ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Область изобретения
Настоящее изобретение относится к продуцирующему L-треонин микроорганизму и способу получения L-треонина посредством ферментации с данным микроорганизмом.
Более конкретно, изобретение относится к мутантному микроорганизму со способностью к высокой продукции L-треонина, у которого расположенный на хромосоме микроорганизма ген fadR посредством применения системы сайт-специфической гомологичной рекомбинации, подобно гену cre и участку loxp, выключен так, что экспрессия оперона aceBAK в течение биосинтеза L-треонина частично не репрессирована, и к способу получения L-треонина посредством ферментации с данным мутантом.
Описание связанной области
L-треонин представляет собой разновидность незаменимых аминокислот и его широко применяют в качестве пищевых и кормовых добавок, а также как фармацевтическое и сырьевое вещество для синтеза некоторых лекарственных средств. Его получают посредством ферментации с искусственными мутантами родов Escherichia, Coryneform bacteria, Seratia и Providencia. Например, в публикации патента Японии № S56-10037 описан способ получения L-треонина с применением принадлежащего роду Escherichia штамма, требующего для роста диаминопимелиновую кислоту и метионин и обладающего в системе биосинтеза треонина устойчивостью к ингибированию треонином по принципу обратной связи. В выложенной заявке на выдачу патента Японии № S58-224684 описан способ получения L-треонина с применением принадлежащего роду Brevibacterium штамма, резистентного к S-(2-аминоэтил)-L-цистеину and α-амино-β-гидроксивалериановой кислоте и требует для роста L-изолейцин и L-лизин. В выложенной заявке на выдачу патента Кореи №87-8022 описан способ получения L-треонина с применением принадлежащего роду Escherichia штамма, с дополнительной устойчивостью, по меньшей мере, к одному веществу, выбранному из группы, состоящей из рифампицина, лизина, метионина, аспарагиновой кислоты и гомосерина, и со сниженной способностью разлагать L-треонин. В выложенной заявке на выдачу патента Японии № H2-219582 описан способ получения L-треонина с применением принадлежащего роду Providencia штамма, устойчивого к α-амино-β-гидроксивалериановой кислоте, L-этионину, тиоизолейцину, окситиамину и сульфагуанидину и требующего для роста L-лейцин, а также с низкой потребностью в L-изолейцине.
Однако указанные выше известные способы обладают теми недостатками, что они не в состоянии обеспечить высокий уровень продукции L-треонинина или что для роста необходимо добавлять дорогие вещества, такие как диаминопимелиновая кислота и изолейцин. Другими словами, применение нуждающихся в диаминопимелиновой кислоте штаммов включает в себя дополнительную ферментацию с диаминопимелиновой кислотой и, таким образом, может увеличивать расходы при получении L-треонинина. В том случае, когда для получения L-треонина применяют штамм, требующий для роста изолейцин, в среду для ферментации необходимо добавлять дорогостоящий изолейцин, что увеличивает расходы.
С целью преодолеть данные недостатки, авторы настоящего изобретения разработали продуцирующий L-треонин штамм Escherichia coli, устойчивый к аналогам L-метионина, L-треонина и L-лизина и к α-аминомаслянной кислоте и требующий для роста метионин и обладающий небольшой потребностью в изолейцине и успешно продуцирующий посредством ферментации L-треонин с более высоким чем предыдущие штаммы выходом. На штамм и способ получения L-треонина с применением указанного штамма выдан патент посредством публикации патента Кореи № 92-8365.
Авторы настоящего изобретения провели исследования для получения штамма со способностью продуцировать L-треонин с более высоким выходом и сосредоточили усилия на гене fadR, регуляторном факторе для экспрессии оперона, ассоциированного с разрушением жирных кислот (оперон fad), и оперона глиоксилатного шунта (оперон aceBAK). Известно, что белок fadR репрессирует экспрессию оперона fad и оперона aceBAK, когда концентрация жирных кислот в клетке высокая, но он увеличивает экспрессию связанного с биосинтезом жирных кислот оперона, когда концентрация жирных кислот в клетке низкая (J.E. Cronan Jr. and D. Lapore, E.coli and Salmonella, vol 1, pp. 211-214). Оперон aceBAK обеспечивает синтез оксалоацетата из пирувата, включающий в себя преобразование изоцитрата, промежуточного соединения цикла лимонной кислоты (цикл TCA), в глиоксилат, глиоксилата в малат, а малата в оксалоацетат. Тем временем цикл лимонной кислоты обеспечивает биосинтез оксалоацетата из пирувата посредством восьми (8) путей, а в ходе процесса, промежуточные соединения преобразуются в другие метаболические промежуточные соединения, включающие в себя диоксид углерода. В данном контексте, если экспрессия оперона aceBAK снижена, тогда ингибирован глиоксилатный цикл, увеличивая скорость преобразования углеродного источника в другие метаболические промежуточные соединения через цикл лимонной кислоты, что приводит к сниженной продукции L-треонина в сравнении с теми же самыми количествами источника углерода.
