ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
1. Область техники
Изобретение касается компьютерных способов идентификации и расчета малых молекул-миметиков эритропоэтина.
2. Описание предшествующего уровня техники
Эритропоэтин (ЕРО) является первичным регулятором пролиферации и дифференцировки незрелых эритроидных клеток. ЕРО производится в печени зародыша и почке взрослого в ответ на гипоксию (низкое содержание кислорода в крови и ткани). Он циркулирует в потоке крови, где находит ЕРО-рецепторы (EPOR) на коммитированных клетках-предшественниках в костном мозге и других кроветворных тканях. Рекомбинантный человеческий эритропоэтин (rHuEPO) широко используется при терапии пациентов с анемией вследствие хронической почечной недостаточности, химиотерапии рака и AZT-лечения.
Рецепторы ЕРО относятся к суперсемейству рецепторов цитокина, которое включает рецепторы для других кроветворных факторов роста, таких как интерлейкины (IL), колониестимулирующих факторов (CSF), а также гормона роста пролактина и цилиарного нейротрофического фактора (ciliary neurotrophic factor) (CNTF). Структурное строение этого семейства рецепторов состоит из трех модулей: лиганда, связывающего внеклеточный домен, короткой трансмембранной области и большого цитоплазматического домена. Предполагают, что внеклеточный домен данного суперсемейства включает два дискретных домена, каждый из которых содержит приблизительно 100 остатков, которые складываются в сандвич, состоящий из 7 антипараллельных β-цепей с топологией Ig неизменного домена. Все члены семейства имеют два характеристических мотива в своих внеклеточных доменах: пару консервативных дисульфидных мостиков в N-терминальном домене и WSXWS блок (где Х является любым аминокислотным остатком) в С-терминальном домене. Для большинства членов этого суперсемейства рецепторов существенной является олигомеризация одной или более полипептидных цепей для образования комплексов рецепторов с высоким сродством. Показано, что гомодимерный комплекс является активной формой hGHR, и похожую модель предлагают для рецепторов G-CSF, пролактина и ЕРО.
Эритропоэтин вызывает димеризацию двух молекул рецепторов ЕРО, результатом которой является последующее фосфорилирование цитоплазматических доменов посредством ассоциации с двумя молекулами тирозинкиназы (JAK2) для инициации каскада-событий, которые ведут к соответствующим биологическим проявлениям. Показав важность эритропоэтина, желательно иметь возможность идентификации молекул, способных связывать рецепторы ЕРО и вызывать реакцию, обычно вызываемую посредством ЕРО.
Из публикаций WO 96/40772 и WО 94/25055 и статьи Wrighton (1996), Science, том 273, стр. 458-462, которые можно считать уровнем техники, известны способы идентификации молекул, которые присоединяются к рецепторам ЕРО и действуют как миметики ЕРО, но ни в одной из этих публикаций не говорится о кристаллической структуре частицы и о координатах атомов, т.е. отсутствует информация, указанная в таблице.
КРАТКОЕ СОДЕРЖАНИЕ ИЗОБРЕТЕНИЯ
Данное изобретение представляет способы идентификации молекул, которые присоединяются к рецепторам ЕРО и действуют как миметики ЕРО. Предпочтительные миметики ЕРО, идентифицированные с использованием способа данного изобретения, действуют как агонисты рецепторов ЕРО в одном или более in vitro и in vivo биологических исследованиях активности ЕРО. Предпочтительными миметиками являются молекулы, не имеющие пептидных связей, то есть не являющиеся пептидными миметиками. Предпочтительные пептидные миметики имеют 15 или более, предпочтительно 10 или более, аминокислот.
Способы данного изобретения обеспечивают идентификацию и разработку молекул с конкретной структурой. Способы уверяют в использовании точной структурной информации, полученной из рентгеновских кристаллографических исследований внеклеточного домена рецептора ЕРО (аминокислоты от 1 до 225) в комплексе с пептидом, ЕМР1 (Пептид 1 - миметик ЕРО; описанный ниже), который действует как миметик ЕРО. Эти кристаллографические данные позволяют идентифицировать атомы в пептидном миметике, которые важны для связывания рецептора ЕРО и димеризации. Более важно, что эти данные определяют трехмерное пространство важных контактных атомов. Другие молекулы, которые включают часть, в которой, атомы имеют похожее трехмерное окружение, аналогичное для некоторых или для всех этих контактных атомов, вероятно, также способны действовать как миметики ЕРО. Кроме того, можно использовать структурную информацию для разработки или идентификации молекул, имеющих даже большую ЕРО-активность, чем описанный здесь пептидный миметик.
Далее представлены подробности предпочтительных вариантов настоящего изобретения, снабженные чертежами и описанием. Вместе с тем, что подробности данного изобретения известны, многочисленные дополнительные новшества и изменения будут очевидны специалистам.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 представлена блок-схема, изображающая первый способ идентификации потенциальных миметиков эритропоэтина с использованием компьютерной системы.
На фиг. 2 представлена блок-схема, изображающая второй способ идентификации потенциальных миметиков эритропоэтина с использованием компьютерной системы.
Аналогичные номера ссылок и обозначений на различных чертежах указывают аналогичные элементы.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Представленные в данном описании предпочтительные варианты и примеры правильнее считать иллюстрациями, а не ограничениями настоящего изобретения.
Ниже описана кристаллическая структура малого пептида - миметика ЕРО, связанного с внеклеточной частью рецептора ЕРО. Пептид ЕМР1 (GGTYSCHFGPLTWVCKPQGG; SEQ ID NО:l) отличается наличием внутримолекулярного дисульфидного мостика. Некоторые очевидные признаки говорят о том, что ЕРМ1 может действовать как миметик ЕРО. Например, ЕРМ1 конкурирует с ЕРО в исследованиях по связыванию рецепторов и индуцирует пролиферацию клеточных линий, составленных таким образом, что они легко реагируют на ЕРО. И ЕРО, и пептид вызывают аналогичный каскад стадий фосфорилирования и прогресс клеточного цикла в ЕРО-чувствительных клетках. Кроме того, ЕРМ1 демонстрирует значительные эритропоэтические эффекты на мышах, что контролируют посредством двух различных in vivo исследований процесса возникновения красных кровяных клеток. Эти данные при их объединении строго подтверждают представление, что пептидный лиганд, который имеет последовательность, не имеющую отношения к последовательности ЕРО, способен связывать рецептор ЕРО и индуцировать конформацию или устройство агониста рецептора ЕРО.
Разработка малых молекул - миметиков
Структура диаметра ЕМР1 демонстрирует, что молекула, существенно меньшая, чем природный гормон, может действовать как агонист и вызывать подходящую биологическую реакцию. Предполагают, что пептид имеет существенно меньшую поверхность взаимодействия с рецептором, чем природный гормон. Определяющие факторы связывания в рецепторе ЕРО образуют почти плоскую поверхность, которая в основном гидрофобна по природе, без полостей или заряженных остатков, которые могут помогать в расчете взаимодействия малой молекулы - лиганда с рецептором.
Этот упрощенный каркас взаимодействий, показанный присутствующими здесь структурными данными, можно использовать для идентификации дополнительных миметиков ЕРО. Атомы ЕМР1, которые важны для связывания рецептора ЕРО и образования димерного рецептора ЕРО, включают атомы, вовлеченные в контакт между ЕМР1 (пептид) и ЕВР (рецептор ЕРО), и атомы, вовлеченные в контакт между двумя молекулами ЕМР1 в димерном комплексе (пептид-пептидные контакты). Кроме того, в противоположность перечисленным в таблице 2, в каждом пептиде важными являются следующие ЕМР1-ЕМР1 гидрофобные контакты: ТyrP4, CysР6, PheP8, TRPP13 и CysP15. Важны также следующие ЕМР1-ЕВР гидрофобные взаимодействия в каждом пептиде: ТуrP4, PheP8 и ТrрP13 и СуsP15. Специалисту также будет понятно, что не все имеющиеся в существенных контактных остатках атомы должны присутствовать в миметике. В действительности, это только те несколько атомов из важных контактов с рецептором ЕРО, которые фактически важны для активности миметика. Специалисты способны идентифицировать эти важные атомы на основе модели димерного комплекса ЕМР1-ЕРО, который можно сконструировать, используя имеющиеся структурные данные.
Предпочтительные миметики включают атомы в положениях, аналогичных положениям рецептора ЕРО, контактирующих с атомами ЕМР1. Даже более предпочтительные миметики структурно похожи на димер ЕМР1, обнаруженный в описанной ниже структуре. Это происходит потому, что димеризация ЕМР1 является важным фактором в димеризации рецептора ЕРО.
Способ данного изобретения использует компьютерные способы идентификации соединений, имеющих необходимую структуру. Более подробно, для определения пептидных и непептидных кандидатов в миметики посредством компьютерных способов данное изобретение применяет трехмерные координаты подмножества атомов в пептиде GGTYSCHFGPLTWVCKPQGG, при совместной кристаллизации пептида с частью рецептора эритропоэтина, включающей аминокислоты с 1 по 225 рецептора эритропоэтина человека.
Эти компьютерные способы распадаются на два широких класса: способы базы данных и способы компьютерного расчета. В способах базы данных интересующее соединение сравнивают со всеми соединениями, имеющимися в базе данных химических структур, и идентифицируют соединения, структуры которых некоторым образом похожи на рассматриваемое соединение. Структуры в базе данных основаны на экспериментальных данных, полученных способами ЯМР или рентгеновской кристаллографии, или являются моделированными трехмерными структурами на основе двумерных данных (то есть последовательность). В способах компьютерного расчета модели соединений, структуры которых некоторым образом похожи на интересующее соединение, генерируют при помощи компьютерной программы, используя информацию, полученную из известных структур, например, данных рентгеновской кристаллографии и/или теоретических закономерностей. Такие способы разработки могут построить соединение с нужной структурой либо соединяя атом-за-атомом, либо посредством сборки имеющихся фрагментов малых молекул.
Успех обоих способов (базы данных и расчета) в идентификации соединений с активностью, аналогичных рассматриваемому соединению, зависит от идентификации функционально уместной части рассматриваемого соединения. Для лекарственных препаратов функционально уместная часть называется фармакофором. Фармакофор - окружение структурных признаков и функциональных групп, важных для биологической активности, например, ЕРО-активности.
Не все идентифицированные соединения, имеющие нужный фармакофор, действует как ЕРО-миметики. Фактическую активность можно окончательно определить только путем измерения активности соединения в подходящих биологических опытах. Однако способы данного изобретения предельно полезны, так как их можно применять для значительного снижения количества соединений, подлежащих исследованию с целью идентификации действительного миметика.
Димеризация рецепторов ЕРО важна для активности. Следовательно, предпочтительные миметики основаны на структуре димера ЕМР1, так как он присоединяется к димеру рецептора ЕРО. Таким образом, предпочтительные миметики включают важные контакты от обоих RWJ 61233 пептидов, присутствующих в описанной ниже структуре. Такие миметики способствуют димеризации рецептора ЕРО.
Программы, подходящие для построения предсказанных трехмерных структур из двумерных данных, включают Concord (Tripos Associated, St. Louis, МО), 3-D Builder (Chemical Design Ltd., Oxford, U.K.), Catalyst (Bio-CAD Corp., Mountain View, CA) и Daylight (Abbott Laboratories, Abbott Park, IL).
Программы, подходящие для поиска трехмерных баз данных для идентификации молекул, несущих нужный фармакофор, включают: MACCS-3D и ISIS/3D (Molecular Design Ltd., San Leandro, CA), ChemDBS-3D (Chemical Design Ltd., Oxford, U. K.) и Sybyl/3DB Unity (Tripos Associates, St. Louis, МО).
Программы, подходящие для выбора и разработки фармакофора, включают: DISCO (Abbott Laboratories, Abbott Park, IL), Catalyst (Bio-CAD Corp., Mountain View, CA) и ChemDBS-3D (Chemical Design Ltd., Oxford, U.K.).
Базы данных химических структур доступны от Cambridge. Crystallographic Data Centre (Cambridge, U.K.) и Chemical Abstracts Service (Columbus, ОН).
Программы компьютерного расчета включают Ludi (Biosym Technologies Inc., San Diego, CA) и Aladdin (Daylight Chemical Information Systems, Irvine CA).
Специалисты поймут, что расчет миметика может потребовать незначительной структурной деформации или подгонки рассчитанной или идентифицированной способами данного изобретения химической структуры.
Химические соединения, идентифицированные или рассчитанные способами данного изобретения, в основном можно синтезировать химическими способами и затем исследовать на ЕРО-активность, используя любой из способов, описанных ниже. Способы данного изобретения особо полезны потому, что их можно применять для существенного увеличения количества потенциальных миметиков, которые следует проверить на ЕРО-активность.
Это изобретение можно осуществить на аппаратуре, или программном обеспечении, или комбинации обоих. Однако предпочтительно воплощать данное изобретение на компьютерных программах, выполняемых на программируемых компьютерах, каждые из которых включают процессор, систему хранения информации (включая энергозависимое и энергонезависимое запоминающие устройства и/или запоминающие элементы), по крайней мере, одно устройство ввода и, по крайней мере, одно устройство вывода. Систему программирования применяют к введенным данным, выполняя описанные выше функции и получая выводимую информацию. Выводимую информацию подают известным способом на одно или более устройств вывода. Компьютер может быть, например, персональным компьютером, микрокомпьютером или рабочей станцией обычного назначения.
Для взаимодействия с компьютерной системой каждую программу предпочтительно пишут на процедурном или проблемно-ориентированном языке высокого уровня. Однако, если требуется, программы можно выполнять на языке ассемблера или машинном языке. В любом случае язык может быть транслированным или интерпретированным языком.
Каждую такую компьютерную программу предпочтительно хранят на носителях данных или устройствах (например, ROM или магнитная дискета), читаемых основным компьютером или программируемым компьютером специального назначения, для ее установки и действия компьютера, когда носитель или устройство читается компьютером для выполнения описанных здесь процедур. Можно также считать, что заявляемая система выполнена как читаемый компьютером носитель информации, совместимый (configured) с компьютерной программой, где носитель информации в данной конфигурации заставляет компьютер действовать специфическим и предварительно определенным образом для выполнения описанных здесь функций.
На фиг. 1 приведена блок-схема, показывающая первый способ идентификации потенциальных миметиков эритропоэтина с использованием компьютерной системы. Способ использует программируемый компьютер, включающий процессор, систему хранения информации, по крайней мере, одно устройство ввода и, по крайней мере, одно устройство вывода, и включает стадии:
(1) Ввод в программируемый компьютер через устройство ввода данных, содержащих трехмерные координаты подмножества атомов в пептиде GGTYSCHFGPLTWVCKPQGG при совместной кристаллизации пептида с частью рецептора эритропоэтина, включающей аминокислоты с 1 по 225 рецептора, генерируя при этом набор оценочных данных (Стадия 100);
(2) Сравнение, используя процессор набора оценочных данных с компьютерной базой данных химических структур, хранящейся в системе хранения информации компьютера (Стадия 102);
(3) Выбор из базы данных химических структур, имеющих структурно аналогичную набору оценочных данных часть, используя программу, подходящую для поиска трехмерных баз данных для идентификации молекул, несущих нужный фармакофор (такой, как описанные выше или эквивалентные) (Стадия 104);
(4) Вывод на выводное устройство выбранных химических структур, имеющих структурно аналогичную набору оценочных данных часть (Стадия 106).
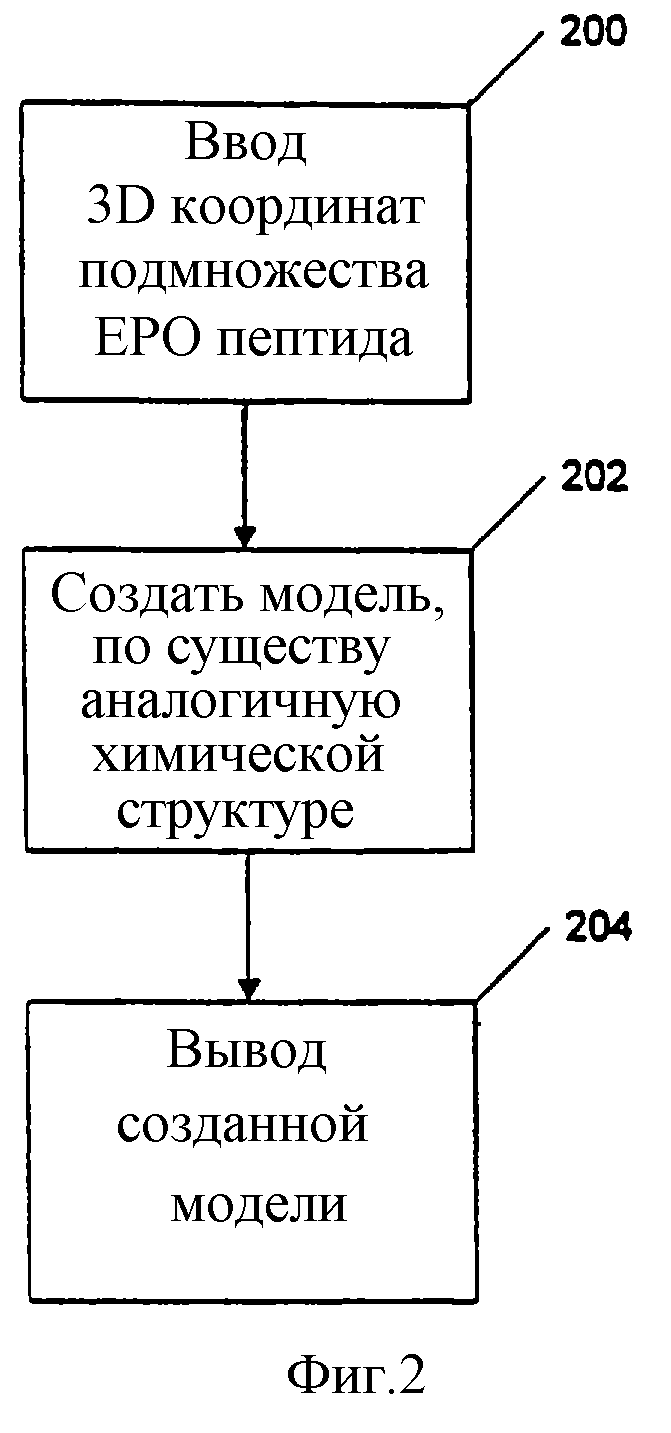
На фиг. 2 приведена блок-схема, показывающая второй способ идентификации потенциальных миметиков эритропоэтина с использованием компьютерной системы. Способ использует программируемый компьютер, включающий процессор, систему хранения информации, по крайней мере, одно устройство ввода и, по крайней мере, одно устройство вывода, и включает стадии:
(1) Ввод в программируемый компьютер через устройство ввода данных, содержащих трехмерные координаты подмножества атомов в пептиде GGTYSCHFGPLTWVCKPQGG при совместной кристаллизации пептида с частью рецептора эритропоэтина, включающей аминокислоты с 1 по 225 рецептора, генерируя при этом набор оценочных данных (Стадия 200);
(2) Создание модели химической структуры, имеющей структурно аналогичную набору оценочных данных часть, используя программу, подходящую для расчета моделей химических структур (Стадия 202);
(3) Вывод на выводное устройство созданной модели (Стадия 204).
Подтверждение биологической активности
Для определения может ли молекула, идентифицированная с использованием способов данного изобретения, действовать как миметик ЕРО, необходимо выполнить одно или более in vitro или in vivo исследований ЕРО-активности. Например, молекулы-миметики должны обладать способностью стимулировать. пролиферацию клеток TF-1 (Kitamura и др., J. Cell Physiol. 140:323, 1985) или клеток B6Sut (Greenberger и др., Ргос. Natl. Acad. Sci. USA 80:2931, 1983), но предпочтительно не стимулировать пролиферацию клеток, которые не несут рецепторы ЕРО. Таким образом, предпочтительные миметики не стимулируют пролиферацию клеток Мо7е (Avanzi и др., Br. J. Haematol. 69:359, 1988).
Потенциальные миметики можно также исследовать на мышиной модели эритропоэза. В данном исследовании потенциальные миметики вводят нормальным мышам, которые проявляют эндогенные базальные уровни ЕРО. Подсчитывают ретикулоциты, предпочтительно способом цитометрии в потоке, определяя, увеличивает ли кандидат в миметики уровень ретикулоцитов. Повышение содержания ретикулоцитов указывает, что кандидат в миметики стимулирует эритропоэз. Так как использованные в этом исследовании мыши уже обеспечивают экспрессию ЕРО, это исследование может быть относительно нечувствительным. В качестве альтернативы потенциальные миметики можно исследовать в биологических опытах на мышах с эксипоксией-полицитемией (exhypoxic-polycythemic). В этом исследовании полицитемию индуцируют посредством выдерживания мышей в камере с пониженным давлением для снижения содержания ЕРО. Подготовленным мышам можно вводить потенциальные миметики ЕРО. Включение в кровь 59Fe служит в качестве меры эритропоэза. Этот эритропоэз может быть присущ кандидатам в миметики.
Описанные выше опыты являются примерами подходящих исследований. Полезны также другие исследования ЕРО-активности, известные специалистам.
Для определения биологической активности кандидата в миметики предпочтительно определять биологическую активность при нескольких концентрациях кандидата в миметики. Активность при данной концентрации кандидата в миметики можно сравнить с активностью самого ЕРО.
