[0001] Данное изобретение относится к иммуногенной композиции и способу ее получения.
Предпосылки изобретения
[0002] Несмотря на глубокие усилия не существует безопасной, лечебной вакцины против ВИЧ. Изобретатели уделяли внимание различным этапам жизненного цикла ВИЧ. На сегодняшний день исследование не обнаружило композицию, которая бы стимулировала эффективный иммунный ответ против подавляющего иммунитет ретровируса ВИЧ-1 и ВИЧ-2. Большинство ВИЧ вакцин используют части оболочечных поверхностных гликопротеинов (gp160, gp120 и gp41) вируса с попыткой индуцировать продукцию нейтрализующих антител против оболочечных спайков вируса. Некоторые из них были успешными в продуцировании высоких титров антител. В основе этого подхода лежит принцип, что антитела, связывающиеся с гликопротеинами, будут нейтрализовать вирус и предотвращать инфекцию. Функционирующая иммунная система сможет затем активировать систему комплемента, которая функционирует каскадом, лизируя и уничтожая вирус. Тем не менее, вирус способен избегать иммунной системы с проворством и простотой. На сегодняшний момент не существует отдельных публикаций, посвященных анализу конкретного примера индивидуума, зараженного СПИДом, проявляющего приемлемый иммунный ответ и элиминирование вируса. Таким образом, не существует маркера для иммунитета с заболеванием ВИЧ.
[0003] Ряд лекарственных средств и композиций (например, AZT (зидовудин), ddl (диданозин), ddC (зальцитабин), d4T (ставудин) и ЗТС (ламивудин)) ингибирует обратную транскрипцию. Эти 2′,3′-дидеоксинуклеозидные аналоги могут быть эффективными против определенных штаммов, но они чувствительны к геномной изменчивости ВИЧ (Deeks, Ch. 6). Эти медикаменты также сталкиваются с проблемами токсичности, стоимости, схем комплексного лечения, межлекарственного взаимодействия, а также лекарственной резистентности.
[0004] Вакцины к патогенам более ограничены в масштабах своего действия, чем противомикробная терапия. Антибиотик может иметь множество подтвержденных применений для различных бактериальных инфекций из различных источников. Вакцина, тем не менее, если она эффективна, лишь защищает индивидуум от заражения специфической болезнью. Однако границы могут быть расширены, если вакцина состоит из патогена из родственного инфекционного элемента. Классическим примером этого является противооспенная вакцина, которая является вакцинией. Этот вирус имеет сходство с коровьей оспой и может быть получен из пассажа коровьей оспы и/или натуральной оспы через векторы животных (Flint, 2004, р.6). В 19-м веке Эдвард Дженнер (Edward Jenner) отметил, что пациенты, которые перенесли коровью оспу, обладали иммунитетом к натуральной оспе. Вслед за этим наблюдением он разработал вакцину, полученную из коровьей оспы, для предотвращения натуральной оспы. Противооспенная вакцина по Дженнеру использует один патоген для того, чтобы вызвать иммунный ответ к двум патогенам (Wagner, et al., pp.102-108).
[0005] Доступные в настоящее время вакцины попадают в одну из восьми широких категорий: (1) живая аттенюированная (ослабленная) (вакцина Сэбина против полиомиелита, корь, свинка, краснуха, желтая лихорадка, ветряная оспа (ветрянка), бацилла Кальметта-Герена (туберкулез), брюшной тиф (Salmonella typhi), бешенство (для собак и других животных)); (2) инактивированная цельно-вирионная или бактериальная (бешенство (для людей), грипп, гепатит А, коклюш (Bordatella pertussis), паратиф (сальмонеллез) (Salmonella paratyphi), сыпной тиф (Rickettsia prowazekii), чума (Yersinia pestis)); (3) субъединичная (гепатит А или В); (4) инактивированный токсин или анатоксин (столбняк, дифтерия); (5) вакцина по Дженнеру (натуальная оспа); (6) рекомбинантная живая (бешенство для животных с использованием вектора коровьей оспы); (7) конъюгированная (менингит), и (8) очищенный капсулярный полисахарид (менингит (Haemophilus грипп) и Streptococcal pneumonia).
[0006] Живые вакцины вызывают острую инфекцию в организме хозяина. Тем самым гуморальная и клеточная ветви иммунной системы отвечают координированным ритмическим образом для искоренения инфекции. В результате, возможен длительный, если не пожизненный, иммунитет. Другое преимущество живой вакцины может быть осуществлено, если вектор экскретируется иммунизированным хозяином. Неиммунизированный пациент может заразиться инфекцией и, следовательно, стать иммунизированным. Случайный и часто смертельный результат может иметь место, если неиммунизированный пациент был неподходящим кандидатом. Хотя концепция "коллективного иммунитета" может быть реализована с живым вектором (Levinson, pp.247-243). Зачастую необходима лишь одна вакцинация. Живые вакцины обычно состоят из ослабленного, невирулентного или относительно невирулентного вектора. Неудобством данного метода вакцинации является возможность возникновения обратной мутации, что компенсирует вирулентность организма. Более того, некоторые индивидуумы будут подвергаться действию относительно авирулентного вектора часто из-за скрытого иммунологического заболевания, сопутствующей болезни или предшествующего состояния. Классическим примером этого может быть введение противооспенной вакцины пациенту с экземой или псориазом, но по другим показателям имеющему хорошее состояние здоровья. Эти пациенты зачастую развивали рассеянную скоротечную форму болезни и становились жертвами вакцины.
[0007] Убитые цельно-вирионные или полученные из бактерий вакцины характеризуются большим коэффициентом безопасности. Инфекционное заболевание не будет возникать по причине вакцины, если вирус потерял способность к репликации. Таким образом, эффект "коллективного иммунитета" неприменим к вакцине Солка таким же образом, как и к вакцине Сэбина. Недостатком убитых цельно-вирионных вакцин является то, что они, в общем, вызывают слабый иммунный ответ, если вообще он имеет место. Без репликации патогена иммунологическое распознание часто вообще не осуществляется. Дополнительным недостатком является отсутствие системного ответа на убитый или неспособный к репликации вектор. Вакцина Солка для полиомиелита, полученная генной инженерией инактивированная вакцина, не приводит к иммунитету слизистых оболочек. Другими словами IgA ответ не осуществляется. Более того, цитотоксический Т-клеточный ответ не имеет места или зачастую неэффективен из-за отсутствия внутриклеточной репликации инактивированными векторами. Без внутриклеточной репликации, которая происходит в случае живых векторных вакцин, вирусные белки не проникают в путь: цитозольная протеасома, ТАР, эндоплазматический ретикулум, Гольджи, что необходимо для ассоциации вирусных эпитопов с белком MHC-I. Эпитопы патогена, присутствующие в окружении белков MHC-I, вызывают CD8* (или Th-1) ответы.
[0008] Убитые вакцины подвергаются альтернативному иммунному ответу на патогенные эпитопы. Интернализация убитой вакцины происходит в результате либо эндоцитоза, либо фагоцитоза. Какой бы механизм захвата не использовался, в результате появляется внутриклеточная органелла, известная как эндоцитозная везикула или фагосома. Мембрана везикулы происходит от плазматической мембраны и содержимое просвета содержит цитоплазму и материал, имеющий внеклеточное происхождение. Посредством действия протонных насосов на мембране везикулы, атомы водорода активно транспортируются в везикулу, закисляя содержимое. Эти везикулы далее сливаются с липосомами, которые содержат различные ферменты, активирующиеся в кислотной среде. Полученные фаголизосомы сбрасывают содержимое везикул, образуя различные пептиды и гликопротеины. В данной структуре фрагменты, имеющие патогенное происхождение, контактируют с белками MHC-II. Эти белки синтезируются в цитоплазматическом ретикулуме и транспортируются в фаголизосомы посредством аппарата Гольджи. Белки MHC-II взаимодействуют преимущественно с клетками CD4+, проявляя Th-2 опосредованный иммунный ответ, который ограничен по области применения сферой иммунологии, в которой он встречается. Тем самым убитые или инактивированные векторы вызывают гуморальный ответ или ответ антител, и иммунитет слизистых оболочек не осуществляется, несмотря на то, что они вводятся через слизистую. Кроме того, убитые векторы обычно требуют многократных введений вакцины, и отмеченный ответ иммунологической памяти обычно и менее длительный, чем наблюдающийся в случае живого вектора (Parham, 2005, pp.67-96; Levinson, pp.393-412; Kaufmann; 1997, pp.37-45).
[0009] Субъединичные вакцины направляют иммунологический ответ к основному структурному компоненту вторгающегося организма. Так как не происходит репликация, то существует большой коэффициент безопасности. Безопасность имеет свою цену: слабый, слегка определяемый иммунологический ответ, который является первично Th-2 опосредованным.
[0010] Он отличается от Th-1 опосредованного иммунного ответа, который является предпочтительным для всех находящихся внутри клетки патогенов, включая вирусы. Т-клетки отвечают на антигены только в контексте молекул МНС (главный комплекс гистосовметимости) на антиген-презентующих клетках (В-клетки, макрофаги и дендритные клетки) (Peter Parham, 2005, Ch. 3, pp.67). Более подробно, Th-1 ответ зависит от презентации антигена, связанного с белком MHC-I. Внутриклеточно расположенные организмы разрушаются с помощью пути ТАР-протеасомы (Peter Parham, 2005, Ch. 3, pp.67-96). Клетка направляет протеолиз в этот путь при помощи конъюгирования белка со множественными остатками убиквитина посредством иерархической серии ферментов (E1, E2 и Е3) (Krauss, pp.101-113; Parham, 2005, pp.81).
[0011] Вакцины, основанные на нуклеотидах (ДНК или РНК), используют гены ВИЧ. Клеточная транскрипция в хозяине и механизм трансляции продуцирует белки ВИЧ. Иммунологический ответ на эти белки является предсказуемым. Сама по себе нуклеиновая кислота не является центром предположительного иммунного ответа, но вызывает клеточные эффекторные механизмы, созданные для ее разрушения, зачастую оказывая влияние на эффективность вакцины. Вирусная нуклеиновая кислота распознается различными компонентами врожденного иммунного ответа как "чужеродная". Перед транскрипцией и трансляцией начальных фрагментов вирусной нуклеиновой кислоты хозяин элиминирует ее. При введении должным образом ВИЧ заболевание не возникает. Будучи потенциально эффективными, основанные на нуклеотидах вакцины должны выполнять некоторые этапы. Они включают, но не ограничиваются, следующее. Первый этап представляет собой клеточный захват интактной, немодифицированной вирусной нуклеиновой кислоты. Второй этап представляет собой уклонение от множества цитоплазматических ферментов хозяина, направленных на разрушение нуклеиновой кислоты, произошедшей от патогена. Также направляются многие компоненты врожденного иммунного ответа. Врожденный ответ на вирусную нуклеиновую кислоту не вызывает иммунитет. Тем самым, защитный эффект вакцинации не может быть реализован, если вирусная нуклеиновая кислота разрушена цитоплазмой или нуклеоплазмой в клетке. Третий этап представляет собой ассимиляцию вирусной нуклеиновой кислоты в ядре. Это может быть достигнуто либо путем прохождения через поры ядра, что является высоко регулируемым процессом, либо прорыва через ядерную мембрану, что потенциально может нарушить нормальное функционирование клетки и препятствовать предполагаемому иммунному ответу. Четвертый этап представляет собой встраивание вирусной нуклеиновой кислоты в ДНК хозяина. Пятый этап представляет собой транскрипцию вирусного гена(генов). Шестой этап представляет собой трансляцию вирусных генов в цитоплазму хозяина. Все эти этапы могут завершаться в строго иммунологической области до возникновения клеточного ответа. Завершение этих этапов, тем не менее, неравноценно эффективному иммунологическому ответу. Нуклеотидные вакцины недоступны коммерчески. Таким образом, существует необходимость в ВИЧ вакцине с высоким уровнем безопасности и эффективности.
Описание изобретения
[0012] Данное изобретение представляет собой иммуногенную композицию или вакцину и способ получения иммуногенной композиции, обладающей характеристиками высокой безопасности субъединичной вакцины, комбинированными с эффективностью живой вакцины, которая способна вызывать Th-1 опосредованный иммунный ответ. Альтернативно традиционным подходам данное изобретение основано на условно живом вирусе; то есть, вирус, неспособный к репликации другим способом, способен к репликации на протяжении ограниченного промежутка времени при добавлении экзогенного белка, который замещает белок, недоступный из-за модификации или делеции соответствующей генной последовательности, кодирующей этот белок в вирусном геноме (или "условно живом"). Один вариант осуществления данного изобретения использует нокаутированный вирион, в котором мишенью являются один или несколько специфических вирусных белков. В каждом варианте осуществления данного изобретения дефицит белка, соответствующего "нокаутированной" генной последовательности-мишени, может быть добавлен экзогенно. Полагают, что предопределенное количество экзогенно добавленного белка-мишени может быть необходимо для того, чтобы вирион, неспособный к репликации другим способом, достигал желаемого определенного или ограниченного во времени и количественно уровня репликации. Количество, время полужизни, внутриклеточная концентрация, внутриклеточная локализация и конформационная структура экзогенно добавленного белка, обеспеченного в коктейле с условно живым вирусом, будут регулировать кинетические параметры репликации иммуногенной композиции или вакцины.
[0013] В понимании объема данного изобретения термин "включающий" и его производные, использованные в данном описании, подразумевает под собой не ограничивающие термины, которые определяют наличие сформулированных признаков, элементов, компонентов и/или этапов, но не исключают наличия других, не сформулированных признаков, элементов, компонентов и/или этапов. Вышеупомянутое также относится к словам, имеющим сходное значение, такие как термины "содержащий", "имеющий" и их производные.
[0014] Один аспект данного изобретения заключается в том, что хозяин является незащищенным по отношению к полному или практически полному набору иммуногенов, включающих патоген в контексте инфекции, что может оказаться нормальным по отношению к иммунной системе. Этот подход охватывает эффективность, широту иммунологического ответа и длительную память живого вирусного вектора. В зависимости от контекста введенной композиции или вакцины может также реализовываться коллективный иммунитет. Кроме того, данное изобретение осуществляет безопасность убитой или субъединичной вакцины, потому что полученный вирион будет неспособным к репликации отсутствие экзогенного белка, соответствующего модифицированному гену белка репликации или соответствующей мРНК.
[0015] Данное изобретение может рассматриваться как имеющее два компонента, каждый из которых является иммуногенным. Первый компонент представляет собой интактный вирион, модифицированный в вирусной ДНК или мРНК для того, чтобы иметь дефективные последовательности, лишенные части или цельного гена(генов) или мРНК, кодирующих один или несколько белков. Альтернативно гены, кодирующие белки-мишени, могут быть замещены нуклеиновыми кислотами, неспособными к трансляции. Второй компонент представляет собой/представляют собой добавление одного или нескольких экзогенных белков, соответствующих тем, которые не кодируются в вирусной ДНК или РНК последовательности. Иммуногенность первого компонента является бифазной. При введении без дополнения дефицитным белком(белками) реализуется иммуногенная композиция или вакцина, основанная на цельном вирусном вирионе, неспособном к репликации. При введении с дополнением экзогенным белком можно получить условно живой вирион, контролируемый во времени и лимитированный в репликации, с улучшенной безопасностью. Как только прекращается репликация, останется неспособный к репликации и неинфекционный вирион. Тем самым, исходный пункт и конечный пункт данной композиции является одинаковым: цельный вирион, неспособный к репликации, который может функционировать как иммуногенная композиция.
[0016] В одном варианте данного изобретения Th-1 ответ будет проявляться и направляться на один или несколько компонентов интактного вириона. Введенный или добавленный белок(белки) в концепции иммунного ответа интактного хозяина представляет субъединичную вакцину. Так как первый компонент является бифазным, иммуногенная композиция представляет собой в действительности три компонента вакцины, введенных одновременно: (1) цельный интактный неспособный к репликации вирус; (2) (условно) живой вирион (с временно контролируемым временем полужизни, измеренным в часах или днях); и (3) субъединичная вакцина(ы). Теоретически каждый компонент вакцины будет служить в качестве вспомогательного средства для двух других, усиливая общую иммуногенность формуляции вакцины. Множество векторных вакцин, таких как MMR и DPT, проявляли позитивные ответы на каждый компонент. Каждый вектор трехвалентных вакцин усиливает иммуногенность двух других.
[0017] Один вариант осуществления данного изобретения обеспечивает быстродействующую систему для образования условно живой вакцины в комбинации без тканевой культуры. Антигены, произошедшие от клеточной культуры, подвергаются риску быть ассимилированными тканевыми антигенами, к которым может иметь место непредвиденный иммунный ответ, что ограничит эффективность вакцины. Более того, вирус в клеточной культуре будет продолжать мутировать в ответ на клеточное окружение хозяина. Клеточная культура неточно отображает иммунные системы интактного хозяина. Вирус, произошедший от клеточной культуры, не отражает полевые или клинические изоляты. Продолжающаяся вирусная мутация в тканевой культуре непредсказуемо ведет к недостатку репродуктивности, компрометируя контроль качества в репликации вириона и получение вакцинной продукции. Таким образом, тканевой вирус является условно оптимальным для введения вакцины.
[0018] Данное изобретение не полагается на живой рекомбинантный носитель. Начальные иммунные ответы к рекомбинантным векторам ограничены частично или в целом вектором носителя самим по себе, что ставит под угрозу эффективность. Последующие вакцинные провокации живого рекомбинантного носителя вызывают быстрый адаптивный иммунный ответ на вектор носителя, уничтожающий вектор и антигенный материал, полученный генной инженерией. Концепция "первичный антигенный стимул" не дает возможности более широкому иммунному ответу с последующими вакцинными провокациями. Повторные вакцинные провокации усиливают приобретенный иммунный ответ в терминах специфичности и надежности, но ответ проявляется только к тем антигенам, по отношению к которым изначально был направлен иммунологический ответ. Таким образом, рекомбинантные векторы являются антигенно ограничивающими в иммунном распознавании и ограниченными одним введением. Другими словами, рекомбинантная векторная система позволяет отдельное применение к индивидууму, так как после первого представления индивидуум будет развивать иммунитет к самому вектору. Кроме того, рекомбинантные векторные системы представляет иммунной системе широкий спектр антигенных материалов, к которым иммунологический ответ не является желаемым. Вектор сам по себе является источником этой антигенной ловушки. Потенциальное количество антигенов, включающих вектор носителя сам по себе, значительно превышает количество генетически встроенных материалов, к которым иммунный ответ является желательным. Более того, наружные белки рекомбинантного вектора представляют собой первые антигены, действию которых подвергается иммунная система. "Повторные дозы" рекомбинантных векторов не являются полезными. Данное изобретение преодолевает эти недостатки, с которыми в настоящее время сталкиваются вакцины, веденные в рекомбинантные векторные системы.
Краткое описание графических материалов
[0019] Схематическое изображение линейного генома ВИЧ-1 с кодирующими последовательностями ВИЧ генов, изображенных как открытые прямоугольники.
[0020] Схематическое изображение линейного генома ВИЧ-2 с кодирующими последовательностями ВИЧ генов, изображенных как открытые прямоугольники.