Следовательно, для получения штамма, способного к образованию высокой концентрации L-треонина посредством увеличения экспрессии aceBAK, необходимо выключать ген fadR.
Кроме того, в том случае, когда антибиотик-маркер интегрирован в хромосому, данный маркер нельзя применять снова для выключения другого гена. Таким образом, необходимо удалить маркер из хромосомы, и для выключения гена fadR авторы настоящего изобретения применяют систему сайт-специфической рекомбинации cre/loxp.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения разработали способный к высокой продукции L-треонина штамм посредством выключения гена fadR на его хромосоме и при ферментации с указанным штаммом успешно получили значительно увеличенный выход L-треонина.
В первом аспекте настоящее изобретение относится к продуцирующему L-треонин микроорганизму, у которого выключен расположенный на хромосоме микроорганизма ген fadR.
Во втором аспекте настоящее изобретение относится к способу получения продуцирующего L-треонин микроорганизма, включающему в себя конструирование кассеты для выключения гена fadR или его фрагмента, введение конструкции в исходный продуцирующий L-треонин микроорганизм так, чтобы произошла рекомбинация между чужеродной конструкцией и геном fadR, расположенным на хромосоме микроорганизма, и отбор мутантного микроорганизма, в котором ген fadR выключен.
В третьем аспекте настоящее изобретение относится к способу получения L-треонина посредством ферментации, включающей в себя культивирование микроорганизма, у которого выключен расположенный на хромосоме микроорганизма ген fadR, и очистку L-треонина из культуры.
В четвертом аспекте настоящее изобретение относится к кассете для выключения гена fadR или его фрагмента, сконструированной посредством вставки фланкированного на обоих концах участком loxp антибиотика-маркера в ген fadR, где участок loxp применяют для удаления антибиотика-маркера из хромосомы мутанта с произошедшей интеграцией посредством экспрессии гена cre, кодирующего сайт-специфическую для loxp рекомбиназу.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 представлена конструкция рекомбинантной плазмиды по настоящему изобретению pT7Blue/fadR.
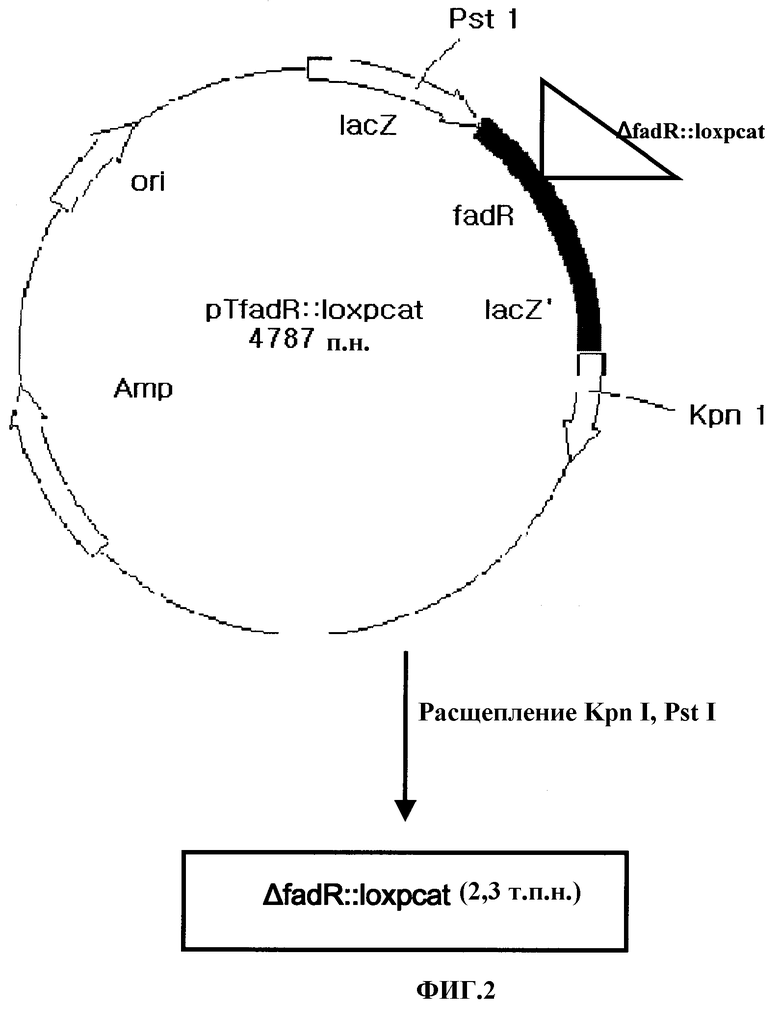
На фиг.2 представлена конструкция фрагмента ДНК ΔfadR::loxpcat из рекомбинантной плазмиды pT7fadR::loxpcat.
