Область техники, к которой относится изобретение
Описывается ДНК-последовательность для генной терапии заболеваний сосудов. Существенными элементами ДНК-последовательности являются активаторная последовательность, промоторный модуль и ген активного вещества. Активаторная последовательность специфически к клетке активируется в гладкомышечных клетках, активированных эндотелиальных клетках, активированных макрофагах или активированных лимфоцитах. Это активирование специфически к клеточному циклу регулируется промоторным модулем. Активное вещество представляет собой ингибитор роста гладкомышечных клеток и/или свертывания. Описанную ДНК-последовательность включают в вирусный или невирусный вектор, дополненный лигандом со сродством к летке-мишени.
1). Заболевания сосудов за счет гладкомышечных клеток
Гладкомышечные клетки сосудов преобладающе локализованы в средней оболочке артериальных кровеносных сосудов и участвуют в локальной и системной регуляции кровяного давления. В случае неповрежденного, здорового сосуда эти гладкомышечные клетки находятся в состоянии покоя при делении клетки (R. Ross, Nature, 362, 801 (1993)). Повреждения сосудов приводят к перемещению гладкомышечных клеток во внутреннюю оболочку стенки сосуда, где они пролиферируют (образование новой внутренней оболочки) и образуют внеклеточные матричные компоненты.
Относящуюся к внутренней оболочке пролиферацию гладкомышечных клеток рассматривают как существенный компонент при возникновении артериосклероза (J. S. Forrester и др., Am. Coll. Cardiol., 17, 758 (1991)). Далее, эта пролиферация гладкомышечных клеток приводит к повторному стенозу (сужению) сосудов после ангиопластических операций, как также после увеличения размеров полости суженных сосудов (R.S. Schwartz и др., Am. Coll. Cardiol., 20, 1284 (1992), M.W. Liu и др., Circulation, 79, 1374 (1989)).
Как известно, артериосклероз, так же как стеноз и повторный стеноз, сосудов приводит в конечном итоге к тромбозу сосуда и тем самым зачастую к угрожающему жизни инфаркту.
До сих пор, однако, еще нет никакой терапии, которая путем ингибирования роста гладкомышечных клеток предотвращала бы стенозы сосудов. Правда, известно, что гепарин может ингибировать пролиферацию гладкомышечных клеток (Cochran и др., J. Cell. Physiol., 124, 29 (1995)), однако с помощью гепарина нельзя в достаточной мере предотвращать возникновение стенозов. Таким образом, существует потребность в новых способах для предотвращения роста гладкомышечных клеток в поврежденных сосудах и тем самым устранения опасности инфаркта. При этом используют знание о генах и молекулах, которые регулирующе вмешиваются в рост гладкомышечных клеток.
Так, известно, что протоонкоген c-myb, так же как cdc2-киназа и "ядерный антиген пролиферируюших клеток" (PCNA) принимает участие в пролиферации гладкомышечных клеток. Путем введения антисмысловых к c-myb олигонуклеотидов (Simons и др., Nature, 359, 67 (1992)), так же, как антисмысловых к сdс2-киназе олигонуклеотидов в комбинации с антисмысловыми к PCNA олигонуклеотидами (Morishita и др., Proc. Natl. Acad. Sci., 90, 8474 (1993)) непосредственно после и локально в место повреждения сосуда можно предотвращать пролиферацию гладкомышечных клеток у крысы.
Подобных результатов достигают в случае крысы и свиньи путем введения известного в качестве онкогенного супрессора гена ретинобластомы (Rb), также и здесь непосредственно после и локально в место повреждения сосуда. Для того чтобы предотвратить инактивирование вырабатываемого геном ретинобластомы продукта путем фосфорилирования применяют точечно мутированный Rb-ген (обмен Thr-246, Ser-601, Ser-605, Ser-780, Ser-786, Ser-787, Ser-800 на Ala; Thr-350 на Аrg и Ser-804 на Glu (Hamel и др., Mol. Cell. Biol., 12, 3431 (1992)), который кодирует нефосфорилируемую конститутивно активную форму Rb-протеина. Этот мутированный Rb-ген встраивают в рекомбинантный аденовирус с неполной репликацией и этот вектор вводят локально (Chang и др., Science, 267, 518 (1995)).
В другом эксперименте применяют рекомбинантный аденовирус с неполной репликацией, в котором ген вводят в тимидинкиназу вируса простого герпеса (AV-HS-TK). Продукт, вырабатываемый геном киназы, способен фосфорилировать предшественник активного вещества ("пропрепарат") ганцикловир и таким образом превращать в неуклеозидный аналог, который ингибирует синтез ДНК.
Генный вектор AV-HS-TK вводят спустя 7 дней после повреждения сосуда, однако здесь также локально в место повреждения, и затем ежедневно в течение 14 дней интраперитонеально инъецируют ганцикловир. Путем этой обработки в случае крысы достигают отчетливого ингибирования роста гладкомышечных клеток (Guzman и др. , Proc. Natl. Acad. Sci., 91, 10732 (1994)). Путем такого же рода обработки достигают подобных результатов также в случае свиньи (Ohno и др. , Science, 265, 781 (1994)). Однако здесь вектор вводят непосредственно после повреждения сосуда, а введение ганцикловира осуществляют ежедневно в течение 6 дней.
В целом эти опыты показывают возможность с помощью генотерапевтических мер, которые представляют собой вмешательство в процессы деления гладкомышечных клеток, предотвращать стеноз после повреждения сосудов.
Недостатком известных из литературы способов, однако, является то, что эффективные вещества (векторы) нужно вводить локально в место повреждений сосудов, причем, возможно, даже время от времени нужно подвергать обтурации соответствующий участок сосуда, чтобы предотвращать "смывание" векторов. Такого рода инвазивные вмешательства проводят, правда, обычным образом в рамках увеличения размеров полости суженных сосудов, однако они требуют значительных расходов и со своей стороны представляют собой значительную угрозу для пациента вследствие опасности тромбозов и эмболии.
Между прочим, несмотря на локальное введение векторов, нет гарантии в том, что они трансдуцируют только пролиферирующие гладкомышечные клетки. Трансдукция находящихся вблизи или удаленных клеток может приводить к побочным воздействиям (в том числе к трансформации клеток и индукциям опухолей), как это в настоящее время обсуждается широким кругом специалистов (Friedmann, Science, 244, 1275 (1989); Plummer, Scrip Magazine, III/29 (1995)).
В качестве альтернативы введению вышеописанных векторов системное (например, внутривенное или оральное) введение цитостатических препаратов для ингибирования пролиферации гладкомышечных клеток в место заболевания сосуда оказывает только незначительное и временное действие, с другой стороны, вызывает опасность повреждения эндотелия и приводит к значительным острым, а также хроническим побочным воздействиям.
2). Тромбозы
Тромбозы представляют собой все еще трудно излечиваемое, отчасти угрожающее жизни осложнение метаболических заболеваний, как артериосклероз, заболевания артериальных и венозных сосудов и локальные, так же как системные иммуно-реактивные синдромы (см. обзоры Philipps и др. , Blood, 71, 831 (1988); Harker, Biomed. Progr., 8, 17 (1995)).
Хотя ряд антикоагулянтов, антитромботических средств, фибринолитических препаратов и ингибиторов агрегации тромбоцитов уже давно находит применение в клинической практике и новые вещества проходят клинические испытания, до сих пор угрожающие жизни осложнения тромбоза невозможно в достаточной мере ни предотвращать, ни сдерживать (White, Scrip Magazine, 4, 6 (1994); Antiplatelet Trialists' Collaboration BMJ, 308, 81 (1994)).
Таким образом, существует потребность в новых лекарственных средствах для предотвращения и терапии тромбозов (BMJ, 305, 567 (1992); Vinazzer, Biomedical Progress, 6, 17 (1993)). Причиной значительной части тромбозов являются активированные или поврежденные эндотелиальные клетки. Они сами или стимулированные для пролиферации за счет факторов роста в крови гладкомышечные клетки сами по себе или в совокупности с активированными макрофагами, лимфоцитами, тромбоцитами и гранулоцитами вызывают активирование системы свертывания (Nemerson, Blood, 71, 1 (1988)).
Это активирование приводит в конечном счете к образованию фибрина, активированию и агрегации тромбоцитов и к образованию обогащенных фибрином или обогащенных тромбоцитами сужающих или закупоривающих кровеносные сосуды сгустков крови, тромбозов. Такого рода тромбозы в области артериальной сосудистой системы приводят к инфарктам, которые, например, в случае сердца или мозга, являются угрозой для жизни.
Применяемая до сих пор терапия с помощью антитромботических средств (как гепарин или фракции гепарина), антикоагулянтов (как кумарин), ингибиторов агрегации тромбоцитов (как аспирин) и фибринолотических препаратов (как стреп-токиназа, урокиназа или тканевые плазминогенные активаторы (tPA)), правда, оказывает подтвержденное многочисленными клиническими испытаниями профилактическое действие на угрожающие тромбозы и терапевтическое действие на существующие тромбозы, это действие, однако, является недостаточным. Причина этого в значительной мере заключается в том факте, что действие применяемых терапевтических средств не ограничивается местом заболевания, то есть тромбозом, а они действуют системно. Обусловленные тем самым кровотечения, таким образом, ограничивают как увеличение дозы, так и продолжительность применения.
3). Общее описание изобретения
Предметом изобретения является активное вещество (то есть лекарственное средство), которое можно вводить пациенту как локально, так и также системно и которое:
- влияет только на находящиеся в стадии деления гладкомышечные клетки, подавляет пролиферацию гладкомышечных клеток после повреждений сосудов или поражений сосудов и, таким образом, препятствует стенозу или повторному стенозу сосудов;
- ингибирует свертывание крови только в месте возникшего тромбоза, то есть в месте активированных и пролиферирующих эндотелиальных клеток, гладкомышечных клеток внутренней оболочки кровеносных сосудов, макрофагов и/или лимфоцитов; или
- ингибирует как пролиферацию гладкомышечных клеток, так и локально ингибирует там тромбоз.
Основной составной частью этого активного вещества является конструкция ДНК, которая состоит из следующих элементов:
(ДНК во всем тексте настоящей заявки используют в виде общего понятия как для комплементарной (кДНК), так и для геномной ДНК-последовательности)
Основной элемент этого активного вещества представляет собой регулируемый клеточным циклом промоторный модуль.
Под регулируемым клеточным циклом промоторным модулем нужно понимать, например, нуклеотидную последовательность CDE-CHR-Inr- (см. ниже). Существенной функцией промоторного модуля является ингибирование функции активаторной последовательности в GO/Gl-фазе клеточного цикла и специфической к клеточному циклу экспрессии в S/G2-фазе и, таким образом, в пролиферирующих клетках.
