Изобретение относится к биотехнологии, к генной инженерии, микробиологической и медицинской промышленности и представляет собой сконструированную in vitro рекомбинантную плазмиду, которая обеспечивает синтез HBsAg, способ конструирования этой рекомбинантной плазмиды и дрожжевой штамм - продуцент поверхностного антигена вируса гепатита В (HBsAg), содержащий сконструированную in vitro плазмиду, которая обеспечивает синтез HBsAg.
Гепатит В (ВГВ) - это широко распространенное инфекционное заболевание, которое характеризуется поражением печени. Ежегодно в мире регистрируется около 50 млн. больных только с острой ВГВ-инфекцией. ВГВ отличается исключительно высокой инфекционностью, а также высокой устойчивостью вируса в условиях окружающей среды и в известных режимах дезинфекции. У 2-10% заболевших острым гепатитом В инфекция принимает характер хронический. Больные ВГВ, как в острой, так и в хронической форме, являются носителями инфекции. Передача вируса происходит при гемотрансфузии, различных парентеральных манипуляциях, половых контактах, вследствие бытовых контактов с больными хроническим гепатитом. При заражении новорожденных от матерей, больных гепатитом В, возникает врожденная ВГВ-инфекция.
При хроническом течении ВГВ-инфекции продолжительные обострения болезни чередуются с ее ремиссией. В последующие годы у 20% больных развивается цирроз печени, в 30% случаев - первичный рак печени (Dego S.F. Eurobiologiste, 1995, v.29, h.63-65). По оценкам экспертов ВОЗ общая численность больных хроническим гепатитом В превышает 1 млрд. человек (Ghendon Y.Z. WHO strategy for control Hepatitis B. In: Control of virus disease, N.-Y., 1993, p.141-164). В Российской Федерации заболеваемость гепатитом В растет: с 18,1 на 100 тыс. населения в 1992 г. до 35,8 в 1996 г. (Аммосов А.Д. Гепатит В, "Вектор-Бест", Кольцово, 2000). Гепатит В давно стал общегосударственной проблемой (экономический ущерб в России составляет не менее 2 млрд. рублей в год) (Борисова В.Н, Мельников В.А. Информ. бюллетень "Вакцинация", 1999, 4).
Учитывая многообразие путей передачи гепатита В и огромное число источников этой инфекции (больные хроническими гепатитами, острыми формами инфекции и особенно носители HBsAg), наиболее перспективным средством профилактики этой инфекции является вакцинация. Вакцинация является единственным средством прерывания естественного механизма передачи вируса от матери-носительницы HBsAg к новорожденному ребенку. Кроме того, иммунизация против гепатита В обеспечивает защиту от заражения вирусом гепатита D. Многолетний опыт применения вакцины против гепатита В в рамках национальных календарей профилактических прививок в ряде стран мира свидетельствует о том, что эта мера обеспечивает снижение заболеваемости гепатитом В и носительства вируса не только у детей и подростков, но и у взрослого населения в 10-20 раз (Бектимиров Т.А. и др. Информ. бюллетень "Вакцинация", 1999, 4).
Благодаря тому, что поверхностные белки ВГВ (HBs белки) обладают сильными протективными иммуногенными свойствами и обеспечивают вируснейтрализующую активность антител класса IgG, первая профилактическая вакцина против гепатита В была сделана на основе очищенного и инактивированного HBsAg из плазмы хронических носителей вируса гепатита В. И хотя такие вакцины все еще применяются в некоторых странах мира, их распространение в целом невелико и постоянно снижается, т.к. требует длительной и дорогостоящей проверки на остаточную инфекционность (Stevens C.E. et al. In: "Viral hepatitis and liver disease", 1984, p.275-291). Эта проблема решается при использовании вакцин, созданных на основе рекомбинантного HBs белка, экспрессированного в клетках дрожжей. В настоящее время для профилактики гепатита В используются рекомбинантные дрожжевые вакцины, характеризующиеся слабой реактогенностью, полной безопасностью и выраженной протективной активностью (Бектимиров ТА. и др. Информ. бюллетень "Вакцинация", 1999, 4).
Первая отечественная рекомбинантная дрожжевая вакцина против гепатита В была создана НПК "Комбиотех Лтд.". Рекомбинантная дрожжевая вакцина против гепатита В производства НПК "Комбиотех Лтд." представляет собой поверхностный антиген вируса гепатита В (HBsAg), выделенный из штамма-продуцента Saccharomyces cerevisiae, сорбированный на геле алюминия гидроксида.
