Изобретение относится к ретровирусным векторам, включая вектор, который подвергается промоторной конверсии (ПроКон вектор). Векторная система применима в качестве носителя для доставки гена при целевой генной терапии.
Предпосылки создания изобретения
Использованию ретровирусных векторов для генной терапии уделялось большое внимание, и в настоящее время это является методом выбора для переноса терапевтических генов в ряде усовершенствованных методик как в США, так и в Европе (Kotani et al., 1994). Однако в большинстве этих методик необходимо, чтобы инфицирование целевых клеток ретровирусным вектором, несущим терапевтический ген, происходило in vitro, и успешно инфицированные клетки затем возвращают больному индивидууму (Rosenberg et al., 1992; обзор см. у Anderson, 1992). Такие ex vivo методики генной терапии являются идеальными в медицине для коррекции состояний, при которых целевая клеточная популяция может быть легко выделена (например, лимфоциты). Дополнительно, ех vivo инфицирование целевых клеток дает возможность введения больших количеств концентрированного вируса, который может быть строго и безопасно испытан перед использованием.
К сожалению, единственный раздел возможного применения генной терапии состоит из целевых клеток, которые могут быть легко выделены, культивированы и затем снова интродуцированы. Кроме того, сложная технология и связанная с ней высокая стоимость ех vivo генной терапии, по существу, мешают ее широкому использованию по всему миру. Легко осуществимая и экономически эффективная генная терапия в будущем будет нуждаться в in vivo методе, при котором вирусный вектор или клетки, продуцирующие вирусный вектор, непосредственно вводятся пациенту в виде инъекции или простой имплантацией продуцирующих ретровирусный вектор клеток.
При этом виде метода in vivo, конечно, возникает ряд новых проблем. Во-первых, и прежде всего, нужно обратиться к соображениям безопасности. Вирус будет продуцироваться, возможно, в результате имплантации продуцирующих вирус клеток, и не будет возможности предварительно проверить продуцируемый вирус. Важно осознать конечный риск, связанный с использованием таких систем, также как и стараться получить новые системы, которые снижают этот риск до минимума.
Ретровирусные векторные системы состоят из двух компонентов (фиг.1):
1) сам ретровирусный вектор, являющийся модифицированным ретровирусом (векторной плазмидой), в котором гены, кодирующие вирусные белки, были замещены терапевтическими генами, которые нужно перенести в целевые клетки. Так как замещение генов, кодирующих вирусные белки, существенно нарушает функционирование вируса, оно должно быть восполнено вторым компонентом в системе, который обеспечивает недостающие вирусные белки модифицированному вирусу.
Вторым компонентом является:
2) клеточная линия, которая продуцирует большие количества вирусных белков, у которой, однако, отсутствует способность продуцировать репликационный компонент вируса. Эта клеточная линия известна как упаковывающая клеточная линия-носитель и состоит из клеточной линии, трансфицированной второй плазмидой, несущей гены, обеспечивающие модифицированному ретровирусному вектору способность к упаковке.
Для получения упакованного вектора векторную плазмиду трансфицируют в упаковывающую клеточную линию. При этих условиях модифицированный ретровирусный геном, включающий вставленные терапевтические и маркерные гены, транскрибируется из векторной плазмиды и упаковывается в модифицированные ретровирусные частицы (рекомбинантные вирусные частицы). Этот рекомбинантный вирус затем используется для инфицирования целевых клеток, в которых векторный геном и любые маркерные или терапевтические гены становятся интегрированными в целевую клеточную ДНК. Клетка, инфицированная такой рекомбинантной вирусной частицей, не может продуцировать новый векторный вирус, так как в этих клетках отсутствуют вирусные белки. Однако ДНК вектора, несущего терапевтические и маркерные гены, интегрируется в ДНК клетки и может теперь экспрессироваться в инфицированной клетке.
По существу, случайная интеграция провирусной формы ретровирусного генома в геном инфицированной клетки (Varmus, 1988) приводила к идентификации ряда клеточных протоонкогенов посредством их инсерционной активации (Varmus, 1988; van Lohuizen and Berns, 1990). Возможность того, что подобный механизм может вызывать рак у пациентов, которых лечили ретровирусными векторами, несущими терапевтические гены, предназначенные для лечения существовавшего заболевания, поставила вновь и вновь возникающую этическую проблему. Многие исследователи должны согласиться с тем, что возможность репликации дефектного ретровирусного вектора, такого, как все используемые в настоящее время, интегрирующего в или рядом с клеточным геном, участвующим в регуляции клеточной пролиферации, исчезающе мала. Однако, в основном, признано, что взрывной рост популяции репликационно компетентного ретровируса от единственного инфицирования со временем обеспечит достаточное число интеграции, чтобы сделать такую фенотипическую интеграцию очень реально возможной.
Ретровирусные векторные системы оптимизируются так, чтобы свести к минимуму шансы репликации компетентного вируса, когда он присутствует. Однако хорошо документально подтверждено, что число рекомбинаций между компонентами ретровирусной векторной системы может привести к воспроизведению потенциально патогенного репликационно компетентного вируса, и был сконструирован ряд генераций векторных систем для сведения к минимуму этого риска рекомбинации (обзор сделан у Salmons and Gunzburg, 1993).
Дополнительное соображение при рассмотрении использования генной терапии in vivo, как с точки зрения безопасности, так и с чисто практической точки зрения, состоит в направленности ретровирусных векторов. Ясно, что терапевтические гены, несомые векторами, не должны без разбора экспрессироваться во всех тканях и клетках, а скорее только в нужных целевых клетках. Это особенно важно, если гены, которые нужно перенести, являются генами токсина, нацеленными на удаление конкретных опухолевых клеток. Удаление других, не целевых клеток было бы, безусловно, нежелательно.
Ранее был описан ряд ретровирусных векторных систем, которые должны дать возможность направленной доставки несомых терапевтических генов (Salmons and Gunzburg, 1993). Большинство этих методов включают или лимитирование инфицирования предопределенными типами клеток или использование гетерологичных промоторов для управления экспрессией связанных гетерологичных терапевтических или маркерных генов в специфических типах клеток. Использованы гетерологичные промоторы, которые должны управлять экспрессией связанных генов только в типе клеток, в которых этот промотор в норме активен. Эти промоторы предварительно встраивались в сочетании с маркерным или терапевтическим геном в структуру ретровирусных векторов вместо gag, pol или env генов.
