Область техники, к которой относится изобретение
Настоящее изобретение относится к термостабильным ДНК-полимеразам, способным более эффективно включать рибонуклеозидтрифосфаты. В изобретении описаны методы и способы выделения таких полимераз. Ферменты по изобретению могут применяться для различных целей, в частности для секвенирования нуклеиновых кислот. Таким образом, изобретение также относится к улучшенным методам анализа последовательности нуклеиновых кислот.
Предпосылки создания изобретения
Секвенирование ДНК обычно включает получение четырех популяций фрагментов одноцепочечной ДНК, имеющих одно определенное окончание и одно вариабельное окончание. Вариабельное окончание обычно заканчивается на конкретных основаниях нуклеотидов [на гуанине (Г), аденине (А), тимидине (Т) либо на цитозине (Ц)]. Каждый из четырех различных наборов фрагментов разделяют на основе их длины. С этой целью используют полиакриламидный гель с высокой разрешающей способностью. Каждая полоса на таких гелях соответствует конкретному нуклеотиду в последовательности ДНК, показывая тем самым его положение в последовательности.
Часто применяемым методом секвенирования ДНК является дидезокси- или цепь-терминирующий метод секвенирования, который включает ферментативный синтез цепи ДНК (Sanger и др., 1977, Ргос. Natl. Acad. Sci. 74: 5463). Обычно осуществляют четыре различные реакции синтеза, каждая из которых должна закончиться на определенном основании (Г, А, Т или Ц) путем включения соответствующего терминирующего цепь нуклеотида, такого как дидезоксинуклеотид. Продукты реакции легко определить, поскольку каждая полоса соответствует только Г, А, Т либо Ц.
В дидезокси- или цепь-терминирующем методе отжигают короткий одноцепочечный праймер, соответствующий одноцепочечной матрице. Праймер удлиняют на его 3’-конце путем включения дезоксинуклеотидов (дНТФ) до включения дидезоксинуклеотида (ддНТФ). После включения ддНТФ удлинение прекращается. Однако для гарантии правильности репликации ДНК ДНК-полимеразы обладают более выраженной способностью включать естественные для них субстраты, т.е. дНТФ, и не включать аналоги нуклеотидов, называемые несвойственными нуклеотидами. При синтезе ДНК рибонуклеотиды (рНТФ) рассматриваются как несвойственные нуклеотиды, поскольку в отличие от ддНТФ рНТФ обычно не являются естественными субстратами для ДНК-полимеразы in vivo. В клетке это свойство способствует снижению включения аномальных оснований, таких как дезоксиинозинтрифосфат (дИТФ) или рНТФ, в растущую цепь ДНК.
Двумя наиболее часто используемыми методами автоматического секвенирования являются секвенирование с использованием окрашенного праймера и окрашенного терминатора. Эти методы пригодны для использования с флуоресцентно меченными фрагментами. Хотя секвенирование также может быть осуществлено с использованием радиоактивно меченных фрагментов, секвенирование, основанное на флуоресценции, является более предпочтительным. В целом при секвенировании с окрашенным праймером флуоресцентно меченный праймер применяют в комбинации с немечеными ддНТФ. Процесс осуществляется с использованием четырех реакций синтеза и до получения на геле четырех полос для каждой подлежащей секвенированию матрицы (соответствующих продукту терминации, который специфичен для каждого основания). После удлинения праймера подлежащие секвенированию смеси, содержащие продукты терминации с включенными дидезоксинуклеотидами, обычно анализируют с использованием электрофореза на геле для секвенирования ДНК. После электрофоретического разделения флуоресцентно меченные продукты вырезают с помощью лазера из основания геля и флуоресценцию определяют с использованием соответствующего монитора. В автоматических системах при прохождении реакционных смесей через матрикс геля во время электрофореза детектор сканирует основание геля для обнаружения, какой из меченых фрагментов используется (Smith и др., 1986, Nature 321: 674-679). В модификации этого метода каждый из четырех праймеров метят различным флуоресцентным маркером. По завершении четырех различных реакций секвенирования реакционные смеси объединяют и объединенные реакционные смеси подвергают гель-анализу для каждой полосы, посредством чего индивидуально выявляют различные флуоресцентные метки (соответствующие четырем различным продуктам терминации, которые специфичны для каждого основания).
В другом варианте применяют метод секвенирования с использованием окрашенного терминатора. В этом методе для включения дНТФ применяют ДНК-полимеразу и флуоресцентно меченные ддНТФ на наращиваемом конце ДНК-праймера (Lee и др., 1992, Nucleic Acid Research 20: 2471). Преимуществом этого способа является отсутствие необходимости синтезировать меченные красителем праймеры. Кроме того, реакции с использованием окрашенного терминатора являются более удобными, поскольку при этом все четыре реакции могут быть осуществлены в одной и той же пробирке. Ранее были описаны модифицированные термостабильные ДНК-полимеразы, обладающие пониженной способностью различать ддНТФ (см. публикацию европейской заявки ЕР-А-655506 и заявку на патент США 08/448223). Примером модифицированной термостабильной ДНК-полимеразы является мутантная форма ДНК-полимеразы из Т. aquaticus, имеющая остаток тирозина в положении 667 (вместо остатка фенилаланина), т.е. так называемая F667Y-мутантная форма ДНК-полимеразы Taq. AmpliTaq® FS, которая производится фирмой Hoffmann-La Roche и продается через фирму Perkin Elmer, снижает количество ддНТФ, необходимое для эффективного секвенирования нуклеиновой кислоты-мишени, в несколько сотен или несколько тысяч раз. AmpliTaq® FS представляет собой мутантную форму ДНК-полимеразы из Т. aquaticus, имеющую мутацию F667Y и дополнительный остаток аспарагиновой кислоты в положении 46 (вместо остатка глицина; G46D- мутация).
Таким образом, существует необходимость в термостабильных ДНК-полимеразах, применяемых для альтернативных методов синтеза нуклеиновых кислот, пригодных для точного и эффективного с точки зрения стоимости анализа нуклеотидной последовательности в ДНК. Также существует необходимость в разработке методов, основанных на флуоресценции, для которых не требуется применение дидезоксинуклеотидов. Настоящее изобретение направлено на решение этих задач.
Краткое изложение сущности изобретения
Настоящее изобретение относится к зависимым от матрицы термостабильным ДНК-полимеразам, которые включают характерную аминокислотную последовательность-мотив SerGlnIleXaaLeuArgXaa (SEQ ID NO: 1), причем "Хаа" в положении 4 этой последовательности представляет собой остаток любой аминокислоты, кроме остатка глутаминовой кислоты (Glu), а "Хаа" в положении 7 этой последовательности представляет собой остаток валина (Val) или остаток изолейцина (Ilе). При выражении в однобуквенном коде аминокислот эта последовательность-мотив может быть обозначена как S Q I X L R V/I, где "X" в положении 4 этой последовательности представляет собой остаток любой аминокислоты, кроме остатка глутаминовой кислоты. Способность термостабильных ДНК-полимераз, имеющих аминокислотную последовательность, включающую указанную последовательность-мотив, где "X" в положении 4 этой последовательности-мотива не является остатком глутаминовой кислоты, ограничивать включение рибонуклеотидов по сравнению с ранее описанными термостабильными полимеразами понижена. Для растущей цепи ДНК рибонуклеотиды являются несвойственными нуклеотидами. Таким образом, первым предметом изобретения являются новые ферменты, способные включать несвойственные аналоги оснований, такие как рибонуклеотиды, в растущую цепь ДНК и на несколько порядков более эффективные, чем выявленные ранее термостабильные ДНК-синтезирующие ферменты. Гены, кодирующие эти ферменты, также относятся к настоящему изобретению, равно как и рекомбинантные векторы экспрессии и клетки-хозяева, включающие эти векторы. С помощью таких трансформированных клеток-хозяев могут быть получены большие количества очищенных термостабильных ферментов полимераз.
В соответствии с настоящим изобретеним была выявлена область или последовательность-мотив внутри аминокислотной последовательности термостабильных ДНК-полимераз, увеличивающая способность полимераз включать рибонуклеотиды, сохраняя при этом способность правильно включать дезоксирибонуклеотиды. Изменения в этой области, например замена одной или нескольких аминокислот (например, введение их путем сайтнаправленного мутагенеза), приводят к получению термостабильного фермента полимеразы, который способен синтезировать РНК или РНК/ДНК-химеры либо гибридную цепь на матрице ДНК.
Другим предметом изобретения являются усовершенствованные методы и композиции, предназначенные для определения последовательности нуклеиновой кислоты-мишени, при этом необходимость в терминирующих цепь ддНТФ отпадает. С помощью представленных в настоящем описании усовершенствованных методов рибонуклеотиды (рНТФ) включаются в продукты удлинения праймеров. Поскольку ферменты, являющиеся предметом изобретения, точно и эффективно включают как рНТФ, так и дНТФ, в реакциях секвенирования могут использоваться смеси обоих нуклеотидов. После удлинения праймера вновь синтезированные олигонуклеотидные продукты могут быть расщеплены по местам включения рНТФ методами, известными в данной области техники, например путем гидролиза, что приводит тем самым к получению популяции фрагментов, пригодных для фракционирования и анализа последовательности стандарными методами, такими как гель-электрофорез. Эти методы, основанные на применении новых термостабильных ферментов полимераз, представлены в данном описании. Таким образом, этот предмет изобретения относится к термостабильным ДНК-полимеразам, которые отличаются тем, что полимераза включает критический мотив SerGlnIleXaaLeuArgXaa (SEQ ID NO: 1), где "Хаа" в положении 4 может представлять собой остаток любой аминокислоты, кроме остатка глутаминовой кислоты (Glu), а "Хаа" в положении 7 представляет собой остаток валина (Val) или остаток изолейцина (Ilе).
Другим предметом изобретения являются модифицированные полимеразы, представленные в настоящем описании, которые включают рибонуклеотиды или аналоги, содержащие гидроксильную группу или другой заместитель в положении 2’, которые обычно отсутствуют у дезоксирибонуклеотидов. Эти нуклеотиды могут быть по-разному помечены, что создает альтернативные варианты обычному применению дидезоксинуклеотидов для целей секвенирования ДНК.
Мутантные термостабильные полимеразы по изобретению способны более эффективно включать несвойственные нуклеотиды, в частности рибонуклеотиды, по сравнению с соответствующими ферментами дикого типа. В предпочтительном варианте осуществления изобретения подлежащий включению несвойственный нуклеотид может быть аналогом терминирующего цепь основания, таким как 2’-гидрокси-3’-дезоксиАТФ (кордицепинтрифосфат), являющийся "риботерминатором"-аналогом АТФ, или нетерминирующим цепь нуклеотидом, таким как рНТФ.
Другим предметом изобретения явялются мутантные термостабильные полимеразы, которые способны более эффективно включать несвойственные нуклеотиды, в частности рибонуклеотиды, чем соответствующие ферменты дикого типа. Таким образом, этот предмет изобретения относится к рекомбинатным термостабильным ДНК-полимеразам, каждая из которых характеризуется тем, что (а) в своей нативной форме полимераза включает аминокислотную последовательность SerGlnIleGluLeuArgXaa (SEQ ID NO: 2), где "Хаа" в положении 7 этой последовательности обозначает остаток валина (Val) или остаток изолейцина (Ilе); (б) в аминокислотной последовательности рекомбинантного фермента, предпочтительно в положении 4 этой последовательности, получают мутацию, при которой остаток глутаминовой кислоты в положении 4 представляет собой остаток другой аминокислоты, предпочтительно остаток глицина; и (в) способность рекомбинантного фермента ограничивать включение рибонуклеотидов и аналогов рибонуклеотидов по сравнению с нативной формой этого фермента понижена.
Другим предметом изобретения являются полимеразы, которые предназначены для фрагментации продуктов амплификации и продуктов удлинения праймера, причем такие фрагментированные продукты могут быть пригодны для использования в методах, основанных на гибридизации, и для различных стратегий определения последовательности.
Ферменты по настоящему изобретению и кодирующие их гены могут применяться для создания композиций, пригодных для осуществления реакций секвенирования ДНК и включающих смесь обычных нуклеотидов и по крайней мере один рибонуклеотид или аналог рибонуклеотида. В предпочтительном варианте осуществления изобретения несвойственный нуклеотид представляет собой рибонуклетид, а концентрация рибонуклеотида ниже концентрации соответствующего дезоксирибонуклеотида, т.е. отношение рНТФ:дНТФ составляет 1:1 или менее. Ферменты по изобретению также пригодны для продажи в виде наборов, которые также могут включать любой из дополнительных элементов, необходимых для осуществления реакции секвенирования нуклеиновой кислоты, такой как, например, дНТФ, рНТФ, буферы и/или праймеры.
Подробное описание изобретения
Настоящее изобретение относится к новым и улучшенным модифицированным термостабильным ДНК-полимеразам, композициям и наборам, охарактеризованным в формуле изобретения. Ферменты по изобретению более эффективно включают несвойственные нуклеозидтрифосфаты в сравнении с ранее известными полимеразами или соответствующими полимеразами дикого типа, из которых получают эти новые полимеразы. Согласно изобретению также предлагаются последовательности ДНК, кодирующие эти модифицированные ферменты, векторы для экспрессии модифицированных ферментов и клетки с такими внедренными в них векторами. Ферменты по изобретению пригодны для практического применения в новых методах секвенирования ДНК, которые обладают рядом преимуществ по сравнению с методами секвенирования ДНК, известными из уровня техники.
Ниже для облегчения понимания сущности настоящего изобретения определены некоторые понятия.
Понятие "обычный", если оно упоминается в связи с основаниями нуклеиновой кислоты, нуклеозидтрифосфатами или нуклеотидами, относится к таковым, которые встречаются в естественных условиях в указанном полинуклеотиде (например, для ДНК это дАТФ, дГТФ, дЦТФ и дТТФ). Кроме того, в реакциях синтеза ДНК in vitro, таких как секвенирование, вместо дЦТФ часто используют с7дГТФ и дИТФ (хотя они включаются с меньшей эффективностью). В целом они могут быть обобщены понятием дезоксирибонуклеозидтрифосфаты (дНТФ).