Лучший способ(ы) для выполнения данного изобретения
Введение
[0021] Данное изобретение основано на условно живом вирионе; то есть, вирионе, модифицированном так, чтобы неспособный другим образом к репликации был способен к репликации на протяжении ограниченного промежутка времени при добавлении экзогенного белка, который замещает белок, недоступный из-за модификации (или делеции) соответствующей генной последовательности, кодирующей этот белок в вирусном геноме. Вирус по определению не является живой или мертвой структурой. Он наилучшим образом характеризуется как являющийся способным к репликации или неспособным к репликации. В данном изобретении живой вирус относится к вирусу, способному к репликации. Один аспект данного изобретения представляет собой иммуногенную композицию, включающую вирусную ДНК или РНК, представляющую полный вирусный геном, в котором, по меньшей мере, один ген белка репликации или соответствующая мРНК модифицирована для того, чтобы компенсировать неспособность к репликации вирусной ДНК или РНК; эта модифицированная вирусная ДНК или РНК затем инкапсулируется вирусными белками, которые являются самособирающимися в бесклеточной системе экспрессии, формируя условно живой вирион. Способ получения этого условно живого вириона включает этапы обеспечения, по меньшей мере, одной вирусной молекулы ДНК или РНК, представляющей полный геном, амплификацию вирусной ДНК или РНК, модификацию вирусной ДНК или РНК, по меньшей мере, в одном гене белка репликации или соответствующей мРНК, отбор амплифицированной и модифицированной вирусной ДНК или РНК, переупаковку отобранных ДНК или РНК в бесклеточной системе экспрессии, пригодной для самосборки вирусных частиц, и отбор желаемого количества полученных условно живых вирионов. Альтернативный способ получения этого условно живого использует традиционную систему клеточной культуры. В данном способе вирион, модифицированный, по меньшей мере, в одном гене белка репликации или соответствующей мРНК, может быть культивирован в условиях, пригодных для вирусной репликации с добавлением экзогенного белка, соответствующего, по меньшей мере, одному гену белка репликации или соответствующей мРНК. Таким образом, четвертый аспект данного изобретения представляет собой формулирование вакцины с применением вириона, не способного к репликации, в комбинации с белками цельного вируса, фрагментами белка, производными белка или их комбинациями. Вакцина, созданная любым из способов, будет иметь тройные иммуногенные свойства, которые проявляются 1) цельным интактным не способным к репликации вирусом; 2) условно живым вирионом, временно оживленном при добавлении белковых добавок; и 3) белковой добавкой самой по себе, действующей как субъединичная вакцина. Дополнительным признаком вакцины, сформулированной с условно живым вирионом, созданной с бесклеточной системе, является то, что не существует векторов, вносящих свой вклад в вызываемый иммунологический ответ вакцины при ее введении.
Предпочтительная нуклеотидная поледовательность-мишень гена белка репликации или соответствующая мРНК
[0022] Предпочтительно для варианта осуществления, направленного на ВИЧ, нуклеоитдная последовательность(и)-мишень(и) расположена в центральном участке генома ВИЧ и необходима для вирусной репликации. Другая нуклеотидная последовательность(и) согласно данному изобретению может быть мишенью делеции или замещеннной генетической информацией, не способной к трансляции. Такие изменения включают, но не ограничиваются, следующее: оболочечные гликопротеины, gp120 и gp41, ферменты, кодируемые ретровирусами (протеаза, обратная транскриптаза, интеграза и РНКазаН), Nef и длинные концевые повторяющиеся последовательности, настолько длинные, что полная модификация приводит к вириону, не способному к репликации. В целом, тем не менее, для целей данного применения, ген белка репликации представляет собой ген, который может быть модифицирован или удален для того, чтобы компенсировать некомпетентность репликации вириона, при нахождении в интактном хозяине. Таким образом, ген белка репликации или соответствующая мРНК для целей данного применения означает последовательность нуклеиновой кислоты, кодирующую белок. Тем не менее, белок(белки), недостающий в транскрипции вирусного генома, может быть добавлен экзогенно. Это будет приводить к активной "нормальной" вирусной репликации в интактном хозяине. Также для целей данного изобретения модифицирование (или модификация) гена белка репликации должно быть истолковано широко, так чтобы включать делецию или мутацию, например, настолько длинную, чтобы вирион компенсировал бы некомпетентность репликации в отсутствие экзогенного белка репликации, как описано далее в описании изобретения. В одном варианте осуществления ВИЧ по данному изобретению, например, вирусные белки-мишени представляют собой Vif, Vpr, Vpu (ВИЧ-1), Tat экзон 1, и Vpx (ВИЧ-2). Относительно маленькие по размеру, они также кодированы, частично, не перекрывающимися сегментами и представляют собой все необходимые белки для вирусной репликации. Удаление одного или нескольких из этих неперекрывающихся геномных сегментов будет приводить к неспособности вируса к репродукции in vivo за исключением добавления экзогенного источника дефективного или дефицитного белка.
[0023] Нуклеотидная последовательность vif локализована 3′ к роl нулкеотидной последовательности и 5′ к vpr нуклеотидной последовательности. В некоторых вирусных изолятах существует небольшое перекрывание между нуклеотидными последовательностями на 3′ конце роl и 5′ конце vif. 3′ концевая нуклеотидная последовательность vif перекрывается с 5′ концом vpr. Белок vif кодируется одним экзоном. Неперекрывающийся сегмент vif между роl и vpr может быть селективно удален, что компенсирует vif дефективный вирус без негативного влияния на продукты транскрипции и трансляции pol и vpr.
[0024] Белок Vif (вирусный белок инфективности) включен в ВИЧ-1 и ВИЧ-2 вирионы посредством взаимодействия с вирусной РНК и нуклеопротеиновыми комплексами. Vif составляет приблизительно 216 аминокислот в длину. Vif не является структурным белком и является Rev зависимым и, таким образом, продуцируется поздно в жизненном цикле вируса. Vif дефектные вирионы in vitro являются в 103 раз менее заразными, чем интактный вирус с функциональным vif геном. Vif дефективные вирионы in vivo являются не способными к репликации, Vif имеет множество функций, включая, но не ограничиваясь, следующее:
1) повышает вирусную инфективность;
2) усиливает сборку вириона;
3) содействует синтезу вирусной ДНК с помощью фермента обратной транскриптазы;
4) противодействует клеточному белку CEM15/APOBEC3G (аполипопротеин В РНК-обрабатывающий фермент или аполипопротеин В РНК-катализирующий фермент); APOBEC3G, компонент врожденной иммунной системы, представляет собой цитидиновую деаминазу;
5) содержит ингибиторную последовательность (INS), которая предотвращает преждевременный ядерный экспорт вирусной РНК в цитоплазму;
6) вызывает структурные изменения плазматической мембраны;
7) вносит вклад в нарушение регуляции цитокинов, ингибирует фагоцитоз и ограничивает рассеивание клеток;
8) защищает вирусную РНК от внутриклеточного разрушения РНКазой;
9) временно регулирует активность фермента протеазы.
[0025] Условно живой вирус, в котором неперекрывающаяся нуклеотидная последовательность для vif срощена, не способен к вирусной репликации и инфицированию. Белок Vif производится в избытке по сравнению с тем, что необходимо в инфицированной клетке. Большая часть избытка белка Vif ассимилируется в неинфицированных клетках, где этот избыток вызывает много, если не большинство нарушений регуляции цитокинов и, тем самым, иммуносупрессорный эффект. Экзогенный запас Vif белка не только ограничивает внутриклеточную репликацию, но также ограничивает vif иммуносупрессию.
[0026] Нуклеотидная последовательность vpr локализована 3′ к нуклеотидной последовательности vif и 5′ к Tat экзон 1 нуклеотидной последовательности. В большинстве вирусных изолятов 5′ конец перекрывается vpr с vif, а 3′ конец перекрывается с tat экзоном 1. Между 5′ и 3′ перекрывающимися сегментами существует неперекрывающийся сегмент, который может быть селективно удален. Дефективный мутант vpr не способен к активной репликации в интактном хозяине. Нуклеотидные последовательности vif и tat экзон 1 остаются интактными с селективным удалением промежуточного неперекрывающегося сегмента.
[0027] Белок Vpr (вирусный белок r) представляет собой поздний генный продукт как ВИЧ-1, так и ВИЧ-2. Vpr является rev зависимым. Vpr встроен в вирионы посредством взаимодействия с вирусным белком р6, который отщеплен от большего Gag полипептида.
[0028] Vpr имеет множество функций, включая следующие, но не ограничиваясь ними:
1) сигнал ядерной локализации;
2) координирует ВИЧ геномную экспрессию с помощью Tat белка;
3) блокирует клеточное деление инфицированных Т-клеток в G2 фазе клеточного цикла;
4) блокирует клеточный цикл в Gi фазе неинфицированных Т-клеток;
5) растворимый Vpr задерживает неинфицированные цитотоксические CD8 Т-клетки, специфические для ВИЧ антигенов, в G2 фазе клеточного цикла;
6) ограничивает В-клеточную соматическую гипермутацию, необходимую для созревания афинности рецептора антитела;
7) усиливает гетерогенность вирусной популяции путем увеличения вирусной мутации;
8) усиливает активность р300, коактиватора с гистонацетилазной активностью, который регулирует генную транскрипцию;
9) активирует факторы транскрипции NF-IL-6 и NF-kB;
10) взаимодействует с и коактивирует внутриклеточный глюкокортикоидный рецептор;
11) взаимодействует с и контролирует экспрессию целого ряда других клеточных белков, включая, но не ограничиваясь, следующие: Spl, p53, Rb (гиперфосфорилирование), TFIIB, факторы ядерного транспорта импортин-α и нуклеопорин Роm21 и человеческий гомолог MOV34; кооперативное взаимодействие белка Vpr и клеточных белков, p53 и Spl имеет позитивный эффект на генную транскрипцию ВИЧ-1;
12) вызывает изменения клеточного цитоскелета;
13) вызывает выпячивания ядерной мембраны, что, возможно, вносит свой вклад в ядерную локализацию;
14) вызывает дисфункции в проницаемости мембран митохондрий;
15) в ВИЧ-2 Vpr обнаруживает встраивание Vpx в вирионы; Vpx является вспомогательным белком, обнаруженным только в ВИЧ-2, необходимым для полной вирусной активности.
[0029] Условно живой вирион, из которого удален неперекрывающийся сегмент нуклеотидной последовательности vpr, не способен к активной репликации и инфицированию в интактном хозяине. В ВИЧ инфицированной клетке, белок Vpr производится в количестве, которое превышает необходимое для активной репликации. Избыток белка Vpr ассимилируется неинфицированными иммунными клетками и функционирует частично для подавления клеточной функции. В вакцине, основанной на vpr дефективном условно живом вирусе, экзогенная добавка белка Vpr ограничивает как вирусную репликацию, так и индуцированную вирусом иммунную супрессию.
[0030] Нуклеотидная последовательность vpu локализована между 3′ концом tat экзон 1 нуклеотидной последовательностью и 5′ концом env нуклеотидной последовательностью. В большинстве вирусных изолятов не существует перекрывающейся нуклеотидной последовательности tat и vpu. В отличие от перекрывающегося сегмент vpu и env обнаружен в большинстве вирусных штаммов. Неперекрывающийся сегмент vpu может быть селективно удален, что компенсирует vpu дефективность без негативного влияния на продукты транскрипции и трансляции env.
[0031] Белок Vpu обнаружен лишь в ВИЧ-1 и четырех штаммах SIV. Первичная аминокислотная последовательность Vpu является наименьшей из белков, кодируемых геномом ВИЧ, и варьирует по длине от 77 до 86 аминокислот. Она преимущественно локализована в RER (ретикуло-эндотелиальная система)/аппарате Гольджи. Белок Vpu не встроен в зрелые вирионы. Функциональный эквивалент, по меньшей мере, частично, белка Vpu в ВИЧ-2 приписывался гликопротеину Env. Продукция белка Vpu является Rev зависимой и, тем самым, отмечается позднее в цикле вирусной репликации. Мессенджер РНК, кодирующей белок Vpu, является бицистронным. В большинстве вирусных изолятов инициирующий кодон Vpu не является точной последовательностью Kozak или кодирует аминокислоту, отличную от метионина. Для многих, если не для большинства, вирусных изолятов инициирующая последовательность белка Vpu мРНК является A/GCCAATGG. (Последовательность Kozak, которая намного легче распознается с помощью рибосомального аппарата хозяина, представляет собой 5′-ACCAUGG-3′.) Инициирующий кодон гликопротеина Env представляет собой остаток метионина в точной последовательности Kozak. Протекающее считывание "Leaky scanning" позволяет направить вирус в транскрипционный аппарат хозяина для получения соответсвующего соотношения (1/10) белков Vpu/Env.
[0032] Хотя ВИЧ-2 не кодирует белок Vpu, Vpu-подобная активность обнаружена в субъединице gp36 TM. Если отбор и разделение функции является коррелятом эволюции и маркера созревания, то ВИЧ-2 является менее дифференцированным, чем ВИЧ-1. В филогенетическом древе более сложные организмы развиваются позднее в эволюции жизни и лучше приспособлены. В эволюционных терминах ВИЧ-2 является боле "поздним" вирусом. Таким образом, ВИЧ-2 должен быть более легкой мишенью, чем ВИЧ-1. Действительно обнаружено, что ВИЧ-2 менее заразный и менее вирулентный, чем ВИЧ-1. Таким образом, белок Vpu может быть генетическим маркером, отслеживающим ВИЧ на протяжении промежутка времени.
[0033] Белок Vpu (вирусный белок U) имеет множество функций, включая, но не ограничиваясь, следующее:
1) временно контролирует репликацию ВИЧ;
2) иммуносупрессия;
3) экспрессия Vpu усиливает вирусную экспрессию, мутацию и распространение в хозяине;
4) снижает экспрессию клеточного CD4 в хозяине;
5) вносит вклад в нарушение регуляции цитокинов и иммунологическую дисфункцию;
6) способствует злокачественной трансформации в ВИЧ;
7) усиливает высвобождение вириона;
8) снижает клеточную экспрессию MHC-I.
[0034] Условно живой вирион с удалением неперекрывающегося сегмента нуклеотидной последовательности vpu не способен к активной репликации и инфицированию интактного хозяина. Продукция белка Vpu в ВИЧ-инфицированной клетке превышает потребность клеток для вирусной репликации. Большая часть избытка белка Vpu ассимилируется другими неинфицированными иммунологическими клетками. Иммуносупрессия, присущая белку Vpu, в первую очередь относится к неинфицированным клеткам, которые получили белок Vpu. В vpu дефицитном условно живом вирусном векторе экзогенный белок Vpu является контролирующим фактором вирусной репликации. Кроме того, иммуносупрессия белка Vpu будет подобным образом контролироваться количеством и временем полужизни доставленного белка Vpu.
[0035] Нуклеотидная последовательность vpx представляет собой 3′ к нуклеотидной последовательности vif и 5′ к нуклеотидной последовательности vpr. Перекрывающаяся геномная последовательность между нуклеотидными последовательностями vif и vpx обнаружена в большинстве ВИЧ-2 штаммов. В отличие от этого vpx и vpr нуклеотидные последовательности большинства изолятов являются неперекрывающимися. Удаление неперекрывающейся нуклеотидной последовательности vpx компенсирует дефективность vpx вируса без негативного влияния на продукты транскрипции и трансляции Vif и Vpr белков.
[0036] Белок Vpx (вирусный белок х) упакован в вирионы посредством взаимодействия с р6 доменом Gag. Vpx обнаружен только в ВИЧ-2 и четырех штаммах SIV. Встраивание Vpx в интактный вирион опосредуется дилейциновым мотивом на N-концевом домене р6. Приблизительно эквимолярные количества Gag и Vpx включены в ВИЧ-2 вирионы. Vpx является маленьким гидрофобным белком приблизительно 100 аминокислот длиной с тремя амфипатическими α-спиралями. Vpx структурно относится к белку Vpr, но функционально отличается от него. Vpx не хватает сигнала ядерного экспорта, и он не вовлечен в задержку клеточного цикла.
[0037] Vpx имеет множество функций, включающих следующие, но не ограничиваясь:
1) сигнал ядерной локализации (СЯЛ, или NLS);
2) Vpx облегчает ВИЧ-2 инфицирование неделящихся клеток;
3) облегчает ВИЧ репликацию в макрофагах;
4) облегчает связывание вириона на цитоплазматической стороне плазматической мембраны;
5) влияет на презентацию антигена MHC-II;
6) усиливает активность обратной транскриптазы.
[0038] Условно живой вирион, дефицитный по нуклеотидной последовательности для vpx, не жизнеспособен в интактном хозяине. Инфицированная ВИЧ-2 клетка производит Vpx в избытке по сравнению с тем, что требуется. Большая часть избытка белка Vpx ассимилируется в неинфицированных иммунных клетках. Иммуносупрессия, связанная с Vpx, возникает в большей части в неинфицированных клетках, в цитоплазмы которых встроен белок Vpx. Экзогенная доставка белка Vpx в такой композиции на основе условно живого вириона делает возможным не только ограничение внутриклеточной репликации вируса в инфицированных клетках, но также ограниченную иммуносупрессию, вызванную Vpx в неинфицированных клетках.
[0039] Нуклеотидная последовательность tat (трансактиватор фактора транскрипции) экзон 1 локализована от 3′ до vpr нуклеотидной последовательности и от 5′ первоначально до vpu нуклеотидной последовательности. Перекрывающийся сегмент между vpr и tat экзон 1 обнаруживается в большинстве вирусных изолятов. Неперекрывающийся сегмент включает остаток нуклеотидной последовательности tat экзон 1. В большинстве изолятов эта нуклеотидная последовательность tat экзон 1 не перекрывается с vpu в ВИЧ-1. Vpu нуклеотидная последовательность не встроена в геном ВИЧ-2. Вирион, дефективный по tat (экзон 1), не способен к репликации в интактном хозяине.
[0040] Полный ВИЧ-1 Tat белок кодируется двумя отдельными экзонами. Посредством альтернативного сплайсинга, две формы белка Tat продуцируются в ВИЧ-инфицированных клетках. Первые 72 аминокислоты (NH2 домен) белка Tat являются необходимыми для вирусной репликации и кодируются одним транскриптом экзона. Второй экзон кодирует СООН-концевой домен, охватывающий аминокислоты 73-101. То есть, одна форма белка Tat отражает нуклеотидную последовательность только одного экзона, кодирующего NH2 домен, и составляет 72 аминокислоты длиной. Другая форма является продуктом обоих экзонов и составляет в длину 101 аминокислоту (один штамм ВИЧ заболевания имеет 86 аминокислоты белка Tat). СООН-концевой домен необходим для того, чтобы белок Tat приводил в действие многие иммуномодулирующие воздействия. Таким образом, альтернативная вакцина может кодировать только амино-концевой экзон белка Tat, кодируемого нуклеотидной последовательностью Tat экзон 1.
[0041] Белок Tat рано экспрессируется в цикле вирусной репликации и является независимым от rev. Белок Tat не включен в интактный вирион.
[0042] Белок Tat имеет множество функций, включающих, но не ограничиваясь, следующее:
1) вызывает активацию NF-KB;
2) ингибирует клеточную (у хозяина), но не вирусную, мРНК трансляцию;
3) истощает внутриклеточный циклин Т как в инфицированных, так и неинфицированных Т-клетках;
4) регулирует нижележащие этапы bcl-2 и вызывает апоптоз в неинфицированных гематопоэтических клетках;
5) регулирует вышележащие этапы bcl-2 в ВИЧ-инфицированных макрофагах, прерывая апаптоз;
6) вызывает нейрональную смерть в центральной и периферической нервной системе;
7) снижает способность участия клеток в организации кластеров Т-клеток;
8) активирует В-клетки и вызывает В-клеточную лимфому;
9) вызывает синтез иммуноглобулина путем стимулирования высвобождения IL-6;
10) ингибирует активность CD26 или дипептидиламинопептидазы IV на мембранах Т-клеток, блокируя обратную активацию Т-клеток;
11) блокирует фаголизосомальное слияние в моноцитах;
12) ингибирует экспрессию IL-2 и IL2R в CD4-клетках;
13) усиливает воспалительное окислительно-восстановительное состояние (окислительный стресс);
14) усиливает активность фактора некроза опухоли (TNF);
15) стимулирует высвобождение TGF-бета (дополнительная иммуносупрессия);
16) угнетает транскрипцию генов МНС I;
17) активирует пути JNK и ERK/MAPK в неинфицированных CD4-клетках;
18) стимулирует хемотаксис моноцитов;
19) угнетает промотор бета 2-микроглобулина;
20) ингибирует синтез IL-12;
21) вызывает ВИЧ-1 ко-рецепторный синтез (CCR5 и CXCR4) в неинфицированных, но Tat трансфецированных клетках, усиливая чувствительность неинфицированных макрофагов и Т-клеток к вирусу ВИЧ (облегчает инфективность вирусных штаммов, тропичных как к макрофагам, так и к Т-клеткам);
22) гиперактивирует Т-клетки посредством пути CD28;
23) усиливает рост саркомы Капоши;
24) ингибирует пролиферацию неинфицированных лимфоцитов в ответ на специфические антигены;
25) защищает ВИЧ-инфицированные Т-клетки от апоптоза, вызванного активацией;
26) вызывает апоптоз неинфицированных Т-клеток;
27) ингибирует цитотоксичность природных киллеров (NK);
28) регулирует вышележащую TRAIL продукцию в макрофагах;
29) повышает экспрессию TRAIL в неинфицированных моноцитах;
30) защищает ВИЧ инфицированные моноциты от TRAIL-опосредованного апоптоза;
31) регулирует вышележащие IL-4 рецепторы на В-клетках;
32) вызывает деменцию ВИЧ;
33) ухудшает функцию дендритных клеток;
34) уменьшает рецепторы маннозы на инфицированных и неинфицированных клетках;
35) усиливает транскрипцию вируса ВИЧ, по меньшей мере, в тысячу раз посредством связывания белка с пусковым участком генома, реагирующим на трансактивацию (TAR) на 5′ конце мРНК ВИЧ; специфически взаимодействует с петлевой областью в стволе TAR элемента;
36) усиливает активность клеточного комплекса РНК-полимеразы II в вирусной транскрипции.