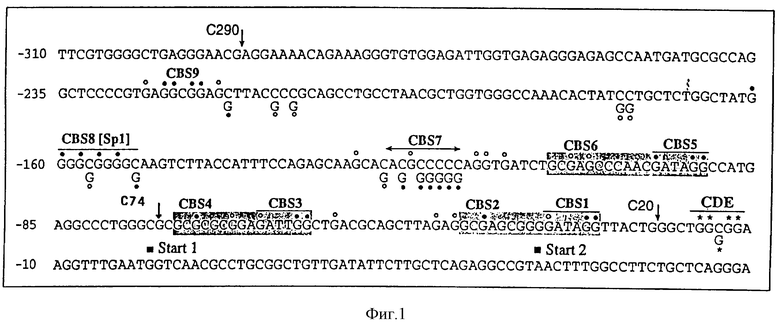
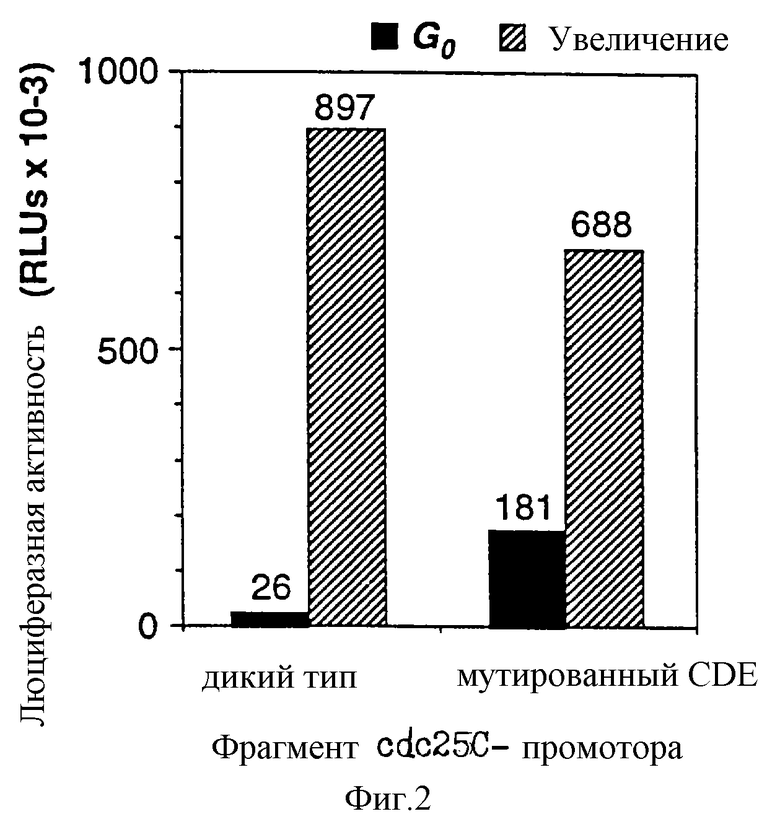
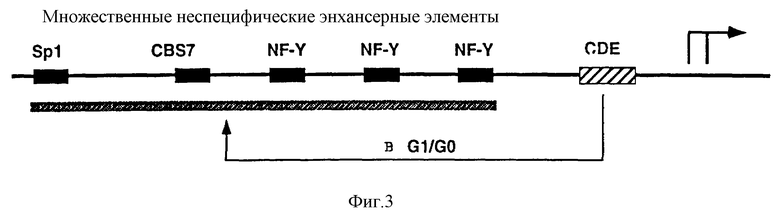
Промоторный модуль CDE-CHR-Inr обнаружен при детальном исследовании G2-специфической экспрессии человеческого промотора cdc25C. Исходным моментом являлось нахождение репрессорного элемента (зависимый от клеточного цикла элемент CDE), который является ответственным за "отключение" промотора в Gl-фазе клеточного цикла (Lucibello и др., EMBO J., 14, 132 (1995)). Путем геномного диметилсульфатного (ДМС) футпринтинга и функционального анализа (фиг. 1, 2) можно показать, что CDE Gl-специфически связывает репрессор ("CDE-связывающий фактор", CDF) и тем самым приводит к ингибированию транскрипции в непролиферирующих (GO) клетках. Локализованный в области базального промотора CDE в своей репримирующей функции зависит от "обратно активирующей (против хода транскрипции) последовательности" (UAS). Это приводит к выводу, что CDE-связывающий фактор подавляет активирующее транскрипцию действие связанных на 5'-конце активаторных протеинов зависимым от клеточного цикла образом, то есть в непролиферирующих клетках, а также в Gl-фазе клеточного цикла (фиг.3).
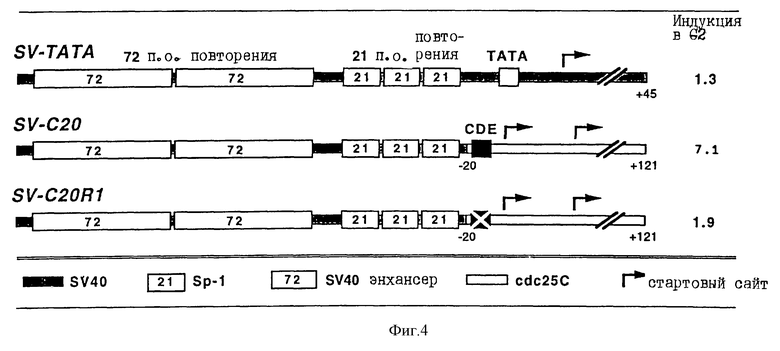
Этот вывод можно подтвердить следующим экспериментом: слияние вирус-специфического, не регулирующего клеточный цикл раннего энхансера SV40 с минимальным промотором cdc25 (состоящим из CDE и расположенной на 3'-конце стартовой области) приводит к отчетливой регуляции клеточным циклом химерного промотора (фиг.4). Последующие исследования энхансера cdc25C показывают, что в случае регулируемых зависимых от клеточного цикла CDF факторов транскрипции речь идет о NF-Y (CBF) (Dom и др., Cell, 50, 863 (1987); Van Hujisduijnen и др., ЕМВО J., 11, 3119 (1990); Coustry и др., J. Biol. Chem., 270, 468 (1995)), Sp1 (Kadonaga и др., TIBS, 11, 10 (1986)) и, возможно, новом связывающем CBS 7 факторе транскрипции (CIF). Другим представляющим интерес заключением этого исследования является обнаружение того, что NF-Y внутри энхансера cdc25C эффективно активирует транскрипцию только в сочетании по меньшей мере с одним другим комплексом NF-Y или с CIF. Как NF-Y, так и Sp1 относятся к классу обогащенных глутамином активаторов, что является важным указанием на механизм репрессии (например, взаимодействие соответственно интерференция с определенными базальными факторами транскрипции или TAFS).
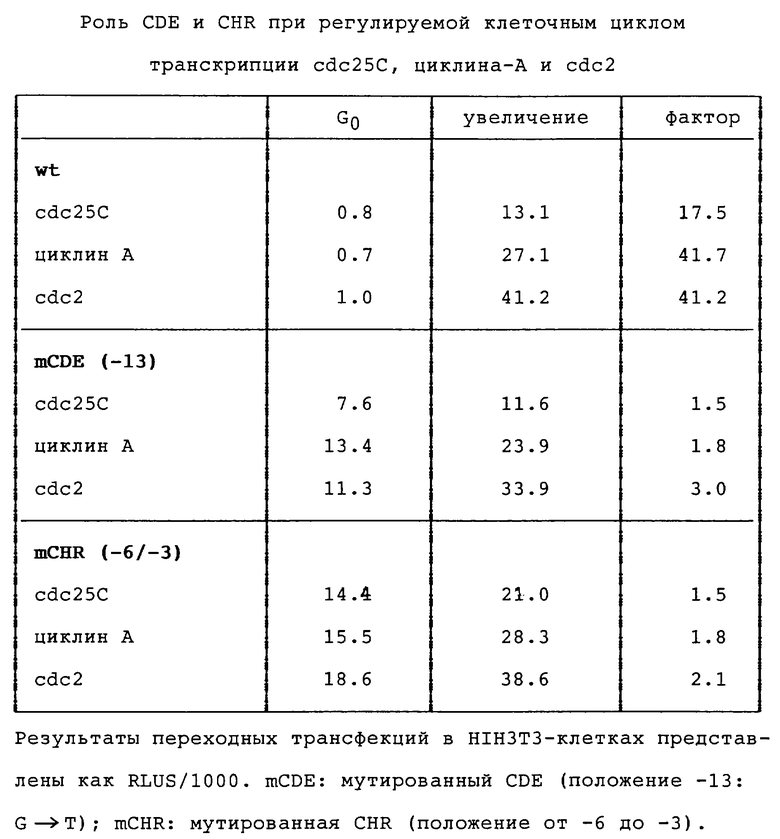
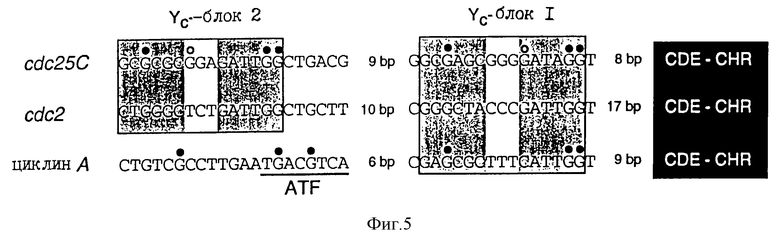
Сравнение промоторных последовательностей cdc25C, циклина А и cdc2 показывает гомологию в нескольких областях (фиг.5). Во всех трех промоторах (имеющиеся отклонения функционально безотносительны) сохраняется не только CDE, но и также соседние Ус-блоки. Все эти области, как и следовало ожидать, показывают связывание протеина ин виво в случае CDE зависимым от клеточного цикла образом. Кроме того, можно видеть, что все три промотора дерегулируются за счет мутации CDE (таблица 1). Аналогичное подобие наблюдают при сравнении cdc25C, циклина А и cdc2.
Последовательности также отчетливы (фиг.5) в области непосредственно у 3'-конца CDE ("область, гомологичная гену клеточного цикла"; CHR). Эта область функционально так же важна, как и CDE (таблица 1), однако не выявляется в экспериментах ин виво по диметилсульфатному футпринтингу. Возможным объяснением этого является взаимодействие фактора с маленьким участком ДНК. Результаты экспериментов по "анализу на резкое изменение электрофоретической подвижности" (EMSA) указывают на то, что CDE и CHR вместе связываются в протеиновый комплекс, CDF. Эти наблюдения указывают на то, что репрессия через посредство CDF обогащенных глутамином активаторов протекает по зачастую встречающемуся механизму регулируемой клеточным циклом транскрипции.
Для регуляции промотора cdc25C имеет значение, однако, не только CDE-CHR-область, но и также один из сайтов инициации (положение + 1) внутри нуклеотидной последовательности базального промотора (положения ≤ -20 до ≥ +30, см. фиг. 1). Мутации в этой области, которая включает ин витро сайт связывания фактора транскрипции YY-1 (Seto и Shenk, Nature, 354, 241 (1991); Usheva и Shenk, Cell, 76, 1115 (1994)), приводят к полной дерегуляции. Принимая во внимание близость CDE-CHR к базальному промотору, таким образом, очень вероятно взаимодействие CDF с базальным комплексом транскрипции.
В качестве активаторной последовательности (обратно активирующая (против хода транскрипции) последовательность = UAS) нужно понимать нуклеотидную последовательность (промоторную или энхансерную последовательность), с которой взаимодействуют образовавшиеся или активные в клетке-мишени факторы транскрипции. В качестве активаторной последовательности можно использовать CMV-энхансер, CMV-промотор (европейский патент 073177 В1), промотор SV40 или любую другую известную специалисту промоторную или энхансерную последовательность. Согласно настоящему изобретению, однако, к предпочтительным активаторным последовательностям причисляют такие ген-регуляторные последовательности, соответственно элементы генов, которые особенно кодируют образовавшиеся в гладкомышечных клетках, активированных эндотелиальных клетках или в активированных макрофагах или лимфоцитах протеины.
Под активным веществом нужно понимать ДНК-последовательность протеина, который может вызывать в месте образования терапевтический эффект, то есть ингибирование пролиферации гладкомышечных клеток, свертывания или (в случае двух активных веществ) ингибирование пролиферации и одновременно свертывания. При выборе нуклеотидной последовательности для активаторной последовательности и активного вещества руководствуются клеткой-мишенью и желательным активным веществом.
