В настоящем изобретении предлагается ослабленный вирус, который происходит от модифицированного вируса коровьей оспы Ankara и который характеризуется потерей способности к своей репродукции путем репликации в клеточных линиях человека. В нем дополнительно описываются рекомбинантные вирусы, которые происходят от данного вируса, и применение вируса или его рекомбинантных вариантов в качестве лекарства или вакцины. Дополнительно предлагается способ индукции иммунного ответа даже у больных с нарушенным иммунитетом, больных с исходно существующим иммунитетом к вирусу вакцины или больных в процессе проведения противовирусной терапии.
ИЗВЕСТНЫЙ УРОВЕНЬ ТЕХНИКИ
Модифицированный вирус коровьей оспы Ankara (MVA) относится к вирусу коровьей оспы, члену рода Orthopoxvirus семейства Poxiviridae. MVA был создан в результате 516 серийных пассажей вируса коровьей оспы (CVA) в эмбриональных фибробластах цыплят линии Ankara (в качестве обзора смотри Mayr, A., et al. Infection 3, 6-14 [1975]). Вследствие данных многочисленных пассажей полученный вирус MVA потерял приблизительно 31 тысячу оснований своей геномной последовательности и, следовательно, был описан как вирус с крайне ограниченными клетками-хозяевами, относящимися к клеткам птиц (Meyer, H. et al., J. Gen. Virol. 72, 1031-1038 [1991]). На различных животных моделях было показано, что полученный MVA был практически авирулентным (Mayr, A. & Danner, K. [1978], Dev. Biol. Stand. 41:225-34). Кроме того, данный штамм MVA был протестирован при клинических испытаниях в качестве вакцины для иммунизации против оспы человека (Mayr et al., Zbl. Bakt. Hyg. I, Abt. Org. B 167, 375-390 [1987], Stickl et al., Dtsch. med. Wschr. 99, 2386-2392 [19974]). В данное исследование было вовлечено более 120000 людей, включая пациентов с высоким риском заболевания, и было доказано, что по сравнению с вакцинами, основанными на вирусе коровьей оспы, MVA имеет ослабленную вирулентность или инфекционность, сохраняя в то же время хорошую иммуногенность.
В последующие десятилетия были созданы генно-инженерные конструкции MVA для применения их в качестве вирусного вектора для экспрессии рекомбинантных генов или в качестве рекомбинантной вакцины (Sutter, G. et al. [1994], Vaccine 12:1032-40).
В данном отношении наиболее удивительным является то, что даже несмотря на то, что Mayr et al. в 1970-е годы показали, что MVA является крайне ослабленным и авирулентным у человека и млекопитающих, в настоящее время в некоторых представленных работах (Blandchard et al., 1998, J Gen Virol 79, 1159-1167; Carroll & Moss, 1997, Virology 238, 198-211; Altenberger, патент США 5185146; Ambrosini et al., 1999, J Neurosci Res 55(5), 569) показано, что в клеточных линиях человека и млекопитающих MVA не является полностью ослабленным, так как в данных клетках может происходить остаточная репликация. Предполагается, что результаты, представленные в данных публикациях, получены с различными штаммами MVA, так как использованные вирусы существенно различаются по своим свойствам, особенно по характеру своего роста в различных клеточных линиях. Характер роста рассматривается в качестве индикатора ослабленности вируса. Обычно штамм вируса считается ослабленным, если он потерял свою способность или только снизил свою способность к своей репродукции путем репликации в клетках-хозяевах. Указанный выше факт, что MVA не является полностью не способным к репликации в клетках человека и млекопитающих, ставит вопрос о том, насколько абсолютна безопасность MVA как вакцины для человека или вектора для рекомбинантных вакцин.
Особенно для вакцины, а также для рекомбинантной вакцины баланс между эффективностью и безопасностью вирусного вектора вакцины является крайне важным.
ЦЕЛЬ ИЗОБРЕТЕНИЯ
Таким образом, целью изобретения является обеспечение новыми штаммами, характеризуемыми повышенной безопасностью, для разработки более безопасных продуктов, таких как вакцины или лекарства. Более того, дополнительной целью является обеспечение средствами для улучшения существующего режима вакцинации.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Для достижения перечисленных выше целей в соответствии с предпочтительным осуществлением настоящего изобретения предлагаются новые вирусы коровьей оспы, которые способны к репродукции путем репликации в клетках и в клеточных линиях животных, не являющихся человеком, особенно в фибробластах эмбрионов цыплят (CEF) и в линии клеток почки детенышей хомячка BHK (ECACC 85011433), но не способны к репродукции путем репликации в клеточных линиях человека.
Известные штаммы вирусов коровьей оспы способны к репродукции путем репликации в, по меньшей мере, некоторых клеточных линиях человека, в частности в клеточной линии кератиноцитов человека HaCat (Boukamp et al. 1988, J Cell Biol 106(3): 761-71). Репликация в клеточной линии HaCat предполагает репликацию in vivo, в частности репликацию in vivo у человека. Конечно, в разделе примеров показано, что все протестированные известные штаммы вируса коровьей оспы, которые проявляют способность к остаточному воспроизведению путем репликации в HaCat, также реплицируются in vivo. Таким образом, изобретение предпочтительно относится к вирусам коровьей оспы, которые не способны к репродукции путем репликации в клеточной линии HaCat человека. Наиболее предпочтительно изобретение касается штаммов вируса коровьей оспы, которые не способны к репродукции путем репликации в любой из следующих клеточных линий человека: клеточной линии аденокарциномы шейки матки человека HeLa (ATCC № CCL-2), линии клеток почки эмбриона человека 293 (ECACC № 85120602), клеточной линии остеосаркомы кости человека 143B (ECACC № 91112502) и клеточной линии HaCat.
Характер роста или амплификации/репликации вируса обычно выражается отношением количества вирусов, продуцируемых инфицированной клеткой (выход), к количеству, первоначально используемому для исходного инфицирования клеток (вход) ("амплификационное отношение"). Отношением между выходом и входом, равным "1", определяется такой амплификационный статус, при котором количество вируса, продуцируемого инфицированными клетками, является тем же самым, что и количество, исходно используемое для инфицирования клеток. Данный статус указывает на тот факт, что инфицированные клетки являются пермиссивными для вирусной инфекции и репродукции вируса. Амплификационное отношение, составляющее менее 1, т.е. снижение амплификации ниже уровня входа является индикатором недостаточности репликации и, таким образом, индикатором ослабленности вируса. Следовательно, для изобретателей особый интерес представляет идентификация и, в конечном итоге, выделение штамма, который характеризуется амплификационным отношением менее 1 в нескольких клеточных линиях человека, в частности во всех клеточных линиях человека 143B, HeLa, 293 и HaCat.
Таким образом, термин "не способный к репродукции путем репликации" обозначает, что вирус в соответствии с изобретением характеризуется амплификационным отношением менее 1 в клеточных линиях человека, таких как клеточные линии 293 (ECACC № 85120602), 143B (ECACC № 91112502), HeLa (ATCC № CCL-2) и HaCat (Boukamp et al. 1988, J Cell Biol 106(3): 761-71), в условиях, описанных в примере 1 настоящего описания, для некоторых конкретных штаммов MVA. Предпочтительно, чтобы амплификационное отношение для вируса в соответствии с изобретением составляла 0,8 или менее в каждой из перечисленных выше линиях HeLa, HaCat и 143B.
В примере 1 и в таблице 1 показано, что вирусы в соответствии с настоящим изобретением не способны к репродукции путем репликации ни в одной из клеточных линий 143B, HeLa и HaCat. Особый штамм в соответствии с настоящим изобретением, который был использован в примерах, был положен на хранение в Европейскую коллекцию клеточных культур под номером V00083008. Данный штамм обозначается во всем описании как "MVA-BN".
Известные штаммы MVA проявляют способность к остаточной репликации в, по меньшей мере, одной из протестированных клеточных линий человека (фиг.1, пример 1). Все известные штаммы вирусов коровьей оспы проявляют способность к, по меньшей мере, некоторой репликации в клеточной линии HaCat, в то время как штаммы MVA в соответствии с настоящим изобретением, в частности MVA-BN, не проявляют способности к воспроизведению путем репликации в клетках HaCat. В более подробном изложении MVA-BN характеризуются амплификационным отношением от 0,05 до 0,2 в линии клеток почки эмбриона человека 293 (ECACC № 85120602). В клеточной линии остеосаркомы кости человека 143B (ECACC № 91112502) отношение находится в диапазоне от 0,0 до 0,6. Для клеточной линии аденокарциномы шейки матки человека HeLa (ATCC № CCL-2) и клеточной линии кератиноцитов человека HaCat (Boukamp et al. 1988, J Cell Biol 106(3): 761-71) амплификационное отношение находится в диапазоне от 0,04 до 0,8 и от 0,02 до 0,8, соответственно. MVA-BN характеризуется амплификационным отношением от 0,01 до 0,06 в клетках почки африканской зеленой мартышки (CV1: ATCC № CCL-70). Таким образом, MVA-BN, который является образцом штамма в соответствии с настоящем изобретением, не способен к воспроизведению путем репликации ни в одной из протестированных клеточных линий.
Амплификационное отношение MVA-BN равняется величине, несомненно, выше 1 в фибробластах эмбрионов цыплят (CEF: первичные культуры) или в линии клеток почки детенышей хомячка BHK (ATCC № CRL-1632). Как отмечалось выше, отношение, составляющее более "1", указывает на репродукцию путем репликации, так как количество вирусов, продуцируемых инфицированными клетками, увеличивается по сравнению с количеством вирусов, которое использовалось для инфицирования клеток. Следовательно, вирус может легко размножаться и амплифицироваться в первичных культурах CEF с отношением выше 500 или в клетках BHK с отношением выше 50.
В конкретном осуществлении настоящего изобретения изобретение касается производных вируса, депонированного под № ECACC V0083008. "Производные" вирусов, депонированных под № ECACC V00083008, обозначают вирусы, проявляющие по существу те же самые репликационные характеристики, что и депонированный штамм, но имеющие отличия в одной или более частей своего генома. Вирусы, имеющие те же самые "репликационные характеристики", что и депонированный вирус, представляют собой вирусы, которые реплицируются со сходными амплификационными отношениями, что и депонированный штамм, в клетках CEF и клеточных линиях BHK, HeLa, HaCat и 143B, и которые проявляют сходную репликацию in vivo при определении на модели трансгенной мыши AGR129 (смотри ниже).
В предпочтительном осуществлении штаммы вируса коровьей оспы в соответствии с настоящим изобретением, в частности MVA-BN и его производные, характеризуются отсутствием способности реплицироваться in vivo. В контексте настоящего изобретения "отсутствие способности реплицироваться in vivo" относится к вирусам, которые не реплицируются в клетках человека и мышиной модели, описанной ниже. "Отсутствие способности реплицироваться in vivo" может быть определено предпочтительно на мышах, которые не способны продуцировать зрелые B- и T-клетки. Примером таких мышей являются трансгенные мыши модели AGR129 (полученные от Mark Sutter, Institute of Virology, University of Zurich, Швейцария). Данная линия мышей характеризуется направленными генетическими повреждениями в генах рецептора IFN типа I (IFN-α/β) и типа II (IFN-γ) и в RAG. Из-за данных повреждений мыши не имеют системы IFN и не способны продуцировать зрелые B- и T-клетки и в результате этого характеризуются существенно нарушенным иммунитетом и высокой восприимчивостью к репликации вирусов. Вместо мышей AGR129 может быть использована любая другая линия мышей, которая не способна продуцировать зрелые B- и T-клетки и в результате этого характеризуется существенно нарушенным иммунитетом и высокой восприимчивостью к репликации вирусов. В частности, вирусы в соответствии с настоящим изобретением не убивают мышей AGR129 в течение периода времени, по меньшей мере, 45 дней, более предпочтительно в течение, по меньшей мере, 60 дней, наиболее предпочтительно в течение 90 дней после инфицирования мышей путем внутрибрюшинного введения 107 БОЕ вируса. Предпочтительно, чтобы вирусы, которые проявляют "отсутствие способности реплицироваться in vivo", дополнительно характеризовались тем, что ни одного вируса не могло выявляться в органах или тканях мышей AGR129 через 45 дней, предпочтительно через 60 дней и наиболее предпочтительно через 90 дней после инфицирования мышей путем внутрибрюшинного введения 107 БОЕ вируса. Подробная информация об испытаниях по инфицированию мышей AGR129 и тестах, которые применяются для определения того, может ли вирус выявляться в органах или тканях инфицированных мышей, может быть найдена в разделе примеров.
В предпочтительном осуществлении штаммы вируса коровьей оспы в соответствии с настоящим изобретением, в частности MVA-BN и его производные, характеризуются более высокой иммуногенностью по сравнению с известным штаммом MVA-575 при определении на модели мышей с летальным инфицированием. Подробности данного эксперимента описаны в примере 2, представленном ниже. Вкратце, в такой модели невакцинированные мыши умирают после инфицирования репликативными компонентами штаммов вирусов коровьей оспы, такими как штамм L929 TK+ Western Reserve или IHD-J. В контексте описания модели с летальным заражением, "заражением" обозначается инфицирование репликативными компонентами вирусов коровьей оспы. Через четыре дня после заражения мышей обычно забивают и определяют титр вирусов в яичниках с помощью стандартных тестов образования бляшек с применением клеток VERO (для более подробного описания смотри раздел примеров). Титр вирусов определяют у невакцинированных мышей и у мышей, вакцинированных вирусной вакциной в соответствии с настоящим изобретением. Более конкретно, вирусы в соответствии с настоящим изобретением характеризуются тем, что в данном тесте после вакцинации 102 TCID50/мл вирусами в соответствии с настоящим изобретением титры вирусов в яичнике снижены, по меньшей мере, на 70%, предпочтительно, по меньшей мере, на 80%, более предпочтительно, по меньшей мере, на 90% по сравнению с невакцинированными мышами.
