Настоящее изобретение относится к промотору гена человеческого эндоглина или его функциональным частям и к его применению для получения лекарственного средства.
Одной из важных проблем генной терапии является регулирование транскрипции и трансляции эффекторного гена, который встраивается в клетку. На уровне транскрипции этот контроль делается возможным за счет добавления против хода транскрипции к промоторной или энхансерной последовательности - кодирующей последовательности эффекторного гена.
Под "промоторной последовательностью" понимают последовательность, представляющую собой сегмент гена, с которым могут связываться регуляторные белки - так называемые факторы транскрипции, которые в целом активируют транскрипцию эффекторного гена в прямом направлении. Эти области, которые лежат в направлении транскрипции, называют "нижними" последовательностями, в то время как последовательности, которые располагаются в противоположном направлении, называют "верхними" последовательностями. Под "эффекторным геном" обычно понимают ген, являющийся структурным геном, генный продукт которого обладает, например, нужным эффектом в смысле действия при генной терапии.
Такие промоторные или энхансерные последовательности могут быть неклеточно-специфическими, клеточно-специфическими, вирус-специфическими, метаболически-специфическими или специфическими для клеточного цикла. Примеры таких промоторных последовательностей и их применение, например, для генной терапии при различных заболеваниях описываются в заявках на патент WO 96/06940, WO 96/06938, WO 96/06941 и WO 96/06939. Кроме того, в этих заявках на патент приводятся методы и примеры соединения этих промоторных последовательностей, например, с целью клеточно-специфического и специфического для клеточного цикла управления эффекторным геном.
В зависимости от выбора и комбинации промоторов последние являются причиной большей или меньшей рестрикции и/или более или менее сильной траскрипции эффекторного гена, который зависит от этих промоторов.
Эндотелиальная клетка является примером выгодной клетки-мишени для генной терапии, с одной стороны, потому что эндотелиальные клетки откровенно доступны для генных конструкций, которые инъецируют в сердечно-сосудистую систему, и, с другой стороны, потому что они непосредственно вовлекаются в развитие и ход ряда заболеваний, таких как опухолевые заболевания, воспаления, аллергии, аутоиммунные болезни, реакции отторжения органов и нарушения кровообращения и коагуляции, а также в процессы заживления и/или непосредственно примыкают к месту расположения этих нарушений.
Как правило, мишенные клеточно-специфические промоторы являются промоторами генов для тех белков, которые образуются особенно энергично или в исключительно большой степени в релевантной клетке-мишени. В случае эндотелиальной клетки примером одного из таких белков является эндоглин.
Эндоглин представляет собой не передающий сигнал рецептор TGFβ (Gougos et al., J.Biol. Chem., 265, 8361 (1990), Cheifetz, J. Biol. Chem., 267, 19027 (1992), Moren et al., BBRC 189, 356 (1992)). Хотя он встречается в небольшом количестве в нормальном эндотелии, он в возросшей степени экспрессируется в пролиферирующем эндотелии (Westphal et al., J. Invest. Derm., 100, 27 (1993), Burrows et al., Pharmac. Ther., 65, 155 (1994)). Доступная информация в отношении промотирующей силы и клеточной специфичности отсутствует. Несмотря на тот факт, что эндоглиновый ген известен уже примерно 4 года (Bellon et al., (1993)), до сих пор не удалось выделить эндоглиновый промотор.
Bellon и др. (Eur. J. Immunol., 23, 2340 (1993)) описали кДНК-последовательность человеческого эндоглина, в то время как мышиный эндоглин описан Ge и др. (Gene, 138, 201 (1994)). Хотя имеется информация о последовательности для части 5’-нетранслируемой области эндоглинового гена, ничего не известно о функции этой области или о промоторной области.
Другим эндотелиальным клеточно-специфическим белком является VEGF-рецептор. В этом случае различают два рецептора (Plate et al. Int. J. Cancer, 59, 520 (1994)): с одной стороны, VEGF-рецептор 1 (flt-1) (de Vries et al., Science, 255, 989 (1992)), который содержит fms-подобную тирозинкиназу в цитоплазматической части, и VEGF-рецептор 2 (flk-1, KDR) (Terman et al., BBRC, 187, 1579 (1992)), который содержит тирозинкиназу в цитоплазматической части. Оба рецептора обнаруживаются почти исключительно на эндотелиальных клетках (Senger et al., Cancer Metast. Rev., 12, 303 (1993)).
