Изобретение относится к биотехнологии, генной инженерии, микробиологической промышленности, ветеринарии и сельскому хозяйству и представляет собой дрожжевой штамм - продуцент иммунного интерферона быка, содержащий сконструированную in vitro плазмиду, которая обеспечивает синтез внутриклеточного иммунного интерферона быка.
Иммунный интерферон, гамма-интерферон, относится к обширной группе эволюционно родственных белков, получивших название интерферонов. Образование гамма-интерферона происходит в Т-лимфоцитах и ЕК-клетках (естественных киллерах) под воздействием соответственно вирусов и опухолевых клеток. Гамма-интерферон активирует специфический клеточный иммунитет, стимулирует цитотоксичность макрофагов, повышает устойчивость организма к различным инфекциям. Показано, что бычий, свиной иммунные интерфероны повышают бактерицидную и фунгицидную активность макрофагов и являются эффективными препаратами для лечения различных заболеваний животных (Kishko I.H., Vasilenko M.I. Mikrobiol. Z., 1998, v.60, p.65-75; Zhang Z.D. et al., Arch. Virol., 2002, v.147, p.2157-2167). Использование интерферонов для лечения заболеваний различной этиологии имеет существенные преимущества по сравнению с традиционными антибиотиками и химиотерапевтическими препаратами за счет широкого спектра действия, обусловленного активацией иммунной системы, и из-за отсутствия побочных эффектов. По мнению экспертов Всемирной Организации Здравоохранения цитокиновая терапия особенно актуальна в связи с опасностью появления в продуктах питания патогенных микроорганизмов, устойчивых к антибиотикам (Lowenthal J.W. et al., Vet. Immun. Immunopathol, 1999, v.15, р.183-188). Кроме того, гамма-интерферон является противоопухолевым агентом, активирует антинеопластическую функцию макрофагов. Показано, что рекомбинантный гамма-интерферон быка подавляет in vitro репликацию вируса лейкоза крупного рогатого скота (Sentsui H. et al., Cytokine, 2001, v.16, p.227-231). Учитывая, что это заболевание является широко распространенным, необходимость подобного лекарственного средства для ветеринарии и сельского хозяйства не вызывает сомнения.
Однако систематические исследования спектра биологического действия гамма-интерферона, как и его применение в качестве лекарственного препарата затруднены, так как в настоящее время в России не существует препаратов гамма-интерферона быка.
Перспективным подходом для получения гамма-интерферона быка в значительных количествах является использование микроорганизмов в качестве продуцентов этого препарата. Существуют штаммы Escherichia coli, синтезирующие гамма-интерферон быка (Kashima Т. et al., J.Vet.Med.Sci. 1999, v.61, p.171-173). Но Escherichia coli является условно патогенным микроорганизмом, и препараты гамма-интерферона, полученные из бактериальных клеток, могут содержать эндотоксины.
Все вышеизложенное свидетельствует о перспективности создания штаммов-продуцентов гамма-интерферона быка на основе других микроорганизмов, в частности дрожжей. Применение непатогенных микроорганизмов (дрожжей), не содержащих токсических и пирогенных факторов, в качестве продуцентов белков животных позволяет использовать рекомбинантные белки в ветеринарной практике. Созданный ранее в лаборатории биохимической генетики БиНИИ СПбГУ штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-1223 - внутриклеточный продуцент иммунного интерферона быка синтезировал около 10 мг рекомбинантного интерферона на литр культуры (Мясников А.Н.. и др. Патент РФ, SU 1660388, 1991).
Использование для гетерологичной экспрессии метилотрофных дрожжей Pichia pastoris представляет особый интерес. Достоинствами этих дрожжей является накопление значительной биомассы при культивировании на недорогих минеральных питательных средах и более высокий уровень синтеза рекомбинантных белков (Sreekrishna К. et al., J.Basic.Microbiol, 1988, v.28, p.265-278). В настоящее время ген иммунного интерферона быка клонирован в составе векторов, обеспечивающих продукцию рекомбинантного белка клетками дрожжей Pichia pastoris и секрецию синтезированного белка в культуральную жидкость (Николаев Я.В. и др. Цитокины и воспаление, 2002, т.1, стр.28-29). Однако не менее перспективным представляется использование штаммов, накапливающих рекомбинантный белок внутри клетки в качестве пробиотической кормовой добавки.
