Изобретение откосится к области детекции последовательностей нуклеиновых кислoт. Настоящее изобретение касается способа "детекции мутации любого типа в заданном представляющем интерес сайте нуклеиновой кислоты. Настоящее изобретение связано со способом, названным как "анализ со сдвигом терминации", также известным как "тест на специфическую, терминацию" или "сопоставление смещенных концов", а сокращенно он может быть обозначен как "способ STA".
Практическое применение заявляемого настоящим изобретением способа включает диагностику генетических заболеваний, диагностику инфекционных заболеваний, судебно-медицинские манипуляции, установление отцовства и картирование геномов, при том условии, что сайт, в котором детектируется мутация, заранее известен.
За последнее десятилетие были идентицифированы гены, участвующие в наследственной предрасположенности к развитию рака, а также было охарактеризовано значительное количество связанных с раком генных мутаций. Тесты на диагностику этих мутаций могут позволить более точно оценивать индивидуальный риск развитая рака. Ранняя диагностика связанной с раковыми заболеваниями мутации является одной из целей настоящего изобретения.
Существует четыре основных типа мутаций. Первый тип - это точковые мутации, обусловленные заменой одного нуклеотида в нормальной последовательности ДНК. В большинстве случаев такая мутация обусловливает сдвиг рамки считывания в кодирующей цепи, что приводит к преждевременной терминации синтеза нормального белка. Характерным примером является точковая мутация в гене АРС, обнаруживаемая у пациентов с семейным диагнозом адено-полипоза (Kinzler et al. , 1991, Science 253, 661-665; Joslyn et al., 1991, Cell 66, 601-613, Nishisho et al., 1991, Science, 253, 665-669). Второй тип представлен мутациями типа вставок, когда один или множество нуклеотидов включаются в нормальную последовательность ДНК. Третий тип мутаций - делении, при которых один нуклеотид или множество нуклеотидов делегируются из нормальной последовательности ДНК. Оба этих типа мутаций (вставки и делении) могут обусловливать такие серьезные изменения, как сдвиг рамки считывания, преждевременную терминацию синтеза белка и включение или утрату одной или множества аминокислот в полипептиде. Четвертый тип - транслокация гена, которая происходит тогда, когда фрагмент одного гена включается в другой ген. Примером такого явления служит хромосома Philadelphia, наблюдаемая у пациентов с хроническим миелоидным лейкозом (Konopka et al., 1984, Cell 37, 1035). Изменения в структуре белка обусловливают серию аномалий в клетке, которые могут приводить к развитию рака.
Выявление одной мутированной ДНК среди тысяч нормальных ДНК является трудоемким процессом. Метод химического или каталитического секвенирования ДНК, нацеленный на прямое "считывание" последовательности ДНК выделенного образца, применяется как наиболее точный метод для исследовательской лаборатории с точки зрения анализа и идентификации генных мутаций. Однако применение такого метода секвенирования в медицине непрактично из-за нехватки специалистов соответствующей квалификации, которые должны проводить такие анализы, из-за его трудоемкости, высокой стоимости закупок потребного оборудования и реагентов, необходимых для осуществления реакций секвенирования, равно как и из-за значительного времени, требуемого для проведения такого анализа. Наконец, другим недостатком метода секвенирования является то, что для этой процедуры требуется значительное количество ДНК-матрицы, которое бывает трудно получить из 10 мл образца крови, которое обычно получают от пациента.
Примерами стандартных подходов к идентификации мутаций являются: выявление ПДРФ - полиморфизма длин рестрикционных фрагментов (Botstein et al., 1980, Amer. J. Human Genet., 32, 314-331; White et al., 1988, Sci. Amer., 258, 40-48); выявление ОЦКП - одноцепочечного конформационного полиморфизма (Howell et al., 1994, Amer. J. Hum. Genet., 55, 203-206); проведение гибридизации с аллель-специфичными олиго-нуклеотидными зондами (Studencki et al., 1985, Amer. J. Human Genet., 37, 42-51; Saiki et al., Nature, 324, 163-166); тест с лигированием олигонуклеотидов (Landgrun et al., 1990, Science, 241, 1077-1080); и аллель-специфичная ПЦР (АС-ПЦР) (Wu et al., 1989, Proc. Nati. Acad. Sci. USA, 86, 2757-2760; Okayama et al, 1989, J. Lab. Clin. Med., 114, 105-113).
Некоторые из этих методов применимы только для выявления точковых мутаций. Некоторые другие методы могут быть применены с целью выявления только вставок или делеций, которые могут, например, разрушать или создавать сайты рестрикционного расщепления, но при этом не пригодны для выявления мутаций, затрагивающих отдельные нуклеотиды. Например, точковые мутации, которые не затрагивают рестрикционные сайты, пропускаются при применении таких методов, как ПДРФ. В других метода необходимым является обеспечение оптимизации условий с точки зрения специфичности гибридизации конкретного зонда. Кроме того, для всех упоминавшихся выше методов требуется специальное лабораторное оборудование, такое как устройство для гель-электрофореза и оборудование для гибридизации, а также значительные трудозатраты и время.
Также известен ряд методов выявления мутаций, основанных на достройке затравок (Mohan et al. , 1991, Proc. Natl. Acad. Sci. USA, 83, 1143-1147; Prezant et al., 1992, Human Mutat., 1, 159-164; Fahy et al., 1997, Nucl. Acid Res. , 25, 3102-3109; патенты США 5846710 и 5888819 за 1998 г.). Такими методами являются: достройка затравок тионуклеотидами: достройка затравок начиная от олигонуклеотида, фланкирующего мутантный нуклеотид помеченным нуклеотидом, комплементарным мутировавшему нуклеотиду; и достройка затравок помеченным дидезоксинуклеотидным терминатором, комплементарным мутантному нуклеотиду.
Такие методы выявления мутаций, основанные на достройке затравок, являются быстрыми, простыми в осуществлении и могут быть применены в клинической практике. Однако у этих методов имеется по крайней мере два недостатка. Во-первых, все эти методы основаны на включении только одного помеченного нуклеотида в достраиваемую от затравки цепь. Включение только одного варианта помеченного нуклеотида, выбираемого из А, С, G, Т или U, или помеченного дидезоксинуклеотида, обеспечивает выявление только конкретной точковой мутации, которая специфична в отношении того нуклеотида, который комплементарен данному помеченному нуклеотиду, который включен в такой тест.
