttgatccaaa agggattttg 420
cgacaaatca caatcaatga tcttccagtt ggtcgttctg tagatgaaac tttacgtctg 480
attcaagctt ttcaatttgt cgacaatcac ggtgaagtat gtccggccaa ttggcagcca 540
ggatctgaag caatcaaacc tggagtgaaa gaaagcaaag cgtacttcga aaagcactga 600
<210> 95
<211> 199
<212> PRT
<213> Dirofilaria immitis
<400> 95
Met Thr Leu Ala Gly Ser Lys Ala Phe Ile Gly Gln Pro Ala Pro Asn
1 5 10 15
Phe Lys Thr Thr Ala Val Val Asn Gly Asp Phe Lys Glu Ile Ser Leu
20 25 30
Cys Gln Phe Lys Gly Lys Tyr Val Val Leu Phe Phe Tyr Pro Leu Asp
35 40 45
Phe Thr Phe Val Cys Pro Thr Glu Ile Ile Ala Phe Ser Asp Arg Leu
50 55 60
Ala Glu Phe Lys Lys Leu Asp Val Ala Val Met Ala Cys Ser Thr Asp
65 70 75 80
Ser His Phe Ser His Leu Ala Trp Ile Asn Thr Asp Arg Lys Met Gly
85 90 95
Gly Leu Gly Gln Met Asn Ile Pro Ile Leu Ala Asp Thr Asn His Thr
100 105 110
Ile Ser Arg Ala Tyr Gly Val Leu Lys Glu Asp Asp Gly Ile Ala Tyr
115 120 125
Arg Gly Leu Phe Ile Ile Asp Pro Lys Gly Ile Leu Arg Gln Ile Thr
130 135 140
Ile Asn Asp Leu Pro Val Gly Arg Ser Val Asp Glu Thr Leu Arg Leu
145 150 155 160
Ile Gln Ala Phe Gln Phe Val Asp Asn His Gly Glu Val Cys Pro Ala
165 170 175
Asn Trp Gln Pro Gly Ser Glu Ala Ile Lys Pro Gly Val Lys Glu Ser
180 185 190
Lys Ala Tyr Phe Glu Lys His
195
<210> 96
<211> 1389
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic polynucleotide
<400> 96
atggaagaga aagtagtgga actaacgcat aattggagtg ccgatcagtg ggattggcct 60
ttacagcata atgatgatgt ggttaaagtg accaatacga atgataagtt tgaagtaggt 120
ttggacgcat cattctttac gccgaaggaa atcgaggtga aggtttgtgg cgataatttg 180
gttattcact gtagacatga aacacgcact gatcaatatg gtgaaattaa acgtgaaatt 240
agtcgcactt ataaactccc atcggatgta gatacaaaga cattaacatc aaatttgact 300
aaacgaggac atctggttat tgctgccaaa aagaaggcat aaaacaaact tttcatagtt 360
cttggcttag cgcttctttt tgttgcatta ccttccgcat cagaatcaca agaagagact 420
gtatcttttg aagaaagcga cgaagattat gaagacgata gtgaagatca aacaaaagaa 480
gaggaacatt caaaagagga agatcattca gaagaacacg acgatcattc agctgaagac 540
gataaatttg taactaaagg aaaatttgtt gaaagtgacg gcaagatgaa gcattgcaaa 600
acccatgaag cttgctatga tcaacgtgaa ccacaatcgt ggtgcatatt aaaaccgcat 660
cagtcatgga cacaaagagg ttgtttctgc gaatcaaaaa aacatgcatg cgttatcgaa 720
cgaaaaagtg gcgacaaatt ggaatattcg tattgctcac cccgaaaaaa ctggcagtgt 780
tcatacgatt aaactcttgc tggaagcaaa gcattcattg gtcaaccggc ccctaatttc 840
aaaacaacag cggttgtgaa tggcgatttc aaggaaattt cactttgtca gttcaaagga 900
aaatatgtgg tcctcttctt ttatcctctc gatttcactt tcgtttgccc aacagagata 960
attgcttttt ctgatcgtat tgcggagttc aaaaaattag atgtagctgt tatggcatgc 1020
tcaactgatt cacatttttc acaccttgca tggataaata ctgaccgaaa aatgggtggc 1080
ctcggtcaga tgaatatacc aattcttgct gataccaatc atacaattag tagggcatat 1140
ggcgtgctca aggaagatga tggcattgct taccgtggat tattcatcat tgatccaaaa 1200
gggattttgc gacaaatcac aatcaatgat cttccagttg gtcgttctgt agatgaaact 1260
ttacgtctga ttcaagcttt tcaatttgtc gacaatcacg gtgaagtatg tccggccaat 1320
tggcagccag gatctgaagc aatcaaacct ggagtgaaag aaagcaaagc gtacttcgaa 1380
aagcactga 1389
<210> 97
<211> 462
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 97
Met Glu Glu Lys Val Val Glu Leu Thr His Asn Trp Ser Ala Asp Gln
1 5 10 15
Trp Asp Trp Pro Leu Gln His Asn Asp Asp Val Val Lys Val Thr Asn
20 25 30
Thr Asn Asp Lys Phe Glu Val Gly Leu Asp Ala Ser Phe Phe Thr Pro
35 40 45
Lys Glu Ile Glu Val Lys Val Cys Gly Asp Asn Leu Val Ile His Cys
50 55 60
Arg His Glu Thr Arg Thr Asp Gln Tyr Gly Glu Ile Lys Arg Glu Ile
65 70 75 80
Ser Arg Thr Tyr Lys Leu Pro Ser Asp Val Asp Thr Lys Thr Leu Thr
85 90 95
Ser Asn Leu Thr Lys Arg Gly His Leu Val Leu Ala Ala Lys Lys Lys
100 105 110
Ala Met Asn Lys Leu Phe Ile Val Leu Gly Leu Ala Leu Leu Phe Val
115 120 125
Ala Leu Pro Ser Ala Ser Glu Ser Gln Glu Glu Thr Val Ser Phe Glu
130 135 140
Glu Ser Asp Glu Asp Tyr Glu Asp Asp Ser Glu Asp Gln Thr Lys Glu
145 150 155 160
Glu Glu His Ser Lys Glu Glu Asp His Ser Glu Glu His Asp Asp His
165 170 175
Ser Ala Glu Asp Asp Lys Phe Val Thr Lys Gly Lys Phe Val Glu Ser
180 185 190
Asp Gly Lys Met Lys His Cys Lys Thr His Glu Ala Cys Tyr Asp Gln
195 200 205
Arg Glu Pro Gln Ser Trp Cys Ile Leu Lys Pro His Gln Ser Trp Thr
210 215 220
Gln Arg Gly Cys Phe Cys Glu Ser Lys Lys His Ala Cys Val Ile Glu