Понятие "система экспрессии" относится к последователностям ДНК, содержащим требуемую кодирующую последовательность и функционально связанные контролирующие последовательности, так что хозяева, трансформированные этими последовательностями, обладают способностью продуцировать кодируемые протеины. Для осуществлениия трансформации система экспрессии может быть включена в вектор, хотя пригодная ДНК также может быть включена в хромосому хозяина.
Понятие "ген" относится к последовательности ДНК, которая включает контролирующие и кодирующие последовательности, необходимые для воспроизводимого получения биологически активного полипептида или предшественника. Полипептид может кодироваться полноразмерной последовательностью гена или любой имеющей достаточную длину частью кодирующей последовательности, чтобы сохранить ферментативную активность.
Понятие "клетка(и)-хозяин(ева)" относится как к одноклеточным прокариотическим или эукариотическим организмам, таким как бактерии, дрожжи и актиномицеты, так и к отдельным клеткам из растений или животных высших отрядов, если они способны расти в культуре клеток.
Используемое в настоящем описании понятие "реакционная смесь для секвенирования ДНК" относится к реакционной смеси, которая включает элементы, необходимые для реакции секвенирования ДНК. Таким образом, реакционная смесь для секвенирования ДНК пригодна для использования в методе секвенирования ДНК для определения последовательности нуклеиновой кислоты-мишени, хотя вначале реакционная смесь может быть неполной, чтобы пользователь мог контролировать инициацию реакции секвенирования. В этом случае реакция может быть начата после того, как будет добавлен последний элемент, такой как фермент, который вводится для получения полной реакционной смеси для секвенирования ДНК. Обычно реакционная смесь для секвенирования ДНК должна содержать буфер, пригодный для полимеризации, нуклеозидтрифосфаты и по крайней мере один несвойственный нуклеотид. Реакционная смесь также может содержать праймер, который может удлиняться на мишени полимеразой, полимеразу и нуклеиновую кислоту-мишень. Праймер, равно как и один из нуклеотидов, обычно включает содержащий метку, например, флуоресцентную фрагмент, который может быть обнаружен. Обычно реакционнная смесь представляет собой смесь, которая включает четыре обычных нуклеотида и по крайней мере один несвойственный нуклеотид. В предпочтительном варианте осуществления полимераза представляет собой термостабильную ДНК-полимеразу, а несвойственный нуклеотид представляет собой рибонуклеотид.
Понятие "олигонуклеотид", используемое в данном описании, относится к молекуле, состоящей из двух или нескольких дезоксирибонуклеотидов или рибонуклеотидов, предпочтительно более чем из трех и обычно более чем из десяти. Точный размер олигонуклеотида может зависеть от многих факторов, включая основную функцию или применение олигонуклеотида.
Олигонуклеотиды могут быть получены любым пригодным методом, включая, например, клонирование и рестрикцию соответствующих последовательностей, и методом прямого химического синтеза, таким как фосфотриэфирный метод Narang и др., 1979, Meth. Enzymol. 68: 90-99; фосфодиэфирный метод Brown и др., 1979, Meth. Enzymol. 68: 109-151; диэтилфосфорамидный метод Beaucage и др., 1981, Tetrahedron Lett. 22: 1859-1862; триэфирный метод Matteucci и др., 1981, J. Am. Chem. Soc. 103: 3185-3191; автоматическими методами синтеза или методом с использованием твердой подложки, описанным в патенте США 4458066.
Понятие "праймер", используемое в данном описании, относится к естественному или синтетическому олигонуклеотиду, который способен служить точкой инициации синтеза при создании условий, при которых начинается удлинение праймера. Праймер предпочтительно представляет собой одноцепочечный олигодезоксирибонуклеотид. Соответствующая длина праймера зависит от предназначения праймера, но обычно составляет от 15 до 35 нуклеотидов. Для коротких молекул праймера обычно необходимы более низкие температуры для образования достаточно стабильных гибридных комплексов с матрицей. Праймер не должен отражать точную последовательность матрицы, но для того, чтобы происходило удлинение праймера, должен обладать достаточной степенью комплементарности для гибридизации с матрицей.
При необходимости праймер может быть помечен путем включения метки, которая может быть обнаружена спектроскопическими, фотохимическими, биохимическими, иммунохимическими или химическими способами. Например, пригодные метки включают 32Р, флуоресцентные красители, электронноплотные реагенты, ферменты (такие, как обычно применяемые в методах ELISA), биотин или гаптены и протеины, для которых имеются антисыворотки или моноклональные антитела.
Понятие "термостабильная полимераза" относится к ферменту, который стабилен при нагревании и который устойчив к нагреванию и сохраняет достаточную активность для последующиего осуществления реакции удлинения праймера при воздействии повышенных температур в течение времени, необходимого для осуществления денатурации двухцепочечных нуклеиновых кислот. Согласно данному описанию термостабильная полимераза пригодна для использования в реакции с температурными циклами, такой как полимеразная цепная реакция (ПЦР). Для термостабильной полимеразы под ферментативной активностью понимают способность соответствующим образом катализировать присоединение нуклеотидов для формирования продуктов удлинения праймера, которые комплементарны к цепи нуклеиновой кислоты-матрицы.
Необходимые для денатурации нуклеиновой кислоты условия нагревания могут зависеть, например, от концентрации соли в буфере, а также состава и длины нуклеиновых кислот, подлежащих денатурации, но обычно они находятся в интервале от приблизительно 90°С до приблизительно 105°С, предпочтительно от 90°С до 100°С, а продолжительнось нагревания в основном зависит от температуры и длины нуклеиновой кислоты, и обычно она составляет от нескольких секунд до четырех минут.
Понятие "несвойственный" или "модифицированный", когда оно характеризует основание нуклеиновой кислоты, нуклеозидтрифосфат или нуклеотид, включает модификацию, производные или аналоги обычных оснований или нуклеотидов, которые встречаются в ДНК в естественных условиях. В частности, согласно данному описанию несвойственные нуклеотиды модифицированы в положении 2’ сахара рибозы по сравнению с обычными дНТФ. Таким образом, хотя рибонуклеотиды (т.е. АТФ, ГТФ, ЦТФ, УТФ, называемые в целом рНТФ) и явялются встречающимися в естественных условиях нуклеотидами РНК, в контексте данного описания эти рибонуклеотиды являются несвойственными нуклеотидами, поскольку имеют в положении 2’ сахара гидроксильную группу, которая отсутствует в дНТФ. Аналоги рибонуклеотидов, содержащие заместители в положении 2’, такие как 2’-фтор-, 2’-аминозамещенные аналоги, подпадают под объем изобретения. Кроме того, аналоги рибонуклетидов могут быть модифицированы в положении 3’, например, путем замещения нормальной гидроксильной группы на водородную группу (3’-дезокси), что приводит к получению аналога-терминатора рибонуклеотида. Такие нуклеотиды также включены в объем понятия "несвойственные нуклеотиды".
Поскольку ДНК обычно состоит из дНТФ, включение рНТФ должно рассматриваться как несвойственное, и, следовательно, рНТФ должен рассматриваться как несвойственное основание. Таким образом, в предпочтительном варианте изобретения при осуществлении методов удлинения ДНК-праймера, включая методы секвенирования ДНК, в качестве продуктов получают нуклеиновые кислоты, которые содержат как обычные, так и несвойственные нуклеотиды, но в основном они содержат обычные нуклеотиды, которые представляют собой дНТФ.
Несвойственные основания могут быть флуоресцентно меченными, например, флуоресцином или родамином; нефлуоресцентно меченными, например, биотином; меченными изотопами, например, 32Р, 33Р или 35S или немечеными.
Для пояснения сущности изобретения в описании приведены примеры специфических термостабильных ферментов ДНК-полимераз по изобретению, однако эти ссылки не должны рассматриваться как ограничивающие объем изобретения. В предпочтительном варианте термостабильные ферменты по изобретению применяли в различных методах секвенирования нуклеиновых кислот, хотя новые термостабильные полимеразы, приведенные в настоящем описании, могут применяться для любой цели, где такая ферментативная активность является необходимой или желательной. Фермент также может применяться в реакциях амплификации, таких как ПЦР.
Термостабильные полимеразы по изобретению отличаются тем, что каждая из них содержит критический мотив SerGlnIleXaaLeuArgXaa (SEQ ID NO: 1), где "Хаа" в положении 4 этой последовательности представляет собой остаток любой аминокислоты, кроме остатка глутаминовой кислоты (Glu), а "Хаа" в положении 7 этой последовательности представляет собой остаток валина (Val) или остаток изолейцина (Ilе). Гены, кодирующие термостабильные полимеразы, которые имеют остаток глутаминовой кислоты в положении 4 этого мотива, могут быть модифицированы согласно настоящему описанию с получением пригодных модифицированных полимераз. Эти модифицированные термостабильные полимеразы отличаются тем, что по сравнению с соответствующими нативными ферментами или ферментами дикого типа они имеют модификацию в аминокислотной последовательности-мотиве SerGlnIleGluLeuArgXaa (SEQ ID NO: 2), где "Хаа" в положении 7 этой последовательности обозначает остаток валина (Val) или остаток изолейцина (Ilе); т.е. этот мотив был модифицирован путем замещения остатка глутаминовой кислоты в положении 4 остатком другой аминокислоты. Критический мотив термостабильной ДНК-полимеразы по настоящему изобретению приведен ниже с использованием стандартного однобуквенного кода аминокислот (Lehninger, Biochemistry, New York, Worth Publishers Inc., 1970, стр.67).
SEQ ID NO: 1 SerGlnIleXaaLeuArgXaa, где "Хаа" в положении 4 представляет собой остаток любой аминокислоты, кроме остатка глутаминовой кислоты (Glu), а "Хаа" в положении 7 представляет собой остаток валина (Val) или остаток изолейцина (Ilе).
Кодирующие последовательности генов, равно как и протеины, содержащие эту критическую аминокислотную последовательность, где Хаа в положении 4 не является остатком глутаминовой кислоты (Glu), относятся к полимеразе, способность которой ограничивать включение рНТФ понижена, и включены в объем настоящего изобретения. Внутри критического мотива могут быть осуществлены дополнительные модификации остатков других аминокислот, предпочтительно остатков аминокислот, выбранных из группы, включающей глутамин (Gln или Q), лейцин (Leu или L) или аргинин (Arg или R).
Настоящее изобретение может использоваться для получения термостабильных ДНК-полимераз с улучшенными свойствами путем определенной модификации в последовательности гена, кодирующего термостабильную ДНК-полимеразу. В предпочтительном варианте изобретения последовательность гена и кодируемый фермент происходят из видов рода Thermus, хотя эубактерии, не относящиеся к роду Thermus, также включены в настоящее изобретение, как это более подробно описано ниже. Аналогично этому, ввиду высококонсервативной природы выявленного в настоящем исследовании критического мотива, новые термостабильные ДНК-полимеразы могут быть обнаружены на основе их гомологии, например, с Taq-полимеразой. Такие термостабильные полимеразы включены в объем настоящего изобретения при условии, что их аминокислотная последовательность включает мотив S Q I Х L R V/I, где Х обозначает остаток любой аминокислоты, кроме остатка глутаминовой кислоты, и эта аминокислотная последовательность в целом гомологична (идентичность последовательности) по крайней мере приблизительно на 39%, предпочтительно по крайней мере приблизительно на 60%, более предпочтительно по крайней мере приблизительно на 80% аминокислотной последовательности нативной Taq-полимеразы. Вся длина последовательности этой Taq-полимеразы приведена в WO 89/06691 и зарегистрирована под регистрационным номером Р90556 в базе данных запатентованных последовательностей GENESEQ или под регистрационным номером М26480 в базе данных последовательностей EMBL и под регистрационным номером А33530 в базе данных последовательностей PIR.
Примером термостабильных ДНК-полимераз по настоящему изобретению являются рекомбинантные производные нативных полимераз из организмов, приведенных в таблице 1. В таблице 1 приведена конкретная последовательность критического мотива и положение остатка "X" для каждой из этих нативных полимераз. Поскольку каждая термостабильная ДНК-полимераза является уникальной, положение аминокислот критического мотива является определенным для каждого фермента. Для перечисленных ниже полимераз аминокислотный остаток в положении "X" критического мотива S Q I Х L R V/I представляет собой глутаминовую кислоту. Молекулярная масса предпочтительных полимераз по настоящему изобретению составляет от 85000 до 105000 Да, более предпочтительно от 90000 до 95000 Да. Аминокислотная последовательность этих полимераз состоит из приблизительно 750-950 остатков аминокислот, предпочтительно из 800-850 остатков аминокислот. Полимеразы по настоящему изобретению могут состоять из приблизительно 540 или более аминокислот и включают, по крайней мере, полимеразный домен и часть, соответствующую 3’-5’-экзонуклеазному домену (полученная полимераза может обладать или не обладать 3’-5’-экзонуклеазной активностью), и возможно части 5’-3’-экзонуклеазного домена, который расположен на первой трети аминокислотной последовательности многих полноразмерных термостабильных полимераз.
Для термостабильных ДНК-полимераз, не приведенных в таблице 1, выбор соответствующей глутаминовой кислоты, подлежащей модификации, является простым, если определен критический мотив или консенсусный мотив в аминокислотной последовательности.