На фиг.3 показана хромосомная рекомбинация гена fadR по настоящему изобретению посредством анализа саузерн-блоттинга.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В соответствии с настоящим изобретением применяемый для ферментации микроорганизм характеризуется выключением гена fadR, регуляторного фактора для экспрессии оперона fad и оперона aceBAK. Конкретно, в данном изобретении для получения L-треонина посредством ферментации применяют мутантный микроорганизм, у которого расположенный на хромосоме ген fadR приведен в состояние выключения посредством гомологичной рекомбинации так, что экспрессия оперона aceBAK белком fadR не репрессирована и продолжает оказывать благоприятное воздействие в ходе биосинтеза L-треонина.
Для настоящего изобретения можно применять несколько прокариотических и эукариотических микроорганизмов со способностью к продукции L-треонина. Типичные примеры пригодных микроорганизмов включают в себя в качестве не ограничивающих примеров мутантные штаммы, принадлежащие родам Escherichia, Coryneform bacteria, Seratia и Providencia, ранее применяемым в данной области техники для получения L-треонина посредством ферментации. В предпочтительном осуществлении настоящего изобретения микроорганизмы включают в себя в качестве неограничивающих примеров продуцирующий L-треонин штамм Escherichia coli, устойчивый к аналогам L-метионина, L-треонина и L-лизина и к α-аминомаслянной кислоте и требующий для роста метионин и обладающий небольшой потребностью в изолейцине. В другом предпочтительном осуществлении настоящего изобретения микроорганизмы включают в себя мутанты, у которых в дополнение к собственным генам, кодирующим фофсофенолпируваткарбоксилазу (ppc) и входящие в состав треонинового оперона ферменты, в хромосомную ДНК дополнительно введена, по меньшей мере, одна копия гена, выбранного из экзогенного гена ppc и включенных в состав треонинового оперона генов thrA, thrB и thrC.
Расположенный на хромосоме продуцирующий L-треонин микроорганизма ген fadR можно выключать соответствующим способом выключающей мутации, известным специалистам в данной области. «Выключающая мутация» здесь обозначает проводимое in vitro нарушение в природной хромосомной ДНК, как правило, в кодирующей белок области так, что в природную последовательность вставляется часть чужеродной ДНК, легко, хотя и не обязательно, обеспечивающая доминантный селективный маркер. Выключающая мутация в кодирующей белок области предотвращает экспрессию белка дикого типа, что обычно приводит к потере функции, обеспечиваемой данным белком.
Процедуру выключения можно проводить посредством смешивания кассеты для выключения с культурой продуцирующего L-треонин микроорганизма, компетентного для захвата ДНК.
Хотя микроорганизм способен трансформироваться от природы, предпочтительно, чтобы клетки любым пригодным способом можно было приводить в состояние компетентности для захвата ДНК (см., например, LeBlanc et al. Plasmid 28, 130-145, 1992; Pozzi et al. J. Bacteriol. 178, 6087-6090, 1996). «Кассета для выключения» относится к фрагменту природной хромосомной ДНК, несущей часть чужеродной ДНК, которая может нести селективный маркер. В одном осуществлении «кассеты для выключающих мутаций» создают посредством прерывания фрагмента геномной ДНК чужеродной частью ДНК и замещением хромосомной копии последовательности дикого типа кассетой для выключения. В данном осуществлении протокол выключения включает в себя клонирование фрагмента чужеродной ДНК в ДНК-мишень так, что на 5'- и 3'-концах кассеты для выключения остаются содержащие участок ДНК-мишени «хвосты». Для эффективной рекомбинации и/или генной конверсии концы должны составлять, по меньшей мере, 50 пар оснований, а предпочтительно более чем от 200 до 500 пар оснований. Для удобства, клонированная в ДНК-мишень чужеродная ДНК также содержит селективный маркер, например ген устойчивости к антибиотику. Когда ДНК-мишень разрушена устойчивым к антибиотику геном, выбор трансформантов проводят на содержащих подходящие уровни соответствующего антибиотика чашках с агаром. После трансформации часть захвативших кассету для выключения клеток посредством хвостов геномной ДНК в кассете претерпевают гомологичную рекомбинацию или генную конверсию, что приводит к замещению геномной последовательности дикого типа на кассету для выключения. События выключающей рекомбинации легко подтвердить, например, посредством гибридизации по Саузерну или более удобно - посредством ПЦР.
