Область техники, к которой относится изобретение
Настоящее изобретение относится к способу индукции мейоза в половых клетках и к использованию некоторых химических соединений и лекарственных средств, включающих такие соединения, для стимуляции мейоза ин виво, экс виво и ин витро.
Предпосылки изобретения
Мейоз является уникальным и основным событием половых клеток, на котором основано половое размножение. Мейоз включает в себя два мейотических деления. В течение первого деления, перед расхождением пар хромосом в две дочерние клетки, имеет место обмен между материнскими и отцовскими генами. Они содержат только половину набора (1n) хромосом и 2с ДНК. Второе мейотическое деление протекает без синтеза ДНК. По этой причине это деление приводит к образованию гаплоидных половых клеток только с 1с ДНК.
Мейотические события сходны в мужских и женских половых клетках, но время осуществления и процессы дифференциации, которые приводят к образованию яйцеклетки или сперматозоида основательно различаются. Все женские половые клетки входят в профазу первого мейотического деления на ранних этапах развития, часто до рождения, но все подвергаются задержке в виде поздних ооцитов в профазе (стадия диктиогенеза) вплоть до овуляции по достижении половой зрелости. Поэтому у женщины с раннего развития имеется линия ооцитов, которая исчерпывается только с истощением линии. Мейоз у женщин заканчивается только после оплодотворения и имеет своим результатом только одну яйцеклетку и два абортивных полярных тельца на одну половую клетку. В противоположность этому, только некоторые мужские половые клетки проходят мейоз при достижении половой зрелости и покидают стволовую популяцию зародышевых клеток в течение жизни. Однажды начавшись, мейоз в мужских клетках протекает без существенной задержки и образует 4 сперматозоида.
О механизме, который контролирует инициацию мейоза у мужчин и женщин, известно совсем немного. Новые исследования свидетельствуют, что в ооцитах фолликулярные пурины, гипоксантин или аденозин могут нести ответственность за задержку мейоза (Downs, S.M. et al. Dev. Biol. 82 (1985) 454-458; Eppig, J. J. et al. Dev. Biol. 119 (1986) 313-321; и Downs, S.M. Mol. Reprod. Dev. 35 (1993) 82-94). Наличие диффундирующего вещества, регулирующего мейоз, впервые описано Byskov et al. для культуральной системы эмбриональных мышиных гонад (Byskov, A.G. et al. Dev. Biol. 52 (1976) 193-200). Вещество, активирующее мейоз (ВАМ), выделяется эмбриональным мышиным яичником, в котором мейоз происходит постоянно, а вещество, препятствующее мейозу (ВПМ), обнаружили в морфологически дифференцированных семенниках, содержащих покоящиеся, немейотические половые клетки. Было предположено, что относительные концентрации ВАМ и ВПМ регулируют начало, остановку и возобновление мейоза в мужских и в женских половых клетках (Byskov, A.G. et al. in The Physiology of Reproduction (eds. Knobil, E. and Neill, J. D. , Raven Press, New York (1994)). Очевидно, что если мейоз можно регулировать, можно контролировать размножение. Поэтому, если необходима стимуляция мейоза ооцита, один из возможных путей гарантированного достижения этого состоит в том, чтобы количество ВАМ, присутствующего в окружении ооцита, превышало количество имеющегося ВПМ. В принципе, это можно было бы достигнуть введением ВАМ, путем стимуляции секреции ВАМ или путем блокирования биотрансформации уже имеющегося ВАМ.
Сущность изобретения
Ранее установили, что введение некоторых стеринов, известных в качестве промежуточных соединений в биосинтезе холестерина, приводит к стимуляции мейоза. Удивительно, но оказалось, что введение некоторых соединений, о которых известно, что они подавляют биосинтез холестерина, может также стимулировать мейоз. Хотя основной механизм (или механизмы) понятен не вполне, следует ожидать, что, по крайней мере, в некоторых случаях, подавление включает в себя ингибирование одного или нескольких ферментов, вовлеченных в биоконверсию одного или нескольких предшественников холестерина. В результате ингибирования, концентрация предшественников, образованных на стадиях, предшествующих стадии, которая была подавлена, будет увеличиваться. Один или несколько из этих предшественников может - прямо или косвенно - обусловить стимуляцию мейоза.
