Область техники
Настоящее изобретение относится к новому модифицированному белку орнитиндекарбоксилазе и его применению.
Предшествующий уровень техники
Путресцин (или 1,4-бутандиамин) представляет собой важное сырье для получения полиамида-4,6, включая нейлон-4,6, и главным образом производится в промышленном масштабе путем гидрирования сукцинонитрила, который производится из акрилонитрила путем добавления циановодорода. Для химического синтеза этого соединения требуются невозобновляемые продукты переработки нефти в качестве сырья и относительно высокая температура и давление в многостадийной и многореакторной конструкции, а также применение дорогостоящих каталитических систем. Кроме того, поскольку эти сырьевые материалы являются высокотоксическими и легковоспламеняющимися, известные способы химического синтеза являются неблагоприятными для окружающей среды. Соответственно, в качестве альтернативы для способа химического получения требуется способ получения путресцина из возобновляемого источника углерода на основе биомассы. Недавно способ биохимической продукции путресцина при помощи экологически безвредных микроорганизмов получил значительное внимание. Путресцин представляет собой разновидность полиамина, который обнаружен у широкого спектра организмов в диапазоне от бактерий до животных и растений. Известно, что концентрация путресцина в Е.coli является чрезвычайно высокой, составляя приблизительно 2,8 г/л. Кроме того, микроорганизмы обладают потенциально хорошей устойчивостью к высоким концентрациям полиаминов, и, таким образом, они способны расти и выживать в присутствии высоких концентраций полиаминов. Например, сообщалось о том, что Corynebacterium glutamicum может расти даже в присутствии больше чем 30 г/л кадаверина. Соответственно, непрерывно продолжаются исследования по применению микроорганизмов в продукции промышленно доступных высококонцентрированных полиаминов. Тем не менее, исследования по продукции полиаминов с использованием микроорганизмов недостаточно продвинулись в направлении их промышленной применимости. Таким образом, сохраняется потребность в конструировании штамма, способного продуцировать полиамины с высоким выходом (Qian ZG, et al., Biotechnol Bioeng, 104: 651-662, 2009; Schneider J, et al., Appl Microbiol Biotechnol, 88: 859-868, 2010).
В то же время, орнитиндекарбоксилаза (ODC) представляет собой фермент, обнаруженный у большинства микроорганизмов, которые превращают орнитин в путресцин. ODC в Е.coli как правило образует гомодимер, и активные сайты образуются на границе димера. Для механизма реакции ODC требуется пиридоксальфосфат (PLP) в качестве кофактора, и PLP образует основание Шиффа по лизиновому остатку активного сайта фермента, который позже замещается субстратом орнитином, который подвергается декарбоксилированию. При образовании путресцина ODC вновь образует основание Шиффа с PLP.
Когда ODC, вводимый в штамм, продуцирующий путресцин, рода Corynebacterium, представляет собой белок, кодируемый геном speC Е.coli, тогда его активность, как сообщается, является очень низкой. Таким образом, для конструирования штамма, продуцирующего путресцин с высоким выходом, очень важным является улучшение ODC, которая представляет собой фермент, вовлеченный в конечную стадию пути биосинтеза путресцина. До настоящего времени исследования мутации осуществлялись только в отношении структуры или реакционного механизма белка ODC, и отсутствуют сообщения об увеличении ее активности.
Описание изобретения
Техническая задача
Авторы настоящего изобретения приложили множество усилий для улучшения белка ODC, который играет важную роль в продукции путресцина, но демонстрирует низкую активность. В результате они обнаружили новый сайт для мутации, и осуществили мутацию по этому сайту для получения модифицированного белка ODC, обладающего улучшенной путресцин-продуцирующей активностью, и также обнаружили, что когда модифицированный белок ODC вводят в продуцирующий путресцин микроорганизм, тогда микроорганизм способен продуцировать путресцин с высоким выходом, таким образом, завершая настоящую заявку на изобретение.
Техническое решение
Задача настоящего изобретения заключается в том, чтобы предложить новый модифицированный белок орнитиндекарбоксилазу (ODC).
Еще одна задача настоящего изобретения заключается в том, чтобы предложить полинуклеотид, кодирующий модифицированный белок ODC, вектор, включающий полинуклеотид, и трансформант, трансформированный этим вектором.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить способ получения путресцина, включающий стадию взаимодействия L-орнитина, смеси, содержащей L-орнитин, или L-орнитиновой культуральной жидкости с модифицированным белком ODC.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить рекомбинантный микроорганизм, который обладает улучшенной путресцин-продуцирующей активностью благодаря замене на модифицированный белок ODC у микроорганизма Corynebacterium sp, обладающего путресцин-продуцирующей активностью.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить способ получения путресцина, включающий стадии культивирования микроорганизма Corynebacterium sp, обладающего улучшенной путресцин-продуцирующей активностью благодаря введению модифицированного белка ODC; и выделения путресцина из культуры, полученной на вышеприведенной стадии.
Полезные эффекты
Модифицированный белок орнитиндекарбоксилаза в соответствии с настоящим изобретением обладает путресцин-конвертазной активностью, которая в 21 раз превышает такую активность у нативной формы. Когда модифицированный белок орнитиндекарбоксилазу вводят в путресцин-продуцирующий штамм, тогда продуктивность путресцина значительно улучшается. Таким образом, этот способ может широкого использоваться для эффективного массового производства путресцина в качестве альтернативы известному пути химического синтеза.
Описание графических материалов
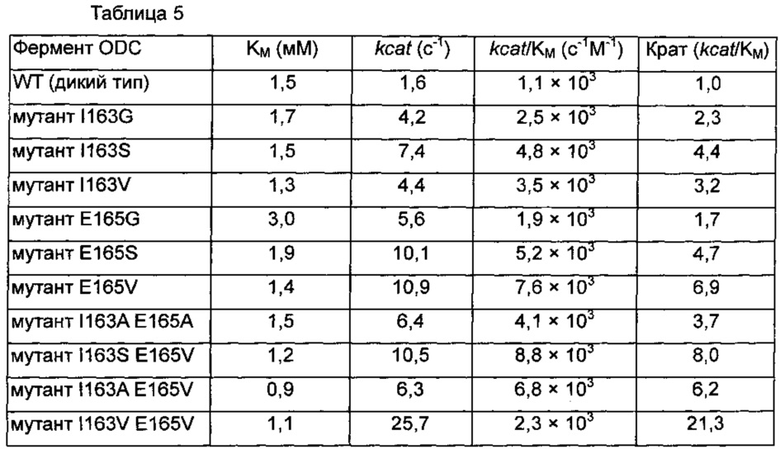
Фиг. 1 демонстрирует сравнение путресцин-конвертазной активности между нативным белком ODC, полученным из Е.coli, и белком ODC, имеющим мутацию I163A или Е165А. Подробно, рН увеличивается по мере прохождения реакции превращения, и когда увеличение рН проверяют при помощи фенолового красного, тогда обнаруживают увеличение поглощения. Обнаружено, что белок ODC, имеющий I163A или Е165А, или обе мутации, демонстрирует превосходную путресцин-конвертазную активность по сравнению с нативным белком ODC.
Лучший вариант
Для решения вышеприведенных задач, в настоящем изобретении предложен новый модифицированный белок ODC, причем этот модифицированный белок ODC имеет мутацию по одному или более чем одному аминокислотному остатку, выбранному из группы, состоящей из аминокислотного остатка изолейцина в положении 163 и аминокислотного остатка глутаминовой кислоты в положении 165 от N-конца орнитиндекарбоксилазы (ODC), имеющей аминокислотную последовательность, представленную в SEQ ID NO: 1.
Используемый здесь термин ''орнитиндекарбоксилаза (ODC)'' относится к ферменту, который катализирует следующую реакцию, представляющую собой первую стадию синтеза полиамина из орнитина, и последнюю стадию пути синтеза путресцина. При получении путресцина с использованием L-орнитина в качестве субстрата, пиридоксальфосфат (PLP) действует в качестве кофактора.
Схема реакции
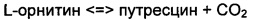
В настоящем изобретении орнитиндекарбоксилаза (ODC) может представлять собой в частности ODC, полученную из Е.coli, и конкретней ODC, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 1, полученную из Escherichia coli.
