Настоящее изобретение относится к способу получения вектора экспрессии, кодирующего адаптированную рекомбиназу, где указанная адаптированная рекомбиназа осуществляет рекомбинацию ассиметричных участков-мишеней в LTR провирусной ДНК ретровируса, встроенного в геном клетки-хозяина. Такие адаптированные рекомбиназы, распознающие ассиметричные участки-мишени в LTR провирусной ДНК, представляют собой средства для вырезание провируса из генома клетки-хозяина. Настоящее изобретение, кроме того, относится к использованию адаптированных рекомбиназ для получения фармацевтических композиций для снижения вирусной нагрузки у индивида, инфицированного ретровирусом. Также изобретение относится к способу оптимизации лечения ретровирусной инфекции у индивида in vitro, включающему адаптацию рекомбиназ, специфично распознающих и рекомбинирующих последовательности ассиметричных участков-мишеней в провирусной ДНК ретровируса у инфицированного индивида.
ОБЛАСТЬ ТЕХНИКИ
Ретровирусные инфекции такие как, например, инфекции вируса иммунодефицита человека (ВИЧ), до сих пор остаются одними из наиболее важных и наиболее распространенными заболеваниями человека.
Что касается СПИД, то носителем ВИЧ, яляющегося ретровирусом, вызывающим СПИД, по оценкам являются 39,5 миллионов человек. Последние данные указывают, что даже в 2006 году произошло приблизительно 4,3 миллионов новых случаев инфицирования, а в некоторых областях мира скорость распространения инфекции с 2004 года увеличилась более чем на 50%. Кроме того, по данным AIDS Epidemic Update 2006, публикуемого WHO, (декабрь 2006), в 2006 году от связанных со СПИД заболеваний умерли приблизительно 2,9 миллиона человек.
Основными задачами прикладной противоретровирусной терапии являются снижение частоты заболеваемости ВИЧ и смертности, улучшение качества жизни, восстановление и сохранение функционирования иммунной системы и максимальная и длительная супрессия вирусной нагрузки. В настоящее время противоретровирусная терапия, а более точно, схемы лечения против ВИЧ в основном основаны на ингибиторах вирусных ферментов и молекулах, ингибирующих слияние вируса с клеткой.
В этом отношении в настоящее время существуют четыре доступных класса лекарственных средств против ВИЧ, которые используют для лечения СПИД. Эти классы лекарственных средств направлены на конкретные стадии в процессе репликации ВИЧ.
Первым классом активных средств являются ингибиторы слияния (FI), которые работают вне клеток-хозяев с целью предотвращения слияния ВИЧ с этими клетками, его проникновения в эти клетки и инфицирование этих клеток. Соответствующим подходом является предотвращение связывания ВИЧ с клеткой-мишенью посредством CD4-рецепторов и корецепторов, называемых CCR5 или CXCR4, на поверхности клеток-мишеней.
Три других класса активных средств действуют внутри клетки. Так называемые нуклеозидные ингибиторы обратной транскриптазы (NRTI), ненуклеозидные ингибиторы обратной транскриптазы (NNRTI) и ингибиторы протеаз (PI) используют для предотвращения репликации вируса внутри клеток-хозяев после их инфицирования ВИЧ.
Примерами NRTI и NNRTI, препятствующих образованию копий генетической информации ВИЧ (продуцирующих, таким образом, так называемую провирусную ДНК), являются 3TC (ламивудин, эпивир), абакавир (зиаген), AZT (зидовудин, ретровир), d4T (ставудин, зерит), ddC (зальцитабин, хивид), ddI (диданозин, видекс/видекс EC), FTC (эмтрицитабин, эмтрива), эфавиренз (сустива) и невирапин (вирамун).
Мишенью PI является фермент ВИЧ протеаза, вовлеченная в сборку вируса. Примерами активных средств являются ампренавир (агенераза), атазанавир (реатаз), фосампренавир (телзир), индинавир (криксиван), лопинавир, нелфинавир (вирасепт), ритонавир (норвир) и саквинавир (инвираза/фортоваза).
Один из типов используемого в настоящее время комбинированного лечения, включающего использование более одного активного средства, представляет собой высокоактивную противоретровирусную терапию (ВААРТ), направленную на вирусную обратную транскриптазу, протеазу и слияние (Gulick et al., 1997; Lalezari et al., 2003). Использование такой терапии привело к преобразованию инфекции ВИЧ-1 в хроническое заболевание, которое снижает смертность инфицированных индивидов.
Однако одним из недостатков всех современных подходов лечения является то, что они только подавляют жизненный цикл вируса, не устраняя инфекции. Основным препятствием при таких способах лечения является образование долгоживущих резервуаров ВИЧ-1, в частности в латентно инфицированных покоящихся CD4+ T-клетках (Chun et al., 1998; Finzi et al., 1997), что требует пожизненной ВААРТ.
К сожалению, у все более увеличивающегося количества пациентов долговременная ВААРТ сопровождается значительными неблагоприятными побочными эффектами, включая токсичность для митохондрий, липодистрофию, сахарный диабет и остеопороз (Dybul et al., 2002). Значительная токсичность лекарственных средств часто приводит к нечеткому следованию схеме лечения, что приводит к недостаточному ингибированию репликации вируса. Как следствие возникают новые штаммы вируса ВИЧ-1, которые устойчивы к супрессирующей терапии (Little et al., 2002). Вследствие увеличивающегося количества устойчивых штаммов ВИЧ необходимы новые активные средства, которые в настоящее время находятся на стадии разработки. Кроме того, для улучшенного контроля ВИЧ-1 иследуют новые мишени в вирусах и новые подходы ингибирования (Donzella, 1998; Chiu et al., 2005; Hazuda et al., 2004; Hauber et al., 2005).
Обсуждаемый в данной области альтернативный подход заключается в нацеливании на провирус, встроенный в геном клетки-хозяина. Например, вырезание провирусной ДНК из генома хозяина могло бы предотвратить дальнейшую репликацию ВИЧ и такое вырезание отличается от современных способов тем, что обладает возможностью уничтожать даже латентный вирус, присутствующий в геноме хозяина.
Одним из классов белков, которые рассматривают для применения в этом альтернативном подходе, являются сайт-специфические рекомбиназы (Flowers et al., 1997). Сайт-специфические рекомбиназы в природе опосредуют множество функций, начиная от перестановки генов до сегрегации геномов, например, такого как вырезание, инверсия или интеграция определенных сегментов ДНК (обзор представлен в Stark et al., 1992).
Одной из простейших и наиболее исследованных рекомбиназ является рекомбиназа Cre бактериофага P1, которая разделяет геномные димеры до мономеров путем рекомбинации двух идентичных участков двухцепочечной ДНК со специфической последовательностью (Hoess & Abremski, 1985). Рекомбиназа Cre нашла широкое применение в генетических исследованиях на мышах (Nagy, 2000). Рекомбиназа Cre представляет собой белок массой 38 кДа, название которого связано с его функцией, так как он вызывает рекомбинацию (causes recombination) (Sternberg & Hamilton, 1981). Необходимым условием для рекомбинации является расположение двух участков рекомбинации, распознаваемых Cre в антипараллельной ориентации, которые затем связываются четырьмя идентичными субъединицами Cre, связывающимися с образованием кольца, в котором каждая субъединица контактирует с двумя соседними субъединицами и половиной участка в одном участке рекомбинации (Hoess & Abremski, 1985). Участок рекомбинации, распознаваемый Cre, представляет собой последовательность двухцепочечной ДНК длиной 34 п.н., известную как loxP (из участка кроссинговера (locus of crossing over (x)), P1; Sternberg & Hamilton, 1981), которая является палиндромной за исключением восьми центральных пар оснований (называемых спейсером), которые задают направленность участка.
Некоторые системы сайт-специфической рекомбинации, включая систему Cre/loxP, функционируют без вспомогательных белков или кофакторов, в широком спектре условий в клетке. Однако так как сайт-специфические рекомбиназы функционируют на основе специфических взаимодействий субъединиц фермента рекомбиназы с распознаваемыми последовательностями-мишенями ДНК, то использование этих ферментов ограничено необходимостью того, что области-мишени ДНК должны содержать соответствующим образом расположенные участки-мишени (Lewandoski, 2001). До настоящего времени рекомбиназы дикого типа, которая в качестве ее последовательностей-мишеней ДНК распознает природные ретровирусные последовательности, не было найдено.
В последние годы был проведен глубокий мутационный и структурный анализ сайт-специфических рекомбиназ с изменением их свойств для достижения лучшего понимания сложных механизмов действия этих ферментов (обзор представлен в van Duyne, 2001; и Coates et al., 2005). Большое число исследований были сосредоточены на рекомбиназе Cre с целью анализа ее способности к эволюции. Несколько исследований показали, что если в участке распознавания рекомбиназы Cre, а именно, loxP, изменены несколько нуклеотидов, то специфичность распознавания мишени Cre может изменяться (Buchholz & Stewart, 2001; Santoro & Schultz, 2002; Rufer & Sauer, 2002). Дополнительные исследования направлены на конструирование мутантных участков-мишеней loxP, содержащих последовательности из LTR ВИЧ-1, с конструированием возможных участков-мишеней для использования Cre в качестве противовирусной стратегии. (Lee & Park, 1998; Lee et al., 2000). До настоящего времени, однако, было невозможно получить рекомбиназу, которая бы распознавала природные ассиметричные последовательности ВИЧ в качестве последовательностей-мишеней ДНК.
Способ направленной эволюции представляет собой мощный способ для выбора ферментов с измененной специфичностью (рассмотрено в Yuan et al., 2005; и Johannes & Zhao, 2006). Сначала этот способ использовали для выделения улучшенных ферментов на основе РНК путем селекции молекулы РНК с измененными субстратными участками. Использование способа на основе ПЦР позволяет скринировать очень большие библиотеки и выделять из совокупности кандидатов удачные кодирующие области. В отличие от этого, при направленной эволюции белков скрининг и выделение улучшенных мутантов, которых идентифицируют по изменениям в свойствах белка, необходим способ восстановления последовательности нуклеиновой кислоты, кодирующей белок. Связь между белком и его кодирующей последовательностью часто поддерживают посредством компартментализации. Таким образом, скрининг библиотек в направленной эволюции белков был ограничен подходами "один за другим", которые сохраняют компартменты, а преимущества, связанные со скринингом совокупности кандидатов, были недоступны.
Это ограничение было устранено благодаря созданию способов, позволяющих перекрестно сливать белки с соответствующими матричными РНК (мРНК), используя слитые конструкции мРНК-белок и рибосомный дисплей. Таким образом, функциональный скрининг белков с улучшенными свойствами связан с прямым получением соответствующих кодирующих молекул, а скринингу in vitro подвергали большие совокупности (например, смотри Buchholz et al., 1998). Дополнительного улучшения направленной эволюции белков достигли посредством так называемой эволюции на основе сшитых с субстратом белков (SLiPE; Buchholz & Stewart, 2001), где субстрат рекомбиназы помещали на ту же молекулу ДНК, что и кодирующую область белка. Таким образом, если в компартменте экспрессировалась рекомбиназа, ее действие изменяло субстрат ДНК, следующий за ее собственной кодирующей областью. Таким образом, библиотеку можно было скринировать в качестве совокупности с помощью ПЦР с амплификацией только кодирующих областей-кандидатов, которые следовали за измененным субстратом. Это обеспечивает удобный скрининг больших библиотек для быстрого получения удачных кодирующих областей. Этот способ использовали для изменения ДНК-специфичности рекомбиназы Cre и адаптации ее к новому распознаваемому участку-мишени (Buchholz & Stewart, 2001).
Однако основной помехой использования любой рекомбиназы для вырезания ретровирусной ДНК является необходимость симметричных участков-мишеней для рекомбиназы, которые, как правило, не встречаются в провирусной ДНК по меньшей мере дважды, чтобы использовать известные в настоящее время рекомбиназы.
Таким образом, учитывая возможность сайт-специфических рекомбиназ и необходимость создания терапевтического подхода для лечения СПИД, при котором устранялся бы провирус ВИЧ-1 из генома клетки-хозяина, задача настоящего изобретения состоит в создании способа получения адаптированной рекомбиназы, которая осуществляет рекомбинацию ассиметричных участков-мишеней в LTR провирусной ДНК ретровируса, встроенного в геном клетки-хозяина, таким образом, вырезая провирус из генома клетки-хозяина.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В соответствии с первым аспектом настоящее изобретение относится к способу получения вектора экспрессии, кодирующего адаптированную рекомбиназу, которая осуществляет рекомбинацию ассиметричных участков-мишеней в LTR провирусной ДНК, введенной в геном клетки-хозяина, причем способ включает стадии:
(a) определения последовательности LTR провирусной ДНК путем идентификации в ней последовательностей, гомологичных с последовательностями левой половины участка и правой половины участка известных участков-мишеней рекомбиназ по меньшей мере на 30%, причем гомологичные последовательности разделены спейсером из 5-12 нуклеотидов, и гомологичные последовательности LTR с наибольшей гомологией с известным участком-мишенью представляют ассиметричную последовательность-мишень;
(b) получения двух синтетических последовательностей, в которых первая синтетическая последовательность соответствует последовательности асимметричной последовательности-мишени стадии (а), гомологичной левой половине участка указанного известного участка-мишени плюс последовательность спейсера, и обозначена как "последовательность половины участка 1", а вторая синтетическая последовательность соответствует последовательности спейсера плюс последовательность ассиметричной последовательности-мишени стадии (а), гомологичная правой половине участка, и обозначена как "последовательность половины участка 2";
(c) определения нуклеотидов в синтетических последовательностях стадии (b), которые отличаются от соответствующих гомологичных последовательностей левой половины участка и правой половины участка известного гомологичного участка-мишени стадии (а);
(d) получения первой подгруппы двух последовательностей-мишеней на основе синтетических последовательностей стадии (b), в которой первая последовательность-мишень в первой подгруппе содержит инвертированный повтор, состоящий из последовательности половины участка 1 стадии (а) и последовательности половины участка 1', разделенных последовательностью спейсера, а вторая последовательность-мишень в первой подгруппе содержит инвертированный повтор, состоящий из последовательности половины участка 2' и последовательности половины участка 2 стадии (b), разделенных последовательностью спейсера, при этом последовательности половины участка 1' и 2' представляют инвертированные повторы соответствующих последовательностей половины участка 1 и 2 стадии (b);
(e) получение второй подгруппы последовательностей-мишеней на основе последовательностей-мишеней первой подгруппы стадии (d), в которой каждую последовательность половин участков вместе с соответствующей последовательностью спейсера последовательностей-мишеней первой подгруппы стадии (d) используют для получения независимой последовательности-мишени второй подгруппы, формируя инвертированный повтор на основании выбранной последовательности половины участка так, чтобы последовательность спейсера разделяла обе последовательности, образуя инвертированный повтор, где последовательности обеих последовательностей половин участков, происходящие от одной последовательности-мишени первой подгруппы стадии (d) при синтезе и до их использования изменяют с получением инвертированного повтора, образующего полную последовательность-мишень так, что в последовательности левой половины участка часть нуклеотидов, отличающихся от соответствующей гомологичной последовательности половины участка известного участка-мишени стадии (а), замещена природными нуклеотидами, находящимися в известном участке-мишени, а в последовательности правой половины участка оставшиеся нуклеотиды, отличающиеся от соответствующей гомологичной левой половины участка, замещены природными нуклеотидами, находящимися в известном участке-мишени так, что в обеих последовательностях половин участков, происходящей из одной из последовательностей-мишеней первой подгруппы стадии (d), взятых вместе, можно найти все отличающиеся нуклеотиды, тогда как ни одна из указанных последовательностей половин участков отдельно не содержит всех отличающихся нуклеотидов;
(f) получения дополнительных подгрупп последовательностей-мишеней на основе последовательностей-мишеней из второй подгруппы, полученных на стадии (e), посредством пошагового повторения процесса стадии (e), каждый раз получая новую подгруппу последовательностей-мишеней до тех пор, пока последовательности половин участков, образующие инвертированные повторы в каждой полученной последовательности-мишени содержат один, два или три нуклеотида, отличающихся от соответствующей гомологичной последовательности половины участка известного участка-мишени;
(g) использования направленной молекулярной эволюции для рекомбиназы, распознающей известный гомологичный участок-мишень, выбранный на стадии (a), с использованием в качестве субстрата последовательностей-мишеней из конечной подгруппы, полученной на стадии (f), содержащих один, два или три нуклеотида, отличающихся от соответствующей гомологичной последовательности половины участка указанного известного гомологичного участка-мишени;
(h) перестановки в библиотеках рекомбиназ, полученных на стадии (g);
(i) использования направленной молекулярной эволюции для библиотеки с перестановками, полученной на стадии (h), с использованием последовательностей-мишеней следующей подгруппы более высокого порядка в соответствии со стадией (f);
(j) повторения стадий (h) и (i) по меньшей мере до тех пор, пока не будет получена посредством направленной молекулярной эволюции одна рекомбиназа, активная в отношении асимметричной последовательности-мишени в LTR ретровирусной ДНК стадии (а);
(k) выделения из библиотеки нуклеиновой кислоты по меньшей мере одной рекомбиназы, полученной на стадии (j); и
(l) клонирования нуклеиновой кислоты, полученной на стадии (k) в подходящем векторе экспрессии.
В предпочтительном варианте осуществления рекомбиназа, к которой на стадиях (g) и (i) применяют направленную молекулярную эволюцию, происходит из семейства сериновых интеграз или из семейства тирозиновых интеграз, и, предпочтительно, рекомбиназа представляет собой модифицированную рекомбиназу Cre фага P1, модифицированную рекомбиназу FLP дрожжей или модифицированную рекомбиназу Dre фага D6. Предпочтительно, направленная молекулярная эволюция представляет собой эволюцию на основе сшитых с субстратом белков (SLiPE). Вектор экспрессии, кодирующий адаптированную рекомбиназу, предпочтительно, представляет собой ретровирусный вектор, лентивирусный вектор, спумавирусный вектор или аденовирусный вектор.
Во втором аспекте настоящее изобретение относится к медицинскому применению вектора экспрессии, кодирующего адаптированную рекомбиназу, или к взрослой стволовой клетке, содержащей указанный вектор экспрессии, для получения фармацевтической композиции, снижающей вирусную нагрузку у индивида, инфицированного ретровирусом. Фармацевтическую композицию можно вводить индивиду для лечения инфекции, вызванной широким спектром ретровирусов, например, таких как ВИЧ. Фармацевтическую композицию по настоящему изобретению, предпочтительно, используют вместе с сопутствующим введением другого активного средства для высокоактивной противоретровирусной терапии (ВААРТ) или для сопутствующего или дополнительного введения при терапии с целью общей активации иммунитета или специфической активации экспрессии генов провируса.
В соответствии с третьим аспектом настоящее изобретение относится к способу оптимизации лечения ретровирусной инфекции у индивида in vitro, причем способ включает стадии:
(a) определения последовательности нуклеиновой кислоты ретровирусной ДНК, находящейся в образце крови пациента;
(b) сканирования последовательности LTR в последовательности стадии (а) на известные последовательности рекомбинации, для которых получены специфические адаптированные рекомбиназы;
(c) в случае если по меньшей мере одна из указанных известных последовательностей рекомбинации существует, то получения соединения, выбранного из группы, состоящей из вектора экспрессии, содержащего нуклеиновую кислоту адаптированной рекомбиназы, специфически распознающей указанную известную последовательность рекомбинации, указанной адаптированной рекомбиназы, слитого белка, содержащего аминокислотную последовательность указанной адаптированной рекомбиназы, или взрослой стволовой клетки, содержащей указанный вектор экспрессии, в виде фармацевтической композиции для снижения вирусной нагрузки у индивида; в противном случае идентификации последовательностей, гомологичных последовательностям левой половины участка и правой половины участка известных участков-мишеней рекомбиназ по меньшей мере на 30%, причем гомологичные последовательности разделены спейсером из 5-12 нуклеотидов, которые обозначают как "ассиметричные последовательности-мишени";
(d) проведение стадий (b)-(l) указанного выше способа получения вектора экспрессии, кодирующего адаптированную рекомбиназу, осуществляющую специфическую рекомбинацию асимметричной последовательности-мишени, идентифицированной в LTR провирусной ДНК; и
(e) получения вектора экспрессии, полученного на стадии (d), белка или слитого белка, экспрессированных с указанного вектора экспрессии, или стволовой клетки, трансфицированной или инфицированной указанным вектором экспрессии в виде фармацевтической композиции, снижающей вирусную нагрузку у индивида.
В предпочтительном варианте осуществления направленную молекулярную эволюцию, используемую на стадии (d), применяют для адаптированной рекомбиназы, уже распознающей участок-мишень, отличающийся от участка-мишени рекомбиназы дикого типа.
