ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка заявляет приоритет по предварительной заявке на патент США №61/621975, поданной 9 апреля 2012 года, которая включена в данный документ посредством ссылки во всей своей полноте.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Технологии амплификации нуклеиновых кислот обеспечили средство для понимания сложных биологических процессов, выявления, идентификации и количественного определения патогенных и непатогенных организмов, судебно-криминалистических анализов, исследований взаимосвязи заболеваний и выявления трансгенных объектов - генетически модифицированных организмов и т.д. Полимеразная цепная реакция (ПЦР) представляет собой общеизвестную зависимую от термоциклирования технологию амплификации нуклеиновых кислот, используемую для амплификации ДНК, состоящую из циклов повторяемого нагревания и охлаждения реакционной смеси для плавления ДНК и ферментативной репликации ДНК с использованием ДНК-полимеразы. Количественная ПЦР в режиме реального времени (qPCR) представляет собой методику, используемую для количественного определения числа копий данной последовательности нуклеиновой кислоты в биологическом образце. В настоящее время в qPCR используют выявление продуктов реакции в режиме реального времени в ходе реакции и сравнивают профиль амплификации с амплификацией контрольных образцов, которые содержат известное количество нуклеиновых кислот в начале каждой реакции (или известное относительное количество нуклеиновых кислот по отношению к неизвестной тестируемой нуклеиновой кислоте). Результаты контрольных образцов используют для построения калибровочных кривых, как правило, на основе логарифмического участка калибровочных кривых реакции амплификации. Эти значения используют для интерполирования количества неизвестных нуклеиновых кислот на основе сравнения их кривых амплификации с количеством в стандартных контрольных образцах.
В дополнение к ПЦР, существуют системы амплификации, независимые от термоциклирования, или технологии изотермической амплификации нуклеиновых кислот, в том числе, без ограничения: реакция амплификации с внесением одноцепочечных разрывов и достройкой (NEAR), амплификация по типу «катящегося кольца» (RCA), геликаза-зависимая амплификация (HDA), амплификация с формированием петель (LAMP), амплификация с замещением цепей (SDA), транскрипционно-опосредованная амплификация (ТМА), самоподдерживающаяся репликация последовательностей (3SR), амплификация, основанная на последовательности нуклеиновых кислот (NASBA), изотермическая амплификация с одним праймером (SPIA), система Qβ-репликазы и рекомбиназная полимеразная амплификация (RPA).
NEAR-амплификация имеет сходство с термоциклированием при ПЦР. Подобно ПЦР, при NEAR-амплификации используются олигонуклеотидные последовательности, комплементарные целевым последовательностям, называемые праймерами в ПЦР и матрицами в NEAR. Кроме того, NEAR-амплификация целевых последовательностей приводит к логарифмическому увеличению числа копий целевой последовательности так же, как это происходит в стандартной ПЦР. В отличие от стандартной ПЦР, NEAR-реакция развивается изотермически. В стандартной ПЦР температуру повышают, чтобы разделить две цепи ДНК. В NEAR-реакции в целевую последовательность нуклеиновой кислоты вводят одноцепочечные разрывы в определенных сайтах одноцепочечных разрывов, присутствующих в тестируемом образце. Полимераза проникает в сайт одноцепочечного разрыва и начинает синтез цепи, комплементарной нуклеотидной целевой последовательности, в которую был внесен одноцепочечный разрыв (добавленной экзогенной ДНК), наряду с замещением существующей комплементарной цепи ДНК. Процесс репликации с замещением цепей устраняет необходимость в повышенной температуре. На этой стадии молекулы матрицы/праймера отжигают с замещенной комплементарной последовательностью из добавленной экзогенной ДНК. Теперь полимераза производит достраивание с 3'-конца матрицы, создавая цепь, комплементарную ранее замещенной цепи. Вторую олигонуклеотидную матрицу/праймер затем отжигают с вновь синтезированной комплементарной цепью и достраивают, получая ДНК-дуплекс, который содержит последовательность распознавания для фермента, вносящего одноцепочечный разрыв. Затем эта цепь подлежит внесению одноцепочечного разрыва с последующей достройкой с замещением цепи с помощью полимеразы, что ведет к получению ДНК-дуплекса, который имеет сайты одноцепочечного разрыва по обе стороны от исходной целевой ДНК. Сразу после его синтеза молекула продолжает амплифицироваться экспоненциально благодаря репликации замещенных цепей с новыми молекулами матриц. Кроме того, амплификация также протекает линейно от каждой молекулы продукта благодаря повторным операциям синтеза путем ник-трансляции в сайтах одноцепочечного разрыва, введенных с помощью матриц. Результатом является очень быстрое увеличение интенсивности амплификации целевого сигнала, намного более быстрое, чем в случае термоциклирования при ПЦР, при этом результаты амплификации появляются в течение менее чем десяти минут.
Однако количественное определение является проблематичным. Оптимальная производительность системы NEAR в режиме реального времени зависит от образования и амплификации специфического продукта. Известно, что в системах NEAR в реакциях с помощью ферментов образуются неспецифические фоновые продукты на существенных уровнях в дополнение к специфическому продукту. Эти фоновые продукты могут служить амплифицируемыми объектами, и их образование может препятствовать образованию специфического продукта. Хотя можно сконструировать детекторные зонды, специфичные к желаемой целевой молекуле (и, таким образом, специфический продукт можно выявить в смешанном фоне), неспецифические фоновые продукты на существенных уровнях блокируют компоненты реакции, которые могли быть в противном случае использованы для амплификации специфического продукта. Таким образом, блокировка компонентов реакции в связи с образованием неспецифических фоновых продуктов приводит к реакции, которая является субоптимальной. Это особенно нежелательно, когда целевая нуклеиновая кислота изначально характеризуется очень низким относительным содержанием и когда для надежного выявления целевой молекулы требуется высокооптимизированная реакция. Кроме того, субоптимальная реакция может не отображать истинное количество целевой нуклеиновой кислоты, даже если оно может быть выявлено. Было бы выгодно создавать оптимизированные NEAR-реакции, в которых устраняется амплификация неспецифических фоновых продуктов. Это могло бы обеспечить реакцию, которая подходит для количественного определения с помощью системы на основе калибровочных кривых или с помощью относительного количественного определения.
Кроме того, обычной практикой является оценка NEAR-реакций с помощью масс-спектрометрии. Высокие уровни фоновых продуктов могут мешать интерпретации данных масс-спектрометрии. Если, например, реакционная смесь содержит фоновые продукты, один или несколько продуктов, полученных в результате неспецифической амплификации (от родственных, но неодинаковых целевых молекул), и специфический продукт, было бы сложно выявить эти полученные из матрикса продукты среди фоновых продуктов. Устранение фоновых продуктов приводит к четкому определению производительности/специфичности конкретного анализа.
Дополнительно, высокие уровни фоновых продуктов могут препятствовать оптимальной амплификации в заведомо дуплексных или мультиплексных реакциях. В то время как многочисленные дифференциально меченные детекторные зонды совместимы с выявлением в режиме реального времени, до сих пор существует проблема ограничений для реагентов из-за образования неспецифических продуктов. Это особенно справедливо для дуплексных или мультиплексных реакций, поскольку в этих реакциях используется более чем две матрицы/праймера, которые потенциально могут формировать смешанные группы фоновых продуктов. Реакционная система NEAR, в которой устраняется амплификация фоновых продуктов, также обеспечивает условия для выявления в заведомо дуплексных или мультиплексных реакциях в режиме реального времени. Было бы очень выгодным обеспечить средство для устранения амплифицируемых фоновых продуктов, максимизируя, таким образом, потенциал образования специфических продуктов в NEAR-реакциях. Было бы желательно, если бы количественный результат мог быть представлен благодаря точному контролю хода реакции в режиме реального времени.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Как описано ниже, в настоящем изобретении представлены композиции и способы для выявления целевого олигонуклеотида в образце в режиме реального времени, которые снижают или устраняют образование фоновых продуктов, позволяя проводить количественное определение целевого олигонуклеотида в образце. Эти способы совместимы с целевыми олигонуклеотидами, амплифицируемыми с помощью NEAR-реакции. Настоящее изобретение по меньшей мере частично основано на обнаружении того, что специфические продукты в одноплексных NEAR-реакциях могут быть образованы без образования фоновых продуктов. Реакционные композиции и способы предусматривают относительное количественное определение в неизвестных тестируемых образцах, дуплексные реакции и мультиплексные реакции и создание калибровочных кривых для абсолютного количественного определения неизвестных тестируемых образцов.
В одном аспекте настоящее изобретение обеспечивает способ количественного определения специфического продукта в реакции амплификации с внесением одноцепочечных разрывов и достройкой, при этом способ включает: приведение целевой молекулы нуклеиновой кислоты в контакт в практически изотермических условиях с полимеразой с недостатком экзонуклеазной активности, двумя или более олигонуклеотидными праймерами/матрицами, каждый из которых специфически связывается с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты, ферментом, вносящим одноцепочечный разрыв, и выявляемым полинуклеотидным зондом, где каждый из олигонуклеотидного праймера/матрицы имеет один или несколько 2'-модифицированных нуклеотидов в последовательности, комплементарной целевой молекуле нуклеиновой кислоты; образование ампликонов, имеющих по меньшей мере часть указанной целевой молекулы нуклеиновой кислоты; и выявление сигнала, специфичного для гибридизации олигонуклеотидного зонда с целевой молекулой нуклеиновой кислоты или ее ампликоном, где сигнал указывает на количество целевой молекулы нуклеиновой кислоты, присутствующей в образце, или ее ампликона.
В другом аспекте настоящее изобретение обеспечивает способ выявления множества различных продуктов реакции, получаемых в ходе одной реакции, при этом способ включает: приведение целевой молекулы нуклеиновой кислоты в контакт в практически изотермических условиях с полимеразой с недостатком экзонуклеазной активности, двумя или более олигонуклеотидными праймерами/матрицами, каждый из которых специфически связывается с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты, ферментом, вносящим одноцепочечный разрыв, и выявляемым полинуклеотидным зондом, где каждый из олигонуклеотидного праймера/матрицы имеет один или несколько 2'-модифицированных нуклеотидов в последовательности, комплементарной целевой молекуле нуклеиновой кислоты; образование ампликонов, имеющих по меньшей мере часть указанной целевой молекулы нуклеиновой кислоты; и выявление сигнала, специфичного для гибридизации олигонуклеотидного зонда с целевой молекулой нуклеиновой кислоты или ее ампликоном, где сигнал указывает на количество целевой молекулы нуклеиновой кислоты, присутствующей в образце, или ее ампликона.
В конкретном аспекте настоящее изобретение обеспечивает способ количественного определения специфического продукта в реакции амплификации с внесением одноцепочечных разрывов и достройкой, при этом способ включает: приведение целевой молекулы нуклеиновой кислоты в контакт в практически изотермических условиях с полимеразой с недостатком экзонуклеазной активности, двумя или более олигонуклеотидными праймерами/матрицами, каждый из которых специфически связывается с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты, ферментом, вносящим одноцепочечный разрыв, и выявляемым полинуклеотидным зондом, где каждый из олигонуклеотидных праймеров/матриц имеет по меньшей мере приблизительно 5 смежных нуклеотидов с 2'-O-метильной модификацией, которые расположены на 3'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты (например, 3'-конце олигонуклеотида), или прилегают к нему; образование ампликонов, имеющих по меньшей мере часть указанной целевой молекулы нуклеиновой кислоты; и выявление сигнала, специфичного для гибридизации олигонуклеотидного зонда с целевой молекулой нуклеиновой кислоты или ее ампликоном, где сигнал указывает на количество целевой молекулы нуклеиновой кислоты, присутствующей в образце, или ее ампликона.
В одном аспекте настоящее изобретение обеспечивает способ контроля реакции амплификации с внесением одноцепочечных разрывов и достройкой в режиме реального времени, при этом способ включает: приведение тестируемого образца в контакт в практически изотермических условиях с полимеразой с недостатком экзонуклеазной активности, двумя или более олигонуклеотидными праймерами/матрицами, каждый из которых специфически связывается с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты, ферментом, вносящим одноцепочечный разрыв, и выявляемым полинуклеотидным зондом, где каждый из олигонуклеотидных праймеров/матриц имеет один или несколько 2'-модифицированных нуклеотидов в последовательности, комплементарной целевой молекуле нуклеиновой кислоты; образование ампликонов, имеющих по меньшей мере часть указанной целевой молекулы нуклеиновой кислоты; и выявление сигнала в режиме реального времени с количественным определением, таким образом, целевой(целевых) молекулы(молекул) нуклеиновой кислоты.
В другом аспекте настоящее изобретение обеспечивает способ контроля в режиме реального времени целевой молекулы нуклеиновой кислоты в NEAR-реакции, при этом способ включает: приведение целевой молекулы нуклеиновой кислоты в контакт в практически изотермических условиях с полимеразой с недостатком экзонуклеазной активности, двумя или более олигонуклеотидными праймерами/матрицами, каждый из которых специфически связывается с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты, ферментом, вносящим одноцепочечный разрыв, специфичным для гетеродуплексов ферментом, вносящим одноцепочечный разрыв, и выявляемым полинуклеотидным зондом, где каждый из олигонуклеотидных праймеров/матриц имеет один или несколько 2'-модифицированных нуклеотидов в последовательности, комплементарной целевой молекуле нуклеиновой кислоты; образование ампликонов, имеющих целевую последовательность, связывающуюся с выявляемым олигонуклеотидным зондом; и выявление сигнала в режиме реального времени с количественным определением, таким образом, целевой молекулы нуклеиновой кислоты.
В еще одном аспекте настоящее изобретение обеспечивает способ контроля в режиме реального времени целевой молекулы нуклеиновой кислоты в тестируемом образце, при этом способ включает: приведение целевой молекулы нуклеиновой кислоты в контакт в практически изотермических условиях с полимеразой, двумя или более олигонуклеотидными праймерами/матрицами, каждый из которых специфически связывается с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты, ферментом, вносящим одноцепочечный разрыв, репаративным ферментом или ферментом для исправления ошибок и выявляемым полинуклеотидным зондом, где каждый из олигонуклеотидных праймеров/матриц имеет один или несколько 2'-модифицированных нуклеотидов в последовательности, комплементарной целевой молекуле нуклеиновой кислоты; образование ампликонов, имеющих целевую последовательность, связывающуюся с выявляемым олигонуклеотидным зондом, и выявление сигнала в режиме реального времени с количественным определением, таким образом, целевой молекулы нуклеиновой кислоты.
В еще одном аспекте настоящее изобретение обеспечивает набор для выявления целевой последовательности в NEAR-реакции, при этом набор содержит один или несколько олигонуклеотидных праймеров/матриц, которые специфически связываются с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты и имеют один или несколько 2'-модифицированных нуклеотидов в последовательности, комплементарной целевой молекуле нуклеиновой кислоты, и инструкцию для применения олигонуклеотидного праймера/матрицы в способах по настоящему изобретению.
В одном аспекте настоящее изобретение обеспечивает выделенный олигонуклеотид, имеющий в направлении 5'-3' первый участок и второй участок, где первый участок имеет последовательность распознавания для фермента, вносящего одноцепочечный разрыв; где второй участок имеет по меньшей мере 9 или более нуклеотидов (например, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 или более смежных нуклеотидов), специфически связывающихся с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты; и где второй участок имеет один или несколько 2'-модифицированных нуклеотидов. В других вариантах осуществления выделенный олигонуклеотид является одним из представленных на фигуре 1.
