Область техники, к которой относится изобретение
Настоящее изобретение относится к получению L-треонина с использованием микроорганизмов. Более конкретно, настоящее изобретение относится к способу получения L-треонина с высоким выходом с использованием микроорганизма, в котором ген (tdc) треониндегидратазы геномной ДНК микроорганизма частично дезактивируют с использованием рекомбинантной технологии, вследствие чего существенно повышается выход L-треонина.
Описание уровня техники
L-треонин, являющийся представителем незаменимых аминокислот, широко используется в качестве кормовых и пищевых добавок, а также в качестве жидких сред и синтетических материалов медицинского и фармацевтического назначения. L-треонин получают ферментацией с использованием синтетических мутантов - производных Escherichia Coli, Corynobacterium, Serratia, и Providencia дикого типа. Известно, что такие мутанты включают устойчивые к аналогам аминокислот и лекарственным препаратам мутанты, а также их синтетические формы, наделенные ауксотрофными свойствами по диаминопимелиновой кислоте, метионину, лизину или изолейцину (выложенная заявка на патент Японии № hei 2-219582, Appl., Microbiolo. Biotechnol., 29, 550-553 (1988) и корейская патентная публикация №92-8365). В заявке на патент Кореи №90-22965 описывается продуцирующий L-треонин штамм TF4076 (KFCC10718), являющийся ауксотрофным по метионину и устойчивым к действию аналогов треонина (AHV: α-амино-β-гидроксивалериановая кислота), аналогов лизина (АЕС: S-(2-аминоэтил)-L-цистеин), аналогов изолейцина (α-аминомасляная кислота) и метиониновых аналогов (этионин).
Общий подход к повышению уровня экспрессии конкретного гена предусматривает использование плазмиды, делающей большее число копий с целью увеличения числа генов в микроорганизме (Sambrook et al., Molecular cloning, Second Edition, 1989, 1.3-1.5). Ген-мишень интегрируется в плазмиду, а микроорганизм-хозяин трансформируется рекомбинантной плазмидой с целью увеличения числа генов в микроорганизме-хозяине в соответствии с копийностью плазмиды. Об определенном успехе в подходе такого типа, предпринятом для увеличения продуктивности треонина, сообщается в US Patent №5538873. Однако большинство методов, использующих такие рекомбинантные плазмиды, характеризуются сверхпродукцией конкретного гена, что нежелательно для микроорганизма-хозяина и создает проблемы, связанные с неустойчивостью плазмиды, вследствие чего может происходить потеря плазмиды в ходе культивирования рекомбинантного штамма.
Для решения такой проблемы были предложены методы, предусматривающие добавление антибиотиков в культурную среду или использование плазмиды с регулируемой экспрессией (Sambrook et al., Molecular cloning, Second Edition, 1989, 1.5-1.6&1.9-1.11). Согласно методу, в котором для получения конкретного продукта используют плазмиду с регулируемой экспрессией, культивирование клеток проводят в условиях отсутствия экспрессии на стадии роста с целью минимизации ущерба для микроорганизма-хозяина, и после полного развития микроорганизма индуцируется временная (транзиторная) экспрессия. Однако большинство плазмид с регулируемой экспрессией может использоваться лишь в том случае, когда конечный продукт представляет собой белок.
Продукция первичных метаболитов тесно связана с ростом микроорганизмов, вследствие чего трудно повысить выход первичных метаболитов, если на стадии роста не происходит экспрессии генов-мишеней. Именно это происходит при продуцировании треонина, являющегося первичным метаболитом.
Для компенсации такого недостатка специфический ген биосинтеза треонина включают в хромосомную ДНК с целью продуцирования треонина (US Patent №5939307). Однако такой подход предусматривает замену хромосомного гена на ген с заменой на индуцибельный промотор, и трудно ожидать, что это может существенно увеличить экспрессию гена треонинового оперона.