Другими эндотелиальными клеточно-специфическими рецепторными тирозинкиназами являются tie-1 или tie-2 (Partanen et al., Mol. Cell. Biol., 12, 1698 (1992), Schnurch und Risau, Development, 119, 957 (1993), Dumont et al., Oncogene, 7, 1471 (1992)) и рецептор В61 (Eck-рецептор) (Bartley et al. Nature, 368, 558 (1994), Pandey et al.. Science, 258, 567 (1995), van der Geer et al., Ann. Rev. Cell. Biol., 10, 251 (1994)).
Другими эндотелиальными клеточно-специфическими белками являются: молекула В61, которая представляет лиганд для рецептора В61 (Hoizman et al. J. Am. Soc. Nephrol., 4, 466 (1993), Bartley et al., Nature, 368, 558 (1994)), эндотелин, в частности эндотелин В (O’Reilly et al., J. Cardiovasc. Pharm., 22, 18 (1993), Benatti et al., J. Clin. Invest., 91, 1149 (1993), O’Reilly et al., BBRC, 193, 834 (1993)), промоторная последовательность которого описана в работе Benatti et al., J. Clin. Invest., 91, 1149 (1993), эндотелин 1 (Yanasigawa et al.. Nature, 332, 411 (1988)), промоторная последовательность которого описана в работе Wilson et al., Mol. Cell. Biol., 10, 4654 (1990), эндотелиновые рецепторы, в частности рецептор эндотелина В (Webb et al. Mol. Pharmacol., 47, 730 (1995), Haendler et al. J. Cardiovasc. Pharm., 20, 1 (1992)), маннозо-6-фосфатные рецепторы (Perales et al., Eur. J. Biochem., 226, 225 (1994)), чьи промоторные последовательности описаны в работах Ludwig и др. (Gene, 142, 311 (1994)), Oshima и др. (J. Biol. Chem., 263, 2553 (1988)) и Pohlmann и др. (PNAS USA, 84, 5575 (1987)), и фактор фон Виллебранда (vWF), промоторная последовательность которого описана в работах Jahroudi и Lynch (Mol. Cell. Biol., 14, 999 (1994)), Ferria и др. (Biochem. J., 293, 641 (1993)) и Aird и др. (PNAS USA, 92, 4567 (1995)).
Другими эндотелиальными клеточно-специфическими белками являются IL-1 в форме, например, IL-1α и IL-1β, которые продуцируются активированными эндотелиальными клетками (Warner et al. J. Immunol., 139, 1911 (1987)) и промоторные последовательности которых описаны в работах Hangen et al. Mol. Carcinog., 2, 68 (1986), Turner et al. J. Immunol., 143, 3556 (1989), Fenton et al. J. Immunol., 138, 3972 (1987), Bensi et al. Cell Growth Diff., 1, 491 (1990), Hiscott et al., Mol. Cell. Biol., 13, 6231 (1993) и Mori et al., Blood, 84, 1688 (1994), рецептор IL-1, промоторная последовательность которого описана в работе Ye et al., PNAS USA, 90, 2295 (1993), и фактор васкулярной клеточной адгезии (VCAM-1), причем экспрессия VCAM-1 в эндотелиальных клетках активируется липополисахаридами, TNF-α (Neish et al., Mol. Cell. Biol., 15, 2558 (1995)), IL-4 (Iademarco et al., J. Clin. Invest., 95, 264 (1995)) и IL-5 (Marni et al., J. Clin. Invest., 92, 1866 (1993)). Промоторная последовательность VCAM-1 описана в работах Neish et al., Mol. Cell. Biol., 15, 2558 (1995), Ahmad et al., J. Biol. Chem., 270, 8976 (1995), Neish et al., J. Exp. Med., 176, 1583 (1992), Iademarco et al., J. Biol. Chem., 267, 16323 (1992) и Cybulsky et al., PNAS USA, 88, 7859 (1991).