[0043] Специфические примеры иммуногенной композиции, основанной на условно живом вирионе, и способы ее получения изложены ниже с применением белка Tat. Хотя для специалиста в данной области будет очевидно, что любые модификации или альтернативные варианты осуществления возможны, и данные специфические примеры приводятся лишь с целью иллюстрации и не ограничивают изобретение, только если это не оговаривается отдельно.
[0044] Например, условно живой вирион, в котором удалена неперекрывающаяся нуклеотидная последовательность tat экзон 1, является не способным к вирусной репликации и инфицированию. Один аспект следующего варианта осуществления представляет собой не только ограниченную репликацию условно живого вириона, но также ограниченную иммуносупрессорную функцию белка Tat как иммуногена и в вирусной транскрипции. Белок Tat является высококонсервативным среди штаммов ВИЧ. Более того, белок Tat является высоко иммуносупрессорным, и его эффекты были документально подтверждены. Клетка, инфицированная вирусом ВИЧ и активно реплицирующаяся, производит многие вирусные компоненты, которые не ассимилируются в интактном вирионе или используются для вирусной репликации. Белок Tat в таких клетках производится в избытке, по сравнению с тем, что необходимо для репликации. Функцией избытка белка Tat является подавление иммунной системы хозяина. Экзогенная поставка белка Tat для tat дефективного условно живого вириона будет обеспечивать ограниченную репликацию вируса ВИЧ и ограниченную Tat-опосредованную иммуносупрессию (см., например, Rubartelli, et al.).
[0045] Белок ВИЧ Tat может быть подразделен на несколько определенных регионов, каждый из которых обладает специфическими физическими, стерическими и электростатическими свойствами. Короткая из двадцати аминокислот последовательность, состоящая из "ядерного" домена Tat, особенно аминокислоты 21-40, необходима для размножения ВИЧ in vitro. Белок Tat кодируется в ВИЧ двумя отдельными экзонами. Таким образом, цельный интактный вирус, не способный к репликации, может быть получен с помощью сплайсинга. Например, первый экзон может быть изменен или удален, второй экзон может быть изменен или удален, или оба экзона могут быть изменены или удалены. Первые 72 аминокислоты (NH2 домен) белка Tat являются необходимыми для вирусной репликации ВИЧ и кодируются одним транскриптом экзона. Второй экзон кодирует СООН-концевой домен, включающий аминокислоты 73-101. Таким образом, вариант осуществления данного изобретения может быть основан на нуклеотидной последовательности ВИЧ, имеющей только один экзон, кодирующий NH2 домен, и составляющий 72 аминокислоты в длину. СООН-концевой домен необходим для того, чтобы белок Tat проявил множество иммуномодулирующих эффектов. Таким образом, другой аспект данного изобретения может включать нуклеотидную последовательность ВИЧ, кодирующую только карбоксил-концевой экзон белка Tat. Следующий аспект данного изобретения может включать мутированную сплайсингом нуклеотидную последовательность на одном или обоих экзонах. Второй экзон белка Tat перекрывается внутри гена env полностью. Нуклеотидная последовательность rev экзон 2 полностью включена в нуклеотидные последовательности tat экзон 2. Должны быть созданы специальные условия для сохранения функции генов env и rev экзона 2. В одном варианте осуществления, сайты сплайсинга (как 5′, 3′, так и оба 3′ и 5′ сайта сплайсинга могут быть компенсированы нефункциональным терминированием транскрипции tat экзона 2) для нуклеотидной последовательности tat экзон 2 могут быть мутированы таким образом, что сплайсинг этих сайтов является невозможным, но не происходит никаких значительных изменений, если это имеет место, в аминокислотной последовательности гена env или rev экзон 2.
[0046] Альтернативно, определенные специфические бессмысленные или нонсенс-нуклеотидные последовательности для tat компенсируют неспособность вируса к репликации. Эти последовательности, кодированные в итнактную последовательность РНК ВИЧ, могут быть использованы в интактной вирусной структуре. Замещение глицина на цистеиновый остаток в аминокислотном положении номер 22 (C22G) или 30 (C30G) белка Tat отменяет Tat-опосредованную трансактивацию LTR ВИЧ. Замещение цистеинового остатка номер 31 на глицин снижает, но не полностью ингибирует вирусную трансактивацию ВИЧ Tat. Это будет частично приемлемо для вириона, кодирующего только экзон I с описанным выше замещением цистеина (C31G), в нем вирусная репликация будет протекать внутриклеточно, хотя и медленными темпами. Без экзона II большинство иммуносупресорных эффектов белка Tat будут утрачены (Wang, et al.).
[0047] Tat-дифицитныые вирионы могут быть получены любым из множества способов. Как обсуждается в общих чертах в патенте США №7132271 Lau, который включен с помощью ссылки. Методики для получения стабильных Tat-дефицитных мутантов могут включать, но не ограничиваться, те, что включены в ссылки: случайный или сайт-направленный мутагенез (например, Deng, et al.; Busby, et al.), делеция гена-мишени ("генный нокаут") (например. Camper, et al.; Aguzzi, et al.), трансфекция с tat-антисмысловыми полинуклеотидами (например, Lee et al.) и трансфекция с tat-доминантным негативным мутантным геном. Таким образом, Tat-опосредованные иммунологические ответы могут отменяться с помощью делеции и/или мутации нуклеотидной последовательности(последовательностей), кодирующих биоактивный белок Tat без изменения структуры интактного вириона, пока белок Tat не включен в интактный вирион.
[0048] В иммуногенные композиции, в которых полностью отсутствует неперекрывающаяся нуклеотидная последовательность для белка Tat, кодируемая tat экзоном 1 или кодирующая мутированный, усеченный или другим образом лишенный эффективности белок Tat, может быть добавлено определенное количество белка Tat, или он может быть введен экзогенно отдельно с вакциной самой по себе. Это будет способствовать внутриклеточной вирусной репликации на желаемый промежуток времени (например, часы) пока не будет истощен экзогенный белок Tat путем вирусной репликации, диссеминирован во внеклеточную среду или разрушен клеточными ферментами. Необходимо, чтобы экзогенный Tat белок был бы в своей нативной неокисленной форме для поддержания его способности трансактивировать вирус. Снабжаемый белок Tat может принимать одну из нескольких форм, которые могут использоваться независимо, конкурентно или последовательно:
1) полная последовательность из 101 аминокислоты;
2) короткая, но все еще эффективная последовательность из 86 аминокислот;
3) усеченная NH272 аминокислотная последовательность, кодируемая экзоном I;
4) другие усеченные аминокислотные последовательности, кодируемые экзоном I, обладающие способностью к трансактивации, как описано выше, с ядерным доменом белка Tat;
5) мутированные последовательности номер 1, 2, 3 или 4, указанные выше, демонстрирующие способность к репликации; или
6) комбинация вышеупомянутого, не ограниченная в относительной или абсолютной концентрациях или временной рамке введения;
7) мессенджер РНК, кодирующей белок Tat или транскрипционно биологически активный фрагмент.
[0049] Путем включения ограниченного количества экзогенного белка Tat отдельно с условно живым вирионом (например, тем, что испытывает недостаток в способности самому продуцировать белок Tat), добавленный белок Tat действует как субъединичная вакцина, которая контролирует вирусную репликацию. Иммунологический ответ к белку Tat как гуморальный, так и клеточный отмечался у пациентов с ВИЧ, и он обратно коррелировал с прогрессированием заболевания. По аналогии с другими мультивалентными вакцинами, такими как вакцина против дифтерии, коклюша и столбняка, пертуссиновый компонент проявляет функцию адъюванта для компонентов дифтерии и столбняка, вероятно путем усиления местного неспецифического воспаления. Подобным образом, условно живой вирион может действовать как иммунный стимулятор для экзогенного белка Tat в форме субъединичной вакцины или противоположным образом.
[0050] Как только ограниченное количество экзогенного добавленного белка Tat израсходуется, остается инактивированный внутриклеточный и внеклеточный ВИЧ вирион, не способный к репликации. Этот вирион имеет структурные компоненты инфекционного, способного к репликации компонента ВИЧ вириона. Недостающий белок Tat является регуляторный белком, вовлеченным в вирусную репликацию и иммунологическую супрессию; белок Tat не является компонентом вируса ВИЧ. Таким образом, данное изобретение обеспечивает внутриклеточную репликацию, в конце концов, не способного к репликации вируса.
[0051] В целом, этот пример данного изобретения представляет собой иммуногенную композицию, в которой часть или все нуклеотидные последовательности, кодирующие белок Tat, модифицированы (т.е., включая делецию или специфическую мутацию). В зависимости от способа введения, она может включать один или оба экзона, кодирующие Tat. Белок Tat включен в сферу режима вакцинирования для того, чтобы обеспечить протекание репликации ВИЧ. Эта репликация будет короткоживущей и будет терминироваться при истощении по белку Tat. Этот вариант осуществления данного изобретения, описанный выше, представлен только для примера, а не для ограничения.
Выбор исходного материала и штамма(ов) вируса ВИЧ
[0052] Классически вакцина для одного патогена содержит один, два или возможно три отдельных, но связанных вектора. Например, вакцина Салка и Сабина являются трехвалентными. Этот подход не будет применяться для таких заболеваний, как ВИЧ, с их характерными популяционными демографическими показателями (квазивиды) и избытком задокументированных штаммов и циркулирующих рекомбинантных форм вируса. Формулирование вакцины с иммуногенной композицией, в целом, хорошо известно из уровня техники.
[0053] Две ветви данного изобретения могут быть осуществлены отдельно. Далее изложен аспект изобретения для получения не способного к репликации вириона, включающий следующие этапы: (1) обеспечение или отбор вирусной ДНК или РНК молекул, представляющих полный вирусный геном для вирусного штамма(ов) интереса; (2) изолирование вирусных нуклеиновых кислот, если это необходимо; (3) модификация нуклеиновой кислоты; (4) амплификация нуклеиновой кислоты; (5) объединение цельного вириона, не способного к репликации, то есть, переупаковка собранной нуклеиновой кислоты в систему экспресси, пригодную для самосборки вирусных частиц; (6) собирание подвергшихся самосборке условно живых вирионов; и (7) необязательно добавление экзогенного белка(белков) репликации, соответствующих модифицированному гену(генам) или соответственной мРНК.
[0054] Живые векторы ВИЧ могут быть закуплены и являются источниками материала для вакцины от NIH. Тем не менее, эти вирусные изоляты несовершенны по многим характеристикам, что отмечается у активно инфицированных пациентов, потому что они пассируются через множество клеточных линий in vitro. Достаточно типичен тон факт, что непрерывные клеточные линии (т.е., клеточные линии, которые не имеют предела возможного числа митотических делений) используются как культурная среда из-за их универсальной способности и низкой цены, четко определенных питательных потребностей и всеобщей прогнозируемости. Прогнозируемость неперывных клеточных культур определяется тремя параметрами: (1) большое количество митозов; (2) короткая G1 фаза клеточного цикла позволяет произойти клеточному делению за часы или минуты; и (3) непрерывная мутация. Вирус, тем не менее, быстро адаптируется к окружающей среде хозяина. Непрерывные человеческие Т-клеточные линии, такие как SupTI, H9, Jurkat или A3.01, можно также получить по Программе исследования СПИД и референтных реагентов Национального Института Здоровья (NIH AIDS Research and Reference Reagent Program) или из Американской коллекции типовых культур, обе в Роквиле, MD Laboratory. Адаптированные вирусы ВИЧ могут размножаться в этих непрерывных клеточных линиях, но большинство вирусных изолятов человеческого происхождения не размножаются в таких условиях (Michael, et al.).
[0055] Классическая вирусология проводит отличия между вирусом "дикого типа" и мутированным или другим способом измененным вирусным материалом. Действительно, вирус "дикого типа" может не быть, и зачастую не является вирусом, синонимичным изолированному из интактного хозяина. Тем самым необходимо отличать лабораторно полученный "дикий тип", обычно полученный путем пассирования через неперывные клеточные культуры и вирусные изоляты, от интактного природного хозяина. Последние имеют название полевых или клинических изолятов и демонстрируют структурные и генетические свойства, искомые в вакцине. Таким образом, вирус, описанный как полевой или клинический изолят из интактного хозяина, отличается от вируса из клеточных культур.
[0056] В интактном хозяине вирус ВИЧ населяет множество сфер, систем органов и/или гистологических тканей и экскретируется в различных клеточных жидкостях. Настоящий вирус ВИЧ, также как интактные последовательности РНК и ДНК, может быть восстановлен из инфицированных пациентов на всех стадиях спектра заболевания, даже до острого ретровирусного синдрома (т.е., того, что происходит у большинства пациентов через 30 дней после инфицирования). Особенно, вирус адаптируется к окружающему пространству хозяина, с полужизнью в шесть часов, типичный вирус ВИЧ продуцируется и секретируется клетками в той же ткани, которая, в конечном счете, повторно инфицирует. Таким образом, вирусные культуры в различных системах органов одного пациента часто демонстрируют едва заметные, но важные генотипические и фенотипические отличия, что необходимо для вирусной репликации в инфицированной ими ткани.
[0057] Это представляет собой экстраполяцию основного принципа Дарвина, что организм будет адаптироваться к своему окружению или погибать. Иммунологическая среда хозяина-человека разделена на различные отдельные биосферы или компартменты (все из которых становятся ВИЧ-инфицированными), включая, но не ограничиваясь, следующее: лимфоидная ткань, ассоциированная с кишечником (GALT), лимфоидная ткань, ассоциированная с бронхами (BALT), лимфоидная ткань, ассоциированная с кожей (SALT), лимфоидная ткань, ассоциированная с молочной железой (MALT), и лимфоидная ткань, ассоциированная с конъюктивой (CALT). Лимфоциты и другие клеточные компоненты, а также другие молекулярные компоненты иммунной системы неравномерно распределены в соматических тканях (Parrish, et al.). Иммунная нагрузка на вирус ВИЧ, тем самым, отличается в этой специфической ткани или органе происхождения. Генотипическая и фенотипическая экспрессия вируса будет отражать иммунное окружение, в котором он размножается.
[0058] Первичный способ передачи ВИЧ представляет собой половой. То есть, семенная, вагинальная и ректальная жидкости интактных хозяев являются логическими источниками для вирусных полевых или клинических изолятов для получения вакцины. Хорошо известны способы сбора образцов путем цервиковагинального орошения. Также был в общих чертах описан отбор вручную цервикальных секретов. Это альтернативный способ получения либо цельных способных к репликации вирионов, вирусной РНК либо ДНК. Также обычно проводится изолирование вируса из семенной жидкости (Michael, et al., 1999, Ch. 17). Способ культивирования ВИЧ-1 в человеческой сперме является стандартным в промышленности (Michael, et al., 1999, Ch. 8). В конечном итоге, процесс отбора и обработки ректальных секретов был описан в литературе (Michael, et аl., 1999.Ch.35).
[0059] Определение, изолирование и развитие вируса ВИЧ может быть проведено в различных инфицированных тканях, включая, но не ограничиваясь, следующее: человеческие моноциты/макрофаги, Т-клетки и ткань центральной нервной системы (Michael, et al., 1999, Ch. 9 и 10). Культивирование ВИЧ и развитие могут быть дополнены митоген-стимулированными мононуклеарными клетками периферической крови (РВМС) "нормальных" неинфицированных здоровых доноров (Michael, et al., 1999, Ch. 1). Этот способ, несмотря на направленные усилия по отношению ко многим ВИЧ-вакцинам и лекарствам, является рискованным. Вирус будет продолжать мутировать в клеточной культуре и будет быстро принимать генотипические и фенотипические характеристики (случайное распространение генетических мутаций в популяции), что отличает его от начального тканевого изолята. Культуры также могут быть ненадежными, часто требующими прошествия 30 дней до вирусной репликации.
[0060] Исходные материалы для изолирования вирусных нуклеиновых кислот могут быть разделены на две широкие категории: (1) богатые клетками; и (2) обедненные клетками. Существует некоторое перекрывание этих категорий. Обедненный клетками изолят может быть получен из начальной богатой клетками культуры. Богатые клетками начальные материалы включают, но не ограничиваются, следующее: (1) цельная кровь или фракции крови; (2) костный мозг; (3) тканевые препараты, свежие, замороженные, залитые парафином или приготовленные другим образом; (4) in vitro культивируемые клетки; (5) тампоны, пропитанные жидкостями и клетками из тканей; и (6) смыв из бронхов. Обедненные клетками начальные материалы включают, но не ограничиваются, следующее: (1) плазма крови; (2) сыворотка крови; (3) моча; (4) слюна; (5) супернатанты клеточной культуры; и (6) стул (Botho Bowien, et al.).
[0061] Вирусы, включая ВИЧ, могут быть изолированы из многих категорий начальных материалов. Тем не менее, изолирование вирусной ДНК из богатых клетками материалов будет осложнено совместным очищением ДНК хозяина и вируса. Технология, основанная на ПЦР, как обсуждается ниже, может направлять, изолировать и амплифицировать вирусную нуклеиновую кислоту из богатых клетками культур, но это требует большого количества нуклеиновой кислоты как матрицы, и это требование может ингибировать ПЦР (Bowien, et al., Ch. 5). Вирусная ДНК/РНК в богатой клетками среде является как ассоциированной с клетками, так и бесклеточной. Во внутриклеточном компартменте вирусные нуклеиновые кислоты могут быть интегрированы в геном хозяина или связаны с белками хозяина и/или вируса как в цитоплазменном, так и ядерном компартментах. В конце концов, вирусные нуклеиновые кислоты частично или полностью могут быть обнаружены в богатой клетками системе безбелковой внеклеточной среды. Тем самым, в богатой клетками среде источник и содержание вирусной нуклеиновой кислоты ДНК неоднородны.
[0062] Бесклеточные жидкости организма ограничивают, но не полностью, устраняют загрязнения ДНК хозяина. Содержание вирусной ДНК во многих обедненных клетками изолятах характеризуется низким титром, что требует концентрирования нуклеиновых кислот до изолирования и амплификации.
[0063] Эритроциты млекопитающих лишаются ядер сразу после попадания в циркуляцию, и, таким образом, имеют очень мало ДНК. Митохондриальная ДНК обнаруживается в митохондриях, но в интактных клетках, имеющих ядро, митохондриальная ДНК составляет очень малую фракцию общей клеточной ДНК. Кровь человека содержит приблизительно в 1000 раз больше эритроцитов, чем лейкоцитов, которые имеют ядра. Таким образом, если кровь используется как селективная среда для вирусного изолирования и амплификации, необходимо вначале удалить эритроциты.
[0064] Это можно сделать с помощью гипотонического шока, так как красные кровяные клетки разрушаются в гипотонической среде быстрее, чем белые кровяные клетки. Альтернативно, с помощью центрифугирования в градиенте плотности Фрикола можно отделить мононуклеарные клетки (лимфоциты и моноциты) от эритроцитов. Третий способ состоит из центрифугирования цельной крови при 3300 g в течение десяти минут при комнатной температуре. Этот способ разделяет кровь на три точно определенные фракции: (1) фракция, обогащенная белыми кровяными клетками, известная как лейкоцитная пленка; (2) плазма крови; и (3) красные кровяные клетки (Bowien, et al., Ch. 2). Лейкоцитная пленка будет богатым клетками источником, пригодным для выделения вирусной нуклеиновой кислоты, и фракция плазмы крови будет служить в качестве обедненной клетками среды, также пригодной для выделения вирусной нуклеиновой кислоты.
[0065] Отбор вирусных штаммов логически аналогичен тем штаммам, которые являются врожденными в популяции. Как упомянуто выше, отдельный клон вируса не будет репрезентативным для эпидемии ВИЧ. Другие факторы, которые должны приниматься во внимание, включают, но не ограничиваются, иммуногенность и патогенность индивидуальных штаммов ВИЧ. Оптимальная вакцина будет преимущественно включать элементы, которые наиболее близко воспроизводят действительно инфекционную частицу или ее часть. Это должно отражаться в наличии квазивидов, различных генотипически и фенотипически, найденных у интактного хозяина. Вирионы, используемые для производства вакцины, могут происходить из любого тканевого источника, но семенная, вагинальная и/или ректальная ткань являются преимущественными.
