Настоящее изобретение относится к новому мутантному штамму Е.Coli с супрессированной продукцией ферментативных органических кислот (особенно, уксусной) в процессе аэробного культивирования в среде с добавлением глюкозы. Уменьшение продукции органических кислот данным мутантным штаммом позволяет использовать его в качестве эффективного штамма-хозяина для получения рекомбинантного белка с помощью ферментации при высокой клеточной плотности.
2. Уровень техники
Е. coli широко используется в качестве штамма-хозяина для получения полезных рекомбинантных белков с помощью аэробной ферментации при высокой клеточной плотности. В ростовую среду добавляют значительное количество глюкозы для обеспечения высокой плотности роста клеток-хозяев так же, как и для надлежащей экспрессии рекомбинантного гена, поскольку глюкоза недорога и является легко утилизируемым источником углерода и энергии. Основная проблема при аэробной ферментации с большой плотностью клеток - образование ферментативных кислотных побочных продуктов, из которых наиболее важным является ацетат. Продукция таких побочных соединений, особенно ацетата, представляет собой важный фактор, ограничивающий высокую плотность роста клеток, и соответственно выход рекомбинантного белка (Han et al., Biotechnol. Bioeng., 39, 663 (1992); Luli et al., Appl. Environ. Microbiol., 56, 1004 (1990)).
Для решения указанных проблем используются технологии культивирования "фид-бэтч" (feed-batch) (Fieschko et al., Chem. Eng. Commun., 45 (1986); Ohta et al., J. Ferment. Bioeng., 75, 155 (1993); Yang, Y. Biotechnol., 23, 271 (1992)), методы, основанные на удалении продуцируемых органических кислот из культуры (Landwall et al., Y. Gen Microbiol., 103, 345 (1977); Meyer et al. , Proceedings of the 3rd European Congress on Biotechnology, (1984); MacDonald, Appl. Environ. Microbiol., 56, 640 (1990)) или изменениях состава среды (Holmes, Curr. Topics cell. Regul. 28, 69 (1986); Han et al., Biotechnol. Bioeng., 41, 316 (1993); Reiling, J. Biotechnol. 2, 191 (1985); Mori et al., J. Ferment. Technol., 50, 519 (1972)). Однако указанные методы также имеют ограничения, заключающиеся в том, что медленная скорость роста клеток и пониженная метаболическая активность культур, обусловленная условиями выращивания, часто приводит к снижению выхода рекомбинантного белка, и, кроме того, контроль подачи питательных веществ затруднен и подвержен ошибкам (Chou et al., Biotechnol. Bioeng., 44, 952 (1994)), а соли, добавляемые в культуральную среду для контроля рН, часто ингибируют рост клеток-хозяев и/или образование продукта (Jensen et al. , Biotechnol. Bioeng., 36, 1 (1990)).
При метаболизме глюкозы штаммами Е. coli в аэробных условиях избыток углерода, превышающий возможности ТСА-цикла, конвертируется с образованием уксусной кислоты, которая экскретируется из клетки (Majewski & Domach, Biotechnol. Bioeng. , 35, 732 (1990)). Выделяемая уксусная кислота ингибирует рост штамма-хозяина и образование нужного рекомбинантного белка. Ацетат образуется из ацетилкофермента А в результате последовательного действия фосфотрансацетилазы (pta) и ацетаткиназы (ack). Показано, что мутационная инактивация (путем делеции генов pts и ask) ацетатобразующих ферментов приводит к образованию уменьшенного, но все еще значительного количества ацетата. Мутация также приводит к аккумуляции лактата и пирувата (Diaz-Ricci et al. , Biotechnol. Bioeng., 38, 1318 (1991)) на более высоком уровне по сравнению с родительским штаммом. Эти результаты позволяют предположить, что существует альтернативный путь образования ацетата, а инактивация известных путей биосинтеза ацетата обусловливает образование из углерода другой органической кислоты.
Таким образом, существует необходимость продолжения исследований, направленных на получение нового штамма-хозяина с пониженной продукцией органических кислот и соответственно с повышенным выходом нужных рекомбинантных белков.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Объектом настоящего изобретения является новый штамм Е. coli с супрессированной продукцией органической кислоты, при этом его рост и образование рекомбинантного белка не ингибируются.
Изобретение также относится к новому штамму Е. coli JL 1506 с супрессированной продукцией органических кислот в процессе роста в среде, содержащей глюкозу, в аэробных условиях.
Еще одним объектом изобретения является способ получения белка, который подразумевает использование штамма Е. coli JL 1506 в качестве хозяйской системы экспрессии.
Приведенные выше и другие объекты, характерные особенности и возможные пути осуществления изобретения будут легко поняты специалистами в данной области исследований из приведенного ниже детального описания.
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Новый штамм Е. coli JL 1506 может быть получен следующим образом.