225 230 235 240
Arg Lys Ser Gly Asp Lys Leu Glu Tyr Ser Tyr Cys Ser Pro Arg Lys
245 250 255
Asn Trp Gln Cys Ser Tyr Asp Met Thr Leu Ala Gly Ser Lys Ala Phe
260 265 270
Ile Gly Gln Pro Ala Pro Asn Phe Lys Thr Thr Ala Val Val Asn Gly
275 280 285
Asp Phe Lys Glu Ile Ser Leu Cys Gln Phe Lys Gly Lys Tyr Val Val
290 295 300
Leu Phe Phe Tyr Pro Leu Asp Phe Thr Phe Val Cys Pro Thr Glu Ile
305 310 315 320
Ile Ala Phe Ser Asp Arg Ile Ala Glu Phe Lys Lys Leu Asp Val Ala
325 330 335
Val Met Ala Cys Ser Thr Asp Ser His Phe Ser His Leu Ala Trp Ile
340 345 350
Asn Thr Asp Arg Lys Met Gly Gly Leu Gly Gln Met Asn Ile Pro Ile
355 360 365
Leu Ala Asp Thr Asn His Thr Ile Ser Arg Ala Tyr Gly Val Leu Lys
370 375 380
Glu Asp Asp Gly Ile Ala Tyr Arg Gly Leu Phe Ile Ile Asp Pro Lys
385 390 395 400
Gly Ile Leu Arg Gln Ile Thr Ile Asn Asp Leu Pro Val Gly Arg Ser
405 410 415
Val Asp Glu Thr Leu Arg Leu Ile Gln Ala Phe Gln Phe Val Asp Asn
420 425 430
His Gly Glu Val Cys Pro Ala Asn Trp Gln Pro Gly Ser Glu Ala Ile
435 440 445
Lys Pro Gly Val Lys Glu Ser Lys Ala Tyr Phe Glu Lys His
450 455 460
<210> 98
<211> 23
<212> PRT
<213> Dirofilaria immitis
<400> 98
Ala Ser Glu Ser Gln Glu Glu Thr Val Ser Phe Glu Glu Ser Asp Glu
1 5 10 15
Asp Tyr Glu Asp Asp Ser Glu
20
<210> 99
<211> 24
<212> PRT
<213> Dirofilaria immitis
<400> 99
Phe Val Glu Ser Asp Gly Lys Met Lys His Cys Lys Thr His Glu Ala
1 5 10 15
Cys Tyr Asp Gln Arg Glu Pro Gln
20
<210> 100
<211> 26
<212> PRT
<213> Dirofilaria immitis
<400> 100
Asn Trp Ser Ala Asp Gln Trp Asp Trp Pro Leu Gln His Asn Asp Asp
1 5 10 15
Val Val Lys Val Thr Asn Thr Asn Asp Lys
20 25
<210> 101
<211> 24
<212> PRT
<213> Dirofilaria immitis
<400> 101
Gly Glu Val Cys Pro Ala Asn Trp Gln Pro Gly Ser Glu Ala Ile Lys
1 5 10 15
Pro Gly Val Lys Glu Ser Lys Ala
20
<210> 102
<211> 627
<212> DNA
<213> Dirofilaria immitis
<400> 102
atgagttata agctgacata tttccctatc cgaggcttgg ctgagccaat ccgcttactt 60
ctcgttgatc aaggcataaa gtttactgac gaacacattc ctaaggatga tttcgtttcc 120
ataaaatcac aatttcaatt tgggcaattg ccttgtttct acgatggcga tcagcaaatt 180
gttcaatccg gtgccattct gagacatctt gcccgcaaat tcaatttgaa tggtgaaaat 240
aatgcggaaa caacatatgt cgacatgttc tatgagggca ttcgtgacct acacagcaaa 300
tacacaagaa tgatctatga ggcttatgaa actcaaaagg atccttttat aaaaaatata 360
ttgccacaag aattggcgaa acttgaaaaa ttgctggcta ctcgagacaa tggaaaaaat 420
ttcattctcg gagacaagat ttcattcgca gattacgtcc tttttgaaga gcttgacgtt 480
cagcaaatct tagatccaca ctgcttggag aaatttccat tgttgaaggc gtttcatcaa 540
cgattgggag ataaaccaaa aattaaggaa tattgcgcga agcgcaatgc atcaaagatg 600
ccagtgaatg gaaatgggaa gcagtaa 627
<210> 103
<211> 208
<212> PRT
<213> Dirofilaria immitis
<400> 103
Met Ser Tyr Lys Leu Thr Tyr Phe Pro Ile Arg Gly Leu Ala Glu Pro
1 5 10 15
Ile Arg Leu Leu Leu Val Asp Gln Gly Ile Lys Phe Thr Asp Glu His
20 25 30
Ile Pro Lys Asp Asp Phe Val Ser Ile Lys Ser Gln Phe Gln Phe Gly
35 40 45
Gln Leu Pro Cys Phe Tyr Asp Gly Asp Gln Gln Ile Val Gln Ser Gly
50 55 60
Ala Ile Leu Arg His Leu Ala Arg Lys Phe Asn Leu Asn Gly Glu Asn
65 70 75 80
Asn Ala Glu Thr Thr Tyr Val Asp Met Phe Tyr Glu Gly Ile Arg Asp
85 90 95
Leu His Ser Lys Tyr Thr Arg Met Ile Tyr Glu Ala Tyr Glu Thr Gln
100 105 110
Lys Asp Pro Phe Ile Lys Asn Ile Leu Pro Gln Glu Leu Ala Lys Leu
115 120 125
Glu Lys Leu Leu Ala Thr Arg Asp Asn Gly Lys Asn Phe Ile Leu Gly
130 135 140
Asp Lys Ile Ser Phe Ala Asp Tyr Val Leu Phe Glu Glu Leu Asp Val
145 150 155 160
Gln Gln Ile Leu Asp Pro His Cys Leu Glu Lys Phe Pro Leu Leu Lys
165 170 175
Ala Phe His Gln Arg Leu Gly Asp Lys Pro Lys Ile Lys Glu Tyr Cys
180 185 190
Ala Lys Arg Asn Ala Ser Lys Met Pro Val Asn Gly Asn Gly Lys Gln
195 200 205
<210> 104
<211> 71
<212> PRT
<213> Brugia malayi
<220>
<221> misc_feature
<222> (11)..(11)
<223> Xaa can be any naturally occurring amino acid
<400> 104
Asp Asn Ala Leu Asp Arg Ala Asp Ala Ala Xaa Glu Lys Val Arg Gln
1 5 10 15
Met Thr Asp Lys Leu Glu Arg Ile Glu Glu Glu Leu Arg Asp Thr Gln
20 25 30
Lys Lys Met Met Gln Thr Glu Asn Asp Leu Asp Lys Ala Gln Glu Asp
35 40 45
Leu Ala Val Ala Asn Thr Asn Leu Glu Glu Lys Glu Lys Lys Val Gln
50 55 60
Glu Val Lys Phe Lys Lys Phe
65 70
<210> 105
<211> 71
<212> PRT
<213> Dirofilaria sp.