В соответствии с этим, в самом широком аспекте, настоящее изобретение относится к способу стимуляции мейоза половых клеток, который включает в себя введение указанным клеткам ин виво, экс виво или ин витро эффективного количества соединения, которое вызывает накопление эндогенного вещества, активирующего мейоз, до уровня, при котором индуцируется мейоз.
В соответствии с предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половых клеток млекопитающих.
В соответствии с другим предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половых клеток человека.
В соответствии со следующим предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза ооцита.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза мужских половых клеток.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу контрацепции для применения у женщин.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу лечения бесплодия путем стимуляции образования мейотических ооцитов, так чтобы иметь увеличенное количество мейотических ооцитов при достижении гонадотропинами пика в процессе овуляции.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу лечения бесплодия у мужчин путем стимуляции образования сперматозоидов из мужских зародышевых клеток.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половой клетки, который включает в себя введение соединения, которое проявляет свойства, активирующие мейоз при испытании хотя бы одного из способов, описанных в примерах настоящего описания.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половой клетки, который включает в себя введение амфотерицина В.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половой клетки, который включает в себя введение аминогуанидина.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половой клетки, который включает в себя введение 3β,5α,6β-тригидроксихолестана.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половой клетки, который включает в себя введение мелатонина.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половых клеток, который включает в себя введение производного мелатонина.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половых клеток, который включает в себя введение 6-хлормелатонина.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половых клеток, который включает в себя введение 5-метокситриптамина.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половых клеток, который включает в себя введение агониста мелатонина.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к способу стимулирования мейоза половых клеток, который включает в себя введение любого соединения, выбранного из нижеследующего списка:
4α-цианометил-5α-холестан-3β-ол;
4α-нитро-5α-холестан-3β-ол;
4α-амино-5α-холестан-3β-ол;
4α-формил-5α-холестан-3β-ол;
4α-аминометил-5α-холестан-3β-ол;
4α-(2-циано)этинил-5β-холестан-3α-ол;
4-алленил-5α-холестан-3β-ол;
4α-оксиран-5α-холестан-3β-ол;
4α-(3,3-дихлор)винил-5α-холестан-3β-ол;
4α-(3,3-дибром)винил-5α-холестан-3β-ол;
4α-(дифторметил)-5α-холестан-3β-ол;
4-циклододецил-2,6-диметилморфолин;
4-тридецил-2,6-диметилморфолин;
4-додецил-2,6-диметилморфолин;
4-[3-[4-(1,1-диметилэтил)фенил]-2-метилпропил]-2,6-диметилморфолин;
4-[3-[4-(1,1-диметилэтил)фенил] -2-метилпропил] -2,6-диметилморфолин-N-оксид;
цис-4-[3-[4-(1,1-диметилпропил)фенил] -2-метилпропил] -2,6-диметилморфолин;
1-[3-[4-(1,1-диметилэтил)фенил]-2-метилпропил]пиперидин;
8-аза-4α, 10-диметил-транс-декал-3β-ол;
N-бензил-8-аза-4α, 10-диметил-транс-декал-3β-ол;
N-бензил-8-аза-4α, 10-диметил-транс-декал-3β-ол-N-оксид;
N-(1,5,9-триметилдецил)-8-аза-4α, 10-диметил-транс-декал-3β-ол;
N-(1-метилдодецил)-8-аза-4α, 10-диметил-транс-декал-3β-ол;
N-[6-(4-трет-бутилфенил)-1,5-диметил] гексил-8-аза-4α, 10-диметил-транс-декал-3β-ол;
15-аза-24-метилен-D-гомохолеста-8,14-диен-3β-ол;
4-хлор-α-[4-[2-(диэтиламино)этокси]фенил]-α-(4-метил-фенил)бензолэтанол;
1,4-бис(2-хлорбензиламинометил)циклогексан;
6-амино-2-пентилтиобензотиазол;
24,25-иминоланостерин;
аминотриазол;
нистатин;
25-азахолестерин;
25-аза-24,25-дигидрозимостерин;
25-азахолестанол;
(20R)-22,25-диазахолестерин;
(20S)-22,25-диазахолестерин;
24-азахолестерин;
25-аза-24,25-дигидроланостерин; и
23-азахолестерин.