Предлагаемую согласно изобретению конструкцию ДНК известным специалисту образом дополняют до вектора; так, например, вводят в вирусный вектор (см. D. Jolly, Cancer Gene Therapy, 1, 51 (1994)), или, однако, дополняют до плазмиды. Вирусные векторы или плазмиды можно комплексовать с коллоидными дисперсиями, так, например, с липосомами (Farhood и др., Annal. of the New York Academy of Sciences, 716, 23 (1994)) или, однако, с конъюгатами полилизин-лиганд (Curiel и др., Annals of the New York Academy of Sciences, 716, 36 (1994)). Точно также можно получать лекарственные средства с помощью обычных вспомогательных для получения лекарственных средств веществ.
Такого рода вирусные или невирусные векторы можно дополнять лигандом, который обладает сродством к связыванию мембранной структуры с выбранной клеткой-мишенью. При выборе лиганда, таким образом, руководствуются выбором клетки-мишени (см. п.4.4-и последующие и п.5.4 и последующие). Предлагаемое согласно изобретению активное вещество подробнее поясняется, руководствуясь следующими примерами.
4). Активное вещество для ингибирования пролиферации гладкомышечных клеток:
4.1. Выбор активаторной последовательности гладкомышечных клеток
В качестве активаторных последовательностей согласно настоящему изобретению предпочтительно нужно выбирать ген-регуляторные последовательности, соответственно элементы генов, которые кодируют особенно в гладкомышечных клетках образовавшиеся протеины. Этими генами, например, являются следующие:
- тропомиозин (Tsukahara и др., Nucleic Acid Res., 22, 2318 (1994); Novy и др., Cell Motility and Cytosceleton, 25, 267 (1993); Wilton и др., Cytogenetics and Cell Genetics, 68, 122 (1995));
- α-актин (Sartorelli и др., Gens and Developm., 4, 1811 (1990); Miwa и др., Nucleic Acids Res, 18, 4263 (1990));
- α-миозин (Kelly и др., Can. J. Physiol. and Pharm., 72, 1351 (1994); Moussavi и др., Mol. Cell. Biochem., 128, 219 (1993));
- рецепторы факторов роста, как, например, фактор роста, полученный из тромбоцитов (PDGF), FGF (Rubin и др., Int. Congress Ser., 925, 131 (1990); Ross, Ann. Rev. Med., 38, 71 (1987);
- рецепторы ацетилхолина (Dutton и др., PNAS USA, 90, 2040 (1993); Durr и др., Eur. J. Biochem., 224, 353 (1994));
- фосфофруктокиназа-А (Gekakis и др., Biochemistry, 33, 1771, (1994); Tsujino и др., J. Biol. Chem., 264, 15334 (1989); Castella-Escola и др., Gene, 91, 225 (1990));
- фосфоглицератмутаза (Nakatsuji и др. , Mol. Cell. Biol., 12, 4384 (1992));
- тропонин С (Lin и др., Mol. Cell. Biol., 11, 267 (1991));
- десмин (Li и др. , J. Biol. Chem., 266, 6562 (1991); Neuromuscular Disorders, 3, 423 (1993));
- миогенин (Funk и др., PNAS USA, 89, 9484 (1992); Olson, Symp. Soc. Exp. Biol. , 46, 331 (1992); Zhon и др., Mol. Cell. Biol., 14, 6232 (1994); Atchley и др., PNAS USA, 9JL, 11522 (1994));
- рецепторы эндотелина A (Hosoda и др., J. Biol. Chem., 26 7, 18797 (1992); Orelly и др., J. Cardiovasc, Pharm., 22, 18 (1993); Hayzer и др., Am. J. Med. Sci. 304, 231 (1992); Haendler и др., J. Cardiovasc. Pharm., 20, 1 (1992));
- VEGF; VEGF образуется гладкомышечными клетками, особенно в гипоксических условиях (Berse и др., Mol. Biol. Cell, 3, 211 (1992); Finkenzeller и др. , ВВРС, 208, 432 (1995); Tischer и др., ВВРС, 165, 1198 (1989); Leung и др., Science, 246, 1306 (1989); Ferrara и др., Endoc. Fev., 13, 18 (1992)).
Промоторные последовательности генов для этих протеинов указываются в следующих работах:
- тропомиозин (Gooding и др., Embo. J., 13, 3861 (1994));
- α-актин (Shimizu и др., J. Biol. Chem., 270, 7631 (1995); Sartorelli и др., Genes Dev., 4, 1811 (1999));
- α-миозин (Kurabayashi и др., J. Biol. Chem., 265, 19271 (1990); Molkentin и др., Mol. Cell. Biol., 14, 4947 (1994));
- рецептор PDGF (Pistritto и др., Antibiot. Chemother., 46, 73 (1994));
- рецептор FGF (Myers и др., J. Biol. Chem., 270, 8257 (1995); Johnson и др. , Adv. Cencer Res., 60, 1 (1993); Chellaiah и др., J. Biol. Chem., 269, 11620 (1994); Yu и др., Hum. Mol. Genetics, 3, 212 (1994); Wang и др., ВВРС, 203, 1781 (1994); Murgue и др., Cancer Res., 54, 5206 (1994); Avraham и др., Genomics, 21, 656 (1994); Burgess и др. , Ann. Rev. Biochem., 58, 575 (1989));
- MRF-4 (Naidu и др., Mol. Cell. Biol., 15, 2707 (1995));
- фосфофруктокиназа A (Gekakis и др., Biochem., 33, 1771 (1994));
- фосфоглицератмутаза (Makatsuji и др. , Mol. Cell. Biol., 12, 4384 (1992)) ;
- тропонин С (Ip и др., Mol. Cell. Biol., 14, 7517 (1994); Parmacek и др., Mol. Cell. Biol., 12, 1967 (1992));
- миогенин (Salminen и др., J. Cell. Biol., 115, 905 (1991); Durr и др., Eur. J. Biochem. , 224, 353 (1994); Edmondson и др., Mol. Cell. Biol., 12, 3665 (1992));
- рецепторы эндотелина A (Hosoda и др., J. Biol. Chem., 267, 18797 (1992); Li и Paulia, J. Biol. Chem., 266, 6562 (1991));
- десмин (Li и др., Neuromisc. Dis., 3, 423 (1993); Li и Capetanaki, Nucl. Acids Res., 21, 335 (1993));
- VEGF.
Ген-регуляторными последовательностями VEGF-гена являются следующие:
- промоторная последовательность VEGF-гена (5'-фланкирующая область) (Michenko и др. , Cell. Mol. Biol. Res., 40, 35 (1994); Tischer и др., J. Biol. Chem., 266, 11947 (1991)); или
- энхансерная последовательность VEGF-гена (3'-фланкирующая область) (Michenko и др., Cell. Mol. Biol. Res., 40, 35 (1994)); или
- c-Src-ген (Mukhopadhyay и др., Nature, 375, 577 (1995) 7 Bonham и др., Oncogene, 8, 1973 (1993); Parker и др., Mol. Cell. Biol., 5, 831 (1985); Anderson и др., Mol. Cell. Biol., 5., 1122 (1985)); или
- V-Src-ген (Mukhodpadhyay и др., Nature, 375, 577 (1995); Anderson и др. , Mol. Cell. Biol. , 5, 1122 (1985); Gibbs и др., J. Virol., 53, 19 (1985));
- "искусственные" промоторы; факторы семейства спираль-петля-спираль (HLH) (MyoD, Myf-5, миогенин, MRF4 (обзор Olson и Klein, Genes Dev., 8, 1 (1994)) описываются в качестве специфических к мышцам активаторов транскрипции. Далее, к специфическим к мышцам факторам транскрипции относятся протеин "цинковый палец" GATA-4 (Arceci и др., Mol. Cell. Biol., 13, 2235 (1993); Ip и др., Mol. Cell. Biol., 14, 7517 (1994)), а также группы факторов транскрипции MEF-2 (Yu и др., Gene Dev., 6, 1783 (1992)).
HLH-Протеины, а также GATA-4 проявляют специфическую к мышце транскрипцию не только с промоторами специфических к мышце генов, но и также в гетерологичном окружении, а также с искусственными промоторами. Такого рода искусственными промоторами, например, являются следующие:
- множественные копии сайтов связывания (ДНК) специфических к мышце HLH-протеинов, как Е-блок (MyoD)
(например, 4хAGCAGGTGTTGGGAGGC) (Weintraub и др. , PNAS, 87, 5623 (1990));
- множественные копии сайта связывания ДНК GATA-4 α-миозин-гена вируса синдрома Франклина
(например, 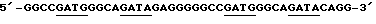 ) (Molkentin и др., Mol. Cell. Biol., 14, 4947 (1994)).
) (Molkentin и др., Mol. Cell. Biol., 14, 4947 (1994)).
4.2. Выбор активного вещества гладкомышечных клеток
В качестве активного вещества согласно изобретению нужно понимать ДНК-последовательность, экспрессированный протеин которой ингибирует пролиферацию гладкомышечных клеток. К этим ингибиторам клеточного цикла относятся, например, ДНК-последовательности следующих протеинов:
а) Ингибирующие протеины:
- протеин ретинобластомы (pRb=pllO) или родственные р107 и р130 протеины (La Thangue, Curr. Opin. Cell Biol., 6, 443 (1994));
- р53-протеин (Prives и др., Genes Dev., 1, 529 (1993));
- p21 (WAF-1)-протеин (El-Deiry и др., Cell, 75, 817 (1993));
- р16-протеин (Serrano и др. , Nature, 366, 704 (1993); Kamb и др., Science, 264, 436 (1994); Nobori и др., Nature, 368, 753 (1994));
- другие ингибиторы cdK (обзор Pines, TIBS, 19, 143 (1995));
- протеин GADD45 (Papathanasiou и др. , Mol. Cell. Biol., 11, 1009 (1991); Smith и др., Science, 266, 1376 (1994));
- bak-протеин (Farrow и др., Nature, 374, 731 (1995); Chittenden и др., Nature, 374, 733 (1995); Kiefer и др., Nature, 374, 736 (1995)).
Для того чтобы предотвратить быструю внутриклеточную инактивацию этих игнибиторов клеточного цикла, нужно предпочтительно применять такие гены, которые обладают мутациями сайтов инактивации экспримированных протеинов, при этом не ухудшая этим их в отношении их функции.
Протеин ретинобластомы (pRb/pllO) и родственные протеины р107 и р130 инактивируются за счет фосфорилирования. Таким образом, предпочтительно используют pRb/p110-, p107- или р130-кДНК-последовательность, которая мутирована таким образом, что участки фосфорилирования кодированного протеина заменены нефосфорилируемыми аминокислотами.
Согласно Hamel и др. (Mol. Cell. Biol., 12, 3431 (1992)) кДНК-последовательность протеина ретинобластомы (р110) за счет обмена аминокислот в положениях 246, 350, 601, 605, 780, 786, 787, 800 и 804 становится более нефосфорилируемой, однако ее активность связывания с большим Т-антигеном не ухудшается. Например, аминокислоты Thr-246, Ser-601, Ser-605, Ser-780, Ser-786, Ser-787 и Ser-800 заменяются на Ala, аминокислота Thr-350 заменяется на Arg и аминокислота Ser-804 заменяется на Glu.
Аналогичным образом мутируется ДНК-последовательность протеина р107 или протеина р130.