Ретровирусный длинный концевой повтор (ДКП) на концах этих генов несет ретровирусный промотор, который, в основном, является неспецифичным в том, что он может управлять экспрессией в многих различных типах клеток (Majors, 1990). Сообщалось о промоторной интерференции между ДКП промотором и гетерологичными внутренними промоторами, такими как тканевые специфичные промоторы, описанные выше. К тому же известно, что ретровирусные ДКП несут сильные энхансеры, которые могут, или независимо, или в сочетании с ретровирусным промотором, влиять на экспрессию клеточных генов вблизи сайта интеграции ретровируса. Этот механизм, как было показано, отвечает за онкогенность у животных (van Lohuizen and Berns). Эти два наблюдения побудили на разработку самоинактивирующихся векторов (СИН), в которых ретровирусные промоторы функционально инактивируются в целевых клетках (РСТ WO 94/29437). Дополнительные модификации этих векторов включают вставку промоторных генных кластеров в область ДКП для создания двойных копий векторов (РСТ WO 89/11539). Однако в обоих этих векторах гетерологичные промоторы, вставленные или в структуру вектора, или в ДКП область, непосредственно связаны с маркерным/терапевтическим геном.
В ранее описанном СИН векторе, упомянутом выше, несущем делетированную 3'ДКП (РСТ WO 94/29437), используется, кроме того, сильный гетерологичный промотор, такой как промотор цитомегаловируса (ЦМВ), вместо ретровирусного 5'ДКП промотора (5'ДКП без U3) для управления экспрессией векторной конструкции в упаковывающей клеточной линии. Гетерологичная сигнальная последовательность полиаденилирования также включена в 3'ДКП (РСТ WO 94/29437).
Предметом данного изобретения является конструкция нового ретровирусного вектора, который может использоваться в качестве безопасного переносчика генов для целевой генной терапии со сниженной способностью подвергаться рекомбинации с упаковывающей конструкцией. Этот новый вектор несет гетерологичный промотор и/или регуляторные элементы в 3'ДКП, которые после инфицирования становятся удвоенными и транслоцируются в 5'ДКП в целевых клетках, контролируя во времени экспрессию маркерных/терапевтических генов, не связанных непосредственно с промотором, но все же вставленных в структуру вектора. Этот вектор не подвергается самоинактивации, но взамен промотор обменивается, что дало появление названия РrоСоn (ПроКон) по Promoter Conversion - промоторной конверсии.
Так как промоторное превращение не приводит к самоинактивации, ретровирусный вектор будет транскрипционно активен в целевых клетках. Однако обе ДКП будут состоять в большей степени из гетерологичных промоторных/энхансерных последовательностей в целевых клетках. Это будет снижать вероятность того, что интегрированный вектор в целевых клетках будет подвергаться инактивации через длительные промежутки времени, как было описано для общепринятых векторов (Xu et al., 1989), а также будет снижать возможность рекомбинации с эндогенными ретровирусными последовательностями с генерацией потенциально патогенного репликационно компетентного вируса, увеличивая безопасность системы.
В этом изобретении 5'ДКП ретровирусной векторной конструкции не модифицирована, и экспрессия вирусного вектора в упаковывающих клетках регулируется нормальным ретровирусным промотором U3. Возможно нормальное ретровирусное полиаденилирование, и не включены никакие гетерологичные сигнальные последовательности полиаденилирования в 3'ДКП. Это важно для разработки стратегии генной терапии in vivo, так как нормальная физиологическая регуляция посредством нормального вирусного промотора и, возможно также, с участием нормальной вирусной регуляции полиаденилирования будет существовать в течение длительных периодов in vivo, пока несущие клетки продуцируют рекомбинантный вирус.
Дополнительная модификация этого нового ретровирусного вектора ускоряет включение клеточных последовательностей вместо гетерологичного промотора и/или регуляторных элементов. Это должно дать возможность более высокой селективности для сайт-специфичной рекомбинации с клеточными последовательностями для направления интеграции ретровирусных векторов в конкретные сайты в геноме клетки-хозяина (Sailer, 1994).
Для получения предшествующего и других объектов изобретение представляет ретровирусный вектор, испытывающий промоторную конверсию, включающую 5'ДКП область структуры U3-R-U5; одну или более последовательностей, выбираемых из кодирующих и некодирующих последовательностей; и 3'ДКП область, состоящую из полностью или частично делетированной U3 области, в которой делетированная U3 область заменяется полилинкерной последовательностью, за которой следует R и U5 область.
Указанная полилинкерная последовательность несет по крайней мере один уникальный рестрикционный сайт и содержит предпочтительно по крайней мере одну вставку фрагмента гетерологичной ДНК. Указанный фрагмент гетерологичной ДНК предпочтительно выбирается из регуляторных элементов и промоторов, которые предпочтительно являются специфичными для целевых клеток по их экспрессии, но могут также быть фрагментом ДНК без регуляторной функции.
Указанный фрагмент гетерологичной ДНК является предпочтительно гомологичным по отношению к одной или более клеточных последовательностей. Регуляторные элементы и промоторы предпочтительно могут регулироваться транс-активирующими молекулами.
Дополнительные объекты, признаки и преимущества будут очевидны из последующего описания и предпочтительных осуществлений изобретения.
Регуляторные элементы, специфичные для целевых клеток, и промоторы выбираются из одного или более элементов из группы, состоящей из специфичных регуляторных элементов и промоторов сывороточного кислотного белка (СКБ=WAP), мышиного вируса опухоли молочных желез (МВОМЖ=MMTV), β-лактоглобулина и казеина, специфичных регуляторных элементов и промоторов поджелудочной железы, включая регуляторные элементы и промоторы карбоангидразы II и β-глюкокиназы, специфичные лимфоцитные регуляторные элементы и промоторы, включая регуляторные элементы и промоторы иммуноглобулина и МВОМЖ специфичные лимфоцитные регуляторные элементы и промоторы и МВОМЖ специфичные регуляторные элементы и промоторы, придающие чувствительность к глюкокортикоидным гормонам или направляющие экспрессию в молочные железы. Указанные регуляторные элементы и промоторы предпочтительно регулируют экспрессию по крайней мере одной из кодирующих последовательностей указанного ретровирус-ного вектора. ДКП области выбираются из по крайней мере одного элемента из группы, состоящей из ДКП вируса мышиной лейкемии (MLV=ВМЛ), мышиного вируса опухоли молочных желез (МВОМЖ), вируса мышиной саркомы (MSV=ВМС), вируса иммунодефицита обезьян (SIV=ВИО), вируса иммунодефицита человека (ВИЧ), вируса Т-клеточной лейкемии человека (ВТЛЧ=HTLV), вируса иммунодефицита кошек (ВИК= FIV), вируса лейкемии кошек (ВЛК=FELV), вируса лейкемии крупного рогатого скота (ВЛКРС = BLV) и обезьяньего вируса Мэзон-Файзера (ОВМФ=MPMV).