В различных вариантах осуществления аспектов, определенных в данном документе, олигонуклеотид (например, олигонуклеотидный праймер/матрица, выделенный олигонуклеотид) содержит модифицированный нуклеотид, в том числе 2'-модифицированный нуклеотид. В различных вариантах осуществления любого аспекта, определенного в данном документе, 2'-модификация представляет собой один или несколько из следующего: 2'-O-метил, 2'-метоксиэтокси, 2'-фтор, 2-гидроксил, 2-аллил, 2'-O-[2-(метиламино)-2-оксоэтил], 4'-тио, 4, -СН2-O-2'-мостик, 4'-(СН2)2-O-2'-мостик, 2'-LNA, 2'-алкил и 2'-O-(N-метилкарбамат), или модифицированный нуклеотид содержит аналог основания. В различных вариантах осуществления любого аспекта, определенного в данном документе, один или несколько 2'-модифицированных нуклеотидов расположены на 3'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты (например, на 3'-конце олигонуклеотида), или прилегают к нему. В других вариантах осуществления любого аспекта, определенного в данном документе, один или несколько 2'-модифицированных нуклеотидов расположены на 5'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты. В различных вариантах осуществления любого аспекта, определенного в данном документе, один или несколько 2'-модифицированных нуклеотидов, расположенных на 5'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты, отделены от сайта одноцепочечного разрыва с помощью 1, 2, 3, 4, 5 или более немодифицированных нуклеотидов. В различных вариантах осуществления любого аспекта, определенного в данном документе, два или более 2'-модифицированных нуклеотида являются смежными (2, 3, 4, 5 или более). В других вариантах осуществления любого аспекта, определенного в данном документе, два или более 2'-модифицированных нуклеотида чередуются с немодифицированными нуклеотидами. В различных вариантах осуществления любого аспекта, определенного в данном документе, последовательность распознавания для фермента, вносящего одноцепочечный разрыв, представляет собой 5'-GAGTC-3'. В различных вариантах осуществления любого аспекта, определенного в данном документе, 5 смежных нуклеотидов с 2'-O-метильной модификацией расположены на 3'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты (например, на 3'-конце олигонуклеотида), или прилегают к нему. В других вариантах осуществления любого аспекта, определенного в данном документе, 5 смежных нуклеотидов с 2'-O-метильной модификацией расположены на 5'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты. В других вариантах осуществления любого аспекта, определенного в данном документе, 2 или более нуклеотида с 2'-O-метильной модификацией, чередующихся с немодифицированными нуклеотидами, расположены на 5'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты (т.е. конкретному целевому участку).
В различных вариантах осуществления любого аспекта, определенного в данном документе, на этапе выявления не выявляется ампликон нецелевой молекулы. В различных вариантах осуществления любого аспекта, определенного в данном документе, способ выполняют в режиме реального времени. В определенных вариантах осуществления любого аспекта, определенного в данном документе, этап образования ампликонов выполняют в режиме реального времени (например, для определения количества целевой молекулы, присутствующей в реакционной смеси).
В различных вариантах осуществления любого аспекта, определенного в данном документе, способ представляет собой полуколичественный и/или количественный способ порогов для определения количества молекулы нуклеиновой кислоты, присутствующей в биологическом образце до амплификации. В различных вариантах осуществления любого аспекта, определенного в данном документе, расположение одного или нескольких 2'-модифицированных нуклеотидов ближе к 5'-концу последовательности, комплементарной целевой молекуле нуклеиновой кислоты, увеличивает время выявления в амплификации. В различных вариантах осуществления любого аспекта, определенного в данном документе, способ дополнительно включает применение олигонуклеотидных праймеров/матриц в определенных соотношениях для обеспечения повышенной степени выявления продуктов реакции, получаемых из различных количеств исходного целевого материала. Было установлено, что увеличение соотношения олигонуклеотидных праймеров/матриц, имеющих один или несколько 2'-модифицированных нуклеотидов на 3'-конце последовательности распознавания, и олигонуклеотидных праймеров/матриц, имеющих один или несколько 2'-модифицированных нуклеотидов на 5'-конце последовательности распознавания, сужает кривую сигнала и изменяет наклон кривой.
В различных вариантах осуществления любого аспекта, определенного в данном документе, способ дополнительно включает применение модификатора скорости амплификации для обеспечения повышенной степени выявления продуктов реакции, получаемых из различных количеств исходного целевого материала. В различных вариантах осуществления любого аспекта, определенного в данном документе, целевая молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты ДНК или РНК. В различных вариантах осуществления любого аспекта, определенного в данном документе, выявляемый зонд представляет собой SYBR Green или молекулярный маяк. В различных вариантах осуществления любого аспекта, определенного в данном документе, выявляемый зонд представляет собой неамплифицируемый выявляемый полинуклеотидный зонд, имеющий по меньшей мере 10 нуклеотидов, комплементарных целевой последовательности, выявляемый фрагмент и молекулу, подавляющую активность полимеразы, где молекула, подавляющая активность полимеразы, предотвращает амплификацию зонда с помощью полимеразы в условиях, поддерживающих полимеразную активность в остальных случаях.
В различных вариантах осуществления любого аспекта, определенного в данном документе, тестируемый образец содержит патоген. В различных вариантах осуществления любого аспекта, определенного в данном документе, патоген представляет собой вирус, бактерию, дрожжи или гриб. В различных вариантах осуществления любого аспекта, определенного в данном документе, тестируемый образец представляет собой биологический образец. В различных вариантах осуществления любого аспекта, определенного в данном документе, биологический образец представляет собой клетку, образец ткани или биологическую жидкость (например, мочу, сперму, влагалищный секрет или кал). В различных вариантах осуществления любого аспекта, определенного в данном документе, тестируемый образец представляет собой пробу из окружающей среды.
Настоящее изобретение обеспечивает композиции и способы для выявления целевой молекулы нуклеиновой кислоты, амплифицированной с помощью NEAR-реакции. Композиции и изделия, определенные настоящим изобретением, выделяли или иным образом изготовляли с использованием примеров, представленных ниже. Другие отличительные признаки и преимущества настоящего изобретения будут очевидны из подробного описания и из формулы изобретения.
Определения
В данном раскрытии "включает", "включающий", "содержащий" и "имеющий" и т.п. могут иметь значение, определенное для них в патентном праве США, и могут означать "включает в себя", "включающий в себя" и тому подобное; "состоящий по существу из" или "состоит по существу из" также имеет значение, определенное в патентном праве США, и термин является неограничивающим, позволяя присутствие большего, чем то, что упомянуто, до тех пор, пока основные или новые характеристики того, что упомянуто, не изменятся благодаря присутствию большего, чем то, что упомянуто, но исключает варианты осуществления из предшествующего уровня техники.
Под "молекулой, подавляющей активность полимеразы" подразумевают фрагмент, связанный с полинуклеотидной матрицей/праймером, который предотвращает или существенно снижает продвижение полимеразы по полинуклеотидной матрице. Предпочтительно, фрагмент встроен в полинуклеотид. В одном предпочтительном варианте осуществления фрагмент предотвращает продвижение полимеразы по матрице.
Под "полимеразной достройкой" подразумевают продвижение полимеразы вперед от доступной 3'-гидроксильной группы, при котором происходит встраивание поступающих мономеров по принципу комплементарности по отношению к противоположным им нуклеотидам в полинуклеотидной цепи матрицы.
Под "полимеразой с недостатком экзонуклеазной активности" подразумевают ДНК-зависимую ДНК-полимеразу или РНК-зависимую ДНК-полимеразу, которая не обладает 5'-3'-экзонуклеазной активностью или которая обладает такой активностью на практически невыявляемых уровнях.
Под "аддуктом нуклеотида" подразумевают фрагмент, который ковалентно связан или иным образом присоединен к стандартному нуклеотидному основанию.
Применяемый в данном документе термин "выявляемый полинуклеотидный зонд" относится к любому по меньшей мере частично одноцепочечному полинуклеотиду, меченному выявляемым фрагментом с участком последовательности, комплементарным по меньшей мере одной цепи целевой последовательности, который высвобождает выявляемый сигнал из выявляемого фрагмента при связывании с целевой последовательностью, тогда как образование сигнала этим выявляемым фрагментом зависит от расщепления выявляемого полинуклеотидного зонда посредством неспецифической 5'-3'-экзонуклеазной активности. Примером "выявляемого полинуклеотидного зонда", применяемого в данном документе, является, без ограничения, флуоресцентный зонд типа молекулярного маяка, описанный в предшествующем уровне техники.
Применяемый в данном документе термин "нуклеиновая кислота" относится к дезоксирибонуклеотидам, рибонуклеотидам или модифицированным нуклеотидам и полимерам на их основе в одно- или двухцепочечный форме. Термин охватывает нуклеиновые кислоты, содержащие известные аналоги нуклеотидов или модифицированные каркасные остатки или связи, которые являются синтетическими, встречающимися в природе и не встречающимися в природе, которые имеют такие же свойства связывания, как и эталонная нуклеиновая кислота, и которые превращаются в ходе метаболизма аналогично эталонным нуклеотидам. Примеры таких аналогов включают, без ограничения, 2'-модифицированные рибонуклеотиды (например, 2'-O-метилрибонуклеотиды, 2'-F-нуклеотиды).
Применяемый в данном документе "модифицированный нуклеотид" относится к нуклеотиду, который имеет одну или несколько модификаций нуклеозида, нуклеинового основания, пентозного кольца или фосфатной группы. Например, модифицированные нуклеотиды исключают рибонуклеотиды, содержащие аденозинмонофосфат, гуанозинмонофосфат, уридинмонофосфат и цитидинмонофосфат, и дезоксирибонуклеотиды, содержащие дезоксиаденозинмонофосфат, деокси-гуанозинмонофосфат, дезокситимидинмонофосфат и дезоксицитидинмонофосфат. Модификации включают таковые, встречающиеся в природе, которые являются результатом модификации ферментами, модифицирующими нуклеотиды, такими как метилтрансферазы. Модифицированные нуклеотиды также включают синтетические или не встречающиеся в природе нуклеотиды. Нуклеотиды с синтетическими или не встречающимися в природе модификациями в включают в себя таковые с 2'-модификациями, например, содержащие 2'-алкил, такой как 2'-O-метил и 2'-метоксиэтокси, 2'-фтор, 2'-гидроксил (РНК), 2'-аллил, 2'-O-[2-(метиламино)-2-оксоэтил], 4'-тио, 4'-СН2-O-2'-мостик, 4'-(СН2)2-O-2'-мостик, 2-LNA и 2'-O-(N-метилкарбамат), или таковые, содержащие аналоги оснований.
Под "заменой основания" подразумевают введение заместителя полимера на основе нуклеиновых оснований, которое не вызывает существенного нарушения гибридизации между комплементарными нуклеотидными цепями.
Под "специфическим продуктом" подразумевают полинуклеотидный продукт, полученный в результате гибридизации олигонуклеотидных матриц с комплементарной целевой последовательностью и последующей достройки целевой последовательности, опосредованной полимеразой.
Под "реакцией амплификации с внесением одноцепочечных разрывов и достройкой" подразумевают чередующиеся циклы внесения одноцепочечных разрывов и достройки, приводящие к амплификации полинуклеотида, представляющего интерес.
Под "практически изотермическими условиями" подразумевают нахождение при одной температуре или в узком диапазоне температур, существенные изменения в котором не происходят. В одном варианте осуществления реакцию, выполняемую в практически изотермических условиях, выполняют при температуре, которая изменяется лишь на приблизительно 1-5°С (например, изменяется на 1, 2, 3, 4 или 5 градусов). В другом варианте осуществления реакцию выполняют при одной температуре в пределах рабочих параметров используемого прибора.
Под "ферментом, вносящим одноцепочечный разрыв" подразумевают полипептид, способный распознавать конкретную структуру в двухцепочечных молекулах нуклеиновой кислоты и связываться с ней, а также разрушать фосфодиэфирную связь между соседними нуклеотидами в одной цепи при связывании с ее распознаваемой конкретной структурой с предоставлением, таким образом, свободной 3'-гидроксильной группы в концевом нуклеотиде выше сайта одноцепочечного разрыва, от которой может быть произведена достройка с помощью полимеразы с недостатком экзонуклеазной активности.
Под "сайтом одноцепочечного разрыва" подразумевают положение "разрушенной" фосфодиэфирной связи в одной цепи двухцепочечной молекулы нуклеиновой кислоты, подвергаемой гидролизу с помощью фермента, вносящего одноцепочечный разрыв.
Под "ампликоном" подразумевают полинуклеотид или множество полинуклеотидов, образующихся в ходе амплификации полинуклеотида, представляющего интерес. В одном из примеров ампликон образуют в ходе полимеразной цепной реакции.
Под "полуколичественным" подразумевают обеспечение оценки относительного количества на основе внутреннего контроля.
Под "количественным методом порогов" подразумевают обеспечение оценки количества на основе превышения либо отсутствия превышения количества сравнительного стандарта.
Под "модификаторами скорости амплификации" подразумевают средство, способное воздействовать либо на скорость полимеразной достройки, либо на скорость внесения одноцепочечных разрывов с помощью фермента, вносящего одноцепочечный разрыв, либо на то и другое.
Под "контролем реакции" подразумевают выявление хода реакции. В одном варианте осуществления контроль хода реакции включает в себя выявление полимеразной достройки и/или выявление завершенной NEAR-реакции.
"Выявление" относится к выявлению присутствия, отсутствия или количества аналита, подлежащего выявлению.
Под "выявляемым фрагментом" подразумевают композицию, которая при связывании с молекулой, представляющей интерес, делает последнюю выявляемой с помощью спектроскопических, фотохимических, биохимических, иммунологических или химических средств. Например, применимые метки включают радиоактивные изотопы, магнитные гранулы, металлические гранулы, коллоидные частицы, флуоресцентные красители, электронно-плотные реагенты, ферменты (например, обычно используемые в ELISA), биотин, дигоксигенин или гаптены.
Под "фрагментом" подразумевают часть молекулы нуклеиновой кислоты. Эта часть содержит предпочтительно по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90% от полной длины эталонной молекулы нуклеиновой кислоты или полинуклеотида. Фрагмент может содержать 5, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90 или 100 нуклеотидов.
"Гибридизация" означает образование водородных связей, которое может представлять собой уотсон-криковское, хугстиновское или обратное хугстиновское образование водородных связей между комплементарными нуклеиновыми основаниями. Например, аденин и тимин являются комплементарными нуклеиновыми основаниями, которые образуют пару путем образования водородных связей.
Под "выделенным полинуклеотидом" подразумевают нуклеиновую кислоту (например, ДНК), которая не содержит генов, которые во встречающемся в природе геноме организма, из которого получена молекула нуклеиновой кислоты по настоящему изобретению, фланкируют данный ген. Термин, следовательно, включает, например, рекомбинантную ДНК, которая встроена в вектор, в автономно реплицирующиеся плазмиду или вирус или в геномную ДНК прокариота или эукариота или которая существует в виде отдельной молекулы (например, в виде кДНК или фрагмента геномной ДНК или кДНК, полученных с помощью ПЦР или расщепления рестрикционной эндонуклеазой) независимо от других последовательностей. Кроме того, термин включает молекулу РНК, транскрибированную из молекулы ДНК, а также рекомбинантную ДНК, которая является частью гибридного гена, кодирующего дополнительную полипептидную последовательность.