В одном осуществлении выключающая мутация по настоящему изобретению включает в себя следующие ниже процедуры. Геномную ДНК выделяют из способного продуцировать L-треонин штамма и традиционными способами, с применением геномной ДНК в качестве матрицы, проводят ПЦР для амплификации гена fadR. Полученный таким образом продукт ПЦР, ген fadR, клонируют в пригодную плазмиду или другие векторы и полученный рекомбинантный вектор посредством трансдукции вводят в клетку-хозяина, такую как E.coli. После того, как трансформант выращивают для пролиферации в культуральной среде, выделяют рекомбинантный вектор с генами fadR. В ген fadR выделенного рекомбинантного вектора(ов), подлежащего конструированию, в качестве рекомбинантного вектора для выключения вставляют фрагмент гена устойчивости к антибиотику, а рекомбинантный вектор для выключения посредством трансформации вводят в клетку-хозяина, а клетку-хозяина культивируют в пригодной культуральной среде. Затем размножившийся рекомбинантный вектор выделяют из полученного трансформанта и посредством обработки подходящим рестрикционным ферментом получают фрагмент кассеты для выключения гена fadR. Затем данный фрагмент традиционным способом, таким как электропорация, вводят хозяину, способному продуцировать L-треонин. Как результат, с применением устойчивости к антибиотику выделяют хозяина, со свойствами выключения гена fadR в течение повторного выращивания, а ген fadR дикого типа на хромосоме хозяина замещается фрагментом гена fadR, содержащим антибиотик-маркер.
Специалисты в данной области поймут, что кассеты для выключения и участки ДНК по данному изобретению или их фрагменты можно получать посредством обычных способов клонирования. Предпочтительными являются способы амплификации ПЦР с применением олигонуклеотидных праймеров, направленных к любой подходящей области любой из описанных здесь последовательностей. Способы амплификации ПЦР широко известны в данной области. См., например, PCR Protocols: A Guide to Method and Application, Ed. M. Innis et al., Academic Press (1990) или патент США № 4889818, включенный сюда в качестве ссылки. ПЦР включает в себя геномную ДНК, пригодные ферменты, праймеры и буферы, и ее удобно проводить в DNA Thermal Cycler (Perkin Elmer Cetus, Norwalk, Conn. USA). Положительный результат ПЦР определяют, например, посредством выявления фрагмента ДНК соответствующего размера после электрофореза в агарозном геле.
В предпочтительном осуществлении настоящего изобретения в качестве продуцирующего L-треонин микроорганизма применяют продуцирующие L-треонин штаммы Escherichia coli, устойчивые к аналогам L-метионина, L-треонина и L-лизина и к α-аминомаслянной кислоте и требующие для роста метионин, и обладающие небольшой потребностью в изолейцине.
Аналоги L-метионина включают в себя в качестве неограничивающих примеров D,L-этионин, норлейцин, α-метилметионин и L-метионин-D,L-сульфоксимин. Аналоги L-треонина включают в себя в качестве неограничивающих примеров α-амино-β-гидроксивалериановую кислоту и D,L-треонингидроксамат.
Аналоги L-лизина включают в себя в качестве неограничивающих примеров S-(2-аминоэтил)-L-цистеин и δ-метил-L-лизин.
В особенно предпочтительном осуществлении в настоящем изобретении в качестве продуцирующего L-треонин микроорганизма применяют новый мутант KCCM 10236 Escherichia coli с выключением гена fadR на хромосоме и способный к продукции большего количества L-треонина, чем родительский штамм KCCM 10236. Данный мутант получают трансформацией посредством электропорации кассеты для выключения ΔfadR::loxpcat в KCCM 10236 Escherichia coli, устойчивый к аналогам L-метионина, L-треонина и L-лизина и к α-аминомаслянной кислоте и требующий для роста метионин и обладающий небольшой потребностью в изолейцине. Кассету для выключения ΔfadR::loxpcat вырезают из конструкции рекомбинантной плазмиды pT7fadR::loxpcat, полученной из конструкции рекомбинантной плазмиды pT7Blue/fadR. Новый мутант обозначили как FTR 1201 Escherichia coli и 13 сентября 2002 года согласно Будапештскому договору депонировали в Korean Culture Center of Microorganisms под инвентарным номером KCCM-10422.
Мутации в KCCM 10236 Escherichia coli получали из KCCM TF4076 (KFCC 10718) Escherichia coli, требующего метионин и устойчивого к аналогу L-треонина (например, α-амино-β-гидроксивалериановой кислоте), аналогу L-лизина (например, S-(2-аминоэтил-L-цистеину), аналогу L-изолейцина (например, α-аминомаслянной кислоте), аналогу метионина (например, этионину) и т.п.
KCCM TF4076 Escherichia coli описан в публикации патента Кореи № 92-8365, полностью включенной сюда в качестве ссылки. KCCM 10236 Escherichia coli отличается от KCCM TF4076 Escherichia coli тем, что он несет два гена фосфоенолпируваткарбоксилазы (ppc) и два оперона L-треонина (thr). Ген ppc и оперон thr, происходящие из хромосомы KCCM TF4076 Escherichia coli, амплифицировали посредством полимеразной цепной реакции и дополнительно встроили в хромосомы KCCM TF4076 Escherichia coli для получения KCCM 10236 Escherichia coli.