В другом предпочтительном варианте осуществления адаптированную рекомбиназу, полученную указанными выше способами, включают в коллекцию специфически адаптированных рекомбиназ.
Изобретение также относится к коллекции адаптированных рекомбиназ, в которой каждая распознает особые участки-мишени в геноме провируса.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторами настоящего изобретения впервые предлагается способ, в котором используется подход комбинаторной эволюции на основе сшитых с субстратом белков для получения адаптированной рекомбиназы, распознающей ассиметричную последовательность в последовательности провируса, интегрированного в геном клеток-хозяев.
Направленная эволюция представляет собой лабораторный способ, используемый для выбранных молекул с получением мутаций и идентификации последующих адаптаций к новым свойствам. Таким образом, этот способ является мощным способом отбора ферментов с измененной специфичностью (Yuan et al., 2005, Johannes & Zhao, 2006). Одним из необходимых условий направленной эволюции является наличие исходного фермента, который обладает остаточной активностью для успешной эволюционной стратегии (Bloom et al., 2005). Маловероятно, что на основании неактивного в отношении субстрата фермента могут быть получены варианты, которые будут активны в отношении мишени с минимальным родством.
Способ по настоящему изобретению также можно использовать для адаптации модифицирующих ДНК белков в целом. Однако рекомбиназы являются предпочтительными. Под термином "белок, модифицирующий ДНК" подразумевают любой белок, активность которого приводит к изменению последовательности или структуры нуклеиновой кислоты. Примеры модифицирующих ДНК белков, подходящих для адаптации способом по настоящему изобретению включают белки, вовлеченные в гомологичную рекомбинацию, экзонуклеазы, ДНК-метилазы, ДНК-лигазы, рестрикционные эндонуклеазы, топоизомеразы, транспозазы и резолвазы.
Хотя возможность адаптированных рекомбиназ, полученных посредством направленной эволюции хорошо известна (например, смотри Collins et al., 2003), однако ни один из способов до настоящего времени не привел к успешному применению свойств рекомбиназы для рекомбинации природной ретровирусной последовательности. В статье Flowers et al. (1997) указано, что Cre может снижать вирусную нагрузку в клетках, если вирус содержит участки loxP дикого типа. В статье Lee et al. (2000) дополнительно показано, что Cre может рекомбинировать последовательность спейсера, выбранную из генома ВИЧ-1. Основной проблемой этих подходов являлось, как правило, отсутствие в LTR ВИЧ-1 симметричных участков-мишеней, которые распознаются природными рекомбиназами, например, такими как рекомбиназа Cre, распознающая симметричный инвертированный повтор длиной 13 п.н. со спейсером 8 п.н. Это ограничение описано в статье авторов Saraf-Levy et al. (2006), которые разработали гетероспецифические рекомбиназы, каждая из которых связывалась с одной половиной последовательности-мишени, отличающейся по последовательности. Авторы показали, что при таких условиях две различные рекомбиназы вместе могут рекомбинировать асимметричный участок-мишень. Однако основным недостатком этого подхода является необходимость наличия в клетке-мишени двух различных рекомбиназ. В совокупности ни один из разработанных до настоящего времени в уровне техники подходов не смог дать одну рекомбиназу, которая могла бы рекомбинировать природную последовательность ретровирусного генома.
Авторы настоящего изобретения были первыми, кто понял, что рекомбиназы можно адаптировать к распознаванию ассиметричных участков-мишеней, отличающихся от природных симметричных участков-мишеней, разделяя субстрат на ряд новых подгрупп с меньшими различиями по сравнению с исходной мишенью и постадийно адаптируя рекомбиназы для распознавания этих подгрупп (смотри фиг.1). Кроме того, в дальнейшем, комбинаторный подход позволит отбирать функциональные молекулы, распознающие в данной последовательности асимметричный участок-мишень. Таким образом, используя новый подход прохождения при направленной молекулярной эволюции через промежуточные субстраты, авторы изобретения могут получать ферменты с сильно отличающейся новой специфичностью к ассиметричным мишеням.
Способ по настоящему изобретению, заключающийся в получении вектора экспрессии, кодирующего адаптированную рекомбиназу, которая осуществляет рекомбинацию асимметричных участков-мишеней в LTR провирусной ДНК, встроенной в геном клетки-хозяина, позволяет получать векторы экспрессии, кодирующие адаптированные рекомбиназы, которые рекомбинируют асимметричные участки-мишени, отличающиеся от участка-мишени рекомбиназы дикого типа. Способ по изобретению включает стадии:
(a) определения последовательности LTR провирусной ДНК путем идентификации в ней последовательностей, гомологичных последовательностям левой половины участка и правой половины участка известных участков-мишеней рекомбиназ по меньшей мере на 30%, при этом гомологичные последовательности разделены спейсером из 5-12 нуклеотидов, а гомологичные последовательности LTR, которые наиболее гомологичны известному участку-мишени, представляют собой ассиметричную последовательность-мишень;
(b) получения двух синтетических последовательностей, где первая синтетическая последовательность соответствует последовательности ассиметричной последовательности-мишени стадии (а), которая гомологична левой половине участка указанного известного участка-мишени плюс последовательность спейсера, и ее обозначают как "последовательность половины участка 1", и где вторая синтетическая последовательность соответствует последовательности спейсера плюс последовательность асимметричной последовательности-мишени стадии (а), которая гомологична правой половине участка, и ее обозначают как "последовательность половины участка 2";
(c) определения нуклеотидов в синтетических последовательностях стадии (b), отличающихся от соответствующих гомологичных последовательностей левой половины участка и правой половины участка известного гомологичного участка-мишени стадии (а);
(d) получения первой подгруппы из двух последовательностей-мишеней на основе синтетических последовательностей стадии (b), где первая последовательность-мишень в первой подгруппе содержит инвертированный повтор, состоящий из последовательности половины участка 1 стадии (а) и последовательность половины участка 1', разделенных последовательностью спейсера, и где вторая последовательность-мишень в первой подгруппе содержит инвертированный повтор, состоящий из последовательности половины участка 2' и последовательность половины участка 2 стадии (b), разделенных последовательностью спейсера, где последовательности половин участков 1' и 2' представляют инвертированные повторы соответствующих последовательностей половин участков 1 и 2 стадии (b);
(e) получение второй подгруппы последовательностей-мишеней на основе последовательностей-мишеней из первой подгруппы стадии (d), где каждую из последовательностей половин участков вместе с соответствующей последовательностью спейсера последовательностей-мишеней в первой подгруппе стадии (d) используют для получения независимой последовательности-мишени второй подгруппы, формируя инвертированный повтор на основании выбранной последовательности половины участка так, что последовательность спейсера разделяет обе последовательности, формируя инвертированный повтор, где последовательности обеих половин участков, полученные из одной из последовательностей-мишеней первой подгруппы стадии (d) при синтезе и до их использования изменяют с получением инвертированного повтора, формирующего полную последовательность-мишень так, что в последовательности левой половины участка часть нуклеотидов, отличающихся от соответствующей гомологичной последовательности половины участка известного участка-мишени стадии (а), замещена природными нуклеотидами, находящимися в известном участке-мишени, а в последовательности правой половины участка оставшиеся нуклеотиды, отличающиеся от соответствующей гомологичной левой половины участка, замещены природными нуклеотидами, находящимися в известном участке-мишени так, что в обеих последовательностях половин участков, происходящей из одной из последовательностей-мишеней первой подгруппы стадии (d), взятых вместе, можно найти все отличающиеся нуклеотиды, тогда как ни одна из указанных последовательностей половин участков отдельно не содержит всех отличающихся нуклеотидов;
(f) получения дополнительных подгрупп последовательностей-мишеней на основе последовательностей-мишеней из второй подгруппы, полученных на стадии (e), посредством пошагового повторения процесса стадии (e), каждый раз получая новую подгруппу последовательностей-мишеней, до тех пор пока последовательности половин участков, формирующие инвертированные повторы в каждой полученной последовательности-мишени содержат один, два или три нуклеотида, отличающихся от соответствующей гомологичной последовательности половины участка известного участка-мишени;
(g) применения направленной молекулярной эволюции для рекомбиназы, распознающей известный гомологичный участок-мишень, выбранный на стадии (a), с использованием в качестве субстрата последовательностей-мишеней из конечной подгруппы, полученной на стадии (f), содержащих один, два или три нуклеотида, отличающихся от соответствующей гомологичной последовательности половины участка указанного известного гомологичного участка-мишени;
(h) перестановки в библиотеках рекомбиназ, полученных на стадии (g);
(i) применения направленной молекулярной эволюции для библиотеки с перестановками, полученной на стадии (h), с использованием последовательностей-мишеней следующей подгруппы более высокого порядка в соответствии со стадией (f);
(j) повторения стадий (h) и (i) по меньшей мере до получения посредством направленной молекулярной эволюции одной рекомбиназы, которая активна в отношении ассиметричной последовательности-мишени в LTR ретровирусной ДНК стадии (а);
(k) выделения из библиотеки нуклеиновой кислоты по меньшей мере одной рекомбиназы, полученной на стадии (j); и
(l) клонирования нуклеиновой кислоты, полученной на стадии (k) в подходящем векторе экспрессии.
На стадии (a) способа по настоящему изобретению определяют последовательность LTR провирусной ДНК, например, посредством секвенирования ДНК с использованием терминирующих цепь ингибиторов (Sanger et al., 1977). Однако если последовательность LTR ретровирусной ДНК, встроенной в геном хозяина, уже определена, то эту стадию можно пропустить. Кроме того, можно использовать известные последовательности, доступные в базах данных последовательностей. На основе информации последовательности проводят компьютерный анализ информации последовательности с идентификацией в ней последовательностей, которые гомологичны по меньшей мере на 30%, соответственно, последовательностям левой половины участка и правой половины участка известных участков-мишеней известных рекомбиназ, которые разделены подходящим спейсером из 5-12 нуклеотидов.
Как используют в настоящем документе термин "рекомбиназа" относится к белку, вовлеченному в рекомбинацию. По существу рекомбиназы распознают и связывают две специфические последовательности ДНК, обозначенные как "участки рекомбинации" или "участки-мишени " и опосредуют рекомбинацию между этими двумя участками-мишенями. Таким образом, под термином "рекомбиназа" понимают любой белковый компонент любой системы рекомбинации, которая опосредует перестановку ДНК в конкретном локусе ДНК. Природные рекомбиназы распознают симметричные участки-мишени, состоящие из двух идентичных последовательностей, обозначенных как "половина участка" приблизительно из 9-20 п.н., формирующих инвертированный повтор, где последовательности половин участков разделены последовательностью спейсера из 5-12 п.н.
Следует отметить, что в настоящем изобретении, а также в данной области термины "последовательность-мишень", "участок-мишень" и "участок рекомбинации" используют взаимозаменяемо.
В отличие от природных рекомбиназ, распознающих симметричные участки-мишени, способ по настоящему изобретению относится к адаптированным рекомбиназам, распознающим непалиндромные участки-мишени, разделенных спейсером. В отличие от природных в последовательностях ассиметричных участков-мишеней не формируется инвертированный повтор. Таким образом, адаптированная рекомбиназа, способная распознавать ассиметричный участок-мишень, может распознавать и рекомбинировать участки-мишени, состоящие из половин участков различной последовательности.
В асимметричном участке-мишени последовательности, обозначаемые как "левая половина участка" и "правая половина участка", соответственно, определены по их гомологии с левой и правой половинами участка известного участка-мишени. Последовательность, расположенную между последовательностями, гомологичными левой и правой половинам участка известного участка-мишени, обозначают как спейсер.
Однако если последовательности расположены в LTR, который гомологичен только последовательности левой или правой половины участка известного участка-мишени, тем не менее эти последовательности можно использовать в практическом осуществлении настоящего изобретения. Размер участка-мишени, с которым взаимодействует рекомбиназа, природная последовательность-мишень которой демонстрирует гомологию с последовательностями в LTR, известна специалисту. Например, если гомология с последовательностью-мишенью, распознаваемой рекомбиназой Cre, находится в последовательности LTR, асимметричный участок-мишень для распознавания рекомбиназой Cre должен состоять из 34 нуклеотидов с двумя последовательностями половин участков из 13 нуклеотидов каждый, разделенных спейсером из 8 нуклеотидов. Таким образом, гомологичная последовательность в LTR в зависимости от гомологии с последовательностью известного участка-мишени определена как левая или правая половина участка или спейсер асимметричного участка-мишени. Таким образом, последовательности с гомологией с левой половиной участка известной последовательности-мишени определены как левая половина участка, последовательности с гомологией с правой половиной участка известной последовательности-мишени определены как правая половина участка. Исходя из этого определения, другие части ассиметричных участков-мишеней определены на основе структуры известного участка-мишени. Таким образом, определив, например, последовательность правой половины участка в LTR по гомологии с участком loxP (распознаваемым рекомбиназой Cre), можно легко определить другие последовательности, соответствующие спейсеру и левой половине участка асимметричной последовательности-мишени. Например, последовательность спейсера определяют, отсчитывая 8 нуклеотидов до 5'-конца последовательности, определенной как последовательность правой половины участка, тогда как последовательность левой половины участка подобным образом определяют, отсчитывая 13 нуклеотидов до 5'-конца ранее определенной последовательности спейсера.
Гомология в данном случае, а также во всей заявке, означает сходство или идентичность последовательностей, предпочительно, идентичность. Предпочтительным сравнением для выявления гомологии является сравнение по меньшей мере двух последовательностей с использованием стандартных известных в данной области способов, включая в качестве неограничивающих примеров, алгоритм локальной гомологии Smith & Waterman (1981), алгоритм выравнивания по гомологии Needleman & Wunsch (1970) или способ поиска сходства Pearson & Lipman (1988). Для целей настоящей заявки, если не указано иначе, гомологию последовательностей, предпочтительно, определяют с использованием компьютерного алгоритма ClustalW, доступного от European Bioinformatics Institute (EBI).
Ввиду необходимости в двух идентичных участках-мишенях, которые должны присутствовать в геноме провируса, чтобы позволить рекомбиназе вырезать последовательность между этими двумя участками-мишенями, на стадии (a) способа по настоящему изобретению сканируют последовательности провирусной ДНК, которые в геноме присутствуют по меньшей мере дважды. Такие последовательности представляют собой, например, последовательности LTR провирусной ДНК. Таким образом, предпочтительно, сканируют последовательность LTR, так как 5'-LTR и 3'-LTR провирусной ДНК являются идентичными. Асимметричный участок-мишень, присутствующий в 5'-LTR, также присутствует в 3'-LTR и, таким образом, позволяет вырезать провирусную ДНК, расположенную между LTR.
Из последовательностей, идентифицированных в последовательности LTR с достаточной гомологией с известными последовательностями участков-мишеней предпочтительно выбирают те, которые обладают наибольшей гомологией с последовательностью участка-мишени известных рекомбиназ. Однако также можно выбирать последовательности, отличные от последовательностей с наибольшей гомологией.
Следует отметить, что возможности способа по настоящему изобретению позволяют адаптировать даже рекомбиназы, которые распознают ассиметричные участки-мишени менее чем с 30% гомологии с известными участками-мишенями. Однако для гарантии наличия остаточной рекомбинационной активности для соответствующего асимметричного участка-мишени, предпочтительно сканировать на последовательности с гомологией с последовательностями левой половины участка и правой половиной участка известных участков-мишеней известных рекомбиназ по меньшей мере 30%. В дополнительных предпочтительных вариантах осуществления следует сканировать на последовательности с гомологией с последовательностями левой половины участка и правой половины участка известных участков-мишеней известных рекомбиназ в размере 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80%, более предпочтительно, 85%, особенно предпочтительно, 90% и, наиболее предпочтительно, 95%.
В предпочтительном варианте осуществления настоящего изобретения последовательность, выбранная в LTR, обладает гомологией с симметричными участками-мишенями loxP, распознаваемыми сайт-специфической рекомбиназой Cre. В более предпочтительном варианте осуществления последовательность асимметричного участка-мишени представляет собой 5'-ACAACATCCTATTACACCCTATATGCCAACATGG-3' (SEQ ID NO:2) (фиг.2A).
На стадии (b) выбранные в LTR последовательности, гомологичные с известным участком-мишенью, используют для получения двух синтетических последовательностей. Эти синтетические последовательности получают в виде олигонуклеотидных последовательностей известными в данной области способами. Последовательность первой синтетической последовательности (обозначаемой как "последовательность половины участка 1") соответствует последовательности выбранного в LTR провируса асимметричного участка-мишени, гомологичной левой половине участка известного участка-мишени, и включает последовательность между последовательностями, представляющими собой левую и правую половину участка, которую обозначают как спейсер. Вторая синтетическая последовательность соответствует последовательности спейсера и последовательности выбранного асимметричного участка-мишени в LTR провируса, гомологичной правой половине участка известного участка-мишени. Эту вторую олигонуклеотидную последовательность обозначают как "последовательность половины участка 2".
Последовательность спейсера в обеих синтетических последовательностях, предпочтительно, идентична и соответствует последовательности LTR, представляющей собой последовательность спейсера асимметричного участка-мишени или определенной как последовательность спейсера асимметричного участка-мишени. Однако в дополнительном варианте осуществления последовательности спейсера обеих синтетических последовательностей содержат одно или два изменения в последовательности. Изменения предпочтительно представляют собой замены нуклеотидов, которые сохраняют исходную длину последовательности спейсера.
На стадии (c) посредством выравнивания последовательностей и сравнения последовательностей в "последовательности половины участка 1" и "последовательности половины участка 2", соответственно, первой подгруппы синтетических последовательностей стадии (b), определяют нуклеотиды, отличающиеся от последовательностей соответствующих гомологичных последовательностей левой половины участка и правой половины участка выбранной известной мишени. В этом контексте, последовательность "последовательности половины участка 1" сравнивают с соответствующей природной половиной участка, которая предпочтительно представляет собой последовательность левой половины участка, тогда как последовательность "последовательности половины участка 2" сравнивают с другой половиной участка, формирующей палиндромный природный участок-мишень, который предпочтительно представляет собой последовательность правой половины участка.
Этот способ не обязательно проводить после стадии (b) и до стадии (d) способа по изобретению, но его также можно проводить на другой стадии способа, после стадии (a) и до стадии (e).
На стадии (d) с использованием синтетических последовательностей стадии (b) получают первую подгруппу из двух последовательностей-мишеней. Последовательности-мишени первой подгруппы представляют собой палиндромные олигонуклеотидные последовательности со структурой симметричного участка-мишени. Эти искусственные симметричные участки-мишени синтезируют на основе последовательностей половин участков стадии (b), дополняя отсутствующую последовательность половины участка в каждой олигонуклеотидной последовательности в виде инвертированного повтора, где для дополнения второй последовательности половины участка с противоположного конца последовательности спейсера используют последовательность "последовательности половины участка 1" и "последовательности половины участка 2", соответственно. Таким образом, первая последовательность-мишень в первой подгруппе (обозначаемая как "последовательность-мишень 1") содержит инвертированный повтор, состоящий из "последовательности половины участка 1" и повторенной с инверсией "последовательности половины участка 1'", разделенных последовательностью спейсера, тогда как вторая последовательность-мишень в первой подгруппе (обозначаемая как "последовательность-мишень 2") содержит инвертированный повтор, состоящий из повторенной с инверсией "последовательности половины участка 2'" и "последовательности половины участка 2", разделенных последовательностью спейсера (смотри фиг.1). В "последовательности-мишени 1" последовательность расположена следующим образом: 5'-"последовательность половины участка 1"-спейсер-"последовательность половины участка 1'"-3', в "последовательности-мишени 2" последовательность расположена следующим образом: 5'-"последовательность половины участка 2'"-спейсер-"последовательность половины участка 2"-3'.
Подобно тому, что указано выше на стадии (b), последовательности спейсера в каждой из двух синтетических последовательностях-мишенях первой подгруппы предпочтительно являются идентичными и соответствуют последовательности LTR, представляющей собой последовательность спейсера ассиметричного участка-мишени или определенной как последовательность спейсера ассиметричного участка-мишени. Однако в дополнительном варианте осуществления последовательности спейсера могут содержать одно или два изменения в последовательности, возникающие в результате замен нуклеотидов.
Как правило, эта стадия представляет собой первое разделение последовательностей ассиметричного участка-мишени, выбранных для адаптации конкретной рекомбиназы (смотри фиг.1). Последовательности на этой стадии получают, располагая симметричные участки-мишени, полученные из половин участков ассиметричного участка-мишени, выбранного для адаптации конкретной рекомбиназы (смотри фиг.1). Как следствие, каждая мутация (т.е. различие с участком-мишенью, распознаваемым рекомбиназой дикого типа), находящаяся на одной половине участка указанного ассиметричного участка-мишени, теперь распределена между симметричными последовательностями-мишенями в первой подгруппе.