Структурные данные
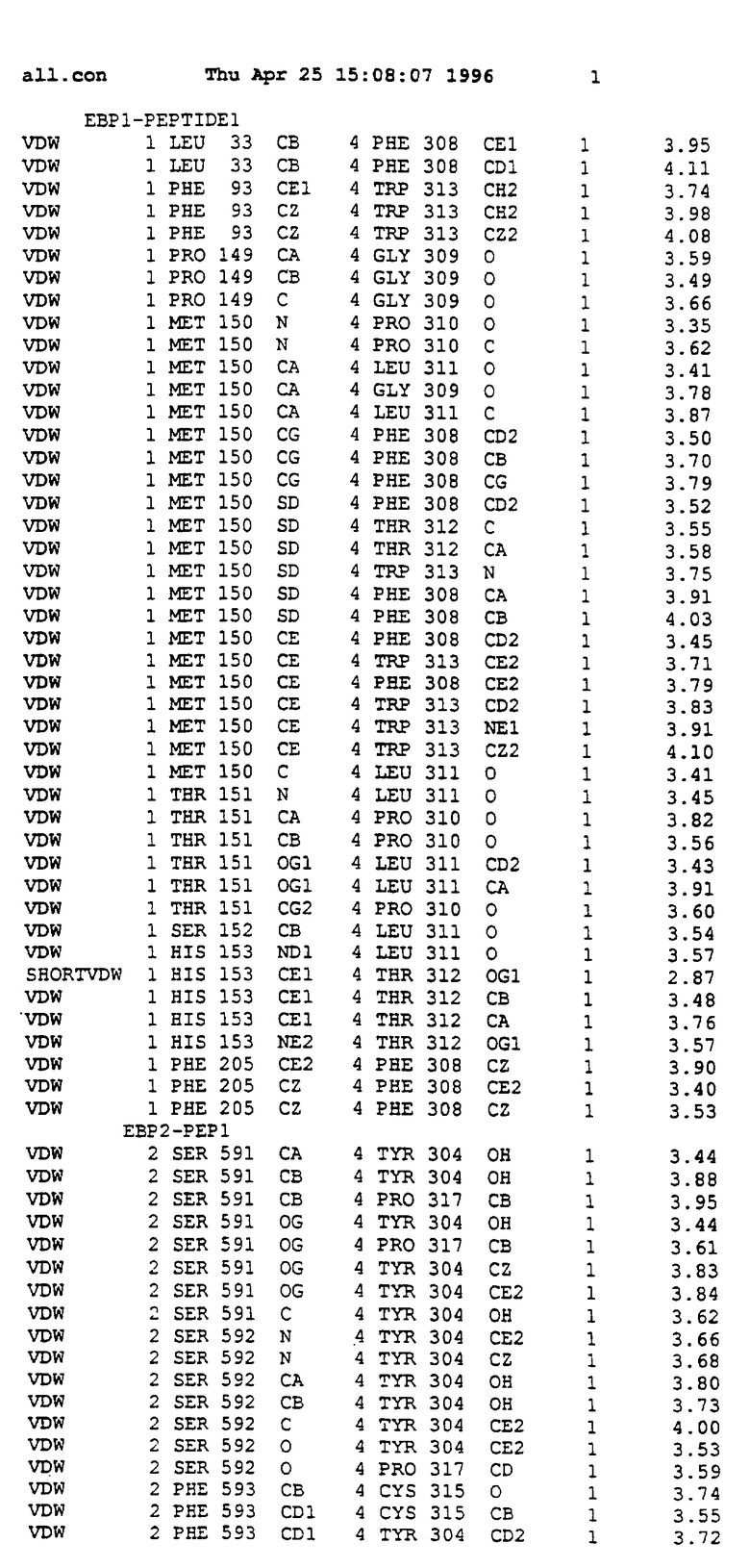
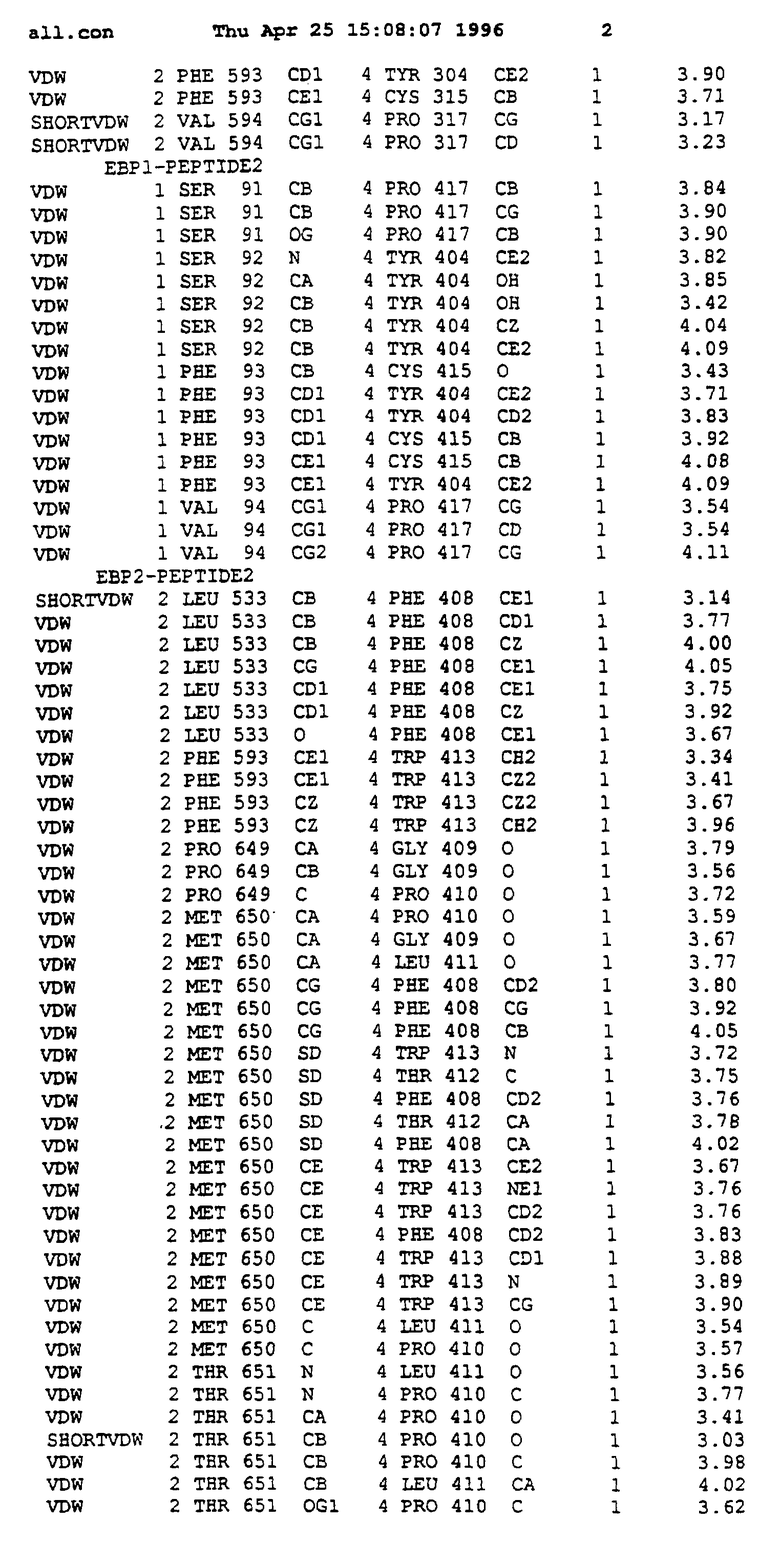
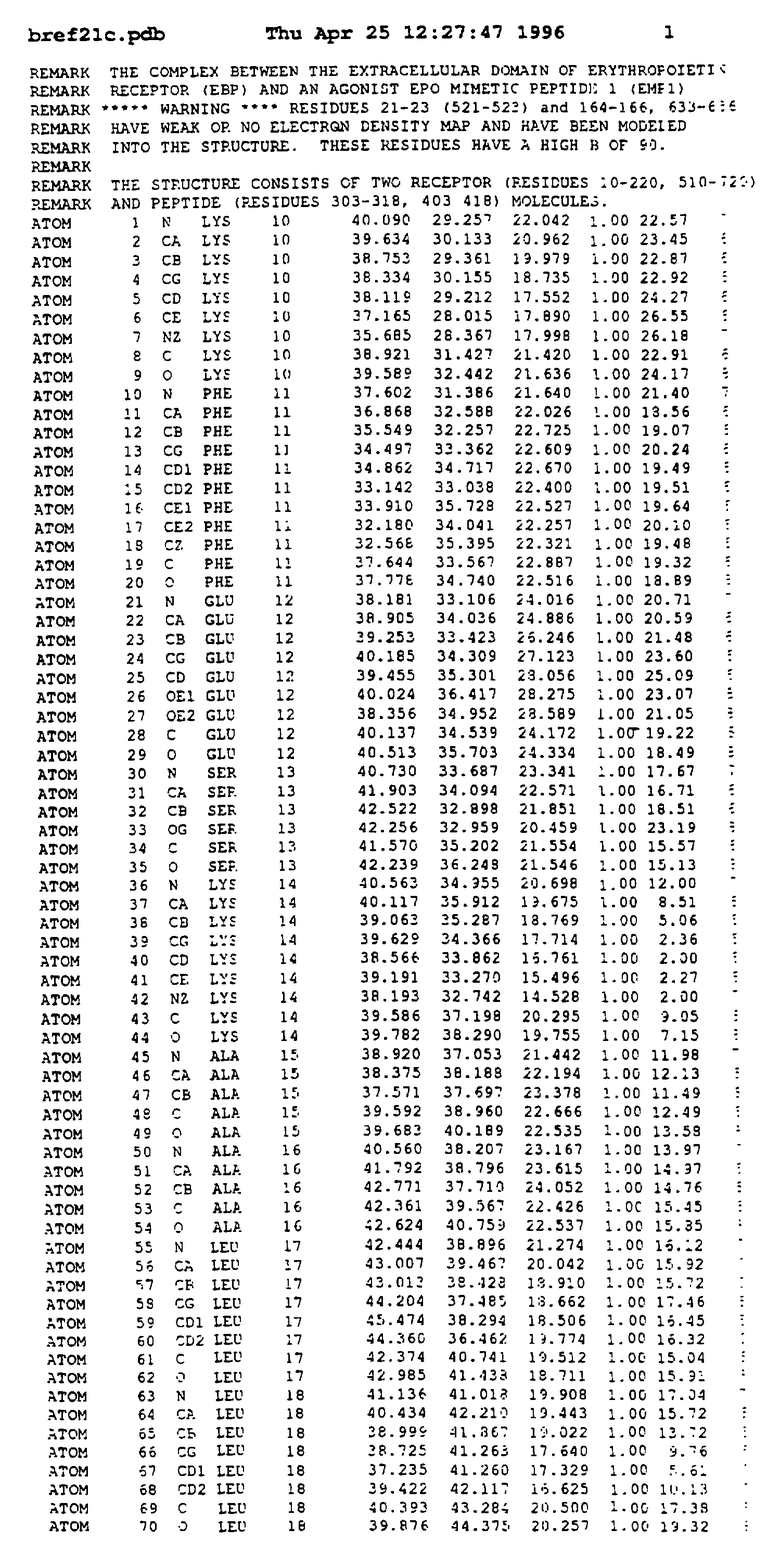
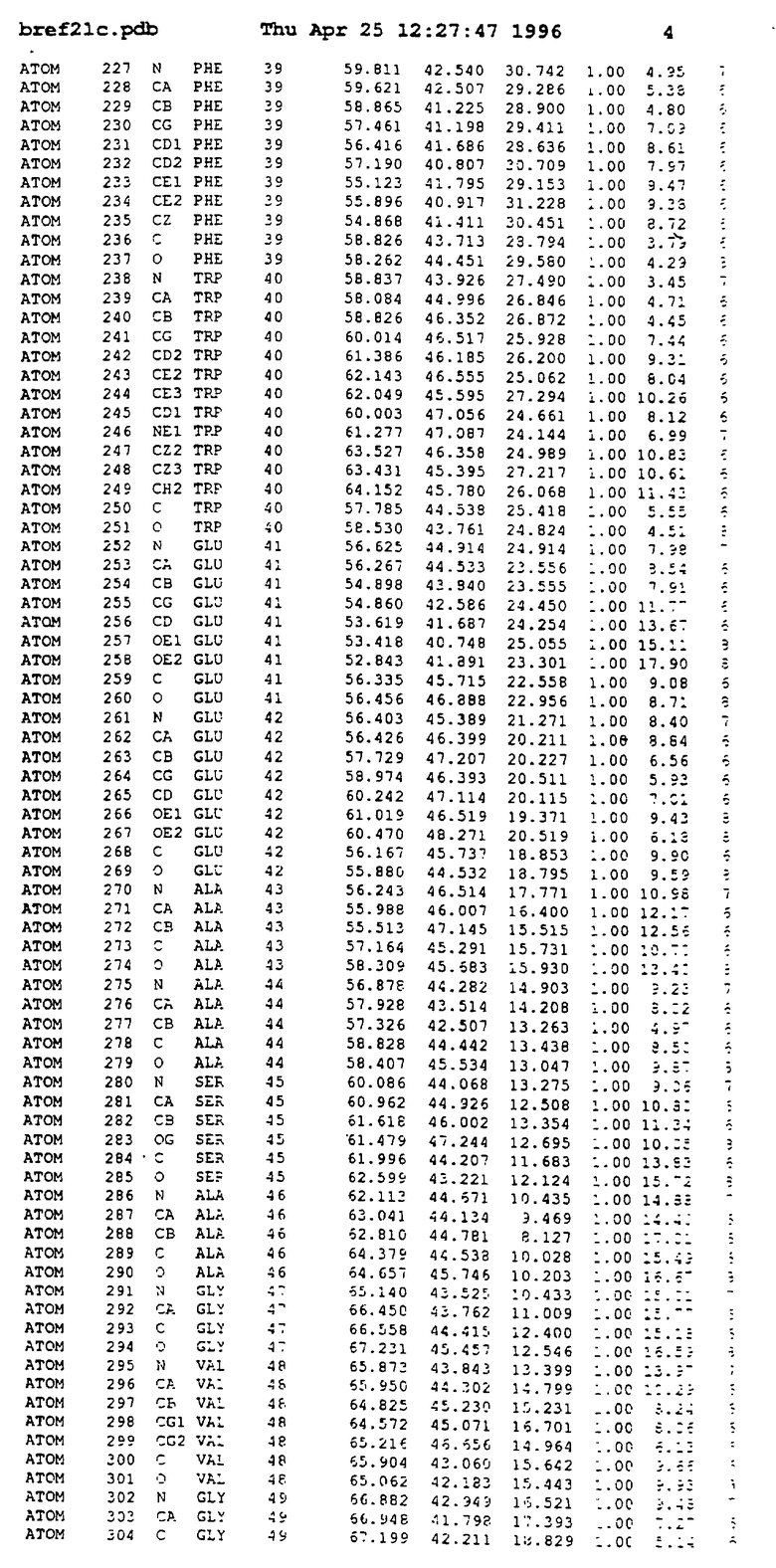
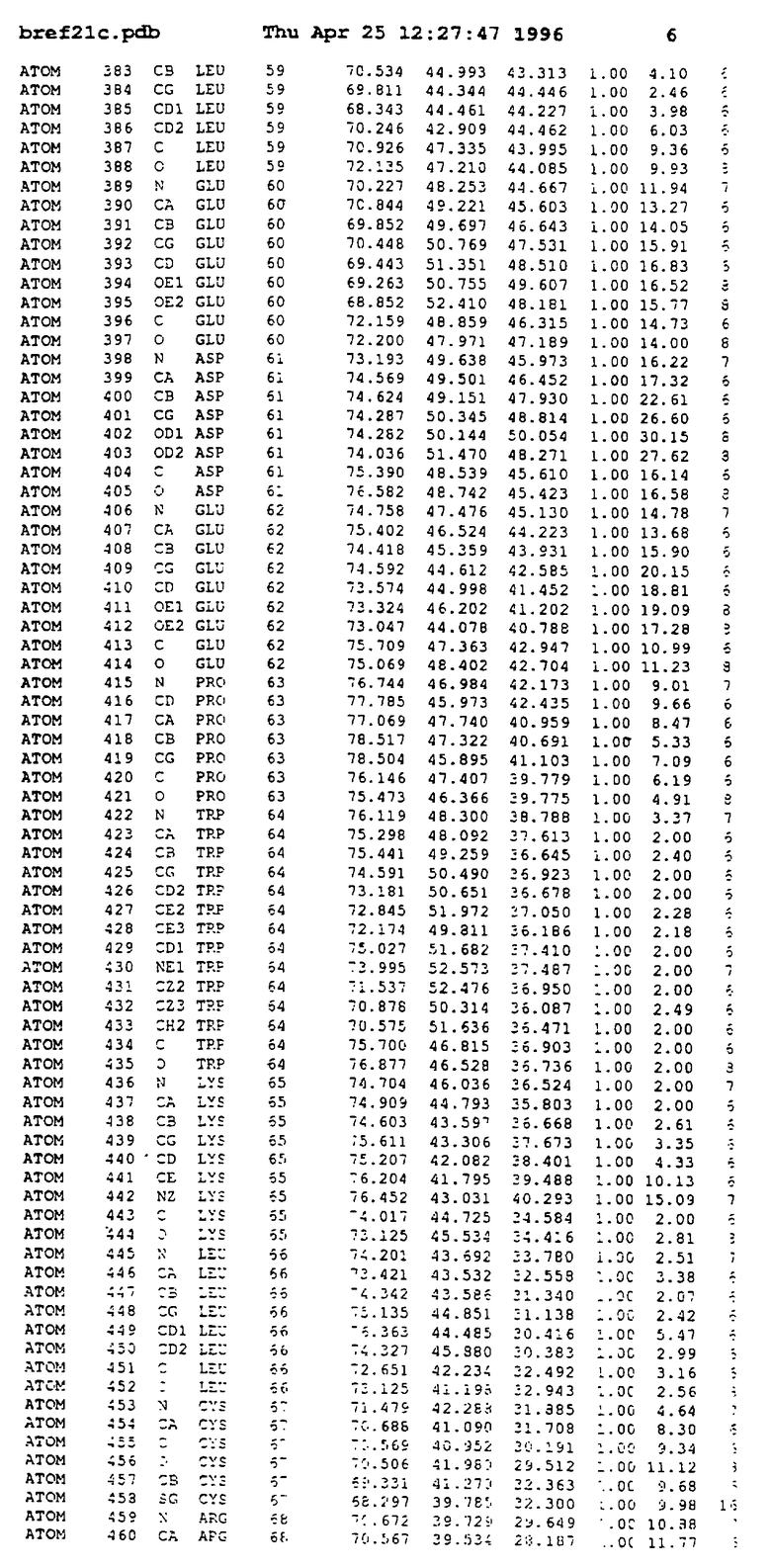
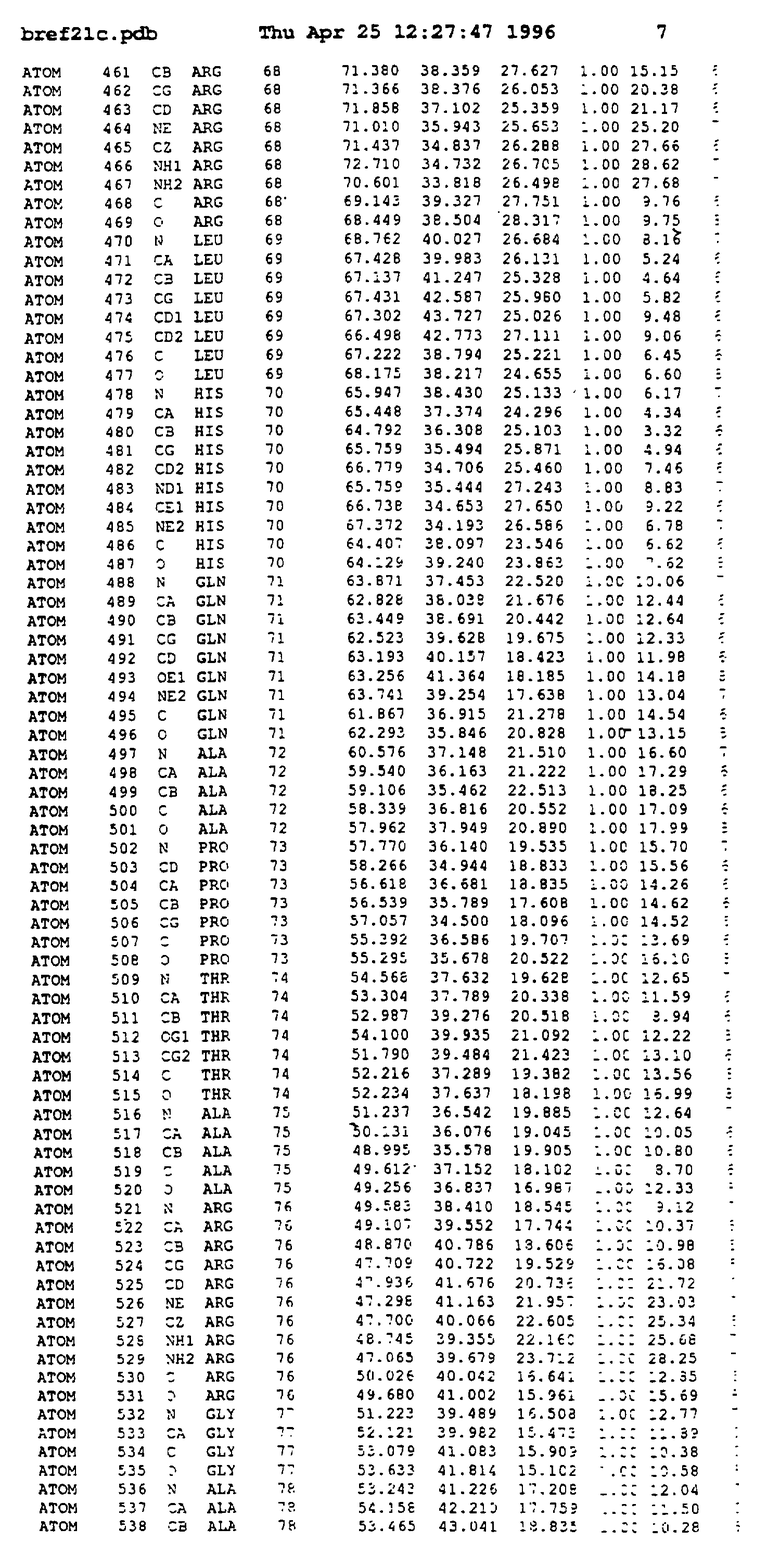
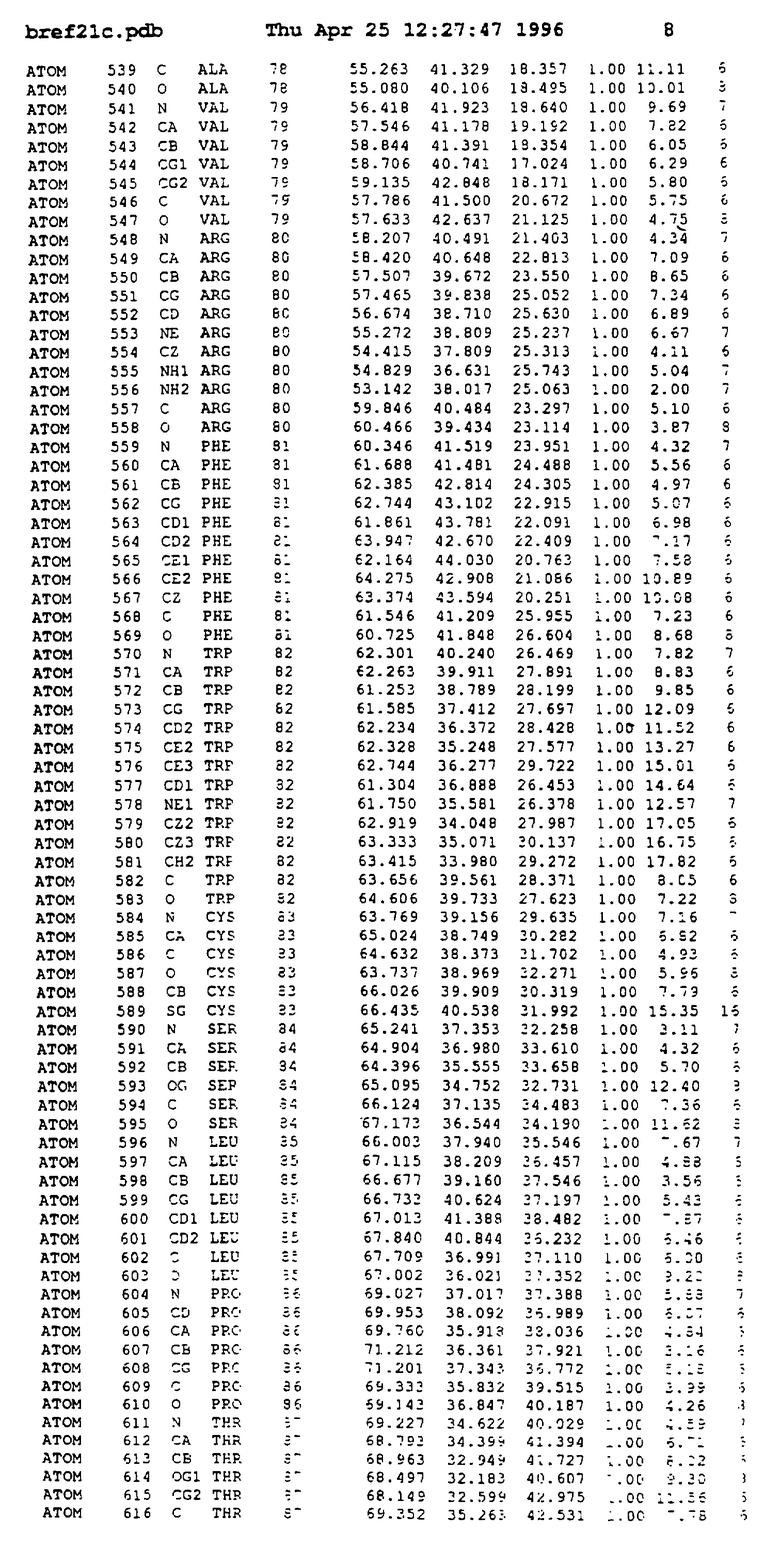
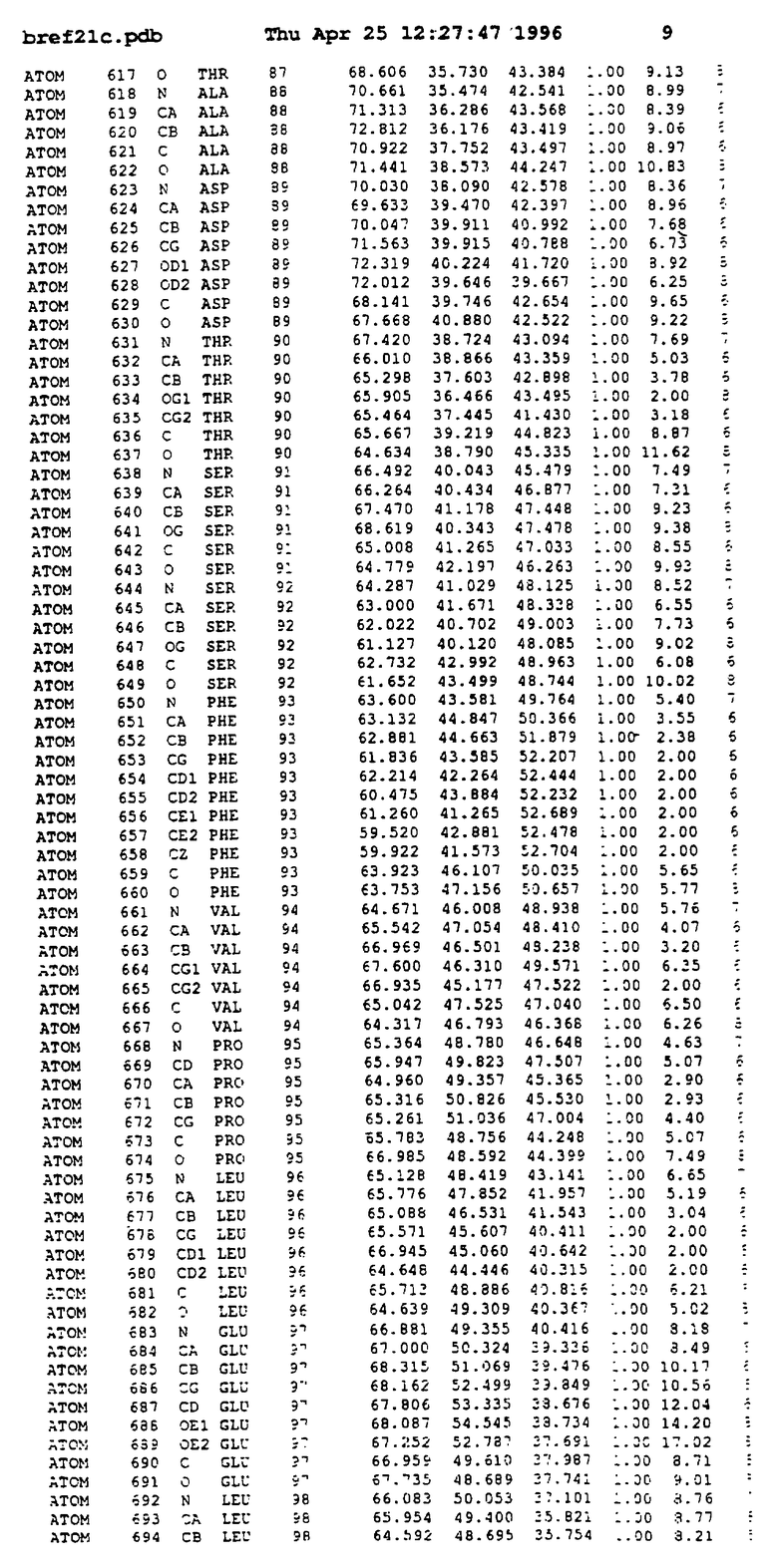
