Изобретение относится к биотехнологии, генной инженерии, микробиологической и медицинской промышленности и представляет собой сконструированную in vitro рекомбинантную плазмидную ДНК, которая обеспечивает синтез фибробластного интерферона человека в клетках дрожжей Saccharomyces cerevisiae, способ конструирования этой плазмидной ДНК и дрожжевой штамм - продуцент фибробластного интерферона человека, содержащий эту плазмиду.
Фибробластный интерферон человека, бета-интерферон, относится к обширной группе эволюционно родственных белков, получивших название интерферонов. Образование интерферонов стимулируется воздействием вирусов на различные группы клеток. Интерфероны способны активировать противовирусные защитные механизмы клетки. Клинические испытания показали возможность использования бета-интерферона для лечения ряда вирусных и аутоиммунных заболеваний человека (Ершов Ф.И. Система интерферона в норме и патологии, 1996). Получены обнадеживающие результаты применения бета-интерферона при рассеянном склерозе (Paty D.W. et al. Neurology, 1993, v.43, р.662-667). Однако систематические исследования спектра биологического действия бета-интерферона, как и его применение, в клинике затруднены, так как высокоочищенные препараты этого белка практически отсутствуют. Видоспецифичность интерферонов не позволяет использовать в клинических испытаниях препараты бета-интерферона других млекопитающих. Использование в качестве источника бета-интерферона донорской крови или культуры фибробластов не обеспечивает нужных количеств белка, а кроме того, несет опасность заражения гепатитом и ВИЧ.
Перспективным подходом для получения бета-интерферона человека в значительных количествах является использование микроорганизмов в качестве продуцентов этого белка. Существует ряд штаммов Escherichia соli (кишечной палочки), продуцирующих бета-интерферон человека. Уровень продукции колеблется от 1,5 до 6000 млн ед. биологически активного бета-интерферона на литр культуры и составляет в последнем случае несколько десятков миллиграммов (Taniguchi Т. et al. Nature, 1980, v.285, p.547-549; Goeddel D.V. ct al. Nucl. Acids Res. , 1980, v.8, p.4057-4074; Remaut E. et al. Nucl. Acids Res., 1983, v.7, p.4677-4688). Содержание бета-интерферона в клетках может достигать около 2% суммарного клеточного белка. Однако Escherichia coli являются условно патогенными для человека, и препараты бета-интерферона, полученные из клеток кишечной палочки, могут содержать эндотоксины. Полное освобождение бета-интерферона от примеси эндотоксинов, являющееся обязательным условием применения его в клинической практике, значительно затрудняет процедуру очистки рекомбинантного белка. Например, в настоящее время зарегистрирован и разрешен к применению препарат рекомбинантного бета-интерферона человека, экспрессированного в Escherichia соli, производимый фирмой "Шеринг". Длительное применение его может приводить к нежелательным побочным эффектам, обусловленным условной патогенностью штамма-продуцента (Neilley L.K. et al. Neurology, 1996, v.46, р.552-554; Brod S.A. et al. Neurology, 1996, v.46, p.1633-1638; Huber S. et al. Schweiz. Med. Wochenschr., 1996, v.126, p.1475-1481).
Все вышеизложенное свидетельствует о перспективности создания штаммов-продуцентов бета-интерферона человека на основе непатогенных микроорганизмов, в частности дрожжей Saccharomyces cerevisiae. Дрожжи-сахаромицеты - эукариотические микроорганизмы, хорошо изученные генетически. Они не содержат токсинов, имеют короткий жизненный цикл, их легко культивировать. Кроме того, экспрессированные в них белки высших эукариота претерпевают нормальные процессы созревания, гликозилирования и т.д. Ранее в нашей лаборатории был получен штамм-продуцент секреторного фибробластного бета-интерферона человека на основе штамма дрожжей Saccharomyces cerevisiae GRF18 (MATalpha leu2-3,l12 his3-11,15), трансформированного плазмидой pAC137(IFN-b). Экспрессия гена бета-интерферона человека находилась под контролем гибридного дрожжевого промотора PHО5-MFalphal, обеспечивающего регуляцию синтеза бета-интерферона неорганическим фосфатом и секрецию продукта в культуральную среду. Клетки вышеупомянутого штамма ВКПМ-У876 синтезируют только 500 мкг бета-интерферона внутриклеточно и около 100 мкг белка секретируют в культуральную жидкость (на 1 литр культуры клеток дрожжей). Кроме того, углеводный компонент белков, экскретируемых дрожжами-сахаромицетами, существенно отличается от такового гликопротеинов высших эукариота (Kukuruzinska M. A. et al. Ann. Rev. Biochem., 1987, v. 56, p. 915-944). В связи с тем, что углеводная часть гликопротеинов является сильной антигенной детерминантой, использование рекомбинантного бета-интерферона человека, секретируемого дрожжами Saccharomyces cerevisiae, возможно только для лабораторных целей (Ballon C.E. J.Biol.Chem, 1970, v.245, p.1197-1203).