Штамм Е. coli MG 1655 (Guyer et al., Cold Spring Harbor Sym. Quant. Biol. , 45, 135 (1981)), полученный в Центре Исходных культур Е. coli в Йельском Университете, США, подвергают мутагенезу этилметансульфонатом (EMS) в соответствии со стандартной методикой (Miller, Краткое введение в бактериальную генетику. Cold Spring Harbor Laboratory (1992)) и скринируют мутанты, дефектные по путям образования органических кислот в анаэробных условиях. Однако все исследованные мутанты все еще образуют в значительном количестве органические кислоты. Было предварительно установлено, что мутанты, способные к росту в анаэробных условиях, могут продуцировать органические кислоты.
Таким образом, авторы изобретения продолжили работу по скринингу мутантов, которые не способны к росту в анаэробных условиях. Один из выявленных мутантов, обозначенный JL 1031, сохраняет скорость роста родительского штамма, в то время как образование им органической кислоты из глюкозы в аэробных условиях культивирования значительно снижено. Мутацию, обусловившую указанный выше фенотип, переносят в немутантный родительский штамм MG 1655 путем трансдукции с помощью бактериофага Р1, и полученный штамм JL 1506 имеет тот же самый фенотип, что и JL 1031.
Приведенные выше полезные свойства мутантного штамма, полученного согласно настоящему изобретению, позволяют использовать данный штамм в качестве хозяйской системы экспрессии для получения рекомбинантного белка. Тип рекомбинантных белков, которые могут быть получены при использовании штамма Е. coli JL 1506 в качестве хозяйской системы экспрессии, практически не ограничен. Тем не менее указанную систему более целесообразно применять к белкам, чья экспрессия контролируется промоторами, чувствительными к катаболитной репрессии, например промоторами lac, tac и т. д. Однако хозяйская система согласно настоящему изобретению может иметь преимущества для получения и других белков, экспрессия которых находится под контролем промоторов, не чувствительных к катаболитной репрессии, при сравнении ее с другими системами на основе подходящих штаммов Е. coli.
Трансформация Е. coli JL 1506 чужеродным геном, кодирующим рекомбинантный белок, может быть осуществлена любыми методами, хорошо известными для специалиста в данной области исследований. Способы и условия трансформации, которые не ограничиваются настоящим изобретением, могут быть определены специалистом без затруднений в зависимости от типа рекомбинантного белка, размера гена, кодирующего данный белок, селективного маркера и т. д.
Культивирование трансформанта Е. coli JL 1506 может осуществляться в любой подходящей культуральной среде в должных условиях, которые могут быть определены специалистом в данной области исследований.
Новый мутант Е. coli JL 1506 в соответствии с настоящим изобретением может быть получен путем обработки родительского штамма MG 1655 подходящим мутагеном и отбором мутантов с нужными свойствами. Могут быть использованы химические вещества или ультрафиолетовое облучение в качестве мутагенов. Согласно настоящему изобретению для получения мутанта могут использоваться методики генных манипуляций.
Мутация, вызывающая описанный выше фенотип штамма JL 1506, генетически картирована в положении 51-52 min на хромосоме Е. coli.
Отдельная мутация, определяющая фенотип нормального аэробного роста, на утрату способности к росту в анаэробных условиях в минимальной среде с глюкозой ранее не описана и впервые генерирована авторами изобретения.
Авторы изобретения обозначили мутантный генотип как "glf-1031" (дефект ферментации глюкозы) и установили, что мутант JL 1031 имеет генотип glf-1031.
Чтобы предотвратить какие-либо возможные мутации (мутацию) в генах, отличных от glf, авторы изобретения трансдуцировали генотип glf-1031 в MG 1655 с помощью фага Р1 с получением Е. coli JL 1506.
Настоящее изобретение более детально описано в примерах, при этом в примерах 1-3 описывается выделение Е. coli JL 1031 и конструирование JL 1506, в примерах 4 и 6 - катаболитная репрессия глюкозой в штаммах JL 1506 и MG 1655.
Далее, с целью исследования возможности использования мутанта Е. coli JL 1506 как хозяйской системы для экспрессии рекомбинантных генов, получают штаммы JL 1506/рМКТ2-1 и MG 1655/рМКТ2-1 при трансформации плазмидой рМКТ2-1 (Min et al. , Korean Biochem. J., Nucleic Acid Res., 16, 5075 (1988)), несущей нативный ген бета-галактозидазы (lac Z), штаммов JL 1506 и MG 1655 соответственно (примеры 7 и 8).
В примерах 9 и 10 оценивается экспрессия белка при использовании Е. coli JL 1506 в качестве хозяйской системы, при этом культивируют трансформанты Е. coli JL 1506, MG 1655 или С 600, содержащие плазмиду pHTl (Huh et al., Korean Biochim. J., 23, 459 (1990)), включающую кДНК липокортина человека под контролем промотора try (Amann et al., Gene, 40, 183 (1985)).