Протеин р53 инактивируется в клетке либо за счет связывания со специальными протеинами, как, например, MDM2, или вследствие олигомеризации р53 через дефосфорилированный С-концевой серин-392 (Schikawa и др., Leukemia и Lymphoma, 11, 21 (1993); Brown, Annals of Oncology, 4, 623 (1993)). Таким образом, предпочтительно используют ДНК-последовательность протеина р53, который укорочен на С-конце на серин-392.
б) Цитостатические или цитотоксические протеины
В качестве активного вещества, далее, нужно понимать ДНК-последовательность, которая экспрессирует (выражает) цитостатический или цитотоксический протеин.
К такого рода протеинам относятся, например, следующие:
- перфорин (Lin и др., Immunol. Today, 16, 194 (1995));
- гранзим (Smyth и др., Immunol. Today, 16, 202 (1995));
- фактор некроза опухоли (Porter, Tib Tech., 9, 158 (1991); Sidhu и др., Pharmac. Ther., 57, 79 (1993)); в особенности
-α-фактор некроза опухоли (Beutler и др., Nature, 320, 584 (1986); Kriegler и др., Cell, 53, 45 (1988));
-β-фактор некроза опухоли (Gray и др., Nature, 312, 721 (1984); Li и др. , J. Immunol., 138, 4496 (1987); Aggarwal и др., J. Biol. Chem., 260, 2334 (1985)).
в) Ферменты
В качестве активного вещества, однако, также нужно понимать ДНК-последовательность фермента, который инактивный предшественник цитостатического препарата превращает в цитостатический препарат.
Такого рода ферменты, которые расщепляют инактивные предшественники веществ (пропрепараты) до активных цитостатических препаратов (лекарственные средства), и возможные пропрепараты и лекарственные средства уже наглядно описаны Deonarain и др., (Br. J. Cancer, 70, 786 (1994)), Mullen (Pharmac. Ther., 63, 199 (1994)) и Harris и др. (Gene Ther., 1, 170 (1994)).
Например, можно применять ДНК-последовательность одного из следующих ферментов:
- тимидинкиназа вируса простого герпеса
(Garapin и др., PNAS USA, 76, 3755 (1979); Vile и др., Cancer Res., 53, 3860 (1993); Wagner и др., PNAS USA, 78, 1441 (1981); Moelten и др., Cancer Res., 46, 5276 (1986); J. Natl. Cancer Inst., 82, 297 (1990));
- тимидинкиназа вируса ветряной оспы
(Huber и др. , PNAS USA, 88, 8039 (1991); Snoeck, Int. J. Antimicrob. Agents, 4, 211 (1994));
- бактериальная нитроредуктаза
(Michael и др., FEMS Microbiol. Letters, 124, 195 (1994); Bryant и др., J. Biol. Chem. , 266, 4126 (1991); Watanabe и др., Nucleic Acids Res., 18, 1059 (1990));
- бактериальная β-глюкуронидаза
(Jefferson и др., PNAS USA, 83, 8447 (1986));
- растительная β-глюкуронидаза из сухих зерновых культур
(Schulz и др., Phytochemistry, 26, 933 (1987));
- человеческая β-глюкуронидаза
(Bosslet и др., Br. J. Cancer, 65, 234 (1992); Oshima и др., PNAS USA, 84, 685 (1987));
- человеческая карбоксипептидаза (СВ), например:
СВ-А тучной клетки
(Reynolds и др., J. Clin. Invest, 89, 273 (1992));
СВ-В поджелудочной железы
(Yamamoto и др., J. Biol. Chem., 267, 2575 (1992);
Catasus и др., J. Biol. Chem., 270, 6651 (1995));
- бактериальная карбоксипептидаза
(Hamilton и др. , J. Bacteriol., 174, 1626 (1992); Osterman и др., J. Protein Chem., 11, 561 (1992));
- бактериальная β-лактамаза
(Rodriges и др. , Cancer Res., 55, 63 (1995); Hussain и др., J. Bacteriol., 164, 223 (1985); Coqe и др., Embo J., 12, 631 (1993));
- бактериальная цитозиндезаминаза
(Mullen и др., PNAS USA, 89, 33 (1992); Austin и др., Mol. Pharmac., 43, 380 (1993); Danielson и др., Mol. Microbiol., 6, 1335 (1992));
- человеческая каталаза, соответственно пероксидаза
(Ezurum и др., Nucl. Acids Res., 21, 1607 (1993));
- фосфатаза, в особенности
человеческая щелочная фосфатаза
(Gum и др., Cancer Res., 50, 1085 (1990));
человеческая кислая фосфатаза простаты
(Sharieff и др. , Am. J. Hum. Gen., 49, 412 (1991); Song и др., Gene, 129, 291 (1993); Tailor и др., Nucl. Acids Res., 18, 4928 (1990));
- кислая фосфатаза типа 5
(Gene, 130, 201 (1993));
- оксидаза, в особенности
человеческая лизилоксидаза
(Kimi и др., J. Biol. Chem., 270, 7176 (1995));
человеческая кислая D-аминооксидаза
(Fukui и др., J. Biol. Chem., 267, 18631 (1992));
- пероксидаза, в особенности
человеческая глутатион-пероксидаза
(Chada и др., Genomics, 6, 268 (1990); Ishida и др., Nucl. Acids Res., 15, 10051 (1987));
человеческая эозинофильная пероксидаза
(Ten и др. , J. Exp. Med., 169, 1757 (1989); Sahamaki и др., J. Biol. Chem., 264, 16828 (1989));
- человеческая пероксидаза щитовидной железы
(Kimura, PNAS USA, 84, 5555 (1987)).
Для облегчения секреции указанных ферментов, возможно, содержащуюся в ДНК-последовательности гомологичную сигнальную последовательность можно заменять на гетерологичную улучшающую внеклеточное выделение сигнальную последовательность.
Так, например, сигнальную последовательность β-глюкуронидазы (положение в ДНК ≤ 27 до 93; Oshima и др., PNAS, 84, 685 (1987)) можно заменять сигнальной последовательностью человеческого иммуноглобулина (положение в ДНК ≤ 63 до ≥ 107; Riechmann и др., Nature, 332, 323 (1988)).
Далее, предпочтительно нужно выбирать кДНК таких ферментов, которые за счет точечных мутаций в более незначительной степени накапливаются в лизосомах. Такого рода точечные мутации описываются, например, для β-глюкуронидазы (Shipley и др., J. Biol. Chem., 268, 12193 (1993)).
4.3. Комбинация одинаковых или разных активных веществ гладкомышечных клеток
Предметом изобретения, далее, является активное вещество, в котором имеется комбинация ДНК-последовательностей нескольких одинаковых активных веществ (А, А) или разных активных веществ (А, Б). Для экспрессии двух ДНК-последовательностей предпочтительно промежуточно включают кДНК "внутреннего рибосомного аминоацильного сайта" (IRES) в качестве регуляторного элемента. Такого рода IRES описываются Mountford и Smith (TIG, 11, 179 (1995)), Kaufman и др., Nucl. Acids Res., 19, 4485 (1991); Morgan и др., Nucl. Acids Res. , 20, 1293 (1992) и Dirks и др., Gene, 129, 247 (1993); Pelletier и Sonenberg, Nature, 334, 320 (1988); и др., Bio Techn., 12, 694 (1994).
Так, кДНК IRES-последовательности полиовируса (положение ≤ 140 до ≥ 630 5'-UTR (Pelletier и Sonenberg, Nature, 334, 320 (1988)) можно применять для связывания ДНК антитромботического вещества А (по 3'-концу) и ДНК антитромботического вещества Б (по 5'-концу).
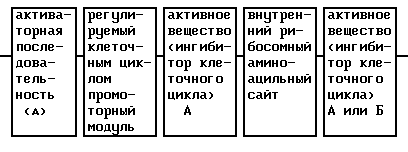
Такого рода активное вещество в зависимости от сочетания обладает аддитивным или синергическим действием согласно изобретению.
Вследствие относящегося к внутренней оболочке роста гладкомышечных клеток, так же как за счет их апоптоза или некроза, как результат воздействия ингибитора цикла может активироваться система свертывания и могут возникать тромбозы. Такого рода тромбозы нужно предотвращать путем профилактического введения ингибитора свертывания (аспирин, гепарин или другое антитромботическое средство). Введение ингибитора свертывания осуществляют системно, то есть орально или парентерально.
Однако зачастую побочные действия ингибитора свертывания препятствуют достаточной концентрации в месте относящегося к внутренней оболочке роста гладкомышечных клеток.
Таким образом, профилактика тромбоза благодаря такого рода ингибиторам свертывания является ненадежной (Pukac, Am. J. Pathol., 139, 1501 (1991)).
Следующим предметом изобретения является то, что заявляемое в изобретении активное вещество дополнительно к активному веществу, которое представляет собой ингибитор клеточного цикла, в качестве другого элемента содержит ДНК-последовательность активного вещества, которое представляет собой ингибитор свертывания.
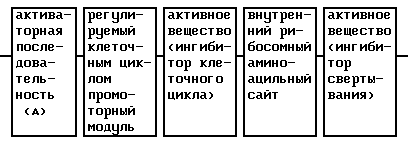
Экспрессия ингибитора свертывания регулируется таким же образом, как и экспрессия ингибитора клеточного цикла, за счет активаторной последовательности и регулируемого клеточным циклом репрессорного модуля. Одновременная экспрессия как ингибитора клеточного цикла, так же как ингибитора свертывания, регулируется предпочтительно элементом гена "внутренним рибосомным аминоацильным сайтом" (IRES).
В качестве ингибитора свертывания можно использовать гены, например, активаторов плазминогена (РА), как тканевый PA (tPA) или урокиназа-подобный PA (uPA), протеин С, антитромбин-III, ингибитор метаболического пути тканевого фактора или гирудин. ДНК-Последовательности этих ингибиторов свертывания описываются ниже:
- активатор тканевого плазминогена (tPA)
(Sasaki и др., Nucl. Acids Res., 16, 5695 (1988); Pennica и др., Nature, 301, 214 (1983); Wei и др., DNA, 4, 76 (1985); Harris и др., Mol. Biol. Med. , 3, 279 (1986));
- плазминогенный активатор урокиназного типа (uРА)
(Miyake и др. , J. Biochem., 104, 643 (1988); Nelles и др., J. Biol. Chem., 262, 5682 (1987));
- гибриды tPA и uPA
(Kalyan и др. , Gene, 68, 205 (1988); Devries и др., Biochem. 27, 2565 (1988));
- протеин С (Foster и др., PNAS, 82, 4673 (1985));
- гирудин
(Maerki и др., Semin. Thromb. Hemostas., 17, 88 (1991); De Taxis du Poet и др., Blood Coag. Fibrin., 2, 113 (1991); Harvey и др., PNAS USA, 83, 1084 (1986); Sachhieri и др., европейский патент 0324712 В1; европейский патент 0142860 В1);
- ингибиторы серин-протеиназы (серпины), как, например, C-lS-ингибитор
(Bock и др., Biochem., 25, 4292 (1986); Davis и др., PNAS USA, 83, 3161 (1986); Que, BBPC, 137, 620 (1986); Rauth и др., Proteine Sequences and Data Analysis, 1, 251 (1988); Carter и др., Eur. J. Biochem., 173, 163 (1988); Tosi и др., Gene, 42, 265 (1986); Carter и др., Eur. J. Biochem., 197, 301 (1991); Eldering и др., J. Biol. Chem., 267, 7013 (1993));
- 1-антитрипсин
(Tosi и др., Gene, 42, 265 (1986); Graham и др., Hum. Genetics, 85, 381 (1990); Hafeez и др. , J. Clin. Invest., 89, 1214 (1992); Тикунова и др., Биоорганическая химия, 17, 1694 (1991); Kay и др., Human Gene Ther., 3, 641 (1992); Lemarchand и др., Mol. Biol., 27, 1014 (1993); Lambach и др., Human Mol. Gen., 7, 1001 (1993));
- антитромбин-III
(Stackhouse и др., J. Biol. Chem., 258, 703 (1983); Olds и др., Bioshem. , 32, 4216 (1993); Laue и др., Nucl. Acids Res., 22, 3556 (1994));
- ингибитор метаболического пути тканевого фактора
(TFPI)
(Enjyoji и др. , Genomics, 17, 423 (1993); Wun и др., J. Biol. Chem., 263, 6001 (1988); Girard и др., Thromb. Res., 55, 37 (1989)).