Основой для ретровирусного вектора является или вектор BAG (Price et al. , 1987) или вектор LXSN (Miller and Rosman, 1989).
Кодирующая последовательность предпочтительно выбирается из одного или более представителей группы, состоящей из маркерных генов, терапевтических генов, антивирусных генов, противоопухолевых генов, генов цитокинов.
Указанные маркерные и терапевтические гены предпочтительно выбираются из одного или более представителей группы, состоящей из гена β-галактозидазы, неомицинового гена, гена тимидинкиназы вируса герпеса простого, гена пуромицина, цитозиндезаминазного гена, гена гигромицина, гена секретируемой щелочной фосфатазы, гена гуанинфосфорибозилтрансферазы (gpt) гена, гена алкогольдегидрогеназы и гипоксантинфосфорибозилтрансферазного (ГФРТ) гена.
Другое осуществление изобретения представляет собой изменение или частичную делению по крайней мере одной ретровирусной последовательности, необходимой для интеграции ретровирусов.
В следующем осуществлении изобретения представлена ретровирусная система, включающая ретровирусный вектор, который описан выше, в качестве первого компонента, и упаковывающую клеточную линию, несущую по крайней мере одну ретровирусную или рекомбинантную ретровирусную конструкцию, кодирующую протеины, необходимые для указанного ретровирусного вектора, нуждающегося в упаковке.
Упаковывающая клеточная линия несет рекомбинантные ретровирусные конструкции, кодирующие эти ретровирусные белки, которые не кодируются в указанном ретровирусном векторе. Упаковывающая клеточная линия предпочтительно выбирается из членов группы, состоящей из psi-2, psi-Crypt, psi-AM, GP+E-86, PA317 и GP+envAM-12, или из любой из линий, суперинфицируемых рекомбинантными конструкциями, дающих возможность экспрессии поверхностных белков от других оболочечных вирусов.
Еще одно осуществление изобретения включает использование упаковывающей клеточной линии, несущей рекомбинантную ретровирусную конструкцию, дефектную по интегразной функции.
После введения ретровирусного вектора этого изобретения, который описан выше, в ретровирусную упаковывающую клеточную линию и инфицирования целевой клетки, как описано выше, получен ретровирусный провирус, у которого указанный полилинкер и любые последовательности, вставленные в указанный полилинкер в 3'ДКП, становятся удвоенными в процессе обратной транскрипции в инфицированной целевой клетке и появляются в 5'ДКП, а также в 3'ДКП получающегося в результате провируса.
Изобретение включает также мРНК ретровирусного провируса по этому изобретению и любую РНК, полученную из ретровирусного вектора по этому изобретению.
Дополнительное воплощение этого изобретения представляет нетерапевтический способ введения гомологичных и/или гетеро-логичных нуклеотидных последовательностей в клетки человека или животных in vitro и in vivo, включающий трансфицирование упаковывающей клеточной линии ретровирусной векторной системы по этому изобретению ретровирусным вектором по этому изобретению и инфицирование целевой клеточной популяции рекомбинантными ретровирусами, продуцируемыми упаковывающей клеточной линией. Нуклеотидные последовательности выбираются из одного или более представителей группы, состоящей из генов или частей генов, кодирующих белки, регуляторных последовательностей и промоторов.
Ретровирусный вектор, ретровирусная векторная система и ретровирусный провирус, а также их РНК используются для получения фармацевтической композиции для генной терапии у млекопитающих, включая человека. Кроме того, они используются для целевой интеграции в гомологичные клеточные последовательности.
Промоторная конверсия
В данном изобретении используется принцип промоторной конверсии, типичной для ретровирусов.
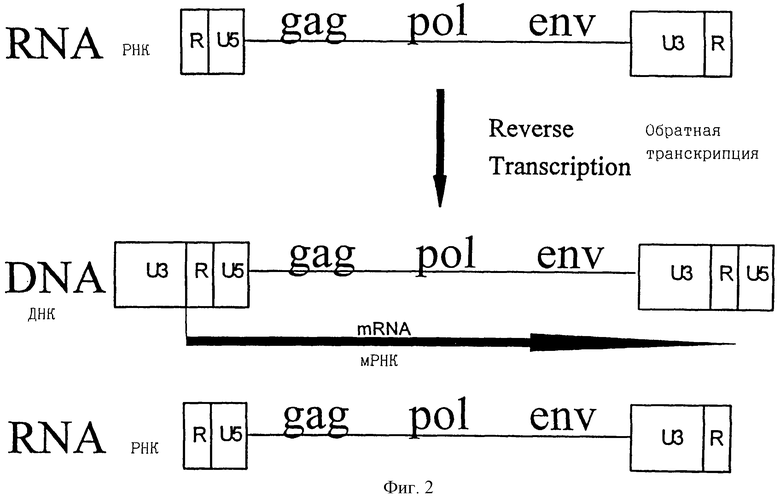
Геном ретровирусов состоит из молекулы РНК со структурой R-U5-gag-pol-env-U3-R (фиг.2). Во время процесса обратной транскрипции область U5 дуплицируется и помещается на правом конце генерируемой молекулы ДНК, тогда как U3 область дуплицируется и помещается на левом конце получаемой молекулы ДНК (фиг. 2). Получающаяся структура U3-R-U5 называется ДКП (длинный концевой повтор) и является, таким образом, идентичной и повторена на обоих концах структуры ДНК или провируса (Varmus, 1988). Область U3 на левом конце провируса несет промотор (см. ниже). Промотор управляет синтезом транскрипта РНК, начинающимся на границе между левой U3 и областями R и кончающейся на границе между правой R и U5 областью (фиг.2). Эта РНК упаковывается в ретровирусные частицы и переносится в целевые клетки, которые инфицируют. В целевой клетке геномная РНК снова подвергается обратной транскрипции, как описано выше.