В настоящем изобретении способ получения ODC (орнитиндекарбоксилаза) может быть осуществлен путем применения различных способов, известных в области техники. Например, ODC может быть получена способом генного синтеза, включающим оптимизацию кодона для получения фермента с высоким выходом из Е.coli, который обычно используется в экспрессии ферментов, и способом скрининга полезных источников ферментов методами биоинформатики на основе информации о геноме микроорганизма, но не ограничиваясь этими способами.
Используемый здесь термин ''модифицированный белок ODC'' относится к белку ODC, в котором одна или более чем одна аминокислота в аминокислотной последовательности белка ODC добавлена, удалена или заменена. В частности, модифицированный белок ODC относится к белку, у которого активность эффективно увеличена путем модификации белка ODC по сравнению с белком дикого типа. В настоящем изобретении модификация может быть осуществлена, без ограничения, с использованием любого общего способа улучшения ферментов, известного в области техники, и примеры способа представляют собой стратегии, такие как рациональный дизайн и направленная эволюция. Например, стратегия рационального дизайна может включать мутацию по аминокислоте в конкретном сайте (сайт-направленный мутагенез), и стратегия направленной эволюции может включать случайный мутагенез. Кроме того, природная(ые) модификация(и) могут осуществляться по аминокислотному(ным) остатку(кам) в положении 163 и/или в положении 165 в SEQ ID NO: 1 без манипуляции извне. Использованные здесь термины ''модифицированный белок ODC'', ''мутант ODC'' и ''мутант speC'' могут быть использованы взаимозаменяемо.
В частности, модифицированный белок ODC по настоящему изобретению может иметь модификацию(и) аминокислотного остатка изолейцина в положении 163 и/или аминокислотного остатка глутаминовой кислоты в положении 165 от N-конца орнитиндекарбоксилазы (ODC), полученной из Escherichia coli и имеющей аминокислотную последовательность, представленную в SEQ ID NO: 1. Например, глутаминовая кислота в положении 165 может быть заменена на аланин, глицин, серии или валин, или изолейцин в положении 163 может быть заменен на аланин, глицин, серии или валин. Кроме того, модифицированный белок ODC может иметь двойную модификацию изолейцина в положении 163, а глутаминовая кислота в положении 165, в которой изолейцин в положении 163 и глутаминовая кислота в положении 165 могут быть заменены на аминокислоту, выбранную из группы, состоящей из аланина, валина, серина и глицина, соответственно. В частности, изолейцин в положении 163 и глутаминовая кислота в положении 165 могут быть заменены на аланин-аланин, аланин-валин, серин-валин или валин-валин, соответственно.
В воплощениях настоящего изобретения, когда обнаружили, что различные комбинации мутаций по аминокислотам в положениях 163 и 165 ODC дикого типа приводят к увеличению продукции путресцина, то эти положении стали рассматривать как очень важные в получении мутанта ODC, обладающего улучшенной продуктивностью в отношении путресцина. В частности, когда аминокислоты, находящиеся в важном для мутаций сайте, заменяли на небольшие аминокислотные остатки (аланин, серии, валин или глицин), тогда продуктивность в отношении путресцина увеличивалась.
Кроме того, модифицированный белок ODC по настоящему изобретению может состоять из любой из аминокислотных последовательностей от SEQ ID NO: 34 по SEQ ID NO: 57, и в частности, любой из аминокислотных последовательностей от SEQ ID NO: 34 по SEQ ID NO: 42, SEQ ID NO: 45, и SEQ ID NO: 49 и SEQ ID NO: 57, которые представляют собой аминокислотную последовательность модифицированного белка ODC, в которой изолейцин или глутаминовая кислота в положении 163 или 165, соответственно, от N-конца заменена на небольшой остаток. Модифицированный белок ODC может включать любой полипептид, имеющий гомологию на 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 97% или 99% или больше чем 99% с вышеприведенными последовательностями, пока он несет вышеприведенную модификацию и обладает путресцин-конвертазной активностью, превосходящей активность для дикого типа.
Используемый здесь термин ''гомология'' относится к процентной доли идентичности между двумя полинуклеотидными или полипептидными группировками. Соответствие между последовательностями от одной группировки до другой может быть определено при помощи известного в области техники способа. Например, гомология может быть определена при помощи информации о непосредственном выравнивании последовательностей для двух полинуклеотидных молекул или двух полипептидных молекул с использованием компьютерной программы, которая легко доступна и способна представлять информацию о выравнивании последовательностей. Дополнительно, гомология может быть определена путем гибридизации полинуклеотидов в условиях образования стабильной двойной цепи в гомологичных областях и затем расщепления гибридизированной цепи при помощи нуклеазы, специфической в отношении единичной цепи, для определения размера расщепленного фрагмента.
Используемый здесь термин ''гомологичный'' относится к корреляции между белками, где все грамматические формы и вариации написания включают белки суперсемейства и другие полученные для данного вида гомологичные белки, имеющие ''общее эволюционное происхождение''. Такие белки (и кодирующие их гены) обладают гомологией последовательности, отражаемой высокой степенью сходства последовательностей. Тем не менее, в общем применении и в настоящем изобретении, когда термин ''гомогенность'' скорректирован прилагательным, таким как ''очень высокая'', тогда он относится к сходству последовательностей, а не к общему эволюционному происхождению.
Используемый здесь термин ''сходство последовательностей'' относится к степени идентичности или гомологии между нуклеотидными последовательностями или аминокислотными последовательностями белков, которые могут иметь общее эволюционное происхождение или не иметь такого. В конкретном воплощении, когда совпадение полипептидов между двумя аминокислотными последовательностями составляет по меньшей мере 21% для фиксированной длины аминокислотной последовательности (в частности, по меньшей мере приблизительно 50% и конкретней приблизительно 75%, 90%, 95%, 96%, 97% или 99%), тогда эти две последовательности являются ''по существу гомологичными'' или ''по существу схожими''. По существу гомологичные последовательности могут быть идентифицированы путем сравнения последовательностей с использованием стандартного программного обеспечения, используемого в базе данных или, например путем осуществления эксперимента с гибридизацией по Саузерну в строгих условиях, определенных для конкретной системы. Определенные условия, подходящие для гибридизации, находятся в объеме обычных способов в области техники (например, смотри Sambrook et al., 1989, ниже).
В конкретном воплощении настоящего изобретения осуществляли структурный анализ белка ODC, полученного из Е.coli, и основываясь на структурной информации, осуществляли мутагенез при помощи стратегии рационального дизайна. Конструировали и осуществляли мутации (V156, D160, I163, Е165, Q691) для расширения входного участка пути для прохождения субстрата в активный сайт и мутации (N153, D309) для стабилизации PLP, который представляет собой кофактор, связывающийся с активным сайтом (примеры 1 и 2). Подробно, когда изолейцин в качестве аминокислоты в положении 163 и глутаминовая кислота в качестве аминокислоты в положении 165 от N-конца заменяли на аланин путем модификации, заменяющей объемный остаток во входной области пути на небольшой остаток аланин, тогда обнаружили, что активность белка значительно увеличивалась (пример 3). В то же самое время, белки ODC, имеющие 6 других типов мутантов V156A, D160A, Q691A, N153D, N153E и D309E для стабилизации PLP, демонстрировали очень низкую активность или небольшую активность по сравнению с диким типом. Таким образом, можно видеть, что изолейцин в положении 163 и глутаминовая кислота в положении 165 в белке ODC, полученном из Е.coli (SEQ ID NO: 1), представляют собой важные остатки, которые увеличивают активность белка. Мутации осуществляли путем сайт-направленного мутагенеза с использованием праймеров, приведенных в таблице 1, и ПЦР (полимеразная цепная реакция).
Кроме того, в конкретном воплощении настоящего изобретения изолейцин в положении 163 и глутаминовую кислоту в положении 165 заменяли на другие небольшие остатки, серии, валин или глицин, дополнительно к аланину, для оптимизации модификаций соответствующих остатков (пример 4 и таблица 4). Соответствующие аминокислотные остатки в положениях 163 и 165 заменяли на глицин (G), серии (S) или валин (V). В результате, когда аминокислотный остаток в положении 163 заменяли на серии, и аминокислотный остаток в положении 165 заменяли на валин, тогда величина kcat/KM увеличивалась в 4,4 раза и 6,9 раз, соответственно, по сравнению с диким типом (таблица 5). На основе этого результата два остатка изменяли спонтанно и проверяли активность ODC. Активность увеличивалась до 8 раз больше, чем активность для дикого типа путем комбинации I163S и E165V, что демонстрировало самую высокую активность при единичной мутации. С другой стороны, активность увеличивалась до 21,3 раз больше, чем активность для дикого типа путем замены обоих аминокислотных остатков в положениях 163 и 165 на валин (пример 4 и таблица 5).