В соответствии с очередным предпочтительным вариантом осуществления, настоящее изобретение относится к применению соединения, которое вызывает накопление эндогенного вещества, активирующего мейоз, до уровня, при котором индуцируется мейоз, для получения лекарственного средства, индуцирующего мейоз.
В соответствии с очередным вариантом осуществления, настоящее изобретение относится к применению соединения, которое проявляет свойства, активирующие мейоз, при испытании хотя бы одного из способов, описанных в примерах настоящего описания, для получения лекарственного средства, индуцирующего мейоз.
В соответствии с еще одним предпочтительным вариантом осуществления, настоящее изобретение относится к использованию амфотерицина В для получения лекарственного средства, индуцирующего мейоз.
В соответствии с еще одним предпочтительным вариантом осуществления, настоящее изобретение относится к использованию аминогуанидина для получения лекарственного средства, индуцирующего мейоз.
В соответствии с еще одним предпочтительным вариантом осуществления, настоящее изобретение относится к использованию 3β,5α,6β-тригидрохолестана для получения лекарственного средства, индуцирующего мейоз.
В соответствии с еще одним предпочтительным вариантом осуществления, настоящее изобретение относится к использованию мелатонина для получения лекарственного средства, индуцирующего мейоз.
В соответствии с еще одним предпочтительным вариантом осуществления, настоящее изобретение относится к использованию производного мелатонина для получения лекарственного средства, индуцирующего мейоз.
В соответствии с еще одним предпочтительным вариантом осуществления, настоящее изобретение относится к использованию 6-хлормелатонина для получения лекарственного средства, индуцирующего мейоз.
В соответствии с еще одним вариантом осуществления, настоящее изобретение относится к использованию 5-метокситриптамина для получения лекарственного средства, индуцирующего мейоз.
В соответствии с еще одним вариантом осуществления, настоящее изобретение относится к использованию агониста мелатонина для получения лекарственного средства, индуцирующего мейоз.
В соответствии с еще одним предпочтительным вариантом осуществления, настоящее изобретение относится к использованию соединения, выбранного из нижеследующего списка, для получения лекарственного средства, индуцирующего мейоз:
4α-цианометил-5α-холестан-3β-ол;
4α-нитро-5α-холестан-3β-ол;
4α-амино-5α-холестан-3β-ол;
4α-формил-5α-холестан-3β-ол;
4α-аминометил-5α-холестан-3β-ол;
4α-(2-циано)этинил-5α-холестан-3β-ол;
4-алленил-5α-холестан-3β-ол;
4α-оксиран-5α-холестан-3β-ол;
4α-(3,3-дихлор)винил-5α-холестан-3β-ол;
4α-(3,3-дибром)винил-5α-холестан-3β-ол;
4α-(дифторметил)-5α-холестан-3β-ол;
4-циклододецил-2,6-диметилморфолин,
4-тридецил-2,6-диметилморфолин;
4-додецил-2,6-диметилморфолин;
4-[3-[4-(1,1-диметилэтил)фенил]-2-метилпропил]-2,6-диметилморфолин;
4-[3-[4-(1,1-диметилэтил)фенил] -2-метилпропил] -2,6-диметилморфолин-N-оксид;