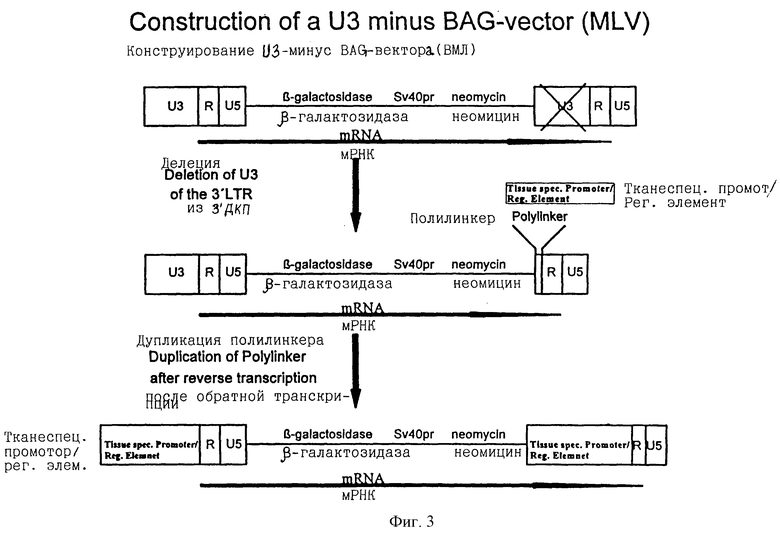
В соответствии с этим изобретением сконструирован ретровирусный вектор, в котором правая область U3 изменена (фиг.3), но остается нормальной структура левой U3 (фиг.3);
вектор может нормально транскрибироваться в РНК с использованием нормального ретровирусного промотора, расположенного в левой U3 области (фиг.3). Однако генерируемая РНК будет содержать только измененную правую структуру U3. В инфицированной целевой клетке после обратной транскрипции эта измененная U3 структура будет помещаться на обоих концах ретровирусной структуры (фиг.3).
Если измененная область несет полилинкер (см. ниже) вместо U3 области, то может быть легко вставлен любой промотор, включая те, которые управляют специфической экспрессией, такие как промотор СКВ (см. ниже). Этот промотор затем будет использоваться исключительно в целевой клетке для экспрессии связанных генов, которые несет ретровирусный вектор. Альтернативно или дополнительно в полилинкер могут быть вставлены сегменты ДНК, гомологичные одному или более клеточных последовательностей с целью нацеливания генов путем гомологичной рекомбинации (см. ниже).
В соответствии с этим изобретением термин "полилинкер" используется для короткой вытянутой или искусственно синтезированной ДНК, которая несет ряд уникальных сайтов рестрикции, дающих возможность легкой вставки любого промотора или сегмента ДНК. Термин "гетерологичный" используется для любой комбинации последовательностей ДНК, которые обычно в природе не обнаруживают тесно связанными.
Экспрессия генов регулируется промоторами. В отсутствие функции промотора ген не будет экспрессироваться. Нормальный ретровирусный промотор ВМЛ не является строго селективным в том, что он активен в большинстве типов клеток (Majors, 1990). Однако существует ряд промоторов, которые проявляют активность только в очень специфичных типах клеток. Такие тканеспецифичные промоторы будут идеальными кандидатами для регуляции экспрессии гена при ретровирусных векторах, ограничивая экспрессию терапевтических генов конкретными целевыми клетками.
В упаковывающей клеточной линии экспрессия ретровирусного вектора регулируется нормальным неселективным ретровирусным промотором, заключенным в области U3 (фиг. 3). Однако, как только вектор проникает в целевую клетку, происходит промоторная конверсия, и терапевтический или маркерный ген, например β-галактозидазный, экспрессируется от тканеспецифичного промотора выбора, введенного в полилинкер (фиг.3). В систему не только может быть практически введен тканевой специфический промотор, обеспечивающий селективную доставку широкого ряда различных типов клеток, но к тому же после превращения структура и свойства ретровирусного вектора больше не похожи на структуру и свойства вируса. Это, конечно, имеет чрезвычайно большое значение с точки зрения безопасности, так как обычные или современные ретровирусные векторы легко подвергаются генетической рекомбинации с ретровирусной упаковывающей конструкцией и/или эндогенными ретровирусами с получением потенциально патогенных вирусов. Векторы с промоторной конверсией (ПроКон) не похожи на ретровирусы, так как они после конверсии больше не несут ретровирусные промоторы U3, что, таким образом, снижает возможность генетической рекомбинации.
Ретровирусная промоторная структура находится в U3 области ДКП. ДКП несут сигнальные последовательности, которые позволяют им интегрировать в геном целевой клетки. Интеграция ретровирусных провирусов может также способствовать патогенетическим изменениям (van Lohuizen and Berns, 1990). В одном из осуществлений этого изобретения векторы ПроКон могут нести модифицированные ДКП, которые больше не содержат сигнальные последовательности, необходимые для интеграции. И это опять повышает потенциальную безопасность этих векторных систем.
Обеспечение направленной доставки гена
В соответствии с другим аспектом данного изобретения ретровирусный вектор используется для направленной интеграции в целевые клетки. Интеграция провирусного варианта ДНК ретровирусного генома в целевую клетку является главным преимуществом в отношении использования ретровирусов в качестве векторов по сравнению с другими вирусами, такими как аденовирусы, так как это дает возможность долгосрочной стабильной экспрессии перенесенных генов. Однако случайная природа этого факта интеграции также является главным недостатком использования ретровирусных векторов, так как это повышает возможность инсерционной (ин)активации клеточных генов опухолевых супрессоров или протоонкогенов и, таким образом, индукции опухолей (van Lohuizen and Berns, 1990).
Гомологичная рекомбинация была успешно применена для того, чтобы направить интеграцию трансфицированной или микро-инъекцированной ДНК в специфичные локусы ДНК и обычно используется при создании "истощенных" трансгенных мышей или животных (обзор сделан у Capecchi, 1989; Bradley et al., 1992; Morrow and Kucherlapati, 1993). К сожалению эффективность переноса ДНК такими чисто физическими методами чрезвычайно низка. В противоположность этому опосредованный ретровирусный перенос генов очень эффективен, есть возможность инфицирования почти 100% популяции клеток. Сочетание ретровирусного переноса гена с гомологичной рекомбинацией должно дать возможность конструирования идеальной системы для локус-направленной интеграции.