В целом, увеличенные активности мутантов фермента ODC относятся к повышению величины kcat/KM вследствие повышения величины kcat, нежели чем уменьшения величины KM. Предполагается, что структура фермента ODC изменяется для увеличения скорости превращения в продукт путресцин, нежели чем связывающей аффинности субстрата орнитина для фермента.
В настоящем изобретении активность фермента ODC анализируют путем использования реакции, превращающей орнитин в путресцин. Конкретно, когда фермент ODC превращает одну молекулу орнитина в путресцин, тогда расходуется одна молекула воды и образуется одна молекула диоксида углерода и один ион ОН- вместе с путресцином. Таким образом, общий рН увеличивается. Когда увеличенный рН измеряют при 559 нм с использованием фенолового красного, представляющего собой индикатор рН, тогда поглощение увеличивается пропорционально увеличению рН во время реакции. Это свойство используют для опосредованного измерения продуцируемого количества путресцина.
Используемый здесь термин ''орнитин'' относится к основной аминокислоте, которая играет важную роль в орнитиновом цикле и, в частности, L-орнитин широко обнаруживается в растениях, животных и микроорганизмах. В общем, орнитин играет важную роль в связи с циклом мочевины у организма, имеющего орнитиновый цикл. Кроме того, орнитин может подвергаться взаимопревращению в аргинин, глутаминовую кислоту и пролин в организме, и он переносит аминогруппы на α-кетокислоту и глиоксалевую кислоту. Орнитин представляет собой субстрат, продуцирующий амин (путресцин) посредством орнитиндекарбоксилазы, и из него синтезируется полиамин. В настоящем изобретении орнитин может представлять собой в частности L-орнитин, который может быть использован в качестве субстрата для орнитиндекарбоксилазы.
Используемый здесь термин ''путресцин'' представляет собой вещество, продуцируемое путем декарбоксилирования орнитина или гидролиза агматина. Путресцин может быть обнаружен при гнилостном разложении, но также обычно обнаруживается в нормальном компоненте в организме. Путресцин представляет собой полиамин, входит в состав рибосом и способствует клеточному росту или синтезу РНК. В промышленности путресцин представляет собой важный сырьевой материал для продукции полиамида-4,6, включая нейлон-4,6, и существует постоянная потребность в исследованиях его массового производства.
В еще одном аспекте настоящего изобретения предложен полинуклеотид, кодирующий модифицированный белок ODC по настоящему изобретению.
Используемый здесь термин ''полинуклеотид'' охватывает молекулы ДНК и РНК, и нуклеотид в качестве основной единицы полинуклеотида включает природный нуклеотид, а также аналог с модифицированным сахаром или основанием.
В еще одном аспекте настоящего изобретения предложен вектор, включающий полинуклеотид, кодирующий модифицированный белок ODC по настоящему изобретению.
Используемый здесь термин ''вектор'' относится к любому носителю для клонирования и/или переноса оснований в клетку-хозяин. Вектор может представлять собой репликон для того, чтобы дать возможность для репликации фрагментов, комбинированных с другими фрагментами ДНК. ''Репликон'' относится к любой генетической единице, действующей в качестве самореплицирующейся единицы для репликации ДНК in vivo, то есть, реплицируемой путем саморегуляции (например, плазмида, фаг, космида, хромосома и вирус). Термин ''вектор'' может включать вирусные и невирусные носители для введения нуклеотидов в клетку хозяин in vitro, ex vivo или in vivo, и может также включать минисферическую ДНК. Например, вектор может представлять собой плазмиду без последовательности бактериальной ДНК. Удаление последовательностей бактериальной ДНК, богатых областью CpG, осуществляли для уменьшения сайленсинга трансгенной экспрессии и для того, чтобы способствовать непрерывной экспрессии вектора плазмидной ДНК. Термин ''вектор'' может также включать транспозон или искусственную хромосому.
В настоящем изобретении вектор представляет собой вектор, включающий полинуклеотид, кодирующий модифицированный белок ODC по настоящему изобретению, и он может представлять собой, но без ограничения, вектор, способный к репликации и/или экспрессии полинуклеотида в эукариотической или прокариотической клетке, включающей клетку млекопитающего (например, клетку человека, обезьяны, кролика, крысы, хомяка, мыши и т.п.), клетку растения, дрожжевую клетку, клетку насекомого или бактериальную клетку (например, Е.coli и т.п.). В частности, вектор может представлять собой вектор, который функционально связан с надлежащим промотором для обеспечения экспрессии полинуклеотида в клетке хозяине, и включает по меньшей мере один селективный маркер. Конкретней, вектор может находиться в форме, в которой полинуклеотид вводят в фаг, плазмиду, космиду, минихромосому, вирусный или ретровирусный вектор.
Система рЕТ с использованием промотора 11, обычно используемая в области техники, хорошо известна, и могут быть использованы различные системы экспрессии, известные в области техники, но не ограничиваясь ими. В настоящем изобретении, в частности, вектор, включающий полинуклеотид, кодирующий модифицированный белок ODC, может представлять собой вектор рЕТ28а.
В конкретном воплощении настоящего изобретения полинуклеотид, кодирующий сайт-направленный модифицированный белок ODC, встраивали в вектор рЕТ28а при помощи ПЦР. В этом способе получали векторы, экспрессирующие модифицированный ODC (speC), pET28a-speC_I163A, pET28a-speC_I163G, pET28a-speC_I163S, pET28a-speC_I163V, pET28a-speC_E165A, pET28a-speC_E165S, pET28a-speC_E165G, pET28a-speC_E165V, pET28a-speC_I163A_E165A, pET28a-speC_I163S_E165V, pET28a-speC_I163A_E165V, и pET28a-speC_I163V_E165V, и мутации подтверждали при помощи анализа последовательности.
В еще одном аспекте настоящего изобретения предложен трансформант, трансформированный вектором.
В настоящем изобретении трансформант не ограничен конкретным образом до тех пор, пока модифицированный ODC в соответствии с настоящей заявкой на изобретение способен экспрессироваться путем введения вектора.
Трансформант может представлять собой бактериальные клетки, такие как трансформированные Е.coli, Corynebacterium, Streptomyces, Salmonella typhimurium и т.п.; дрожжевые клетки; клетки грибов, такие как pichia pastoris и т.п.; клетки насекомых, такие как клетки Drosophila, Spodoptera Sf9 и т.п.; клетки животных, такие как СНО (клетки яичника китайского хомячка), SP2/0 (клетки мышиной миеломы), человеческие лимфобластоидные клеточные линии, COS, NSO (клетки мышиной миеломы), 293Т, клетки меланомы Боуэса, НТ-1080, ВНК (клетки почки новорожденного хомяка), НЕК (человеческие эмбриональные клетки почек) или PERC.6 (человеческие эмбриональные клетки сетчатки); или клетки растений.
В еще одном аспекте настоящего изобретения предложен способ получения путресцина, включающий стадию взаимодействия L-орнитина, смеси, содержащей L-орнитин, или L-орнитиновой культуральной жидкости с модифицированным белком ODC.
L-Орнитин, модифицированный белок ODC и путресцин являются теми же самыми, как описано выше.
В настоящем изобретении вещество, взаимодействующее с модифицированным белком ODC, для получения путресцина может представлять собой L-орнитин, смесь, содержащую L-орнитин, или L-орнитиновую культуральную жидкость. Смесь, содержащая L-орнитин, относится к смеси отдельно существующего L-орнитина и других компонентов, а L-орнитиновая культуральная жидкость относится к культуральной жидкости, в которой L-орнитин продуцируется, или его количество увеличивается во время ферментации, и, таким образом, включен L-орнитин в количестве, достаточном для реакции, но не ограничивается этим количеством.
Например, способ получения L-орнитина путем ферментации и получаемая культуральная жидкость раскрыты в патенте США №3668072, включенном сюда путем ссылки (Е.coli, АТСС 21104).
В настоящем изобретении модифицированный белок ODC может представлять собой очищенный модифицированный белок ODC или культуральную жидкость микроорганизмов, содержащую модифицированный белок ODC. В частности, микроорганизм, используемый для приготовления культуральной жидкости микроорганизмов, может представлять собой микроорганизм, экспрессирующий модифицированный белок ODC по настоящему изобретению, и конкретней он может представлять собой микроорганизм-трансформант, трансформированный вектором, включающим полинуклеотид, кодирующий модифицированный белок ODC по настоящему изобретению.