Использование иммунопробиотической кормовой добавки имеет определенные преимущества по сравнению с лекарственными препаратами. Дрожжи являются ценным белково-витаминным продуктом и широко используются в животноводстве и птицеводстве в качестве кормовых добавок к основному рациону. Белки, синтезируемые дрожжами, усваиваются легче, чем белки животного происхождения, кроме того, повышают биологическую ценность других кормов. Фосфор и кальций, находящиеся в составе дрожжей, способствуют нормальному развитию костной ткани; витамины группы В, содержащиеся в дрожжевых клетках, являются регуляторами метаболизма жиров. Кроме того, было показано, что липополисахариды, входящие в состав клеточной стенки микроорганизмов, активизируют систему местного иммунитета и, взаимодействуя с факторами регуляции интерферонов, индуцируют продукцию собственных цитокинов (Navarro L., David M., J.Biol.Chem, 1999, v. 274, р.35535-35538; Coates N.J., McColl S.R., J.Immunol, 2001, v.166, p.5176-5182).
Известно, что штаммы Bacillus subtilis, синтезирующие α2-интерферон человека, при пероральном введении сохраняются в желудочно-кишечном тракте экспериментальных животных не менее 96 часов и обеспечивают синтез рекомбинантного белка (Бакулина Л.Ф. и др. Биотехнология, 2001, № 2, стр.48-56). Это позволяет с твердой уверенностью ожидать, что штамм дрожжей Pichia pastoris - продуцент гамма-интерферона быка в аналогичных условиях будет обеспечивать выработку рекомбинантного белка в организме животных, что будет сопровождаться специфическим повышением бактерицидной, фунгицидной и антинеопластической активности макрофагов животных. При этом особенности создания штаммов-продуцентов исключают возможность попадания в клетки дрожжей последовательности бактериальной ДНК. Использование такой иммунопробиотической кормовой добавки является также экономически выгодным, так как позволяет исключить трудоемкие процедуры по выделению и очистке рекомбинантного интерферона. Эксперименты по применению штаммов дрожжей Pichia pastoris, синтезирующих гормон роста птиц, в качестве кормовой добавки в птицеводстве оказались успешными (Chen C.M. et al., 2000, Life Sci., v.67, p.2103-2115).
Задачей, на решение которой направлены предлагаемые изобретения, является создание штамма дрожжей Pichia pastoris PS105(pBIG)-, синтезирующего внутриклеточный иммунный интерферон быка, получение рекомбинантной плазмидной ДНК pBIG, обеспечивающей биосинтез внутриклеточного иммунного интерферона быка и разработка способа конструирования рекомбинантной плазмидной ДНК pBIG.
Для решения поставленной задачи разработан способ конструирования рекомбинантной плазмидной ДНК pBIG, обеспечивающей биосинтез внутриклеточного иммунного интерферона быка, в результате создан штамм дрожжей Pichia pastoris PS105(pBIG) - продуцент внутриклеточного иммунного интерферона быка.