Термины "выделенный", "очищенный" или "биологически чистый" относятся к материалу, в разной степени свободному от компонентов, которые обычно сопровождают его, как обнаруживается в его нативном состоянии. "Выделять" указывает на степень отделения от исходного источника или окружения. "Очищать" указывает на степень отделения, превышающую таковую для выделения. "Очищенный" или "биологически чистый" белок является в достаточной степени свободным от других материалов, так что любые примеси не оказывают существенного влияния на биологические свойства белка или не вызывают другие неблагоприятные последствия. То есть нуклеиновая кислота или пептид по настоящему изобретению являются очищенными, если они по сути свободны от клеточного материала, вирусного материала или культуральной среды, будучи полученными с помощью методик рекомбинантной ДНК, или химических предшественников или других химических веществ, будучи синтезированными химически. Чистоту и однородность, как правило, определяют с использованием методов аналитической химии, например, электрофореза в полиакриламидном геле или высокоэффективной жидкостной хроматографии. Термин "очищенный" может обозначать, что нуклеиновая кислота или белок дает по сути одну полосу в геле для электрофореза. Для белка, который может быть подвергнут модификациям, например, фосфорилированию или гликозилированию, различные модификации могут давать различные выделенные белки, которые можно очищать по отдельности.
Как применяется в данном документе, "получение", как и "получение средства", включает синтезирование, закупку или приобретение средства иным способом.
Под "эталонным" подразумевают стандартное или контрольное состояние. Как очевидно для специалиста в данной области, подходящим эталоном является тот, где элемент изменяют для того, чтобы определить эффект элемента.
Под "гибридизацией" подразумевают образование пары между комплементарными полинуклеотидными последовательностями (например, гена, описанного в данном документе) или их частями в условиях различной жесткости с формированием двухцепочечной молекулы. (См., например, Wahl G.М. и S.L. Berger (1987) Methods Enzymol. 152:399; Kimmel A.R. (1987) Methods Enzymol. 152:507).
Под "субъектом" подразумевают млекопитающее, в том числе, без ограничения, человека или млекопитающее, отличное от человека, такое как бык, лошадь, собака, овца или кошка.
Под "целевой молекулой нуклеиновой кислоты" подразумевают полинуклеотид, подлежащий анализу. Такой полинуклеотид может представлять собой смысловую или антисмысловую цепь целевой последовательности. Термин "целевая молекула нуклеиновой кислоты" также относится к ампликонам исходной целевой последовательности.
Диапазоны, представленные в данном документе, понимаются как подразумевающие все значения внутри диапазона. Например, диапазон от 1 до 50 понимают как включающий любое число, комбинацию чисел или поддиапазон из группы, включающей 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 или 50.
Если специально не оговорено или не очевидно из контекста, применяемый в данном документе термин "или" следует понимать как включающий. Если специально не оговорено или не очевидно из контекста, применяемые в данном документе формы единственного числа понимают как означающие единственное или множественное число.
Если специально не оговорено или не очевидно из контекста, применяемый в данном документе термин "приблизительно" следует понимать как обозначающий величину в диапазоне нормального допустимого отклонения в данной области техники, например, в пределах 2 стандартных отклонений от среднего значения. Термин "приблизительно" можно понимать как обозначающий величину в пределах 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1%, 0,5%, 0,1%, 0,05% или 0,01% от заявленного значения. Если иное не ясно из контекста, все числовые значения, представленные в данном документе, изменяются с помощью термина "приблизительно".
Приведение перечня химических групп в любом определении переменной в данном документе включает определения этой переменной в качестве любой отдельной группы или комбинации перечисленных групп. Приведение варианта осуществления для переменной или аспекта в данном документе включает данный вариант осуществления в качестве любого отдельного варианта осуществления или в комбинации с любыми другими вариантами осуществления или их частями.
Любые композиции или способы, представленные в данном документе, могут быть объединены с одним или несколькими из любых других композиций и способов, представленных в данном документе.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На фигуре 1 изображены иллюстративные структуры объектов, подавляющих активность полимеразы. Черный = стабилизирующая последовательность, синий = последовательность распознавания для фермента, вносящего одноцепочечный разрыв, зеленый = спейсерная последовательность для фермента, вносящего одноцепочечный разрыв, красный = последовательность распознавания, специфичная для целевой молекулы, А = аденин, Т = тимин, G = гуанин, С = цитозин, U = урацил, mX = основание РНК с 2'-O-метильной модификацией. Подчеркнутое(подчеркнутые) основание(основания) определяет(определяют) модифицированный сегмент последовательности.
На фигурах 2А-2С изображена оценка динамического диапазона синтетической многомерной целевой ДНК Clavibacter michiganensis sepidonicus (Cms). Показаны иллюстративные результаты титрования синтетической "многомерной" целевой ДНК Cms и выявления с помощью флуориметрического маяка. NEAR-анализ для Cms, выполняемый без ввода целевой нуклеиновой кислоты, указан как контроль без целевой молекулы (NTC). Фигура 2А представляет собой график, показывающий, что сигнал в контролях без целевой молекулы (NTC) подавлен в реакционных смесях, содержащих праймер/матрицу с 2'-O-метильной модификацией. Фигура 2В представляет собой график, показывающий, что калибровочная кривая отображает широкий динамический диапазон с применением реакционных смесей с матрицами с 2'-O-метильной модификацией. Фигура 2С представляет собой сравнение иллюстративных данных масс-спектрометрии, показывающее устранение неспецифических продуктов амплификации в контролях без целевой молекулы (NTC) в системе анализа Cms с применением праймеров/матриц с 2'-O-метильной модификацией (правая панель) по сравнению с немодифицированными праймерами/матрицами (левая панель). Для определения эффекта праймеров/матриц с 2'-O-метильной модификацией в отношении образования фоновых продуктов образцы (10 мкл) реакционных смесей из контроля без целевой молекулы, изображенного на фигуре 2А, проанализировали с помощью ВЭЖХ/масс-спектрометрии. Данные масс-спектрометрии четко демонстрируют, что в присутствии праймеров/матриц с 2'-O-метальной модификацией выявляли присутствие только ожидаемых видов молекул, и продукты из смешанного фона, образующиеся в присутствии немодифицированных праймеров/матриц, устранены.
На фигуре 3 изображено устранение матрицами/праймерами с 2'-O-метильной модификацией сигнала фона в NEAR-анализе с использованием выявления с помощью SYBR Green. Показаны иллюстративные данные амплификации с использованием праймеров/матриц 2'-O-метильной модификацией и устранение неспецифических продуктов амплификации. Фигура 3А представляет собой график, показывающий, что в контролях без целевой молекулы (NTC) наблюдался существенный сигнал, что указывало на образование фонового продукта в отсутствие целевой ДНК. Фигура 3В представляет собой график, показывающий, что в реакционных смесях, содержащих матрицы с 2'-O-метильной модификацией, сигнал в контролях без целевой молекулы (NTC) был подавлен.
На фигуре 4 изображено устранение матрицами/праймерами с 2'-O-метильной модификацией сигнала фона в NEAR-анализе с использованием выявления с помощью молекулярного маяка. Показаны иллюстративные данные амплификации с использованием праймеров/матриц с 2'-O-метильной модификацией и устранение неспецифических продуктов амплификации. Фигура 4А представляет собой график, показывающий, что в контролях без целевой молекулы (NTC) наблюдался существенный сигнал, что указывало на образование фонового продукта в отсутствие целевой ДНК. Фигура 4В представляет собой график, показывающий, что в реакционных смесях, содержащих матрицы с 2'-O-метильной модификацией, сигнал в контролях без целевой молекулы (NTC) был подавлен.
На фигуре 5 изображены иллюстративные объекты, подавляющие активность полимеразы, которые при помощи праймеров/матриц с 2'-O-метильной модификацией или определенных соотношений праймеров/матриц с 2'-O-метильной модификацией можно применять для регуляции как времени до выявления, так и эффективности реакции, производя, таким образом, "настройку" реакции. Показаны схематические изображения иллюстративных матриц/праймеров с 2'-O-метильной модификацией для настройки конкретной реакции, в том числе праймера/матрицы, имеющего блок из пяти нуклеотидов с 2'-O-метильной модификацией на 3'-конце (матрица "Концевая"; слева) и праймера/матрицы, имеющего блок из пяти нуклеотидов с 2'-O-метильной модификацией, начинающийся с 3-го нуклеотида после сайта одноцепочечного разрыва (матрица "Одноцепочечный разрыв +2"; справа). Каждое условие настройки включает конкретное соотношение прямых и обратных матриц в каждом наборе матриц, имеющих различные структуры.
На фигуре 6 изображены графики амплификации, демонстрирующие полезность матриц/праймеров с 2'-O-метильной модификацией для "настройки" конкретной реакции. Показаны иллюстративные данные амплификации с использованием праймеров/матриц с 2'-O-метильной модификацией. Все реакционные смеси (в двух повтори остях) содержали 10000 геномных эквивалентов ДНК Cms. Каждое условие настройки представляет конкретное соотношение прямых и обратных матриц в каждом наборе матриц, имеющих различные структуры. Красные круги демонстрируют сдвиг времени до выявления для каждого условия настройки. Кроме того, логарифмическая фаза для каждого условия сокращалась, и наклон кривой изменялся.
На фигуре 7 изображена конструкция праймеров/матриц из двух наборов (TS3 и TS6), применяемых в NEAR-анализе для амплификации фрагмента гена кукурузы ADH1. Специфичные для целевой молекулы участки в последовательностях праймеров/матриц из наборов TS3 и TS6 существенно длиннее (15-17 оснований), чем в праймерах/матрицах из наборов для типичных NEAR-анализов (от 9 до 12 оснований) из предшествующего уровня техники. В наборе праймеров/матриц TS3 перед блоком из 5 последовательных нуклеотидов с 2'-O-метильной модификацией, прилегающих к 3'-концу, находится вышерасположенный участок из нуклеотидов с 2'-O-метильной модификацией, чередующихся с немодифицированными нуклеотидами, начинающийся с нуклеотида с 2'-O-метильной модификацией на 5 или на 4 нуклеотида ниже сайта одноцепочечного разрыва, соответственно. В отличие от TS3, в каждом из праймеров/матриц из набора TS6 есть только пять нуклеотидов с 2'-O-метильной модификацией, которые образуют блок из последовательных нуклеотидов, прилегающих к немодифицированному 3'-концевому нуклеотиду.
На фигуре 8 показаны графики амплификации для анализа ADH1 с использованием двух наборов праймеров/матриц (TS3 и TS6), записанные в канале выявления для красителя SYBR Green.
На фигуре 9 показаны графики амплификации для реакционных смесей из того же анализа, записанные в канале для ROX.
На фигуре 10 изображены записанные графики амплификации для реакционных смесей из NTC-анализа ADH1. При сравнении результатов, показанных в фигурах 8 и 9, становится очевидным, что только набор праймеров/матриц TS3 обеспечивает получение ADH1-специфичного ампликона, тогда как сигнал, образуемый набором TS6, главным образом основан на неспецифической амплификации фоновых продуктов, выявляемых только с помощью SYBR Green.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем изобретении представлены композиции и способы, применимые для количественного определения целевой молекулы нуклеиновой кислоты в изотермической реакции. В конкретных вариантах осуществления настоящее изобретение обеспечивает композиции и способы для количественного определения целевой молекулы нуклеиновой кислоты в NEAR-реакции (например, в режиме реального времени).
Настоящее изобретение по меньшей мере частично основано на неожиданном обнаружении того, что олигонуклеотидные праймеры-матрицы, содержащие 2'-модифицированный нуклеотид (например, с 2'-O-метильной, 2'-фтормодификацией), снижают интенсивность незаконной амплификации с помощью производных ДНК-полимеразы I Bst с недостатком 5'-3'-экзонуклеазной активности или устраняют ее.
NEAR-реакция
NEAR-реакцию применяли в качестве реакции с анализом результатов по конечной точке, которая предусматривает неколичественное выявление целевых олигонуклеотидов. Обычный NEAR-анализ включает (1) целевую молекулу нуклеиновой кислоты; (2) две молекулы олигонуклеотидов, которые являются аналогами молекул праймеров в ПЦР, называемые "матрицами-праймерами", содержащие некоторое количество нуклеотидов, комплементарных целевой молекуле нуклеиновой кислоты, и сайт, который может быть расщеплен ферментом, вносящим одноцепочечный разрыв; (3) dNTP; (4) полимеразу с недостатком 5'-3'-экзонуклеазной активности, производящую замещение цепей; и (5) фермент, вносящий одноцепочечный разрыв. Современные способы количественного определения в NEAR-реакции, в частности, в режиме реального времени, являются неприемлемыми, отчасти из-за незаконной амплификации нецелевых молекул, присутствующих в образце, что может затруднять выявление целевых последовательностей в обычной NEAR-реакции. Например, существует постоянная нежелательная амплификация в NEAR-реакциях, которая приводит к образованию выявляемого сигнала в отсутствие целевой молекулы или с сигналами, которые неточно отражают количество целевой молекулы нуклеиновой кислоты, присутствующей в реакционной смеси. Хотя обеспечивается выявление продукта по конечной точке, обеспечить контроль реакции в режиме реального времени не удается.
Настоящее изобретение обеспечивает модифицированные олигонуклеотидные праймеры/матрицы, которые решают проблему точного определения количества целевой молекулы нуклеиновой кислоты в NEAR-реакции. Это особенно применимо для определения количества целевой молекулы нуклеиновой кислоты в NEAR-реакции в режиме реального времени. Настоящее изобретение по меньшей мере частично основано на обнаружении того, что олигонуклеотидные праймеры-матрицы, содержащие 2'-модифицированный нуклеотид (например, с 2'-O-метильной, 2'-фтормодификацией), снижают интенсивность незаконной амплификации или устраняют ее без предотвращения достройки этих модифицированных праймеров-матриц в целях амплификации специфического продукта. Олигонуклеотидные праймеры/матрицы по настоящему изобретению применимы в NEAR-реакциях, включающих один или несколько из упомянутых выше компонентов NEAR.