Гипотетически, когда в том же положении имеется мутация другого типа или отличающейся природы, например, когда А заменяется на С или на GT, или на ТСТ, при безграничном количестве других возможных мутаций, то эти известные методы потребуют проведения по крайней мере трех отдельных тестов с использованием помеченных нуклеотидов G, С и А, С другой стороны, единый тест может быть проведен с использованием, по-разному помеченных нуклеотидов вкупе с разделением в геле и выявлением по отдельности мутантных нуклеотидов G, С и А с помощью отдельных маркеров. Однако проведение трех раздельных тестов требует по крайней мере втрое большего объема проб крови, которые обычно получают от пациента. Такой высокий объем пробы крови или усложненная процедура разделения в геле не только приводят к удорожанию такого, теста, а также увеличивают затраты времени и труда, но, что более важно, повышается вероятность ошибок, что обусловливается вероятностью неправильного маркирования пробирок из-за многочисленности этапов, из которых состоит такое тестирование. Таким образом, данные методы, основанные на достройке затравок, непригодны для тестирования большого числа образцов или для проведения рутинного тестирования в клинических условиях.
Во-вторых, чувствительность данных методов, основанных на достройке затравок, нуждается в повышении. Поскольку получаемая в этих методах цепь, являющаяся достроенной затравкой, включает только один помеченный нуклеотид или помеченный дидезоксинуклеотид, то полученные сигналы варьируются, а их сила зависит от типа использовавшейся химической метки. Однако в целом такой сигнал оказывается слабым.
Таким образом, имеется необходимость в выявлении мутаций с помощью быстрого, относительно дешевого, с невысокими трудозатратами и клинически эффективного способа, с помощью которого можно было бы выявлять (детектировать) мутацию любого типа по нуклеотиду, находящемуся в заданном конкретном положении, на основании мощного и точного детекционного сигнала.
Резюме изобретения
Настоящее изобретение было сформулировано перед описанием потребностей данной области.
Даже при том, что настоящее изобретение имеет общие черты, с некоторыми преимуществами, характерными основным методам, основанным на достройке затравок, такие как простота, собственно, тестирования мутации в конкретном сайте, тем не менее, настоящее изобретение представляет способ, который преодолевает те недостатки, которые характерны для методов достройки затравок и были описаны выше. Настоящее изобретение предоставляет возможности широкого применения в выявлении и идентификации всех типов мутаций. Оно является экономичным, времясберегающим и менее трудоемким по сравнению с имеющимися стандартными методами.
Некоторые из ключевых преимуществ настоящего изобретения по отношению к описанным выше методам таковы: (1) возможность детекции мутаций всех типов в единственной реакционной пробирке без необходимости применения метода электро-форетического разделения фрагментов в геле; 2) высокий уровень чувствительности детекции за счет получения мощного сигнала, вырабатываемого за счет включения множественных помеченных нуклеотидов в цепь, достраиваемую от затравки; и (3) высокая степень точности, обусловливаемая тем, что два или три различных типа маркеров нуклеотидов или аналогов нуклеотидов могут быть включены в достраиваемую от затравки цепь одновременно. Эти преимущества представляют возможность применения настоящего изобретения для рутинного тестирования присутствия генетической мутации в любых клинических условиях, основываясь на заявляемом простом способе. Также настоящее изобретение может быть легко адаптировано для проведения автоматизированного скрининга большого числа образцов.
Настоящее изобретение касается способа детекции любой мутации, происходящей в заданном сайте (нуклеотиде-мишени) в известной нуклеотидной последовательности, в одной реакции. Заявляемый способ выявления мутации основан на анализе с достройкой затравки. Предпочтительно эта затравка комплементарна интересующей нуклеотидной последовательности по положению, непосредственно примыкающему к заданному нуклеотиду, и специфическим образом гибридизует с ней с образованием дуплекса таким образом, что нуклеотид-мишень в анализируемой нуклеотидной кислоте оказывается неспаренным основанием, расположенным непосредственно с 3'-конца данной затравки. Реагенты для реакции достройки затравки включают один тип непомеченного терминаторного нуклеотида (или, что необязательно, при отсутствии соответствующего нуклеотида) наряду с тремя типами помеченных (или, что необязательно, по-разному помеченных или непомеченных) нетерминаторных нуклеотидов, при том, что нуклеотид-терминатор комплементарен нуклеотиду-мишени по заданному положению анализируемой нуклеиновой кислоты. При этом помеченные нетерминаторные нуклеотиды некомплементарны нуклеотиду-мишени. Включение терминаторного нуклеотида с 3'-конца используемой затравки, комплементарного нуклеотиду-мишени анализируемой нуклеиновой кислоты, будет обусловливать терминацию реакции достройки затравки без дальнейшего включения любого из помеченных нетерминаторных нуклеотидов. Если нуклеотид-мишень изменен вследствие мутации, то помеченные нетерминаторные нуклеотиды в зависимости от данной последовательности будут встраиваться в данную затравку. Таким образом, любой сигнал метки, выявляемый в данной затравке, указывает на то, что в заданном сайте нуклеиновой кислоты произошла мутация.
Объектом настоящего изобретения является представление способа детекции или количественного анализа нуклеиновой кислоты-мишени в образце, включающего:
(a) получение затравки, комплементарной последовательности, непосредственно примыкающей к нуклеотиду-мишени в заданном положении анализируемой нуклеиновой кислоты, которая служит матрицей;
(b) обработку образца, содержащего анализируемую нуклеиновую кислоту, в случае, если она является двухцепочечной, таким образом, чтобы получить неспаренные нуклеотиды, охватывающие конкретный сайт, или прямое применение стадии (с), если анализируемая нуклеиновая кислота является одноцепочечной;
(c) отжиг затравки, полученный в соответствии с (а), с нуклеиновой кислотой-мишенью по (b) в условиях высокой степени жесткости с получением дуплекса "затравка/нуклеиновая кислота", при том, что нуклеотид-мишень в последовательности анализируемой нуклеиновой кислоты является первым неспаренным нуклеотидом, находящимся сразу за 3'-концом данной затравки;
(d) смешивание дуплекса "затравка/нуклеиновая кислота" по (с) с реагентами реакции достройки затравки, включающими: (i) один тип терминаторного нуклеотида или необязательно "нуклеотидный пропуск", который комплементарен нуклеотиду-мишени по заданному положению анализируемой нуклеиновой кислоты; и (ii) три типа нетерминаторных нуклеотидов, которые не совпадают с терминаторным нуклеотидом по п.(i), причем по крайней мере один из этих типов помечен выявляемым маркером;
(e) проведение реакции достройки затравки каталитическими или химическими способами, при том, что включение упомянутого терминаторного нуклеотида или нетерминаторного нуклеотида в затравку зависит от идентичности неспасенного нуклеотида, находящегося в нуклеиновой кислоте-матрице сразу за 3'-концом затравки, и при том, что включение упомянутого терминаторного нуклеотида в последовательность, комплементарного упомянутому нуклеотиду-мишени анализируемой нуклеиновой кислоты, будет обеспечивать терминацию упомянутой достройки затравки без возможности включения любого из помеченных нетерминаторных нуклеотидов в состав затравки, при том, что упомянутая затравка не помечена, а также при том, что, когда нуклеотид-мишень изменен на любой иной нуклеотид, тогда один из нетерминаторных нуклеотидов, помеченных упомянутым выявляемым маркером, или, необязательно, непомеченных каким-либо маркером нуклеотидов, если в качестве метода детекции применяется масс-спектрометрия, который комплементарен мутантному нуклеотиду, в зависимости от конкретной последовательности, включают в состав затравки в упомянутой реакции достройки затравки; и
(f) установление присутствия и идентичности нуклеотида в заданном положении анализируемой нуклеиновой кислоты путем детекции включенного в данную затравку помеченного нетерминаторното нуклеотида.