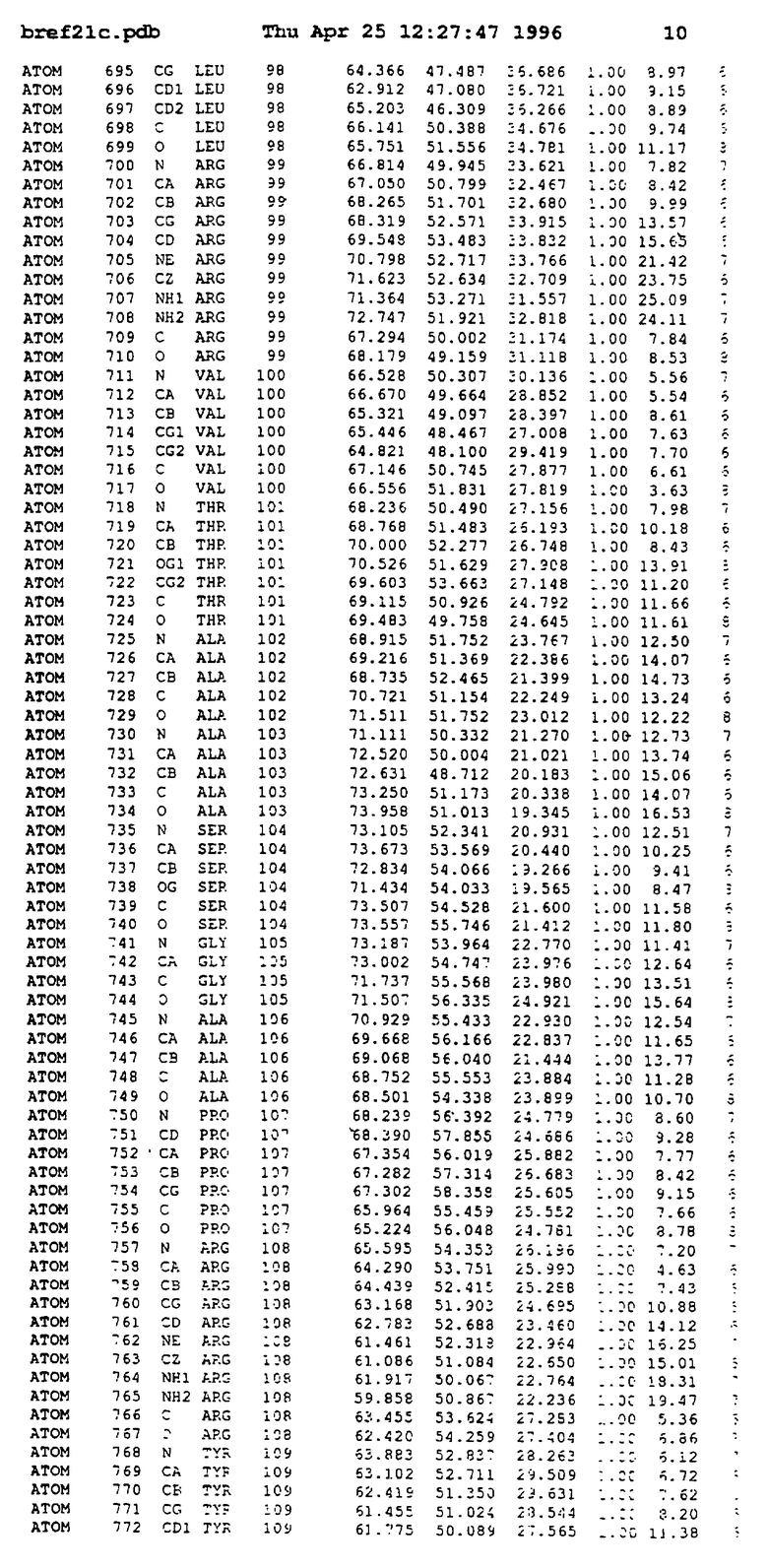
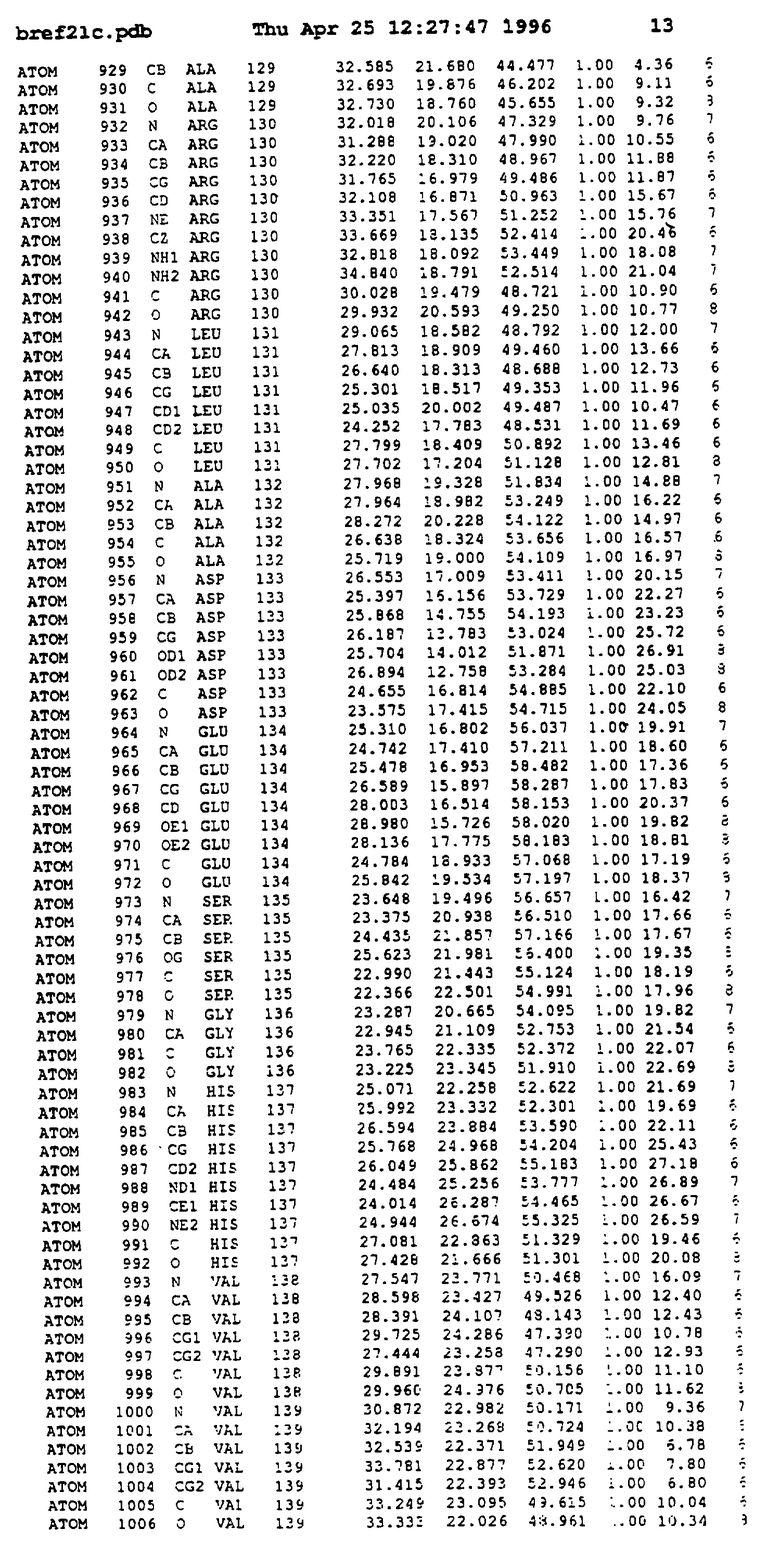
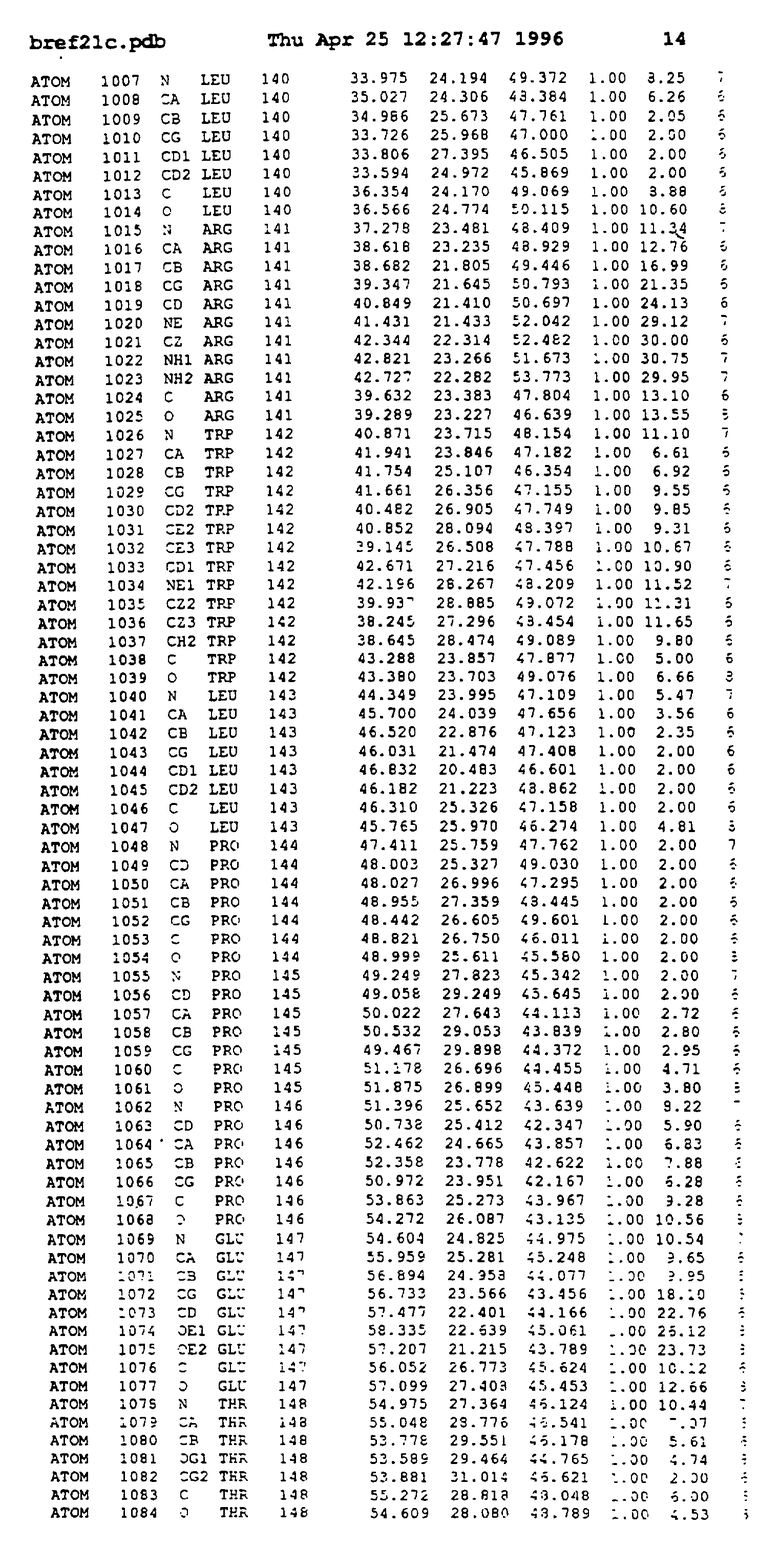
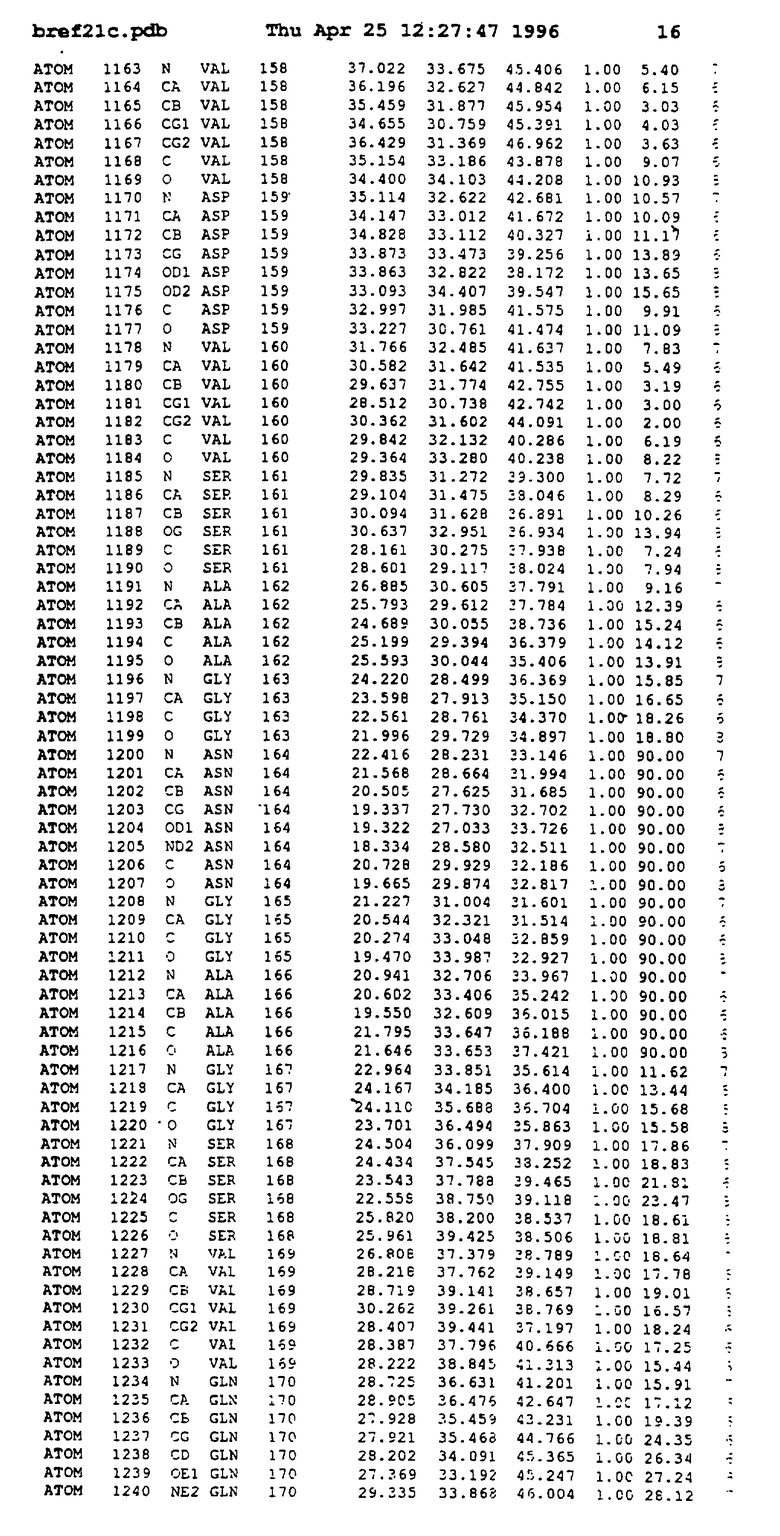
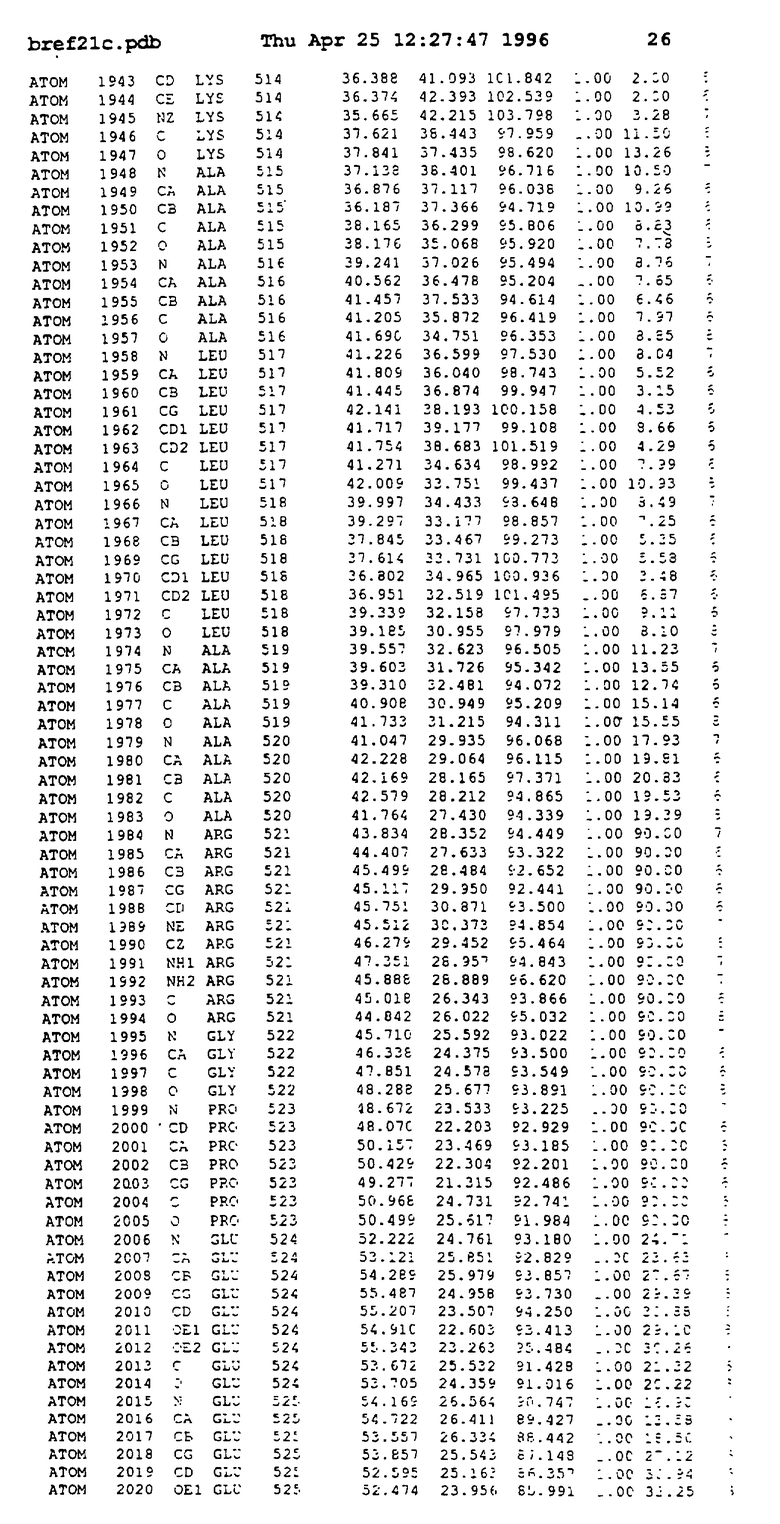
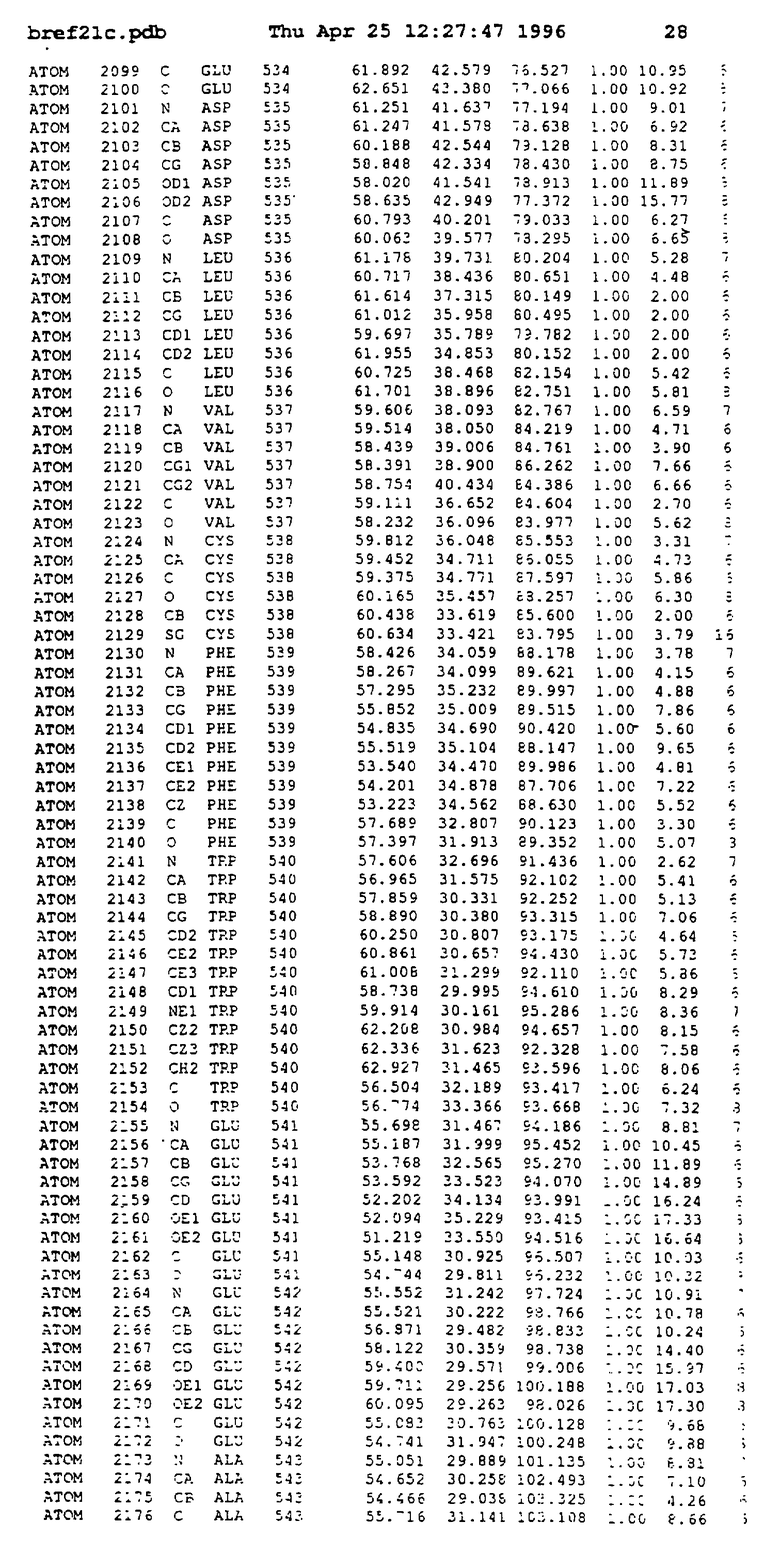
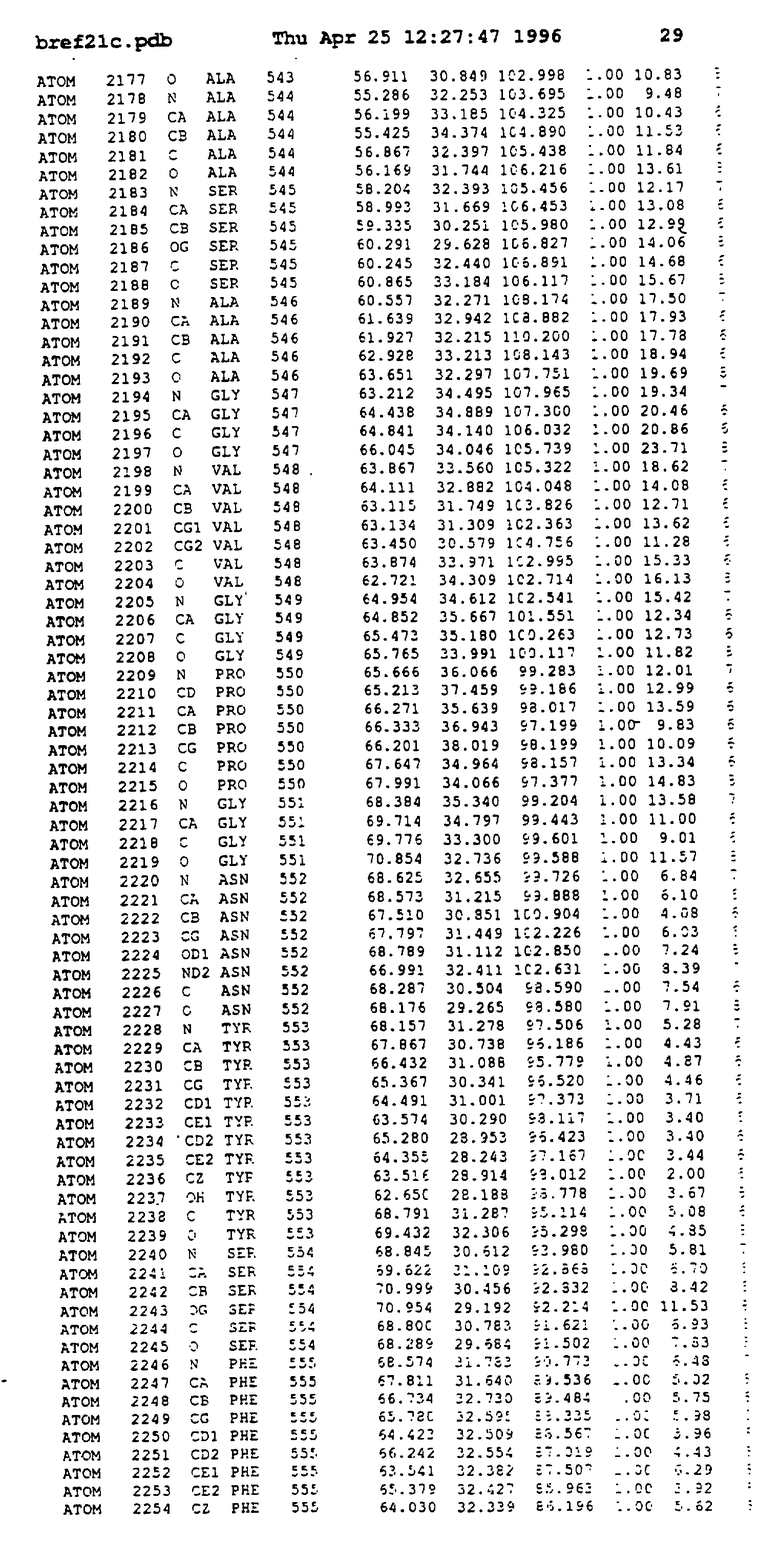
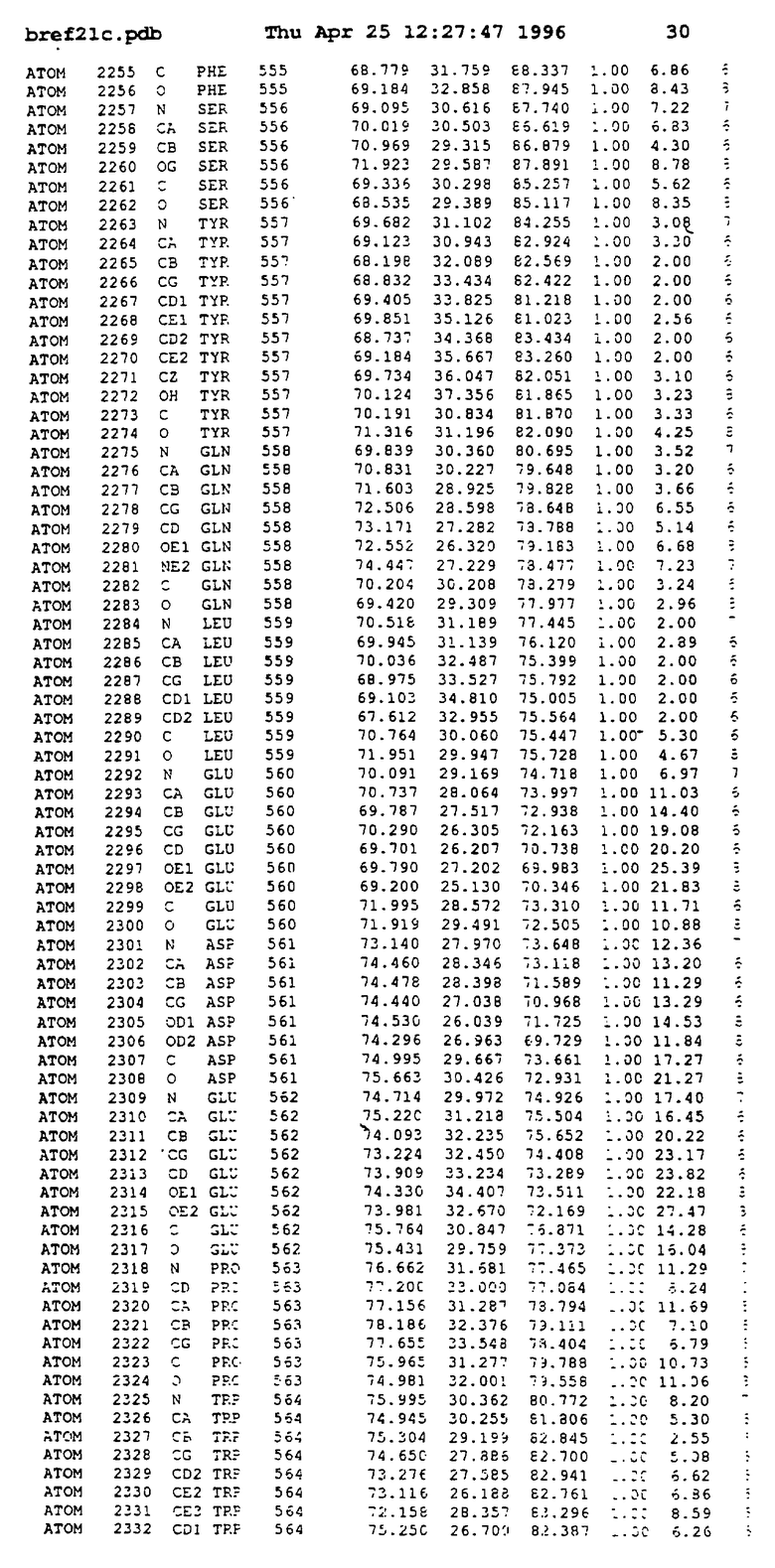