Когда бета-галактозидаза или липокортин продуцируется трансформантами, выход белка примерно в два раза выше у штамма JL 1506 по сравнению с контрольными штаммами дикого типа MG 1655 или С 600.
При осуществлении настоящего изобретения используются следующие эксперименты.
1) Генетические эксперименты, такие как трансдукция, конъюгация и генное картирование, которые проводятся согласно методикам Миллера (Миллер, Краткий курс бактериальной генетики, Колд Спринг Харбор (1992)).
2) Активность бета-галактозидазы определяют по методу Мина (Min et al., Nucleic Acid Res. , 16, 5075 (1988)). Так, штамм, несущий ген, кодирующий бета-галактозидазу, культивируют в среде LB, содержащей 40 мкг/мл ампициллина, при 37oС и 200 rpm в течение 12-16 ч. В процессе культивирования добавляют 1 мМ IPTG (изопропил-1-тио-бета-Д-галактопиранозида) для индукции экспрессии lac Z. Образцы культурального бульона отбирают через определенные интервалы и для дезинтеграции клеток добавляют Z-буфер, хлороформ и 0.1% SDS. Дезинтегрированные клетки оставляют на водяной бане при 28oС в течение 5 мин, затем для остановки реакции добавляют ONPG. После центрифугирования реакционной смеси измеряют поглощение супернатанта при 240 нм с помощью спектрофотометра (Pharmacia LKB-Ultraspec III).
3) Активность бета-лактомазы измеряют иодиновым методом (Нап et al., Biotechnol., Bioeng., 39, 653 (1992)). Так, клетки из культурального бульона (5 мл) суспендируют в отмывающем буфере (0.05 М Na2HPO4, 0.05 М КН2РO4, 0.014 М NaCl, 0.01 М H2SO4 в 0.1 М фосфатном буферном растворе (PBS) (5 мл) и центрифугируют. Преципитировавшие клетки суспендируют в 0.1 М PBS (5 мл) и разрушают в ультразвуковом гомогенизаторе US-150T в течение 5 мин. Затем материал центрифугируют при 4oС и 15000 rpm для получения клеточного гомогената.
В пробирку для тестирования (А) добавляют 20 мМ бензилпенициллина (0.25 мл), 0.1 М PBS (1.25 мл) и клеточного гомогената и полученную смесь выдерживают при 30oС с течение 20 мин. В контрольную пробирку (В) вносят 0.1 М PBS (2 мл). В тестируемые пробирки добавляют иодин (2.5 мл), оставляют их на 10 мин и измеряют поглощение при 490 нм.
4) Липокортин идентифицируют электрофорезом в полиакриламидном геле в присутствии SDS как полосу размером 37 кДа (Davis, Ann. Rev. NY Acad. Sci., 121, 404 (1964)). Для количественного определения используют создающий изображение денситометр (Bio-Rad GS-670).
4-1) Количественное определение липокортина, продуцируемого клеткой. Культуральный бульон после аэробного культивирования в течение 12 ч вносят в среду LB (20 мл), содержащую 1% глюкозы, до концентрации 2%, так что поглощение при 600 нм составляет 0.1, и продолжают культивирование в течение 10 ч. Культуральный бульон (1 мл) смешивают с 0.5 мл раствора SDS (дистиллированная вода, 4.0 мл; 0.5 М Трис-Cl, 1.0 мл; глицерол, 0.8 мл; SDS, 10%; β-меркаптоэтанол, 0.4 мл; 0.1% бромфенолового синего, рН 6.8) и нагревают при 95oС в течение 5 мин. Двадцать микролитров реакционной смеси отбирают и подвергают электрофорезу в полиакриламидном геле в присутствии 10% SDS (SDS-PAGE).
4-2) Количественное выявление липокортина среди растворимых белков. Культивирование осуществляют в соответствии с указанной выше методикой (см. стадию 4-1). Культуральный бульон (3 мл) отбирают через 6 и 10 ч и центрифугируют для сбора клеток. Клетки суспендируют в Трис-буфере (1 мл), содержащем 5 мМ ЭДТА, и разрушают в ультразвуковом гомогенизаторе. Супернатант используют как фракцию растворимых белков. К 20 мкл супернатанта добавляют эквивалентное количество SDS, и полученную смесь обрабатывают, как описано на стадии 4-1. Количественное содержание липокортина определяют методом SDS-PAGE в присутствии 10% SDS.
Согласно настоящему изобретению получен новый мутант Е. coli JL 1506, не способный утилизировать глюкозу в анаэробных условиях и обладающий супрессированной продукцией органической кислоты при культивировании с высокой плотностью клеток в аэробных условиях. Мутант может быть эффективно использован в качестве полезной хозяйской системы для получения рекомбинантного белка. Кроме того, он может быть использован для получения новых мутантов путем блокирования генов, вовлеченных в образование органической кислоты.