Изолирование вирусных нуклеиновых кислот
[0066] Изолирование вирусной нуклеиновой кислоты РНК или ДНК из инфицированной ткани может быть достигнуто с помощью целого ряда хорошо известных технологий. Начальные этапы, если необходимо изолировать вирусную ДНК, состоят из ферментативной или медикаментозной деградации материала клеточной стенки, если она присутствует, и детергентный лизис клеточной мембраны. После клеточного разрушения белки как хозяина, так и вируса отделяют от нуклеиновой кислоты.
[0067] Свежесобранные ткани и клетки являются идеальными для изолирования нуклеиновых кислот. Сберегание тканей и клеток подвергает риску цельность нуклеиновой кислоты. Если необходимо длительное сберегание рекомендуется использовать бумажный фильтр или замораживание ДНК при -20°С в ТЕ буфере при рН 8. Среда сберегания ДНК должна быть лишена воды и загрязнений. Длинный период сберегания биологический жидкостей, таких как семенная жидкость, хотя не является преимущественным, может быть достигнут при -20 - -80°С (Bowien, et al., Ch. 2).
[0068] Описаны две очень простые технологии для изолирования ДНК из клеток:
(1) инкубация клеточных лизатов при высоких температурах (например, 90°С в течение 20 минут) и (2) расщепление протеиназой К. Обе технологии ограничены по применению и часто подвергается риску многочисленных загрязнений (Bowien, et al., Ch. 2).
[0069] Биологические ткани могут быть созданы из однородной композиции до выделения нуклеиновых кислот с использованием ротор-статорных гомогенизаторов. Альтернативно, мельница для смешивания может дробить и гомогенизировать клетки и ткани до выделения нуклеиновых кислот (Bowien, et al, Ch. 2).
[0070] Молекулярная структура, электростатический характер и коэффициент диффузии РНК и ДНК достаточно сходны. Таким образом, многие способы изолирования ДНК будут подвергаться риску загрязнения примесями РНК. Обработка РНКазой А будет удалять РНК. Раствор РНКазы А должен быть подвергнут тепловой обработке перед использованием для удаления любых загрязняющих соединений с ДНКазной активностью. РНКаза, свободная от ДНКазы, также коммерчески доступна. РНКаза Н может быть включена в процедуры изолирования ДНК на различных этапах, включая исходную среду и/или конечный продукт (Bowien, et al., Ch. 2).
[0071] Способы органической экстракции, состоящие из фенола или смесей фенол/хлороформ, определены в литературе (Bowien, et al. Ch. 2). Способ Блоттинг по Саузерну является еще более усовершенствованным способом, используемым для определение нуклеиновых кислот ВИЧ, и состоит из экстракционной смеси фенола/хлороформа/изоамилового спирта (соотношение 25:24:1) и также описан в литературе. Рибонуклеаза (РНКаза) может быть добавлена для расщепления РНК при приготовлении изолята вирусной ДНК. Дальнейшее изолирование интактной вирусной ДНК из вирусных ДНК фрагментов и ДНК хозяина может быть завершена ценрифугированием в градиенте (Michael, et al., Ch. 9 и 10).
[0072] Способы "высаливания" вирусных нуклеиновых кислот являются другой альтернативой. Клеточный лизат помещают в гипертоническую среду, что облегчает осаждение белков и других загрязняющих соединений. Центрифугирование удаляет преципитаты, а вирусную ДНК восстанавливают с помощью второго этапа спиртовой преципитации. Очищение ДНК и количественный выход являются временами непредсказуемыми в этом способе (Bowien, et al., Ch. 2).
[0073] Центрифугирование через градиент плотности цезия хлорида/этидия бромида может отделять вирусную ДНК, обнаруженную в клеточном лизате, полученном спиртовым осаждением. Центрифугирование требует нескольких часов, и нить ДНК эктрагируют изопропанолом для удаления этидия бромида. ДНК затем осаждают спиртом. Этот способ дает на выходе высокое качество ДНК, но он неавтоматизирован, поэтому длительный по времени, относительно дорогостоящий и может быть неприемлемым для промышленного применения из-за вариабильности человека (Bowien, et al., Ch. 2). Будучи удаленной, однако, и найденной идеальной для формуляции вакцины, модификация нуклеиновой кислоты может быть проведена для удаления последовательностей-мишеней.
[0074] Другой способ изолирования представляет собой способ селективной абсорбции нуклеиновых кислот на силикагеле в присутствии высоких концентраций диссоциирующих солей. Они включают, но не ограничиваются, следующее: гуанидин гидрохлорид, гуанидин изотиоцианат, натрия йодид и натрия перхлорат. Эта методика эффективно отделяет ДНК от РНК, но другие клеточные загрязняющие соединения нуждаются в отмывке до того, как ДНК высокой степени очистки и качества может быть элюирована из частиц силикагеля с низкосолевым буфером. Основанные на силикагеле методики предлагаются некоторыми компаниями в виде наборов (Bowien, et al., Ch. 2).
[0075] Способы анионного обмена, основанные на электростатическом взаимодействии между негативно заряженными фосфатами нуклеиновой кислоты и позитивно заряженными поверхностными молекулами на субстрате, используются для изолирования вирусной ДНК. Использование твердофазной анионообменной хроматографии вирусной ДНК будет связываться с субстратом при низкосолевых условиях. Загрязнения, такие как РНК и белки, отделяют с использованием среды - солевых буферов. ДНК затем элюируют с высокосолевым буфером, и ДНК обладает высоким качеством, относительно свободная от загрязнений. Элюированную ДНК затем восстанавливают с помощью спиртового осаждения, и она пригодна для геномной модификации и амплификации (Bowien, et al., Ch. 2).
[0076] Фильтровальную бумагу, пропитанную соединениями с известной функцией стабилизации и изоляции ДНК, можно применять для хранения ДНК перед модификацией и амплификацией. На фильтровальной бумаге находятся соединения, которые лизируют клетки, обладают бактерицидной способностью, ингибируют ДНК деградацию, такую как окисление, и связывают нуклеиновые кислоты. ДНК остается связанной с фильтровальной бумагой до элюирования. Этот метод позволяет хранить ДНК при комнатной температуре несколько лет без ее значительного повреждения или ухудшения (Bowien, et al., Ch. 2).
[0077] Как упомянуто, кровь может быть источником геномной нуклеиновой кислоты. Общепринятые антикоагулянты, такие как гепарин и EDTA, могут мешать методикам выделения ДНК, и поэтому их нужно избегать, если кровь не нужно хранить. QIAGEN® изготовляет наборы ДНК крови QIAamp® для выделения ДНК из цельной крови. Для этой методики не нужно центрифугировать и отделять фракции цельной крови. В альтернативном способе коммерчески доступными являются наборы тканей DNeasy® и основанная на силикагеле мембранная технология. Также QIAGEN® производит наборы для анионообменной технологии для выделения ДНК из крови и клеточной культуры ДНК. Наконец, с помощью вирусного набора QIAamp® UltraSens® от QIAGEN® можно выделить ДНК ВИЧ из плазмы крови и сыворотки (Botho Bowien, et al., Ch. 5).
[0078] Вирусная РНК может быть предпочтительной ДНК в определенных вариантах осуществления. ДНК предпочтительна, если применяется из-за врожденной нестабильности РНК. Если используется кровь, то до выделения РНК красные кровяные клетки хозяина и тромбоциты необходимо удалить из вирусного источника. Красные кровяные клетки, как упомянуто выше, содержат немного нуклеиновой кислоты и являются бедными источниками для выделения вирусной нуклеиновой кислоты. Удаление эритроцитов упрощает выделение РНК, поскольку соотношение rbcs к wbcs составляет 1000 к 1. Применяют те же способы для выполнения этой методики, обсуждаемые с выделением ДНК выше, но они включают, но не ограничиваются: (1) гипотонический шок с последующим центрифугированием и (2) центрифугирование в градиенте плотности в фиколле.
[0079] В общем, обедненный клетками материал, поэтому, является предпочтительным богатому клетками материалу, если целью является выделение вирусной РНК. Это ограничило бы лабораторную методику, если намеченная вирусная РНК будет внеклеточной. Как обсуждалось выше для ДНК, внеклеточная нуклеиновая кислота может быть неспособной к неинфекционной репликации. Идеальный источник вирусной РНК отражает те из вирусной ДНК, жидкости тела, передающие вирус при половом контакте, главном способе передачи ВИЧ сегодня. РНК клеточного происхождения из такой обедненной клетками жидкости организма была бы более типичной для компетентных к репликации инфекционных вирионов и была бы предпочтительной, по мнению автора. Вирусная РНК, происходящая из обедненной клетками среды, может быть связанной с клеткой, свободной от клетки или комбинацией этих двух вариантов. Полученная концентрация вирусной РНК, не взирая на источник, ожидается низкой, что делает необходимым ультрацентрифугирование, ультрафильтрацию или осаждение (Bowien, et al., Ch. 6).
[0080] Клеточная РНК из не-ВИЧ инфицированных тканей состоит из трех отдельных пулов: (1) рибосомальная РНК; (2) трансферная РНК и (3) мРНК, МРНК несет генетическую информацию, полученную в ДНК. Фракция мРНК наименьшая из этих трех, но является необходимым компонентом для разработки, основанной на РНК иммуногенной композиции или вакцины. Из общей РНК в типичной клетке млекопитающих только 1-5% представляет собой мРНК (Bowien, et al., Ch. 6). Экспрессия РНК в клетке достаточно изменчива. В ВИЧ инфицированных клетках можно выделить четвертый пул РНК клеточного происхождения, состоящий из гетерогенной смеси вирусной РНК. Вирусная РНК в таких клеточных линиях является либо однонитевой, диплоидной (соединенной вместе только в определенных последовательностях с 5′ конца), или связанной с комплементарной ей ДНК в дуплексе РНК/ДНК. Встречаются также молекулы двухнитевой РНК, принимающие спиральную структуру, более или менее похожую на двойную спираль Уотсона и Крика. Молекулы однонитевой РНК включают нерасщепленные, отдельно расщепленные или многократно расщепленные последовательности нуклеиновой кислоты. Нерасщепленная РНК может иметь или может не иметь 5′ кэп и 3′ полиаденилированный (поли-А последовательности) хвост клеточного происхождения. В частности в клетке, инфицированной вирусом ВИЧ, содержание мРНК варьирует во времени и зависит от экспрессии белка Rev. После экстракции вирусной РНК обогащение фракции мРНК можно выполнить добавлением олиго(dT)-целлюлозы. Это можно использовать для связывания и отделения поли(А) хвостов эукариотических мРНК. Это облегчает отделение мРНК от ДНК, рРНК и тРНК.
[0081] Процесс сбора образца и обработки может влиять на получение мРНК в течение секунд. Идеально мРНК, изолированная для получения вакцины, должна отражать мРНК, полученную in vivo. Однако клеточная смерть и ферментативная деградация РНК ферментами РНКазы клеточного и вирусного происхождения может быстро разрушить фракцию мРНК. Аналогично, процессинг образцов и обработка могут индуцировать или подавлять экспрессию определенных вирусных генов. Поэтому мРНК необходимо стабилизировать перед какими-либо методиками изолирования нуклеиновых кислот. Быстрое замораживание в жидком азоте или в этаноле и сухом льду применяли для стабилизации мРНК с ненадежными результатами.
[0082] Раньше предпочитали инактивацию РНКаз клеточного или вирусного происхождения в лабораторном процессе выделения РНК. Ферменты РНКазы являются повсеместными в клетке, обычно не нуждаются в кофакторах функций, относительно стабильны, высоко эффективны и часто инактивируются с трудом. Лизис клетки для получения вирусной нуклеиновой кислоты впоследствии высвобождает внутриклеточные РНКазы. Хаотропные агенты, включая гуанидина изотиоцианат и гуанидина гидрохлорид, непосредственно инактивируют РНКазы. Также расщепление контаминирующей ДНК можно выполнять с помощью ДНКазы I (Bowien, et al., Ch. 6). Обработку ДНКазой I можно выполнять в начале, середине или конце какого-либо лабораторного протокола, включающего изолирование РНК, но обычно должна следовать за обработкой РНКазами, инактивирующими соединение.
[0083] Смесь меркаптоэтанола, саркозила и гуанидина тиоцианата применяли для инактивации РНКаз и очистки вирусной РНК от тканевых проб при рН 7,0. Затем добавляли ацетат натрия при рН 4,0 и кислый фенол, позволяя РНК осадиться со спиртом.
[0084] Соединения, консервирующие РНК, такие как стабилизирующий РНК реагент RNAIater®, коммерчески доступны. Он позволяет хранить образец ткани до изолирования мРНК продолжительные периоды времени. Другим примером является система РНК крови PAXgene® для стабилизации и очистки РНК. Этот продукт препятствует генной транскрипции (Bowien, et al., Ch. 2).
[0085] Если используют богатую клетками среду для изолирования РНК, то клеточный лизис протеиназой К в носителе, содержащем ингибитор РНКазы натрия додецил сульфат (SDS), является относительно легкой методикой. ДНК можно удалить ДНКазой I. Органическая экстракция, сопровождаемая спиртовым осаждением, или четко определенные способы на основе диоксида кремния или анионного обмена будут удалять какую-либо лишнюю контаминирующую ДНКазу. Отделение вирусной РНК для геномной РНК можно выполнять центрифугированием или гелевым электрофорезом (Bowien, et al., Ch. 6).
[0086] Альтернативно, вышеупомянутые хаотропные агенты не только инактивируют РНКазы, но также разрушают клетки. Органическая экстракция продолжает хаотропную экстракцию и включает одну или более из следующих определенных методик: (1) спиртовое осаждение; (2) LiCl осаждение; (3) CsCl градиенты плотности; (4) способы на основе диоксида кремния; (5) способы анионного обмена и (6) гибридная селекция (Bowien, et al., Ch. 6).
[0087] РНК ВИЧ, конъюгированная с ВИЧ нуклеокапсидным белком, стабильна на протяжении приблизительно 2-3 часов. Определение количества и амплификация РНК ВИЧ является технически затрудненным, но может выполняться с коммерчески доступными испытаниями, такими как испытание разветвленной ДНК от Chiron®, RT-ПЦР (полимеразная цепная реакция с обратной транскрипцией) испытание Amplicor® от Roche® и NASBA (амплификация, основанная на последовательности нуклеиновых кислот) система амплификации с помощью Organon-Teknika®. NASBA может селективно амплифицировать РНК в композициях, контаминированных ДНК. NASBA может обойти, по меньшей мере, один этап очистки, отделяющий вирусную РНК от ДНК. Меньшее число выполненных этапов приводит к упрощению лабораторной методики и более высокому проценту точности геномной амплификации (Nelson Michael, et al., 1999, Ch. 16).
[0088] РНК нуклеиновые кислоты ВИЧ можно выявлять и изолировать из многообразия тканей и in vitro клеточных линий с помощью процесса нозерн-блоттинг. Для этой технологии можно применять либо ДНК, либо РНК. Но более успешным отмечают ДНК (Michael, et al., 1999, Ch. 10).
[0089] Коммерчески доступные реагенты, такие как Trizol®, приемлемы для экстракции РНК из образцов ткани. Технологию на основе диоксида кремния, такую как наборы QIAamp®, можно применять в клеточных лизатах или бесклеточных образцах для отделения и очистки РНК (Bowien, et al., Ch. 5).
[0090] Вирусную РНК можно также изолировать и концентрировать из образцов стула через микроконцентратор, такой как мининабор вирусной РНК QIAamp® (Bowien, et al., Ch. 5).
[0091] Определены другие установленные способы для выделения и стабилизация РНК ВИЧ. Они включают, но не ограничиваются, катионные детергенты, такие как Catrimox®, применяемый на образцах цельной крови, и мининабор RNeasy®, который можно применять для изолирования вирусной РНК из крови после хранения при комнатной температуре в течение нескольких месяцев.
[0092] Вирусная РНК типично сгибается и принимает собственные вторичные структуры. У ВИЧ вирусные РНК дуплексы выполняются с помощью молекулярной связи в консервативной области на 5' конце (Flint, et al., 2004, Ch. 7). Обратная транскрипция через эти вторичные, а в случае ВИЧ, третичные и четвертичные структуры может быть затруднена. Для этой цели приемлемы коммерчески доступные ферменты обратной транскрипции, такие как Omniscript® и Sensiscript® (Bowien, et al., Ch. 6).
[0093] Было бы разумно предположить, что генотипические и фенотипические характеристики вируса будут определены частично типом клетки хозяина, в которого он вторгается и в котором реплицируется. Первичные резервенты репликации ВИЧ включают макрофаги и Т-клетки, прежде всего найденные в лимфоидной ткани. In situ гибридизация (ISH) позволяет идентификацию, оценку концентрации и внутриклеточной локализации специфических нуклеиновых кислот, включая ДНК и мРНК, а также внутриклеточных белков. ДНК, мРНК и белок можно определить одновременно в отдельной клетке, что позволяет исследователю увязать содержание генома и геномную экспрессию на внутриклеточном уровне.
[0094] ISH является относительно интенсивной по сравнению с процессом in situ ПЦР, описанным ниже. Специалист в данной области с помощью ISH может определить концентрации мРНК до 20 копий на клетку. Большинство лабораторий, выполняющих ISH, более ограничены по порогу идентификации мРНК, определяя не менее чем 100 копий на клетку. Хотя процесс in situ гибридизации определили, не хватает объединенного подхода ко всем типам клеток с ВИЧ. Не смотря на это, известны традиционные методики.
[0095] In situ полимеразная цепная реакция позволяет идентифицировать и амплифицировать внутриклеточную ДНК и РНК. Она может оказаться предпочтительной в получении вакцины, поскольку устраняются многие этапы в изолировании нуклеиновых кислот (упрощение лабораторных методик, облегчение очистки нуклеиновых кислот и увеличение количества извлекаемой нуклеиновой кислоты), а идентифицированная нуклеиновокислотная последовательность будет параллельна последовательности инфицирующих, репликационно компетентных вирионов. Большинство полученных вирионов ВИЧ являются неинфекционными и репликационно некомпетентными. В какой-либо системе, in vitro или in vivo, неизбежно произойдет контаминация инфекционных репликационно компетентных вирионов неинфекционными некомпетентными вирионами. Иммунологический ответ, направленный на неинфекционные репликационно некомпетентные вирионы, не принесет пользы или, в худшем случае, может неблагоприятно влиять на хозяина.
[0096] Понятие "первичный антигенный стимул" четко определено для гриппа А, сегментированного РНК "минус-цепь" вируса в семействе ортомиксовирусов. Первичный ответ хозяина на патогенный организм блокирует дополнительный иммунологический ответ на этот организм, пока антигенно полностью отличный штамм не инфицирует хозяина (Parham, 2005, Ch. 8). Понятие "первичный антигенный стимул" можно очень хорошо применять для других патогенов, включая ВИЧ. Оптимально, начальная вакцина (субъединица, живая, условно репликационно компетентная, рекомбинантная или иная) должна быть близко параллельной актуальным инфицирующим организмам, а не дефектным вирионам, которые могут служить иммунологической приманкой, мешающей соответствующему иммунному ответу хозяина, и блокировать последующий иммунный ответ на подобные патогены.
[0097] Использование одного или более из вышеупомянутых источников для обеспечения изолирования нуклеиновой кислоты ВИЧ - модификации и амплификации будут следующими для разработки композиции или вакцины. Один вариант осуществления данного изобретения далее охватит отделение инфекционных репликационно компетентных вирионов от неинфекционных репликационно некомпетентных вирионов перед изолированием нуклеиновых кислот. Это можно будет выполнить путем изолирования инфицированных клеток бедной клетками среды в качестве источника(ов) нуклеиновой кислоты ВИЧ. Такое изолирование можно выполнить путем центрифугирования жидкостей организма. In situ гибридизация и in situ ПЦР затем позволят идентификацию и амплификацию предпочтительных нуклеиновых кислот. При обратной транскрипции также можно выполнить in situ ПЦР. Фрагменты мРНК менее 1,5 тысячи оснований можно подвергнуть RT-ПЦР in situ с технологией, доступной на сегодня. Это не позволило бы RT (обратную транскрипцию) полного генома ВИЧ, который имеет длину 10 тысяч оснований. Однако перекрывание или последовательные фрагменты мРНК после in situ RT-ПЦР можно лигировать ДНК лигазами для получения интактного ДНК генома ВИЧ, приемлемого для модификации и амплификации (Michael, et al., 1999, Ch. 18).