В предпочтительном осуществлении вирусы коровьей оспы в соответствии с настоящим изобретением, в частности MVA-BN и его производные, пригодны для иммунизации путем первичного/повторного введения вакцины. Существуют многочисленные сообщения, предполагающие, что режимы первичной/повторной иммунизации с применением MVA в качестве вектора для доставки индуцируют слабые иммунные ответы и уступают режиму первичной иммунизации с помощью ДНК и повторной иммунизации MVA (Schneider et al., 1998, Nat. Med. 4; 397-402). Во всех данных исследованиях применялись штаммы MVA, которые отличаются от вирусов коровьей оспы в соответствии с настоящим изобретением. Для объяснения слабого иммунного ответа при применении MVA для первичного и повторного введения была выдвинута гипотеза о том, что антитела, выработанные против MVA при первичном введении, нейтрализуют MVA, вводимые при повторной иммунизации, предотвращая эффективное повышение иммунного ответа. В противоположность этому режимы с первичной иммунизацией ДНК и повторной иммунизацией MVA, как сообщается, являются прекрасными в плане генерации антител с высокой авидностью, потому что при данном режиме способность ДНК эффективно примировать иммунный ответ сочетается со свойствами MVA усиливать данный ответ при повторной иммунизации в отсутствие ранее существующего иммунитета по отношению к MVA. Ясно, что, если ранее существующий иммунитет к MVA и/или вирусу коровьей оспы предотвращает усиление иммунного ответа после повторной иммунизации, то применение MVA в качестве вакцины или лекарства должно иметь ограниченную силу, особенно у больных, которые были вакцинированы против оспы. Однако в соответствии с дополнительным осуществлением вирус коровьей оспы в соответствии с настоящим изобретением, в частности MVA-BN и его производные, также как соответствующие рекомбинантные вирусы, включающие гетерологичные последовательности, могут быть использованы для эффективных первичного примированного и последующего усиливающего иммунных ответов у интактных животных, а также у животных с предсуществующим иммунитетом по отношению к вирусам оспы. Таким образом, вирус коровьей оспы в соответствии с настоящим изобретением индуцирует, по меньшей мере, по существу тот же уровень иммунитета при режимах первичной иммунизации вирусом коровьей оспы/повторной иммунизации вирусом коровьей оспы по сравнению с режимами первичной иммунизации ДНК/повторной иммунизации вирусом коровьей оспы.
Вирус коровьей оспы рассматривается как индуцирующий, по меньшей мере, по существу тот же уровень иммунитета при режимах первичной иммунизации вирусом коровьей оспы/повторной иммунизации вирусом коровьей оспы по сравнению с режимами первичной иммунизации ДНК/повторной иммунизации вирусом коровьей оспы, если CTL (ЦТЛ) ответ при измерении в одном из следующих двух тестов ("тест 1" и "тест 2"), предпочтительно в обоих тестах, является, по меньшей мере, по существу тем же самым при режимах первичной иммунизации вирусом коровьей оспы/повторной иммунизации вирусом коровьей оспы по сравнению с режимами первичной иммунизации ДНК/повторной иммунизации вирусом коровьей оспы. Более предпочтительно, чтобы CTL ответ после первичной иммунизации вирусом коровьей оспы/повторной иммунизации вирусом коровьей оспы был выше, по меньшей мере, в одном из тестов по сравнению с режимами первичной иммунизации ДНК/повторной иммунизации вирусом коровьей оспы. Наиболее предпочтительно, чтобы CTL ответ был выше в обоих из последующих тестов.
Тест 1: для первичного введения вируса коровьей оспы/повторного введения вируса коровьей оспы проводят первичную иммунизацию 6-8 недельных мышей BALB/c (H-2d) путем внутривенного введения 107 TCID50 вируса коровьей оспы в соответствии с настоящим изобретением, экспрессирующего мышиный политоп, как описано в Thomson et al., 1988, J. Immunol. 160, 1717, и повторно иммунизируют тем же количеством того же вируса, вводимого тем же путем через три недели. Для данной цели необходимо создать конструкцию рекомбинантного вируса коровьей оспы, экспрессирующий указанный политоп. Способы создания конструкций таких рекомбинантных вирусов известны специалисту в данной области техники и описываются более подробно ниже. В случае режимов первичной иммунизации ДНК/повторной иммунизации вирусом коровьей оспы первичную вакцинацию проводят путем внутримышечного введения мышам 50 мкг ДНК, экспрессирующей тот же самый антиген, что и вирус коровьей оспы; при повторном введении вводят вирус коровьей оспы точно тем же путем, что и в случае первичной иммунизации вирусом коровьей оспы/повторной иммунизации вирусом коровьей оспы. Плазмидная ДНК, экспрессирующая политоп, описана также в указанной выше публикации Thomson et al. В случае обоих режимов развитие CTL ответа против эпитопов SYIPSAEKI, RPQASGVYM и/или YPHFMPTNL определяют через две недели после повторного введения. Определение CTL ответа предпочтительно осуществляют путем использования анализа ELISPOT, как описано Schneider et al., 1998, Nat. Med. 4, 397-402, и представлено в разделе примеров ниже для одного конкретного вируса в соответствии с настоящим изобретением. Вирус в соответствии с настоящим изобретением характеризуется в данном эксперименте тем, что иммунный CTL ответ против указанных выше эпитопов, который индуцируется первичным введением вируса коровьей оспы/повторным введением вируса коровьей оспы, является по существу тем же самым, предпочтительно, по меньшей мере, тем же самым, что и индуцируемый первичным введением ДНК/повторным введением вируса коровьей оспы при оценке по числу клеток, продуцирующих IFN-γ/106 клеток селезенки (смотри также экспериментальный раздел).
Тест 2: данный тест в основном соответствует тесту 1. Однако вместо использования в/в введения 107 TCID50 вируса коровьей оспы, как в случае теста 1, в данном тесте 108 TCID50 вируса коровьей оспы в соответствии с настоящим изобретением вводят подкожно для первичной иммунизации и для повторной иммунизации. Вирус в соответствии с настоящим изобретением характеризуется в данном эксперименте тем, что иммунный CTL ответ против эпитопов, указанных выше, который индуцируется первичным введением вируса коровьей оспы/повторным введением вируса коровьей оспы, является по существу тем же самым, предпочтительно, по меньшей мере, тем же самым, что и индуцируемый первичным введением ДНК/повторным введением вируса коровьей оспы при оценке по числу клеток, продуцирующих IFN-γ/106 клеток селезенки (смотри также экспериментальный раздел).
Сила CTL ответа, измеренная в одном из тестов, представленных выше, соответствует уровню защиты.
Таким образом, вирусы в соответствии с настоящим изобретением особенно подходят для целей вакцинации.
В целом вирус в соответствии с настоящим изобретением характеризуется наличием, по меньшей мере, одного из следующих свойств:
(i) способностью к репродукции путем репликации в фибробластах эмбрионов цыплят (CEF) и в клеточной линии BHK, но отсутствием способности к репродукции путем репликации в клеточной линии HaCat человека,(ii) отсутствием способности к репликации in vivo,
(iii) индукцией более высокого иммунитета по сравнению с известным штаммом MVA-575 (ECACC V00120707) в модели летального заражения и/или
(iv) индукцией, по меньшей мере, по существу того же самого уровня иммунитета при режимах первичной иммунизации вирусом коровьей оспы/повторной иммунизации вирусом коровьей оспы по сравнению с режимами первичной иммунизации ДНК/повторной иммунизации вирусом коровьей оспы.
Предпочтительно вирус коровьей оспы в соответствии с настоящим изобретением имеет, по меньшей мере, два из указанных выше свойств, более предпочтительно, по меньшей мере, три из указанных выше свойств. Наиболее предпочтительными являются вирусы коровьей оспы, имеющие все указанные выше свойства.
В дополнительном осуществлении изобретение касается набора для вакцинации, включающего вирус в соответствии с настоящим изобретением для первичной вакцинации ("примирования") в одном флаконе/контейнере и для повторной вакцинации ("бустинга") во втором флаконе/контейнере. Вирус может представлять собой нерекомбинантный вирус коровьей оспы, т.е. вирус коровьей оспы, который не содержит гетерологичных нуклеотидных последовательностей. Примером такого вируса коровьей оспы является MVA-BN и его производные. В альтернативном варианте вирус может представлять собой рекомбинантный вирус коровьей оспы, который содержит дополнительные нуклеотидные последовательности, которые являются гетерологичными по отношению к вирусу коровьей оспы. Как указано в других разделах описания, гетерологичные последовательности могут кодировать эпитопы, которые индуцируют ответ иммунной системы. Таким образом, можно применять рекомбинантный вирус коровьей оспы для вакцинации против белков или агентов, включающих указанный эпитоп. Вирусы могут быть включены в составы, как более подробно показано ниже. Количество вируса, которое может быть использовано для каждой вакцинации, указано выше.
Специалисту в данной области техники известно как можно получить вирусы коровьей оспы, имеющие, по меньшей мере, одно из следующих свойств:
- способность к репродукции путем репликации в фибробластах эмбрионов цыплят (CEF) и в линии клеток почек детенышей хомячка BHK, но отсутствие способности к репродукции путем репликации в клеточной линии кератиноцитов человека HaCat,
- отсутствие способности к репликации in vivo,
- индукция более высокого иммунитета по сравнению с известным штаммом MVA-575 в модели летального заражения и/или
- индукция, по меньшей мере, по существу того же самого уровня иммунитета при режимах первичной иммунизации вирусом коровьей оспы/повторной иммунизации вирусом коровьей оспы по сравнению с режимами первичной иммунизации ДНК/повторной иммунизации вирусом коровьей оспы.
Способ получения такого вируса может включать следующие стадии:
(i) введение известного штамма вируса коровьей оспы, предпочтительно MVA-574 или MVA-575 (ECACC V00120707) в клетки, отличные от клеток человека, в которых вирус способен к репродукции путем репликации, где клетки, отличные от клеток человека, предпочтительно выбраны из клеток CEF и клеточной линии BHK,
(ii) выделение/повышение концентрации вирусных частиц из данных клеток и
(iii) анализ того, будет ли полученный вирус иметь, по меньшей мере, одно из желаемых биологических свойств, как указано выше,
где указанные выше стадии могут необязательно повторяться до тех пор, пока не будет получен вирус с желаемыми репликативными характеристиками. Изобретение дополнительно относится к вирусам, полученным данным способом в соответствии с настоящим изобретением. Способы того, как могут быть определены желаемые биологические свойства, объяснены в других частях настоящего описания.
Применяя данный способ, изобретатели идентифицировали и выделили в результате нескольких циклов очистки клона штамм в соответствии с настоящим изобретением, начиная с пассажа 575 изолята MVA (MVA-575). Данный новый штамм соответствует штамму с депонентным номером ECACC V00883008, указанным выше.
Характер роста вирусов коровьей оспы в соответствии с настоящим изобретением, в частности характер роста MVA-BN, указывает на то, что штаммы в соответствии с настоящим изобретением существенно опережают любой другой охарактеризованный к настоящему времени изолят MVA в отношении ослабленности действия в клеточных линиях человека и отсутствия способности к репликации in vivo. Штаммы в соответствии с настоящим изобретением являются, следовательно, идеальными кандидатами для разработки более безопасных продуктов, таких как вакцины или лекарства, как будет описано ниже.
В одном осуществлении вирус в соответствии с настоящим изобретением, в частности MVA-BN и его производные, применяют в качестве вакцины против вирусных заболеваний человека, сопровождающихся сыпью, таких как оспа. В дополнительном осуществлении вирус в соответствии с настоящим изобретением может быть рекомбинантным, т.е. может экспрессировать гетерологичные гены, такие как, например, антигены или эпитопы, гетерологичные по отношению к вирусу, и может, таким образом, быть пригодным в качестве вакцины для индукции иммунного ответа против гетерологичных антигенов или эпитопов.
Термин "иммунный ответ" обозначает реакцию иммунной системы при поступлении в организм чужеродного вещества или микроорганизма. По определению иммунный ответ подразделяют на специфическую и неспецифическую реакции, хотя обе они тесно переплетаются. Неспецифический иммунный ответ представляет собой немедленную защиту от широкого разнообразия чужеродных веществ и инфекционных агентов. Специфический иммунный ответ представляет собой защиту, возникающую после лаг-периода после первого заражения организма веществом. Специфический иммунный ответ является высокоэффективным и определяет тот факт, что индивидуум, который переболел конкретной инфекцией, становится защищенным против данной конкретной инфекции. Таким образом, повторное инфицирование тем же самым или очень близким инфекционным агентом вызывает намного более сглаженные симптомы или совсем не вызывает симптомов, так как уже имеется "предсуществующий иммунитет" в отношении данного агента. Такой иммунитет и иммунологическая память, соответственно, сохраняется в течение длительного времени, в некоторых случаях даже в течение всей жизни. Соответственно, индукция иммунологической памяти может быть использована при вакцинации.
"Иммунная система" обозначает комплекс органов, вовлеченных в защиту организма от чужеродных веществ и микроорганизмов. Иммунная система включает клеточную составляющую, включающую несколько типов клеток, таких как, например, лимфоциты и другие клетки, происходящие от белых клеток крови, и гуморальную составляющую, включающую небольшие пептиды и факторы комплемента.