В других вариантах осуществления настоящее изобретение обеспечивает олигонуклеотидные праймеры-матрицы, содержащие 2'-модифицированный нуклеотид (например, с 2'-O-метильной, 2'-фтормодификацией), который расположен на 3'-конце праймера-матрицы или прилегает к нему. Неожиданно, нуклеотиды с 2'-O-метильной модификацией, расположенные в 3'-концевом участке праймера-матрицы, не только являются эффективными затравочными субстратами для производных ДНК-полимеразы I Bst с недостатком 5'-3'-экзонуклеазной активности в изотермических реакциях амплификации ДНК, но применение таких модифицированных праймеров-матриц полностью подавляет неспецифическую амплификацию с участием димеров праймеров. Это особенно неожиданно, поскольку общепринятый подход в области изотермической амплификации ДНК предполагает, что модифицированные нуклеотиды (например, рибонуклеотиды с 2'-O-метильной модификацией, немодифицированные рибонуклеотиды) могут быть введены только в 5'-концевой участок праймера/матрицы далеко от 3'-конца, поскольку размещение 2'-O-метила, а также рибонуклеотидов в пределах 6 нуклеотидов от 3'-конца праймера, как было продемонстрировано, ингибирует достройку праймера с помощью ДНК-полимераз (в качестве ссылки включена заявка на патент от Amersham и патент от Qiagen). Производные ДНК-полимеразы I Bst с недостатком 5'-3' экзонуклеазной активности, применяемые в NEAR и других методиках изотермической амплификации (LAMP), принадлежат к polA-типу бактериальных ДНК-полимераз, участвующих в процессах репарации ДНК с низкой точностью синтеза. В отличие от этого, высокоточную репликацию генома у бактерий катализируют холоферменты ДНК-полимеразы III типа DNAE и POLC, которые используют исключительно РНК-праймеры для инициирования репликации ДНК. В опубликованных источниках из предшествующего уровня техники различие между РНК- и ДНК-праймерами считалось одним из механизмов предотвращения вмешательства ферментов ДНК-полимераз I с высокой частотой ошибок в высокоточную репликацию генома. В связи с этим неожиданное обнаружение того, что производные ДНК-полимеразы I Bst могут эффективно использовать 2'-модифицированные рибонуклеотиды в качестве праймеров для синтеза ДНК, является примечательным и парадоксальным.
Конструкция праймера-матрицы
Иллюстративные структуры объектов, подавляющих активность полимеразы, в направлении 5'-3' содержат стабилизирующую последовательность, последовательность распознавания для фермента, вносящего одноцепочечный разрыв, спейсерную последовательность для фермента, вносящего одноцепочечный разрыв, и последовательность распознавания, специфичную для целевой молекулы, при этом последовательность распознавания, специфичная для целевой молекулы, содержит один или несколько 2'-модифицированных нуклеотидов (например, содержащих 2'-O-метил, 2'-метоксиэтокси, 2'-фтор, 2'-аллил, 2'-O-[2-(метиламино)-2-оксоэтил], 2'-гидроксил (РНК), 4'-тио, 4'-СН2-O-2'-мостик, 4'-(СН2)2-O-2'-мостик, 2'-LNA и 2'-O-(N-метилкарбамат)). Не ограничиваясь теорией, предположили, что встраивание одного или нескольких 2'-модифицированных нуклеотидов в участки распознавания делает эти модифицированные участки непригодными для того, чтобы служить в качестве матрицы для полимеразной достройки в неспецифических межмолекулярных и/или внутримолекулярных комплексах, образуемых посредством взаимодействий праймеров/матриц (например, в случае образования димеров праймеров), и, следовательно, ослабляет или устраняет сигнал фона в изотермической амплификации. 2'-модифицированный нуклеотид предпочтительно имеет основание, которое образует пару оснований с целевой последовательностью. В конкретных вариантах осуществления два или более 2'-модифицированных нуклеотида (например, 2, 3, 4, 5 или более 2'-модифицированных нуклеотидов) в участке распознавания, специфичном для целевой молекулы, являются смежными (например, блок из модифицированных нуклеотидов). В некоторых вариантах осуществления блок из 2'-модифицированных нуклеотидов расположен на 3'-конце участка распознавания, специфичного для целевой молекулы. В других вариантах осуществления блок из 2'-модифицированных нуклеотидов расположен на 5'-конце участка распознавания, специфичного для целевой молекулы. Если блок из 2'-модифицированных нуклеотидов расположен на 3'-конце участка распознавания, специфичного для целевой молекулы, 2'-модифицированные нуклеотиды могут быть отделены от сайта одноцепочечного разрыва с помощью одного или нескольких немодифицированных нуклеотидов (например, 2, 3, 4, 5 или более 2'-немодифицированных нуклеотидов). Заявители обнаружили, что расположение одного или нескольких 2'-модифицированных нуклеотидов или блока из 2'-модифицированных нуклеотидов изменяет кинетику амплификации. Если один или несколько 2'-модифицированных нуклеотидов или блок из 2'-модифицированных нуклеотидов расположены на 5'-конце участка распознавания или вблизи от него или близко к сайту одноцепочечного разрыва, в реакциях амплификации в режиме реального времени демонстрируется уменьшенное время до выявления. Кроме того, кривая сигнала сужается, и наклон кривой изменяется. Заявители также выявили, что в участках распознавания длиной свыше 12 нуклеотидов один блок из 5 последовательных 2'-модифицированных нуклеотидов является недостаточным для подавления неспецифической амплификации и, следовательно, весь участок распознавания, расположенный на расстоянии до 4 или 5 нуклеотидов ниже сайта одноцепочечного разрыва, должен быть замещен 2'-модифицированныгми нуклеотидами, чередующимися с модифицированными нуклеотидами.
В связанном варианте осуществления соотношения олигонуклеотидных праймеров/матриц, имеющих один или несколько 2'-модифицированных нуклеотидов, можно применять для изменения времени до выявления и/или эффективности реакции для "настройки" реакций, что приводит к предсказуемому контролю над кинетикой амплификации. Увеличение соотношения олигонуклеотидных праймеров/матриц, имеющих один или несколько 2'-модифицированных нуклеотидов на 3'-конце последовательности распознавания, и олигонуклеотидных праймеров/матриц, имеющих один или несколько 2'-модифицированных нуклеотидов на 5'-конце последовательности распознавания, сужает кривую сигнала и изменяет наклон кривой. Предпочтительным является наличие возможности для "настройки" реакции с обеспечением средства для регуляции как времени до выявления, так и эффективности реакции. Относительное количественное определение с использованием внутреннего контроля требует выполнения двух важных условий. Во-первых, целесообразно иметь возможность изменять время до выявления в реакции, создавая неконкурентные условия реакции. Таким образом, при воздействии на контрольную реакцию с целью получения возможности выявления в более поздний момент времени (относительно целевой молекулы, представляющей интерес) контрольная реакция не препятствует реакции со специфической целевой молекулой, представляющей интерес, даже если целевая молекула, представляющая интерес, изначально характеризуется низким относительным содержанием. Во-вторых, для обеспечения расчета истинного относительного содержания требуется, чтобы контрольная реакция и реакция со специфической целевой молекулой имели равноценную эффективность. Путем контроля эффективности каждой реакции с помощью условий "настройки" реакции можно делать равноценными, что позволяет проводить удовлетворительные расчеты для относительного количественного определения. Настройку реакций можно применять для обеспечения равноценности эффективности амплификации целевой нуклеиновой кислоты и амплификации эталонной нуклеиновой кислоты (например, внутреннего стандарта) в количественной ПЦР (qPCR). Кроме того, кривые амплификации целевой нуклеиновой кислоты и внутреннего стандарта могут быть изменены таким образом, что выявление их продуктов амплификации будет разделено во времени, с обеспечением при этом такой же эффективности амплификации целевой нуклеиновой кислоты и амплификации внутреннего стандарта. Благодаря применению конкретных комбинаций и соотношений олигонуклеотидных структур в реакционной смеси можно создать условия, которые позволяют настроить производительность реакции.
В различных вариантах осуществления пары праймеров/матриц сконструированы в конфигурации "стебель-петля". 5'-конец олигонуклеотидного праймера/матрицы содержит самокомплементарный участок, которая формирует по меньшей мере часть стебля. В некоторых вариантах осуществления настоящего изобретения стебель дополнительно охватывает по меньшей мере часть последовательности распознавания для фермента, вносящего одноцепочечный разрыв, или всю ее. В других различных вариантах осуществления настоящего изобретения последовательность распознавания для фермента, вносящего одноцепочечный разрыв, в праймерах-матрицах не является частью двухцепочечной структуры стебля, но находится в пределах главным образом одноцепочечной петли. Этот сайт распознавания для фермента, вносящего одноцепочечный разрыв, связан на 3'-конце с не имеющем вторичной структуры сайтом, включающим в себя сайт одноцепочечного разрыва, который связан на 3'-конце с последовательностью, комплементарной целевой последовательности. При необходимости последовательность, комплементарная целевой последовательности, может иметь вторичную структуру или может не иметь вторичной структуры. Наличие или отсутствие вторичной структуры, которая может содержать самокомплементарный участок, будет определяться для оптимизации конкретного NEAR-анализа.
В одном варианте осуществления способы по настоящему изобретению обеспечивают реакционную смесь для NEAR, которая содержит стандартные компоненты для NEAR, но также содержит фермент, способный вносить одноцепочечный разрыв в нуклеотид РНК, если он присутствует в гетеродуплексе с комплементарной цепью ДНК. В одном примере расщепляемый нуклеотид РНК будет присутствовать в нити из 4-15 нерасщепляемых нуклеотидов РНК (т.е. O-2-Ме-РНК) в направлении 5'-конца целевого комплементарного участка РТО, и 3'-конец олигонуклеотидной матрицы будет иметь 3'-концевой "кэп". Только после полной надлежащей гибридизации олигонуклеотидной матрицы молекула, расщепляющая гетеродуплекс (т.е. РНКаза Н), будет способна расщеплять основание РНК, предоставляя 3'-конец для достройки от него с помощью фермента ник-трансляции и позволяя проходить NEAR-реакции до завершения. Нарушение связывания с матрицей (димеры праймеров, частичная гибридизация с нецелевыми молекулами и т.д.) не будет приводить к формированию гетеродуплекса РНК-ДНК и, таким образом, предотвратит прохождение NEAR-реакции. Эти матрицы будут амплифицироваться только после связывания с комплементарной нуклеотидной последовательностью благодаря удалению 3'-"кэпа" для полимеразной достройки. Это приведет к повышению уровня специфичности и чувствительности NEAR-реакции.
Олигонуклеотидные матрицы по настоящему изобретению включены в реакционную смесь для NEAR, которая содержит (1) целевую молекулу нуклеиновой кислоты; (2) две молекулы олигонуклеотидных матриц, содержащие некоторое количество олигонуклеотидов, комплементарных целевой молекуле нуклеиновой кислоты, и сайт, который может быть расщеплен ферментом, вносящим одноцепочечный разрыв и содержит 4-15 нуклеотидов РНК, один из которых способен подвергаться действию РНКазы; (3) dNTP; (4) полимеразу, производящую замещение цепей; (5) фермент, вносящий одноцепочечный разрыв; и (6) фермент, вносящий одноцепочечный разрыв в РНК в гетеродуплексе ДНК-РНК, и 3'-концевой кэп для полимеразной достройки. Таким образом, настоящее изобретение обеспечивает способ применения этих компонентов для количественного определения целевой молекулы нуклеиновой кислоты.
Способ включает приведение целевой молекулы нуклеиновой кислоты в контакт в практически изотермических условиях с полимеразой, двумя олигонуклеотидными матрицами, каждая из которых специфически связывается с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты, ферментом, вносящим одноцепочечный разрыв, и ферментом, вносящим одноцепочечный разрыв в гетеродуплекс ДНК-РНК (например, РНКазой Н), и 3'-концевым кэпом для полимеразной достройки; образование выявляемого ампликона, содержащего по меньшей мере часть олигонуклеотидной матрицы, которая связывается с целевой последовательностью.
Целевые молекулы нуклеиновых кислот
Способы и композиции по настоящему изобретению применимы для идентификации целевой молекулы нуклеиновой кислоты в тестируемом образце. Целевые последовательности амплифицируют практически из любых образцов, которые содержат целевую молекулу нуклеиновой кислоты, включающих, без ограничения, образцы, содержащие грибы, споры, вирусы или клетки (например, прокариоты, эукариоты). В конкретных вариантах осуществления при помощи композиций и способов по настоящему изобретению выявляют Clavibacter michiganensis subsp. michiganensis, Clavibacter michiganensis subsp. sepedonicus, Pseudomonas syringae pv Tomato, Xanthomonas campestris pv Vesicatoria, Alternaria spp., Cladosporium spp., Fusahum oxysporum, Verticilium dahlia, Pseudomonas currugata, Erwina carotovora и Ralstonia solanacearwn. Иллюстративные тестируемые образцы включают биологические жидкости (например, кровь, сыворотку, плазму, амниотическую жидкость, мокроту, мочу, спинномозговую жидкость, лимфу, слезную жидкость, кал или желудочный сок), тканевые экстракты, культуральные среды (например, жидкость, в которой клетка, такая как клетка патогена, была выращена), пробы из окружающей среды, сельскохозяйственную продукцию или другие продукты питания и их экстракты, ДНК-метки для идентификации. При необходимости образец очищают перед включением в NEAR-реакцию с помощью любого стандартного способа, как правило, применяемого для выделения молекулы нуклеиновой кислоты из биологического образца.
В одном варианте осуществления олигонуклеотидные праймеры/матрицы амплифицируют целевую нуклеиновую кислоту патогена для выявления присутствия патогена в образце. Иллюстративные патогены включают грибы, бактерии, вирусы и дрожжи. Такие патогены могут быть выявлены с помощью идентификации молекулы нуклеиновой кислоты, кодирующей белок патогена, такой как токсин, в тестируемом образце. Иллюстративные токсины включают, без ограничения, афлатоксин, холерный токсин, дифтерийный токсин, токсин сальмонеллы, шига-токсин, токсин Clostridium botulinum, эндотоксин и микотоксин. Для путей применения в отношении окружающей среды тестируемые образцы могут включать воду, жидкие экстракты с воздушных фильтров, образцы почвы, строительные материалы (например, гипсокартон, потолочные плитки, стеновые плиты, ткани, обои и напольные покрытия), мазки из окружающей среды или любой другой образец.
В одном из вариантов осуществления, раскрытых в данном документе, олигонуклеотидные праймеры/матрицы амплифицируют целевую нуклеиновую кислоту растения, применяемого в качестве внутреннего контроля в экспериментах по молекулярной селекции, направленных на улучшение, например, устойчивости растений к засухе, устойчивости растений к гербицидам, к хищническому истреблению вредными насекомыми. Одним из примеров такой целевой нуклеиновой кислоты внутреннего контроля, применяемым на практике в данном документе, является ген ADH1 (алкогольдегидрогеназы 1) кукурузы.
Целевые молекулы нуклеиновых кислот включают в себя двухцепочечные и одноцепочечные молекулы нуклеиновых кислот (например, ДНК, РНК и другие полимеры на основе нуклеиновых оснований, известные в данной области техники, способные к гибридизации с молекулой нуклеиновой кислоты, описанной в данном документе). Молекулы РНК, применимые для выявления с помощью выявляемого олигонуклеотидного зонда или выявляемого олигонуклеотидного праймера/матрицы по настоящему изобретению, включают в себя, без ограничения, двухцепочечные и одноцепочечные молекулы РНК, которые содержат целевую последовательность (например, информационную РНК, вирусную РНК, рибосомальную РНК, транспортную РНК, микроРНК и предшественники микроРНК и миРНК или другие РНК, описанные в данном документе или известные в данной области техники). Молекулы ДНК, применимые для выявления с помощью выявляемого олигонуклеотидного зонда или олигонуклеотидного праймера/матрицы по настоящему изобретению включают в себя, без ограничения, двухцепочечную ДНК (например, геномную ДНК, плазмидную ДНК, митохондриальную ДНК, вирусную ДНК и синтетическую двухцепочечную ДНК). Целевые молекулы нуклеиновой кислоты в виде одноцепочечной ДНК включают, например, вирусную ДНК, кДНК и синтетическую одноцепочечную ДНК или другие типы ДНК, известные в данной области техники.