Другими эндотелиальными клеточно-специфическими промоторами являются синтетические активирующие последовательности, при этом синтетические активирующие последовательности, которые составлены из олигомеризованных сайтов связывания для факторов транскрипции, которые преимущественно или избирательно активны в эндотелиальных клетках, например фактора транскрипции GATA-2, сайтом связывания которого в гене эндотелина 1 является 5’-ТТАТСТ-3’ (Lee et al., Biol. Chem. 266, 16188 (1991), Dorfmann et al., J. Biol. Chem., 267, 1279 (1992) и Wilson et al., Mol. Cell. Biol., 10, 4854 (1990)), также могут использоваться в качестве альтернативы природным эндотелийспецифическим промоторам и специфический для головного мозга эндотелиальный глюкозо-1-переносчик, при этом эндотелиальные клетки головного мозга характерно, весьма энергично экспрессируют этот переносчик для осуществления трансэндотелиального переноса D-глюкозы в головной мозг (Gerhart et al., J. Neurosci. Res., 22, 464 (1989)). Промоторная последовательность описана в работе Murakami и др. (J. Biol. Chem., 267, 9300 (1992)).
Будучи довольно специфическими для эндотелиальных клеток, некоторые из этих промоторов, например промотор для гена фактора фон Виллебранда или для гена для VEGF-рецептора 1 (flk-1), имеют, однако, только относительно низкую активность. Хотя активность таких "слабых" промоторов можно увеличить путем соединения их с основным промотором (например, SV40) или энхансером, обычно такая процедура приводит в дальнейшем к сопутствующему падению специфичности.
Целью настоящего изобретения является поэтому поиск промотора, который является как сильным, так и эндотелийспецифическим.
К удивлению, обнаружилось, что этими свойствами среди прочих обладает промотор эндоглинового гена.
Настоящее изобретение относится, следовательно, к промотору гена человеческого эндоглина или его функциональным частям и вариантам. Обнаружено, что этот промотор имеет протяженность максимум в 2415 пар оснований (см. табл.1, послед. №1) и преимущественно включает нуклеотидную последовательность 1-2378, в том числе инициирующую последовательность.
Чтобы охарактеризовать промотор по настоящему изобретению, промоторную последовательность эндоглинового гена или ее части связывают с репортерным геном (например, геном, кодирующим фермент люциферазу) в плазмиде pGL3 (Promega) и эндотелиальные клетки (клеточная линия ECV-304) и для сравнения клетки цервикальной карциномы (клеточная линия HeLa) трансфектируют этой конструкцией. Обнаружено, к удивлению, что эндоглиновый промотор почти в 80 раз сильнее, чем промотор vWF. Это удивляет, так как vWF, как упоминалось ранее, экспрессируется эндотелийспецифически и, следовательно, можно было бы ожидать, что активность эндоглинового промотора будет такой же, как y vWF-промотора. Также обнаружено, что эндоглиновый промотор примерно в 30 раз активнее в эндотелиальных клетках, чем в клетках цервикальной карциномы. Это удивляет, поскольку vWF-промотор, который также является эндотелийспецифическим, обладает схожей активностью в клетках цервикальной карциномы и в эндотелиальных клетках. Следовательно, эндоглиновый промотор по настоящему изобретению явно превосходит промотор vWF-гена как в отношении силы, так и в отношении специфичности для эндотелия.
Также обнаружено, что части промоторной последовательности по настоящему изобретению также показывают сильную специфическую для эндотелиальных клеток активность.
Так, получают приведенные ниже значения относительной (по отношению к активности промотора SV40 в качестве стандарта) активности в эндотелиальных клетках для вырезанных из 5’-конца конструкции промотора эндоглинового гена.

Термин "функциональные части промотора" понимается как обозначающий все неполные последовательности промотора по изобретению, которые обладают промоторной активностью, в частности неполные последовательности приблиз. от 1 до приблиз. 2378, приблиз. от 36 до приблиз. 2378, приблиз. от 470 до приблиз. 2378 и приблиз. от 948 до приблиз. 2378, а также неполные последовательности приблиз. от 36 до приблиз. 2415, приблиз. от 470 до приблиз. 2415 и приблиз. от 948 до приблиз. 2415, предпочтительно - неполные последовательности приблиз. от 470 до приблиз. 2415 и приблиз. от 470 до приблиз. 2378. Неполные последовательности, обладающие промоторной активностью, также простираются, например, приблиз. от 1310 до приблиз. 2415, приблиз. от 1310 до приблиз. 2378, приблиз. от 1847 до приблиз. 2415 и приблиз. от 1847 до приблиз. 2378.