Мы исследовали возможность введения длинных гомологичных отрезков ДНК в ретровирусные векторы в различные положения для того, чтобы вызывать интеграцию путем гомологичной рекомбинации (Saller, 1994). Оценивали как конверсию гена, так и гомологичную рекомбинацию. Используя клеточную линию, несущую единственную копию HSV-tk гена в качестве мишени, нам удалось разорвать мишень с частотой в 15 раз выше, чем ранее сообщалось другими исследователями (Ellis and Bernstein, 1989). Клонирование рекомбинированных фрагментов ДНК демонстрировало наличие как целевой tk последовательности, так и ретровирусного вектора (Saller, 1994).
Для направленной интеграции сегменты ДНК, гомологичные клеточным последовательностям, встраиваются в полилинкер ProCon векторов. После инфицирования целевых клеток и обратной транскрипции эти последовательности появятся на 5' терминальном конце провируса. Терминальные гомологии, как было показано, благоприятствуют гомологичной рекомбинации (Bradley, 1991) в изогенных клеточных последовательностях (Bradley, 1991). Инфицирование целевых клеток, которые несут мутантные варианты гомологичной последовательности, должно приводить к рекомбинации и, таким образом, репарации мутантной последовательности. Или только гомологичные последовательности будут рекомбинировать в клеточный геном, или будет встраиваться полный вектор (Saller, 1994). Этот класс векторов не только имеет потенциал для использования в генной репарации, он может также применяться для регуляции интеграции ретровирусных векторов, несущих терапевтические гены в конкретные локусы в геноме, о которых известно, что они не несут активных генов. Это будет значительно снижать возможность инсерционной активации или инактивации, как описано выше, и будет, таким образом, способствовать безопасности использования ретровирусных векторов.
Следующие примеры дополнительно проиллюстрируют изобретение. Эти примеры, однако, никоим образом не предназначены для ограничения объема данного изобретения, так как будут ясны очевидные модификации, и еще и другие модификации и замены будут видны специалисту.
Методы генной инженерии, использованные при осуществлении данного изобретения, являются стандартными методиками, хорошо известными специалистам, и детально описаны, например в "Molecular Cloning" (Sambrook et al., 1989) и в "A Practical Guide to Molecular Cloning" (Perbal, 1984).
Краткое описание фигур, упоминаемых в последующих примерах:
фиг.1 - ретровирусная векторная система;
фиг.2 - ретровирусный геном, обратная транскрипция;
фиг.3 - принцип ProCon (промоторной конверсии);
фиг.4 - анализ ПЦР, проба на ВМЛ;
фиг.5 - анализ ПЦР, проба на МВОМЖ;
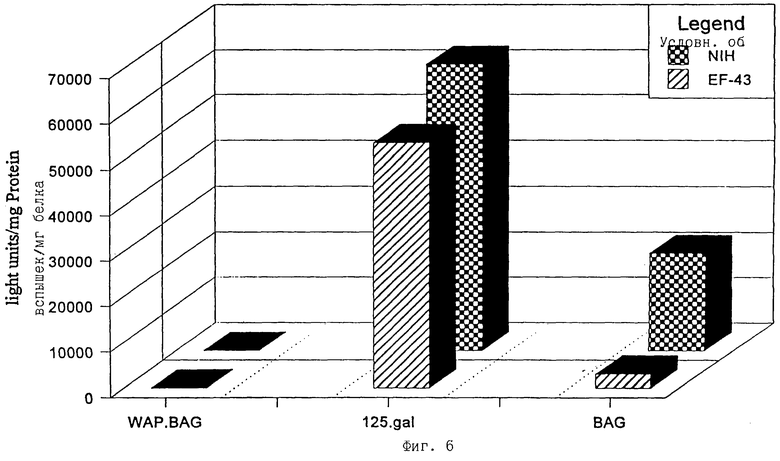
фиг.6 - экспрессия β-галактозидазы в инфицированных клетках NIH и EF43;
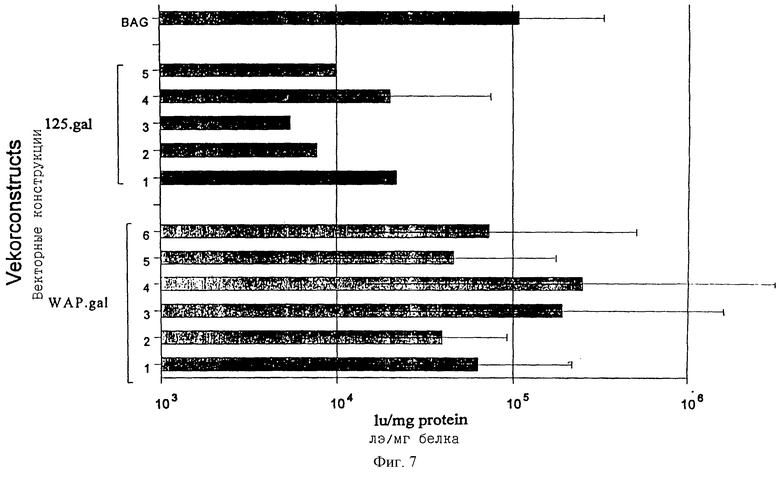
фиг. 7 - экспрессия β-галактозидазы в инфицированных основных клетках молочных желез от беременных мышей;
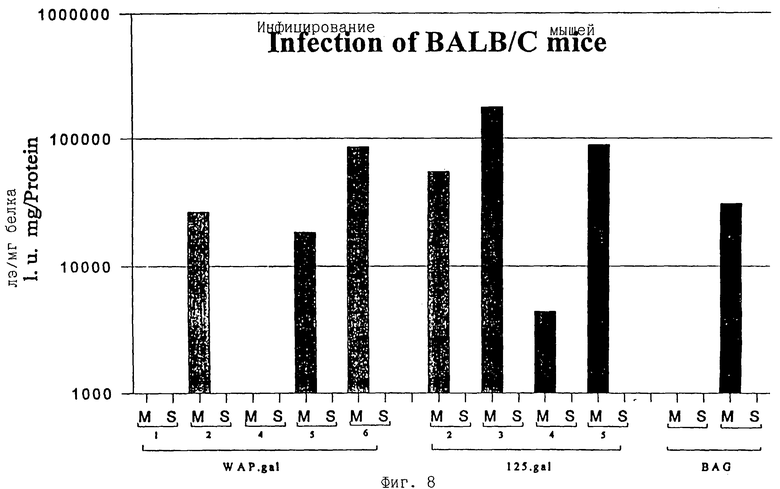
фиг.8 - экспрессия β-галактозидазы после инъекции вируса в молочную железу и кожу беременных мышей Balb/c;
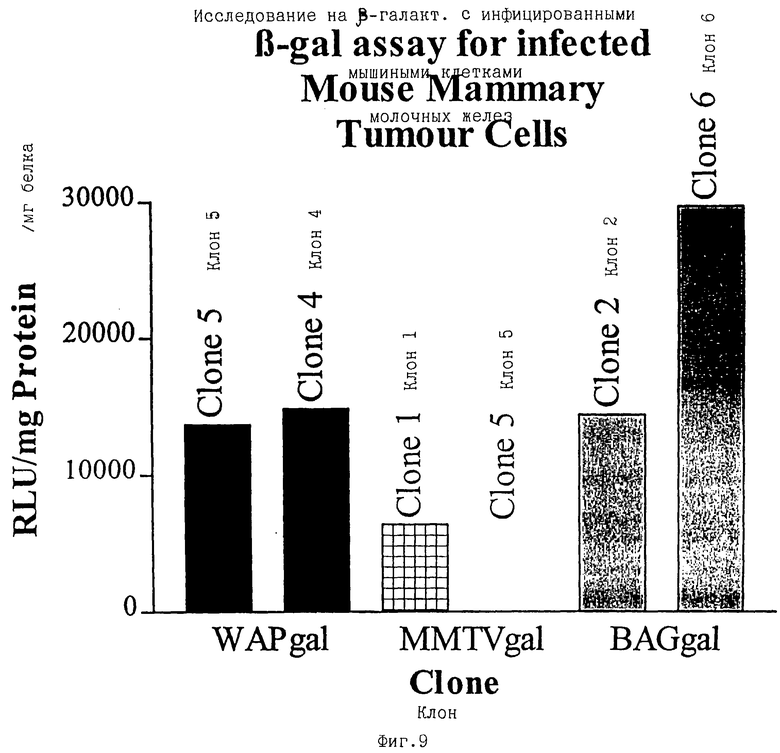
фиг.9 - экспрессия β-галактозидазы в инфицированных клетках опухоли молочных желез;
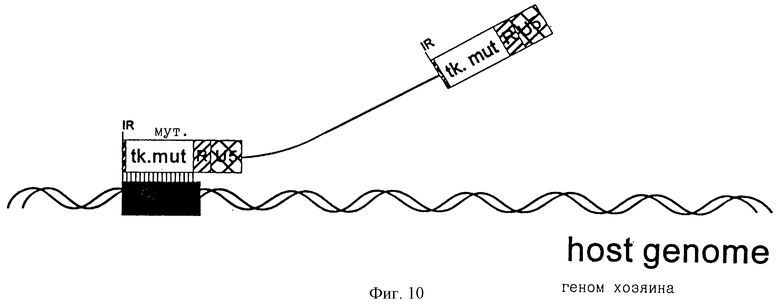
фиг.10 - направленная интеграция ретровирусного вектора путем гомологичной рекомбинации.
Пример 1
Специфическая экспрессия в молочных железах после инфицирования векторами ПроКон, несущими специфические для молочных желез промоторы