"Вакцинация" обозначает то, что организм нагружают инфекционным агентом, например, ослабленной или инактивированной формой указанного инфекционного агента, для индукции специфического иммунитета. Термин вакцинация также охватывает заражение организма рекомбинантными вирусами коровьей оспы в соответствии с настоящим изобретением, в частности, рекомбинантным MVA-BN и его производными, экспрессирующими антигены или эпитопы, которые являются гетерологичными по отношению к вирусу. Примеры таких эпитопов даются в других частях описания и охватывают, например, эпитопы белков, происходящих от других вирусов, таких как вирус Денге, вирус гепатита С, ВИЧ или эпитопы, происходящие от белков, которые ассоциированы с развитием опухолей и рака. После введения рекомбинантного вируса коровьей оспы в организм эпитопы экспрессируются и презентируются иммунной системе и может индуцироваться специфический иммунный ответ против данных эпитопов. Организм, таким образом, является иммунизированным против агента/белка, содержащего эпитоп, который кодируется рекомбинантным вирусом коровьей оспы.
"Иммунитет" обозначает частичную или полную защиту организма от заболеваний, вызываемых инфекционным агентом, обусловленную успешной ликвидацией предшествующего инфицирования указанным инфекционным агентом или его характерной частью. Иммунитет основан на существовании, индукции и активации специализированных клеток иммунной системы.
Как подчеркивалось выше, в одном осуществлении изобретения рекомбинантные вирусы в соответствии с настоящим изобретением, в частности, рекомбинантный MVA-BN и его производные, содержат, по меньшей мере, одну гетерологичную последовательность нуклеиновой кислоты. Термин "гетерологичная" применяют здесь далее для любого сочетания последовательностей нуклеиновой кислоты, которое в обычных условиях не находят в близкой связи с вирусом в природе, такой вирус называют также "рекомбинантный вирус".
В соответствии с дополнительным осуществлением настоящего изобретения гетерологичные последовательности представляют собой предпочтительно антигенные эпитопы, которые выбраны из любого источника, не являющегося вирусом коровьей оспы. Наиболее предпочтительно, чтобы указанный рекомбинантный вирус экспрессировал один или более антигенных эпитопов от Plasmodium falciparum, Mycobacteria, Influenza virus, от вирусов, выбранных из семейства Flaviviruses, Paramyxoviruses, вирусов гепатита, вирусов иммунодефицита человека, или от вирусов, вызывающих геморрагическую лихорадку, таких как Hantaviruses или Filoviruses, т.е. вируса Эбола или Марбург.
В соответствии с еще одним осуществлением, но также в дополнение к указанному выше выбору антигенных эпитопов, гетерологичные последовательности могут быть выбраны из других источников оспы и коровьей оспы. Данные вирусные последовательности могут быть применены для модификации спектра хозяев или иммуногенности вируса.
В дополнительном осуществлении вирус в соответствии с настоящим изобретением может кодировать гетерологичный ген/нуклеиновую кислоту, экспрессирующие терапевтическое соединение. "Терапевтическое соединение", кодируемое гетерологичной нуклеиновой кислотой в вирусе, может представлять собой, например, терапевтическую нуклеиновую кислоту, такую как антисмысловая нуклеиновая кислота, или пептид, или белок с желаемой биологической активностью.
В соответствии с дополнительным предпочтительным осуществлением экспрессия гетерологичной последовательности нуклеиновой кислоты находится предпочтительно, но не исключительно, под транскрипционным контролем промотора вируса оспы, более предпочтительно промотора вируса коровьей оспы.
В соответствии с еще одним осуществлением вставка гетерологичной последовательности нуклеиновой кислоты осуществляется в область вирусного генома, не являющуюся необходимой. В другом предпочтительном осуществлении изобретения гетерологичную последовательность нуклеиновой кислоты вставляют в природно существующий сайт делеции генома MVA (раскрытый в патенте PCT/EP96/02926). Методы вставки гетерологичных последовательностей в геном вируса оспы известны специалистам в данной области техники.
В соответствии с другим дополнительным предпочтительным осуществлением изобретение включает также геном вируса, его рекомбинантные варианты или его функциональные части. Такие вирусные последовательности могут быть использованы для идентификации или выделения вируса или его рекомбинантных вариантов, например, с применением ПЦР, методов гибридизации или с помощью традиционных тестов ELISA. Более того, такие вирусные последовательности могут экспрессироваться экспрессионным вектором с получением кодируемого белка или пептида, которые затем могут дополнять делеционные мутанты вируса, у которых отсутствует вирусная последовательность, содержащаяся в экспрессионном векторе.
"Функциональная часть" вирусного генома обозначает часть полной геномной последовательности, которая кодирует физический объект, такой как белок, домен белка, эпитоп белка. Функциональная часть вирусного генома также описывает части полной геномной последовательности, которые кодируют регуляторные элементы или части таких элементов с индивидуализированной активностью, такие как промотор, энхансер, цис- или транс-действующие элементы.
Рекомбинантный вирус в соответствии с настоящим изобретением может быть применен для введения гетерологичной последовательности нуклеиновой кислоты в клетку-мишень, причем указанная последовательность является либо гомологичной, либо гетерологичной по отношению к клетке-мишени. Введение гетерологичной последовательности нуклеиновой кислоты в клетку-мишень может быть применено для получения in vitro гетерологичных пептидов или полипептидов и/или полных вирусов, кодируемых указанной последовательностью. Данный способ включает инфицирование клетки-хозяина рекомбинантным MVA, культивирование инфицированной клетки-хозяина в подходящих условиях и выделение и/или повышение концентрации пептида, белка и/или вируса, продуцируемых указанной клеткой-хозяином.
Более того, способ введения гомологичной или гетерологичной последовательности в клетки может быть применен для терапии in vitro и предпочтительно in vivo. Для терапии in vitro выделенные клетки, которые предварительно (ex vivo) инфицировали вирусом, вводят в организм живого животного для индукции иммунного ответа. Для терапии in vivo вирус или его рекомбинантные варианты прямо вводят в организм живого животного для индукции иммунного ответа. В данном случае клетки вокруг места инокуляции становятся инфицированными вирусом или его рекомбинантными вариантами непосредственно in vivo.
Так как вирус в соответствии с настоящим изобретением характеризуется крайне ограниченным ростом в клетках человека и обезьян и, таким образом, крайне ослабленным, он является идеальным средством для лечения широкого круга млекопитающих, включая человека. Следовательно, в настоящем изобретении предлагается также фармацевтическая композиция и вакцина, например, для индукции иммунного ответа в организме живого животного, включая человека. Вирус изобретения является также безопасным при любых других протоколах терапии.
Фармацевтическая композиция может обычно включать один или более фармацевтически приемлемых и/или допустимых носителей, добавок, антибиотиков, консервантов, адъювантов, разбавителей и/или стабилизаторов. Такими вспомогательными веществами могут быть вода, физиологический раствор, глицерин, этанол, увлажняющие или эмульгирующие агенты, вещества, буферирующие pH, или тому подобное. Подходящими носителями являются обычно большие, медленно метаболизирующиеся молекулы, такие как белки, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, сополимеры аминокислот, агрегаты липидов или тому подобное.
Для получения вакцин вирус или его рекомбинантные варианты в соответствии с изобретением переводят в физиологически приемлемую форму. Это может быть сделано, основываясь на опыте работы с получением вакцин вируса оспы для вакцинации против натуральной оспы (как описано Stickl, H. et al. [1974] Dtsch. med. Wschr. 99, 2386-2392). Например, очищенный вирус с титром 5х108 TCID50/мл в приблизительно 10 мМ Трис, 140 мМ NaCl, pH 7,4 хранят при -80°С. Для приготовления вакцины для уколов, например, 102-108 вирусных частиц лиофилизуют в 100 мл забуференного фосфатом физиологического раствора (PBS) в присутствии 2% пептона и 1% альбумина человека в ампуле, предпочтительно в стеклянной ампуле. Альтернативно, вакцина для уколов может быть получена с помощью поэтапной лиофилизации вируса в композиции. Данная композиция может содержать дополнительные добавки, такие как маннит, декстран, сахар, глицин, лактозу или поливинилпирролидон, или другие добавки, такие как антиоксиданты или инертный газ, стабилизаторы или рекомбинантные белки (например, сывороточный альбумин человека), подходящие для введения in vivo. Стеклянную ампулу затем запаивают и ее можно хранить при температуре между 4°С и комнатной в течение нескольких месяцев. Однако, когда отсутствует необходимость использования ампулы, ее хранят предпочтительно при температуре ниже -20°С.
Для вакцинации или терапии лиофилизат может быть растворен в от 0,1 до 0,5 мл водного раствора, предпочтительно физиологического раствора или Трис буфера и введен либо системно, либо местно, т.е. с помощью парентерального, внутримышечного или любого другого пути введения, известного специалисту в данной области техники. Путь введения, доза и количество введений может быть оптимизировано специалистом в данной области техники известным способом.
Дополнительно в соответствии с другим осуществлением вирус согласно настоящему изобретению особенно полезен для индукции иммунных ответов у животных с нарушенным иммунитетом, например, у обезьян, инфицированных SIV (CD4 < 400/мкл крови), или у людей с нарушенным иммунитетом. Термин "нарушенный иммунитет" описывает состояние иммунной системы индивидуума, у которого возникает только неполные иммунные ответы или который характеризуется сниженной эффективностью защиты от инфекционных агентов. Еще более интересным и находящимся в соответствии с еще одним дополнительным осуществлением является тот факт, что вирус в соответствии с настоящим изобретением может усиливать иммунные ответы у животных или людей с нарушенным иммунитетом даже при наличии у данных животных или людей предсуществующего иммунитета к вирусу оспы. Особенно интересным было то, что вирус в соответствии с настоящим изобретением может усиливать иммунные ответы также у животных или людей, проходящих курс антивирусной терапии, например, терапии против ретровирусов. "Антивирусная терапия" включает концепции лечения, направленные на уничтожение или подавление вирусной инфекции, включая, например, (i) применение аналогов нуклеотидов, (ii) применение ингибиторов ферментативной активности вирусов или их сборки или (iii) применение цитокинов для влияния на иммунные ответы хозяина.
В соответствии с еще одним дополнительным осуществлением вакцина является особенно, но не исключительно, пригодной в области ветеринарии, например, для иммунизации против инфекции, вызываемой вирусом оспы животных. У мелких животных инокуляция для иммунизации предпочтительно выполняется парентерально или интраназально, в то время как у более крупных животных или у людей предпочтительной является подкожная, внутримышечная или пероральная инокуляция.
Изобретатели нашли, что инъекция вакцины, содержащей эффективную дозу только 102 TCID50 (дозы инфицирования в тканевой культуре) вируса в соответствии с настоящим изобретением, уже достаточна для индукции полного иммунитета против вируса коровьей оспы дикого типа при контрольном заражении им мышей. Это является особенно неожиданным, так как такая высокая степень ослабленности вируса в соответствии с настоящим изобретением должна, как ожидалось, влиять негативно и, в результате этого, снижать его иммуногенность. Такое предположение основывается на представлении о том, что для индукции иммунного ответа антигенные эпитопы нуждаются в презентации иммунной системе в достаточном количестве. Вирус, который является крайне ослабленным, и, таким образом, не реплицируемым, может презентировать только очень небольшое количество антигенных эпитопов, то есть столько, сколько он сам включает. Данное количество антигена, которое несут вирусные частицы, не рассматривается как достаточное для индукции сильного иммунного ответа. Однако вирус в соответствии с изобретением стимулирует, даже при очень низкой эффективной дозе только в 102 TCID50, сильный и защищающий иммунный ответ в модели мышь/заражение вирусом коровьей оспы. Вирус в соответствии с настоящим изобретением, таким образом, характеризуется неожиданной и даже увеличенной иммуногенностью по сравнению с другими охарактеризованными к настоящему времени штаммами MVA. Такая высокая иммуногенность делает вирус в соответствии с настоящим изобретением и любую вакцину, связанную с ним, особенно пригодными для применения у животных и людей с нарушенным иммунитетом.
В соответствии с еще одним осуществлением изобретения вирус применяют в качестве адъюванта. "Адъювант" в контексте настоящего описания обозначает усилитель специфического иммунного ответа в вакцинах. "Применение вируса в качестве адъюванта" обозначает включение вируса в уже существующую вакцину для дополнительной стимуляции иммунной системы больного, который получает вакцину. Иммунизирующий эффект антигенного эпитопа в большинстве вакцин часто усиливается при добавке так называемого адъюванта. Адъювант костимулирует иммунную систему путем индукции более сильной специфической иммунной реакции против антигенного эпитопа вакцины. Данная стимуляция может регулироваться факторами системы неспецифического иммунитета, такими как интерферон и интерлейкин. Следовательно, в дополнительном осуществлении изобретения вирус применяют у млекопитающих, включая человека, для активации, поддержки или подавления иммунной системы и предпочтительно для активации иммунного ответа против любой антигенной детерминанты. Вирус может быть также применен для поддержки иммунной системы в случае увеличенной восприимчивости к инфекциям, как в случае стресса.
Вирус, используемый в качестве адъюванта, может быть нерекомбинантным вирусом, т.е. вирусом, который не содержит гетерологичную ДНК в своем геноме. Примером данного типа вируса является MVA-BN. В альтернативном варианте, вирус, используемый в качестве адъюванта, является рекомбинантным вирусом, содержащим в своем геноме последовательности гетерологичной ДНК, которые не присутствуют в вирусном геноме в природе. Для применения в качестве адъюванта рекомбинантная вирусная ДНК вируса предпочтительно содержит и экспрессирует гены, которые кодируют иммуностимулирующие пептиды или белки, такие как интерлейкины.