В еще одном аспекте настоящего изобретения предложен микроорганизм, обладающий улучшенной путресциновой продуктивностью, который получают путем замены на модифицированный белок ODC у микроорганизма Corynebacterium sp., обладающего путресциновой продуктивностью.
Используемый здесь термин ''микроорганизм'' включает все из микроорганизмов дикого типа и естественным образом или искусственно генетически модифицированных микроорганизмов, и он может представлять собой микроорганизм, обладающий конкретным ослабленным или усиленным механизмом вследствие встраивания чужеродного гена, или усиления или ослабления активности эндогенного гена.
Используемый здесь термин ''микроорганизм Corynebacterium sp., обладающий путресциновой продуктивностью'' относится к микроорганизму Corynebacterium sp., который обладает естественной путресциновой продуктивностью или вследствие модификации. Известно, что путресцин включен в культуру микроорганизма Corynebacterium sp. Тем не менее, его путресциновая продуктивность является очень низкой, и гены или механизмы, вовлеченные в продукцию, еще пока не выявлены. Таким образом, ''микроорганизм Corynebacterium sp., обладающий путресциновой продуктивностью'' в настоящем изобретении относится к самому нативному штамму или микроорганизму Corynebacterium sp., в который встроен чужеродный ген, вовлеченный в механизм продукции путресцина, или активность эндогенного гена усилена или ослаблена таким образом, чтобы микроорганизм обладал улучшенной путресциновой продуктивностью.
Используемый здесь термин ''микроорганизм Corynebacterium sp.'' может в частности представлять собой Corynebacterium glutamicum, Corynebacterium ammoniagenes, Brevibacterium lactofermentum, Brevibacterium flavum, Corynebacterium thermoaminogenes, Corynebacterium efficiens и т.п., но не ограничиваться ими. Конкретней, микроорганизм Corynebacterium sp. В настоящем изобретении может представлять собой Corynebacterium glutamicum, у которого рост и выживание клеток почти не поражаются даже при воздействии высокой концентрации путресцина. Например, микроорганизм Corynebacterium sp. может представлять собой Corynebacterium glutamicum штамм KCCM11240P (KCCM11138P ΔNCgI1469), который модифицирован таким образом, что обладает ослабленной активностью NCgI1469 по сравнению с его эндогенной активностью, таким образом, обладая улучшенной продуктивностью по путресцину, но не ограничиваясь им. Штамм KCCM11240P представляет собой сверхэкспрессирующий путресцин штамм, полученный путем делеции гена, кодирующего NCgI1469 для блокирования пути биосинтеза N-ацетилпутресцина из путресцина, который раскрыт в Международной заявке на патент № WO 2013/105827.
В конкретном воплощении настоящего изобретения на основе микроорганизма Corynebacterium sp. (KCCM11240P (KCCM11138P ΔNCgI1469)), обладающего улучшенной путресциновой продуктивностью за счет ослабления активности NCgI1469 по сравнению с его эндогенной активностью, получали модифицированный штамм путем замены speC дикого типа на мутант ODC M63S/E165V(speC), обладающий увеличенной путресцин-конвертазной активностью в хромосоме (пример 6). Модифицированный штамм обозначен как Corynebacterium glutamicum СС01-0578 и депонирован в Корейском центре культур микроорганизмов (KCCM) 10 июня 2013 года под инвентарным номером KCCM11425P в соответствии с Будапештским договором.
В еще одном аспекте настоящего изобретения предложен способ получения путресцина, включающий стадии культивирования микроорганизма Corynebacterium sp., обладающего улучшенной путресцин-продуцирующей активностью благодаря замене на модифицированный белок ODC в соответствии с настоящим изобретением; и выделения путресцина из культуры, полученной на вышеприведенной стадии.
Микроорганизм Corynebacterium sp.может представлять собой в частности Corynebacterium glutamicum, и конкретней штамм Corynebacterium glutamicum СС01-0578 (инвентарный номер: KCCM11425P).
Используемый здесь термин ''культура'' относится к выращиванию микроорганизма в искусственным образом контролируемых условиях окружающей среды. В настоящем изобретении способ получения путресцина с использованием микроорганизма Corynebacterium sp. может быть осуществлен с использованием способа, широко известного в области техники. В частности, примеры способа выращивания включают процесс выращивания партиями (batch process) и процесс периодического выращивания с добавлением субстрата (fed batch process) или непрерывный повторяющийся процесс периодического выращивания с добавлением субстрата (repeated fed batch process), но не ограничиваются ими.
Среда, используемая для выращивания, должна подходящим образом удовлетворять требованиям конкретных штаммов. Культуральные среды для микроорганизма Corynebacterium sp.известны (например, Manual of Methods for General Bacteriology. American Society for Bacteriology. American Society for Bacteriology. Washington D.C., USA, 1981). В качестве источника углерода в среде могут быть использованы сахара и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза, масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло, жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота, спирты, такие как глицерин и этанол, и органические кислоты, такие как уксусная кислота и т.п. Эти вещества могут быть использованы индивидуально или в виде смеси. В качестве источника азота может быть использован пептон, дрожжевой экстракт, мясной экстракт, экстракт солода, кукурузный экстракт, соевая мука и мочевина или неорганические соединения, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония, и эти вещества также могут быть использованы индивидуально или в виде смеси. В качестве источника фосфора может быть использован дигидрофосфат калия или вторичный кислый фосфат калия или соответствующая содержащая натрий соль. Кроме того, культуральная среда может включать соль металла, такую как сульфат магния или сульфат железа, который важен для роста, и наконец, важные способствующие росту вещества, такие как аминокислоты и витамины, могут быть использованы дополнительно к вышеупомянутым веществам. Подходящий предшественник может быть добавлен в культуральную среду. Вышеприведенные вещества могут быть достаточным образом вводиться в культуру партиями или непрерывно.
рН культуры может быть скорректирован при помощи подходящего основного соединения, такого как гидроксид натрия, гидроксид калия или аммиак, или кислотного соединения, такого как ортофосфорная кислота или серная кислота. Пенообразование может быть скорректировано при помощи пеногасителя, такого как полигликолевый эфир жирной кислоты. Аэробное состояние культуральной среды может поддерживаться путем введения кислорода или содержащих кислород газовых смесей (например воздуха). Температура культивирования может в общем составлять от 20°С до 45°С, в частности от 25°С до 40°С. Культивирование может продолжаться до тех пор, пока продукция путресцина не достигнет желаемого максимума, и может обычно достигаться в течение от 10 часов до 160 часов. Путресцин может высвобождаться в культуральную среду или удерживаться в клетке.
Способ получения путресцина по настоящему изобретению включает стадию выделения путресцина из клетки или культуры. Способ выделения путресцина из клетки или культуры может быть осуществлен с использованием подходящего известного в области техники способа, например центрифугирования, фильтрования, анионообменной хроматографии, кристаллизации и ВЭЖХ (высокоэффективной жидкостной хроматографии), но не ограничивающегося ими.
Вариант осуществления изобретения
Далее настоящее изобретение будет описано более подробно со ссылкой на примеры. Тем не менее, эти примеры приведены только для иллюстративных задач, и не предполагается, что изобретение ограничивается этими примерами.
Пример 1. Структурный анализ ODC (орнитиндекарбоксилаза) и конструирование ее мутанта
В общем, известно, что Е.coli имеет два типа ODC. Один из них представляет собой индуцируемую ODC (speF), экспрессия которой индуцируется при кислом рН, а другой представляет собой конститутивную ODC (speC), вовлеченную в продукцию диамина, такого как путресцин (Applebaum DM, et al., Biochemistry, 16: 1590-1581, 1977). Из них в качестве гена-мишени выбрали spec, который представляет собой конститутивную ODC, вовлеченную в продукцию путресцина.