Примеры предпочтительных вариантов осуществления изобретения
Настоящее изобретение может быть осуществлено, как ниже указано в примерах. Однако данные примеры приведены только для иллюстрации изобретения и не ограничивают его объема, который охватывается формулой изобретения. В примерах используются весовые доли и проценты, если специально не оговорено иное.
Пример 1
Штамм Е. coli MG-1655 дикого типа обрабатывают EMS. Мутантные клетки помещают на чашки с агаровой средой Лурия-Бертрани (LB: триптон, 10 г; дрожжевой экстракт, 5 г; NaCl, 5 г; дистиллированная вода, 1 л) и инкубируют при 37oС до образования отдельных колоний. Колонии переносят методом реплик на два набора чашек с глюкозой (конечная концентрация - 1%) и минимальной средой М 56 (КН2РO4, 10.6; Na2HPO4, 17.4 г; 10% MgSO4•7H2O, 17.4 г; 1% (NH4)2SO4, 2мл; 0.05% FeSO4•7H2O, 2 мл; дистиллированная вода, 1 л, рН 7.0). Один набор чашек инкубируют в аэробных условиях, а другой в анаэробных условиях в специальном устройстве (BBL) при 37oС. Мутантный штамм Е. coli JL 1031 идентифицируют как колонию, обнаруживающую нормальный рост в аэробных условиях, но не способную к росту в анаэробных условиях даже после 48 ч.
Пример 2
Для локализации позитивного маркера в непосредственной близости от мутации (экспериментально обозначенной glf-1031, см. пример 4 для более подробного объяснения), обусловливающей неспособность JL 1031 к анаэробному росту (для описания этого свойства мутанта используется аббревиатура Glf), вблизи мутации glf-1031 встраивают транспозон Тn 10. Для этого MG 1655 (Glf+) инфицируют фагом лямбда NK 561 (Foster, Cell, 23, 215 (1981)) и помещают на чашки со средой LB/Tet (15 мкг/мл). Полученную популяцию колоний, устойчивых к тетрациклину (Тс-r), используют для размножения фага Plvir. Штамм JL 1031 инфицируют лизатом Р1 и помещают на чашки со средой LB/Tet для получения Тс-r трансдуктантов. Методом реплик трансдуктанты переносят на чашки с минимальной средой с глюкозой и отбирают колонию, способную к росту на этой среде в анаэробных условиях (Glf+). P1 лизат, полученный при использовании трансдуктантов Glf, применяют для трансдукции JL 1031 (Glf+) с целью придания ему Тс-r. Среди исследованного 61 Тс-r трансдуктанта 16 оказались неспособными к росту в анаэробных условиях в минимальной среде с глюкозой. Полученные результаты показывают, что транспозон Тn 10 (ZZZ-1051::Tn 10) расположен около glf-1031, и 26.2% (16/61) мутантов котрансдуцибельны. Один из оставшихся 45 трансдуктантов Glf-ZZZ-1031::Tn 10 выбирают и обозначают как JL 1501 (glf-1031, ZZZ-1051::Tn 10).
Пример 3
Мутацию glf-1031 повторно трансдуцируют в MG-1655, используя JL 1501 в качестве донора, выбирают случайным образом один из полученных трансдуктантов Glf-ZZZ-1051::Tn 10, обозначают JL 1506 (glf-1031, ZZZ-1051::Tn 10) и используют для дальнейшего описания, как указано ниже. Штамм Е. coli JL 1506 депонирован в соответствии с Будапештским договором о депонировании микроорганизмов для целей патентной процедуры 19 октября в Корейской коллекции типовых культур, находящейся в GERI, KIST, PO Box 115, Юсон Теджон, под регистрационным номером КСТС 0204ВР.
Hfr-конъюгация и P1-трансдукционный анализ показывают, что ZZZ-1051::Tn 10 котрансдуцибельна с nup С-3146::Тп 10, локализованной в положении 51.75 min на хромосоме Е. coli. Частота реверсии glf-1031, определенная как частота реверсии на минимальной среде с глюкозой в анаэробных условиях, составляет 8•10-3.
Пример 4
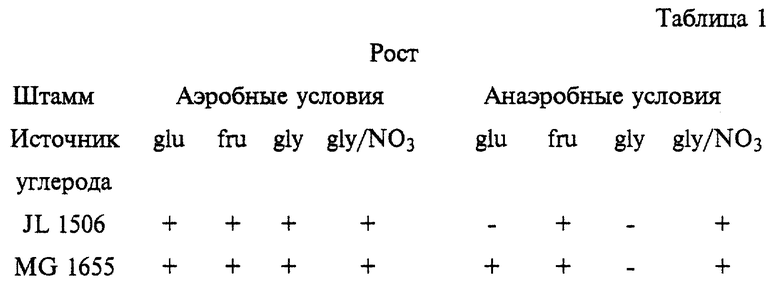
Штамм JL 1506 и родительский штамм MG 1655 тестируют на их способность расти на среде М56 с добавлением различных источников углерода до конечной концентрации 1% как в аэробных, так и в анаэробных условиях. Для этого указанные штаммы помещают на чашки с LB и инкубируют в аэробных условиях при 37oС в течение ночи. Культуры после ночного выращивания методом реплик переносят на двойной набор чашек с подходящей минимальной средой с добавлением сахара. Один набор чашек инкубируют в аэробных условиях, а второй - в анаэробных условиях в специальном устройстве (BBL). Определяют рост аэробных культур через 18 ч и анаэробных через 48 ч. Способность указанных штаммов утилизировать различные источники углерода в аэробных и анаэробных условиях отражена в таблице 1.
