Изобретение относится к ДНК, кодирующей мутантную альфа-изопропилмалат синтазу, а также к способу получения штамма Escherichia coli - продуцента L-лейцина, содержащего мутантную альфа-изопропилмалат синтазу, и способу получения L-лейцина с использованием указанного штамма. L-лейцин является незаменимой аминокислотой, которая может использоваться в виде питательной добавки в пищу человека или корм животных, применяться в качестве реагента или вещества в медицине, фармацевтической или химической промышленности или в качестве ростового фактора при получении других аминокислот, таких как лизин.
В прошлом L-лейцин получали методом ферментации, первоначально используя микроорганизмы, принадлежащие к роду Brevibacterium, Corynebacterium или Serratia, либо их мутанты, продуцирующие L-лейцин (Amino acid fermentation, JAPAN SCIENTIFIC SOCIETY'S PRESS, pp.397-422, 1986).
Наиболее высокий уровень накопления L-лейцина был достигнут при использовании штамма VKPM В-2736 Brevibacterium flavium. На содержащих сахарозу средах за 72 часа ферментации в лабораторном ферментере данный штамм продуцирует L-лейцин с концентрацией до 26 грамм/литр (смотри Авторское свидетельство СССР 1394711). Штамм Brevibacterium lactofermentum 34 продуцирует до 34 г/л L-лейцина на среде, содержащей глюкозу (смотри Appl. Environ. Microbiol., 51, стр. 1024 (1986)).
Как указывалось выше, эффективность получения L-лейцина была несколько повышена, однако для обеспечения в будущем растущих потребностей в L-лейцине существует необходимость разработать более эффективный и экономичный способ получения L-лейцина.
С другой стороны, как показывают данные генетического анализа и результаты исследования большого числа генетического материала, в качестве эффективных продуцентов L-лейцина потенциально могут использоваться принадлежащие к роду Escherichia микроорганизмы, отличающиеся высокой скоростью роста. Тем не менее, имеется мало свидетельств, в которых описывается получение L-лейцина с использованием бактерий, принадлежащих к роду Escherichia.
В качестве штаммов бактерии рода Escherichia, продуцирующих L-лейцин, известен штамм, обладающий устойчивостью к бета-тиенилаланину и бета-гидроксилейцину (смотри публикацию японского патента 62-34397 (1987), и штамм, обладающий устойчивостью к 4-азалейцину или 5,5,5-трифторлейцину (смотри выложенную японскую патентную заявку 8-70879 (1996).
Тем не менее, до настоящего времени не известно ни бактерий рода Escherichia, обладающих устойчивостью к L-лейцину, ни какой-либо зависимости между устойчивостью к L-лейцину и способностью продуцировать L-лейцин.
КРАТКОЕ ИЗЛОЖЕНИЕ ИЗОБРЕТЕНИЯ
Созданное с учетом вышеизложенного настоящее изобретение имеет целью повысить эффективность получения L-лейцина с использованием штамма бактерии, принадлежащей к роду Escherichia, а также обеспечить эффективный и экономичный способ получения L-лейцина.
В результате тщательных исследований, направленных на достижение указанной выше цели, авторы настоящего изобретения установили, что утрата чувствительности альфа-изопропилмалат синтазы (далее IPMS) к ингибированию ее активности L-лейцином по типу обратной связи обеспечивает более высокую эффективность получения L-лейцина, и, таким образом, совершили настоящее изобретение.
Более точно в настоящем изобретении предложены:
1. Мутантная альфа-изопропилмалат синтаза (IPMS) (также именуемая далее как "IPMS согласно настоящему изобретению") с аминокислотной последовательностью, приведенной в перечне последовательностей под 2, тип замещения у которой выбран из группы, включающей:
(a) замещение остатка треонина в положении 482,
(b) замещение остатка глутаминовой кислоты в положении 386,
(c) замещение остатка пролина в положении 428,
(d) замещение остатка глицина в положении 479,
(e) замещение остатка глицина в положении 462;
у которой чувствительность к ингибированию L-лейцина по типу обратной связи утрачена.
2. Мутантная альфа-изопропилмалат синтаза по п. 1, отличающаяся тем, что упомянутое замещение выбрано из группы, включающей:
(a) замещение остатка треонина в положении 482 остатком изолейцина,
(b) замещение остатка глутаминовой кислоты в положении 386 остатком лизина,
(c) замещение остатка пролина в положении 428 остатком пролина,
(d) замещение остатка цистеина в положении 479 остатком глицина,
(e) замещение остатка глицина в положении 462 остатком аспарагиновой кислоты.
3. Мутантная альфа-изопропилмалат синтаза по п.п. 1, отличающаяся тем, что в ней произведена делеция, замещение, инсерция или добавление одного или нескольких аминокислотных остатков, при этом активность альфа-изопропилмалат синтазы и утрата чувствительности IPMS к ингибированию по типу обратной связи в указанном белке такие же, как в IPMS по п.1.
Также в настоящем изобретении предложена ДНК, кодирующая мутантную IPMS. Предпочтительной ДНК (далее также именуемой - "ДНК согласно настоящему изобретению") является ДНК, кодирующая мутантную IPMS, в которой тип замещения выбран из группы, включающей:
(а') замещение остатка треонина в положении 482 остатком изолейцина, (b') замещение остатка глутаминовой кислоты в положении 386 остатком лизина,
(с') замещение остатка пролина в положении 428 остатком лейцина,
(d') замещение остатка глицина в положении 479 остатком цистеина, и
(е') замещение остатка глицина в положении 462 остатком аспарагиновой кислоты.