В предпочтительном варианте на стадии (b) анализируемый нуклеотид непосредственно примыкает к нуклеотиду, который должен быть идентифицирован в заданном положении, а тот нуклеотид, который должен быть идентифицирован, является неспаренным нуклеотидом, находящимся в заданном положении непосредственно за 3'-концом образованного дуплекса. В предпочтительном варианте данного способа на стадии (d) дуплекс по (с) контактируют с по крайней мере одним помеченным нетерминаторным нуклеотидом и по крайней мере с одним непомеченным терминаторным нуклеотидом. С другой стороны, на стадии (d) дуплекс по (с) контактируют с нетерминаторными нуклеотидами, при том, что каждый из таких нетерминаторных нуклеотидов помечают одной и той же или различными метками.
В другом предпочтительном варианте описанный выше способ может быть осуществлен на практике так, чтобы определяемым матрицей ферментом являлась ДНК-полимер а за I E.coli или ее т.н. "фрагмент Кленова", ДНК-полимер а за фага Т4, ДНК-полимераза фага Т7, ДНК-полимераза Thermophilus aquaticus, ретровирусная обратная транскриптаза или их сочетания.
В других предпочтительных вариантах нуклеиновой кислотой по настоящему изобретению является дезоксирибонуклеиновая кислота, рибонуклеиновая кислота или сополимер дезоксирибонуклеиновой кислоты и рибонуклеиновой кислоты. Затравкой является олигодезоксирибонуклеотид, олигорибонуклеотид или сополимер дезоксирибонуклеиновой кислоты и рибонуклеиновой кислоты. Если матрицей является дезоксирибонуклеиновая кислота, то затравкой является олигодезоксирибонуклеотид, олигорибонуклеотид или сополимер дезоксирибонуклеотидов и рибонуклеотидов, а определяемым типом матрицы ферментом является ДНК-полимераза. Предпочтительно, если матрицей является рибонуклеиновая кислота, то затравка является олигодезоксирибонуклеотидом, олигорибонуклеотидом или сополимером дезоксирибонуклеотидов и рибонуклеотидов, а определяемым типом матрицы ферментом является обратная транскриптаза. Предпочтительно, если матрицей является дезоксирибонуклеиновая кислота, то затравкой является олигорибонуклеотид, а ферментом является РНК-полимераза. Предпочтительно, если матрицей является рибонуклеиновая кислота, то затравка является олигорибонуклеотидом, а определяемым типом матрицы ферментом является РНК-репликаза.
В описанном выше способе на стадии (d) дуплекс по (с) контактируют с по крайней мере одним помеченным нетерминаторным нуклеотидом и с по крайней мере одним терминаторным нуклеотидом, который помечают не так, как нетерминаторный нуклеотид. Кроме того, на стадии (е) выявляют метку включенного помеченного нетерминаторного нуклеотида и метку по крайней мере одного терминаторного нуклеотида, который помечен отличным от нетерминаторного нуклеотида способом.
В соответствии со способом по настоящему изобретению анализируемая нуклеиновая кислота была синтезирована каталитически in vivo, синтезирована каталитически in vitro или синтезирована некаталитическим путем. В другом варианте способа по настоящему изобретению олигонуклеотидная затравка была синтезирована каталитически in vivo, синтезирована каталитически in vitro или синтезирована некаталитическим путем. Кроме того, олигонуклеотидная затравка может включать одну или большее число составляющих, которая обеспечит аффинное отделение затравки от невключившегося реагента и (или) от анализируемой нуклеиновой кислоты.
В частности, в предпочтительном варианте олигонуклеотидная затравка включает биотин, который обеспечивает аффинное отделение затравки от невключенного реагента и (или) от анализируемой нуклеиновой кислоты за счет связывания биотина со стрептавидином, которые присоединяют к твердой подложке. В другом варианте настоящего изобретения последовательность олигонуклеотидной затравки включает последовательность ДНК, которая обеспечивает аффинное отделение затравки от невключенного реагента и (или) от анализируемой нуклеиновой кислоты за счет спаривания оснований с комплементарной последовательностью, имеющейся в составе нуклеиновой кислоты, присоединенной к твердой подложке. Еще в одном варианте настоящего изобретения анализируемая нуклеиновая кислота включает одно или большее число составляющих, которые обеспечивают аффинное отделение анализируемой нуклеиновой кислоты от невключенного реагента и (или) от затравки. Анализируемая нуклеиновая кислота может включать биотин, который обеспечивает аффинное отделение анализируемой нуклеиновой кислоты от невключенного реагента и (или) от затравки за счет связывания биотина со стрептавидином, который присоединяют к твердой подложке.
В способе по настоящему изобретению последовательность анализируемой нуклеиновой кислоты включает последовательность ДНК, которая обеспечивает аффинное отделение анализируемой нуклеиновой кислоты от невключенного реагента и (или) от затравки за счет спаривания с комплементарной последовательностью, имеющейся в составе нуклеиновой кислоты, присоединенной к твердой подложке. Олигонуклеотидная затравка может быть помечена с помощью выявляемого маркера. Олигонуклеотидная затравка может быть помечена с использованием выявляемого маркера, который бы отличался от любого выявляемого маркера, присутствующего в составе реагента или прикрепленного к анализируемой нуклеиновой кислоте. Анализируемая нуклеиновая кислота может быть помечена с помощью выявляемого маркера. Анализируемая нуклеиновая кислота предпочтительно помечена с использованием выявляемого маркера, который отличается от любого из выявляемых маркеров, присутствующих в составе реагента или присоединенного к затравке.
В другом варианте настоящего изобретения анализируемая нуклеиновая кислота включает ненативный аналог нуклеотида. Ненативными нуклеотидными аналогами являются дезоксиинозин или 7-деаза-2'-дезоксигуанозин. Анализируемая нуклеиновая кислота может быть синтезирована с применением полимеразной цепной реакции.
В другом способе настоящего изобретения образец включает геномную ДНК организма, ее РНК-транскрипты или кДНК, синтезированную на матрице этих РНК-транскриптов. Данный образец может включать внегеномную ДНК организма, ее РНК-транскрипты или кДНК, синтезированную на матрице этих РНК-транскриптов. В способе по настоящему изобретению затравка может быть предпочтительно отделена от анализируемой нуклеиновой кислоты после реакции достройки затравки на стадии (d), названном выше, с использованием подходящих денатурирующих условий. Предпочтительно денатурирующими условиями являются нагревание, обработка щелочью, формамидом, мочевиной, глиоксалем, ферментами и их сочетания. Еще более предпочтительно, денатурирующие условия создаются обработкой 0,2 N едкого натра.