Ген ppc катализирует формирование оксалоацетата из фосфоенолпирувата. По существу, фосфоенолпируват представляет собой предшественник оксалоацетата, являющегося промежуточным соединением пути биосинтеза треонина. Таким образом, KCCM 10236 Escherichia coli тем, что он может усиливать экспрессию гена ppc и трех генов, вовлеченных в биосинтез треонина из аспартата, thrA, thrB и thrC, кодирующих аспартаткиназу I/гомосериндегидрогеназу, гомосеринкиназу и треонинсинтазу соответственно, что приводит к увеличению продукции L-треонина.
Геномную ДНК выделяют из KCCM 10236 Escherichia coli. Для амплификации гена fadR проводят ПЦР с применением геномной ДНК в качестве матрицы. Продукты ПЦР вносят в агарозный гель и подвергают электрофорезу. Выделенный таким образом ген fadR или фрагмент его ДНК клонируют в соответствующий вектор. Для получения выключения гена fadR в область гена fadR конструируемого рекомбинантного вектора вставляют фрагмент ДНК, дающего устойчивость к антибиотику гена.
Из полученного рекомбинантного вектора выделяют кассету для выключения гена fadR, содержащую фрагмент ДНК для устойчивости к антибиотику. Для получения мутантного штамма со способностью к высокой продукции L-треонина выделенную кассету для выключения гена fadR традиционными способами, такими как электропорация, трансформируют в продуцирующий L-треонин микроорганизм, такой как KCCM 10236 Escherichia coli.
Как применяют в получении выключения гена fadR или его фрагментов, термин «вектор» означает молекулу нуклеиновой кислоты, способную нести связанный с ней чужеродный ген fadR или фрагмент его ДНК и вводящую ген fadR или фрагмент ДНК в клетки-хозяева. Примерами векторов являются плазмиды, космиды, вирусы и бактериофаги, происходящие из природных источников или синтезированные рекомбинантным способом.
Получение L-треонин с применением мутантного продуцирующего L-треонин микроорганизма по изобретению, в котором находящийся на хромосоме микроорганизма ген fadR приведен в состояние выключения, можно проводить обычным способом культивирования хозяйских клеток, таких как бактерии и дрожжи. В качестве среды, применяемой для получения L-треонина, можно применять любую из синтетических сред и натуральных сред, до тех пор, пока она содержит источники углерода, источники азота, неорганические вещества и следовые количества питательных веществ, необходимых штамму.
Примеры источников углерода включают в себя углеводороды, такие как глюкоза, фруктоза, лактоза, меласса, гидролизат целлюлозы, грубый гидролизат сахара и гидролизат крахмала, органические кислоты, такие как пировиноградная кислота, уксусная кислота, фумаровая кислота, яблочная кислота и молочная кислота и спирты, такие как глицерин и этанол. Примеры источников азота включают в себя аммиак, различные неорганические соли (такие как хлорид аммония, сульфат аммония, ацетат аммония и фосфат аммония), аммонийные соли органических кислот, амины, пептон, мясной экстракт, жидкий кукурузный экстракт, гидролизат казеина, гидролизат соевого жмыха, различные ферментированные клетки и их гидролизованные вещества.
Примеры неорганических веществ включают в себя дигидрофосфат калия, двухосновный гидрофосфат калия, фосфат магния, сульфат магния, хлорид магния, хлорид натрия, сульфат железа, сульфат марганца, сульфат меди, хлорид кальция и карбонат кальция.
Культивирование проводят в аэробных условиях, например, посредством перемешивания культуры или центрифугирования культуры при аэрации. Температура культивирования находится в диапазоне от 20 до 40°C, предпочтительно от 28 до 37°C. pH среды находится в диапазоне от pH 5 до 9, предпочтительно около нейтрального. Регулирование pH осуществляют посредством применения карбоната кальция, органической или неорганической кислоты, щелочного раствора, аммиака, буфера pH и т.д.
Как правило, L-треонин образуется и накапливается в культуре с 1 по 7 сутки культивирования. Необязательный способ культивирования включает в себя, например, любой из непрерывного режима, полунепрерывного режима и серийного режима.
После завершения культивирования такие преципитаты как клетки, удаляют из культуры, а L-треонин можно выделить из культуры посредством сочетания ионообменной хроматографии, концентрирования, высаливания и т.д. Например, выделение L-треонина из культурального бульона проводят указанным ниже способом. Культуральный бульон, из которого удаляют клетки, соляной кислотой доводят до pH 2. Затем раствор бульона пропускают через сильно кислую ионообменную смолу, а адсорбированное вещество элюируют разбавленным водным аммиаком. Аммиак выпаривают и затем полученный раствор конденсируют. Добавляют спирт для концентрирования, а затем собирают кристаллы, сформированные при охлаждении, а затем можно получать L-треонин.