В связи с этим сущность настоящего изобретения состоит в инактивации треонин-дегидратазного (tdc) гена в хромосоме, участвующей в одном из путей деградации треоинина, с целью ингибирования деградации треонина при сохранении других видовых функций микроорганизма-хозяина и, тем самым, обеспечения повышенного выхода треонина.
Большинство современных генно-инженерных методов, используемых для повышения выхода треоинина, сфокусированы на биосинтетическом пути его получения, начинающегося с оксалоацетата. Однако в настоящем изобретении также предусматривается использование активности треонин-дегидратазного (tdc) гена, вовлеченного в путь деградации треонина, с целью эффективного и существенного увеличения выхода L-треонина.
Раскрытие изобретения
Цель настоящего изобретения, направленная на решение отмеченных выше проблем, заключается в разработке способа высокоэффективного продуцирования L-треонина, исключающего проблемы, связанные с нестабильностью плазмиды и ингибированием микробного роста, возникающие при использовании штаммов, несущих рекомбинантную плазмиду, и обеспечивающего существенное увеличение продуктивности L-треонина в результате подавления активности треонин-дегидратазы.
Для достижения целей настоящего изобретения разработана рекомбинантная плазмида, включающая инактивированный ген tdc (треониндегидратаза).
Для достижения целей настоящего изобретения, в соответствии с одним из его воплощений, tdc-оперон треонин-продуцирующего микроорганизма может инактивироваться путем расщепления сайта в tdc В и tdc С tdc-оперона и вставки кассеты с антибиотическим маркером в сайт расщепления. Предпочтительным треонин-продуцирующим микроорганизмом является штамм E.coli.
В соответствие с другим воплощением настоящего изобретения штамм E.coli может обладать устойчивостью в отношении аналогов треонина, аналогов лизина, аналогов изолейцина и аналогов метионина. Согласно другому воплощению настоящего изобретения одна или более дополнительных копий каждого из генов фосфоенолпируваткарбоксилазы (ррс) и треонинового оперона интегрируются в хромосомную ДНК штамма E.coli. Предпочтительно использовать E.coli с названием pGmTN-PPC12 и каталожным № КССМ-10326.
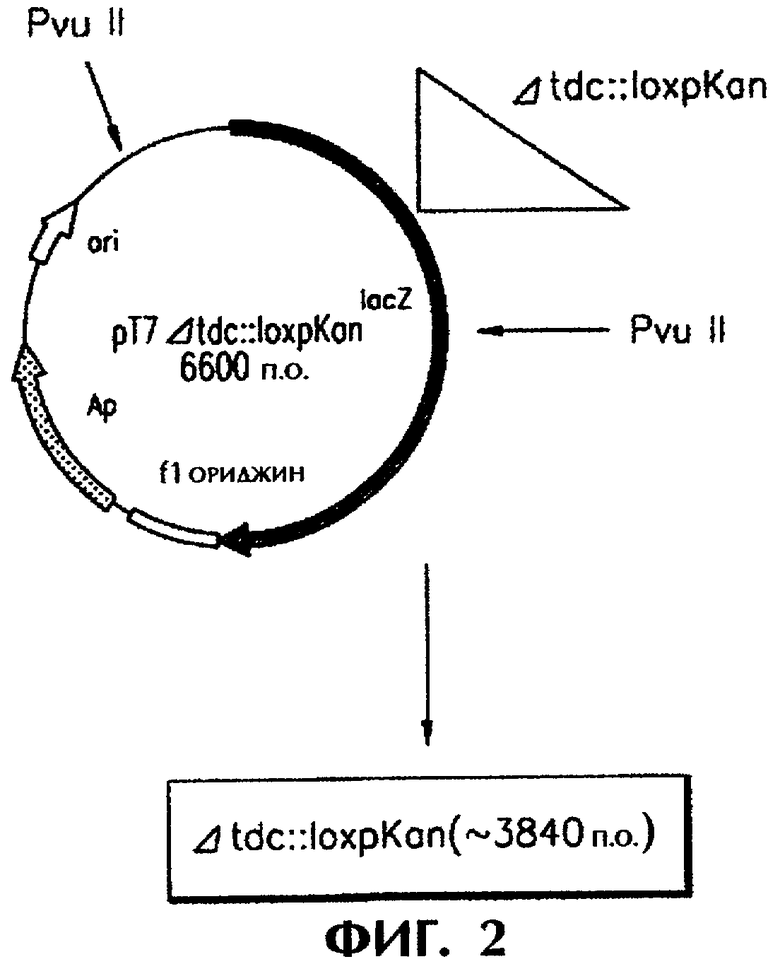
Кроме этого, настоящее изобретение обеспечивает штамм E.coli, трансформированный описанной выше рекомбинантной плазмидой. Фрагмент гена tdc В и tdc С, содержащий антибиотический маркер, который происходит из рекомбинантной плазмиды, может быть вставлен в геном штамма E.coli. Предпочтительный фрагмент гена tdc В и tdc С, включающий антибиотическитй маркер, представляет собой Δtdc::loxpKan, изображенный на фигуре 2. Наиболее предпочтительный штамм E.coli имеет название TRN212 и каталожный номер КССМ-10353.