<400> 105
Met Ile Thr Met Thr Val Asp Asn Ala Leu Asp Arg Ala Asp Ala Ala
1 5 10 15
Glu Glu Lys Val Arg Gln Met Thr Asp Lys Leu Glu Arg Ile Glu Glu
20 25 30
Glu Leu Arg Asp Thr Gln Lys Lys Met Met Gln Thr Glu Asn Asp Leu
35 40 45
Asp Lys Ala Gln Glu Asp Leu Ala Val Ala Asn Thr Asn Leu Glu Glu
50 55 60
Lys Glu Lys Lys Val Gln Glu
65 70
<210> 106
<211> 504
<212> PRT
<213> Brugia malayi
<400> 106
Met Asn Arg Thr Thr Leu Ile Leu Phe Phe Ile Ile Leu Ser Asn Thr
1 5 10 15
Ile Thr Val Ile His Gly Tyr Val Arg Gly Cys Tyr Tyr Thr Asn Trp
20 25 30
Ala Gln Tyr Arg Asp Gly Glu Gly Lys Phe Leu Pro Gly Asn Ile Pro
35 40 45
Asn Gly Leu Cys Thr His Ile Leu Tyr Ala Phe Ala Lys Val Asp Glu
50 55 60
Leu Gly Asp Ser Lys Pro Phe Glu Trp Asn Asp Glu Asp Thr Glu Trp
65 70 75 80
Ser Lys Gly Met Tyr Ser Ala Val Thr Lys Leu Arg Glu Thr Asn Pro
85 90 95
Gly Leu Lys Val Leu Leu Ser Tyr Gly Gly Tyr Asn Phe Gly Ser Ala
100 105 110
Ile Phe Thr Gly Ile Ala Lys Ser Ala Gln Lys Thr Glu Arg Phe Ile
115 120 125
Lys Ser Ala Ile Ala Phe Leu Arg Lys Asn Asn Phe Asp Gly Phe Asp
130 135 140
Leu Asp Trp Glu Tyr Pro Val Gly Val Ala Glu Glu His Ala Lys Leu
145 150 155 160
Val Glu Ala Met Lys Thr Ala Phe Val Glu Glu Ala Lys Thr Ser Gly
165 170 175
Lys Gln Arg Leu Leu Leu Thr Ala Ala Val Ser Ala Gly Lys Gly Thr
180 185 190
Ile Asp Gly Ser Tyr Asn Val Glu Ser Leu Gly Lys Asn Phe Asp Leu
195 200 205
Leu Phe Leu Met Ser Tyr Asp Leu His Gly Ser Trp Glu Lys Asn Val
210 215 220
Asp Leu His Gly Lys Leu His Pro Thr Lys Gly Glu Val Ser Gly Ile
225 230 235 240
Gly Ile Phe Asn Thr Glu Phe Ala Ala Asp Tyr Trp Ala Ser Lys Gly
245 250 255
Met Pro Lys Glu Lys Ile Ile Ile Gly Ile Pro Met Tyr Ala Gln Gly
260 265 270
Trp Thr Leu Asp Asn Pro Ser Glu Thr Ala Ile Gly Ala Ala Ala Ser
275 280 285
Arg Pro Ser Ser Ala Ser Lys Thr Asn Pro Ala Gly Gly Thr Ala Ser
290 295 300
Tyr Trp Glu Ile Cys Lys Tyr Leu Lys Glu Gly Gly Lys Glu Thr Val
305 310 315 320
His Gln Glu Gly Val Gly Ala Tyr Met Val Lys Gly Asp Gln Trp Tyr
325 330 335
Gly Tyr Asp Asn Glu Glu Thr Ile Arg Ile Lys Met Lys Trp Leu Lys
340 345 350
Glu Lys Gly Tyr Gly Gly Ala Phe Ile Trp Ala Leu Asp Phe Asp Asp
355 360 365
Phe Thr Gly Lys Ser Cys Gly Lys Gly Pro Tyr Pro Leu Leu Asn Ala
370 375 380
Ile Ser Ser Glu Leu Glu Gly Glu Ser Glu Asn Pro Glu Ile Thr Thr
385 390 395 400
Glu Glu Pro Ser Ile Thr Glu Thr Glu Ala Tyr Glu Thr Asp Glu Thr
405 410 415
Glu Glu Thr Ser Glu Thr Glu Ala Tyr Asp Thr Asp Glu Thr Glu Glu
420 425 430
Thr Ser Glu Thr Glu Ala Thr Thr Tyr Asp Thr Asp Glu Thr Glu Gly
435 440 445
Gln Glu Cys Pro Glu Arg Asp Gly Leu Phe Pro His Pro Thr Asp Cys
450 455 460
His Leu Phe Ile Gln Cys Ala Asn Asn Ile Ala His Val Met Gln Cys
465 470 475 480
Pro Ala Thr Thr Phe Phe Asn Asp Ala Ile Lys Val Cys Asp His Met
485 490 495
Thr Asn Ala Pro Asp Thr Cys Ile
500
<210> 107
<211> 454
<212> PRT
<213> Dirofilaria sp.
<400> 107
Met Tyr Phe His Leu Glu Gly Asp Gly Lys Phe Leu Pro Glu Asn Leu
1 5 10 15
Pro Lys Gly Leu Cys Thr His Leu Leu Tyr Ala Phe Ala Lys Val Asp
20 25 30
Gln Leu Gly Leu Ser Gln Ala Phe Glu Trp Asn Asp Glu Asp Thr Glu
35 40 45
Trp Ser Lys Gly Met Tyr Ser Arg Val Leu Glu Leu Lys Asn Thr Asp
50 55 60
Pro Glu Leu Lys Leu Leu Leu Ser Tyr Gly Gly Tyr Asn Phe Gly Ser
65 70 75 80
Ala Leu Phe Thr Ala Leu Ala Lys Ser Ala Glu Lys Arg Lys His Phe
85 90 95
Ile Glu Ser Ala Leu Glu Phe Leu Arg Lys Asn Lys Phe Asp Gly Phe
100 105 110
Asp Leu Asp Trp Glu Tyr Pro Leu Gly Ala Val Glu Glu His Ala Lys
115 120 125
Leu Val Glu Glu Met Lys Ala Ala Phe Met Asn Glu Ala Lys Arg Ser
130 135 140
Gly Asn Gln Gln Leu Leu Leu Thr Ala Ala Val Ser Ala Gly Lys His
145 150 155 160