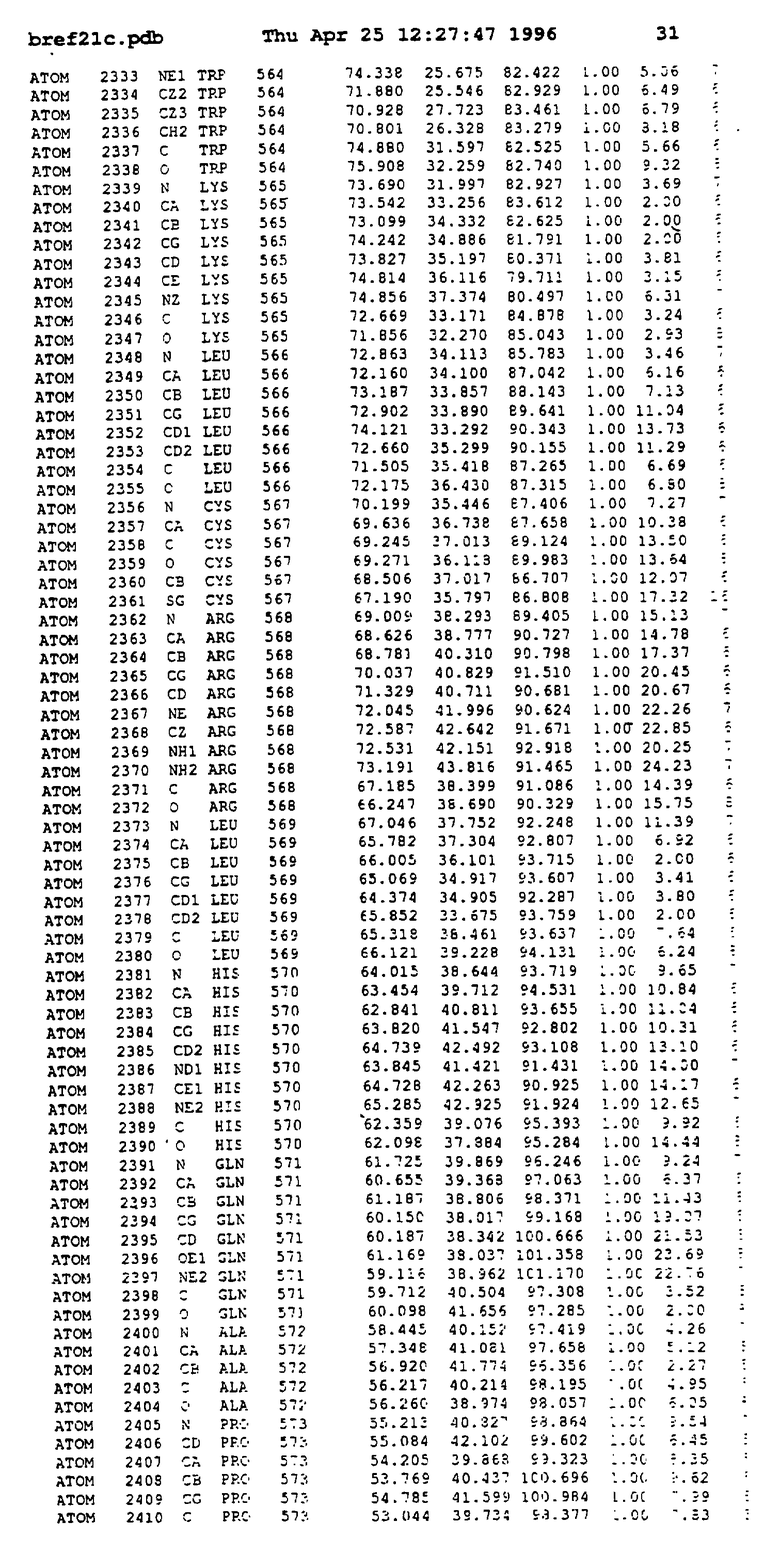
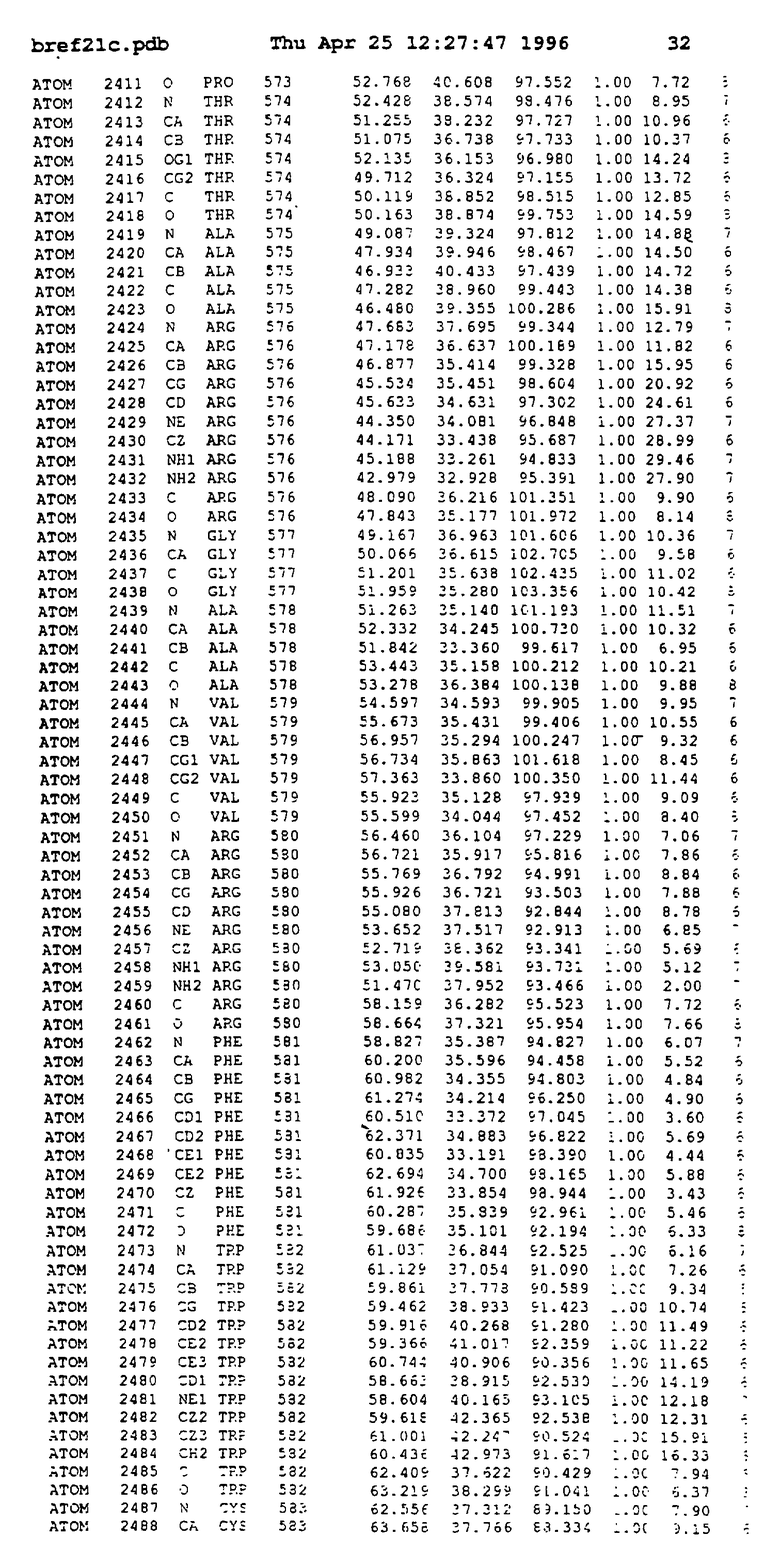
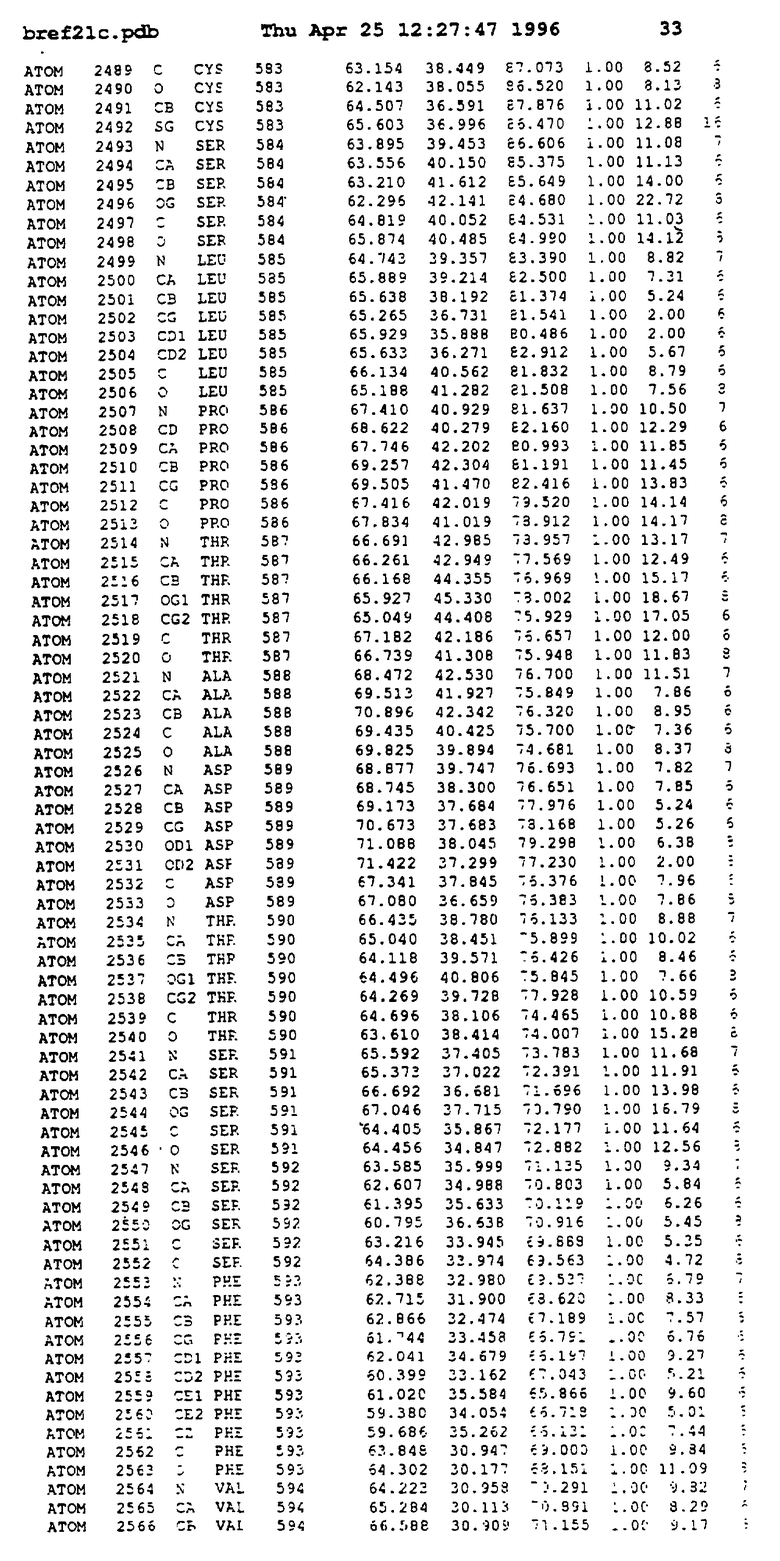
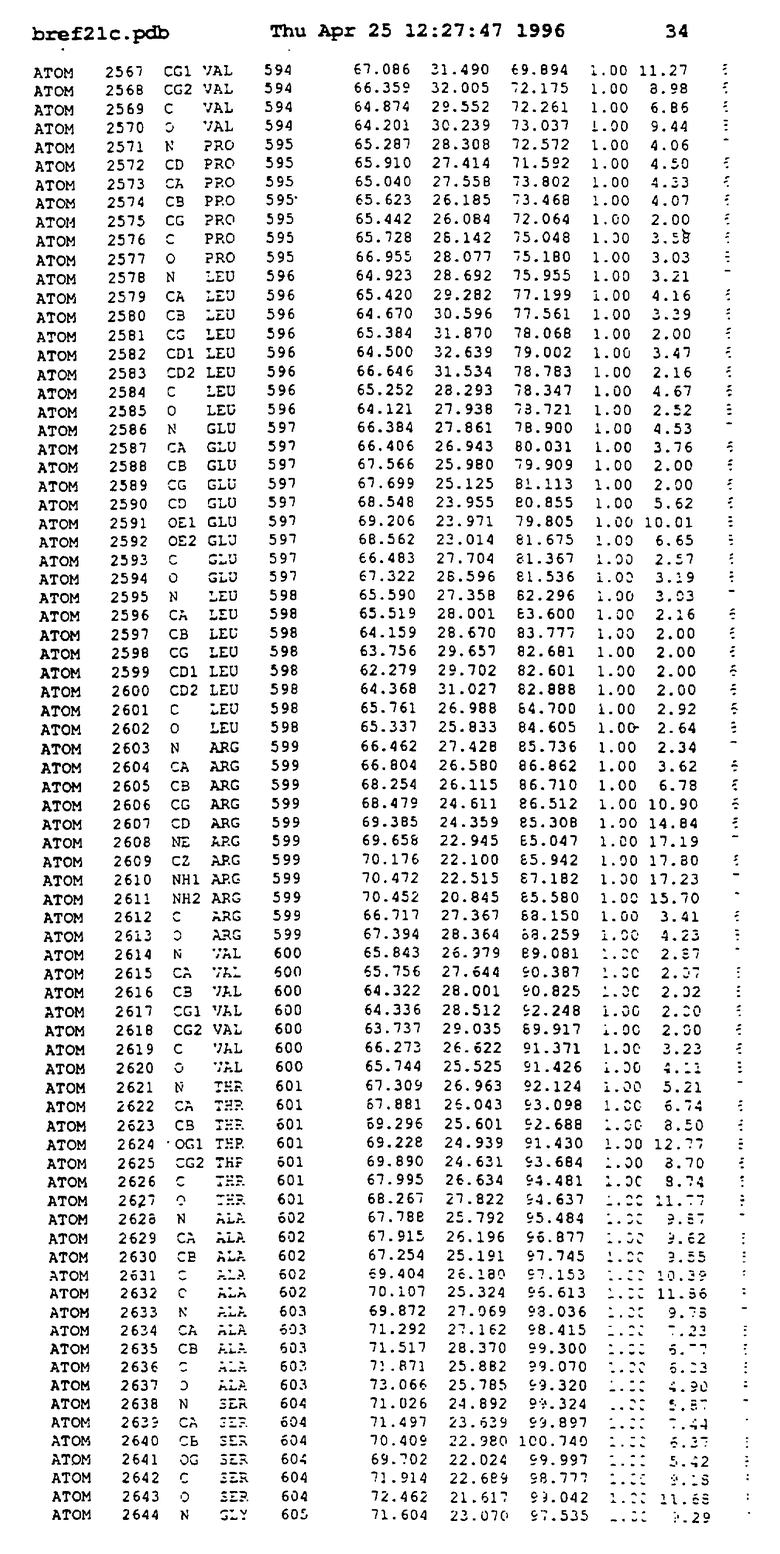
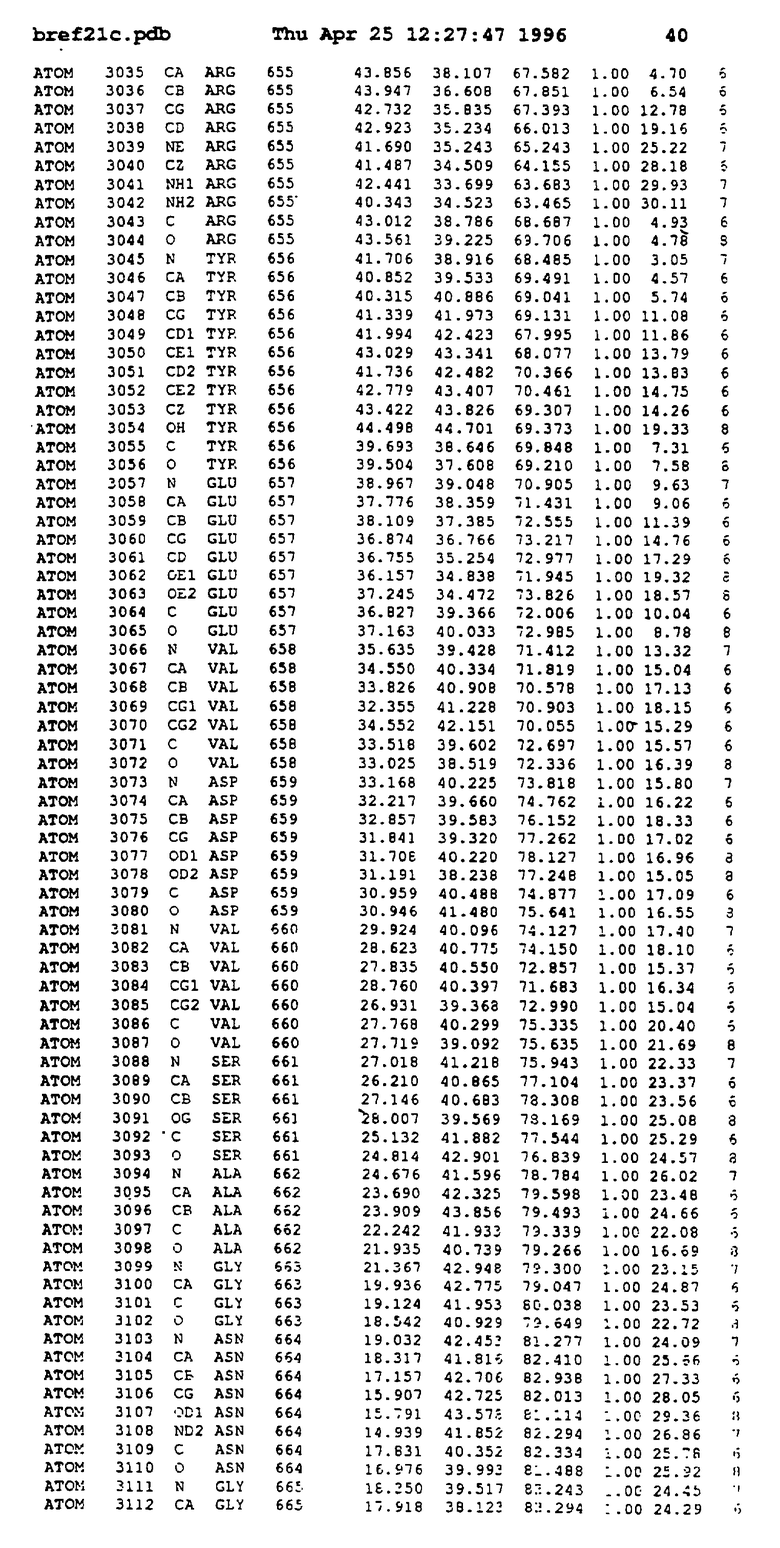
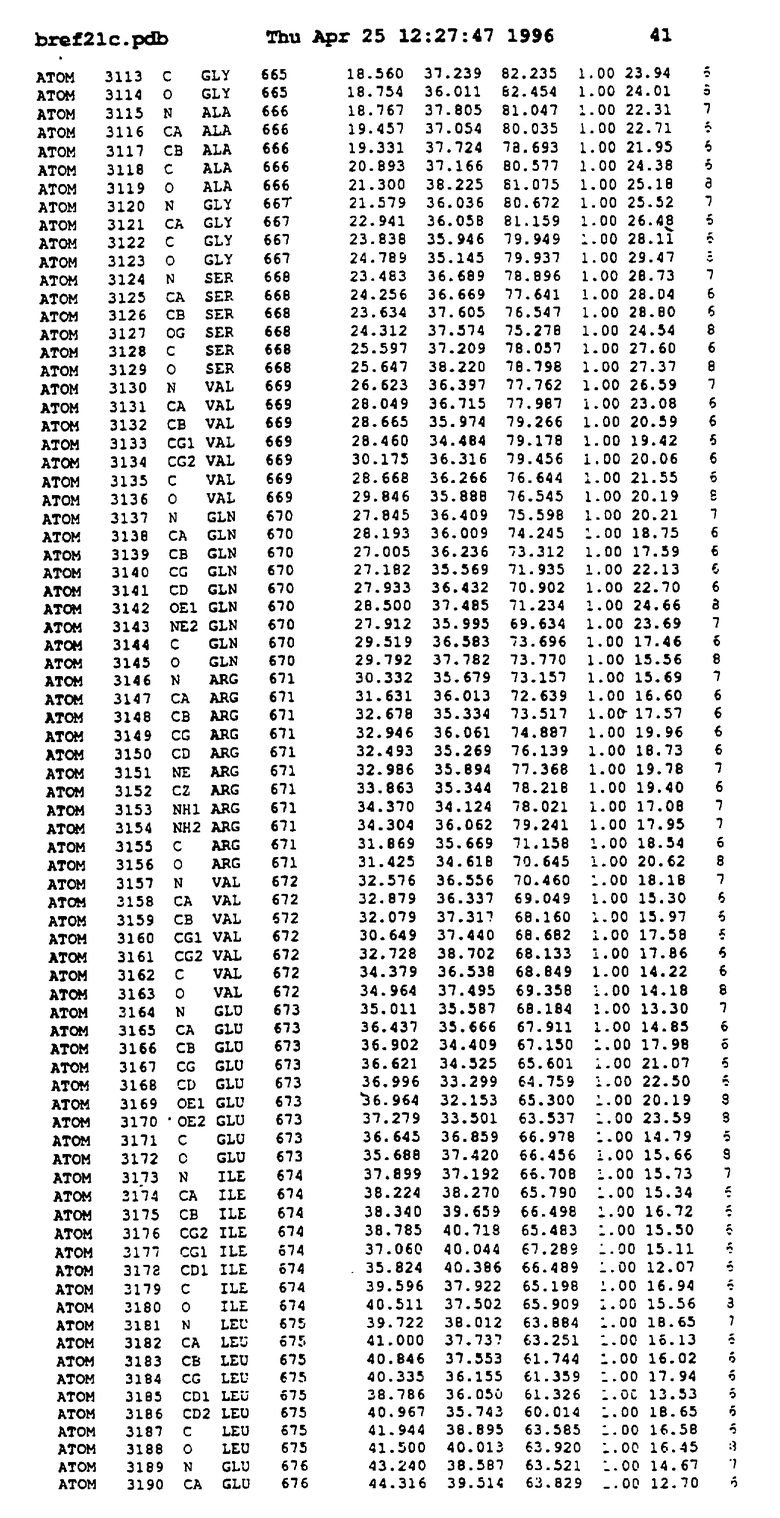
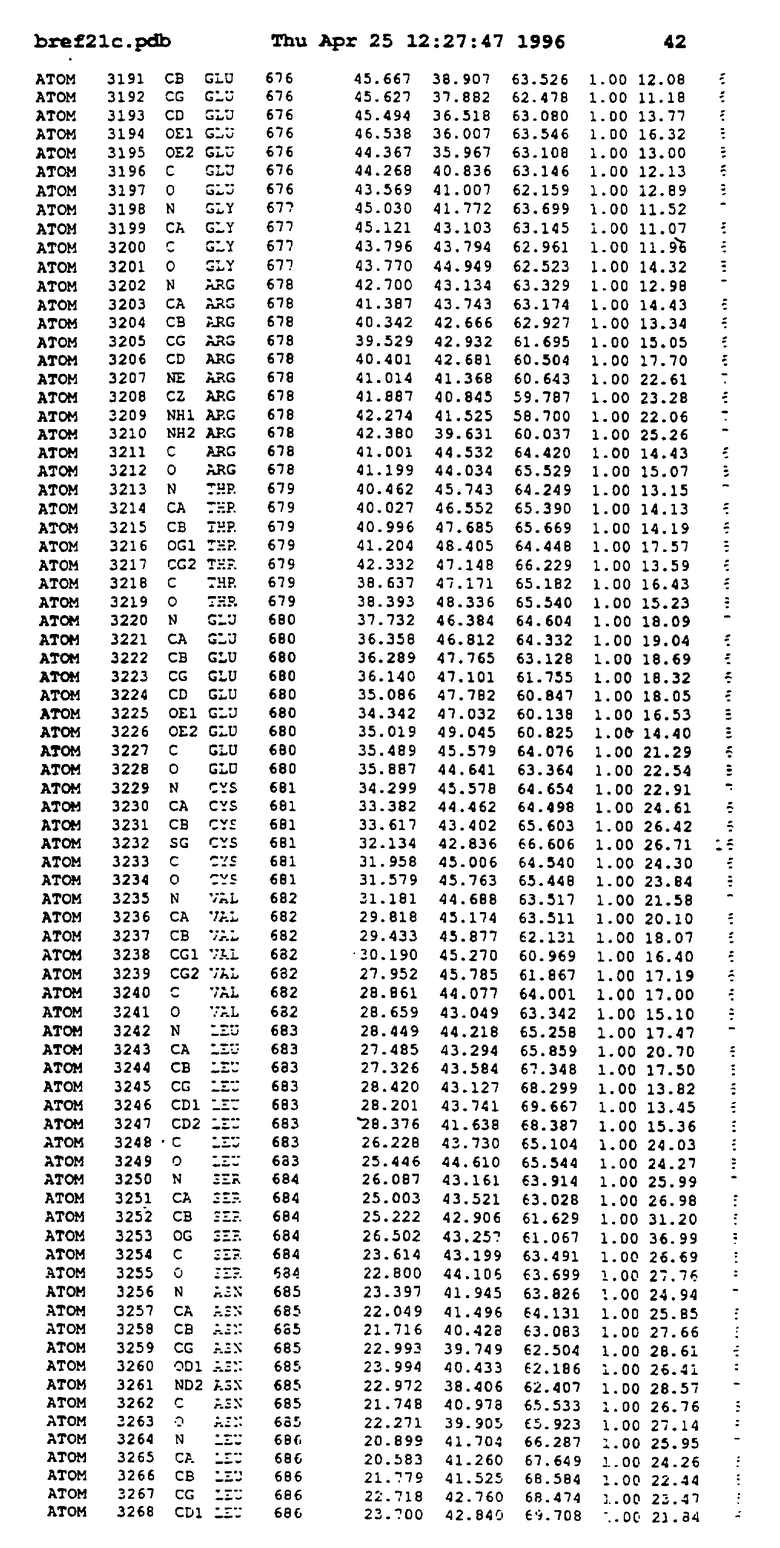
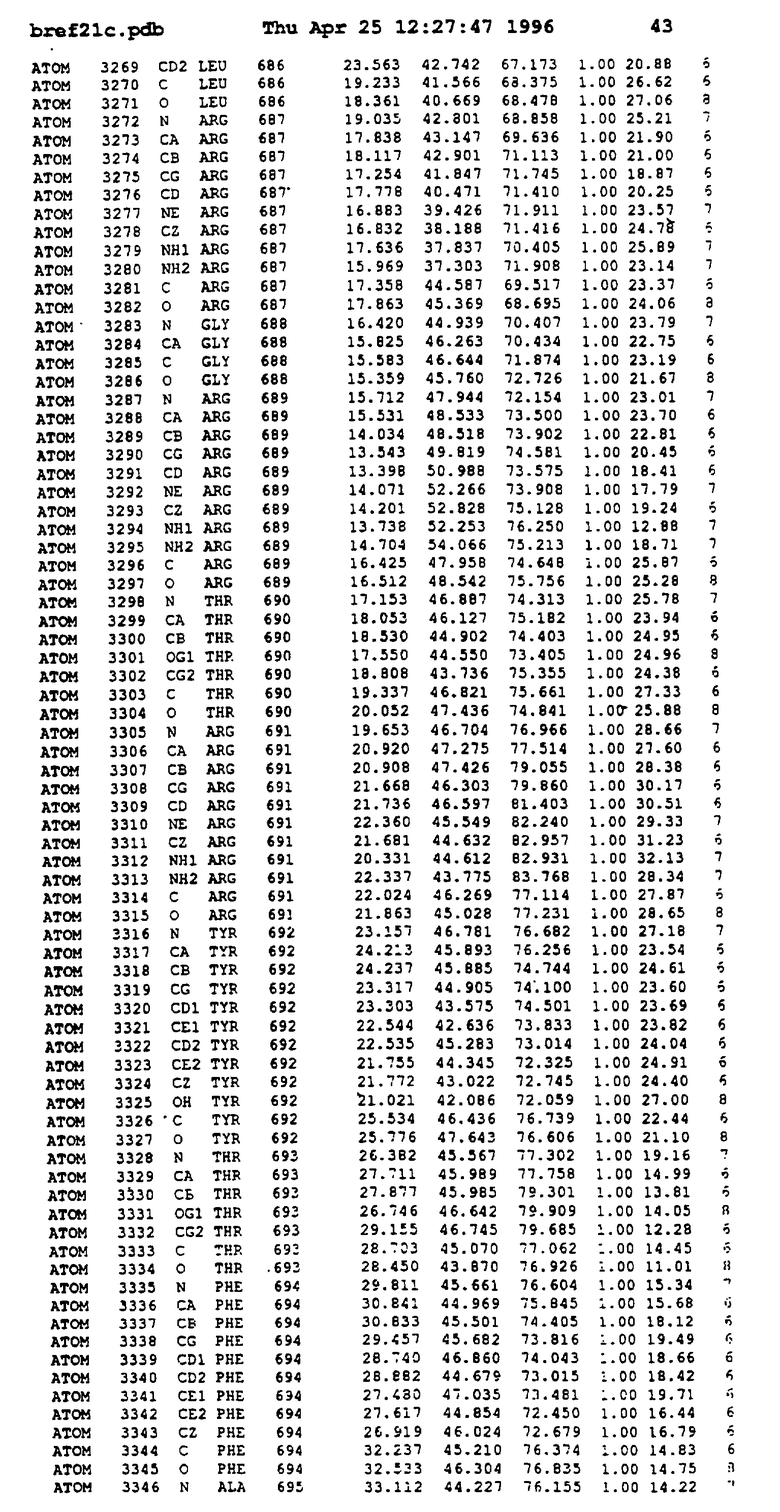
Координаты для аминокислот с 1 по 225 рецептора ЕРО человека, связанного с пептидом ЕРМ1, приведены в приложении в стандартном формате Brokhaven базы данных (standart Brookhaven database format). В это приложение включен также перечень Ван дер Ваальсовых взаимодействий. Эти координаты можно использовать в расчете и идентификации миметиков ЕРО согласно способам данного изобретения.