Цель настоящего изобретения достигается с помощью способа получения L-треонина, в котором используют указанный выше штамм E.coli, имеющий название TRN212 и каталожный номер КССМ-10353.
Далее приводится более подробное описание настоящего изобретения.
Известно несколько путей метаболизма L-треонина (Neihardt FC et al. (eds) Escherichia coli и Salmonella: Cellular and Molecular Biology, 2nd edn. ASM Press., Washington D.C., pp.369-370), три из которых являются основными: первый путь с участием треонин-дегидрогеназы, катализирующей разложение треонина с образованием α-амино-β-кетобутирата, который затем превращается в ацетил-СоА и глицин или подвергается дальнейшему разложению с образованием аминоацетона и превращением в пируват; второй путь с участием треониндегидратазы, дающей α-кетобутират, который метаболизируется до пропионил-СоА и далее до сукцинил-Со-А, являющегося интермедиатом цикла трикарбоновых кислот (ЦТК); и третий путь, протекающий с участием треонинальдолазы.
Настоящее изобретение отличается тем, что с целью максимального накопления L-треонина в пути метаболизма L-треонина подавляется активность треонин-дегидратазы.
В способе продуцирования L-треонина согласно настоящему изобретению в качестве штамма, продуцирующего L-треонин, используют штамм pGmTN-PPC12 (КССМ-10136, Korean Patent Application №01-6976, поданная заявителем по настоящей заявке), производный от E.coli TF4076 (KFCC10718, Korean Patent Application №90-22965). Штамм pGmTN-PPC12 (КССМ-10136) получали инсерцией гена фосфоенолпируваткарбоксилазы (ррс) из хромосомы штамма-продуцента L-треонина E.coli TF4076 (KFCC10718) с использованием полимеразной цепной реакции (ПЦР) и треонинового оперона, клонированного из той же хромосомы, в хромосому штамма-хозяина E.coli TF4076, вследствие чего в штамме TF4076 содержится удвоенное количество гена ррс и треонинонового оперона относительно их количества в хромосомной ДНК. Штамм pGmTN-PPC12 способен усиливать экспрессию гена ррс, участвующего в конверсии фосфоенолпирувата (PEP) в предшественник биосинтеза треонина, оксалоацетат, а также генов, кодирующих ферменты, участвующие в синтезе треонина из оксалоацетата, включающих thrA (аспартокиназа I-гомосериндегидрогеназа), thrB (гомосеринкиназа) и thrC (треонинсинтаза) и, таким образом, способен повышать выход продукции L-треонина. Штамм pGmTN-PPC12 депонирован в Korean Collection for Type Cultures (KCTC) 5 января 2001 г. и ему присвоен каталожный номер КССМ 10236.