Мутантный штамм JL 1506 не способен к росту на среде с глюкозой в анаэробных условиях, в то время как родительский штамм MG 1655 в этих условиях проявляет нормальную способность к росту. Однако мутантный штамм растет нормально в анаэробных условиях на минимальной среде с добавлением глицерола/NО3 - (20 мМ). Приведенные результаты свидетельствуют о том, что мутант JL 1506 не дефектен по анаэробному дыхательному пути, но может быть дефектен по пути ферментации глюкозы. По этим причинам мутантный локус в JL 1506 произвольно обозначен glf (от англ. Glucose fermentation defective - дефектный по ферментации глюкозы).
Пример 5
Сравнивают способность мутантного штамма JL 1506 и контрольного штамма MG 1655 продуцировать органические кислоты в процессе аэробного роста в среде с глюкозой. Культуры каждого штамма после ночного выращивания в аэробных условиях на среде LB переносят во флаконы объемом 250 мл, содержащие 50 мл среды М56 с глюкозой (конечная концентрация глюкозы 0.5%), при первоначальной оптической плотности 0.01 при 600 нм. Культуры инкубируют в ротационном шейкере при 37oС и 200 rpm. Аликвоты культур собирают каждые 3 ч. С помощью HPLC определяют поглощение при 600 нм для оценки продукции органических кислот и ростовых характеристик. Для анализа органических кислот 1 мл культурального образца центрифугируют повторно после смешивания с эквивалентным объемом 5 мМ H2SО4. Полученную надосадочную жидкость фильтруют через мембранный фильтр (Gelman, 0.2 мкм) и хранят при -20oС до анализа. 10 мкл фильтрата подвергают HPLC (Spectra Physics Со.) на колонке Aminex HPX-87H (300•7.8 мм; Bio-Rad) для количественного определения органических кислот. В качестве элюента используют 0.01 М H2SО4 при скорости потока 0.4 мл/мин. Используют УФ-детектор (длина волны - 210 нм).
При сравнении максимальной концентрации каждой органической кислоты, продуцируемой каждой культурой, получают следующие результаты: JL 1506 продуцирует 0.3 мМ уксусной кислоты, 0.8 мМ муравьиной кислоты и 0.3 мМ молочной кислоты, в то время как MG 1655 продуцирует 20.6 мМ уксусной кислоты, 14.9 мМ муравьиной кислоты и 0.4 мМ молочной кислоты. Таким образом, у мутанта JL 1506, полученного согласно изобретению, продукция органической кислоты существенным образом супрессирована. Особенно важно, что он продуцирует только 1% уксусной кислоты по сравнению с контрольным штаммом дикого типа.
Пример 6
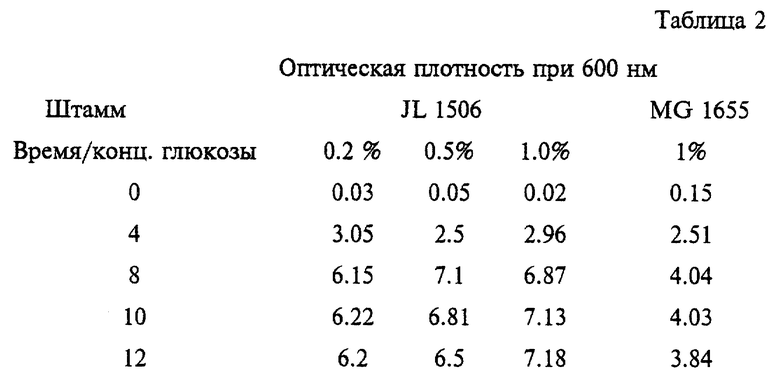
Проводят сравнительную оценку влияния глюкозы на рост штаммов JL 1506 и MG 1655 в среде LB с различными концентрациями глюкозы. Культуры после ночного выращивания в среде LB в аэробных условиях переносят во флаконы объемом 125 мл, содержащие 10 мл среды LB с добавлением глюкозы до конечной концентрации 0.2%, 0.5% и 1%. Культуры инкубируют в ротационном шейкере при 37oС и 200 rpm. Для определения способности к росту указанных штаммов каждый час измеряют поглощение культурального бульона при 600 нм. Результаты исследований приведены в таблице 2.