цис-4-[3-[4-(1,1-диметилпропил)фенил] -2-метилпропил] -2,6-диметилморфолин;
1-[3-[4-(1,1-диметилэтил)фенил]-2-метилпропил]пиперидин;
8-аза-4α, 10-диметил-транс-декал-3β-ол;
N-бензил-8-аза-4α, 10-диметил-транс-декал-3β-ол;
N-бензил-8-аза-4α, 10-диметил-транс-декал-3β-ол-N-оксид;
N-(1,5,9-триметилдецил)-8-аза-4α, 10-диметил-транс-декал-3β-ол;
N-(1-метилдодецил)-8-аза-4α, 10-диметил-транс-декал-3β-ол;
N-[6-(4-трет-бутилфенил)-1,5-диметил] гексил-8-аза-4α, 10-диметил-транс-декал-3β-ол;
15-аза-24-метилен-D-гомохолеста-8,14-диен-3β-ол;
4-хлор-α-[4-[2-диэтиламино)этокси]фенил]-α-(4-метилфенил)бензолэтанол;
1,4-бис(2-хлорбензиламинометил)циклогексан;
6-амино-2-пентилтиобензотиазол;
24,25-иминоланостерин;
аминотриазол; нистатин;
25-азахолестерин;
25-аза-24,25-дигидрозимостерин;
25-азахолестанол;
(20R)-22,25-диазахолестерин;
(20S)-22,25-диазахолестерин;
24-азахолестерин;
25-аза-24,25-дигидроланостерин и
23-азахолестерин.
Подробное описание изобретения
С некоторых пор стало известно о существовании вещества, активирующего или стимулирующего мейоз. Однако до недавнего времени оставалась неизвестной идентичность вещества или веществ, активирующих мейоз.
В перспективе существует несколько возможностей влиять на мейоз. В соответствии с предпочтительным вариантом осуществления настоящего изобретения выбранные соединения применимы для стимуляции мейоза. В соответствии с другим предпочтительным вариантом осуществления настоящего изобретения выбранные соединения применимы для стимуляции мейоза у человека. Таким образом, выбранные соединения являются многообещающими для использования в качестве вещества, регулирующего фертильность. Можно надеяться, что обычное побочное действие на соматические клетки, до сих пор известное при использовании гормональных контрацептивов, которые основаны на эстрогенах и/или гестагенах, не будет обнаружено у настоящего изобретения. Для использования в качестве контрацептивного средства у женщин мейоз можно индуцировать так, чтобы как можно быстрее вызвать возобновление мейоза в ооцитах, в то время пока они все еще находятся в растущем фолликуле, до достижения гонадотропинами овуляционного пика. У женщин возобновление мейоза можно, например, индуцировать спустя неделю, предшествующую окончанию менструации. Перезрелые ооциты, полученные после овуляции, весьма вероятно, не будут оплодотворены. Маловероятно, чтобы было затронуто течение нормального менструального цикла. В этой связи важно отметить, что синтез прогестерона в культуре человеческих клеток гранулезы (соматические клетки фолликула) не подвержен влиянию присутствующего вещества, индуцирующего мейоз, тогда как применение эстрогенов и гестагенов, используемых до сих пор в качестве гормональных контрацептивов, отрицательно влияет на синтез прогестерона.
Продемонстрирована также стимуляция мейоза в мужских половых клетках. В соответствии с этим настоящее изобретение может быть также полезным при лечении мужского бесплодия.