Способ по настоящему изобретению может быть реализован на практике с использованием нуклеиновой кислоты, происходящей от любого организма, включая растения, микроорганизмы, вирусы или птицы. Этот организм может быть позвоночным или беспозвоночным животным. Предпочтительно таким организмом является млекопитающее. Еще более предпочтительно таким млекопитающим является человек. Таким млекопитающим также могут быть лошадь, собака, корова, кошка, свинья или овца.
Эти и другие объекты настоящего изобретения будут полностью ясны из нижеследующего описания настоящего изобретения, прилагаемых чертежей и следующей далее формулы изобретения.
Краткое описание чертежей
На фиг. 1А-1С схематически изображен предпочтительный вариант способа выявления мутации по настоящему изобретению. "L" обозначает нуклеотид дикого типа, который может являться A, G, С, Т или U; "L*" обозначает непомеченный терминаторный нуклеотид, такой как дидезоксинуклеотид, который комплементарен нуклеотиду L. "M" обозначает мутацию в сайте L, а мутантным нуклеотидом является А, G, С, Т или U. "W" обозначает нуклеотид, комплементарный M, и он может являться А, G, С, Т или U, помеченный выявляемым маркером. "n" обозначает один или множество нуклеотидов или аналогов нуклеотидов, включая A, G, С, Т или U. "у" обозначает нуклеотид или аналог нуклеотида, включая А, G, С, Т или U, помеченный выявляемым маркером и комплементарный М или n.
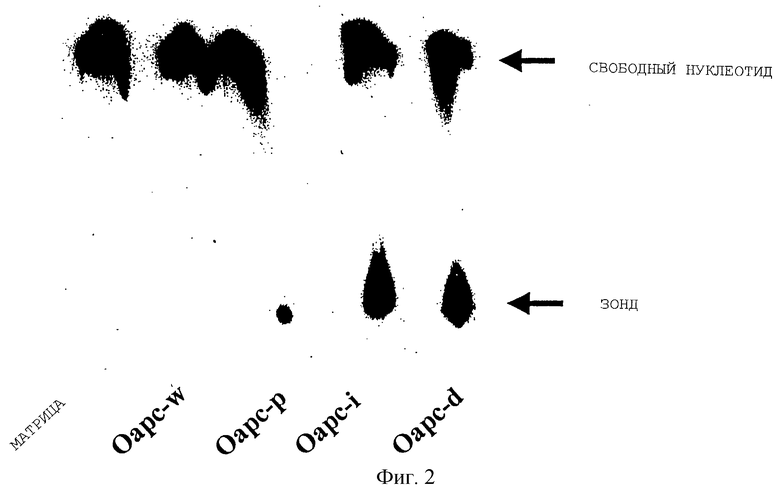
Фиг. 2. 1 мкл реакционной смеси реакции STA наносят на полоску для тонкослойной хроматографии и затем эту полоску обрабатывают растворителем, содержащим 1 М NaCl и 1 М НС1. Затем полоску высушивают при комнатной температуре в течение 10 минут и экспонируют на пленку Kodak в течение 30 минут, после чего пленку проявляют автоматически. Матрицы, используемые в каждом SТА-тесте, помечают внизу каждой полоски. Стрелка ввepxy обозначает свободные нуклеотиды, а нижняя стрелка указывает на цепь достроенной затравки с включенным в нее помеченным [α-32P]дЦТФ.
Подробное описание изобретения
По использованию в данном тексте термины "нуклеиновая кислота" или "нуклеотид" могут соответствовать дезоксирибонуклеиновой кислоте, рибонуклеиновой кислоте или сополимеру дезоксирибонуклеиновой кислоты и рибонуклеиновой кислоты. Образец нуклеиновых кислот может быть нативным или синтетическим. Образец нуклеиновой кислоты может являться встречающейся в естественных условиях нуклеиновой кислотой и может быть получен от любого организма. Некоторыми примерами организмов, к которым может быть применен способ по настоящему изобретению, являются растения, микроорганизмы, вирусы, птицы, позвоночные животные, беспозвоночные животные, млекопитающие, человек, лошади, собаки, коровы, кошки, свиньи или овцы. Являющаяся мишенью нуклеиновая кислота может встречаться в природе или может быть синтезирована каталитически in vivo, синтезирована каталитически in vitro или синтезирована химическим путем.
Образец, содержащий представляющие интерес нуклеиновые(ую) кислоты(у), может включать геномную ДНК организма, ее РНК-транскрипты или кДНК, полученные на матрице таких РНК-транскриптов. Образец, содержащий представляющие интерес нуклеиновые(ую) кислоты(у), также может включать внегеномную ДНК организма, ее РНК-транскрипты или кДНК, полученные на матрице таких РНК-транскриптов. Также представляющие интерес нуклеиновые кислоты (нуклеиновая кислота) могут быть синтезированы методом полимеразной цепной реакции.
Представляющая интерес нуклеиновая кислота может включать ненативные аналоги нуклеотидов, такие как дезоксиинозин или 7-деаза-2'-дезоксигуанозин. Эти аналоги дестабилизируют ДНК-дуплексы и могут обеспечивать отжиг затравки и реакции достройки в образце двухцепочечных образцов без полного разделения их цепей.
Представляющая интерес нуклеиновая кислота может включать одну или большее число составляющих, которые обеспечивают аффинное отделение анализируемой нуклеиновой кислоты от невключенного реагента и (или) от затравки. Например, анализируемая нуклеиновая кислота может включать биотин, который обеспечивает аффинное отделение анализируемой нуклеиновой кислоты от невключенного реагента и (или) от затравки за счет связывания биотина с авидином и его аналогом, которые присоединены к твердой подложке. Последовательность анализируемой нуклеиновой кислоты может включать последовательность ДНК, которая обеспечивает аффинное отделение анализируемой нуклеиновой кислоты от невключенного реагента и (или) от затравки за счет спаривания оснований с комплементарной последовательностью, имеющейся в составе нуклеиновой кислоты, присоединенной к твердой подложке. Анализируемая нуклеиновая кислота может быть помечена выявляемым маркером; этот выявляемый маркер может отличаться от любого выявляемого маркера, присутствующего в составе реагента или присоединенного к затравке.
В этом отношении термин "нормальный нуклеотид" определяется как дикий тип или ранее известный стандартный нуклеотид, мутация которого идентифицируется по положению нуклеотида. Под "стандартным нуклеотидом" понимается любой нуклеотид, который может являться диким типом или известным мутантным вариантом, лишь бы этот нуклеотид был известен и было бы желательно идентифицировать его вариант. Таким образом, например, нормальный нуклеотид может являться известным нуклеотидом дикого типа, для которого подразумевается мутация в этом положении. С другой стороны, известный нуклеотид может являться известным мутантным нуклеотидом, для которого присутствие нуклеотида дикого типа является
искомым по данному положению. Напротив, известный нормальный нуклеотид может являться известным мутантом, для которого искомым является иной мутантный вариант. Следовательно, способ по настоящему изобретению может быть применен в отношении любой известной последовательности, которая может быть использована для установления присутствия любого другого варианта нуклеотида в данном сайте.