4.4. Выбор лиганда гладкомышечных клеток
В качестве лиганда в коллоидных дисперсиях, например, полилизин-лиганд-конъюгаты, предпочтительны вещества, которые связываются с поверхностью гладкомышечных клеток. Сюда относятся антитела или фрагменты антител, направленные против мембранных структур гладкомышечных клеток, как, например:
- антитело 10F3
(Printseva и др., Exp. Cell Res., 169, 85 (1987); Am. J. Path., 134, 305 (1989)); или
- антитело против актина
(Desmonliere и др., Comptes Rendus des Seances de la Soc. de Biol. и de ses Filiales, 182, 391 (1988)); или
- антитела против рецепторов ангиотензина-II
(Butcher и др., ВВРА, 196, 1280 (1993)); или
- антитела против рецепторов факторов роста
(обзор Mendelsohn, Prog. All., 45, 147 (1988); Sato и др., J. Nat. Canс. Inst., 81, 1600 (1989); Hynes и др., ВВА, 1198, 165 (1994));
или антитела, направленные, например, против
- EGF-рецепторов
(Fan и др., Cancer Res., 53, 4322 (1993); Bender и др., Cancer Res., 52, 121 (1992); Aboud-Pirak и др., J. Nat. Cancer Inst., 80, 1605 (1988); Sato и др., Mol. Biol. Med., 1, 511 (1983); Kawamoto и др., PNAS, 80, 1337 (1983));
- или против PDGF-рецепторов
(Yu и др. , J. Biol. Chem., 269, 10668 (1994); Kelly и др., J. Biol. Chem. , 266, 8987 (1991); Bowen-Pope и др. , J. Biol. Chem., 257, 5161 (1982));
- или против FGF-рецепторов
(Vanhalteswaran и др., J. Cell. Biol., 115, 418 (1991); Zhan и др., J. Biol. Chem., 269, 20221 (1994));
- или антитела против рецепторов эндотелина-А.
Мышиные моноклональные антитела предпочтительно нужно использовать в гуманизированной форме. Гуманизирование осуществляют способом, описанным Winter и др., Nature, 349, 293 (1991), и Hoogenbooms и др., Rev. Tr. Transfus. Hemobiol., 36, 19 (1993). Фрагменты антител получают согласно уровню техники, например, способом, описанным Winter и др., Nature. 349, 293 (1991); Hoogenboom и др., Rev. Tr. Transfus. Hemobiol., 36, 19 (1993); Girol, Mol. Immunol. , 28, 1379 (1991) или Huston и др., Int. Res. Immunol., 10, 195 (1993).
К лигандам относятся, далее, все активные вещества, которые связываются с мембранными структурами или мембранными рецепторами гладкомышечных клеток (обзор Pusztai и др. , J. Pathol., 169, 191 (1993); Harris, Current Opin. Biotechnol. , 2, 260 (1991)). Например, сюда относятся факторы роста или их фрагменты, соответственно их частичные последовательности, которые связываются с экспрессированными гладкомышечными клетками рецепторами, как, например:
- PDGF
(Westermark и др. , Cancer Res., 51, 5087 (1991); Ponten и др., J. Invest. Dermatol., 102, 304 (1994));
- EGF
(Modjtahedi и др., Int. J. Oncol., 4, 277 (1994);
Carpentar и др., J. Biol. Chem., 265, 7709 (1990);
- TGF-β
(Segarini, BBA, 1155, 269 (1993));
- TGF-α
(Salomon и др., Cancer Cells, 2, 389 (1990));
- FGF
(Burgess и др., Annu. Rev. Biochem., 58, 575 (1989));
- эндотелин-А
(Oreilly и др., J. Cardiovasc. Pharm., 22, 18 (1993)).
4.5. Получение биологически активного вещества гладкомышечных клеток
Получение предлагаемого согласно изобретению активного вещества описывается подробнее, руководствуясь следующими примерами:
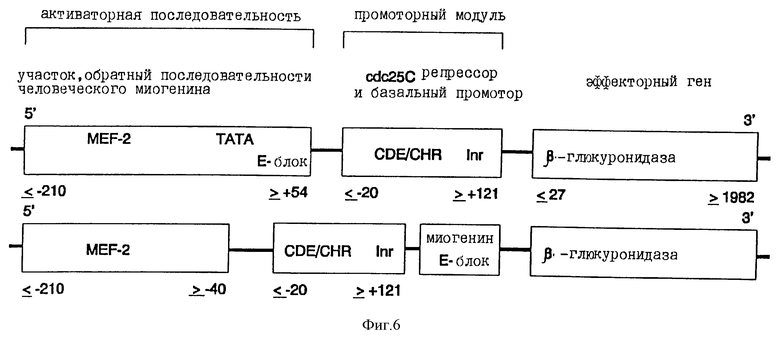
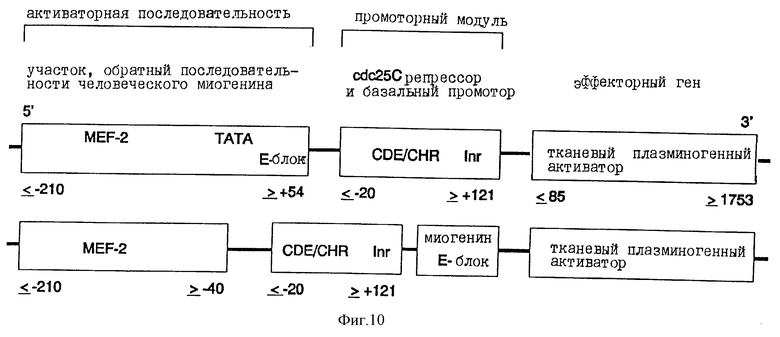
а). Конструкция химерного промотора миогенин-промотор CDE-CHR-Inr
Человеческий миогениновый промотор (положение ≤ -210 до ≥ +54 ДНК-последовательности, опубликованной Salmin и др. , J. Cell. Biol., 115, 905 (1991)) своим 3'-концом связывается с 5'-концом CDE-CHR-Inr-модуля человеческого cdc25C-гeнa (положение ≤ -20 до ≥ +121 последовательности, опубликованной Lucibello и др., EMBO J., 14, 132 (1995)) (см. фиг.6). Связывание осуществляется с помощью известных специалисту и имеющихся в продаже ферментов. Кроме того, применяют различные фрагменты миогениновой промоторной последовательности (см. фиг. 6). Так, применяют содержащую ТАТА-блок ДНК-последовательность миогенинового промотора. Однако равным образом можно применять также промоторную последовательность в положении ≤ -210 до ≥ -40 (см. фиг.6).
б). Конструкция плазмиды, содержащей основную (центральную) составную часть активного вещества
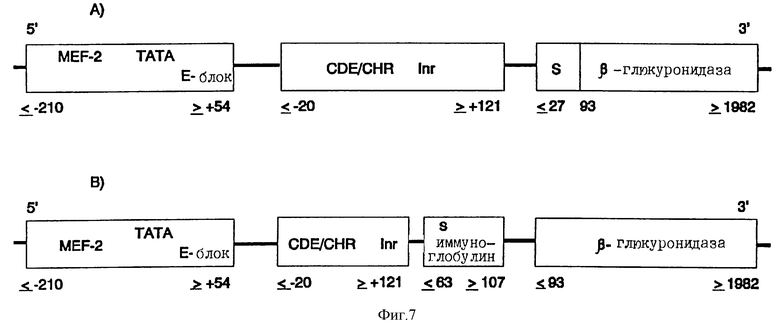
Таким образом полученную химерную контрольную единицу транскрипции промоторного модуля миогенина ее 3'-концом связывают с 5'-концом ДНК, которая содержит полную кодирующую область человеческой β-глюкуронидазы (положение в ДНК ≤ 27 - ≥ 1982 последовательности, опубликованной Oshima и др., PNAS USA, 84, 684 (1987)). (см. фиг.6).
Эта ДНК также содержит необходимую для секреции сигнальную последовательность (22 N-концевые аминокислоты). Для облегчения клеточного выделения эту сигнальную последовательность предпочтительно обменивают на сигнальную последовательность иммуноглобулина (положение ≤ 63 до ≥ 107; Riechmann и др. , Nature, 332, 323 (1988)) (фиг.7). Таким образом полученные контрольные единицы транскрипции и ДНК человеческой β-глюкуронидазы клонируют в pUC18/19 или производимых от Bluescript плазмидных векторов, которые можно использовать непосредственно или в коллоидных дисперсионных системах для введения ин виво. Альтернативно, химерные гены можно трансферировать в вирусные векторы или другие пригодные векторы и вводить путем инъекции.
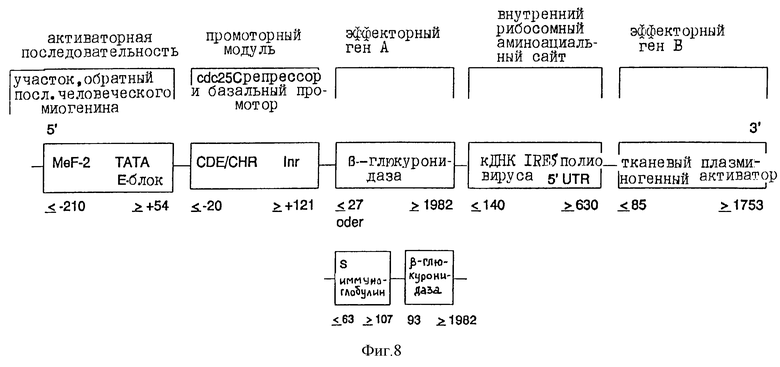
в). Конструкция плазмиды, содержащей гены β-глюкуронидазы, как также тканевый плазминогенный активатор
Полученный как в п.2) элемент β-глюкуронидазы миогенинового репрессорного модуля его 3'-концом связывают с 5'-концом кДНК "внутреннего рибосомного аминоациального сайта" полиовируса (положение ≤ 140 до ≥ 630 5'-UTR-элемента Pelletier и Sonenberg, Nature, 334, 320 (1988)). С его 3'-концом снова связывают 5'-конец ДНК тканевого плазминогенного активатора (положение ≤ 85 до ≥ 1753; Pennica и др., Nature, 301, 214 (1983)) (фиг.7). Затем всю конструкцию клонируют в pUC17/19 или производимых от Bluescript плазмидных векторах, которые можно использовать непосредственно или в коллоидальных дисперсионных системах для переноса ин виво. Альтернативно, химерные гены можно трансферировать в вирусные векторы или другие пригодные векторы (фиг. 8).
5). Активное вещество для ингибирования свертывания
5.1. Выбор активаторной последовательности для игнибирования или свертывания
В качестве активаторных последовательностей согласно изобретению предпочтительно нужно применять ген-регуляторные последовательности, соответственно элементы из генов, которые кодируют обнаруживаемые в гладкомышечных клетках, в активированных эндотелиальных клетках, в активированных макрофагах или в активированных лимфоцитах протеины.