Leu Leu Asp Gln Ser Tyr Lys Val Leu Ser Leu Ala Lys Asn Phe Asp
165 170 175
Leu Leu Phe Leu Met Ser Tyr Asp Leu His Gly Ser Trp Glu Asn Lys
180 185 190
Val Asp Leu His Ala Lys Leu Tyr Pro Thr Lys Gly Glu Thr Phe Gly
195 200 205
Leu Gly Leu Phe Asn Thr Glu Phe Ala Ala Glu Tyr Trp Val Ser Lys
210 215 220
Gly Met Pro Lys Gln Lys Leu Leu Leu Gly Leu Pro Thr Tyr Gly Arg
225 230 235 240
Gly Trp Thr Leu Lys Asn Pro Ser Glu Thr Met Leu Gly Ala Ala Ser
245 250 255
Leu Gly Ala Ser Ala Ala Ser Thr Thr Asn Pro Ala Gly Gly Ser Ala
260 265 270
Ala Tyr Trp Glu Leu Cys Lys Tyr Leu Lys Lys Gly Gly Lys Glu Thr
275 280 285
Leu Asp Lys Gln Gly Leu Gly Ala Tyr Met Val Gln Glu Asn Gln Trp
290 295 300
Tyr Ser Tyr Asp Asn Glu Glu Thr Leu Lys Leu Lys Met Lys Trp Leu
305 310 315 320
Lys Lys Lys Gly Tyr Gly Gly Ala Phe Leu Trp Ser Leu Asp Phe Asp
325 330 335
Asp Phe Lys Gly Ala Ser Cys Gly Lys Gly Pro Tyr Pro Leu Leu Ser
340 345 350
Ala Leu Asn Arg Glu Leu Gly Ser Lys Leu Ala Ala Ser Ala Lys Ser
355 360 365
Leu Gln Ile Thr Pro Pro Pro Ser Glu Leu Ala Glu Asn Val Val Leu
370 375 380
Lys Met Thr Thr Ser Val Lys Pro Thr Glu Gln Phe Phe Asp Leu Lys
385 390 395 400
Cys Pro Glu Ser Ser Gly Leu Phe Gln Tyr Pro Ser Asp Cys His Met
405 410 415
Phe Leu Leu Cys Ala His Asp Tyr Ser Tyr Val Met Ala Cys Pro Pro
420 425 430
Asn Thr Phe Phe Asn Asp Asn Ser Lys Val Cys Asp His Met Met Asn
435 440 445
Ala Ala Gly Thr Cys Lys
450
<210> 108
<211> 125
<212> PRT
<213> Brugia malayi
<400> 108
Met Asn Lys Leu Leu Ile Ala Phe Gly Leu Ile Ile Leu Thr Val Thr
1 5 10 15
Leu Pro Cys Met Ser Gln Ser Asp Asp Glu Phe Asp Asp Glu Ser Ser
20 25 30
Gly Ala Asp Glu Gly Gly Asp Gly Ser Glu Gly Gly Asp Glu Tyr Val
35 40 45
Thr Lys Gly Glu Phe Val Glu Thr Asp Gly Lys Lys Lys Glu Cys Thr
50 55 60
Ser His Glu Ala Cys Tyr Asp Gln Arg Glu Pro Gln Ala Trp Cys Arg
65 70 75 80
Leu Asp Glu Asn Gln Ser Trp Thr Asp Lys Gly Cys Phe Cys Asp Asp
85 90 95
Lys Leu His Ser Cys Val Ile Glu Arg Lys Asn Ser Gly Lys Leu Glu
100 105 110
Tyr Ser Tyr Cys Ala Pro Gln Glu Ser Trp Gln Cys Ser
115 120 125
<210> 109
<211> 144
<212> PRT
<213> Dirofilaria sp.
<400> 109
Met Asn Lys Leu Phe Leu Phe Leu Gly Leu Val Leu Leu Phe Val Ser
1 5 10 15
Ser Phe Ser Ala Ser Glu Ser Glu Ser Glu Asp Val Thr Phe Glu Glu
20 25 30
Ser Asp Glu Asp Tyr Glu Glu Asp Glu Asp Asn Ser Glu Asp Gln Asn
35 40 45
Asn Gln Ser Asn Glu His Asp Asp His Pro Thr Glu Asp Asp Glu Tyr
50 55 60
Val Thr Lys Gly Glu Phe Val Glu Ser Asp Gly Lys Met Lys His Cys
65 70 75 80
Glu Thr His Glu Ala Cys Tyr Asp Gln Arg Glu Pro Gln Ser Trp Cys
85 90 95
Leu Leu Lys Pro His Gln Ser Trp Thr Gln Arg Gly Cys Phe Cys Glu
100 105 110
Ser Lys Lys His Ala Cys Val Leu Glu Arg Lys Ser Gly Asp Lys Leu
115 120 125
Glu Tyr Ser Tyr Cys Ser Pro Arg Asn Asn Trp Gln Cys Ser Tyr Asp
130 135 140
<210> 110
<211> 153
<212> PRT
<213> Brugia malayi
<400> 110
Met Lys Tyr Phe Ile Phe Leu Ser Ile Gly Leu Ile Ala Ala Ala Ser
1 5 10 15
Ala Gln Arg Glu Ala Gln Leu Pro Gln Pro Glu Ile Pro Pro Phe Leu
20 25 30
Ser Gly Ala Pro Ser His Val Val Lys Gln Phe Phe Asp Leu Leu Lys
35 40 45
Ala Asp Glu Ser Lys Thr Asp Pro Gln Thr Glu Ala Asp Ile Glu Ala
50 55 60
Phe Ile Arg Arg Leu Gly Gly Asp Tyr Gln Thr Arg Phe Glu Gln Phe
65 70 75 80
Lys Gln Glu Ile Lys Lys Glu Lys Ala Gln Tyr Glu Lys Ile His Gln
85 90 95
Ala Ala Leu Leu Lys Phe Ser Pro Ala Ala Arg Glu Ala Asp Ala Lys
100 105 110
Met Ser Ala Ile Ala Asp Ser Thr Gln Leu Thr Asn His Gln Lys Thr
115 120 125
Glu Gln Ile Lys Ala Ile Met Asp Ser Leu Ser Glu Ala Val Arg Lys
130 135 140
Glu Ile Leu Glu Gly Phe Asn Ser Gln
145 150
<210> 111
<211> 116
<212> PRT
<213> Dirofilaria sp.