Целью предлагаемых объектов изобретения является создание штамма дрожжей Saccharomyces cerevisiae, синтезирующего бета-интерферон человека внутриклеточно.
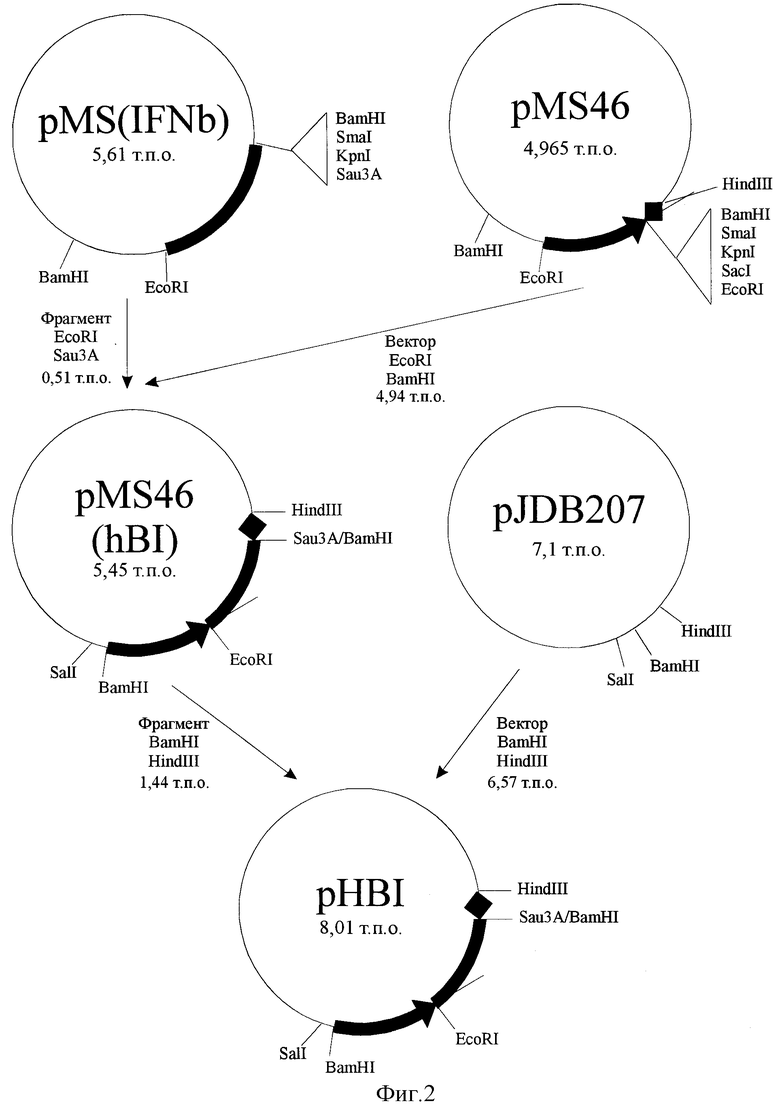
Плазмида pHBI, обеспечивающая синтез бета-интерферона человека трансформированными ею клетками дрожжей, состоит из следующих элементов:
- фрагмент ДНК челночного бактериально-дрожжевого вектора pJDB207 размером 6,57 т.п.о., ограниченный сайтами рестрикции BamHI и HindIII, включающий бактериальный ген устойчивости к ампициллину, бактериальную область инициации репликации, фрагмент дрожжевой двухмикронной плазмиды и ген LEU2 дрожжей;
- BamHI-EcoRI - фрагмент 5'-нeкодирующей области дрожжевого гена РНО5 размером 0,54 т.п.о., содержащий область, обеспечивающую активацию транскрипции этого гена в отсутствие неорганического фосфата в культуральной среде, область инициации транскрипции;
- EcoRI-Sau3a - фрагмент размером 0,51 т.п.о., содержащий кодирующую часть гена бета-интерферона человека за исключением области, кодирующей сигнальный пептид;
- Sau3a-PstI - фрагмент гена РНО5 дрожжей размером 0,38 т.п.о., содержащий область терминации транскрипции этого гена;
- PstI-HindIII - фрагмент полилинкерного участка плазмиды pUC19 размером 0,01 т.п.о.