На стадии (e) способа по настоящему изобретению последовательности-мишени первой подгруппы используют для получения второй подгруппы последовательностей-мишеней. Каждую из последовательностей половин участков и соответствующих последовательностей-мишеней спейсера в первой подгруппе стадии (d) независимо используют для получения новой последовательности-мишени (т.е. формируя вторую подгруппу), образуя инвертированный повтор на основе выбранной последовательности половины участка так, что последовательность спейсера разделяет обе последовательности, формируя инвертированный повтор (смотри фиг.1). Таким образом, последовательность левой половины участка "последовательности-мишени 1" используют для получения первого нового симметричного участка-мишени, последовательность правой половины участка "последовательности-мишени 1" используют для получения второго нового симметричного участка-мишени, последовательность левой половины участка "последовательности-мишени 2" используют для получения третьего нового симметричного участка-мишени и, наконец, последовательность правой половины участка "последовательности-мишени 2" используют для получения четвертого нового симметричного участка-мишени, где каждый раз в новый симметричный участок-мишень включают последовательность спейсера исходной последовательности-мишени (смотри фиг.1). При этом, что является крайне важным, последовательности половин участков, полученные из каждой из последовательностей-мишеней первой подгруппы, при синтезе и перед их использованием для получения инвертированного повтора изменяют, что дает полностью новую последовательность-мишень (например, такую как при проведении замен нуклеотидов). В последовательности левой половины участка новой последовательности-мишени (происходящей из последовательности-мишени первой подгруппы) часть нуклеотидов, отличающихся от соответствующей гомологичной последовательности половины участка известной последовательности-мишени стадии (а) заменяют природными нуклеотидами, находящимися в известной последовательности-мишени, а в соответствующей последовательности правой половины участка (происходящей из той же последовательности-мишени первой подгруппы) оставшиеся нуклеотиды, отличающиеся от соответствующей гомологичной левой половины участка, заменяют природными нуклеотидами, находящимися в известной последовательности-мишени (смотри фиг.1). Нуклеотиды, измененные в левой половине участка, отличаются от нуклеотидов, измененных в правой половине участка, так, что отличающиеся нуклеотиды присутствуют только на одной из двух последовательностях половин участков, происходящих из одной последовательности-мишени первой подгруппы: или в последовательности-мишени, формируемой на основе левой половины участка, или в последовательности-мишени, формируемой на основе правой половины участка (смотри фиг.1). Кроме того, изменение последовательности проводят так, что каждый отличающийся нуклеотид один раз сохраняется в одной из половин участка. Наконец, ни одна из половин последовательностей половин участков в одиночку не должна содержать все отличающиеся нуклеотиды.
Как уже указано выше на стадиях (b) и (d), последовательности спейсера в каждой из двух синтетических последовательностях-мишенях новой подгруппы, происходящие из синтетической последовательности-мишени подгруппы более высокого порядка, предпочтительно, являются идентичными и соответствуют последовательности LTR, представляющей собой последовательность спейсера ассиметричного участка-мишени или определенной как последовательность спейсера ассиметричного участка-мишени. Однако в дополнительном варианте осуществления последовательности спейсера могут содержать одно или два изменения в последовательности, возникающие в результате замен нуклеотидов.
С используется этого подхода, количество мутаций (т.е. различий с участком-мишенью, распознаваемым рекомбиназой дикого типа) в последовательностях-мишенях, представляющих каждую подгруппу, является меньшим, чем в исходной ассиметричной последовательности-мишени, но все мутации все еще представлены в одной из последовательностей-мишеней (смотри фиг.1).
Как используется в настоящем документе термин "отличающийся нуклеотид" относится к нуклеотиду в асимметричной последовательности-мишени, идентифицированному или определенному в LTR или в последовательности-мишени из подгруппы, полученной по настоящему изобретению, который отличается (т.е. является другим) от нуклеотида, присутствующего в том же положении в соответствующей гомологичной последовательности известной гомологичной симметричной последовательности-мишени известной рекомбиназы, выбранной на стадии (a) способа по настоящему изобретению. В данном контексте термины "отличающиеся нуклеотиды" и "мутации" используют взаимозаменяемо.
Пример получения подгрупп последовательностей-мишеней в общих чертах представлен на фиг.1. Более конкретный пример на основе асимметричной последовательности в LTR провируса ВИЧ представлен на фиг.2A.
На стадии (f) на основе последовательностей-мишеней второй подгруппы получают дополнительные подгруппы последовательностей-мишеней посредством пошагового повторения процесса стадии (e), т.е., разделяя последовательности-мишени на соответствующие последовательности половин участков и получая на основе этих последовательностей половин участков после изменения последовательности половины участка, полученной из последовательности-мишени второй подгруппы новые палиндромные структуры, каждый раз получая новую подгруппу последовательностей-мишеней, до тех пор, пока последовательности половин участков, используемые для получения инвертированных повторов, содержат один, два или три нуклеотида, отличающиеся от соответствующей гомологичной последовательности половины участка известного участка-мишени.
Авторы настоящего изобретения обнаружили, что рекомбиназы можно адаптировать с использованием направленной молекулярной эволюции с использованием в качестве субстрата последовательностей-мишеней, если последовательность-мишень, используемая в качестве субстрата, отличается от природной последовательности-мишени не более чем на 3 нуклеотида. Таким образом, получение подгрупп различных порядков, описанное выше, служит для уменьшения количества отличающихся нуклеотидов на последовательность-мишень до 3 или менее (смотри фиг.1). Пошаговое уменьшение количества отличающихся нуклеотидов, в конечном счете, дает ряд подгрупп последовательностей-мишеней различных порядков с уменьшенными количествами отличающихся нуклеотидов, с получением конечной подгруппы, которую можно использовать в качестве субстрата для направленной молекулярной эволюции. При получении различных подгрупп и, таким образом, уменьшении количества отличающихся нуклеотидов, различия с участком-мишенью, распознаваемым рекомбиназой дикого типа, распределены между несколькими последовательностями-мишенями, которые не содержат более 3 из этих отличающихся нуклеотидов каждая, хотя при этом последовательности-мишени конечного порядка, рассматриваемые в целом, все еще представляют все отличающиеся нуклеотиды (смотри фиг.1).
На основе второй подгруппы последовательностей-мишеней, полученных на стадии (e), получают третью подгруппу с последующей четвертой, пятой, шестой и тому подобное подгруппой, если необходимо. Однако если последовательности-мишени второй подгруппы по-прежнему содержат более трех отличающихся нуклеотидов, необходимым, как правило, является получение только третьей подгруппы. Этот же подход используют при получении последующих подгрупп, которые являются необходимыми только тогда, когда у последовательности-мишени предыдущей подгруппы сохраняются более трех отличающихся нуклеотидов. Следует отметить, что в одном из вариантов осуществления подгруппы последовательностей-мишеней получают до тех пор, пока последовательности-мишени конечной подгруппы не будут содержать один отличающийся нуклеотид. Таким образом, в зависимости от количества отличающихся нуклеотидов в каждой последовательности половины участка, количество подгрупп, полученных для каждой последовательности половины участка асимметричного участка-мишени, может отличаться. Например, для распределения отличающихся нуклеотидов между несколькими последовательностями-мишенями так, чтобы одна последовательность-мишень не содержала более 3 из этих отличающихся нуклеотидов, для последовательности левой половины участка необходимым может являться получение только двух подгрупп, тогда как для правой половины участка нужно получить три или четыре подгруппы.
Правило получения дополнительных подгрупп последовательностей-мишеней для уменьшения количества отличающихся нуклеотидов до количества меньшего трех проиллюстрировано на фиг.1.
На стадии (g) для рекомбиназы, распознающей известный гомологичный участок-мишень стадии (а), используют способ направленной молекулярной эволюции с использованием в качестве субстрата полученных на стадии (f) последовательностей-мишеней конечной подгруппы, содержащих один, два или три нуклеотида, отличающихся от соответствующей гомологичной последовательности половины участка указанного известного гомологичного участка-мишени.
Как используется в настоящем документе термин "конечная подгруппа" относится к последней подгруппе, полученной на стадии (f). В зависимости от количества отличающихся нуклеотидов в асимметричном участке-мишени и количества подгрупп, которые необходимо получить для уменьшения количества отличающихся нуклеотидов на последовательность-мишень меньше 3, "конечная подгруппа" может соответствовать любой подгруппе, например второй, третьей, четвертой или более поздней подгруппе и может отличаться для последовательностей половин участков асимметричной последовательности-мишени в LTR.
Способы направленной молекулярной эволюции, также называемые лабораторной эволюцией или эволюцией in vitro, хорошо известны в данной области (для обзора смотри Yuan et al., 2005 и ссылки в ней; Johannes & Zhao, 2006).
На первой стадии направленной молекулярной эволюции известными в данной области способами получают библиотеки последовательностей случайным образом мутированных рекомбиназ, например, посредством ПЦР пониженной точности и перестановки ДНК (рассмотрено, например в Yuan et al., 2005), или способами, описанными в международной патентной заявке WO 02/44409. Плазмиды каждой библиотеки, содержащей мутантную рекомбиназу, также содержат одну из последовательностей-мишеней конечной подгруппы, полученной на стадии (f). После трансфекции полученной плазмидной библиотеки в подходящие клетки, рекомбиназе позволяют экспрессироваться и проводят направленную молекулярную эволюцию, как известно специалисту в данной области.
В предпочтительном варианте осуществления направленная молекулярная эволюция, используемая на стадии (g) способа по настоящему изобретению, представляет собой эволюцию на основе сшитых с субстратом белков (SLiPE; Buchholz & Stewart, 2001; международная патентная заявка WO 02/44409). Эволюцию на основе сшитых с субстратом белков проводят как подробно описано в примерах. В кратком изложении последовательности-мишени, полученные на стадии (f), клонируют в плазмиде (так называемый эволюционный вектор) вместе со случайным образом мутированной кодирующей последовательностью рекомбиназы. Случайные мутации вносят посредством ПЦР с пониженной точностью (смотри Buchholz & Stewart, 2001). Затем полученную плазмидную библиотеку трансфицируют в клетки E. coli для последующей экспрессии рекомбиназы. Уровни экспрессии можно изменять с использованием индуцибельного промотора, направляющего экспрессию рекомбиназы. После инкубации в течение ночи плазмидную ДНК выделяют из клеток и обрабатывают NdeI с расщеплением плазмид, которые не рекомбинировали и затем только рекомбинированные плазмиды амплифицируют с праймерами. Продукт ПЦР рекомбинированной формы плазмиды дает полосу 1,7 т.п.н. Продукт ПЦР расщепляют ферментами BsrGI и XbaI и субклонируют обратно в сходным образом расщепленном эволюционном векторе для следующего цикла эволюции.
На стадии (h) библиотеки рекомбиназ, полученные на стадии (g), комбинируют и подвергают перестановкам. Технология перестановки ДНК известна в данной области (для обзора смотри Minshull & Stemmer, 1999; Stemmer, 1994).
Затем комбинированную и подвергнутую перестановкам библиотеку клонируют в новом поколении векторов, содержащих последовательности-мишени следующей подгруппы более высокого порядка, полученной на стадии (f).
Как используется в настоящем документе термин "следующая подгруппа более высокого порядка" относится к подгруппе, используемой для получения некоторой подгруппы. Например, третья подгруппа получена на основе последовательностей-мишеней второй подгруппы. Таким образом, начиная с третьей подгруппы "следующей подгруппой более высокого порядка" будет вторая подгруппа. Кроме того, следующей подгруппой более высокого порядка, начиная от второй подгруппы, будет первая подгруппа. Это же используют для третьей, четвертой (и так далее) подгрупп, и следующей подгруппой более высокого порядка, начиная с этой первой подгруппы, является ассиметричная последовательность-мишень с последовательностью из LTR.
На стадии (i) способ направленной молекулярной эволюции применяют для библиотеки с перестановками, полученной на стадии (h), с использованием последовательности-мишени следующей подгруппы более высокого порядка в соответствии со стадией (f). На этой стадии можно использовать тот же способ направленной молекулярной эволюции как и способ направленной молекулярной эволюции, используемый ранее на стадии (g), но также на этой стадии способа по настоящему изобретению можно использовать другой способ направленной молекулярной эволюции. Примеры других способов направленной молекулярной эволюции описаны, например, в Yuan et al. (2005).
Предпочтительно, способ эволюции на основе сшитых с субстратом белков также используют для комбинированных или подвергнутых перестановкам библиотек.
Эта стадия дает рекомбиназы, распознающие и рекомбинирующие последовательности-мишени, несущие комбинацию (и таким образом, увеличенное количество) мутаций из различных последовательностей-мишеней подгруппы более низкого порядка. Авторы настоящего изобретения были первыми, кто показал, что комбинация мутаций из различных библиотек подгруппы последовательностей-мишеней более низкого порядка приводит к синергическому действию и приводит к получению рекомбиназ, которые теперь рекомбинируют последовательности-мишени подгруппы более высокого порядка, демонстрируя, что стратегию эволюции перехода через промежуточные продукты можно использовать для достижения желаемой активности.
На стадии (j) повторяют стадии (h), т.е. комбинирования и перестановки библиотек рекомбиназ, и (i), т.е. применения направленной молекулярной эволюции для комбинированных и подвергнутых перестановки библиотек, до тех пор, пока получают, по меньшей мере, одну рекомбиназу, которая активна в отношении ассиметричной последовательности-мишени, присутствующей в LTR провирусной ДНК.
Например, в способе, где для получения последовательностей-мишеней только с одним, двумя или тремя нуклеотидными изменениями необходимо получение трех подгрупп последовательностей-мишеней, комбинируют и подвергают перестановкам библиотеки рекомбиназ, подвергнутые эволюции, например, для третьей подгруппы последовательностей-мишеней и для этой библиотеки с перестановками применяют направленную молекулярную эволюцию с использованием последовательностей-мишеней второй подгруппы. Затем подвергнутые таким образом эволюции библиотеки рекомбиназ комбинируют и подвергают перестановкам и для этой библиотеки применяют направленную молекулярную эволюцию с использованием последовательностей-мишеней первой подгруппы. На следующей (конечной) стадии для эволюции посредством направленной молекулярной эволюции библиотеки рекомбиназ, содержащей рекомбиназы, распознающие последовательности-мишени из первой подгруппы, используют асимметричную последовательность-мишень в LTR провирусной ДНК стадии (а) с получением, по меньшей мере, одной рекомбиназы, которая активна в отношении ассиметричной последовательности-мишени в LTR ретровирусной ДНК.
На этой стадии способ направленной молекулярной эволюции предпочтительно представляет собой способ эволюции на основе сшитых с субстратом белков.
На стадии (k) из библиотеки выделяют нуклеиновую кислоту по меньшей мере одной рекомбиназы с активностью в отношении асимметричной последовательности-мишени в LTR ретровирусной ДНК стадии (а). Нуклеиновую кислоту выделяют из соответствующей плазмиды в библиотеке с использованием соответствующих рестрикционных ферментов. Способ расщепления рестрикционными эндонуклеазами известен специалистам в данной области. Затем нуклеиновую кислоту, кодирующую рекомбиназу, можно выделить известными способами, такими как электрофорез в геле.
Нуклеиновую кислоту можно хранить (предпочтительно, при температурах ниже -80°C) можно, необязательно, клонировать на стадии (l) в векторе экспрессии для использования в дальнейшем анализе, в способах экспрессии белка или для введения индивиду для лечения СПИДа. Подходящие векторы экспрессии определены ниже.
Полученные в результате рекомбиназы, предпочтительно, тестируют в клетках млекопитающих, чтобы убедиться в их активности в среде клеток млекопитающих. Кроме того, для получения удовлетворительной экспрессии в клетках млекопитающих рекомбиназы можно оптимизировать для экспрессии в этих клетках (например, оптимизация использования кодонов с использованием хорошо известных в данной области способов. Например, смотри Shimshek et al., 2002), или к нуклеиновой кислоте адаптированной рекомбиназы можно добавлять сигнальные последовательности, необходимые для направления белка в ядро клетки млекопитающего, такие как последовательность NLS (Macara, 2001).
В предпочтительном варианте осуществления настоящего изобретения известная рекомбиназа, последовательность-мишень которой используют на стадии (a) и к которой на стадиях (g) и (i) применяют направленную молекулярную эволюцию, относится к семейству сериновых интеграз. Предпочтительные рекомбиназы, принадлежащие семейству сериновых интеграз выбраны из группы, состоящей из интегразы phiC31 (Combes et al., 2002), любого компонента систем рекомбинации Gin или Hin, резолвазы Tn3 (Krasnow & Cozzarelli, 1983) или любого другого представителя больших сериновых рекомбиназ, Rag1, Rag2 или другого компонента системы рекомбинации VDJ или их вариантов.
В другом предпочтительном варианте осуществления указанная рекомбиназа относится к семейству тирозиновых интеграз. Предпочтительные рекомбиназы, относящиеся к семейству тирозиновых интеграз, выбраны из группы, состоящей из Cre фага P1 (Abremski et al., 1983, 1984), рекомбиназы FLP дрожжей (Volkert & Broach, 1986), Dre фага D6 (Sauer & McDermott, 2004), рекомбиназы R плазмиды Zygosaccharomyces rouxii pSR1, рекомбиназы A плазмиды Kluveromyces drosophilarium pKD1, рекомбиназы плазмиды Kluveromyces waltii pKW1, Tnp1 транспозона Bacillus Tn4430 любого компонента системы рекомбинации λ Int или их вариантов.
Рекомбиназы семейства тирозиновых интеграз характеризуются наличием в качестве нуклеофила активного центра, используемого для расщепления ДНК, тирозина, тогда как рекомбиназы из семейства сериновых интеграз вместо тирозина используют серин.
В данном контексте термин «вариант» относится к белкам, которые получены из указанных выше белков посредством делеции, замены и/или добавления аминокислот и которые сохраняют некоторые или все из функций, свойственных белку, из которого они получены.
В предпочтительном варианте осуществления известная рекомбиназа представляет собой химерную рекомбиназу, полученную, например, посредством "перестановки в семействе" как описано в Crameri et al. (1998). Необходимым условием для использования перестановки в семействе является значительная гомология между рекомбиназами, используемыми для получения химерных рекомбиназ. Примером химерной рекомбиназы, которую можно использовать в настоящем изобретении, является химерная рекомбиназа, состоящая из последовательностей рекомбиназы Cre и рекомбиназы Dre, соответственно.
В более предпочтительном варианте осуществления рекомбиназа представляет собой рекомбиназу Cre, распознающую симметричный участок-мишень из 34 п.н., известный как loxP. Участок loxP (а также другие участки рекомбинации рекомбиназ дикого типа) является палиндромным с двумя повторами из 13 п.н., разделенными восемью центральными парами оснований, которые представляют собой так называемый спейсер, который задает направленность участка. Рекомбинация происходит посредством расщепления в последовательности спейсера. В зависимости от относительного расположения и ориентации двух принимающих участие участков loxP, Cre катализирует интеграцию, вырезание или перестановку ДНК (Hoess & Abremski, 1985).
Для вырезания провирусной ДНК из генома клетки-хозяина ассиметричная последовательность-мишень, идентифицированная на стадии (a), предпочтительно, расположена в 5'-LTR и 3'-LTR провируса.
Авторы настоящего изобретения в последовательности LTR ДНК провируса ВИЧ идентифицировали последовательность, которая на 50% гомологична участку loxP. Эту последовательность обозначили как loxLTR (SEQ ID NO:1) и она лежит в LTR штамма первичного ВИЧ-1 TZB0003 (Blackard et al., 1999), являясь частью его модулирующей области U3 (положение в последовательности от -262 до -229; где участок инициации транскрипции представляет собой +1). Участок loxltr представляет собой ассиметричную последовательность длиной 34 п.н., которая на 50% сходна последовательности с последовательностью участка loxP с четырьмя несоответствиями в левом элементе, шестью в правом элементе и полностью отличающимся спейсером (смотри фиг.2A).
Используя связанную с субстратом направленную эволюцию и последовательности loxLTR в качестве субстрата авторы настоящего изобретения смогли получить адаптированную рекомбиназу, которая рекомбинирует эту асимметричную последовательность-мишень ДНК, присутствующую в длинном концевом повторе ВИЧ-1. Впервые была получена единственная родственная Cre сайт-специфическая рекомбиназа, которая рекомбинирует асимметричный участок-мишень. В отличие от рекомбиназ, известных в данной области, рекомбиназа по настоящему изобретению распознает участки-мишени, которые являются ассиметричными и очень отличаются от исходного участка-мишени природной рекомбиназы Cre. Как показано авторами настоящего изобретения, получение адаптированных рекомбиназ, которые специфически направлены к асимметричным последовательностям в LTR ВИЧ-1, позволяет вырезать соответствующий провирус из участка его интеграции в хромосому.