В число конкретных примеров ДНК согласно настоящему изобретению входит ДНК с нуклеотидной последовательностью, приведенной в перечне последовательностей под 1, при этом такая последовательность имеет мутацию, выбранную из группы, включающей:
(i) мутацию цитозина в положении 1445 на тимин,
(ii) мутацию гуанина в положении 1156 на аденин,
(iii) мутацию цитозина в положении 1283 на тимин,
(iv) мутацию гуанина в положении 1435 на тимин и
(v) мутацию гуанина в положении 1385 на аденин.
В настоящем изобретении также предложен способ получения штамма бактерии - продуцента L-лейцина, в котором указанный штамм получают путем трансформации штамма бактерии вышеописанной ДНК.
Предпочтительно микроорганизм согласно настоящему изобретению относится к роду Escherichia. Более предпочтительно микроорганизм согласно настоящему изобретению представляет собой Escherichia coli.
В настоящем изобретении также предложен способ получения L-лейцина путем культивирования штамма бактерии - продуцента L-лейцина в питательной среде с последующим выделением L-лейцина из культуральной жидкости, в котором используют штамм, полученный вышеуказанным способом.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет описано подробно.
<1> IPMS согласно настоящему изобретению.
Альфа-изопропилмалат синтаза (IPMS) является ферментом, участвующим в первой реакции синтеза L-лейцина и осуществляющим конденсацию ацетильной группы ацетил-кофермента А с альфа-кетоизовалератом с образованием альфа-изопропилмалата. IPMS согласно настоящему изобретению является мутантная альфа-изопропилмалат синтаза, у которой чувствительность к ингибированию L-лейцина по типу обратной связи утрачена. Более конкретно, IPMS согласно настоящему изобретению является мутантная альфа-изопропилмалат синтаза с аминокислотной последовательностью, приведенной в перечне последовательностей под 2, тип замещения у которой выбран из группы, включающей:
(a) замещение остатка треонина в положении 482,
(b) замещение остатка глутаминовой кислоты в положении 386,
(c) замещение остатка пролина в положении 428,
(d) замещение остатка глицина в положении 479,
(e) замещение остатка глицина в положении 462;
у которой чувствительность к ингибированию L-лейцина по типу обратной связи утрачена.
Более конкретно, IPMS согласно настоящему изобретению является мутантная альфа-изопропилмалат синтаза, у которой упомянутое замещение выбрано из группы, включающей:
(a) замещение остатка треонина в положении 482 остатком изолейцина,
(b) замещение остатка глутаминовой кислоты в положении 386 остатком лизина,
(c) замещение остатка пролина в положении 428 остатком пролина,
(d) замещение остатка цистеина в положении 479 остатком глицина,
(e) замещение остатка глицина в положении 462 остатком аспарагиновой кислоты.
Также IPMS согласно настоящему изобретению является мутантная альфа-изопропилмалат синтаза, у которой произведена делеция, замещение, инсерция или добавление одного или нескольких аминокислотных остатков, при этом активность альфа-изопропилмалат синтазы и утрата чувствительности IPMS к ингибированию L-лейцином по типу обратной связи в указанном белке такие же, как в вышеописанной IPMS. К подобным вариантам (мутантам) IPMS также относятся варианты (мутанты), которые могут существовать в естественных условиях в виду природного разнообразия и видовых различий.
<2> ДНК согласно настоящему изобретению
ДНК, кодирующая IPMS дикого типа, согласно настоящему изобретению имеет мутацию, благодаря которой происходит утрата чувствительности IPMS, кодируемой этой ДНК, к ингибированию L-лейцином по типу обратной связи.
Выражение "утрата чувствительности к ингибированию лейцином по типу обратной связи" означает, что подавление по типу обратной связи снижается. Степень снижения подавления по типу обратной связи можно определить путем измерения степени снижения подавления активности IPMS под воздействием L-лейцина и ее сравнения с таковой у дикого штамма или родительского штамма.
Примерами IPMS служат ферменты, полученные из бактерий, принадлежащих к роду Escherichia, в особенности IPMS, полученная из Е. coli. Примерами мутаций, вызывающих утрату чувствительности IPMS к ингибированию L-лейцином по типу обратной связи, служат следующие типы замещений (а) - (е) в аминокислотной последовательности, приведенной в перечне последовательностей под 2:
(a) замещение остатка треонина в положении 482,
(b) замещение остатка глутаминовой кислоты в положении 386,
(c) замещение остатка пролина в положении 428,
(d) замещение остатка глицина в положении 479 и
(e) замещение остатка глицина в положении 462.
Предпочтительно замещения выбирают из группы, включающей:
(а') замещение остатка треонина в положении 482 остатком изолейцина,
(b') замещение остатка глутаминовой кислоты в положении 386 остатком лизина,
(с') замещение остатка пролина в положении 428 остатком лейцина,
(d') замещение остатка глицина в положении 479 остатком цистеина, и
(е') замещение остатка глицина в положении 462 остатком аспарагиновой кислоты.