В соответствии с дополнительным осуществлением предпочтительно, чтобы вирус был инактивированным при использовании в качестве адъюванта или добавлении к другой вакцине. Инактивация вируса может быть осуществлена, например, с помощью прогревания или химических агентов, как известно в данной области техники. Предпочтительно, чтобы вирус инактивировался с помощью β-проприолактона. В соответствии с данным осуществлением изобретения инактивированный вирус может быть добавлен к вакцинам против многочисленных инфекционных или пролиферативных заболеваний для увеличения иммунного ответа больного на данное заболевание.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩЕСТВА ИЗОБРЕТЕНИЯ
Изобретение среди прочего включает следующее по одиночке или в сочетании:
вирус коровьей оспы, имеющий, по меньшей мере, одно из следующих свойств - способность к репродукции путем репликации в фибробластах эмбрионов цыплят (CEF) и в линии клеток почек детенышей хомячка BHK, но отсутствие способности к репродукции путем репликации в клеточной линии кератиноцитов человека HaCat,
- отсутствие способности к репликации in vivo,
- индукция более высокого иммунитета по сравнению с известным штаммом MVA-575 в модели летального заражения и/или
- индукция, по меньшей мере, по существу того же самого уровня иммунитета при режимах первичной иммунизации вирусом коровьей оспы/повторной иммунизации вирусом коровьей оспы по сравнению с режимами первичной иммунизации ДНК/повторной иммунизации вирусом коровьей оспы.
Вирус, как описано выше, где вирус не способен к репродукции путем репликации в любой из следующих клеточных линий человека: линии клеток почки эмбриона человека 293, клеточной линии остеосаркомы кости человека 143B и клеточной линии аденокарциномы шейки матки человека HeLa.
Вирус, как описано выше, положенный на хранение в Европейскую коллекцию клеточных культур (ECACC) Salisbury (UK) под номером V00083008 и его производные.
Вирус, как описано выше, включающий, по меньшей мере, одну последовательность гетерологичной нуклеиновой кислоты.
Вирус, как описано выше, где указанная последовательность гетерологичной нуклеиновой кислоты выбрана из последовательности, кодирующей, по меньшей мере, один антиген, антигенный эпитоп и/или терапевтическое соединение.
Геном или его функциональные части, происходящие от вируса, как указано выше. Фармацевтическая композиция, включающая вирус, как указано выше, и/или геном, и/или его функциональную часть, как указано выше, и фармацевтически приемлемый носитель, разбавитель и/или добавку.
Вакцина, включающая вирус, как указано выше, и/или геном, и/или его функциональную часть, как указано выше.
Вирус, как указано выше, геном и/или его функциональная часть, как указано выше, композиция, как указано выше, или вакцина, как указано выше, в качестве лекарства для влияния на иммунный ответ, предпочтительно для его индукции, у живого животного, включая человека.
Вирус, как указано выше, фармацевтическая композиция, как указано выше, вакцина, как указано выше, или вирус, как указано выше, где вирус, композицию или вакцину вводят в терапевтически эффективных количествах при первой инокуляции ("примирующая инокуляция") и при второй инокуляции ("бустерная инокуляция").
Применение вируса, как указано выше, и/или генома, как указано выше, для получения лекарства или вакцины.
Способ введения гомологичной и/или гетерологичной последовательностей нуклеиновой кислоты в клетки-мишени, включающий инфицирование клеток-мишеней вирусом, включающим гетерологичные последовательности, как указано выше, или трансфекцию клетки-мишени геномом, как указано выше.
Способ получения пептида, белка и/или вируса, включающий
- инфицирование клетки-хозяина вирусом, как указано выше,
- культивирование инфицированной клетки-хозяина в подходящих условиях и
- выделение и/или повышение концентрации пептида, и/или белка, и/или вируса, продуцируемых указанной клеткой-хозяином.
Способ влияния на иммунный ответ, предпочтительно его индукции, в организме живого животного, включая человека, включающий введение вируса, как указано выше, генома и/или его функциональной части, как указано выше, композиции, как указано выше, или вакцины, как указано выше, животному или человеку, подвергаемому лечению.
Способ, как указано выше, включающий введение, по меньшей мере, 102 TCID50 (дозы инфицирования тканевой культуры) вируса.
Способ, как указано выше, в котором вирус, композицию или вакцину вводят в терапевтически эффективных количествах при первой инокуляции ("примирующая инокуляция") и при второй инокуляции ("бустерная инокуляция").
Способ, как указано выше, где животное имеет нарушенный иммунитет.
Способ, как указано выше, где животное имеет уже существующий иммунитет по отношению к вирусу оспы.
Способ, как указано выше, где животное подвергается противовирусной терапии.
Способ, где животное подвергается противовирусной терапии, отличающейся тем, что противовирусная терапия представляет собой терапию против ретровирусов.
Применение вируса, как указано выше, его генома и/или функциональной части, как указано выше, в качестве адъюванта.
Способ увеличения специфического иммунного ответа против антигена и/или антигенного эпитопа, включенного в вакцину, включающий введение в качестве адъюванта вируса, как указано выше, или генома, как указано выше, в организм живого животного, включая человека, подвергаемого лечению вакциной.
Вирус, как указано выше, или геном, как указано выше, в качестве адъюванта.
Клетка, предпочтительно клетка человека, содержащая вирус, как указано выше, или геном, или его функциональную часть, как указано выше.
Способ получения вируса коровьей оспы, как указано выше, включающий следующие стадии:
- введение обычно доступного штамма вируса коровьей оспы, предпочтительно MVA-575 в клетки, отличные от клеток человека, в которых вирус способен к репродукции путем репликации, где клетки, отличные от клеток человека, предпочтительно выбраны из клеток CEF и клеточной линии BHK,
- выделение/повышение концентрации вирусных частиц из данных клеток и
- анализ того, будет ли полученный вирус иметь, по меньшей мере, одно из биологических свойств, как указано выше,
где указанные выше стадии могут необязательно повторяться до тех пор, пока не будет получен вирус с желаемыми репликативными характеристиками.
Набор для примированной/бустерной иммунизации, включающий вирус, как указано выше, вакцину, как указано выше, или вирус в качестве лекарства, как указано выше, для первой инокуляции ("примирующая инокуляция") в первом флаконе/контейнере и для второй инокуляции ("бустерная инокуляция") во втором флаконе/контейнере.
Применение вируса, как указано выше, композиции, как указано выше, и/или вакцины, как указано выше, для получения вакцины, где вирус, композицию или вакцину вводят при примированной инокуляции и где тот же вирус или вакцину вводят при бустерной инокуляции.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фигура 1: Кинетика роста различных штаммов MVA в клетках разных линий. В части A) результаты сгруппированы по тестированным штаммам MVA, тогда как в части B) результаты сгруппированы по тестированным линиям клеток. B) Количество вирусов, выделенных из линии клеток после четырех дней (D4) культивирования, определяли с помощью теста по бляшкам и выражали в виде отношения вирусов, выделенных через 4 дня, к исходному прививочному материалу на 1-й день (D1).
Фигура 2: Защита от летального заражения коровьей оспой, обеспечиваемая вакцинацией MVA-BN или MVA-575. Защиту измеряли по снижению овариальных титров, определенных с помощью стандартного теста по бляшкам через 4 дня после контрольного заражения.
Фигура 3: Индукция CTL и защита от заражения гриппом, обеспечиваемая применением различных режимов примирования-повторной иммунизации.
3A: Индукция CTL ответов на 4 различных H-2d ограниченных эпитопа после вакцинации различными сочетаниями ДНК или вакцинами MVA-BN, кодирующими политоп мыши. Мышей BALB/c (по 5 на группу) вакцинировали либо ДНК (внутримышечно), либо MVA-BN (подкожно), и мышей подвергали бустерным иммунизациям спустя три недели. CTL ответы на 4 различных эпитопа, кодируемых вакцинами (TYQRTRALV, грипп; SYIPSAEKI, P. Berghei; YPHFMPTNL, Cytomegalovirus; PRQASGVYM, LCV) определяли с помощью анализа ELISPOT через 2 недели после бустерных иммунизаций.
3B: Индукция CTL ответов на 4 различных эпитопа после вакцинации различными сочетаниями ДНК или вакцинами MVA-BN, кодирующими политоп мыши. Мышей BALB/c (по 5 на группу) вакцинировали либо ДНК (внутримышечно), либо MVA-BN (внутривенно), и мышей подвергали бустерным иммунизациям спустя три недели. CTL ответы на 4 различных эпитопа, кодируемых вакцинами (TYQRTRALV, грипп; SYIPSAEKI, P. Berghei; YPHFMPTNL, Cytomegalovirus; PRQASGVYM, LCV) определяли с помощью анализа ELISPOT через 2 недели после бустерных иммунизаций.
3C: Частота T-клеток, специфичных в отношении пептида и MVA, после гомологичной первичной иммунизации с применением оптимальной дозы (1 х 108 TCID50) рекомбинантного MVA-BN, вводившейся подкожно. Группы из 8 мышей вакцинировали двумя инъекциями сочетаний, как указано на фигуре. Через две недели после последней вакцинации измеряли количество специфичных в отношении пептида спленоцитов с помощью анализа INF-гамма ELISPOT. Столбики представляют среднее количество специфичных пятен плюс/минус стандартное отклонение от среднего.
Фигура 4: Заражение SIV обезьян, вакцинированных MVA-BN nef или MVA-BN.
Фигура 5: Выживаемость вакцинированных обезьян после инъекции SIV.
Фигура 6: Титры антител обезьян против MVA-BN. Титры антител каждого животного показаны в форме различных значков, а средний титр показан в виде темного прямоугольника.
Фигура 7: Уровни SIV у обезьян с нарушенным иммунитетом (CD4<400 мл крови) после вакцинации MVA-BN, кодирующим tat. Обезьяны предварительно получали три вакцинации либо MVA-BN, либо MVA-BN nef (0, 8, 16 недели), и обезьян инфицировали патогенным изолятом SIV (22 неделя). На 100, 102 и 106 неделю (указано стрелками) обезьян вакцинировали MVA-BN tat.
Фигура 8: Уровни SIV у обезьян, подвергнутых антиретровирусной терапии и терапевтическим вакцинациям с применением MVA-BN. Три группы обезьян (n=6) инфицировали SIV и лечили ежедневно PMPA (отмечено черной линией). На 10 и 16 недели животных вакцинировали (отмечено стрелками) либо смесями рекомбинантного MVA, либо физиологическим раствором.
Фигура 9: Гуморальный ответ, индуцированный против SIV, после инфицирования и вакцинаций рекомбинантным MVA. Три группы (n=6) обезьян инфицировали патогенным изолятом SIV (неделя 0) и затем подвергали антиретровирусной терапии (PMPA, отмеченной жирной линией). Обезьян вакцинировали смесями рекомбинантного MVA или физиологическим раствором на 10 и 16 недели. Антитела против SIV определяли с применением лизатов инфицированных T-клеток в качестве антигена с помощью стандартного ELISA.
Фигура 10. Гуморальный ответ, индуцированный против MVA у инфицированных SIV обезьян, подвергнутых антиретровирусной терапии. Три группы (n=6) обезьян инфицировали патогенным изолятом SIV (неделя 0) и затем подвергали антиретровирусной терапии (PMPA, отмеченной жирной линией). Обезьян вакцинировали смесями рекомбинантного MVA или физиологическим раствором на 10 и 16 недели. Антитела против MVA определяли с помощью стандартного ELISA с поглощением для MVA.
Фигура 11. Индукция антител против MVA после вакцинации мышей различными вакцинами против оспы. Уровень антител, выработанных против MVA после вакцинаций MVA-BN (0 и 4 недели), сравнивали с традиционными штаммами коровьей оспы, Elstree и Wyeth, вводившимися посредством скарификации хвоста (0 неделя), MVA-572 (0 и 4 недели) и MVA-BN и MVA-572, вводившимися в форме pre-Elstree вакцины. MVA-572 был помещен на хранение в Европейскую коллекцию культур клеток животных как ECACC V94012707. Титры определяли с помощью ELISA с поглощением, рассчитывали путем линейной регрессии с применением линейной части графика и определяли как разведение, которое давало оптическую плотность 0,3. * MVA-BN: MVA-BN значимо (p>0,05) отличается от MVA-572: MVA-572.
ПРИМЕРЫ
Следующие примеры будут дополнительно иллюстрировать настоящее изобретение. Специалисту в данной области техники должно быть хорошо понятно, что предлагаемые примеры никоим образом не могут быть интерпретированы как ограничивающие применимость технологии, предлагаемой данным изобретением на данных примерах.
Пример 1
Кинетика роста нового штамма MVA в отдельных линиях клеток и репликация in vivo
(i) Кинетика роста в линиях клеток:
Для характеристики нового выделенного штамма согласно настоящему изобретению (далее обозначаемого как MVA-BN) сравнивали кинетику роста данного нового штамма с таковой других штаммов MVA, которые уже были охарактеризованы.
Эксперимент проводили путем сравнения кинетики роста следующих вирусов в перечисленных ниже первичных клетках и линиях клеток:
MVA-BN (Вирусное хранение #23, 18. 02. 99 сырой, титрован в концентрации 2,0x107 TCID50/мл);
MVA, охарактеризованный Altenburger (патент США 5185146) и далее обозначаемый как MVA-HLR;
MVA (пассаж 575), охарактеризованный Anton Mayr (Mayr, A., et al. [1975] Infection 3; 6-14) и далее обозначаемый как MVA-575 (ECACC V00120707); и
MVA-Vero, охарактеризованном в Международной патентной заявке PCT/EP01/02703 (WO 01/68820)(Вирусное хранение, пассаж 49, #20, 22.03.99 сырой, титрован в концентрации 4,2x107 TCID50/мл).
Применяли следующие первичные клетки и линии клеток:
Для инфицирования различные клетки высевали в 6-луночные планшеты при концентрации 5х105 клеток/лунка и инкубировали в течение ночи при 37°С, 5% CO2 в DMEM (Gibco, Cat. № 61965-026) плюс 2% FCS. Среду клеточной культуры удаляли, и клетки инфицировали при приблизительно 0,05 moi в течение одного часа при 37°C, 5% CO2 (для инфицирования принимали, что количество клеток удваивалось за ночь). Количество вирусов, использованных для каждого инфицирования клеток разного типа, составляло 5х104 TCID50, и это будет обозначаться как ввод. Клетки затем промывали 3 раза DMEM, после чего добавляли 1 мл DMEM, 2% FCS, и планшеты оставляли инкубироваться в течение 96 часов (4 дней) при 37°C, 5 % CO2. Данное инфицирование останавливали замораживанием планшет при -80°C, готовых к анализу титрованием.