Однако созданные штаммы дрожжей Saccharomyces cerevisiae - продуценты поверхностного антигена вируса гепатита В отличаются низкой продуктивностью. Уровень продукции составляет 5 мг HBsAg на литр культуры (Друца В.Л. и др. Патент РФ RU 2088664, 1997).
Использование для гетерологичной экспрессии метилотрофных дрожжей Pichia pastoris представляет особый интерес, т.к. дает возможность существенно повысить выход рекомбинантных белков (Sudbery Р.Е. Сurr. Opin. Biotechnol., 1996, v.7, p.517-524).
Задачей предлагаемых объектов изобретения является создание рекомбинантной плазмиды, обеспечивающей синтез HВsAg, способа ее конструирования и штамма дрожжей Pichia pastoris, синтезирующего поверхностный антиген вируса гепатита В (HBsAg).
Для решения этой задачи разработан способ конструирования плазмиды, обеспечивающей синтез HBsAg в клетках дрожжей, в результате создан штамм дрожжей Pichia pastoris, синтезирующий поверхностный антиген вируса гепатита В (HBsAg).
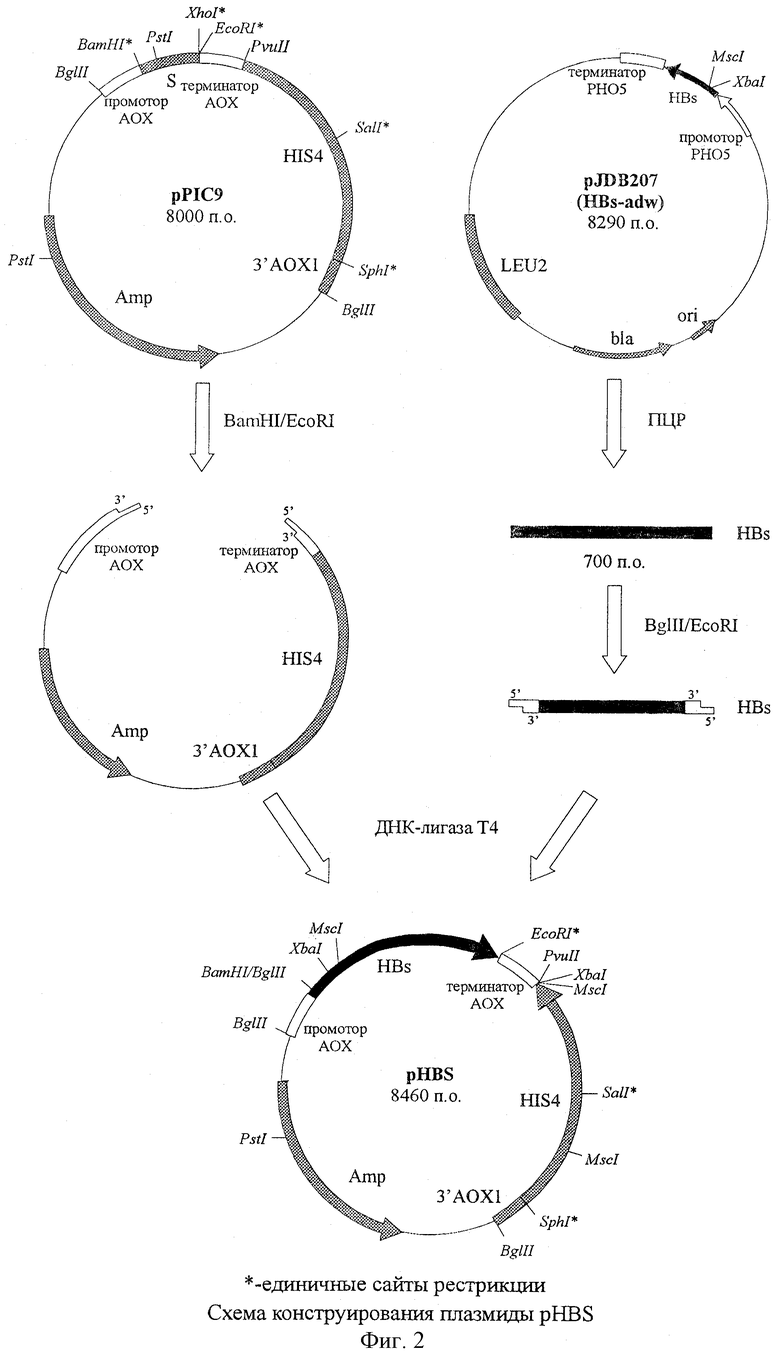
Плазмида pHBS, обеспечивающая синтез HBs белка трансформированными ею клетками дрожжей, состоит из следующих элементов:
- EcoRI-BamHI фрагмент плазмидной ДНК бифункционального бактериально-дрожжевого вектора рРIС9 размером 7,76 т.п.о., включающий бактериальный ген устойчивости к ампициллину, бактериальную область инициации репликации, ген HIS4 дрожжей; фрагмент 5'-некодирующей области дрожжевого гена АОХ1 размером 0, 95 т.п.о., содержащий область, обеспечивающую активацию транскрипции этого гена в присутствии метанола в качестве источника углерода в культуральной среде; фрагмент гена АОХ1 размером 0,33 т.п.о., содержащий область терминации транскрипции этого гена; фрагмент 3'-нетранслируемой области гена АОХ1 размером 0,76 т.п.о.;
- BglII-EcoRI - фрагмент размером 0,70 т.п.о., содержащий кодирующую часть гена HBs белка (adw серотипа).