Структура комплекса ЕВР-ЕМР1
Обеспечивают экспрессию в Escherichia coli внеклеточного фрагмента рецептора ЕРО человека (ЕРО, связывающий протеин, ЕВР), состоящего из 1-225 остатков, и чистят, как описано (Johnson и др., Protein Express. Purif. 7: 104, 1996). Получают кристаллы комплекса ЕРО с ЕМР1 ромбоидной формы в пространственной группе P212121 с параметрами ячейки
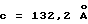 ,
,
с двумя ЕВР и двумя молекулами пептида в асимметрических единицах и 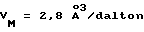 (Matthews, J. Mol. Biol. 33:491, 1968). Кристаллическую структуру определяют способом многократного изоморфного замещения (MIR), используя два производных с тяжелым атомом (таблица 1). Остатки 1-2 и 19-20 каждого пептида, также как остатки 1-9, 21-23, 164-166, 221-225 молекулы рецептора I и остатки 1-9, 21-23, 133-135, 221-225 молекулы рецептора II, имеют малую электронную плотность (или не имеют вовсе) и исключены из анализа структуры.
(Matthews, J. Mol. Biol. 33:491, 1968). Кристаллическую структуру определяют способом многократного изоморфного замещения (MIR), используя два производных с тяжелым атомом (таблица 1). Остатки 1-2 и 19-20 каждого пептида, также как остатки 1-9, 21-23, 164-166, 221-225 молекулы рецептора I и остатки 1-9, 21-23, 133-135, 221-225 молекулы рецептора II, имеют малую электронную плотность (или не имеют вовсе) и исключены из анализа структуры.
Важный разрыв в электронной плотности, который влияет на интерпретацию структуры, имеет место для трех остатков (Arg21-Gly22-Pro23), которые связывают амино-терминальную α-спираль с первой β-цепью в D1 обеих молекул рецептора. Диаграмма молекулярной упаковки показывает близость второго не определенного кристаллографически димера в кристалле, что дает две возможности присоединения этого линкера из трех остатков. Конкретный выбор присоединения линкера основан на структуре другого независимого комплекса ЕВР-пептид при более высоком разрешении 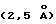 , который имеет аналогичную молекулярную упаковку, но для которого ясна электронная плотность для этих трех остатков. К настоящему времени отсутствуют экспериментальные данные, подтверждающие, существует ли эта N-терминальная α-спираль в растворе или является артефактом кристаллизационной упаковки. Примечательно, что эту спиральную область не наблюдают в опубликованных структурах hGHbp (начинается на остатке 32; de Vos и др. , Science 255:306, 1992), PRIL (начинается на остатке 2, без какой-либо вторичной структуры до первой β-цепи, остаток 6; Somers и др., Nature 372:478, 1994), INF-yRα (начинается на остатке 17; Walter и др., Nature 376: 230, 1995) или фактора ткани (начинается на остатке 3, без какой-либо определенной вторичной структуры до первой β-цепи, остаток 11; Muller и др., Nature 370:662, 1994).
, который имеет аналогичную молекулярную упаковку, но для которого ясна электронная плотность для этих трех остатков. К настоящему времени отсутствуют экспериментальные данные, подтверждающие, существует ли эта N-терминальная α-спираль в растворе или является артефактом кристаллизационной упаковки. Примечательно, что эту спиральную область не наблюдают в опубликованных структурах hGHbp (начинается на остатке 32; de Vos и др. , Science 255:306, 1992), PRIL (начинается на остатке 2, без какой-либо вторичной структуры до первой β-цепи, остаток 6; Somers и др., Nature 372:478, 1994), INF-yRα (начинается на остатке 17; Walter и др., Nature 376: 230, 1995) или фактора ткани (начинается на остатке 3, без какой-либо определенной вторичной структуры до первой β-цепи, остаток 11; Muller и др., Nature 370:662, 1994).
Мономер ЕВР загибается в два домена, D1 и D2, которые образуют L-форму с длинными осями доменов, расположенными примерно под углом 90o друг к другу; общие молекулярные размеры составляют 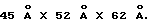 N-терминальный домен (D1, остатки 10-114) и С-терминальный домен (D2, остатки 119-220) соединены коротким (на четыре остатка) α-спиральным линкером. Оба домена по общей топологии ближе к доменам типа Fibronectin type-III (FBN-III), чем к Ig доменам (Bork и др., J. Mol. Biol. 242:309, 1994). Складка FBN-III составлена из двух антипараллельных β-сложенных листов, состоящих из цепей А, В, Е и цепей G, F, С и С', и обнаружена в двух доменах рецепторов человеческого гормона роста (de Vos и др., Science 255:306, 1992) и пролактина (Somers и др., Nature 372:478), D1 и D2 домены α-цепи рецептора γ-интерферона (IFN-γRα) (Walter и др., Nature 376:230, 1995), D2 домен CD4 (Wang и др. , Nature 348:411, 1990; Ryu и др., Nature 348:419, 1990), двух доменах фактора ткани (Muller и др., Biochemistry 33:10864, 1994; Harols и др., Nature 370:662, 1994), в третьем повторе тенацина фибронектинового типа (Leagy и др., Science 258:987, 1992) и D2 домене протеина chaperon protein PapD (Holmgren и др., Nature 342:248, 1989). FBN-III топология отличается от Ig константного домена сдвигом цепи D от одного β-листа (цепи А, В, Е и D) к другому (цепи G, F, С, С'), где его определяют как С' цепь. Суперпозиция эквивалентных коровых (сердцевинных) остатков β-листа (β-sheet core residues) доменов D1 и D2 в ЕВР дает среднеквадратичное отклонение (RMS)
N-терминальный домен (D1, остатки 10-114) и С-терминальный домен (D2, остатки 119-220) соединены коротким (на четыре остатка) α-спиральным линкером. Оба домена по общей топологии ближе к доменам типа Fibronectin type-III (FBN-III), чем к Ig доменам (Bork и др., J. Mol. Biol. 242:309, 1994). Складка FBN-III составлена из двух антипараллельных β-сложенных листов, состоящих из цепей А, В, Е и цепей G, F, С и С', и обнаружена в двух доменах рецепторов человеческого гормона роста (de Vos и др., Science 255:306, 1992) и пролактина (Somers и др., Nature 372:478), D1 и D2 домены α-цепи рецептора γ-интерферона (IFN-γRα) (Walter и др., Nature 376:230, 1995), D2 домен CD4 (Wang и др. , Nature 348:411, 1990; Ryu и др., Nature 348:419, 1990), двух доменах фактора ткани (Muller и др., Biochemistry 33:10864, 1994; Harols и др., Nature 370:662, 1994), в третьем повторе тенацина фибронектинового типа (Leagy и др., Science 258:987, 1992) и D2 домене протеина chaperon protein PapD (Holmgren и др., Nature 342:248, 1989). FBN-III топология отличается от Ig константного домена сдвигом цепи D от одного β-листа (цепи А, В, Е и D) к другому (цепи G, F, С, С'), где его определяют как С' цепь. Суперпозиция эквивалентных коровых (сердцевинных) остатков β-листа (β-sheet core residues) доменов D1 и D2 в ЕВР дает среднеквадратичное отклонение (RMS) 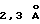 для 77 Сα пар, что существенно больше, чем соответствующие перекрывания для hGHbp
для 77 Сα пар, что существенно больше, чем соответствующие перекрывания для hGHbp 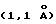 и PRLR
и PRLR  , и отражает различие в подклассе складки между двумя доменами ЕВР.
, и отражает различие в подклассе складки между двумя доменами ЕВР.
В D1 короткая α-спираль (остатки 10-20) предшествует первому β-сандвичу, что лучше описывается как гибрид FBN-III складки и Ig складки (остатки 24-114), чем строгой FBN-III топологией. В этой складке h-типа (Wang и др., Nature 348: 411, 1990; Ryu и др., Nature 348:419, 1990) С' цепь длинная и взаимодействует сначала с цепью С, а затем изменяет направление, взаимодействуя с цепью Е (где С' изменяет свое обозначение на цепь D), образуя β-сандвич четыре-на-четыре цепи. D1 содержит два сохраненных дисульфидных мостика, связывающих Суs28 (βА) с Суs38 (βВ) и Суs67 (βC') с Суs83 (βЕ). Количество остатков между цистеиновыми парами, которые образуют два дисульфидных мостика, равно 9 и 15 для ЕВР, по сравнению с 9 и 10 в GHR и PRLR. Более длинная связь между цепями C' и Е дает возможность второй половине цепи С' стать цепью D. Эта топология h-типа обнаружена в одной из двух доменов GHR s-типа. На остатке Аsn52, который расположен по направлению к концу петлеобразной области, связывающей цепи βВ и βC, присутствует потенциальный сайт гликозилирования. Хотя Аsn52 не гликозилирован в этом бактериально экспрессированном протеине, внешняя полость вокруг боковой цепи Аsn52 легко приспосабливает карбогидратную часть.
Спиральный линкер (остатки 115-118) связывает D1 и D2 (торсионные углы φ, ψ для междоменного спирального линкера для Ile115, Asn116, Glu117 и Val118 составляют -50o, -27o, -76o, -21o, -99o, 26o и -151o, 38o, соответственно) и наблюдается в других членах этого семейства рецепторов, hGHbp, PRLR, IFN-γRα и фактора ткани. В ЕВР ассоциация доменов ограничена смешанным ассортиментом водородных связей, гидрофобных взаимодействий и одним соляным мостиком (между Аrg32 и Asp122) из 11 остатков D1 и 12 остатков D2 с полностью потерянной поверхностью [Площадь молекулярной поверхности, потерянной вследствие взаимодействия, рассчитывают, применяя программу MS (Connolly, J. Appl. Crystallog., 16:439, 1983), используя 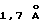 сферический образец и стандартные атомные радиусы (как описано в работе Davies и др., Ann. Rev. Biochem. 59:439, 1990). Между приведенными и другими опубликованными величинами (de Vos и др., Science 255:306, 1992) могут иметься некоторые разногласия вследствие использования разных алгоритмов (Connolly) по сравнению с Lee и др. , J. Mol. Biol., 55:379, 1971) и радиусов образцов. Для ясности все сообщенные здесь величины рассчитаны одинаковым способом для лучшего сравнения рецепторов], равной
сферический образец и стандартные атомные радиусы (как описано в работе Davies и др., Ann. Rev. Biochem. 59:439, 1990). Между приведенными и другими опубликованными величинами (de Vos и др., Science 255:306, 1992) могут иметься некоторые разногласия вследствие использования разных алгоритмов (Connolly) по сравнению с Lee и др. , J. Mol. Biol., 55:379, 1971) и радиусов образцов. Для ясности все сообщенные здесь величины рассчитаны одинаковым способом для лучшего сравнения рецепторов], равной  для двух доменов.
для двух доменов.
D2 (остатки 119-220) складывается в стандартную FBN-III (s-тип) топологию с одним свободным цистеином и без дисульфидных мостиков, согласующуюся с GHR и PRLR, которые имеют три и два дисульфидных мостика, соответственно, в D1 и ни одного в D2. После α-спирального линкера D2 начинается с нерегулярного витка (остатки 118-126), который содержит Pro124, сохраняющиеся в структурах hGHbp, PRLR, фактора ткани и IFN-γRα, и основан на сравнительном анализе последовательностей в большинстве рецепторов цитокина класса-I и класса-II (Bazan, Proc. Natl. Acad. Sci. USA 87:6934, 1990). Этот короткий виток заканчивается Gly124, который имеет положительный φ (φ, ψ=52o, 40o), согласующийся с эквивалентными торсионными углами А1а136 и А1а101 в hGHbp (φ, ψ= 63o, 68o) и PRLR (φ, ψ=58o, 38o). Область Pro124 образует аналогично расположенную выпуклую конформацию, примыкающую и параллельную соответствующей выпуклости, содержащей мотив WSXWS. Последовательность WSXWS образует большую модифицированную β-выпуклость (Richardson, Adv. Prot. Chem. 34:167, 1981) и расположена в обширной области цепи, непосредственно предшествующей βG-цепи, которая, по всей вероятности, обычно связана с областью присоединения мембраны (membrane spanning region) EPOR.
Четвертичная структура комплекса составлена из двух пептидов и двух рецепторов, которые образуют Т-образное соединение. Нековалентный пептидный димер взаимодействует с двумя молекулами рецептора, образуя почти совершенную симметричную структуру с 2 складками. После суперпозиции D2 от двух молекул ЕВР в димере центры масс двух доменов D1 отстоят только на 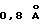 , что достаточно для нарушения совершенной двухскладчатой симметрии. Отдельная суперпозиция соответствующих D1 и D2 каждого рецептора в димере дает в результате среднеквадратичные отклонения (RMS)
, что достаточно для нарушения совершенной двухскладчатой симметрии. Отдельная суперпозиция соответствующих D1 и D2 каждого рецептора в димере дает в результате среднеквадратичные отклонения (RMS)  (105 D1 Сα пар) и
(105 D1 Сα пар) и 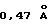 (105 D1 Сα пар).
(105 D1 Сα пар).
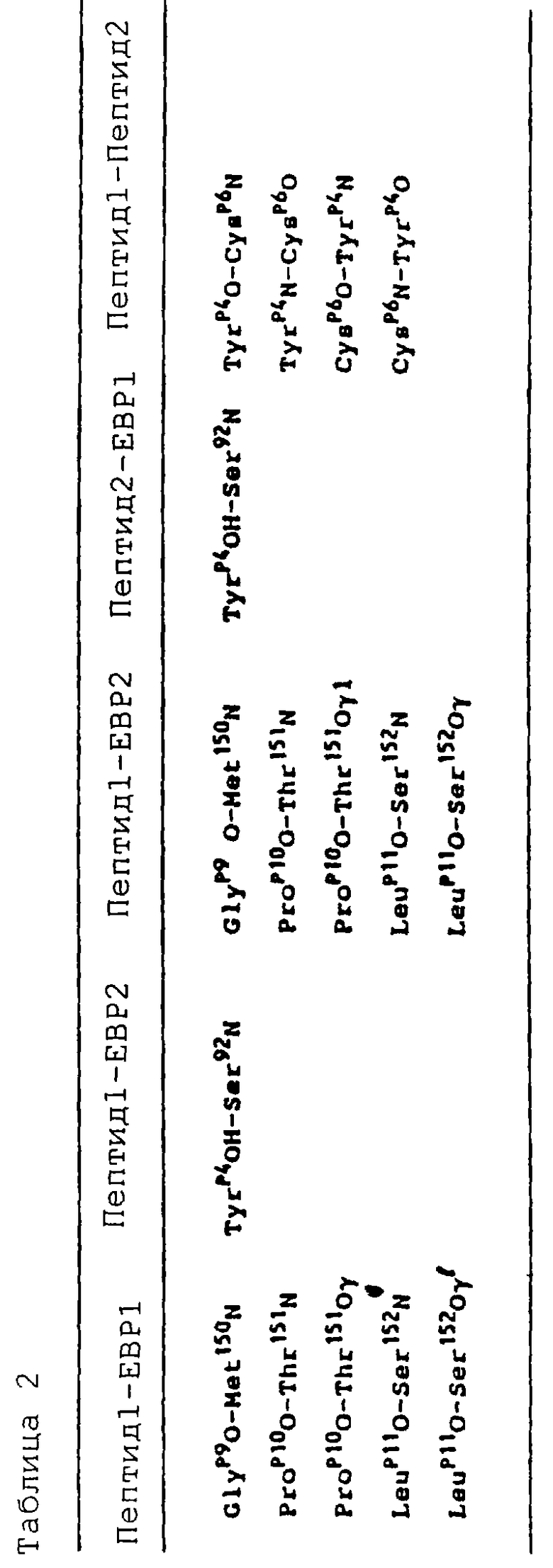
Циклический ЕМР1 содержит один дисульфидный мостик между CysP6 и CysP15, который связывает короткие β-цепи (остатки 4-7 и 13-16), которые соединены слегка искаженным β-изгибом типа 1 [ProP10 (i+1) и LeuP11 (i+2) β-изгиба имеет φ, ψ= -62, -38o и -99o, -60o, соответственно. Карбонильный кислород LeuP11 имеет водородную связь с ЕВР, которая дает отклонение величины ψ от нормы 0±30o (i+2) в стандартном β-изгибе типа 1], состоящем из остатков GlyP9-ProP10-LeuP11-ThrP11. Каждый пептид очень близко ассоциирован с другими своим пептидным партнером и в этом взаимодействии теряет 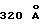 своей молекулярной поверхности
своей молекулярной поверхности 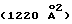 (Connelly, J. Appl. Crystallogr. 16:439, 1983; Davies и др., Ann. Rev. Biochem. 59:439, 1990; Richards, J. Mol. Biol. 55: 379, 1971). Четыре водородные связи между основными цепями двух пептидов дают образование четырехнитевого антипараллельного β-сложенного листа (таблица 2). Две симметричные гидрофобные сердцевины (cores) объединены димеризацией пептидов и охвачены дисульфидными мостиками и боковыми цепями ТуrP4, PheP8 и ТrрP13. Конструкция каждой гидрофобной сердцевины походит на ящик (блок) (box), который располагает по углам ароматические кольца PheP8, TrpP13 и ТуrP4 (от другого пептида) и дисульфидный мостик (CysP6-CysP15). Два глициновых остатка на каждом конце пептида не структурированы.