Независимо от точного положения внутри термостабильной ДНК-полимеразы, замещение остатка глутаминовой кислоты (Glu) на остаток другой аминокислоты внутри последовательности-мотива SerGlnIleGluLeuArgXaa (SEQ ID NO: 2), где "Хаа" в положении 7 этой последовательности обозначает остаток валина (Val) или остаток изолейцина (Ilе), в полимеразном домене позволяет получить термостабильные полимеразы, способные к эффективному включению несвойственных нуклеотидов. В предпочтительном варианте глутаминовую кислоту заменяют на аминокислоту, имеющую ненагруженную полярную R-группу, такую как глицин, серин, цистеин, треонин, или на аминокислоту, имеющую небольшую неполярную R-группу, такую как, например, аланин. В наиболее предпочтительном варианте остаток глутаминовой кислоты заменяют на остаток глицина (G). Программы, позволяющие установить аминокислотную и нуклеотидную последовательность, могут быть приобретены у фирмы Genetics Computer Group, 575 Science Drive, Медисон, шт. Висконсин. Программы, пригодные для конкретного, определенного в данном описании мотива, включают, например, "GAP", "BESTFIT" и "PILEUP", способствующие выявлению точной последовательности подлежащей модификации области.
Как видно из приведенной таблицы 1, существуют две необходимые формы консервативной последовательности-мотива SerGlnIleGluLeuArgXaa (SEQ ID NO: 2) внутри полимеразного домена термостабильных ДНК-полимераз этих термофильных организмов. Последовательность-мотив SerGlnIleGluLeuArgVal (SEQ ID NO: 3) присутствует в нативных термостабильных полимеразах таких видов рода Thermus, как, например, Thermus aquaticus, Thermus caldophilus, Thermus thermophilus, Thermus flavus и Thermus filiformis, а также видов Thermus spsl7 и Z05. Последовательность-мотив SerGlnIleGluLeuArgVal (SEQ ID NO: 3) также присутствует в полимеразном домене других термостабильных ДНК-полимераз, получаемых, например, из Thermosipho africanus и из различных штаммов Bacillus, таких как Bacillus caldotenax и Bacillus stearothermophilus. Последовательность-мотив SerGlnIleGluLeuArgIle (SEQ ID NO: 4), например, присутствует в нативных термостабильных полимеразах Thermotoga maritima, Thermotoga neapolitana и Anaerocellum thermophilum.
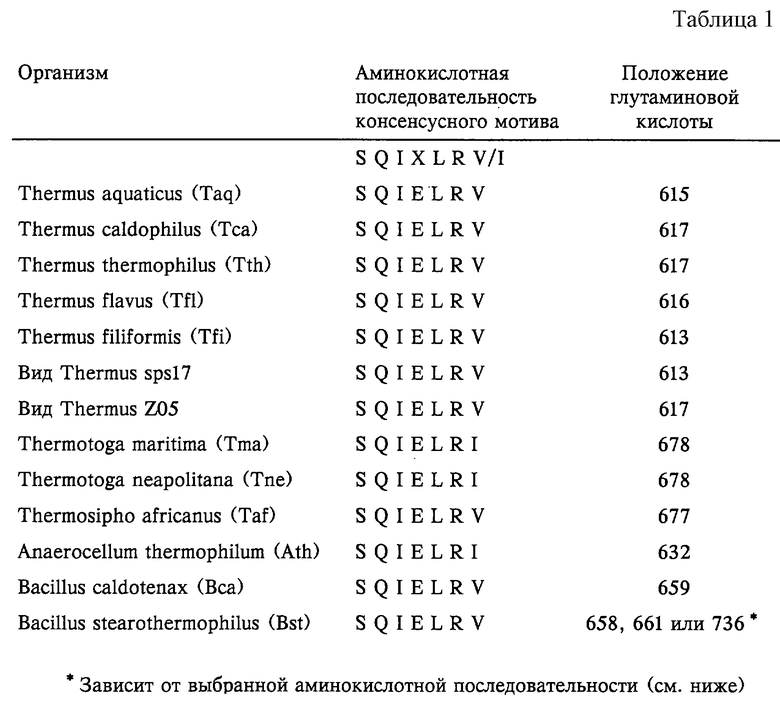
Полная последовательность нуклеиновой кислоты и аминокислотная последовательность для каждой из Taq-, Tth-, Z05-, sps17-, Tma- и Taf-полимераз описана в патенте США 5466591. Последовательность ДНК-полимеразы из Tсa, Tfl, Tne, Ath, Bca и Bst описана в следующих публикациях: Tсa - в базе данных последовательностей EMBL под регистрационным номером U62584 (см. также у Kwon, 1997, Mol. Cells 7(2): 264-271); Tfl - у Akhmetzjanov и Vakhitov, 1992, Nucleic Acids Research 20(21): 5839; Tne - в WO 97/09451 и в WO 96/41014; Ath - в базе данных последовательностей EMBL под регистрационным номером Х98575 (более подробно штамм Ath описан у Rainey и др., 1993, J. Bacteriol. 175(15): 4772-4779); Bst - у Uemori и др., 1993, J. Biochem. 113: 401-410 и в базе данных последовательностей EMBL под регистрационным номером U23149 (см. также у Phang и др., 1995, Gene 163: 65-68). Аминокислотные последовательности Bst-полимеразы, содержащие "Е" в критическом мотиве в положении 658, также описаны в опубликованном японском патенте 05/304964 А, опубликованной европейской заявке ЕР-А-699760 и у Aliotta и др., 1996, Genet. Anal. 12: 185-195; последовательность также может быть получена из базы данных последовательностей EMBL под регистрационным номером U33536. Последовательность, как описано в Gene 163: 65-68 (1995), содержит остаток "Е" в положении 661 критического мотива. Вса-полимераза описана у Uemori и др., 1993, J. Biochem. 113: 401-410 и в базе данных последовательностей EMBL под регистрационным номером D12982. Термостабильная ДНК-полимераза из Thermus filiformis (см. FEMS Microbiol. Lett. 22: 149-153, 1994; также депонированная в АТСС под номером 43280) может быть восстановлена с использованием методов, описанных в патенте США 4889818, а также на основе информации о последовательности, представленной в таблице 1. Каждая из вышеуказанных последовательностей и публикаций включены в настоящее описание в качестве ссылки. Гомология (идентичность последовательности) между аминокислотной последовательностью нативной формы Taq-полимеразы, как указано в WO 89/06691, и последовательностью указанной ранее Tfl-полимеразы составляет более 87,4%. Соответствующая гомология с Tth-полимеразой составляет 87,4%, с Тса-полимеразой составляет 86,6%, с Bst-полимеразой (регистрационный номер U23149) составляет 42,0%, с Вса-полимеразой составляет 42,6% и с Ath-полимеразой составляет 39,7%.
Как видно из таблицы 1, критический мотив является заметно консервативным у термостабильных ДНК-полимераз. В случае, когда "X" обозначает остаток глутаминовой кислоты, изменение гена, кодирующего полимеразу, позволяет получить фермент по изобретению, который легко включает рНТФ по сравнению, например, с Taq-полимеразой, у которой критический мотив не модифицирован. Следовательно, изобретение относится к классу ферментов, который также включает, например, термостабильную ДНК-полимеразу, и к соответствующему гену и векторам экспрессии из Thermus oshimai (Williams и др., 1996, Int. J. Syst. Bacteriol. 46(2): 403-408); Thermus silvanus и Thermus chliarophilus (Tenreiro и др., 1995, Int. J. Syst. Bacteriol. 45(4): 633-639); Thermus scotoductus (Tenreiro и др., 1995, Res. Microbiol. 146(4): 315-324); Thermus brockianus (Munster, 1986, Gen. Microbiol. 132: 1677) и Thermus ruber (Loginov и др., 1984, Int. J. Syst. Bacteriol. 34: 498-499; также депонированные в АТСС под номером 35948). Кроме того, изобретение включает, например, модифицированные формы термостабильных ДНК-полимераз и соответствующие гены и векторы экспрессии из Thermotoga elfii (Ravot и др., 1995, Int. J. Syst. Bacteriol. 45: 312; также депонированные в DSM под номером 9442) и Thermotoga thermarum (Windberger и др., 1992, Int. J. Syst. Bacteriol. 42: 327; также депонированные в DSM под номером 5069). Каждая из вышеуказанных последовательностей и публикаций включены в настоящее описание в качестве ссылки.
В предпочтительном варианте подлежащий модификации критический мотив находится внутри аминокислотной последовательности LeuAspTyrSerGlnIleGluLeuArgValLeuAlaHisLeuSer (SEQ ID NO: 5). Таким образом, одним из предметов изобретения является получение мутантных термостабильных ДНК-полимераз, проявляющих существенно увеличенную эффективность по включению несвойственных нуклеотидов при использовании матрицы. В особенно предпочтительном примере последовательность полимеразы включает LeuAspTyrSerGlnIleGlyLeuArgValLeuAlaHisLeuSer (SEQ ID NO: 6). Такие термостабильные ДНК-полимеразы особенно пригодны в таких процессах, как секвенирование ДНК, синтез РНК на основе ДНК и синтез in vitro ДНК, имеющих в качестве заместителей рНТФ.
Получение термостабильных ДНК-полимераз с повышенной эффективностью по включению несвойственных оснований может быть осуществлено с помощью таких способов, как сайт-направленный мутагенез. См., например, публикацию Sambrook и др., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, 1989, второе издание, глава 15.51, "Oligonucleotide-Mediated Mutagenesis". Например, мутация, заключающаяся в замене "А" на "G" во втором положении кодона, кодирующего остаток глутаминовой кислоты 615 в последовательности гена ДНК-полимеразы Thermus aquaticus (Taq) (см. SEQ ID NО: 7), приводит к более чем 500-кратному увеличению эффективности включения несвойственных нуклеотидов согласно данному описанию, сохраняя при этом способность фермента медиировать ПЦР в присутствии обычных нуклеотидов, т.е. дНТФ.
Эта конкретная мутация в ДНК-полимеразе Taq приводит к замене аминокислоты Е (глутаминовая кислота) на G (глицин). Хотя эта конкретная замена аминокислоты существенно изменяет способность фермента включать несвойственные нуклеотиды, следует ожидать, что замена остатка глутаминовой кислоты на остатки любых других аминокислот, такие как, например, остатки серина, цистеина, треонина, аланина, валина или лейцина, будет оказывать такое же действие. Следовательно, настоящее изобретение включает и другие аминокислотные заместители, которые замещают Е615, хотя мутант E615G является предпочтительным. Таким образом, в основе изобретения лежит тот факт, что четвертый остаток аминокислоты в мотиве, представленном в SEQ ID NO: 1, не является остатком глутаминовой кислоты.
Сайт-направленный мутагенез также может быть осуществлен путем сайт-специфического праймернаправленного мутагенеза. Этот способ в настоящее время является стандартным в данной области техники, и его осуществляют с использованием синтетического олигонуклеотидного праймера, комплементарного одноцепочечной ДНК фага, которая подлежит мутированию, за исключением ограниченного ошибочного спаривания, представляющего собой требуемую мутацию. В целом в качестве праймера для прямого синтеза цепи, комплементарной плазмиде или фагу, применяют синтетический олигонуклеотид, а образовавшуюся двухцепочечную ДНК трансформируют в бактерию-хозяина, поддерживающую фаг. Образовавшаяся бактерия может быть проанализирована, например, методом секвенирования ДНК или путем гибридизации с зондом с целью выявить те бляшки, которые несут требуемую измененную последовательность гена. В альтернативном варианте могут применяться методы "рекомбинантной ПЦР", описанные в PCR Protocols, San Diego, Academic Press, под ред. Innis и др., 1990, глава 22, озаглавленная "Recombinant PCR", Higuchi, стр.177-183.
Как показано в таблице 1, глутаминовая кислота в критическом мотиве Taq-полимеразы сохраняется в других термостабильных ДНК-полимеразах, однако может быть локализована в другом, но близком положении аминокислотной последовательности. Изменение консервативной глутаминовой кислоты в SEQ ID NO: 2 термостабильных ДНК-полимераз из представителей рода Thermus и ДНК-полимераз из родственных родов Thermotoga, Thermosipho и Anaerocellum должно также усиливать способность полимеразы по эффективному включению несвойственных нуклеотидов по сравнению с Taq-полимеразой, содержащей SEQ ID NO: 2.
Изменение остатка глутаминовой кислоты в критическом мотиве других термостабильных ДНК-полимераз может быть осуществлено с использованием принципов и способов, применяемых в сайт-направленном мутагенезе. В GeneBank или в базе данных SwissProt/PIR существует несколько последовательностей ДНК-полимеразы Bacillus stearothermophilus. Эти последовательности имеют высокую степень родства, однако несколько отличаются друг от друга, но каждая из них содержит идентичную критическую последовательность-мотив SerGlnIleGluLeuArgXaa (SEQ ID NO: 2), где "Хаа" в положении 7 этой последовательности обозначает остаток валина (Val) или остаток изолейцина (Ile), хотя и в различных положениях последовательности.
Основываясь на опубликованной информации о доступных аминокислотных и нуклеотидных последовательностях термостабильных ДНК-полимераз, представленных в данном описании, с помощью стандартных методов рекомбинации также можно конструировать химерные полимеразы, которые состоят из доменов, происходящих из различных термостабильных ДНК-полимераз. В патентах США 5466591 и 5374553 описаны методы обмена различными функциональными сегментами термостабильных полимераз, такими как 5’-3’-экзонуклеазный домен, 3’-5’-экзонуклеазный домен и полимеразный домен, с получением новых ферментов. Предпочтительные химерные термостабильные ферменты полимеразы включают 5’-3’-экзонуклеазный домен, 3’-5’-экзонуклеазный домен и полимеразный домен, причем один из доменов происходит от другой полимеразы, и полимеразный домен содержит критический мотив SerGlnIleXaaLeuArgXaa (SEQ ID NO: 1), где "Хаа" в положении 4 этой последовательности представляет собой остаток любой аминокислоты, кроме остатка глутаминовой кислоты (Glu), а "Хаа" в положении 7 этой последовательности представляет собой остаток валина (Val) или изолейцина (Ile). Примерами таких химерных молекул являются химерные ферменты Taq/Тmа, строение которых приведено в таблице 2. Как видно из этой таблицы, полимеразный домен этих химерных ферментов Taq/Tma содержит мутацию в указанном выше критическом мотиве.