Сущность изобретения пояснена чертежами, где:
фиг.1 - схема плазмидной ДНК pHBS с рестрикционной картой (знаком * отмечены единичные сайты рестрикции; общий размер плазмиды 8,46 т.п.о.);
фиг.2 - схема конструирования плазмиды pHBS.
Ранее была получена плазмида pJDB207(HBs-adw) (Peterson D.L., Smirnov M. N. Structure of hepatitis В proteins // FIRCA, 5-26471, 1992). Эту плазмиду используют в качестве матрицы для синтеза гена HBs белка (adw серотипа) при помощи ПЦР. В качестве прямого праймера служит олигонуклеотид 5'-gaagatctaaaagaatggagaacatcacat, содержащий сайт для рестриктазы BglII. В качестве обратного праймера используют олигонуклеотид 5'-ggaattccttaaatgtatacccaaagacaa, который содержит сайт для рестриктазы EcoRI. Синтезированный ген HBs белка (adw серотипа) размером 0,70 т.п.о. выделяют при помощи электрофореза в 0,7% агарозном геле, обрабатывают рестриктазами BglII и EcoRI и лигируют с плазмидой рРIС9 ("Invitrogen"), предварительно обработанной рестриктазами BamHI и EcoRI.
Полученной лигазной смесью трансформируют клетки штамма DH5α Escherichia coli (F,/endA1 hsdR17 (rk - mk +) supE44 thi-1 recA1 gyrA (Nalr) relAl Δ(lacZYA-argF)U169 deoR (φ80dlacΔ(lacZ)M15), с помощью рестрикционного анализа отбирают трансформанты, содержащие плазмиду pHBS. Для доказательства идентичности гена HBs белка (adw серотипа), синтезированного при помощи ПЦР, нативному гену HBs белка (adw серотипа) проводят определение его нуклеотидной последовательности (Sanger F.S. et al. Proc. Natl. Acad. Sci., 1977, v.74, p.5463-5467).
Выбор конструкции плазмиды для продукции HBs белка обусловлен следующими причинами. Плазмида pHBS получена на основе челночного бактериально-дрожжевого интегративного вектора рРIС9. В результате трансформации линеаризованной плазмидой и последующей гомологичной рекомбинации происходит встраивание экспрессионной кассеты в хромосому дрожжей Pichia pastoris, что обеспечивает стабильной поддержание клонированного гена HBs белка. В состав плазмиды входит ген HIS4 дрожжей, что позволяет селективно отбирать трансформантов при использовании в качестве рецишгентов штаммов дрожжей с мутациями в этом гене.
Экспрессия гена HBs белка в составе плазмиды pHBS находится под контролем промотора гена АОХ1, содержащим области, обеспечивающие активацию транскрипции в присутствии метанола в культуральной среде, а также область инициации транскрипции. Промотор гена АОХ1 относится к числу наиболее сильных дрожжевых промоторов. Уровень экспрессии генов, находящихся под контролем АОХ1 промотора, эффективно регулируется источниками углерода. Транскрипция гена АОХ1 полностью блокирована при выращивании дрожжей на среде с глюкозой, на среде с глицерином наблюдается только базальный уровень экспрессии гена. Использование метанола в качестве единственного источника углерода значительно усиливает экспрессию гена АОХ1 и, следовательно, генов, находящихся под контролем АОХ1 промотора. Это позволяет регулировать синтез HBs белка в клетках дрожжей. На необходимость подобной регуляции указывают данные, свидетельствующие о токсичности HBs белка для клеток дрожжей (Chen D.-C. et al. Сurr. Genet., 1995, v.27, p.201-206).