[0098] Коммерчески доступны ферменты обратной транскриптазы, такие как Superscript II®, который недостаточен по активности РНКазы Н (деградация однонитевой РНК в подвергшемся обратной транскрипции РНК/ДНК гетеродуплексе), и поэтому более эффективен при амплификации ДНК. Он способен к обратному считыванию относительно длинных молекул мРНК и может применяться для рутинной RT амплификации.
[0099] Отжигающие температуры для обратной транскрипции и ДНК амплификации математически определили для in situ гибридизации и in situ ПЦР. Вторично отжигающие температурные параметры также можно определить с термоциклером, разработанным с блоком температурного градиента для быстрого эмпирического определения отжигающих температур или "Touchdown PCR" (Michael, et al., 1999, Ch. 18).
[0100] Количество нуклеиновых кислот, идентифицированных и амплифицированных при in situ ПЦР и ISH, характерно намного меньше, чем способов ПЦР на основе раствора, обсуждаемых ниже. Исследователи в данной области заключили, что норма, лимитирующая фактор при ISH и in situ ПЦР, представляет собой сложность прохождения праймерами через клеточные мембраны. Это можно преодолеть несколькими способами, включая примеры: (1) подвержение клеток тепловому шоку, что временно повышает проницаемость мембраны для макромолекул; (2) сцепление праймеров с пептидами, проникающими через клеточную мембрану (СРР) и/или (3) комбинация (1) и (2).
Способы определения количества ДНК и РНК и оценивание чистоты
[0101] Лабораторные способы для оценивания концентрации РНК и ДНК и чистоты стандартизированы и достаточно похожи. Концентрацию РНК можно определить, измеряя поглощение при 260 нм (А260) с помощью кварцевых кювет, которые позволяют ультрафиолетовому излучению проходить с минимальной дисторсией и абсорбцией в спектрофотометр. Значение рН 7,0 во всей методике может гарантировать достоверность и воспроизводимость. Соотношение значений абсорбции при 260 и 280 нм обеспечивают оценку чистоты РНК. Наборы являются коммерчески доступными, такими как набор мРНК Oligotex® для определения количества мРНК (Bowien, et al., 2003, Ch. 6; Nicholl, Ch. 3).
[0102] Количественное определение концентрации ДНК и чистоты также можно выполнить с помощью спектрофотометра, выполняющего измерение абсорбции при 260 нм в кварцевой кювете. Анализ в агарозном геле также можно применять для определения количества ДНК. Как и с РНК, чистоту ДНК можно определить, измеряя соотношение А260/А280 (Bowien, et al., Ch. 7; Nicholl, 2002, Ch. 3).
[0103] Полимеразную цепную реакцию можно применять не только для амплификации ДНК и/или РНК последовательностей, а также для удаления праймеров, ферментов, солей, буферов, нуклеотидов и других контаминант. Наборы, такие как набор ПЦР для очистки 96 QIAquick®, позволяют ПЦР очистку ДНК и представляют собой технологию на основе диоксида кремния - гель - мембраны. Альтернативный способ очищения ДНК представляет собой методику реакционного очищающего набора MinElute®. Эта методика также основана на диоксиде кремния - гелевой - мембранной технологии (Bowien, et ai., Ch. 7).
[0104] Очистку РНК можно выполнить с помощью свободной от РНКазы ДНКазой I, и она является коммерчески доступной, например наборы RNeasy® и мининабор РНК крови QIAamp® для очистки РНК, которые основаны на диоксид кремния - мембране, спин-колоночной технологии, избегающей необходимость обработки ДНКазой (Bowien, et al., Ch. 6).
[0105] Актуальную последовательность молекулы ДНК можно выполнить двумя четко определенными в литературе способами. Секвенирующий способ Максама-Гилберта основан на серии "футлярных" фрагментов и включает радиомечение ДНК с помощью 32P на 5′ конце каждой нити. Секвенирующий способ Сангера-Коулсона (дидеокси или ферментативный) использует фрагмент Кленова ДНК полимеразы и праймер для обеспечения 3′ конца для ДНК полимеразы (Nicholl, Ch. 3).
[0106] Любой из вышеупомянутых способов можно применять для секвенирования интактной вирусной нуклеиновой кислоты, модифицированной нуклеиновой кислоты, применяемой в зависимом от репликации вирионе, а также нуклеиновокислотной последовательности расщепленного фрагмента. Кроме того, можно определить нуклеиновокислотную последовательность, кодирующую компонент субъединицы вакцинного вектора.
Модификация нуклеиновых кислот
[0107] Как только выбранная вирусная нуклеиновая кислота изолирована, можно начинать модификацию последовательности. Как отмечено выше, модификация, как используется здесь, может включать делецию, а также мутацию. Можно использовать либо РНК, либо ДНК, но ДНК была бы предпочтительной. ДНК более стабильна, легче амплифицируется ex vivo, а мутацию ДНК можно выполнить более эффективно. Можно также применять матрицу РНК вирусного генома. Фермент обратной транскриптазы, РНК-зависимая ДНК полимераза, дает комплементарную нить ДНК от РНК. Альтернативно РНК можно модифицировать делецией перекрывающих и/или неперекрывающих сегментов. ПЦР самостоятельно может ввести точечные мутации, делеции или вставки в ДНК (Flint, et al., 2004, Ch. 2).
[0108] Получение условно живых вирионов можно совершить применением ферментов рестрикции бактериального или иного происхождения для выщепления желательных последовательностей из интактного вирусного генома. Для вырезания генетической последовательности намеченных белков можно использовать комплемент ферментов рестрикции. В этом процессе можно будет идентифицировать генетическую последовательность, окружающую кодон намеченного белка.
[0109] Ферменты рестрикции продуцируются бактерией как защита против вирусной инфекции. Идентифицированы и коммерчески доступны более чем 200 ферментов рестрикции; около 100 из этих ферменты обычно используются исследователями. Каждый фермент рестрикции связывается с ДНК и распознает специфическую нуклеотидную последовательность, названную последовательностью распознавания. Фермент разрезает обе нити ДНК в последовательности распознавания в специфической картине расщепления. Это сопровождается этапом очистки модифицированной нуклеиновокислотной последовательности.
[0110] Фрагменты, образованные применяемыми ферментами рестрикции, могут иметь "тупые" концы, 3′ торчащие концы или 5′ торчащие концы. Модификацию к отрезанной ДНК можно выполнить перед повторным отжигом концов. Например, фермент терминальная деоксинуклеотидил трансфераза (TdT) неоднократно и беспорядочно добавляет нуклеотиды к каким-либо доступным 3′ концам нематричным способом (Nicholl, 2002, Ch. 4). Он включает торчащий, тупоконечный и углубленный 3′ конец. Как только два концы последовательности ДНК опять соединяются вместе (например, добавлением лигазы), может быть создан нокаутный ВИЧ вирион, который служит основой данного изобретения. Другие модифицирующие ДНК ферменты, такие как экзонуклеаза, которые разрушают 5′ и/или 3′ конец ДНК, также можно применять с этой целью.
[0111] В литературе четко определены четыре дополнительные применяемые нуклеазы (ВаI 31, экзонуклеаза III, деоксирибонуклеаза I [ДНКаза I] и S1-нуклеаза), каждая из которых отличается по локализации и способу активности и обеспечивает молекулярному биологу точные разрезающие инструменты для работы. Фосфатные группы можно добавить или удалить с конца молекулы ДНК. Фермент щелочная фосфатаза отщепляет концевую фосфатную молекулу ДНК, а фермент полинуклеотидкиназа добавляет фосфатные группы на конец ДНК.
[0112] Другой фермент концевая трансфераза (концевая деоксинуклеотидил трансфераза) неоднократно добавляет нуклеотиды на любой открытый 3′ конец ДНК, включая торчащий, тупоконечный и углубленный 3′ конец. После удаления намеченной нуклеиновокислотной последовательности(ей) из вирусного генома оставшиеся фрагменты ДНК можно соединить в функциональную молекулу ферментом ДНК лигазой. Такой вирусный геном с делецией последовательностей, необходимых для получения добавочного белка (и/или структурального, и/или ферментативного), можно амплифицировать с помощью ПЦР или другим способом амплификации последовательности.
[0113] Для гарантирования и усиления чистоты модифицированной нуклеиновой кислоты могут следовать несколько этапов. Они включают, но не ограничиваются, центрифугирование, гелевый электрофорез, секвенирование нуклеиновой кислоты и обратную транскрипцию геномной последовательности с идентификацией всех белков, транскрибированных и транслированных. Последний процесс можно выполнить в бесклеточном питательном бульоне или в in vitro клеточной культуре, такой как полиморфонуклеарные клетки крови или непрерывная линия клеток, такая как HeLa клетки.
[0114] Удаление неперекрывающегося сегмента(ов) намеченного белка(ов) приведет к транскрипции обрезанного, нефункционального белка(ов). Только эти намеченные белки будут неблагоприятно затронуты, потому что каждый удаленный сегмент кодирует только часть одного белка. Тем не менее, это нарушит и инактивирует вирионы ВИЧ. Полученные вирионы будут репликационно некомпетентными и неинфекционными. Как обсуждалось в другом месте, репликация будет нуждаться в экзогенном источнике дефицитного белка(ов). Удаление перекрывающихся сегментов будет неблагоприятно влиять на все белки, частично кодируемыме перекрывающимися сегментами.
[0115] Применяя обычно доступные методики молекулярной биологии, ДНК можно разрезать по точно определенным намеченным зонам, таким как те последовательности, что кодируют для неперекрывающихся частей vif, vpr, vpu, tat экзон 1 и vpx кодоны. Одна или более мутация, неблагоприятно влияющая на один или более структуральный, ферментативный или добавочный белок, делает вирион некомпетентным (Flint, et al., 2004, Ch. 20). Эти мутации могут представлять собой одно или более замещение основания, делецию основания (смежную или несмежную) или делецию нуклеотидной последовательности(ей). В контексте условно живого вириона будут применять удаление неперекрывающихся и/или перекрывающихся генных сегментов намеченного белка(ов). Точечные мутации в самом и самого не обеспечивают достаточных параметров безопасности из-за склонности возникновения обратной мутации, позволяющей вирусу стать репликационно компетентным.
Амплификация нуклеиновой кислоты
[0116] ПЦР технология на основе раствора, первоочередный способ нуклеиновокислотной амплификации, обсуждаемый ниже, не дифференцирует источник вирусных нуклеиновых кислот. Кроме того, эта технология не зависит от источника нуклеиновых кислот. В большинстве применений ПЦР не является механизмом идентификации нуклеиновых кислот, очисткой или изолированием. Исключением является in situ ПЦР (обсуждаемая выше), которая не позволяет внутриклеточную идентификацию нуклеиновых кислот, очистку и изолирование.
[0117] Полимеразная цепная реакция (ПЦР) дает возможность исследователю селективно амплифицировать ДНК последовательности какого-либо организма миллион раз или более. Эта методика полагается на выбор праймеров из двух консервативных областей вирусного генома. Большинство процессов используют праймер тРНК, связывающий сайт, расположенный в непосредственном соседстве к 5′ LTR (длинным концевым повтором), и gag нуклеотидную последовательность и мРНК сигнальный сайт полиаденилирования на R/U5 месте соединения 3' LTR. Так амплифицируют 9 тысяч оснований вирусной ДНК, обобщая все кодирующие области для структурных, ферментативных и добавочных белков и U3 и R доменов 3′′ RTR. Инфекционные про-вирионы не могут быть реализованы без интактных 5′ и 3′ LTR. Отдельная амплификация LTR областей, не включенная в GWh реакцию, может быть преобразована отдельной амплификацией и лигирование ДНК может быть использовано для получения интактной геномной последовательности с обоими LTR (Michael, et al., 1999, Ch. 12; Nicholl, 2002, Ch. 7; Specter, et al.).
[0118] Имеются другие способы амплификации нуклеиновой кислоты, кроме ПЦР, и включают, но не ограничиваясь, следующие: амплификация, основанная на последовательности нуклеиновой кислоты (NASBA) и опосредованная транскрипцией амплификация (ТМА). Кроме того, стандартная амплификация смещением (SDA), цепная реакция лигазы (LCR), технология циклирующего зонда (СРТ), и клевазный тест (Cleavase) используются для амплификации нуклеиновой кислоты. Определение вирусных нуклеиновых кислот можно также выполнить этими способами. Также существуют другие лабораторные методики, направленные только на отдельную амплификацию без увеличения числа последовательностей нуклеиновой кислоты, и они включают, но не ограничиваются, следующие: технологии иммуноферментного анализа (EIA), разветвленная цепь ДНК (bDNA) и испытание гибридизационной защиты (HPA)1 и методики передачи энергии посредством флуоресцентного резонанса (FRET) (Specter, et al.). Идентификация нуклеиновой кислоты с помощью методики отдельной амплификации может предшествовать амплификации нуклеиновой кислоты. Это может модернизировать лабораторные методики.
[0119] Все способы амплификации, без учета методики, преимущественно проводятся с клеточным ферментом урацил-Н-гликозилазой (UNG). Это уменьшает ПЦР остаточную контаминацию. Предобработка ПЦР реакционной смеси UNG в течение 10 минут при комнатной температуре расщепляет и вырезает остатки урацила из ДНК молекулы. Затем тепловой инактивацией удаляют любую остаточную UNG. Этот процесс обеспечивает геномную чистоту (Michael, et al., 1999, Ch. 15).
[0120] Классически процесс ПЦР использует Taq полимеразу, которую получают из термофильной бактерии Thermus aquaticus, которая населяет теплые источники (Nicholl, 2002, Ch. 7). Другие подобно функционирующие полимеразы в данный момент стали доступными, что увеличило скорость и точность геномной репликации (Michael, et al., 1999, Ch. 15). Другие ДНК полимеразные ферменты включают Pwo, Tth и HotTub ДНК полимеразу, которые применялись и могут быть использованы, когда присутствуют загрязнения.
[0121] Технологии GWH в реальном времени, такие как зонды и системы определения последовательности, могут обеспечивать тот факт, что изолирование ПЦР и методики амплификации будут проходить с минимальным риском лабораторной контаминации (Bowien, et al., Ch. 8).
[0122] Амплификация нуклеиновой кислоты может быть проведена до или после модификации нуклеиновой кислоты и очистки модифицированной последовательности нуклеиновой кислоты.
Сборка условно живых вирионов
[0123] Вирусная ДНК или РНК после модификации (например, удаления обозначенных последовательностей, других мутаций и т.п.) должна быть переупакована. Это можно выполнить в бесклеточной системе экспрессии/среде или клеточной культуре. В случае интактных вирионов in vitro и in vivo процесс обратной транскрипции начинается до слияния вируса с клеточной мембраной. В случае суб-вирусной частицы вирусная РНК подвергается обратной транскрипции с помощью фермента обратной транскрипции, который включен в капсидную ядерную область. Поэтому ДНК, РНК или обе нуклеиновые кислоты могут быть начальным этапом для сборки вирусных частиц (Flint, et al., 2004, Ch. 4, и Арр. А).
[0124] Вирусная структура диктует правильно организованную предсказуемую последовательность самосборки. Нуклеокапсидный белок является основанием для капсидного белка. Аналогично, капсидный белок является основой для матричного белка. Матричный белок является основой для gp120/gp41 тримеров. В сайте вирусной сборки приблизительно 10% gag полипротеинов (gag-pol) несут трансляционные продукты ретровирусных ферментов, протеазы (PR), обратной транскриптазы (RT) и тинтегразы (IN) на 3′ или СООН концах gag-pol белка.
[0125] Изначально две нити РНК связаны специфическими последовательностями (начальный сайт димеризации и димерная связующая структура) на 5′ LTR вируса. Этот процесс происходит in vitro и инициирует вирусную сборку. Связывание двух РНК нитей облегчает конформационные изменения в РНК, что позволяет происходить следующему этапу. NC белок (р7) затем покрывает вирусную РНК. Добавочные белки Vif и Nef связываются с нуклеокапсидным комплексом вирусной РНК и встраиваются в интактный вирион. Приблизительно 2000 молекул р7 обнаруживается в интактным вирионом на поверхности вирусного генома. Связывание NC сегментов (р7) с диплоидным вирусным РНК геномом запускает его связывание с р6 белком, который прикреплен к его Vpr (ВИЧ-1 и ВИЧ-2) и Vpx (ВИЧ-2) добавочным белкам. Приблизительно 2,000 р6 белков ассимилированы в каждый вирион.
[0126] Вирусные кодируемые ферменты, RT, IN, РНКаза Н и протеаза, затем нековалентно связываются с диплоидным вирусным РНК/р7/р6Λ/рr/Vрх(ВИЧ-2) комплексом. Приблизительно 10 копий каждого фермента включено в каждый вирион. Вирусный капсидный белок связывается с повсеместно встречающимся клеточным белком циклофилином А (СурА), который демонстрирует цис-транс пептидил - пролил изомеразную активность. Связывание СурА с р24 происходит при капсидной последовательности 87 His-Ala-Gly-Pro-lle-Ala 92. Происходит конформационное изменение в капсидном белке, облегчая следующий этап.
[0127] Диплоидный вирусный PHK/p7/p6/Vpr/Vpx(BH4-2) комплекс является основанием для сборки капсидного (р17 или СА) белка с СурА молекулой. Приблизительно 2000 молекул р17/вириона необходимо для завершения следующего этапа. Только 200 СурА молекул встраиваются в каждый ВИЧ вирион. Идеально соотношения собирающихся вирусных и клеточных белков должно отражать финальную композицию интактного вирион. ВИЧ-2 вирион не ассимилирует СурА молекулу и поэтому этот этап не является необходимым для сборки ВИЧ-2. Собранный капсидный белок/СурА комплекс принимает циллиндрическую форму и является основанием для сборки матричного белка (р24).
[0128] Матричный белок формирует икосаэдр вокруг капсидного циллиндра. Приблизительно 2000 мономеров матричного белка самопроизвольно собираются для формирования икосаэдра. Эта структура является основанием для ассиммиляции gp41 молекул. Кристаллическая структура МА является трехмерной и тримеризация МА структурного белка оказывается консервативным свойством антивирусов (A. Cimarelli, et al.). Молекулы gp41 самопроизвольно собираются в гомотримеры на внешней поверхности икосаэдра матричного белка. Молекула gp120 является самым внешним белком вириона и является последним из тех, которые должны собраться в вирусе. Тример gp41 принимает трехмерную структуру и проявляет электростатические свойства, которые совпадают с gp120 молекулой. Взаимодействие gp41/gp120 может быть связано с "мячом для гольфа" (gp120), сидящим на верхушке мишени (gp41). Семьдесят два тримера gp41/gp120 формируют внешнюю белковую оболочку вируса.
[0129] Взаимодействия вирусной РНК с белком, а также взаимодействия вирусного белка с вирусным белком и вирусного белка с клеточным белком хозяина опосредованы нековалентной связью, такой как Вандерваальсовы силы, водородной связью и диполь-дипольным моментом. Эти межмолекулярные взаимодействия определяют порядок сборки вириона и конечную трехмерную структуру вириона.
[0130] На цитоплазматической стороне плазматической мембраны ВИЧ инфицированной клетки Gag полипротеин и Gag-Pol полипротеины аккумулируются в соотношении 10 к 1. Фермент протеаза расщепляет эти полипротеины в течение и после, но не до процесса вирусного бадинга. Протеазная активность возникает после того, как про-вирион приобретает клеточную оболочку. Отмечается, что в каждом ВИЧ вирионе ассимилируется приблизительно 2000 копий р7 и р24 белка. Приблизительно 90% р7 и р24 белков происходят от Gag полипротеинов и приблизительно 10% от Gag-Pol полипротеина. Расщепление этих полипротеинов происходит в течение бадинга в интактной клеточной оболочке, и поэтому все индивидуальные белки должны быть ассимилированы в интактном вирионе.
[0131] Как вывод, 2000 копий р7 и р24 и приблизительно 1800 происходят от Gag полипротеина и 200 от Gag-Pol полипротеина. В этой точке сборки вириона потеря мономеров индивидуального белка маловероятна из-за расщепления полипротеинов в оболочке бадинг вируса и повсеместной высокой эффективности сборки вириона. Поэтому разумно допустить, что 2000 копий р6 и р17 включаются в каждый ВИЧ вирион. Gag и Gag - Pol полипротеин кодирует в направлении от 5′ к 3′ одну копию р17, р24, р7, р6 и с Gag - Pol одну копию каждого из вирусных ферментов. Таким образом, до 200 копий каждого из вирусных ферментов могут встраиваться в каждый вирион. В бесклеточной системе экспрессии для самосборки вирусных частиц соотношения вирусных белков, включая ферменты, должны быть в разумной степени согласующимися.