В целом, целевая последовательность для выявления имеет длину от 10 до 100 нуклеотидов (например, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100 нуклеотидов). Содержание GC в целевой молекуле нуклеиновой кислоты выбирают составляющим меньше, чем приблизительно 45, 50, 55 или 60%. Желательно выбирать целевую последовательность и ферменты, вносящие одноцепочечный разрыв, таким образом, что целевая последовательность не содержит сайтов одноцепочечных разрывов для любых ферментов, вносящих одноцепочечный разрыв, которые будут включены в реакционную смесь.
Выявляемые олигонуклеотидные зонды
Настоящее изобретение обеспечивает количественное определение целевых молекул нуклеиновых кислот или их ампликонов в NEAR-реакции с использованием неамплифицируемых выявляемых полинуклеотидных зондов, содержащих по меньшей мере одну молекулу, подавляющую активность полимеразы (например, модификацию нуклеотида или другой фрагмент, который делает олигонуклеотид способным к связыванию с целевой молекулой нуклеиновой кислоты, но неспособным поддерживать достройку матрицы с использованием выявляемого олигонуклеотидного зонда в качестве мишени). Не желая ограничиваться теорией, предположили, что наличие одного или нескольких фрагментов, которые не позволяют продвижение полимеразы, вероятно, вызывает подавление активности полимеразы посредством добавлений к каркасу олигонуклеотида, отличных от нуклеиновых кислот, или из-за потери скорости репликативной полимеразы (т.е. посредством С3-спейсера, поврежденных оснований ДНК, другого спейсерного фрагмента, оснований с модификацией O-2-Ме). Эти конструкции, таким образом, предотвращают или снижают интенсивность незаконной амплификации зонда в ходе NEAR-реакции. Это отличает их от обычных детекторных зондов, которые должны добавляться в конце NEAR-реакции для предотвращения их амплификации.
Обычные детекторные зонды оказались непрактичными для количественной оценки в NEAR-реакции в режиме реального времени. Если обычные детекторные зонды включены в NEAR-реакцию, эти обычные детекторные зонды амплифицируются одновременно с целевой молекулой. Амплификация этих детекторных молекул скрывает должные целевые ампликоны от выявления в связи с количеством исходных молекул детекторного зонда в начале реакции.
Настоящее изобретение обеспечивает неамплифицируемый выявляемый полинуклеотидный зонд, который содержит по меньшей мере одну молекулу, подавляющую активность полимеразы. Молекула, подавляющая активность полимеразы, по настоящему изобретению включает, без ограничения, модификацию нуклеотида или другой фрагмент, который блокирует достройку праймера-матрицы с помощью репликативных ДНК-полимераз, тем самым предотвращая амплификацию выявляемых молекул; но может обеспечить правильную гибридизацию или расположение нуклеотидов для целевой молекулы или амплифицированных копий целевой молекулы. В одном из вариантов осуществления выявляемый олигонуклеотидный зонд по настоящему изобретению содержит 3-углеродный спейсер (С3-спейсер), который предотвращает или снижает интенсивность незаконной амплификации детекторной молекулы.
В одном из вариантов осуществления выявляемый олигонуклеотидный зонд по настоящему изобретению представляет собой олигонуклеотид в форме шпильки, содержащий выявляемый фрагмент. В другом варианте осуществления неамплифицируемый выявляемый полинуклеотидный зонд представляет собой олигонуклеотид в форме шпильки, который содержит флуорофор на одном конце и гасящий краситель на противоположном конце. Петля шпильки содержит последовательность, комплементарную целевой последовательностью и способную к гибридизации с ней. Стебель шпильки образуют путем отжига комплементарных последовательностей плеч, расположенных по обе стороны от петли. Флуорофор и гасящая молекула ковалентно связаны на противоположных концах каждого плеча. Если выявляемый олигонуклеотидный зонд находится в шпилечной конфигурации, флуоресцентная и гасящая молекулы находятся близко друг к другу, что, таким образом, приводит к резонансному переносу энергии флуоресценции (FRET) и гашению флуоресценции флуорофора. Когда выявляемый олигонуклеотидный зонд встречается с целевой молекулой, происходит гибридизация; структура петли преобразуется в конформацию дуплекса с целевой молекулой, что вызывает разделение молекулы флуорофора и гасящей молекулы, в результате чего происходит флуоресценция (Tyagi et al. Nature Biotechnology 14: March 1996, 303-308).
Выявляемые олигонуклеотидные зонды являются специфичными для целевой последовательности. В одном из вариантов осуществления выявляемый олигонуклеотидный зонд содержит одно или несколько модифицированных нуклеотидных оснований, обладающих повышенным сродством связывания с комплементарным нуклеотидом. Примеры оснований включают, без ограничения, закрытые нуклеиновые, кислоты (LNA), амидиты нуклеиновых кислот с 2'-фтормодификацией и амидиты РНК с модификацией 2'ОМе (также функционирующие в качестве молекул, подавляющих активность полимеразы). Выявляемые олигонуклеотидные зонды по настоящему изобретению можно синтезировать с различными цветными флуорофорами и можно конструировать для гибридизации практически с любой целевой последовательностью. С учетом его исключительной специфичности неамплифицируемый выявляемый полинуклеотидный зонд по настоящему изобретению применяют для выявления одной целевой молекулы нуклеиновой кислоты в образце или применяют в комбинации с выявляемыми олигонуклеотидными зондами, каждый из которых связывает отличную целевую молекулу нуклеиновой кислоты. Соответственно, неамплифицируемые выявляемые полинуклеотидные зонды по настоящему изобретению можно применять для выявления одной или нескольких целевых молекул нуклеиновой кислоты в одной и той же реакции, что позволяет производить одновременное количественное определение этих целевых молекул. Настоящее изобретение охватывает применение таких флуорофоров в сочетании с выявляемыми олигонуклеотидными зондами, описанными в данном документе.
Применение неамплифицируемых выявляемых полинуклеотидных зондов
Неамплифицируемые выявляемые полинуклеотидные зонды применимы в способах для количественной оценки целевой молекулы нуклеиновой кислоты в реакции амплификации с внесением одноцепочечных разрывов и достройкой (NEAR). Способ включает приведение целевой молекулы нуклеиновой кислоты в контакт в практически изотермических условиях с полимеразой, двумя олигонуклеотидными праймерами/матрицами, каждый из которых специфически связывается с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты, ферментом, вносящим одноцепочечный разрыв, и выявляемым олигонуклеотидным зондом в присутствии подходящего буфера и dNTP; образование ампликонов, содержащих по меньшей мере часть указанной целевой молекулы нуклеиновой кислоты; и определение уровня целевой молекулы нуклеиновой кислоты, присутствующей в реакционной смеси, посредством количественной оценки олигонуклеотидного зонда, гибридизирующегося с целевой молекулой нуклеиновой кислоты, в режиме реального времени в ходе реакции на основе интенсивности флуоресценции молекул зонда в реакционной смеси. Предпочтительно, такие способы применимы для контроля NEAR в режиме реального времени.
В целом, неамплифицируемые выявляемые полинуклеотидные зонды по настоящему изобретению включены в реакционную смесь для NEAR, которая содержит (1) целевую молекулу нуклеиновой кислоты; (2) две молекулы олигонуклеотидных матриц, содержащие некоторое количество олигонуклеотидов, комплементарных целевой молекуле нуклеиновой кислоты, и сайт, который может быть расщеплен ферментом, вносящим одноцепочечный разрыв; (3) dNTP; (4) полимеразу, производящую замещение цепей; и (5) фермент, вносящий одноцепочечный разрыв. Таким образом, настоящее изобретение обеспечивает способ применения этих компонентов для количественной оценки целевой молекулы нуклеиновой кислоты.
NEAR-анализ
Настоящее изобретение обеспечивает выявление целевых молекул нуклеиновых кислот, амплифицированных в NEAR-анализе. Такой анализ известен в данной области техники и описан в данном документе. См., например, публикацию заявки на патент США №2009/0081670, заявку по РСТ №2009/012246 и патенты США №7112423 и №7282328, каждый из которых включен в данный документ во всей своей полноте. Полимеразы, применимые в способах, описанных в данном документе, способны катализировать встраивание нуклеотидов с достройкой от 3'-гидроксильного конца олигонуклеотида (например, олигонуклеотидного праймера/матрицы или другого праймера), связанного с целевой молекулой нуклеиновой кислоты. Такие полимеразы включают те, которые являются термофильными, и/или те, которые способны к замещению цепей. Полимеразы, применимые в способах, описанных в данном документе, не обладают 5'-3'-экзонуклеазной активностью, которая в противном случае разрушала бы замещенную цепь одноцепочечной нуклеиновой кислоты. Полимеразы также обладают активностью обратной транскриптазы (например, производные ДНК-полимеразы Bst (большой фрагмент), ДНК-полимераза Therminator, ДНК-полимераза Therminator II). Иллюстративные полимеразы включают, без ограничения, большие фрагменты ДНК-полимеразы I Bst, ДНК-полимеразу I Е. coli (фрагмент Кленова), фрагмент Кленова (3'-5'-экзо-), ДНК-полимеразу Т4, ДНК-полимеразу Т7, ДНК-полимеразу Deep VentR. (экзо-), ДНК-полимеразу Deep VentR, Therminator, ДНК-полимеразу Therminator II, ДНК-полимеразу AmpliTherm, ДНК-полимеразу SP6. Следующие неограничивающие примеры обратных транскриптаз (RT) могут быть использованы в реакциях способа по настоящему изобретению для повышения производительности при выявлении последовательности РНК: OmniScript (Qiagen), Sensi Script (Qiagen), MonsterScript (Epicentre), Transcriptor (Roche), RT HIV (Ambion), Superscript III (Invitrogen), ThermoScript (Invitrogen), Thermo-X (Invitrogen), ImProm II (Promega).
Фермент, вносящий одноцепочечный разрыв, связывается с последовательностью распознавания в двухцепочечной ДНК и расщепляет одну цепь двухцепочечной спирали. Ферменты, вносящие одноцепочечный разрыв, могут производить расщепление выше или ниже сайта распознавания для них или в пределах сайта распознавания для фермента. В способах, раскрытых в данном документе, для запуска повторяющихся циклов внесения одноцепочечных разрывов в ДНК-субстрат и достройки от одноцепочечного разрыва с помощью полимеразы для проведения экспоненциальной амплификации целевого фрагмента нуклеиновой кислоты между праймерами-матрицами можно применять только те ферменты, вносящие одноцепочечный разрыв, которые расщепляют верхнюю цепь ниже сайта распознавания. В идеальном случае фермент, вносящий одноцепочечный разрыв, функционирует при тех же условиях реакции, что и полимераза. В предпочтительном варианте осуществления настоящего изобретения фермент, вносящий одноцепочечный разрыв, является термостабильным и активным от 50°С до 60°С. Иллюстративные ферменты, вносящие одноцепочечный разрыв, применимые в способах, раскрытых в данном документе, включают, без ограничения, Nt.BspQI (NEB), Nt.BspD6I, Nt.BsmAI (NEB), Nt.AlwI (NEB), Nt.BbvCI (NEB), N.Bst9I (Sibenzyme) и Nt.BstNBI (NEB).
Реакционная смесь для NEAR обычно содержит нуклеотиды, такие как, например, дидезоксирибонуклеозидтрифосфаты (dNTP). Реакцию можно также проводить в присутствии dNTP, которые содержат выявляемый фрагмент, в том числе, без ограничения, радиоактивную метку (например, 32Р, 33Р, 125I, 35S), фермент (например, щелочную фосфатазу), флуоресцентную метку (например, изотиоцианат флуоресцеина (FITC)), биотин, авидин, дигоксигенин, антигены, гаптены или флуорохромы. Реакционная смесь для NEAR дополнительно включает некоторые соли и буферы, которые обеспечивают активность фермента, вносящего одноцепочечный разрыв, и полимеразы.
NEAR-реакцию предпочтительно проводят в практически изотермических условиях, когда температура реакции является более или менее постоянной в ходе реакции амплификации. Поскольку температура не должна изменяться циклически от верхней температуры до нижней температуры, NEAR-реакцию можно проводить в условиях, в которых осуществить обычную ПЦР было бы трудно. Как правило, реакцию проводят при температуре, составляющей приблизительно от 35°С до 90°С (например, при 35, 37, 42, 60, 65, 70, 75, 80 или 85°С). Поддержание температуры с высокой степенью точности предпочтительно не является существенным. Некоторая изменчивость температуры является приемлемой.
Для понижения температуры плавления олигонуклеотидов также можно применять модификаторы температуры плавления (Tm) и скорости реакции, такие как (без ограничения) этиленгликоль и глицерин. Кроме того, для изменения скорости реакции можно применять модификаторы скорости реакции с участием ДНК-полимеразы (такие как dNTP и концентрация магния), что приводит к большей точности количественного определения.
Настоящее изобретение обеспечивает способы контроля NEAR-реакции в режиме реального времени с использованием стратегии NEAR-амплификации, описанной выше и в патентах US 007112423 В2 и US 20090017452 A1. В одном варианте осуществления в количественной NEAR используется амплификация целевых нуклеиновых кислот наряду с контрольной амплификацией известного количества. Количество целевой нуклеиновой кислоты может быть рассчитано в виде абсолютного количественного определения или относительного количественного определения (полуколичественного) на основе источника контроля (экзогенного или эндогенного контроля).
Количественную оценку неизвестной нуклеотидной последовательности можно осуществить либо путем сравнения порогового значения из логарифмической фазы амплификации для неизвестной последовательности с таковым для серии известных целевых последовательностей в отдельной группе реакционных смесей или в той же реакционной смеси; или путем получения порогового значения от внутреннего эндогенного или экзогенного продукта совместной амплификации, которое указывает либо на положительный результат (если значение для неизвестной последовательности превышает пороговое значение), либо на отрицательный результат (если значение для неизвестной последовательности не превышает пороговое значение).
Пути применения
Настоящее изобретение обеспечивает контроль изотермической амплификации в NEAR-реакции в режиме реального времени, который может обеспечить количественную меру количества исходной целевой нуклеиновой кислоты. Композиции и способы по настоящему изобретению применимы в диагностике для человека, где желателен быстрый количественный ответ (например, выявляемая амплификация в течение до 15, 10, 9, 8, 7, 6, 5 мин. или менее). В конкретных вариантах осуществления настоящее изобретение обеспечивает применение анализа на основе NEAR-реакции в диагностике для человека в клинических условиях. В других вариантах осуществления настоящее изобретение обеспечивает применение анализа на основе NEAR-реакции в диагностике при выездной работе, где доступ к оборудованию для термоциклирования отсутствует или будет чрезмерно дорогим. В других вариантах осуществления настоящее изобретение обеспечивает применение анализа на основе NEAR-реакции в условиях для научной работы, где желательны быстрые количественные ответы.
Наборы
Настоящее изобретение также обеспечивает наборы для амплификации целевой молекулы нуклеиновой кислоты. Такие наборы применимы для выявления или количественной оценки целевой нуклеиновой кислоты в биологическом образце, полученном от субъекта. Наборы по настоящему изобретению могут содержать, например, одну или несколько полимераз, прямые и обратные праймеры-матрицы и один или несколько ферментов, вносящих одноцепочечный разрыв, описанных в данном документе. Если амплификации подлежит одна целевая последовательность, то в набор могут быть включены один или два фермента, вносящих одноцепочечный разрыв. Если амплификации подлежат несколько целевых последовательностей и праймеры-матрицы, сконструированные для этих целевых последовательностей, содержат сайты для фермента, вносящего одноцепочечный разрыв, для одного и того же фермента, вносящего одноцепочечный разрыв, тогда могут быть включены один или два фермента, вносящих одноцепочечный разрыв. Если праймеры-матрицы распознаются разными ферментами, вносящими одноцепочечный разрыв, то в набор могут быть включены несколько ферментов, вносящих одноцепочечный разрыв, как, например, 3 или более.