<400> 111
Met Ile Leu Glu Gln Leu Glu Val Pro Pro Phe Leu Val Gly Ala Pro
1 5 10 15
Gln Ser Val Ile Lys Gln Phe Tyr Asp Leu Leu Lys Ala Asp Glu Thr
20 25 30
Lys Thr Asp Ala Gln Thr Glu Ala Asp Val Glu Ala Phe Ile Asn Arg
35 40 45
Leu Gly Gly Thr Tyr Lys Thr Arg Phe Asp Gln Phe Lys Gln Glu Ile
50 55 60
Lys Gln Gly Lys Ala Ala Tyr Glu Arg Leu His Gln Gln Ala Val Ala
65 70 75 80
Lys Phe Ser Lys Glu Ala Arg Glu Ala Asp Ala Lys Met Ser Ala Ile
85 90 95
Ala Asp Ser Pro Ser Leu Thr Thr Gln Gln Lys Thr Gln Gln Ile Gln
100 105 110
Ala Ile Met Asp
115
<210> 112
<211> 220
<212> PRT
<213> Brugia malayi
<400> 112
Met Leu Phe Phe Val Ile Phe Ser Ile Thr Ile Ala Val Val Ala Gly
1 5 10 15
Phe Glu Cys Pro Gly Gly Arg Leu Thr Pro Gln Gln Arg Lys Asp Ile
20 25 30
Val Arg Gln Asn Asn Lys Phe Arg Ser Leu Leu Ile His Gly Lys Leu
35 40 45
Lys Asn Arg Asn Gly Thr Tyr Met Pro Arg Gly Lys Asn Met Leu Leu
50 55 60
Leu Lys Trp Ser Cys Gln Leu Glu Asn Ser Ala Gln Arg Trp Ala Asn
65 70 75 80
Gln Cys Val Phe Gly His Ser Pro Arg Asn Gln Arg Gln Gly Ile Gly
85 90 95
Glu Asn Val Tyr Ala Tyr Trp Ser Ser Glu Ser Val Glu Lys Leu Arg
100 105 110
Asn Thr Ala Gly Thr Glu Ala Gly Lys Ser Trp Trp Ser Glu Leu Pro
115 120 125
Lys Leu Tyr Lys Gln Asn Pro Ser Asn Asn Leu Thr Asp Asp Val Ala
130 135 140
Arg Gln Gly Val Leu His Phe Thr Gln Met Ala Trp Gly Lys Thr His
145 150 155 160
Lys Ile Gly Cys Gly Ile Ala Thr Asn Cys Asp Gly Gly Arg Thr Leu
165 170 175
Ile Ala Ile Cys His Tyr Ser Pro Ala Gly Asn Met Leu Lys Glu Leu
180 185 190
Ile Tyr Glu Leu Gly Glu Pro Cys Lys Thr Asp Ser Asp Cys Asn Thr
195 200 205
Lys Lys Cys Ala Lys Lys Ser Gly Leu Cys Arg Lys
210 215 220
<210> 113
<211> 152
<212> PRT
<213> Dirofilaria immitis
<400> 113
Lys Leu Glu Lys Ser Ala Gln Asn Trp Ala Asn Met Cys Val Phe Gly
1 5 10 15
His Ser Pro Ser Ser Glu Arg Arg Gly Ile Gly Glu Asn Val Tyr Ala
20 25 30
Tyr Trp Ser Ser Gly Ser Val Arg Asp Leu Lys Lys Thr Ala Gly Thr
35 40 45
Asp Ala Gly Arg Leu Trp Trp Ser Glu Leu Glu Lys Tyr Val Ser Asp
50 55 60
Asn Pro Ser Asn Asn Leu Thr Ser Glu Val Ala Met Glu Asn Ile Leu
65 70 75 80
His Phe Thr Gln Met Ala Trp Gly Glu Thr Tyr Lys Leu Gly Ser Gly
85 90 95
Val Asp His Asn Ile Val Met Val Ala Arg Thr Leu Val Phe Ile Cys
100 105 110
His Tyr Phe Pro Gly Gly Asn Met Val Lys Asp Leu Ile Tyr Glu Leu
115 120 125
Gly Asn Pro Cys Lys His Asn Lys Asp Cys Arg Thr Lys Arg Cys Ser
130 135 140
Ala Lys Ser Gly Leu Cys Lys Lys
145 150
<210> 114
<211> 115
<212> PRT
<213> Brugia malayi
<400> 114
Met Pro Tyr Phe Thr Ile Asp Thr Asn Ile Pro Gln Asn Ser Ile Ser
1 5 10 15
Ser Ala Phe Leu Lys Lys Ala Ser Asn Val Val Ala Lys Ala Leu Gly
20 25 30
Lys Pro Glu Ser Tyr Val Ser Ile His Val Asn Gly Gly Gln Ala Met
35 40 45
Val Phe Gly Gly Ser Glu Asp Pro Cys Ala Val Cys Val Leu Lys Ser
50 55 60
Ile Gly Cys Val Gly Pro Lys Val Asn Asn Ser His Ala Glu Lys Leu
65 70 75 80
Tyr Lys Leu Leu Ala Asp Glu Leu Lys Ile Pro Lys Asn Arg Cys Tyr
85 90 95
Ile Glu Phe Val Asp Ile Glu Ala Ser Ser Met Ala Phe Asn Gly Ser
100 105 110
Thr Phe Gly
115
<210> 115
<211> 115
<212> PRT
<213> Dirofilaria immitis
<400> 115
Met Pro Tyr Phe Thr Ile Asp Thr Asn Ile Pro Gln Asp Arg Val Ser
1 5 10 15
Asp Ala Phe Leu Lys Lys Ala Ser Ser Thr Val Ala Lys Ala Leu Gly
20 25 30
Lys Pro Glu Ser Tyr Val Ser Ile His Val Asn Gly Gly Gln Ala Met
35 40 45
Thr Phe Gly Gly Ser Thr Asp Pro Cys Ala Val Cys Val Leu Lys Ser
50 55 60
Ile Gly Ser Val Gly Pro Ser Val Asn Asn Ser His Cys Glu Lys Leu
65 70 75 80
Tyr Lys Leu Leu Ala Asp Glu Leu Lys Ile Pro Lys Asn Arg Cys Tyr
85 90 95
Ile Glu Phe Val Asp Ile Asn Ala Ser Ala Met Gly Phe Asn Gly Ser
100 105 110
Thr Phe Gly
115
<210> 116
<211> 120
<212> PRT
<213> Brugia malayi
<400> 116
Met Pro Leu Ile Thr Leu Ala Ser Asn Val Pro Ala Ser Arg Phe Pro
1 5 10 15
Ser Asp Phe Asn Val Gln Phe Thr Glu Leu Met Ala Lys Met Leu Gly
20 25 30
Lys Pro Thr Ser Arg Ile Leu Leu Leu Val Met Pro Asn Ala Gln Leu
35 40 45
Ser His Gly Thr Thr Glu Asn Pro Ser Cys Phe Thr Val Val Lys Ser
50 55 60
Ile Gly Ser Phe Ser Ala Asp Lys Asn Ile Glu Tyr Ser Ser Leu Ile
65 70 75 80
Ser Glu Phe Met Lys Lys Thr Leu Asp Ile Asp Pro Ala His Cys Ile
85 90 95
Ile His Phe Leu Asn Leu Asp Pro Glu Asn Val Gly Cys Asn Gly Thr
100 105 110
Thr Met Lys Glu Leu Met Lys Lys
115 120
<210> 117
<211> 78
<212> PRT
<213> Dirofilaria sp.