Однако настоящее изобретение не ограничивается промотором, изображенным в послед. №1, и его функциональными частями, но также включает варианты, которые обладают промоторной активностью. Варианты такого рода содержат, например, делеции, добавления, инсерции и/или замещения одного или нескольких оснований, предпочтительно приблиз. от 1 до приблиз. 50 оснований, в частности - приблиз. от 1 до приблиз. 25 оснований, особенно - приблиз. от 1 до приблиз. 5 оснований. Промоторную активность можно легко измерить, например, с использованием описанного люциферазного анализа.
Настоящее изобретение также относится к нуклеотидной конструкции, которая содержит,
а) по меньшей мере, одну нуклеотидную последовательность промотора по изобретению (компонент а)) и в соответствующем случае,
б) по меньшей мере, один эффекторный ген (компонент б)), причем транскрипция этого эффекторного гена активируется компонентом а).
Компонент а) предпочтительно располагается против хода транскрипции компонента б).
Настоящее изобретение также относится к нуклеотидной конструкции, в которой промоторная последовательность эндоглинового гена по изобретению соединяется с другой мишенной клеточно-специфической, вирусно-специфической, специфической для метаболизма или специфической для клеточного цикла промоторной последовательностью и, по меньшей мере, с одним эффекторным геном, в которой эта комбинация промоторных последовательностей регулирует активацию транскрипции, по меньшей мере, одного эффекторного гена.
Нуклеотидная конструкция по изобретению состоит предпочтительно из ДНК. Термин "нуклеотидная конструкция" понимается как обозначающий искусственные структуры, которые составляются из нуклеиновой кислоты и которые можно транскрибировать в клетках-мишенях. Их предпочтительно встраивают в вектор, например в невирусные векторы, такие как плазмиды, или вирусные векторы. Специалист в этой области техники хорошо знаком с получением невирусных векторов и вирусных векторов.
Настоящее изобретение также относится к клеткам, которые укрывают нуклеотидную конструкцию по настоящему изобретению.
Вообще, выбор эффекторного гена зависит от болезни, от которой лечат с помощью генной конструкции.
Примеры таких эффекторных генов для лечения опухолевых заболеваний, лейкозов, аутоиммунных болезней, аллергии, артрита, воспалительных заболеваний, отторжения органов, реакций "транспланта против хозяина", заболеваний свертывающей системы крови сердечно-сосудистых заболеваний, анемии, инфекций или поражения ЦНС подробно описаны в заявках на патент WO 96/06940, WO 96/06938, WO 96/06941 и WO 96/06939.
Например, эффекторные гены, соответствующие настоящему изобретению, кодируют цитокин, хемокин, фактор роста, рецептор для цитокина, рецептор для хемокина или рецептор для фактора роста, а также антагонист цитокина, белок, который индуцирует цитостаз, цитотоксичность или апоптоз, антитело или фрагмент антитела, ингибитор ангиогенеза, коагулирующий фактор, ингибитор свертывания крови, фибринолитический белок, фермент, который расщепляет предшественника лекарственного средства, образуя посредством этого лекарственное средство, белок, который оказывает влияние на кровообращение, или антиген инфекционного патогенного микроорганизма, который вызывает иммунную реакцию.
Нуклеотидная конструкция по изобретению также может содержать два или большее число идентичных или различных эффекторных генов, которые соединяются друг с другом через промоторные последовательности или внутренние рибосомные аминоацильные сайты (IRES). Их примеры приводятся в упомянутых выше заявках на патент.
Нуклеотидную конструкцию по настоящему изобретению можно использовать, например, для экспрессии гена только клеточно-специфическим для эндотелиальных клеток способом или клеточно-специфическим для эндотелиальных клеток и специфическим для метаболизма способом, клеточно-специфическим для эндотелиальных клеток и специфическим для клеточного цикла и/или клеточно-специфическим для эндотелиальных клеток и вирусно-специфическим, причем ген предпочтительно представляет ген, который кодирует фармакологически активное соединение или фермент, который расщепляет неактивный предшественник лекарственного средства с образованием посредством этого активного лекарственного средства.