По использованию в данном тексте термин "затравка" или "олигонуклеотидная затравка" обозначает олигонуклеотид, который способен выполнять роль инициирующей "точки" в синтезе в случае помещения его в такие условия, которые способствуют синтезу продукта достройки затравки, который комплементарен нуклеотидной цепи (матрице), в присутствие различных факторов, таких как, например, нуклеотиды и ферменты, такие как ДНК-полимераза, при подходящих величинах рН и температуры.
С другой стороны, термин "затравка" определяется как произвольный фрагмент нуклеиновой кислоты, получаемый из любого источника. Например, затравка может быть получена в результате фрагментации более крупных фрагментов нуклеиновых кислот, таких как геномная ДНК, кДНК или ДНК, которые были получены в методе ПЦР. Другими словами, природа затравки не ограничивается тем, как эта затравка была получена, т.е. была ли она получена путем фрагментации нативной или синтетически полученной нуклеиновой кислоты, или путем синтеза нуклеотидной затравки. Более того, затравка может быть олигодезоксирибонуклеотидом, сополимером олигодезоксирибонуклеотидов и олигорибонуклеотидов, сополимером рибонуклеотидов или сополимером дезоксирибонуклеотидов и рибонуклеотидов. Затравка может быть нативной или синтетической. Олигонуклеотидная затравка может быть синтезирована либо каталитически in vivo, либо каталитически in vitro, либо in vitro некаталитическим путем. Затравка может быть помечена выявляемым маркером: такой выявляемый маркер может отличаться от любого выявляемого маркера, присутствующего в составе реагента или присоединенного к анализируемой нуклеиновой кислоте. Кроме того, затравка должна включать последовательность, соответствующую фланкирующей последовательности по специфичному анализируемому сайту, соседнему по отягощению к идентифицируемому нуклеотиду и находящемуся выше его.
Кроме того, затравка должна быть способна гибридизовать или претерпевать отжиг с нуклеотидами, имеющимися в составе анализируемой нуклеиновой кислоты. Одним из путей осуществления желательной гибридизации является наличие определяемой матрицей затравки, которая в существенной степени комплементарна или полностью комплементарна известной нуклеотидной последовательности.
Олигонуклеотидная затравка может включать одну или несколько составляющих, которые обеспечивают аффинное отделение этой затравки от невключенного реагента и (или) от анализируемой нуклеиновой кислоты. Такими аффинными составляющими являются, тем самым не ограничиваясь, дигитонин, намагниченные шарики и лиганды, такие как белковые лиганды, включая антитела. Предпочтительно таким составлядщим является биотин. В случае использования биотина затравка, включающая биотин, обеспечивает аффинное отделение этой затравки от невключенного реагента и (или) от анализируемой нуклеиновой кислоты за счет связывания биотина с авидином и его аналогом, который присоединен к твердой подложке. Последовательность олигонуклеотидной затравки может включать последовательность ДНК, которая обеспечивает аффинное отделение данной затравки от невключенното реагента и (или) от анализируемой нуклеиновой кислоты за счет спаривания основания с комплементарной последовательностью, имеющейся в нуклеиновой кислоте, прикрепленной к твердой подложке.
По использованию в данном тексте термин "реакция достройки затравки" обозначает такие условия реакции, при которых осуществляется реакция направляемого матрицей синтеза нуклеиновой кислоты. Условия протекания реакции направляемой матрицей достройки затравки, в частности, могут быть созданы в присутствие подходящего зависимого от типа матрицы фермента. Некоторыми такими подходящими зависимыми от типа матрицы ферментами являются ДНК-полимеразы. ДНК-полимераза может представлять ряд типов. Однако ДНК-полимераза должна соответствовать типам затравки и матрицы. Например, могут быть использованы ДНК-полимераза E. coli или ее "фрагмент Кленова", ДНК-полимераза Т4, ДНК-полимераза Т7 ("секвеназа"), ДНК-полимераза T.aquaticus или ретровирусная обратная транскриптаза. В некоторых вариантах способа также могут быть использованы РНК-полимеразы, такие как РНК-полимераза фагов Т3 -или Т7. В зависимости от типа полимеразы должны использоваться различные условия, а для реакций гибридизации и достройки затравки могут требоваться различные температурные режимы.
По использованию в данном тексте термин "цепь, достроенная от затравки" определяет ту цепь, которая была образована "напротив" матрицы в составе дуплекса после добавления затравки. Предпочтительно достройку затравки терминируют путем связывания терминатора с матрицей.
По использованию в данном тексте термин "матрица" обозначает нуклеиновую кислоту, выключая двухцепочечную ДНК, одноцепочечную ДНК и РНК или их модификации, которая может иметь любуго длину или последовательность.
По использованию в данном тексте термин "терминатор", или "терминатор цепи", или "терминаторный нуклеотид" обозначает нуклеотид, такой как A, G, С, Т или U, или аналог, который эффективно терминирует реакцию достройки затравки тогда, когда он встраивается в цепь, достраиваемую от затравки, находящуюся "напротив" матричной цепи. Предпочтительно, терминатор является дидезоксинуклеотидом. Также предпочтительным является то, чтобы терминатор был либо вовсе не помечен, либо был помечен так, чтобы он отличался от метки нетерминаторного нуклеотида. По использованию в данном тексте термин "терминатор", или "терминатор цепи", или "терминаторный нуклеотид" обозначен в единственном числе, но это не означает, что используется одна молекула нуклеиновой кислоты. Единственное число термина "терминатор", скорее относится к тому типу нуклеотида или аналога нуклеотида, который используется в данном способе. Например, если терминатор является ddA (дидезоксиаденин), то все ddA в совокупности рассматриваются в единственном числе, а не каждая из молекула ddA по отдельности. С другой стороны, "терминатор" может соответствовать отсутствию конкретного типа нуклеотида, чтобы в результате достройка затравки останавливалась из-за отсутствия конкретного нуклеотида в данном локусе. Например, если желательным является, чтобы реакция достройки затравки останавливалась напротив нуклеотида С в составе матрицы, то не являющиеся терминаторными нуклеотиды А, Т и G должны быть включены в реакцию достройки затравки, в то время как являющийся комплементарным С нуклеотид G должен быть из нее изъят. Таким образом, отсутствие комплементарного нуклеотида должно обусловить терминацию реакции достройки затравки, что по результату сходно, например, с добавлением терминаторного дидезоксинуклеотида.