В качестве продуцента HBs белка используют штамм PS103(pHBS). Штамм PS103(pHBS) получен при трансформации штамма дрожжей PS103(his4 pep4::ADE2) плазмидой pHBS. Штамм PS103 несет мутацию в гене HIS4, что позволяет селективно отбирать трансформантов, несущих плазмиду pHBS. Мутация в гене РЕР4 приводит к отсутствию активности протеаз А и В, а также карбоксипептидазы Y в клетках дрожжей, что сопровождается повышением стабильности гетерологичных рекомбинантных белков (Hisch H. H. et al. In: Walton E.F., Yarranton G.T. Eds. Molecular and Cell Biology of Yeast, 1989, p.134-200).
Штамм дрожжей Pichia pastoris PS103(pHBS) характеризуется следующими признаками.
Морфологические признаки
Клетки округлой, слегка овальной формы, размером 5-10 мкм, часть клеток имеет на поверхности почки или соединена с дочерними клетками.
Культуральные признаки
Клетки хорошо растут на полной органической среде YEPD - 2% пептона, 1% дрожжевого экстракта, 2% глюкозы или 1% глицерина.
Кроме того, клетки хорошо растут на минеральной среде SC: 1,34% Yeast Nitrogen Base ("Difco", США), 2% глюкозы (1% глицерина, 0,5% метанола), а также на других синтетических средах для дрожжей.
При росте на твердых средах клетки образуют гладкие, круглые колонии с матовой поверхностью, светло-кремового цвета, край неровный.
При росте в жидких средах образуют интенсивную ровную суспензию. Культура имеет характерный запах метилотрофных дрожжей.
Физиолого-биохимические признаки
Клетки растут в пределах от 4 до 37o. Оптимальной температурой выращивания является 30o. При росте в аэробных условиях клетки незначительно защелачивают среду. Оптимум рН для роста составляет 4,5-6,5.
В качестве источника углерода клетки могут использовать многие простые соединения, такие как глюкоза, глицерин, метанол.
В качестве источника азота клетки могут использовать минеральные соли в аммонийной форме, аминокислоты, мочевину.
Клетки способны к аэробному и анаэробному росту.
Существенными признаками штамма является отсутствие потребности в гистидине.
Способ получения плазмиды pJDB207(HBs-adw) проиллюстрирован следующим примером.
Пример 1
Клетки бактерий Escherichia coli, содержащие плазмиду pJDB207(HBs-adw), выращивают при 37o в течение ночи в 1 л питательной среды LB (1% пептона, 0,5% дрожжевого экстракта, 1% хлористого натрия), содержащей ампициллин в концентрации 50 мг/л. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут при 4o, суспендируют в 20 мл 25 мМ трис-хлоридного буфера (рН 8,0), содержащего 10 мМ ЭДТА и 50 мМ глюкозы, добавляют 30 мг лизоцима и инкубируют 10 минут при комнатной температуре. Далее добавляют 40 мл 0,2 М гидроокиси натрия, содержащей 1% додецилсульфата натрия, осторожно перемешивают и инкубируют в течение 10 минут при 4o. Раствор нейтрализуют добавлением 30 мл 3 М ацетата натрия (рН 5,0) и выдерживают в течение 10 минут при 4o. После этого центрифугируют при 14000 об/мин в течение 40 минут при 4o. К супернатанту добавляют 0,6 объема изопропилового спирта, выдерживают 20 минут при комнатной температуре и центрифугируют при 14000 об/мин в течение 20 минут при 20o. Полученный осадок промывают 70% этиловым спиртом, высушивают в вакууме и растворяют в 4 мл дистиллированной воды. Далее добавляют 4,2 г хлористого цезия и 0,36 мл раствора бромистого этидия (10 мг/мл). Полученный раствор выдерживают в течение 1 часа при 4o, затем центрифугируют при 15000 об/мин в течение 15 минут. Супернатант центрифугируют при 70000 об/мин в течение 16 часов в центрифуге TL100 ("Beckman"). После центрифугирования отбирают полосу плазмидной ДНК (нижнюю из двух флюоресцирующих в ультрафиолетовом свете полос), дважды экстрагируют бромистый этидий равным объемом изоамилового спирта, разбавляют в два раза дистиллированной водой и осаждают плазмидную ДНК двумя объемами этилового спирта и 1/15 объема 3 М ацетата натрия (рН 5,0). Осадок собирают центрифугированием при 10000 об/мин в течение 10 минут, промывают 70% этиловым спиртом и растворяют в 0,5-1 мл буфера ТЕ (10 мМ трис-хлоридный буфер, рН 8,0, содержащий 1 мМ ЭДТА). Концентрацию плазмидной ДНК определяют по поглощению раствора при длине волны 260 нм. Чистоту препарата контролируют при помощи электрофореза в 0,7% агарозном геле в буфере ТВЕ (0,1 М трис-боратный буфер, рН 8,3, содержащий 1 мМ ЭДТА).