Примером ДНК, кодирующей IPMS дикого типа, служит ДНК, кодирующая IPMS, полученную из бактерий, принадлежащих к роду Escherichia. В особенности таким примером служит ДНК, кодирующая аминокислотную последовательность, приведенную в перечне последовательностей под 2, а также ДНК с нуклеотидной последовательностью, приведенной в перечне последовательностей под 1. В число ДНК согласно настоящему изобретению включены те из последовательностей, у которых имеется мутация в нуклеотидной последовательности, вызывающая описанное выше замещение аминокислотных остатков. Может быть использован любой кодон, соответствующий замещенному аминокислотному остатку, независимо от его вида при условии, что он кодирует идентичный аминокислотный остаток.
В число конкретных примеров ДНК согласно настоящему изобретению входят ДНК с нуклеотидной последовательностью, приведенной в перечне последовательностей под 1, при этом у такой последовательности имеется мутация, выбранная из группы, включающей:
(i) мутацию цитозина в положении 1445 на тимин,
(ii) мутацию гуанина в положении 1156 на аденин,
(iii) мутацию цитозина в положении 1283 на тимин,
(iv) мутацию гуанина в положении 1435 на тимин и
(v) мутацию гуанина в положении 1385 на аденин.
Кроме того, IPMS в различных бактериях и бактериальных штаммах могут иметь различия в последовательностях, однако в число ДНК согласно настоящему изобретению также включены ДНК, кодирующие такие виды IPMS, у которых имеется замещение, делеция или инсерция аминокислотного остатка/остатков в положении/положениях, не зависящих от активности ферментов. Иными словами, в число ДНК согласно настоящему изобретению также включена ДНК, кодирующая белок, имеющий аминокислотную последовательность мутантной IPMS, при этом у такой последовательности имеется делеция, замещение, инсерция или добавление одного или нескольких аминокислотных остатков, активность IPMS сохранена, но подавление активности L- лейцином по типу обратной связи в упомянутом белке утрачено таким же образом, как в мутантной IPMS. В их число входят ДНК, мутации у которых могут происходить естественным путем, в частности мутации, основанные на различиях между отдельными микроорганизмами, их видами и родами, содержащими IPMS (мутанты или варианты).
Способ получения ДНК, кодирующей мутантные IPMS, заключается в следующем.
(1) Получение гена IPMS дикого типа
Предпочтительным примером микроорганизма, донора такой ДНК, содержащего ген IPMS дикого типа или ген IPMS с иной мутацией описанного выше типа, служит микроорганизм, принадлежащий к роду Escherichia. В особенности, могут использоваться микроорганизмы, описанные в книге, автором которой является Neidhardt и др. (смотри Neidhardt, F.С. et al., Escherichia coli and Salmonella Typhimurium, American Society for Microbiology, Washington D.C., 1208, таблица 1). В частности, примерами служат штаммы Е. coli K-12, JM109 и МС1061. В случае когда в качестве донорского микроорганизма для ДНК используется штамм дикого типа, содержащий ген IPMS, может быть получена ДНК, содержащая ген IPMS дикого типа.
Ниже будет описан пример получения ДНК, содержащей ген IPMS. Во-первых, выращивают Е. coli, содержащую ген IPMS дикого типа, например, штамм К-12, с целью получения культуры. Процесс выращивания описанного выше микроорганизма может осуществляться обычным методом твердофазного культивирования, однако предпочтительно выращивание осуществляют методом жидкофазного культивирования с оценкой эффективности в процессе сбора бактерий. Для этого может использоваться среда, в которую добавлен один и несколько источников азота, таких как дрожжевой экстракт, пептон, мясной экстракт, кукурузный настой и вытяжка из соевых бобов или зерен пшеницы, а также одна или несколько неорганических солей, таких как дигидрофосфат калия, гидрофосфат калия, сульфат магния, хлорид натрия, хлорид магния, хлорид железа (II), сульфат железа (II) или хлорид марганца и дополнительно в соответствующем количестве добавлены сахара, витамины и аналогичные вещества. Предпочтительно, чтобы исходный уровень рН среды находился в пределах от 6 до 8. Культивирование осуществляют в течение от 4 до 24 часов при температуре от 30 до 42oС, предпочтительно около 37oС, методом глубинного культивирования с аэрацией и перемешиванием, методом качалочного культивирования, стационарного культивирования или аналогичным.
Полученную таким образом культуру в течение 5 минут подвергают центрифугированию, например, со скоростью 3000 оборотов в минуту с целью получения дебриса штамма К-12 Е. coli. Из дебриса может быть получена хромосомная ДНК, например, по методу Saito и Miura (смотри Biochem. Biophys. Acta., 72, 619 (1963) или по методу К. S. Kirby (смотри Biochem. J., 64, 405(1956)).
С целью отделить ген IPMS от полученной таким способом хромосомной ДНК создают библиотеку хромосомной ДНК. Во-первых, с целью получения смеси различных фрагментов хромосомную ДНК подвергают частичному расщеплению с помощью соответствующего фермента рестрикции. Могут использоваться самые разнообразные виды ферментов рестрикции при условии, что процесс расщепления регулируется временем реакции вырезания и тому подобным образом. Например, для обеспечения расщепления хромосомной ДНК ее инкубируют различные периоды времени (от 1 минуты до 2 часов) в присутствии Sau3AI с концентрацией фермента от 1 до 10 единиц/мл при температуре не ниже 30oС, предпочтительно 37oС.