Настоящее изобретение будет раскрыто на основе нескольких конкретных примеров осуществлений, которые, однако, не рассматриваются как ограничивающие.
Пример 1
Конструирование рекомбинантной плазмиды и выключение гена fadR
Геномную ДНК выделяли из продуцирующего треонин штамма Escherichia coli KCCM 10236 с применением QIAGEN Genomic-tip System. Фрагмент ДНК (приблизительно 0,8 т.п.н.), включающий в себя ORF (открытую рамку считывания) гена fadR, амплифицировали посредством ПЦР с применением выделенной в качестве матрицы геномной ДНК и пары олигонуклеотидов 5'-TCG CGG AAG AGT ACA TTA TTG-3' (прямой праймер) и 5'-ATC GGC GCA AAG AAG TCC-3' (обратный праймер). ПЦР проводили в количестве 30 циклов, каждый из которых состоял из денатурации в течение 30 секунд при 94°C, отжига в течение 30 секунд при 55°C и удлинения в течение 60 секунд при 72°C по порядку.
Продукт ПЦР вносили в 1,0% агарозный гель и подвергали электрофорезу. Соответствующую 0,8 т.п.н гена fadR полосу вырезали из геля и элюировали. Для конструирования рекомбинантной плазмиды pT7Blue/fadR (см. фиг.1) полученный ген fadR в течение ночи при температуре 16°C лигировали в участок EcoRV клонирующего вектора pT7Blue (Novagen Inc., USA). Полученную плазмидную конструкцию трансформировали в NM522 Escherichia coli. Трансформированный штамм помещали на содержащую 50 мг/л карбенициллина твердую среду и культивировали в течение ночи при температуре 37°C.
Сформированные колонии собирали платиновой петлей и инокулировали в 3 мл содержащей карбенициллин жидкой среды. После культивирования в течение ночи плазмидные ДНК выделяли из культуры с применением QIAGEN Mini Prep Kit. Экстракт плазмидной ДНК расщепляли рестрикционным ферментом SacII и подтверждали клонирование гена fadR. Проверенные плазмиды pT7Blue/fadR расщепляли SacII и сформированные таким образом фрагменты ДНК подвергали электрофорезу в 0,8% агарозном геле. Полосу приблизительно 3,8 т.п.н. вырезали из геля и элюировали. У фрагмента ДНК из 3,8 т.п.н. обработкой ферментом Кленова формировали тупые концы. Для конструирования рекомбинантной плазмиды pT7ΔfadR::loxpcat длиной приблизительно 5,0 т.п.н. (см. фиг.2) полученный фрагмент ДНК по тупым концам лигировали с включающим область loxp фрагментом длиной приблизительно 1,1 т.п.н. гена устойчивости к хлорамфениколу, полученным расщеплением плазмиды ploxpcat2 (Gosset et al. A family of removable cassettes designed to obtain Ab-resistance-free genomic modifications of E. col, Gene 247, pp. 255-264) рестрикционным ферментом HincII.
Рекомбинантной плазмидой pT7ΔfadR::loxpcat трансформировали NM522 Escherichia coli. Полученные трансформанты штрихами наносили на чашку с твердой средой, содержащей 50 мг/л карбенициллина и 15 мг/л хлорамфеникола и в течение ночи культивировали при 32°C. Сформированные колонии собирали платиновой петлей и инокулировали в 3 мл содержащей карбенициллин и хлорамфеникол жидкой среды.
После культивирования в течение ночи с применением QIAGEN Mini Prep Kit выделяли плазмидные ДНК. Экстракт плазмидной ДНК расщепляли рестрикционными ферментами KpnI и PstI и сформированные таким образом фрагменты ДНК подвергали электрофорезу в 0,7% агарозном геле. Полосы длиной приблизительно 2,2 т.п.н. вырезали из геля и элюировали (см. фиг.2). Полученный фрагмент ДНК длиной приблизительно 2,2 т.п.н., ΔfadR::loxpcat посредством электропорации трансформировали в KCCM 10236 Escherichia coli и трансформированный KCCM 10236 Escherichia coli штрихами наносили на содержащую хлорамфеникол среду с агаром. У выбранных колоний проверяли продукцию L-треонина в колбе.
Пример 2
Продукция L-треонина выбранными штаммами в колбе Эрленмейера
Тридцать колоний, выбранных в примере 1, культивировали в колбе Эрленмейера, в которую добавляли приведенную в таблице 1 среду для титрования треонина, и сравнивали продукцию L-треонина.
Каждую колонию культивировали на среде LB с агаром в инкубаторе с перемешиванием при 32°C.
Одну платиновую петлю культуры инокулировали в 25 мл среды для титрования и культивировали при перемешивании при 32°C и 250 об./мин в течение 48 часов. Результаты титрования представлены в таблице 2 ниже.