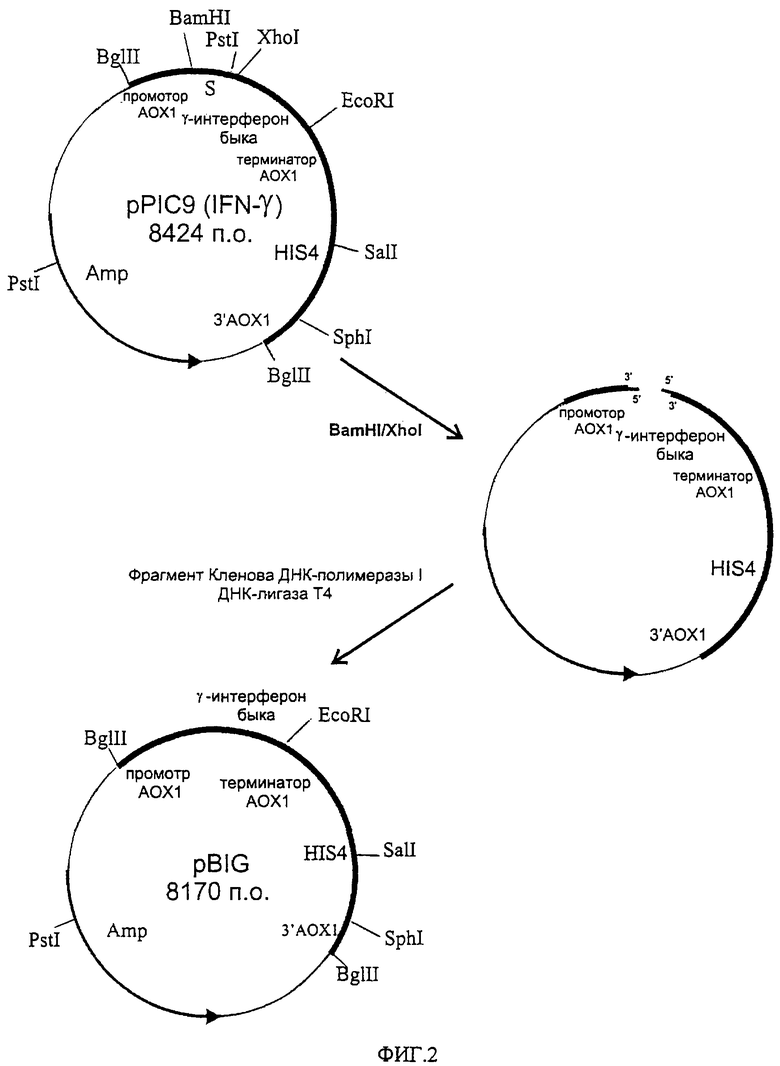
Рекомбинантная плазмидная ДНК pBIG, обеспечивающая биосинтез внутриклеточного иммунного интерферона быка трансформированными ею клетками дрожжей, состоит из следующих элементов:
- BamHI-EcoRI - фрагмент плазмидной ДНК бифункционального бактериально-дрожжевого вектора рРIС9 размером 7,75 т.п.о. (из которого удалена препрообласть гена MFα дрожжей Saccharomyces cerevisiae размером 0,25 т.п.о., обеспечивающая секрецию рекомбинантного белка в культуральную среду), включающий бактериальный ген устойчивости к ампициллину, бактериальную область инициации репликации, ген HIS4 дрожжей; фрагмент 5'-некодирующей области дрожжевого гена АОХ1 размером 0, 95 т.п.о., содержащий область, обеспечивающую активацию транскрипции этого гена в присутствии метанола в качестве источника углерода в культуральной среде; фрагмент гена АОХ1 размером 0,33 т.п.о., содержащий область терминации транскрипции этого гена; фрагмент 3'-нетранслируемой области гена АОХ1 размером 0,76 т.п.о.;
- XhoI-EcoRI - фрагмент размером 0,42 т.п.о., содержащий кодирующую часть гена гамма-интерферона быка за исключением области, кодирующей сигнальный пептид;
- уникальные сайты распознавания следующих рестриктаз: EcoRI - 1,39 т.п.о.; XbaI - 2,20 т.п.о.; SalI - 3,34 т.п.о.; SphI - 4,96 т.п.о; PstI - 7,35 т.п.о.
Схема плазмидной ДНК pBIG с рестрикционной картой изображена на фиг.1. Общий размер плазмиды pBIG - 8170 п.о. (5,39 MDa).