Затем с целью получения рекомбинантной ДНК полученные фрагменты ДНК сшивают с векторной ДНК, способной к автономной репликации в клетках бактерий, принадлежащих к роду Escherichia. В частности, с целью полного переваривания, вырезания и расщепления векторной ДНК ее при температуре не ниже 30 oС в течение не менее 1 часа, предпочтительно от 1 до 3 часов, подвергают воздействию фермента рестрикции с концентрацией от 1 до 100 единиц/мл, например BamHI, продуцирующего конечную нуклеотидную последовательность, являющуюся комплементарной по отношению к последовательности, продуцированной ферментом рестрикции Sau3AI, использованным для вырезания хромосомной ДНК. Далее с целью получения рекомбинантной ДНК смесь фрагментов хромосомной ДНК, полученную описанным выше способом, смешивают с расщепленной и разрезанной векторной ДНК, которую при температуре от 4 до 16oС в течение не менее 1 часа, предпочтительно от 4 до 24 часов, подвергают воздействию ДНК-лигазы, предпочтительно ДНК-лигазы Т4 с концентрацией фермента от 1 до 100 единиц/мл.
Полученную рекомбинантную ДНК используют для трансформации микроорганизма, принадлежащего к роду Escherichia, например мутантного штамма, дефектного по IPMS, такого как штамм Escherichia coli, К-12, предпочтительно штамм JE7627 (ponB704, dacB12, pfv +, tonA2, dapA, lysA, str, malA38, metBl, ilvH611, leuA371, ргоА3, lac-3, tsx-76) с целью получения библиотеки хромосомной ДНК. Трансформация может быть осуществлена, например, по методу D. М. Morrison (смотри Methods in Enzymology 68, 326 (1979)) или по методу, в котором с целью повышения проницаемости ДНК клетки-реципиенты бактерий подвергают воздействию хлорида кальция (смотри Mandel, М. и Higa, A., J. Mol. Biol. , 53, 159 (1970)). Штамм JE7627 имеется в наличии в Национальном институте генетики (Mishima-shi, Shizuoka-ken, Japan).
Из полученной библиотеки хромосомной ДНК получают штамм бактерии, содержащий рекомбинантную ДНК гена IPMS, который выбирают из штаммов с повышенной активностью IPMS или штаммов, в которых ауксотрофность, являющаяся результатом дефекта в гене IPMS, комплементирована. Например, для мутантного штамма с дефектом в IPMS требуется L-лейцин. Таким образом, когда в качестве хозяина используется мутантный штамм, дефектный по IPMS, фрагмент ДНК, содержащий ген IPMS, может быть получен путем выделения штамма бактерии, становящегося способным к росту, на среде, не содержащей L-лейцина, и извлечения рекомбинатной ДНК из штамма бактерии.
Получить подтверждение того, содержится ли фактически рекомбинантная ДНК, в которой ген IPMS клонирован, в штамме-кандидате, содержащем рекомбинантную ДНК, которая содержит ген IPMS, можно путем приготовления клеточного экстракта из штамма-кандидата и получения из него неочищенного ферментного раствора с целью удостовериться, увеличилась ли активность IPMS. Процедура измерения активности ферментов IPMS может быть осуществлена по методу Kohlaw и др. (смотри J. Biol. Chem., 244, 2218 (1969)).
Рекомбинантная ДНК, в которой осуществлена инсерция ДНК, содержащей ген IPMS, в векторную ДНК, может быть выделена из описанного выше штамма бактерии, например, по методу Р. Guerry и др. (смотри J. Bacteriol., 116, 1064 (1973)) или по методу D.В. Clewell (смотри J. Bacteriol., 110, 667 (1972)).
Получение гена IPMS дикого типа может быть также осуществлено путем получения хромосомной ДНК из штамма, содержащего ген IPMS, по методу Saito и Miura или аналогичному методу, а амплификация гена IPMS - методом полимеразной цепной реакции (ПЦР) (смотри White, Т.J. и др.; Trends Genet., 5, 185 (1989)). Затравки ДНК для реакции амплификации должны быть комплементарны обеим 3'-концевым областям двухцепочечной молекулы ДНК, содержащей область гена IPMS или часть его. В случае, когда амплификации подвергается лишь область, занятая частью гена IPMS, в качестве затравок необходимо использовать фрагменты ДНК, дающие возможность выделить из библиотеки хромосомных ДНК фрагмент ДНК, содержащий ген IPMS целиком. В случае, когда амплификации подвергается область, содержащая весь ген IPMS целиком, продукты ПЦР, включающие фрагменты ДНК, содержащие амплифицированный ген IPMS, подвергают электрофорезу в геле агарозы, из которого затем извлекают нужный фрагмент ДНК. Таким образом, может быть извлечен фрагмент ДНК, содержащий ген IPMS.
Соответствующие затравки ДНК могут быть получены на основе, например, последовательности, известной для Е. coli (регистрационный номер EMBL D10483 или АЕ000117). В частности, предпочтительными являются затравки, способные амплифицировать область, содержащую 1572 нуклеотида, кодирующих ген IPMS. Синтез затравок может быть осуществлен обычным методом, таким как фосфорамидитный метод (смотри Tetrahedron Letters, 22, 1859 (1981)) с использованием промышленно производимого синтезатора ДНК (например, синтезатора ДНК модели 380В производства Applied Biosystems). Кроме того, ПЦР может быть осуществлена с использованием промышленно производимой установки ПЦР (например, термоблока для проведения реакций модели PJ2000 производства Takara Shuzo Co., Ltd.) в соответствии с методикой, указанной поставщиком.