Предпочтение отдается применению нуклеотидной конструкции по настоящему изобретению для получения лекарственного средства для лечения перечисленных выше заболеваний, причем указанное получение фармацевтического средства обычно включает клонирование нуклеотидной конструкции в подходящий вектор, который затем, например, вводят пациенту. Специалист в этой области техники хорошо знаком с другими применениями промотора по настоящему изобретению или нуклеотидной конструкции по настоящему изобретению.
Приведенный ниже пример вместе с таблицей и чертежами предназначен для более подробного описания изобретения без ограничений его.
Описание таблицы и чертежей
Таблица 1. Последовательность промотора человеческого эндоглина. Пара оснований 1 соответствует области последовательности, которая располагается дальше всего от 5’. Высококонсервативная последовательность A1u располагается в области пар оснований 1360-1666, в то время как гомология с документально подтвержденной кДНК M.musculus начинается в 3’-области с пары оснований 2300, а документально подтвержденная часть кДНК Н.sapiens (5’-нетранслируемая область) начинается у пары оснований 2379.
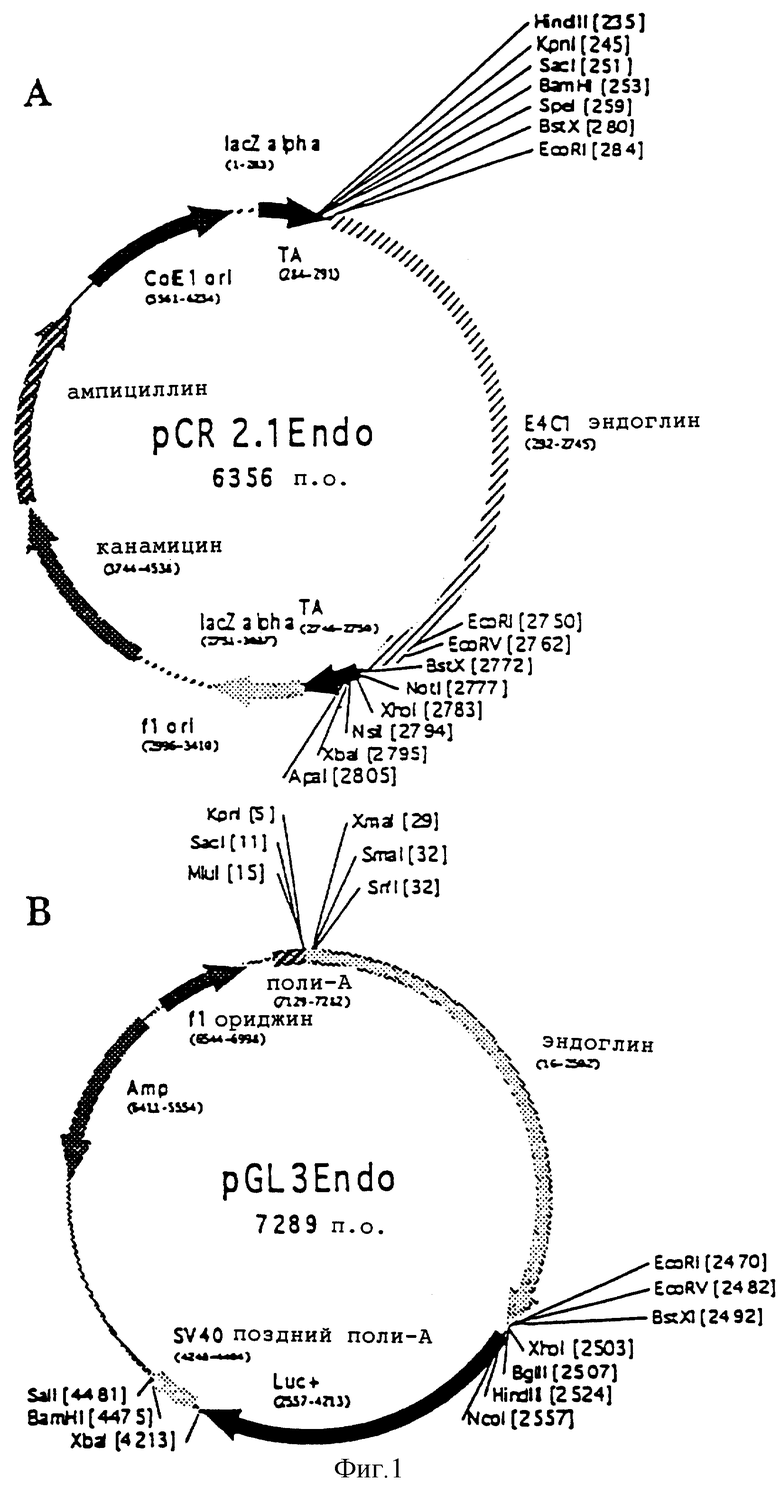
Фиг.1. Клонирование промотора человеческого эндоглина. А: фрагмент промотора человеческого эндоглина, который получен с помощью PCR, лигируют к сайту клонирования ТА вектора pCR 2.1 (Invitrogen). В: фрагмент, содержащий промотор человеческого эндоглина, исключают из конструкции pCR 2.1 Endo ферментами M1uI и Xhol и клонируют в люциферазный репортерный вектор pGl3 (Promega).