Схема плазмидной ДНК pHBI с рестрикционной картой изображена на фиг. 1. Общий размер плазмиды 8,0 т.п.о.
Для достижения цели используют способ конструирования плазмиды, обеспечивающей синтез бета-интерферона в клетках дрожжей. Схема конструирования плазмиды pHBI приведена на фиг. 2. Плазмиду pMS(IFNb), полученную ранее в нашей лаборатории (Авторское свидетельство SU 1530749 А1, 1989, Останин К.В. , Мясников А. Н. , Смирнов М.Н., Машко С.В., Лебедева М.И. Рекомбинантная плазмидная ДНК рАС137 IFN-бета, обеспечивающая синтез и секрецию фибробластного интерферона человека, способ ее получения и штамм дрожжей Saccharomyces cerevisiae - продуцент фибробластного интерферона человека), обрабатывают рестриктазами EcoRI и Sau3a, с помощью электрофореза в 0,8% агарозном геле выделяют фрагмент 0,51 т.п.о., содержащий ген бета-интерферона человека без сигнального пептида. Этот фрагмент лигируют с плазмидой pMS46 (Останин К.В. и др. Биополимеры и клетка, 1988, т.4, с.272-279), гидролизованной рестриктазами EcoRI и BamHI. Для рестриктазы BamHI подбирают условия ограниченного гидролиза. При помощи электрофореза в агарозном геле выделяют фрагмент размером 4,94 т.п.о. и используют его в реакции лигирования. Полученной лигазной смесью трансформируют клетки штамма DH5α Escherichia coli (F' /endAl hsdR17 (rk -mk +) supE44 thi-1 recAl gyrA (Nalr)reiAl Δ(IacZYA-argF)U169 doR θ80dlacΔ(lacZ)M15) и с помощью рестрикционного анализа отбирают трансформанты, содержащие плазмиду pMS46(HBI). Данную плазмиду обрабатывают рестриктазами ВаmHI и HindIII и при помощи электрофореза в агарозном геле выделяют фрагмент размером 1,44 т.п.о. Данный фрагмент лигируют с плазмидой pJDB207 (Beggs J.D., Nature, 1978, v.275, р.104), предварительно обработанной рестриктазами ВаmHI и HindIII. Полученной лигазной смесью трансформируют клетки штамма DH5α Escherichia coli и с помощью рестрикционного анализа отбирают те трансформанты, которые содержат плазмиду pHBI.
Выбор конструкции плазмиды для продукции бета-интерферона обусловлен следующими причинами. Плазмида pHBI получена на основе челночного бактериально-дрожжевого вектора pJDB207, который является одним из наиболее стабильных и высококопийных векторов дрожжей. В его состав входит ген LEU2 дрожжей, что позволяет селективно отбирать трансформантов при использовании в качестве реципиентов штаммов дрожжей с мутациями в этом гене. Плазмиды, созданные на основе pJDB207, способны поддерживать рост трансформантов на среде без лейцина только в том случае, если в клетке содержится 100-200 копий плазмиды.
Экспрессия гена бета-интерферона человека в составе плазмиды pHBI находится под контролем промотора гена РНО5, содержащего области, обеспечивающие активацию транскрипции при отсутствии неорганического фосфата в культуральной среде, а также область инициации транскрипции. Промотор гена РНО5 относится к числу наиболее сильных дрожжевых промоторов. Уровень экспрессии генов, находящихся под контролем РНО5 промотора, эффективно регулируется экзогенным неорганическим фосфатом. При концентрации неорганического фосфата в культуральной среде 1 г/л наблюдается только базальный уровень экспрессии. Это позволяет регулировать синтез бета-интерферона в клетках дрожжей. На необходимость подобной регуляции указывают полученные нами данные о токсичности бета-интерферона для клеток дрожжей.