Что касается гена IPMS, амплифицированного методом ПЦР, в случае когда он лигирован с векторной ДНК, способной к автономной репликации в клетках бактерий, принадлежащих к роду Escherichia, и введен в клетки бактерий, принадлежащих к роду Escherichia, облегчается осуществление таких операций, как введение мутации в ген IPMS. Используемая векторная ДНК, способ трансформации и способ подтверждения наличия гена IPMS аналогичны способам, применяемым в описанных выше операциях.
Данные о выделении гена IPMS смотри у Hertberg, K.M. и др., Gene, 8, стр. 135-152 (1980), Davis, M.G. и др., J. Bacteriol., 129, стр. 1078-1090 (1977) и т.д.
Как упоминалось выше, способ получения гена IPMS может быть использован для получения мутантных генов в том случае, когда микроорганизм, содержащий IPMS дикого типа, подвергают мутагенезу с целью получения мутантного штамма, продуцирующего мутантную IPMS, а мутантный ген получают из мутантного штамма.
(2) Введение мутации в ген IPMS
Примером осуществления мутации, такой как замещение, инсерция или делеция аминокислотных остатков служит метод рекомбинантной ПЦР (смотри Higuchi, R., стр. 61, "PCR Technology" (Издательство Erlich, H.A. Eds., Stockton press (1989)), а также метод сайт-направленного мутагенеза (смотри Kramer, W. и Frits, H.J., Meth. in Enzymol, 154, 350 (1987); Kunkel Т.А. и др., Meth. in Enzymol, 154, 367 (1987)). С помощью данных мтодов в нужном месте может быть вызвана целевая мутация.
Кроме того, при химическом синтезе целевого гена в соответствующий ген возможно ввести определенную мутацию или произвольную мутацию.
Помимо этого предлагается метод, в процессе осуществления которого ген IPMS на хромосоме или плазмиде непосредственно обрабатывается гидроксиламином (смотри Hashimoto, Т. и Sekiguchi, М. , J. Bacteriol., 159, 1039 (1984)). В качестве альтернативы возможно использовать метод, согласно которому содержащую ген IPMS бактерию, принадлежащую к роду Escherichia, подвергают облучению ультрафиолетом, или метод, основанный на обработке с помощью химического агента, такого как N-метил-N'-нитрозогуанидин или азотистая кислота. С помощью данных методов может быть вызвана произвольная мутация.
Что касается метода отбора мутантного гена, сначала рекомбинантную ДНК, включающую фрагмент ДНК, содержащий ген IPMS и векторную ДНК, непосредственно подвергают мутационной обработке гидроксиламином или другим веществом. Затем ею трансформируют, например, штамм Е. coli W3110. Затем трансформированные штаммы культивируют на минимальной среде, такой как М9, содержащей в качестве аналога L-лейцина 4-аза-D,L-лейцин или 3-гидрокси-D,L-лейцин. Штаммы, несущие рекомбинантную ДНК, содержащую ген IPMS дикого типа, не способны синтезировать L-лейцин, и их рост подавляется, так как IPMS, экспрессированная из рекомбинантной ДНК, ингибируется аналогом L-лейцина. В отличие от этого штамм, несущий рекомбинантную ДНК, содержащую ген IPMS, в котором ингибирование активности L-лейцином нарушено, содержит мутантный фермент, кодированный геном IPMS, в упомянутой выше рекомбинантной ДНК и не ингибируемый аналогом L-лейцина. Таким образом, он должен обладать способностью к росту на минимальной среде, в которую добавлен аналог L-лейцина. Данное свойство может быть использовано для отбора штамма, обладающего способностью к росту в присутствии аналога L-лейцина, то есть штамма, несущего рекомбинантную ДНК, содержащую мутантный ген IPMS, ингибирование активности в котором нарушено.
Полученный таким способом мутантный ген может быть введен в качестве рекомбинантной ДНК в соответствующий микроорганизм-хозяин и экспрессирован. Таким образом, может быть получен микроорганизм, у которого IPMS утратила чувствительность к ингибированию L-лейцином по типу обратной связи. Хозяином предпочтительно является микроорганизм, принадлежащий к роду Escherichia, примером которого служит Е. coli.
В качестве альтернативы фрагмент с мутантным геном IPMS может быть извлечен из рекомбинантной ДНК и вставлен в другой вектор.
Предпочтительной векторной ДНК, которая может быть использована в настоящем изобретении, является плазмидная векторная ДНК, примерами которой служат векторы pUC19, pUC18, pBR322, pHSG299, pHSG298, pHSG399, pHSG398, RSF1010, pMW119, pMW118, pMW219 и pMW218. Кроме того, могут быть также использованы векторы на основе фаговой ДНК.
Затем с целью эффективной экспрессии мутантного гена IPMS перед последовательностью ДНК, кодирующей мутантную IPMS, может быть встроен другой действующий в таких микроорганизмах промотор, такой как lac, trp и PL, либо может быть использован содержащийся в гене IPMS промотор как таковой или после его амплификации.