Полученную плазмиду pJDB207(HBs-adw) используют в качестве матрицы для синтеза гена HBs белка (adw серотипа) при помощи ПЦР.
К 0,1 мкг плазмидной ДНК, растворенной в 5 мкл буфера ТЕ, добавляют 1 мкл 0,5 М NaOH и нагревают при 85o в течение 3 минут. Затем пробу быстро переносят в лед, добавляют 1 мкл 0.5 М НСl и далее используют в ПЦР.
В качестве прямого праймера служит олигонуклеотид 5'-gaagatctaaaagaatggagaacatcacat, содержащий сайт для рестриктазы BglII. В качестве обратного праймера используют олигонуклеотид 5'-ggaattccttaaatgtatacccaaagacaa, который содержит сайт для рестриктазы EcoRI.
Проба для ПЦР содержит 5 мкл матрицы, 30 рМ каждого праймера (по 2 мкл), 10 мкл 10-кратного раствора дезоксинуклеотидтрифосфатов (дНТФ), содержащего 1,25 мМ каждого дНТФ (дАТФ, дТТФ, дГТФ, дЦТФ), 10 мкл 10-кратного буфера для ПЦР (100 mM КСl, 100 mМ (NH4)2SO4, 200 mМ Трис-HCl, рН 8,8, 20 mM MgSO4, 1% тритон Х-100). В пробу добавляется дистиллированная Н2О до конечного объема 100 мкл.
Далее пробу прогревают 5 мин при 95oС, охлаждают, добавляют 2,5 ед. (2,5 мкл) Вент-ДНК-полимеразы ("BioLabs") и проводят 50 циклов ПЦР в следующих условиях: 1 мин при 95o (плавление цепей ДНК), 1 мин при 43o (отжиг праймеров), 1 мин при 72o (полимеразная реакция). После окончания ПЦР пробу инкубируют при 72o 5 мин.
Для доказательства того, что в ходе ПЦР синтезируется ген HBs белка, проводят электрофорез 0,7% агарозном геле в буфере ТВЕ. Реакционную смесь вносят в лунки агарозного геля и проводят разделение фрагментов ДНК в течение 1-2 часов в условиях, описанных выше. По окончании разделения вырезают полоску геля, содержащую фрагмент ДНК размером 0,70 т.п.о., соответствующий гену HBs белка. Выделение ДНК из агарозного геля проводят по методике, разработанной фирмой QIAGEN. Полоску геля с фрагментом ДНК помещают в пробирку и добавляют раствор QX1 (300 мкл на 100 мг геля). Пробу нагревают до 50o, добавляют реактив QIAEX (10 мкл на 5 мкг ДНК) и инкубируют при 50o в течение 10 минут, периодически перемешивая. Далее центрифугируют 30 секунд при 15000 об/мин, супернатант отбрасывают, осадок дважды экстрагируют растворами QX2 и QX3, удаляют супернатант центрифугированием при 15000 об/мин в течение 30 секунд. Осадок высушивают на воздухе, растворяют в 20 мкл буфера ТЕ, центрифугируют 30 секунд при 15000 об/мин, супернатант переносят в новую пробирку.
Гидролиз фрагмента ДНК, содержащего синтезированный ген HBs белка, рестриктазами BglII и EcoRI проводят в 10 мМ трис-хлоридном буфере (рН 7,5), содержащем 50 мМ хлористого натрия, 10 мМ хлористого магния и 1 мМ дитиотреитол. К 5 мкг плазмидной ДНК в объеме 20 мкл добавляют по 5 ед. каждой рестриктазы, после чего пробу инкубируют в течение 25 часов при 37o.