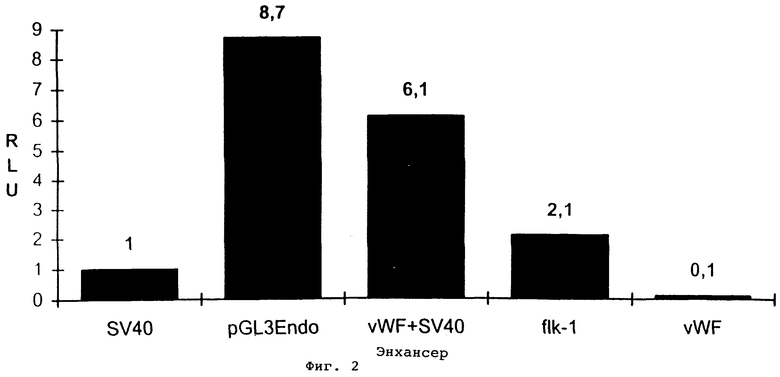
Фиг.2. Люциферазная активность различных эндотелиальных клеточно-специфических промоторов. Все промоторы клонируют в pGl3 и все величины стандартизируют по отношению к основному промотору SV40. SV40: основной промотор SV40 без энхансера. PGL3Endo: эндоглиновый промотор (см. фиг.1В). Энхансер vWF+SV40: промотор фактора фон Виллебранда, усиленный энхансером SV40 в обратном направлении, flk-1: промотор для VEGF-рецептора flk-1 (-224/начиная от ATG). vWF: промотор фактора фон Виллебранда (-487/+247) без дополнительных энхансеров.
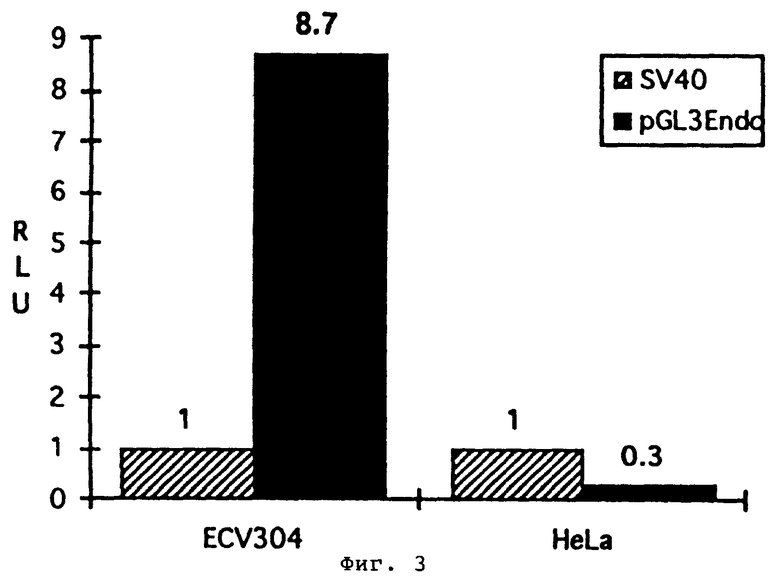
Фиг.3. Люциферазная активность промотора человеческого эндоглина в эндотелиальных и в неэндотелиальных клетках. Конструкции те же, что на фиг.2. Стандартизированную относительно основного промотора SV40 активность эндоглинового промотора в эндотелиальной клеточной линии ECV304 сравнивают с его активностью в клеточной линии HeLa цервикального рака. При этом анализе активность в клетках ECV304 приблизит в 29 раз выше, чем в клетках HeLa.