По использованию в данном тексте термин "нетерминирующий нуклеотид" или "нетерминатор" или "нецепочечный терминатора обозначает нуклеотид, который не обусловливает терминацию реакции достройки тогда, когда он включается в состав цепи, достраиваемой от затравки. Предпочтительно, по крайней мере, один нетерминаторный нуклеотид в реакции достройки цепи помечают. Также в данном тексте использование термина "нетерминирующий нуклеотид" или "нетерминатор" или "нецепочечный терминатор" в единственном числе не обозначает, что он соответствует единственному нуклеотиду. Более того, единственное число термина "нетерминирующий нуклеотид" обозначает тип нуклеотида или аналога нуклеотида, который используется в данном способе. Например, если терминатором является G, то все нуклеотиды G в совокупности обозначаются единственным числом, что не указывает на единственную молекулу G.
По использованию в данном тексте термин "мутант" или "мутация" обозначает любой нуклеотид в составе матричной цепи, который отличается от нуклеотида дикого типа или нормального нуклеотида. Мутация, которая может быть идентифицирована в соответствии со способом по настоящему изобретению, может являться мутацией любого типа, включая мононуклеотидную мутацию, вставку, делецию или генную транслокацию, лишь бы при этом нуклеотид в составе матрицы, непосредственно противостоящий нуклеотиду, находящемуся сразу за 3'-концом затравки, был изменен.
По использованию в данном тексте термин "метка" ("маркер") обозначает любую молекулу, которая присоединена к терминаторному или нетерминаторному нуклеотиду с целью получения выявляемого сигнала. Метка может быть радиоактивной, хемолюминесцентной, являться белковым лигандом, таким как антитело, или флуоресцентной группой, при том, что для каждого типа нетерминирующих нуклеотидов могут быть использованы различные флуоресцентные группы. Такие флуоресцентные группы должны обладать свойствами испускать спектры, которые различимы при спектроскопии.
С другой стороны, способ выявления уровня включения нуклеотида в достраиваемый от затравки продукт может быть применен на основании масс-спектрометрии в соответствии с описываемым в патенте США 5885775, который включен здесь для сведения в виде библиографической ссылки.
По использованию в данном тексте выражение "условия гибридизации высокой степени жесткости" обозначает такие условия гибридизации нуклеотидов, как, тем самым не ограничиваясь, условия промывки в 0,lxSSC при 42oС. Базовые характеристики условий гибридизации можно найти в руководстве по молекулярной биологии, таком как Ausubel et al., 1994, "Current Protocols in Molecular Biology", Greene & Wiley Publ., которое включено здесь для сведения в виде библиографической ссылки.
По использованию в данном тексте термин "тонкослойная хроматография (ТСХ)" может быть осуществлена на бумаге на основе целлюлозных продуктов, но может быть также выполнена на любом другом веществе, которое способствует точному разделению молекул с образованием однотипного слоя. Этими веществами являются, тем самым не исчерпываясь, неорганические соединения, такие как кремнезем, окись алюминия, "диатомейная земля" или силикат магния. Органическими соединениями являются, тем самым не ограничиваясь, целлюлоза, полиамид или порошок полиэтилена. Методы тонкослойной хроматографии в принципе описаны в руководствах по химии, таких как Freifelder, 1982, "Physical Biochemistry - Applications to Biochemistry and Molecular Biology", 2d ed., Freeman & Co., которое включено здесь для сведения в виде библиографической ссылки, особенно в части его главы 8, в которой рассматриваются хроматографические методы и, в частности, метод тонкослойной хроматографии, стр. 229-232.
Для специалиста в данной области техники должно быть ясно, что терминаторный нуклеотид может быть помечен меткой, отличающейся от таковой у нетерминаторного нуклеотида, что может быть затем использовано для различения встраивания терминаторного или нетерминаторного нуклеотида в состав цепи, достраиваемой от затравки. Пример терминатора, как "нуклеотидного пропуска", в настоящей заявке приведен только для целей проиллюстрировать ее, но такое иллюстративное описание не должно рассматриваться как в чем-либо ограничивающее заявляемое изобретение. По-разному помеченный или непомеченный терминатор также заявляется настоящим изобретением, лишь бы только метка такого терминатора отличалась от метки нетерминаторного нуклеотида.
Также для специалиста в данной области техники должно быть ясно, что при том, что последовательность матрицы известна, по крайней мере, частично, затравка может быть сконструирована таким образом, чтобы связываться с матрицей так, чтобы имело место связывание матрицы и затравки. Также для специалиста в данной области техники должно быть ясно, что способ по настоящему изобретению может быть осуществлен с использованием нескольких затравок, помещенных в одну или несколько тест-пробирок.
Свойством способа по настоящему изобретению является то, что в нем может быть генерирован мощный сигнал, если нетерминаторные нуклеотиды однотипно помечены, за счет чего обусловливается "аддитивный сигнальный эффект" после включения нескольких помеченных нетерминаторных нуклеотидов в состав достраиваемой от затравки цепи в случае, когда в заданном сайте имеется мутация. Это создает преимущественный по мощности сигнал по сравнению с традиционными методами выявления сигналов, при которых включается только одна метка в расчете на каждую достраиваемую цепь. Точность детекции выявляется тогда, когда сигналы считываются по различным меткам, специфичным в отношении различных терминаторов или нетерминаторных нуклеотидов.
Нижеследующие примеры призваны проиллюстрировать настоящее изобретение, но не в чем-либо его ограничить.
ПРИМЕРЫ
Пример 1
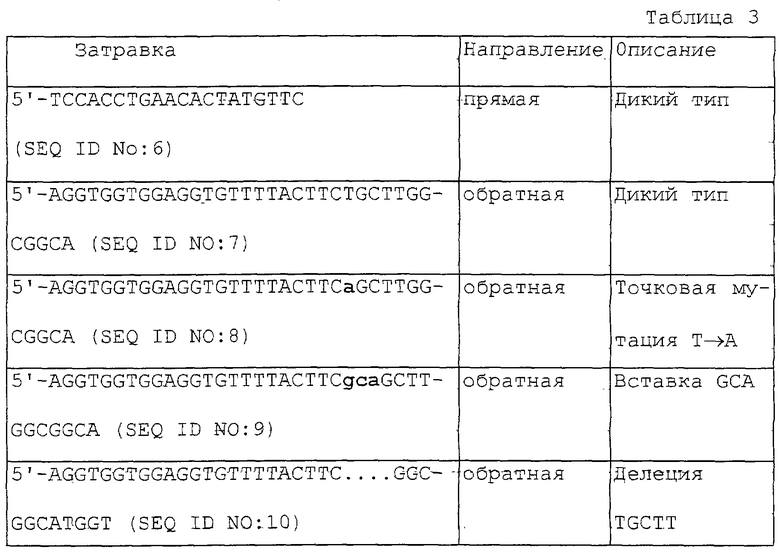
Последовательность гена АРС человека была выбрана в качестве последовательности-мишени для STA-теста (по способу терминации со сдвигом) по настоящему изобретению. Олигонуклеотиды, соответствующие нуклеотидам 4317-4347 последовательности гена АРС дикого типа и три различных мутантных варианта были синтезированы и использованы в качестве матриц. Использовавшиеся в STA-тесте затравки приведены в табл. 1.