а) Гладкомышечные клетки
Примеры активаторных последовательностей генов в гладкомышечных клетках уже указаны в п.4.1.
б). Активированные эндотелиальные клетки
Примеры протеинов, которые особенно образуются в активированных эндотелиальных клетках, описываются Burrows и др., (Pharmac. Therp., 64, 155 (1994)) и Plate и сотр., (Brain Pathol., 4, 207 (1994)). В особенности к этим интенсивно образующимся в эндотелиальных клетках протеинам относятся, например, следующие:
- специфический к мозгу эндотелиальный переносчик глюкозы-I; эндотелиальные клетки мозга отличаются очень сильной экспрессией этого переносчика, чтобы осуществить трансэндотелиальную транспортировку D-глюкозы в мозг (Gerhart и др., J. Neurosci. Res., 22, 464 (1989)). Промоторная последовательность описывается Murakami и др., J. Biol. Chem., 267, 9300 (1992);
- эндоглин; эндоглин, по-видимому, является не переносящим сигнала рецептором TGF-β (Gougos и др., J. Biol. Chem., 265, 8361 (1990); Cheifetz, 1. Biol. Chem., 267, 19027 (1992); Moren и др., ВВРС, 189, 356 (1992)). В незначительных количествах он встречается в нормальном эндотелии, однако усиленно экспримируется пролиферирующим эндотелием (Westphal и др., J. Invest. Derm. , 100, 27 (1993); Burrows и др., Pharmac. Ther., 64, 155 (1994)). Промоторная последовательность описывается Bellon и др. (Eur. J. Immunol., 23, 2340 (1993)) и Ge и др. (Gene, 138, 201 (1994)).
- VEGF-рецепторы:
Различают два рецептора (Plate и др., Int. J. Cancer, 59, 520 (1994)):
- VEGF-рецептор-1 (flt-1)
(Vries и др., Science, 255, 989 (1992))
(в цитостатической части содержит fms-подобную тирозинкиназу); и
- VEGF-рецептор-2 (flk-1, KDR)
(Terman и др., ВВРС, 187, 1579 (1992))
(содержит в цитоплазмической части тирозинкиназу).
Оба рецептора почти исключительно встречаются в эндотелиальных клетках (Senger и др., Cancer Metast. Rev., 12, 303 (1993));
- другие специфические к эндотелию рецепторные тирозинкиназы:
- til-1 или til-2
(Partanen и др. , Mol. Cell Biol., 12, 1698 (1992);  и Risau, Development, 119, 957 (1993); Dumont и др., Oncogene, 7, 1471 (1992));
и Risau, Development, 119, 957 (1993); Dumont и др., Oncogene, 7, 1471 (1992));
- В61-рецептор (Eck-рецептор)
(Bartley и др., Nature, 368, 558 (1994); Pandey и др., Science, 268, 567 (1995); van der Geer и др., Ann. Rev. Cell Biol., 10, 251 (1994));
- B61
Молекула B61 представляет собой лиганд В61-рецептора
(Hoizman и др., J. Am. Soc. Nephrol., 4, 466 (1993); Bartley и др., Nature, 368, 558 (1994));
- эндотелии, в особенности:
- эндотелин В
(Oreilly и др., J. Cardiovasc. Pharm., 22, 18 (1993); Benatti и др., J. Clin. Invest., 91, 1149 (1993); O'Reilly и др., ВВРС, 193, 834 (1993)). Промоторная последовательность описывается Benatti и др., J. Clin. Invest., 91, 1149 (1993);
- эндотелин-1
(Yanasigawa и др. , Nature, 332, 411 (1988)). Промоторная последовательность описывается Wilson и др., Mol. Cell. Biol., 10, 4654 (1990);
- рецепторы эндотелина, в особенности рецептор эндотелина-В
(Webb и др. , Mol. Pharmacol., 47, 730 (1995); Haendler и др., J. Cardiovasc. Pharm., 20, 1 (1992));
- манноза-6-фосфат-рецепторы
(Perales и др., Eur. J. Biochem., 226, 225 (1994)).
Промоторные последовательности описываются Ludwig и др.
(Gene, 142, 311 (1994)), Oshima и др. , (J. Biol. Chem., 263, 2553 (1988)) и Pohlmann и др., (PNAS USA, 84, 5575 (1987));
- фактор Виллебранда;
промоторная последовательность описывается Jahroudi и Lynch (Mol. Cell. Biol. , 14, 999 (1994); Ferreira и др., Biochem. J., 293, 641 (1993)) и Aird и др. (PNAS USA, 92, 4567 (1995));
- интерлейкин-lα, интерлейкин-1β.
Интерлейкин-1 продуцируется активированными эндотелиальными клетками (Wamer и др., J. Immunol., 139, 1911 (1987)). Промоторные последовательности описываются Hangen и др. , Mol. Carcinog., 2, 68 (1986), Turner и др., J. Iimnunol. , 143, 3556 (1989); Fenton и др., J. Immunol., 138, 3972 (1987); Bensi и др., Cell. Growth. Diff., 1, 491 (1990); Hiscott и др., Mol. Cell. Biol., 13, 6231 (1993) и Mori и др., Blood, 84, 1688 (1994);
- рецептор интерлейкина-1;
промоторная последовательность описывается Ye и др., PNAS USA, 90, 2295 (1993);
- адгезионная молекула сосудистой клетки (VCAM-1). Экспрессия VCAM-1 в эндотелиальных клетках активируется липополисахаридами, α-фактором некроза опухоли (Neish и др., Mol. Cell. Biol., 15, 2558 (1995)); интерлейкином-4 (Iademarko и др., J. Clin. Invest., 95, 264 (1995)) и интерлейкином-1 (Marni и др., J. Clin. Invest., 92, 1866 (1993)).
Промоторная последовательность VCAM-1 описывается Neish и др., Mol. Cell. Biol., 15, 2558 (1995); Ahmad и др., J. Biol. Chem., 270, 8976 (1995); Neish и др., J. Exp. Med., 176, 1583 (1992); Iademarco и др., J. Biol. Chem. , 267, 16323 (1992) и Cybulsky и др., PNAS USA, 88, 7859 (1991);
- синтетическая активаторная последовательность. В качестве альтернативы природным специфическим к эндотелию промоторам также можно применять синтетические активаторные последовательности, которые состоят из олигомеризованных сайтов связывания факторов транскрипции, предпочтительно или селективно активных в эндотелиальных клетках. Примером их является фактор транскрипции GATA-2, сайтом связывания которого в эндотелине-1 является ген 5'-ТТАТСТ-3' (Lee и др. , Biol. Chem., 266, 16188 (1991); Dorfmann и др., J. Biol. Chem. , 267, 1279 (1992) и Wilson и др., Mol. Cell. Biol., 10, 4854 (1990)).
в). Активированные макрофаги и/или активированные лимфоциты
В качестве активаторной последовательности согласно настоящему изобретению, кроме того, нужно понимать промоторные последовательности генов протеинов, которые интенсивно образуются при иммунной реакции в макрофагах и/или лимфоцитах. Сюда относятся, например:
- интерлейкин-1
(Bensi и др., Gene, 52, 95 (1987); Fibbe и др., Blut., 59, 147 (1989));
- рецептор интерлейкина-1
(Colotta и др., Immunol. Today, 15, 562 (1994); Sims и др., Clin. Immunol. Immunopath., 72, 9 (1994); Ye и др., PNAS USA, 90, 2295 (1993));
- интерлейкин-2
(Jansen и др. , Cell, 39, 207 (1994); Ohbo и др., J. Biol. Chem., 270, 7479 (1995));
- рецептор интерлейкина-2 (Semenzato и др., Int. J. Clin. Lab. Res., 22, 133 (1992));
- γ-интерферон
(Kirchner, DMW, 111, 64 (1986); Lehmann и др., J. Immunol., 153, 165 (1994));
- интерлейкин-4
(Paul, Blood, 77, 1859 (1991); te Velde и др., Blood, 76, 1392 (1990));
- рецептор интерлейкина-4
(Vallenga и др., Leukemia, 7, 1131 (1993); Gelizzi и др., Int. Immunol., 2, 669 (1990));
- интерлейкин-3
(Frendl, Int. J. Immunopharm., 14, 421 (1992));
- интерлейкин-5
(Azuma и др., Nucl. Acid Res., 14, 9149 (1986); Yokota и др., PNAS, 84, 7388 (1987));
- интерлейкин-6
(Brack и др., Int. J. Clin. Lab. Res., 22, 143 (1992);
- LIF
(Metcalf, Int. J. Cell Clon. , 9, 95 (1991); Samal, ВВА, 1260, 27 (1995));
- интерлейкин-7
(Joshi и др., 21, 681 (1991));
- интерлейкин-10
(Benjamin и др., Leuk. Lymph., 12, 205 (1994); Fluchiger и др., J. Exp. Med., 179, 91 (1994));
- интерлейкин-11
(Yang и др., Biofactors, 4, 15 (1992));
- интерлейкин-12
(Kiniwa и др., J. Clin. Invest., 90, 262 (1992), Gatelay, Cancer Invest. , 11, 500 (1993));
- интерлейкин-13
(Punnonen и др. , PNAS, 90, 3730 (1993); Muzio и др., Blood, 83., 1738 (1994));
- GM-CSF
(Metcalf, Cancer, 15, 2185 (1990));
- рецептор GM-CSF
Nakagawa и др., J. Biol. Chem., 269, 10905 (1994));
- адгезионные протеины, как интегрин, бета-2-протеин
(Nueda и др., J. Biol. Chem., 268, 19305 (1993)).
Промоторные последовательности этих протеинов известны из следующих публикаций:
- рецептор интерлейкина-1
(Ye и др., PNAS USA, 90, 2295 (1993));
- интерлейкин-lα.