Однако специалисту в данной области очевидно, что могут быть получены другие адаптированные сайт-специфические рекомбиназы, которые рекомбинируют апериодические участки-мишени, находящиеся в геноме ретровирусного провируса, встроенного в геном клетки-хозяина. Например, последовательности-кандидаты можно определять на основе гомологии с существующей последовательностью loxLTR. Примеры таких последовательностей LTR с гомологией с последовательностью loxLTR, полученных из LTR других штаммов ВИЧ, представлены на фиг.2A.
Провирусная ДНК, встроенная в геном клетки-хозяина, предпочтительно, представляет собой ДНК ретровируса. Ретровирусы включают большое и разнообразное семейство оболочечных РНК-содержащих вирусов. Отличительным свойством семейства является его стратегия репликации, которая включает в виде обязательных стадий обратную транскрипцию вирусной РНК в линейную двухцепочечную ДНК и последующую интеграцию этой ДНК (провирусной ДНК) в геном клетки-хозяина. Ретровирусы подразделяют на семь групп, определенных по эволюционному родству. Пять из этих групп (ретровирус альфа, бета, дельта, эпсилон и гамма) представляют ретровирусы с онкогенным потенциалом, а другие две группы представляют собой лентивирусы и спумавирусы. Патогенные для человека вирусы T-клеточного лейкоза человека типа I и типа II (HTLV-I и HTLV-II) принадлежат группе дельта-ретровирусов, тогда как вирусы СПИД, вирусы иммунодефицита человека типа 1 и типа 2 (ВИЧ-1 и ВИЧ-2), принадлежат группе лентивирусов (для обзора смотри стандартный учебник "Retroviruses" Coffin JM, Hughes SH, Varmus HE (Eds.) 1997, Cold Spring Harbor Laboratory Press, New York).
В предпочтительном варианте осуществления провирусная ДНК, встроенная в геном клетки-хозяина представляет собой ДНК ретровируса, выбранного из группы, состоящей из вируса опухоли молочной железы мыши (MMTV), вируса обезьян Мэйзена-Пфейзера (MPMV), вируса T-клеточного лейкоза человека типа I (HTLV-I), вируса T-клеточного лейкоза человека типа II (HTLV-II), вируса T-клеточного лейкоза обезьян типа I (STLV-I), вируса T-клеточного лейкоза обезьян типа II (STLV-II), вируса лейкоза коров (BLV), вируса лейкоза кошек (FeLV) и вируса лейкоза мышей Молони (MoMLV).
В дополнительном предпочтительном варианте осуществления ретровирус представляет собой лентивирус, выбранный из группы, состоящей из вируса иммунодефицита человека типа 1 (ВИЧ-1), вируса иммунодефицита человека типа 2 (ВИЧ-2), вируса иммунодефицита обезьян (SIV), вируса иммунодефицита кошек (FIV), вируса иммунодефицита коров (BIV), вируса Maedi-visna (MVV), вируса инфекционной анемии лошадей (EIAV) и вируса артрита-энцефалита коз (CAEV).
В более предпочтительном варианте осуществления асимметричная последовательность-мишень, идентифицированная на стадии (a) способа по настоящему изобретению расположена в 5'-LTR и в 3'-LTR провируса ВИЧ. Предпочтительно, указанная ассиметричная последовательность-мишень, расположенная в 5'-LTR и в 3'-LTR провируса ВИЧ, обладает последовательностью, приведенной как SEQ ID NO:1. Эту последовательность обозначают как loxltr.
В предпочтительном варианте осуществления способ направленной молекулярной эволюции, используемый в способе по настоящему изобретению, представляет собой способ эволюции на основе сшитых с субстратом белков (SLiPE; Buchholz & Stewart, 2001; смотри также WO 02/44409).
В способе по настоящему изобретению нуклеиновая кислота, кодирующая по меньшей мере одну адаптированную рекомбиназу, которая активна в отношении асимметричной последовательности-мишени в LTR ретровирусной ДНК, клонируют в векторе экспрессии. Векторы экспрессии представляют собой генетические конструкции для экспрессии белков, кодируемых нуклеиновыми кислотами в векторе. Такие векторы экспрессии могут представлять собой либо самореплицирующиеся внехромосомные векторы, либо векторы, которые интегрируются в геном хозяина. Как правило, такие векторы экспрессии содержат регулирующую транскрипцию и трансляцию нуклеиновую кислоту, функционально связанную с нуклеиновой кислотой, кодирующей адаптированную рекомбиназу по настоящему изобретению.
Термин "контрольные последовательности" относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательность в конкретном организме-хозяине. Контрольные последовательности, подходящие для эукариот, включают, например, промотор, необязательно, последовательность оператора и участок связывания рибосомы. Известно, что в эукариотических клетках используют промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота является "функционально связанной", когда ее размещают в функциональной связи с другой последовательностью нуклеиновой кислоты. Например, промотор или энхансер функционально связаны с кодирующей последовательностью, если они воздействуют на транскрипцию последовательности; или участок связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, чтобы способствовать трансляции. Связывание проводят посредством лигирования по подходящим участкам рестрикции. Если такие участки отсутствуют, используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с состоянием техники. Нуклеиновая кислота, регулирующая транскрипцию и трансляцию, как правило, должна подходить для клетки-хозяина, используемой для экспрессии адаптированной рекомбиназы. В данной области известно множество подходящих векторов экспрессии и подходящих регуляторных последовательностей для множества клеток-хозяев.
Вектор экспрессии, используемый в настоящем изобретении, может представлять собой ретровирусный вектор, лентивирусный вектор, спумавирусный вектор или аденовирусный вектор. Однако в предпочтительном варианте осуществления вектор экспрессии представляет собой лентивирусный вектор, выбранный из группы, состоящей из, полученных из ВИЧ-1, SIV, FIV или EIAV лентивирусных векторов. Лентивирусные векторы описаны, например, в Schambach et al. (2006).
В предпочтительных вариантах осуществления настоящего изобретения вектор экспрессии содержит клеточный, бактериальный, вирусный или гибридный промотор.
В основном, для целей настоящего изобретения, промотор может являться конститутивным или индуцибельным промотором. Кроме того, промоторы могут представлять собой природный промотор, такой как бактериальный, клеточный или вирусный промотор или гибридный промотор. Гибридные промоторы, которые сочетают элементы более чем одного промотора, известны в данной области и могут использоваться в настоящем изобретении. Кроме того, промотор, используемый по настоящему изобретению, также может представлять собой производное природного промотора. Как используется в настоящем документе "производное" природного промотора может представлять собой сочетание цис-действующих элементов, полученных из промоторов или последовательностей различного происхождения или, альтернативно, может быть получен посредством делеции или мутации цис-действующих элементов в конкретном природном промоторе (Edelman et al., 2000; Alper et al., 2006; Hartenbach & Fussenegger, 2006).
В более предпочтительном варианте осуществления настоящего изобретения конститутивный промотор или его производное выбраны или получены из группы, состоящей из промоторов цитомегаловируса, вируса саркомы Рауса, родственных вирусу лейкоза мышей ретровирусов, гена фосфоглицерокиназы, вируса некроза селезенки мышей или фактора элонгации 1 альфа человека.
В дополнительном более предпочтительном варианте осуществления настоящего изобретения, индуцибельный промотор или его производное выбраны или получены из группы, состоящей из LTR или его производных, полученных из лентивирусов, спумавирусов и дельта-ретровирусов.
В данном контексте термин "LTR" относится к 5'- и 3'- длинным концевым повторам провируса с промоторной функцией (для обзора смотри учебник "Retroviruses" (Coffin JM, Hughes SH, Varmus HE (Eds.) 1997, Cold Spring Harbor Laboratory Press, New York)).
Предпочтительно, индуцибельный промотор или его производное выбраны или получены из LTR или его производных, полученных из ВИЧ-1, ВИЧ-2, MVV, EIAV, CAEV, SIV, FIV, BIV, HTLV-I и HTLV-II.
Настоящее изобретение, кроме того, относится к способу получения адаптированной рекомбиназы, где указанный способ включает указанный выше способ получения вектора экспрессии, кодирующего адаптированную рекомбиназу, и дополнительную стадию экспрессии адаптированной рекомбиназы или слитого полипептида, содержащего аминокислотную последовательность указанной адаптированной рекомбиназы, с нуклеиновой кислоты, кодирующей рекомбиназу, встроенной в вектор экспрессии, полученный указанным выше способом получения вектора экспрессии, кодирующего адаптированную рекомбиназу, в подходящей клетке-хозяине.
Экспрессию нуклеиновой кислоты, кодирующей адаптированную рекомбиназу, клонированную в векторе экспрессии в соответствии со стадией (l) способа получения вектора экспрессии, кодирующего адаптированную рекомбиназу, можно проводить, например, с применением систем экспрессии бактерий, насекомых или млекопитающих. Однако также можно использовать другие системы экспрессии, известные в данной области. Способы введения экзогенной нуклеиновой кислоты хозяевам млекопитающим, насекомым или бактериям, а также другим хозяевам, также хорошо известны в данной области, и изменяются в зависимости от используемой клетки-хозяина. Способы включают опосредованную декстраном трансфекцию, осаждение фосфатом кальция, опосредованную полибреном трансфекцию, слияние протопластов, электропорацию, заражение вирусом, инкапсуляцию полинуклеотида(ов) в липосомы и прямую микроинъекцию ДНК в ядра.
Слитые белки получают хорошо известными в данной области способами. Например, вектор экспрессии, в который клонируют нуклеиновую кислоту, кодирующую адаптированную рекомбиназу, уже содержит нуклеиновую последовательность, кодирующую второй полипептид или белок. Клонируя нуклеиновую кислоту, кодирующую адаптированную рекомбиназу в рамку с последовательностью второго полипептида или белка, обе последовательности экспрессируют в виде слитого белка.
Клетки-хозяева, используемые для экспрессии адаптированной рекомбиназы с вектора экспрессии, предпочтительно, представляют, собой клетки-хозяева, включающие прокариотические клетки, например, такие как бактериальные клетки или дрожжевые клетки, или эукариотические клетки, например, такие как клетки насекомых или клетки млекопитающих.
Настоящее изобретение дополнительно относится к способу получения трансформированной взрослой стволовой клетки, где указанный способ включает указанный выше способ получения вектора экспрессии, кодирующего адаптированную рекомбиназу, и дополнительную стадию введения вектора экспрессии, полученного указанным выше способом получения вектора экспрессии, кодирующего адаптированную рекомбиназу, в подходящую взрослую стволовую клетку in vitro.
В дополнительном аспекте настоящее изобретение относится к нуклеиновой кислоте, получаемой указанным выше способом по настоящему изобретению.
Как используется в настоящем документе "нуклеиновая кислота" представляет собой полимерное соединение, состоящее из ковалентно связанных субъединиц, называемых нуклеотидами. Нуклеиновая кислота включает полирибонуклеиновую кислоту (РНК) и полидезоксирибонуклеиновую кислоту (ДНК), которые обе могут являться одноцепочечными или двухцепочечными. ДНК включает кДНК, геномную ДНК, синтетическую ДНК и полусинтетическую ДНК.
В широком аспекте нуклеиновая кислота, полученная способом по настоящему изобретению, представляет собой нуклеиновую кислоту, кодирующую адаптированную рекомбиназу, где адаптированная рекомбиназа рекомбинирует асимметричные последовательности-мишени в ДНК провируса, встроенного в геном клетки-хозяина, что приводит к вырезанию провируса из генома клетки-хозяина, где ассиметричные участки-мишени отличаются от участков-мишеней рекомбиназы дикого типа.
В одном из вариантов осуществления нуклеиновая кислота, полученная способом по настоящему изобретению содержит последовательность нуклеиновой кислоты SEQ ID NO:2, и распознает и рекомбинирует последовательность loxltr, SEQ ID NO:1.
Также в определение нуклеиновой кислоты включены варианты последовательности нуклеиновой кислоты, полученной способом по настоящему изобретению, которые обладают по меньшей мере 50% сходством, предпочтительно, по меньшей мере 60% сходством, более предпочтительно, по меньшей мере 70% сходством, еще более предпочтительно, по меньшей мере 80% сходством, и наиболее предпочтительно, по меньшей мере 90% сходством с последовательностью нуклеиновой кислоты SEQ ID NO:2. В некоторых вариантах осуществления сходство составляет до 91, 92, 93, 94, 95, 96, 97, 98 или 99%.
В дополнительном аспекте настоящее изобретение также относится к вектору экспрессии, полученному указанным выше способом по настоящему изобретению.
Дополнительным аспектом изобретения является белок адаптированной рекомбиназы, полученный указанным выше способом по настоящему изобретению.
Как применяют в настоящем документе термин "белок" включает белки, полипептиды и пептиды. Как понятно специалистам в данной области, последовательности нуклеиновой кислоты по изобретению можно использовать для получения белковых последовательностей.
В предпочтительном варианте осуществления белок адаптированной рекомбиназы содержит аминокислотную последовательность SEQ ID NO:3. Эту рекомбиназу обозначили как Tre из-за рекомбиназы LTR (LTR recombinase).
Также в вариант осуществления белков адаптированных рекомбиназ включены аминокислотные варианты аминокислотной последовательности SEQ ID NO:3. Предпочтительно варианты обладают последовательностью, сходной по меньшей мере на 70%, более предпочтительно, последовательностью, сходной по меньшей мере на 80%, а наиболее предпочтительно, последовательностью, сходной по меньшей мере на 90% с последовательностью SEQ ID NO:3. В некоторых вариантах осуществления сходство достигает 91, 92, 93, 94, 95, 96, 97, 98 или 99%. В отношении нуклеиновых кислот сходство в данном контексте означает гомологию или идентичность последовательности, с предпочитаемой идентичностью. Это сходство определяют с использованием стандартных известных в данной области способов, которые приведены выше для гомологии нуклеиновой кислоты.
В дополнительном предпочтительном варианте осуществления белок адаптированной рекомбиназы также можно получать в виде слитого полипептида, с использованием хорошо известных в данной области способов. В предпочтительном варианте осуществления белок адаптированной рекомбиназы связывают со вторым полипептидом. Предпочтительно, слитый полипептид получают указанным выше способом по настоящему изобретению, где адаптированную рекомбиназу связывают со вторым полипептидом.
Предпочтительно, белок адаптированной рекомбиназы получают в виде слитого полипептида для увеличения экспрессии. В дополнительном предпочтительном варианте осуществления белок адаптированной рекомбиназы получают в виде слитого полипептида для обеспечения введения полипептида в живые клетки. Как правило, очищенные белки не могут проникать в клетки, так как они не способны проходить через клеточные мембраны вследствие их размера. Однако слияние с белками специфических пептидных последовательностей может приводить к захвату этих слитых белков клетками. Затем в клетке белок может осуществлять свое действие. Посредством этого подхода сайт-специфические рекомбиназы, включающие рекомбиназу Cre, успешно доставляли в клетки (Peitz et al., 2002). Проходящая в клетки рекомбиназа дополнительно описана в Nolden et al. (2006) и Lin et al. (2004). Таким образом, эту стратегию можно использовать для доставки адаптированных рекомбиназ в клетки для удаления провируса из инфицированных клеток.
Таким образом, второй полипептид в слитом полипептиде может содержать сигнальный пептид. Сигнальный пептид может представлять собой домен белковой трансдукции, такой как пептид TAT или пептид из третьей спирали гомеодомена Antennapedia (Derossi et al., 1994, 1996; Vives et al., 1997; Vives, 2003; Richard et al., 2005) или NLS (последовательность ядерной локализации) для доставки слитого полипептида в ядро эукариотической клетки (Macara, 2001).
Дополнительный аспект настоящего изобретения относится к взрослой стволовой клетке, полученной указанным выше способом получения трансформированной взрослой стволовой клетки по настоящему изобретению. Стволовые клетки, предпочтительно, инфицируют или трансфицируют вектором экспрессии по изобретению.
В предпочтительном варианте осуществления взрослая стволовая клетка представляет собой стволовую клетку из гемопоэтической линии дифференцировки, экспрессирующую адаптированную рекомбиназу, указанный выше слитый полипептид или содержащую указанный выше вектор экспрессии.
Гемопоэтические стволовые клетки (HSC) представляют собой происходящие из костного мозга CD34+ клетки, которые можно очищать из периферической крови доноров с мобилизацией G-CSF (например, инфицированные ВИЧ пациенты) посредством обычного лейкафереза (Scherr & Eder, 2002). Затем генетически модифицированные in vitro клетки реинфузируют пациентам.
В настоящее время в данной области термин "стволовые клетки" означает клетки, которые (a) обладают способностью к самоподдержанию и (b) способностью формировать, по меньшей мере, один, а чаще ряд специализированных типов клеток, вследствие их способности к ассиметричному делению (Donovan & Gearhart, 2001). Взрослые стволовые клетки можно выделять из различных тканей взрослого, т.е. у различных индивидуумов. Такие стволовые клетки в настоящее время в данной области обозначают как "мультипотентные взрослые стволовые клетки". Основное отличие между эмбриональными плюрипотентными стволовыми клетками и взрослыми мультипонентными стволовыми клетками заключается в количестве дифференцированных тканей, которые можно получать из соответствующих клеток.
В дополнительном предпочтительном варианте осуществления настоящее изобретение относится к первичным CD4+ клеткам (клетки крови) ВИЧ-инфицированных пациентов.
В дополнительной стадии способа по настоящему изобретению вектор экспрессии, содержащий последовательность нуклеиновой кислоты, кодирующую адаптированную рекомбиназу, белок рекомбиназы, слитый белок или взрослую стволовую клетку, полученную способами по настоящему изобретению получают в виде фармацевтической композиции для снижения вирусной нагрузки у индивида, инфицированного ретровирусом.
Еще одним объектом изобретения является фармацевтическая композиция, полученная указанным выше способом. Фармацевтическая композиция, предпочтительно, находится в форме раствора, подходящего для внутривенного применения (вливания).
Фармацевтический препарат может дополнительно содержать один или несколько фармацевтически приемлемых носителей, эксципиентов и/или вспомогательных средств. Носители, эксципиенты и вспомогательные средства, подходящие для использования в фармацевтической композиции известны в данной области.
Фармацевтическая композиция по настоящему изобретению при введении индивиду уменьшает вирусную нагрузку у индивида, инфицированного ретровирусом, ниже 5000 эквивалентов генома/мл плазмы, предпочтительно, ниже 500 эквивалентов генома/мл плазмы, а более предпочтительно, ниже 50 эквивалентов генома/мл плазмы.
Таким образом, фармацевтическая композиция по настоящему изобретению, содержащая вектор экспрессии, кодирующий адаптированную рекомбиназу (или адаптированную рекомбиназу в виде белка или слитого полипептида или стволовой клетки, содержащей вектор экспрессии) способна снижать вирусную нагрузку у индивида, инфицированного ретровирусом, устраняя генетический резервуар ретровирусов в клетках-хозяевах, таким образом, предотвращая дальнейшие жизненные циклы вируса.
Как используется в настоящем документе термин "вирусная нагрузка" относится к эквивалентам РНК ВИЧ (например, геномам), которые связаны с 1 мл плазмы пациента (Dybul et al., 2002). Таким образом, вирусную нагрузку определяют посредством измерения содержания вирусной ДНК в образце, взятом у пациента. В настоящее время, существуют три основных доступных типа анализа вирусной нагрузки:
1) обратная транскрипция-полимеразная цепная реакция с РНК ВИЧ (ОТ-ПЦР): Amplicor™ HIV-1 Monitor Test; Roche Diagnostics
2) ответвление цепи ДНК (bDNA): Versant™ HIV RNA Assay; Bayer Diagnostics; и
3) амплификация на основе последовательности нуклеиновой кислоты (NASBA): NucliSens™ Assay; bioMerieux.
В предпочтительном варианте осуществления фармацевтическая композиция по настоящему изобретению способна снижать вирусную нагрузку у индивида, инфицированного ретровирусом, ниже 5000 эквивалентов генома/мл плазмы, предпочтительно, ниже 500 эквивалентов генома/мл плазмы, а более предпочтительно, ниже 50 эквивалентов генома/мл плазмы. Полагают, что пациент с вирусной нагрузкой ниже 5000 эквивалентов генома/мл плазмы относительно хорошо адаптирован к медицинскому лечению. Однако целью современной терапии СПИД является уменьшение вирусной нагрузки ниже предела детекции анализов вирусной нагрузки, который в настоящее время составляет приблизительно ниже 50 эквивалентов генома/мл плазмы.