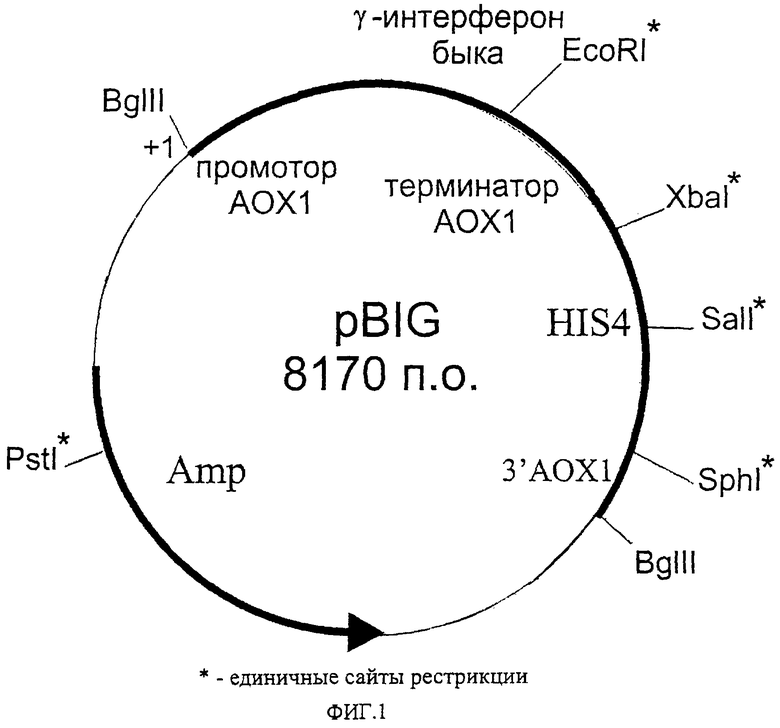
Способ конструирования плазмиды, обеспечивающей биосинтез внутриклеточного иммунного интерферона в клетках дрожжей проиллюстрирован схемой конструирования плазмиды pBIG, приведенной на фиг.2.
Из плазмиды pPIC9 (IFN-γ) (Николаев Я.В. и др. Цитокины и воспаление, 2002, т.1, стр.28-29) с помощью гидролиза рестриктазами BamHI и XhoI вырезают препрообласть гена MFα дрожжей Saccharomyces cerevisiae, обеспечивающую секрецию гамма-интерферона в культуральную среду, затем лигазной смесью трансформируют клетки Escherichia coli и отбирают клоны, содержащие рекомбинантную плазмидную ДНК pBIG.
Выбор конструкции плазмиды для продукции иммунного интерферона (гамма-интерферона) быка обусловлен следующими причинами. Плазмида pBIG получена на основе челночного бактериально-дрожжевого интегративного вектора рРIС9. В результате трансформации линеаризованной плазмидой и последующей гомологичной рекомбинации происходит встраивание экспрессионной кассеты в хромосому дрожжей Pichia pastoris, что обеспечивает стабильное поддержание клонированного гена гамма-интерферона. В состав плазмиды входит ген HIS4 дрожжей, что позволяет селективно отбирать трансформантов при использовании в качестве реципиентов штаммы дрожжей с мутациями в этом гене.
Экспрессия гена гамма-интерферона быка в составе плазмиды pBIG находится под контролем промотора гена АОХ1, содержащим области, обеспечивающие активацию транскрипции в присутствии метанола в культуральной среде, а также область инициации транскрипции. Промотор гена АОХ1 относится к числу наиболее сильных дрожжевых промоторов. Уровень экспрессии генов, находящихся под контролем АОХ1 промотора, эффективно регулируется источниками углерода. Транскрипция гена АОХ1 полностью блокирована при выращивании дрожжей на среде с глюкозой, на среде с глицерином наблюдается только базальный уровень экспрессии гена. Использование метанола в качестве единственного источника углерода значительно усиливает экспрессию гена АОХ1 и, следовательно, генов, находящихся под контролем АОХ1 промотора. Это позволяет регулировать синтез гамма-интерферона быка в клетках дрожжей. Регулируемая экспрессия клонированного гена позволяет существенно снизить метаболическую нагрузку на клетку дрожжей.
В качестве продуцента гамма-интерферона используют штамм PS105(pBIG). Штамм PS105(pBIG) получен при трансформации штамма дрожжей PS99 (his4 рер4::РНO85) плазмидной ДНК pBIG. Штамм PS99 несет мутацию в гене HIS4, что позволяет селективно отбирать трансформантов, несущих плазмидную ДНК pBIG. Мутация в гене РЕР4 приводит к отсутствию активности протеаз А и В, а также карбоксипептидазы Y в клетках дрожжей, что сопровождается повышением стабильности гетерологичных рекомбинантных белков (Hisch H.H. et al. In: Walton E.F., Yarranton G.T., Eds., Molecular and Cell Biology of Yeast, 1989, p.134-200).