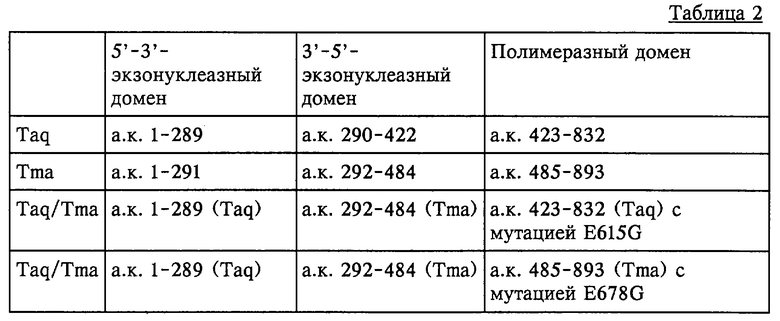
Плазмида рС1 в соответствии с Будапештским договором была депонирована в АТСС 17 июля 1996 г. и получила регистрационный номер 98107. Плазмида рС1 содержит ген, кодирующий термостабильную ДНК-полимеразу с мутацией в кодоне, кодирующем остаток глутаминовой кислоты в положении 615 аминокислотной последовательности нативной Taq-полимеразы, что приводит к получению мутантной формы Taq-полимеразы, которая имеет остаток глицина в положении 615 (Е615G-мутантная Taq-полимераза). Этот депозит позволяет применять альтернативные способы для получения термостабильных ДНК-полимераз, обладающих повышенной эффективностью по включению несвойственных аналогов нуклеотидов. В примере I показано, что применение фланкирующих сайтов рестрикции, пригодных для субклонирования мутации E615G, позволяет создать другие термостабильные ферменты ДНК-полимеразы. Поскольку для многочисленных термостабильных ДНК-полимераз известна полная последовательность гена, основываясь на информации о последовательности критического мотива, приведенной в описании, для введения мутации Е615 можно использовать другие известные специалистам в данной области техники способы, такие как расщепление рестриктазами и замещение фрагмента или сайт-специфический мутагенез in vitro.
Модифицированный ген или фрагмент гена, полученный сайт-специфическим мутагенезом, может быть получен из плазмиды или фага стандартными способами и лигирован с вектором экспрессии для последующего культивирования и выделения образовавшегося фермента. Для практического осуществления изобретения пригодны многочисленные клонирующие и экспрессирующие векторы, включая системы, пригодные для млекопитающих и бактерий, и они описаны, например, у Sambrook и др., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, второе издание, 1989. Для удобства в настоящем изобретении приведен пример использования промотора PL, происходящего из фага лямбда (Shimatake и др., 1981, Nature 292: 128). Применение этого промотора, в частности, описано в патентах США 4711845 и 5079352.
Термостабильные ДНК-полимеразы по настоящему изобретению обычно выделяют из микроорганизмов, таких как Е.coli, которые были трансформированы вектором экспрессии, функционально связанным с геном, кодирующим ДНК-полимеразу дикого типа или модифицированную термостабильную ДНК-полимеразу. Примером пригодного микроорганизма-хозяина является штамм Е. coli DG116, описанный у Lawyer и др., 1993, PCR Methods and Applications 2: 275-287, и этот штамм также может быть получен из американской коллекции типовых культур (American Type Culture Collection) под регистрационным номером АТСС 53601. Методы очистки термостабильной полимеразы также описаны, например, у Lawyer и др., 1993, PCR Methods and Applications 2: 275-287.
Для специалиста в данной области техники очевидно, что вышеуказанные термостабильные ДНК-полимеразы, обладающие увеличенной эффективностью по включению несвойственных нуклеотидов, наиболее легко могут быть получены с использованием методов, основанных на технологии рекомбинантной ДНК. Необходимость получения одного из ферментов по настоящему изобретению или производного либо гомолога этих ферментов предпологает создание рекомбинантной формы фермента, которое обычно включает конструирование вектора экспрессии, трансформацию клетки-хозяина этим вектором и культивирование трансформированной клетки-хозяина в условиях, при которых такая экспрессия будет происходить. Способы получения векторов экспрессии, трансформации и культивирования трансформированной клетки-хозяина хорошо известны в данной области техники и подробно описаны, например, у Sambrook и др., 1989, см. выше.
Настоящее изобретение относится к термостабильным ДНК-полимеразам, которые вместе с рибонуклеозидтрифосфатами пригодны для осуществления многочисленных функций, включая амплификацию нуклеиновой кислоты, методы определения и секвенирования ДНК. Применение рибонуклеотидов при секвенировании позволяет исключить использование цепь-терминирующих аналогов, имеющих высокую стоимость, таких как ддНТФ, и, что является важным фактором, облегчает получение новых продуктов амплификации, пригодных не только для анализа последовательности ДНК, но и для других типов анализа, таких как электрофорез или гибридизация, без необходимости последующего проведения реакций секвенирования ДНК.
Ранее уже было подтверждено, что пирофосфатаза улучшает результаты секвенирования при использовании как мезофильных полимераз, так и термостабильных ДНК-полимераз за счет снижения количества накапливающихся продуктов удлинения, полученных в результате пирофосфоролизиса. При этом до начала цикла секвенирования необходимо использовать методы, позволяющие вводить дополнительный фермент в реакцию секвенирования. Однако наиболее существенное преимущество настоящего изобререния состоит в том, что для секвенирования ДНК не требуется пирофосфатаза. Таким образом, применение новых, представленных в данном описании ферментов устраняет необходимость в дополнительных расходах по введению второго фермента в смесь для реакции секвенирования.
При применении ферментов по настоящему изобретению реакции амплификации и секвенирования объединяют, что способствует экономии времени и материалов, а также упрощает весь анализ. Эти, а также другие преимущества достигаются, в первую очередь, благодаря тому, что включение как обычных нуклеотидов, так и рибонуклеотидов и их аналогов в продукт удлинения праймера позволяет получить химерную цепь РНК/ДНК, чувствительную к гидролизу РНК. Обработка не оказывает влияния на скелет ДНК и приводит к получению популяции фрагментов нуклеиновой кислоты, каждый из которых заканчивается в том положении, где был встроен рибонуклеотид вместо соответствующего дНТФ. Гидролиз легко осуществляют различными способами, включающими, но не ограничиваясь ими, щелочной гидролиз (например, обработка NaOH в конечной концентрации 0,2 М, как описано ниже в примере VI), нагревание или ферментативная обработка РНКазой (под ред. Vogel и др., Informational Macromolecular, New York, Academic Press, 1963, глава, написанная Berg и др., озаглавленная "The Synthesis of Mixed Polynucleotides Containing Ribo- and Deoxyribonucleotide by Purified Preparation of DNA Polymerase from E.coli", стр.467-483).
В предпочтительном варианте настоящее изобретение относится к новым и улучшенным композициям, наиболее пригодным для методов секвенирования ДНК. Новые ферменты, приведенные в данном описании, могут использоваться для методов секвенирования нуклеиновых кислот, основанных на применении либо окрашенных терминаторов, либо окрашенных праймеров, а также для других методов секвенирования. Как описано ранее, методы терминации цепи обычно требуют использования матрицы для удлинения праймера в присутствии терминирующих цепь нуклеотидов, что приводит к созданию неполных фрагментов, которые затем разделяют по размеру. В стандартном дидезокси-секвенировании для терминации цепи используются дидезоксинуклеозидтрифосфаты и ДНК-полимераза, такая как фрагмент Кленова из штамма E.coli Pol I (Sanger и др., см. выше).
Таким образом, основная методика дидезокси-секвенирования включает (I) отжиг олигонуклеотидного праймера, соответствующего матрице; (II) удлинение праймера с помощью ДНК-полимеразы в четырех различных реакционных смесях, в каждой из которых содержится один меченый нуклеотид или меченый праймер, смесь немеченых дНТФ и один терминирующий цепь ддНТФ; (III) разделение четырех наборов продуктов реакций с помощью, например, гель-электрофореза на денатурирующем полиакриламидном геле с высокой разрешающей способнстью/мочевине, капиллярного разделения или других способов разделения; и (IV) получение авторадиографического изображения геля, которое может быть проанализировано для получения заключения о последовательности. В альтернативном варианте для получения информации о последовательности ДНК могут применяться масс-спектрометрические методы или методы, основанные на гибридизации, с использованием флуоресцентно меченных праймеров или нуклеотидов.
Доступность термоустойчивых полимераз, таких как Taq-полимераза, привела к улучшению метода секвенирования (см. патент США 507216) и к его модификациям, называемым "цикличным секвенированием". При цикличном секвенировании повторяют циклы нагревания и охлаждения, что позволяет получить многочисленные продукты удлинения каждой молекулы-мишени (см. Murray, 1989, Nucleic Acids Research, 17: 8889). Эта асимметричная амплификация последовательностей-мишеней, комплементарных последовательности матрицы, в присутствии дидезокси-терминаторов цепи приводит к получению семейства продуктов удлинения всех возможных длин.
После денатурации продукта реакции удлинения ДНК-матрицы множественные циклы отжига праймера и удлинения праймера осуществляют в присутствии дидезокси-терминаторов. Термостабильные ДНК-полимеразы имеют несколько преимуществ при цикличном секвенировании; они устойчивы к жестким температурам отжига, которые необходимы для специфической гибридизации праймера с нуклеиновыми кислотами-мишенями, а также устойчивы к множеству циклов высокотемпературной денатурации, которые осуществляются в каждом цикле, т.е. к температуре 90-95°С. По этой причине различные формы ДНК-полимеразы AmpliTaq® были включены в наборы для цикличного секвенирования с использованием Taq, которые реализуются через фирму Perkin Elmer, Норуолк, шт. Коннектикут.
Однако способность ДНК-полимеразы Taq ограничивать включение несвойственных нуклеотидов, таких как ддНТФ, связана с проблемой ее использования для цикличного секвенирования, поскольку ддНТФ или флуоресцентно меченные ддНТФ должны быть включены в качестве терминаторов цепи. В целом до создания настоящего изобретения секвенирование ДНК с помощью термостабильных ДНК-полимераз требовало наличия смеси терминирующих цепь нуклеотидов, обычно дидезоксинуклеотидов, в высокой концентрации с целью гарантировать получение популяции продуктов удлинения, которая включает фрагменты, имеющие все возможные длины, вплоть до длины в несколько сотен оснований. Часто для снижения стоимости этого процесса применяли протоколы с использованием очень низких концентраций обычных дНТФ, что делало реакции неэффективными. Эти реакционные смеси, включающие низкую концентрацию дНТФ и высокую концентрацию ддНТФ, создавали условия, при которых термостабильная полимераза испытывает существенный недостаток нуклеотидных субстратов.
Несмотря на появление модифицированных ферментов, таких как ДНК-полимераза AmpliTaq® FS, позволяющих увеличить концентрацию дНТФ до более оптимальных уровней, для ферментов-прототипов все еще были необходимы дорогостоящие ддНТФ для секвенирования ДНК. В противоположность этому настоящее изобретение относится к ферментам, которые не только позволяют увеличивать концентрацию дНТФ, но и исключают применение дорогостоящих ддНТФ, поскольку вместо них в растущую цепь включают рНТФ. Способность новых ферментов эффективно осуществлять частичное замещение рибонуклеотидами облегчает получение ступеней для секвенирования ДНК при отсутствии отдельной реакции по включению терминирующего нуклеотида.
Выбор несвойственных нуклеотидных аналогов, пригодных для использования в методах секвенирования ДНК, ранее определялся способностью термостабильной ДНК-полимеразы включать эти аналоги. К сожалению, эти нуклеотидные аналоги являются довольно дорогостоящими. Например, цена ддНТФ приблизительно в 25 раз превышает цену как рНТФ, так и дНТФ. Поскольку ранее термостабильные ДНК-полимеразы не обладали способностью эффективно включать рНТФ в растущую цепь ДНК при использовании матрицы, такие рибонуклеотиды, которые являются легко доступными и недорогими, не могли использоваться для секвенирования ДНК с участием термостабильной ДНК-полимеразы. Настоящее изобретение позволяет исключить необходимость в ддНТФ в реакциях секвенирования ДНК. Таким образом, одиним из предметов изобретения является метод секвенирования ДНК, который является существенно менее дорогостоящим по сравнению с ранее применяемыми методами терминации цепи.
Присутствие марганца в реакции удлинения праймера оказывать влияние на способность полимеразы точно включать правильные пары нуклеотидных оснований. Марганец может применяться для усиления некорректного спаривания оснований или для снижения ограничения по инсерции нуклеотидного аналога. Ранее исследователи применяли марганец с той целью, чтобы индуцировать мутагенез при репликации или амплификации ДНК. Таким образом, марганец может влиять на правильность реакции полимеризации, а также на выход продукта реакции. Образовавшаяся последовательность может оказаться неправильной или же - в методе секвенирования ДНК - полученная информация может оказаться неверной. Способы согласно настоящему изобретению не требуют включения двухвалентного катиона марганца в смесь для реакции секвенирования с целью усиления способности полимеразы встраивать несвойственный нуклеотид. В противоположность известным из уровня техники ДНК-полимеразам в настоящем изобретении описан критический мотив, входящий в полимеразный домен, контролирующий способность фермента различать 2’-замещенные и незамещенные нуклеотиды, не используя для этого марганец.
Ферменты по настоящему изобретению не требуют для секвенирования высоких концентраций несвойственных аналогов оснований. До создания настоящего изобретения при осуществлении секвенирования, основанном на терминации цепи ДНК (см. также патент США 5075216 на имя Innis и др.), несвойственные аналоги оснований и соответствующие традиционные основания обычно присутствовали в соотношении (например, ддАТФ:дАТФ) приблизительно от 1,3:1 до 24:1. Для сравнения термостабильные полимеразы, описанные в настоящем изобретении, позволяют снизить отношение несвойственных аналогов оснований к обычным основаниям в сто или в несколько тысяч раз. Отношение рНТФ:дНТФ, равное 1:1 или ниже, в сочетании с новыми ферментами, предствленными в данном описании, является достаточным для анализа последовательности ДНК. В предпочтительном варианте отношение рНТФ:дНТФ снижают до менее чем 1:8. Отношение 2’-замещенного нуклеотида к соответствующему естественному дНТФ может составлять даже 1:80 или 1:200, что зависит от конкретного плана эксперимента и требуемой длины фрагментов.