В качестве продуцента бета-интерферона используют штамм p4-l-GRF18(pHBI). Штамм p4-l-GRF18(pHBI) получен при трансформации штамма дрожжей p4-l-GRF18 (MATalpha leu2-3,112 his3-11,15 pho3 pep4) плазмидой pHBI. Штамм p4-l-GRF18 несет мутацию в гене LEU2, что позволяет селективно отбирать трансформантов, несущих плазмиду pHBI. Делеционно получали мутацию в гене PEP4, приводящую к отсутствию активности протеаз А и В, а также карбоксипептидазы Y в клетках дрожжей, что сопровождается повышением стабильности гетерологичных рекомбинантных белков (Hisch H.H. et al. In: Walton E.F., Yarranton G.T., Eds., Molecular and Cell Biology of Yeast, 1989, p.134-200).
Штамм дрожжей Saccharomyces cerevisiae p4-l-GRF18(pHBI) характеризуется следующими признаками.
Морфологические признаки.
Клетки округлой, слегка овальной формы, размером 5-10 мкм, часть клеток имеет на поверхности почки или соединена с дочерними клетками.
Культуральные признаки.
Клетки хорошо растут на полных органических средах:
ПЕП - 2% пептона, 2% глюкозы,
ПЕПФо - 2% пептона, 2% глюкозы, 0,1% однозамещенного фосфата калия,
YEPD - 2% пептона, 2% глюкозы, 1% дрожжевого экстракта.
Кроме того, клетки хорошо растут на минеральной среде Sd: 0,67% Yeast Nitrogen Base ("Difco", США), 2% глюкозы, 50 мг/л гистидина, а также на других синтетических средах для дрожжей, содержащих 50 мг/л гистидина.
При росте на твердых средах клетки образуют гладкие, круглые колонии с матовой поверхностью, светло-кремового цвета, край ровный.
При росте в жидких средах образуют интенсивную ровную суспензию. Культура имеет характерный запах дрожжей.
Физиолого-биохимические признаки.
Клетки растут в пределах от 4oС до 37oС. Оптимальной температурой выращивания является 30oС. При росте в аэробных условиях клетки значительно закисляют среду. Оптимум рН для роста составляет 3,5-5,5.
В качестве источника углерода клетки могут использовать многие простые соединения, такие как глюкоза, сахароза, глицерин.
В качестве источника азота при условии добавки гистидина клетки могут использовать минеральные соли в аммонийной форме, аминокислоты, мочевину.
Клетки способны к аэробному и анаэробному росту.
Существенными признаками штамма является ауксотрофность по гистидину и отсутствие потребности в лейцине.
Способ конструирования плазмиды pHBI проиллюстрирован следующими примерами.
ПРИМЕР 1.
Клетки бактерий Escherichia coli, содержащие плазмиду pMS(IFNb), выращивают в течение ночи в 1 л питательной среды LB (1% пептона, 0,5% дрожжевого экстракта, 1% хлористого натрия), содержащей ампициллин в концентрации 50 мг/л. Клетки собирают центрифугированием при 5000 об./мин в течение 10 минут при 4oС, осадок суспендируют в 20 мл 25 мМ трис-хлоридного буфера (рН 8,0), содержащего 10 мМ ЭДТА и 50 мМ глюкозы. К суспензии добавляют 30 мг лизоцима и инкубируют 10 минут при комнатной температуре. Далее добавляют 40 мл 0,2 М раствора NaOH, содержащего 1% додецилсульфата натрия, осторожно перемешивают и инкубируют в течение 10 минут при 4oС. Раствор нейтрализуют добавлением 30 мл 3 М ацетата натрия (рН 5,0) и выдерживают в течение 10 минут при 4oС. После этого центрифугируют при 14000 об./мин в течение 40 минут при 4oС. К супернатанту добавляют 0,6 объема изо-пропилового спирта, выдерживают 20 минут при комнатной температуре и центрифугируют при 14000 об./мин в течение 20 минут при 20oС. Полученный осадок промывают 70% этиловым спиртом, высушивают в вакууме и растворяют в 4 мл дистиллированной воды. Далее добавляют 4,2 г хлористого цезия и 0,36 мл раствора бромистого этидия (10 мг/мл). Полученный раствор выдерживают в течение 1 часа при 4oС, затем центрифугируют при 15000 об. /мин в течение 15 минут. Супернатант центрифугируют при 70000 об./мин в течение 16 часов в центрифуге TL-100 ("Beckman"). После центрифугирования отбирают полосу плазмидной ДНК (нижнюю из двух флюоресцирующих в ультрафиолетовом свете полос), дважды экстрагируют бромистый этидий равным объемом изо-амилового спирта, разбавляют в два раза дистиллированной водой и осаждают плазмидную ДНК двумя объемами этилового спирта и 1/15 объема 3 М ацетата натрия (рН 5,0). Осадок собирают центрифугированием при 10000 об./мин в течение 10 минут, промывают 70% этиловым спиртом и растворяют в 0,5-1 мл буфера ТЕ (10 мМ трис-хлоридный буфер, рН 8,0, содержащий 1 мМ ЭДТА). Концентрацию плазмидной ДНК определяют по поглощению раствора при длине волны 260 нм. Чистоту препарата контролируют при помощи электрофореза в 0,7% агарозном геле в буфере ТВЕ (0,1 М трис-боратный буфер, рН 8,3, содержащий 1 мМ ЭДТА).