В одном аспекте настоящее изобретение обеспечивает набор для амплификации нуклеиновых кислот, содержащий ДНК-полимеразу, первый праймер-матрицу, второй праймер-матрицу, фермент, вносящий одноцепочечный разрыв, со специфичностью к сайту распознавания для фермента, вносящего одноцепочечный разрыв, в праймерах-матрицах и дезоксинуклеотидтрифосфаты (dNTP) (например, в буферном растворе, содержащем компоненты, достаточные для амплификации). В различных вариантах осуществления каждый из первого праймера-матрицы и второго праймера-матрицы имеет последовательность 3'-концевого специфичного участка распознавания, комплементарную или практически комплементарную целевой последовательности, где концевой специфичный участок распознавания содержит один или несколько 2'-модифицированных нуклеотидов; последовательность 5'-концевого хвоста, содержащую сайт распознавания для фермента, вносящего одноцепочечный разрыв, выше последовательностей 3'-концевого специфичного участка распознавания, и стабилизирующую последовательность выше (5') сайта связывания для фермента, вносящего одноцепочечный разрыв.
В одном аспекте наборы по настоящему изобретению содержат гомогенную смесь всех компонентов для NEAR-реакции, в том числе, без ограничения, dNTP, прямые и обратные праймеры-матрицы, фермент, вносящий одноцепочечный разрыв, полимеразу, выявляемый полинуклеотидный зонд, специфичный для целевой молекулы, буфер и стабилизаторы реакции, за исключением целевой нуклеиновой кислоты.
Наборы по настоящему изобретению могут также содержать один или несколько компонентов в любом количестве отдельных контейнеров, пакетов, пробирок (например, на <0,2 мл, 0,2 мл, 0,6 мл, 1,5 мл, 5,0 мл, >5,0 мл), флаконов, титрационных микропланшетов (например, на <96 лунок, 96 лунок, 384 лунки, 1536 лунок, >1536 лунок), ArrayTape и т.п., или компоненты могут быть объединены в различных комбинациях в таких контейнерах. В различных вариантах осуществления набор дополнительно включает пару олигонуклеотидных праймеров-матриц, способных к связыванию и амплификации эталонной последовательности. В других вариантах осуществления набор включает стерильный контейнер, содержащий олигонуклеотидные праймеры-матрицы; такие контейнеры могут быть коробками, ампулами, бутылками, флаконами, пробирками, мешками, сумками, блистерными упаковками или другой подходящей формой контейнеров, известных в данной области техники. Такие контейнеры могут быть изготовлены из пластика, стекла, ламинированной бумаги, металлической фольги или других материалов, применимых для хранения нуклеиновых кислот.
Компоненты набора могут, например, присутствовать в одном или нескольких контейнерах, например, все компоненты могут находиться в одном контейнере, или, например, ферменты и матрицы могут находиться в отдельных контейнерах. Компоненты могут, например, быть высушенными (например, сухой остаток), лиофилизированными (например, сухой осадок) или находиться в стабильном буфере (например, химически стабилизированном, термически стабилизированном). Сухие компоненты могут, например, быть получены путем лиофилизации, вакуумной сушки и сушки при помощи центрифужной сушилки и/или сушки при окружающей температуре. В различных вариантах осуществления полимераза и ферменты, вносящие одноцепочечный разрыв, находятся в лиофилизированной форме в одном контейнере, а матрицы являются либо лиофилизированными, подвергнутыми сублимационной сушке, или находятся в буфере в другом контейнере. В некоторых вариантах осуществления полимераза, ферменты, вносящие одноцепочечный разрыв, и матрицы находятся в лиофилизированной форме в одном контейнере. В других вариантах осуществления полимераза и фермент, вносящий одноцепочечный разрыв, могут быть разделены по разным контейнерам.
Наборы могут дополнительно включать, например, dNTP, применяемые в реакции, или модифицированные нуклеотиды, кюветы или другие контейнеры, применяемые для реакции, или флакон с водой или буфером для повторной гидратации лиофилизированных компонентов. Применяемый буфер может, например, подходить для активности как полимеразы, так и фермента, вносящего одноцепочечный разрыв.
Наборы по настоящему изобретению могут также содержать инструкции для выполнения одного или нескольких способов, описанных в данном документе, и/или описание одной или нескольких композиций или реагентов, описанных в данном документе. Инструкции и/или описания могут быть в печатной форме и могут быть включены во вкладыш для набора. Набор также может включать в себя письменное описание адреса в Интернете, по которому предоставляют такие инструкции или описания.
Наборы могут дополнительно включать реагенты, применяемые в способах выявления (например, в режиме реального времени или по конечной точке), такие как, например, габридизационные зонды или красители, связывающиеся с ДНК. Наборы могут дополнительно включать реагенты, применяемые в способах выявления, такие как:, например, реагенты, применяемые для FRET, устройства для анализа в горизонтальном потоке, указатели уровня, флуоресцентный краситель, коллоидные частицы золота, латексные частицы, молекулярный маяк или полистирольные гранулы. Компоненты выявления могут быть встроены в устройство для анализа в горизонтальном потоке. Устройство для анализа в горизонтальном потоке можно применять в месте наблюдения.
В практическом осуществлении настоящего изобретения используют, если не указано иное, обычные методики молекулярной биологии (в том числе методики рекомбинантных молекул), микробиологии, клеточной биологии, биохимии и иммунологии, которые вполне находятся в пределах компетенции специалиста в данной области. Такие методы подробно описаны в литературе, такой как "Molecular Cloning: A Laboratory Manual", second edition (Sambrook, 1989); "Oligonucleotide Synthesis" (Gait, 1984); "Animal Cell Culture" (Freshney, 1987); "Methods in Enzymology", "Handbook of Experimental Immunology" (Weir, 1996); "Gene Transfer Vectors for Mammalian Cells" (Miller and Calos, 1987); "Current Protocols in Molecular Biology" (Ausubel, 1987); "PCR: The Polymerase Chain Reaction" (Mullis, 1994); "Current Protocols in Immunology" (Coligan, 1991). Эти методики применимы к получению полинуклеотидов и полипептидов по настоящему изобретению, и, таким образом, могут рассматриваться в подготовке и практическом осуществлении настоящего изобретения. Особенно применимые методики для конкретных вариантов осуществления будут обсуждаться в следующих разделах.
Следующие примеры предложены таким образом, чтобы обеспечить специалиста в данной области полным раскрытием и описанием подготовки и применения анализа, скрининга и терапевтических способов по настоящему изобретению, и не предназначены для ограничения объема того, что авторы настоящего изобретения считают своим изобретением.
ПРИМЕРЫ
В настоящее время NEAR-реакцию применяют для быстрого и изотермического выявления присутствия или отсутствия целевого олигонуклеотида в образце. В связи с техническими ограничениями обычные способы NEAR не подходят для количественной оценки целевых олигонуклеотидов в режиме реального времени по меньшей мере частично по причине незаконной амплификации нецелевых молекул в образце, которая препятствует выявлению и точному количественному определению целевых ампликонов. Настоящее изобретение обеспечивает композиции и способы, которые преодолевают эти ограничения путем обеспечения выявляемых праймеров/матриц, которые не поддаются незаконной амплификации. В одном из вариантов осуществления в количественном анализе NEAR используют праймер, содержащий одну или несколько модификаций 2'-O-Ме, который предотвращает или снижает интенсивность незаконной амплификации нецелевых молекул в ходе NEAR-реакции. В настоящее время план анализа на основе NEAR-амплификации ограничен очень короткими участками в целевой нуклеиновой кислоте, которые имеют по меньшей мере один встречающийся в природе сайт распознавания для фермента, вносящего одноцепочечный разрыв, в непосредственной близости. Синтез с замещением цепей, инициированный в этих сайтах одноцепочечного разрыва, обеспечивает одноцепочечные целевые молекулы ДНК, с которыми праймеры-матрицы с короткими участками, специфичными для целевой молекулы, могут связываться и начинать циклы внесения одноцепочечных разрывов/полимеразной достройки в реакциях амплификации целевой молекулы. Настоящее изобретение обеспечивает композиции и способы, которые преодолевают это ограничение путем использования праймеров-матриц с более длинными участками, специфичными для целевой молекулы, которые вследствие этого способны проникать в цепь в диапазоне от 50°С до 60°С во время первой фазы реакции амплификации без помощи синтеза с замещением цепей. Более длинные участки, специфичные для целевой молекулы, в праймерах-матрицах привносят недостаток, заключающийся в предоставлении большей полезной площади для формирования неспецифических гибридов ДНК с достраиваемыми 3'-концами, что может запустить синтез неспецифических продуктов амплификации. Композиции в настоящем изобретении ослабляют этот недостаток путем распространения размещения 2'-модифицированных нуклеотидов за пределы 3'-концевого блока из пяти последовательных модифицированных нуклеотидов с покрытием всего участка, специфичного для целевой молекулы, с использованием альтернативной последовательности из 2'-модифицированных и немодифицированных нуклеотидов.
Пример 1. Олигонуклеотидные праймеры-матрицы, содержащие нуклеотиды с 2'-О-метильной модификацией, ослабляют или устраняют сигнал фона в NEAR-амплификации
Если NEAR-амплификацию выполняют без ввода целевой нуклеиновой кислоты (т.е. в контролях без целевой молекулы; NTC), сигнал образуется, несмотря на отсутствие матрицы. Таким образом, образование сигнала фона способно снижать точности количественного определения целевой нуклеиновой кислоты с помощью NEAR-амплификации. Было высказано предположение, что сигнал фона образовывался, в частности, посредством формирования димеров праймеров олигонуклеотидными праймерами/матрицами. Не ограничиваясь теорией, предполагают, что структуры, подавляющие активность полимеразы, содержащие 2'-модифицированные нуклеотиды, можно применять для ослабления или устранения межмолекулярных и/или внутримолекулярных взаимодействий праймеров/матриц (например, формирования димеров праймеров), и, таким образом, ослабления или устранения сигнала фона в NEAR-анализе.
Иллюстративные структуры объектов, подавляющих активность полимеразы, в направлении 5'-3' содержат стабилизирующую последовательность, последовательность распознавания для фермента, вносящего одноцепочечный разрыв, спейсерную последовательность для фермента, вносящего одноцепочечный разрыв, и последовательность распознавания, специфичную для целевой молекулы, при этом последовательность распознавания, специфичная для целевой молекулы, содержит один или несколько 2'-модифицированных нуклеотидов (например, рибонуклеотиды с 2'-O-метильной модификацией). Если в последовательности распознавания, специфичной для целевой молекулы, присутствуют два или более 2'-модифицированных нуклеотида, то 2'-модифицированные нуклеотиды могут быть смежными (например, 2, 3, 4, 5 или более 2'-модифицированных нуклеотидов). Титрование синтетической двухцепочечной целевой молекулы ДНК Clavibacter michiganensis sepidonicus (Cms) оценивали с помощью выявления посредством флуоресцентного маяка. Целевую ДНК подвергали серийному разведению из исходного раствора синтезированной "многомерной" ДНК в 250 пар оснований, которую сконструировали с целевой последовательностью и одним сайтом одноцепочечного разрыва.
Сигнал в контролях без целевой молекулы (NTC) был подавлен в реакционных смесях, содержащих матрицы с 2'-O-метильной модификацией (фигура 2А). Калибровочная кривая отображает широкий динамический диапазон с использованием реакционных смесей с матрицами с 2'-O-метильной модификацией (фигура 2В). Образцы (10 мкл) реакционных смесей из контроля без целевой молекулы проанализировали с помощью ВЭЖХ/масс-спектрометрии, и подтвердили подавление образования фоновых продуктов амплификации (фигура 2С). Спектры, полученные в реакциях с применением немодифицированных олигонуклеотидов, показали сложный спектр, образованный несколькими продуктами амплификации, полученными из неспецифических фоновых продуктов, вместе с непрореагировавшими матрицами (фигура 2С, левая панель). Спектры, полученные в реакциях с применением олигонуклеотидов с 2'-O-метильной модификацией, показали простой спектр, образованный непрореагировавшими матрицами без присутствия неспецифических фоновых продуктов (фигура 2С, правая панель).
Для изучения эффекта реакционных смесей, содержащих матрицы с 2'-O-метильной модификацией, в отношении амплификации биологического образца геномную ДНК Clavibacter michiganensis sepidonicus (Cms) применяли в качестве целевой ДНК. Амплифицированные продукты выявляли с помощью SYBR Green, который выявляет двухцепочечную ДНК (фигуры 3А и 3В), или молекулярных маяков, которые выявляли специфический продукт (фигуры 4А и 4В). Стандартные реакции выполняли с олигонуклеотидными ДНК-матрицами (фигуры 3А и 4А), а реакции, включающие матрицы с 2'-O-метильной модификацией (DNAble), выполняли с олигонуклеотидами, содержащими блок из 5 смежных нуклеотидов с 2'-O-метильной модификацией на 3'-конце в последовательности распознавания, специфичной для целевой молекулы (фигуры 3В и 4В). Сигнал в контролях без целевой молекулы (NTC) был подавлен в реакционных смесях, содержащих матрицы с 2'-O-метильной модификацией (фигуры 3В и 4В), тогда как в контролях без целевой молекулы (NTC) наблюдался существенный сигнал (фигуры 4А и 4В), что указывало на образование фонового продукта в отсутствие целевой ДНК.
Таким образом, эти результаты указывают на то, что праймеры, содержащие нуклеотиды с 2'-O-метильной модификацией, ослабляют или устраняют сигнал фона в NEAR-амплификации.
Пример 2. Размещение нуклеотидов с 2'-O-метильной модификацией в олигонуклеотидных праймерах/матрицах изменяет время до выявления и эффективность NEAR-реакций
Иллюстративные объекты, подавляющие активность полимеразы, имеющие нуклеотиды с 2'-O-метильной модификацией в различных положениях внутри участка специфичности, применяли в реакциях NEAR-амплификации, и изучали их кинетику реакции. В частности, изученные праймеры/матрицы содержали пару олигонуклеотидов, имеющих блок из пяти нуклеотидов с 2'-O-метильной модификацией, расположенных на 3'-конце участка специфичности или на 5'-конце участка специфичности (на 2 нуклеотида ниже сайта одноцепочечного разрыва) (фигура 5). Стандартные реакции выполняли в двух повторностях с блоком из нуклеотидов с 2'-O-метильной модификацией, расположенным на 3'-конце или начинающимся с 3-го нуклеотида после сайта одноцепочечного разрыва и охватывающим 5 оснований, или со смесью этих двух структур, как указано. Целевая ДНК была геномной ДНК Clavibacter michiganensis sepidonicus (Cms). Выявление было основано на молекулярном маяке в конечной концентрации 100 нМ.