Ланоста-8,24-диен-3β-ол (ланостерин), который лишен каких-либо свойств, активирующих мейоз, является первичным продуктом циклизации при синтезе стерина в клетках млекопитающих. Последовательный биосинтез холестерина проходит через серии стадий, как то, деметилирований, окислений, восстановлений и замещений двойных связей, которые все контролируются ферментативно. Выделены и охарактеризованы только некоторые из ферментов, контролирующих эти стадии. Первым продуктом, образованным с холестеновым скелетом, является 4,4-диметилхолеста-8,14,24-триен-3β-ол, который идентичен с активирующим мейоз соединением, выделенным из жидкости фолликул человека (A.G.Byskov et al. Nature 374 (1995) 559-562). Последующее восстановление двойной связи в положении 14 образует 4,4-диметилхолеста-8,24-диен-3β-ол, который идентичен с активирующим мейоз соединением, выделенным из семенников быка (A.G.Byskov et al. Nature 374 (1995) 559-562). Постадийное удаление метильных групп в положении 4 образует 4-метилхолеста-8,24-диен-3β-ол и холеста-8,24-диен-3β-ол (зимостерин), которые, оба, обладают активирующими мейоз свойствами. Последующее перемещение двойной связи из положения 8 в положение 5 образует холест-5-ен-3β-ол (холестерин), который не обладает свойствами, активирующими мейоз. Ингибирование любого из упомянутых ферментов, которые активны в сериях реакций, описанных выше, будет вызывать накопление только что приведенных промежуточных соединений с активирующими мейоз свойствами, индуцируя таким образом мейоз в присутствующих половых клетках.
Ряд соединений, известных из литературы, был описан в качестве ингибиторов одного или нескольких ферментов, вовлеченных в преобразование, ин виво, ланоста-8,24-диен-3β-ола в холестерин, что привело к пересмотру в данной области (Mercer, E.I. Prog. Lipid Res. 32 (1993) 357-416). Известно, что амфотерицин подавляет последние стадии синтеза эргостерина в грибах (Coulon, J. et al. Can. J. Microbiol. 32 (1986) 738-742) и применяется в клинике в качестве противогрибкового средства. Обнаружено, что в крысиной печени, ин витро, холестентриол подавляет деметилирование, в положении 4, промежуточных соединений в биосинтезе холестерина и, таким образом, вызывает накопление 4,4-диметилхолеста-8-ен-3-ола и 4-метилхолеста-8-ен-3-ола (Scallen, T.J. et al. J. Biol.Chem. 246 (1971) 3168-3174).
Количество активного вводимого агента этого изобретения определяется в соответствии с целью лечения сведущими в этой области специалистами. Количество зависит от отсутствия специфического агента, о котором идет речь, от конкретного способа введения (напр., ин виво, экс виво или ин витро) и от других факторов.
Композиции, в соответствии с настоящим изобретением, для введения активных агентов, могут быть в форме таблеток, капсул, порошков, растворов или суспензий. В таких композициях активные агенты могут быть объединены с носителями, вспомогательными средствами и носителями, обычно используемыми в этой области.
Системное действие на живое животное может быть достигнуто путем перорального введения или путем инъекции, или инфузии стерильных растворов активных агентов, в соответствии с настоящим изобретением, растворов, которые получали в соответствии с известными в этой области способами. Системный эффект может быть достигнут также путем ингаляции или путем введения через нос порошка или аэрозоля, содержащего активный агент.
Введение активного агента, в соответствии с настоящим изобретением, в изолированную клетку, напр., в ооцит или в мужскую половую клетку, может быть достигнуто путем выдерживания клетки в среде, используемой обычно для сохранения клеток соответствующего вида, с последующим добавлением в среду активного агента.
Настоящее изобретение более подробно иллюстрируется нижеследующими примерами, которые, однако, не должны рассматриваться в качестве ограничения объема защиты. Признаки, раскрытые в предшествующем описании и в нижеследующих примерах, могут служить материалом, порознь или в сочетании, для осуществления изобретения в различных его формах.
Примеры
МАТЕРИАЛЫ И МЕТОДЫ
Испытание веществ, активирующих мейоз, в ооцитном тесте.