Анализ титрованием (иммуноокраска специфичным в отношении вируса коровьей оспы антителом)
Для титрования количества вируса клетки для тестирования высевали в 96-луночные планшеты в RPMI (Gibco, Cat. № 61870-010), 7% FCS, 1% антибиотик/противогрибковый агент (Gibco, Cat. № 15240-062) в концентрации 1x104 клеток/лунка и инкубировали в течение ночи при 37°C, 5% CO2. 6-Луночные планшеты, содержащие материал экспериментов инфицирования, подвергали заморозке/оттаиванию 3 раза, и получали разведения от 10-1 до 10-12 с применением ростовой среды RPMI. Разведения вируса вносили к клеткам для тестирования и инкубировали в течение пяти дней при 37°C, 5% CO2 для обеспечения развития CPE (цитопатического эффекта). Клетки для тестирования фиксировали (смесь ацетон/метанол 1:1) в течение 10 мин, промывали PBS и инкубировали с поликлональным антителом, специфичным в отношении вируса коровьей оспы (Quartett Berlin, Cat. № 9503-2057) при разведении 1:1000 в буфере для инкубации в течение одного часа при комн. темп. После промывки дважды с помощью PBS (Gibco, Cat. № 20012-019) добавляли конъюгированное с HPR антитело против кролика (Promega Mannheim, Cat. № W4011) в разведении 1:1000 в буфере для инкубации (PBS, содержащем 3% FCS) в течение одного часа при комн. темп. Клетки вновь дважды промывали PBS и инкубировали с окрашивающим раствором (свежеприготовленным 10 мл PBS+200 мкл насыщенного раствора о-дианизидина в 100% этаноле+15 мкл H2O2) до проявления коричневых пятен (два часа). Окрашивающий раствор удаляли, и добавляли PBS для остановки реакции окрашивания. Каждую лунку, проявившую коричневое пятно, отмечали как позитивную в отношении CPE, и рассчитывали титр по формуле Кербера (на основе анализа TCID50) (Kaerber, G. 1931. Arch. Exp. Pathol. Pharmakol. 162, 480).
Вирусы применяли для инфицирования дублированных рядов, с одной стороны, CEF и BHK, которые, как ожидалось, являются пермиссивными для MVA, и, с другой стороны, CV-1, Hela, 143B и HaCat, которые, как ожидалось, являются непермиссивными для MVA, при низкой множественности инфицирования (moi), т.е. 0,05 инфицирующих единиц на клетку (5х104 TCID50). После этого вирусный прививочный материал удаляли, и клетки три раза промывали для удаления всех оставшихся неабсорбированными вирусов. Инфицированные клетки оставляли суммарно на четыре дня, когда получали вирусные экстракты и затем титровали на клетках CEF. В таблице 1 и на фигуре 1 показаны результаты теста титрования, где величины представлены в виде общего количества вирусов, образовавшихся через 4 дня после инфицирования.
Было показано, что в клетках CEF (фибробластах куриного эмбриона) хорошо амплифицируются все вирусы, что можно было ожидать, поскольку CEF является пермиссивной линией клеток для всех MVA. Кроме того, было показано, что все вирусы хорошо амплифицируются в BHK (линии клеток почек хомячка). MVA действовал наилучшим образом, поскольку BHK является пермиссивной линией клеток.
Что касается репликации в клетках Vero (линия клеток почек обезьяны), MVA-Vero, как и ожидалось, амплифицировался хорошо, а именно 1000-кратно по сравнению с вводом. MVA-HRL, а также MVA-575 амплифицировались хорошо, с 33-кратным и 10-кратным повышением по отношению к вводу, соответственно. Лишь MVA-BN, как было обнаружено, не амплифицируется так же хорошо в данных клетках, как другие, а именно, наблюдалось лишь 2-кратное превышение ввода.
Что касается репликации в клетках CV1 (линии клеток почек обезьяны), было обнаружено, что MVA-BN очень сильно подавляется в данной линии клеток. Было показано 200-кратное снижение по сравнению с вводом. Сходным образом, MVA-575 не амплифицировался выше уровня ввода и также была показана слегка негативная амплификация, а именно 16-кратное снижение по сравнению с вводом. Наилучшим образом амплифицировался MVA-HLR с 30-кратным повышением по сравнению с вводом, и за ним следует MVA-Vero с 5-кратным повышением по сравнению с вводом.
Наиболее интересным было сравнение кинетики роста различных вирусов в линиях клеток человека. В отношении репродуктивной репликации в клетках 143B (линия клеток рака кости человека) было показано, что MVA-Vero является единственным, проявляющим амплификацию по сравнению с вводом (3-кратное повышение). Все другие вирусы не амплифицировались по отношению к вводу, но имелась большая разница между MVA-HLR и MVA-BN и MVA-575. MVA-HLR был "пограничным" (1-кратное снижение по отношению к вводу), а в отношении MVA-BN было показано наибольшее подавление (30-кратное снижение по отношению к вводу), за которым следует MVA-575 (59-кратное снижение по отношению к вводу). В итоге MVA-BN является наивысшим в отношении подавления в клетках 143B человека.
Более того, в отношении репликации в клетках HeLa (клетках рака шейки матки человека) было показано, что в данной линии клеток MVA-HLR хорошо амплифицируется, даже лучше, чем в пермиссивных клетках BHK (HeLa = 125-кратное повышение по сравнению с вводом; BHK = 88-кратное повышение по сравнению с вводом). MVA-Vero также амплифицировался в данной линии клеток (27-кратное повышение по сравнению с вводом). Однако MVA-BN, а также, в меньшей степени, MVA-575 подавлялись в данных линиях клеток (MVA-BN = 29-кратное снижение по сравнению с вводом, а MVA-575 = 6-кратное снижение по сравнению с вводом).
Что касается репликации в клетках HaCat (клеточной линии кератиноцитов человека), было показано, что MVA-HLR хорошо амплифицируется в данной линии клеток (55-кратное повышение по сравнению с вводом). MVA-Vero адаптированный и MVA-575 проявляли амплификацию в данной линии клеток (1,2 и 1,1-кратное повышение по сравнению с вводом, соответственно). Однако MVA-BN был единственным, который проявлял подавление (5-кратное снижение по сравнению с вводом).
В заключение, можно утверждать, что MVA-BN является наиболее подавляемым вирусным штаммом данной группы вирусов: MVA-BN проявляет предельное подавление в линиях клеток человека, показывая отношение амплификации от 0,05 до 0,2 в клетках почек эмбриона человека (293: ECACC No. 85120602) (данные не включены в таблицу 1), он дает далее отношение амплификации приблизительно 0,0 в клетках 143B; отношение амплификации приблизительно 0,04 в клетках HeLa; отношение амплификации приблизительно 0,22 в клетках HaCat. Кроме того, MVA-BN проявляет отношение амплификации приблизительно 0,0 в клетках CV1. Лишь в клетках Vero может наблюдаться амплификация (отношение 2,33), но не в той же степени, как в пермиссивных линиях клеток, таких как BHK и CEF (сравни с таблицей 1). Таким образом, MVA-BN является единственным штаммом MVA, демонстрирующим отношение амплификации ниже 1 во всех линиях клеток человека 143B, Hela, HaCat и 293.
MVA-575 проявляет профиль, сходный с MVA-BN, но не подавляется, как MVA-BN.
MVA-HLR хорошо амплифицируется во всех тестированных линиях клеток (за исключением клеток 143B), таким образом, его можно рассматривать как компетентный в отношении репликации во всех тестированных линиях клеток, за исключением клеток 143B. В одном случае он амплифицируется в линии клеток человека (HeLa) даже лучше, чем в пермиссивной клеточной линии (BHK).
MVA-Vero проявляет амплификацию во всех линиях клеток, но в меньшей степени, чем это демонстрирует MVA-HLR (при игнорировании результата с 143B). Тем не менее, с точки зрения подавления его нельзя рассматривать как относящегося к тому же "классу", что и MVA-BN и MVA-575.
2. Репликация in vivo
Принимая во внимание то, что некоторые штаммы MVA явно реплицируются in vitro, проверяли способность различных штаммов MVA реплицироваться in vivo с применением модели трансгенных мышей AGR129. У данной линии мышей имеются направленные повреждения генов рецептора типа I IFN (IFN-α/β) и типа II (IFN-γ) и в гене RAG. Вследствие данных повреждений у мышей отсутствует система IFN, и они не способны продуцировать зрелые B- и T-клетки, и по существу имеют тяжелое нарушение иммунитета и высокочувствительны к реплицируемому вирусу. Группы из шести мышей иммунизировали (в/б) 107 БОЕ MVA-BN, MVA-HLR или MVA-572 (применявшихся у 120000 человек в Германии) и контролировали ежедневно на предмет клинических проявлений. Все мыши, вакцинированные MVA-HLR или MVA-572, погибли в пределах 28 и 60 дней, соответственно. При вскрытии трупов были обнаружены общие признаки тяжелой вирусной инфекции в большинстве органов, и, с помощью стандартного теста по бляшкам, в яичниках был выявлен MVA (108 БОЕ). В противоположность этому, мыши, вакцинированные той же дозой MVA-BN (соответствующему хранящемуся штамму ECACC V00083008) выживали на протяжении более 90 дней, и MVA не удавалось извлечь из органов или тканей.
Взятые вместе, данные исследований in vitro и in vivo четко показывают, что MVA-BN подавлен в значительно большей степени, чем исходный и имеющийся в продаже штамм MVA-HLR.
Пример 2
Иммунологические данные и данные in vivo
Данные эксперименты были предприняты для сравнения различных доз и режимов вакцинации MVA-BN с другими MVAs.
2.1. Разные штаммы MVA различаются по своей способности стимулировать иммунный ответ
Способные к репликации штаммы коровьей оспы индуцируют сильные иммунные ответы у мышей, а в высоких дозах являются летальными. Хотя MVA являются сильно ослабленными и обладают сниженной способностью к репликации в клетках млекопитающих, между различными штаммами MVA имеются различия в степени ослабления. Действительно, MVA-BN, по-видимому, является более ослабленным по сравнению с другими штаммами MVA, даже по сравнению с исходным штаммом MVA-575. Для определения того, влияет ли данное различие в степени ослабления на эффективность MVA в индукции защитных иммунных ответов, производили сравнение различных доз MVA-BN и MVA-575 на модели летального контрольного заражения коровьей оспой. Степень защиты измеряли по снижению титров коровьей оспы в яичниках через 4 дня после контрольного заражения, поскольку это обеспечивало количественную оценку действия различных доз и штаммов MVA.
Модель летального контрольного заражения
Свободных от данного патогена 6-8-недельных самок мыши BALB/c (H-2d) (n=5) иммунизировали (в/б) различными дозами (102, 104 или 106 TCID50/мл) либо MVA-BN, либо MVA-575. MVA-BN и MVA-575 размножали на клетках CEF, очищали в сахарозе и готовили в трис, pH 7,4. Через три недели мыши получали повторную инъекцию той же дозы и того же штамма MVA с последующим через две недели летальным контрольным заражением (в/б) способным к репликации штаммом коровьей оспы. В качестве способного к репликации штамма вируса коровьей оспы (обозначаемого как "rVV") применяли либо штамм WR-L929 TK+, либо штамм IHD-J. Контрольные мыши получали вакцину-плацебо. Защиту измеряли по снижению титров в яичниках, определенных через 4 дня после контрольного заражения с помощью стандартного теста пятен. Для этого мышей забивали на 4 день после контрольного заражения, яичники выделяли, гомогенизировали в PBS (1 мл), и титры вируса определяли с помощью стандартного теста по бляшкам с применением клеток VERO (Thomson et al., 1998, J. Immunol. 160: 1717).
Мыши, вакцинированные двумя иммунизациями 104 или 106 TCID50/мл MVA-BN или MVA-575, были полностью защищены, судя по 100% снижению титров rVV в яичниках через 4 дня после контрольного заражения (фиг. 2). Вирус контрольного заражения был удален. Однако при меньших дозах наблюдались различия в степени защиты, обеспечиваемой MVA-BN или MVA-575. Мыши, получавшие две иммунизации 102 TCID50/мл MVA-575, не были защищены, судя по высоким титрам rVV в яичниках (в среднем 3,7 х 107 БОЕ +/- 2,11 х 107). Напротив, у мышей, вакцинированных той же дозой MVA-BN, индуцировалось значительное снижение (96%) титров rVV в яичниках (в среднем 0,21 х 107 БОЕ +/- 0,287 х 107). У контрольных мышей, получавших вакцину-плацебо, средний титр вируса составлял 5,11 х 107 БОЕ(+/- 3,59 х 107) (фиг. 2).
Оба штамма MVA индуцируют защитные иммунные ответы у мышей против летального контрольного заражения rVV. Хотя оба штамма MVA одинаково эффективны при более высоких дозах, MVA-BN является более эффективным, чем его родительский штамм MVA-575 в индукции защитного иммунного ответа против летального контрольного заражения rVV, что может быть связано с повышенным ослаблением MVA-BN по сравнению с MVA-575.