(Connelly, J. Appl. Crystallogr. 16:439, 1983; Davies и др., Ann. Rev. Biochem. 59:439, 1990; Richards, J. Mol. Biol. 55: 379, 1971). Четыре водородные связи между основными цепями двух пептидов дают образование четырехнитевого антипараллельного β-сложенного листа (таблица 2). Две симметричные гидрофобные сердцевины (cores) объединены димеризацией пептидов и охвачены дисульфидными мостиками и боковыми цепями ТуrP4, PheP8 и ТrрP13. Конструкция каждой гидрофобной сердцевины походит на ящик (блок) (box), который располагает по углам ароматические кольца PheP8, TrpP13 и ТуrP4 (от другого пептида) и дисульфидный мостик (CysP6-CysP15). Два глициновых остатка на каждом конце пептида не структурированы.
Пептидный димер вставлен в глубокую щель между молекулами рецептора ЕВР. Часть каждого пептидного мономера взаимодействует с обеими молекулами рецептора. Сайты связывания каждого ЕВР практически идентичны вследствие действующих на связь пептидного димера 2-кратных симметричных взаимодействий. Четыре основных контактных области на ЕВР поступают от сегментов на четырех петлевых участках (L1, L3, L5, L6), которые соединяют цепи А и В (33-34 остатки L1) и F и G (90-94 остатки L3) в D1 и цепи В и С (148-153 остатки L5) и F и G (203-205 остатки L6) в D2. Общая закрытая молекулярная поверхность в структуре пептид-ЕВР составляет  для двух пептидов и двух ЕВР, соответственно. Взаимодействие пептид-ЕВР можно разделить на отдельные гидрофобные (67%) и полярные (33%) области. Гидрофобная сердцевина образована между пептидом и рецептором и включает Phe93, Met150 и Phe205 от молекулы ЕВР и пептидный гидрофобный блок (box), состоящий из PheP8 и ThrP13 от одного пептида и TyrP4 и СysP15 от другого пептида. Полярные взаимодействия локализованы в основном по дну связывающей щели и осуществляются, главным образом, с L5 в D2. Пять из шести водородных связей происходят между основной цепью остатков β-изгиба GlyP9, ProP10 и LeuP11 от одного пептида с гидроксилом основной и боковой цепи сохраненного ТyrP4, который пересекает другой свой пептидный партнер, взаимодействуя с петлей L3 (таблица 2). Взаимодействие ЕВР-ЕВР делает на удивление малый вклад в общую стабильность комплекса, где межрецепторная закрытая молекулярная поверхность составляет только
для двух пептидов и двух ЕВР, соответственно. Взаимодействие пептид-ЕВР можно разделить на отдельные гидрофобные (67%) и полярные (33%) области. Гидрофобная сердцевина образована между пептидом и рецептором и включает Phe93, Met150 и Phe205 от молекулы ЕВР и пептидный гидрофобный блок (box), состоящий из PheP8 и ThrP13 от одного пептида и TyrP4 и СysP15 от другого пептида. Полярные взаимодействия локализованы в основном по дну связывающей щели и осуществляются, главным образом, с L5 в D2. Пять из шести водородных связей происходят между основной цепью остатков β-изгиба GlyP9, ProP10 и LeuP11 от одного пептида с гидроксилом основной и боковой цепи сохраненного ТyrP4, который пересекает другой свой пептидный партнер, взаимодействуя с петлей L3 (таблица 2). Взаимодействие ЕВР-ЕВР делает на удивление малый вклад в общую стабильность комплекса, где межрецепторная закрытая молекулярная поверхность составляет только 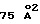 , вносимых Leu175 и Аrg178 от каждой молекулы рецептора.
, вносимых Leu175 и Аrg178 от каждой молекулы рецептора.
ЕМР1 является одной из семейства последовательностей, которые содержат несколько сохраненных остатков кроме цистеина. Наиболее структурно значимыми из этих остатков для согласования, по-видимому, являются ТurP4 и TrpP13, которые наряду с дисульфидным мостиком вносят основной вклад в гидрофобную сердцевину пептид-пептидного взаимодействия. Кроме того, эти два ароматических остатка играют центральную роль во взаимодействии пептид-рецептор и в димеризации рецептора.
Димеризация ЕВР в растворе
Для исследования взаимодействия ЕМР1 с ЕВР в растворе авторы применяли бифункциональный сульфидрильный реактивный кросслинкер DPDPB [1,4-ди-(2'-пиридилдитиопропионамидо) бутан] в попытке стабилизировать пептид-зависимую димерную структуру. Выбор кросслинкера основан на предварительных экспериментах с амин-реактивными кросслинкерами, которые, как было обнаружено, инактивируют ЕВР. ЕВР содержит один свободный сульфидрил (Сys181) в D2, который является потенциально активным в связывании реагентов (сам кросслинкер DPDPB не инактивирует ни способность ЕВР связывать ЕРО, ни пролиферационные свойства ЕМР1). Димерный ЕВР продукт образуется посредством совместной инкубации ЕМР1, DPDPB и ЕВР. Количество димерного продукта увеличивается с концентрацией пептида, а в отсутствие пептида существенного количества димерного продукта не наблюдают. DPDPB-кросссвязанные продукты, образованные по дисульфидобменной реакции, должны быть легко обратимыми посредством восстановления, как видно для ковалентно связанного EMPl-опосредованного димера. Кроме того, авторы разработали ковалентно связанную димерную форму ЕМР1, которая демонстрирует повышенную биологическую способность (Johnson и др., готовится к публикации). Остатки Cys181 в D2 димера ЕВР отстоят друг от друга на 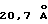 (расстояние Sγ-Sγ), что приблизительно соответствует длине кросслинкера DPDPB
(расстояние Sγ-Sγ), что приблизительно соответствует длине кросслинкера DPDPB 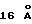 (и приблизительно
(и приблизительно 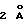 на длину связи с каждого конца). Таким образом, ЕМР1 опосредует образование растворимого комплекса димера ЕВР в растворе, совместимого с кристаллической структурой.
на длину связи с каждого конца). Таким образом, ЕМР1 опосредует образование растворимого комплекса димера ЕВР в растворе, совместимого с кристаллической структурой.
Мотив WSXWS
Последовательность WSAWS (остатки 209-213), соответствующая блоку WSXWS, находится в β-выпуклости (Richardson, Adv. Prot. Chem. 34:167, 1981; Chan и др., Protein Science, 2:1574, 1993), непосредственно предшествуя
β-цепи G в D2. Остатки в этом мотиве не взаимодействуют с лигандом, не играют роли во взаимодействиях рецептор-рецептор и расположены на противоположной стороне поверхности раздела рецептор-рецептор и рецептор-лиганд. Блок WSXWS представляет только сегмент организации комплексных взаимодействий, который включает несколько других сохранившихся боковых цепей от четырехнитевого β-листа в D2. Системы с индольным кольцом Тrр209 и Тrр212 направлены в внешней поверхности β-листа и только частично подвержены действию растворителя, в то время как боковая цепь А1а202 направлена прямо в раствор. Амиды и гидроксилы обоих Ser213 и Ser213 образуют водородные связи с основной цепью остатков 198 и 196 соседней цепи F во взаимодействии типа псевдо β-листа, которое походит на модифицированную большую β-выпуклость (Richardson, Adv. Prot. Chem. 34:167, 1981; Chan и др.. Protein Science, 2: 1574, 1993), где взаимодействие типа β-листа осуществляет скорее гидроксил боковой цепи, чем кислород карбонила. Структура β-выпуклости определяет положение двух остатков Тrр, которые разворачиваются в стороне от четырех остатков с той же стороны β-листа, а не с противоположных сторон, как в нормальном β-листе или вытянутых цепных структурах. Гуанидиниум-группа (guanidinium group) Arg197 из цепи F, центральный остаток (Richardson, Adv. Prot. Chem. 34:167, 1981; Chan и др., Protein Science, 2:1574, 1993) в выпуклости, расположена точно между двумя индольными кольцами Тrр, образуя вытянутую π-катионную систему (Kumpf и др., Science 261:1708, 1993). Центр пиррольного кольца Тrp209, Ns Аrg197 и центр бензольного кольца Тrр212 расположены на прямой линии с тремя плоскостями сопряженных систем, упакованных параллельно друг другу на расстоянии приблизительно 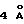 . Кроме того, алифатическая часть боковой цепи Аrg199 имеет гидрофобные взаимодействия с индольным кольцом Трr209, завершая чередующуюся упаковку двух ароматических и двух положительно заряженных аминокислотных остатков. Боковая цепь Glu157 образует водородную связь с Аrg197, предположительно помогая ориентировать гуанидиниум-группу, и добавляет системе некоторую специфичность и стабилизацию.
. Кроме того, алифатическая часть боковой цепи Аrg199 имеет гидрофобные взаимодействия с индольным кольцом Трr209, завершая чередующуюся упаковку двух ароматических и двух положительно заряженных аминокислотных остатков. Боковая цепь Glu157 образует водородную связь с Аrg197, предположительно помогая ориентировать гуанидиниум-группу, и добавляет системе некоторую специфичность и стабилизацию.
По-видимому, когда этот линейный мотив WSXWS, идентифицированный из сравнительного анализа последовательностей рецепторов цитокина, представляет только компонент более сложной конформационной единицы, он вносит существенную структурную особенность в D2. Как предположено ранее, ароматические остатки обладают стабилизирующим действием и играют роль складных ядер в структурах антипараллельных β-сандвичей (Finkelstein и др., Protein Eng. 6: 367, 1993). Амино-ароматическая параллельная упаковка между гуанидиниум-группой (guanidinium group) аргинина и ароматическими кольцами является особенностью в протеиновых структурах (Burley и др., Adv. Prot. Chem., 39:125, 1988; Flocco и др., J. Mol. Biol., 235:709, 1994), а параллельная тройная упаковка π-катионных систем встречается редко (Kim и др., Biochemistry, 32: 8465, 1993), хотя наблюдается в других рецепторах цитокина класса-1, hGHbp и PRLR.
Структурные эквиваленты мотива WSXWS в hGHbp (YGEFS) и PRLR (WSAWS) включены в множество π-катионных взаимодействий, даже более запутанных и сложных. π-Катионная система распространяется в hGHbp и PRLR, включая дополнительный ароматический остаток (Тrр186 для hGHbp и Тrр156 для PRLR) из петлевой области, который связывает βC и βС' в D2, и положительно заряженный остаток (Аrg211 для hGHbp и Аrg147 для PRLR), который упаковывается между Тrр и вторым ароматическим остатком. Дополнительный остаток Аrg берется либо из цепи βF, как в hGHbp (Аrg211) или из βС, как в PRLR (Аrg147); глутаминовый остаток, который связан водородными связями и ориентирует аргинин, также переключает (switches) цепи. Сравнительные анализы последовательностей предполагают, что этот переключатель Arg-Gln может быть общим для других членов семейства рецепторов цитокина класса-1. Протяженная π-катионная система в hGHbp и PRLR состоит из пяти положительно заряженных и трех ароматических остатков, упакованных стопкой в порядке чередования, который включает Lys215, Tyr222, Arg213, Phe225, Arg211, Trp186, Lys179 для hGHbp и Lys185, Trp191, Arg183, Trp194, Arg147, Trp156, Lys149 для PRLR. Первое трио ароматика-Аrg-ароматика отстоит примерно на 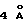 , как в ЕВР, а вторая система упакована ближе с расстояниями примерно
, как в ЕВР, а вторая система упакована ближе с расстояниями примерно 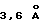 , что согласуется с π-π-взаимодействием (Burley и др., Adv. Prot. Chem., 39:125, 1988; Flocco и др. , J. Mol. Biol., 235:709, 1994). Внешние лизины также используют алифатические части своих боковых цепей для образования гидрофобных взаимодействий с ароматическими кольцами. По данным сравнительного анализа последовательностей с другими членами подсемейства рецепторов цитокина класса-1, такие структурно-протяженные π-катионные системы могут существовать в рецепторах человеческого тромбопиетина, IL-6 и цилиарного нейротрофического фактора (ciliary neurotrophic factor) и в рецепторе человеческого IL-4, что основано на структурном моделировании (Gustchina и др. Proteins 21:140, 1995). Хотя IFN-γRα и фактор ткани не имеют мотива WSXWS, соответствующие последовательности TTEKS (остатки 213-217) для IFN-γRα (Walter и др., Nature 376: 230, 1995) и KSTDS (остатки 201-205) для фактора ткани (Muller и др., Biochemistry 33: 10864, 1994; Harlos и др., Nature 370:662, 1994) сохраняют очень похожую β-выпуклость. Вероятная последовательность среди этих пяти рентгеновских структур указывает, что серин или треонин в положениях 2 и 5 сохраняют обычный набор водородных связей между гидроксилами своих боковых цепей и основной цепью соседней цепи. Только hGHbp является не содержащим гидроксил остатком в положении 2, a Ser226 еще сохраняет эквивалентное взаимодействие. Мутация Ser226 в А1а аннулирует связывание hGHbp с hGH, и его экспрессия на клеточной поверхности резко снижается (Baumgartner и др., J. Biol. Chem. , 269:29094, 1994). В GMCSFRα и IL-2Rβ точковые мутации остатков серина вызывают существенное снижение экспрессии на клеточной поверхности, но слабо действуют (или не действуют вовсе) на связывание лигандов (Ronco и др. , J. Biol. Chem. 269:277, 1994; Miyazaki и др., EMBO Journal 10:3191, 1991).
, что согласуется с π-π-взаимодействием (Burley и др., Adv. Prot. Chem., 39:125, 1988; Flocco и др. , J. Mol. Biol., 235:709, 1994). Внешние лизины также используют алифатические части своих боковых цепей для образования гидрофобных взаимодействий с ароматическими кольцами. По данным сравнительного анализа последовательностей с другими членами подсемейства рецепторов цитокина класса-1, такие структурно-протяженные π-катионные системы могут существовать в рецепторах человеческого тромбопиетина, IL-6 и цилиарного нейротрофического фактора (ciliary neurotrophic factor) и в рецепторе человеческого IL-4, что основано на структурном моделировании (Gustchina и др. Proteins 21:140, 1995). Хотя IFN-γRα и фактор ткани не имеют мотива WSXWS, соответствующие последовательности TTEKS (остатки 213-217) для IFN-γRα (Walter и др., Nature 376: 230, 1995) и KSTDS (остатки 201-205) для фактора ткани (Muller и др., Biochemistry 33: 10864, 1994; Harlos и др., Nature 370:662, 1994) сохраняют очень похожую β-выпуклость. Вероятная последовательность среди этих пяти рентгеновских структур указывает, что серин или треонин в положениях 2 и 5 сохраняют обычный набор водородных связей между гидроксилами своих боковых цепей и основной цепью соседней цепи. Только hGHbp является не содержащим гидроксил остатком в положении 2, a Ser226 еще сохраняет эквивалентное взаимодействие. Мутация Ser226 в А1а аннулирует связывание hGHbp с hGH, и его экспрессия на клеточной поверхности резко снижается (Baumgartner и др., J. Biol. Chem. , 269:29094, 1994). В GMCSFRα и IL-2Rβ точковые мутации остатков серина вызывают существенное снижение экспрессии на клеточной поверхности, но слабо действуют (или не действуют вовсе) на связывание лигандов (Ronco и др. , J. Biol. Chem. 269:277, 1994; Miyazaki и др., EMBO Journal 10:3191, 1991).
Полагают, что сохранение мотива WSXWS в EPOR или его эквивалента в других рецепторах цитокина класса-1 является существенным для биологической активности, и, следовательно, считается частью сайта связывания рецептора (Yoshimura и др., J. Biol. Chem. 267:11619, 1992; Quelle, Mol. Cell. Biol. 12: 4553, 1992). Для EPOR систематическое исследование 100 мутаций последовательности WSAWS демонстрирует, что большинство мутаций двух триптофанов и серина дают в результате молекулы, которые не достигают клеточной поверхности, но сохраняются в эндоплазматическом ретикулуме (Hilton и др., Proc. Natl. Acad. Sci. USA 92:190, 1995; Hilton и др., J. Biol. Chem., 271:4699, 1996). Кроме того, мутация А1а211 в Glu в последовательности WSAWS дает лучший перенос от ER к Golgi и 3-5-кратное увеличение количества молекул EPOR на клеточной поверхности по сравнению с природным типом (Hilton и др., Proc. Natl. Acad. Sci. USA 92:190, 1995; Hilton и др., J. Biol. Chem., 271:4699, 1996). Эти результаты подтверждают вывод авторов, что последовательность WSXWS играет важную роль в структуре и складчатости D2 в EPOR и других родственных рецепторах.
Сравнение со структурами других комплексов рецепторов цитокина
Общая четвертичная структура комплекса пептид-ЕВР существенно отличается от эквивалентного окружения в комплексе hGH-hGHbp. Несимметричная природа структуры отдельного четырехцепного пучка лиганда гормона роста дает асимметричную гомодимеризацию рецептора, которая соответствует 159o развороту между рецепторами по сравнению с почти совершенным 2-складчатым разворотом (180o) для комплекса ЕВР-пептид. Третичная структура доменов в ЕВР и hGHbp тоже в некоторой степени различна. Если эквивалентные D2 домены ЕВР и hGHbp наложить друг на друга, их соответствующие D1 домены отличаются 12o поворотом и  трансляцией.
трансляцией.
Механизм присоединения hGH к его рецептору хорошо изучен (Wells, Curr. Opin. Cell Biol. 6:163, 1994; Clackson и др.. Science 267:383, 1995) и является последовательным. Результатом начального связывания с высоким сродством (nМ) гормона с одним рецептором является потеря поверхности на рецепторе площадью 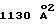 . Второй hGHbp2 имеет существенно меньшую поверхность взаимодействия (de Vos и др., Science 255:306, 1992) со вторым сайтом связывания на hGH и взаимодействует только с предварительно образованным комплексом 1: 1, давая площадь потерянной поверхности
. Второй hGHbp2 имеет существенно меньшую поверхность взаимодействия (de Vos и др., Science 255:306, 1992) со вторым сайтом связывания на hGH и взаимодействует только с предварительно образованным комплексом 1: 1, давая площадь потерянной поверхности 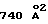 с hGH и
с hGH и  с первым hGHbpl (de Vos и др., Science 255:306, 1992; Wells, Curr. Opin. Cell Biol. 6:163, 1994; Clackson и др., Science 267:383, 1995). Детерминанты связывания каждого hGHbp состоят из шести петель узнавания (L1-L6), три из которых (L1-L3) идут с одного конца β-сандвичевой структуры в D1, одна от внутридоменного линкера и две от D2.
с первым hGHbpl (de Vos и др., Science 255:306, 1992; Wells, Curr. Opin. Cell Biol. 6:163, 1994; Clackson и др., Science 267:383, 1995). Детерминанты связывания каждого hGHbp состоят из шести петель узнавания (L1-L6), три из которых (L1-L3) идут с одного конца β-сандвичевой структуры в D1, одна от внутридоменного линкера и две от D2.
Хотя эти два комплекса рецептора, ЕВР-ЕМР1 и hGH-hGHbp, имеют разные димерные структуры, которые, возможно, представляют в этом случае отличия размера и формы природного и синтетического лиганда, оба рецептора имеют эквивалентные петли узнавания лиганда, L1, L3, L5 и L6 для ЕВР и L1-L6 для hGHbp. Неактивный PRLR, закомплексованный только с одной молекулой hGH, также использует такие же контактные петли (L1-L6) (Somers и др., Nature 372: 478, 1994). Основываясь на схожести сайтов узнавания в hGHbp и PRLR, можно ожидать, что сайт связывания ЕВР, если присоединяется природный лиганд ЕРО, будет распространяться, включая две дополнительные петли, L2 и L4, которые включают остатки 59-63 (L2) между цепями С и С', и остатки 110-118 (L4) от карбоксильного конца βG D1 и междоменного линкера. Эти шесть петель в областях ЕВР, hGHbp и PRLR находятся в структурно эквивалентных положениях, но различаются по размеру, аминокислотному составу и конформации, хотя части взаимодействия каждой петли (боковая сторона или верхушка) остаются одинаковыми, L1, L2, L3, L5 взаимодействуют главным образом своими верхушками, a L6 боковой стороной. В ЕВР L5 на три остатка короче, чем в hGHbp и PRLR, тогда как петля L6 на три и четыре остатка длиннее, чем в hGHbp и PRLR, соответственно. Петля L2 тоже различна (от 6 до 10 остатков) для трех рецепторов, но в ЕВР не участвует в пептидной связи, а в hGHbp частично разупорядочена, хотя она не контактирует с гормоном. С одной стороны, эта ситуация аналогична для областей, определяющих комплементарность (CDR's) в антителах, где изменения длины и последовательности шести петель связывания обеспечивают специфичность для различных антигенов, в то время как сам каркас остается постоянным (Wilson и др., Ciba Foundation Symposium. Wiley, Chichester, 1991, Vol. 159, р.13).