В ретровирусном векторе вируса мышиной лейкемии (ВМЛ), известном как BAG (Price et al., 1987) β-галактозидазный ген регулируется смешанным (т.е. не тканеспецифичным) промотором ВМЛ в U3 области ДКП (фиг.3). По данному изобретению было сконструировано производное BAG вектора, в котором промотор ВМЛ (U3), расположенный в 3'ДКП (фиг.3), удалялся и заменялся полилинкером, причем указанный полилинкер дает возможность легкого введения гетерологичных промоторов. Вектор BAG, лишенный U3, экспрессируется с промотора ВМЛ (U3) в 5'ДКП, если он введен в упаковывающую клеточную линию. В результате перегруппировок, происходящих в ретровирусном геноме во время его жизненного цикла, после инфицирования его целевых клеток, полилинкер будет дублироваться на обоих концах ретровирусного генома, как описано выше. Таким образом может быть сконструирован вектор, в котором экспрессия β-галактозидазного гена BAG будет контролироваться любым гетерологичным промотором, вставленным в полилинкер в целевой клетке (фиг.3).
По принципу, представленному выше, следующие специфичные промоторы были вставлены в область полилинкера или модифицированный вектор BAG:
некоторые субрайоны промотора мышиного вируса опухоли молочных желез (МВОМЖ), включая область, которая придает чувствительность к глюкокортикоидным гормонам, и область, содержащую элемент, который управляет экспрессией в молочных железах;
промотор сывороточного кислотного протеина (СКП). Этот промотор контролирует экспрессию СКП, так что он продуцируется только в молочных железах беременных и лактирующих грызунов.
Контроль экспрессии β-галактозидазного гена промоторами, вставленными в полилинкер, оценивали путем исследований по инфицированию с использованием сконструированных МВОМЖ и СКП ретровирусных векторов для инфицирования различных клеток.
Чтобы получить ретровирусные векторные частицы, ПроКон векторы МВОМЖ и СКП трансфицировали в упаковывающую клеточную линию GP+E86 (Markowitz et al. , 1988). После селекции по устойчивости к неомицину, которая кодировалась вектором, были получены стабильные популяции и клоны рекомбинантных продуцирующих ПроКон вирус клеток. Содержащий вирус супернатант от этих популяций использовали для инфицирования линии клеток молочной железы мыши EF43 (Gunzburg et al. , 1988), а также клеточной линии мышиных фибробластов (Jainchill et al. , 1969). Через четыре дня после инфицирования целевые клетки лизировали, и количественное определение β-галактозидазы не выявило экспрессии в каком-либо типе клеток, инфицированных ПроКон векторами, несущими СКП, и хорошую экспрессию в обоих типах клеток от ПроКон вектора, несущего МВОМЖ (фиг.6). Этот результат согласуется с тем, что СКП промотор функционирует только in vivo в поздние сроки беременности и во время лактации, но не в наиболее простых системах культуры клеток молочных желез in vitro, которые представлены клетками EF43. Чтобы исследовать, будут ли несущие СКП ПроКон векторы активны в сложных системах клеточной культуры, произведенной из основных клеток молочных желез, основные органоиды от мышей с 8-10-дневной беременностью (фиг. 7) или из опухолей молочных желез (фиг.9) брали в культуру и инфицировали супернатантом от таких же стабильно трансфицируемых клеточных линий. Оба вектора, несущие промоторные фрагменты как СКП, так и МВОМЖ, были активны в этих основных клетках (фиг.7) и клетках, происходящих из опухоли молочных желез (фиг.9), что показано с помощью β-галактозидазной активности.