Объекты, модифицирующие скорость реакции, имеющие нуклеотиды с 2'-O-метильной модификацией в различных положениях внутри участка специфичности олигонуклеотидного праймера/матрицы, отображали различную кинетику амплификации (фигура 6). В реакциях, в которых применяются праймеры/матрицы, имеющие блок из пяти нуклеотидов с 2'-O-метильной модификацией на 3'-конце, показано уменьшенное время до выявления (матрица "Концевая"; 170 с) по сравнению с праймером/матрицей, имеющим блок из пяти нуклеотидов с 2'-O-метильной модификацией, начинающийся с 3-го нуклеотида после сайта одноцепочечного разрыва (матрица "Одноцепочечный разрыв +2"; 430 с). Таким образом, предположили, что соотношения двух олигонуклеотидных праймеров/матриц можно применять для регуляции времени до выявления и/или эффективности реакции для "настройки" реакций. В реакциях с разными соотношениями матрица "Концевая" : матрица "Одноцепочечный разрыв +2" показано промежуточное время до выявления, значение которого находится между значениями для данных двух матриц (фигура 6). Кроме того, с увеличением соотношения матрица "Концевая" : матрица "Одноцепочечный разрыв +2" кривая сужалась, а наклон кривой изменялся. Таким образом, было показано, что расположение 2'-модифицированных нуклеотидов в олигонуклеотидных праймерах/матрицах и соотношение олигонуклеотидных праймеров/матриц с по-разному расположенными 2'-модифицированными нуклеотидами изменяли время до выявления и эффективность NEAR-реакций. Настоящее изобретение по меньшей мере частично основано на данных обнаружениях.
Пример 3. Полное подавление неспецифической амплификации в NEAR-анализах с использованием праймеров-матриц с длинными участками, специфичными для целевой молекулы
NEAR-анализ для количественного определения гена алкогольдегидрогеназы 1 (ADH1) кукурузы разработали с использованием двух альтернативных наборов прямых и обратных праймеров-матриц (TS3 и TS3). Подходящий сайт распознавания для фермента, вносящего одноцепочечный разрыв, не мог быть расположен в пределах 500 нуклеотидов выше или ниже целевого участка последовательности в геномной ДНК кукурузы. Оба набора праймеров-матриц отличаются более длинными участками, комплементарными целевой последовательности (16 и 19 нуклеотидов, соответственно), способными к гибридизации с целевой ДНК, опосредованной проникновением в цепь. В первом наборе (TS3) комплементарные целевой последовательности участки прямых и обратных праймеров-матриц содержат блок из 5 последовательных рибонуклеотидов с 2'-O-метильной модификацией непосредственно выше 3'-концевого дезоксинуклеотида. Остальная часть участка, комплементарного целевой последовательности, содержит последовательность чередующихся немодифицированных дезоксинуклеотидов и рибонуклеотидов с 2'-O-метильной модификацией, начинающуюся на пять нуклеотидов (прямой праймер-матрица) или четыре нуклеотида (обратный праймер-матрица) ниже сайта одноцепочечного разрыва. Второй набор (TS6) праймеров-матриц отличается только блоком из пяти рибонуклеотидов с 2'-O-метильной модификацией, прилегающих к 3'-концевому немодифицированному дезоксинуклеотиду, в то время как остальная часть участка, комплементарного целевой последовательности, содержит только немодифицированные дезоксинуклеотиды.
Десять микролитров реакционных смесей для NEAR помещали в 50 мМ Tris с рН 8,0, 15 мМ (NH4)2SO4, 15 мМ Na2SO4 и 15 мМ MgSO4 с применением 3,84 Ед ДНК-полимеразы 1 Bst Warmstart 2.0 (NEB), 10000 копий синтетической целевой ДНК ADH1 кукурузы, 0,3 мМ dNTP, 3 Ед фермента, вносящего одноцепочечный разрыв, Nt.BstNBI, 200 нМ ROX/BHQ-меченого зонда для ADH1 типа молекулярного маяка, 0,5Х красителя SYBR Green (LifeTechnologies), 1000 нМ обратного праймера-матрицы из TS3 или TS6 и 100 нМ прямого праймера-матрицы из TS3 или TS6. Группу контрольных реакционных смесей без целевой ДНК (NTC) составляли из тех же компонентов без синтетической целевой ДНК ADH1 кукурузы. Все реакционные смеси инкубировали при 56°С в течение 15 минут, и флуоресцентные сигналы регистрировали при 520 нм (SYBR Green) и 610 нм (ROX).
Сравнивают графики амплификации для реакционных смесей, содержащих целевую ДНК, в каналах выявления для SYBR Green (фигура 8) и ROX (фигура 9) с графиками амплификации для реакционных смесей из NTC в канале выявления для SYBR Green (фигура 10).
Результаты, представленные в данном документе, получены с помощью следующих способов и материалов, если не указано иное.
Реакции NEAR-амтификации
Реакционные смеси (50 мкл) содержали 15 мМ MgSO4, 0,3 мМ dNTP, 19,2 единиц полимеразы Bst, 15 единиц n.BstNBI, 1000 нМ матрицы 1 и 200 нМ матрицы 2. Целевая ДНК представляла собой геномную ДНК Clavibacter michiganensis sepidonicus (Cms) или "многомерную" ДНК на основе последовательностей Cms. Матрицы и целевую молекулу предварительно проинкубировали вместе при 56°С в течение 30 секунд в общем объеме 10 мкл. Мастер-микс из оставшихся компонентов реакции предварительно инкубировали при 56°С в течение 30 секунд в общем объеме 40 мкл. Мастер-микс объединяли с матрицами и целевой молекулой и инкубировали при 56°С в течение 10 минут с флуоресцентной детекцией (SYBR Green или молекулярные маяки), данные которой собирали каждые 10 секунд в ходе инкубации. Реакционные смеси "убивали нагреванием" на этапе, продолжавшемся 2 минуты при 95°С, с последующим возвращением к комнатной температуре. Эквиваленты порогового цикла (Ct) определяли для каждой реакции на основе формулы для подбора кривой в программном обеспечении Biorad IQ5, и значения откладывали на графике с помощью Microsoft Excel. Выполняли линейную регрессию и определяли коэффициент корреляции (R2).
Другие варианты осуществления
Из вышеприведенного описания будет очевидно, что в настоящее изобретение, описанное в данном документе, можно внести изменения и модификации, чтобы приспособить его к различным путям применения и условиям. Такие варианты осуществления также находятся в пределах объема прилагаемой формулы изобретения.