Таким образом, поскольку ферменты по настоящему изобретению легко включают несвойсвенные нуклеотиды, такие как 2’-замещенные нуклеотиды, нет необходимости усиливать их способность по включению рНТФ с помощью высокой концентрации рНТФ и ограничения концентрации соответствующего дНТФ. Следовательно, методы по настоящему изобретению позволяют применять оптимальные концентрации дНТФ в сочетании с небольшими количествами рНТФ.
Когда в соответствующем методе, таком как секвенирование с окрашенным праймером, применяют модифицированные ферменты полимеразы по настоящему изобретению, хорошие результаты по секвенированию ДНК получают при концентрации каждого дНТФ 50-500 мкМ. Предпочтительно концентрация дНТФ составляет от 100 до 300 мкМ. При использовании этих интервалов соответствующий рНТФ может присутствовать приблизительно в такой же концентрации, что и дНТФ, или в более низкой. Предпочтительно концентрация рНТФ составляет приблизительно от 0,1 мкМ до 100 мкМ, наиболее предпочтительно приблизительно от 2,5 мкМ до 25 мкМ.
Концентрация рНТФ, пригодная для применения модифицированных ферментов по настоящему изобретению, легко может быть определена обычным специалистом в данной области техники путем титрования и оптимизации экспериментов. Необходимое количество рНТФ или аналога определяется типом эксперимента, и на него может влиять размер и чистота мишени, а также выбор буфера и конкретные виды фермента.
Отношение рНТФ:дНТФ будет определять частоту, с которой рНТФ встраиваются в растущий олигонуклеотид. Поскольку в каждом встроенном рНТФ может происходить гидролиз, отношение рНТФ:дНТФ можно регулировать таким образом, чтобы обеспечить пользователю гибкость в возможности увеличения или уменьшения размера образовавшихся фрагментов.
Как хорошо известно, ДНК представляет собой полимер, синтезированный из дНТФ. Каждый дезоксинуклеозидтрифосфат включает сахар рибозу, который имеет гидроксильную группу в 3’-положении и водород в 2’- положении. Рибонуклеотиды также содержат гидроксильную группу в 3’-положении сахара. Однако рНТФ отличаются от дНТФ тем, что в 2’-положении сахара атом водорода замещен на вторую гидроксильную группу. В данном случае рНТФ являются примером, на котором выявляется способность ферментов по изобретению точно включать 2’-замещенные нуклеотиды. Однако соединения по изобретению не ограничены применением несвойственных нуклеотидов, которые представляют собой рибонуклеотиды. Модификация последовательности термостабильной полимеразы в критическом домене, приведенном в данном описании, позволяет матрице направленно включать альтернативные 2’-замещенные нуклеотиды, такие как 2’-гидроксил,3’-дезоксинуклеотиды и нуклеотиды, замещенные в 2’-положении фтором или аминогруппой.
Как описано в приведенных ниже примерах, включение 3’-дезокси, 2’-гидроксиАТФ, который в данном описании обозначен также как кордицепинтрифосфат, облегчается наличием второй мутации в термостабильной полимеразе, способность которой ограничивать включение нуклеотида, содержащего дезоксигруппу в 3’-положении рибозы, снижена. Такие ферменты ранее были описаны, например, в ЕР-А-655506 и в заявке на патент США 08/448223, поданной 23 мая 1995 г. Депозит АТСС 69820, депонированный в соответствии с Будапештским договором 10 мая 1995 г., обеспечивает получение гена, кодирующего модифицированную термостабильную ДНК-полимеразу Thermus aquaticus, способность которой ограничивать включение аналогов, таких как ддНТФ, снижена. Дидезоксинуклеотиды имеют заместитель в 3’-положении по сравнению с обычными дНТФ. Таким образом, в сочетании с настоящим изобретением двойная мутация, примером которой в данном описании является Е615G, F667Y-мутант Taq-полимеразы, позволяет использовать нуклеотидные аналоги, которые содержат заместитель в 3’- и 2’-положениях рибозы по сравнению с дНТФ (см. примеры III и V).
Конкретным применением изобретения является метод секвенирования с использованием рНТФ, где секвенирующий праймер является меченным с помощью выявляемой флуоресцентной или радиоактивной метки. В отличие от ддНТФ включение немодифицированного рНТФ не приводит к такому действию, как терминация цепи. Реакционная смесь для секвенирования ДНК включает как рНТФ, так и дНТФ в сочетании с ферментом по изобретению, что приводит к получению смеси беспорядочно замещенных продуктов удлинения праймера, чувствительных к расщеплению по 3’-5’-фосфодиэфирной связи между рибонуклеотидом и соседним дезоксирибонуклеотидом. После удлинения праймера, например, при ПЦР-амплификации или при цикличном секвенировании и до разделения продуктов удлинения праймера, например, с помощью гель-электрофореза реакционную смесь либо обрабатывают щелочью, нагревают, обрабатывают рибонуклеазой, либо другими способами гидролизуют продукты удлинения по каждому встречающемуся рибонуклеотиду. Для каждого меченого продукта удлинения праймера только наиболее крупный 5’-фрагмент, который является промежуточным продуктом удлинения меченого праймера, обнаруживается на секвенирующем геле. Для данной мишени анализ полученного секвенирующего геля дает ступени секвенирования, т.е. серии обнаруживаемых сигналов в Г-, А-, Т- и Ц-линиях, соответствующих нуклеотидной последовательности мишени. Полученные ступени секвенирования предоставляют такую же информацию, что и обычный метод, предполагающий использование ддНТФ или основанный на использовании рНТФ и новых термостабильных полимераз, представленных в данном описании. Таким образом, при применении настоящего изобретения дорогостоящие ддНТФ для секвенирования ДНК больше не требуются (см. пример VI).
В альтернативном методе секвенирования применяют терминирующие цепь рибонуклеотиды. В этом варианте в качестве терминаторов применяют 2’-гидрокси,3’-дезоксинуклеотиды, такие как кордицепинтрифосфат. Эти аналоги рНТФ могут быть флуоресцентно меченными, и их применяют для секвенирования ДНК. У Lee и др., см. выше, описано применение окрашенных терминаторов ддНТФ. В ЕР-А-655506 и в заявке на патент США 08/448223, поданной 23 мая 1995 г., описаны модифицированные ферменты для применения с ддНТФ. Термостабильная ДНК-полимераза, содержащая как модификацию, присутствующую в ДНК-полимеразе FS AmpliTaq® (см. выше), так и таковую, представленную в SEQ ID NO: 1, где X не обозначает глутаминовую кислоту (Е), как описано ранее, может применяться для эффективного включения меченых аналогов рНТФ при осуществлении реакции секвенирования, которая основана на терминации цепи. Этот процесс может быть автоматизирован и не требует синтеза меченных красителем праймеров. Кроме того, поскольку реакции с окрашенным терминатором позволяют осуществлять в одной и той же пробирке все четыре реакции, они более удобны, чем методы с окрашенным праймером. 2’-гидрокси,3’-дезоксинуклеотиды могут быть синтезированы из коммерчески доступных 3’-нуклеотидов (3’-дА, 3’-дЦ, 3’-дГ и 3’-дТ, например, поставляемых фирмой Sigma Chemical Corporation, Ст. Луис, шт. Миссури) и при добавлении 5’-трифосфата, как описано у Ludwig, Biophosphates and Their Synthesis Structure, Metabolism and Activity, под ред. Bruzik и Stec, Амстердам, Elsevier Science Publishers, 1987, стр.201-204.
Помимо применения в новых методах секвенирования, модифицированные ферменты, представленные в данном описании, могут применяться для различных целей в молекулярной биологии. В одном из примеров осуществления изобретения модифицированные ферменты применяют в реакционных смесях для амплификации, содержащих как обычные, так и несвойственные нуклеотиды, например дНТФ и по крайней мере один обнаруживаемый в результате мечения рНТФ, метки, которые включают, например, флуоресцентные метки или радиоизотопы. Управляемый матрицей синтез комплементарной цепи приводит к получению ДНК-продукта, содержащего рибонуклеозидмонофосфат в различных положениях по его длине. Нагревание и/или обработка щелочью гидролизуют продукт удлинения нуклеиновой кислоты на каждом рибонуклеотиде. Таким образом, получают семейство сегментов ДНК, где каждый фрагмент содержит один меченый остаток на своем 3’-конце. Размер образовавшихся фрагментов нуклеиновой кислоты может быть изменен путем регулирования отношения и количества рНТФ, включенного в реакцию.
Амплификация мишени с использованием рНТФ и ферментов по настоящему изобретению создает многочисленные преимущества, обусловленные конкретным применением. В описанном выше методе при использовании меченого рНТФ все полученное семейство фрагментов оказывается меченым с равной интенсивностью: одна метка на один олигонуклеотидный фрагмент. Для достижения оптимальных результатов при осуществлении таких методов, как обнаружение нуклеиновой кислоты с использованием олигонуклеотидного зонда, определенным образом фиксированного на тонком слое силикона, обычно необходимо, чтобы амплифицированная мишень была случайным образом фрагментирована в пределах фиксированного и воспроизводимого размера с целью ограничить образование вторичных структур для контроля кинетики гибридизации. Кроме того, для выявления гибридизации при упорядоченном расположении тысяч зондов на тонком слое силикона может оказаться предпочтительным, чтобы фрагменты нуклеиновой кислоты были мечены с равной интенсивностью. Согласно настоящему изобретению были разработаны способы получения семейств фрагментов, удовлетворяющих этому стандарту, облегчая тем самым применение альтернативных форматов выявления, таких, как методы, основанные на использовании тонких слоев, описанные, например, у Cronin и др., 1996, Human Mutation, 7: 244-255.
В другом примере предполагается применение одного меченого праймера и одного немеченого праймера в смеси для реакции амплификации, которая включает термостатабильную полимеразу по изобретению, а также рНТФ и дНТФ, что позволяет одновременно улучшить реакции амплификации и секвенирования. Для этого метода требуется, чтобы осуществлялись четыре различные реакции амплификации, по одной для каждого рНТФ. Таким образом, например, поскольку фермент по изобретению пригоден для амплификации мишени, например, с помощью ПЦР или других способов амплификации, полученный продукт, если он присутствует, может быть обнаружен обычными способами, такими как гель-электрофорез или гибридизация с зондом при использовании части продукта реакции. Эти методы определения не приводят к гидролизу включенных рибонуклеотидов, а поведение РНК/ДНК-химерных цепей будет таким, как это ожидается для обычного продукта амплификации нуклеиновой кислоты. Если требуемый продукт обнаружен, оставшаяся часть этой же реакционной смеси может быть обработана щелочью и проанализирована с помощью гель-электрофореза с целью определить последовательность нуклеиновой кислоты. Таким образом, после обнаружения продукта последующая реакция секвенирования не является необходимой. Эта упрощенная методика обеспечивает экономию затрат времени и материалов и характеризуется повышенной точностью благодаря уменьшению стадий: обнаруженный продукт и является секвенированным продуктом.
Кроме того, также может применяться аналогичная методика с четырьмя мечеными рНТФ и одним биотинилированным (меченным биотином) праймером. После амплификации продукт разлагают с помощью щелочного гидролиза, а праймер, связанный с продуктами, удаляют путем взаимодействия с покрытыми стрептавидином гранулами. Захваченные гранулами продукты затем анализируют на секвенирующем геле. Эта модификация позволяет осуществлять реакцию секвенирования в одной пробирке, исключая тем самым необходимость в четырех различных амплификациях.
В соответствии с другим предметом изобретения ферменты, представленные в данном описании, могут применяться для получения РНК с использованием ДНК-матрицы или для получения замещенной ДНК для медиируемой щелочью стерилизации без применения обычных стерилизующих агентов, таких как урацил-N-гликозилаза (UNG), как описано в международной заявке WO 92/01814.
В приведенных примерах осуществления изобретения термостабильная полимераза также содержит мутацию в 5’-3’-экзонуклеазном домене, которая приводит к значительному ухудшению экзонуклеазной активности. Модифицированные формы Taq-полимеразы описаны в патенте США 5466591. В одном из вариантов осуществления изобретения кодон, кодирующий остаток глицина (G) в положении 46 аминокислотной последовательности, замещали кодоном, кодирующим аспарагиновую кислоту (D). Полученный таким образом фермент обладает преимуществом при использовании в реакциях цикличного секвенирования благодаря пониженной 5’-3’-экзонуклеазной активности и является предпочтильным для применения по настоящему изобретению. По сравнению с ферментом дикого типа наличие мутации G46D не оказывает влияния на полимеразный домен аминокислотной последовательности и на полимеразную активность.
Согласно изобретению предлагаются также наборы для секвенирования нуклеиновых кислот, содержащие термостабильную полимеразу по настоящему изобретению, что является примером коммерческого применения изобретения.
Такие наборы обычно включают дополнительные реагенты для секвенирования ДНК, такие как, например, рНТФ, дНТФ и соответствующие буферы. Если рНТФ являются немечеными, также может быть включен меченый праймер.
Ниже изобретение проиллюстрировано на примерах, не ограничивающих его объем.