<400> 117
Met Phe Ala Leu Ile Asp Val Ala Arg Tyr Glu Thr Ile Lys Tyr Pro
1 5 10 15
Thr Pro Leu Ile Lys Ser Ile Gly Ser Phe Ser Ala Asp Lys Asn Ile
20 25 30
Lys Tyr Ser Ala Ser Ile Ser Glu Phe Ile Lys Lys Thr Leu Gly Ile
35 40 45
Asp Pro Ala His Cys Leu Ile His Phe Phe Asn Leu Asp Pro Glu Asp
50 55 60
Val Gly Cys Asn Gly Thr Thr Met Lys Glu Leu Met Lys Lys
65 70 75
<210> 118
<211> 161
<212> PRT
<213> Brugia malayi
<400> 118
Met Met Ser Thr Met Ser Ile Lys Glu Gly Leu Leu Val Ile Leu Leu
1 5 10 15
Ser Leu Phe Leu Phe Asp Thr Thr Ala Leu Ile His Arg Arg Glu Ile
20 25 30
Pro His Met Glu Ser Lys Gly Gln Met Gln Arg Gly Gln Val Leu Leu
35 40 45
Gly Gly Trp Gln Glu Arg Ser Pro Glu Asp Asn Glu Ile Leu Glu Leu
50 55 60
Leu Pro Ser Val Leu Thr Lys Val Asn Gln Gln Ser Asn Asp Glu Tyr
65 70 75 80
His Leu Met Pro Ile Lys Leu Leu Lys Val Ser Ser Gln Val Val Ala
85 90 95
Gly Val Lys Tyr Lys Met Glu Val Gln Val Ala Arg Ser Glu Cys Lys
100 105 110
Lys Ser Ala Ser Glu Gln Val Asn Leu Lys Thr Cys Lys Lys Leu Glu
115 120 125
Gly His Pro Asp Gln Val Met Thr Leu Glu Val Trp Glu Lys Pro Trp
130 135 140
Glu Asp Phe Leu Gln Val Asn Ile Leu Glu Thr Lys Val Leu Ser Ser
145 150 155 160
Val
<210> 119
<211> 162
<212> PRT
<213> Dirofilaria sp.
<400> 119
Met Thr Met Leu Val Ser Ile Lys Asp Gly Ser Tyr Leu Val Leu Leu
1 5 10 15
Leu Leu Phe Ser Leu Ile Thr Leu Val Gln Leu Arg Glu Ala Lys Glu
20 25 30
Ala Lys Val Thr Gly Ile Met Glu Ser Lys Thr Asn Glu Arg Pro His
35 40 45
Glu Val Leu Leu Gly Gly Trp Glu Glu Arg Asp Pro Gln Asp Glu Glu
50 55 60
Ile Leu Glu Leu Leu Pro Ser Ile Leu Thr Lys Val Asn Glu Gln Ser
65 70 75 80
Asn Asp Glu Tyr Tyr Leu Val Pro Ile Lys Leu Leu Lys Val Ser Ser
85 90 95
Gln Val Val Ala Gly Val Lys Tyr Lys Met Asp Val Gln Val Ala Arg
100 105 110
Ser Glu Cys Lys Lys Ser Ser Asn Glu Lys Val Asp Leu Lys Thr Cys
115 120 125
Lys Lys Leu Gly His Pro Asp Gln Val Met Thr Leu Glu Ile Trp Glu
130 135 140
Lys Pro Trp Glu Asn Phe Leu Arg Val Asn Ile Leu Glu Thr Lys Val
145 150 155 160
Leu Pro
<210> 120
<211> 208
<212> PRT
<213> Brugia malayi
<400> 120
Met Ser Tyr Lys Leu Thr Tyr Phe Pro Ile Arg Gly Leu Ala Glu Pro
1 5 10 15
Ile Arg Leu Val Leu Val Asp Gln Gly Ile Lys Phe Thr Asp Asp Arg
20 25 30
Ile Asn Ala Ser Asp Trp Pro Ser Met Lys Ser His Phe His Phe Gly
35 40 45
Gln Leu Pro Cys Leu Tyr Asp Gly Asp His Gln Ile Val Gln Ser Gly
50 55 60
Ala Ile Leu Arg His Leu Ala Arg Lys His Asn Leu Asn Gly Gly Asn
65 70 75 80
Glu Leu Glu Thr Thr His Ile Asp Met Phe Cys Glu Gly Val Arg Asp
85 90 95
Leu His Thr Lys Tyr Thr Lys Met Ile Tyr Gln Ala Tyr Asp Thr Glu
100 105 110
Lys Asp Ser Tyr Ile Lys Asp Ile Leu Pro Val Glu Leu Ala Lys Phe
115 120 125
Glu Lys Leu Leu Ala Thr Arg Asp Asp Gly Lys Asn Phe Ile Leu Gly
130 135 140
Glu Lys Ile Ser Tyr Val Asp Phe Val Leu Phe Glu Glu Leu Asp Ile
145 150 155 160
His Gln Ile Leu Asp Pro His Cys Leu Asp Lys Phe Pro Leu Leu Lys
165 170 175
Ala Tyr His Gln Arg Met Glu Asp Arg Pro Gly Leu Lys Glu Tyr Cys
180 185 190
Lys Gln Arg Asn Arg Ala Lys Ile Pro Val Asn Gly Asn Gly Lys Gln
195 200 205
<210> 121
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa denotes Val or Phe
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> Xaa denotes Ser or Glu
<220>
<221> MISC_FEATURE
<222> (7)..(7)
<223> Xaa denotes Asp or Glu
<220>
<221> MISC_FEATURE
<222> (8)..(8)