Штамм дрожжей Pichia pastoris PS105(pBIG) характеризуется следующими признаками.
Морфологические признаки.
Клетки округлой, слегка овальной формы, размером 5-10 мкм, часть клеток имеет на поверхности почки или соединена с дочерними клетками.
Культуральные признаки.
Клетки хорошо растут на полной органической среде YEPD - 2% пептона, 1% дрожжевого экстракта, 2% глюкозы или 1% глицерина.
Кроме того, клетки хорошо растут на минеральной среде SC: 1,34% Yeast Nitrogen Base (“Difco”, США), 2% глюкозы (1% глицерина, 0,5% метанола), а также на других синтетических средах для дрожжей.
При росте на твердых средах клетки образуют гладкие, круглые колонии с матовой поверхностью, светло-кремового цвета, край неровный.
При росте в жидких средах образуют интенсивную ровную суспензию. Культура имеет характерный запах метилотрофных дрожжей.
Физиолого-биохимические признаки.
Клетки растут в пределах от 4 до 37°С. Оптимальной температурой выращивания является 30°С. При росте в аэробных условиях клетки незначительно защелачивают среду. Оптимум рН для роста составляет 4,5-6,5.
В качестве источника углерода клетки могут использовать многие простые соединения, такие как: глюкоза, глицерин, метанол.
В качестве источника азота клетки могут использовать минеральные соли в аммонийной форме, аминокислоты, мочевину.
Клетки способны к аэробному и анаэробному росту.
Существенным признаком штамма является отсутствие потребности в гистидине.
Способ конструирования плазмидной ДНК pBIG проиллюстрирован следующим примером.
ПРИМЕР.
Клетки бактерий Escherichia coli, содержащие плазмиду pPIC9(IFN-γ) выращивают при 37°C в течение ночи в 1 л питательной среды LB (1% пептона, 0,5% дрожжевого экстракта, 1% хлористого натрия), содержащей ампициллин в концентрации 50 мг/л. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут при 4°С, суспендируют в 20 мл 25 мМ трис-хлоридного буфера (рН 8,0), содержащего 10 мМ ЭДТА и 50 мМ глюкозы, добавляют 30 мг лизоцима и инкубируют 10 минут при комнатной температуре. Далее добавляют 40 мл 0,2 М гидроокиси натрия, содержащей 1% додецилсульфата натрия, осторожно перемешивают и инкубируют в течение 10 минут при 4°С. Раствор нейтрализуют добавлением 30 мл 3 М ацетата натрия (рН 5,0) и выдерживают в течение 10 минут при 4°C. После этого центрифугируют при 14000 об/мин в течение 40 минут при 4°C. К супернатанту добавляют 0,6 объема изопропилового спирта, выдерживают 20 минут при комнатной температуре и центрифугируют при 14000 об/мин в течение 20 минут при 20°C. Полученный осадок промывают 70% этиловым спиртом, высушивают в вакууме и растворяют в 4 мл дистиллированной воды. Далее добавляют 4,2 г хлористого цезия и 0,36 мл раствора бромистого этидия (10 мг/мл). Полученный раствор выдерживают в течение 1 часа при 4°С, затем центрифугируют при 15000 об/мин в течение 15 минут. Супернатант центрифугируют при 70000 об/мин в течение 16 часов в центрифуге TL100 (“Beckman”). После центрифугирования отбирают полосу плазмидной ДНК (нижнюю из двух флюоресцирующих в ультрафиолетовом свете полос), дважды экстрагируют бромистый этидий равным объемом изоамилового спирта, разбавляют в два раза дистиллированной водой и осаждают плазмидную ДНК двумя объемами этилового спирта и 1/15 объема 3 М ацетата натрия (рН 5,0). Осадок собирают центрифугированием при 10000 об/мин в течение 10 минут, промывают 70% этиловым спиртом и растворяют в 0,5-1 мл буфера ТЕ (10 мМ трис-хлоридный буфер, рН 8,0, содержащий 1 мМ ЭДТА). Концентрацию плазмидной ДНК определяют по поглощению раствора при длине волны 260 нм. Чистоту препарата контролируют при помощи электрофореза в 0,7% агарозном геле в буфере ТВЕ (0,1М трис-боратный буфер, рН 8,3, содержащий 1 мМ ЭДТА).