Животные
Неполовозрелых мышиных самок (B6D2-F1, линия C57B1/6J) выдерживали при контролируемом освещении (14 ч света, 10 ч темноты) и температуре, а пищу и воду предоставляли по желанию. Когда животные достигали веса 13-16 граммов (что соответствует 20-22-дневному возрасту после родов), им делали однократную инъекцию (внутрибрюшинно) менопаузного гонадотропина человека (Humegon, Organon, Нидерланды), содержащего приблизительно 20 ИЕ ФСГ и 20 ИЕ ЛГ (Ziebe, S. et al. Hum. Reprod. 8 (1993) 385-388). 48 часами позже животных забивали путем смещения шейных позвонков.
Сбор и культивирование ооцитов
Удаляли яичники, помещали их в ГК-среду (см. ниже) и освобождали от инородной ткани. Среда для сбора и культивирования состояла в основе из минимальной среды Игла (Flow, США), содержащей 4 мМ гипоксантина (ГК), 3 мг/мл бычьего сывороточного альбумина, 0,23 мМ пирувата натрия, 2 мМ глутамина, 100 Е/мл пенициллина и 100 мкг/мл стрептомицина (все Sigma, США). Эту среду обозначили ГК-среда. Такую же среду, но без ГК, использовали в качестве контрольной среды.
Влияние испытываемых соединений на мейоз ооцитов изучали на скоплении включенных ооцитов (СВО, Опыт А) и на денудированных ооцитах (ДО, Опыт В). СВО получали путем пункции антральных фолликулов из яичников под рассекающим микроскопом, используя иглу 27. Отбирали скопления включенных ооцитов (СВО) одинакового размера и, перед использованием в Опыте А, трижды промывали свежей ГК-средой. Ооциты освобождали от скоплений клеток, т.е. денудировали ооциты, ДО для использования в Опыте В получали путем осторожной промывки СВО через пипетку с тонким контролируемым отверстием. СВО в Опыте А и ДО в Опыте В культивировали в 4-луночных чашках Петри (Nunclon, Дания) в 0,5 мл ГК-среды, содержащей испытуемое соединение в концентрации, представленной в таблице, за исключением контролей, где культивирование осуществляли в контрольной среде. Каждая лунка содержала 35-50 ооцитов. Испытуемые культуры готовили при различных концентрациях испытываемых соединений, как указано в таблицах.
Культуры содержали при 37oС и 100%-ной влажности с 5% СО2 в атмосфере в течение 24 часов.
Инициация ооцитов, Опыт С(А) и С(В)
Опыт С(А) и С(В) осуществляли, соответственно, так же как и Опыт А и Опыт В, за тем только исключением, что ооциты помещали в среду, содержащую испытуемое соединение, в течение периода времени (период инициации) от 5 мин до 3-х ч с начала опыта. После периода инициации, ооциты переносили в контрольную среду и продолжали культивирование до 22 часов после начала опыта.
Исследование ооцитов
К концу 24-часового периода культивирования подсчитывали, под микроскопом с инверсией, снабженным устройством для контрастной дифференциальной интерференции, число ооцитов с зародышевым пузырьком (ЗП) или с разрушенным зародышевым пузырьком (ЗПР) и зародышевым пузырьком с полярными тельцами (ПТ). Вычисляли процентную долю ооцитов с ЗПР в общем числе ооцитов и процентную долю ооцитов с ПТ среди ЗПР. Результаты, полученные в Опытах А, В, С(А) и С(В), подсчитанные в качестве единиц активности ВАМ, приведены в таблицах по каждому примеру. Одну единицу активности ВАМ, ЕВАМ, определяли как: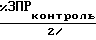
Количество единиц активности ВАМ, ЕВАМ, вычисляли как: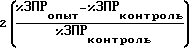
ПРИМЕР 1
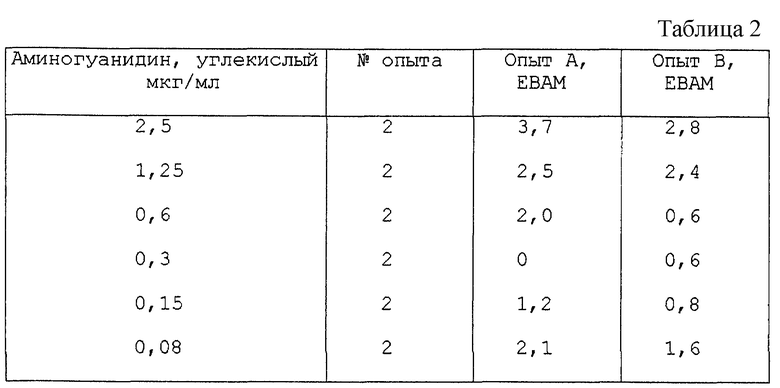
Активация мейоза в ооцитах, с использованием холестан-3β,5α,6β-триола.