Пример I
Экспрессия гена модифицированной Taq-полимеразы с пониженной способностью ограничивать включение несвойственных нуклеотидов
С-концевая часть аминокислотной последовательности ДНК-полимеразы Taq кодирует домен, включающий сайт полимеразной активности (Lawyer и др., 1993, PCR Methods and Applications 2: 275-287). Фрагмент ДНК, содержащий эту область, выделяли из полноразмерного гена Taq и осуществляли мутагенез с помощью ПЦР-амплификации в присутствии марганца (Leung и др., Technique 1(1): 11-15). В этом примере все рестриктазы были получены от фирмы New England Biolabs, Беверли, шт. Массачусетс. Фрагменты, содержащие мутации, расщепляли рестриктазами PstI и BglII и клонировали в плазмиде, экспрессирующей Taq, в данном случае в плазмиде pLK102, которую предварительно расщепляли PstI и BglII. Плазмида pLK102 представляет собой модифицированную форму плазмиды pSYC1578, которая экспрессирует Taq (Lawyer и др., см. выше). Фрагмент HincII/EcoRV, локализованный на 3’-области, кодирующей полимеразу, удаляли для создания плазмиды pLK101. PstI-BglII-фрагмент длиной 898 пар оснований затем удаляли из pLK101 и заменяли коротким олигонуклеотидным дуплексом PstI-EcoRV-BglII для создания плазмиды pLK102. Таким образом, в результате этой делеции было удалено 900 пар оснований с 3’-конца pol-гена ДНК Taq с их замещением более коротким участком ДНК.
Полученными экспрессирующими плазмидами трансформировали штамм Е.coli N1624 (описанный у Gottesman, 1973, J. Mol. Biol. 77: 531; он также может быть получен из Е.coli Genetic Stock Center в Йельском Университете, номер штамма CGSC 5066) и полученных трансформантов подвергали скринингу на способность более эффективно по сравнению с ферментом дикого типа включать рНТФ. Используя эту методику, выявляли мутант С1, способный более эффективно включать рНТФ.
Для определения, какая часть гена Taq-полимеразы ответственна за измененный фенотип, содержащую мутацию плазмиду, экспрессирующую Taq, названную рС1 и выделенную из мутанта С1, расщепляли различными рестриктазами и образовавшиеся рестрикционные фрагменты субклонировали в плазмиде pLK101, несущей ген ДНК-полимеразы Taq дикого типа, замещая рестрикционные фрагменты, не содержащие мутаций. Анализ образовавшихся субклонов показал, что мутация, отвечающая за фенотип, находилась в рестрикционном фрагменте NheI-BamHI длиной 265 пар оснований.
Осуществляли анализ последовательности ДНК этой области плазмиды рС1, используя набор ABI PRISMa Dye Terminator Cycle Sequencing Core Kit, в который входит ДНК-полимераза FS AmpliTaq®, поставляемая фирмой Applied Biosystems, Фостер Сити, Калифорния, и система для секвенирования Applied Biosystems, Model 373A DNA Sequencing System. Анализ последовательности выявил две миссенс-мутации в гене Taq-полимеразы между сайтами Nhel и BamHI. Мутация аминокислоты в положении 615 привела к тому, что остаток глутаминовой кислоты (Е) был замещен остатком глицина (G), а другая мутация в положении 653 привела к замещению остатка аланина (А) на треонин (Т). Нумерацию начинают с кодона, кодирующего первый остаток метионина зрелого протеина, как описано в патенте США 5079352. Мутация E615G привела к замене GAG на GGG в кодоне 615. Мутация А653Т привела к замене GCC на АСС в кодоне 653. Плазмида С1 в штамме-хозяине Е.coli T1624 была депонирована в соответствии с Будапештским договором в АТСС 17 июля 1996 года и получила регистрационный номер 98107.
Две точечные мутации анализировали раздельно путем субклонирования каждой отдельно в гене Taq-полимеразы дикого типа, используя рекомбинантную ПЦР (под. ред Innis и др., PCR Protocols, San Diego, Academic Press, 1990, глава 22, озаглавленная "Pecombinant PCR", автор Higuchi, стр.177-183). Образовавшиеся продукты экспрессии анализировали с целью определить, какая из мутаций E615G или А653Т либо обе ответственны за фенотип, для которого характерно включение рибонуклеотидов. Результаты эксперимента показали, что за мутантный фенотип ответственна исключительно мутация E615G.
Для дальнейшего анализа и количественной оценки эффективности включения аналогов нуклеотидов ПЦР-фрагмент BamHI-Nhel длиной 265 пар оснований, содержащий мутацию E615G, клонировали в векторе экспрессии Taq, обозначенном pRDA3-2. Вектор экспрессии pRDA3-2 содержал полноразмерный ген Taq, функционально связанный с промотором фага-лямбда PL. Экзонуклеазный домен гена Taq в этом векторе содержал точечную мутацию в кодоне, кодирующем глицин, т.е. аминокислотный остаток в положении 46, которая снижает 5’-3’-экзонуклеазную активность. Однако последовательность гена внутри полимеразного домена вектора экспрессии pRDA3-2 идентична последовательности гена Taq дикого типа. Плазмида pRDA3-2 полностью описана в патенте США 5466591, где плазмида обозначена как "клон 3-2". Плазмиду pRDA3-2 расщепляли с помощью BamHI и NheI и ПЦР-фрагмент длиной 265 пар оснований стандартными способами лигировали с вектором.
Полученной плазмидой pLK108 трансформировали штамм Е.coli DG116 (Lawyer и др., 1993, см. выше, этот штамм также может быть получен из американской коллекции типовых культур под номером АТСС 53606). Плазмида pLK108 кодирует термостабильную ДНК-полимеразу, обозначенную в настоящем описании как G46D, E615G-Taq. С помощью рекомбинантной ПЦР путем комбинации мутаций E615G и P667Y во фрагменте BamHI-NheI получали мутант G46D, E615G, F667Y-Taq. Этот фрагмент клонировали в плазмиде pRDA3-2 для создания плазмиды pLK109. Экспрессируемый плазмидами pLK108 и pLK109 протеин термостабильной ДНК-полимеразы выделяли в соответствии с методом, описанным у Lawyer и др., 1993, см. выше, хотя стадии хроматографии были исключены. Представляющую интерес последовательность подтверждали с помощью анализа последовательности ДНК. Во вставке (инсерции) pLK108 была обнаружена дополнительная мутация в последовательности; однако эта мутация не изменяла аминокислотную последовательность протеина.
После частичной очистки активность модифицированного фермента определяли в соответствии с методом оценки активности, описанным у Lawyer и др., 1989, J. Biol. Chem. 264: 6427-6437. Активность модифицированного фермента оценивали следующим образом: одна единица фермента соответствует 10 нмолям продукта, синтезированного в течение 30 минут. ДНК-полимеразная активность фермента дикого типа прямо пропорциональна концентрации фермента, обеспечивающей включение до 80-100 пмолей дЦМФ (разбавление фермента составляет до 0,12-0,15 единиц на реакцию). Активность мутантов E615G, G46D и E615G, F667Y, G46D прямо пропорциональна концентрациям, обеспечивающим включение до 0,25-3 пмолей дЦМФ (разбавление фермента составляет от 6×10-4 до 5×10-3 единиц на реакцию). Этот препарат фермента использовали для оценки включения и для реакций секвенирования, описанных в примерах III-V. Для примеров II и VI фермент очищали в соответствии с методами, описанными у Lawyer и др. (см. выше).
Пример II
Анализ сравнительной эффективности включения
Относительную способность G46D- и G46D, E615G-Taq включать рНТФ определяли путем измерения количества [α-32Р]рНТФ, которое каждый из ферментов может включить на ДНК-матрице из активированной спермы лосося при ограниченной концентрации этого фермента. Для количественной оценки включения рАТФ реакционную смесь готовили таким образом, чтобы конечные концентрации в 50 мкл реакционной смеси составляли 12,5 мкг ДНК активированной спермы лосося, полученной по описанной ниже методике, 200 мкМ каждого из дЦТФ, дГТФ и дТТФ (фирма Perkin Elmer, Норуолк, шт. Коннектикут), 100 мкМ [α-32Р]рАТФ, 1 мМ β-меркаптоэтанола, 25 мМ N-трис[гидроксиметил]метил-3-аминопропансульфоновой кислоты (TAPS), pH 9,5 при 20°С, 50 мМ КСl и 2,25 мМ MgCl2.
Аналогичную смесь для анализа получали для количественной оценки включения рЦТФ, рГТФ и рУТФ. В каждом случае рНТФ были радиоактивно меченными и присутствовали в концентрации 100 мкМ, а каждый из трех остальных дНТФ (дАТФ, дГТФ и дТТФ для рЦТФ, дАТФ, дЦТФ и дТТФ для рГТФ и дАТФ, дЦТФ и дГТФ для рУТФ) присутствовал в концентрации 200 мкМ. В качестве стандарта также измеряли включение каждым ферментом соответстующего [α-32Р]дНТФ. Смесь для анализа в этих опытах была аналогична таковой, применяемой в опыте по включению рНТФ, которая описана выше, за исключением того, что каждый [α-32Р]рНТФ был заменен 100 мкМ соответствующего [α-32Р]дНТФ. Неочищенную ДНК спермы лосося в количестве 1 г/л, полученную от фирмы Worthington Biochemical (Фрихолд, шт. Нью Йорк), активировали путем инкубации в 10 мМ трис-HCl, рН 7,2, 5 мМ MgCl2 при температуре 2-8°С в течение 96 часов. Затем добавляли ЭДТК и NaCl до концентрации 12,5 мМ и 0,1 М соответственно. Затем ДНК экстрагировали фенолом/хлороформом и далее осаждали этанолом и ресуспендировали в 10 мМ трис, 1 мМ ЭДТК, рН 7,5. Затем препарат активированной ДНК подвергали диализу по отношению к этому же буферу.
45 мкл каждой реакционной смеси разливали в виде аликвотных количеств в пять пробирок объемом 0,5 мл (например, в пробирки Эппендорфа), внося в каждую 5’-меченные предшественники нуклеотидов. Таким образом, каждую полимеразу G46D-Taq и G46D, E615G-Taq оценивали дважды, при этом одну пробирку оставляли в качестве отрицательного контроля. Смесь, полученную в результате реакции полимеризации, из каждого опыта в двух пробирках смешивали сначала с 5 мкл либо полимеразы G46D-Taq (0,02 единицы), либо G46D, E615G-Taq (0,002 единицы). В качестве контроля уровня фона к реакционной смеси, применяемой в качестве отрицательного контроля, вместо фермента добавляли 5 мкл буфера для разбавления фермента.
Каждую реакционную смесь в течение короткого интервала времени центрифугировали и инкубировали в течение 10 минут при температуре 75°С. Реакции прекращали, добавляя 10 мкл 60 мМ ЭДТК, и хранили на льду. Для каждого образца аликвоты объемом 50 мкл из 60 мкл реакционной смеси разбавляли 1 мл 2 мМ ЭДТК, 50 мкг/мл фрагментированной ДНК спермы лосося. ДНК осаждали с помощью трихлоруксусной кислоты (ТХУ), используя стандартные методы и собирая на фильтровальный диски типа GF/C (фирма Whatman, Кент, Великобритания). Количество включенного [α-32Р]-меченного нуклеотида или рибонуклеотида количественно оценивали с помощью жидкостной сцинтилляционной спектрометрии и затем подсчитывали количество включенных пмолей. Количество пмолей каждого рНТФ, включенное каждым ферментом, соотносили с количеством пмолей соответствующего [α-32Р]дНТФ, включенного каждым ферментом. В табл.3 приведены полученные данные.
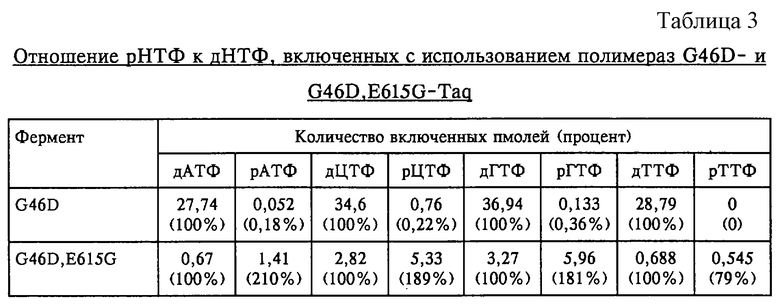
Эти результаты показывают, что полимераза G46D, E615G включает рибонуклеотиды более чем в 500 раз эффективнее по сравнению с тем, как это может осуществлять полимераза G46D (например, эффективнее для рГТФ в 181:0,36=502 раза, для рЦТФ в 189:0,22=859 раз и для рАТФ в 210:0,18=1166 раз).
Таким образом, миссенс-мутация в гене полимеразы в кодоне 615 приводит к получению нового фенотипа: к получению термостабильной ДНК-полимеразы, обладающей способностью помимо дезоксирибонуклеотидов эффективно включать рибонуклеотиды.
Пример III
Анализ сравнительной эффективности включения 3’-дезоксиАТФ (кордицепина)
Относительную способность полимераз G46D-; G46D, E615G-; G46D, E615G, F667Y- и G46D, F667Y-Taq включать 3’-дезоксиаденозин-5’-трифосфат (кордицепинтрифосфат) определяли путем измерения количества [α-32Р]кордицепинтрифосфата, которое каждый фермент может включить на ДНК-матрице из активированной спермы лосося при ограниченной концентрации этого фермента. Для количественной оценки включения [α-32Р] кордицепинтрифосфата опыт проводили таким образом, чтобы конечные концентрации в 50 мкл реакционной смеси составляли: 12,5 мкг ДНК активированной спермы лосося, 200 мкМ каждого из дЦТФ, дГТФ и дТТФ, 50 мкМ дАТФ (фирма Perkin Elmer), 50 мкМ [α-32P]-3’дАТФ/3’-дАТФ (фирма New England Nuclear, Sigma), 1 мМ β-меркаптоэтанола, 25 мМ N-трис[гидроксиметил]метил-3-аминопропансульфоновой кислоты (TAPS), pH 9,5 при 20°С, 55 мМ КСl и 2,25 мМ MgCl2.