До настоящего времени выявили структуры ODC бактерий Vibrio и Lactobacillus. Предположили, что из них ODC Е.coli (speC) обладает структурой, напоминающей структуру ODC 30а Lactobacillus. Таким образом, на основе 3D структуры ODC Lactobacillus, выравнивание аминокислотной последовательности ODC (speC) Е.coli осуществляли с использованием программы GeneDoc (Momany С, et al., J Mol Biol, 4: 849-854, 1995). В результате сравнения аминокислотных последовательностей идентичность последовательностей между speC Е.coli и ODC Lactobacillus 30а составляла 53%, и сходство последовательностей между ними составляло 65%, свидетельствуя о том, что два фермента весьма похожи друг на друга. Таким образом, на основе структуры ODC 30а Lactobacillus (PDB ID: 10RD), приведенной в базе данных белков RCSB, осуществляли гомологичное моделирование структуры speC Е.coli. В результате общие скелеты белков почти идентичны друг другу, и аминокислотные последовательности активного сайта, вовлеченного в связывание с PLP (пиридоксальфосфат), были также почти идентичными друг другу.
Результат анализа структур двух ферментов продемонстрировал, что ODC существует в клетках в виде димера, их активный сайт формируется на димерной границе, и входная область пути для проникновения субстрата в активный сайт является узкой. Таким образом, для расширения входной области для эффективного проникновения в активный сайт и быстрого превращения в продукт сконструирована модификация для замены объемных остатков во входной области на небольшие остатки (V156, D160, I163, Е165, Q691).
Дополнительно, для стабилизации кофактора PLP, связывающегося с активным сайтом, также конструировали мутацию для остатков, окружающих активный сайт (N153, D309).
Пример 2. Клонирование и экспрессия гена ODC (speC) Е.coli
Для экспрессии гена speC Е.coli использовали векторную систему рЕТ28а (Novagen), обычно используемую для экспрессии ферментов. Сначала ген speC амплифицировали при помощи ПЦР с использованием хромосомы Е.coli W3110 дикого типа в качестве матрицы и праймеров, приведенных в следующей таблице 1. Фрагмент гена, полученный путем амплификации при помощи ПЦР и вектор рЕТ28а, обрабатывали ферментами рестрикции NdeI и XhoI (37°С, 3 часа), и затем фрагмент гена speC встраивали в вектор рЕТ28а при помощи общего способа лигирования.

Полученную таким образом мутацию экспрессирующего speC вектора (pET28a-speC) подтверждали при помощи секвенирующего анализа.
Остатки-мишени в примере 1 заменяли на небольшой остаток аланин, соответственно. Для стабилизации PLP каждый остаток, окружающий активный сайт, модифицировали по-разному в соответствии с положением, связывающемся с PLP.
ПЦР осуществляли с использованием полученного вектора pET28a-speC в качестве матрицы и праймеров, приведенных в таблице 1 и следующей таблице 2. Во-первых, для мутации гена speC первичную ПЦР осуществляли для прямых (5') и обратных (3') областей относительно мутантной области, и затем осуществляли вторичную ПЦР для лигирования двух фрагментов ПЦР. Например, в случае speC V156A прямую область амплифицировали при помощи ПЦР с использованием speC_start (NdeI)_5 (SEQ ID NO: 2) и speC_V156A_3 (SEQ ID NO: 5) в качестве праймеров, и обратную область амплифицировали при помощи ПЦР с использованием speC_V156A_5 (SEQ ID NO: 4) и speC_stop (XhoI)_3 (SEQ ID NO: 3) в качестве праймеров. Вторичную ПЦР осуществляли с использованием двух фрагментов ПЦР, полученных при помощи первичной ПЦР в качестве матрицы и speC_start (NdeI)_5 (SEQ ID NO: 2) и speC_stop (XhoI)_3 (SEQ ID NO: 3) в качестве праймеров. Полученный наконец ген speC V156A встраивали в вектор рЕТ28а тем же самым образом как фрагмент гена speC. Другие мутантные фрагменты также вводили в вектор рЕТ28а при помощи ПЦР с использованием праймеров, приведенных в таблице 2, тем же самым образом как приведено выше, соответственно.
Мутации полученных таким образом мутантных векторов экспрессии speC (pET28a-speC_V156A, pET28a-speC_D160A, pET28a-speC_I163A, pET28a-speC_E165A, pET28a-speC_Q691A, pET28a-speC_N153D, pET28a-speC_N153E, pET28a-speC_D309E) подтверждали при помощи секвенирующего анализа.
Пример 3. Измерение активности мутантных Ферментов ODC (speC)
3-1. Получение мутантных Ферментов ODC
Каждый из мутантных векторов pET28a-speC, полученных в примере 2, трансформировали в Е.coli, имеющую тип гена DE3, для получения штамма, экспрессирующего фермент ODC.
Экспрессию мутантного вектора pET28a-speC осуществляли со ссылкой на руководство для системы рЕТ (Novagen). Подробно, единичные колонии соответствующих штаммов отбирали из сред для высевания LB и инокулировали в 3 мл жидкой среды LB (+50 мкг/мл канамицина) с последующей инкубацией при 37°С и 200 об./мин в течение 16 часов. Культуру повторно инокулировали в 15 мл свежей среды LB (+50 мкг/мл канамицина) и инкубировали в тех же самых условиях до достижения OD600, составляющей приблизительно 0,6. Затем немедленно добавляли IPTG (изопропилтиогалактозид) до конечной концентрации 0,5 мМ и инкубировали при 18°С и 180 об./мин в течение 20 часов для того, чтобы индуцировать экспрессию фермента.
После индуцирования экспрессии фермента полученные клетки подвергали ультразвуковой обработке и центрифугировали. Получающийся в результате супернатант использовали для первичного тестирования активности. Дополнительно, для характеристики фермента его очищали и затем подвергали вторичному тестированию активности. Фермент выделяли при помощи колонки Ni-NTA с использованием His-tag (гистидиновой метки), которую связывали с ферментом в векторе рЕТ. При очистке использовали набор Chelating Excellose spin kit (Bioprogen). Полученные таким образом ферменты ODC (дикого типа и мутантный speC), экспрессированные в растворенной форме, пропускали через 8% SDS PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), и таким образом выделяли из супернатанта.
3-2. Измерение активности мутантных ферментов ODC (speC)
Для оценки путресцин-конвертирующей активности посредством ODC с использованием орнитина в качестве субстрата измеряли активности ферментов ODC (speC дикого типа и мутантного speC), полученных в примере 3-1. Тестирование активности ODC для исследования путресцин-конвертирующей активности осуществляли со ссылкой на критерии, о которых ранее сообщалось (Vienozinskiene J, et al., Anal Biochem, 146: 180-183, 1985).
To есть, когда фермент ODC превращает одну молекулу орнитина в путресцин, тогда потребляется одна молекула воды и она молекула диоксида углерода, и один ион ОН- продуцируется вместе с путресцином. Таким образом, общее значение рН увеличивается (реакционная схема 1). Когда увеличенное значение рН измеряют при 559 нм с использованием фенолового красного, тогда измеряют поглощение индикатора рН. Поглощение увеличивается пропорционально увеличению рН. Количество путресцина опосредовано измеряли с использованием этого свойства.
Реакционная схема 1
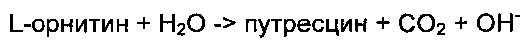
Для первичного тестирования активности ферментов ODC количественно измеряли общее количество белка в супернатантах до очистки и концентрации в супернатантах корректировали аналогичным образом. Реакционный раствор готовили с использованием 30 мкг супернатанта фермента, 10 мМ орнитина и 1,25 мкМ PLP, и затем 40 мкМ фенолового красного использовали для контроля изменения рН.
В результате измерения активность I163A и Е165А мутантных ферментов ODC продемонстрировала более высокую скорость продукции путресцина, нежели чем для дикого типа. Активность оставшихся 6 типов мутантов ODC V156A, D160A, Q691A, N153D, N153E и D309E продемонстрировала небольшие изменения поглощения при 559 нм (смотри фиг. 1).
Для характеристики двух мутантных ферментов ODC I163A и Е165А, которые были выбраны при первичном скрининге, их очищали при помощи His-tag и количественно измеряли, и затем измеряли скорость превращения в путресцин в зависимости от концентрации орнитина. Фермент ODC использовали в концентрации 10 мкг, и орнитин использовали в концентрации от 0,15 мМ до 10 мМ. В этом диапазоне изменение рН измеряли с использованием фенолового красного.