В данном контексте, авторы настоящего изобретения были первыми, кто показал, что адаптированную рекомбиназу можно использовать для вырезания полноразмерной провирусной ДНК из генома клетки-хозяина млекопитающего без отрицательного воздействия на жизнеспособность клетки (смотри фиг.9). После обработки инфицированных клеток адаптированной рекомбиназой по настоящему изобретению (1) ретровирусная ДНК в геноме хозяина и (2) почкование вирусных частиц из инфицированных клеток более не детектируются, что демонстрирует, что вирусный резервуар в покоящихся клетках разрушен.
Фармацевтическая композиция, предпочтительно, снижает вирусную нагрузку ретровирусов, выбранных из группы, состоящей из вируса опухоли молочной железы мышей (MMTV), вируса обезьян Мэйзена-Пфейзера (MPMV), вируса T-клеточного лейкоза человека типа I (HTLV-I), вирус T-клеточного лейкоза человека типа II (HTLV-II), вирус T-клеточного лейкоза обезьян типа I (STLV-I), вируса T-клеточного лейкоза обезьян типа II (STLV-II), вируса лейкоза коров (BLV), вируса лейкоза кошек (FeLV) и вирус лейкоза мышей Молони (MoMLV).
В дополнительном предпочтительном варианте осуществления ретровирус для лечения фармацевтической композицией по настоящему изобретению представляет собой лентивирус. Указанный лентивирус предпочтительно выбран из группы, состоящей из вируса иммунодефицита человека типа 1 (ВИЧ-1), вируса иммунодефицита человека типа 2 (ВИЧ-2), вируса иммунодефицита обезьян (SIV), вируса иммунодефицита кошек (FIV), вируса иммунодефицита коров (BIV), вируса Maedi-visna (MVV), вируса инфекционной анемии лошадей (EIAV) и вируса артрита-энцефалита коз (CAEV).
Однако специалисту в данной области очевидно, что настоящее изобретение также можно использовать при ретровирусных инфекциях другими ретровирусами, чем те которые указаны выше.
Кроме того, индивид, инфицированный ретровирусом, которому вводят фармацевтическую композицию, выбран из группы, состоящей из людей, приматов, обезьян, крупного рогатого скота, лошадей, коз, овец и домашних кошек. Однако предпочтительно индивид представляет собой человека.
В предпочтительном варианте осуществления фармацевтическая композиция предназначена для совместного введения с другими активными средствами высокоактивной противоретровирусной терапии (ВААРТ).
Высокоактивная противоретровирусная терапия ВААРТ представляет собой комбинированное лечение, направленное на вирусную обратную транскриптазу, протеазу и слияние (Gulick et al., 1997; Lalezari et al., 2003).
В другом предпочтительном варианте осуществления фармацевтическая композиция предназначена для совместного или последовательного введения для лечения, направленного на общую активацию иммунной системы или специфическую активацию экспрессии генов провируса.
Смысл лечения, направленного на активацию иммунной системы основан на гипотезе, что преднамеренная активация латентно инфицированных ВИЧ клеток может ускорить уничтожение персистирующих вирусных резервуаров. Уничтожение происходит посредством иммунной элиминации посредством программируемой гибели клеток, активно экспрессирующих продукты ВИЧ-1 (проапоптотические) (Kulkosky & Bray, 2006). Общей активации иммунной системы (активация иммунных клеток, включая покоящиеся клетки), как правило, достигают, например, посредством введения иммунотоксинов, цитокинов (например, IL-2) или активирующих T-клетки антител (например OKT3).
Ввиду того факта, что активация иммунной системы, проводимая для преднамеренной активации устойчивых к ВААРТ латентных резервуаров, к сожалению потерпела неудачу в отношении окончательного устранения ВИЧ-1 и возобновления связанных с вирусном симптомов (для обзора смотри Kulkosky & Bray 2006; Marcello, 2006; Shehu-Xhilaga et al., 2005) вследствие того факта, что общая активация T-клеток по-видимому также индуцирует репликацию вируса и увеличивает количество потенциальных клеток-мишеней с ВИЧ-1 выше уровня, который может сдерживать ВААРТ (Fraser et al., 2000), для лечения ВИЧ необходимы дополнительные способы специфичного лечения. Один из подходов представляет собой активацию транскрипции неактивных в иных случаях вирусных геномов. Специфической активации генной экспрессии латентных провирусов можно достичь посредством введения форболового эфира простратина или цитокин IL-7 человека, которые оба по-видимому реактивируют латентный ВИЧ-1 в отсутствие клеточной пролиферации (Marcello, 2006). Кроме того, селективной транскрипционной активности ВИЧ-1 также можно достичь посредством ингибиторов гистоновой деацетилазы, таких как, например, вальпроевая кислота, которая в результате индуцирует почкование ВИЧ-1 из покоящихся клеток в отсутствие клеточной активации (Marcello, 2006; Lehrman et al., 2005).
Однако лечение с общей активацией иммунной системы или специфической активацией экспрессия генов провирусов или сходные стратегии лечения получают значительное преимущество при одновременном удалении провирусной ДНК, таким образом, снижая у пациента пул инфицированных клеток.
В дополнительном аспекте настоящее изобретение относится к способу оптимизации лечения ретровирусной инфекции у индивида in vitro.
Способ включает стадии:
(a) определения последовательности нуклеиновой кислоты ретровирусной ДНК, находящейся в образце крови пациента;
(b) сканирования последовательности LTR у последовательности стадии (а) на известные последовательности рекомбинации для которых специфические адаптированные рекомбиназы уже получены;
(c) в случае когда по меньшей мере одна из указанных известных последовательностей рекомбинации существует, получения соединения, выбранного из группы, состоящей из вектора экспрессии, содержащего нуклеиновую кислоту адаптированной рекомбиназы, специфически распознающей указанную известную последовательность рекомбинации, указанной адаптированной рекомбиназы, слитого белка, содержащего аминокислотную последовательность указанной адаптированной рекомбиназы, или взрослой стволовой клетки, где указанный вектор экспрессии получают в виде фармацевтической композиции для снижения вирусной нагрузки у индивида; в противном случае идентификации последовательности с гомологией с последовательностями левой половины участка и правой половины участка известных участков-мишеней рекомбиназ, по меньшей мере, 30%, где гомологичные последовательности разделены подходящим спейсером из 5-12 нуклеотидов, и обозначения ее как "ассиметричной последовательности-мишени";
(d) проведение стадий (b)-(l) указанного выше способа получения вектора экспрессии, для получения вектора экспрессии, кодирующего адаптированную рекомбиназу, осуществляющую специфическую рекомбинацию ассиметричной последовательности-мишени, идентифицированной в LTR провирусной ДНК; и
(e) получения вектора экспрессии, полученного на стадии (d), белка или слитого белка, экспрессированных с указанного вектора экспрессии, или стволовой клетки, трансфицированной или инфицированной указанным вектором экспрессии в виде фармацевтической композиции для снижения вирусной нагрузки у индивида.
В указанном способе предоставлена адаптированная рекомбиназа, использующаяся для лечения пациента, страдающего ретровирусной инфекцией, которая специфически адаптирована для рекомбинации специфических последовательностей-мишеней ретровируса, которым инфицирован пациент. Ввиду разнообразия последовательностей, обнаруженных в последовательностях LTR различных ретровирусов, рекомбиназа, адаптированная для рекомбинации специфической ассиметричной последовательности-мишени конкретного ретровируса не обязательно распознает последовательности-мишени в провирусной ДНК других ретровирусов и, таким образом, является менее эффективной или даже не эффективной для вырезания другой провирусной ДНК, чем провирусная ДНК, для которой она адаптирована. Однако для решения этой проблемы и получения индивидуально адаптированных рекомбиназ для лечения ретровирусной инфекции по существу у каждого индивида, авторы настоящего изобретения предоставляют способ in vitro, позволяющий оптимизировать эффективность рекомбиназы для введения посредством выбора или адаптации рекомбиназы, которая специфически распознает последовательность-мишень в конкретном геноме провируса, которым инфицирован индивид.
На стадии (a) способа оптимизации лечения ретровирусной инфекции in vitro определяют последовательность нуклеиновой кислоты ретровирусной ДНК, присутствующей в образце крови пациента. На данной первой стадии ретровирус, которым инфицирован пациент, предпочтительно, определяют посредством секвенирования ДНК генома вирусных частиц в полученном у пациента образце крови с использованием терминирующих цепь ингибиторов (Sanger et al., 1977). Также можно использовать другие известные специалистам средства, предоставляющие информацию о последовательности ретровируса.
На стадии (b) способа in vitro последовательность LTR, полученную на стадии (a), сканируют на известные последовательности рекомбинации, для которых уже получены специфические адаптированные рекомбиназы. Этот поиск проводят, например, с использованием компьютерного анализа последовательностей, как описано выше.
Если LTR вирусных частиц, находящихся в образце крови, содержит по меньшей мере одну последовательность, соответствующую известной асимметричной последовательности-мишени, для которой адаптированная рекомбиназа уже существует, тогда соединение, выбранное из группы, состоящей из вектора экспрессии, содержащего нуклеиновую кислоту адаптированной рекомбиназы, специфически распознающей указанный известный участок рекомбинации, указанной адаптированной рекомбиназы, слитого белка, содержащего аминокислотную последовательность указанной адаптированной рекомбиназы или взрослой стволовой клетки, трансфицированной или инфицированной вектором экспрессии, как определено выше, получают в виде фармацевтической композиция для снижения вирусной нагрузки у пациента. Это обеспечивает быстрое и индивидуальное лечение конкретной ретровирусной инфекции эффективной рекомбиназой.
Однако если такой адаптированной рекомбиназы ранее получено не было, на стадии (c) последовательность LTR вирусных частиц, найденную в образце крови, используют для идентификации в ней последовательностей с гомологией с последовательностями левой половины участка и правой половины участка известных участков-мишеней рекомбиназы, по меньшей мере 30%, где гомологичные последовательности разделены спейсером из 5-12 нуклеотидов. Как указано выше, такие гомологичные последовательности обозначены как "ассиметричная последовательность-мишень". Подробное описание, предоставленное выше для стадии (a) способа получения вектора экспрессии, кодирующего адаптированную рекомбиназу, также применимо и здесь.
На стадии (d) способа in vitro, проводят стадии (b)-(l) способа получения вектора экспрессии, кодирующего адаптированную рекомбиназу, описанную выше, с получением вектора экспрессии, кодирующего адаптированную рекомбиназу, осуществляющую специфическую рекомбинацию асимметричной последовательности-мишени, идентифицированной в LTR ретровируса, находящегося в образце крови. Это позволяет получить новую рекомбиназу, рекомбинирующую провирусную ДНК ретровируса, от которого страдает пациент, в пределах очень короткого промежутка времени.
Подробное описание, предоставленное выше для стадий (b)-(l) способа получения вектора экспрессии, кодирующего адаптированную рекомбиназу, также применимо и здесь.
В заключение, на стадии (e) способа in vitro полученный вектор экспрессии, кодирующий рекомбиназу, специфически распознающую и рекомбинирующую провирусную ДНК ретровируса, которым инфицирован пациент, получают в виде фармацевтической композиции для снижения вирусной нагрузки у указанного пациента. В данном контексте в дополнительном варианте осуществления фармацевтическая композиция предпочтительно может содержать индивидуально адаптированную рекомбиназу в виде белка или слитого белка, экспрессируемого с вектора экспрессии или в стволовой клетке, трансфицированной или инфицированной указанным вектором экспрессии. В более предпочтительном варианте осуществления фармацевтическая композиция содержит вектор экспрессии, содержащий нуклеиновые кислоты, кодирующие индивидуально адаптированную рекомбиназу.
Рекомбиназу, к которой в указанном выше способе in vitro применяют способ направленной молекулярной эволюции, выбирают из рекомбиназы дикого типа или уже адаптированной рекомбиназы. Однако рекомбиназа предпочтительно представляет собой адаптированную рекомбиназу (распознающую немного отличающуюся от той, которую выявили в LTR данной провирусной ДНК, ассиметричную последовательность-мишень). Выбор зависит от гомологии асимметричных последовательностей-мишеней. Хотя направленную молекулярную эволюцию можно применять для рекомбиназы (дикого типа или адаптированной), распознающей последовательность-мишень с низкой гомологией с последовательностью в LTR провирусной ДНК, это может потребовать дополнительных циклов для получения специфически адаптированной рекомбиназы и, таким образом, будет требовать больших затрат времени. Так как более вероятно, что существующие адаптированные рекомбиназы распознают ассиметричную последовательность-мишень с близкой гомологией последовательности с последовательностями, найденными в LTR, направленную молекулярную эволюцию предпочтительно применяют для таких адаптированных рекомбиназ. Меньшие различия в последовательностях-мишенях требуют меньше циклов эволюции для "изменения" специфичности рекомбиназы (ранее адаптированной для распознания другой последовательности-мишени) к последовательности-мишени в LTR, таким образом, сохраняя время и обеспечивая более быстрое лечение ретровирусной инфекции.
В предпочтительном варианте осуществления способа in vitro эффективность уже существующей адаптированной рекомбиназы, идентифицированной на стадии (c) способа in vitro, или рекомбиназы, специфически адаптированной на стадии (d), необязательно тестируют in vitro в отношении вырезания провирусной ДНК ретровируса, которым заражен пациент. Это тестирование проводят до получения указанных рекомбиназ в виде фармацевтической композиции.
Тестирование эффективности рекомбиназ проводят для гарантии того, что соответствующая рекомбиназа распознает и рекомбинирует специфичную ассиметричную последовательность-мишень также и в клетках млекопитающих. Это тестирование предпочтительно проводят в клетках млекопитающих, таких как клетки HeLa, клетки PM1, T-клетки Jurkat, T-клетки CEM и моноцитарные клетки периферической крови (PMNC).
На дополнительной необязательной стадии способа оптимизации лечения ретровирусной инфекции у индивида in vitro полученную в результате адаптированную рекомбиназу включают в коллекцию специфически адаптированных рекомбиназ. Эта стадия обеспечивает получение коллекции различных адаптированных рекомбиназ, где каждая распознает специфичные последовательности-мишени, отличающиеся от последовательностей-мишеней остальных. Растущее количество адаптированных рекомбиназ в этой коллекции увеличивает вероятность того, что адаптированная рекомбиназа доступна, которая распознает последовательность-мишень, близкородственную последовательностям в последовательности LTR провирусной ДНК, для которых нужна рекомбиназа. Таким образом, при росте количества рекомбиназ в коллекции вероятность того, что рекомбиназу дикого типа будет нужно адаптировать в требующем большего времени способе, снижается, тогда как вероятность, того, что возможно более быстрое получение (т.е. требующее меньше циклов эволюции) специфически адаптированной рекомбиназы, начиная с другой более близкородственной адаптированной рекомбиназы, растет.
В последнем аспекте настоящее изобретение относится к коллекции адаптированных рекомбиназ, где каждый из представителей указанной коллекции получен способами по настоящему изобретению.
Таким образом, коллекция адаптированных рекомбиназ по настоящему изобретению предпочтительно содержит векторы экспрессии, кодирующие адаптированные рекомбиназы, бактерии, содержащие векторы, содержащие нуклеиновую кислоту, кодирующую адаптированную рекомбиназу, адаптированные рекомбиназы в форме белка или вирусные векторы, содержащие нуклеиновую кислоту, кодирующую адаптированные рекомбиназы, полученные в соответствии с надлежащей практикой организации производства.
В предпочтительном варианте осуществления коллекция по настоящему изобретению содержит векторы экспрессии, кодирующие нуклеиновые кислоты, кодирующие специфически адаптированные рекомбиназы. Затем векторы экспрессии лиофилизируют или хранят при температурах, по меньшей мере, -70°C. Хранение бактерий, белков или вирусных векторов осуществляют способами, хорошо известными в данной области.
Специалисту в данной области очевидно, что способы по настоящему изобретению, описанные выше, включают циклические процессы и характеристики каждого цикла или всего цикла могут быть автоматизированы.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Фиг.1. Стратегия разделения асимметричных последовательностей-мишеней. Закрашенные серым стрелки изображают инвертированные повторы симметричного участка-мишени, разделенные последовательностью спейсера (черная полоса). Цифры и буквы в ассиметричном участке-мишени представляют нуклеотиды, отличающиеся от гомологичного симметричного участка-мишени. Изогнутые стрелки представляют получение инвертированных повторов на основе последовательностей половин участков и последовательности спейсера. Заштрихованные серым рамки в участке-мишени во второй и третьей подгруппах представляют нуклеотиды из гомологичного симметричного участка-мишени, замещающие отличающийся нуклеотид. Последовательность спейсера в последовательностях-мишенях различных подгрупп может отличаться от последовательности спейсера гомологичной последовательности-мишени (и, таким образом, она изображена с использованием черной штриховки на белом фоне). Диаграмма только иллюстрирует замену нуклеотидов. Замены других нуклеотидов и другого количества нуклеотидов равно возможны и предпочтительны. Сокращения: hs1 = последовательность половины участка 1; hs2= последовательность половины участка 2, hs1' = инвертированный повтор, полученный из последовательности половины участка 1; hs2' = инвертированный повтор, полученный из последовательности половины участка 2.
Фиг.2. Стратегия комбинаторной направленной эволюции. (A) Изображены участки-мишени рекомбиназы, используемые при способе связанной с субстратом комбинаторной эволюции. Основания, выделенные серым, представляют собой основания, отличающиеся от loxP (SEQ ID NO:35). Левая и правая последовательности loxltr длиной 13 п.н. использовали для формирования палиндромных подгрупп loxltr1 и loxltr2 длиной 34 п.н., соответственно. loxltr1a и 1b получены из loxltr1 и характеризуются двумя мутациями в половине участка, где каждая соответствует плечам палиндромна. loxltr2a и 2b получены из loxltr2 и характеризуются тремя мутациями в половине участка, каждая. (B) Приведено общее представление 126 циклов связанной с субстратом направленной эволюции, проводимых для получения рекомбиназы Tre. Количество циклов эволюции для каждой подгруппы loxltr показано внутри стрелок с конечным количеством циклов, представленным на острие стрелки. Активность библиотеки рекомбиназ на первом и последнем циклах участка-мишени представлены в виде рестрикционного анализа плазмидной ДНК под соответствующими мишенями (расщепление BsrGI/XbaI при индукции L-арабинозой 200 мкг/мл). Нерекомбинированные (верхняя полоса длиной 4,9 т.п.н.) и рекомбинированные (нижняя полоса длиной 4,2 т.п.н.) приведены в виде линии с двумя треугольниками и одним треугольником, соответственно.
Фиг.3. Приведено представление стратегии направленной эволюции, используемой для эволюции специфичных для loxltr рекомбиназ. Эволюционные векторы pEVO-loxltr основаны на векторе pBAD33 и содержат два прямо повторяющихся участка-мишени рекомбиназы (loxltr). Рекомбинация по loxltr приводит к удалению промежуточной области, содержащей уникальный участок рестрикции NdeI и успешные кандидаты можно найти с применением ПЦР с праймерами P1 (SEQ ID NO:19) и P2 (SEQ ID NO:20) после расщепления NdeI. ПЦР приводит к полосе 1,7 т.п.н., которую можно использовать для обратного субклонирования успешных рекомбиназ в исходном векторе для следующего цикла. Рестрикционное расщепление пула плазмид посредством NdeI гарантирует удаление нерекомбинантных плазмид (не показано на фигуре) и, таким образом, устраняет нефункционирующие рекомбиназы.
Фиг.4. Картирование мутаций эволюционных рекомбиназ на вторичной структуре Cre. Встречаемость мутаций показана указанным цветовым кодом. Мутации аминокислот, встречающиеся более одного раза, добавляются к столбцу. Структурные элементы Cre представлены в виде полос для α-спиралей (от A до N) и стрелок для β-слоев (от 1 до 5) на основе кристаллической структуры рекомбиназы Cre. Каталитический тирозин обозначен как "Y". Закрашенные треугольники обозначают аминокислоты, формирующие специфичные контакты с ДНК. Гипервариабельные кодоны представлены указанием положения аминокислоты сверху столбцов.
Фиг.5. Представлена аминокислотная последовательность рекомбиназы Tre (SEQ ID NO:3). Последовательность NLS, добавленная для локализации Tre в клетках млекопитающих в ядре подчеркнута. Мутации, выделенные светло-серым и темно-серым (а/к 131, 244, 245, 259 и 262), изображают контактирующие и не контактирующие с ДНК основания на основе кристаллической структуры Cre, соответственно. Номер аминокислоты, соответствующий последовательности Cre и анализу мутаций, представленному на фиг.3, показан над мутациями.