45 мкл каждой реакционной смеси разливали в виде аликвотных количеств в девять пробирок объемом 0,5 мл, т.е. каждую реакцию проводили либо с G46D-; G46D, E615G-; G46D, E615G, F667Y-; либо с G46D, F667Y-Taq в двух повторностях, при этом одну пробирку оставляли без фермента в качестве отрицательного контроля. Смесь, полученную в результате реакции полимеризации, для каждого опыта в двух пробирках смешивали сначала с 5 мкл (0,058 единиц) полимеразы G46D-Taq. Такую же операцию проводили с полимеразой G46D, E615G-Taq (0,0025 единиц), полимеразой G46D, E615G, F667Y-Taq (0,0034 единицы) или полимеразой G46D, F667Y-Taq (0,083 единицы). В качестве контроля уровня фона в одну оставшуюся пробирку вместо фермента вначале добавляли буфер для разбавления фермента.
Каждую реакционную смесь в течение короткого интервала времени центрифугировали и инкубировали в течение 10 минут при 75°С. Реакции прекращали, добавляя 10 мкл 60 мМ ЭДТК, и хранили на льду. Каждый образец аликвоты объемом 50 мкл из 60 мкл реакционной смеси разбавляли 1 мл 2 мМ ЭДТК, 50 мкг/мл фрагментированной ДНК спермы лосося. ДНК осаждали с помощью ТХУ, используя стандартные методы и собирая на фильтровальные диски типа GF/C (фирма Whatman, Кент, Великобритания). Количество включенного [α-32Р]-меченного нуклеотида оценивали с помощью жидкостной сцинтилляционной спектрометрии, после чего подсчитывали количество включенных пмолей. Количество пмолей [α-32Р] кордицепинтрифосфата, включенное каждым ферментом, соотносили с количеством единиц каждого фермента, примененного в опыте, для получения количества пмолей, включенных единицей фермента. В табл.4 приведены полученные данные.

Эти результаты показывают, что обе мутации E615G и F667Y необходимы для эффективного включения молекулы кордицепина в ДНК.
Пример IV
Секвенирование ДНК на основе щелочного разложения с использованием ДНК-полимеразы G46D, E615G-Taq.
В этом примере показано применение модифицированной полимеразы по изобретению для секвенирования на основе щелочного разложения с использованием ДНК, частично замещенной рНТФ. Отношение рНТФ к дНТФ в реакционных смесях составляло от 1:80 до 1:8. Реакции удлинения праймера осуществляли в буфере, состоящем из 50 мМ Bicine (N,N-бис(2-гидроксиэтил) глицин); рН 8,3), 25 мМ КОАс и 2,5 мМ MgCl2. Проводили четыре отдельные реакции, по одной для каждого из четырех рНТФ. Каждая реакционная смесь (50 мкл) содержала по 200 мкМ каждого из дАТФ, дЦТФ, дГТФ и дТТФ (фирма Perkin Elmer) и по 0,09 пмолей одноцепочечной ДНК-матрицы М13mр18 (фирма Perkin Elmer), которую отжигали с 5’-[32P]-меченным DG48 (Lawyer и др., 1993, PCR Methods and Applications 2: 275-287). Реакционные смеси также содержали 2,5, 2,5, 2,5 или 25 мкм рАТФ, рЦТФ, рГТФ или рУТФ соответственно.
Каждую из этих четырех реакций начинали путем добавления 7 единиц ДНК-полимеразы G46D, E615GB-Taq и инкубировали в течение 10 минут при температуре 75°С. Реакции прекращали добавлением 10 мкл 60 мМ ЭДТК и помещали на лед. 20 мкл каждой реакционной смеси добавляли к 80 мкл 50 мМ Bicine (рН 8,3), 25 мМ КОАс и 2,5 мМ MgCl2. Продукты расщепления получали добавлением 7 мкл 1 н. NaOH и инкубированием в течение 15 минут при температуре 98°С. Реакции нейтрализовали добавлением 7 мкл 1 н. НСl. Каждую реакционную смесь осаждали, добавляя 312 мкл 95%-ного этанола и 10 мкл 3 М ацетата натрия (рН 4,8). Реакционные смеси микроцентрифугировали в течение 15 минут для сбора осадка, супернатант удаляли, дебрис промывали 500 мкл 70%-ного этанола и сушили. Каждый дебрис ресуспендировали в 5 мкл 0,5-кратного стоп-буфера (поставляемого фирмой Perkin Elmer, Норуолк, шт. Коннектикут, который содержит 95% формамида, 20 мМ ЭДТК и 0,05% бромфенола синего), выдерживали при температуре 98°С в течение 3 минут и непосредственно загружали на секвенирующий гель для ДНК, состоящий из предварительно подвергнутого электрофорезу 6%-ного полиакриламида/8 M мочевины, и осуществляли электрофорез. Гель сушили и снимали на рентгеновскую пленку. Полученная пленка показала выраженную секвенирующую лестницу, которая дает избыток в 100 оснований правильной последовательности.
Пример V
Секвенирование ДНК с использованием ДНК-полимеразы G46D, E615G, F667Y-Taq и 3’-дезоксинуклеотидтрифосфатов
В этом примере описано применение модифицированной полимеразы G46D, E615G, F667Y-Taq для секвенирования ДНК с использованием 3’-дезоксинуклеотидтрифосфатов. Этот эксперимент осуществляли с использованием 3’-дезоксиАТФ; однако он также может быть осуществлен с использованием других 3’-дезоксинуклеотидов. Реакции удлинения праймера осуществляли в буфере, состоящем из 50 мМ Bicine (рН 8,3), 25 мМ КОАс и 2,5 мМ MgCl2. Каждая реакционная смесь (50 мкл) содержала по 200 мкМ каждого из дАТФ, дЦТФ, дГТФ и дТТФ (фирма Perkin Elmer) и по 0,09 пмолей одноцепочечной ДНК-матрицы М13mр18 (фирма Perkin Elmer), которую отжигали с 5’-[32Р]-меченным DG48 (Lawyer и др., 1993, PCR Methods and Applications 2: 275-287). Реакционные смеси также содержали 0, 0,1, 0,25, 0,5, 1 или 5 мкм 3’-дезоксиАТФ.
Каждую из реакций начинали путем добавления 7 единиц ДНК-полимеразы G46D, E615G, F667Y-Taq и инкубировали в течение 10 минут при 75°С. Реакции прекращали добавлением 10 мкл 60 мМ ЭДТК и помещали на лед. 30 мкл каждой реакционной смеси осаждали этанолом и ресуспендировали в стоп-буфере, выдерживали при температуре 98°С в течение 3 минут и непосредственно загружали на секвенирующий гель для ДНК, состоящий из предварительно подвергнутого электрофорезу 6%-ного полиакриламида/8 М мочевины, и осуществляли электрофорез. Гель сушили и снимали на рентгеновскую пленку. Полосы, которые содержали продукты реакции, проведенной в присутствии кордицепина, свидетельствовали о наличии ясно выраженных лестниц терминации. Полосы, содержащие наибольшее количество кордицепина, т.е. 5 мкМ, показали лестницу терминации, в которой в среднем полосы были короче по длине по сравнению с полосами, соответствующими более низким уровням кордицепина. Полосы, которые содержали продукты реакции, проведенной в отсутствии кордицепина, свидетельствовали о наличии продукта, имеющего полную длину, и показали отсутствие лестницы терминации. Эти результаты показывают, что мутантный фермент способен включать кордицепин, и включение этой молекулы в продукт удлинения праймера вызывает терминацию. Этот метод также может применяться для создания лестницы секвенирования ДНК с использованием 3’-дезоксиЦТФ, 3’-дезоксиГТФ и 3’-дезоксиУТФ.
Пример VI
ПЦР-секвенирование с окрашенным праймером с использованием ДНК-полимеразы G46D, E615G-Taq
Этот метод иллюстрирует применение модифицированной полимеразы по изобретению для секвенирования с окрашенным праймером с использованием рибонуклеозидтрифосфатов (рНТФ) в ПЦР при соотношении рНТФ:дНТФ не более 1:30. Проводили четыре отдельные реакции, каждую с использованием рНТФ. Реакции секвенирования на основе ПЦР проводили в буфере, состоящем из 25 мМ трис-НСl (рН 9), 5,0 мМ MgCl2 и 10%-ного глицерина (объем/объем). Каждая реакционная смесь также содержала по 500 мкМ каждого из дАТФ, дЦТФ, дГТФ и дТТФ (фирма Perkin Elmer), матрицу, представляющую собой 5×106 копий/мкл плазмиды pBSM13+ (фирма Stratagene), линеаризованной в результате обработки эндонуклеазой рестрикции XmnI, и 0,05 единиц/мкл ДНК-полимеразы G46D, E615G-Taq. Реакционные смеси рибо-АТФ (10 мкл) содержали 2,5 мкМ АТФ (фирма Pharmacia Biotech), 0,1 мкл праймера JOE M13 Reverse Dye Primer (фирма Perkin Elmer) и 0,1 мкМ праймера ASC46 (5’-CGCCATTCGCCATTCAG). Реакционные смеси рибо-ЦТФ (10 мкл) содержали 2,5 мкМ ЦТФ (фирма Pharmacia Biotech), 0,1 мкл праймера FAM M13 Reverse Dye Primer (фирма Perkin Elmer) и 0,1 мкМ праймера ASC46. Реакционные смеси рибо-ГТФ (20 мкл) содержали 2,5 мкМ ГТФ (фирма Pharmacia Biotech), 0,1 мкл праймера TAMRA M13 Reverse Dye Primer (фирма Perkin Elmer) и 0,1 мкМ праймера ASC46. Реакционные смеси рибо-УТФ (20 мкл) содержали 16 мкМ УТФ (фирма Pharmacia Biotech), 0,1 мкл праймера ROX M13 Reverse Dye Primer (фирма Perkin Elmer) и 0,1 мкМ праймера ASC46.
Каждую из четырех реакционных смесей помещали в предварительно нагретую (75°С) термоячейку для проведения ПЦР типа Perkin Elmer GeneAMP® PCR System 9600 и подвергали 30 циклам: выдержка при температуре 95°С в течение 10 секунд, при температуре 55°С в течение 10 секунд, постепенное понижение температуры до 65°С в течение 1 минуты и выдержка при 65°С в течение 5 минут. В результате каждой из реакций с участием рАТФ и рЦТФ получали по 6×1011 копий меченного красителем амплифицированного продукта длиной 300 пар оснований, а в результате реакций с участием рГТФ и УТФ получали по 1,2х1012 копий меченного красителем амплифицированного продукта длиной 300 пар оснований.