Дополнительно, как описывалось выше, мутантный ген может быть введен в векторную ДНК, способную к автономной репликации, при этом его вводят в микроорганизм-хозяин, в котором он содержится в качестве экстрахромосомной ДНК, такой как плазмида. В качестве альтернативы мутантный ген может быть интегрирован в хромосому микроорганизма-хозяина методом трансдукции с использованием транспозона (смотри Berg, D. Е. и Berg, С.М., Bio/Technol., 1,417 (1983)), Mu-фагов (смотри выложенный японский патент No. 2-109985) или гомологичной рекомбинации (смотри "Experiments in Molecular Genetics", Cold Spring Harbor Lab. (1972)).
<3> Способ получения штамма бактерии - продуцента L-лейцина.
Способом получения штамма бактерии - продуцента L-лейцина согласно настоящему изобретению является способ получения штамма бактерии - продуцента L-лейцина путем трансформации с помощью ДНК согласно настоящему изобретению.
Трансформация с помощью ДНК согласно настоящему изобретению может быть осуществлена в соответствии с обычными и известными способами. Например, фрагмент, включающий ДНК согласно настоящему изобретению, сшивают с вектором, функционирующим в микроорганизме-хозяине (подлежащем трансформации) с целью получения рекомбинантной ДНК, а рекомбинантную ДНК вводят в микроорганизм-хозяин. Вектор может быть соответствующим образом выбран в зависимости от микроорганизма-хозяина. Введение рекомбинантной ДНК осуществляют в соответствии с обычными и известными способами. Например, возможно использовать способ, согласно которому клетки-реципиенты (как сообщается, Escherichia coli К-12) подвергают обработке хлоридом кальция с целью повышения их проницаемости для ДНК (смотри Mandel, M. и Higa, A., J. Mol. Biol., 53, 159 (1970)), а также способ, согласно которому компетентные клетки (как сообщается, Bacillus subtilis) получают из клеток на стадии пролиферации с целью введения в них ДНК (смотри Duncan, C.H., Wilson, G.F. и Young, F. E., Gene, 1, 153 (1977)). Также в качестве альтернативы возможно применить способ, в котором с целью введения рекомбинантной ДНК в реципиенты ДНК, такие как Bacillus subtilis, актиномицеты и дрожжи, клетки-реципиенты ДНК преобразуют в протопласты или сферопласты, легко инкорпорирующие рекомбинантную ДНК (смотри Chang, S. и Choen, S.N., Molec. Genet, 168, 11 (1979); Bibb, M.J, Ward, J.M. и Hopwood, O.A., Nature, 274, 398 (1978); Hinnen, A., Hicks, J.В. и Fink, G.R, Proc. Natl. Acad. Sci. USA, 75, 1929 (1978)). Также может быть использован способ электроимпульсов (смотри выложенную японскую патентную заявку 2-207791 (1990)). Соответствующий способ введения может быть выбран в зависимости от микроорганизма-хозяина. В частности, примерами вектора и способа введения служат описанные выше в разделе <2>.
Используемый в данном описании оборот "обладающий способностью продуцировать L-лейцин" означает способность к накоплению L-лейцина в подходящей для этого среде предпочтительно в таком количестве, что L-лейцин может быть легко извлечен из такой среды.
Бактерии, использующиеся в настоящем изобретении, принадлежат предпочтительно к роду Escherichia. Их примером служит E. coli. Примером бактерии, принадлежащей к роду Escherichia и обладающей способностью к продукции L-лейцина, могут служить бактерии, обладающие устойчивостью к аналогу лейцина, такому как бета-2-тиенилаланин, 3-гидроксилейцин, 4-азалейцин и 5,5,5-трифторлейцин, описанным в японской патентной публикации 62-34397 (1987 г.) и выложенной японской патентной заявке 8-70879 (1996 г.), а также бактерия, которая может быть выведена методом генной инженерии, как это описано в заявке WO96/06926.
В бактерии, принадлежащей к роду Escherichia, L-лейцин синтезируется в специальном пути биосинтеза, который ответвляется от конечного интермедиата (2-кетоизовалериата) валиновой биосинтетической системы, В бактерии, относящейся к роду Escherichia, конечная стадия биосинтеза L-валина и L-лейцина осуществляется группой ферментов, кодированных опероном ilvGMEDA и опероном leuABCD соответственно.
Оперон leuABCD включает гены leuA, leuB, leuC и leuD. При этом ген leuA кодирует IPMS, ген leuB кодирует бета-изопропилмалат дегидрогеназу, ген leuC и ген leuD кодируют альфа-изопропилмалат изомеразу. Из перечисленных ферментов IPMS катализирует реакцию синтеза альфа-изопропилмалат, из альфа-кетоизовалериата, альфа-изопропилмалат изомераза катализирует реакцию изомеризации альфа-изопропилмалата в бета-изопропилмалат, а бета-изопропилмалат дегидрогеназа катализирует реакцию дегидрогенизации бета-изопропилмалата в альфа-кетоизокапроновую кислоту, являющуюся конечным промежуточным продуктом биосинтеза L-лейцина.