Согласно настоящему изобретению ген треониндегидратазы (tdc), участвующей в пути метаболизма треонина, из pGmTN-PPC12 (КССМ-10136) подвергают инактивации, тем самым повышая выход L-треонина.
Известно, что треониндегидратаза представляет собой оперон, экспрессируемый в условиях низкого уровня содержания кислорода и высокой концентрации треонина. Экспрессия tdc-оперона также индуцируется при низких содержаниях глюкозы, что имеет место во второй половине ферментации. В связи с этим для обнаружения высокопроизводительных штаммов-продуцентов треонина необходимо инактивировать треониндегидратазу.
Краткое описание чертежей
Фигура 1 изображает способ конструирования рекомбинантной плазмиды pT7Blue/tdc.
Фигура 2 изображает способ конструирования рекомбинантной плазмиды pT7Δtdc::loxpKan и фрагмента ДНК Δtdc:: loxpKan; a
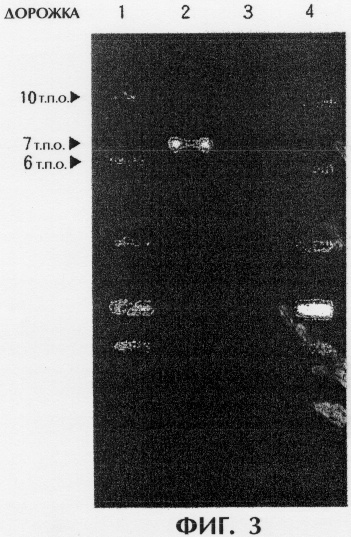
на фигуре 3 представлены результаты анализа саузерн-блоттинг, полученные визуальным сканированием с использованием системы формирования изображений FLA-5000 (FUJIFILM), где зоны 1 и 4 показывают размер маркеров, а зоны 2 и 4 относятся к TRN212(KCCM-10353) и хозяйскому штамму pGmTN-PPC12.
Осуществление изобретения
Описание предпочтительного варианта реализации изобретения
Ниже приведено подробное описание способа получения L-треонина согласно настоящему изобретению.
1. Конструирование рекомбинантной плазмиды
Хромосомную ДНК выделяли из штамма-продуцента треонина. Используя хромосомную ДНК в качестве матрицы, получили рекомбинантную плазмиду pT7Blue/tdc, включающую tdc В и tdc A tdc оперона. Для этой цели можно использовать любой клонирующий вектор без всякого ограничения, однако предпочтительным является клонирующий вектор рТ7Вluе.
Рекомбинантную плазмиду pT7Δtdc::loxpKan, содержащую дефектный ген tdc, получали интеграцией канамицин-устойчивого фрагмента гена, содержащего сайты lox р, в рекомбинантную плазмиду pT7Blue/tdc.
2. Интеграция рекомбинантной плазмиды и скрининг
Е.coli трансформировали рекомбинантной плазмидой pT7Δtdc::loxpKan. Плазмидную ДНК выделяли и переваривали в присутствии рестрикционного фермента, и фрагменты ДНК (Δtdc::loxpKan) выделяли из перевара электрофорезом. Этот процесс проиллюстрирован на фигуре 2.
Штамм-продуцент треонина подвергали трансформации полученным фрагментом ДНК Δtdc::loxpKan. Трансформацию осуществляли посредством гомологической рекомбинации, позволяющей проводить замену природной области гена tdc штамма Е.coli фрагментом рекомбинантной ДНК pT7Δtdc::loxpKan.
Полученный в результате трансформант инокулировали на канамицин-содержащую твердую среду с целью сбора колоний. Наконец, получали целевой рекомбинантный штамм, содержащий инактивированный ген tdc.
Ген tdc в рекомбинантном штамме остается дефицитным, несмотря на пролиферацию штамма, при отличной продуктивности L-треонина, превышающей на 20% продуктивность штамма-хозяина.