(Hangen и др., Mol. Carcinog., 2, 68 (1986); Turner и др., J. Immunol., 143, 3556 (1989); Mori и др., Blood, 84, 1688 (1994));
- интерлейкин-lβ
(Fenton и др., J. Immunol., 138, 3972 (1987); Bensi и др., Cell Growth. Diff., 1, 491 (1990); Turner и др., J. Immunol., 143, 3556 (1989); Hiscott и др., Mol. Cell. Biol., 13, 6231 (1993));
- интерлейкин-2
(Fujita и др., Cell, 46, 401 (1986); Наmа и др., J. Exp. Med., 181, 1217 (1995); Kant и др. , Lymph. Rec. Interact., 179 (1989); Kamps и др., Mol. Cell. Biol., 10, 5464 (1990); Williams и др., J. Immunol., 141, 662 (1988); Brunvand, FASEB J., 6, А 998 (1992));
- рецептор интерлейкина-2
(Ohbo и др. , J. Biol. Chem., 270, 7479 (1995); Shibuya и др., Nucl. Acids Res. , 18, 3697 (1990); Lin и др., Mol. Cell. Biol., 13, 6201 (1993); Williams и др., J. Immunol., 141, 662 (1988));
- γ-интерферон
(Ye и др., J. Biol. Chem., 269, 25728 (1994));
- интерлейкин-4
(Rooney и др., EMBO J., 13, 625 (1994); Hama и др., J. Exp. Med., 181, 1217 (1995); Li-Weber и др., J. Immunol, 153, 4122 (1994); Min и др., J. Immunol., 148, 1913 (1992); Abe и др., PNAS, 89, 2864 (1992));
- рецептор интерлейкина-4
(Beckmann и др., Chem. Immunol., 51, 107 (1992); Ohara и др., PNAS, 85, 8221 (1988));
- интерлейкин-3
(Mathey-Prevot и др., PNAS USA, 87, 5046 (1990); Cameron и др., Blood, 83, 2851 (1994); Arai и др., Lymphokine Res., 9, 551 (1990));
- рецептор интерлейкина-3 (α-субъединица);
Miyajima и др., Blood, 85, 1246 (1995); Rapaport и др., Gene, 137, 333 (1993); Kosugi и др., ВВРС, 208, 360 (1995));
- рецептор интерлейкина-3 (β-субъединица)
(Gorman и др., J. Biol. Chem., 267, 15842 (1992); Kitamura и др., Cell, 66, 1165 (1991); Hayashida и др., PNAS USA, 87, 9655 (1990));
- интерлейкин-5
(Lee и др., J. Allerg. Clin. Immunol., 94, 594 (1994); Kauhansky и др., J. Immunol., 152, 1812 (1994); Staynov и др., PNAS USA, 92, 3606 (1995));
- интерлейкин-6
(Lu и др. , J. Biol. Chem., 270, 9748 (1995); Gruss и др., Blood, 80, 2563 (1992); Ray и др., PNAS, 85, 6701 (1988); Droogmans и др., DNA-Sequence, 3, 115 (1992); Mori и др., Blood, 84, 2904 (1994); Liberman и др., Mol. Cell. Biol., 10, 2327 (1990); Ishiki и др., Mol. Cell. Biol., 10, 2757 (1990));
- интерлейкин-7
(Pleiman и др. , Mol. Cell. Biol., 11, 3052 (1991); Lapton и др., J. Immunol., 144, 3592 (1990));
- интерлейкин-8
(Chang и др. , J. Biol. Chem., 269, 25277 (1994); Sprenger и др., J. Immunol., 153, 2524 (1994));
- интерлейкин-10
(Kim и др., J. Immunol., 148, 3616 (1992); Platzer и др., DNA-Sequence, 4, 399 (1994); Kube и др., Cytokine, 7, 1 (1995));
- интерлейкин-11
(Yang и др., J. Biol. Chem., 269, 32732 (1994));
- GM-CSF
(Nimer и др. , Mol. Cell. Biol., 10, 6084 (1990); Staynov и др., PNAS USA, 92, 3606 (1995); Koyano-Nakayawa и др., Int. Immunol., 5, 345 (1993); Ye и др., Nucl. Acids Res., 22, 5672 (1994));
- рецептор GM-CSF (α-цепь)
(Nakagawa и др., J. Biol. Chem., 269, 10905 (1994));
- рецептор макрофаг-колониестимулирующего фактора (M-CSF)
(Yue и др., Mol. Cell. Biol., 13, 3191 (1993); Zhang и др., Mol. Cell. Biol., 14, 373 (1994));
- тип I и тип II макрофаговых сапрофитных рецепторов
(Moulton и др., Mol. Cell. Biol., 14, 4408 (1994));
- интерлейкин-13
(Staynov и др., PNAS USA, 92, 3606 (1995));
- LIF
(Gough и др., Ciba Found. Symp., 167, 24 (1992); Stahl и др., Cytokine, 5, 386 (1993));
- интерфероновый регуляторный фактор-1, промотор которого стимулируется интерлейкином-6, также гамма- или бета-интерфероном
(Harrock и др., ЕМВО J., 13, 1942 (1994));
- реактивный промотор γ-интерферона
(Lamb и др., Blood, 83, 2063 (1994));
- γ-интерферон
(Hardy и др., PNAS, 82, 8173 (1985));
- MAC-1
(Dziennis и др., Blood, 85, 319 (1995); Bauer и др., Hum. Gene Ther., 5, 709 (1994); Hickstein и др., PNAS USA, 89, 2105 (1992));
- LFA-1
(Nueda и др., J. Biol. Chem., 268, 19305 (1993); Agura и др., Blood, 79, 602 (1992); Cornwell и др., PNAS USA, 90, 4221 (1993));
- р150, 95
(Noti и др., DNA and Cell. Biol., 11, 123 (1992); Lopezcabrera и др., J. Biol. Chem., 268, 1187 (1993)).
5.2. Выбор активного вещества для ингибирования свертывания
В качестве активного вещества согласно настоящему изобретению нужно понимать ДНК-последовательность, которая кодирует протеин, прямо или косвенно ингибирующий агрегацию тромбоцитов, или фактор свертывания крови, или стимулирующий фибринолиз.
Такого рода активное вещество обозначают как ингибитор свертывания. В качестве ингибиторов свертывания можно использовать гены, например, активаторов плазминогена (РА), так, тканевый PA (tPA), или урокиназоподобный PA (uPA), или протеин С, антитромбин-III, C-IS-ингибитор, α-1-антитрипсин, ингибитор метаболического пути тканевого фактора (TFPI) или гирудин. ДНК-последовательности этих ингибиторов свертывания уже описаны в разделе 4.3.
5.3. Комбинация двух одинаковых или разных активных веществ для ингибирования свертывания
Предметом изобретения, далее, является активное вещество, в котором встречается сочетание ДНК-последовательностей: двух одинаковых, ингибирующих свертывание веществ (А, А), или двух различных, ингибирующих свертывание веществ (А, Б). Для экспрессии обеих ДНК-последовательностей предпочтительно промежуточно включают кДНК "внутреннего рибосомного аминоацильного сайта" (IRES) в качестве регуляторного элемента.
Такого рода IRES описываются, например, Montford и Smith (TIG, 11, 179 (1995); Kaufman и др. , Nucl. Acids Res., 19, 4485 (1991); Morgan и др., Nucl. Acids Res. , 20, 1293 (1992); Dirks и др., Gene, 128, 247 (1993); Pelletier и Sonenberg, Nature, 334, 320 (1988) и Sugitomo и др., Bio-Techn., 12, 694 (1994).
Так, можно использовать кДНК IRES-последовательности полиовируса (положение ≤ 140 до ≥ 630 5'-UTR (Pelletier и Sonenberg, Nature, 334, 320 (1988)) для связывания ДНК антитромботического вещества А (по 3'-концу) и ДНК антитромботического вещества Б (по 5'-концу).
Такого рода сочетание двух одинаковых или разных генов способствует аддитивному действию (в случае одинаковых генов) или синергическому действию выбранных антитромботических веществ.
5.4. Выбор лиганда для ингибирования свертывания
В качестве лиганда вирусных или невирусных векторов, например, в коллоидных дисперсиях, содержащих полилизин-лиганд-конъюгаты, предпочтительны вещества, которые связываются с поверхностью гладкомышечных клеток, или пролиферирующих эндотелиальных клеток, или активированных макрофагов и/или лимфоцитов.
а). Лиганды гладкомышечных клеток
Примеры лигандов, которые связываются с гладкомышечными клетками, уже указаны в разделе 4.4.
б). Лиганды активированных эндотелиальных клеток
Сюда относятся согласно изобретению антитела или фрагменты антител, направленные против мембранных структур эндотелиальных клеток, которые описываются, например, Burrows и др. (Pharmac. Ther., 64, 155 (1994)), Hughes и др. (Cancer Res. , 49, 6214 (1989)) и Maruyama и др. (PNAS USA, 87, 5744 (1990)). В особенности сюда относятся антитела против VEGF-рецепторов.
Мышиные моноклональные антитела предпочтительно нужно использовать в гуманизированной форме. Гуманизирование осуществляют описанным в разделе 4.4. образом. Фрагменты антител получают согласно уровню техники, например, описанным в разделе 4.4 способом.
К лигандам, далее, относятся все активные вещества, которые связываются с мембранными структурами или мембранными рецепторами эндотелиальных клеток. Например, сюда относятся вещества, которые содержат концевую маннозу, далее, интерлейкин-1 или факторы роста или их фрагменты, соответственно их частичные последовательности, которые связываются с рецепторами, экспрессируемыми эндотелиальными клетками, как, например, PDGF, bFGF, VEGF, TGF-β (Pusztain и др., J. Pathol., 169, 191 (1993)).
Далее, к таким лигандам относятся адгезионные молекулы, которые связываются с активированными и/или пролиферирующими эндотелиальными клетками. Такого рода адгезионные молекулы, как, например, SLex, LFA-1, MAC-1, LECAM-1 или VLA-4, уже описаны (обзоры Augustin-Voss и др., J. Cell. Biol., 119, 483 (1992); Pauli и др., Cancer Metast. Rev., 9, 175 (1990); Honn и др., Cancer Metast. Rev., 11, 353 (1992)).
в). Лиганды активированных макрофагов и/или активированных лимфоцитов
К лигандам согласно изобретению относятся, далее, вещества, которые специфически связываются с поверхностью иммунных клеток. Сюда относятся антитела или фрагменты антител, направленные против мембранных структур иммунных клеток, которые описываются, например, Powelson и др., Biotech. Adv., 11, 725 (1993).
Далее, к лигандам также относятся моноклональные или поликлональные антитела или фрагменты антител, которые с помощью своих постоянных доменов связываются с Fc - или μ-рецепторами иммунных клеток (Rojanasakul и др., Pharm. Res., 11, 1731 (1994)).
Здесь также предпочтительно используют мышиные моноклональные антитела в гуманизированной форме (см. раздел 4.4.) и фрагменты получают, например, с помощью указанной в разделе 4.4 методики.
К лигандам, далее, относятся все вещества, которые связываются с мембранными рецепторами на поверхности иммунных клеток. Например, сюда относятся факторы роста, как цитокины, EGF, TGF, FGF или PDGF или их фрагменты, соответственно их частичные последовательности, которые связываются с рецепторами, экспрессируемыми такого рода клетками.
5.5. Получение активного вещества для ингибирования свертывания
Получение предлагаемого согласно изобретению активного вещества описывается подробнее, руководствуясь следующими примерами.
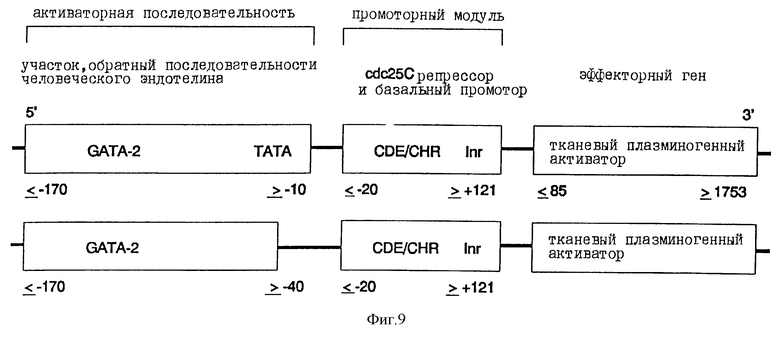
а). Конструкция химерного промотора эндотелина-1-CDE-CHR-Inr
Человеческий промотор эндотелин-1 (положение ≤ -170 до ≥ -10; Wilson и др., Mol. Cell. Biol., 10, 4854 (1990)) или укороченный на ТАТА-блок вариант (положение ≤ -170 до ≥ -40) по его 3'-концу связывают с 5'-концом CDE-CHR-Inr-модуля (положение ≤ -20 до ≥ +121) человеческого cdc25C-гена (Lucibello и др. , EMBO J. , 14, 132 (1995)) (фиг.9). Связывание осуществляют с помощью известных специалисту и имеющихся в продаже ферментов.