Гидролиз плазмиды pMS(IFNb) рестриктазами EcoRI и Sau3a проводят в 50 мМ трис-хлоридном буфере (рН 7,5), содержащем 100 мМ хлористого натрия, 10 мМ хлористого магния и 0,02% Triton Х-100. К 50 мкг плазмидной ДНК в объеме 100 мкл добавляют по 50 ед. каждой рестриктазы, после чего пробу инкубируют в течение 6-12 часов при 37oС. Полноту гидролиза контролируют при помощи электрофореза в 0,7% агарозном геле. Реакционную смесь вносят в лунки агарозного геля и проводят разделение фрагментов ДНК в течение 1-2 часов в условиях, описанных выше. По окончании разделения вырезают полоску геля, содержащую фрагмент ДНК размером 0,51 т.п.о., соответствующий гену бета-интерферона человека без сигнальной последовательности.
Выделение ДНК из агарозного геля проводят по методике, разработанной фирмой QIAGEN. Полоску геля с фрагментом ДНК помещают в пробирку и добавляют раствор QX1 (300 мкл на 100 мг геля). Пробу нагревают до 50oС, добавляют реактив QIAEX (10 мкл на 5 мкг ДНК) и инкубируют при 50oС в течение 10 минут, периодически перемешивая. Далее центрифугируют 30 секунд при 15000 об. /мин, супернатант отбрасывают, осадок дважды экстрагируют растворами QX2 и QX3, удаляют супернатант центрифугированием при 15000 об./мин в течение 30 секунд. Осадок высушивают на воздухе, растворяют в 20 мкл буфера ТЕ, центрифугируют 30 секунд при 15000 об./мин, супернатант переносят в новую пробирку.
Выделение плазмиды pMS46 и гидролиз ее рестриктазой EcoRI проводят в условиях, описанных выше для плазмиды pMS(IFNb). Линеаризованную плазмидную ДНК выделяют по описанной выше методике фирмы QIAGEN, исключая стадию нагревания. Далее плазмиду подвергают неполному гидролизу рестриктазой ВаmHI в 10 мМ трис-хлоридном буфере (рН 8,0), содержащем 100 мМ хлористого калия, 10 мМ хлористого магния и 0,02% Triton Х-100. К 50 мкг плазмидной ДНК в объеме 100 мкл добавляют 50 ед. рестриктазы BamHI и инкубируют при 37oС. Контроль за ходом реакции осуществляют каждые 20 минут при помощи электрофореза в 0,7% агарозном геле. Полученные фрагменты ДНК разделяют электрофорезом в 0,7% геле агарозы и выделяют фрагмент размером 4,94 т.п.о. по приведенной выше методике.
Для получения плазмиды pMS46(hBI) проводят лигирование плазмиды pMS46, гидролизованной рестриктазами EcoRI и BamHI, и фрагмента плазмиды pMS(IFNb), соответствующего гену бета-интерферона человека. Для этого смешивают 0,45 мкг ДНК вектора и 0,1 мкг ДНК встройки в 10 мкл 70 мМ трис-хлоридного буфера (рН 7,6), содержащего 5 мМ дитиотреитола, 5 мМ MgCl2, 1 мМ АТФ, добавляют 10 ед. ДНК-лигазы фага Т4 и инкубируют при 14oС в течение ночи.