[0132] Матричный белок облегчает ядерное мечение комплекса предварительной интеграции и мечение плазматической мембраны только что транскрибированных gag полипротеинов. Матричный белок в gag полипротеине связывается с клеточным миристоиловым компонентом. Это позволяет происходить направленным изменениям в матричном белке. Nef белок также миристоилируется в процессе вирусной сборки, поляризуя цитоплазматическую сторону плазматической мембраны. В описанной выше методике избегают включение миристоилового компонента в матрикс и белок Nef. Мистроиловые компоненты добавляют после проникновения вируса в намеченную клетку, и они не являются компонентами интактного вириона. Миристоиловый компонент в интактном вирионе не должен подвергаться ядерной локализации PIC. Поэтому бесклеточная среда, используемая для получения ВИЧ вириона, должна быть лишена миристоиловых компонентов или подобных жирнокислотных соединений. Ферменты, которые катализируют миристилирование, также должны быть удалены.
[0133] Матричный белок связывается специфически с внутренним цитоплазматическим доменом gp41. Гликопротеин gp41 нековалентно прикрепляет gp120. Слияние плазматической мембраны вокруг бадинг-вириона(вирионов) вначале осуществляется в незрелой, неинфицированной вирусной частице. Вирусный фермент протеаза затем продолжает расщеплять gag и gag-pol полипротеины, давая в результате инфекционную частицу.
[0134] Кроме того, процесс самосборки может регулироваться модулированием следующих параметров:
РН;
осмолярность;
температура;
относительная пропорция вирусных белков;
порядок добавления вирусных белков;
включение содействующих или ингибиторных невирусных соединений;
интенсивность, частота и длительность света, особенно света в ультрафиолетовом диапазоне.
[0135] Предпочтительно, рН, осмолярность и температура должны отражаться на внутриклеточной среде: (рН=7), осмолярность (=280 мосм), температура =37°С. Ультрафиолетового излучения, в частности, при 260 нм необходимо избегать. При этой длине волны наблюдаются конформационные изменения как в РНК, так и ДНК. В частности, тиминовые димеры наблюдаются в ДНК как результат нахождения в ультрафиолетовом излучении при 260 нм. Относительные пропорции вирусных белков должны отражать соотношение белковых мономеров в интактном вирионе. Порядок добавления вирусных белков зависит от желаемого конечного продукта, но, в общем, для поддержания порядка в системе, внутренние структурные белки типично являются начальным пунктом, тогда как конечным пунктом являются самые внешние структурные белки. В общем, последовательность Env гена и Gag гена в направлении от 3′ к 5′ кодирует вирусные белки, отражая это расположение от внутреннего к внешнему краю. Вирусные кодируемые ферменты, а также определенные добавочные белки (Vpr в ВИЧ-1 и ВИЧ-2 и Vpx в ВИЧ-2) включаются в капсидное ядро, но не являются структурными. Включение этих белков является необходимым для этой композиция или вакцины, так как рассматривается условно живой вирион, не способный к репликации. В композиции, способной к репликации, также необходимы эти ферменты, кодируемые вирусом, и вышеупомянутые добавочные белки.
[0136] Получение вириона может катализироваться ферментом протеазой, кодируемым вирионом, который расщепляет Gag полипротеин и Gag-Pol полипротеин на индивидуальные белковые компоненты в определенной по времени последовательности, что оптимально облегчает внутриклеточную вирусную продукцию. Nef белок также расщепляется протеазой в процессе и после бадинга. Поэтому включение этого фермента с Gag полипротеином (или Gag полипротеином и Gag-Pol полипротеином в соотношении 10:1) является альтернативным способом получения вируса (по сравнению с последовательным добавлением каждого белка).
[0137] Вирусная самосборка не является АТФ или ГТФ затратным процессом. Следовательно, каждый этап следует логично из предыдущего этапа, давая в результате состояние с меньшей энтропией. Энтропия - это количество возможных расположений элементов в любой системе. Это мера случайности или дисперсии. Без затраты энергии материя распадается на структуры с меньшей энтропией. Для поддержания изменчивости должна потребляться энергия, и живые клетки очень отличаются по их источникам энергии, что приводит к поддержанию этого соотношения дисперсии/недисперсии. Вирусы не являются живыми структурами. Они не производят или потребляют АТФ или ГТФ, тесно зависят от машинерии клеточной транскрипции и трансляции хозяина.
[0138] Энтропия, как она обычно рассматривается, не применяется к вирусам. За исключением наиболее сложных вирусов, которые могут представлять мост между вирусами и бактериями, вирусные структуры принимают одно из двух возможных низкоэнтропийных состояния: икосаэдральное или спиралевидное. Внешняя структура ВИЧ является икосаэдральной структурой, характеризующейся двадцатью треугольными гранями и двадцатью ребрами, и может рассматриваться с двукратной, трехкратной или пятикратной осью вращения симметрии. Хотя gp120 и gp41 гликопротеины являются внешними или поверхностными белками ВИЧ вируса, нижележащий матричный белок определяет икосаэдральную структуру.
[0139] Икосаэдральные симметричные структуры основаны на числе треугольников, Т, числе структурных единиц на грань. Минимальное количество субъединиц для самосборки в икосаэдр составляет 60. В случае только 60 субъединиц каждая должна быть идентична для получения икосаэдра. В этой модели Т=1. Если существует более 60 единиц в вирусном структурном белке, каждая единица или субъединица находится в квазиэквивалентном положении, которое определяется свойствами нековалентного связывания этих субъединиц. Хотя различные структурные окружающие условия могут определять больший икосаэдр, свойства нековалентного связывания субъединиц подобны (но не обязательно идентичны, что характерно для наиболее простой структуры из 60 субъединиц). Регулярность и точная подгонка молекул в любой структуре делает возможным внутренние молекулярные структуры, водородную связь, диполь-дипольные взаимодействия и вандерваальсовы силы.
[0140] Эластичность субъединичного белка(белков), который составляет внешнюю или внутреннюю структуры, дает другое измерение самосборке вирусного капсида. Структурные комплементарности между смежными капсидными мономерами, а также координированные электрические взаимодействия определяют сформированную мульти-субъединичную белковую структуру. Каждая субъединица, поэтому, может иметь множество доменов, каждый со своей собственной трехмерной структурой с каждым доменом, принимающим определенную ориентацию по отношению к другим доменам.
[0141] Поэтому правило количества треугольников по отношению к некоторым вирусам может не применяться. При рассмотрении отдельных эластичных белковых доменов каждый рассматривается как отдельная структура, и применяется правило числа треугольников. Без расходования энергии вирусная сборка следует определенной последовательности, что приводит к структуре с наименьшей энтропией. Кристаллическая структура МА является трехмерной и тримеризация МА структурного белка является консервативным свойством антивирусов (Cimarelli, et at.).
[0142] Структурные, ферментативные и добавочные генные продукты, необходимые для получения вириона, могут быть получены in vitro. Генетический перенос с применением родового ретровирусного вектора (RV) является хорошо описанным способом генной трансдукции. С использованием как 5′, так и 3′ LTR, сигнального сайта упаковки (ψ сайт) и полипуринового тракта генный вектор может быть встроен в клеточную культуру, такую как дрожжи, Е. coli или стабильная клеточная линия, такая как HeLa (Michael, 1998, Ch. 24). Генетические последовательности, кодирующие один или более маркерных белков, могут быть включены в ретровирусный вектор как экзоген. Нуклеокапсидный (р7), р6, капсидный, матричный, gp41 и gp120 структурный белки могут быть получены в клеточной культуре генного переноса и выделены. Аналогично, геномная последовательность для ретровирусных ферментов и добавочных белков, включенная в интактный вирион, может быть встроена в клеточную культуру и выделена.
[0143] Белки Tat и Rev не являются необходимыми для вирусной сборки, после бадинг не являются структурными белками, и поэтому генетические последовательности, кодирующие эти белки, не нуждаются в расщеплении в тканевой культуре для получения интактного вириона. Nef белок упакован в ВИЧ вирионы, где вирусная протеаза его расщепляет. ВИЧ провирусный ДНК синтез менее эффективен без Nef белка. Nef белок, однако, не является необходимым для вирусной репликации, созревания и бадинга. Предпочтительно белковые компоненты вируса, генетически кодированные в линиях с дефектом упаковки, будут добавлены последовательным образом, что является аналогичным нормальной вирусной сборке и будет включать Nef белок.
[0144] Точная последовательность сборки ВИЧ вириона, начинающаяся с наиболее внутренней структуры и оканчивающаяся в наиболее наружной структуре, определяет последовательность белков и РНК, которая соблюдается в сборке условно живых интактных вирионов. Вирусные компоненты в соответствующих соотношениях добавляют в одном варианте осуществления последовательным образом, что отражает природный процесс самосборки. Избыток белков удаляется центрифугированием или другим процессом до следующего этапа. Вирион является технически репликационно некомпетентным, так как геномная информация, кодирующая одну или более белков, необходимых для репликации в интактном хозяине, удалена.
[0145] В описанном выше варианте осуществления может быть использована бесклеточная система. Поэтому оболочка не будет частью вирусной структуры. Оболочка является приобретенной после сборки вириона и до бадинга на цитоплазматической стороне плазматической мембраны.
[0146] Вакцина против гепатита В является аналогичной в части вышеупомянутых понятий нормального оболочечного вируса, независящего от оболочки для сборки вириона, структуры и стабильности. Вакцину против гепатита В получают в культуре дрожжей, и она содержит один вирусный структурный белок: поверхностный антиген вируса гепатита В. Этот структурный белок спонтанно собирается в стабильные вирусоподобные частицы. Эти частицы лишены оболочки, но остаются стабильными и иммуногенными. Интересно, что гепатит В кодирует фермент обратную транскриптазу, подобную ВИЧ. Гепатит В является ДНК вирусом, а ВИЧ является РНК вирусом.
[0147] Альтернативно, вакцина может быть получена в клеточной линии как стабильной, так и любой другой. Геном условно живого вириона, как упоминалось выше, может быть расщеплен в клеточной линии (стабильной или нестабильной). Это может осуществляться с помощью ферментной рестрикции, как обсуждалось выше, в получении субъединичного компонента по данному изобретения. Экзогенный белок(белки), не кодированный в последовательности нуклеиновой кислоты, может быть введен дополнительно для облегчения и регуляции репликации. Альтернативно, интактный условно живой вирион с добавленным модифицированным белком репликации может быть помещен in vitro в тканевую культуру. Это дублирует вакцинную методику, описанную выше, в тканевой культуре. Репликация вируса будет регулироваться частично с помощью количества и времени полужизни экзогенно добавленного белка. Альтернативно, биологически активные белки или белковые фрагменты модифицированной генной последовательности могут быть добавлены к in vitro тканевой культуре, инфицированной условно живым вирионом.
[0148] Применение in vitro клеточной линии или культуры для культивирования ВИЧ ведет к сборке вирусных структур, которые будут нести генотипические и фенотипические отличия от ВИЧ вирионов, полученных из естественной среды или хозяина (например, человека). Это является возможным аспектом второго способа с точки зрения применения. Второй способ, однако, требует меньшего количества этапов и может протекать в стабильной клеточной культуре на неопределенно долгое время, если обеспечиваются соответствующие питательные вещества и вся клеточная культура благоприятна для продолжительной клеточной репликации.
Получение экзогенного белка (Субъединицы)
[0149] Условно живой вирусный вирион будет нуждаться в экзогенной поддержке дефицитными белками репликации. Этот белок репликации может быть получен в клеточной культуре с помощью генного переноса. Встраивание белка(белков) в клетку хозяина и вирусную частицу можно выполнить с помощью сцепления белка с проникающим в клетку пептидом. Проникающий в клетку пептид (СРР) является олигомером, состоящим из 5-40 аминокислот, который способен к прохождению через плазматическую мембрану клетки и внутриклеточной доставке множества конъюгированных биоактивных соединений. Ряд механизмов, включая эндоцитоз, описан в литературе для объяснения механизма действия (СРР). Доставляемый груз может быть ковалентно или нековалентно прикреплен к СРР. (Gellissen; Langel)
[0150] В идеале последовательность нуклеиновой кислоты, кодирующая один или более белков, по которым дефицитен условно живой вирион, получают из интактной нуклеиновой кислоты того же вирусного источника. В одном варианте осуществления нуклеотидную последовательность, кодирующую два или более смежных белка, отщепляют от интактной последовательности нуклеиновой кислоты. Перекрывающиеся и неперекрывающиеся сегменты удаляют и поэтому возможно использование в in vitro векторе экспрессии для получения полной аминокислотной последовательности этих белков. Если только один белок является модифицированным или дефицитным в вирусном векторе, то только неперекрывающийся считывающий сегмент этого белка удаляют. Эта неперекрывающаяся последовательность нуклеиновой кислоты не будет достаточной для генного переноса. Однако расщепление перекрывающейся и неперекрывающейся последовательности, кодирующей один белок, будет делать возможным генный перенос. Источник генетического материала в этом примере может или может не быть приемлемым для условно живого вирусного вектора. Обеспечение белками, которые кодируются как перекрывающимися, так и неперекрывающимися сегментами в тканевой культуре будет приводить к вирусной репликации и сборке. В in vitro клеточной линии, в которой один или более белков, которые кодируются дефицитными нуклеиновыми кислотами, в конкретном вирионе дополняются с помощью продукции белка другим вирионом в той же клетке или той же клеточной культуре, ассимиляция дефицитного белка(белков) вирионом будет происходить транс-образом, облегчая вирусную репликацию и сборку.
[0151] В случае изолированной нуклеиновой кислоты, кодирующей один или более модифицированных белка, должен быть выбран приемлемый вектор экспрессии. Используются классические плазмиды, кольцевые двухнитевые ДНК молекулы, содержащиеся в экстрахромосомном сайте в цитоплазме клетки. Плазмиды представляют собой маленькие молекулы, содержащие источник репликации для того, чтобы позволить копирование ДНК, произвольный маркер для визуализации вектора, и один или более уникальных сайт рестрикции рестрикционной эндонуклеазой, что позволяет вставить намеченную ДНК для крупномасштабного производства. Плазмиды обычно не являются необходимыми для выживания клетки, но зачастую они обеспечивают выборочные характеристики, позволяющие организму выжить в условиях, отличных от идеальных. Определены некоторые природно встречающиеся плазмиды и они являются доступными для лабораторных методик генного переноса. Другие коммерчески доступные плазмиды представляют собой продукт методик генного переноса, и не обнаруживаются за пределами лаборатории.
[0152] Плазмиды обнаружены только у прокариотических организмов в условиях, которые испытывают недостаток ядерных мембран. Поэтому транскрипция и трансляция происходят одновременно. Посттранскрипционная модификация не может происходить у прокариот. Без посттранскрипционной и посттрансляционной модификаций белковые последовательности, кодируемые вирусами, которые инфицируют млекопитающих, такие как ВИЧ, могут принимать структуру у прокариота, которая значительно отличается от той, которая наблюдается у нормального хозяина. Поэтому эти белки, продуцируемые плазмидами и прокариотами, такими как Е. coli, могут не быть функциональными при ассимилировании в нормальной эукариотической клетке-хозяине. Вирусные белки из плазматической мембраны в прокариотической системе экспрессии могут требовать дополнительные этапы модификации до встраивания в интактный вирион (Desmond S.Т. Nicholl, Ch. 5).
[0153] Эукариотический организм, который наиболее часто используется в генетической инженерии, представляет собой дрожжи Saccharomyces cerevisiae. Он обычно используется для массовой продукции вакцины против гепатита В, которая состоит из одного структурного белка вируса, поверхностного антигена гепатита В. (Nicholl, Ch. 5) Saccharomyces cerevisiae посттранскрипционная и посттрансляционная модификация белков строго параллельна посттранскрипционной модификации белков в клетках млекопитающего.
[0154] Бактериофаги использовались для переноса ДНК в Е. coli. Бактериофаги представляют собой вирусы, которые инфицируют бактерии. Другие векторы для генного переноса состоят из плазмидных последовательностей, связанных с нуклеиновой кислотой бактериофага, известны как космиды. Эта технология четко определена в литературе (Nicholl, Ch. 5).
[0155] Эукариотические клетки делают возможной посттранскрипционную и посттрансляционную модификацию белков. Поэтому они являются предпочтительными системами экспрессии для вирусных белков, инфицирующих млекопитающих. В дрожжах множество генетически сконструированных векторов, включая, но не ограничиваясь следующими: дрожжевые эписомальные плазмиды, дрожжевые интегральные плазмиды, дрожжевые репликативные плазмиды, дрожжевые центромерные плазмиды и дрожжевые искусственные хромосомы, описаны в литературе и могут использоваться для получения ВИЧ вирусных белков in vitro. Кроме того, также известны бактериальные искусственные хромосомы (bас). Вас испытывают, однако, недостаток в аппарате посттранскрипционной и посттрансляционной модификации (Nicholl, Ch. 5).
[0156] Плазмиды представляют собой идеальный механизм экстрахромосомной белковой продукции, но, в основном, они ограничены только прокариотическими организмами. Эктрахромосомное расположение, а также способность одной клетки ассимилировать множество идентичных плазмид, делает возможной непрерывную продукцию белка. Экстрахромосомное расположение выносит намеченную последовательность нуклеиновой кислоты за предел контроля хромосомы организма. Транскрипция бактериальной хромосомы находится под контролем промотеров. Промотеры, однако, контролируют только генетические последовательности в цис-положении. Промотеры поэтому не контролируют транскрипцию плазмид.
[0157] Плазмидная ДНК, встроенная в клеточные культуры млекопитающего, обычно приводит либо к деградационной потере плазмиды, либо к интеграции плазмиды в хромосому хозяина, и поэтому она находится под контролем хромосомы хозяина. Большинство хромосом хозяина является неактивными в клеточной транскрипции (гетерохроматин). Вставка плазмиды в или рядом с хроматином будет приводить к потере плазмидной генетической экспрессии. (Klug. et al.)
[0158] Отмечено одно исключение из плазмидной интеграции в клетки млекопитающего. Плазмида, содержащая источник репликации вируса Эпштейна-Барра, вирусно кодируемый ядерный антиген вируса Эпштейна-Барра (EBNA-1), связывающий сайт EBNA-1, и произвольный маркер обеспечивают платформу для такой плазмиды. При удалении плазмидного источника репликации и замещении его случайными частями человеческого генома плазмидный вектор, полученный таким образом, при введении в in vitro клеточную культуру млекопитающего, остается экстрахромосомным по своему расположению и реплицируется автономно. Нуклеотидная последовательность для одного или более ВИЧ белков может быть расщеплена в этой плазмиде. Плазмида, помещенная в эукариотическую клеточную культуру, будет ассимилироваться в цитоплазме. Ядерное мечение плазмиды не будет происходить. Транскрипция, трансляция и посттрансляционная модификация вирусных генов будет происходить без ядерного контроля, и поэтому последовательным образом в присутствии EBNA-1, добавленного экзогенно (RLP Adams). Это является подходящим механизмом для in vitro продукции ВИЧ вирусных белков, если используется клеточно ассоциированная среда.
[0159] Может также рассматриваться изолирование плазмидного вектора, который не является коммерчески доступным. Приемлемой культуральной средой для роста бактериальных клеточных культур для изолирования плазмид является среда Лурия-Бертани (LB). Коммерческие наборы, такие как щелочной лизис быстрой экстракции (R.E.A.L.) Prep 96 Kits, позволяют проводить быстрое изолирование плазмид, космид, bас и фаговых искусственных хромосом. Способы на основе диоксида кремния являются также надежными способами для изолирования ДНК плазмид (Bowien, et al., Ch. 3). Процесс, начинающийся с плазмидного изолирования, до in vitro плазмидного конструирования, очистки и коммерциализации, описан в литературе (Botho Bowien, et al., Ch. 4).