Гидролиз плазмиды pPIC9(IFN-γ) рестриктазами BamHI и XhoI проводят в 10 мМ трис-хлоридном буфере (рН 8,0), содержащем 100 мМ хлористого калия, 5мМ хлористого магния, 0,02% тритона Х-100 и 0,1 мг/мл бычьего сывороточного альбумина.
К 5 мкг плазмидной ДНК в объеме 20 мкл добавляют по 5 ед. каждой рестриктазы, после чего пробу инкубируют в течение 2 часов при 37°C.
Выделение линеаризованной плазмидной ДНК из агарозного геля проводят по методике, разработанной фирмой QIAGEN. Полоску геля с фрагментом ДНК помещают в пробирку и добавляют раствор QX1 (300 мкл на 100 мг геля). Пробу нагревают до 50°C, добавляют реактив QIAEX (10 мкл на 5 мкг ДНК) и инкубируют при 50°C в течение 10 минут, периодически перемешивая. Далее центрифугируют 30 секунд при 15000 об/мин, супернатант отбрасывают, осадок дважды экстрагируют растворами QX2 и QX3, удаляют супернатант центрифугированием при 15000 об/мин в течение 30 секунд. Осадок высушивают на воздухе, растворяют в 20 мкл буфера ТЕ, центрифугируют 30 секунд при 15000 об/мин, супернатант переносят в новую пробирку.
Выделенную линейную ДНК обрабатывают фрагментом Кленова ДНК полимеразы I, обеспечивающим достраивание укороченных 3'-концов. Реакцию проводят в 50 мМ трис-хлоридном буфере, содержащем 10 мМ хлористого магния, 1 мМ дитиотреэтола, 50 мкг/мл бычьего сывороточного альбумина, по 1ммоль дезоксирибонуклеотидов и 1 ед. фермента на 1 мкг ДНК. Реакционную смесь инкубируют 15 мин при 37°С, затем инактивируют фермент нагреванием при 65°С в течение 10 мин.
Для получения плазмиды pBIG проводят лигирование 1 мкг плазмиды pPIC9(IFN-γ), гидролизованной рестриктазами BamHI и XhoI и обработанной фрагментом Кленова ДНК полимеразы I в 10 мкл 70 мМ трис-хлоридного буфера (рН 7,6), содержащего 5 мМ дитиотреэтола, 5 мМ хлористого магния, 1 мМ АТФ, добавляя 10 ед. ДНК-лигазы фага Т4 и инкубируя при 14°С в течение ночи.
Полученной лигазной смесью трансформируют клетки штамма DH5α Escherichia coli (F'/endA1 hsdR17 (r
Из выросших отдельных клонов трансформантов выделяют плазмидную ДНК при помощи методики, использованной для получения плазмиды pPIC9(IFN-γ), за исключением того, что клетки Escherichia coli выращивают в 10 мл LB и соответственно объемы всех растворов уменьшают в 100 раз. Кроме того, вместо стадии центрифугирования в градиенте плотности хлористого цезия проводят обработку ДНК панкреатической РНКазой. Для этого нуклеиновые кислоты, осажденные изопропиловым спиртом, растворяют в 100 мкл буфера ТЕ, добавляют 10 мкл раствора РНКазы (1 мг/мл) и инкубируют 30 минут при 37°С.