Выделение плазмиды рРIС9 проводят в условиях, аналогичных для плазмиды pJDB207(HBs-adw). Гидролиз ее рестриктазами BamHI и EcoRI осуществляют в условиях, описанных для расщепления синтезированного при помощи ПЦР гена HBs белка. Линеаризованную плазмидную ДНК выделяют по описанной выше методике фирмы QIAGEN, исключая стадию нагревания.
Для получения плазмиды pHBS проводят лигирование плазмиды рРIС9, гидролизованной рестриктазами BamHI и EcoRI, и BglII/EcoRI фрагмента гена HBs белка, синтезированного при помощи ПЦР. Для этого смешивают 0,5 мкг ДНК вектора и 0,1 мкг ДНК встройки в 10 мкл 70 мМ трис-хлоридного буфера (рН7,6), содержащего 5 мМ дитиотреитола, 5 мМ хлористого магния, 1 мМ АТФ, добавляют 10 ед. ДНК-лигазы фага Т4 и инкубируют при 14o в течение ночи.
Полученной лигазной смесью трансформируют клетки штамма DH5α Escherichia coli (F'/endAl hsdR17 (rk - mk +) supE44 thi-1 recAl gyrA (Nalr ) relAl Δ(lacZYA-argF)U169 deoR (φ80dlacΔ(lacZ)M15) Escherichia coli. Для этого клетки Escherichia coli выращивают в 100 мл среды LB при 37o до достижения культурой густоты клеточной суспензии, соответствующей 0,4-0,6 ед. оптической плотности при длине волны 550 нм. Клеточную суспензию охлаждают в ледяной бане, центрифугируют при 5000 об/мин в течение 10 минут при 4o. Клетки супендируют в 100 мл 10 мМ хлористого натрия, собирают центрифугированием в тех же условиях. Далее клетки суспендируют в 50 мл 75 мМ хлористого кальция, выдерживают в ледяной бане в течение 40 минут, осаждают центрифугированием в тех же условиях и суспендируют в 1 мл 75 мМ хлористого кальция. К суспензии компетентных клеток добавляют глицерин до конечной концентрации 15%, разделяют на аликвоты и хранят при -70o. Перед трансформацией суспензию компетентных клеток размораживают в ледяной бане, добавляют лигазную смесь и инкубируют в ледяной бане в течение 40 минут. Далее клетки подвергают действию теплового шока при 42o в течение 2 минут, после чего инкубируют в 1,5 мл среды LB при 37o в течение 1 часа. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут и высевают на чашки Петри со средой LB, содержащей 2% агара и 50 мг/л ампициллина. Чашки инкубируют при 37o в течение 12-16 часов.
Из выросших отдельных клонов трансформантов выделяют плазмидную ДНК при помощи методики, использованной для получения плазмиды pJDB207(HBs-adw), за исключением того, что клетки Escherichia coli выращивают в 10 мл LB, и соответственно объемы всех растворов уменьшают в 100 раз. Кроме того, вместо стадии центрифугирования в градиенте плотности хлористого цезия проводят обработку ДНК панкреатической РНКазой. Для этого нуклеиновые кислоты, осажденные изопропиловым спиртом, растворяют в 100 мкл буфера ТЕ, добавляют 10 мкл раствора РНКазы (1мг/мл) и инкубируют 30 минут при 37o.
Далее проводят гидролиз полученной плазмидной ДНК рестриктазой XbaI и рестриктазой MscI. При последующем электрофорезе в 0,7% агарозном геле искомая плазмида pHBS дает в первом случае фрагменты 1,41 и 7,05 т.п.о., а во втором - 1,40, 1,59 и 5,47 т.п.о. Из выявленного таким образом клона препаративно выделяют плазмиду pHBS так же, как описано для плазмиды pJDB207(HBs-adw), и гидролизуют рестриктазой BglII в 50 мМ трис-хлоридном буфере (рН 7,5), содержащем 100 мМ хлористого натрия, 10 мМ хлористого магния. BglII-фрагмент плазмиды рРIС9, размером 5,63 т.п.о, выделяют по описанной выше методике фирмы QIAGEN, исключая стадию нагревания и используют его для трансформации клеток дрожжей, как описано в примере 2.
Пример 2
Для получения штамма дрожжей Pichia pastoris - продуцента HBs белка, клетки дрожжей штамма PS 103 трансформируют плазмидой pHBS.