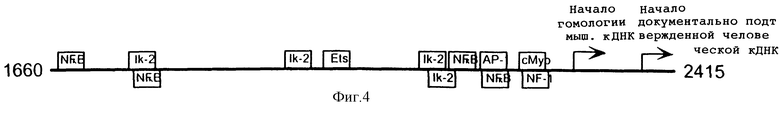
Фиг.4. Предполагаемые сайты связывания для факторов транскрипции на эндоглиновом промоторе. Показана только область между последовательностью Alu и началом кДНК. Все сайты связывания располагаются на плюс(+)-цепи.
Пример
Для клонирования промотора используют набор для прогулки по ДНК PromoterFinder™ (Clontech). С использованием этого набора фрагмент приблизит. в 2,4 тысячи пар оснований документально подтвержденной последовательности из 5’-нетранслируемой области кДНК человеческого эндоглина амплифицируют в два этапа PCR с помощью двух генспецифических затравок
E1: GCTGGGCTGGAGTTGCTGTCCGAAGGATC (послед.№2) и
Е2: AATGGATGGCAGTCACAGCAGCAGTCCTG (послед. №3).
Условия PCR в этом случае следующие.
1. PCR: Затравка Е1 - 25 с при 94°С, 25 с при 94°С, 20 с при 63°С, 4 мин при 68°С, 39 циклов - 4 мин при 68°С.
2. (Гнездовая) PCR: Затравка Е2 - 25 с при 94°С, 25 с при 94°С, 20 с при 61°С, 4 мин при 68°С, 26 циклов - 4 мин при 68°С.
Полимераза: Система для PCR с длинной матрицей Expand™ (Boehringer Mannheim).
Фрагмент PCR очищают на спин-колонках QIAquick™ (Qiagen) и вставляют в клонирующий вектор ТА (набор Original ТА Cloning™ (Invitrogen)). Эту конструкцию pCR 2.1Endo (см. фиг.1А) секвенируют и клонированную область идентифицируют как 5’-область в 2415 пары оснований гена человеческого эндоглина (последовательность, см. табл.1).
Клонированную область из этого вектора клонируют в люциферазный репортерный вектор, т.е., pGL3 (Promega), и испытывают его на промоторную активность, как конструкцию pGL3Endo (см. фиг.1Б), в клетках HeLa и ECV304.
Клетки трансфектируют или с помощью метода DEAE/декстран (на основе метода Sompayrac et al., PNAS, 78, 7575 (1981)), или с использованием LipofectAMINE™ (Gibco BRL). В качестве стандарта трансфектируют основной промотор SV40 так же, как конструкцию pGL3Endo; также трансфектируют промотор flk-1-(VEGF) (-225/начиная с ATG), а также промотор фактора фон Виллебранда (vWF) (-487/+247) с или без энхансера SV40. Эта последняя vWF-промоторная конструкция, содержащая энхансер SV40, отличается тем, что ее активность заметно выше, чем активность промотора дикого типа, в то время как ее селективность, хотя и снижается, но не уничтожается. Все конструкции клонируют в pGL3 и осуществляют анализ с люциферазой, как описано Herber и др. (Oncogene, 9, 1295 (1994)) и Lucibello и др. (EMBO J., 14, 132 (1995)).
Люциферазная активность различных промоторов в клетках ECV304 (фиг.2) показывает, что клонированный фрагмент 5’-области эндоглинового гена обладает промоторной активностью. Эта активность весьма высока, когда ее сравнивают с активностью других типичных эндотелиальных клеточно-специфических промоторов. Она в четыре раза выше, чем активность промотора flk-1, и более чем в восемь раз выше активности промотора vWF. Активность конструкции pGL3Endo является более высокой даже тогда, когда промотор vWF усиливают энхансерной последовательностью SV40. Эти данные подтверждают, что клонированная область является промотором гена человеческого эндоглина.
На фиг.3 показывается сравнение активности эндоглинового промотора в клетках ECV304 с его активностью в клеточной линии цервикального рака HeLa. При стандартизации по основному промотору SV40 активность в клетках ECV304 примерно в 29 раз выше, чем в клетках HeLa. Это указывает на то, что клонированный промотор не только активен в эндотелиальных клетках, но также является селективным для этих клеток.