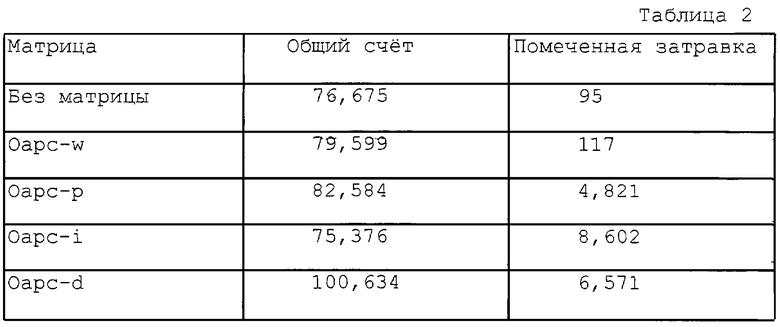
STA: каждую реакцию STA проводили в 20 мкл буфера (10 мМ Трис-HCl, рН= 7,5; 50 мМ КС1 и 5 мМ MgCl2), содержащем 50 нг олигонуклеотидной матрицы, 1 мкМ затравки, 2 ед. ДНК-полимеразы, 1 мкл [α-32P]-помеченного dCTP (250 мКи/мл, 3000 Ки/ммоль Dupont New England Nuclear), dATP, dTTP и 1 мкл непомеченного ddGTP. Реакционную смесь инкубировали при 37oС в течение 30 минут и нагревали до 100oС на 3 минуты. 1 мкл реакционной смеси наносили на бумажную полоску TI, предназначенную для осуществления тонкослойной хроматографии (TRIM USA, MD). Полоски выдерживали в течение 10 минут в растворе, содержащем 1 М НСl и 1 М NaCl. Затравки полностью отделяли от невключенного нуклеотида на полоске TI с помощью следующей процедуры. Помеченную затравку визуализовали авторадиографически и радиоактивность подсчитывали с помощью бекмановского счетчика сцинтилляций (Beckman, LS-5000). Авторадиограмма показана на фиг. 2, а результаты подсчета соответствующей авторадиограммы приведены в табл. 2.
В подсчетах, приведенных в табл. 2, правая колонка соответствует включению [α-32P]dCTP в достраиваемый от затравки олигонуклеотид дикого типа - Oapc-w, - играющий роль матрицы. Первым неспаренным нуклеотидом после отжига затравки является С, который по принципу комплементарности узнается терминаторным ddG. После того, как реакция направляемой матрицей достройки затравки началась, терминатор ddGB вскоре встроился с 3'-конца затравки в качестве первого достроечного нуклеотида, и тем самым дальнейшее включение помеченных нуклеотидов было заблокировано этим включенным ddG. В результата затравка оказалась достроена единственным нуклеотидом, который и оказался терминаторным нуклеотидом. Поскольку отсутствует возможность связывания с матрицей других нуклеотидов после присоединения терминатора, то реакция достройки затравки останавливается. Подсчет радиоактивности для образца Oapc-w показал, что ее величина была сходной с таковой, определенной для образца без какой-либо матрицы, т.е. для фонового контроля.
Для сравнения взята матрица Оарс-р, являющаяся олигонуклеотидом с точковой мутацией. Матрицу формировали путем замещения С последовательтности дикого типа на мутантный Т по первому неспаренному нуклеотиду с 3'-конца матрицы дикого типа. В этом случае dATP вместо терминатора ddG по принципу комплементарности замыкает мутантный нуклеотид Т после начала реакции достройки затравки. Реакция достройки затравки останавливается после того, как терминатор ddG встраивается в положение, находящееся "напротив" остатка С в матричной цепи.
В мутантном олигонуклеотиде, охватывающем мутацию типа вставки - Оарс-i, в соответствии с табл. 1 и 2, - первым неспаривающимся нуклеотидом является Т, который не комплементарен терминатору ddG. В этом случае затравку достраивают путем связывания dATP "напротив" этого Т в цепи, достраиваемой от затравки, и затем затравку далее удлиняют добавлением помеченного [α-32P]-dCTP, dATP, двух помеченных [α-32P]-dCTP и dATP. Последующее включение ddG с участием нуклеотидполимеразы на участке первого встретившегося С терминирует процесс достройки. Конечный итогом является то, что три нуклеотида [α-32P]-dCTP оказались включенными в цепь, достраиваемую от затравки.
Как и в случае с мутацией типа вставки, олигонуклеотид делеционной мутации Оарс-d (табл. 1 и 2) - был протестирован с использованием заявляемых реагентов и способа STA. Затравку достраивали включением нуклеотидов в следующем порядке: два [α-32P]-dCTP, dATP - и терминировали с ddG. Таким образом, в цепь, достраиваемую от затравки, включались два [α-32P]-dCTP. Эти данные дают отчетливое доказательство тому, что заявляемый способ STA может позволить выявлять все типы мутаций. В способе STA может быть идентифицировано присутствие мутации любого типа путем проведения единственного теста.
В частности, в случаях мутаций типов делеции и вставки (Oapc-i и Oapc-d) в достраиваемую от затравки цепь происходило встраивание множества помеченных нуклеотидов. Такое множественное мечение обусловливает резкое усиление чувствительности детекции. Кроме того, чувствительность способа может быть дополнительно повышена путем использования различных нуклеотидов, помеченных одним и тем же выявляемым маркером. Например, все нетерминаторные нуклеотиды могут быть помечены в достраиваемой затравке так: [α-32P]-СТР, [α-32P]-АТР и [α-32P]-TTP.
Неоднократное мечение также представляет возможность пометить нетерминаторные нуклеотиды с помощью разных выявляемых маркеров с целью распознавания каждого из нетерминаторных нуклеотидов. Например, нуклеотиды могут быть помечены различными флуоресцентными красителями, а затравка затем достраивается с включением разных флуоресцентных меток. Детекция различных сигналов в одно и то же время будет способствовать повышению точности теста STA. Такие преимущественные свойства неоднократного мечения, связанные с заявляемыми реагентом и способом STA, обеспечивают более высокий уровень чувствительности и точности по сравнению с методами, известными в данной области техники.
Пример 2
В качестве тестируемого образца использовали ПЦР-продукты гена АРС человека. Фрагмент гена АРС был амплифицирован с помощью ПЦР с использованием стандартного протокола ПЦР. В качестве матрицы исполъзовали кДНК гена АРС человека. Затравки, использовавшиеся для ПЦР, приведены в табл. 3.