Клетки дрожжей выращивают в 100 мл среды YEPD при 30o до достижения культурой оптической плотности, соответствующей 2-4 ед. поглощения при длине волны 600 нм. Клетки дважды промывают стерильной водой, после чего суспендируют в 0,3 мл 100 мМ раствора ацетата лития и инкубируют при 30o в течение 30 минут. К 50 мкл полученной суспензии клеток добавляют 0,1-1 мкг плазмидной ДНК, 50 мкг ДНК спермы лосося, предварительно денатурированной нагреванием (10 минут при 100o) и 0,3 мл раствора 100 мМ ацетата лития, содержащего 40% полиэтиленгликоля 4000. Далее пробу инкубируют 30 минут при 30o и 20 минут при 42o, помещают на 15 секунд в ледяную баню и центрифугируют 10 секунд при 10000 об/мин. Клетки суспендируют в 1 мл стерильной воды и высевают на твердую среду SC. Клоны трансформантов вырастают через 2-3 суток. Выросшие клоны пересевают на чашки со средой SC, содержащей 2% глюкозу, отдельными колониями, затем перепечатывают на среду ММ (1,34% Yeast Nitrogen Base ("Difco", США), 0,5% метанола, 2% агара ("Difco", США) для отбора трансформантов, не растущих на среде с метанолом, что свидетельствует об интеграции чужеродного гена в локус AOXI (фенотип Met-).
Для анализа продукции HBs белка клетками трансформантов фенотипа Met- их выращивают при 30o в 100 мл жидкой среды BMGY (2% пептона, 1% дрожжевого экстракта, 1% глицерина, 10 мл 1М калий-фосфатного буфера, рН 6,0, 1,34% Yeast Nitrogen Base ("Difco", США) до стационарной фазы роста в течение 2 суток. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут, супернатант сливают и переносят всю биомассу в 20 мл жидкой среды BMMY (2% пептона, 1% дрожжевого экстракта, 0,5% метанола, 10 мл 1 М калийфосфатного буфера, рН 6,0, 1,34% Yeast Nitrogen Base ("Difco", США) для индукции экспрессии HBs белка. Индукцию проводят при 30o в течение 4 суток.
По окончании индукции клетки дрожжей собирают центрифугированием, суспендируют в равном объеме буфера для вскрытия (50 мМ Na-фосфатный буфер рН 8,0, 5 мМ ЭДТА, 2 мМ PMSF, 0,5% Твин-20), добавляют стеклянные шарики (⊘ 0,1 мм) и разрушают на дезинтеграторе "Dyno-Mill" в течение 10 минут, далее центрифугируют 10 мин при 3000 об/мин, для удаления неразрушенных клеток. В супернатанте определяют присутствие HBsAg иммуноферментным методом и при помощи электрофореза в полиакриламидном геле.
Для определения содержания HBsAg методом иммуноферментного анализа используют тест-систему Вектогеп B-HBs-антиген-стрип (ЗАО "Вектор-Бест"). Тестируемые пробы разводят в соотношениях от 1:10 до 1:100 10 мМ К-фосфатным буфером, рН 6,7 и помещают в лунки планшета для иммуноферментного анализа. Далее пробы инкубируют при 37o в течение часа, лунки промывают 5 раз буфером ФСБ-Т (10 мМ К-фосфатный буфер, рН 6,7, содержащий 137 мМ NaCl, 2,7 мМ KCl, 0,05% Твин-20) и вносят в лунки раствор моноклональных антител к HBsAg, меченных пероксидазой хрена. Затем пробы инкубируют при 37o в течение часа, промывают буфером ФСБ-Т, добавляют по 100 мкл субстрата ТМБ (тетраметилбензидин), выдерживают в темноте в течение 15-20 минут при комнатной температуре. Реакцию останавливают 50 мкл 1 М Н2SO4 и измеряют оптическую плотность при 450 нм.
Согласно полученным таким образом данным клетки дрожжей штамма PS103(pHBS) синтезируют HBsAg, дающий положительную реакцию с моноклональными антителами к нативному HBsAg. В то же время в клетках дрожжей штамма PS103, трансформированных плазмидой рРIС9, отсутствует белок, иммунологически родственный нативному HBsAg.