[0160] Могут использоваться альтернативные способы изолирования индивидуальных белковых компонентов ВИЧ вириона. Супернатант in vitro клеточной культуры, такой как HeLa клетки, инфицированные ВИЧ, могут быть отделены от клеточной культуры, и индивидуальные вирусные белки идентифицированы с помощью гелевого электрофореза, центрифугирования или других способов. Альтернативно, очищенная клеточная культура может быть гомогенизирована до отделения индивидуальных ВИЧ белков. Оба способа, несмотря на приемлемость, не являются предпочтительными из-за контаминирующего материала и вышеупомянутых генотипических и фенотипических отличий между полевыми изолятами и in vitro ВИЧ клеточными культурами.
[0161] Дополнительно химический синтез белков можно выполнить с множеством аминокислотных последовательностей.
[0162] Пути конструирования векторов известны специалистам в данной области (например, как проиллюстрировано в Патенте США №7132271). Примеры включают с использованием химически и ферментативно синтезированной ДНК, фрагменты вирусной кДНК или намеченные гены. Кроме того, трансфекция клеточной культуры проводится стандартными способами, например способ DEAE-дектсран (McCutchen и Pagano), методика кальция фосфата (Graham et al), или любым другим способом, известным из уровня техники, включая, но не ограничиваясь следующим: микроинженерия, липофекция и электропорация. (Sambrook et al.) Выбирают трансфектанты, имеющие недостаточную репликацию или другую активность. Для простоты отбора маркерный ген, такой как неомицин фосфотрансфераза II, устойчивая к ампициллину или G418, может быть включен в вектор, несущий антисмысловой или мутантный ген. Когда включают маркерный ген, трансфектант может быть выбран для экспрессии маркерного гена (например, устойчивого к антибиотику), культивирован и далее испытан на выбранную активность.
[0163] В хозяине, совместно инфицированном двумя или несколькими штаммами ВИЧ, выявлены циркулирующие рекомбинантные формы, состоящие из нуклеотидных сегментов различных вирусов. Также отмечена совместная зависимость нежизнеспособных вирионов кодировать белки, которые совпадают с дефицитными белками, каждый из которых кодирует, приводя к репликации и развитию одного или более в других случаях нежизнеспособных вирионов (Flint, et al., 2004, Ch. 20). Существует параллель между эукариотическими и ВИЧ вирионами, которые являются диплоидными. Эукариоты обладают ядерной мембраной, и типично имеют диплоидной набор хромосом. Поэтому, дефицитный белок. Колируемый на одной хромосоме, может не воздействовать на жизнеспособность организма, если его комплиментарная хромосома кодирует недефицитный белок.
[0164] ВИЧ является диплоидным вирионом (в отличие от большинства вирусов, которые являются гаплоидными), и подобно эукариотическим организмам нуклеотидные последовательности РНК нитей не должны быть и зачастую не являются идентичными. В клетке, инфицированной более чем одним штаммом ВИЧ, существует множество возможностей для одного штамма, чтобы обойти дефективные белки, кодирующие один или более других штаммов. Чем больше количество различных штаммов, совместно инфицирующих клетку, тем больше возможностей существует для развития нежизнеспособных штаммов. Это объясняет, частично, как вирусная мутация обеспечивает выживание вирусов.
[0165] Данное изобретение строится на вышеописанных явлениях, которые являются характеристикой ВИЧ вирусной эволюции. Вирусный геном, дефективный по одному или более белкам, будет становиться жизнеспособным, если эти дефективные белки обеспечиваются другим источником. В одном варианте осуществления данного изобретения множество вирусных штаммов будут удалять ту же намеченную последовательность нуклеиновой кислоты, как описано выше.
[0166] Альтернативная in vitro методика получения условно живого вириона включает совместное инфицирование тканевой культуры первым условно живым вирионом и вторым вирионом (условно живым или каким-либо другим), который включает последовательность нуклеиновой кислоты, расщепленную от первого условно живого вириона. Это делает возможным вирусную репликацию. Однако рекомбинаторное событие в такой культуре может происходить, потенциально позволяя появиться репликационно компетентному вектору.
[0167] Некоторые добавочные вирусные белки, включая Vpr, Vif и Vpx (и Vpx в ВИЧ-2), обнаружены в интактном вирионе. Они могут поддерживать один или несколько циклов вирусной репликации и могут быть в достаточном количестве для образования соответствующего Th-1 ответа в иммунологической памяти и последующего иммунитета в условно живом вирионе, дефицитном по последовательности нуклеиновой кислоты(кислот) для Vpr, Vif, Vpx или комбинации. Как только снабжение дефицитными белками исчерпывается, репликация прекращается. Поэтому условно живые вирионы, дефицитные по последовательности нуклеиновой кислоты(кислотам) для Vpr, Vif, Vpx или комбинации, могут или могут не нуждаться в поддержке экзогенным белком в некоторых условиях. Количество циклов репликации вируса в конечном итоге продолжает регулироваться и контролироваться количеством и временем полужизни дефицитного белка(белков), но в данном способе может быть не включено добавление ни одного экзогенного белка.
[0168] В одном варианте осуществления данного изобретения нокаутный вирион для vpr последовательности метят и удаляют, как описано выше. Но так как каждая ВИЧ вирусная частица ассимилирует приблизительно 100 копий Vpr белка, экзогенно добавленный Vpr белок может или может не быть необходимым (Cohen, et al., Ch. 16). Как только поддержка Vpr белками исчерпывается, вирусная инфективность, вирулентность и репликация будет подвержена серьезному риску. Интактная иммунная среда здорового хозяина будет проявлять соответствующий ответ и устранять внутриклеточный реплицирующий вирус.
[0169] Возможно, что вакцина или композиция вводится как чистый или в значительной мере чистый вирион совместно с экзогенно добавленным белком, или как фармацевтическая формуляция или препарат, необязательно с адъювантами или другими композициями.
[0170] Формуляции, применяемые на практике по данному изобретению как для ветеринарного, так и для человеческого использования включают нокаутированные вирионы совместно с экзогенно добавленным белком репликации, как описано выше, вместе с одним или более фармацевтически приемлемыми носителями и необязательно другими терапевтическими ингредиентами. Белковые носители должны быть "фармацетвически приемлемыми" в том смысле, что они должны быть совместимы с другими ингредиентами формуляции и не опасны для реципиента, их принимающего. Сцепление белковых носителей (например, белки комплемента) известны в фармакологии.
[0171] Желательно, чтобы формуляция не включала другие соединения, с которыми ВИЧ вирион, как известно, несовместим. В соответствии с современными фармакологическими стандартами способы включают этап доведения до взаимодействия условно живого вириона и экзогенно добавленного белка репликации с носителем, который может включать один или более добавочных ингредиентов. Формуляции, пригодные для введения с помощью инъекции, соответственно включают стерильные водные растворы для вакцины, причем эти растворы являются преимущественно изотоническими крови реципиента. Такие формуляции могут быть приготовлены для получения фармакологически приемлемого стерильного водного раствора.
[0172] В одном варианте осуществления данного изобретения дефицитный белок будет соединяться с пептидом, проникающим в клетку, таким как Пенетратин, фрагмент Tat белка (аминокислоты 48-60) Транспортан, пептиды, основанные на сигнальной последовательности, аргининовые полипептиды, pVEC и/или амфифильные модельные пептиды. Соединение таких пептидов, как перечисленные выше, хорошо известно специалистам в данной области и они будут облегчать проникновение намеченного дефицитного белка через плазматическую мембрану клеток хозяина (UIo Langel).
[0173] В другом варианте осуществления данное изобретение стоит на основе знаний о том, что убиквитинированные белки коррелировали с множеством клеточных функций, включая, но не ограничиваясь следующим: процессинг и презентация антигенов Т-клеткам. (Krauss). Экзогенно добавленные белки могут быть полиубиквитинированными. Для облегчения проникновения в протеасомный путь интактный вирион может также быть полиубиквитинированным. Поэтому конъюгирование экзогенно добавленных вирусных белков и/или условно живого вириона с убиквитином будет направлять экзогенно добавленный белок и/или условно живой вирион в протеасомный путь, приводя к Th-1 иммунному ответу на основе MHC-I к одному или нескольким эпитопам на этом белке и/или вирусном векторе. В объеме данного изобретения существуют варианты осуществления, в которых часть экзогенного белка является убиквитинированной; например, половина экзогенного белка может быть убиквитинированной.
Альтернативные варианты осуществления, основанные на расщепленных нуклеотидных последовательностях
[0174] Любая возможная комбинация последовательного удаления и лигирования используется настолько долго, чтобы сформировался условно живой вирион. Любой добавочный или регуляторный белок, аномальный по последовательному удалению, может быть обеспечен экзогенно, что позволяет вирусной репликации проходить внутриклеточно в "нормальной", но ограниченной манере. В настоящее время описано пятнадцать (15) условно живых вирионов с ВИЧ-1 и 15 условно живых вирионов с ВИЧ-2, которые являются исключительно примерами и не служат для ограничения объема изобретения.
[0175] В примере и в простейшем варианте осуществления перекрывающаяся и неперекрывающаяся геномная последовательность, кодирующая vif, vpr, tat (экзон 1), vpu (ВИЧ-1) и vpx (ВИЧ-2), может быть удалена индивидуально или в комбинации с другой, применяя ферменты рестрикции. Перекрывающиеся сегменты vif с pol и vpu с env не должны быть удалены, оставляя эти гены и белки, которые ими кодируются, интактными. Используя эту систему нокаута, пять отдельных белков (Vif, Vpr, Tat (экзон 1), Rev экзон 1 и Vpu в ВИЧ-1 или Vpx в ВИЧ-2), которые могут быть или могут не быть включены в капсулу интактного вириона, могут быть добавлены экзогенно.
[0176] В альтернативном варианте осуществления используется иммуногенная композиция, лишенная неперекрывающейся геномной последовательности tat экзона 1 с vpr. Чистый Tat белок, кодируемый экзоном 1 и 2, затем экзогенно добавляют как часть иммуногенной композиции или вакцины. Любая формуляция, в которой нуклеотидную последовательность tat экзона 1 удаляют, будет также приводить к удалению нуклеотидной последовательности rev экзона 1. Rev экзон 1 полностью перекрывается с tat экзоном 1 в большинстве вирусных изолятов. Поэтому два белка необходимо добавить к вирусной композиции, Tat и Rev. Предпочтительно добавляют чистые Tat и Rev белки, которые кодируются двумя отдельными экзонами. Tat экзон 2 полностью перекрывает rev экзон 2 и оба полностью перекрывают env. В еще одном варианте осуществления иммуногенную композицию, лишенную геномной последовательности rev экзона 1, удаляют и чистый Rev белок, кодируемый экзоном 1 и 2, а также чистый Tat белок будут экзогенно добавлены.
[0177] Интронная последовательность, локализована между 3′ концом tat или rev экзон 1 и 5′ концом vpu белка предпочтительно не будет расщепляться любой из выше- или нижеперечисленных вакцин. Полиуриновый путь, важный в обратной транскрипции РНК ВИЧ генома, обнаружен в этой интронной последовательности. В ВИЧ-2 подобная последовательность обнаружена между 3′ концом vpx нуклеотидной последовательности и 5′ концом vpr нуклеотидной последовательности. Аналогичным образом сплайсинг этой последовательности не будет предпочтительным.
[0178] Удалением только одного неперекрывающегося генного сегмента можно развить четыре отдельных условно живых вирионов для ВИЧ-1 и четыре для ВИЧ-2. Белки, кодируемые усеченными генами, могу быть введены экзогенно.
[0179] Если два неперекрывающихся генных сегмента удалить, возможно шесть условно живых вирионов. Белки репликации, кодируемые удаленным геномом, могут быть добавлены экзогенно. Если геномы для двух последовательных белков удалены, то перекрывающиеся сегменты этих двух геномов могут также быть удалены. Это будет увеличивать безопасность и простоту устройства и производства, так как только два "разреза" вирусной последовательности нуклеиновой кислоты будет необходимо сделать вместо четырех, и только один повторный процесс отжига - вместо двух. То есть, удаление непоследовательных неперекрывающихся сегментов будет обычно требовать больших количеств "разрезов" и больших повторных отжигов.
[0180] Удаление трех неперекрывающихся генных сегментов дает на выходе четыре возможных условно живых вирионов. Белки, кодируемые удаленным геномом, могут быть добавлены экзогенно.
[0181] При удалении всех четырех неперекрывающихся геномных сегментов в результате остается один живой вирион. Поэтому пятнадцать отдельных условно живых вирионов могут быть созданы с ВИЧ-1 и пятнадцать для ВИЧ-2. Приведенный список отражает частично и/или полностью удаленные геномные последовательности потенциальных композиций.
ВИЧ-1:
vif
vpr
tat экзон 1
vpu
vif и vpr
vif и tat экзон 1
vif и vpu
vpr и tat экзон 1
vpr и vpu
tat экзон 1 и vpu
vif, vpr, и tat экзон 1
vif, vpr, и vpu
vif, tat экзон 1, и vpu
vpr, tat экзон 1, и vpu
vif, vpr, tat экзон 1, и vpu
ВИЧ-2
vif
vpr
tat экзон 1
vpx
vif и vpr
vif и tat экзон 1
vif и vpx
vpr и tat экзон 1
vpr и vpx
tat экзон 1 и vpx
vif, vpr и tat экзон 1
vif, vpr, и vpx
vif, tat экзон 1 и vpx
vpr, tat экзон 1 и vpx
vif, vpr, tat экзон 1 и vpx
Введение и адъюванты
[0182] Иммуногенная композиция или вакцина может быть введена посредством эритроцит - опосредованной микроинъекции. Эритроциты лизируют в гипотонической среде in vitro. Вакцину добавляют в раствор. Мембрана красной кровяной клетки очень пориста в гипотоническом растворе и позволяет большим белкам из внеклеточной среды проникать в клетку. (Doherty, et al.) Красные клетки затем помещают в раствор с нормальной тоничностью (0,9% NaCl). Разрушенные красные клетки секвестируются и разрушаются в селезенке и печени. Антигенпрезентирующие клетки в обоих органах, в частности Купферовские клетки, выстилающие синусоидные капилляры печени, будут захватывать инородный материал и презентировать его соответствующим Т-клеткам.
[0183] В другом варианте осуществления для введения вакцины интактная кожа, орган тела с минимальной иммунологической активностью, может становиться эффекторным органом иммунной системы, если его барьеры нарушены. От двух до трех дней до вакцинации область, получившая вакцину, механически соскабливают, создавая внешнее трение. Триггерные эффекторные клетки и белки как врожденного, так и приобретенного иммунного ответа будут секвестироваться в этот пораженный участок, готовя его к вакцине. Размещения под ультрафиолетовым излучением, которое вызывает иммуносупрессию, необходимо избегать. Вакцину будут вводить внутрикожным путем в стертую кожу. Это приводит к анатомически определяемому иерархическому иммунному ответу, который аналогичен архитектуре лимфатического узла. Это представляет собой предпочтительный способ введения вакцины по изобретению.
[0184] Предпочтительная область вакцинации должна быть выше середины бедра. Отток лимфы из этой области направлен к паховым лимфатическим узлам. Количество паховых лимфатических узлов варьирует от 12 до 20 в количестве и они не только фильтруют лимфу от нижних конечностей, но также лимфу от наружных половых органов, промежности, ягодиц и нижней части заднепроходного канала (Ben Pansky). Это сайты первичной ВИЧ инфекции и распространения при половом пути передачи этой болезни.
[0185] Данное изобретение также предполагает, что адъюванты или другие композиции, предусматриваемые для поддержания иммунного ответа на вакцину, могут быть добавлены ко всем описанным выше вакцинным коктейлям. Такие адъюванты предпочтительно существуют в форме для связывания в коктейле. Такие композиции могут включать, но не ограничиваться, полисахаридами, включающими, по меньшей мере, одну молекулу маннозы, тейхоевой кислоты, зимозана, полисахаридную капсулу Cryptococcus neoformans серотип С, Протамин, гепариназу, фактор яда кобры в форме, адаптированной для усиления продукции С3b, фактор яда кобры в форме dCVF, никель в форме, адаптированной для усиления С3 конвертазной активности, сульфатированные полианионы. Операции с этими адъювантами были описаны ранее в публикации патента США №20050112139, которая включена в данное описание ссылкой. Примеры дополнительных адъювантов могут включать:
Дополнительные адъюванты
(a) Белки теплового шока (HSP): HSP90 ассоциируются с некоторыми различными внутриклеточными белковыми шаперонами для формирования мультимерных белков. Ингибиторы HSP90 приводят к быстрой убиквитинизации и протеасомальной деградации этих ассоциированных белков, включая внутриклеточные патогены или их субъединицы (Hoffman, Ronald). HSP60 и HSP70: Активные иммунные клетки, такие как макрофаги и дендритные клетки (Kaufmann, 2004, Ch. 13).
(b) Повторяющийся экстрадомен А типа III фибронектина: Активные иммунные клетки через распознавание посредством TLR4.
(c) Низкомолекулярные олигосахариды гиалуроновой кислоты: Активаторы дендритных клеток, также опосредованные TLR4 (Kaufman, 2004, Ch. 13).
(d) Полисахаридные фрагменты сульфата гепарина: Индуцируют созревание дендритных клеток посредством TLR4 (Kaufmann, 2004, Ch. 13).
(e) Фибриноген: Вызывает хемокиновую продукцию в макрофагах через посредство TLR4 (Kaufmann, 2004, Ch. 13).
(f) Липополисахариды (LPS): Наиболее сильный иммуностимулятор среди микробных компонентов (Kaufman, 2004, Ch. 13).
(g) Фосфорилхолин (PC): главная антигенная структура, обнаруженная на грам-позитивных бактериях. PC связывается с природными IgM антителами, а также CRP (С-реактивный белок). PC может поэтому активировать систему комплемента и усиливать врожденный иммунный ответ. Связывание CRP с PC является зависимым от кальция. Связывание PC с природными IgM антителами не зависит от кальция.
(h) Мочевая кислота (UA): Клетки человека, подвергающиеся апоптозу, некрозу или другим формам клеточной смерти, высвобождают множество неспецифических сигналов опасности, один из которых является мочевой кислотой. Мочевая кислота стимулирует созревание дендритных клеток и усиливает реактивность CD8 Т-клеток на антигены. UA является природно встречающимся эндогенным адъювантом. Введение UA с вакциной будет усиливать эффективность вакцины.
(i) IgGI и IgGIII антитела, специфические к любым компонентам вакцины (любой компонент): природные клетки-киллеры (NK) характеризуются Fc рецептором, известным как CD16 или FcγRIII, который специфичен для IgGI и IgGIII. Путем конъюгирования вакцины с этими антителами зависимая от антитела клеточно опосредованная цитотоскичность (ADCC) NK-клеток будет усиливаться (Parham, 2003, Ch. 7). Эти рецепторы также обнаруживаются на дендритных клетках (DC). NK- и
DC-клетки подвергаются процессу совместного праймирования и в случае дендритных клеток созревание происходит в результате этих "перекрестных помех". DC-клетки являются мостиком между врожденной и приобретенной или адаптивной иммунной системами. Поэтому IgGI и IgGIII антитела, добавленные к вакцине, будут опсонизировать вакцину путем усиления ответа NK-клеток и DC-клеток, а также облегчения их кооперации по отношению к потенциальному патогену или вакцине (Ferlazzo, et al.; Cooper, et al.; Chiesa, et al.).
Q) Белки комплемента: Разнообразные белки комплемента, в частности С3b, опсонизирует иммуногены, к которым они прикреплены. Это будет усиливать как врожденный, так и адаптивный иммунный ответ (Hoffman, et al).
Заключение
[0186] Анализ и развитие иммуногенной композиции должен включать широкий диапазон доз инактивированных частиц для определения ценности. Исследования на животных должны рассмотреть различия в размере, видах и иммунологических характеристиках; предполагалось, что иммунологические отличия между человеком и животными могут привести исследования на животных к анализу на токсичность. Клинические исследования будут включать, по меньшей мере, стандартную трехфазную модель, ранжируя по безопасности и дозировке в малой популяции, безопасности и иммуногенности во второй фазе нескольких сотен волонтеров, до широкомасштабной фазы эффективности. Клинические исследования должны включать приемлемые исключающие критерии, которые применяются обычно, такие как исключение по другим условиям иммунной супрессии, беременность, активное применение лекарственных средств и т.п.