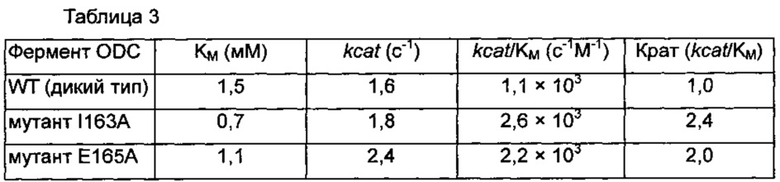
Эти результаты продемонстрировали, что модифицированные ферменты ODC I163A и Е165А, сконструированные в соответствии со структурным анализом ODC, продемонстрировали 53% и 27% уменьшения величины KM по сравнению с диким типом, соответственно, свидетельствуя о том, что их связывающие аффинности в отношении субстрата орнитина увеличивались. Кроме того, активность модифицированного ODC I163A и Е165А демонстрировала 12,5% и 50% увеличения величины kcat по сравнению с WT, соответственно, свидетельствуя о том, что способность превращать орнитин в путресцин также увеличивалась. Наконец, рассчитывали величину kcat/KM, демонстрирующую характеристику ферментативной активности. I163A и Е165А демонстрировали 2,4- и 2-кратные увеличения величины kcat/KM по сравнению с WT, соответственно (таблица 3).
Примеры 4. Оптимизация мутации ODC (speC)
Как подтверждено в примере 3, осуществлены мутации множества небольших аминокислотных остатков по аминокислоте (изолейцину) в положении 163 и аминокислоте (глутаминовой кислоте) в положении 165, которые представляют собой важные остатки для активности ODC. Мутации осуществляли тем же самым образом, как в примере 1, и использованные здесь праймеры приведены в следующей таблице 4. Дополнительно, единичные мутации осуществляли в положениях 163 и 165, соответственно, и затем получали двойные мутанты путем введения в каждое положение комбинацию мутаций, демонстрируя увеличенную активность ODC в соответствии с оценкой.

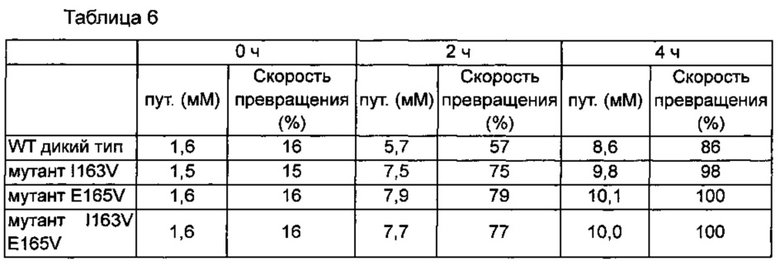
Мутанты ODC, полученные с использованием праймеров в таблице 4, очищали в соответствии со способом в примерах 2 и 3, и измеряли скорости превращения путресцина. Результаты измерения скоростей превращения путресцина полученными мутантами ODC приведены в следующей таблице 5.
Как представлено в таблице 5, когда аминокислотные остатки 163 и 165 вводили путем единичных мутаций глицина (G), серина (S) и валина (V), соответственно, тогда замена остатка 163 на серии и замена остатка 165 на валин демонстрировала 4,4- и 6,9-кратные увеличения величины kcat/KM по сравнению с диким типом, соответственно. На основе этого результата двойные мутации вводили в два остатка, и их активности исследовали. Неожиданно, ни двойная мутация комбинаций I163S и E165V, ни каждая единичная мутация не демонстрировали самую высокую активность, но двойная мутация замены обоих остатков 163 и 165 на валин демонстрировала 21,3-кратное увеличение активности по сравнению с диким типом.
В общем, увеличенные активности мутантов фермента ODC связаны с увеличением величины kcat/KM вследствие увеличения величины kcat, нежели чем уменьшения величины KM, свидетельствуя о том, что фермент ODC мутировал таким образом, что имеет структуру, увеличивающую скорость превращения в продукт путресцин, нежели чем структуру, увеличивающую связывающую аффинность субстрата орнитина для фермента.
Пример 5. Получение мутантного экспрессирующего фермент ODC штамма, использующего орнитин в качестве субстрата, и измерение превращения путресцина
Оценили то, оказывают ли практическое влияние мутантные ферменты ODC, в которых мутации оптимизированы в примере 4, на превращение орнитина в путресцин у микроорганизма.
Подробно, для осуществления эксперимента использовали штаммы, полученные путем введения Е.coli, имеющей генетический тип DE3, полученных мутантных векторов pET28a-speC. Единичные колонии соответствующих штаммов отбирали из сред для высевания LB и инокулировали в 3 мл жидкой среды LB (+50 мкг/мл канамицина), а затем инкубировали при 37°С и 200 об./мин в течение 16 часов. Культуру повторно инокулировали в 25 мл свежей среды LB (+50 мкг/мл канамицина и 0,2% глюкозу), и инкубировали до тех пор, пока OD600 не достигала от 0,5 до 0,6. Затем добавляли 0,5 мМ IPTG для того, чтобы вызвать экспрессию ODC (speC), и инкубировали при 18°С и 200 об./мин в течение 20 часов. Затем центрифугирование осуществляли для удаления супернатанта и сбора клеток. Клетки, полученные в форме осадка, ресуспендировали в 1Х минимальной среде М9 (3,37 мМ Na2HPO4, 2,2 мМ KH2PO4, 0,86 мМ NaCl, 0,94 мМ NH4Cl) для корректирования величины OD600 до 20. Дополнительно, 10 мМ орнитин в качестве субстрата и 0,5 мкМ PLP в качестве кофактора добавляли до конечного объема 10 мл. Реакции давали возможность протекать в условиях 25°С и 200 об./мин при встряхивании, и со временем осуществляли отбор образцов. Концентрацию образовавшегося в результате превращения путресцина определяли при помощи способа количественного измерения путресцина с использованием TNBS (тринитробензолсульфоновая кислота) (Ngo ТТ, et al., Anal Biochem, 160: 290-293, 1987).
В способе с TNBS супернатант, полученный путем центрифугирования образца культуры, разбавляли в 50 раз и использовали для осуществления анализа. Добавляли 1 мл 4 н. NaOH к 0,5 мл разбавленного образца, и затем дополнительно добавляли 2 мл 1-пентанола, и хорошо перемешивали. Центрифугирование осуществляли при 2000 об./мин в течение 5 минут, и затем 1 мл супернатанта добавляли в новую пробирку, содержащую 1 мл 0,1 М Na2B4O7 (рН 8,0), и ее содержимое хорошо перемешивали. Дополнительно добавляли 1 мл 10 мМ TNBS и хорошо перемешивали, и к смеси добавляли 2 мл DMSO (диметилсульфоксид) и перемешивали. Затем осуществляли центрифугирование, и поглощение получающегося в результате супернатанта измеряли при 426 нм.
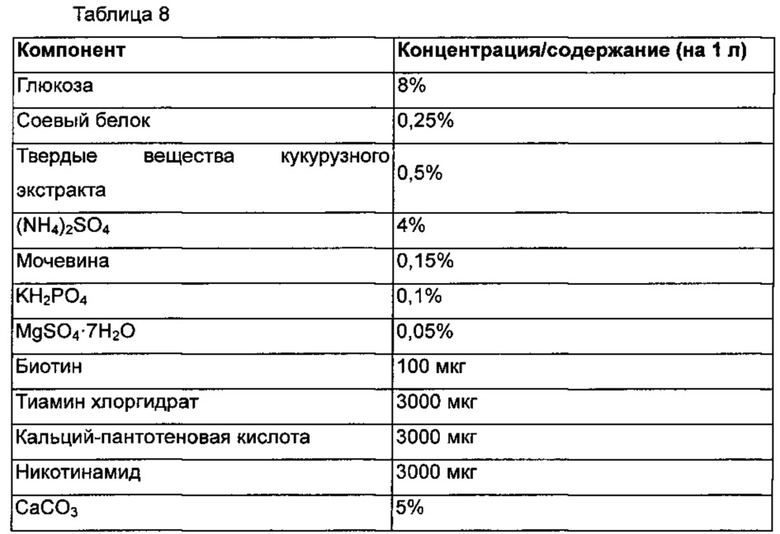
Измеряли превращение в путресцин диким типом и 3 типами мутантов ODC. В результате мутанты ODC демонстрировали приблизительно на 32%-39% увеличенную скорость превращения орнитина в путресцин в образце, собранном через 2 часа по сравнению с диким типом. Существовало небольшое различие в скорости превращения между мутациями ODC, и они не демонстрировали различие в активности, для которой очищенные мутанты ODC демонстрировали значительное различие в эксперименте in vitro. Тем не менее, дикий тип демонстрировал неполную реакцию даже через 4 часа, тогда как мутанты демонстрировали почти полную реакцию в течение 4 часов. В результате увеличенные активности мутантов ODC также подтверждали in vivo у штаммов с конвертирующим ферментом.