Из результатов можно увидеть, что трансформанты по настоящему изобретению продуцируют приблизительно 24-25 г/л L-треонина. Следовательно, трансформированный по настоящему изобретению KCCM 10236 Escherichia coli, где ген fadR выключен, является улучшенным по сравнению со штаммом прототипом KCCM 10236, продуцирующим 23 г/л L-треонина. Кроме того, выявили, что трансформированные микроорганизмы по настоящему изобретению в сравнении со штаммом-прототипом увеличивают выход L-треонина в среде для ферментирования приблизительно вплоть до 8%. Один трансформант из них обозначили как FTR 1201 (KCCM-10422) Escherichia coli.
Пример 3
Подтверждение выключения гена fadR саузерн-блоттингом
Для подтверждения того, выключен ли в выбранных в примере 2 штаммах ген fadR проводили саузерн-блоттинг. Штамм-прототип и штаммы FTR 1201 по настоящему изобретению в течение ночи культивировали в 3 мл содержащей хлорамфеникол жидкой среды. Геномную ДНК выделяли из культуры с применением QIAGEN Genomic Kit 20.
Выделенную геномную ДНК в течение ночи расщепляли рестрикционным ферментом XmnI. Для разделения полученных фрагментов ДНК по размеру проводили электрофорез в 0,7% агарозном геле. После окончания электрофореза молекулы ДНК из агарозного геля переносили на нейлоновую мембрану (YOUNG Sci. Biodyne B Membrane) с применением способа капиллярного переноса (Molecular Cloning vol.1. pp. 6.31-6.38). Мембрану высушивали, а затем молекулы ДНК иммобилизовали на сухой мембране УФ-облучением (120 мДж/см2, SpectroLinkerTM).
Мембрану в течение 2 часов при 55°C обрабатывали раствором для предгибридизации I (Roche № 1093657). После добавления денатурированного зонда ДНК в течение ночи в термостате (BAMBINO 230300) проводили гибридизацию при 55°C.
Применяемый денатурированный зонд ДНК получали как указано ниже. Выделенную плазмиду ploxpcat2 расщепляли рестрикционным ферментом HincII с применением QIAGEN Kit для получения содержащего область loxp фрагмента ДНК длиной приблизительно 1,1 т.п.н., гена устойчивости к хлорамфениколу. Полученный фрагмент ДНК нагревали в воде при 100°C в течение 5 минут и быстро охлаждали на льду в течение 5 минут для получения однонитевых молекул ДНК, меченных DIG при 37°C с применением мечения DIG и набора для выявления (Roche №1093657) для получения меченного DIG-УДФ зонда ДНК.
После окончания гибридизации не специфически гибридизовавшиеся на мембране молекулы ДНК удаляли с применением растворов для отмывания I и II (Roche № 1093657). Мембрану маскировали буферным раствором для предгибридизации 2 (Roche № 1093657) в течение 30 минут при комнатной температуре и в течение 30 минут при комнатной температуре подвергали реакции с антителом к DIG, специфически связывающимся с DIG-УДФ.
Не специфически связавшиеся с мембраной антитела к DIG удаляли раствором для отмывания III (Roche № 1093657), а мембрану окрашивали указанной выше меткой и набором для выявления так, чтобы показалась полоса. Изображения сканировали с применением FLA-5000 Image System (FUJIFLIM). Результаты представлены на фиг.3. У штамма-прототипа KCCM 10236 (дорожка 3) не выявили полосы, так как он не несет гена устойчивости к хлорамфениколу. Напротив, у штамма по настоящему изобретению FTR 1201 (KCCM 10422) (дорожка 2), как и ожидали, выявили полосу длиной приблизительно 2,8 т.п.н. Понятно, что полоса состоит из части гена fadR длиной 1,7 т.п.н. и фрагмента ДНК гена устойчивости к хлорамфениколу приблизительно 1,1, длиной приблизительно 1,1 т.п.н.
Пример 4
Получение L-треонина в ферментере
Продукцию L-треонина штаммом FTR 1201 (KCCM-10422) по настоящему изобретению в 5 л ферментере сравнивали с продукцией L-треонина штаммом-прототипом 10236. Содержимое среды для инокуляции приведено в таблице 3 ниже.
Для инокуляции применяли среду LB, дополненную 10 г/л глюкозой и 0,1 г/л L-метионином. Начальный объем инокулята в ферментере доводили до величины от 3% до 5% начального объема культуры. Глюкозу добавляли шесть раз, каждый раз в момент ее истощения. После каждого добавления концентрация глюкозы составляла 5%. Во время добавления глюкозы также добавляли 1% по массе одноосновного фосфата калия (KH2PO4). Начальный и конечный объемы культуры составляли 1,5 л и 3,0 л соответственно, а концентрация общей глюкозы, добавленной до завершения ферментации, составляла 250 г/л. Ферментацию проводили при аэрации 0,5 vvm и перемешивании при 1000 об./мин при температуре 32°C. pH поддерживали на уровне 7,0 посредством автоматического клапана, обеспечивающего от 25 до 28% газообразного аммония.