Для комплекса hGH-hGHbp показано, что только подмножество 9 из 33 взаимодействующих остатков, которые создают структурный эпитоп рецептора, составляет функциональный эпитоп или "горячую точку" (Wells, Curr. Opin. Cell Biol. 6: 163, 1994; Clackson и др., Science 267:383, 1995), где имеет место связывания с высоким сродством. Этот уменьшенный эпитоп существенно меньше, чем структурный эпитоп, и состоит из остатков (Arg43, Glu44, Ilе103, Trp104, Ile105. Pro106, Asp165 и Тrр169), которые расположены в контактных петлях L1, L3 и L5 с наиболее существенным вкладом (>4,5 ккал/моль) от двух ароматических остатков (Трr104 и Trp169) в L3 и L5 (Wells, Curr. Opin. Cell Biol. 6:163, 1994; Clackson и др., Science 267:383, 1995; Wells, Proc. Natl. Acad. Sci. USA, 93:1, 1996). В ЕВР Phe93 эквивалентен Трr104 в hGHbp, как предполагали ранее (Wells, Curr. Opin. Cell Biol. 6:163, 1994; Clackson и др. , Science 267:383, 1995; Wells, Proc. Natl. Acad. Sci. USA, 93:1, 1996; Jolliffe и др., Nephrol. Dial. Trans. 10:suppl. 2, 28, 1995), но отсутствуют остатки, гомологичные Trp169 в более короткой петле L5. В комплексе ЕВР-ЕМР1 ароматическая боковая цепь пептида Pht98 занимает эквивалентное положение боковой цепи Trp169 в hGHbp. Можно предположить, что если ЕРО связан со своим рецептором, гормон может обеспечить ароматическим остатком гидрофобную сердцевину промежуточной поверхности взаимодействия, и/или L6 в ЕВР может играть более существенную роль в связывании гормона, чем в hGHbp, так как она на три остатка длиннее и содержит ароматический Pht205.
В этих трех структурах рецепторов класса-1 некоторые петли разупорядочены, что имеет место в D2 для ЕВР (остатки 164-166) в ЕВР1 и 133-135 в ЕВР2) и в D1 для обоих, hGHbp (остатки 55-58, 73-78 для hGHbpl и 54-606 73-75 для hGHbp2) и PRLR (остатки 31-33, 84-86). С другой стороны, эти три рецептора цитокина класса-1 несильно различаются по третичным структурам; D1 и D2 имеют во всех трех рецепторах похожие в общих чертах основные области структуры, например: угол между длинными осями двух доменов равен примерно 90 градусам. Такая структура доменов позволяет этим отдельным петлям L1-L6 быть доступными для узнавания и связывания лиганда. В комплексе 2:2 между IFN-γ и его рецептором IFN-γRα класса-2 D1 и D2 располагаются под углом 125 градусов относительно друг друга, что удлиняет рецептор и ограничивает детерминанты связывания, что можно использовать для взаимодействия с гормоном; петля L1 теперь становится закрытой в D1-D2 промежуточной области, хотя другие пять петель (L2-L6) все еще доступны для взаимодействия лигандов. Такую удлиненную междоменную структуру наблюдают также в факторе ткани (Muller и др. , Biochemistry 33:10864, 1994; Harlos и др. Nature 370:662, 1994), который имеет отдаленное отношение к суперсемейству рецепторов цитокина.
Анализ мутаций молекулы ЕВР показывает, что решающими аминокислотными остатками для присоединяющегося ЕРО в большинстве случаев являются Phe93 в петле L3 (Jolliffe и др. , Nephrol. Dial. Trans. 10:suppl 2, 28, 1995). Phe98Ala мутант показывает увеличение величины IC50 приблизительно в 1000 раз по сравнению с природным типом, в то время как другие мутанты (Ser91Ala, Ser92Ala, Val94Ala, Met50Ala и Hisl53Ala) демонстрируют относительно небольшие увеличения величины IC50, только в 2,5-12,5 раза. Боковые цепи Phe93 закрывают  молекулярной поверхности, что является максимальным среди взаимодействующих боковых цепей. В hGHbp соответствующая мутация Тrр104А1а дает увеличение Кb более, чем в 2500 раз по сравнению с природным типом, указывая на эквивалентную важность этого остатка в связывании hGH и его ключевой вклад в гидрофобную сердцевину функционального эпитопа (Wells, Curr. Opin. Cell Biol. 6:163, 1994; Clackson и др., Science 267:383, 1995; Bass и др., Ргос. Natl. Acad. Sci. USA 88:4498, 1991).
молекулярной поверхности, что является максимальным среди взаимодействующих боковых цепей. В hGHbp соответствующая мутация Тrр104А1а дает увеличение Кb более, чем в 2500 раз по сравнению с природным типом, указывая на эквивалентную важность этого остатка в связывании hGH и его ключевой вклад в гидрофобную сердцевину функционального эпитопа (Wells, Curr. Opin. Cell Biol. 6:163, 1994; Clackson и др., Science 267:383, 1995; Bass и др., Ргос. Natl. Acad. Sci. USA 88:4498, 1991).
Роль димеризации в трансдукции сигнала
В структуре комплекса ЕВР-ЕМР1 авторы к своему удивлению наблюдают, что пептид, не имеющий отношения к природному лиганду (по последовательности и, вероятно, по структуре), может индуцировать биологически активную димеризацию рецептора ЕРО, которая стимулирует трансдукцию сигнала и пролиферацию клеток. Из сравнения трех комплексов рецептора цитокина класса-1, структуры которых пока определены, полагают, что, если природный гормон ЕРО, для которого предложена структура четырехцепного пучка (Boissel и др., J. Biol. Chem. , 268:15983, 1993), индуцирует димеризацию рецептора, то с большей вероятностью он имеет сходство с соединением hGH-hGHbp. Это, возможно, предполагает, что для инициации каскада событий, которые продуцируют биологически уместный сигнал, дозволено более одного способа продуктивной межклеточной димеризации для внутриклеточной димеризации цитоплазматических доменов с молекулами JAK2 (Ihle и др., Seminars in Immunology 5:375, 1993; Klingmuller и др., Cell 80:729, 1995). По-видимому, если структура пептид-ЕВР представляет только одно возможное димерное соединение, это стимулирует трансдукцию сигнала.
Показано, что мутантные молекулы EPOR, содержащие одну мутацию Arg в Cys (Arg130 у человека и Arg129 у крысы) в отсутствие ЕРО образуют биологически активные димеры (Yoshimura и др., J. Biol. Chem. 267:11619, 1992; Watowich и др. , Proc. Natl. Acad. Sci. USA 89:2140, 1992; Watowich и др., Mol. Cell. Biol. 14:3535, 1994), предполагая, что внеклеточная гомодимеризация рецептора может быть достаточна сама по себе для трансдукции сигнала. В другой системе показана (Spencer и др., Science 262:1019, 1993), что активацию специфического множества факторов транскрипции можно вызвать путем химического кросс-связывания цитоплазматических доменов модифицированных рецепторов клеточной мембраны, которые не содержат внеклеточных и трансмембранных доменов. Эти рецепторы не относятся к суперсемейству рецепторов цитокина, но иллюстрируют, что олигомеризация играет ключевую роль в активации рецептора, и что основной функциональной ролью внеклеточного домена, связывающего лиганд, является разрешение (в присутствии лиганда) димеризацию или олигомеризацию и вызвать аналогичную ассоциацию цитоплазматических доменов.
Эксперименты по мутагенезу первоначально отводили роль мотива WSXWS в этом процессе клеточной передачи сигнала (Yoshimura и др., J. Biol. Chem. 267: 11619, 1992; Quelle и др., Mol. Cell. Biol. 12:4553, 1992; Chiba и др., Biochem. Biophys. Res. Comm. 184:485, 1992) возможной стимуляции гомодимеризации рецептора. Однако мутанты процессирования (truncation mutants) EPOR (Miura и др., Arch. Biochem. Biophys. 306:200, 1993) не подтверждают эту роль для мотива WSXWS. Структура комплекса ЕВР-ЕМР1 показывает, что мотив WSXWS EPOR, как для комплекса hGH-hGHbp (de Vos и др., Science 255:306, 1992), расположен на противоположной от рецептора димеризации поверхности молекулы. В отсутствие структур с одинаковыми лигандами для внеклеточных доменов EPOR, hGHR и PRLR невозможно определить, происходит ли какое-либо изменение конформации при связывании лиганда, который, возможно, включает блок WSXWS. Не говоря о наличии замечательной структурной особенности в D2 и его очевидной близости к домену присоединения мембраны (membrane spanning domen), нельзя исключать возможные взаимодействия этой области с некоторыми другими молекулами клеточной поверхности, которые так или иначе включены в процесс трансдукции сигнала.
Относительно разработки малых молекул-миметиков
Структура димера ЕМР1 показывает, что пептид, значительно меньший, чем природный гормон, может действовать как агонист и индуцировать соответствующую биологическую реакцию. Можно допустить, что пептид образует существенно меньшую поверхность контакта с рецептором, чем природный гормон. Сайт связывания пептида с ЕВР образует почти плоскую поверхность, в основном гидрофобную по природе, без впадин или заряженных остатков, что обычно существенно для специфической реакции малых молекул-лигандов с сайтом связывания рецептора. Исследование hGHbp (Wells и др., Science 267:383, 1995; Wells, Proc. Natl. Sci. USA 93:1, 1996) показывает, что только небольшая часть наблюдаемых структурных сайтов связывания, так называемый функциональный эпитоп (supra), дает наибольший вклад энергии взаимодействия, и строго подразумевается, что "доведенный до минимума" гормон, предназначенный для взаимодействия с этим сайтом, может образовать достаточно взаимодействий для активации рецептора. Кроме того, ограниченный сайт взаимодействия малого агониста пептида с ЕВР почти точно соответствует меньшему функциональному эпитопу, полученному сканированием аланина в hGH и hGhbbp. Таким образом, применяя различные подходы, авторы пришли к одинаковому выводу, что вносить свой вклад в функциональный эпитоп на рецепторе может небольшое количество ключевых взаимодействий. Понимание этой упрощенной поверхности взаимодействия можно теперь объединить с дополнительными мутационными исследованиями в помощь идентификации наиболее важных остатков в функциональном эпитопе и, следовательно, для обеспечения более практической задачи - разработки лекарственных средств.
Сбор данных, многократное изоморфное замещение (MIR) и статистическая обработка
Кристаллографические данные суммированы в таблице 1. Природные кристаллографические данные получают на многопроводном детекторе поверхности фирмы Simens "Simens multiwire area detector", смонтированном на генераторе "Elliott GX-18 generator", работающем при 40 кВ и 55 мА, с расстоянием кристалл-детектор 120 мм. Два набора производных данных собирают на плоскости для получения изображения "MAR image plate", смонтированной на генераторе фирмы Simens, работающем при 50 кВ и 80 мА, с расстоянием кристалл-изображение 150 мм. Данные интегрируют, сводят к определенному масштабу и обрабатывают, используя программы XENGEN (Howard и др., J. App. Cryst. 20:383, 1987) для природных данных и DENZO/SCALEPACK (Otwinowski и др., SERC Darsbury Laboratory, Warrington, 1993) для производных данных. Рассчитывают начальные фазы аномального рассеивания множественного изоморфного замещения (MIRAS) до 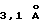 , используя пакет программ PHASES (Furey, American Crystallographic Association Fortieth Anniversary Meeting, New Orleans, LA, 1990) со средним критерием качества 0,64
, используя пакет программ PHASES (Furey, American Crystallographic Association Fortieth Anniversary Meeting, New Orleans, LA, 1990) со средним критерием качества 0,64  . Фазы обрабатывают посредством PHASES, используя протокол "solvent flattening protocol" до среднего критерия качества 0,92
. Фазы обрабатывают посредством PHASES, используя протокол "solvent flattening protocol" до среднего критерия качества 0,92  . Качество карты обычно хорошее и большинство структур комплексов (94%) можно подогнать, используя графическую программу О (Jone и др., Acta Crystallogr. A47:110, 1991). Список аминокислотных остатков проверяют из расположения двух дисульфидных мостиков в D1 и расположения двух атомов Нg от производного ацетата ртути, для которого правильно предположено, что оно связано со свободным остатком Cys181. Пептидное взаимодействие проверяют из другого набора данных от комплекса между ЕВР и йодированным пептидом (ТуrP4 замещает пара-йод-Phe), который дает дифракцию до разрешения
. Качество карты обычно хорошее и большинство структур комплексов (94%) можно подогнать, используя графическую программу О (Jone и др., Acta Crystallogr. A47:110, 1991). Список аминокислотных остатков проверяют из расположения двух дисульфидных мостиков в D1 и расположения двух атомов Нg от производного ацетата ртути, для которого правильно предположено, что оно связано со свободным остатком Cys181. Пептидное взаимодействие проверяют из другого набора данных от комплекса между ЕВР и йодированным пептидом (ТуrP4 замещает пара-йод-Phe), который дает дифракцию до разрешения 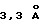 , что по разнице Фурье-функций (Fйод-Fприродн)αMIRAS дает ясное указание расположения атомов йода. Структуру совершенствуют, используя протокол слабого охлаждения "slow-cooling protocol" в X-PLOR 3.1 (Brurynger и др., Acta Crystallogr. А46:585, 1990; Brunger, X-PLOR, Version 3.1: A System for X-ray and NMR, Yale Univ. Press, New Haven, CN, 1992) и восстанавливают, используя Fo-Fc, 3Fo-2Fc и оцененные посредством SIGMAA (Read, Acta Crystallogr. A42:140, 1986) карты электронной плотности. После каждых двух циклов обработки рассчитывают набор моделированных карт, исключающих отжиг (set of simulated anneling omit maps) (7-10%) для уменьшения отклонения рассчитанной модели и восстановления целой структуры. После нескольких циклов обработки рассчитывают индивидуальные температурные коэффициенты и через 10 циклов обработки и построения модели величина R составляет 0,21 для
, что по разнице Фурье-функций (Fйод-Fприродн)αMIRAS дает ясное указание расположения атомов йода. Структуру совершенствуют, используя протокол слабого охлаждения "slow-cooling protocol" в X-PLOR 3.1 (Brurynger и др., Acta Crystallogr. А46:585, 1990; Brunger, X-PLOR, Version 3.1: A System for X-ray and NMR, Yale Univ. Press, New Haven, CN, 1992) и восстанавливают, используя Fo-Fc, 3Fo-2Fc и оцененные посредством SIGMAA (Read, Acta Crystallogr. A42:140, 1986) карты электронной плотности. После каждых двух циклов обработки рассчитывают набор моделированных карт, исключающих отжиг (set of simulated anneling omit maps) (7-10%) для уменьшения отклонения рассчитанной модели и восстановления целой структуры. После нескольких циклов обработки рассчитывают индивидуальные температурные коэффициенты и через 10 циклов обработки и построения модели величина R составляет 0,21 для 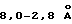 данных с F>1σ (13984 отражений).
данных с F>1σ (13984 отражений).
Средние тепловые параметры для рецептора I, рецептора II и пептидов составляют 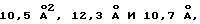 соответственно. Только один неглициновый остаток [Asn164 d ЕВР2] , расположенный в петлевой области в D1, является запрещенным участком в диаграмме "Ramachandran plot". Молекулы растворителя не включены в модель из-за посредственного разрешения
соответственно. Только один неглициновый остаток [Asn164 d ЕВР2] , расположенный в петлевой области в D1, является запрещенным участком в диаграмме "Ramachandran plot". Молекулы растворителя не включены в модель из-за посредственного разрешения  в определении структуры.
в определении структуры.
Связывающие контакты
Связывающие контакты частично суммированы в таблице 2. Взаимодействия типа водородных связей в сайтах связывания комплексов ЕВР-ЕМР1. Вследствие симметричной природы комплекса пептид-1 и пептид-2 имеют эквивалентные взаимодействия с двумя молекулами ЕВР. Взаимодействия типа водородных связей анализируют, применяя HBPLUS (McDonald и др., J. Mol. Biol. 238:777, 1994), основанную на расстоянии ( порог) и геометрических соображений.
порог) и геометрических соображений.
Описано множество вариантов настоящего изобретения. Однако будет понятно, что можно сделать различные модификации, не отклоняясь от духа и области данного изобретения. Таким образом, следует понимать, что данное изобретение не ограничено отдельным иллюстрированным вариантом, а только областью приложенной формулы изобретения.
















Изобретение относится к компьютерным способам идентификации и расчета малых молекул-миметиков эритропоэтина (ЕРО). Способы предназначены для идентификации молекул, которые присоединяются к рецепторам ЕРО и действуют как миметики ЕРО. Предпочтительные миметики ЕРО, идентифицированные с использованием способа данного изобретения, действуют как агонисты рецепторов ЕРО в одном или более in vitro и in vivo биологических исследованиях активности ЕРО. Это позволяет расширить область применения, так как способы можно использовать для существенного увеличения количества потенциальных миметиков, которые следует проверить на ЕРО-активность. 3 с. и 3 з.п. ф-лы, 2 ил., 2 табл.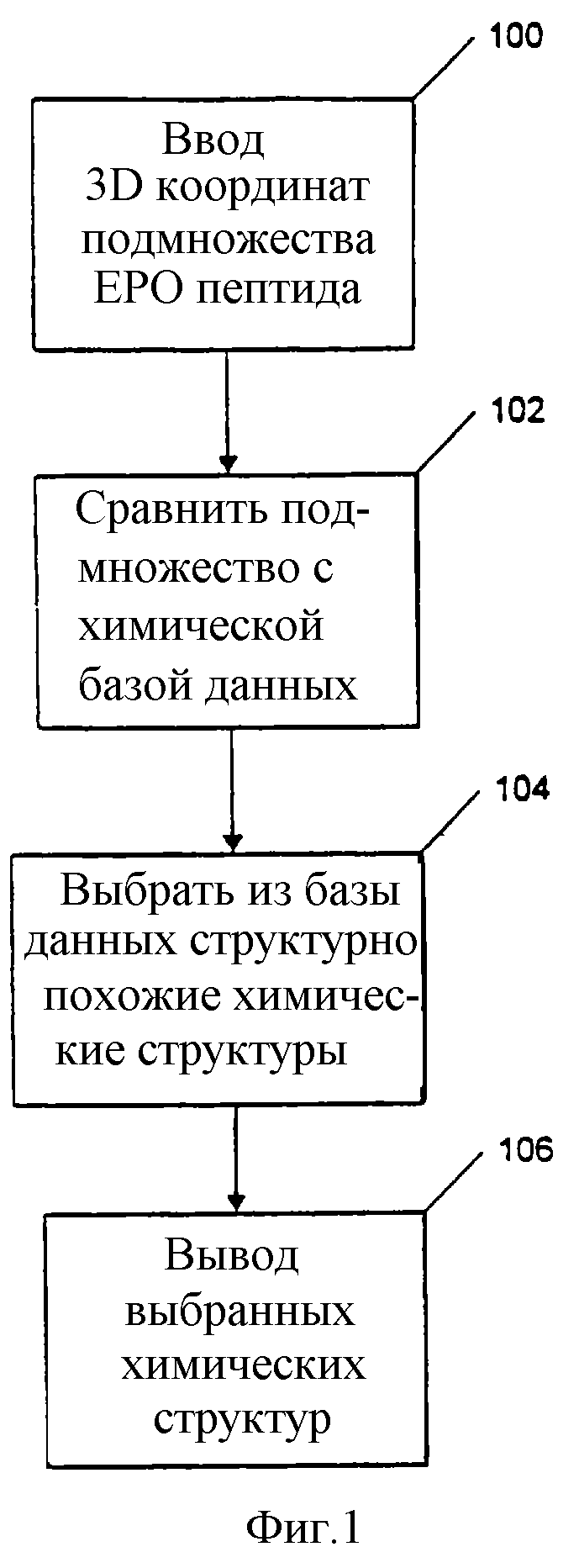
| НЕЙЛОР К | |||
| Как построить свою экспертную систему | |||
| Пер | |||
| с англ | |||
| - М.: Энергоатомиздат, 1991, с | |||
| Устройство для вытяжки и скручивания ровницы | 1923 |
|
SU214A1 |
| УИНСТОН П | |||
| Искусственный интеллект | |||
| - М.: Мир, 1980, с | |||
| СПОСОБ ПОЛУЧЕНИЯ ЧИСТОГО ГЛИНОЗЕМА И ЕГО СОЛЕЙ ИЗ СИЛИКАТОВ ГЛИНОЗЕМА, ПРОСТЫХ ГЛИН И. Т.П. | 1915 |
|
SU280A1 |
| US 5331573 А, 19.07.1994. | |||