Пример 6. Получение продуцирующего путресцин штамма, имеющего мутантную ODC, и измерение продуктивности путресцина
Для проверки того, зависит ли путресциновая продуктивность от того, когда мутанты ODC, обладающие увеличенной путресцин-конвертирующей активностью, практически вводят в штамм, продуцирующий путресцин, измеряли путресциновую продуктивность.
6-1. Получение штамма, продуцирующего путресцин, имеющего мутантную ODC
На основе микроорганизма Corynebacterium sp. (KCCM11240P), который обладает улучшенной путресциновой продуктивностью путем ослабления активности NCgI1469 по сравнению с его эндогенной активностью, получали мутантный штамм путем замены speC дикого типа на ODC (speC), обладающий увеличенной путресцин-конвертирующей активностью в хромосоме.
Штамм Corynebacterium glutamicum (KCCM11240P), обладающий улучшенной путресциновой продуктивностью, представляет собой штамм, раскрытый в Международной заявке на патент № WO 2013/105827, и его получали с использованием микроорганизма Corynebacterium sp. (KCCM11138P), обладающего путресциновой продуктивностью, такой же как у материнского штамма, раскрытого в Международной заявке на патент № WO 2012/077995. Подробней, штамм получали путем клонирования N-концевой и С-концевой областей NCgI1469 в векторе pDZ, основываясь на основной последовательности гена NCgI1469 штамма АТСС13032, введения вектора в микроорганизм Corynebacterium sp. (KCCM11138P), обладающий путресциновой продуктивностью, путем электропорации, и затем высевания штамма на среде, содержащей канамицин (25 мкг/мл), а затем путем отбора. Успешное встраивание в хромосому вектора подтверждали путем отбора голубых колоний на среде, содержащей X-gal (5-бромо-4-хлоро-3-индолил-β-D-галактопиранозид). Первичный штамм с встроенной хромосомой выращивали в питательной среде, а затем распределяли разбавленный штамм на среде, содержащей X-gal и не содержащей антибиотик, и отбора белых колоний, которые появлялись в относительно низком отношении. Наконец, штамм с делецией гена NCgI1469 выбирали при помощи кроссовера. Полученный таким образом окончательный штамм KCCM11138P ΔNCgI1469 представляет собой сверхэкспрессирующий путресцин штамм, обладающий улучшенной путресциновой продуктивностью по сравнению с материнским штаммом KCCM11138P, где штамм KCCM11138P ΔNCgI1469 обладает делецией гена, кодирующего NCgI1469, который представляет собой белок, вовлеченный в путь расщепления путресцина до N-ацетилпутресцина в клетках.
Подробно, фрагменты ДНК мутантов ODC (speC), полученых в примерах 2 и 4 амплифицировали с использованием праймеров speC_start (BamHI)_5 и speC_stop (XbaI)_3, приведенных в следующей таблице 7. В частности, полученные мутантные векторы pET28a-speC (I163S, I163V, I163S E165V) в качестве матриц и два праймера speC_start (BamHI)_5 и speC_stop (XbaI)_3, приведенные в следующей таблице 7, использовали для осуществления ПЦР.

Генные фрагменты, полученные путем ПЦР, и вектор pDZ обрабатывали ферментами рестриктазами BamHI и XbaI (37°С, 3 часа), и затем генные фрагменты мутантов speC встраивали в вектор pDZ при помощи общего способа лигирования, соответственно. Полученные таким образом эти рекомбинантные векторы для хромосомного встраивания (pDZ-speC_I163S, pDZ-speC_I163V, pDZ-speC_I163S E165V) подтверждали путем секвенирующего анализа.
Для получения штаммов, в которых мутанты speC встроены в хромосому, каждый из полученных рекомбинантных векторов pDZ-speC_I163S, pDZ-speC_I163V, и pDZ-speC_I163S E165V, трансформировали в штамм KCCM11240P путем электропорации и затем распределяли на среде для высевания BHIS (37 г/л сердечно-мозгового экстракта, 91 г/л сорбита и 2% агар на 1 L + 25 мкг/мл канамицина).
Успешное встраивание в хромосому вектора определяли путем проверки появления голубых колоний на твердой среде, содержащей X-gal (5-бромо-4-хлоро-3-индолил-β-D-галактопиранозид). Первичный штамм со встраиванием в хромосому выращивали в питательной среде с встряхиванием (30°С, 8 часов) с последующим серийным разбавлением и распределением на твердой среде, содержащей X-gal. Большинство колоний были голубыми, тогда как белые колонии появлялись с относительно низким отношением. Из отобранных колоний наконец получали штаммы, имеющие мутанты speC в хромосоме путем вторичного кроссовера. Эти штаммы наконец идентифицировали путем секвенирующего анализа мутантов. Идентифицированные штаммы были обозначены как KCCM11240P::speC_I163S, KCCM11240P::speC_I163V и KCCM11240P::speC_I163S E165V. Из них, KCCM11240P::speC_I163S E165V был обозначен как Corynebacterium glutamicum СС01-0578 и депонирован в Корейском центре культур микроорганизмов (KCCM) 10 июня 2013 года под инвентарным номером KCCM11425P в соответствии с Будапештским договором.
6-2. Измерение продуктивности по путресцину штамма, продуцирующего путресцин, имеющего мутант ODC
Для исследования действия мутанта ODC (speC) на продуктивность по путресцину штамма, продуцирующего путресцин, штаммы, полученные в примере 6-1, оценивали в отношении продуктивности по путресцину.
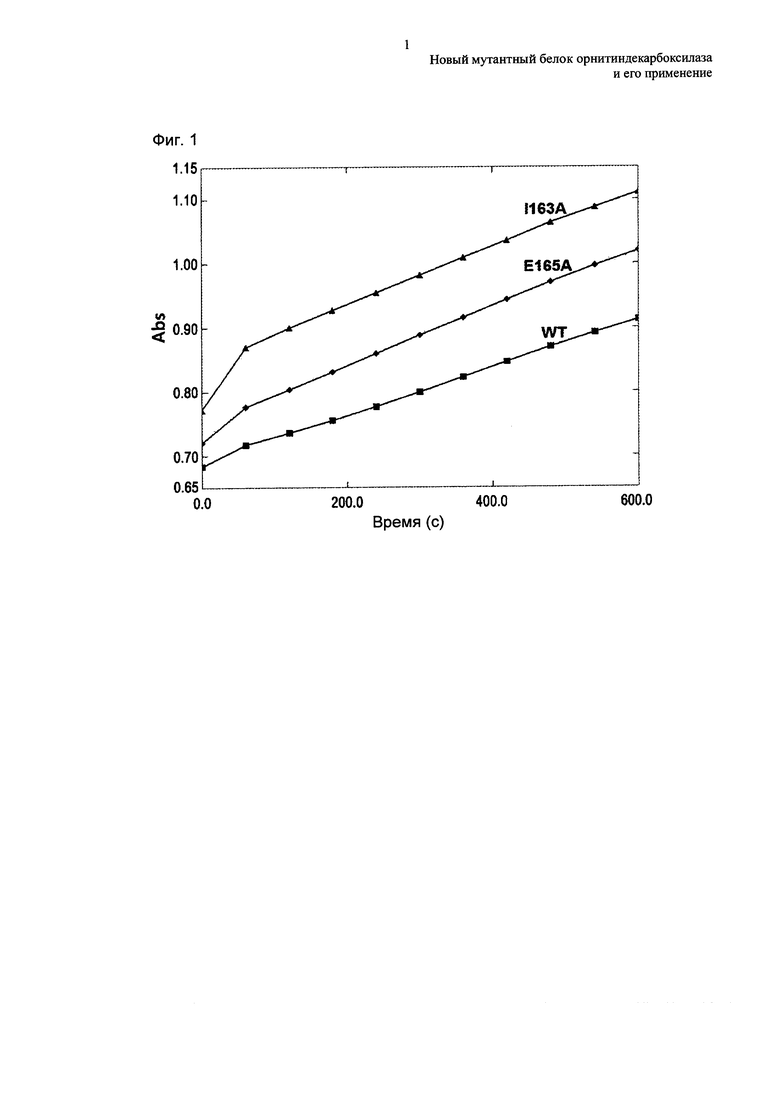
Подробно, полученные штаммы выращивали в среде для высевания СМ, содержащей 1 мМ аргинин (1% глюкоза, 1% полипептон, 0,5% дрожжевой экстракт, 0,5% мясной экстракт, 0,25% NaCl, 0,2% мочевина, 100 мкл 50% NaOH, 2% агар, рН 6,8 на 1 л) при 30°С в течение 16 часов, и затем петлю с клеточной культурой инокулировали в 25 мл титрованной среды в соответствии со следующей таблицей 8, и выращивали при встряхивании при 200 об./мин при 30°С в течение 24 часов. Все полученные штаммы выращивали с добавлением 1 мМ аргинина в среду во время ферментации.