Как можно видеть из таблицы 2, штамм JL 1506 обнаруживает повышенную способность к росту при увеличении концентрации глюкозы, в то время как штамм MG 1655 обнаруживает пониженную способность к росту с 8-ого часа культивирования за счет накопления образуемых им органических кислот.
Более того, степень максимального роста JL 1506 (поглощение 7.18) почти в два раза (поглощение 4.04) превышает таковую родительского штамма.
Пример 7
Для тестирования возможности штамма JL 1506 служить хозяйской системой экспрессии рекомбинантного белка определяют продуктивность плазмид, кодирующих белки, в штаммах JL 1506 и MG 1655. Плазмиду рМКТ2-1 (Min et al., Nucleic Acid Res., 16, 5075 (1988)), содержащую гены β-галактозидазы (lacZ) и β-лактамазы (bla), переносят в штаммы JL 1506 и MG 1655. Полученные трансформанты JL 1506/рМКТ2-1 и MG1655/pMK2-1 выращивают в 250 мл флаконах, содержащих 10 мл среды LB с глюкозой (0.5%) при 37oС и 200 rpm. Определяют поглощение при 600 нм для оценки клеточного роста, активности ферментов β-галактозидазы и β-лактамазы и стабильности плазмид.
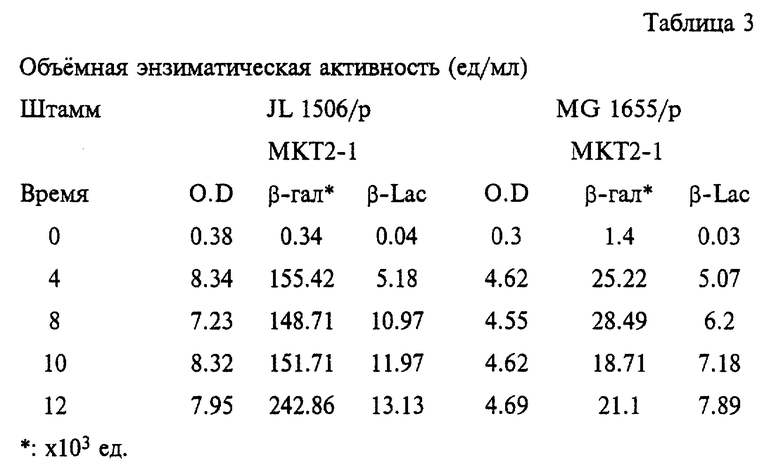
В результате исследований устанавливают, что штамм JL1506/pMKT2-l, который получен из штамма JL 1506 согласно настоящему изобретению, сохраняет плазмиду стабильной (на уровне 100%) после субкультивирования в течение 18 генераций.
Профили ростовой и энзиматической активностей приведены в таблице 3.
Конечная O. D. (12 ч) культуры JL 1506/p MKT2-1 составляет 7.95, в то время как культуры MG 1655/p MKP2-1 - 4.69. Согласно данным O.D. выход клеток JL 1506/p MKT2-1 примерно на 70% больше по сравнению с контрольным штаммом дикого типа, MG 1655/p MKP2-1. Что касается продукции β-галактозидазы, то у заявленного мутантного штамма общая (объемная) активность данного фермента в 11.5 раз выше по сравнению с таковой контрольного штамма дикого типа. Следовательно, мутантный штамм обладает примерно 6, 8 - кратной специфической активностью (ед. активности/О.D.) по сравнению с контрольной. В случае β-лактамазы, кодируемой той же самой плазмидой рМКТР2-1, общая активность мутантного штамма на 66% превышает таковую контрольного. Однако не наблюдается различий в специфической активности между мутантным штаммом и контрольным штаммом дикого типа.
Приведенные результаты свидетельствуют о том, что полученный согласно настоящему изобретению штамм JL 1506 может быть чрезвычайно полезен при использовании в качестве штамма-хозяина для продукции рекомбинантного белка с помощью плазмид, содержащих соответствующие гены.
Пример 8
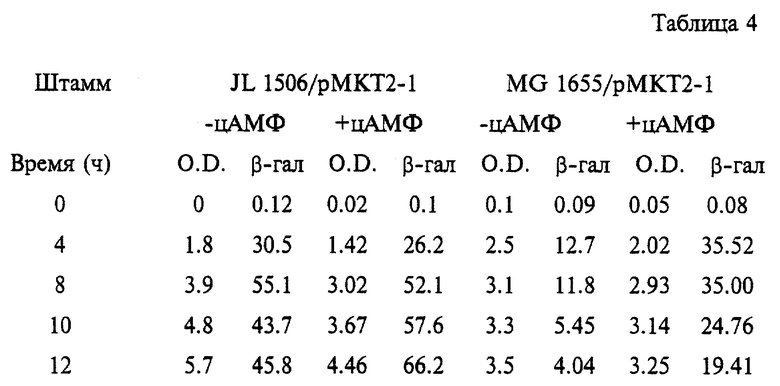
Для определения степени катаболитной репрессии бета-галактозидазы в штамме JL 1506 при росте на среде с глюкозой выявляют его β-галактозидазную и ростовую активность на среде LB/глюкоза с добавлением или в отсутствии цАМФ. Для сравнения штамм MG 1655 выращивают в сходных условиях. Указанные штаммы переносят в 250 мл флаконы, содержащие 20 мл среды LB/глюкоза (0.1%) без или с добавлением цАМФ (конечная концентрация 5 мМ). Культуры инкубируют в ротационном шейкере при 37oС и 200 rpm. Определяют ростовую и бета-галактозидазную активность. Результаты приведены в таблице 4.