Для определения последовательности ДНК амплифицированных продуктов ПЦР без использования разделения с помощью ферментативных реакций секвенирования ДНК реакционные смеси объединяли, обрабатывали щелочью и нагревали, нейтрализовали и осаждали следующим образом. Объединяли 4 мкл каждой из реакционных смесей с использованием АТФ и ЦТФ и 8 мкл каждой из реакционных смесей с использованием ГТФ и УТФ. К объединенным реакционным смесям добавляли 2 мкл 0,25 М ЭДТК (рН 8,0) (конечная концентрация 10 мМ), 10 мкл 1 М NaOH (конечная концентрация 200 мМ) и 14 мкл Н2О, а затем инкубировали при температуре 95°С в течение 5 минут в термоячейке для проведения ПЦР типа Perkin Elmer GeneAMP® PCR System 9600 и нейтрализовали 10 мкм 1 М HCl. Затем объединенную реакционную смесь осаждали добавлением 150 мкл 95%-ного этанола с последующей инкубацией при температуре 4°С в течение 15 минут. Затем ее микроцентрифугировали в течение 15 минут при температуре 4°С для сбора осадка и супернатант удаляли путем аспирации. Дебрис промывали 300 мкл 70%-ного этанола, микроцентрифугировали в течение 5 минут, супернатант удаляли путем аспирации и дебрис сушили. Дебрис ресуспендировали в 6 мкл формамида, 50 мг/мл голубого декстрана (в 25 мМ ЭДТК, 5:1 (объем/объем)) и выдерживали при температуре 90°С в течение 3 минут. 1,5 мкл ресуспендированного дебриса непосредственно загружали на секвенирующий гель, состоящий из предварительно подвергнутого электрофорезу 5%-ного полиакриламида типа Long Ranger (фирма FMC BioProducts) с 6 М мочевиной. Затем проводили электрофорез и анализировали на ДНК-секвенаторе типа Perkin Elmer ABI Prism™ 377 DNA Sequencer в соответствии с инструкцией производителя. Автоматическое определение оснований на основе программного обеспечения для анализа последовательности Perkin Elmer ABI Prism™ 377 Sequencing Analysis показало более чем 99%-ную точность определения последовательности ДНК продукта длиной 300 пар оснований, амплифицированного с использованием ПЦР.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ТЕРМОСТАБИЛЬНАЯ ДНК-ПОЛИМЕРАЗА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 1998 |
|
RU2238978C2 |
| КОМПОЗИЦИИ, СПОСОБЫ И НАБОРЫ ДЛЯ СИНТЕЗА И ОБНАРУЖЕНИЯ НУКЛЕИНОВЫХ КИСЛОТ | 2017 |
|
RU2757416C2 |
| ТЕРМОСТАБИЛЬНАЯ ДНК-ПОЛИМЕРАЗА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ, ФРАГМЕНТ ВЫДЕЛЕННОЙ ДНК, СПОСОБ АМПЛИФИКАЦИИ ДНК, СПОСОБ ВВЕДЕНИЯ МЕТКИ В ДНК И СПОСОБ ОБРАТНОЙ ТРАНСКРИПЦИИ | 1997 |
|
RU2197524C2 |
| СПОСОБ СИНТЕЗА кДНК С ИСПОЛЬЗОВАНИЕМ ТЕРМОСТАБИЛЬНОЙ ДНК-ПОЛИМЕРАЗЫ | 1997 |
|
RU2198222C2 |
| СПОСОБ ГОМОГЕННОЙ ДЕТЕКЦИИ ПО МЕНЬШЕЙ МЕРЕ ОДНОГО ПРОДУКТА ОДНОЦЕПОЧЕЧНОЙ АМПЛИФИКАЦИИ | 2005 |
|
RU2460804C2 |
| ХИМИЧЕСКИ МОДИФИЦИРОВАННЫЙ ТЕРМОСТАБИЛЬНЫЙ ФЕРМЕНТ, СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ АМПЛИФИКАЦИИ СОДЕРЖАЩЕЙСЯ В ОБРАЗЦЕ НУКЛЕИНОВОЙ КИСЛОТЫ, РЕАКЦИОННАЯ СМЕСЬ ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ И НАБОР ДЛЯ ПРОВЕДЕНИЯ ЦЕПНОЙ РЕАКЦИИ | 1996 |
|
RU2174556C2 |
| СПОСОБ АМПЛИФИКАЦИИ ДНК, СПОСОБ КЛОНИРОВАНИЯ ВТОРОЙ кДНК И СПОСОБ ОБРАТНОЙ ТРАНСКРИПЦИИ ДНК | 1997 |
|
RU2297454C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ХАРАКТЕРИСТИК ПОСЛЕДОВАТЕЛЬНОСТИ МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ ПУТЕМ ОБНАРУЖЕНИЯ И ХАРАКТЕРИСТИКИ ВАРИАЦИЙ ЭТОЙ ПОСЛЕДОВАТЕЛЬНОСТИ | 1998 |
|
RU2264468C2 |
| ФЕРМЕНТАТИВНЫЙ СИНТЕЗ L-НУКЛЕИНОВЫХ КИСЛОТ | 2013 |
|
RU2704833C2 |
| СПОСОБ АМПЛИФИКАЦИИ НУКЛЕИНОВЫХ КИСЛОТ С ИСПОЛЬЗОВАНИЕМ ФОСФОРИЛГУАНИДИНОВЫХ ОЛИГОНУКЛЕОТИДОВ | 2017 |
|
RU2698134C2 |



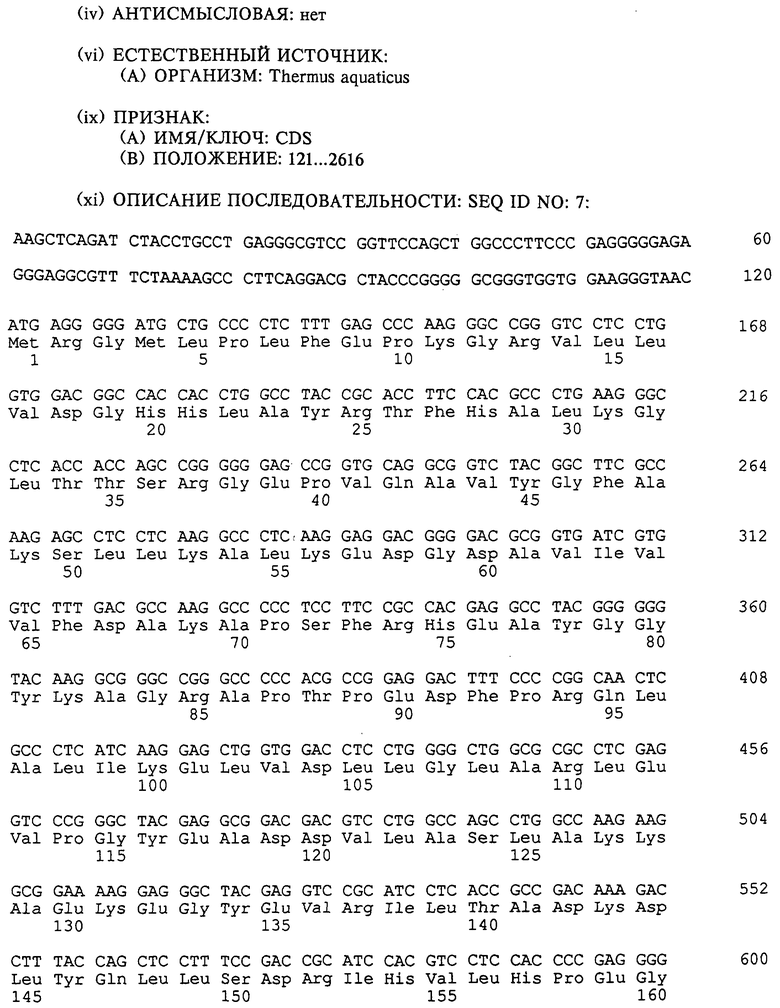
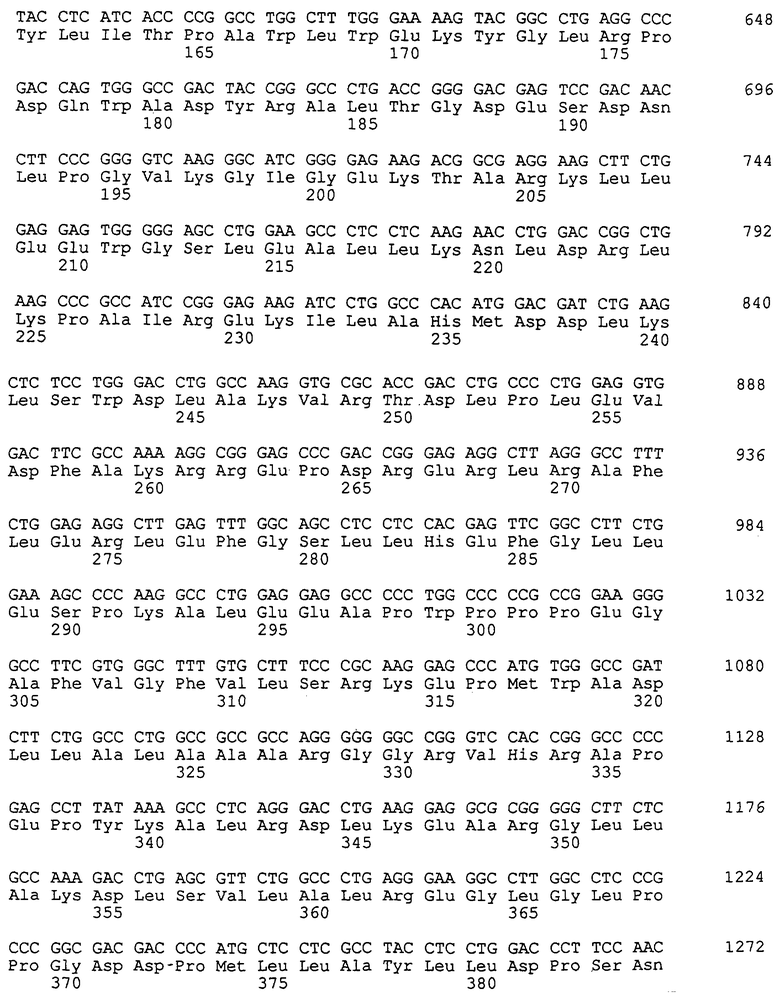
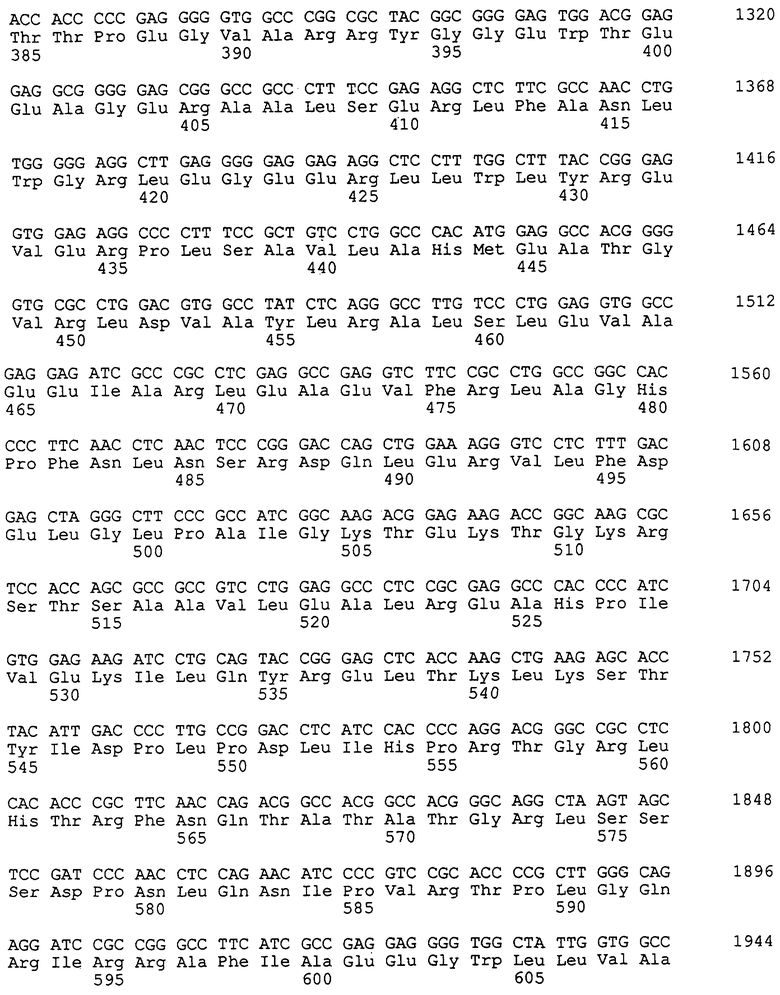
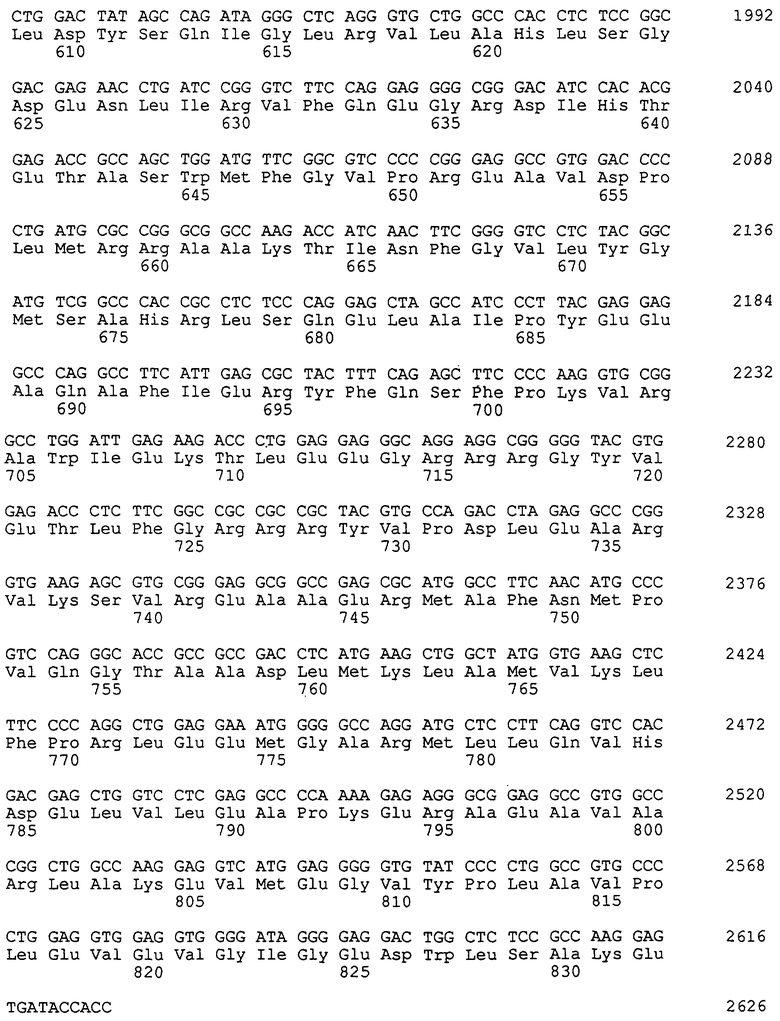



Изобретение относится к области молекулярной биологии и генной инженерии и может быть использовано в различных методах анализа нуклеиновых кислот, связанных с синтезом комплементарных ДНК-последовательностей. Предложена модифицированная форма термостабильной ДНК-полимеразы, получаемая путем замены остатка глутамата в консенсусной последовательности SerGlnIleGluLeuArgVal/Ile природного термостабильного фермента другим аминокислотным остатком и отбора мутеинов, проявляющих повышенную способность к включению в реакцию несвойственных субстратов, предпочтительно рНТФ. Описаны кодирующие модифицированную ДНК-полимеразу нуклеотидные последовательности, содержащие их векторы и штаммы Е.coli, трансформированные указанными векторами и обеспечивающие получение рекомбинантных форм мутантных ферментов. Предлагаемое применение новой термостабильной ДНК-полимеразы, в том числе в составе наборов и композиций, в реакции секвенирования нуклеиновой кислоты позволяет использовать новые подходы при анализе результатов исследования. 9 с. и 11 з.п. ф-лы, 4 табл.
(а) культивирование штамма по п.11 в условиях, которые усиливают экспрессию термостабильной ДНК-полимеразы, и (б) выделение термостабильной ДНК-полимеразы.
(а) подготовку несвойственного нуклеотида, предпочтительно рНТФ, и соответствующего обычного нуклеотида для реакции секвенирования ДНК, причем несвойственный нуклеотид и соответствующий обычный нуклеотид присутствуют в соотношении менее чем приблизительно 1:1; (б) обработку реакционной смеси со стадии (а) в присутствии термостабильной ДНК-полимеразы по любому из пп.1-8 в условиях, пригодных для удлинения праймера, с получением продуктов удлинения праймера, содержащих несвойственный нуклеотид; (в) обработку продуктов удлинения праймера со стадии (б) в условиях, пригодных для гидролиза этих продуктов удлинения праймера; (г) разделение продуктов реакции со стадии (в) и (д) определение последовательности нуклеиновой кислоты-мишени.
Конвенционный приоритет установлен от 06.08.1996 по пп.1-20 в соответствии с заявкой 60-023.376, поданной в патентное ведомство США.
| Автооператор | 1976 |
|
SU655506A1 |
| ЕР 0699760 А, 06.03.1996 | |||
| ЕР 0701000 А, 13.03.1996 | |||
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Автоматический огнетушитель | 0 |
|
SU92A1 |