Чтобы исследовать, были ли активны несущие СКП и МВОМЖ ПроКон векторы in vivo и была ли ограничена экспрессия β-галактозидазы in vivo молочной железой, содержащую рекомбинантный ПроКон вирус среду инъецировали in situ в молочные железы или кожу мышей с 8-10-дневной беременностью. Через пять дней мышей забивали, готовили экстракты клеток и выполняли исследование на β-галактозидазу. Несущие как фрагмент СКП, так и МВОМЖ ПроКон векторы экспрессировались только в молочных железах, но не в коже (cf М и S на фиг.8). Таким образом, in vivo регуляторные элементы от обоих промоторов ограничивают экспрессию молочными железами, тогда как in vitro регуляторные элементы от промотора СКП сохраняют свою строгую тканевую специфичность, а регуляторные элементы от промотора МВОМЖ - нет.
Эти ПроКон векторы, несущие специфические тканевые промоторы и регуляторные элементы, будут применимы для регулировки экспрессии терапевтических генов в предназначенных типах клеток, тканей и органах. Потенциальные терапевтические гены включают меллитиновый, который обладает анти-ВИЧ и противоопухолевым действием, и гены, которые программируют клетки на гибель, включая тимидинкиназный, гуанинфосфорибозилтрансферазный и цитозиндезаминазный гены, цитохрома Р450, а также гены, участвующие в регуляции клеточного цикла, такие как SDI/WAF-1/CIP-1.
Пример 2
Подтверждение промоторной конверсии в клетках, инфицированных ПроКон вектором, который исходно несет промотор МВОМЖ в 3'ДКП
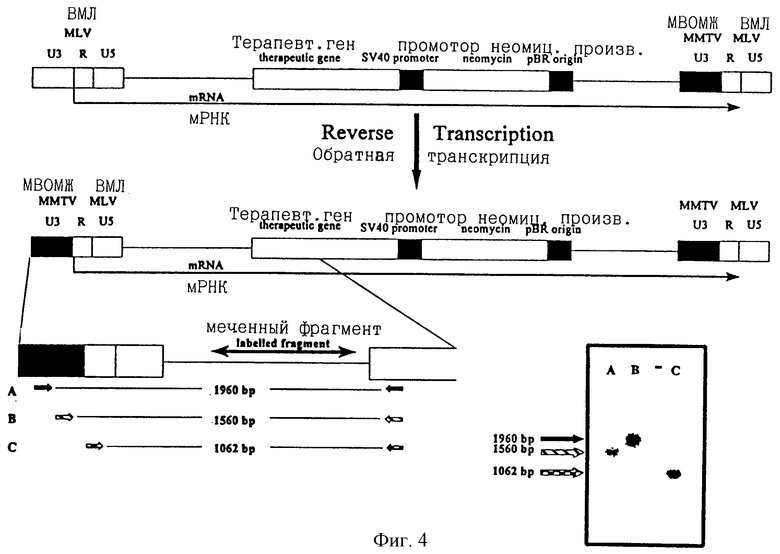
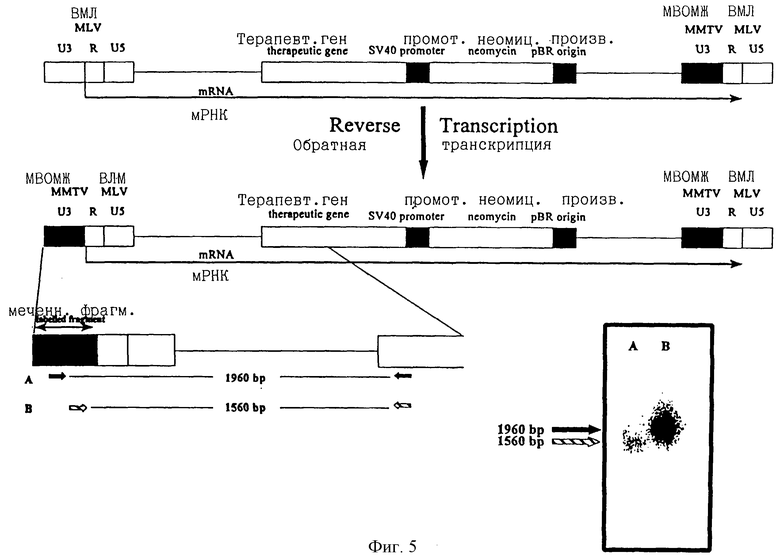
ПроКон вектор, несущий промоторную область из мышиного вируса опухоли молочных желез (МВОМЖ), трансфицировали в упаковывающую клеточную линию и полученные частицы рекомбинантного вектора использовали для инфицирования адаптированной линии клеток человеческой карциномы желчного пузыря (EJ). Селекцию инфицированных клеточных клонов производили в среде, содержащей аналог неомицина G418 (так как вектор несет ген устойчивости к неомицину, регулируемый внутренним промотором SV40). ДНК получали из одного из инфицированных клонов и нетрансфицированных и материнских EJ клеток и использовали для полимеразных цепных реакций (ПЦР). ПЦР выполняли с использованием двух праймеров, которые специфично распознаются и связываются с последовательностями МВОМЖ (А, В на фиг.4 и 5) или R областью ВМЛ (С на фиг.4) ДКП вместе с праймером, расположенным в маркерном гене (фиг.4 и 5). Так как праймер маркерного гена располагается только в прямом направлении последовательности МВОМЖ (или R области ВМЛ), если произошла промоторная конверсия, положительный сигнал ПЦР, полученный с праймерами МВОМЖ в сочетании с праймером маркерного гена, является показателем этого. На фиг.4 показаны продукты ПЦР с использованием праймеров А, В и С после гибридизации с меченым фрагментом из последовательности ВМЛ, подтверждающей, что все три продукта ПЦР имеют происхождение из ВМЛ. На фиг.5 показаны продукты ПЦР с использованием праймеров А и В и гибридизованные со специфической пробой МВОМЖ, что опять подтверждает то, что произошла промоторная конверсия.