[0187] В дополнение к путям введения, описанным в деталях выше, введение может осуществляться различными путями, например перорально, транбуккально, трансмукозально, сублингвально, назально, ректально, вагинально, интраокулярно, внутримышечно, внутрилимфатически, внутривенно, подкожно, чрескожно, внутрикожно, внутрь опухоли, местно, транспульмонарно, с помощью ингаляции, инъекции или имлантации и т.п. Различные формы композиции могут включать, без ограничения, капсулу, гель, таблетку, энтеральную капсулу, инкапсулированную частицу, порошок, суппозиторий, инъекцию, мазь, крем, имплантат, пластырь, жидкость, ингаляцию или спрей, системную, местную или другую пероральную среду, растворы, суспензии, инфузии и т.п. Так как некоторыми из первых мишеней для инфицирования ВИЧ являются эпителиальные клетки и клетки Лангерганса в коже и ректальной и вагинальной слизи, то предпочтительным вариантом осуществления доставки является дермальные в комбинации с ректальными и/или вагинальными суппозиториями. ВИЧ передается преимущественно через ректальный и вагинальный половые контакты. Поэтому введение вакцины в виде ректального и/или вагинального суппозитория будет предпочтительной методикой введения. Кроме того, данное изобретение может быть комбинировано с другими терапевтическими агентами, такими как цитокины, включая природные, рекомбинантные и мутированные формы, фрагменты, белки слияния и другие аналоги и производные цитокинов, смесями, другими биологически активными агентами и вспомогательными веществами для формуляций и т.д. Специалисты в данной области понимают, что для инъекции может быть приемлема формуляция в водных растворах, таких как раствор Рингера или солевой буфер. Липосомы, эмульсии и растворители являются другими примерами носителей для доставки. Пероральное введение должно нуждаться в носителях, приемлемых для капсул, таблеток, жидкостей, пилюль и т.п., таких как сахароза, целлюлоза и т.п.
[0188] Таким образом, в заключение, данное изобретение основано на условно живом вирионе; то есть, вирионе, модифицированном таким образом, что неспособный другим образом к репликации, он становится репликационно компетентным на ограниченное время при добавлении экзогенного белка, который замещает белок, который является недоступным из-за модификации (или делеции) соответствующей генетической последовательности, кодирующей этот белок в вирусном геноме. Вирус по определению не является живой или мертвой структурой. Он лучше всего характеризуется как репликационно компетентный или репликационно некомпетентный. В данном изобретении живой вирус относится к репликационно компетентному вектору. Один аспект данного изобретения представляет собой иммуногенную композицию, включающую вирусную ДНК или РНК, представляющую полный вирусный геном, в котором, по меньшей мере, один ген белка репликации или соответствующая мРНК модифицированы для компенсации неспособности вирусной ДНК или РНК к репликации; эти модифицированные вирусные ДНК или РНК затем инкапсулируются вирусными белками, которые самопроизвольно собираются в бесклеточной системе экспрессии, формируя условно живой вирион. Способ для получения этого условно живого вириона включает этапы обеспечения, по меньшей мере, одной вирусной ДНК или РНК молекулы, представляющей полный геном, амплифицирование вирусной ДНК или РНК, модифицирование вирусной ДНК или РНК в, по меньшей мере, одном гене белка репликации или соответствующей мРНК, отбор амплифицированной и модифицированной вирусной ДНК или РНК, переупаковка отобранной ДНК или РНК в бесклеточной системе экспрессии, пригодной для самосборки вирусных частиц, и отбор желаемого количества полученных условно живых вирионов. Альтернативный способ для получения этого условно живого вириона использует традиционную систему клеточной культуры. В этом способе вирион, модифицированный в, по меньшей мере, одном гене белка репликации или соответствующей мРНК, может быть культивирован в условиях, пригодных для вирусной репликации с добавлением экзогенного белка, соответствующего, по меньшей мере, одному гену белка репликации или соответствующей мРНК. Поэтому четвертый аспект данного изобретения является формулирование вакцины с применением репликационно некомпетентного вириона в комбинации с цельными вирусными белками, белковыми фрагментами, белковыми производными или их комбинациями. Вакцина, созданная любым из способов, будет иметь тройные иммуногенные свойства, которые проявляются в 1) цельном интактном репликационно некомпетентном вирусе; 2) условно живом вирионе, временно оживленном добавлением белковых добавок; и 3) белковой добавкой самой по себе, действующей как субъединичная вакцина. Дополнительной характеристикой вакцины, сформулированной с условно живым вирионом, созданным в бесклеточной системе, является то, что не присутствует ни один вектор, который бы вносил свой вклад в проявление иммуногенного ответа вакцины при ее введении.
[0189] Вышеприведенные примеры должны рассматриваться как примеры вариантов осуществления, и ни в коем случае не ограничивающие данное изобретение. Таким образом, когда описание, приведенное выше, относится к конкретным вариантам осуществления, то необходимо понимать, что модификации могут быть сделаны без отрыва от смысла приведенного.
Литература
Aguzzi A, S. Brandner, U. Sure, et al., Brain Pathology, 1994, 4:3 20.
Bour, Stephan, et al., 2003, The HIV-1 Vpu Protein: A Multifunctional Enhancer Of Viral Particle Release, Microbes And Infections, Vol.5, pp.1029-1039.
Bowien, Botho, et al., 2003, Nucleic Acids Isolation Methods, Ch. 2, pp.7-19; Ch. 5, pp.53-59; Ch. 6, pp.61-80; Ch. 7, pp.81-94.
Busby, S, et al., J. Mol Biol 154:197209 (1982).
Campbell, Mary К., et al., 2006, Biochemistry, 5th Ed., Ch. 12, pp.320-321 Camper, S.A., et al.. Biology of Reproduction, 199552:246257.
Chiesa, Mariella Delia, et al., Pathogen-Induced Private Conversations Between Natural Killer and Dendritic Cells, Trends in Microbiology, Vol. 13, No. 3, March, 2005.
Cimarelli, A. et al., 2002, "Biomedicine And Diseases: Review Assembling The Human Immunodeficiency Virus Type 1" Cellular And Molecular Life Sciences, Vol.59,pp.1166-1184.
Cohen, P. T-. et al., The AIDS Knowledge Base, 3rd Ed., Ch. 16, pp.153.
Cooper, Megan A. et al., "NK Cell and DC Interactions/' Trends in Immunology, Vol.25,Iss. 1,2004, pp.47-52.
Deeks, Steven, The Medical Management of Aids, Ch. 6 (6th ed. 1999) Deng, W.P., J.A.Nickoloff, Analytical Biochemistry, 200:81 88 (1992) Doherty, F. J., et al., 1992, Intracellular Protein Degradation, Ch. 2, pp.9-14.
Ferlazzo, Guido, et al., 2004, NK Cell Compartments and Their Activation by Dendritic Cells, J of.
Flint, S. J., et al., 2000, Principles of Virology, 2nd Ed., App.A, pp.836.
Flint, S. J., et al., 2004, Principals of Virology, 2nd Ed., Ch. 1, pp.6; Ch. 2, pp.51-53.
Ch. 4, pp.83-125; Ch. 7, pp.217-250; Ch. 20, pp.763-768; App.A, pp.835-837.
Gellissen, Gerd, 2005, Production of Recombinant Proteins, Ch. 2, pp.7-37 and Ch. 11, pp.233-251.
Graham, et al., 1973, J. Virol. 33:739748.
Hoffman, Ronald, et al., 2005, Hematology: Basic Principals and Practice, 4th Ed., Ch. 43, pp.720-733; Ch. 55, pp.80-982.
Hout, David R., et al., 2004, Vpu: A Multifunctional Protein that Enhances the Pathogenesis of Human Immunodeficiency Virus Type 1, Current HIV Research, V1. 2, pp.255-277.
Immunology, Dec 1, 2003, pp.1333-1339.
Immunodeficiency Virus Type-1 Tat Protein, Virology, Vol.257, pp.502-510.
Kaufmann, Stefan, H.E., 1997, Host Response to Intracellular Pathogens, Ch. 3, pp.37-45.
Kaufmann, Stefan, H.E., 2004, The Innate Immune Response to Infection, Ch. 13, pp.260.
Klug, William S. et al., 2006, Concepts of Genetics, 8th Ed., Ch. 4, pp.66-99 Krauss, Gerhard, 2003, Biochemistry of Signal Transduction and Regulation, 3rd Ed., Ch. 2, pp.101-113.
Langel, UIo, 2002, Cell-Penetrating Peptides Processes and Applications, Ch. 1-8, pp.1-162.
Langel, UIo1 2007, Handbook of Cell-Penetrating Peptides, 2nd Ed., Ch. 1, pp.5-23.
Lee, et al., Virology 192:380 385 (1993).
Levinson, Warren, Medical Microbiology & Immunology, 2004, Ch. 36, pp.237-243.
McCutchen and Pagano, 1968, J. Natl. Cancer Inst, 41:351 357.
Michael, Nelson, et al., 1998, HIV Protocols, Ch.24, pp.227-230.
Michael, Nelson, et al., 1999, HIV Protocols, Ch. 1, pp.3-10; Ch. 8, pp.51-57; Ch. 9&10, pp.61-81; Ch. 12, pp.89-98; Ch. 17, pp.151-164; Ch. 18, pp.165-196; Ch. 19, pp.185-196; Ch. 24, pp.227-230; Ch. 35, pp.323-327.
Nicholl, Desmond ST., 2002, An Introduction to Genetic Engineering, 2nd Ed., Ch. 2, pp.11-26; Ch. 3, pp.27-41; Ch. 4, pp.43-53; Ch. 7, pp.115-130.
Pansky, Ben,1996, Review of Gross Anatomy, 6th Ed., Ch. 6, pp.497-254 Parham, Peter, 2003, The Immune System, 2nd Ed., Ch. 7, pp.204-205.
Parham, Peter, 2005, The Immune System, 2nd Ed., Ch. 3, pp.67-96; Ch. 8, pp.273-274.
Parrish, John A., et al., 1983, Photo Immunology, Ch.. 6, pp.95-130.
Primate Lentivirus Complete Genomes, http://hiv-web tani.gov/content/hiv-db/HTML V2005cornpendium.html RLP Adams, 1991, DNA Replication, Ch. 4, pp.49-62.
Rubartelli, Anna, et al., "HIV-1 Tat: A Polypeptide for all Seasons", J. Immun. Today, Vol.19, Issue 12, p.545 (1998).
Sambrook, et al., Molecular Cloning: A Laboratory Manual, 2nd Edition, (1989).
Specter, Steven et al., 2000, Clinical Virology Manual, 3rd Ed., Ch. 17, pp.169-179 and Ch. 19, pp.188-197.
Wagner, Edward K., et al., Basic Virology, 1999, Ch. 8, pp.102-108.
Wang, Zhongde, et al., 1999, Activation of Bcl-2 Promoter-Directed Gene Expression by the Human, 1999, Vol.257, pp.502-510.
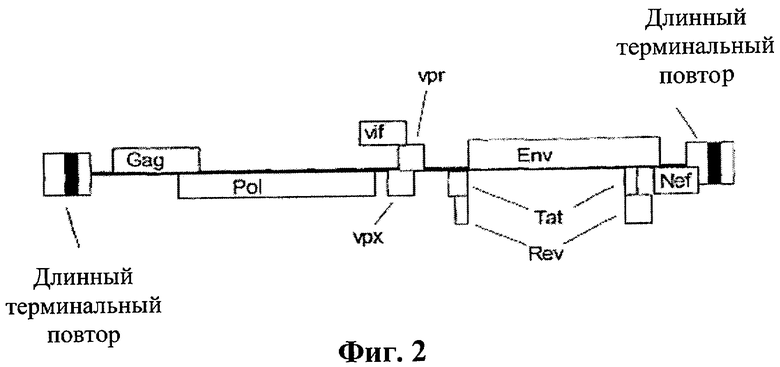
Изобретение относится к биотехнологии. Описан способ получения условно живых вирионов ВИЧ-1, посредством которого вирусную ДНК или РНК модифицируют так, чтобы вирион становился не способным к репликации, если не добавляют белковую добавку к системе экспрессии. Система экспрессии представляет собой либо традиционную клеточную культуру, либо бесклеточную и безвекторную систему экспрессии, пригодную для самосборки вирусных частиц. Обе системы экспрессии нуждаются в добавлении вирусных белков либо для репликации, либо для сборки вириона, не способного к репликации. Условно живой вирион затем используют для создания иммуногенной композиции. Изобретение представляет высокобезопасную иммуногенную композицию, обладающую эффективностью живой вакцины, которая способна вызывать Th-1 опосредованный иммунный ответ. 2 н. и 21 з.п. ф-лы, 2 ил.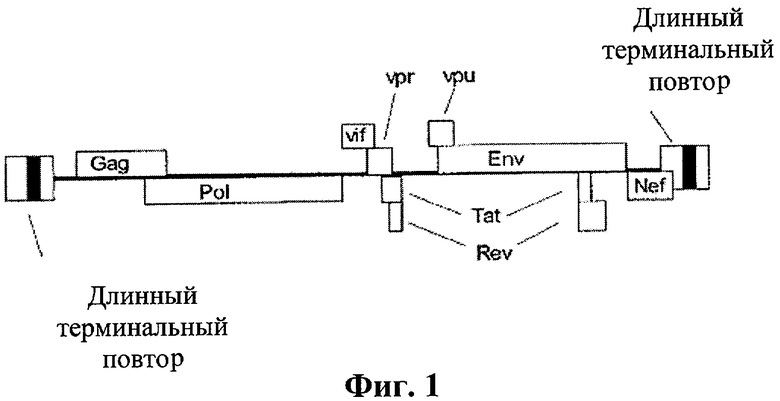
1. Способ получения условно живых вирионов ВИЧ-1, включающий этапы, на которых:
а. очищают и характеризуют ВИЧ-1 изолят, где указанный изолят представляет собой первичный изолят, взятый из биологического образца инфицированного хозяина, где дополнительно указанный изолят характеризуется генотипом;
b. изолируют, амплифицируют и очищают ДНК указанного изолята;
с. модифицируют указанную ДНК указанного изолята, используя технологию делеции гена-мишени для вырезания неперекрывающихся последовательностей, кодирующих ДНК регуляторных генов vif, vpr, vpu и tat экзон 1, где указанные регуляторные гены необходимы для репликации ВИЧ-1 для получения модифицированной ДНК;
d. повторно отжигают модифицированную ДНК для получения последовательности полной длины для вириона, неспособного к репликации;
е. отбирают и очищают указанную модифицированную повторно отожженную ДНК;
f. встраивают указанную модифицированную и повторно отожженную ДНК в бесклеточную систему экспрессии, пригодную для транскрипции-трансляции и сборки вирусных белков, которые кодируются указанной модифицированной и повторно отожженной ДНК в виде условно живых вирионов ВИЧ-1;
g. изолируют, очищают и концентрируют указанные условно живые вирионы ВИЧ-1.
2. Способ по п.1, где указанные условно живые вирионы являются дефицитными по способности к репликации относительно соответствующего вириона, недостаточного по указанной модификации своей вирусной ДНК.
3. Способ по п.1, где в указанном способе дополнительно культивируют указанные условно живые вирионы ВИЧ-1 с ВИЧ-1 белками, которые соответствуют и кодируются указанной вырезанной неперекрывающейся последовательностью ДНК для того, чтобы сделать возможной ограниченную репликацию указанных условно живых вирионов в культуре.
4. Способ по п.3, где в указанном способе дополнительно получают композицию из указанных культивированных условно живых вирионов.
5. Способ по п.4, где указанная композиция составлена в комбинации с фармацевтически приемлемым носителем.
6. Способ по п.5, где указанная композиция составлена в комбинации с, по меньшей мере, одним фармацевтически приемлемым адъювантом.
7. Способ по п.6, где указанный адъювант выбран из группы, состоящей из полисахаридов, включающих, по меньшей мере, одну молекулу маннозы, тейхоевой кислоты, зимозана, полисахаридной капсулы Cryptococcus neoformans серотип С, Протамина, гепариназы, фактора яда кобры в форме, адаптированной для усиления продукции С3b, фактора яда кобры в форме dCVF, никеля в форме, адаптированной для усиления С3 конвертазной активности, сульфатированных полианионов, белков теплового шока, повторяющегося экстрадомена А типа III фибронектина, низкомолекулярных олигосахаридов гиалуроновой кислоты, полисахаридных фрагментов сульфата гепарина, фибриногена, липополисахаридов, фосфорилхолина, мочевой кислоты, антител IgGI и IgGIII, белков комплемента и их комбинаций.
8. Способ по п.1, где указанный биологический образец выбран из группы, включающей семенную ткань, вагинальную ткань и ректальную ткань.
9. Способ по п.1, где указанный биологический образец выбран из группы, включающей семенную жидкость, вагинальную жидкость и ректальную жидкость.
10. Иммуногенная композиция, включающая эффективное количество условно живых вирионов ВИЧ-1, полученных способом по п.1.
11. Иммуногенная композиция по п.10, где указанные условно живые вирионы являются дефицитными по способности к репликации относительно соответствующего вириона, недостаточного по модификации своей вирусной ДНК или РНК.
12. Иммуногенная композиция по п.10, где указанная иммуногенная композиция составлена в виде вакцины, которая способна вызывать Th-1 опосредованный иммунный ответ.
13. Иммуногенная композиция по п.10, где указанная иммуногенная композиция составлена в виде вакцины в комбинации с экзогенным белком (белками), который соответствует указанным вырезанным неперекрывающимся последовательностям ДНК, тем самым способствуя ограниченной репликации указанного условно живого вириона при введении в систему с условиями, приемлемыми для репликации.
14. Иммуногенная композиция по п.10, где указанная иммуногенная композиция составлена как вакцина в комбинации с экзогенным белком(белками), который соответствует указанным вырезанным неперекрывающимся последовательностям ДНК, где экзогенный белок(белки) репликации представляет собой биологически активный фрагмент или производное указанного белка, тем самым способствуя ограниченной репликации условно живого вириона при введении в систему с условиями, приемлемыми для репликации.
15. Иммуногенная композиция по п.10 в комбинации с фармацевтически приемлемым носителем.
16. Иммуногенная композиция по п.10 в комбинации с, по меньшей мере, одним фармацевтически приемлемым адъювантом.
17. Иммуногенная композиция по п.16, где адъювант выбран из группы, состоящей из полисахаридов, включающих, по меньшей мере, одну молекулу маннозы, тейхоевой кислоты, зимозана, полисахаридной капсулы Cryptococcus neofbrmans серотип С, Протамина, гепариназы, фактора яда кобры в форме, адаптированной для усиления продукции С3b, фактора яда кобры в форме dCVF, никеля в форме, адаптированной для усиления С3 конвертазной активности, сульфатированных полианионов, белков теплового шока, повторяющегося экстрадомена А типа III фибронектина, низкомолекулярных олигосахаридов гиалуроновой кислоты, полисахаридных фрагментов сульфата гепарина, фибриногена, липополисахаридов, фосфорилхолина, мочевой кислоты, антител IgGI и IgGIII, белков комплемента и их комбинаций.
18. Иммуногенная композиция по п.10 в комбинации с фармацевтически приемлемым носителем и, по меньшей мере, одним фармацевтически приемлемым адъювантом.
19. Иммуногенная композиция по п.10, где указанная молекула вирусной ДНК или РНК, представляющая вирусный геном, изолирована из ВИЧ-инфицированной ткани.
20. Иммуногенная композиция по п.10, где указанная, по меньшей мере, одна молекула вирусной ДНК или РНК, представляющей вирусный геном, выбрана из группы, включающей ВИЧ-инфицированную семенную, вагинальную и ректальную ткань, и изолирована из интактного хозяина.
21. Иммуногенная композиция по п.10, где указанная, по меньшей мере, одна молекула вирусной ДНК или РНК, представляющей вирусный геном, выбрана из группы, включающей ВИЧ-инфицированную семенную, вагинальную и ректальную жидкость, и изолирована из интактного хозяина.
22. Иммуногенная композиция по п.10, где указанный, по меньшей мере, один ген белка репликации или соответствующая модифицированная мРНК выбраны из последовательностей, выбранных из этих соответствующих белков Vif, Vpr, Vpu, Tat экзон 1, Vpx и их комбинаций.
23. Иммуногенная композиция по п.10, где указанную иммуногенную композицию вводят перорально, трансбукально, трансмукозально, сублингвально, назально, ректально, вагинально, внутриглазным способом, внутримышечно, внутрилимфатически, внутривенно, подкожно, чрескожно, внутрикожно, внутрь опухоли, местно, внутрилегочным способом, с помощью ингаляции, инъекции или имплантации.
| US 20050112143 A1, 26.05.2005 | |||
| RU 2001110116 A, 20.07.2003 | |||
| EA 200300810, 25.12.2003. |