Далее проводят гидролиз полученной плазмидной ДНК рестриктазами PstI и SalI в 50 мМ трис-хлоридном буфере, содержащем 10 мМ хлористого магния, 100 мМ хлористого натрия и 0,1 мг/мл бычьего сывороточного альбумина. При рестрикции искомой плазмиды pBIG и последующем электрофорезе в 0,7% агарозном геле обнаруживаются фрагменты 4,17 и 4,0 т.п.о. Из выявленного таким образом клона препаративно выделяют плазмиду pBIG также, как описано для плазмиды pPIC9(IFN-γ), и гидролизуют рестриктазой BglII в 50 мМ трис-хлоридном буфере (рН 7,5), содержащем 100 мМ хлористого натрия, 10 мМ хлористого магния. BglII-фрагмент плазмиды pBIG, размером 5,77 т.п.о, выделяют по описанной выше методике фирмы QIAGEN, исключая стадию нагревания, и используют его для трансформации клеток дрожжей, как описано в примере 2.
ПРИМЕР 2.
Для получения штамма дрожжей Pichia pastoris - продуцента гамма-интерферона быка клетки дрожжей штамма PS99 трансформируют плазмидной ДНК pBIG.
Клетки дрожжей выращивают в 100 мл среды YEPD при 30°C до достижения культурой оптической плотности, соответствующей 2-4 ед. поглощения при длине волны 600 нм. Клетки дважды промывают стерильной водой, после чего суспендируют в 0,3 мл 100 мМ раствора ацетата лития и инкубируют при 30°C в течение 30 минут. К 50 мкл полученной суспензии клеток добавляют 0,1-1 мкг плазмидной ДНК, 50 мкг ДНК спермы лосося, предварительно денатурированной нагреванием (10 минут при 100°C) и 0,3 мл раствора 100 мМ ацетата лития, содержащего 40% полиэтиленгликоля 4000. Далее пробу инкубируют 30 минут при 30°C и 20 минут при 42°C, помещают на 15 секунд в ледяную баню и центрифугируют 10 секунд при 10000 об/мин. Клетки суспендируют в 1 мл стерильной воды и высевают на твердую среду SC. Клоны трансформантов вырастают через 2-3 суток. Выросшие клоны пересевают на чашки со средой SC, содержащей 2% глюкозу, отдельными колониями, затем перепечатывают на среду ММ (1,34% Yeast Nitrogen Base (“Difco”, США), 0,5% метанола, 2% агара (“Difco”, США) для отбора трансформантов, характеризующихся снижением скорости роста на среде с метанолом, что свидетельствует об интеграции чужеродного гена в локус AOXI (фенотип MetS).
Для анализа продукции гамма-интерферона быка клетками трансформантов фенотипа MetS их выращивают при 30°C в 100 мл жидкой среды MGY (1,34% Yeast Nitrogen Base (“Difco”, США), 1% глицерина, 0,04 мг биотина до стационарной фазы роста в течение 2 суток. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут, супернатант сливают и переносят всю биомассу в 100 мл жидкой среды ММ (1,34% Yeast Nitrogen Base (“Difco”, США), 0,5% метанола, 0,04 мг биотина) для индукции экспрессии гамма-интерферона. Индукцию проводят при 30°C в течение 4 суток. По окончании индукции клетки собирают центрифугированием при 5000 об/мин в течение 10 минут, ресуспендируют в буфере ТЕ, добавляют стеклянные шарики (⊘=0,5 мм) и разрушают на дезинтеграторе (“Braun Metsunger”). Полученную суспензию отделяют от шариков и центрифугируют 10000 об/мин 20 мин. В супернатанте определяют содержание гамма-интерферона при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия и последующей гибридизации со специфическими антителами к гамма-интерферону. Высокая степень гомологии гамма-интерферонов человека и быка позволяет в этом тесте заменить отсутствующие в продаже моноклональные антитела к гамма-интерферону быка антителами к аналогичному белку человека (Мясников А.