Фиг.6. Сравнение последовательности loxltr с другими последовательностями LTR ВИЧ-1. Случайно выбранные 27 последовательностей LTR ВИЧ-1 выравнивали с последовательностью loxltr длиной 34 п.н. и рассчитывали сходство. Результаты представлены в виде процента гомологии последовательности loxltr относительно всех соответствующих LTR в библиотеке. На оси абсцисс приведены инвентарные номера последовательностей LTR ВИЧ-1.
Фиг.7. Активность рекомбиназы Tre в E. coli. (A) Схематическое представление основанного на репортере LacZ анализа. Сайт-специфическая рекомбинация приводит к удалению промотора E. coli, что приводит к отсутствию экспрессии LacZ. (B) Проиллюстрирована специфичность рекомбинации Tre с использованием плазмид с вырезанием основанного на lacZ репортера. Указаны различные сочетания основанных на loxP или loxltr репортерных плазмид и используемые рекомбиназы. В результате удаления после рекомбинации промотора, направляющего экспрессию lacZ, образуются белые колонии. (C) Активность рекомбиназы Tre в отношении различных участков-мишеней после культивирования плазмидной ДНК, соответствующей версиям pEVO различных участков-мишеней, и Tre в течение ночи при концентрации L-арабинозы 50 мкг/мл и расщепление BsrGI и XbaI. Нижняя рекомбинировавшая полоса длиной 4,2 т.п.н. указана как линия с одним треугольником, а верхняя нерекомбинировавшая полоса - как линия с 2 треугольниками. Под дорожками представлен рассчитанный процент рекомбинировавшей плазмиды каждой мишени.
Фиг.8. Анализ на транзиторную рекомбиназную активность в клетках HeLa. (A) В клетки HeLa котрансфицировали плазмиду с указанной рекомбиназой и репортерную плазмиду (смотри фиг.6). Клетки смешивали и через 48 часов после трансфекции окрашивали X-gal. Рекомбинация удаляла находящуюся внутри область терминации и приводила к активации экспрессии lacZ, отраженной синими клетками. (B) Измерение β-галактозидазной активности в лизатах клеток HeLa котрансфицированных плазмидой с указанной рекомбиназой и репортерной плазмидой. β-галактозидазную активность представляют как процент ферментативной активности клеток после параллельной трансфекции рекомбинированными формами репортерных плазмид pSVΔpaX и pSVΔloxltr. (C) Рекомбинация, опосредуемая Tre в стабильной репортерной клеточной линии loxltr HeLa. Клеточную линию трансфицировали отрицательным контролем, экспрессирующим Cre и Tre вектором и через 48 часов после трансфекции измеряли β-галактозидазную активность. (D) Детекция ПЦР рекомбинации в клеточной линии loxltr, описанной в (C) после трансфекции векторами экспрессии Cre и Tre. Показаны продукты ПЦР для нерекомбинировавшей и рекомбинировавшей контрольной плазмиды (pSVloxltr и pSVΔloxltr) и после трансфекции векторами экспрессии Cre и Tre. Нижняя рекомбинировавшая полоса обозначена линией с одним треугольником, а верхняя нерекомбинировавшая полоса линией с двумя треугольниками. Указана длина продуктов ПЦР.
Фиг.9. Анализ активации ВИЧ-1 Tat в клетках HeLa. (A) Схематическая диаграмма репортерного вектора pHIV/T2/LUC, содержащего в области U3 LTR ВИЧ-1 HXB3 последовательность loxltr (обозначенную белыми треугольниками). Другая копия последовательности loxltr, которая отсутствует в контрольном векторе pHIV/T1/LUC, расположена с 3'-конца гена люциферазы. Промотор LTR кодирует в его R области отвечающую на Tat область TAR. Трансактивация Tat отражается увеличенной экспрессией люциферазы светляка. L1 и L2 означают пару праймеров, используемую для детекции рекомбинации посредством ПЦР (черные треугольники). Рекомбинация посредством участков loxltr удаляет кодирующую область люциферазы. (B) Относительная активность LTR ВИЧ-1 после трансфекции клеток HeLa контрольным вектором pHIV/T1/LUC в присутствие и отсутствие Tre. Активность люциферазы светляка измеряли в соответствующих лизатах трансфицированных клеток через 48 часов после трансфекции. Активность корректировали на эффективность трансфекции с использованием вектора LacZ в качестве внутреннего контроля и измеряя β-галактозидазную активность в клеточных лизатах. (C) Относительная активность LTR ВИЧ-1 после трансфекции клеток HeLa вектором pHIV/T2/LUC в присутствие и отсутствие Tre. Клеточные лизаты анализировали как и ранее. (D) Детекция опосредованной Tre рекомбинации в клетках HeLa, транзиторно котрансфицированных экспрессирующей Tat плазмидой и указанными векторами посредством ПЦР. Нижняя полоса длиной 756 п.н. представляет собой рекомбинировавший фрагмент после потери кодирующей области люциферазы и она детектируется только в клетках, трансфицированных pHIV/T2/LUC и вектором экспрессии с Tre.
Фиг.10. Опосредованная Tre рекомбинация провирусной ДНК ВИЧ-1. (A) Схематическая диаграмма провируса NLT2ΔenvPuro. Изображена последовательность loxltr, расположенная в 5'- и 3'-LTR. Последовательность спейсера подчеркнута. Кодирующая область nef замещена геном устойчивости к пуромицину (светло-серая полоса), а специфичные для env последовательности удалены (Δ). Для инфицирования клеток HeLa использовали соответствующие псевдотипы VSV-G. (B) Детекция интегрированного провируса (gag ВИЧ-1; верхняя диаграмма) и рекомбинации области 5'- и 3'-LTR (5'/3'LTR; нижняя диаграмма) посредством количественного ПЦР с детекцией в реальном времени общей геномной ДНК, выделенной из инфицированных и экспрессирующих Tre (Tre) или не содержащих Tre (контрольных) клеток на неделе 4, 6, 8 и 10 после трансфекции. Каждый образец тестировали в трех экземплярах. (C) Анализ высвобождения вирусных частиц и жизнеспособности клеток. Посредством ELISA (верхняя диаграмма) определяли уровни антигена p24Gag в супернатантах культуры на указанных неделях после трансфекции. Представлен процент ингибирования репликации вируса в экспрессирующей Tre культуре по сравнению с контрольными клетками. Одновременно подвергали мониторингу жизнеспособность клеток посредством анализа с alamarBlue (нижняя диаграмма). (D) Детекция экспрессирующих Gag клеток (светло-серая метка) посредством непрямой иммунофлуоресценции в контрольной культуре (Контроль) на неделе 4, или в экспрессирующей Tre культуре на неделе 4, 6, 8 и 12 после трансфекции. Инкубация контрольных клеток со связанным с Cy2 вторичным антителом отдельно служила в качестве контроля отрицательного окрашивания (NC). Ядра визуализировали окрашиванием DRAQ5 (темная метка).
Фиг.11. Экспрессирующие клетки чувствительны к пуромицину. Клетки не содержащей Tre (контроль) или экспрессирующей Tre (Tre) культуры через 8 недель после трансфекции подвергали воздействию 0,5 пг/мл пуромицина и проводили мониторинг жизнеспособности клеток с течением времени посредством анализа с alamarBlue.
Настоящее изобретение будет более подробно описано посредством примеров.
Пример 1
Экспрессирующие рекомбиназу и репортерные плазмиды
Для цели настоящего изобретения с использованием стандартных способов и синтетических двухцепочечных олигонуклеотидов или технологии ПЦР получали различные векторы экспрессии.
кДНК Cre и Tre для экспрессии в клетках млекопитающих клонировали в векторе pIRESneo3 (Clontech) с использованием участков рестрикции NheI и BamHI.
Репортерные вектора с множественными хозяевами pSVpaX и pSVloxltr являются идентичными за исключением прямо повторенных участков loxP или loxltr, фланкирующих область pac, соответственно. pSVpaX был описан ранее (Buchholz et al., 1996). pSVloxltr получали из pSVpaX, замещая участки loxP последовательностями loxltr.
Отвечающую на Tat плазмиду с репортерным геном pHIV/T1/LUC конструировали, лигируя LTR HXB3 ВИЧ-1 между участками SpeI и HindIII, и ген люциферазы светляка между участками HindIII и XhoI эукариотического вектора экспрессии pcDNA3 (Invitrogen). Затем посредством ПЦР в LTR HXB3 вводили последовательность loxltr штамма первичного ВИЧ-1 TZB0003 (инвентарный номер Genbank AF096641). Плазмиду pHIV/T2/LUC получали посредством вставки двухцепочечного кодирующего loxltr олигонуклеотида с 3'-конца гена люциферазы между участками XhoI и ApaI pHIV/T1/LUC. Экспрессирующий Tat ВИЧ-1 вектор pcTat, исходный вектор pBC12/CMV и вектор внутреннего контроля pBC12/CMV/βGal описаны ранее (Malim et al., 1988; Ruhl et al., 1993). Вектор p3Tre получали, лигируя кДНК Tre (SEQ ID NO:2) между участками HindIII и BamRI pcDNA3. Плазмида pNLT2ΔenvPuro представляет собой производное провирусной ДНК pNL4-3 (инвентарный номер GenBank M19921) и ее конструировали, преобразуя посредством ПЦР соответствующие последовательности LTR NL4-3 в последовательность loxltr ВИЧ-1 TZB0003. Затем, удаляли фрагмент гена env длиной 537 п.н. (положение нуклеотидов от 6712 до 7249). Наконец, кодирующую последовательность nef замещали геном устойчивости к пуромицину. Вектор, экспрессирующий гликопротеин оболочки вируса везикулярного стоматита, pCMV-VSV-G, описан (Beyer et al., 2002).
Пример 2
Получение адаптированной рекомбиназы, распознающей и рекомбинирующей ассиметричную последовательность-мишень в LTR ВИЧ-1
I. Определение кандидатных асимметричных последовательностей-мишеней
Для начала процесса эволюции последовательности LTR ВИЧ-1 сканировали сначала на последовательность со сходством с каноническим участком loxP. Выбранная последовательность принадлежит LTR штамма первичного ВИЧ-1 TZB0003 (Blackard et al., 1999) и представляет собой часть его модулирующей области U3 (положение в последовательности от -262 до -229; где участок инициации транскрипции представляет собой +1). Выбранный участок loxltr представляет собой ассиметричную последовательность длиной 34 п.н. с 50% сходством последовательности с loxP с четырьмя несоответствиями в левом элементе, шестью в правом элементе и полностью отличающимся спейсером (смотри фиг.2A).
II. Анализ последовательности loxltr в связанной с субстратом направленной эволюции белков в E. coli
Последовательность loxltr анализировали в связанной с субстратом направленной эволюции белков как описано (Buchholz & Stewart, 2001) (также смотри фиг.3).
Посредством ПЦР с использованием указанных ниже олигонуклеотидов получали фрагменты длиной 690 пар оснований, содержащие участок рестрикции NdeI, фланкированный участками loxltr, с использованием в качестве матрицы плазмиды pEVO-loxP (Buchholz & Stewart, 2001):


Фрагменты ПЦР расщепляли BglII и клонировали в эволюционном векторе, расщепленном тем же ферментом. Корректность последовательности клонов подтверждали с использованием праймера 5'-CAATAACCCTGATAAATG-3' (SEQ ID NO:18) посредством секвенирования.
Исходные библиотеки рекомбиназ, клонируемые в эволюционных векторах pEVO-loxltr, получали посредством ПЦР пониженной точности, как описано в Buchholz & Stewart (2001). Эволюционный вектор основан на векторе pBAD33 (Guzman et al., 1995) и содержит два прямо повторенных участка-мишеней рекомбиназы (loxltrs). Плазмида также содержит промотор araC, направляющий индуцируемую арабинозой экспрессию рекомбиназ и маркер устойчивости к хлорамфениколу (Cm) для размножения в E. coli. Плазмида также обеспечивает удобное клонирование рекомбиназ с использованием участков BsrGI и XbaI, фланкирующих кодирующую область рекомбиназы. Библиотеки трансфицировали в клетки E. coli DH5a и выращивали в жидкой культуре LB, содержащей хлорамфеникол в концентрации 25 мкг/мл. С использованием эволюционного вектора (смотри фиг.3) и Cre архив библиотек мутированных Cre тестировали на рекомбинационную активность, как описано (Buchholz & Stewart, 2001).
Рекомбинация и последующая ПЦР позволяли получать полосу длиной 1,7 т.п.н., отражающую рекомбинацию (смотри фиг.3). Однако Cre, а также библиотека не могла рекомбинировать участки loxltr и продукта ПЦР не получали (данные не показаны), отражая то, что ассиметрия и мутации в loxltr являются слишком сильными, чтобы приводить к рекомбинации.
III. Получение подгрупп ассиметричных последовательностей-мишеней loxltr
Для тестирования того, приводит ли удаление ассиметрии в участке-мишени к рекомбинационной активности, исходную мишень loxltr разделяли на две подгруппы. Получали палиндромные участки-мишени, loxltr1 и loxltr2, на основе исходной ассиметричной последовательности loxltr, с использованием для формирования инвертированного повтора последовательностей левой и правой половины участка, соответственно (смотри фиг.2A). Однако, когда loxltr1 и loxltr2 тестировали на рекомбинацию с применением Cre или библиотеки, рекомбинации не наблюдали (данные не показаны). Таким образом, мутаций в этих участках оставалось слишком много для того, чтобы начальная библиотека продемонстрировала хоть какую-либо активность, и это сделало необходимым дальнейшее разделение loxltr1 и loxltr2 посредством равного разделения мутаций в половинах участков с формированием четырех новых подгрупп, обозначенных loxltr1a, loxltr1b, loxltr2a и loxltr2b (смотри фиг.2A).
IV. Проведение эволюции на основе сшитых с субстратом белков с использованием подгрупп ассиметричных последовательностей-мишеней loxltr
Получали начальные библиотеки рекомбиназ, клонируемые в эволюционных векторах pEVO-loxltr1a, pEVO-loxltr1b, pEVO-loxltr2a и pEVO-loxltr2b, как описано выше (смотри пример 2.II) и независимо трансфицировали в клетки E. coli DH5α, выращиваемые в жидкой культуре LB, содержащей хлорамфеникол в концентрации 25 мкг/мл.
Библиотеки рекомбиназ перед началом циклов эволюции (цикл 1) индуцировали 200 мкг/мл L-арабинозы и концентрацию арабинозы при успешных циклах снижали до тех пор, пока не достигали удовлетворительной рекомбинации при минимальной индукции (0-5 мкг/мл) L-арабинозой.
Плазмидную ДНК, выделяемую после каждого цикла эволюции, расщепляли NdeI и кодирующие рекомбиназы последовательности, которые содержали успешно рекомбинировавший эволюционный вектор, амплифицировали с праймерами P1 (5'-TCTACTGTTTCTCCATA-3'; SEQ ID NO:19) и P2 (5'-TGTCGCCCTTATTCCCT-3'; SEQ ID NO:20) в 50 мкл смеси ПЦР, содержащей 1× буфер для ПЦР, 250 мкМ каждого dNTP, 3 мМ MgCl2 и 1,5 единицы ДНК-полимеразы BioTaq по следующей программе: 35 циклов денатурации при 94°C в течение 60 сек; отжига при 55°C в течение 60 сек и удлинения при 72°C в течение 90 сек. Продукт ПЦР рекомбинированной формы плазмиды давал полосу 1,7 т.п.н. Этот продукт ПЦР расщепляли BsrGI и XbaI и обратно субклонировали в соответствующих векторах pEVO-LTR в течение следующего цикла эволюции. Для всех библиотек библиотеку поддерживали на уровне свыше 25000 отдельных клонов.
Cre рекомбинировала только loxltr1a на очень низких уровнях (данные не показаны), что отражает ее высокую специфичность к мишени. В отличие от этого, экспрессия библиотеки мутантов приводила к низкой рекомбинационной активности во всех подгруппах loxLTR. Разделение мутаций способствовало распознаванию библиотекой Cre и, таким образом, служило исходной точной для последующих циклов направленной эволюции. Повторяющиеся циклы направленной эволюции приводили к обогащению библиотеки рекомбиназ функциональными кандидатами (смотри фиг.2B). Количество циклов эволюции, необходимых для получения эффективных рекомбиназ для каждого loxltr, варьировало в подгруппах loxltr, но в конечном счете эффективную рекомбинационную активность библиотек наблюдали для всех подгрупп.
V. Объединение библиотек и перестановка ДНК для обеспечения рекомбинации следующей подгруппы более высокого порядка
Для тестирования того, позволяет ли комбинаторный подход рекомбинацию следующей подгруппы более высокого порядка, библиотеки 1a и 1b и 2a и 2b, соответственно, объединяли и подвергали перестановкам.
После каждого третьего цикла эволюции проводили перестановку ДНК (Stemmer, 1994) для рекомбинации полученных мутантов Cre in vitro, а также для рекомбинации библиотек рекомбиназ подгрупп более низкого порядка для инициации эволюции относительно участков-мишеней loxltr более высокого порядка. В кратком изложении, фрагменты ДНК, содержащие ген рекомбиназы случайным образом фрагментировали ультразвуком с использованием 12% амплитуды в течение 5 минут с 30 сек включенной импульсации и 5 сек выключенной импульсации с использованием цифрового ультразвукового дезинтегратора 450-D Digital Sonifier из Branson. Полученные посредством ультразвука фрагменты (размером 100-400 п.н.) очищали с использованием набора для очистки ПЦР (Qiagen). Для обратной сборки фрагментов 2 мкг обработанной ультразвуком ДНК добавляли в 50 мкл смесь ПЦР и амплифицировали с использованием той же программы, как указана выше (смотри пример 2.IV). 3 мкл продукта этой реакции ПЦР без использования праймеров использовали в качестве матрицы для следующей ПЦР с праймерами P1 (5'-TCTACTGTTTCTCCATA-3', SEQ ID NO:19) и P3 (5'-GCGGATGAGAGAAGATT-3'; SEQ ID NO:21) в 50 мкл смеси ПЦР с теми же условиями циклов. Библиотеку с перестановками (конечный продукт ПЦР) расщепляли BsrGI и XbaI и обратно клонировали в подходящие векторы pEVO, несущие loxltr1 и loxltr2, соответственно. Комбинирование мутаций из различных библиотек приводило к синергическому действию и приводило к получению рекомбиназ, которые теперь рекомбинировали loxltr1 и loxltr2 (смотри фиг.2B), демонстрируя, что стратегию эволюции перехода через промежуточные продукты можно использовать для достижения желаемой активности.
VI. Объединение библиотек и перестановки ДНК для обеспечения рекомбинации ассиметричного участка-мишени в LTR
Последовательность loxP представляет собой симметричный участок, который связывается двумя мономерами Cre. Таким образом, рекомбиназа, которая рекомбинирует ассиметричный участок-мишень, должна распознавать половины участков различной последовательности. Для тестирования того, можно ли посредством эволюции на основе сшитых с субстратом белков получать рекомбиназу, осуществляющую рекомбинацию ассиметричного участка-мишени loxltr, библиотеки из loxltr1 и loxltr2 объединяли и подвергали перестановке и анализировали на рекомбинацию в эволюционном векторе, несущем последовательность-мишень ассиметричного loxLTR. В первых циклах детектировали очень низкую рекомбинационную активность, которая усиливалась за счет функциональных кандидатов в более поздних циклах (смотри фиг.2B), демонстрируя, что симметрия в участке-мишени не является необходимым условием для реакции сайт-специфической рекомбинации.
После циклов эволюции в общем количестве 126 процесс эволюции остановили и отдельные специфичные для loxLTR рекомбиназы анализировали на их рекомбинационные свойства. С использованием рестрикционного анализа в E. coli функционально проанализировали пятьдесят отдельных рекомбиназ (данные не показаны).
Наиболее активная рекомбиназа (названная Tre из-за рекомбиназы LTR (LTR recombinase)) показала эффективную рекомбинацию участка loxLTR с некоторой остаточной активностью в отношении loxP при тестировании в анализе с репортером на основе lacZ (смотри фиг.7A и B). Для количественного определения специфичности Tre к мишени ее рекомбинационные свойства анализировали в эволюционных векторах всех подгрупп loxLTR (смотри фиг.7C). Как и в анализе с репортером, Tre эффективно рекомбинировала последовательность loxLTR и демонстрировала остаточную активность в отношении loxP (смотри фиг.7C). Tre также демонстрировала эффективную рекомбинацию для loxLTR2b и остаточную активность для loxLTR2, но для loxLTR1a, loxLTR1b, loxLTR1 и loxLTR2a рекомбинации не наблюдали (смотри фиг.7C). Это является необычным, если принимать в расчет, что Tre эволюционировала из этих подгрупп. Это наблюдение подтверждает более ранние открытия, что специфичность к мишеням после начального ослабления в направленной эволюции возвращается через множество циклов генерации (Buchholz & Stewart, 2001; Santoro & Schultz, 2002); Matsumura & Ellington, 2001).