Результаты представлены в таблице 4 ниже.
Штамм-прототип продуцировал 93,5 г/л L-треонина при выходе 37,4% по отношению к потребляемой глюкозе. Напротив, штамм по настоящему изобретению FTR 1201 продуцировал 102 г/л L-треонина при выходе 41% по отношению к потребляемой глюкозе и, таким образом, увеличивал выход треонина по сравнению со штаммом-прототипом приблизительно на 9%.
Продуцирующий L-треонин мутантный микроорганизм по настоящему изобретению, с выключением гена fadR, увеличивает продукцию L-треонина.

Изобретение относится к биотехнологии и представляет собой способ получения L-треонина. Согласно способу культивируют мутантный Е.coli, в котором находящийся на хромосоме Е.coli ген fadR выключен, выделяют полученный L-треонин из культуры. Изобретение относится также к кассете для выключения гена fadR, которая содержит антибиотик-маркер, введенный в кодирующую область гена fadR, и к способу получения мутантного Е.coli, включающего введение в продуцирующий L-треонин Е.coli кассеты для выключения гена fadR и отбор мутантного Е.coli, в котором ген fadR выключен. Изобретение позволяет получать L-треонин с высокой степенью эффективности. 4 н. и 3 з.п. ф-лы, 3 ил., 4 табл.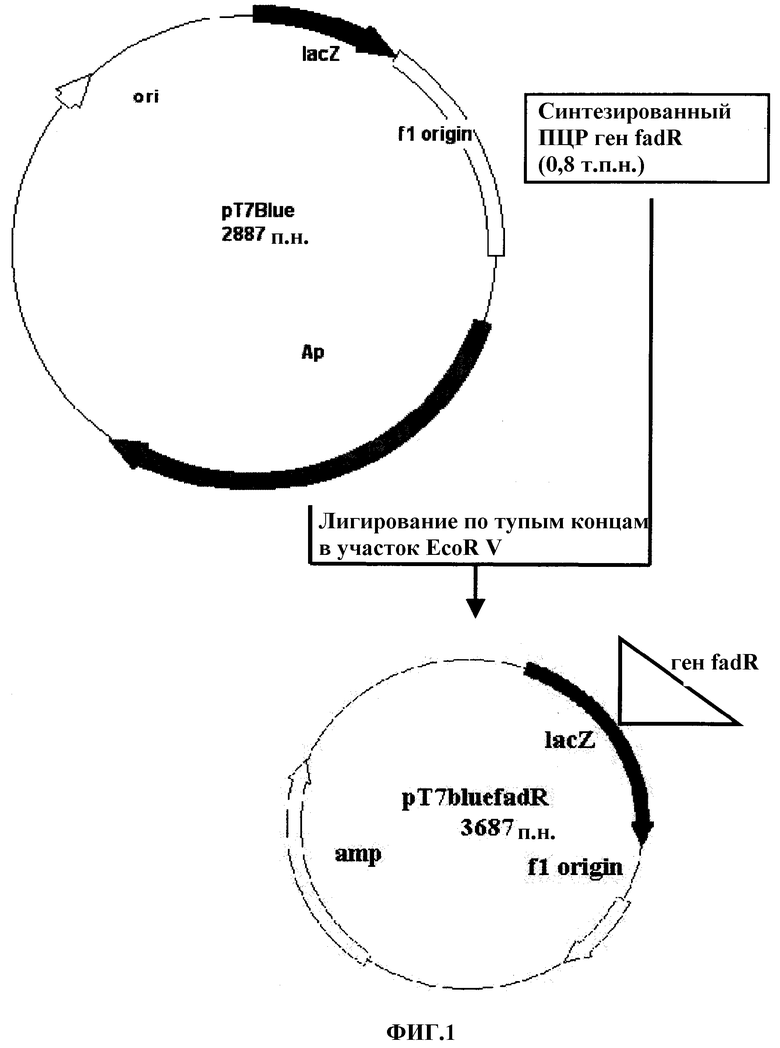
(а) конструирование кассеты для выключения гена fadR, охарактеризованной в п.5;
(b) введение указанной кассеты для выключения гена fadR в продуцирующий L-треонин E.coli, чтобы вызывать гомологичную рекомбинацию между кассетой и расположенным на хромосоме микроорганизма геном fadR;
(c) отбор мутантного E.coli, в котором ген fadR выключен.
| US 5569595, 29.10.1996 | |||
| US 5939307, 17.08.1999 | |||
| ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ АСПАРТОКИНАЗУ III, СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 1993 |
|
RU2113484C1 |