Н. и др. Патент РФ, SU 1660388 А1, 1991). Разделение белков проводят в 15% полиакриламидном геле в стандартной системе буферов (электродный буфер: 25 мМ трис, 192 мМ глицин, 0,1% додецилсульфат натрия, рН 8,3; буфер для геля: 375 мМ трис-хлоридный буфер, рН 8,8). Параллельно проводят разделение белков контрольного штамма, выращенного в идентичных условиях. В качестве стандартов молекулярной массы используют карбоангидразу, ингибитор трипсина, миоглобин, лизоцим. По окончании электрофореза белки ренатурируют, выдерживая гели 15 минут в 10 мМ трис-хлоридном буфере (рН 7,5), содержащем 4М мочевину, 20 мМ ЭДТА, и переносят на нитроцеллюлозную мембрану в 25 мМ трис-192 мМ глициновом буфере (рН 8,3), содержащем 20% метилового спирта, при 30-40 В, в течение 1,5 часов. Далее мембрану выдерживают в буфере TBST (10 мМ трис-хлоридный буфер (рН 8,0), 150 мМ хлористого натрия, 0,05% твин-20, 1% бычьего сывороточного альбумина) в течение 2 часов при 37°C. Затем помещают мембрану в тот же буфер, содержащий разведенные в 1000 раз мышиные моноклональные антитела к гамма-интерферону человека (“Протеиновый контур”, Санкт-Петербург), и инкубируют 2 часа при 37°C. Далее трижды промывают мембрану буфером TBST и инкубируют 1 час при 37°C c разбавленным в 7000 раз конъюгатом видоспецифических антител к иммуноглобулинам мыши и пероксидазы хрена (“Протеиновый контур”, Санкт-Петербург). После отмывки мембраны буфером PBST (58 мМ двузамещенного фосфата натрия, 17 мМ однозамещенного фосфата натрия, 68 мМ хлористого натрия, 0,1% твин-20) добавляют раствор субстратов для пероксидазы: 0,02% DAB (3,'3-диаминобензидин тетрагидрохлорид), 0,006% перекись водорода в 10 мМ трис-хлоридном буфере, рН 7,5. Параллельно окрашивают гели 0,15% раствором кумасси G250 в 25% изопропаноле и 10% уксусной кислоте и отмывают в 10% уксусной кислоте. Уровень синтеза гамма-интерферона быка определяют, сравнивая интенсивность окрашивания полосы рекомбинантного белка с полосой маркерного белка соответствующей молекулярной массы.
Согласно полученным данным клетки дрожжей штамма PS104(pBIG) синтезируют не менее 15-20 мг гамма-интерферона на литр культуры дрожжей, что превышает продуктивность штамма-прототипа ВКПМ Y1223.
Суммируя вышесказанное, можно заключить, что полученный штамм дрожжей Pichia pastoris PS105(pBIG) синтезирует гамма-интерферон быка и может использоваться в ветеринарии и сельском хозяйстве в качестве иммунопробиотической кормовой добавки, повышающей устойчивость животных к различным инфекционным и онкологическим заболеваниям. Уровень синтеза рекомбинантного белка позволяет использовать данный штамм также для выделения гамма-интерферон быка.
Штамм дрожжей Pichia pastoris PS105(pBIG) - продуцент внутриклеточного иммунного интерферона быка депонирован во Всероссийской коллекции промышленных микроорганизмов под номером ВКПМ-У2990.
Изобретение относится к области биотехнологии, генной инженерии, ветеринарии и может быть использовано в микробиологической промышленности и сельском хозяйстве. Путем вырезания рестриктазами BamHI и XhoI из бактериально-дрожжевого вектора pPIC9 препрообласти гена MFα дрожжей Saccharomyces cerevisiae сконструирована плазмидная ДНК pBIG, обеспечивающая биосинтез внутриклеточного гамма-интерферона быка. Получен штамм дрожжей Pichia pastoris PS105(pBIG) - продуцент внутриклеточного иммунного интерферона быка, который депонирован во Всероссийской коллекции промышленных микроорганизмов под номером ВКПМ-У2990.Использование изобретения позволяет получать внутриклеточный гамма-интерферон быка с повышенным выходом. 3 c.п. ф-лы, 2 ил.
| ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS - ПРОДУЦЕНТ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PHIF И СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ | 2000 |
|
RU2180687C1 |
| RU 1660388 С, 20.05.1995 | |||
| УСТАНОВКА СТРЕЛКОВОГО ОРУЖИЯ | 2004 |
|
RU2242694C1 |