VII. Секвенирование эволюционировавших рекомбиназ
Эволюционировавшие рекомбиназы из всех подгрупп и loxLTR секвенировали для мониторинга процесса эволюции. Последовательности выявили кластеризацию мутаций, происходящую из различных подгрупп, которые комбинировали при эволюции и дополнялись новыми кластерами в подгруппах более высокого порядка (смотри фиг.4 и таблицу 1).
В таблице 1 приведены аминокислотные мутации в секвенированных рекомбиназах для указанных подгрупп loxltr эволюционировавших рекомбиназ. Заштрихованные остатки контактируют в кристаллической структуре Cre с ДНК. Гипервариабельные участки (также показанные на фиг.5) показаны жирными буквами для контактирующих с ДНК и не контактирующих с ДНК участков.


Всего, в Tre содержатся 19 аминокислотных замен при сравнении с Cre, где многие из этих мутаций произошли из различных подгрупп (смотри фиг.5).
Пример 3
Характеристика рекомбиназы Tre
I. Рекомбинационные свойства Tre в клетках млекопитающих
Если не указано иначе, клетки HeLa культивировали при 37°C при 5% CO2 в DMEM, содержащей 100 единиц/мл пенициллина и стрептомицина и дополненной 10% ЭТС.
Получали устойчивую к пуромицину репортерную клеточную линию pSVloxltr посредством трансфекции 2 × 106 клеток 8 мкг репортерной плазмиды с использованием реагента для трансфекции Effectene (Qiagen) по инструкциям производителя, и отбора с 5 мкг/мл пуромицина (Invivogen). Полученную стабильную клеточную линию растили в присутствие 3 мкг/мл пуромицина.
Клетки HeLa котрансфицировали плазмидой, экспрессирующей рекомбиназу и репортерной плазмидой в соотношении 1:2 в 6-луночном формате с использованием Effectene (Qiagen). Всего для трансфекций использовали 0,6 мкг ДНК. Проводили параллельные трансфекции рекомбинированной версией репортерных плазмид для подсчета процента рекомбинированных репортерных плазмид.
Рекомбиназную активность оценивали через 48 часов после трансфекции, анализируя относительную β-галактозидазную активность через 48 часов после трансфекции с использованием набора Tropix Galacto-Light kit (Applied Biosystems, Bedford, MA) по протоколу производителя. В кратком изложении, клетки HeLa отмывали PBS через 48 часов после трансфекции и фиксировали 2% формальдегидом и 0,1% глутаральдегидом, пермеабилизировали 0,5% Triton-X 100 и окрашивали в течение ночи при 37°C окрашивающим раствором, содержащим 1 мг/мл X-gal, 2 мМ MgCl2, 5 мМ феррицианид калия и 5 мМ ферроцианид калия. Эффективность рекомбинации корректировали на общее содержание белка в клеточном лизате, измеренное посредством набора для анализа белка BCA (Pierce, Rockford, IL).
Как и при анализах в E. coli Cre эффективно рекомбинировала репортер loxP, но не рекомбинировала loxLTR, а Tre продемонстрировала эффективную рекомбинацию в отношении репортера loxLTR и некоторую остаточную активность в отношении loxP (смотри фиг.8A и B). Для исследования того, может ли Tre рекомбинировать ее мишень в контексте генома, стабильную репортерную клеточную линию loxLTR тестировали на рекомбинацию после трансфекции экспрессирующей Tre плазмидой.
Стабильную клеточную линию loxLTR получали посредством отбора на пуромицине (2 мкг/мл) клеток HeLa, трансфицированных плазмидой pSVloxltr. Для анализа рекомбинации эту клеточную линию трансфицировали 0,8 мкг контроля, плазмид с Cre или Tre с использованием Effectene в 6-луночном формате и анализировали на β-галактозидазную активность через 48 часов после трансфекции с использованием набора Galacto-Light kit (Applied Biosystems, Bedford, MA). β-галактозидазную активность корректировали на эффективность трансфекции, измеряя активность люциферазы renilla в клеточных лизатах.
Анализы ПЦР и измерения β-галактозидазной активности показали, что Tre рекомбинирует последовательности loxltr, упакованные в хроматине (смотри фиг.8C и D).
Для анализов ПЦР клеточную линию loxLTR HeLa трансфицировали соответствующими экспрессирующими рекомбиназу векторами с использованием Effectene. Трансфицированные клетки через 48 часов отмывали PBS и трипсинизировали. Геномную ДНК выделяли с использованием мини-набора QIAamp DNA Blood mini kit (Qiagen) по инструкциям производителя. Геномную ДНК (3 мкг) анализировали на рекомбинацию с использованием праймеров P' (5'-GCCTCGGCCTAGGAACAGT-3'; SEQ ID NO:22) и P" (5'-CCGCCACATATCCTGATCTT-3'; SEQ ID NO:23) с использованием программы ПЦР с 30 циклами денатурации при 95°C в течение 30 сек; отжига при 62°C в течение 30 сек и удлинения при 72°C в течение 40 сек в 50 мкл реакционной смеси ПЦР, содержащей 1× буфер ПЦР, 250 мкМ каждого dNTP, 3 мМ MgCl2 и 1,5 единиц ДНК-полимеразы BioTaq. Продукты ПЦР визуализировали на 0,7% агарозном геле.
II. Анализ опосредованной Tre рекомбинации в контексте LTR ВИЧ-1
Получали репортерные конструкции Tat ВИЧ и тестировали в анализах Tat (смотри фиг.9). Белок Tat представляет собой жизненно важный вирусный трансактиватор, регулирующий направляемую промотором LTR транскрипцию ВИЧ (Emerman & Malim, 1998). Для последующих анализов в вектор с ВИЧ-люциферазным репортером (pHIV/T1/LUC; контрольный вектор) вводили последовательность loxLTR в LTR HXB3 ВИЧ-1 (Ratner et al., 1985). Кроме того, другую копию последовательности loxLTR вставили с 3'-конца кодирующей люциферазу последовательности с получением pHIV/T2/LUC (изображенного на фиг. 9A). Таким образом, в этих векторах трансактивация Tat приводит к экспрессии люциферазы светляка, направляемой промотором LTR.
Для анализов транзиторной трансактивации Tat клетки HeLa трансфицировали в 12-луночном формате с использованием реагента для трансфекции Effectene (Qiagen). Смесь для трансфекции состояла из 0,16 мкг HIV/T2/LUC или HIV/T1/LUC, экспрессирующих люциферазу светляка, 0,16 мкг вектора pIres-neo-рекомбиназа Tre или пустого вектора в качестве отрицательного контроля, 0,08 мкг вектора Tat или pcDNA3 в качестве отрицательного контроля для Tat. Через 48 часов после трансфекции клетки лизировали и оценивали на активность люциферазы светляка с использованием набора для анализа люциферазы светляка и Renilla (Biotium, Inc., Hayward, CA). Активность корректировали на эффективность трансфекции с использованием вектора lacZ в качестве внутреннего контроля и измеряя β-галактозидазную активность в клеточных лизатах.
Когда клетки HeLa котрансфицировали экспрессирующим Tre вектором вместе с вектором Tat и pHIV/T2/LUC, наблюдали трехкратное снижение люциферазной активности (смотри фиг.9C). В отличие от этого снижения экспрессии люциферазы при проведении того же эксперименте с использованием контроля pHIV/T1/LUC не детектировали (смотри фиг.9B).
Для доказательства того, что наблюдаемое снижение экспрессии люциферазы являлось результатом рекомбинации, а не блокирования активности Tat или транскрипции с промотора LTR посредством рекомбиназы, проводили анализ ПЦР, оценивая рекомбинацию при трансактивации Tat (смотри фиг.9D).
Анализ ПЦР рекомбинации в анализе трансактивации Tat проводили с ДНК, выделенной из клеток HeLa, трансфицированных репортерными векторами ВИЧ в присутствие или отсутствие рекомбиназы, с использованием праймеров L1 (5'-GAAGGTGGGTTTTCCAGTCA-3'; SEQ ID NO:24) и L2 (5'-AGGGAAGAAAGCGAAAGGAG-3'; SEQ ID NO:25) с применением программы ПЦР с 30 циклами денатурации при 95°C в течение 30 сек, отжига при 60°C в течение 30 сек и удлинения при 72°C в течение 40 сек в 25 мкл реакционной смеси ПЦР, содержащей 1× буфер ПЦР, 250 мкМ каждого dNTP, 3 мМ MgCl2 и 1,5 единиц ДНК-полимеразы BioTaq. Продукты ПЦР визуализировали на 0,7% агарозном геле. Продукты ПЦР выделяли из геля, очищали с применением набора для экстракции из геля (Qiagen) и использовали для секвенирования.
Вследствие удаления кодирующей последовательности люциферазы рекомбинация приводила к размеру полосы 756 п.н., тогда как в отсутствие рекомбинации размер полосы составляет 2,7 т.п.н. ПЦР в клетках, трансфицированных вектором pHIV/T1/LUC, содержащим только один участок loxltr, давал только одну полосу размером 2,7 т.п.н. и в присутствие, и в отсутствие Tre. В отличие от этого, ПЦР в клетках, трансфицированных вектором pHIV/T2/LUC преимущественно давал нижнюю полосу, демонстрируя, что снижение активации Tat происходило вследствие опосредованного Tre вырезания кассеты люциферазы (смотри фиг.9D). Экстракция фрагментов ПЦР из геля с последующим секвенированием подтвердили точное вырезание фланкированной loxltr последовательности (данные не показаны).
Пример 4
Вырезание полноразмерного провируса из генома инфицированных ВИЧ-1 клеток человека
Анализировали способность Tre вырезать полноразмерный провирус из генома инфицированных ВИЧ-1 клеток человека.
Для тестирования этого конструировали провирусную ДНК, происходящую из штамма ВИЧ-1 NL4-3 (Adachi et al., 1986), которая в своих 5'- и 3'-LTR содержит последовательность loxLTR (приведено на фиг.10A). В этой конструкции рамку считывания nef, необязательную для репликации вируса в клеточной структуре, заменяли геном устойчивости к пуромицину. Кроме того, увеличение общей длины генома компенсировали делецией специфичных для env последовательностей.
Получали вирусные псевдотипы посредством транзиторной котрансфекции клеток 293T данной провирусной ДНК и вектором экспрессии, кодирующим белок оболочки вируса везикулярного стоматита (VSV-G). В кратком изложении, псевдотипы ВИЧ-1 получали посредством транзиторной котрансфекции 1 × 106 клеток 293T 3 мкг pNLT2ΔenvPuro и 3 мкг pCMV-VSV-G в OptiMEM I (Invitrogen) без антибиотиков с использованием 4 мкл полиэтиленимина (PEI; 1 мг/мл) по протоколу производителя (Polysciences, Inc.). На сутки 2 после трансфекции вирусный супернатант собирали и пропускали через фильтры с размером пор 0,2 мкм для гарантии удаления любых вирусных агрегатов и клеток посредством ELISA определяли уровень антигена p24Gag (Innotest HIV p24 Antigen mAb; Innogenetics N. V.).
Затем, 5 × 105 клеток HeLa, культивируемых в DMEM, содержащей 15% эмбриональную телячью сыворотку (Pansystems GmbH) и антибиотики (пенициллин и стрептомицин), инфицировали в шестилуночный планшет посредством спинокуляции (O'Doherty et al., 2000) при 1000 нг/мл псевдотипированного вируса вместе с 1 мкг/мл полибрена (Sigma-Aldrich) при 1200 × g в течении 90 мин при комнатной температуре. Через 24 часа после инфицирования среду для культивирования сменяли и клетки отмывали для удаления свободного вируса. Через 3 суток после инфицирования клеточную культуру дополняли 1 пг/мл пуромицина (Sigma-Aldrich) в течение 2 недель и 0,5 пг/мл пуромицина в течение следующих 4 недель для отбора и клонирования инфицированных высвобождающих вирусные частицы клеток. Среду меняли каждые вторые сутки для удаления мертвых клеток.
Затем инфицированные высвобождающие вирусные частицы клетки трансфицировали плазмидой, экспрессирующей Tre или исходным контрольным вектором, которые оба также несли ген устойчивости к неомицину. Соответствующие пулы устойчивых к неомицину клеток подвергали мониторингу в отношении рекомбиназной активности и продукции вируса.
Общую геномную ДНК выделяли из инфицированных клеток HeLa с использованием набора Blood & Cell Culture DNA Midi kit (Quiagen) по протоколу производителя. Количественная ПЦР с детекцией в реальном времени выявила, что в экспрессирующих Tre клетках по сравнению с контрольными клетками присутствие специфичных для gag последовательностей ВИЧ-1 в клеточном геноме постоянно уменьшалось с течением времени (смотри фиг.10B, верхняя диаграмма). Количественную ПЦР, специфичную для gag ВИЧ-1 (gag-прямой: 5'-ATCAATGAGGAAGCTGCAGAA-3' (SEQ ID NO:26), gag-обратный: 5'-GATAGGTGGATTATGTGTCAT-3' (SEQ ID NO:27), gag-зонд: 5'-FAM-ATTGCACCAGGCCAGATGAGAGAA-TAMRA-3' (SEQ ID NO:28)) и рекомбинации 5'/3' LTR (LTR-прямой: 5'-GATGGTGCTACAAGCTAGTAC-3' (SEQ ID NO:29), LTR-обратный: 5'-CTGTCAAACCTCCACTCTAAC-3' (SEQ ID NO:30), LTR-зонд: 5'-FAM-AAGGAGAGAACAACATCCTATTACAC-TAMRA-3' (SEQ ID NO:31)), проводили следующим образом: 40 циклов, включающих 100 нг геномной ДНК, 300 нМ прямого праймера, 900 нМ обратного праймера и 200 нМ зонда для гибридизации с использованием Platinum qPCR Super Mix UDG (Invitrogen) в системе для ПЦР Applied Biosystems 7500 Fast Real-Time PCR System. Каждый образец тестировали в трех повторениях, а результаты нормализовали с применением амплификации той же геномной ДНК с праймерами для β-глобина человека (HBG-прямой: 5'-CTTAATGCCTTAACATTGTGTATAA-3' (SEQ ID NO:32), HBG-обратный: 5'-GAATATGCAAATAAGCACACATATAT-3' (SEQ ID NO:33), HBG-зонд: 5'-FAM-ACTTTACACAGTCTGCCTAGTACATTAC-TAMRA-3' (SEQ ID NO:34)).
Через 10 недель культивирования, последовательности gag больше уже невозможно было культивировать. Этот результат близко отражался сходным анализом, в котором определяли рекомбинацию 5'- и 3'-LTR (смотри фиг.10B; нижняя диаграмма).
Кроме того, прекращение почкования частиц, которое отслеживали, определяя уровни антигена p24Gag в супернатантах культуры, происходило с немного замедленной кинетикой, что на неделе 12 приводило к 100% ингибированию высвобождения частиц по сравнению с контрольной культурой с отсутствием Tre (смотри фиг.10C, верхняя диаграмма). Параллельный анализ цитотоксичности с alamarBlue (Serotec) не смог выявить какого-либо индуцированного Tre токсического действия на клеточный метаболизм (смотри фиг.10C, нижняя панель).
Визуализация экспрессии Gag посредством лазерной сканирующей иммунофлуоресцентной микроскопии, продемонстрировало полное исчезновение продуцирующих инфицированных клеток в положительной по Tre культуре с течением времени (смотри фиг.10D). Для непрямой иммунофлуоресцентной микроскопии инфицированных ВИЧ-1 клетки HeLa растили на покровных стеклах, отмывали PBS без Ca2+ и Mg2+, а затем фиксировали 3% параформальдегидом в течение 25 мин. Все стадии инкубации проводили при температуре окружающей среды. Через 10 мин инкубации со 100 мМ глицином/PBS, клетки пермеабилизировали в 0,1% Triton X-100/PBS в течение 4 мин и инкубировали в 1% BSA/PBS в течение 30 мин. Белки окрашивали в 1% BSA/PBS с использованием первичных антител, распознающих p55Gag ВИЧ-1 (разведение 1:100, специфичное для Gag ВИЧ-1 моноклональное антитело мыши 183-H12-5C, предоставленное B. Chesebro и K. Wehrly, AIDS Research and Reference Reagent Program, Division AIDS, NIAID, NIH) или антитело кролика к Cre (разведение 1:500) в течение 45 мин с последующими подходящими вторичными антителами козы к антителам мыши или козы к антителам кролика (оба из Rockland Immunochemicals, Inc.; разведение 1:150), связанных с Cy2. Ядра визуализировали окрашиванием DRAQ5 (разведение 1:1000, Alexis Biochemicals) в течение 10 мин. Клетки собирали в среду мовиола и анализировали с использованием лазерного сканирующего микроскопа Zeiss LSM 510 Meta. Примечательно, что повторная обработка этих культур пуромицином на неделе 8 переживалась контрольными клетками, но приводила к тяжелым токсическим эффектам и итоговой смерти клеток в культуре, положительной по Tre (смотри фиг.11). Данное наблюдение можно объяснить опосредованным Tre удалением гена провируса, кодирующего устойчивость к пуромицину (сравни с фиг.10A).