Рекомбинантная плазмида может быть выделена из трансформированного штамма с использованием известных методов, например с помощью раствора щелочи (Sambrook et al., Molecular cloning, vol.1, 1.25-1.28). Так, раствор 1 (50 мМ глюкозы, 25 мМ Трис-HCl и 10 мМ ЭДТА) добавляют для ослабления клеточной мембраны трансформанта, раствор 2 (0,2N NaOH, 1% SDS) добавляют для разрушения клеточной мембраны и денатурации клеточных компонентов - белка и хромосомы, а раствор 3 (5М ацетата калия, уксусная кислота) добавляют для коагуляции всех компонентов, за исключением рекомбинантной плазмиды. Фракцию, содержащую рекомбинантную плазмиду, отделяют центрифугированием, после чего рекомбинантную плазмиду осаждают добавлением этанола и собирают.
Далее настоящее изобретение обсуждается со ссылкой на следующие примеры. Следующие ниже примеры приведены лишь в целях иллюстрации и не ограничивают объем изобретения.
Пример 1: конструирование рекомбинантной плазмиды и нокаут гена tdc
Геномную ДНК pGmTN-PPC (КССМ-10136), штамма-продуцента треонина, выделяли с использованием системы QIAGEN Genomic-tip. Фрагмент гена размером около 3,1 т.п.о. tdc-оперона (5295 п.о.), содержащий tdc В и tdc С, амплифицировали посредством полимеразной цепной реакции (ПЦР) с использованием геномиой ДНК в качестве матрицы. В качестве праймеров использовали 5'-agg agg gga tec ggt atg tct tct gag gcg-3' и 5'-agg agg gaa ttc ate ggc aac agg cac ag-3'. ПЦР осуществляли за 30 циклов амплификации, причем каждый цикл включал денатурацию в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 56°С и достраивание в течение 3 минут 30 секунд при 72°С.
Продукты ПЦР подвергали электрофорезу в 0,7% агарозном геле с целью элюирования полос желаемого размера. Элюированные полосы в течение ночи при 16°С подвергали лигированию по тупым концам с клонирующим вектором pT7Blue (Novagen Со) с получением рекомбинантной плазмиды pT7Blue/tdc (см. фигуру 1). E.coli DH5α трансформировали полученной рекомбинантной плазмидой pT7Blue/tdc, высевали на твердую среду, содержащую 50 мг/л ампициллина и инкубировали при 37°С в течение ночи.
Колонии отбирали с помощью зубочистки из культуры, инокулировали в 3 мл жидкой среды и инкубировали в течение ночи при перемешивании со скоростью 200 об/мин. Плазмидную ДНК выделяли из культуры с использованием мини-препаративного набора QIAGEN и определяли размер плазмидной ДНК. Плазмидную ДНК переваривали рестрикционными ферментами Sma I и Pst I и подвергали электрофорезу в 0,7% агарозном геле для того, чтобы убедиться, что ее ориентация представляет собой tdc R-A-B-C. Идентифицированную плазмидную ДНК pT7Blue/tdc переваривали рестрикционными ферментами Bgl II и Нра I (каждый размером около 1,1 т.п.о.), и перевар наносили на 0,7% агарозный гель с целью элюирования полос размером 4,9 т.п.о. Для получения тупых концов проводили реакцию между элюированными полосами и фрагментом Кленова. Канамицин-устойчивый фрагмент гена размером около 1,7 т.п.о., содержащий сайт lox р, полученный путем реакции плазмиды pUG6 (Ulrich et al., A new efficient gene disruption cassette for repeated use in budding yeast, Nucleic Acids Research, 1996, 24, pp.2519-2524) с рестрикционными ферментами Hinc II и EcoR V, лигировали с изолированным фрагментом pT7Blue/tdc, используя метод лигирования по тупым концам, в результате чего получали рекомбинантную плазмиду pT7Δtdc::loxpKan.