Из числа упомянутых выше реакций, проходящих в процессе биосинтеза L-лейцина, стадией, определяющей скорость, является реакция синтеза альфа-изопропилмалата, катализированная альфа-изопропилмалат синтазой, подверженной ингибированию L-лейцином по типу обратной связи. Таким образом, трансформация с помощью ДНК, кодирующей IPMS, десенсибилизированную к ингибированию по типу обратной связи, может придать микроорганизму способность продуцировать L-лейцин или улучшить его способность продуцировать L-лейцин.
У бактерии, использующейся в настоящем изобретении и относящейся к роду Escherichia, активность одного или нескольких ферментов биосинтеза L-лейцина может быть усилена с помощью обычной мутационной обработки или методами генной инженерии. Такое усиление активности фермента может быть осуществлено за счет введения в плазмиду, фаг или транспозон бактерии, относящейся к роду Escherichia, рекомбинантной ДНК, полученной в результате инсерции фрагмента ДНК, содержащего целый оперон ilvGMEDA и/или оперон leuABCD или их части.
Анализ нуклеотидной последовательности оперона leuABCD был дан в Nucleic Acid Res. , 20, 3305-3308 (1992). Полная последовательность оперона leuABCD зарегистрирована в базе данных (регистрационный номер DDBJ D10483, адрес в интернете: http: //www. ddbj. nig. ac. jp). Фрагмент ДНК, включающий оперон leuABCD, может быть получен за счет амплификации фрагмента ДНК медотом ПЦР (полимеразной цепной реакцией, смотри White, Т.J. и др.; Trends Genet., 5, 185 (1989)), в процессе которой олигонуклеотиды, полученные на основе описанной выше последовательности, используют в качестве затравок, а хромосомная ДНК бактерии, принадлежащей к роду Escherichia, используется в качестве матрицы для ПЦР. В качестве альтернативы оперон leuABCD может быть также получен методом скрининга библиотеки хромосомных ДНК бактерии, принадлежащей к роду Escherichia, на основе гибридизации за счет использования олигонуклеотидного зонда, полученного на основе описанных выше последовательностей.
Полная нуклеотидная последовательность оперона ilvGMEDA и нуклеотидная последовательность области, расположенной перед опероном, описаны в Nucleic Acid Res, 15, 2137-2155 (1987) и Gene, 97, 21-27 (1991) соответственно. Фрагмент ДНК, содержащий оперон ilvGMEDA, может быть получен в результате осуществления ПЦР или гибридизации с использованием олигонуклеотидного зонда или затравок, полученных на основе описанной выше последовательности. В связи с этим в случае, когда с целью получения оперона UvGMEDA используется штамм Escherichia coli К-12 или его производные, предпочтительно использовать штамм с обратной мутацией гена ilvG, рамка в котором восстановлена таким образом, чтобы восстановилась активность синтазы ацетогидроксикислот. Способы получения оперона ilvGMEDA и способ амплификации оперона в клетке бактерии, принадлежащей к роду Escherichia, полностью описаны в заявке WO96/06926 и французском патенте 2627508 соответственно.
<4> Способ получения L-лейцина
С высокой эффективностью L-лейцин может быть получен путем выращивания штамма бактерии, полученной способом, описанным выше, в питательной среде с последующим выделением L-лейцина из культуральной жидкости.
В процессе осуществления способа получения L-лейцина согласно настоящему изобретению выращивание бактерии, принадлежащей к роду Escherichia, извлечение L-лейцина из культуральной жидкости и его очистка могут быть произведены аналогично способу обычной ферментации, которым L-лейцин получают с использованием штаммов бактерий. Среда, используемая для выращивания, может представлять собой синтетическую либо натуральную среду при условии, что она содержит источники углерода и азота и, если необходимо соответствующее количество питательных веществ, необходимых для роста используемой бактерии. К источникам углерода относятся один или несколько различных углеводов, таких как глюкоза и сахароза и различные органические кислоты. В зависимости от типа ассимиляции используемой бактерии, могут использоваться спирты, такие как этанол и глицерин. В качестве источника азота могут быть использованы различные соли аммония, такие как аммиак, сульфат аммония, другие азотистые соединения, такие как амины, натуральный источник азота, такой как пептон, продукт гидролиза соевых бобов или переваренный ферментативный микроб. В качестве минеральных веществ могут быть использованы фосфат калия, сульфат магния, хлорид натрия, сульфат железа (II), сульфат марганца или карбонат кальция.
Выращивание осуществляют предпочтительно в аэробных условиях, таких как качалочная культура, а также аэрационная и перешиваемая культура при температуре от 20 до 40 oС, предпочтительно от 30 до 38oС. Уровень рН культуры обычно составляет от 5 до 9, предпочтительно от 6,5 до 7,2. Уровень рН может быть отрегулирован с помощью аммиака, карбоната кальция, различных кислот, различных щелочей и буферов. Как правило, в результате выращивания в течение от 1 до 3 суток в жидкой среде происходит накопление целевого L-лейцина.
После выращивания нерастворимые вещества, такие как клетки, удаляют из жидкой среды путем центрифугирования и фильтрации через мембрану, после чего целевой L-лейцин может быть собран и очищен методом ионного обмена, концентрации и осаждения.
Микроорганизм согласно настоящему изобретению может использоваться в качестве продуцирующего L-лейцин штамма или в качестве исходного источника для выведения штамма - продуцента L-лейцина. Настоящее изобретение позволяет получать L-лейцин более эффективно по сравнению с ранее известным способом получения L-лейцина с использованием микроорганизмов.