СПИСОК ССЫЛОК











| название | год | авторы | номер документа |
|---|---|---|---|
| АДАПТИРОВАННАЯ РЕКОМБИНАЗА ДЛЯ РЕКОМБИНАЦИИ АСИММЕТРИЧНЫХ УЧАСТКОВ-МИШЕНЕЙ ВО МНОЖЕСТВЕ ШТАММОВ РЕТРОВИРУСОВ | 2011 |
|
RU2617968C2 |
| ХОРОШО ПЕРЕНОСИМАЯ И ВЫСОКОСПЕЦИФИЧЕСКАЯ АДАПТИРОВАННАЯ РЕКОМБИНАЗА ДЛЯ РЕКОМБИНИРОВАНИЯ АСИММЕТРИЧНЫХ УЧАСТКОВ-МИШЕНЕЙ ВО МНОЖЕСТВЕ ШТАММОВ РЕТРОВИРУСОВ | 2015 |
|
RU2706194C2 |
| ИММОРТАЛИЗОВАННАЯ КЛЕТОЧНАЯ ЛИНИЯ ПТИЦ | 2008 |
|
RU2475536C2 |
| ВЕКТОРНАЯ СИСТЕМА НА ОСНОВЕ ASLV | 2010 |
|
RU2566563C2 |
| ТЕЛОМЕРАЗНАЯ ОБРАТНАЯ ТРАНСКРИПТАЗА ПТИЦ | 2007 |
|
RU2415867C2 |
| УСОВЕРШЕНСТВОВАНИЕ ГЕНЕТИЧЕСКИХ КОНСТРУКЦИЙ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АНТИВИЧ ТЕРАПИИ | 2013 |
|
RU2533817C1 |
| КОНСТРУКЦИИ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ ПОЛУЧЕНИЯ БЕЛКОВ | 2021 |
|
RU2840923C1 |
| СТАБИЛЬНЫЕ КЛЕТОЧНЫЕ ЛИНИИ ДЛЯ ПРОДУЦИРОВАНИЯ РЕТРОВИРУСОВ | 2016 |
|
RU2752498C2 |
| САМОИНАКТИВИРУЮЩИЕСЯ АДЕНОВИРУСЫ-ПОМОЩНИКИ ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНЫХ АДЕНОВИРУСОВ С ВЫСОКОЙ ЕМКОСТЬЮ | 2009 |
|
RU2520809C2 |
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ АНТИВИЧ-ТЕРАПИИ | 2010 |
|
RU2426788C1 |
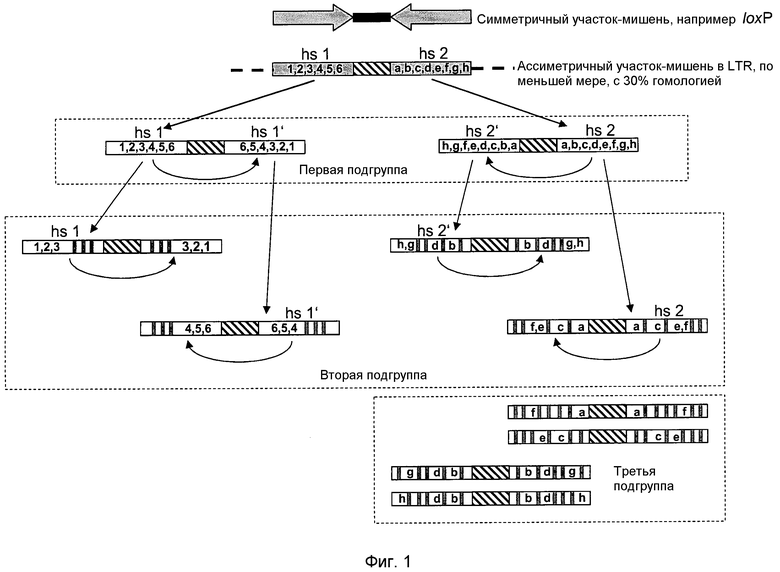
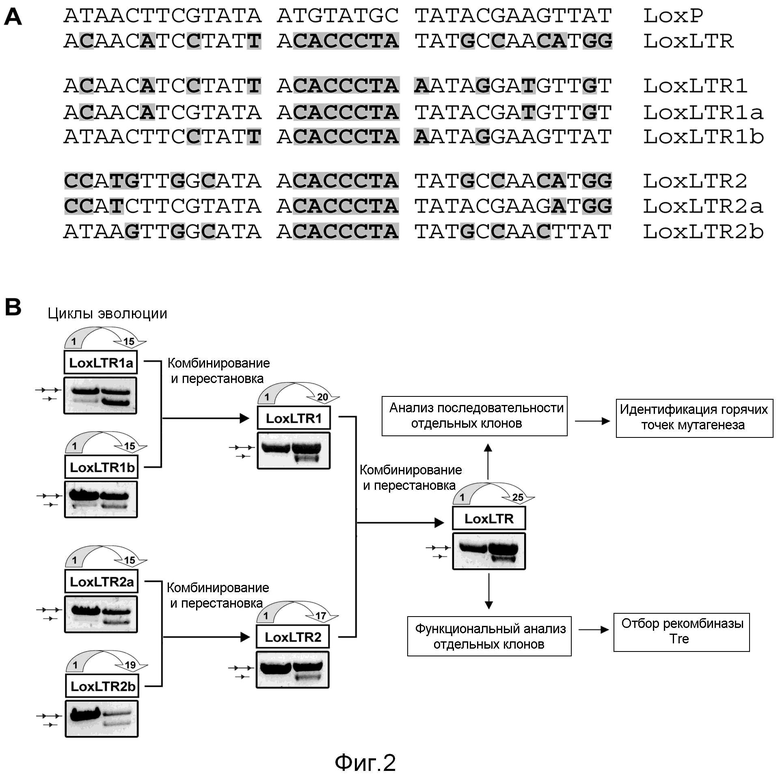



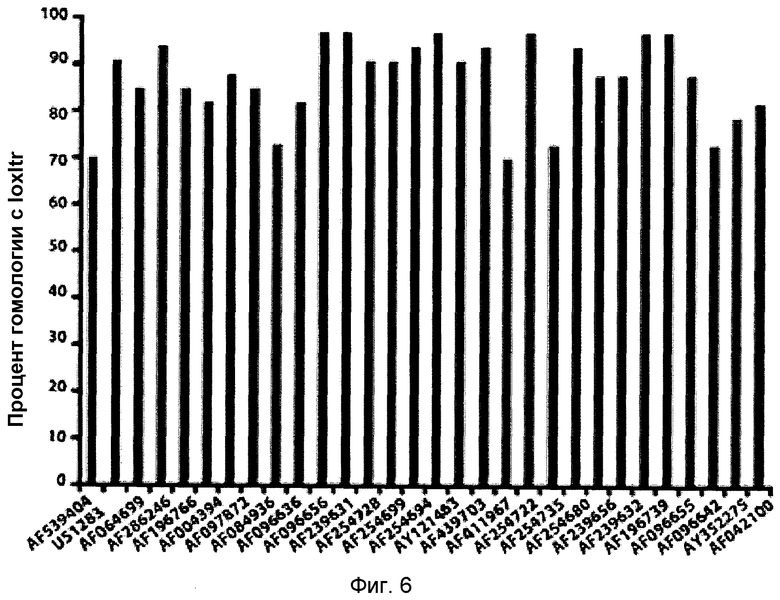



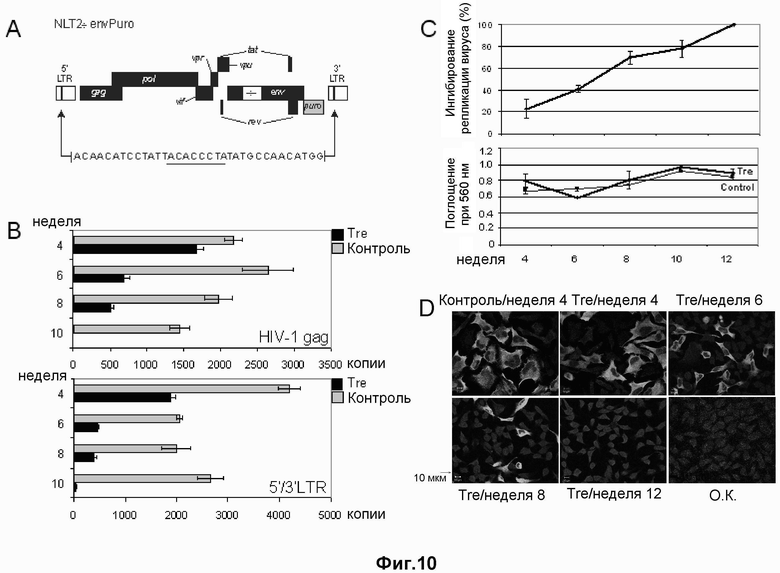

Изобретение относится к области биотехнологии и генной инженерии. Предложен способ получения вектора экспрессии, кодирующего адаптированную рекомбиназу. Указанная адаптированная рекомбиназа рекомбинирует асимметричные участки-мишени в LTR провирусной ДНК ретровируса, встроенного в геном клетки-хозяина, и пригодна в качестве средства для вырезания провируса из генома клетки-хозяина. Настоящее изобретение дополнительно относится к способу оптимизации лечения ретровирусной инфекции у индивида in vitro и к использованию адаптированных рекомбиназ для получения фармацевтических композиций для снижения вирусной нагрузки у индивида, инфицированного ретровирусом. 11 н. и 10 з.п. ф-лы, 11 ил., 1 табл., 4 пр.
1. Способ получения вектора экспрессии, кодирующего адаптированную рекомбиназу, которая рекомбинирует асимметричные последовательности-мишени в LTR провирусной ДНК, встроенной в геном клетки-хозяина, включающий стадии:
(a) определения последовательности LTR провирусной ДНК, идентификации в ней последовательностей, которые гомологичны с последовательностями левой половины участка и правой половины участка известных участков-мишеней рекомбиназ по меньшей мере на 50%, при этом гомологичные последовательности разделены спейсером из 5-12 нуклеотидов, а гомологичные последовательности LTR с наибольшей гомологией с известным участком-мишенью представляют асимметричную последовательность-мишень;
(b) получения двух синтетических последовательностей, где первая синтетическая последовательность соответствует последовательности асимметричной последовательности-мишени стадии (а), гомологичной левой половине участка указанного известного участка-мишени, плюс последовательность спейсера и ее обозначают как "последовательность половины участка 1", и где вторая синтетическая последовательность соответствует последовательности спейсера плюс последовательность асимметричной последовательности-мишени стадии (а), гомологичная правой половине участка, и ее обозначают как "последовательность половины участка 2";
(c) определения нуклеотидов в синтетических последовательностях стадии (b), отличающихся от соответствующих гомологичных последовательностей левой половины участка и правой половины участка известного гомологичного участка-мишени стадии (а);
(d) получения первой подгруппы двух последовательностей-мишеней на основе синтетических последовательностей стадии (b), где первая последовательность-мишень в первой подгруппе содержит инвертированный повтор, состоящий из последовательности половины участка 1 стадии (а) и последовательности половины участка 1′, разделенных последовательностью спейсера, и где вторая последовательность-мишень в первой подгруппе содержит инвертированный повтор, состоящий из последовательности половины участка 2′ и последовательности половины участка 2 стадии (b), разделенных последовательностью спейсера, где последовательности половин участков 1′ и 2′ представляют инвертированные повторы соответствующих последовательностей половин участков 1 и 2 стадии (b);
(e) получение второй подгруппы последовательностей-мишеней на основе последовательностей-мишеней первой подгруппы стадии (d), где каждую из последовательностей половин участков вместе с соответствующей последовательностью спейсера последовательностей-мишеней в первой подгруппе стадии (d) используют для получения независимой последовательности-мишени второй подгруппы, формируя инвертированный повтор на основании выбранной последовательности половины участка так, что последовательность спейсера разделяет обе последовательности, формируя инвертированный повтор, где последовательности обеих последовательностей половин участков, полученные из одной из последовательностей-мишеней первой подгруппы стадии (d), при их синтезе и до их использования изменяют с получением инвертированного повтора, формирующего полную последовательность-мишень так, что в последовательности левой половины участка часть нуклеотидов, отличающихся от соответствующей гомологичной последовательности половины участка известного участка-мишени стадии (а), замещена природными нуклеотидами, находящимися в известном участке-мишени, а в последовательности правой половины участка оставшиеся нуклеотиды, отличающиеся от соответствующей гомологичной левой половины участка, замещены природными нуклеотидами, находящимися в известном участке-мишени так, что в обеих последовательностях половин участков, происходящей из одной из последовательностей-мишеней первой подгруппы стадии (d), взятых вместе, можно найти все отличающиеся нуклеотиды, тогда как ни одна из указанных последовательностей половин участков отдельно не содержит все отличающиеся нуклеотиды;
(f) получения дополнительных подгрупп последовательностей-мишеней на основе последовательностей-мишеней из второй подгруппы, полученных на стадии (е), посредством пошагового повторения процесса стадии (е), каждый раз получая новую подгруппу последовательностей-мишеней, пока последовательности половин участков, формирующие инвертированные повторы в каждой полученной последовательности-мишени, содержат один, два или три нуклеотида, отличающихся от соответствующей гомологичной последовательности половины участка известного участка-мишени;
(g) применения направленной молекулярной эволюции для рекомбиназы, распознающей известный гомологичный участок-мишень, выбранный на стадии (а), с использованием в качестве субстрата последовательностей-мишеней из конечной подгруппы, полученной на стадии (f), содержащих один, два или три нуклеотида, отличающихся от соответствующей гомологичной последовательности половины участка указанного известного гомологичного участка-мишени;
(h) перестановки в библиотеках рекомбиназ, полученных на стадии (g);
(i) применения направленной молекулярной эволюции для библиотеки с перестановками, полученной на стадии (h), с использованием последовательностей-мишеней следующей подгруппы более высокого порядка в соответствии со стадией (f);
(j) повторения стадий (h) и (i) по меньшей мере до получения посредством направленной молекулярной эволюции одной рекомбиназы, которая активна в отношении асимметричной последователыюсти-мишени в LTR ретровирусной ДНК стадии (а);
(k) выделения из библиотеки нуклеиновой кислоты по меньшей мере одной рекомбиназы, полученной на стадии (j); и
(l) клонирования нуклеиновой кислоты, полученной на стадии (k), в подходящем векторе экспрессии.
2. Способ по п.1, где известная рекомбиназа, последовательность-мишень которой используют на стадии (а) и к последовательности-мишени которой применяют направленную молекулярную эволюцию на стадиях (g) и (i), относится к семейству сериновых интеграз или тирозиновых интеграз, где рекомбиназа выбрана из группы, состоящей из Сrе фага Р1, FLP дрожжей и Dre фага D6.
3. Способ по п.1, где асимметричная последовательность-мишень, идентифицированная на стадии (а), локализована в 5'-LTR и 3'-LTR провируса.
4. Способ по п.3, где провирусная ДНК, встроенная в геном клетки-хозяина, представляет собой ДНК ретровируса, выбранного из группы, состоящей из вируса опухоли молочной железы мышей (MMTV), вируса обезьян Мэйзена-Пфейзера (MPMV), вируса Т-клеточного лейкоза человека типа I (HTLV-I), вируса Т-клеточного лейкоза человека типа II (HTLV-II), вируса Т-клеточного лейкоза обезьян типа I (STLV-I), вируса Т-клеточного лейкоза обезьян типа II (STLV-II), вируса лейкоза коров (BLV), вируса лейкоза кошек (FeLV) и вируса лейкоза мышей Молони (MoMLV), где ретровирус предпочтительно представляет собой лентивирус, выбранный из группы, состоящей из вируса иммунодефицита человека типа 1 (ВИЧ-1), вируса иммунодефицита человека типа 2 (ВИЧ-2), вируса иммунодефицита обезьян (SIV), вируса иммунодефицита кошек (FIV), вируса иммунодефицита коров (BIV), вируса Maedi-visna (MW), вируса инфекционной анемии лошадей (EIAV) и вируса артрита-энцефалита коз (CAEV).
5. Способ по п.4, где асимметричная последовательность-мишень, идентифицированная на стадии (а), расположена в 5′-LTR и 3′-LTR провируса ВИЧ и предпочтительно имеет последовательность SEQ ID NO:3.
6. Способ по п.1, где применяемая направленная молекулярная эволюция представляет собой эволюцию на основе сшитых с субстратом белков.
7. Способ по п.1, где вектор экспрессии на стадии (1) представляет собой вектор экспрессии, выбранный из группы, состоящей из ретровирусных векторов, лентивирусных векторов, спумавирусных векторов и аденовирусных векторов, предпочтительно лентивирусный вектор выбран из группы, состоящей из происходящих от ВИЧ-1, SIV, FIV или EIAV лентивирусных векторов.
8. Способ получения адаптированной рекомбиназы, где указанный способ включает способ по любому из пп.1-7 и дополнительную стадию экспрессии адаптированной рекомбиназы или слитого полипептида, содержащего аминокислотную последовательность адаптированной рекомбиназы, кодирующей рекомбиназу нуклеиновой кислоты, встроенной в вектор экспрессии, в подходящей клетке-хозяине, где клетка-хозяин предпочтительно выбрана из группы, состоящей из бактериальных клеток, дрожжевых клеток, клеток насекомых и клеток млекопитающих.
9. Способ получения трансформированной стволовой клетки взрослого организма, включающий способ по любому из пп.1-7 и дополнительную стадию введения вектора экспрессии in vitro в подходящую стволовую клетку взрослого организма.
10. Нуклеиновая кислота, кодирующая адаптированную рекомбиназу, где указанная нуклеиновая кислота имеет последовательность SEQ ID NO:2.
11. Вектор экспрессии, полученный способом по любому из пп.1-7.
12. Адаптированная рекомбиназа для рекомбинации асимметричных последовательностей-мишеней в LTR провирусной ДНК, встроенной в геном клетки-хозяина, полученная способом по п.8.
13. Адаптированная рекомбиназа по п.12, где белок обладает аминокислотной последовательностью SEQ ID NO:3.
14. Слитый полипептид для рекомбинации асимметричных последовательностей-мишеней в LTR провирусной ДНК, встроенной в геном клетки-хозяина, полученный способом по п.8, где адаптированная рекомбиназа связана со вторым полипептидом, где второй полипептид содержит сигнальный пептид, где сигнальный пептид представляет собой домен белковой трансдукции, где домен белковой трансдукции представляет собой пептид ТАТ или пептид из третьей спирали гомеодомена Antennapedia.
15. Стволовая клетка взрослого организма для экспрессии рекомбиназы для рекомбинации асимметричных последовательностей-мишененй в LTR провирусной ДНК, встроенной в геном клетки-хозяина, полученная способом по п.9, которая представляет собой стволовую клетку из гемопоэтической линии дифференцировки.
16. Способ по любому из пп.1-9, дополнительно включающий стадию получения вектора экспрессии, белка, слитого белка или взрослой стволовой клетки в виде фармацевтической композиции для снижения вирусной нагрузки у индивида, инфицированного ретровирусом, где фармацевтическая композиция дополнительно содержит фармацевтически приемлемый носитель, эксципиент и/или вспомогательное средство.
17. Фармацевтическая композиция, полученная способом по п.16, где указанная фармацевтическая композиция снижает вирусную нагрузку у индивида, инфицированного ретровирусом, ниже 5000 эквивалентов генома/мл плазмы, предпочтительно ниже 500 эквивалентов генома/мл плазмы, а более предпочтительно ниже 50 эквивалентов генома/мл плазмы, где ретровирус представляет собой ретровирус, выбранный из группы, состоящей из вируса опухоли молочной железы мышей (MMTV), вируса обезьян Мэйзена-Пфейзера (MPMV), вируса Т-клеточного лейкоза человека типа I (HTLV-I), вируса Т-клеточного лейкоза человека типа II (HTLV-II), вируса Т-клеточного лейкоза обезьян типа I (STLV-I), вируса Т-клеточного лейкоза обезьян типа II (STLV-II), вируса лейкоза коров (BLV), вируса лейкоза кошек (FeLV) и вируса лейкоза мышей Молони (MoMLV), или лентивирус выбран из группы, состоящей из вируса иммунодефицита человека типа 1 (ВИЧ-1), вируса иммунодефицита человека типа 2 (ВИЧ-2), вируса иммунодефицита обезьян (SIV), вируса иммунодефицита кошек (FIV), вируса иммунодефицита коров (BIV), вируса Maedi-visna (MW), вируса инфекционной анемии лошадей (EIAV) и вируса артрита-энцефалита коз (CAEV).
18. Фармацевтическая композиция по п.17, где фармацевтическая композиция предназначена для совместного введения с другими активными средствами для высокоактивной противоретровирусной терапии (НААРТ) или для совместного или последовательного введения для терапии с общей активацией иммунной системы или специфической активацией экспрессии генов провируса.
19. Способ оптимизации лечения ретровирусной инфекции у индивида in vitro, включающий стадии:
(a) определения последовательности нуклеиновой кислоты ретровирусной ДНК, находящейся в образце крови пациента;
(b) сканирования последовательности LTR у последовательности стадии (а) на известные последовательности рекомбинации, для которых специфические адаптированные рекомбиназы уже получены;
(c) в случае когда присутствует по меньшей мере одна из указанных известных последовательностей рекомбинации, тогда стадию получения соединения, выбранного из группы, состоящей из вектора экспрессии, содержащего нуклеиновую кислоту адаптированной рекомбиназы, специфически распознающей указанную известную последовательность рекомбинации, указанной адаптированной рекомбиназы, слитого белка, содержащего аминокислотную последовательность указанной адаптированной рекомбиназы, или взрослой стволовой клетки, содержащей указанный вектор экспрессии, в виде фармацевтической композиции для снижения вирусной нагрузки у индивида; в противном случае, стадию идентификации последовательности с гомологией с последовательностями левой половины участка и правой половины участка известных участков-мишеней рекомбиназ соответственно по меньшей мере 50%, где гомологичные последовательности разделены подходящим спейсером из 5-12 нуклеотидов, и обозначают ее как "асимметричная последовательность-мишень";
(d) проведения стадий (b)-(l) способа по любому из пп.1-7 для получения вектора экспрессии, кодирующего адаптированную рекомбиназу, осуществляющую специфическую рекомбинацию асимметричной последовательности-мишени, идентифицированной в LTR провирусной ДНК по любому из пп.1-17; и
(e) получения вектора экспрессии, полученного на стадии (d), белка или слитого белка, экспрессированного с указанного вектора экспрессии, или стволовой клетки, трансфицированной или инфицированной указанным вектором экспрессии в виде фармацевтической композиции для снижения вирусной нагрузки у индивида,
где до получения указанной рекомбиназы в виде фармацевтической композиции проводят необязательную стадию тестирования in vitro эффективности рекомбиназы, идентифицированной на стадии (с) способа, или специфически адаптированной рекомбиназы, полученной на стадии (d) способа, в отношении вырезания провирусной ДНК ретровируса, которой инфицирован указанный индивид,
и/или предпочтительно способ дополнительно включает стадию включения адаптированной рекомбиназы, полученной на стадии (е), в коллекцию специфически адаптированных рекомбиназ.
20. Способ по п.19, где на стадии (d) направленную молекулярную эволюцию применяют для адаптированной рекомбиназы, уже распознающей участок-мишень, отличающийся от участков-мишеней рекомбиназ дикого типа.
21. Коллекция адаптированных рекомбиназ, подходящая для более быстрой продукции специфически адаптированных рекомбиназ по сравнению с рекомбиназами дикого типа, содержащая по меньшей мере одну адаптированную рекомбиназу по п.12, где представители указанной коллекции получены способом по любому из пп.1-8 или по любому из пп.19-20.
| BUCHHOLZ F | |||
| et al | |||
| Alteration of Cre recombinase site specificity by substrate-linked protein evolution// Nat Biotechnol | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| FLOWERS C.C | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Электрическое сопротивление для нагревательных приборов и нагревательный элемент для этих приборов | 1922 |
|
SU1997A1 |
| LEE Y | |||
| et al | |||
| A novel mutant loxP | |||