Пример 2: скрининг штамма, интегрированного с рекомбинантной плазмидой
E.coli DH5α трансформировали рекомбинантной плазмидой pT7Δtdc::loxpKan, высевали на твердую среду, содержащую 50 мг/л ампициллина и 15 мг/л канамицина и инкубировали при 37°С в течение ночи. Колонии отбирали из культуры, инокулировали в 3 мл жидкой среды, содержащей ампициллин и канамицин, и инкубировали в течение ночи при перемешивании со скоростью 200 об/мин. Плазмидную ДНК выделяли из культуры с помощью мини-препаративного набора QIAGEN и определяли размер плазмиды. Плазмидную ДНК переваривали рестрикционным ферментом и наносили на 0,7% агарозный гель для определения ориентации. Идентифицированную плазмидную ДНК переваривали рестрикционным ферментом Pvu II и подвергали электрофорезу в 0,7% агарозном геле с целью элюирования фрагмента ДНК (Δtdc::loxpKan) размером около 3840 т.п.о. Штамм-продуцент треонина pGmTN-PPC12 трансформировали ДНК-фрагментом Δtdc::loxpKan методом электропорации и высевали на канамицин-содержащую твердую среду с целью скрининга колоний, получая рекомбинантные штаммы с инактивированным геном tdc.
Пример 3: сравнение полученных рекомбинантных штаммов по продуктивности треонина при культивировнии в колбе
Тридцать отдельных колоний рекомбинантных штаммов, культивированных на твердой среде примера 2, содержащей канамицин, подвергали скринингу в Эрленмейровских колбах на продуктивность по L-треонину с использованием сред для титрования треонина. Состав используемой в каждом случае среды для титрования треонина приведен в таблице 1.
Состав титрованной по треонину среды
Отдельные колонии культивировали в инкубаторе на твердой среде LB при 32°С в течение ночи. 25 мл среды для титрования инокулировали петлей каждой культуры и проводили инкубацию в течение 48 часов при 32°С и скорости перемешивания 250 об/мин. Результаты анализа представлены в таблице 2. Было установлено, что 28 из тридцати колоний новых рекомбинантных штаммов демонстрируют отличную продуктивность, включая восемь колоний, продуцирующих 26 г/л или более треонина, в сравнении со штаммом-хозяином, pGmTN-PPC12, продуцирующим 23 г/л треонина. Был зарегистрирован рекомбинантный штамм, обозначенный как "TRN212", обеспечивающий наивысшую продуктивность по L-треонину, 27 г/л, и выход на 17,4% больший, чем штамм-хозяин. Мутантный штамм TRN212 был депонирован 19 февраля 2002 г. в Korean Collection for Type Cultures (КСТС), и ему был присвоен каталожный номер КСТС-10353.
Результаты титрования рекомбинантных штаммов в колбе
Пример 4: Подтверждение нокаута гена tdc с использованием Саузерн-блоттинг-анализа
Саузерн-блоттинг проводили с целью подтверждения специфического нокаута гена tdc в рекомбинантном штамме, полученном в примере 3. Штамм-хозяин pGmTN-PPC12 и рекомбинантный штамм TRN212 (КССМ-10353) инкубировали в течение ночи при перемешивании со скоростью 200 об/мин в 30 мл жидкой среды, и геномную ДНК выделяли с использованием набора QIAGEN Genomic kit 20. Выделенную геномную ДНК переваривали рестрикционным ферментом EcoRI и подвергали электрофорезу в 0,7% агарозном геле с целью разделения фрагментов ДНК по размеру. Разделенные по размеру фрагменты ДНК закрепляли на нейлоновой мембране (YOUNG Sci. Biodyne В membrane) путем капиллярного переноса в течение ночи (Molecular cloning. Vol.1, 6.31-6.38) подвергнутого электрофорезу геля с последующим высушиванием и УФ-облучением (120 мДж/см2, SpectroLinker™) с целью иммобилизации фрагментов ДНК на нейлоновой мембране (Molecular Cloning, Vol.1, 6.45). Фрагменты ДНК, прикрепленные к нейлоновой мембране, подвергали 2-х часовой предгибридизации в предгибризационном буфере I (Roche# 1093657) при 55°С, после чего гибридизовали в гибридизационной печи (BAMBINO 230300) при добавлении ранее полученного денатурированного ДНК-зонда.