б). Конструкция плазмиды, содержащей химерный промотор эндотелин-1-CDE-CHR-Inr в центральной составной части активного вещества
Описанную транскрипционную единицу химерного промоторного модуля эндотелина-1 связывают по ее 3'-концу с 5'-концом ДНК, которая содержит полную кодирующую область тканевого плазминогенного активатора (положение ≤ 85 до ≥ 1753; Pennica и др., Nature, 301, 214 (1983)) (фиг.9). Эта ДНК также содержит необходимую для секреции сигнальную последовательность. Транскрипционные контрольные единицы и ДНК тканевого плазминогенного активатора клонируют в pUC19/19 или производимых от Bluescript плазмидных векторах, которые можно использовать непосредственно или в коллоидальных дисперсионных системах для введения ин виво. Альтернативно, химерные гены трансферируют в вирусные векторы или другие пригодные векторы и инъецируют.
в). Конструкция химерного промотора CDE-CHR-Inr миогенина
Человеческий миогениновый промотор (положение ≤ -210 до ≥ +54 ДНК-последовательности, опубликованной Salmin и др. , J. Cell. Biol., 115, 905 (1991)) по его 3'-концу связывают с 5'-концом CDE-CHR-Inr-модуля человеческого cdc25C-гена (положение ≤ -20 до ≥ +121 последовательности, опубликованной Lucibello и др., ЕМВО J., 14, 132 (1995)) (см. фиг.10). Связывание осуществляют с помощью известных специалисту и имеющихся в продаже ферментов. Кроме того, используют различные фрагменты промоторной последовательности миогенина (см. фиг.10). Так, применяют содержащую ТАТА-блок ДНК-последовательность промотора миогенина. Однако равным образом также можно использовать промоторную последовательность в положении ≤ -210 до ≥ -40.
г). Конструкция плазмиды, содержащей химерный промотор CDE-CHR-Inr миогенина в центральной составной части активного вещества
Таким образом полученную контрольную транскрипционную единицу химерного промоторного модуля миогенина по ее 3'-концу связывают с 5'-концом ДНК, которая содержит полную кодирующую область тканевого плазминогенного активатора (см. фиг. 10). Эта ДНК также содержит необходимую для секреции сигнальную последовательность. Транскрипционные контрольные единицы и ДНК тканевого плазминогенного активатора клонируют в pUC18/19 или производимых от Bluescript плазмидных векторах, которые непосредственно или в коллоидальной дисперсионной системе можно использовать для введения ин виво. Альтернативно, химерные гены можно трансферировать в вирусные векторы или другие пригодные векторы и инъецировать.
д). Конструкция плазмиды, содержащей два гена активных веществ
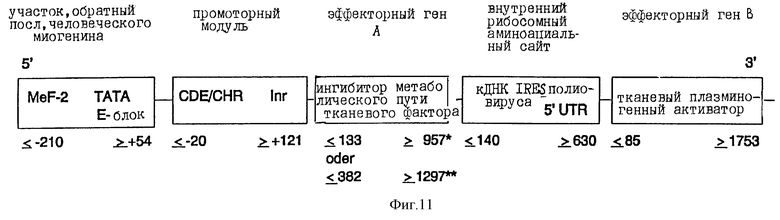
Описанную в п. в) транскрипционную единицу CDE-CHR-Inr миогенина по ее 3'-концу связывают с 5'-концом ДНК ингибитора метаболического пути тканевого фактора (TFPI; положение ≤ 133 до ≥ 957; Wun и др., J. Biol. Chem., 263, 6001 (1988); или положение ≤ 382 до ≥ 1297; Girard и др., Thromb. Res., 55, 37 (1989)). Связывание осуществляют с помощью известных специалисту и имеющихся в продаже ферментов.
3'-конец ДНК TFPI теперь связывают с 5'-концом кДНК внутреннего рибосомного аминоацильного сайта (положение ≤ 140 до ≥ 630; Pelletier и Sonnenberg, Nature., 334, 320 (1988)) и затем его 3'-конец связывают с 5'-концом ДНК тканевого плазминогенного активатора (см. фиг.11). Это таким образом полученное активное вещество, наконец, клонируют в pUC18/19 или производимых от Bluescript плазмидных векторах, которые непосредственно или в коллоидальной дисперсионной системе можно использовать для введения ин виво. Альтернативно, химерные гены можно трансферировать в вирусные векторы или другие пригодные векторы и инъецировать.
6. Действие активного вещества на гладкомышечные клетки и/или на свертывание
Активное вещество согласно настоящему изобретению после локального или системного, предпочтительно внутривенного или внутриартериального введения оказывает преимущественное, если не исключительное, действие на такие гладкомышечные клетки, которые непосредственно доступны вследствие повреждения или травмирования сосуда (в особенности слой эндотелия) и, возможно, после попадания во внутреннюю оболочку объема кровеносного сосуда.
Благодаря сочетанию специфической к ткани активаторной последовательности и регулируемого клеточным циклом репрессорного модуля обеспечивается то, что ингибитор клеточного цикла активируется преобладающе или исключительно в делящихся гладкомышечных клетках.
Благодаря предлагаемому согласно изобретению применению мутированных ингибиторов клеточного цикла обеспечивается их более продолжительное по времени ингибирующее пролиферацию действие.
Активное вещество согласно настоящему изобретению, далее, позволяет осуществлять то, что после локального (например, в ткань, полости тела или промежуточные пространства между тканями) или после системного, предпочтительно внутривенного или внутриартериального введения, преобладающе, если не исключительно, в гладкомышечные клетки, активированные пролиферирующие эндотелиальные клетки, активированные лимфоциты или активированные макрофаги экспрессируют антитромботическое вещество, и оно, таким образом, высвобождается в месте возникновения тромбоза.
Так как активное вещество как за счет его специфичности к клетке, так и также за счет его специфичности к клеточному циклу гарантирует высокую степень безопасности, его также можно использовать в высоких дозировках и в случае необходимости многократно с интервалами от дней или недель для профилактики или терапии закупорки сосудов, вызываемой за счет пролиферирующих гладкомышечных клеток, и/или для профилактики и/или терапии тромбозов.
Пояснение к фигурам 1-11
Фиг.1:
Нуклеотидная последовательность cdc25C-промоторного участка с найденными ин виво сайтами связывания протеина (геномный диметилсульфатный футпринтинг; закрашенный кружок - полная конститутивная защита; незакрашенный кружок - частичная конститутивная защита; звездочка - регулируемая клеточным циклом Gl-специфическая защита; CBS - конститутивный связывающий сайт; CDE - зависимый от клеточного цикла элемент. Окрашенные в серый цвет области указывают YC-блоки (NF-Y-сайты связывания). Стартовые участки маркированы закрашенными квадратами.
Фиг.2:
Дерепрессия сdс25С-промотора специфически в G0 путем мутации cdc.
Фиг.3:
Схематическое изображение регуляции сdс25С-энхансера за счет CDE.
Фиг.4:
G0/G1 - специфическая репрессия 8У40-энхансера за счет CDE.
Фиг.5:
Гомологии в CDE-CHR-области и расположенные у 5'-концов YC-блоки в cdc25C, циклине-А и cdc2-промоторах.
Фиг.6:
Химерная конструкция, состоящая из различных частей человеческого промотора миогенина (Myf-4), слитого на 3'-конце промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческой β-глюкуронидазы (полный кодирующий участок, положение ≤ 27 до ≥ 1982) в качестве эффектора (Oshima и др., PNAS USA, 84, 685 (1987)). Данные о расположении относятся к данным Salminen и др., J. Cell. Biol., 115, 905 (1991) для гена миогенина, соответственно к системе cdc25C, применяемой Lucibello и др., ЕМВО J., 14, 132 (1995).
Фиг.7:
Замена гомологичной сигнальной последовательности β-глюкуронидазы гетерологичной сигнальной последовательностью (человеческого иммуноглобулина). Данные о расположении сигнальной последовательности (MGWSCIILFLVATAT) иммуноглобулина (HuVH-cAMP) относятся к данным Riechmann и др., Nature, 332, 323 (1988).
Альтернатива: Встраивание сигнального пептида иммуноглобулина для лучшего внеклеточного выделения β-глюкуронидазы; см.(Б).
Фиг.8:
Химерная конструкция, состоящая из различных частей человеческого промотора миогенина (Myf-4), слитого на 3'-конце промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческой β-глюкуронидазы, внутреннего рибосомного аминоациального сайта в качестве регулирующей нуклеотидной последовательности и ДНК тканевого плазминогенного активатора. Данные о расположении относятся к таковым Salminen и др., J. Cell. Biol., 115, 905 (1991) для миогенина; Lucibello и др., ЕМВО J., 14, 132 (1995) для элемента CDE/CHR-Inr; Oshima и др. , PNAS USA, 84, 685 (1987) для β-глюкуронидазы; Riechmann и др., Nature, 332? 323 (1988) для сигнальной последовательности иммуноглобулина; Pelletier и Sonenberg, Nature, 334, 320 (1988) для внутреннего рибосомного аминоациального сайта полиовируса и Pennica и др., Nature, 301, 214 (1983) для человеческого тканевого плазминогенного активатора.
Фиг.9:
Химерная конструкция, состоящая из различных частей человеческого промотора эндотелина-1, слитого по 3'-концу промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческого тканевого плазминогенного активатора (полная кодирующая область; Pennica и др., Nature, 301, 214 (1983)) в качестве эффектора. Данные о расположении относятся к таковым Wilson и др., Mol. Cell. Biol., 10, 4854 (1990) для гена эндотелина-1, соответственно к используемой Lucibello и др., ЕМВО J., 14, 132 (1995), системе cdc25C.
Фиг.10:
Химерная конструкция, состоящая из различных частей человеческого промотора миогенина (Myf-4), слитого по 3'-концу промоторного модуля с CDE и CHR репрессорными элементами, а также ДНК человеческого тканевого плазминогенного активатора (полная кодирующая область) в качестве эффектора. Данные о расположении относятся к таковым Salminen и др., J. Cell. Biol., 115, 905 (1991) для гена миогенина, соответственно к используемой Lucibello и др., ЕМВО J. , 14, 132 (1995) системе cdc25C и к данным Pennica и др., Nature, 301, 214 (1983) для тканевого плазминогенного активатора.
Фиг.11:
Химерная конструкция с двумя эффекторными генами.
Данные о расположении для ингибитора метаболического пути тканевого фактора относятся к данным
*) Wun и др., J. Biol. Chem., 263, 6001 (1988);
**) Girard и др., Thromb. Res., 55, 37 (1989);
а данные для внутреннего рибосомного аминоациального сайта (IRES) кДНК основываются на таковых Pelletier и Sonenberg, Nature, 334, 320 (1988).
Изобретение относится к генной терапии и касается генной терапии заболеваний сосудов с помощью специфического к клетке и зависимого от клеточного цикла активного вещества. Сущность изобретения включает ДНК-конструкцию, существенными элементами которой являются активаторная последовательность, промоторный модуль и ген активного вещества. Активаторная последовательность активируется специфически к клетке в гладкомышечных клетках, активированных эндотелиальных клетках, активированных макрофагах или активированных лимфоцитах. Это активирование регулируется специфически к клеточному циклу промоторным модулем. Активное вещество представляет собой ингибитор роста гладкомышечных клеток и/или свертывания (крови). Описанную ДНК-последовательность вставляют в вирусный или невирусный вектор, дополненный лигандом со сродством к клетке-мишени. Преимущество изобретения в том, что активное вещество действует только на находящиеся в стадии деления гладкомышечные клетки, подавляя их пролиферацию после повреждения сосудов. 33 з.п. ф-лы, 11 ил., 1 табл.
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| Экономайзер | 0 |
|
SU94A1 |
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |

