Изобретение относится к нуклеиново-кислотному конструкту для экспрессии активных веществ, которые могут быть активированы протеазами и к его получению и использованию.
Как и области воспаления, новообразования отличаются от окружающих нормальных тканей существенным увеличением в образовании и секрецией протеаз [Schmitt с соавт., Fibrinol. 6, 3 (1992), Cottam с соавт., Int. J. Oncol. 2, 861 (1993), Tryggvason с соавт., Anticancer Res. 12, 235 (1992), Hart, Fibrinol. 6, 11 (1992), Albini с соавт., J. Natl. Cancer Inst. 83, 735 (1991)]. Примерами этих протеаз являются активаторы плазминогена, катепсины и металлопротеиназы матрикса.
Существенная функция этих опухолевых протеаз заключается в растворении внеклеточного матрикса, чтобы позволить опухолевым клеткам вторгаться в инфильтрат и расти в нормальной ткани способом инфильтрации. Вместе с тем, эти протеазы защищают соответствующее новобразование от защитных механизмов данного организма таким образом, чтобы определенные активные соединения, необходимые для защиты, расщеплялись и, тем самым, инактивировались этими протеазами, которые образовала опухоль. Так, например, инактивируются опухолевыми протеазами антитела, цитокины и ростовые факторы, факторы комплемента, факторы коагуляции и медиаторы.
В недавнем прошлом цель, поэтому, состояла в том, чтобы подавить инфильтрирующий рост и метастатический рост опухолей и инактивировать соответствующие защитные механизмы данного организма путем ингибирования протеаз опухолевой клетки [Hocman, Int. J. Biochem. 24, 1365 (1992), Troll с соавт., JNCI 73, 1245 (1984), Ray с соавт., Eur. Respir 7, 2062 (1994), Коор с соавт. , Cancer Res. 54, 4791 (1994), Chiriri с соавт., Int. J. Cancer 58, 460 (1994), Denhardt с соавт., 59, 329 (1993), Melchiori с соавт., Cancer Res. 52, 2353 (1992)]. Однако отчасти по стехиометрическим и фармакокинетическим причинам ранее достигнутый некоторый успех в подавлении протеаз опухолевой клетки оказался небольшим.
Поэтому была предпринята попытка применить соответствующие протеазы опухолевой клетки для активации бактериальных токсинов, таких как α-гемолизин Staphylococcus aureus [Panchal с соавт., Nature Biotechn. 14, 852 (1996)] . Для этого, аминокислотную последовательность, т.е. ХХ-Аrg-Х, инсерцировали в положения 129-132 α-гемолизина и таким способом получали неактивные мутанты, которые только расщепляли и, таким образом, активировали с помощью опухолевых протеаз, таких как катепсин В.
На основании этих результатов были предложены проиммунолизины [Panchal с соавт. , Nature Biotechn. 14, 852 (1996)], проиммунолизины, которые включают антитело, которое присоединяется к α-гемолизину Staphilococcus aureus, который может быть активирован опухолевыми протеазами, или к эквинатоксину II морского анемона, когда соответствующее антитело, определяет специфичность данной клетки-мишени, присоединяющей продукт.
Однако эта предложенная концепция страдает следующими недостатками при ее использовании в терапии рака:
Прежде всего, эти авторы выбрали ксеногенные неэндогенные лизины и/или токсины, которые являются иммуногенными для хозяйского организма (пациента) и как результат те индуцируют иммунную реакцию в данном организме хозяина, чья иммунная реакция нейтрализует и инактивирует конъюгат антитело/токсин.
Во-вторых, известно [Sedlacek с соавт., Antibodies as Carriers of Cytotoxicity, Contrib. to Oncol. 43, Karger Verlag, Munich, 1992], что вследствие их молекулярного размера и реологических условий относительно опухоли опухолеспецифичные антитела и иммунотоксины накапливаются лишь в очень небольших количествах 0,01 - 0,001% от данного антитела или иммунотоксина/г опухоль на опухоли и проникают в опухоль в неполной степени так, чтобы не разрушить все клетки данного новобразования либо разрушить лишь небольшую долю клеток этого новобразования. Кроме того, соответствующий объем раковых антигенов, против которых направлено данное антитело, обычно экспрессируются по-разному среди клеток данного отдельного новобразования, и непостоянен, антиген-негативные раковые клетки легко ускользают от атаки антителами или иммунотоксинами. В дополнение к этому, антигены, которые секретируются клетками данного новобразования, нейтрализуются соответствующими антителами на периферии этого новобразования (Sedlacek с соавт., Monoclonal Antibodies in Tumory Therapy, Contrib. to Oncol., Karger Verlag, 1988).
Поэтому все еще существует большая необходимость в специфичной к клетке-мишени терапии опухолей и воспалений. И поэтому объектом настоящего изобретения является создание активного соединения против новобразований и воспалений, когда активное соединение не проявляет указанных недостатков. Следовательно, настоящее изобретение относится к новому методу, который использует секрецию ферментов в новобразованиях или участках воспаления для осуществления локального высвобождения активных соединений, чьи неактивные предшественники экспрессируются в раковых клетках, клетках, ассоциированных с раком или воспалительными клетками.
Поэтому первая часть содержания настоящего изобретения представляет собой нуклеиново-кислотный конструкт для экспрессии активного вещества, которое активируется с помощью фермента, который высвобождается из клеток млекопитающего, активное вещество которого включает следующие компоненты:
a) как минимум, один промоторный элемент,
b) как минимум, одну последовательность ДНК, которая кодирует активное соединение (протеин В),
c) как минимум, одну последовательность ДНК, которая кодирует аминокислотную последовательность (структурная часть С), которую можно специфически расщепить с помощью фермента, который высвобождается из клетки млекопитающего, и
d) как минимум, одну последовательность ДНК, которая кодирует пептид или протеин (структурную часть D), который связывается с данным активным компонентом (протеин В) через расщепляемую аминокислотную последовательность (структурная часть С) и подавляет эту активность данного активного соединения (протеин В).
В своей простейшей форме эти индивидуальные компоненты выстроены, например, как показано на Фиг. 1. В этом случае экспрессия протеина BCD, кодированного с помощью компонентов b), с) и d), индуцируется в результате активации данной промоторной последовательности [компонент а)]. Данная аминокислотная последовательность С экспрессированного продукта расщепляется затем клеточными ферментами, например, протеазами, в результате из которого высвобождается протеин В, который составляет данное активное соединение. В рамках содержания настоящего изобретения, протеазы или ферменты следует понимать, как представляющие одну или несколько протеаз или ферментов.
В другом варианте осуществления настоящего изобретения указанный фермент представляет собой протеазу, в частности активатор плазминогена, катепсин или металлопротеиназу матрикса. Указанные клетки млекопитающего представляют собой предпочтительно раковые клетки, лейкозные клетки, эндотелиальные клетки, макрофаги, лимфоциты, мышечные клетки, эпителиальные клетки, глиальные клетки, синовиальные клетки или клетки, инфицированные вирусом.
Ферменты предпочтительно высвобождаются в организме с помощью новобразований или раковых клеток, а также с помощью клеток, которые вовлечены в воспалительный процесс [Barrett с соавт., Mammalian Proteases, Academic Press, London 1980; Sedlacek и Möröy, Immune Reactions, Springer Verlag, 1995].
В соответствии с настоящим изобретением компонент с), следовательно, выбирали так, чтобы соответствующий экспрессируемый белок, например BCD, расщеплялся предпочтительно по своей структурной части С с помощью протеаз, которые синтезировались в новообразованиях или секретировались с помощью раковых клеток или воспалительных клеток. Примерами этих протеаз являются активаторы плазминогена, такие как активатор плазминогена урокиназного типа или тканевой активатор плазминогена; катепсины, такие как катепсин В, катепсин D, катепсин L, катепсин Е или катепсин Н, или их предшественники (прокатепсины); металлопротеиназы матрикса (ММР), такие как коллагеназы, например из групп I, II, III, IV или V; стромелизин 1, стромелизин 2 или стромелизин 3; метрилизины; желатиназы, такие как желатиназа А (ММР 2), прожелатиназа В (ММР 9) и прожелатиназа A [Pappot с соавт., Lung Cancer 12, 1 (1995), Schmitt с соавт. , Fibrinolysis 614, 3 (1992), Monsky с соавт., Cancer Biol. 4, 251 (1993), Rochefort с соавт., Medicine/Sciences 7, 30 (1991), Као с соавт. , 46, 1349 (1986), Fridman с соавт., Cancer Res. 55, 2548 (1995), Ray с соавт., Eur. Respir. J. 7, 2062 (1994), Cottam с соавт., Int. J. Oncol. 2, 861 (1993), Tryggvason с соавт., Breast Cancer Res. and Tratm. 24, 209 (1993)]; протеазы поверхности раковой клетки [поверхностно-экспрессируемые протеазы = сепраза; Monsky с соавт., Cancer Res. 54, 5702 (1994)] ; эластаза [Као с соавт., Cancer Res. 46, 1355 (1986)]; простат-специфичный антиген [Lundwall, Biochem Biophys. Res. Commun. 161, 1151 (1989), Riegman с соавт. , Biochem Biophys. Res. Commun. 159, 95 (1989)] или панкреатические трипсиногены [Miszuk-Jamska с соавт. , FEBS Lett. 294, 175 (1991)].
В соответствии со следующим вариантом настоящего изобретения соответствующая нуклеотидная последовательность для компонента b) может быть удлинена путем добавления компонента b'). Этот компонент b') кодирует лиганд (структурная часть В'), который может связывать соответствующее активное соединение с мишенной структурой. Компонент b') выстраивали, например, как показано на Фиг. 2. Экспрессия определенного нуклеиново-кислотного конструкта, корреспонидирующая с белком Фиг. 2, т.е. В'BCD, который связывается с мишенированной структурой через данный лиганд (структурная часть В'). Затем данная структурная часть С расщепляется с помощью клеточных протеаз, высвобождая таким образом данное активное соединение, т.е. белок В'В.
В отдельном варианте осуществления настоящего изобретения указанный белок В и структурная часть D являются частями соответствующих естественных предшественников белковых активных соединений с определенной естественной расщепляемой последовательностью, которая соединяет эти структурные части В и D, будучи замещенной на структурную частью С; в частности, указанная структурная часть D представляет собой соответствующую структурную часть естественного предшественника белкового активного соединения.
Новые нуклеиново-кислотные конструкты составляли предпочтительно из ДНК. Данный термин "нуклеиново-кислотные конструкты" следует понимать как означающий искусственные нуклеиново-кислотные структуры, которые могут быть траснкрибированы в соответствующих клетках-мишенях. Они предпочтительно инсерцированы в вектор, в плазмидные векторы или вирусные векторы, являющиеся особенно предпочтительными.
В зависимости от выбранного промоторного элемента [компонент а)] эти новые нуклеиново-кислотные конструкты экспрессируют структурный ген [компонент b) + с) + d) или b') + b) + с) + d)] либо неспецифично, клеточно-специфично, вирус-специфично, в конкретных метаболических условиях, специфично от клеточного цикла, либо в присутствии тетрациклина. По меньшей мере, два идентичных или различных промоторных элемента могут быть также объединены воедино с целью модификации экспрессии этого структурного гена в зависимости от выбора этих промоторных элементов. Компонент а) предпочтительно активирован в эндотелиальных клетках, в клетках, примыкающих к активированным эндотелиальным клеткам, в мышечных клетках, в лейкозных клетках, в раковых клетках, в глиальных клетках, в лимфоцитах, в макрофагах и/или в синовиальных клетках.
Структурная часть В (протеин В) белка, кодируемого новыми структурными составляющими данного гена, действительно нового активного данного соединения, который высвобожден или активирован путем расщепления этой структурной части С и, таким образом, преобразован из неингибированного состояния, например, в виде протеина BCD или в виде протеина B'BCD, в активное состояние, например, в виде протеина В или в виде протеина В'В.
В соответствии с настоящим изобретением это активное соединение может быть ферментом, который активирует или ингибирует биологический активированный каскад и/или представляет собой активный компонент этого каскада. Примерами биологических активированных каскадов этого рода являются система коагуляции, которая может быть активирована или ингибирована, фибринолиз, который предпочтительно активирован, система комплемента, которая также предпочтительно активирована, или кининовая система, которая также предпочтительно активирована. Это активное соединение может быть также ферментом, который преобразует соответствующий неактивный предшественник фармакологического вещества в активное вещество или которое само по себе является фармакологически активным веществом. Особое предпочтение отдано активному соединению (протеин В), который представляет собой фактор коагуляции, который выбран из тромбина, фактора Va, фактора VIIa, фактора IXа, фактора Ха, TF-фрагментов, активирующих коагуляцию, или фактора XIIа; тромбина, который мутировал в области сайта расщепления Arg-Thr (аминокислотная позиция 327/328); фибринолитического белка, который выбран из урокиназы, tPA или его функциональных гибридов; фактора комплемента, который выбран из CVF, С3b или его функционально расщепленных продуктов; антитромботического белка, который выбран из протеина С, ингибитора С-1S, α1-антитрипсина, гирудина АТ-III, TFPI, PAI-2 или PAI-3; калликреина; цитостатического, цитотоксического или белка, вызывающего воспаление; антиангиогенного белка; иммуномодуляторного белка; противовоспалительного белка; белка, который ослабляет повреждение нервной системы; белка, который ингибирует или нейтрализует нейротоксический эффект TNFα; белка, стимулирующего развитие кровеносных сосудов; гипотензивного белка; противовирусного белка; цитокина; интерферона; фактора некроза опухоли; онкостатина М или LIF; цитокинового рецептора; определенной составляющей цитокинового рецептора, которая находится снаружи на клетке; цитокинового антагониста; ростового фактора; рецептора ростового фактора; определенной составляющей рецептора ростового фактора, которая находится снаружи на клетке; хемокина; ангиостатина; фактора 4 тромбоцитов; TIMP 1, ТIMP 2 или ТIMP 3; нитроредуктазы; β-глюкуронидазы; карбоксипептидазы; β-лактамазы; цитозиндезаминазы; каталазы; пероксидазы; фосфатазы; оксидазы; калликреина или синтазы окиси азота эндотелиальной клетки.
Структурная часть В' белка, кодируемого этим новым структурным геном, составляет новый лиганд для связывания соответствующего активного соединения (протеин В) с мишенированной структурой. Предпочтительной мишенированной структурой является поверхность клеток, предпочтительно рецептор мембраны клетки, антиген мембраны клетки, адгезионная молекула, локализованная на мембране клетки или во внеклеточном матриксе, например, эндотелиальных клеток, в частности, активированных или пролиферирующих эндотелиальных клеток, раковых клеток, мышечных клеток, в частности клеток гладкой мускулатуры, фибробластов, макрофагов, клеток печени, клеток почек, синовиальных клеток, воспалительных клеток, клеток, инфицированных вирусом, бронхиальных эпителиальных клеток, глиальных клеток, лейкозных клеток или клеток других тканей и органов. Особенно предпочтительной мишенированной структурой является поверхность активированных и/или пролиферирующих эндотелиальных клеток.
Другая предпочтительная мишенированная структура составлена компонентами внеклеточного матрикса, например коллагенами [Prockop с соавт., Annu. Rev. Biochem. 64, 403 (1995), Wetzels с соавт., Am. J. Pathol. 139, 451 (1991)]; фиколином [Ichijo с соавт., J. Biol. Chem. 268, 14505 (1993)]; сиалопротеином [Bellahcene с соавт., Cancer Res. 54, 2823 (1994)]; ламинином [von der Mark с соавт., Biochem. Biophys. Acta 823, 147 (1985); Hunt. Expl. Cell Biol. 57. 165 (1989)]; протеогликанами [Schmidtchen с соавт., Biomed. Chromatography 7, 48 (1993)] или тенасцином [Oyama с соавт., Cancer Res. 51, 4853 (1991)].
Этот новый лиганд (структурная часть В') может быть, например, антителом или фрагментом антитела, таким как составляющая, связанная с определенным эпитопом антитела, Fab, Fv, одноцепочечным Fv или Fc, которые специфически связываются с антигеном мембраны клетки или с антигеном на внеклеточном матриксе, или другим пептидом или белком, которые связываются с рецептором на актуальной клеточной мембране. Они включают, например, ростовые факторы, цитокины, интерфероны, фактор некроза опухоли, хемокины, их рецептор-связывающие части последовательностей, пептидные гормоны, ангиотензин, кинин или фолиевую кислоту. Этот лиганд может быть также адгезионной молекулой или ее адгезионной последовательностью, которая связывается с корреспондирующей молекулой на данной клеточной мембране или на внеклеточном матриксе, или составляющей, связывающей клетку-мишень, внеклеточную составляющую Fc-рецептора, гликопротеин вируса, связывающий клетку-мишень, который обладает тропизмом для избранных клеток, или частью последовательности этого гликопротеина, который связывается с этими клетками, или пептидом, с чьей помощью данное активное соединение заякоривается в данной клеточной мембране данной клетки, которая экспрессируют его. Примерами этих заякоривающихся пептидов являются трансмембранные домены рецепторов или белки вирусов или гликофосфолипидные якоря.
Компонент d) кодирует пептид (структурная часть D), который связан с протеином В или протеином В'В через структурную часть С и, таким образом, ингибирует активность протеина В. Компонент d) может быть любой произвольной нуклеиново-кислотной последовательностью. Предпочтительно, однако, чтобы она была составлена из нуклеиново-кислотных последовательностей, которые кодируют эндогенные пептиды или белки, с целью избежать или снизить повреждающее действие иммунной реакции. В другом предпочтительном варианте осуществления настоящего изобретения компоненты b) и d) нового структурного гена кодируют эндогенные белки или пептиды.
Значительное количество белковых активных соединений встречается в природе в форме неактивных предшественников (протеин BSD). Предшественник этого типа активируется с помощью фермента, расщепляющего этот предшественник на структурную часть, которая составляет реальный белок активного соединения (протеин В), и на неактивную структурную часть (структурная часть D). Этот предшественник расщепляется, по меньшей мере, на одну определенную аминокислотную последовательность, например, на так называемую последовательность расщепления (структурная часть S).
Она представляет собой отдельный раздел содержания настоящего изобретения в том, что эта последовательность расщепления (структурная часть S), которая естественно встречается в предшественниках белковых активных соединений, замещается структурной частью С. Это замещение является эффективным для последовательности, кодирующей структурную часть S, будучи замещенной компонентом с), кодирующей структурную часть С, в соответствующей нуклеиново-кислотной последовательности, которая кодирует естественный предшественник (протеин BCD). После добавления компонентов а) и, где подходит, b') получали новый нуклеиново-кислотный конструкт, который включает, например, компоненты a)b')c)d) или a)b)c)d), данную структурную часть С, чей экспрессируемый продукт, т. е. протеин В'BCD или, соответственно, BCD, расщепляли с помощью протеаз образованными в новобразованиях или секретированными раковыми клетками или воспалительными клетками так, чтобы образовалось данное активное соединение, т.е. протеин В'В или В.
В другом варианте осуществления настоящего изобретения, этот новый конструкт включает, по меньшей мере, два идентичных или различных компонента b)c)d) и/или b')b)c)d), компоненты, которые соединены друг с другом через так называемый участок внутреннего входа рибосом (IRES).
Инсерцированный в невирусный вектор или вирусный вектор, этот новый нуклеиново-кислотный конструкт, предназначаемый, в целом, для профилактики и/или терапии расстройств, вводили местно или инъецировали в циркулирующую кровь. Эти нарушения, прежде всего, включают раковые заболевания и воспаления. Такие воспаления могли быть, например, инициированы в результате физико-химического повреждения, инфекции или иммунной реакции против эндогенной или чужеродной ткани.
Поэтому настоящее изобретение, кроме того, относится к определенному использованию нового нуклеиново-кислотного конструкта для получения лекарственного средства для локального или системного введения при профилактике и/или терапии новобразований, лейкемий, аллергий, аутоиммунных заболеваний, инфекций, воспалений, реакций отторжения трансплантата, тромбозов, окклюзии кровяных сосудов, коагуляции крови и нарушениях кровообращения, травмах тканей и/или поражения нервной системы.
Выбор соответствующих компонентов этого нового нуклеиново-кислотного конструкта зависит от определенного заболевания, которое лечат путем введения данного нуклеиново-кислотного конструкта, и может осуществляется следующим образом.
Промоторные последовательности [компонента а)]:
В соответствии с настоящим изобретением особое предпочтение отдано в одном случае промоторным последовательностям [компонент а)], которые представляют собой промоторы и активаторные последовательности, которые могут быть активированы неограничиваемым способом, таким как промотором РНК-полимеразы III, промотором РНК-полимеразы II и т.п., CMV-промотором и CMV-энхансером или SV40-промотором, а в другом случае вирусным промотором и активаторными последовательностями, такими как HBV, HCV, HSV, HPV, EBV, HTLV или HIV.
Например, в случае HIV-промотора полная LTR-последовательность, включающая TAR-последовательность [позиции ≤ -453 - ≥ -80, Rosen с соавт., Cell 41, 813 (1985)] , может быть использована в качестве вирус-специфического промотора.
Метаболически активируемые промоторная и энхансерная последовательности, такие как энхансер, индуцирующий гипоксию, промоторы, которые могут быть активированы специфическим образом в цикле клеточного деления, такие как промоторы гена cdc25C, гена циклин А, гена cdc2, гена Bmyb, гена DHFR или гена E2F-1, или промоторы, активируемые тетрациклином, такими как тетрациклиновый оператор в сочетании с подходящим репрессором, также являются особенно предпочтительными в качестве компонента а).
В соответствии с настоящим изобретением нуклеотидные последовательности, которые после связывания транскрипционных факторов активируют транскрипцию данного структурного гена, которые примыкают к ним по 3'-концу, также использовали в качестве промоторных последовательностей.
Кроме того, промоторы, которые могут быть специфически активированы в цикле клеточного деления, особенно предпочтительны в качестве компонента а). Эти промоторы предпочтительно включают последовательности промоторов или активатора, составленные из промоторов или энхансеров из тех генов, которые предпочтительно кодируют белки в выбранных клетках. Например, промоторы для следующих белков являются предпочтительными для использования в нижеследующих клетках.
Последовательности промотора и активатора, которые активируются в эндотелиальных клетках, такие как мозг-специфичные, эндотелиального глюкозо-I транспортера, эн-дольгина, VEGF-рецептора 2 (flt-1), VEGF-рецептора 2 (flk-1, KDR), tiel-1 или tiel-2, В61-рецептора (Eck-рецептор), B61, эндотелина, особенно эндотелина В и эндотелина 1, рецепторов эндотелина, в частности рецептора эндотелина В, рецепторов маннозо-6-фосфат, фактора von Willebrand, IL-1α, Il-1β, IL-1-рецептора, молекулы межклеточной адгезии (VCAM 1) или синтетические активаторные последовательности.
В качестве альтернативы естественным промоторам, специфичным к эндотелиальным клеткам, можно также использовать созданные синтетические активаторные последовательности, которые составлены из олигомеризованных сайтов связывания для факторов транскрипции, которые предпочтительно или селективно активны в эндотелиальных клетках. Примером является фактор транскрипции GATA 2, чей сайт связывания в гене эндотелин 1 представляет собой 5'-ТТАТСТ-3' [Lee с соавт. , Biol. Chem. 226, 16188 (1991), Dormann с соавт., J. Biol. Chem. 267, 1279 (1992) и Wilson с соавт., Mol. Cell. Biol. 10, 4854 (1990)].
Промоторные или активаторная последовательности, которые активируются в клетках вблизи активированных эндотелиальных клеток, в частности в клетках гладкой мускулатуры, представлены, например, в VEGF-гене. Ген-регуляторные последовательности для гена VEGF представляют собой 5'-фланкирующую область, 3'-фланкирующую область, ген c-Src или ген v-Src.
Рецепторы стероидных гормонов и их промоторные элементы, в частности промотор вируса молочной железы мыши, или промоторные элементы гена, кодирующего тропомиозин, α-актин, α-миозин, рецептор для PDGF, рецептор для FGF, MRF-4, фосфофруктокиназу А, фосфоглицератмутазу, тропонин С, миогены, рецепторы для эндотелина А, десмина или отдельные "искусственные" промоторы, также являются подходящими. Промоторные элементы, с которыми факторы спираль-петля-спираль (HLH) семейства миогенов MyoD, Myf 5 и MRF4 [обзор Olson и Klein, Genes Dev. 8, 1 (1994)] могут связываться в качестве транскрипционных факторов, специфичных для мышц, также является подходящими. Эти транскрипционные факторы, специфичные для мышц, включают также белок типа цинкового пальца GATA-4 [Arceci с соавт., Mol. Cell Biol. 13, 2235 (1993), Ip с соавт. , Mol. Cell Biol. 14 7517 (1994)] и группы транскрипционных факторов MEF [Yu с соавт., Gene Dev. 6, 1783 (1992)].
HLH-белки, а также GATA 4, проявляют сходную транскрипцию, специфичную для мышц, не только с промоторами из мышечно-специфичных генов, но также в ситуации с разнородными элементами, которые представлены "искусственными" промоторами. Примерами таких искусственных промоторов являются множественные копии (ДНК) связывающего сайта мышеч-но-специфичных HLH-белков, таких как Е-блок (myo D), например, 4 х AGCAGGTGTTGGGAGGC [Weintraub с соавт., PNAS 87, 5623 (1990) или множественные копии ДНК-связывающего сайта для GATA 4 гена тяжелой цепи α-миозина, например, 5'-GGCCGATGGGCAGATAGAGGGGGCCGATGGGCAGATAGAGG3' [Molkentin с соавт., Mol. Cell Biol. 14, 4947 (1994)].
Примерами последовательностей промоторов и активатора, которые активируются в лейкозных клетках, являются промоторы для c-myc, HSP-70, bcl-1/cyclin D-1, bcl-2, IL-6, IL-10, TNFα, TNFβ, HOX-11, BCR-Ab1, E2A-PBX-1 или PML-RATA.
Примерами промоторных или активаторной последовательностей, которые активируются в раковых клетках, являются промоторная или активаторная последовательности, которые взаимодействуют с транскрипционными факторами, которые образуются или активируются в раковых клетках. Эти предпочтительные промоторная или активаторная последовательности включают последовательности гена-регулятора или элементы генов, которые кодируют белки, которые образуются, в частности, в клетках карциномы или клетках саркомы. Так, например, промотор белка N-CAM использовали в случае мелкоклеточных бронхиальных карцином, промотор рецептора фактора роста гепатита или L-пластин использовали в случае овариальных карцином, а промотор L-пластина или полиморфного эпителиального муцина (РЕМ) использовали в случае панкреатических карцином.
Последовательности промоторов и активатора, которые активируются в глиальных клетках, представляют собой, в частности, ген-регуляторные последовательности или элементы генов, которые кодируют, например, следующие белки: периаксин, белок, специфичный для Шванновской клетки, глутамин-синтетазу, белок, специфичный для глиальной клетки (глиальный фибриллярный кислый белок = GFAP), белок глиальной клетки S100b, IL-6 (CNTF), рецепторы 5-НТ, TNFα, IL-10, рецептор инсулиноподобного фактора роста I и II или VEGF. Ген-регуляторные последовательности для VEGF-гена уже перечислялись выше.
Примерами последовательностей промоторов и активатора, которые активируются в лимфоцитах и/или макрофагах, являются последовательности промотора и активатора гена, кодирующего цитокины, цитокиновые рецепторы и адгезионные молекулы, и рецепторы для Fc-фрагмента антител. Примерами последних являются: рецептор IL-1, IL-1α, IL-1β, рецептор IL-2, IL-3, рецептор IL-3 (α-субъединица), рецептор IL-3 (β-субъединица), IL-4, рецептор IL-4, IL-5, IL-6, рецептор IL-6, регуляторный фактор 1 интерферона (IRF-1), (промотор IRF-1 активируется с помощью IL-6 в той же мере, как и с помощью IFNγ или IFNβ), IFNγ-респонсивный промотор, IL-7, IL-8, IL-10, IL-11, IFNγ, GM-CSF, рецептор GM-CSF (α-цепь), IL-13, LIF, рецептор макрофагального колониестимулирующего фактора (M-CSF), рецепторы сапрофитного макрофага типа I и II, МАС-1 (лейкоцитарный функционирующий антиген), LFA-1α (лейкоцитарный функционирующий антиген) или р150,95 (лейкоцитарный функционирующий антиген).
Примерами последовательностей промотора и активатора, которые активируются в синовиальных клетках, являются последовательности промотора для металлопротеиназ матрикса (ММР), например для: ММР-1 (интерстициальная коллагеназа), или ММР-3 (стромелизин/транзин). Последние также включают промоторные последовательности для тканевых ингибиторов металлопротеиназ (TIMP), например, TIMP-1, TIMP-2 и TIMP-3.
В соответствии с настоящим изобретением некоторые из промоторных последовательностей, которые были перечислены в качестве примера, можно объединять друг с другом для осуществления наивысшей специфичности клетки-мишени в экспрессии соответствующего нового нуклеиново-кислотного конструкта. Можно также объединить два идентичных промотора. Можно объединить несколько промоторных последовательностей, используя химерные промоторы или гибридные промоторы. Химерный промотор представляет собой объединение вышележащей активаторной последовательности, которая может быть активирована клеточно-специфически, метаболически или вирус-специфически, с нижележащим промоторным элементом, который связывает транскрипционные факторы семейств CDF и CHF или семейств E2F и СНF и может, таким образом, подавлять активацию вышележащей активаторной последовательности в фазах G0 и G1 данного клеточного цикла (Lucibello с соавт., ЕМВО J. 14, 132 (1994)].
Для случая гибридных промоторов, когда ТАТА-бокс промотора является, например, мутантным, эта мутация является компенсаторной по соответствующей мутации в данном гене для белка, связывающего ТАТА, и этот ТАТА-связывающий белок находится под контролем другого промотора.
Нуклеиново-кислотная последовательность[компонент b'], которая кодирует лиганд (структурная часть В'):
В соответствии с настоящим изобретением данный лиганд представляет собой вещество, которое связывает мембранный антиген с рецептором или с адгезионной молекулой на клетке-мишени или которое интегрируется в клеточную мембрану и/или связывается с внеклеточным матриксом. Обзоры по важным цитокинам и ростовым факторам и их рецепторам, адгезионным молекулам и белкам внеклеточного матрикса написали Ayad с соавт., The Extracellular Matrix, Academic Press 1994; Callard с соавт., The Cytokine, Academic Press 1994; Pigott с соавт., The Adhesion Molecule, Academic Press 1994 и Barclay с соавт., The Leukocyte Antigen, Academic Press 1994.
Примерами веществ, которые связываются с рецепторами, являются ростовые факторы, такие как VEGF, PDGF, EGF, TGFα, TGFβ, KGF, SDGF, FGF, IGF, HGF, NGF, BDNF, нейротрофины, BMF, бомбезин, M-CSF, тромбопоэтин, эритропоэтин, SCF, SDGF, онкостатин, PDEGF или эндотелин-1, цитокины, такие как IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-13, IL-14, IL-15, интерфероны α, β и γ, факторы некроза опухолей TNFα и TNFβ, хемокины, такие как RANTES, MCAF, MIP-1α, или MIP-1β, NAP или β-тромбоглобулин, пептидные гормоны, такие как SRH, SIH или STH, MRH или MSH, PRH, PIH или пролактин, LH-RH, FSH-RH, LH/ICSH или FSH, TRH или TSH, CRH или АСТН, ангиотензин, кинины, их гомологи и аналоги, или витамины, такие как фолиевая кислота.
В соответствии с настоящим изобретением данный лиганд может быть также адгезионной молекулой, частью адгезионной молекулы или аналогом адгезионной молекулы, который связывается с соответствующей адгезионной молекулой, которая локализована в данной клеточной мембране или с другой связывающейся структурой, специфической для адгезионной молекулы на клетке-мишени или во внеклеточном матриксе.
Примерами таких адгезионных молекул, которые обладают способностью функционировать в качестве лигандов, являются Lewis Х (для GMP-140), S Lewis Х (для ELAM-1), LFA-1 (для ICAM-1 и ICAM-2), МАС-1 (для ICAM-1), VLA-4 (для VCAM-1), РЕСАМ (для РЕСАМ), витронектин (для соответствующего витронектинового рецептора), GMP-140 (для Lewis X), S Lewis X (для ELAM-1), ICAM-1, ICAM-2 (для LFA-1 и MAC-1), VCAM-1 (для VLA-4), фибронектин (для VLA-4), ламинин (для VLA-6), ламинин (для VLA-1, VLA-2 и VLA-3), фибриноген (для GPIIb-IIIa), B7 (для CD28), CD28 (для В7), CD40 (для CD40L) или CD40L (для CD40).
В соответствии с настоящим изобретением этим лигандом может быть также внеклеточная составляющая Fc-рецептора [Dougherty с соавт., Transfusion Science 17, 121 (1996)]. Кроме того, этим лигандом может быть также молекула антитела или эпитоп-связывающая составляющей молекулы антитела. Соответствующие мышиные моноклональные антитела должны предпочтительно использоваться в форме, приближенной к человеческим. Способ эффективной гуманизации описан у Winter с соавт. , Nature 349, 293 (1991) и Hoogen-booms с соавт., Rev. Tr. Transfus. Hemobiol. 36, 19 (1993).
Фрагменты рекомбинантного антитела получают либо непосредственно из существующих гибридом, либо выделяют из библиотек фрагментов мышиного или человеческого антитела [Winter с соавт., Annu. Rev. Immunol. 12, 433 (1994)], используя метод фаговой индикации [Smith, Science 228, 1315 (1985)]. Фрагменты соответствующего антитела используют затем непосредственно, на генетическом уровне, для последующих манипуляций, например для слияния с другими белками.
С целью получения фрагментов рекомбинантного антитела из гибридом генетическая информация, которая кодирует соответствующие антиген-связывающие домены (VH и VL) данных антител, получали путем выделения мРНК, обратного транскрибирования РНК в кДНК, и последующей амплификации этой кДНК при помощи полимеразной цепной реакции [Saiki с соавт., Science 230, 1350 (1985)] и использования олигонуклеотидов, которые комплементарны по 5' и 3'-концам вариабельных фрагментов (Orlandi с соавт., 1989). Фрагменты VH и VL клонировали затем в бактериальные экспрессионные векторы, например, в форме Fv-фрагментов [Skerra & Plückthun, Science 240, 1038 (1988)], одноцепочечных Fv-фрагментов (scFv) [Bird с соавт., Science 242, 423 (1988), Huston с соавт., PNAS-USA 85, 5879 (1988)] или в виде Fab-фрагментов [Better с соавт., Science 240, 1041 (1988)].
Метод фаговой индикации можно также использовать для выделения новых антительных фрагментов из антительных библиотек (иммунных библиотек или первичных библиотек) мышиного или человеческого происхождения. Для фагиндицированных антительных фрагментов клонировали соответствующие антиген-связывающие домены, в виде слитых белков, покрытых белком g3P из филаментов бактериофага, либо в этот фаговый геном [McCafferty с соавт., Nature 348, 552 (1990)] или в фагемидные векторы [Breitling с соавт., Gene 104, 147 (1991)] в форме scFv-фрагментов [McCafferty с соавт., Nature 348, 552 91990)] или в виде Fab-фрагментов [Hoogenboom с соавт., Nucl. Acid Res. 19, 4133 (1991), Barbas с соавт., PNAS-USA 88, 7978 (1991)]. Антиген-связывающие фаги отбирали на пластиковых сосудах, нагруженных антигеном (промытых) [Marks с соавт. , J. Mol. Biol. 222, 581 (1991)], на конъюгированных с антигеном парамагнитных шариках [Hawkins с соавт., J. Mol. Biol. 226, 889 (1992)] или путем связывания с клеточными поверхностями [Marks с соавт., Bio/Technol. 11, 1145 (1993)].
Иммунные библиотеки получали, подвергая вариабельные антительные фрагменты из В-лимфоцитов иммунизированных животных [Sastry с соавт., PNAS-USA 86, 5728 (1989), Ward с соавт., Nature 341, 544 (1989), Clackson с соавт., Nature 352, 624 (1991)] или пациентов [Mullinax с соавт., PNAS-USA, 87, 8095 (1990), Barbas с соавт., PNAS-USA, 88, 7978 (1991)] PCR-амплификации. Для этого использовали созданные комбинации олигонуклеотидов, которые являются специфичными для мышей [Orlandi с соавт., PNAS-USA, 86, 3833 (1989), Sastry с соавт., PNAS-USA, 86, 5728 (1989)], или для иммуноглобулиновых генов человека [Larrick с соавт., BBRC 160, 1250 (1989)], или для иммуноглобулиновых генных семейств человека [Marks с соавт., Eur. J. Immunol. 21, 985 (1991)].
Первичные библиотеки могут быть получены, например, используя неиммунизированных доноров в качестве источника иммуноглобулиновых генов [Marks с соавт. , J. Mol. Biol. 222, 581 (1991)]. Альтернативно эмбриональная линия иммуноглобулиновых генов может быть получена, используя для получения наборы полусинтетических антител, с определяющей комплементарность областью 3 вариабельных фрагментов, амплифицированными с помощью PCR, использующей вырожденные праймеры [Hoogenboom & Winter, J. Mol. biol. 227, 381 (1992), Barbas с соавт. , PNAS-USA, 89, 4457 (1992), Nissim с соавт., ЕМВО J. 13, 692 (1994), Griffiths с соавт. , EMBO J. 13, 3245 (1994)]. По сравнению с иммунными библиотеками эти так называемые single-pot библиотеки обладают тем преимуществом, что антительные фрагменты против большого числа антигенов могут быть выделены из одной единственной библиотеки [Nissim с соавт., EMBO J. 13, 692 (1994)].
Более того, метод фаговой индикации может быть использован для увеличения сродства антительных фрагментов с новыми библиотеками, которые получили из уже существующих антительных фрагментов путем случайного [Hawkins с соавт. , J. Mol. Biol. 226, 889 (1992), Gram с соавт., PNAS-USA, 89, 3576 (1992)] , основанного на кодоне [Glaser с соавт., J. Immunol. 149, 3903 (1992)] или сайт-направленного мутагенеза [Balint & Larrick, Gene 137, 109 (1993)] , путем перестановки цепей из индивидуальных доменов на фрагменты из первичных наборов [Marks с соавт., Bio/Technol. 10, 779 (1992)] или путем использования бактериальных штаммов-мутаторов [Low с соавт., J. Mol. Biol. 26, 359 (1996)] , и антительных фрагментов, обладающих улучшенными свойствами, которые выделяли путем повторного отбора в строгих условиях [Hawkins с соавт. , J. Mol. Biol. 226, 889 (1992)]. Кроме того, мышиные антительные фрагменты могут быть приближены к человеческим путем постепенного замещения одного из вариабельных доменов на человеческие наборы, а затем отселектированы с исходным антигеном (управляемая селекция) [Jespers с соавт. , Bio/Technol., 12, 889 (1994)]. Альтернативно мышиные антитела гуманизировали путем специфического замещения гипервариабельных областей человеческих антител на корреспондирующие области исходного мышиного антитела [Jones с соавт., Nature 321, 522 (1987)].
В соответствии с настоящим изобретением этим лигандом может быть также нуклеотидная последовательность, кодирующая оболочечный белок или часть оболочечного белка, из вирусов, которые специфически связываются с отобранными клетками через свой оболочечный белок.
Этим лигандом может быть также пептид, с чьей помощью данное активное соединение (протеин В) заякоривается в клеточную мембрану экспрессирующих клеток. Эти якорные пептиды включают соответствующие трансмембранные домены рецепторов, локализованных на клеточной мембране, или вирусные белки, такие как трансмембранная последовательность человеческого макрофагального колониестимулирующего фактора [ДНК позиция ≤ 1485 - ≥ 1554; Cosman с соавт., Behring Inst. Mitt. 83, 15 (1988)] или ДНК-последовательность для сигнальной и трансмембранной областей гликопротеина G человеческого респираторно-синцитиального вируса (RSV) [аминокислоты 1-63 или части их последовательностей, аминокислоты 38-63; Vijaya с соавт., Mol. Cell Biol. 8, 1709 (1988); Lichtenstein с соавт., J. Gen. Virol. 77, 109 (1996)] или ДНК-последовательность для сигнальной и трансмембранной области нейраминидазы вируса гриппа [аминокислоты 7-35 или соответствующая часть последовательности аминокислот 7-27, Brown с соавт., J. Virol. 62, 3824 (1988)].
Тем не менее, соответствующую нуклеотидную последовательность для гликофосфолипидного якоря [обзор по гликофосфолипидным заякоренным мембранным белкам у Ferguson с соавт., (Ann. Rev. Biochem. 57, 285 (1988))] можно также инсерцировать с целью заякорить данное активное соединение в данную клеточную мембрану преобразованных клеток, которые образуют данное активное соединение. Гликофосфолипидные якоря описаны, например, для СЕА [ДНК позиция <893 - > 1079; Berling с соавт., Cancer Res. 50, 6534 (1990)], для N-CAM [Cunningham с соавт.. Science 236, 799 (1987)] и для других мембранных белков, таких как Thy-1 [Clissold, Biochem. J. 281, 129 (1992)] или CD16 [Selvaray с соавт., Nature 333, 565 (1988)].
Определенный выбор соответствующего лиганда зависит, прежде всего, от клетки-мишени, которая преобразована соответствующим нуклеиново-кислотным конструктом. Лиганды активированных эндотелиальных клеток этому примеры. В рамках содержания данного изобретения эти лиганды включают антитела или антительные фрагменты, которые направлены против мембранных структур эндотелиальных клеток, как было описано, например, у Burrows с соавт., Pharmac. Ther. 64, 155 (1994), Hughes с соавт., Cancer Res. 49, 6214 (1989) и Maruyama с соавт., PNAS-USA 87, 5744 (1990). В частности, эти антитела включают антитела против актина, рецепторов ангиотензина II, антитела против рецепторов ростовых факторов, таких как VEGF, FGF, PDGF или EGF, и антитела против адгезионных молекул, например против рецептора виронектина или ICAM 3.
Кроме того, эти лиганды включают все активные соединения, которые связываются с мембранными структурами или мембранными рецепторами на эндотелиальных клетках. Примерами последних являются IL-1 или ростовые факторы, или их фрагменты или части их последовательностей, которые связываются с рецепторами, которые экспрессируются в эндотелиальных клетках, например PDGF, bFGF, VEGF или TGFβ [Pusztain с соавт., J. Pathol. 169, 191 (1993)].
Эти лиганды, кроме того, включают адгезионные молекулы, которые связываются с активированными и/или пролиферирующими эндотелиальными клетками. Адгезионные молекулы этого рода, такие как Slex, LFA-1, MAC-1, LECAM-1, VLA-4 или виронектин, уже были описаны [Augustin-Voss с соавт., J. Cell Biol. 119, 483 (1992), Pauli с соавт., Cancer Metast. Rev. 9, 175 (1990), Honn с соавт. , Cancer Metast. Rev. 11, 353 (1992), Pigott с соавт., The Adhesion Molecule, Academic Press (1994)].
Эти лиганды в рамках содержания этого изобретения включают также, в частности, гликопротеины из оболочек вирусов, которые обладают тропизмом к эндотелиальным клеткам. Примерами этих вирусов являются филовирусы, такие как вирус Марбург со своими оболочечными белками GP (гликопротеин) и sGP (второй гликопротеин) или вирус Эбола, в каждом случае со своими оболочечными белками GP и sG, цитомегаловирус, особенно со своим gВ-белком, вирус простого герпеса типа I, вирус HIV-1, вирус кори, вирус Hantaan, альфавирусы, такие как вирус леса Семлики, вирус
эпидемической геморрагической лихорадки, полиовирус или энтеровирусы, такие как ECHO 9, ECHO 12 и Коксаки В3.
Антитела или антительные фрагменты, которые направлены против мембранных структур мышечных клеток, в частности клеток гладкой мускулатуры, являются примерами лигандов для мышечных клеток. Примерами антител этого рода являются антитело 10F3, антитела против актина, антитела против рецепторов ангиотензина II, антитела против рецепторов ростовых факторов или антител, например, против EGF-рецепторов, против PDGF-рецепторов или против FGF-рецепторов, или антитела против рецепторов эндотелина А.
Эти лиганды, кроме того, включают нуклеотидные последовательности для активных веществ, которые связываются с мембранными структурами или мембранными рецепторами на мышечных клетках [Pusztai с соавт., J. Pathol. 169, 191 (1993), Harris, Curr. Opin. Biotechnol. 2, 260 (1991)]. Примерами этих лигандов являются ростовые факторы или их фрагменты или части их последовательностей, которые связываются с рецепторами, которые экспрессируются в клетках гладкой мускулатуры, например PDGF, EGF, TGFβ, TGFα, FGF или эндотелин А.
Эти лиганды включают также гликопротеины из оболочек тех вирусов, которые обладают тропизмом к мышечным клеткам. Примером этих вирусов является цитомегаловирус [Speir с соавт., Science 265, 391 (1994)].
Примерами лигандов для активированных макрофагов и/или активированных лимфоцитов являются, кроме того, нуклеотидные последовательности, которые кодируют вещества, которые специфически связываются с поверхностью иммунных клеток. Эти вещества включают антитела или антительные фрагменты, которые направлены против мембранных структур иммунных клеток, как это было описано, например, у Powelson с соавт. , Biotech. Adv. 11, 725 (1993) и Barclay с соавт. , The Leucocyte Antigen, Academic Press (1994). Эти лиганды включают также моноклональные или поликлональные антитела или антительные фрагменты, которые связываются с помощью своей антиген-связывающей вариабельной составляющей с рецепторами Fcγ, Fcε или Fсμ иммунных клеток [Rojanasakul с соавт. , Pharm. Res. 11, 1731 (1994)]. Они, кроме того, включают Fc-фрагмент человеческого моноклонального или поликлонального иммуноглобулина.
Эти лиганды включают, кроме того, все вещества, которые связываются с мембранными рецепторами на поверхности иммунных клеток. Эти вещества включают цитокины, такие как IL-1, IL-2, IL-3, IL-4, IL-6, IL-10, TNFα, GM-CSF и M-CSF, а также ростовые факторы, такие как EGF, TGF, FGF, IGF или PDGF, или их фрагменты или части их последовательностей, которые связываются с рецепторами, которые экспрессируются в иммунных клетках [Gallard с соавт., The Cytokine, Academic Press (1994)]. Эти лиганды включают также адгезионные молекулы и другие лиганды, которые связываются с мембранными структурами клетки на макрофагах, и в селезенке, печени, легких и других тканях [Pigott с соавт. , The Adhesion Molecule, Academic Press (1994), Perales с соавт., Eur. J. Biochem. 226, 255 (1994)].
Эти лиганды в рамках содержания настоящего изобретения включают также гликопротеины из оболочек тех вирусов, которые обладают тропизмом по лимфоцитам и/или макрофагам. Примерами этих вирусов, инфицирующих макрофаги, являются HIV-1, в частности те штаммы, которые обладают мутациями в V3-области gp120, которые приводят к увеличению связывания с макрофагами, HIV-2, Хантавирусы, например Пунмалавирус, цитомегаловирус, респираторно-синцитиальный вирус, вирус простого герпеса или филовирусы.
Примерами вирусов, инфицирующих лимфоциты, являются вирус ветряной оспы (VZV), поскольку VZV, в частности, инфицирует Т-клетки, герпесвирус 6 (НН 6), поскольку HHV 6 также, в частности, инфицирует Т-клетки, вирус бешенства, поскольку оболочечный белок вируса бешенства связывается, в частности, с ТН2-клетками, HIV-1, поскольку гликопротеин gp120 связывается предпочтительно с молекулой CD4 Т-клеток, HTLV-II, поскольку HTLV-II, в частности, инфицирует В-клетки, HTLV-I, поскольку HTLV-I, в частности, инфицирует Т-клетки, вирусы гриппа С, поскольку вирусы гриппа С связываются с N-ацетил-9-β-ацетилнейраминовой кислотой (Neu 5,9 Ac), которая преимущественно встречается в В-лимфоцитах и в меньшей мере или вовсе не встречается в Т-лимфоцитах, через гемагглютинин-эстеразный слитой (HEF) белок, вирусы гриппа С, обладающие мутацией в нуклеотидной позиции 872, которая кодирует позицию 284 аминокислотной последовательности этого HEF, в котором, например, треонин замещен на изолейцин, поскольку данный поверхностный белок HEF, который обладает этой мутацией, имеет заметно более сильное сродство по рецептору N-ацетил-9-O-ацетилнейраминовой кислоты, чем вирус дикого типа, продукты расщепления HEF вируса гриппа С, которые содержат структуру по связыванию N-ацетил-9-β-ацетилнейраминовой кислоты. Эта связывающая структура определяется каталитической триадой серин 71, гистидин 368 или 369 и аспарагиновой кислотой 261, вирус Эпштейн-Барра, поскольку, в частности, EBV инфицирует В-клетки, вирус 2 простого герпеса, поскольку, в частности, HSV-2 инфицирует Т-клетки, или вирус кори.
Примеры лигандов для синовиальных клеток и воспалительных клеток, которые упоминались, представляют собой последовательности нуклеиновой кислоты, которые кодируют антитела или антительные фрагменты, которые связываются, по их вариабельным доменам, с мембранными структурами синовиальных клеток или воспалительных клеток. Примерами этих мембранных структур являются виментин [Miettinen с соавт., Am. J. Pathol. 117, 18 (1984)], фибронектин [Wojciak с соавт. , Clin. Exp. Immunol. 93, 108 (1993)] или Fc-рецепторы. Эти лиганды включают также антитела или антительные фрагменты, которые связываются с соответствующим Fc-рецептором с помощью своих константных доменов [Rojanasakul с соавт., Pharm. Res. 11, 1731 (1994)].
Эти лиганды, кроме того, включают все активные соединения, которые связываются с мембранными структурами или мембранными рецепторами на синовиальных клетках. Примерами последних являются цитокины или ростовые факторы, или их фрагменты или части их последовательностей, которые связываются с рецепторами, которые экспрессируются синовиальными клетками, например IL-1-RA, TNFα, IL-4, IL-6, IL-10, IGF или TGFβ [Callard с соавт., The cytokine, Academic Press (1994)].
Примерами лигандов для клеток, инфицированных вирусом, которые упоминались, являются нуклеиново-кислотные конструкты, которые кодируют антитела или антительные фрагменты, которые направлены против этих вирусных антигенов, которые локализованы на клеточной мембране вирус-инфицированных клеток. Антитела этого рода направлены, например, против антигенов HBV, HCV, HSV, HPV, HIV, EBV или HTLV.
Примеры лигандов для клеток печени и клеток других тканей представляют собой все вещества, которые связываются с мембранными структурами или мембранными рецепторами на поверхности клеток печени. Примерами последних являются ростовые факторы, такие как цитокины, EGF, TGF, FGF или PDGF, или их фрагменты или части их последовательностей, которые связываются с рецепторами, которые экспрессируются в клетках этого рода.
Эти лиганды, кроме того, включают лиганды, которые связываются со структурами клеточных мембран, которые для отдельных тканей являются селективными (см. табл. 1).
Эти лиганды и мембранные структуры представлены в обзоре у Perales с соавт., Eur. J. Biochem. 226, 255 (1994).
Данные лиганды, в частности, включают гликопротеины из оболочек вирусов, которые обладают тропизмом к избранным клеткам, таким как бронхиальные эпителиальные клетки (респираторно-синцитиальный вирус), клетки печени (вирус гепатита С), филовирусы, вирус Марбурга через асиалоглико-протеиновый рецептор клеток печени, вирус гепатита В, связывается на клетках печени предпочтительно с рrеS2 и preS1 доменами HBV через асиалогликопротеиновый рецептор, вирус гепатита D, синусоидальные клетки печени, и вирус гепатита В, который связан с HBV через фибронектин.
Примеры лигандов для клеток глии представляют собой последовательности нуклеиновой кислоты, которые кодируют антитела или антительные фрагменты, которые направлены против мембранных структур клеток глии, как было описано, например, у Mirsky с соавт. [Cell and Tissue Res. 240, 723 (1985)], Coakham с соавт. [Prog. Exp. Tumor Res. 29, 57 (1985)] и McKeever с соавт. [Neurobiol. 6, 119 (1991)]. Эти мембранные структуры включают, кроме того, адгезионные молекулы нервной системы, такие как N-CAM, в частности ее полипептидную цепь С [Nybroe с соавт., J. Cell Biol. 101, 2310 (1985)]. Эти лиганды, кроме того, включают все активные соединения, которые связываются с мембранными структурами или мембранными рецепторами глиальных клеток. Примеры этих активных соединений представляют собой инсулиновый и инсулиноподобный ростовой фактор, и те фрагменты из этих ростовых факторов, которые связываются с важными мембранными рецепторами.
Данные лиганды в рамках содержания настоящего изобретения дополнительно включают последовательности нуклеиновой кислоты, которые кодируют гликопротеины оболочек тех вирусов, которые обладают тропизмом к глиальным клеткам.
Эти вирусы включают, например, JRF1-подтип HIV-1 или вирус I простого герпеса.
Примеры лигандов по лейкозным клеткам включают нуклеиново-кислотные конструкты, которые кодируют антитела или антительные фрагменты, которые направлены против мембранных структур лейкозных клеток. Большое число моноклональных антител этого рода уже были описаны для диагностических и терапевтических процедур [Kristensen, Danish Medical Bulletin 41, 52 (1994); Schranz, Therapia Hungarica 38, 3 (1990); Drexler с соавт., Leuk. Res. 10, 279 (1986); Naeim, Dis. Markers 7, 1 (1989); Stickney с соавт., Curr. Opin. Oncol. 4, 847 (1992); Drexler с соавт., Blut 57, 327 (1988); Freedman с соавт. , Cancer Invest. 9, 69 (1991)]. В зависимости от вида лейкоза, моноклональные антитела, или их антиген-связывающие антительные фрагменты, нижеследующей специфичности, являются, например подходящими в качестве лигандов: AML-клетки, обладающие мембранными антигенами CD13, CD14, CD15, CD33, CAMAL и sialosyl-Le; B-CLL-клетки, обладающие мембранными антигенами CD5, CD1c и CD23, а также идиотипы и изотипы мембранных иммуноглобулинов; T-CLL-клетки, обладающие мембранными антигенами CD33, М38, IL-2 рецепторами и Т-клеточными рецепторами; и ALL-клетки, обладающие мембранными антигенами CALLA и CD19, а также лимфома не-Ходжкина.
Эти лиганды, кроме того, включают все активные соединения, которые связываются с мембранными структурами или мембранными рецепторами лейкозных клеток. Примеры этих последних представляют собой ростовые факторы, или их фрагменты или части их последовательностей, которые связываются с рецепторами, которые экспрессируются в лейкозных клетках.
Ростовые факторы такого рода уже были описаны [обзоры у Cross с соавт., Cell 64, 271 (1991); Aulitzky с соавт., Drugs 48, 667 (1994); Moore, Clin. Cancer Res. 1, 3 (1995); Van Kooten с соавт., Leuk. Lymph. 12, 27 (1993)]. Например, они включают IFNα в случае лимфом не-Ходжкина, IL-2, особенно в случае Т-клеточных лейкозов, FGF - в случае Т-клеточного, моноцитарного, миелоидного, эритроцитарного и мегакариобластного лейкозов, TGFβ - в случае лейкозов, или ретиноиды, например, ретиноевую кислоту, в случае промиелоцитарного лейкоза.
Примеры лигандов для раковых клеток включают последовательности нуклеиновой кислоты, которые кодируют антитела, и фрагменты этих антител, которые направлены против мембранных структур раковых клеток. Антитела такого рода приведены в обзоре, например, у Sedlacek с соавт., Соntrib. to Oncol. 32, Karger Verlag, Munich (1988) и Соntrib. to Oncol. 43. Karger Verlag, Munich (1992).
Другие примеры представляют собой антитела против сиалила Льюиса, пептиды на новобразованиях, которые распознаются Т-клетками, белки, экспрессируемые онкогенами, ганглиозиды, такие как GD3, GD2, GM2, 9-O-ацетил-СD3 и фукозил-GM1, антигены групп крови и их предшественники, антигены полиморфного муцина эпителия или антигены на белках "теплового шока".
Последовательность нуклеиновой кислоты [компонент b], которая кодирует активное соединение (протеин В):
Активное соединение (протеин В) в соответствии с настоящим изобретением может быть веществом, которое, например, принимает участие в биологической активации каскада и/или представляет собой активный компонент этого каскада. Эти вещества включают активные соединения, которые активируют каскад коагуляции, например тромбина [MacGillivray с соавт., Ann. N.Y. Acad. Sci. 485, 73 (1986)], тромбина, который мутировал в области сайта расщепления Arg-Thr (аминокислотная позиция 327/328), фактора Va [Cripe с соавт., Biochem. 31, 3777 (1992), Jenny с соавт., PNAS-USA 84, 4846 (1987)], фактора VIIa [O'Hara с соавт. , PNAS-USA 84, 5158 (1987)], фактора IXa [Yoshitake с соавт., Biochem. 24, 3736 (1985)], фактора Ха [Messier с соавт., Gene 99, 291 (1991) или тканевого фактора и их фрагментов, активирующих коагуляцию [Morrissey с соавт. , Cell 50, 29 (1987); Scarpati с соавт., Biochem. 26, 5234 (1987); Spicer с соавт., PNAS-USA 84, 5148 (1987); Rehemtulla с соавт., Thromb. Heamost. 65, 521 (1991)] или которые ингибируют каскад коагуляции, или которые активируют фибринолиз, например, ингибиторы активатора плазминогена PAI-1, PAI-2 и PAI-2, гирудина, протеина С, ингибиторы серинпротеиназы, такие как С-1S-ингибитор, α1-антитрипсин или антитромбин III, ингибитор метаболизма тканевого фактора (TFPI), активаторы плазминогена, такие как урокиназа, активатор тканевого плазминогена (tPA), или его гибриды, или те, которые активируют каскад комплемента, например фактор яда кобры (CVF) или части последовательностей CVF, которые функционально корреспондируют с С3b-фактором комплемента человека, т.е. те, которые обладают способностью связываться с фактором В комплемента и которые, будучи расщепленными с помощью фактора D, составляют С3-конвертазу (эта последовательность ДНК для CVF и часть ее последовательности были описаны у Fritzinger с соавт., Proc. Natl. Acad. Sci. USA 91, 12775 (1994)), фактора С3b комплемента человека (эта последовательность ДНК для С3 или часть ее последовательности была опубликована De Bruijn с соавт., Proc. Natl. Acad. Sci USA 82, 708 (1985), продукты расщепления фактора С3 комплемента человека, который функционально и структурно походит на CVF (такие продукты расщепления были описаны O'Keefe с соавт., J. Biol. Chem. 263, 12690 (1988) или которые активируют кининовую систему, данную систему комплемента и/или данную систему коагуляции, активируемых, например, фактором Хагемана (F XXIa) [Shibuya с соавт., Biochem. Biophys. Acta 1206, 63 (1994), Que с соавт., Biochem. 25, 1525 (1986), Tripodi с соавт. , Nucl. Acid Res. 14, 3146 (1986)] или калликреином [Chen с соавт., Biochem J. 307, 481 (1995), Fukushima с соавт., Biochem. 24, 8037 (1985)].
Данное активное соединение (протеин В) может быть также цитостатическим, цитотоксическим или белком, вызывающим воспаление, таким как перфорин, гранзим, цитокинами, такими как IL-1, IL-2, TL-4, IL-12, IL-3, IL-5, фактором, ингибирующим лейкоз человека (LIF), IL-7, IL-11, IL-13, GM-CSF, G-CSFb или M-CSF, интерферонами, такими как IFNα, IFNβ или IFNγ, TNF, таким как TNFα или TNFβ, онкостатином М, сфингомиелиназой [Jarvis с соавт., PNAS-USA 91, 73 (1994)], магаинином и производными магаинина [Cruciani с соавт. , PNAS-USA 88, 3792 (1991)]; Jacob с соавт., Ciba Found, symp. 186, 197 (1994); Peck-Miller с соавт., Cancer Chemother. Pharmac. 32, 109 (1993)] или хемокинами, такими как RANTES (МСР-2), хемотактическим и активирующим фактором моноцитов (MCAF), IL-8, макрофагальным воспалительным белком 1 (MIP-1α, или MIP-1β) или белком 2, активирующим нейтрофилы (NAP-2).
Данное активное соединение (протеин В) может быть также антиангиогенным белком, таким как ангиостатин, интерферонами, такими как IFNα, IFNβ или IFNγ, тромбоцитарным фактором 4, IL-12, ТIMP-1, TIMP-2 или ТIMP-3.
Данное активное соединение (протеин В) может быть также ферментом, который обладает способностью конвертировать неактивный предшественник фармакологически активного вещества, например цитостатический агент, в собственно активное вещество. Примеры таких активных соединений представляют собой бактериальную нитроредуктазу, β-глюкуронидазу, растительную β-глюкуронидазу, получаемую из Secale cereale, человеческую β-глюкуронидазу, человеческую карбоксипептидазу (СВ), например, СВ-А тучных клеток или СВ-В поджелудочной железы, бактериальную карбоксипептидазу, бактериальную β-лактамазу, бактериальную цитозиндезаминазу, человеческую каталазу или пероксидазу, фосфатазу, в частности человеческую щелочную фосфатазу или человеческую кислую фосфатазу из предстательной железы, кислую фосфатазу тип 5, оксидазу, в частности человеческую лизилоксидазу или человеческую кислую D-аминооксидазу, пероксидазу, в частности человеческую глутатионпероксидазу, пероксидазу эозинофилов человека или пероксидазу щитовидной железы человека.
Данное активное соединение (протеин В) может быть также белком, который влияет на иммунную систему, например белком, обладающим антиаллергическим действием, таким как IFNβ, IFNγ, IL-10, растворимыми IL-4-рецепторами, IL-12 или TGFβ, или белком, который может предотвращать отторжение трансплантированных органов, таким как IL-10, TGFβ, растворимыми IL-1-рецепторами, растворимыми IL-2-рецепторами, антагонистами рецептора IL-2 или растворимыми IL-6-рецепторами, или белком для терапии аутоиммунных заболеваний, опосредованных антителами, например TGFβ, IFNα, IFNβ, IFNγ, IL-12, растворимыми IL-4-рецепторами или растворимыми IL-6-рецепторами, или белком для терапии клеточно-опосредованных аутоиммунных заболеваний, например IL-6, IL-9, IL-10, IL-13, TNFα, IL-4 или TNFβ, или белком для терапии артрита. В соответствии с настоящим изобретением, могут быть также выбраны структурные гены, чей экспрессированный белок прямо или косвенно подавляет воспаление, например в сочленении, и/или стимулировать воссоздание внеклеточного матрикса (хрящевая и соединительная ткань) в сочленении. Эти экспрессированные белки включают, например, антагонисты рецептора IL-1 (IL-1-RA), поскольку IL-1-RA ингибирует связывание IL-1α, и IL-1β, растворимый IL-1-рецептор, поскольку растворимый IL-1-рецептор связывает и инактивирует IL-1, IL-6, поскольку IL-6 повышает секрецию ТIMP и супероксидов и снижает секрецию IL-1 и TNFα благодаря синовиальным клеткам и хондроцитам, растворимый TNF-рецептор связывает и активирует TNF, IL-4, поскольку IL-4 ингибирует образование и секрецию IL-1, TNFα и ММР, IL-10, поскольку IL-10 ингибирует образование и секрецию IL-1, TNFα и ММР и увеличивает секрецию ТIMP, инсулиноподобный ростовой фактор (IGF-1), поскольку IGF-1 стимулирует синтез внеклеточного матрикса, TGFβ, главным образом TGFβ1 и TGFβ2, поскольку TGFβ стимулирует синтез супероксидисмутазы внеклеточного матрикса, или ТIMP (тканевые ингибиторы металлопротеиназ), главным образом TIMP-1, TIMP-2 или TIMP-3.
Данное активное соединение (протеин В) может быть также белком, ослабляющим поражение нервной системы, например ростовым фактором, таким как FGF, фактором роста нервов (NGF), нейротрофическим фактором, производимым мозгом (BDNF), нейротрофином 3 (NT-3), нейротрофином 4 (NT-4) или ресничным нейротрофическим фактором (CNTF), или цитокином, или цитокиновым ингибитором, который обладает способностью ингибировать или нейтрализовать нейротоксическое действие TNFα, например TGFβ, растворимыми TNF-рецепторами, IL-10, поскольку IL-10 подавляет образование IFNγ, TNFα, IL-2 и IL-4, растворимыми IL1-рецепторами, такими как IL-1-рецептор I или IL-1-рецептор II, поскольку растворимые IL-1-рецепторы нейтрализуют активность IL-1, антагонистом рецептора IL-1 или растворимыми IL-6-рецепторами.
Данное активное соединение (протеин В) может быть также белком, который стимулирует ангиогенез, например VEGF или FGF.
Данное активное соединение (протеин В) может, к тому же, быть белком, который снижает кровяное давление, например калликреином или синтазой окиси азота эндотелиальной клетки.
Данное активное соединение (протеин В) может быть также белком для терапии хронических инфекционных заболеваний, например белком, который обнаруживает цитостатический или цитоксический эффекты, или ферментом, который расщепляет предшественник противовирусного или цитотоксического вещества в соответствующее активное вещество, или цитотоксином, обладающим противовирусным действием, или ростовым фактором, обладающим противирусным действием. Примеры представляют собой IFNα, IFNβ, IFNγ, TNFβ, TNFα, IL-1 или TGFβ.
Настоящее изобретение, кроме того, относится к конструкту нуклеиновой кислоты, в котором две идентичные или две отличающиеся последовательности ДНК, которые кодируют идентичные или различающиеся активные соединения (протеин В) [компонент b) и b")], объединены.
С целью обеспечить экспрессию обоих последовательностей ДНК участок внутреннего входа рибосом (IRES) интеркалировали преимущественно в качестве регуляторного элемента между этими двумя структурами. Участок внутреннего входа рибосом делает возможной экспрессию двух последовательностей ДНК, которые сшивали друг с другом через IRES. IRESs этого рода описаны, например, у Montford и Smith TIG 11, 179 (1995); Kaufman с соавт., Nucl. Acids Res. 19, 4485 (1991); Morgan с соавт., Nucl. Acids Res. 20, 1293 (1992); Dirks с соавт. , Gene 128, 247 (1993); Pelletier и Sonenberg, Nature 334, 320 (1988) и Sugimoto с соавт., BioTechn. 12, 694 (1994). Так, например, соответствующая кДНК для IRES-последовательности полиовируса (позиции ≤ 140 - ≥ 630 из 5'-UTR [Pelletier и Sonenberg, Nature 334, 320 (1988)] можно использовать для соединения ДНК компонента с) с ДНК компонента d).
Последовательности нуклеиновой кислоты [компонент с)], которые кодируют расщепляемую протеазой структурную часть С:
В соответствии с настоящим изобретением структурная часть С включает аминокислотную последовательность, которая расщепляется протеазами, которые образуются в новобразованиях или раковыми клетками или воспалительными клетками. Эту последовательность нуклеиновой кислоты для данной структурной части С инсерцировали, например, в соответствующую последовательность нуклеиновой кислоты природного предшественника (протеин BSD, где S представляет собой естественно встречаемую расщепляемую последовательность) важного активного соединения (протеин В), вместо расщепляемой последовательности S, так, чтобы эта рекомбинантная нуклеиновая кислота экспрессировала протеин BCD или В'BCD.
Данную нуклеиновую последовательность, кодирующую структурную часть С, выбирали в зависимости от соответствующей протеазы, которая, преимущественно, секретируется в данном новобразовании или в данном воспалении.
Структурную часть С можно было бы использовать, например, для нижеследующих ферментов [Barret с соавт., Mammalian Proteases, Academic Press, London (1980), Panchal с соавт., Nature Biotechnol. 14, 852 (1996); Pigott с соавт. , Ayad с соавт., The extracellular Matrix, Academic Press (1994); Yoshida с соавт., Int. J. Cancer 63, 863 (1995), Petersen с соавт., J. Biol. Chem. 265, 6104 (1990); Cramer с соавт., J. Urology 156, 526 (1995); Forsgen с соавт. , FEBS Lett. 213, 254 (1987) Zhang с соавт. Chin Chem. 41 1567 (1995)] (см. табл. 2).
Последовательности нуклеиновой кислоты [компонент d) ], которые кодируют структурную часть D:
В соответствии с настоящим изобретением соответствующая последовательность нуклеиновой кислоты [компонент d)], кодирует пептид (структурная часть D), который связывается с соответствующим структурным компонентом (структурная часть В) через структурную часть С, и инактивирует это активное соединение посредством этого связывания.
Предпочтительно, чтобы последовательности нуклеиновой кислоты, использованные для структурной части D, которые кодируют структурную часть, встречающуюся в естественных предшественниках (протеин BSD), со структурной частью S представляли бы естественно расщепляемую последовательность в протеине BSD.
Структуры естественно встречаемых предшественников активных соединений (протеин В) уже рассмотрены, например, у Barret с соавт., Mammalian Proteases, Academic Press, London (1980) для случая факторов коагуляции, факторов комплемента и калликреина, у Callard с соавт., The Cytokine Facts Book, Academic Press (1994) - для случая интерлейкинов, хемокинов и ростовых факторов, и у Denhardt с соавт., Pharmac. Ther. 59, 329 (1993) - для случая тканевых ингибиторов металлопротеиназ (TIMPs).
При отборе активных соединений, которые не имеют каких-либо естественно встречаемых предшественников, и в случае ксеногенных активных соединений необходимо использовать созданные последовательности нуклеиновой кислоты в виде компонента d.), который кодирует любой пептид предпочтительно, однако, из последовательностей нуклеиновой кислоты, которые кодируют те же структурные части D, которые естественно встречены в предшественниках активных соединений человека.
Для того чтобы облегчить секрецию протеина BCD, или В'BCD, который экспрессируется благодаря новой последовательности нуклеиновой кислоты, соответствующую сигнальную последовательность, которая может быть представлена в ДНК-последовательности компонента b), можно заменить гетерологичной сигнальной последовательностью, которая улучшает внеклеточную секрецию. Так, например, можно инсерцировать соответствующую сигнальную последовательность для иммуноглобулина [позиции ДНК ≤ 63 - ≥ 107; Reichmann с соавт., Nature 332, 323 (1988)] или соответствующую сигнальную последовательность для СЕА [позиции ДНК ≤ 33 - ≥ 134, Schrewe с соавт., Mol. Cell Biol. 10, 2738 (1990); Berling с соавт., Cancer Res. 50, 5634 (1990)] или соответствующую сигнальную последовательность гликопротеинов респираторно-синцитиального вируса человека [кДНК аминокислот ≤ 38 - ≥ 50 или 48-65; Lichtenstein с соавт., J. Gen. Virol. 77, 109 (1996)].
Кроме того, с целью усиления трансляции, можно инсерцировать нуклеотидную последовательность GCCACC или GCCGCC [Kozak, J. Cell Biol. 108, 299 (1989)] по 3'-концу соответствующей промоторной последовательности и непосредственно по 5'-концу стартового сигнала (ATG) соответствующей сигнальной последовательности.
Получение новых конструктов нуклеиновой кислоты
Новые нуклеиново-кислотные конструкты были описаны и получены путем сшивки друг с другом индивидуальных компонентов, используя стандартные молекулярные биологические методы.
Применения:
Определенный новый нуклеиново-кислотный конструкт особенно хорошо подходит для лечения заболеваний, которые сопровождаются увеличением локального образования протеаз, таких как раковые заболевания, лейкозы, аллергии, аутоиммунные заболевания, инфекции, воспаления, реакции отторжения трансплантатов, тромбозы и окклюзии кровеносных сосудов и другими нарушениями свертывания крови и кровообращения, и повреждения тканей, включая повреждения центральной нервной системы и повреждение определенной нервной системы. Эффективным является введение местное (например, в определенный участок кожи), через нос, пероральное, через желудочно-кишечный тракт, внутрибронхиальное, внутрипузырное, интравагинальное, в матку, подкожное, внутримышечное, вокругсуставное, внутрисуставное, в цереброспинальную жидкость, в соответствующую ткань мозга, в спинной мозг, в раны, внутрибрюшинное или внутриплевральное, или системное, например, внутривенное, внутриартериальное, внутрипортальное или в сердце.
В общем, данное лекарственное средство включает, где требуется, помимо обычных добавок и вспомогательных веществ, либо соответствующий конструкт нуклеиновой кислоты, либо клетку, которая обладает способностью экспрессировать этот новый нуклеиново-кислотный конструкт. Данное лекарственное средство можно вводить для профилактики или терапии заболевания, как об этом подробно описано выше.
Указанную клетку получают, например, путем трансформации или трансфекции клеток соответствующим новым нуклеиново-кислотным конструктом, используя способы, известные специалистам.
Примерами подходящих клеток являются эндотелиальные клетки, лимфоциты, макрофаги, глиальные клетки, фибробласты, клетки печени, клетки почек, мышечные клетки, клетки костной или хрящевой ткани, перитонеальные клетки, клетки кожи, клетки эпителия, лейкозные клетки и/или раковые клетки.
Эти новые клетки пригодны также для получения белка, который кодируется соответствующим новым нуклеиново-кислотным конструктом и который можно использовать непосредственно в качестве лекарственного средства.
Настоящее изобретение, кроме того, относится к использованию соответствующего нуклеиново-кислотного конструкта для получения рекомбинантно измененной клетки, когда этот нуклеиново-кислотный конструкт вводят в данную клетку, чтобы использовать этот новый нуклеиново-кислотный конструкт для получения белка, который кодируется этим нуклеиново-кислотным конструктом, где этот нуклеиново-кислотный конструкт будет экспрессироваться в подходящей клетке, а соответствующий получаемый белок выделяют, и к клетке, которая является вместилищем для этого нуклеиново-кислотного конструкта. Вышеописанные клетки представляют собой предпочтительные клетки.
Последующий отбор можно, например, осуществить из промоторных последовательностей и структурных генов (для протеина BCD и В'BCD) из вышеприведенных примеров в зависимости от природы и места заболевания и от клетки-мишени, чтобы быть преобразованной.
Терапия новобразований
Промоторы [компонент а)]:
специфичные для эндотелиальной клетки, или специфичные для клеточного цикла, или неспецифичные для клетки, или специфичные для мышечной клетки и специфичные для клеточного цикла, или специфичные для раковой клетки (солидные новобразования, лейкозы)
Лиганды для нижеследующих клеток-мишеней [компонент b')]:
пролиферирующих эндотелиальных клеток или стромы клеток и мышечных клеток, прилежащих к этой эндотелиальной клетке, или раковым клеткам, или лейкозным клеткам.
Структурные гены [компонент b)c)d)]:
для факторов, индуцирующих коагуляцию, для факторов комплемента, для ингибиторов развития кровеносных сосудов, для цитостатических и цитотоксических белков, для индукторов воспаления или для ферментов, активирующих предшественники цитостатических агентов, например для ферментов, которые расщепляют вещества-предшественники (пролекарства), образуя таким образом активные цитостатические агенты (лекарства).
Терапия аутоиммунных заболеваний и воспалений:
Промоторы [компонент а)]:
специфичные для эндотелиальной клетки и специфичные для клеточного цикла, или макрофагально-специфичные, и/или лимфоцитарно-специфичные, и/или специфичные для клеточного цикла, или специфичные для синовиальной клетки, и/или специфичные для клеточного цикла.
Лиганды для нижеследующих клеток-мишеней [компонент b')]:
пролиферирующие эндотелиальные клетки, макрофаги и/или лимфоциты или синовиальные клетки.
Структурные гены [компонент b)c)d)]:
для соответствующей терапии аутоиммунных заболеваний, опосредованных антителом, для ингибиторов клеточной пролиферации, цитостатических или цитотоксических белков, ферментов, активирующих предшественники цитостатических агентов, или для терапии артрита.
Терапия повреждения определенной нервной системы:
Промоторы [компонент а)]:
специфичные для глиальной клетки, специфичные для эндотелиальной клетки и специфичные для клеточного цикла или неспецифичные и специфичные для клеточного цикла.
Лиганды для нижеследующих клеток-мишеней [компонент b')]:
глиальные клетки или пролиферирующие эндотелиальные клетки.
Структурные гены [компонент b)c)d)]:
для ростовых факторов нейронов, например для цитокинов и цитокиновых ингибиторов, которые ингибируют или нейтрализуют данный нейротоксический эффект TNFα.
Терапия патологий системы коагуляции крови и системы кровообращения:
Промоторы [компонент а)]:
неспецифичные для клетки, неспецифичные для клетки и специфичные для клеточного цикла или специфичные для эндотелиальных клеток, клеток гладкой мускулатуры или макрофагов, или специфичные для эндотелиальных клеток, клеток гладкой мускулатуры или макрофагов и специфичные для клеточного цикла.
Лиганды для нижеследующих клеток-мишеней [компонент b')]:
эндотелиальных клеток, пролиферирующих эндотелиальных клеток или соматических клеток вблизи эндотелиальных клеток клеток гладкой мускулатуры или макрофагов.
Структурные гены [компонент b)c)d)]:
для ингибирования коагуляции или содействия фибринолиза, для факторов развития кровеносных сосудов, для гипотензивных пептидов, для антипролиферативного, цитостатического или цитотоксического белка или для фермента, расщепляющего предшественники цитостатических агентов, образующего таким образом цитостатические агенты, для подавления пролиферации клеток гладкой мускулатуры, следующей за повреждением эндотелиального слоя или для белков плазмы крови, таких как C1-инактиватор, сывороточная холинэстераза или α1-антитрипсин.
Терапия хронических инфекционных заболеваний
Промоторы [компонент а)]:
вирус-специфичные, клеточно-специфичные или вирус-специфичные или клеточно-специфичные и специфичные для клеточного цикла.
Лиганды для нижеследующих клеток-мишеней [компонент b')]:
клеток печени, лимфоцитарных и/или макрофагальных, эпителиальных клеток или эндотелиальной клетки.
Структурные гены [компоненты b)c)d)]:
для белка, который проявляет цитостатический или цитотоксический эффекты, фермента, который расщепляет предшественники противовирусного или цитотоксического вещества, образуя таким образом соответствующее активное вещество, или противовирусные белки, такие как противовирусные активные цитокины и ростовые факторы.
Настоящее изобретение поясняется с помощью нижеследующих примеров и фигур, к тому же не ограничивающих его.
Описание представленных фигур
Фиг. 1: Схематическое изображение нового нуклеиново-кислотного конструкта, включающего компоненты а), b), с) и d.).
Фиг. 2: Схематическое изображение нового нуклеиново-кислотного конструкта, который увеличен в результате добавления компонента b').
Фиг. 3: Схематическое изображение нуклеиново-кислотного конструкта для фактора X, активируемого PSA.
Примеры
1. Получение нуклеиново-кислотного конструкта, кодирующего FX, активируемого простат-специфичным антигеном
Касается получения терапевтического агента для лечения метастазов карциномы предстательной железы. Несмотря на хирургическое удаление предстательной железы, которая стала раковой, часто возникают метастазы карциномы предстательной железы, которые в настоящее время, в большинстве своем, не лечатся и которые приводят к смерти пациента. Такие метастазы карциномы предстательной железы индуцируют развитие кровеносных сосудов. Кроме того, метастазы карциномы предстательной железы выделяют тканеспецифичный фермент, т. е. простат-специфичный антиген (PSA). Согласно настоящему изобретению получали нуклеиново-кислотный конструкт, который интродуцировали в пролиферирующие эндотелиальные клетки, что приводило к экспрессии модифицированного фактоpa коагуляции FX. Эта модификация включает замену, в природном гене FX, соответствующей нуклеотидной последовательности для естественного сайта расщепления, расщепление которого дает FXa, активирующий коагуляцию, на нуклеотидную последовательность, кодирующую PSA-специфичный сайт расщепления. В итоге, этот PSA, который выделяется метастазами карциномы предстательной железы, обладает способностью специфически активировать модифицированный FX, который выделяется пролиферирующими эндотелиальными клетками вокруг метастазов, и, таким образом, инициировать коагуляцию, которая ведет к нарушению кровоснабжения этих метастазов, а затем - к некрозу этих метастазов.
Этот нуклеиново-кислотный конструкт для FX, активируемого PSA, получали согласно схеме, которая представлена на Фиг. 3.
ДНК-последовательности индивидуальных компонентов объединяли воедино в направлении 5' к 3' следующим образом:
Компонент а), который содержит соответствующую промоторную последовательность из гена cdc25C [нуклеиновые кислоты: -290 ÷ +121; Lucibello с соавт. , EMBO J. 14, 132 (1995); Zwicker с соавт., Nucl. Acids Res. 23, 3822 (1995); EMBO J. 14, 4514 (1995)], последовательность GCCACC (Kozak, J. Cell Biol. 108, 229 (1989)) и соответствующую кДНК для иммуноглобулинового сигнального пептида [нуклеотидная последовательность ≤ 63 - ≥ 107; Reichmann с соавт. , Nature 332, 323 (1988)], сливали с компонентом b)c)d), который содержит соответствующую кДНК для человеческого FX (нуклеотидная последовательность 1 - ≥ 1468) [Messier с соавт., Gene 99, 291 (1991)], в котором аминокислота мутировала от Аrg до Туг.
Соответствующие индивидуальные компоненты этого конструкта присоединяли через подходящие сайты рестрикции, которые интродуцировали по концам различных элементов путем PCR-амплификации. Такое сшивание является эффективным при использовании ферментов, которые специфичны по сайтам рестрикции, которые известны специалистам в данной области, и по ДНК-лигазам. Эти ферменты можно приобрести коммерческим путем.
Этот нуклеотидный конструкт, который получали таким способом, клонировали в плазмидные векторы pUC 18/19 или векторы, полученные на основе Bluescript.
2. Экспрессия в эмбриональных клетках почки человека
Пролиферирующие эмбриональные клетки почки человека [НЕК 293; Racchi с соавт. , J. Biol. Chem. 268, 5735 (1993)], которые поддерживали в культуре, трансфецировали вышеописанной плазмидой, используя метод, известный специалистам в данной области [Graham и van der Eb, Virol. 52, 456 (1973)].
Мутантный фактор X выделяли очисткой из супернатанта из приблизительно 107 трансфецированных клеток НЕК 293 [Watzke с соавт., J. Clin. Invest. 88, 1685 (1991)] и испытывали в тесте на коагуляцию для фактора Х с добавлением и без добавления PSA. Очищенный PSA получали от Chemicon (Temecula, CA, USA).
В этом тесте дефект коагуляции в человеческой плазме с недостаточностью по FX уравновешивали с помощью функционально активного FXa.
Немутантный (дикого типа) FX (который активируется ядом гадюки Рассела) использовали в качестве положительного контроля. Кроме испытуемой смеси, лишенной PSA, использовали ложный препарат из супернатанта нетрансфецированных клеток НЕК 293 в качестве негативного контроля.
Коагуляционную активность мутантного FX измеряли по времени рекальцификации (плазмы) (Seitz с соавт., Int. J. Cancer 53:514-520, 1993). 100 мкл плазмы, лишенной FX (Behringwerke, Marburg), инкубировали при 37oС в течение 120 с со 100 мкл FX-препарата из соответствующего клеточного супернатанта. Этот FX-препарат содержит PSA в качестве активатора. В случае негативного контроля PSA не добавляли. FX (дикого типа) и яд гадюки Рассела (RVV) использовали в качестве положительного контроля. Реакцию коагуляции усиливали добавлением 100 мкл 0,02 М CaCl2 и определяли в коагулометре.
Получали следующие результаты:
Негативные контроли без какой-либо активации коагуляции дают время коагуляции приблизительно 200 с. Напротив, намного более короткое время, 50 с, получали, когда использовали активированный FX (мутантный FX и PSA или FX дикого типа и RVV).
Из этого можно заключить, что преобразованные клетки НЕК 293 экспрессируют мутантный FX, который в присутствии добавленного PSA уравновешивает коагуляционный дефект плазмы, лишенной FX.
Экспрессия в человеческих эндотелиальных клетках
Эндотелиальные клетки пуповины человека, которые поддерживали в культуре, трансфецировали вышеописанной плазмидой, используя метод, известный специалистам (Lucibello с соавт., EMBO J. 14, 132 (1995)).
С целью проверить специфичность клеточного цикла эндотелиальные клетки синхронизировали в G0/G1, с изъятием метионина, в течение 48 часов. После окрашивания с Hoechst 33258 (Hoechst AG, Frankfurt) в клетках определяли содержание ДНК на активируемом флуоресценцией анализаторе клеток (Lucibello с соавт., EMBO J. 14, 132 (1995)).
В супернатанте из эндотелиальных клеток проверяли экспрессию нуклеиново-кислотного конструкта по аналогии с исследованиями, осуществленными на клетках НЕК 293.
Получали следующие результаты:
Получаемый белок, экспрессируемый трансфецированными эндотелиальными клетками, уравновешивал коагуляционный дефект плазмы, лишенной FX, в противоположность ложным препаратам из супернатанта нетрансфецированных эндотелиальных клеток.
Отчетливо высокую концентрацию мутантного FX можно выявить в супернатанте пролиферирующих, преобразованных эндотелиальных клеток (ДНК > 2S) при сравнении с супернатантом эндотелиальных клеток, которые были синхронизированы в G0/G1 (ДНК = 2S).
Следовательно, вышеописанный нуклеиново-кислотный конструкт связан с геном, мутантным по FX, который экспрессируется в эндотелиальных клетках способом, зависящим от клеточного цикла, и этот мутантный FX может быть активирован с помощью РSА так, чтобы он был причиной коагуляции в плазме, лишенной FX.


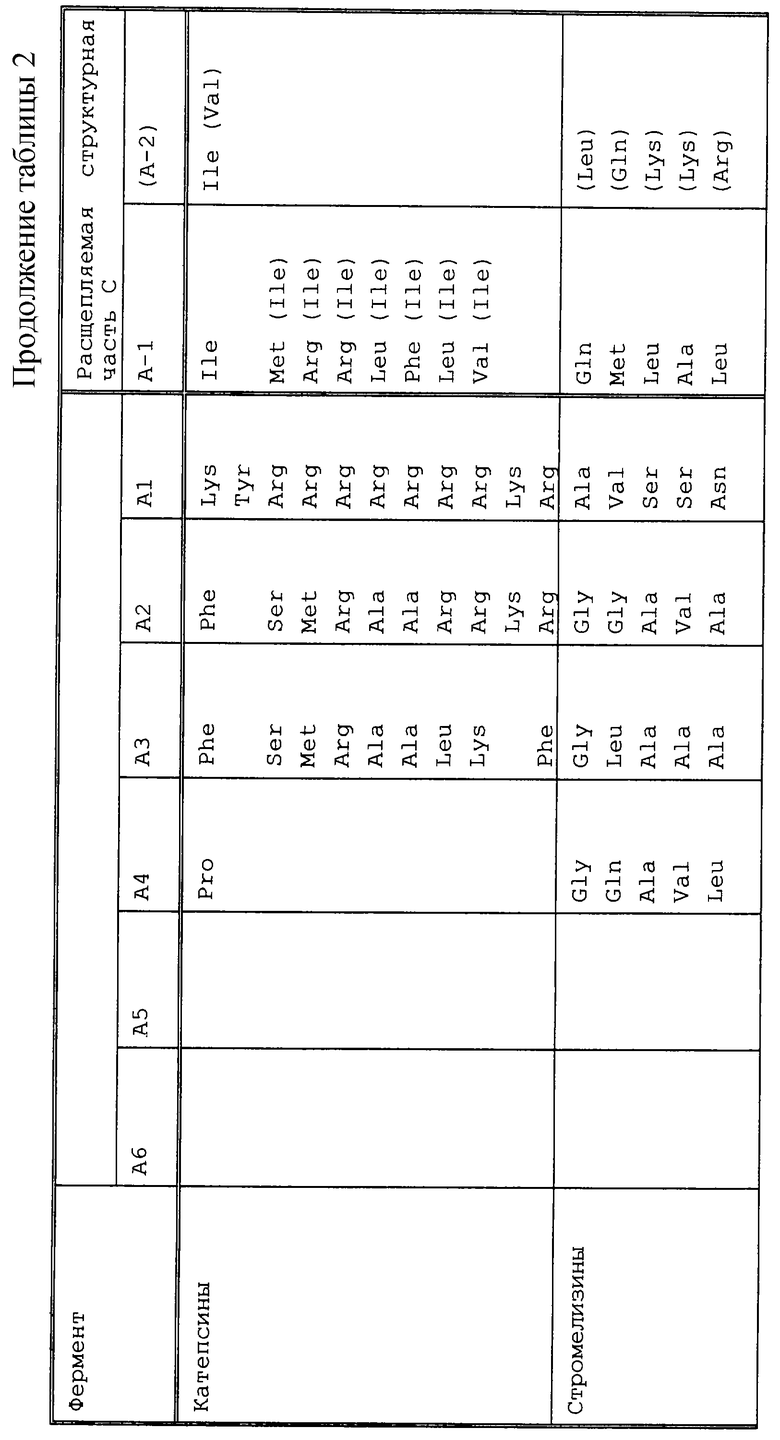
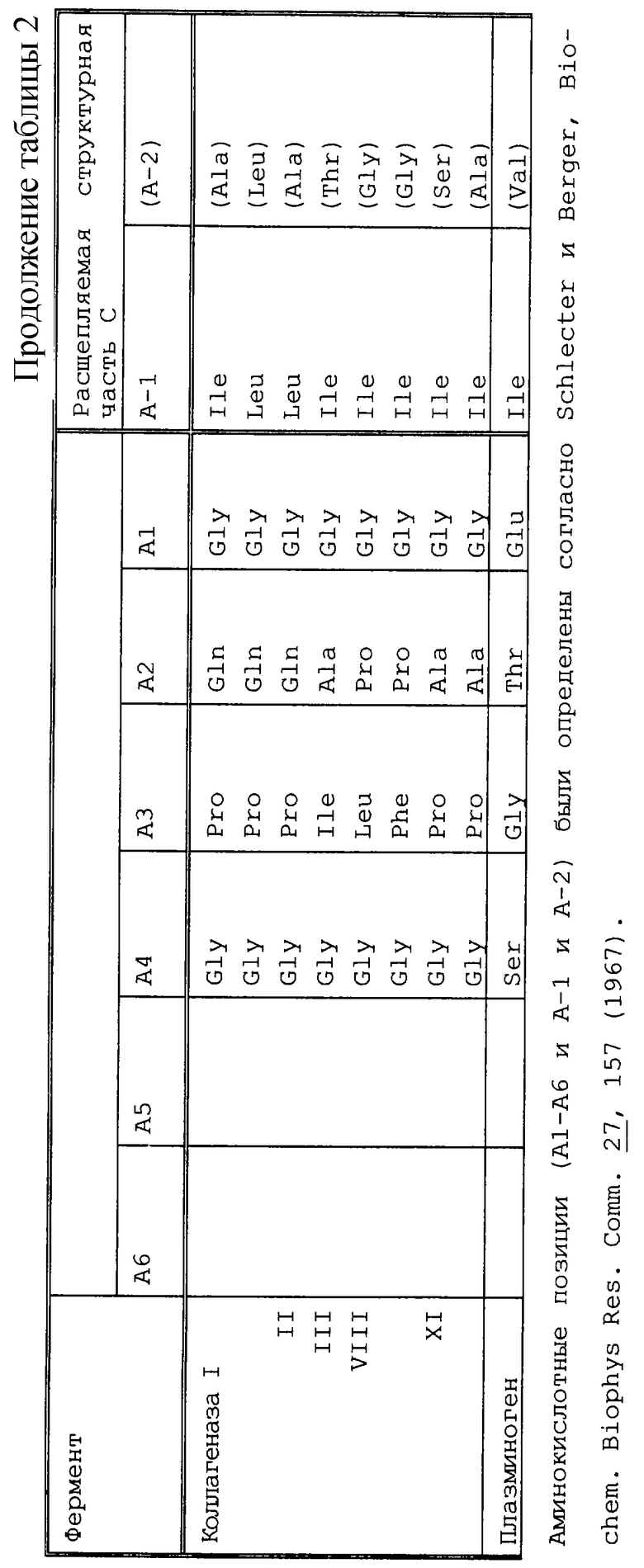
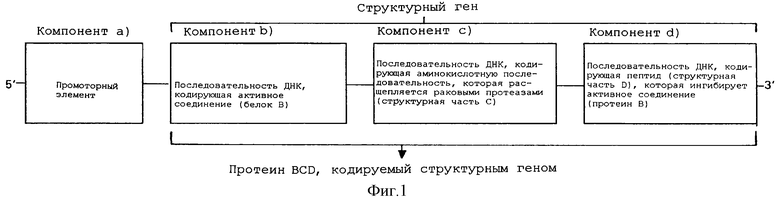


Изобретение относится к области биотехнологии и касается нуклеиново-кислотного конструкта для экспрессии активного вещества, которое активируется с помощью фермента-протеазы, который высвобождается из клеток млекопитающего, и может применяться в генной терапии. Конструкт включает нижеследующие компоненты: а) как минимум, один промоторный элемент, b) как минимум, одну последовательность ДНК, которая кодирует активное соединение (протеин В), с) как минимум, одну последовательность ДНК, которая кодирует аминокислотную последовательность (структурную часть С), которая может быть расщеплена специфически с помощью фермента, который высвобождается из клетки млекопитающего, и d) как минимум, одну последовательность ДНК, которая кодирует пептид или белок (структурная часть D), который связывается с соответствующим активным соединением (протеин В) через расщепляемую аминокислотную последовательность (структурная часть С) и ингибирует активность этого активного соединения (протеин В). Преимущество изобретения заключается в разработке конструкта, который может высвобождать активное вещество с помощью протеаз, вырабатываемых клетками. 2 с. и 14 з.п.ф-лы, 3 ил., 2 табл.
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| MOSCATELLI et al | |||
| Purification of a factor from numan placenta that stimulates capillary endothelial cell protease production, dNA synthesis and migration, PNAS USA, 1986, v | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| Прибор для измерения глубины воды и пройденного судном пути | 1925 |
|
SU2091A1 |