2.2. MVA-BN в режимах примирующей-повторной вакцинации
2.2.1.: Индукция антител против MVA после вакцинации мышей различными вакцинами против оспы
Эффективность MVA-BN сравнивали с другими штаммами MVA и коровьей оспы, ранее применявшимися для подавления оспы. Исследование включало однократные иммунизации с применением штаммов коровьей оспы Elstree и Wyeth, выращенных в клетках CEF и вводимых путем скарификации хвоста, и иммунизации с применением MVA-572, который ранее применяли в программе подавления оспы в Германии. Кроме того, было проведено сравнение MVA-BN и MVA-572 в качестве пре-вакцины с последующей вакцинацией Elstree посредством скарификации хвоста. Для каждой группы использовали восемь мышей BALB/c, и все вакцинации MVA (1x107 TCID50) производили подкожно на 0 и 3 неделях. Спустя две недели после повторной иммунизации мышей подвергали контрольному заражению коровьей оспой (IHD-J), и через 4 дня после контрольного заражения определяли титры в яичниках. Все вакцины и режимы обеспечивали 100% защиту.
Иммунные ответы, индуцированные применением данных различных вакцин или режимов, измеряли у животных перед контрольным заражением. Применяли тесты для измерения уровней нейтрализующих антител, пролиферации T-клеток, продукции цитокинов (IFN-γ против IL-4) и продукции IFN-γ T-клетками. Уровень ответов T-клеток, индуцированных MVA-BN, по результатам измерения с помощью Elispot, был в целом эквивалентен тем, которые вызывались другими MVA и вирусами коровьей оспы, что показывает биологическую эквивалентность. Еженедельный анализ титров антител против MVA после применения разных режимов вакцинации показал, что вакцинации MVA-BN значительно повышали скорость и величину ответа антител по сравнению с другими режимами вакцинации (фиг. 11). Действительно, титры антител против MVA были значительно выше (p>0,05) на 2, 4 и 5 неделях (через 1 неделю после повторной реиммунизации на 4 неделе) при вакцинации MVA-BN по сравнению с вакцинацией мышей MVA-572. После бустерной вакцинации на 4 неделе титры антител были также значительно выше в группе MVA-BN по сравнению с мышами, получавшими однократную вакцинацию штаммов коровьей оспы Elstree или Wyeth. Данные результаты четко показывают, что 2 вакцинации MVA-BN вызывает больший ответ антител по сравнению с классической однократной вакцинацией традиционными штаммами коровьей оспы (Elstree и Wyeth), и подтверждают данные раздела 1.5 о том, что MVA-BN является более иммуногенным, чем другие штаммы MVA.
2.2.2.: Режимы примирования и бустинга MVA вызывают тот же уровень защиты, что и режимы примирования ДНК и бустинга MVA на модели контрольного заражения гриппом.
Оценивали эффективность режимов примирования-бустинга MVA в индукции высокоавидных CTL ответов и сравнивали с режимами примирования ДНК/бустинга MVA, которые, как сообщалось, являются лучшими. Различные режимы оценивали с применением мышиной политопной конструкции, кодируемой либо вектором ДНК, либо MVA-BN, и сравнивали уровни CTL индукции с помощью ELISPOT, а авидность ответа измеряли как степень достигаемой защиты после контрольного заражения гриппом.
КОНСТРУКЦИИ
Плазмидная ДНК, кодирующая политоп мыши (10 эпитопов CTL, включая грипп, овальбумин), была ранее описана (Thomson et al., 1998, J. Immunol. 160: 1717). Данный политоп мыши был вставлен в делеционный сайт II MVA-BN, размножен в клетках CEF, очищен в сахарозе и представлен в трис, pH 7,4.
МЕТОДИКИ ВАКЦИНАЦИИ
В настоящем исследовании конкретного патогена применяли 6-8-недельных самок мыши BALB/c (H-2d). Для анализа ELISPOT использовали группы из 5 мышей, а в экспериментах с контрольным заражением гриппом применяли по 6 мышей на группу. Мышей вакцинировали с применением разных режимов примирования-бустинга с использованием MVA или ДНК, кодирующей политоп мыши, как подробно описано в результатах. Для иммунизаций ДНК мышей анестезировали и затем вводили однократную инъекцию 50 мкг свободной от эндотоксина плазмидной ДНК (в 50 мкл PBS) в четырехглавую мышцу под наркозом. Примирующие иммунизации с применением MVA осуществляли либо путем внутривенного введения 107 БОЕ MVA-BN на мышь, либо путем подкожного введения 107 БОЕ или 108 БОЕ MVA-BN на мышь. Бустинговые иммунизации проводили через три недели после примирующих иммунизаций. Бустинг плазмидной ДНК производили тем же образом, что и примирующую иммунизацию ДНК (смотри выше). Для установления CTL ответов проводили стандартные анализы ELISPOT (Schneider et al., 1998, Nat. Med. 4; 397-402) на спленоцитах через 2 недели после последней бустинговой иммунизации с применением эпитопного пептида CTL гриппа (TYQRTRALV), эпитопного пептида P. Berghei (SYIPSAEKI), эпитопного пептида Cytomegalovirus (YPHFMPTNL) и/или эпитопного пептида LCV (PRQASGVYM).
Для экспериментов с контрольным заражением мышей анестезировали и инфицировали и/н сублетальной дозой распространенного вируса гриппа, Mem71 (4,5 x 105 БОЕ в 50 мл PBS). На 5 день после инфицирования легкие выделяли, и титры вируса определяли в двух параллелях на линии клеток почек собаки Madin-Darby с применением стандартного теста бляшек гриппа.
РЕЗУЛЬТАТЫ:
При применении одной вакцины ДНК CTL индукция на 4 H-2d эпитопа, кодируемых политопом мыши, была плохой, и лишь на два эпитопа P. Berghei (SYIPSAEKI) и вируса лимфоцитарного хориоменингита (PRQASGVYM) можно было выявить слабые ответы. В противоположность этому, при применении режима примирования ДНК и бустинга MVA (107 БОЕ MVA-BN, вводимого подкожно) наблюдалось значительно большая CTL индукция на SLY (8-кратное повышение) и PRQ (3-кратное повышение), и ответы также наблюдались в отношении третьего эпитопа, цитомегаловируса мыши (YPHFMPTNL) (фиг. 3A). Однако применение 107 БОЕ MVA-BN, вводимого подкожно, при гомологичном режиме примирования-бустинга индуцировало тот же уровень ответов, что и ДНК с последующим MVA-BN (фиг. 3A). Неожиданно оказалось, что при применении одной иммунизации MVA-BN (107 TCID50) нет существенной разницы в количестве CTL, индуцированных против трех эпитопов, что указывает на то, что вторичная иммунизация MVA-BN не повышает существенно CTL ответы.
Ранее было показано, что подкожное введение 107 БОЕ MVA является самым неэффективным путем и концентрацией вируса для вакцинации при применении других штаммов MVA, в особенности по сравнению с внутривенными иммунизациями (Schneider et al 1998). Для определения оптимальных режимов иммунизации указанный выше эксперимент был повторен с изменением либо количества вируса, либо способа введения. В одном эксперименте вакцинации 107 БОЕ MVA-BN были осуществлены внутривенно (фиг. 3B). В другом эксперименте 108 БОЕ MVA-BN вводили подкожно (фиг. 3C). В данных экспериментах иммунизации примирования-бустинга MVA-BN индуцировали большие средние количества CTL против всех трех CTL эпитопов по сравнению с режимами примирования ДНК и бустинга MVA. Также в отличие от 107 БОЕ MVA-BN, введенного подкожно, иммунизация 107 БОЕ MVA-BN, введенного внутривенно, и иммунизация 108 БОЕ MVA-BN, введенного подкожно, значительно повышала CTL ответы, четко показывая, что MVA-BN может быть применен для усиления CTL ответов при наличии предсуществующего иммунитета против вектора.
2.2.3.: Эффективность вакцины MVA-BN nef у макак резус инфицированных SIV.
Определение эффективности вакцины MVA-BN nef осуществляют путем оценки количества вируса и задержки заболевания после контрольного заражения вирулентным первичным изолятом SIV. Кроме того, исследованием должно было быть установлено, может ли MVA-BN быть применен для безопасно усиленных иммунных ответов у обезьян с иммунной недостаточностью при наличии предсуществующего иммунитета против MVA.
МЕТОДИКИ ВАКЦИНАЦИИ
Две группы (n=6) макак резус (Macaca malalta) вакцинировали болюсной внутримышечной инъекцией либо одним MVA-BN, либо рекомбинантным MVA-BN nef на 0, 8 и 16 неделях. На 22 неделю всех обезьян подвергали контрольному внутривенному заражению 50 MID50 патогенным, связанным с клетками материалом SIV (1XC) из первичных некультивированных PBMC макак резус. Клиническое состояние животных отслеживали с высокой частотой, и регулярно отбирали образцы крови для измерения виремии, иммунологических показателей и полного спектра гематологических параметров и параметров клинической химии крови. Животных, у которых СПИД развивался в форме болезни, забивали, а выживших обезьян отслеживали в течение 99 недель после вакцинации. На 100 неделе выживших обезьян иммунизировали в/м MVA-BN tat, и животные получали дополнительные иммунизации тем же MVA-BN tat на 102 и 106 неделях.
Не наблюдалось неблагоприятных эффектов после любой вакцинации либо MVA-BN, либо MVA-BN nef. После инфицирования обезьян SIV резко возрастал уровень виремии с максимумом через две недели после инфицирования (фиг. 4). Из-за больших стандартных отклонений в пределах групп не было значимой разницы между средними уровнями SIV в группах, вакцинированных MVA-BN nef или MVA-BN. Однако количество SIV в группе, вакцинированной MVA-BN nef, было в целом в 10 раз ниже по сравнению с контрольной (MVA-BN) группой. Более того, через 35 недель после инфицирования (начало периода наблюдений) лишь 1 из шести обезьян, вакцинированных MVA-BN nef, пришлось умертвить в связи с тяжестью болезни по сравнению с 4 из 6 животных в контрольной группе (фиг. 5). Развитие болезни четко коррелировало с большим количеством вируса, в связи с чем животных наблюдали дополнительно в течение 29 недель после инфицирования. Вакцина MVA-BN nef, очевидно, замедляла прогрессирование болезни по сравнению с контрольной группой, и даже на 46 неделе после инфицирования 5 из 6 животных с MVA-BN nef оставались живы (фиг. 5). Однако к 59 неделе после инфицирования еще двое животных из группы, вакцинированной nef, были умерщвлены, в результате чего в живых осталось пять животных (три из группы MVA-BN nef и два, вакцинированных MVA-BN). Определение титров антител, выработанных против MVA-BN у данных 12 обезьян, четко показало, что MVA-BN может усиливать иммунный ответ даже при наличии предсуществующего иммунитета против MVA (фиг. 6). После примирующей иммунизации MVA-BN или MVA-BN nef у всех обезьян индуцировался ответ антител против MVA со средним титром 1000. Данный ответ антител значительно увеличивался после повторной иммунизации, что четко показывает, что MVA может быть применен для индукции иммунного ответа примированием-бустингом у здоровых обезьян. Данные титры антител постепенно снижаются, хотя к 49 неделе после иммунизации титры выходили на плато, в результате чего средние титры против MVA на 99 неделе равнялись 2000.
Пять выживших обезьян инфицировали SIV и имели иммунодефицит с показателем CD4 ниже 400/мкл крови. Для исследования влияния применяемого MVA-BN у обезьян с нарушенным иммунитетом пять животных вакцинировали три раза MVA-BN tat на 100, 102 и 106 неделях после исходной вакцинации. Первая иммунизация MVA-BN tat значительно повышала ответ антител против MVA у данных обезьян с нарушенным иммунитетом, который дополнительно повышался третьей иммунизацией шесть недель спустя (фиг 6). Данные результаты дополнительно показывают, что MVA-BN может усиливать иммунные ответы при наличии значительного предсуществующего иммунитета к MVA, даже у обезьян с нарушением иммунной системы. Хотя иммунный ответ обезьян усиливался после иммунизации MVA-BN tat, уровни SIV оставались стабильными, что указывает на то, что иммунизации MVA-BN являются безопасными и не влияют на уровень SIV у обезьян с нарушением иммунной системы (фиг. 7).
Данное исследование показало, что MVA-BN может примировать-усиливать иммунные ответы у обезьян резус с нарушением иммунной системы и что иммунизации MVA-BN являются безопасными и не влияют на уровень виремии у животных, инфицированных SIV. Задержка в развитии СПИД-подобной болезни у животных, вакцинированных вакциной MVA-BN nef, указывает на успешное формирование иммунного ответа на nef.
2.2.4.: Терапевтическая вакцинация инфицированных SIV обезьян, подвергнутых противоретровирусному лечению
Терапевтическая вакцина против ВИЧ на основе MVA-BN, по-видимому, может быть применена у индивидуумов, подвергаемых противоретровирусному лечению. В связи с этим в настоящем исследовании изучали безопасность (действие на уровень SIV) и эффективность рекомбинантных MVA, кодирующих множество антигенов SIV (gag, pol, env, rev, tat и nef) у инфицированных SIV обезьян, лечившихся PMPA. PMPA представляет собой нуклеозидный аналог и эффективен против ВИЧ и SIV (Rosenwirth, B. et al., 2000, J Virol 74, 1704-11).
КОНСТРУКЦИИ
Все рекомбинантные конструкции MVA размножали на клетках CEF, очищали в сахарозе и составляли в трис pH 7,4.
МЕТОДИКА ВАКЦИНАЦИИ
Три группы (n=6) обезьян резус (Macaca mulatta) инфицировали 50 MID50 патогенного первичного изолята SIV (1XC) и затем ежедневно лечили PMPA (60 мг/кг, вводимого п/к) в течение 19 недель. На 10 неделю животных вакцинировали рекомбинантным MVA-BN (в/м) или физиологическим раствором, и животные получали идентичные вакцинации спустя 6 недель. Группа 1 получала смесь MVA gag-pol и MVA-env, группа 2 получала MVA-tat, MVA-rev и MVA-nef, а группа 3 получала физиологический раствор. Клиническое состояние животных часто проверялось, и образцы крови регулярно отбирались для измерения виремии, иммунологических показателей и полного спектра гематологических параметров и параметров клинической химии крови.