Штамм дикого типа MG 1655 обнаруживает значительную степень репрессии β-галактозидазы в отсутствие цАМФ и указанный эффект усиливается по мере старения клеток. Добавление цАМФ корригирует эффект катаболитной репрессии, опосредуемый глюкозой. Однако полученный согласно настоящему изобретению мутантный штамм JL 1506 не обнаруживает эффекта катаболитной репрессии и его энзиматическая активность в присутствии цАМФ намного выше таковой MG 1655. Приведенные результаты показывают, что заявленный мутантный штамм JL 1506 отличается чрезвычайно низкой катаболитной репрессией посредством содержащейся в среде глюкозы при экспрессии соответствующих генов, входящих в состав плазмиды.
Пример 9
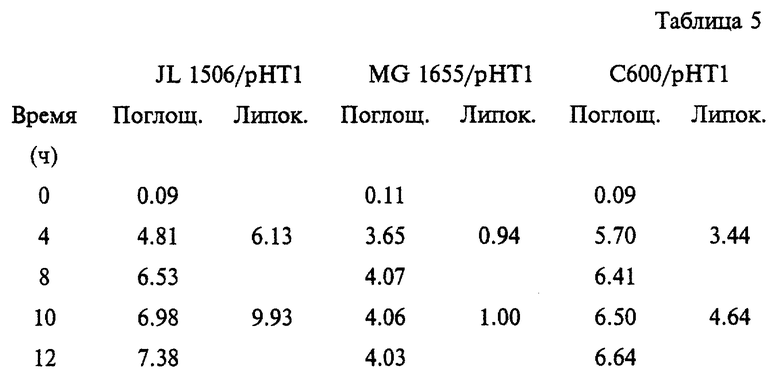
Оценивают и сравнивают выход гетерологичного рекомбинантного белка у культур штаммов JL 1506, MG 1655 и С600 (последние два используются в качестве контроля). Плазмиду pHT1 (Huh et al., Korean Biochem. J., 23 459 (1990)), содержащую кДНК липокортина человека, переносят путем трансформации в заявленный штамм JL 1506, родительский штамм дикого типа MG 1655 и хозяйский штамм С600, который, как отмечено в указанной выше работе, обладает сверхвысокой продукцией белка липокортина. Каждый из указанных трансформантов (JL 1506/pHT1, MG 1655/pHT1 и С600/pHT1) вносят в 250 мл флаконы, содержащие 20 мл среды LB/глюкоза (конечная концентрация 0.1%), и инкубируют в ротационном шейкере при 37oС и 200 rpm. В соответствующие моменты времени культуральный бульон собирают и определяют поглощение при 600 нм для оценки роста. Образцы культур анализируют на количественное содержание общего липокортина в общей белковой фракции. Для этого 1 мл культурального бульона смешивают с 0.5 мл SDS-буфера для образцов [(0.5 М Трис-HCl, рН 6.8, ~1.0 мл; 10% (вес.объем) глицерола, 0.8 мл; 10% (вес/объем) SDS; β-меркаптоэтанол, 0.4 мл; бромфеноловый синий, 0.1% (вес/объем); дистиллированная вода, 4.0 мл)] и нагревают в течение 5 мин при 95oС. 20 мкл образцов исследуют методом PAGE в присутствии 10% SDS. После электрофореза гель окрашивают Кумасси ярко-синим и сканируют на денситометре, создающем изображение (Bio-Rad Model GS-670). Полосу липокортина идентифицируют по известному молекулярному весу (37 кДа) и относительные объемы полос (плотность • область полосы) сравнивают в таблице 5.
Выход клеток мутантного штамма JL 1506, полученного согласно настоящему изобретению, при культивировании в аэробных условиях на среде LB с глюкозой наиболее высокий среди всех тестируемых штаммов. Он также обладает наибольшей продуктивностью липокортина среди всех тестируемых штаммов. В 10-часовом культуральном образце штамма JL 1506/pHT1 липокортина в 9.93 раза больше, чем в 10-часовом образце штамма MG 1655/pHT1. Количество липокортина, продуцируемое заявленным мутантным штаммом, почти в два раза больше такового С600/pHT1.