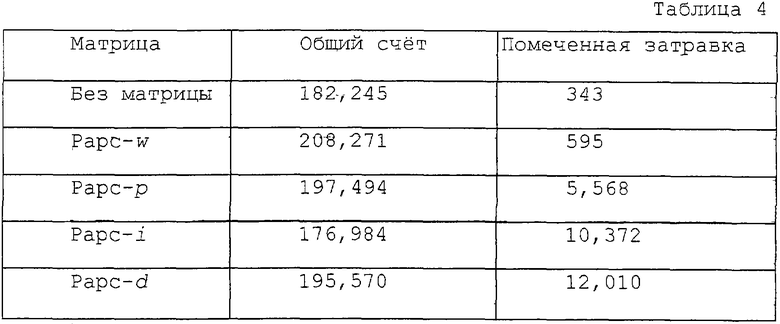
Четыре различных ПЦР-продукта размером примерно 200 пар нуклеотидов создавали путем сочетания затравок. Они представляют собой ATC-w: дикий тип; АРС-р: с включением точковой мутации; APC-i: с включением вставки и APC-d: с включением делеции. ПЦР-продукта вносили в 1%-ный агарозный гель с целью удаления матрицы и свободных нуклеотидов. Затем полученные продукты очищали с помощью реактивов Qiax DNA Purification kit (Qiagen). Затравку для STA конструируют как 5'-AGGTGGTGGAGGTGTTTTACTTC-3' (SEQ ID N0:11), - a реакции STA осуществляли при общем объеме 20 мкл в буфере, содержащем 10 мМ Трис-НСl (рН= 8,3), 50 мМ КС1, 2 мМ MgCl2, 0,05 пкМ двухцепочечного ПЦР-продукта, 5 пкМ затравок, по 20 мкМ dATP, dGTP, 1 мкКи [α-32P] помеченного dCTP, 20 мкМ непомеченного ddTTР и 2 ед. ДНК-полимеразы Taq. Двадцать циклов проводили в амплификаторе (Percin Elmer, Gene-Amp-9600) при таком режиме: 94oС в течение 20 секунд, 55oС в течение 1 минуты. 1 мкл STA-продукта наносили на бумажную полоску Trim (полоска для тонкослойной хроматографии, выпускаемая в Японии фирмой TRIM Corp. ), а радиоактивность подсчитывали в соответствии с описанным в примере 1. Полученные результаты показаны в табл. 4.
Затравка для образцов со всеми тремя типами мутаций достраивалась с включением [α-32P]-dCTP. Интенсивность сигнала, генерируемого достроенной затравкой после включения метки, достоверно коррелирует с количеством помеченных нуклеотидов, которые находятся в составе цепи, достроенной с затравки.
Пример 3
Заявляемые реагент и способ STA были применены в отношении РНК-фрагментов гена АРС человека. ПЦР-продукты гена АРС человека, описанные в примере 2, лигировали в составе клонирующего вектора ТА-3.1 - ТА Cloning kit (Invitrogen). Были сконструированы четыре вектора: они охарактеризованы в табл. 5.
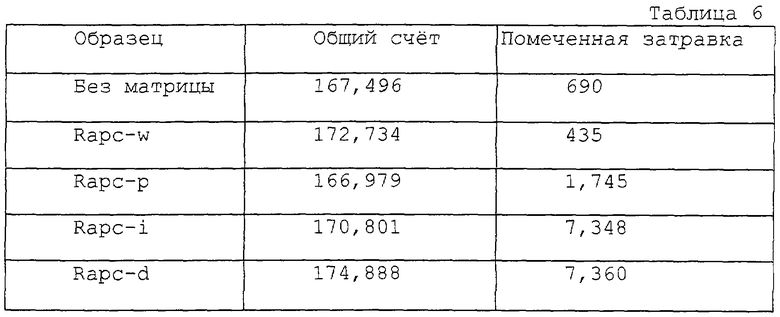
Варианты РНК, соответствующие каждому вектору, были синтезированы с использованием набора реактивов для синтеза РНК in vitro - in vitro RNA Synthesis kit (Promega, WI). РНК синтезировали при 37oС в течение 1 часа в буфере, содержащем 2 мкг вектора и полимеразы Т7. Реакцию останавливали добавлением хлорида лития и 100%-ного этанола. После инкубации при -20oС в течение 15 минут РНК осаждали в центрифуге при 14000 g в течение 15 минут и очищенную РНК ресуспендировали в воде, свободной от РНКазы. По 5 мкг РНК смешивали с SТА-затравкой, описанной в примере 2, при общем объеме 10 мкл буфера, содержащего 10 мМ Трис-НСl (рН=7,6), 50 мМ NaCl и 10 мМ КСl. Смесь денатурировали нагреванием до 65oС в течение 3 минут с последующей остановкой реакции льдом в течение 2 минут. Реакцию STA проводили в соответствии с описанным в примере 1 в буфере, содержащем 1 мкл [α-32P]-помеченного dCTP (250 мКи/мл, 3000 Ки/ммоль: Dupont New England Nuclear), 10 мкМ dATP, dGTP и 10 мкМ непомеченного ddTTP, a также 20 ед. обратной транскриптазы. После инкубации при 40oС в течение 15 минут реакцию останавливали нагреванием до 100oС на 2 минуты. По 1 мкл продукта реакции наносили на полоску Trim и радиоактивность подсчитывали в соответствии с описанным в примере 1. Полученные результаты показаны в табл. 6.
На всех описанных выше стадиях включались химические реакции, манипуляции и протоколы, которые были автоматизированы или могли бы быть автоматизированы. Следовательно, внесение предпочтительного способа реализации настоящего изобретения в операционную систему, подходящим образом запрограммированную для робототехнического устройства, приведет в результате к существенной экономии средств и повышению эффективности предположительно всей диагностической процедуры, которая основана на детекции конкретных нуклеотидных последовательностей или различий последовательностей в нуклеиновых кислотах, производных от биологических образцов.
Все цитаты в данном тексте включены здесь в своем полном объеме в виде библиографических ссылок.



Изобретение относится к молекулярной биологии и может быть использовано для выявления любой мутации в заданном сайте, известной последовательности нуклеиновой кислоты. В основе способа лежит принцип достройки затравки, комплементарной последовательности анализируемого образца, непосредственно примыкающей к нуклеотиду - мишени. При этом в реакционную смесь для достройки затравки вводят терминаторный нуклеотид того типа, который комплементарен нуклеотиду мишени в составе матрицы (указанный тип нуклеотида также может отсутствовать в реакции), и нуклеотиды трех других типов, которые не являются терминаторными и несут выявляемую метку. Прекращение реакции вследствие включения терминаторного нуклеотида свидетельствует об отсутствии мутации в исследуемом сайте; продолжение наращивания цепи за счет меченых нетерминаторных нуклеотидов - о ее наличии. Способ обеспечивает возможность простого, быстрого и экономичного определения мутаций любого типа. 35 з.п. ф-лы, 4 ил., 6 табл.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Мост для дистанционного измерения комплексных сопротивлений | 1972 |
|
SU497527A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Виброгрохот инерционный | 1976 |
|
SU663447A1 |