<223> Xaa denotes Phe or Tyr
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> Xaa denotes Asp or Glu
<220>
<221> MISC_FEATURE
<222> (11)..(11)
<223> Xaa denotes Asp or Ser
<400> 121
Xaa Xaa Glu Ser Asp Glu Xaa Xaa Xaa Asp Xaa
1 5 10
<210> 122
<211> 24
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa is Ser or Thr
<220>
<221> MISC_FEATURE
<222> (8)..(8)
<223> Xaa is Met or Lys
<220>
<221> MISC_FEATURE
<222> (10)..(10)
<223> Xaa is Glu or His
<220>
<221> MISC_FEATURE
<222> (12)..(12)
<223> Xaa is Ser or Lys
<220>
<221> MISC_FEATURE
<222> (13)..(13)
<223> Xaa is Ser or Thr
<400> 122
Phe Val Glu Xaa Asp Gly Lys Xaa Lys Xaa Cys Xaa Xaa His Glu Ala
1 5 10 15
Cys Tyr Asp Gln Arg Glu Pro Gln
20
<210> 123
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa is Glu or Asp
<400> 123
Trp Ser Ala Xaa Gln Trp Asp Trp Pro Leu Gln His
1 5 10
<210> 124
<211> 24
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> Xaa is His or Gln
<220>
<221> MISC_FEATURE
<222> (14)..(14)
<223> Xaa is Thr or Ala
<400> 124
Gly Glu Val Cys Pro Ala Asn Trp Xaa Pro Gly Ser Glu Xaa Ile Lys
1 5 10 15
Pro Gly Val Lys Glu Ser Lys Ala
20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ И СПОСОБЫ ДЕГРАДАЦИИ НЕПРАВИЛЬНО УПАКОВАННЫХ БЕЛКОВ | 2016 |
|
RU2761564C2 |
| ТЕРАПЕВТИЧЕСКИЕ СОЕДИНЕНИЯ И СПОСОБЫ | 2016 |
|
RU2770001C2 |
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2020 |
|
RU2811752C2 |
| ИММУНОГЛОБУЛИН С ОДНИМ ВАРИАБЕЛЬНЫМ ДОМЕНОМ ПРОТИВ F-БЕЛКА PCB | 2016 |
|
RU2730671C2 |
| НЕ ВСТРЕЧАЮЩИЙСЯ В ПРИРОДЕ ВИРУС РЕПРОДУКТИВНО-РЕСПИРАТОРНОГО СИНДРОМА СВИНЕЙ (BPPCC) И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2687150C2 |
| T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, РАСПОЗНАЮЩИЕ МУТАЦИЮ R175H ИЛИ Y220C В P53 | 2020 |
|
RU2830061C2 |
| КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА | 2017 |
|
RU2813829C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНА ПРОТИВ МЕЗОТЕЛИНА ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2714902C2 |
| ОНКОЛИТИЧЕСКИЙ HSV-ВЕКТОР | 2014 |
|
RU2719190C2 |
| АНТИ-αvβ8 АНТИТЕЛА И КОМПОЗИЦИИ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812478C2 |
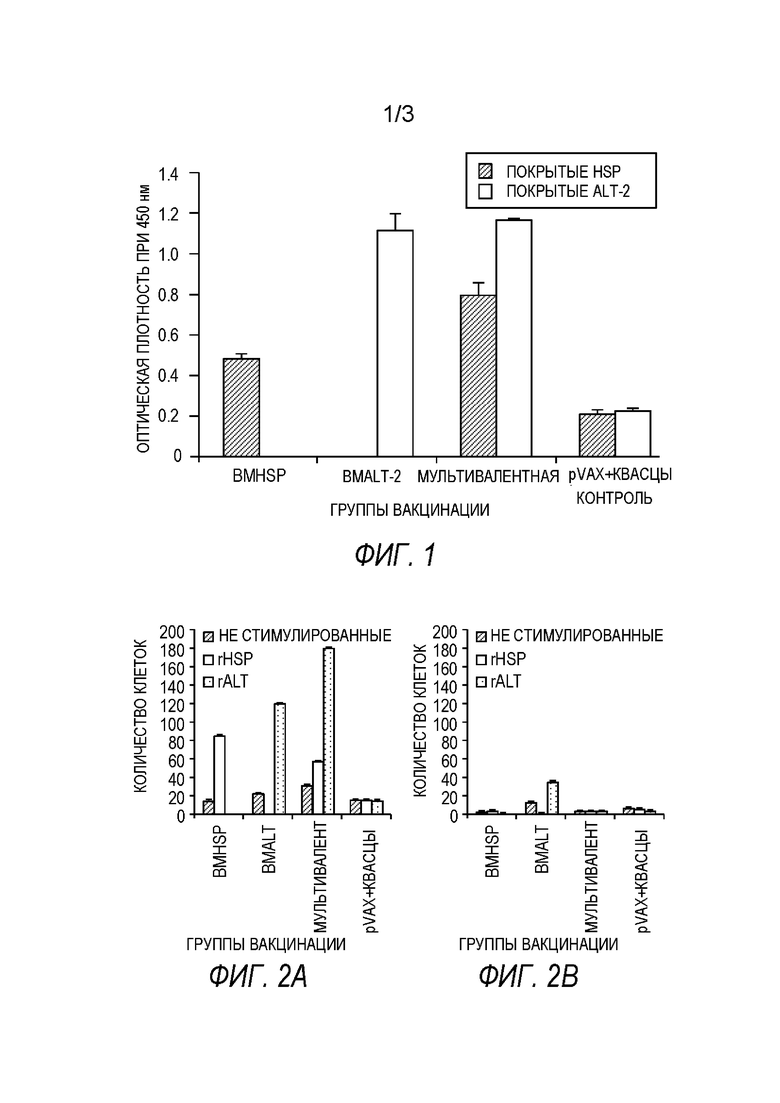
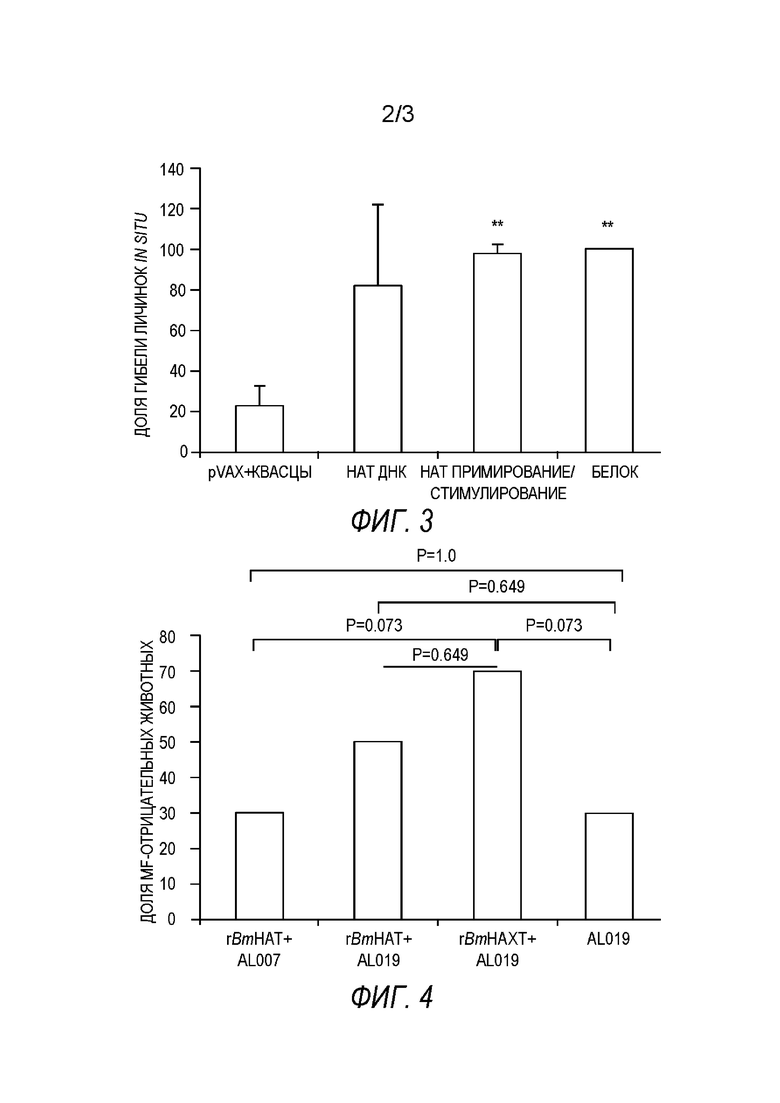
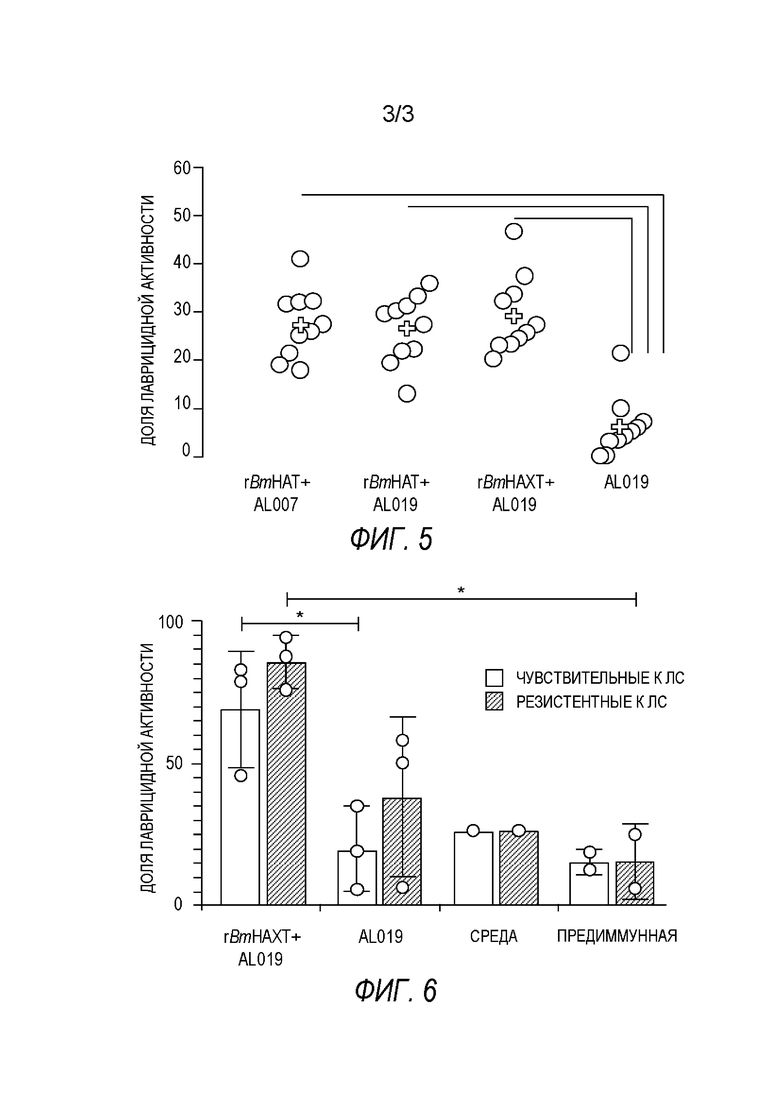
Изобретение относится к биотехнологии. Предложена поливалентная иммуногенная композиция против дирофиляриоза, содержащая два или несколько антигенов из Dirofilaria immitis, где антигены выбраны из многокопийного личиночного транскрипта, малого белка теплового шока (HSP) 12.6, тетраспанина и тиоредоксинпероксидазы 2. Предложены рекомбинантный вектор для доставки молекулы нуклеиновой кислоты в клетку-хозяина и рекомбинантная клетка-хозяин для продуцирования белка. Предложен способ индукции защитного иммунного ответа у субъекта, включающий введение поливалентной иммуногенной композиции, и способ иммунизации животного против дирофиляриоза. Комбинации антигенов, таких как многокопийный личиночный транскрипт (ALT2), тетраспанин (TSP), малый белок теплового шока (HSP) 12.6 и тиоредоксинпероксидаза 2 (TPX-2), обеспечивают >80% защиту от заражения филяриозными нематодами, такими как, например, Dirofilaria immitis. 5 н. и 11 з.п. ф-лы, 6 ил., 22 табл., 14 пр.