Используемый денатурированный зонд получали следующим образом. Плазмиду pUG6, выделенную с использованием набора QIAGEN, переваривали рестрикционными ферментами Hind II и EcoR V с получением канамицин-устойчивого фрагмента гена размером около 1,7 т.п.о., содержащим сайт lox р. Такой фрагмент гена нагревали в кипящей воде при 100°С в течение 5 минут и охлаждали льдом в течение 5 минут с целью генерации одноцепочных ДНК. Проводили реакцию с одноцепочными ДНК при 37°С в течение ночи с использованием набора DIG Labelling and Detection kit (Roche #1093657) с целью получения ДНК-зондов с интегрированным DIG-UDP.
После гибридизации фрагменты ДНК, неспецифически связанные с мембраной, удаляли с использованием промывочных растворов I и II (Roche #1093657). После этого в целях маскировки мембраны обрабатывали в течение 30 минут при комнатной температуре предгибридизационным буфером (Roche #1093657) 2 и в течение 30 минут при комнатной температуре проводили реакцию с анти DIG-антителом, специфически связывающимся с DIG-UTP. Анти-DIG антитело, неспецифически связанное с мембраной, удаляли с использованием промывочного раствора III (Roche #1093657), и мембрану подвергали окрашиванию с помощью набора Labelling and Detection kit (Roche #1093657) до проявления окрашенных полос. Полученные результаты представлены на фигуре 3. В отсутствие канамицинового гена полосы не обнаруживались для штамма-хозяина pGmTN-РРС12 (зона 1). В отличие от этого, как и ожидалось, на дорожке 2 для нового рекомбинантного штамма настоящего изобретения, TRN212 (КССМ-10353), наблюдалась полоса размером около 7 т.п.о. Фрагмент гена tdc размером около 5,3 т.п.о., получающийся при переваривании рестрикционным ферментом EcoR V, и ген устойчивости к канамицину размером около 1,7 т.п.о. составляют около 7,0 т.п.о. Дорожки 1 и 4 представляют маркеры размера.
Пример 5: сравнение продуктивности по L-треонину с использованием ферментеров
Проводили сравнение продуктивности по L-треонину в ферментере емкостью 5 л при использовании штамма-хозяина pGmTN-PPC12 и рекомбинантного штамма TRN212 (КССМ-10353), содержащего инактивированный ген tdc, который был подвергнут скринингу в примере 2 и тесту на подтверждение нокаута гена в примере 4. Состав исходной среды приведен в таблице 3. LB среда, дополненная 10 г/л глюкозы и 0,1 г/л L-метионина, использовалась в качестве посевной среды, а исходный объем инокулята в ферментер задавали как 3-5% об. целевой исходной культуры. Каждый раз глюкозу добавляли до конечной концентрации 5 мас.%, проводили добавление всего 6 раз, при этом добавляли 1 мас.% КН2PO4. Каждое добавление глюкозы определялось исчерпанием глюкозы. Начальный объем культуры составлял 1,5 л, конечный объем - 3,0 л. Общая концентрация глюкозы, добавленной в ходе ферментации, составляла 250 г/л. В ходе ферментации среду перемешивали со скоростью 700-1000 об/мин, температуру контролировали на значении 32 С, рН устанавливали равным 7,0 с помощью 25-28% водного раствора аммиака. Скорость потока воздуха составляла 0,5 объема на объем в минуту (vvm). Полученные результаты представлены в таблице 4. Как следует из таблицы 4, штамм-хозяин TRN212 продуцирует 93,5 г/л L-треонина с выходом 37,4% в расчете на израсходованную глюкозу. В отличие от этого, рекомбинантный штамм TRN212 продуцирует 112 г/л L-треонина с выходом 45,3%, что на 21% выше, чем выход в случае штамма-хозяина TRN212. Кроме того, сходный профиль ферментации наблюдался в присутствии штамма-хозяина и рекомбинантного штамма без снижения потребления сахара в ходе ферментации, что часто происходит с рекомбинантными штаммами за счет ингибирования роста.