Для контроля продукции HBs белка клетками дрожжей штамма PS103(pHBS) используют также метод электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия. Разделение белков проводят в 15% полиакриламидном геле в стандартной системе буферов (электродный буфер: 25 мМ трис, 192 мМ глицин, 0,1% додецилсульфат натрия, рН 8,3; буфер для геля: 375 мМ трис-хлоридный буфер, рН 8,8). Параллельно проводят разделение белков контрольного штамма (PS103, трансформированного плазмидой рРIС9), выращенного в идентичных условиях. В качестве стандартов молекулярной массы используют карбоангидразу, ингибитор трипсина, миоглобин, лизоцим. По окончании электрофореза гели окрашивают 0,15% раствором кумасси G250 в 25% изопропаноле и 10% уксусной кислоте и отмывают в 10% уксусной кислоте. При сравнении спектра белков двух штаммов у штамма PS103(pHBS) обнаруживают появление двух дополнительных белковых полос с молекулярной массой 28 КДа и 24 КДа, что соответствует молекулярной массе гликозилированного и негликозилированного HBs белка. Уровень синтеза HBs белка определяют, сравнивая интенсивность окрашивания полосы рекомбинантного белка с полосой соответствующего белка - стандарта молекулярной массы.
Согласно полученным данным клетки дрожжей штамма PS103(pHBS) синтезируют около 30 мг-40 мг HBs белка на литр культуры дрожжей.
Суммируя вышесказанное можно заключить, что полученный штамм дрожжей Pichia pastoris PS103(pHBS) синтезирует HBsAg в количестве, достаточном для его очистки в лабораторном масштабе. В результате такой очистки может быть получен рекомбинантный HBsAg, пригодный для создания рекомбинантной дрожжевой вакцины против гепатита В. Преимуществом данного продуцента по сравнению с прототипом ВКМП Y-2203 является более высокий уровень продукции HBs белка.
Штамм дрожжей Pichia pastoris PS103(pHBS) - продуцент поверхностного антигена вируса гепатита В (HBsAg) депонирован во Всероссийской коллекции промышленных микроорганизмов под номером ВКПМ Y-2532.
Изобретение относится к биотехнологии, к генной инженерии, микробиологической и медицинской промышленности и представляет собой сконструированную in vitro рекомбинантную плазмиду, которая обеспечивает синтез HBsAg. Дрожжевой штамм-продуцент поверхностного антигена вируса гепатита В (HBsAg), содержащий сконструированную in vitro плазмиду, обеспечивает синтез HBsAg. Полученный штамм дрожжей Pichia pastoris PS103 (pHBS) - продуцент поверхностного антигена вируса гепатита В (HBsAg) депонирован во Всероссийской коллекции промышленных микроорганизмов под номером ВКПМ Y-2532. Продуцируемый плазмидой поверхностный антиген вируса гепатита В (HBsAg) в штамме Pichia pastoris PS103 пригоден для получения вакцины против гепатита В. 3 с.п.ф-лы, 2 ил.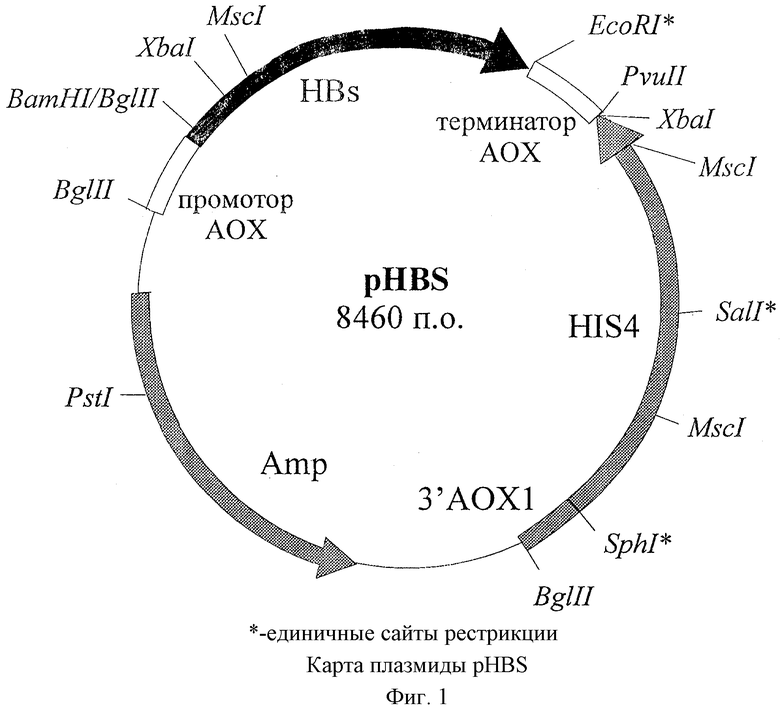
| US 4895800 А1, 23.01.1990 | |||
| RU 1623207 А3, 15.03.1994 | |||
| US 6103519 А1, 15.08.2000. |