1. Поливалентная иммуногенная композиция против дирофиляриоза, содержащая два или несколько антигенов из Dirofilaria immitis, где антигены выбраны из многокопийного личиночного транскрипта, малого белка теплового шока (HSP) 12.6, тетраспанина и тиоредоксинпероксидазы 2.
2. Поливалентная иммуногенная композиция по п. 1, отличающаяся тем, что антигены имеют белковую основу, ДНК основу или их комбинацию.
3. Поливалентная иммуногенная композиция по п. 1, отличающаяся тем, что многокопийный личиночный транскрипт содержит SEQ ID NO: 98 или SEQ ID NO: 99; малый белок теплового шока 12.6 содержит SEQ ID NO: 100 или SEQ ID NO: 81; и тиоредоксинпероксидаза 2 содержит SEQ ID NO: 83 или SEQ ID NO: 101.
4. Поливалентная иммуногенная композиция по п. 3, отличающаяся тем, что многокопийный личиночный транскрипт содержит SEQ ID NO: 93; малый белок теплового шока 12.6 содержит SEQ ID NO: 91; и тиоредоксинпероксидаза 2 содержит SEQ ID NO: 95.
5. Поливалентная иммуногенная композиция по п. 1, отличающаяся тем, что антигены присоединены ковалентно.
6. Поливалентная иммуногенная композиция по п. 1, отличающаяся тем, что она дополнительно содержит адъювант.
7. Рекомбинантный вектор для доставки молекулы нуклеиновой кислоты в клетку-хозяина, содержащий нуклеиновые кислоты, кодирующие поливалентную иммуногенную композицию по п. 1.
8. Рекомбинантная клетка-хозяин для продуцирования белка, содержащая рекомбинантный вектор по п. 7.
9. Способ индукции защитного иммунного ответа у субъекта, включающий введение поливалентной иммуногенной композиции по п. 1 субъекту, тем самым индуцируя защитный иммунный ответ у субъекта.
10. Способ по п. 9, отличающийся тем, что он дополнительно включает введение субъекту одной или нескольких дополнительных доз поливалентной иммуногенной композиции.
11. Способ по п. 9, отличающийся тем, что иммуногенную композицию вводят путем подкожной или внутримышечной инъекции.
12. Способ по п. 9, отличающийся тем, что поливалентную иммуногенную композицию вводят с адъювантом.
13. Способ иммунизации животного против дирофиляриоза, включающий введение субъекту поливалентной иммуногенной композиции по п. 1, где антигены выбраны из антигена многокопийного личиночного транскрипта, содержащего SEQ ID NO: 121 или SEQ ID NO: 122; антигена малого белка теплового шока 12.6, содержащего SEQ ID NO: 81 или SEQ ID NO: 123; антигена тетраспанина, содержащего SEQ ID NO: 82; и антигена тиоредоксинпероксидазы 2, содержащего SEQ ID NO: 83 или SEQ ID NO: 124, тем самым иммунизируя субъект против дирофиляриоза.
14. Способ по п. 13, отличающийся тем, что он дополнительно включает введение субъекту одной или нескольких дополнительных доз поливалентной иммуногенной композиции.
15. Способ по п. 13, отличающийся тем, что иммуногенную композицию вводят путем подкожной или внутримышечной инъекции.
16. Способ по п. 13, отличающийся тем, что поливалентную иммуногенную композицию вводят с адъювантом.
| SHARON M | |||
| et al., Immunisation of mice with fractions derived from the intestines of Dirofilaria immitis, INTERNATIONAL JOURNAL OF PARASITOLOGY, vol | |||
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| Насос | 1917 |
|
SU13A1 |
| Коловратный двигатель | 1924 |
|
SU1459A1 |
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| KLIMOWSKI L | |||
| et al., Molecular cloning, expression and enzymatic activity of a thioredoxin peroxidase from Dirofilaria | |||