Полученной лигазной смесью трансформируют клетки штамма DH5α Escherichia coli. Для этого клетки Escherichia coli выращивают в 100 мл среды LB при 37oС до достижения культурой густоты клеточной суспензии, соответствующей 0,4-0,6 ед. оптической плотности при длине волны 550 нм. Клеточную суспензию охлаждают в ледяной бане, центрифугируют при 5000 об./мин в течение 10 минут при 4oС. Клетки суспендируют в 100 мл 10 мМ хлористого натрия, собирают центрифугированием в тех же условиях. Далее клетки суспендируют в 50 мл 75 мМ хлористого кальция, выдерживают в ледяной бане в течение 40 минут, осаждают центрифугированием в тех же условиях и суспендируют в 1 мл 75 мМ хлористого кальция. К суспензии компетентных клеток добавляют глицерин до конечной концентрации 15%, разделяют на аликвоты и хранят при -70oС. Перед трансформацией суспензию компетентных клеток размораживают в ледяной бане, добавляют лигазную смесь и инкубируют в ледяной бане в течение 40 минут. Далее клетки подвергают действию теплового шока при 42oС в течение 2 минут, после чего инкубируют в 1,5 мл среды LB при 37oС в течение 1 часа. Клетки собирают центрифугированием при 5000 об./мин в течение 10 минут и высевают на чашки Петри со средой LB, содержащей 2% агара и 50 мг/л ампициллина. Чашки инкубируют при 37oС в течение 12-16 часов.
Из выросших отдельных клонов трансформантов выделяют плазмидную ДНК при помощи методики, использованной для получения плазмиды pMS(IFNb), за исключением того, что клетки Escherichia coli выращивают в 10 мл LB, и соответственно объемы всех растворов уменьшают в 100 раз. Кроме того, вместо стадии центрифугирования в градиенте плотности хлористого цезия проводят обработку ДНК панкреатической РНКазой. Для этого нуклеиновые кислоты, осажденные изо-пропиловым спиртом, растворяют в 100 мкл буфера ТЕ, добавляют 10 мкл раствора РНКазы (1 мг/мл) и инкубируют 30 минут при 37oС. Далее проводят гидролиз полученной плазмидной ДНК рестриктазами EcoRI и HindIII в условиях, описанных ранее для расщепления плазмиды pMS(IFNb). При рестрикции искомой плазмиды pMS46(hBI) смесью рестриктаз EcoRI-HindIII и последующего электрофореза в 0,7% агарозном геле обнаруживаются фрагменты 0,89 т.п.о. и 4,56 т.п. о. Из выявленного таким образом клона препаративно выделяют плазмиду pMS46(hBI) размером 5,45 т.п.о. так же, как описано для плазмиды pMS(IFNb).
Для получения плазмиды pHBI с помощью описанных выше методов препаративно выделяют BamHI-HindIII-фрагмент плазмиды pMS46(hBI) размером 1,44 т.п.о., который содержит промотор гена РНО5 дрожжей, ген бета-интерферона человека и терминатор транскрипции гена РНО5 дрожжей, и BamHI-HindIII-фрагмент плазмиды pJDB207 размером 6,57 т.п.о. Так же, как и при получении плазмиды pMS46(hBI), проводят лигирование этих фрагментов и трансформацию клеток Escherichia coli. Клон, содержащий искомую плазмиду, отбирают с помощью расщепления ДНК рестриктазами SalI и HindIII. При этом образуются фрагменты размером 1,72 т. п. о. и 6,29 т.п.о. Из клеток идентифицированного клона препаративно выделяют плазмидную ДНК по приведенным ранее методикам и используют ее для трансформации клеток дрожжей, как описано в примере 2.
ПРИМЕР 2.
Для получения штамма дрожжей Saccharomyces cerevisiae - продуцента бета-интерферона человека, клетки дрожжей штамма p4-l-GRF18 трансформируют плазмидой pHBI. Клетки дрожжей выращивают в 100 мл среды YEPD до достижения культурой оптической плотности, соответствующей 2-4 ед. поглощения при длине волны 600 нм. Клетки дважды промывают стерильной водой, после чего суспендируют в 0,3 мл 100 мМ раствора ацетата лития и инкубируют при 30oС в течение 30 минут. К 50 мкл полученной суспензии клеток добавляют 0,1-1 мкг плазмидной ДНК, 50 мкг ДНК спермы лосося, предварительно денатурированной нагреванием (10 минут при 100oС) и 0,3 мл раствора 100 мМ ацетата лития, содержащего 40% полиэтиленгликоля PEG-4000. Далее пробу инкубируют 30 минут при 30oС и 20 минут при 42oС, помещают на 15 секунд в ледяную баню и центрифугируют 10 секунд при 10000 об./мин. Клетки суспендируют в 1 мл стерильной воды и высевают на твердую среду Sd, содержащую 50 мг/л гистидина. Клоны трансформантов вырастают через 4-6 суток.