Приведение перечня элементов в любом определении переменной в данном документе включает определения этой переменной в качестве любого отдельного элемента или комбинации (или подкомбинации) перечисленных элементов. Приведение варианта осуществления в данном документе включает данный вариант осуществления в качестве любого отдельного варианта осуществления или в комбинации с любыми другими вариантами осуществления или их частями.
Настоящая заявка может быть родственной по отношению к международной заявке на патент № PCT/US 2011/047049, поданной 9 августа 2011 года, которая заявляет приоритет по предварительной заявке на патент США №61/373695, поданной 13 августа 2010 года, полное содержание которых включено в данный документ посредством ссылки.
Все патенты и публикации, упомянутые в данном описании, включены в данный документ посредством ссылки в той же степени, как если бы каждый отдельный патент или публикация были конкретно и отдельно указаны как включенные посредством ссылки.



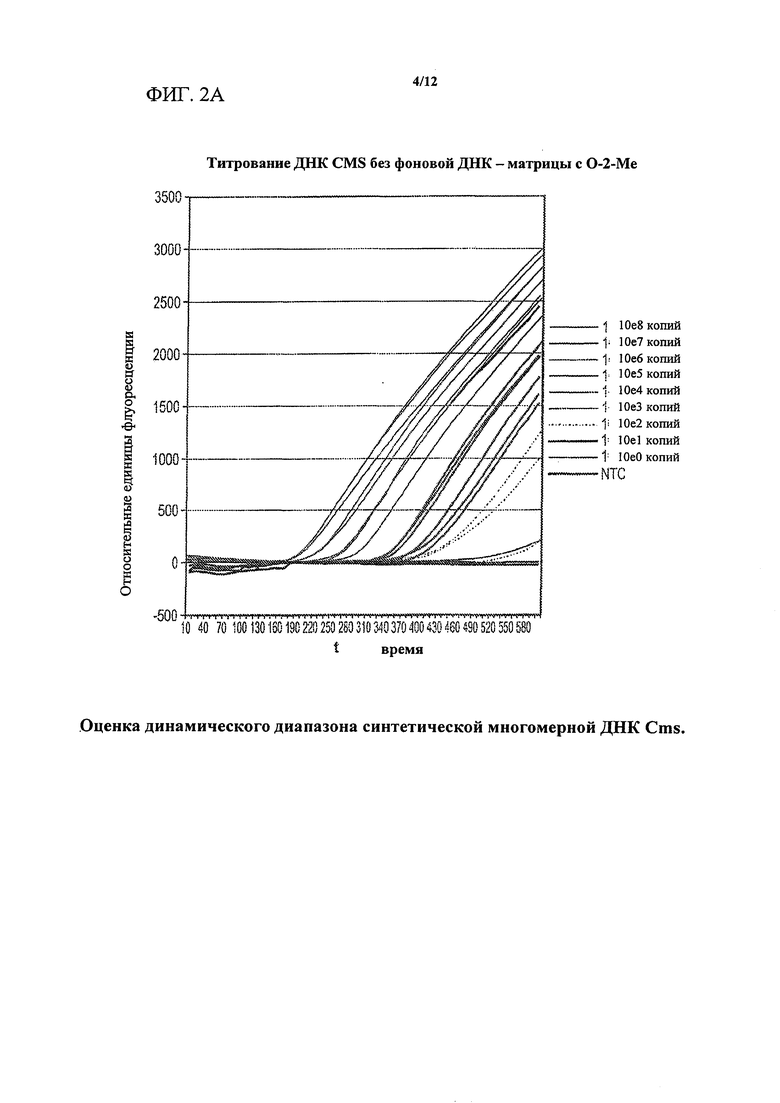
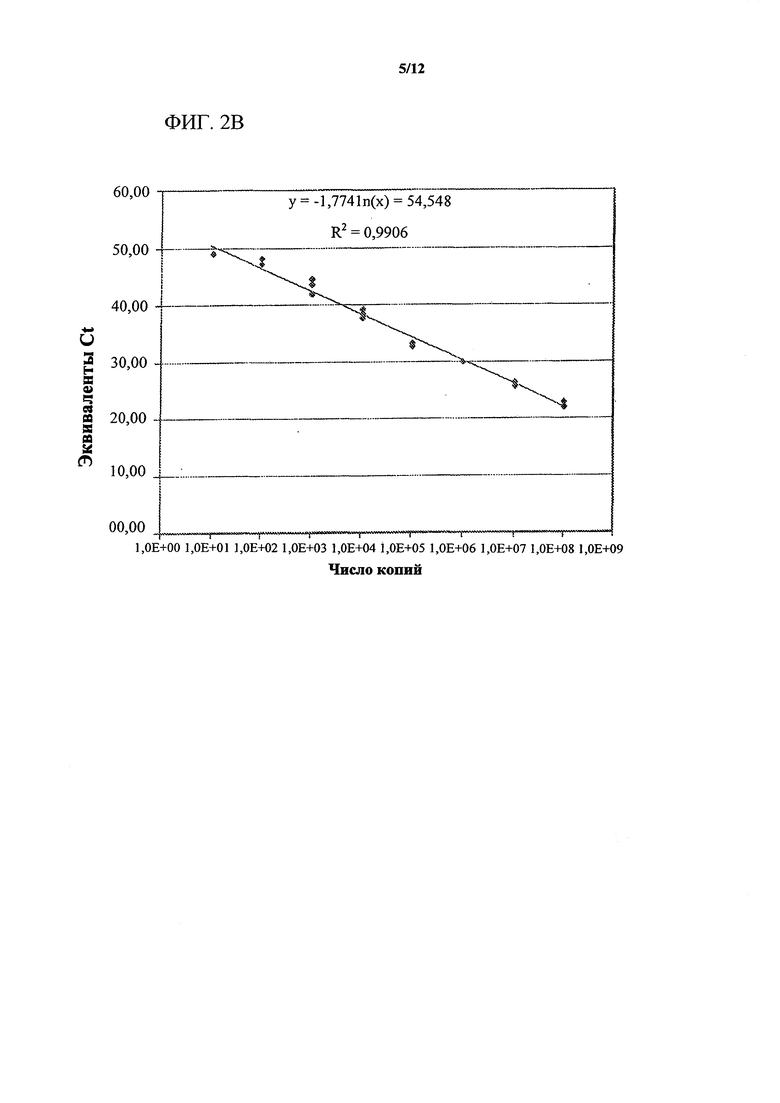

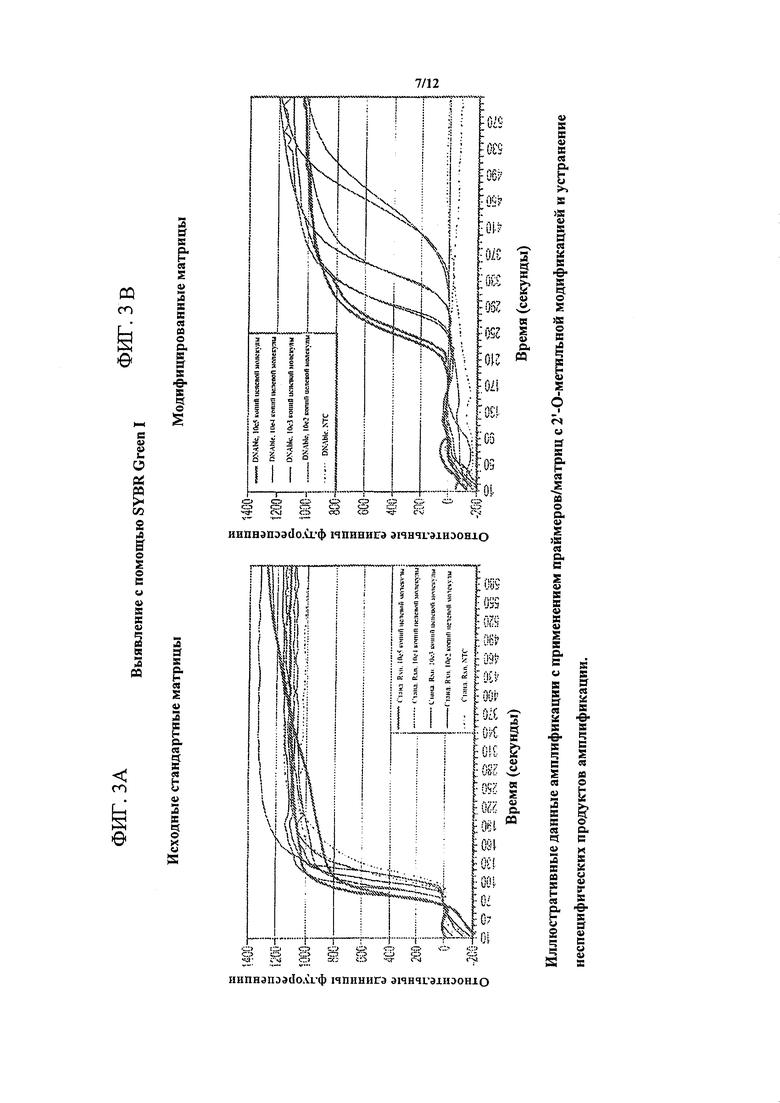
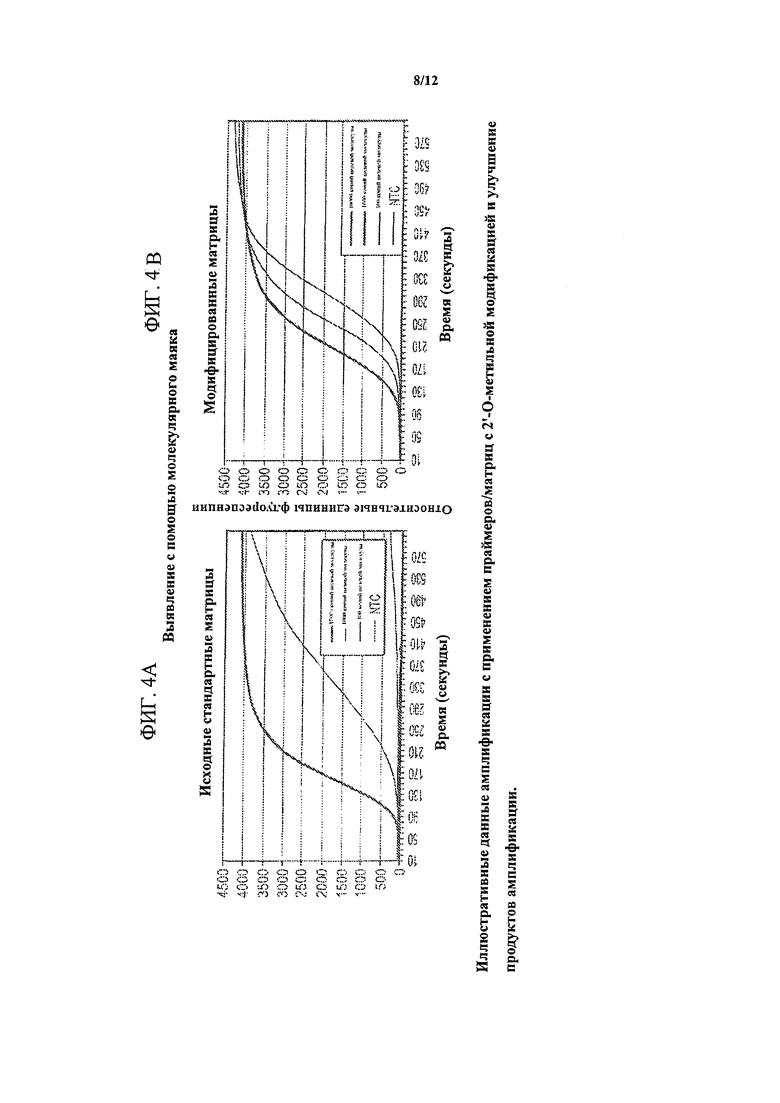

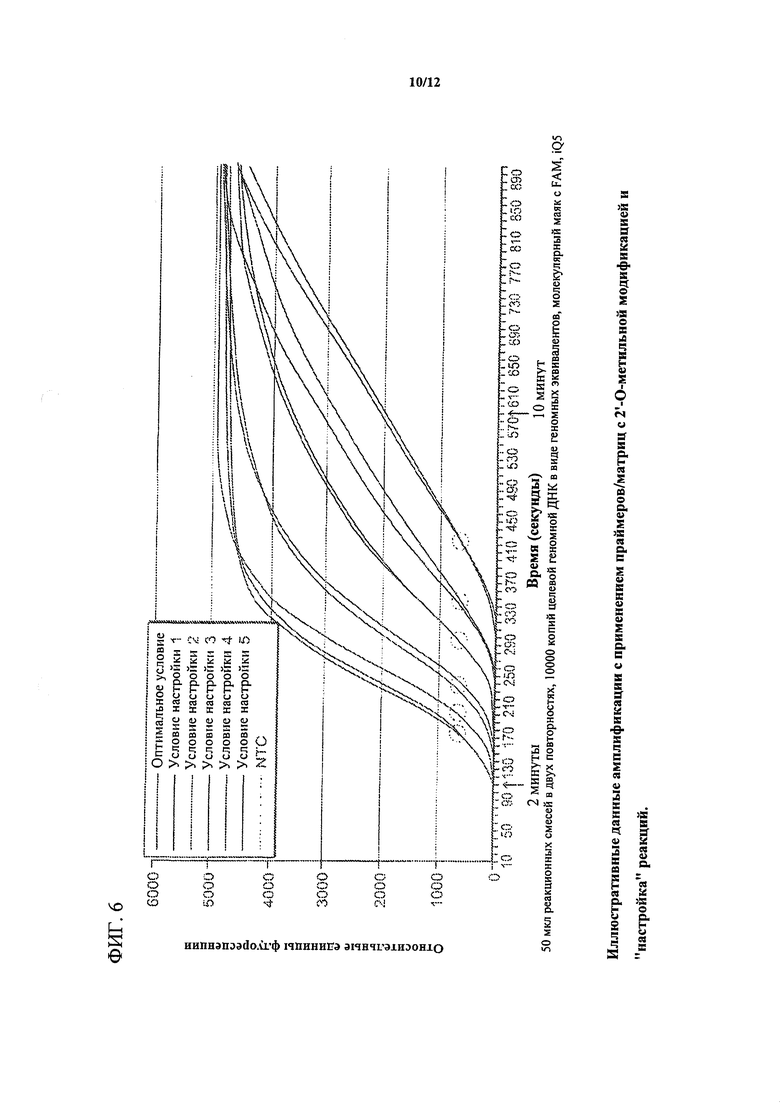
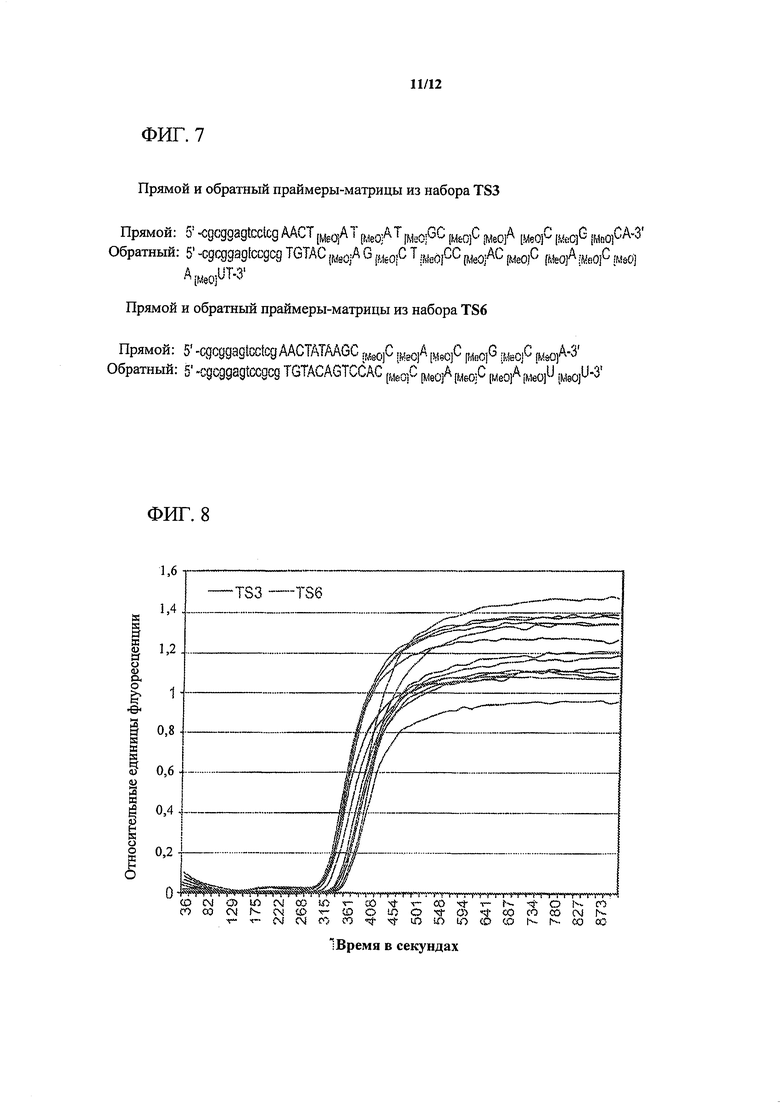
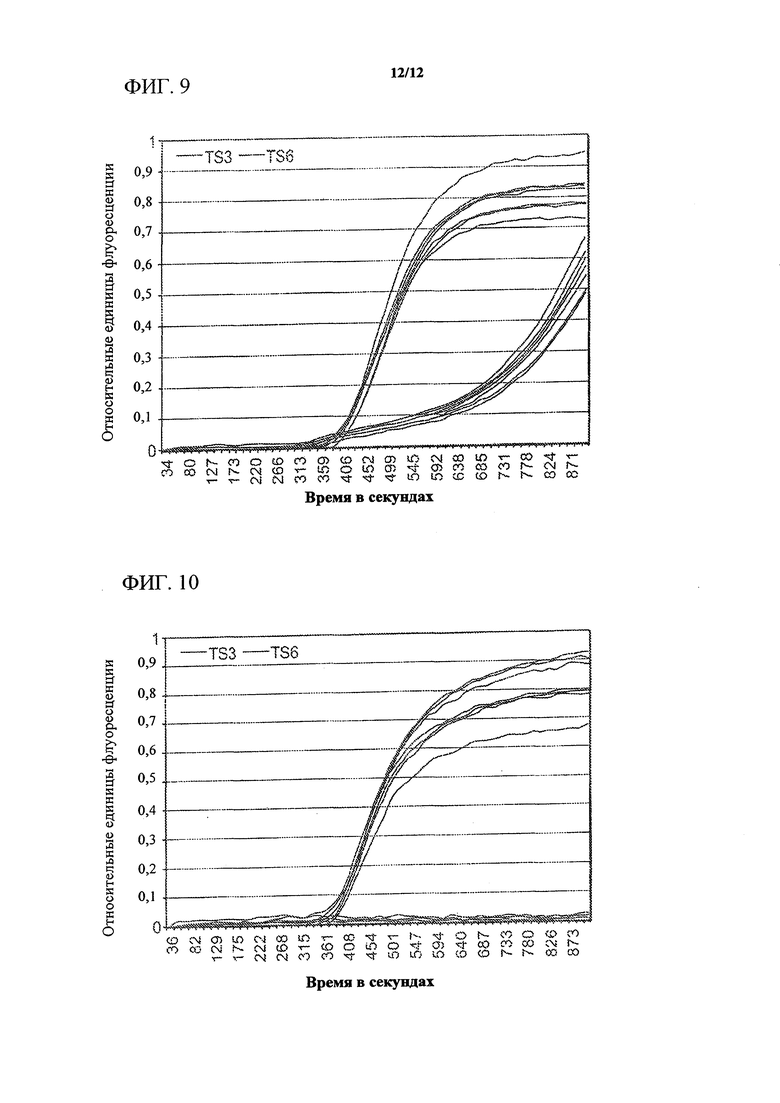
Изобретение относится к области биотехнологии, в частности представлены композиции и способы выявления и количественного определения целевого олигонуклеотида в образце в режиме реального времени. Эти способы совместимы с целевыми олигонуклеотидами, амплифицируемыми с помощью NEAR-реакции. 3 н. и 12 з.п. ф-лы, 10 ил., 3 пр.
1. Способ амплификации полинуклеотида, причем указанный способ включает:
(a) приведение целевой молекулы нуклеиновой кислоты в контакт в практически изотермических условиях с полимеразой, двумя или более олигонуклеотидными праймерами, каждый из которых специфически связывается с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты, и ферментом, вносящим одноцепочечный разрыв, причем каждый из олигонуклеотидных праймеров содержит один или несколько 2'-модифицированных нуклеотидов, расположенных на 3'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты; и
(b) образование ампликонов, содержащих по меньшей мере часть указанной целевой молекулы нуклеиновой кислоты.
2. Способ по п. 1, дополнительно включающий выявление ампликонов.
3. Способ по п. 2, причем указанное выявление включает связывание ампликонов с выявляемым зондом.
4. Способ по п. 1, дополнительно включающий один или несколько 2'-модифицированных нуклеотидов, расположенных на 5'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты.
5. Способ по п. 1, отличающийся тем, что указанные два или более 2'-модифицированных нуклеотида, расположенные на 3’-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты, являются смежными.
6. Способ по п. 1, отличающийся тем, что 5 смежных нуклеотидов с 2'-O-метильной модификацией расположены на 3'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты.
7. Способ по п. 1, дополнительно включающий 5 смежных нуклеотидов с 2'-O-метильной модификацией, расположенных на 5'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты.
8. Способ амплификации пол и нуклеотида, причем указанный способ включает:
(a) приведение целевой молекулы нуклеиновой кислоты в контакт в практически изотермических условиях с полимеразой, двумя олигонуклеотидными праймерами, каждый из которых специфически связывается с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты, и ферментом, вносящим одноцепочечный разрыв, причем каждый из олигонуклеотидных праймеров содержит 5 смежных нуклеотидов с 2'-O-метильной модификацией, расположенных на 3'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты; и
(b) образование ампликонов, содержащих по меньшей мере часть указанной целевой молекулы нуклеиновой кислоты.
9. Способ амплификации полинуклеотида, полученного от патогена, причем указанный способ включает:
(а) приведение полинуклеотида, полученного от патогена, в контакт в практически изотермических условиях с полимеразой, двумя или более олигонуклеотидными праймерами, каждый из которых специфически связывается с комплементарной последовательностью в целевой молекуле нуклеиновой кислоты, и ферментом, вносящим одноцепочечный разрыв, причем каждый из олигонуклеотидных праймеров содержит один или несколько 2'-модифицированных нуклеотидов, расположенных на 3'-конце последовательности, комплементарной целевой молекуле нуклеиновой кислоты; и
(b) образование ампликонов, содержащих по меньшей мере часть указанной целевой молекулы нуклеиновой кислоты.
10. Способ по п. 9, дополнительно включающий выявление ампликона с выявляемым зондом.
11. Способ по п. 9, отличающийся тем, что патоген представляет собой вирус, бактерию, дрожжи или гриб.
12. Способ по п. 9, отличающийся тем, что патоген находится в биологическом образце.
13. Способ по п. 12, отличающийся тем, что биологический образец выбран из группы, состоящей из крови, сыворотки, плазмы, амниотической жидкости, мокроты, мочи, спермы, влагалищного секрета, спинномозговой жидкости, лимфы, слезной жидкости, кала или желудочного сока.
14. Способ по п. 9, отличающийся тем, что патоген находится в пробе из окружающей среды.
15. Способ по п. 4, отличающийся тем, что один или несколько 2’-модифицированных нуклеотидов, расположенных на 5’-конце последовательности, комплементарной целевой нуклеиновой кислоты, отделены от сайта одноцепочечного разрыва с помощью 1, 2, 3, 4, 5 или более немодифицированных нуклеотидов.
| Способ получения покрытий на основе соединения никеля с алюминием | 1985 |
|
SU1420069A1 |
| Шипицына Елена Васильевна | |||
| Оценка методов амплификации нуклеиновых кислот для диагностики трихомониаза | |||
| Журнал акушерства и женских болезней | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Способ подготовки рафинадного сахара к высушиванию | 0 |
|
SU73A1 |