На фиг.4 показаны предполагаемые сайты связывания для факторов транскрипции на эндоглиновом промоторе. Некоторые сильно гомологичные потенциальные сайты связывания, которые могут быть ответственными за селективность и активность промотора и которые включают несколько консервативных сайтов связывания NF-КB, располагаются в области между консервативной Alu-последовательностью и документально подтвержденной кДНК.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКИ ИЗМЕНЕННЫЕ КЛЕТКИ И ИХ ПРИМЕНЕНИЕ ДЛЯ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ | 1998 |
|
RU2205029C2 |
| САМОУСИЛИВАЮЩИЕСЯ ФАРМАКОЛОГИЧЕСКИ КОНТРОЛИРУЕМЫЕ ЭКСПРЕССИОННЫЕ СИСТЕМЫ | 1997 |
|
RU2197993C2 |
| ГЕННАЯ ТЕРАПИЯ ЗАБОЛЕВАНИЙ СОСУДОВ С ПОМОЩЬЮ СПЕЦИФИЧЕСКОГО К КЛЕТКЕ И ЗАВИСИМОГО ОТ КЛЕТОЧНОГО ЦИКЛА АКТИВНОГО ВЕЩЕСТВА | 1995 |
|
RU2197992C2 |
| ОПОСРЕДОВАННАЯ РЕЦЕПТОРАМИ ДОСТАВКА ГЕНОВ С ИСПОЛЬЗОВАНИЕМ ВЕКТОРОВ НА ОСНОВЕ БАКТЕРИОФАГОВ | 1998 |
|
RU2209088C2 |
| НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ, СПОСОБНАЯ ИНГИБИРОВАТЬ АКТИВНОСТЬ IL-6, ПЛАЗМИДНЫЙ ВЕКТОР ДЛЯ ТРАНСФЕКЦИИ В КЛЕТКИ МЛЕКОПИТАЮЩИХ, НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ, ИСПОЛЬЗУЕМАЯ ПРИ ТЕРАПИИ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ (ВАРИАНТЫ) | 1995 |
|
RU2205874C2 |
| РЕКОМБИНАНТНЫЙ ЛЕНТИВИРУСНЫЙ ВЕКТОР, КЛЕТКА-ХОЗЯИН, ТРАНСДУЦИРОВАННАЯ ЛЕНТИВИРУСНЫМ ВЕКТОРОМ, СПОСОБ ЕЕ ТРАНСДУКЦИИ И ПРИМЕНЕНИЕ | 2002 |
|
RU2305708C2 |
| УСОВЕРШЕНСТВОВАНИЕ ГЕНЕТИЧЕСКИХ КОНСТРУКЦИЙ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АНТИВИЧ ТЕРАПИИ | 2013 |
|
RU2533817C1 |
| СРЕДСТВО ДЛЯ ГЕННОЙ ТЕРАПИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2010 |
|
RU2447150C1 |
| УНИВЕРСАЛЬНЫЕ РАКОВОСПЕЦИФИЧНЫЕ ПРОМОТОРЫ И ИХ ИСПОЛЬЗОВАНИЕ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2013 |
|
RU2539764C2 |
| СПОСОБЫ И КОМПОНЕНТЫ ИНДУКЦИИ ОПУХОЛЬ-СПЕЦИФИЧЕСКОЙ ЦИТОТОКСИЧНОСТИ | 1998 |
|
RU2214280C2 |




Изобретение относится к области генной инженерии и может быть использовано в медико-биологической промышленности при получении лекарственных средств для генной терапии. Получена и охарактеризована последовательность промотора гена эндоглина человека, характеризующегося высокой специфичностью в отношении эндотелия определены фрагменты указанной последовательности, которые сохраняют способность выполнять функцию полноразмерного промотора. Предложены нуклеотидные конструкции, в которых под контролем нового промотора или его активного фрагмента находится ген(ы), транскрипцию которого(ых) в эндотелиальных клетках желательно активировать. Путем встраивания предложенных конструкций в подходящие векторы могут быть получены лекарственные средства для лечения широкого круга заболеваний человека. 3 с.и 9 з.п. ф-лы, 4 ил., 1 табл.
| GOUGOS A., LETARTE M., J | |||
| Biol Chem., 265, № 15, р.8361-8364, 1990 | |||
| US 5466596 А, 14.11.1995. |