Примеры
Ниже настоящее изобретение будет более конкретно проиллюстрировано со ссылкой на Примеры.
Пример 1: Получение ДНК, кодирующей мутантную IPMS.
<1> Получение продуцирующих L-лейцин штаммов.
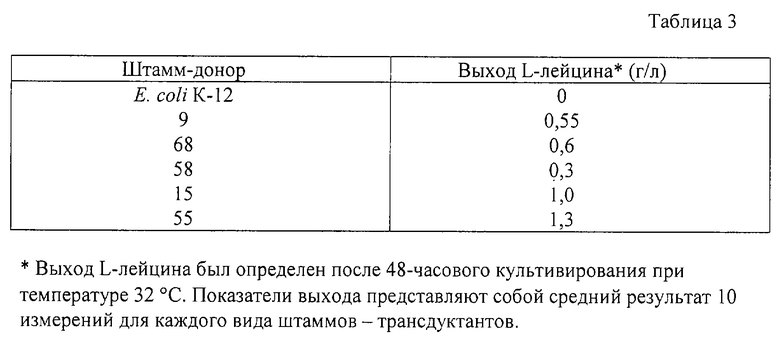
Из стандартного лабораторного штамма Е. coli дикого типа К-12 описанным ниже методом отбора были получены штаммы Escherichia coli, продуцирующие L-лейцин. Клетки штамма Е. coli К-12 в течение 30 мин при температуре 37oС подвергались обработке раствором мутагена, содержащим 0,2 мг/мл N-метил-N'-нитро-N-нитрозогуанидина. Затем клетки были дважды промыты 0,8-процентным раствором NaCl и помещены на чашки с агаризованной средой М9, содержащей 1 мг/мл 4-азалейцина - аналога L-лейцина. Выросшие через 5 суток инкубации при температуре 37oС колонии были собраны и была проверена их способность продуцировать L-лейцин. Продуцентами L-лейцина оказались штаммы 9, 68, 58, 55 и 15.
<2> Получение гена leuA из штаммов - продуцентов L-лейцина.
Методом клонирования in vivo с использованием дефектного бактериофага Mu d5005 (смотри Groisman, Е. А. и др., J. Bacteriol., 168, 357 (1986)) были получены ген leuA из выделенного мутанта 55 - продуцента лейцина и ген leuA штамма Е. coli из дикого типа К-12. Затем перекрывающиеся фрагменты клонированного таким способом гена leuA были амплифицированы методом ПЦР с использованием ДНК-затравок leuA1 и leuA2, leuA3 и leuA4, leuA5 и leuA6, leuA7 и leuA8, lеиА9 и leuA10 соответственно, как это показано в таблице 1.
Такие же затравки ДНК были без предварительного клонирования использованы для амплификации генов leuА, выделенных из хромосом штаммов 9, 68, 58 и 15. Последовательности нуклеотидов полученных фрагментов были определены методом дидезокси-терминации роста цепи.
Выделенные из штаммов 9, 68, 58 и 15 гены leuA содержали мутации, проиллюстрированные в таблице 2.
Клетки штаммов выращивали в течение 10 часов при температуре 32oС в среде, содержащей глюкозу (6%), сульфат аммония (1,5%), дигидрофосфат калия (0,2%), сульфат магния (0,1%), мел (2,5%) и тиамин (0,1 мг/л). Методом дезинтеграции ультразвуком были получены клеточные экстракты, а в качестве ферментного препарата был использован осадок, высаженный сульфатом аммония. Удельная активность IPMS была определена по методу Kohlaw и др. (смотри J. Biol. Chem., 244, 2218 (1969)). I50 - концентрация лейцина, при которой происходит 50-процентное ингибирование активности ферментов. Способность к продукции L-лейцина была определена после 48-часового культивирования в описанной выше среде.
Результаты в суммированном виде приведены в таблице 2.
Пример 2: Продуцирование L-лейцина полученными трансформантами.
Штамм Е. coli C600 (leu-) (смотри Appleyard R.K., Genetics, 39,440-452 (1954) и штамм Е. coli К-12 были подвергнуты трансдукции фагами Р1, выращенными на продуцирующих лейцин штаммах 9, 68, 58, 55 и 15. Были получены трансдуктанты Leu+, подвергнутые испытанию на способность продуцировать L-лейцин. Его результаты приведены в таблице 3.
Перечень последовательностей приведен в конце описания.








Изобретение относится к биотехнологии. Мутантная альфа-изопропилмалат синтаза (IPMS) получена путем делеции, замещения, инсерции или добавления одного или нескольких аминокислотных остатков, при этом свойства мутантной IPMS идентичны природному белку. ДНК, кодирующая IPMS дикого типа, имеет мутацию, позволяющую IPMS утратить чувствительность к ингибированию L-лейцином по типу обратной связи. ДНК трансформирует штамм Escherichia coli, который культивируют для получения L-лейцина. Изобретение позволяет увеличить выход L-лейцина, который используется в качестве пищевой добавки, реагента, ростового фактора. 4 с. и 2 з.п. ф-лы, 3 табл.
| Штамм вниигенетика-10-89-продуцент изолейцина | 1977 |
|
SU751829A1 |
| US 5744331, 28.04.1998. | |||