Состав исходной среды в 5-литровом ферментере
Результаты ферментативного образования треонина рекомбинантными штаммами
При использовании нового рекомбинантного штамма настоящего изобретения при получении L-треонина как количество, так и выход увеличиваются более чем на 20% по сравнению со штаммом-хозяином, при этом в ходе ферментации не снижается потребление сахара, что часто наблюдается при использовании рекомбинантных штаммов за счет ингибирования роста.
Изобретение относится к биотехнологии и представляет собой способ получения L-треонина. Согласно способу ген треониндегидратазы (tdc), присутствующий в геномной ДНК микроорганизма, частично инактивируют с использованием рекомбинантной технологии. Штамм микроорганизма E.coli pGm TN-PPC12 (КСММ-10236) трансформируют фрагментом ДНК Δtdc::loxpKan и получают штамм E.coli TRN212 (КССМ-10353). Данный штамм культивируют для получения L-треонина. Заявленное изобретение позволяет значительно увеличить выход L-треонина. 3 н.п. ф-лы, 3 ил., 4 табл.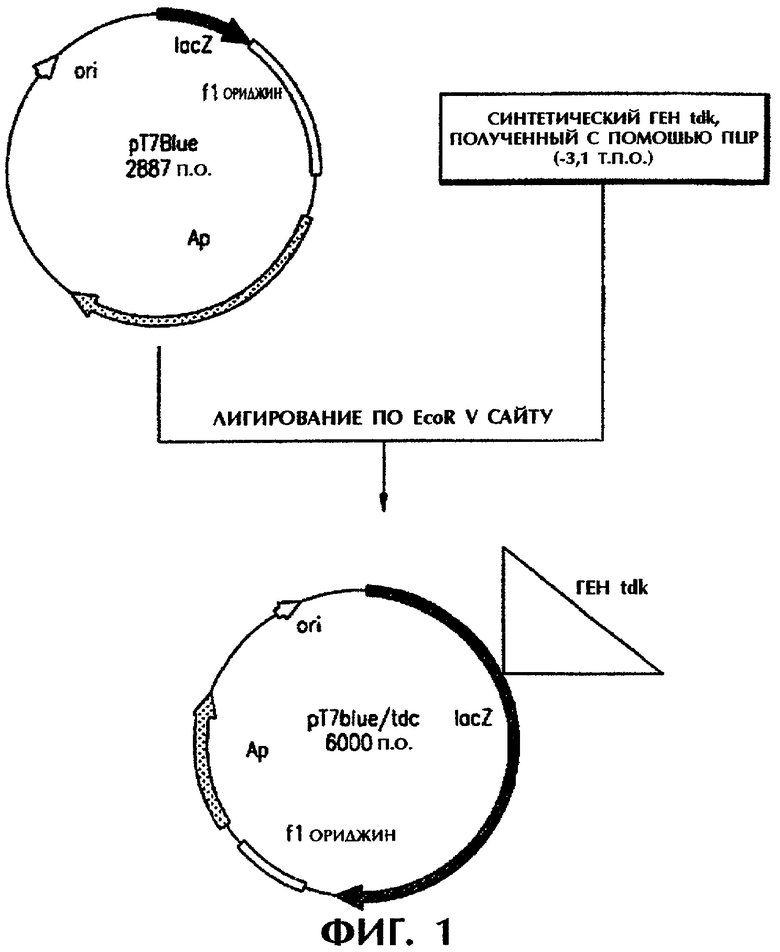
| US 5939307, 17.08.1999 | |||
| US 5538873, 23.07.1996 | |||
| ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ АСПАРТОКИНАЗУ III, СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 1993 |
|
RU2113484C1 |