У всех животных устанавливался высокий уровень SIV с максимумом через 2 недели после инфицирования (фиг. 8). После ежедневного лечения PMPA уровни SIV снижались и стабилизировались на низком уровне к 9 неделе. Как и в предыдущем исследовании, вакцинация MVA на 10 и 16 неделях не влияли на уровни SIV, что свидетельствует о том, что MVA-BN является безопасным вакцинным вектором для животных с нарушением иммунной системы. После прекращения лечения животных PMPA (21 неделя) уровни SIV увеличиваются. Хотя у трех животных в группе 1 уровни SIV были снижены по сравнению с контрольной группой 3, достоверных различий среднего количества SIV между любыми группами после прекращения лечения PMPA не было (фиг. 8). С помощью ELISA для лизатов инфицированных SIV T-клеток было показано, что у животных всех групп возникает ответ антител против SIV к 4 неделе после инфицирования (фиг. 9). Титр антител против SIV в контрольной группе (физиологический раствор) снижался в ходе лечения PMPA и быстро увеличивался после прекращения лечения PMPA, что отражает снижение и последующее повышение в уровне SIV в период противоретровирусной терапии (фиг. 9). Сходный паттерн титра антител против SIV наблюдался в группе 2, получавшей MVA-tat, MVA-rev и MVA-nef, что, возможно, отражает пониженную экспрессию данных регуляторных белков в лизатах инфицированных SIV T-клеток, показанную с помощью ELISA. В противоположность этому, однако, титры антител против SIV в группе 1 возрастали после вакцинаций MVA gag-pol и MVA-env на 10 неделе, что указывает на то, что MVA-BN может усиливать иммунный ответ против SIV у инфицированных (SIV) животных, подвергнутых противоретровирусной терапии. Важно, что титры антител против SIV повышались после повторной иммунизации на 16 неделе, что вновь показывает, что MVA может усиливать иммунные ответы у животных с нарушением иммунной системы, даже при наличии предсуществующего иммунитета против MVA (фиг. 8). Титры антител против MVA в группе 1 также отражали данный паттерн с формированием ответа антител после первичной иммунизации, и этот ответ значительно усиливался после вторичной вакцинации (фиг. 10).
Амплификация вируса выше уровня ввода через 4 дня после инфицирования
Отношение амплификации=выход TCID50 ввод TCID50.
Величины выражены в TCID50.
Источники информации
1. Schneider, J., Gilbert, SC., Blanchard, TJ., Hanke, Т., Robson, KJ., Hannan, CM., Becker, M., Sinden, R., Smith, GL., and Hill, AVS. 1998. Enhanced immunogenicity for CD8+ T cell induction and complete efficacy of malaria DNA vaccination by boosting with modified vaccinia virus Ankara. Nat. Med. 4; 397-402.
2. Thomson, SA., Sherritt, MA., Medveczky, J., Elliott, SL, Moss, DJ., Fernando, GJP., Brown, LE., and Suhrbier, A. 1998. Delivery of multiple CDS cytotoxic T cell epitopes by DNA vaccination. J. Immunol. 160: 1717.
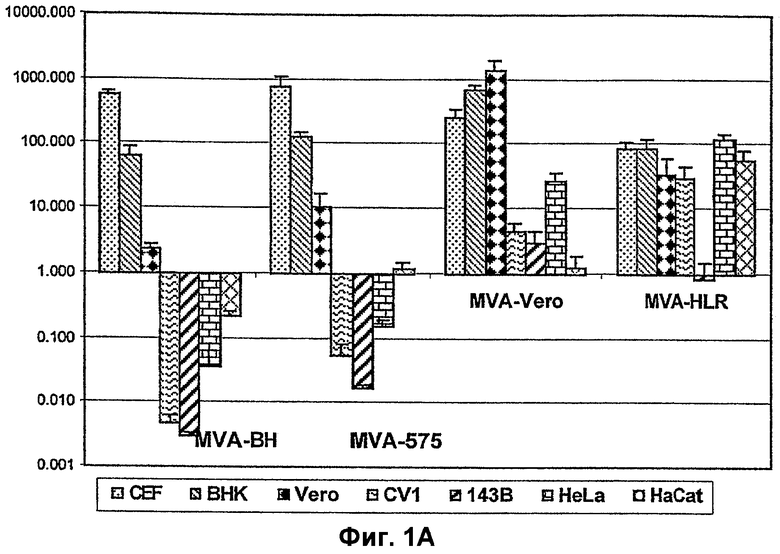
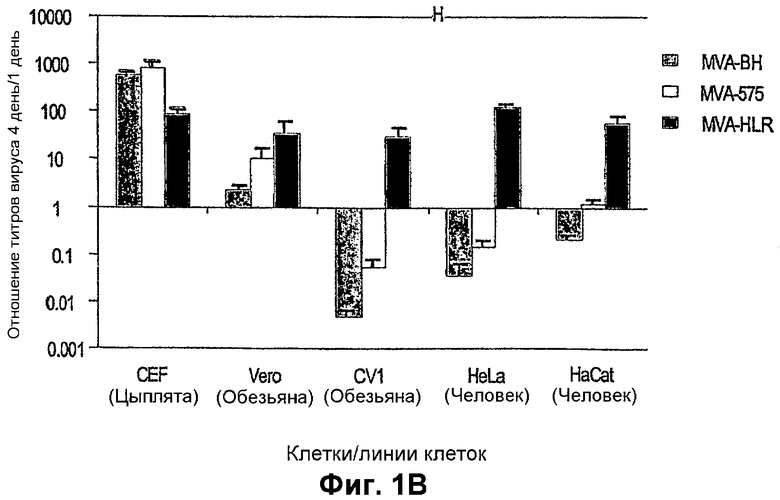
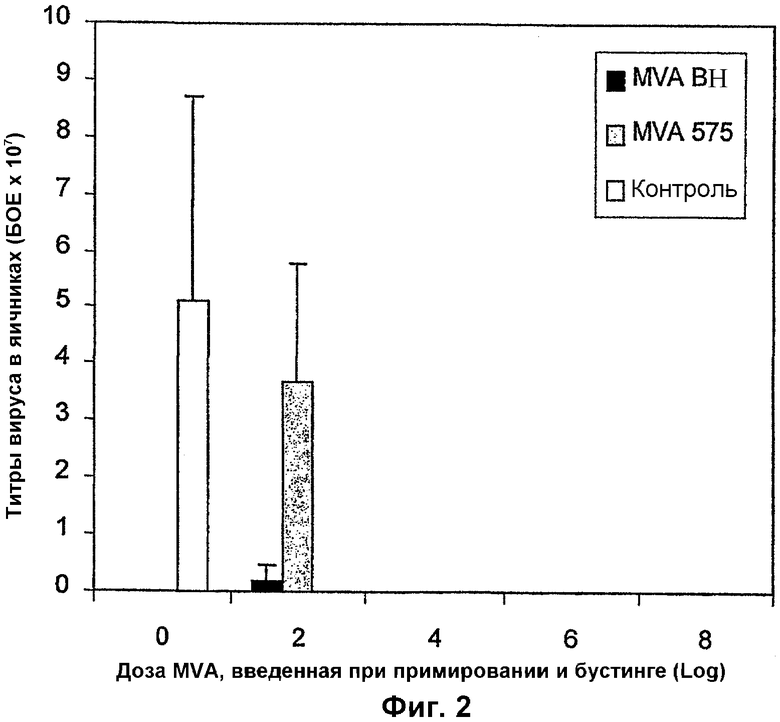