Опыт А и Опыт В осуществляли, как описано выше, используя холестан-3β, 5α, 6β-триол в качестве испытуемого соединения. Результаты представлены в табл.1.
Используемый холестан-3β,5α,6β-триол получали от Sigma (St. Louis, США). Из таблицы видно, что холестан-3β,5α,6β-триол индуцирует возобновление мейоза в ооцитах дозо-зависимым образом.
ПРИМЕР 2
Активация мейоза в ооцитах, с использованием углекислого аминогуанидина.
Опыт А и Опыт В осуществляли, как описано выше, используя, в качестве испытуемого соединения, углекислый аминогуанидин. Результаты представлены в табл.2.
Используемый углекислый аминогуанидин получали от Aldrich Chemical Co., Inc (Milwaukee, Висконсин). Из таблицы видно, что углекислый аминогуанидин возобновляет мейоз в ооцитах дозо-зависимым образом.
ПРИМЕР 3
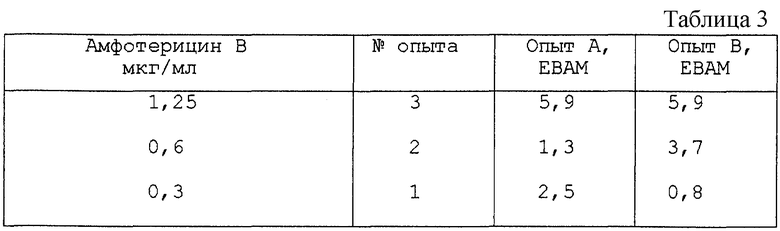
Активация мейоза в ооцитах, с использованием амфотерицина В.
Опыт А и Опыт В осуществляли, как описано выше, используя, в качестве испытуемого соединения, амфотерицин В. Результаты представлены в табл.3.
Амфотерицин В получали от Bristol-Myers Squibb. Из таблицы видно, что амфотерицин В индуцирует возобновление мейоза в ооцитах дозо-зависимым образом.
Амфотерицин токсичен при концентрациях выше 1,25 мкг/мл. Испытывали концентрации до 50 мкг/мл.
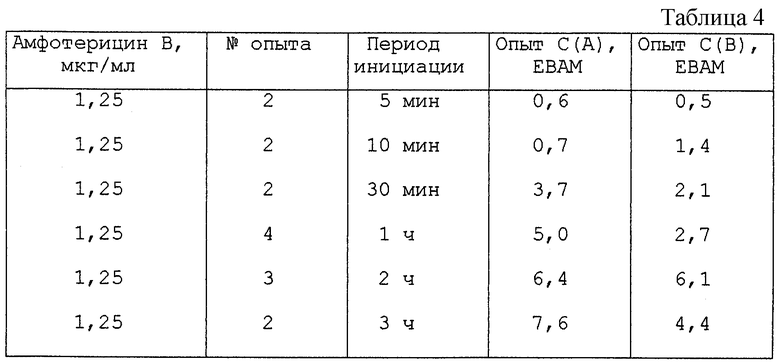
Инициация ооцитов амфотерицином В
Результаты представлены в табл.4.