Для анализа продукции бета-интерферона клетками трансформантов их выращивают в 50 мл жидкой среды ПЕП до стационарной фазы роста. Клетки собирают центрифугированием при 3000 об./мин в течение 10 минут, промывают водой, суспендируют в 1 мл 50 мМ трис-хлоридного буфера (рН 7,5), содержащего 1 мМ ФМСФ (фенилметилсульфонилфторид), добавляют 1 г стеклянных бус (0,5 мм) и разрушают в дезинтеграторе "Braun" в течение 1 минуты при температуре 4oС. Полученный гомогенат центрифугируют при 10000 об./мин в течение 10 минут, осадок промывают 10-кратным объемом того же буфера и суспендируют в 1 мл 50 мМ натрий-фосфатного буфера (рН 7,0), содержащего 2% додецилсульфата натрия и 5% 2-меркаптоэтанола, и инкубируют 5 минут в кипящей водяной бане. По окончании инкубации пробу центрифугируют 15 минут при 12000 об./мин и в супернатанте определяют содержание бета-интерферона при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия и последующей гибридизации со специфическими антителами к бета-интерферону человека. Разделение белков проводят в 15% полиакриламидном геле в стандартной системе буферов (электродный буфер: 25 мМ трис, 192 мМ глицин, 0,1% додецилсульфат натрия, рН 8,3; буфер для геля: 375 мМ трис-хлоридный буфер, рН 8,8). Параллельно проводят разделение белков контрольного штамма p4-1-GRF18, выращенного и вскрытого в идентичных условиях. В качестве стандартов молекулярной массы используют карбоангидразу, ингибитор трипсина, миоглобин, лизоцим фирмы "Sigma". По окончании электрофореза белки ренатурируют, выдерживая гели 15 минут в 10 мМ трис-хлоридном буфере (рН 7,5), содержащем 4 М мочевину, 20 мМ ЭДТА. Далее белки переносят на нитроцеллюлозную мембрану в 25 мМ трис-192 мМ глициновом буфере (рН 8,3), содержащем 20% метилового спирта, при 30-40В, в течение 1,5 часов на приборе фирмы "Bio-Rad". Далее мембрану выдерживают в буфере TBST (10 мМ трис-хлоридный буфер (рН 8,0), 150 мМ хлористого натрия, 0,05% твин-20, 1% бычьего сывороточного альбумина) в течение 2 часов при 37oС. Затем помещают мембрану в тот же буфер, содержащий разведенные в 1000 раз лошадиные поликлональные антитела к бета-интерферону человека ("Boehringer"), и инкубируют 2 часа при 37oС. Далее трижды промывают мембрану буфером TBST и инкубируют 1 час при 37oС с разбавленным в 7000 раз конъюгатом видоспецифических антител к иммуноглобулинам лошади и пероксидазы хрена ("Sigma"). После отмывки мембраны буфером PBST (58 мМ двузамещенного фосфата натрия, 17 мМ однозамещенного фосфата натрия, 68 мМ хлористого натрия, 0,1% Твин-20) ее помещают в раствор субстратов для пероксидазы: 0,02% DAB (3, '3-диаминобензидин тетрагидрохлорид), 0,006% перекись водорода в 10 мМ трис-хлоридном буфере, рН 7,5. Параллельно окрашивают гели 0,15% раствором кумасси G250 в 25% изопропаноле и 10% уксусной кислоте и отмывают в 10% уксусной кислоте. При сравнении спектра белков двух штаммов у штамма p4-l-GRF18(pHBI) обнаруживают появление дополнительной белковой полосы, дающей положительную реакцию с антителами к бета-интерферону человека, с молекулярной массой 20 кДа, что соответствует молекулярной массе бета-интерферона. Уровень синтеза рекомбинантного бета-интерферона определяют, сравнивая интенсивность окрашивания полосы этого белка с полосой стандартного бета-интерферона.
Согласно полученным данным клетки дрожжей штамма p4-l-GRF18(pHBI) синтезируют внутриклеточно около 1 мг бета-интерферона на литр культуры дрожжей.