Пример 3
Конструкция ПроКон векторов для направленной интеграции
Используя тот же самый вектор BAG, описанный в примере 1 выше, можно сконструировать ретровирусный вектор, в котором может быть вставлена последовательность ДНК с гомологией к клеточной последовательности в ДКП. Полученный вектор может использоваться для направленной интеграции или гомологичной последовательности, вставленной в вектор, или всего вектора или его части в гомологичную последовательность, присутствующую в геноме клетки-хозяина.
По принципу, представленному выше, фрагмент тимидинкиназного (tk) гена вируса Herpes simplex (HSV) был вставлен в полилинкерную область модифицированного BAG вектора (tk мутант на фиг.10, Saller, 1994).
Также была закреплена клеточная линия, которая не имеет функциональных копий tk гена млекопитающих, а вместо этого она несет одну копию HSV-tk гена (Saller, 1994). Эта клеточная линия была инфицирована несущим tk BAG вектором, и были отобраны клетки, у которых произошел разрыв HSV-tk гена (фиг. 10).
В вышеприведенных примерах проиллюстрированы принципы конструкции и значение векторов с промоторной конверсией, представляемых данным изобретением.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТРАНСДУЦИРУЮЩИЕ ЦИТОХРОМ Р450 РЕТРОВИРУСНЫЕ ВЕКТОРЫ | 1997 |
|
RU2185821C2 |
| РЕКОМБИНАНТНЫЙ МОДИФИЦИРОВАННЫЙ ВИРУС ОСПОВАКЦИНЫ, ЕГО ИСПОЛЬЗОВАНИЕ (ВАРИАНТЫ) И ИНФИЦИРОВАННАЯ ИМ ЛИНИЯ КЛЕТОК (ВАРИАНТЫ) | 1996 |
|
RU2198217C2 |
| КАПСУЛА ДЛЯ КЛЕТОК, ПРОДУЦИРУЮЩИХ ВИРУСНЫЕ ЧАСТИЦЫ, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ИСПОЛЬЗОВАНИЯ | 1996 |
|
RU2187301C2 |
| РЕКОМБИНАНТНЫЙ МОДИФИЦИРОВАННЫЙ ВИРУС ВАКЦИНЫ АНКАРА, СПОСОБНЫЙ ЭКСПРЕССИРОВАТЬ СТРУКТУРНЫЕ АНТИГЕНЫ ВИРУСА ГЕПАТИТА С | 2002 |
|
RU2270860C2 |
| ИНДУЦИРОВАННАЯ ОЗОНОМ ЭКСПРЕССИЯ ГЕНА В РАСТЕНИЯХ | 1997 |
|
RU2224021C2 |
| ТРАНСДУЦИРУЮЩИЕ ЦИТОХРОМ P450 РЕТРОВИРУСНЫЕ ВЕКТОРЫ | 1997 |
|
RU2223788C2 |
| СРЕДСТВА И СПОСОБЫ ЛЕЧЕНИЯ ВИРУСА ГЕПАТИТА B | 2017 |
|
RU2740802C2 |
| ПРИМЕНЕНИЕ АДАПТИРОВАННЫХ РЕКОМБИНАЗ ДЛЯ ЛЕЧЕНИЯ РЕТРОВИРУСНЫХ ИНФЕКЦИЙ | 2008 |
|
RU2501860C2 |
| УСОВЕРШЕНСТВОВАНИЕ ГЕНЕТИЧЕСКИХ КОНСТРУКЦИЙ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АНТИВИЧ ТЕРАПИИ | 2013 |
|
RU2533817C1 |
| РЕТРОВИРУСНЫЙ ВЕКТОР НА ОСНОВЕ ВИРУСА МЫШИНОГО ЛЕЙКОЗА (MLV) (ВАРИАНТЫ) | 1999 |
|
RU2203321C2 |
Данное изобретение относится к ретровирусному вектору, подвергающемуся промоторной конверсии, включающему 5'ДКП область из структуры U3-R-U5, одну или более последовательностей, выбираемых из кодирующих и некодирующих последовательностей, и 3'ДКП область, содержащую полностью или частично делетированную U3 область. Указанная делетированная U3 область заменяется полилинкерной последовательностью, за которой следует R и U5 область. Ретровирусный вектор подвергается промоторной конверсии и применим в качестве носителя для передачи гена с целью направленной генной терапии. 4 с. и 23 з.п. ф-лы, 10 ил.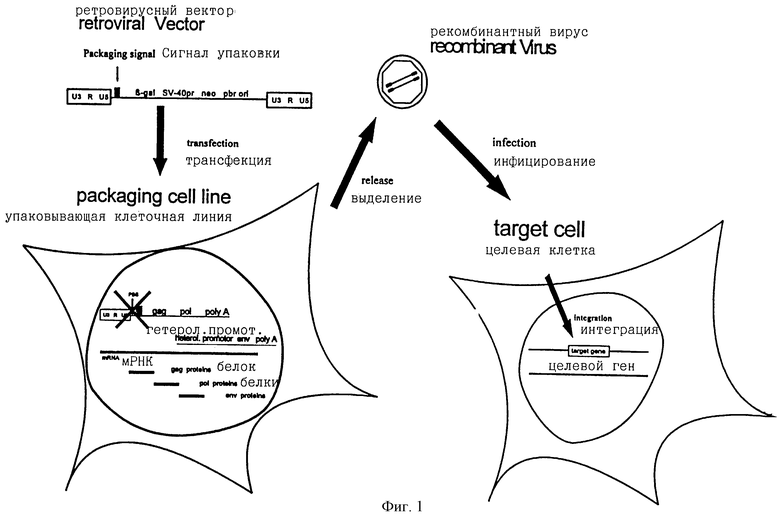
| БУФЕРНОЕ ЗАПОМИНАЮЩЕЕ УСТРОЙСТВО | 1972 |
|
SU415731A1 |