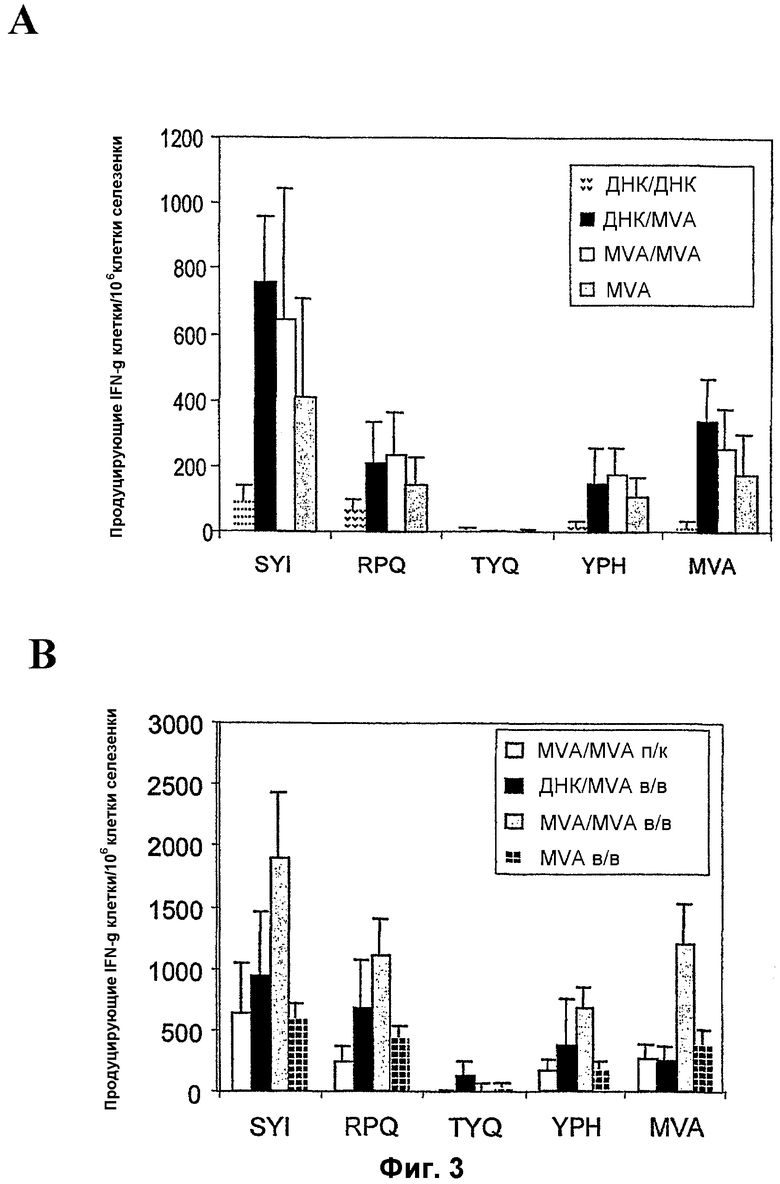
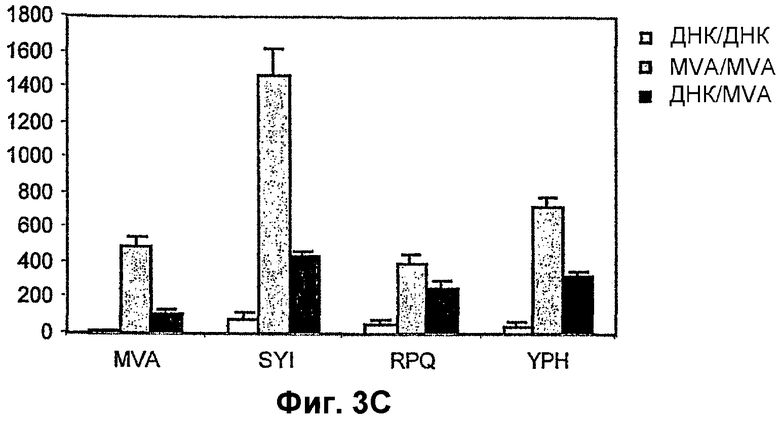
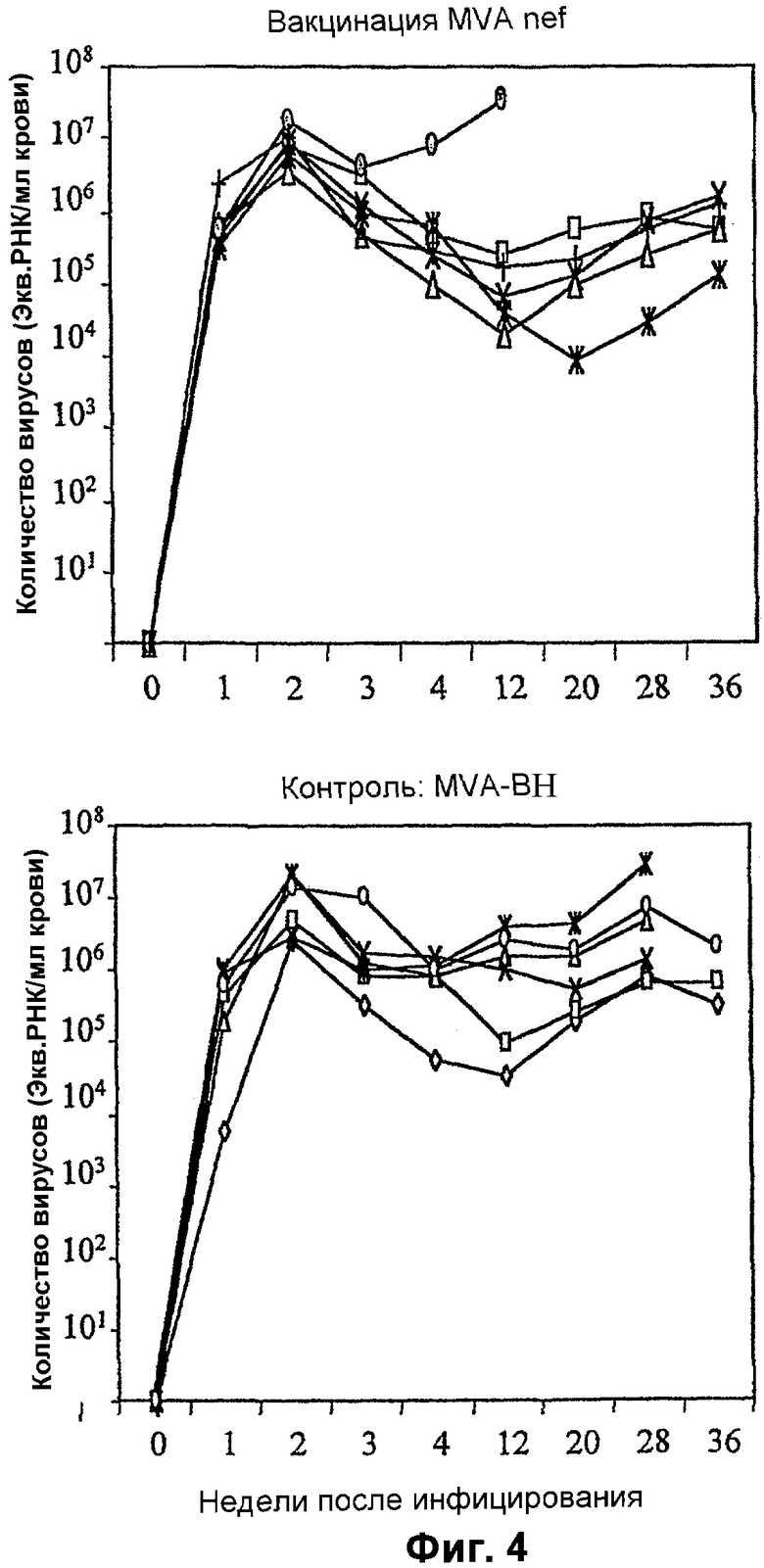
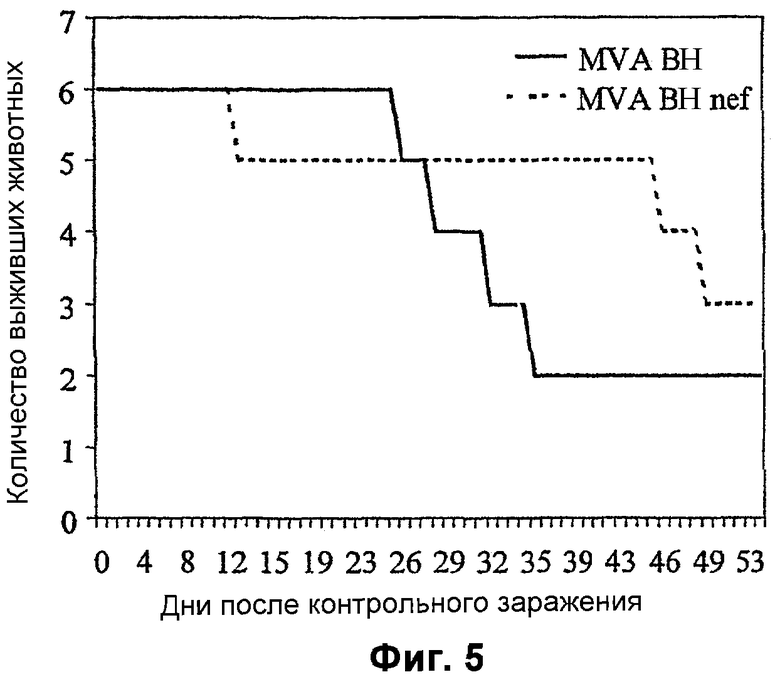

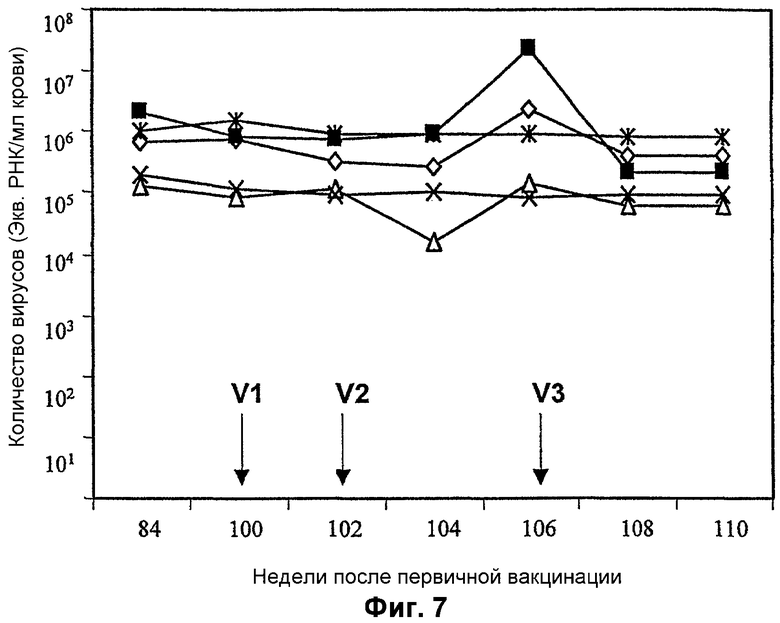
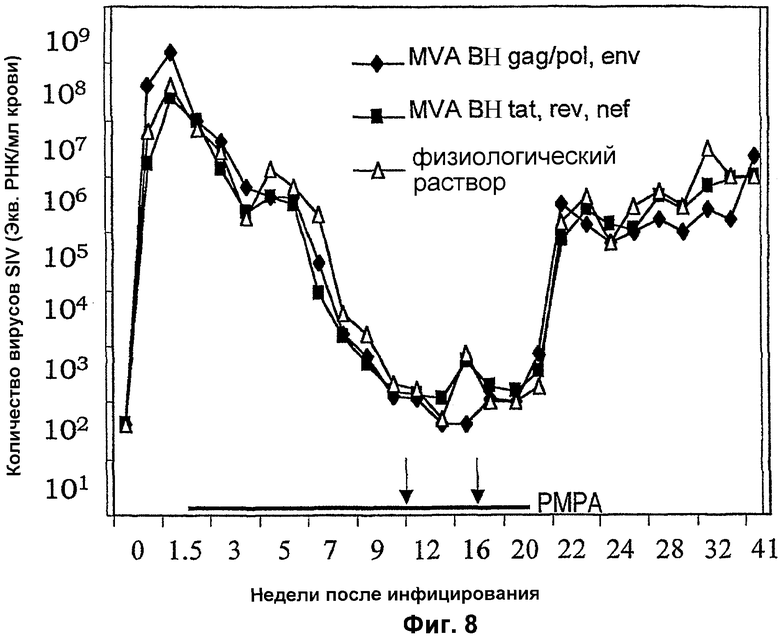
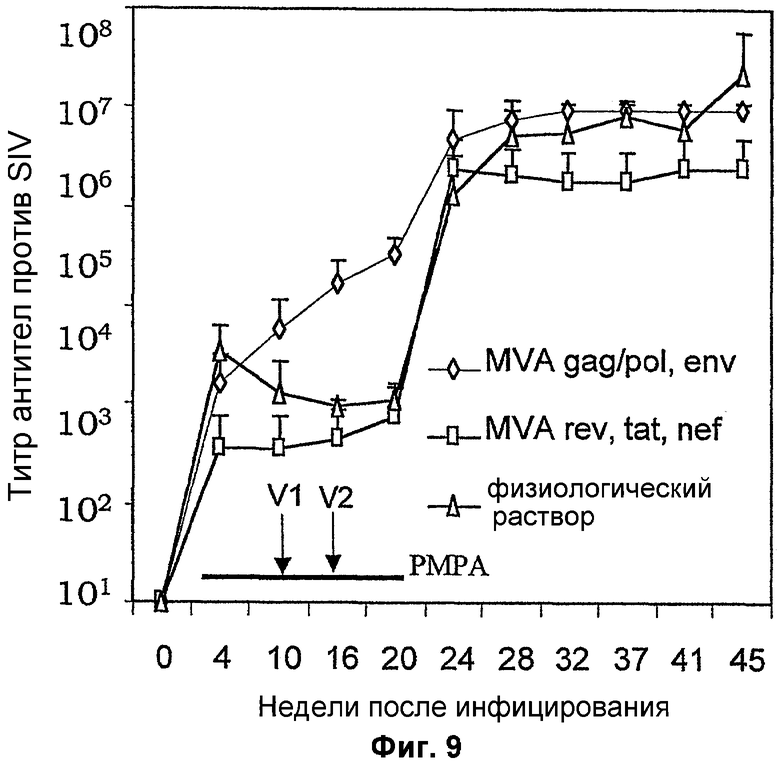
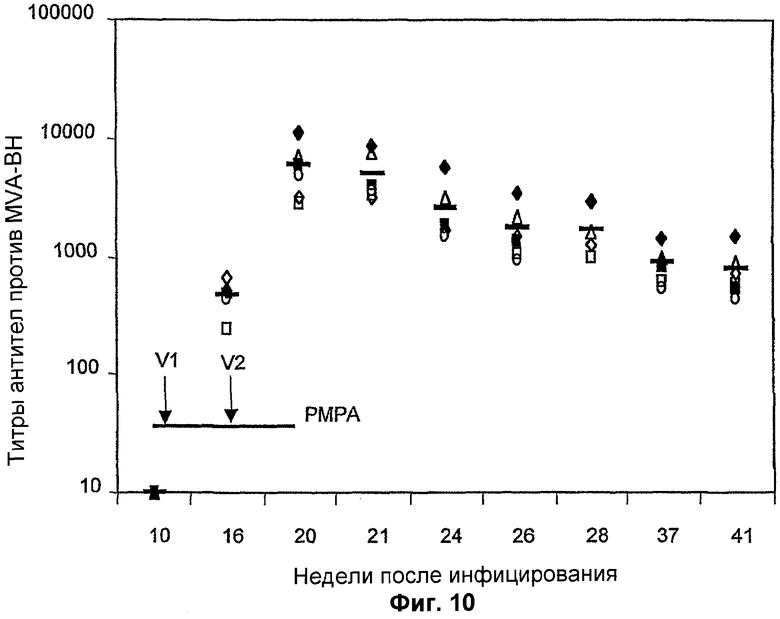
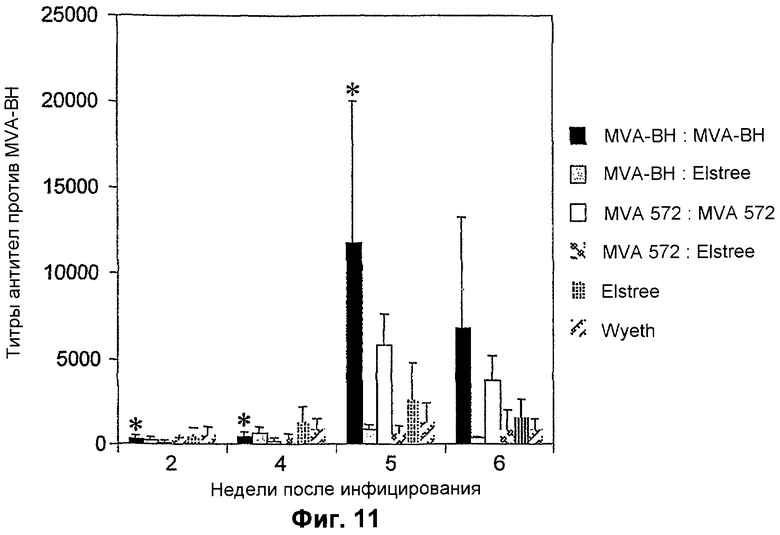
Изобретение относится к области биотехнологии, вирусологии и медицины. Предложен ослабленный вирус, который происходит от модифицированного вируса коровьей оспы Ankara. Вирус характеризуется потерей способности к своей репродукции путем репликации в клеточных линиях человека. Кроме того, описаны применение вируса или его рекомбинантных вариантов в качестве лекарства или вакцины. Дополнительно предлагается способ индукции иммунного ответа даже у больных с нарушенным иммунитетом, у больных с исходно существующим иммунитетом к вирусу вакцины или у больных в процессе проведения противовирусной терапии. Изобретение может быть использовано в медицине и ветеринарии. 14 н. и 72 з.п. ф-лы, 11 ил., 1 табл.
(i) способность к репродукции путем репликации в фибробластах эмбрионов цыплят (CEF) и в линии клеток почек детенышей хомячка ВНК при отсутствии способности к репродукции путем репликации в линиях клеток человека, и
(ii) отсутствие способности к репликации in vivo в мышах с существенным нарушением иммунитета, где мыши с существенным нарушением иммунитета представляют собой мышей, которые не способны продуцировать зрелые В и Т клетки,
в качестве фармацевтического средства для экспрессии в животном, включая человека, терапевтической нуклеиновой кислоты, пептида или белка, в качестве фармацевтического средства для влияния на иммунный ответ и/или для индукции иммунного ответа у животного, включая человека, в качестве средства, действующего как адъювант в отношении животного, включая человека, в качестве средства, активирующего, поддерживающего или подавляющего иммунную систему животного, включая человека, или в качестве вакцины для животного, включая человека.
(a) инфицирование клетки-хозяина вирусом по любому из пп.1-9,
(b) культивирование инфицированной клетки-хозяина в подходящих условиях и
(c) выделение и/или обогащение пептида, белка и/или вируса, продуцируемого указанной клеткой-хозяином.
введение обычного доступного штамма вируса коровьей оспы, предпочтительно MVA 575, в клетки, отличные от клеток человека, в которых вирус способен к репродукции путем репликации, где клетки, отличные от клеток человека, предпочтительно выбраны из клеток CEF и клеточной линии ВНК,
выделение/обогащение вирусных частиц из данных клеток и
анализ того, будет ли полученный вирус иметь биологические свойства, как указано в любом из пп.1 и 2,
где указанные выше стадии могут повторяться до тех пор, пока не будет получен вирус с желаемыми репликативными характеристиками.
| WO 9702355, 23.01.1997 | |||
| US 5185146, 09.02.1993. |