Пример 10
Определяют и сравнивают количество растворимого липокортина в эквивалентных растворимых клеточных фракциях штамма JL 1506 и двух других контрольных штаммов. Используют штаммы и условия культивирования, как описано в примере 9. Собирают образцы культурального бульона (3 мл) через 6 ч и 10 ч культивирования и центрифугируют их при комнатной температуре для сбора клеток. Осадок клеток ресуспендируют в 1 мл буфера Трис-НС1 с ЭДТА (конечная концентрация 5 мМ). После разрушения клеток в ультразвуковом дезинтеграторе и сбора супернатанта в результате центрифугирования при 4oС получают фракции растворимого клеточного белка. Концентрацию белка в них определяют методом Брэдфорда (Bradford, Anal. Biochem., 72, 248 (1976)). Супернатант смешивают с SDS-буфером для образцов, и эквивалентное количество растворимого белка подвергают PAGE в присутствии 10% SDS. Относительную продуктивность по отношению к липокортину определяют, как описано в примере 9. Результаты исследований приведены в таблице 6.
Объем полосы липокортина (плотность x область полосы) MG 1655/рНТ1 через 10 ч культивирования принимают за 1 при сравнительных вычислениях.
На основании содержания растворимого липокортина в расчете на единицу количества растворимых клеточных белков (специфическая продуктивность) установлено, что штамм JL 1506/рНТ1 обладает наивысшей способностью к продукции растворимого липокортина среди всех исследованных штаммов. Он продуцирует липокортина в 4.64 раза больше, чем штамм MG 1655/pHT1. С 600 продуцирует больше растворимого липокортина через 6 ч, чем через 10 ч. JL 1506/pHT1 продуцирует в 2.83 раза (4.64/1.64) больше липокортина, чем 6-часовая культура С600/pHT1. Указанные результаты свидетельствуют, что заявленный штамм JL 1506 может служить эффективным штаммом-хозяином для продукции липокортина.
| название | год | авторы | номер документа |
|---|---|---|---|
| МИКРООРГАНИЗМ С ВЫКЛЮЧЕННЫМ ГЕНОМ fadR НА ХРОМОСОМЕ И СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА ПОСРЕДСТВОМ ФЕРМЕНТАЦИИ С ДАННЫМ МУТАНТОМ | 2003 |
|
RU2305132C2 |
| ЭПИТОП ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА B И ЕГО ПРИМЕНЕНИЕ | 2011 |
|
RU2585525C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pT7ΔpckA::loxpcat ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ L-ТРЕОНИНА В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli FTR2717 (KCCM-10475) - ПРОДУЦЕНТ L-ТРЕОНИНА | 2004 |
|
RU2339699C2 |
| Новый мутантный белок орнитиндекарбоксилаза и его применение | 2014 |
|
RU2669996C2 |
| МИКРООРГАНИЗМ, СПОСОБНЫЙ ПРОДУЦИРОВАТЬ L- АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ПРИМЕНЕНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2013 |
|
RU2600875C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 2002 |
|
RU2288265C2 |
| МИКРООРГАНИЗМ, ПРОДУЦИРУЮЩИЙ О-ФОСФОСЕРИН, И СПОСОБ ПОЛУЧЕНИЯ L-ЦИСТЕИНА ИЛИ ЕГО ПРОИЗВОДНЫХ ИЗ О-ФОСФОСЕРИНА С ЕГО ИСПОЛЬЗОВАНИЕМ | 2011 |
|
RU2536250C1 |
| РЕАГЕНТ ДЛЯ ДИАГНОСТИРОВАНИЯ ИНФЕКЦИИ, ВЫЗВАННОЙ ВИРУСОМ PUUMALA | 1998 |
|
RU2218571C2 |
| НОВАЯ О-АЦЕТИЛГОМОСЕРИН-СУЛЬФГИДРИЛАЗА ИЛИ ЕЕ МУТАНТНЫЙ БЕЛОК И СПОСОБ ПРЕОБРАЗОВАНИЯ В МЕТИОНИН С ИСПОЛЬЗОВАНИЕМ ТАКОВОЙ | 2011 |
|
RU2573928C2 |
| МОДИФИЦИРОВАННЫЙ ПОЛИПЕПТИД, ОБДАДАЮЩИЙ ГОМОСЕРИНАЦЕТИЛТРАНСФЕРАЗНОЙ АКТИВНОСТЬЮ, И ЭКСПРЕССИРУЮЩИЙ ЕГО МИКРООРГАНИЗМ | 2011 |
|
RU2557411C2 |

Изобретение относится к биотехнологии и может быть использовано для получения рекомбинантных белков. Мутант E.coli не способен к росту в анаэробных условиях, утилизирует глюкозу, обладает супрессированной продукцией органической кислоты в аэробных условиях культивирования. Использование мутанта в качестве хозяйской системы позволяет повысить выход рекомбинантных белков при культивировании. 2 с. и 1 з.п. ф-лы, 6 табл.
| US 5028539 A, 02.07.1991 | |||
| 1971 |
|
SU411501A1 | |