Из табл.4 видно, что даже выдерживание с амфотерицином В в течение 5 минут достаточно для запуска возобновления мейоза в ДО и СВО.
ПРИМЕР 4
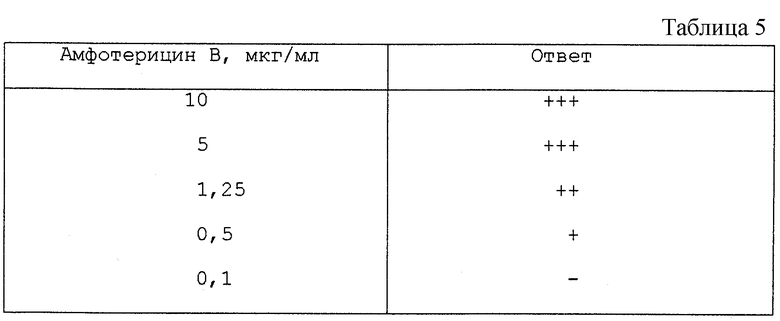
Активация мейоза в мужских половых клетках, с использованием амфотерицина В.
Опытная система состояла из эмбриональных мышиных гонад, 11,5 дней после зачатия. Одну гонаду из каждого эмбриона использовали в качестве контроля, а другую парную гонаду - в качестве испытуемой гонады. Гонады, которые дифференцировались в течение периода выращивания, культивировали в течение 6 дней в химически определенной культуральной среде при обычных условиях культивирования, смотрите Westergaard et al. Fertil. Steril. 41 (1984) 377. К культуральной среде, в которой культивировали испытуемые гонады, добавляли разное количество амфотерицина В, как указано в таблице. После периода выращивания, контрольные гонады содержали исключительно не мейотические половые клетки. Полуколичественный подсчет результатов для опытных гонад, полученный с помощью микроскопирования после окрашивания, дан в табл.5, где "-" означает отсутствие ответа, а "+", "++" и "+++" означают возрастающие ответы.
Используемый амфотерицин В получали от Bristol-Myers Squibb. Из таблицы видно, что амфотерицин активирует мейоз в мужских половых клетках дозо-зависимым образом.
Изобретение относится к биологии, а в дальнейшем может быть использовано в медицине и касается стимулирования мейоза половых клеток млекопитающих. Для этого в качестве стимулятора используют соединение - ингибитор фермента, участвующего в биосинтезе холестерина. В результате происходит накопление предшественников холестерина до уровня, при котором стимулируется мейоз. Способ позволяет повысить эффективность стимулирования мейоза. 9 з.п. ф-лы, 5 табл.
Приоритет по пунктам:
06.03.1995 по пп.1-5;
24.03.1995 по пп.6-10.7
| СКОБЛИНА М.Н | |||
| и др | |||
| Созревание in vitro ооцитов траявной лягушки, одетых фолликулярными оболочками и лишенных их под действием холестерина | |||
| - Онтогенез, 1983, т | |||
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| HAVEL C | |||
| et al | |||
| Regulation of cholesterol syntesis in primary rat hepatocyte culture cells | |||
| Possible regulatory site at sterol demethylation, J | |||
| Biol | |||
| Chem., 1979, № 10, v.254(19), p.9573-9582 | |||
| MULLER-WIELAND D | |||
| et al | |||
| Melatonin inhibits LDL receptor activity and cholesterol synthesis in freshly isolated human mononuclear leukocytes, Biochem | |||
| Biophys | |||
| Res | |||
| Commun, 1994, № 30, v.203(1), p.216-221 | |||
| KRIEGER M | |||
| The use of amphothericin В to detect of cellular cholesterol biosythesis, Anal | |||
| Biochem, 1983, Dec, v.135(2), p.383-391. | |||