При проведении анализа на подавление цитопатической активности вируса везикулярного стоматита в культуре человеческих фибробластов стандартными методами (M. G.Tovey et al., J. Gen. Virol., 1977, 36, 341-344) обнаруживается, что препарат бета-интерферона, синтезированного штаммом дрожжей Saccharomyces cerevisiae p4-l-GRJF18(pHBI), биологически активен. Активность препарата составляет 20-40 млн ед./л.
Суммируя вышесказанное, можно заключить, что полученный штамм дрожжей Saccharomyces cerevisiae p4-l-GRF18(pHBI) синтезирует бета-интерферон человека в количестве, достаточном для его очистки в лабораторном масштабе. В результате такой очистки могут быть получены препараты бета-интерферона, полезные для исследования его биологических свойств и имеющие терапевтическую ценность. Преимуществом данного продуцента по сравнению с прототипом ВКМП-Y-876 является более высокий уровень продукции бета-интерферона и стабильность рекомбинантного белка. Кроме того, он обеспечивает внутриклеточный синтез бета-интерферона, вследствие чего рекомбинантный белок не содержит дополнительных маннозных остатков, характерных для белков, секретируемых дрожжами-сахаромицетами.
Штамм дрожжей Saccharomyces cerevisiae p4-l-GRF18(pHBI) - продуцент бета-интерферона человека депонирован во Всероссийской коллекции промышленных микроорганизмов под номером ВКПМ-Y-2286.
Изобретение относится к биотехнологии, генной инженерии, микробиологической и медицинской промышленности. Сконструированной in vitro рекомбинантной плазмидной ДНК, содержащей кодирующую часть гена бета-интерферона человека (за исключением области, кодирующей сигнальный пептид), трансформируют штамм дрожжей р4-1-GRF18 и получают штамм р4-1-GRF18(pHBI) - продуцент фибробластного интерферона человека. Полученный штамм дрожжей Saccharomyces cerevisiae p4-1-GRF18(pHBI) депонирован во Всероссийской коллекции промышленных микроорганизмов под номером ВКПМ-Y-2286 и обеспечивает синтез фибробластного интерферона человека на более высоком уровне продукции. 3 с.п.ф-лы, 2 ил.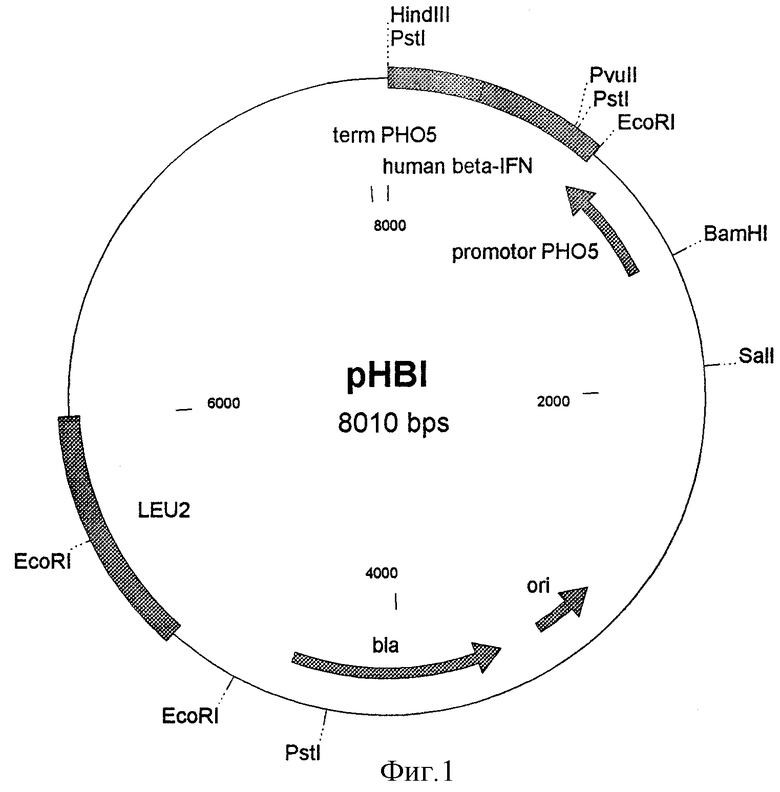
| SU 1830945 A1, 10.08.1999 | |||
| ИНДУКТОР ИНТЕРФЕРОНА РИДОСТИН | 1993 |
|
RU2083221C1 |
| US 5514567, 07.05.1996. | |||