Как представлено в таблице 9, каждый штамм вводили с мутантом ODC (speC), обладающим улучшенной активностью, демонстрирующим 37%-105% увеличение путресциновой продуктивности в течение 24 часов.
Эти результаты демонстрируют что штамм, продуцирующий путресцин, имеющий мутантную ODC, способен продуцировать высокую концентрацию путресцина относительно потребления сахара по сравнению с известным штаммом.

На основе вышеприведенного описания специалисту в данной области техники понятно, что настоящее изобретение может быть осуществлено в отличающейся специфической форме без изменения его технической сущности или его важных характеристик. Таким образом, понятно, что вышеприведенное воплощение не является ограничивающим, а иллюстративным во всех аспектах. Объем изобретения определен в формуле изобретения, нежели чем в предшествующем ей описании, и, таким образом, предполагается, что все изменения и модификации, которые оказываются в пределах границ и объемов формулы изобретения, или эквивалентах таких границ и объемов, охвачены формулой изобретения.




























































Предложены модифицированный белок орнитиндекарбоксилаза (ODC), обладающий улучшенной продуктивностью по путресцину, и его применение. Изолейцин в положении 163 от N-конца ODC, имеющей аминокислотную последовательность, представленную в SEQ ID NO: 1, заменен на аминокислотный остаток, меньший, чем изолейцин, и/или глутаминовая кислота в положении 165 от N-конца ODC, имеющей аминокислотную последовательность, представленную в SEQ ID NO: 1, заменена на аминокислотный остаток, меньший, чем глутаминовая кислота. Предложен полинуклеотид, кодирующий модифицированный белок ODC, обладающий улучшенной путресцин-конвертазной активностью. Также предложены экспрессирующий вектор, содержащий указанный полинуклеотид, и трансформант, трансформированный указанным вектором, и рекомбинантный микроорганизм, обладающий улучшенной путресцин-продуцирующей активностью благодаря замене нативного белка ODC на указанный модифицированный белок ODC у микроорганизма Corynebacterium sp., обладающего путресцин-продуцирующей активностью. Предложены также варианты способа получения путресцина. Группа изобретений обеспечивает продукцию путресцина с высоким выходом. 7 н. и 7 з.п. ф-лы, 1 ил., 9 табл., 6 пр.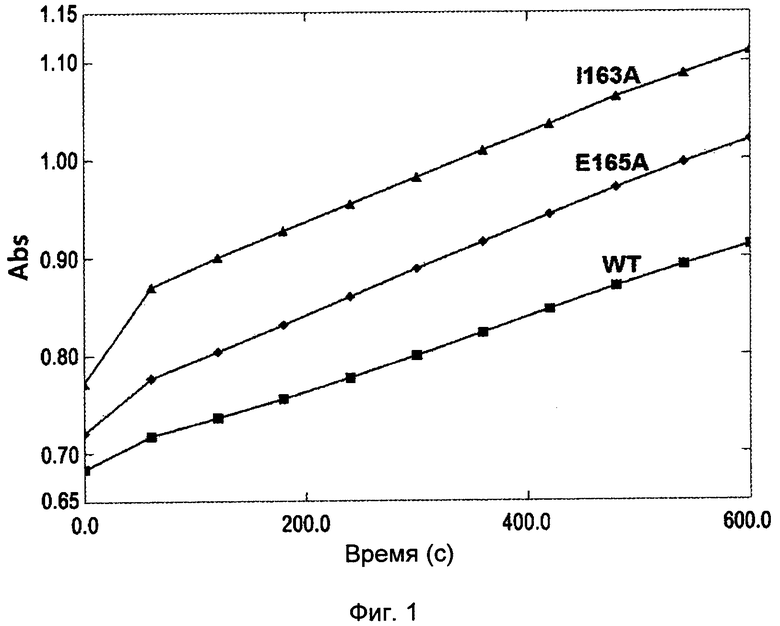
1. Модифицированный белок орнитиндекарбоксилаза (ODC), где изолейцин в положении 163 от N-конца ODC, имеющей аминокислотную последовательность, представленную в SEQ ID NO: 1, заменен на аминокислотный остаток, меньший, чем изолейцин, и/или глутаминовая кислота в положении 165 от N-конца ODC, имеющей аминокислотную последовательность, представленную в SEQ ID NO: 1, заменена на аминокислотный остаток, меньший, чем глутаминовая кислота.
2. Модифицированный белок ODC по п.1, отличающийся тем, что глутаминовая кислота в положении 165 заменена на аланин, глицин, серин или валин.
3. Модифицированный белок ODC по п.1, отличающийся тем, что изолейцин в положении 163 заменен на аланин, глицин, серин или валин.
4. Модифицированный белок ODC по п.1, отличающийся тем, что изолейцин в положении 163 и глутаминовая кислота в положении 165 заменены на аланин-аланин, аланин-валин, серин-валин или валин-валин соответственно.
5. Модифицированный белок ODC по п.1, имеющий любую аминокислотную последовательность в соответствии с SEQ ID NO: 34-57.
6. Полинуклеотид, кодирующий модифицированный белок ODC, обладающий улучшенной путресцин-конвертазной активностью, по любому из пп.1-5.
7. Экспрессирующий вектор, содержащий полинуклеотид, кодирующий модифицированный белок ODC по любому из пп.1-5.
8. Трансформант, трансформированный вектором по п.7, отличающийся тем, что указанный трансформант представляет собой микроорганизм, обладающий путресцин-конвертазной активностью.
9. Способ получения путресцина, включающий стадию взаимодействия L-орнитина, смеси, содержащей L-орнитин, или L-орнитиновой культуральной жидкости с модифицированным белком ODC по любому из пп.1-5.
10. Способ по п.9, отличающийся тем, что модифицированный белок ODC представляет собой очищенный модифицированный белок ODC или культуральную жидкость микроорганизмов, содержащую модифицированный белок ODC.
11. Рекомбинантный микроорганизм, обладающий улучшенной путресцин-продуцирующей активностью благодаря замене нативного белка ODC на модифицированный белок ODC по любому из пп.1-5 у микроорганизма Corynebacterium sp., обладающего путресцин-продуцирующей активностью.
12. Рекомбинантный микроорганизм по п.11, отличающийся тем, что микроорганизм Corynebacterium sp. представляет собой Corynebacterium glutamicum.
13. Способ получения путресцина, включающий стадии:
(а) культивирования рекомбинантного микроорганизма по п.11; и
(б) выделения путресцина из культуры, полученной на стадии (а).
14. Способ по п.13, где трансформант по п.11 представляет собой Corynebacterium glutamicum.
| WO 2006005603 A1, 19.01.2006 | |||
| МИКРООРГАНИЗМ, ОБЛАДАЮЩИЙ СПОСОБНОСТЬЮ К ПРОДУКЦИИ ПУТРЕСЦИНА В ВЫСОКОЙ КОНЦЕНТРАЦИИ, СПОСОБ ПОЛУЧЕНИЯ ЭТОГО МИКРООРГАНИЗМА И СПОСОБ ПРОДУКЦИИ ПУТРЕСЦИНА С ИСПОЛЬЗОВАНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2009 |
|
RU2433180C2 |
| SCHNEIDER J | |||
| ET AL | |||
| Putrescine production by engineered Corynebacterium glutamicum // Appl Microbiol Biotechnol., 2010 Oct; 88(4): 859-68 | |||
| PAKULA A.A | |||
| ET AL | |||
| Genetic analysis of protein stability and function | |||
| Anna | |||
| Rev | |||
| Genet | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |

