Изобретение относится к способу порционной ферментации с подпиткой при использовании специальных систем хозяин-вектор ДНК для эффективного образования рекомбинантных протеинов, в особенности рекомбинантных молекул антител, в частности фрагментов антител, как, например, миниантитела.
В предлагаемых согласно изобретению условиях клетки Е. coli могут расти с максимальной удельной скоростью роста вплоть до очень высоких плотностей клеток. После начала образования рекомбинантного продукта только образовавшийся продукт действует ограничивающе на рост; ограничения роста за счет субстрата или метаболических побочных продуктов не происходит. Таким образом и в связи со специально приспособленными для этой цели и обладающими высокой стабильностью новыми векторами экспрессии можно достигать высоких выходов в единицу времени и на единицу объема рекомбинантных протеинов, которые, в частности, в случае фрагментов антител обладают высокой биологической активностью.
Существенной предпосылкой для эффективного образования рекомбинантных протеинов является культивирование клеток E. coli до высоких плотностей клеток. Для этой цели существуют следующие согласно уровню техники способы культивирования: после неограниченного роста (μ = μmax) в порционной фазе (Batch-Phase) в последующей фазе периодической подпитки (Fed-Batch-Phase) источник углерода обычно (глюкозу или глицерин) добавляют в таком ограниченном количестве, что избегают образования тормозящих рост побочных продуктов, как, например, ацетат, с тем последствием, чтобы рост тогда мог продолжаться вплоть до достижения высоких плотностей клеток, лимитируемый только субстратом (μ < μmax) (например, Riesenberg и др., J. Biotechnol., 20, 17-28 (1991); Strandberg и др., FEMS Microbiol. Rev., 14, 53-56 (1994); Korz и др. , J. Biotechnol. , 39, 59-65 (1995); европейский патент В-0511226). Рост с пониженной нормой роста имеет следствием, естественно, длительные времена ферментации и, таким образом, следовательно, также незначительные выхода в единицу времени на единицу объема. В случае этих ферментации на основании немедленного потребления концентрация источника углерода в культуральном растворе равна почти нулю. После начала образования рекомбинантного продукта ничто не изменяется в лимитируемых субстратом соотношениях.
Известны также периодические культивирования с подпиткой с помощью Е. coli, при которых источник углерода добавляют прерывисто через гораздо большие промежутки времени и тогда в гораздо больших количествах, причем чаще всего используют повышение рO2-значения (величина парциального давления кислорода) в качестве индикатора истощения субстрата для начала следующей дозировки источника углерода (например, Eppstein и др., Biotechnol., 7, 1178-1181 (1989)). Этот образ действий означает частое изменение длительных по времени условий избытка субстрата до условий лимитирования субстрата и таким образом включает метаболические дисбалансы.
В последующем принимают во внимание периодичного культивирования с подпиткой, при которых клетки в фазе порционной подпитки могут расти с максимальной удельной скоростью роста (μ = μmax). Порционные культивирования с подпиткой, при которых после вне линейных (off-line) определений добавляют гораздо большие количества источника углерода через гораздо большие промежутки времени процесса культивирования с целью устранения лимитирования субстрата и которые являются экспериментально применимыми и имеют тот недостаток, что во время всего процесса ферментации постоянно изменяется концентрация источника углерода (например, Pack и др., Biotechnol., 11, 1271-1277 (1993); Hahm и др., Appl. Microbiol. Biotechnol., 42, 100-107 (1994)).
Описываются также порционные культивирования с подпиткой, при которых концентрацию источника углерода замеряют и регулируют внутри линии (on-line), так чтобы избежать лимитирования, хотя этим культивированиям присущи нижеописываемые недостатки, в частности, в области высокой плотности клеток. Недавно описано использование в микробиологических ферментациях, осуществляемых в ферментерах типа котла с мешалкой, автоклавируемого биосенсора глюкозы (M. R. Phelps и др., Biotechnol. Bioeng., 46, 514-524 (1995)). Его используют для культивирования культуры E. coli. Этот in situ-сенсор поставляет с задержкой во времени примерно 2 минуты действительное значение в культуральном растворе. Поставляемый сенсором глюкозы сигнал между прочим зависит от значения рН и парциального давления кислорода (рО2). В области высокой плотности клеток Х>80 г/л сенсор не апробирован. Как показывает опыт, обрастания in situ-зондов с помощью E. coli в условиях очень высоких плотностей клеток могут приводить в значительной степени к дальнейшим ошибочным значениям. Кроме того, сенсор во время процесса протекания ферментации точно не рекалибруется. Другие способы базируются не на измерениях с помощью in situ-сенсора, а основываются, например, на определении источников углерода с помощью анализаторов ввода в потоке (FIA) или в процессе высокоэффективной жидкостной хроматографии в освобожденном от клеток культуральном растворе, который полунепрерывно отбирают из ферментера и подвергают фильтрации или микроцентрифугированию (Kleman и др., Appl. Environ. Microbiol. , 57, 910-917 и 918-923 (1991); Turner и др., Biotechnol. Bioeng., 44, 819-829 (1994)). Предсказанные и обратносвязанные контрольные алгоритмы прогнозирования уменьшают колебания концентрации глюкозы при росте вплоть до Х= 65 г/л (Kleman и др., Appl. Environ. Microbiol., 57, 910-917 (1991)). В области очень высоких плотностей клеток (начиная примерно с 80 г/л до 150 г/л) разделение клеток и питательного раствора становится в возрастающей степени более затруднительным и требующим больших затрат времени, так что также в зависимости от биомассы в ферментере увеличивается задержка во времени определения действительного содержания глюкозы и затрудняется, соответственно становится невозможным поддерживание постоянным уровня глюкозы. В противоположность этому, с постоянной и кратковременной задержкой по времени определяют концентрацию глюкозы в аппаратуре, для использования которой не нужно осуществлять этого отделения клеток (Pfaff и др., Proceedings of the 6th International Conference on Computer Appl. in Biotechnol. Garmisch-Partenkirchen, ФРГ, 1995, с.6-11). Согласно Пфаффу и др., в непосредственной близости от места отбора проб после разбавления культуры ингибитором роста используют FIA с ферментативно-ампериометрическим сенсором глюкозы.
При аэробном культивировании образуются клетки Е. coli, которых не принуждают к субстрат-лимитируемому росту путем режимов дозировки, обычно усиливает это метаболический побочный продукт ацетат (Riesenberg, Curr. Opinion Biotechnol. , 2, 380-384 (1991)), который аккумулируется в питательном растворе и в повышенных количествах действует как ингибитор роста (Pan и др. , Biotechnol. Lett. , 2, 89-94 (1987)). Поэтому до сих пор эти порционные культивирования с подпиткой до высоких плотностей клеток были возможны только при использовании особых штаммов E. coli, в случае которых аккумуляция ацетата снижена за счет целенаправленных генетических изменений, при допустимости других, связанных с этим недостатков. К потомкам E. coli K-12 относятся фосфотрансацетилаза - отрицательные мутанты (Bauer и др., Appl. Environ. Microbiol. , 56, 1296-1302 (1990), Hahm и др., Appl. Microbiol. Biotechnol. , 42, 100-107 (1994)), рост которых, однако, в среде глюкоза - минеральные соли сильно уменьшен. Phelps и др. (см. выше) в качестве хозяина использовали штамм Е. coli TOPP5 для не лимитируемого субстратом культивирования вплоть до получения биомассы Х=85 г/л. Этот штамм Е. coli, который очевидно не сильно аккумулирует ацетат, однако, не является никаким штаммом K-12. Е. coli TOPP5 образует гемолизин и таким образом представляет собой патогенный штамм, который по соображениям безопасности не используют в промышленном секторе в качестве хозяина для образования рекомбинантных ДНК-продуктов. Путем трансформации клеток Е. coli с помощью плазмиды, которая содержит ген для кодирования ацетолактатсинтазы (ALS), благодаря достигнутой переориентации промежуточного обмена веществ можно достичь восстановления аккумуляции ацетата (San и др. , Ann-N-Y-Acad-Sci., 721, 257-267 (1994)). Этому образу действий, однако, присущ недостаток, заключающийся в том, что при использовании ALS-кодирующей плазмиды в сочетании со второй, несущей "производственный" ген плазмидой в условиях высокой плотности клеток обычно появляются нестабильности. За счет нестабильностей плазмид, которые в особенности проявляются усиленно при культивировании до очень высоких плотностей клеток, зачастую снижается эффективность образований рекомбинантных продуктов.
Антитела или фрагменты антител, как Fab', F(ab')2, миниантитела или одноцепочечные Fv, в возрастающей степени приобретают значение в области медицины и биотехнологии. Под миниантителом нужно понимать, далее, согласно изобретению, бивалентный или биспецифический, связанный через псевдошарнирную область одноцепочечный Fv-фрагмент. При этом, как, например, в терапии раковых заболеваний может оказаться важной доступность иметь в распоряжении большие количества антител (примерно 1 г на дозу). В E. coli в настоящее время можно особенно легко и успешно получать моновалентные фрагменты антител или слитые протеины из них, соответственно их мультимерные или мультиспецифические варианты. Эти фрагменты, соответственно варианты имеют маленький размер, связанный с высокой специфической способностью связывания (например,  A., Immunol. Rev., 130, 151-188 (1992); Pack и др., J. Mol. Biol., 246, 28-34 (1995)). Протеины, в частности антитела, однако, должны иметь соответствующую складчатую структуру, чтобы быть биологически и функционально эффективными. Эту проблему нужно принимать во внимание в случае рассмотрения выхода образовавшегося фрагмента антитела на клетку в связи с плотностью клеток. Далее, первичная последовательность антитела играет важную роль при определении выхода in vitro и складчатости in vivo (Knappik А. и
A., Immunol. Rev., 130, 151-188 (1992); Pack и др., J. Mol. Biol., 246, 28-34 (1995)). Протеины, в частности антитела, однако, должны иметь соответствующую складчатую структуру, чтобы быть биологически и функционально эффективными. Эту проблему нужно принимать во внимание в случае рассмотрения выхода образовавшегося фрагмента антитела на клетку в связи с плотностью клеток. Далее, первичная последовательность антитела играет важную роль при определении выхода in vitro и складчатости in vivo (Knappik А. и  A., Protein Engin., 8, 81-89 (1995)). Так, например, Fab-фрагменты экспрессируются в виде нерастворимых цитоплазматических или периплазматических агрегатов и in vitro возвращаются в складки. Так, сообщается о выходах нерастворимых антител примерно от 0,14 г/л в случае низкой плотности клеток (Condra и др. , J. Biol. Chem., 265, 2292-2295 (1990)) и вплоть до примерно 1-2 г/л при средней плотности клеток (Shibui и др., Appl. Microbiol. Biotechnol., 38, 770-775 (1993)). Также бивалентные миниантитела (Pack и др. , Biotechnol. , 11, 1271-1277 (1993)) можно получать в E. coli в биологически-функциональной форме с выходами примерно 0,2 г/л. В случае этих выходов в среднем надлежащим образом возвращаются в складки примерно 5-45%.
A., Protein Engin., 8, 81-89 (1995)). Так, например, Fab-фрагменты экспрессируются в виде нерастворимых цитоплазматических или периплазматических агрегатов и in vitro возвращаются в складки. Так, сообщается о выходах нерастворимых антител примерно от 0,14 г/л в случае низкой плотности клеток (Condra и др. , J. Biol. Chem., 265, 2292-2295 (1990)) и вплоть до примерно 1-2 г/л при средней плотности клеток (Shibui и др., Appl. Microbiol. Biotechnol., 38, 770-775 (1993)). Также бивалентные миниантитела (Pack и др. , Biotechnol. , 11, 1271-1277 (1993)) можно получать в E. coli в биологически-функциональной форме с выходами примерно 0,2 г/л. В случае этих выходов в среднем надлежащим образом возвращаются в складки примерно 5-45%.
В известных E. coli системах образование чужеродного протеина начинается, как правило, пригодным образом после достижения соответствующих плотностей клеток согласно системе экспрессии благодаря регулируемой промоторной системе. В этом случае, например, нужно назвать (i) araBAD-промотор в присутствии АrаС-репрессора (индуцируемый арабинозой) (например, Better и др., Рrос. Natl. Acad. Sci. (USA), 90, 457-461 (1993)); (ii) phoA-промотор (индуцируемый за счет удаления фосфата) (например, Carter и др., Biotechnol. , 10, 163-167 (1992)) и (iii) lac-промоторную систему (индуцируемую с помощью изопропилтиогалактозида [IPTG]) (Pack и др., 1993, цитировано выше). lac-Система, которая, как правило, вызывает успешную экспрессию, однако обладает тем недостатком, что, с одной стороны, может происходить нежелательная основная экспрессия до индукции промотора и, с другой стороны, может наблюдаться нестабильность плазмиды после индукции с помощью изопропилтиогалактозида (IPTG).
В особой форме выполнения изобретения описывается специальный вектор (рНКК), который в качестве чужеродного гена содержит последовательности, кодирующие фрагменты мышиного, соответственного гуманизированного антитела - МАb 425. МАb 425 (АТСС НВ 9629) представляет собой мышиное моноклональное антитело, которое выделено из известной человеческой линии раковых клеток А432 (АТСС CRL 1555) и связано с эпитопом человеческого рецептора эпидермального фактора роста (EGFR, гликопротеин величиной примерно 170 килодальтонов) при ингибировании связи природного лиганда эпидермального фактора роста (EGF). Было установлено, что МАb 425 оказывает цитотоксическое действие на опухолевые клетки, соответственно, может препятствовать их росту (Rodeck и др. , Cancer Res., 47, 3692 (1987)). Из международной заявки WO 92/15683 известны гуманизированные, а также химерные формы МАb 425, включая последовательности ДНК и аминокислотные последовательности их легких и тяжелых цепей.
Целью изобретения, таким образом, является разработка способа получения чужеродных протеинов, в частности фрагментов антител, в рекомбинантных клетках E. coli в условиях высокой плотности клеток (HCDC = культивирование в условиях высокой плотности клеток) с высокими выходами в единицу времени на единицу объема и без существенного ухудшения роста за счет субстратов или метаболитов и без заметных потерь плазмид соответственно за счет нестабильностей плазмид, при обеспечении высокого процента эффективной биологической активности (способность связывания, корректная укладка) экспрессированного протеина.
Предлагаемый согласно изобретению способ представляет собой многостадийный порционный способ, который, в первую очередь, характеризуется тем, что клетки в течение всей порции могут максимально расти (μ = μmax). Так, с помощью описываемого способа в конце концов можно достигать плотности клеток 100-150 г/л (в расчете на сухую биомассу). Рост также незначительно ингибируется путем аккумуляции ацетата, т.к. эта аккумуляция при выбранных условиях неожиданно не особенно выражена, в частности при использовании штаммов E. coli, которые и без того склонны только к уменьшенному образованию ацетата во время ферментации. Это достигается также при ряде других дополнительных мер прежде всего благодаря тому, что в порционной фазе подпитки, заключающей порционную фазу, концентрация источника углерода в среде поддерживается постоянной в определенном диапазоне значений при сохранении неограниченного роста клеток. Благодаря соответствующей форме соответствующего вектора экспрессии также можно почти исключить нежелательную основную экспрессию протеина до начала синтеза протеина за счет регулируемой промоторной системы, точно так же, как отчасти значительную потерю плазмиды, которая, как уже было упомянуто выше, должна обычно наблюдаться в системах экспрессии при использовании сильных промоторов, как, например, 1ас-промоторная система.
В среднем можно достигать выходов протеина от 3 до 5 г/л спустя примерно от 25 до 35 часов протекания всего процесса культивирования. В случае особенно критических из-за их критериев складчатости фрагментов антител, в особенности миниантител, примерно 80% синтезированного материала биологически активно и имеет корректную складчатую структуру.
Таким образом, предметом изобретения является способ получения чужеродного протеина в клетках E. coli, которые трансформируются с помощью чужеродного гена и несущей индуцируемый промотор плазмиды, путем ферментации в условиях высокой плотности клеток через стадии порций и порционной подпитки без всякого ограничения роста благодаря субстратам или метаболическим побочным продуктам, и выделения и очистки экспрессированного протеина из культуральной среды, причем концентрацию на субстратах в фазе порционной подпитки направляют с помощью непрерывно действующей автоматической или полуавтоматической системы анализа и добавления, причем в фазе порционной подпитки (i) концентрация источника углерода в среде поддерживается постоянной в диапазоне от 0,1 г/л до 25 г/л при сохранении неограниченного роста клеток (μ = μmax), (ii) продуцирование чужеродного протеина начинается в указанной фазе порционной подпитки при плотности клеток от 10 до 80 г/л за счет индукции промотора; и (iii) после осуществленной индукции синтеза продукта непрерывно добавляют используемые азот и фосфат, а также соли микроэлементов, причем (iv) во время всей фазы порционной подпитки путем соответствующего введения кислорода в ферментационный бульон величину парциального давления кислорода рO2 устанавливают равной 5-25%.
Предлагаемая согласно изобретению величина необходимой концентрации источника углерода в течение порционной фазы подпитки составляет от 0,1 до 25 г/л, предпочтительно от 0,5 до 15 г/л, в особенности от 1,0 до 5 г/л, соответственно от 1,0 до 3 г/л. Особенно предпочтительная концентрация составляет 1,5 г/л. В качестве источника углерода нужно назвать предпочтительно глюкозу или глицерин или смеси обоих. Согласно изобретению добавление источника углерода осуществляют непрерывно на входе (on-line) с помощью автоматической или полуавтоматической системы добавления и анализа. Предпочтительно используют систему анализа инъекционного потока на входе (FIA).
Добавление используемого азота, предпочтительно аммонийного азота, и фосфата, например диаммонийгидрофосфата или аммонийдигидрофосфата, а также микроэлементов, например растворимых в среде солей бора, марганца, меди, молибдена и кобальта, железа или цинка, осуществляют в порционной фазе подпитки, заключающей порционную фазу, предпочтительно после включения синтеза протеина за счет регулируемого промотора при плотности клеток 50-80 г/л (в расчете на сухую биомассу), предпочтительно примерно при 70 г/л, при общей норме роста от 100 до 150, предпочтительно 140 г/л.
Включение синтеза протеина путем активации регулируемой промоторной системы осуществляют согласно изобретению при плотности клеток 10-80 г/л, предпочтительно 20-60 г/л, в высшей степени особенно предпочтительно в диапазоне значений от 40 до 50 г/л.
Парциальное давление кислорода в течение порционной фазы подпитки составляет 5-25%, предпочтительно 15-25%, в высшей степени особенно предпочтительно 20%.
рН-Значение ферментационной среды согласно изобретению во время всей подпитки устанавливают 6,5-7,0, предпочтительно 6,7-6,9, в особенности 6,8.
Предметом изобретения, далее, является соответствующий способ, при котором используют вектор экспрессии, содержащий чужеродный ген в виде полигенного экспрессирующего кластера, фланкированного двумя терминаторными последовательностями. Эти терминаторные последовательности, в особенности позиционированные, в проксимальном положении успешно предотвращают нежелательную экспрессию протеина до начала за счет промоторной системы. Особенно пригоден терминатор thp (Nohno и др., J. Bacteriol., 170, 4097-4102 (1988)), однако также можно использовать другие известные терминаторные последовательности.
Далее, предметом изобретения является способ, при котором используемый вектор экспрессии дополнительно содержит суицидную систему. Суицидная система продуцирует протеин, который без наличия плазмиды в клетке токсичен для нее. Пригодные суицидные системы известны из литературы. Особенно пригодной согласно изобретению суицидной системой является hok-sok-система (например, Gerdes К., Biotechnol., 6, 1402-1405 (1988)). Для способа в целях эффективного образования рекомбинантных протеинов, в особенности молекул антител, особенно важным является то, что система хозяин-вектор ДНК в области высокой плотности клеток характеризуется высокой плазмидной стабильностью, незначительной рекомбинантной основной экспрессией и высоким образованием продукта. Суицидные системы в сочетании с рекомбинантными, фланкированными с помощью терминаторов экспрессирующими кластерами при этом являются специфичными для вектора.
Далее, предметом изобретения является соответствующий способ, при котором используют чужеродный ген, который кодирует фрагмент антитела, в частности миниантитело.
Далее, предметом изобретения является способ, при котором используют вектор экспрессии с дополнительными, описываемыми ниже признаками. В принципе можно использовать большинство известных, пригодных для технологии рекомбинации и для продуцирования в промышленном масштабе штаммы E. coli. Более предпочтительно находят применение такие штаммы, которые при росте до высокой плотности клеток аккумулируют относительно немного ацетата. Особенно пригодны штаммы, обогащение ацетатом которых составляет величину ниже 5 г/л. Неожиданно оказалось, что благодаря выбранным условиям предлагаемого согласно изобретению способа можно поддерживать особенно низкой аккумуляцию ацетата. В этом отношении особенно пригодным является общеизвестный и имеющийся в продаже штамм E. coli RV308 (ATCC 31608), а также его такие же по действию варианты.
Предметом изобретения, таким образом, в особенности, является соответствующий способ, при котором используют штамм E. coli с аккумуляцией ацетата ниже 5 г/л в культуральной среде во время ферментации.
Предметом изобретения, кроме того, является вектор экспрессии E. coli, пригодный для экспрессии чужеродных протеинов в условиях ферментации при высокой плотности клеток, который обладает следующими признаками:
(i) терминаторная последовательность в проксимальном и дистальном направлении;
(ii) lac-промотор/оператор - система;
(iii) последовательность Шайн-Дальгарно Т7g10;
(iv) pelB или ompA сигнальная последовательность;
(v) последовательность чужеродного гена;
и в предпочтительном варианте осуществления дополнительно включает суицидную систему, в частности hok-sok-суицидную систему.
Согласно изобретению промоторную систему также можно заменить другими пригодными, например вышеуказанными, системами. Точно также изобретение охватывает также другие, одинаково действующие сигнальные и управляемые последовательности.
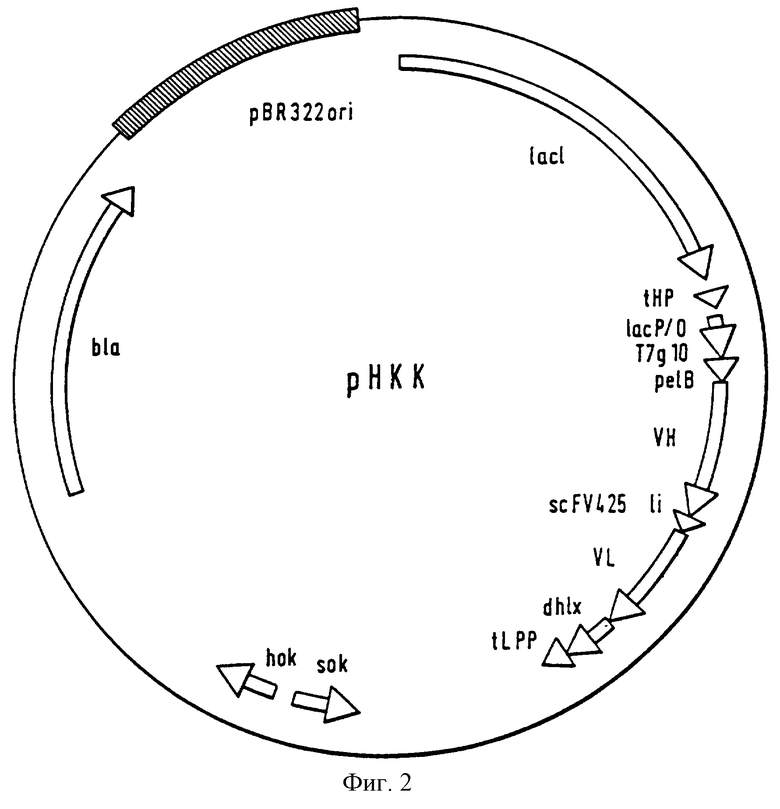
В качестве специальных форм выполнения, наконец, предметом изобретения являются определяемый своей конструкцией вектор экспрессии рНКК (фиг.2), который содержит последовательности для миниантитела, происходящего от МАb 425, а также особый рекомбинантный E. coli-хозяин RV308 [рНКК].
Описание фигур
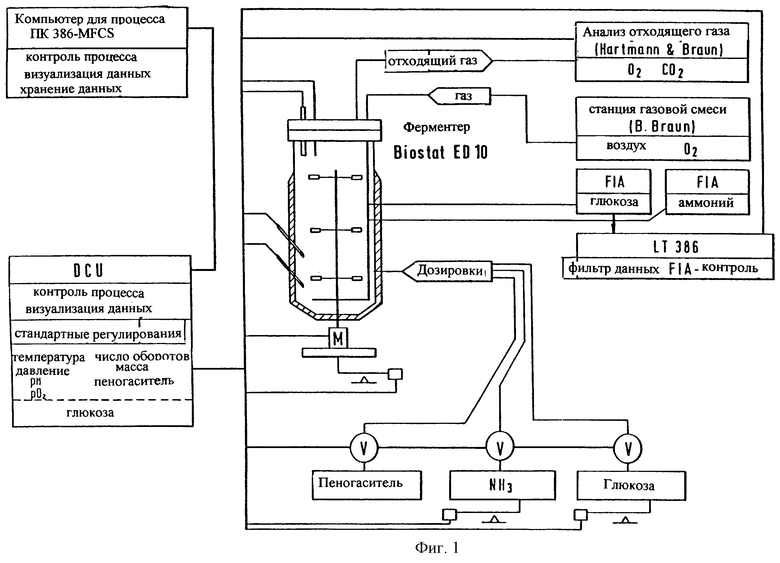
Фиг. 1: Экспериментальная опытная конструкция биореактора для получения протеинов в условиях высокой плотности клеток. Система снабжена измерительным устройством, индикаторным приспособлением, устройствами для контроля и дозировки.
Фиг. 2: Оптимизированный вектор экспрессии рНКК, а также отдельные участки его конструкции. Вектор состоит в основном из отдельных участков известных векторов pASK40, рАК100 и pKGl022.
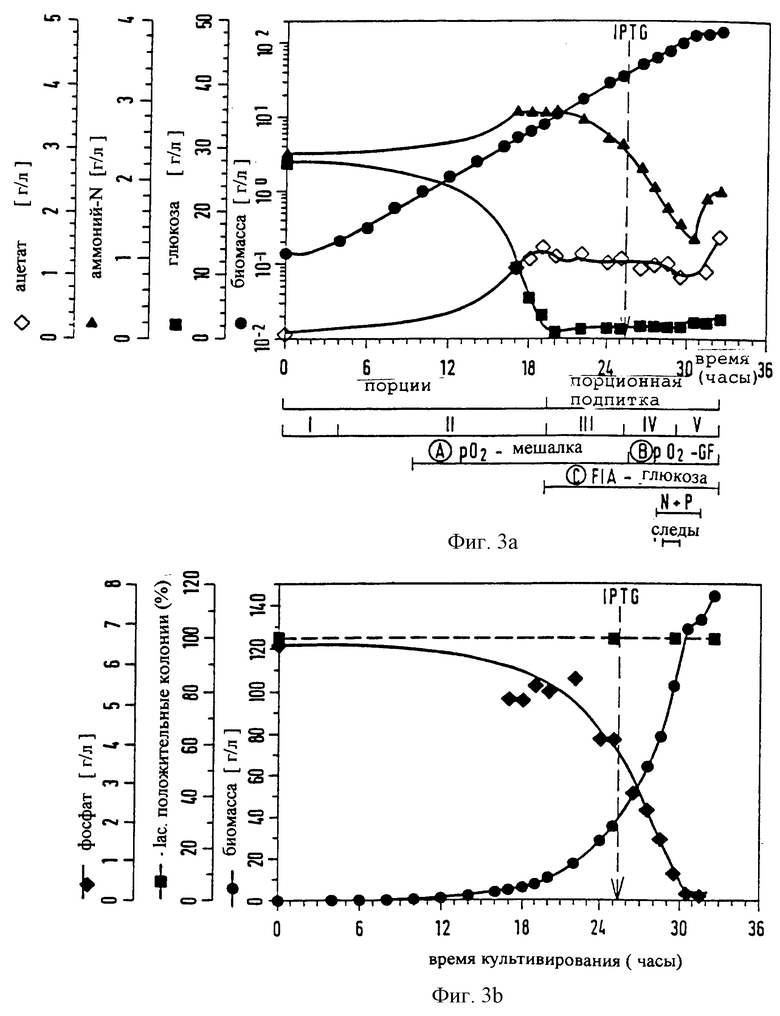
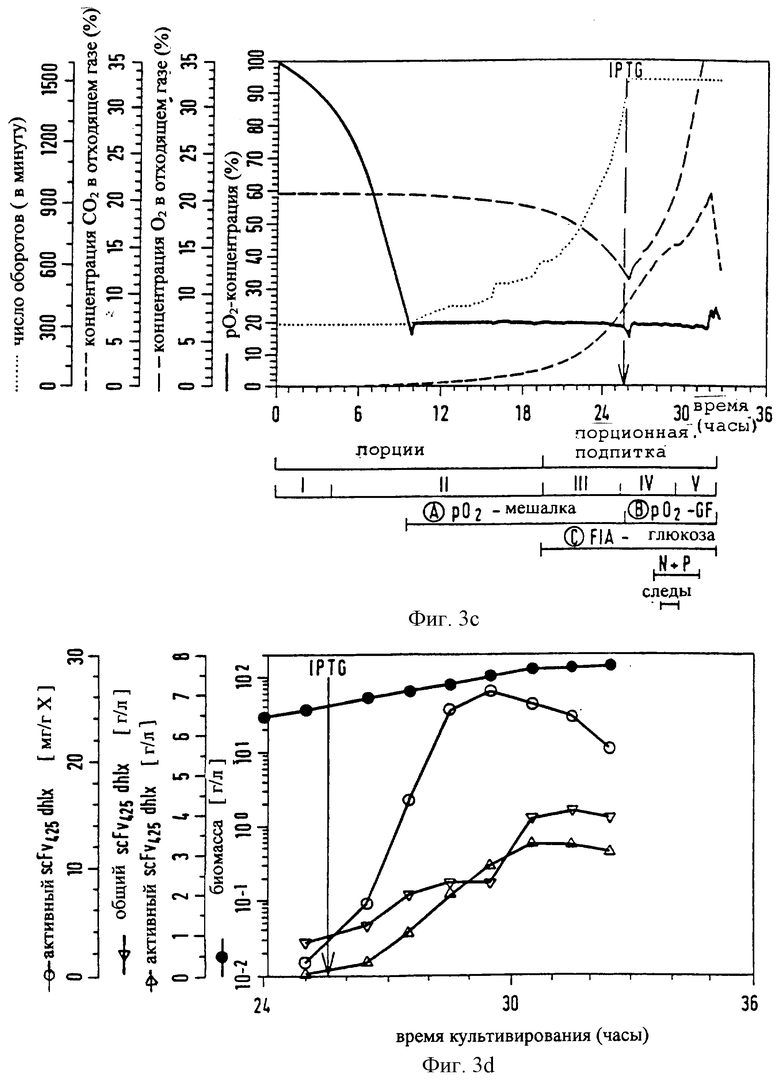
Фиг.3 (a-d): Культивирование при высокой плотности клеток (HCD) рекомбинантной E. coli, например E. coli RV308 [рНКК]: розлив по времени биомассы, глюкозы, аммонийного азота, фосфата, ацетата; скорость мешалки, парциальное давление кислорода; содержание O2 и СО2 в отходящем газе; стабильность плазмиды (выражено в % β-лактамаза-положительных колоний) и образование протеина (здесь: scFv425dhlx). Порционная фаза и фаза порционной подпитки подразделены на 5 подфаз. IPTG-Стрелка указывает старт продуцирования протеина.
В предлагаемом согласно изобретению способе используют трансформированные клетки-хозяева Е. соli. При выборе конструкций плазмид руководствуются родом экспрессированного протеина. Особенно благоприятные признаки таких конструкций описываются ниже. Необходимые для конструкции плазмид и трансформации клеток-хозяев технологии и методы общеизвестны и подробно описываются в литературе (например, Sambrook и др. Лабораторное руководство "Молекулярное клонирование", изд. Cold, Xapбop (1989)). К тому же они представлены в примерах путем указания особых вариантов осуществления изобретения. Исходные плазмиды или части плазмид либо имеются в продаже, либо их можно сразу конструировать по стандартным способам на основе известных схем конструкции.
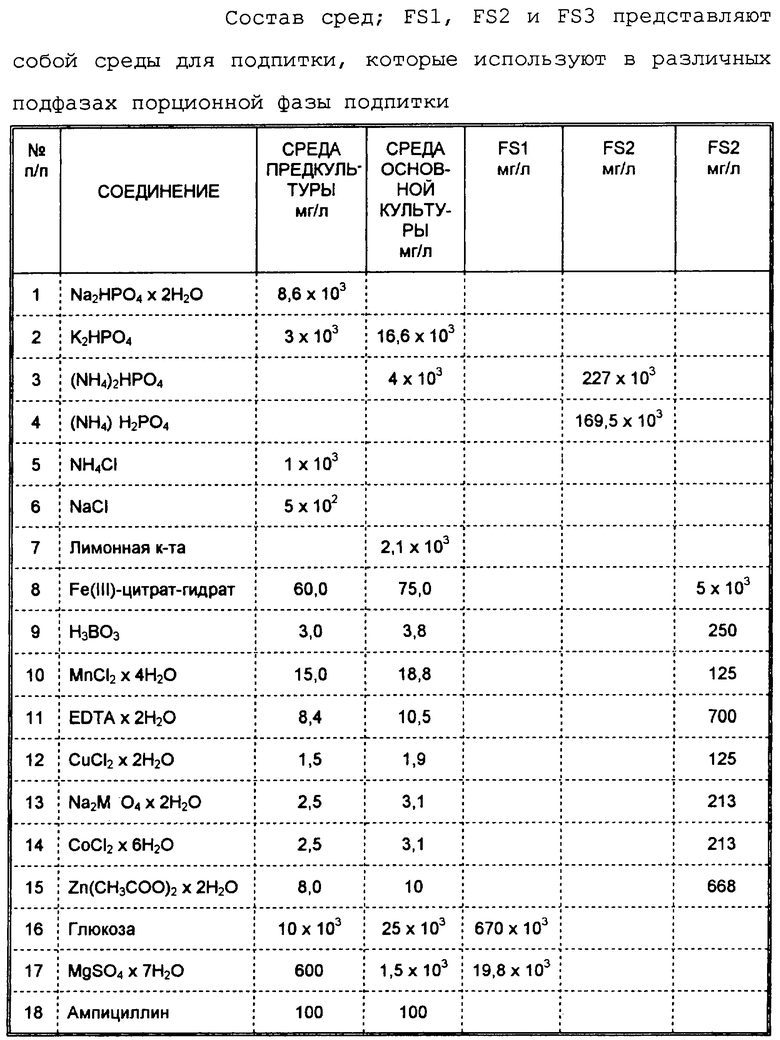
Предварительную порционную фазу типичной согласно изобретению ферментации трансформированных клеток E. coli подразделяют на две подфазы. После инокуляции с помощью соответствующей предкультуры подфаза I характеризуется lac-фазой, в которой клетки адаптируются и норма роста (μ) затем возрастает до μmax. Во время подфазы II клетки растут экспоненциально при μ = μmax. После падения парциального давления кислорода (рO2) от 100% насыщения до значения ниже 5-15% величину парциального давления кислорода путем контролирования скорости рО2-мешалки устанавливают предпочтительно при значении рO2 от 15 до 25%, в частности около 20% (фиг.3с). Это установление (путем пропускания обогащенного чистым кислородом воздуха) нужно осуществлять спустя примерно 6-12 часов после начала ферментации основной культуры. Концентрация глюкозы, которая первоначально составляет предпочтительно 20-35 г/л, снижается вплоть до конца подфазы II, которая также представляет собой конец порционной фазы, предшествующей порционной фазе подпитки. При этом величины не должны ни в коем случае быть ниже 0,1 г/л. Это предотвращают теперь путем соответствующего добавления глюкозы (подфаза III, фиг.3а, начало фазы подпиточной дозировки). Согласно изобретению поддерживают постоянным содержание глюкозы между 0,1 и 25 г/л, предпочтительно, однако, при 1-3 г/л. Для этой цели можно, например, использовать подпиточную питательную среду FS1 (таблица). Т.к. эта область концентрации глюкозы находится в достаточной степени выше Кs-значения для глюкозы (Bergter, "Рост микроорганизмов", с.41, 1983, изд. Gustav Fischer, Йена), клетки могут расти дальше при μmax. Контроль и регулирование концентрации глюкозы согласно изобретению осуществляют с помощью автоматической или полуавтоматической системы.
Особенно пригодными являются (on-line) системы анализа инъекционного потока в непосредственном производстве на входе. Функционирующие за счет чисто ручного или в значительной степени ручного управления системы оказываются неблагоприятными. Порционная фаза подпитки начинается примерно между 15-тым и 22-ым часами, однако в конце концов зависит от различных индивидуальных факторов, таких как температура, состав и концентрация среды, размер реактора и т.д., в особенности, однако, также от рода используемого штамма E. coli. Предпочтительнее примерно спустя 4-8 часов после начала порционной фазы подпитки начинают осуществлять синтез чужеродного протеина. Точный момент времени, однако, в конечном счете выбирают по достигнутой к этому моменту плотности клеток культуры. Плотности клеток от 10 до 80 г/л, предпочтительно 20-60 г/л, являются особенно благоприятными, если можно достигать конечных плотностей клеток от 100 до 150 г/л. Таким образом, вообще благоприятным для начала синтеза протеина является тот момент, когда ко времени индукции имеются примерно 10-60% максимально достигаемой плотности клеток.
Синтез протеина вызывают за счет включения регулируемой промоторной системы. Это включение в зависимости от используемой системы, как правило, осуществляют путем добавления вещества или за счет изменения физической величины, например, в случае предпочтительной lac-системы (промотор, оператор, индуктор) - путем добавления изопропилтиогалактопиранозида (IPTG). Дальнейший рост клеток теперь ограничивается только еще аккумулирующимся продуктом. Поэтому согласно изобретению важным является то, что перед индукцией не может происходить никакой заметной основной экспрессии, которая оказывала бы неблагоприятное влияние на общий рост и, таким образом, на общий выход. Это происходит согласно изобретению благодаря тому, что полигенный экспрессирующий кластер в плазмиде фланкируется активными терминаторными последовательностями.
Начиная с момента добавления глюкозы ферментационная среда обедняется азотом и фосфатом (фиг.3а, b). Для того, чтобы избежать любых ограничений, также осуществляют подпитку азотом и фосфатом путем соответствующего непрерывного способа добавления. Азот добавляют целесообразнее всего в форме аммониевых солей, т.к. таким образом одновременно также можно оказывать влияние на значение рН (6,5-7,0; предпочтительно 6,8). Например, пригоден раствор FS2 (таблица). Подфаза IV характеризуется предлагаемым согласно изобретению подводом микроэлементов (как, например, бор, марганец, железо, кобальт, молибден, олово) в виде их растворимых солей. Добавление осуществляют, как правило, непрерывно с постоянной скоростью. Подфаза V характеризуется уменьшенным ростом, в первую очередь обусловленным аккумуляцией продукта. Кроме того, должно наблюдаться незначительное повышение концентрации ацетата. Однако аккумуляция, соответственно концентрация ацетата в среде неожиданно оказывается низкой. Это также должно объясняться особыми условиями способа. Штаммы E. coli с максимальным накоплением ацетата <5 г/л во время ферментации еще более отчетливо усиливают этот эффект.
Выхода протеина изменяются в зависимости от рода протеина в среднем от 2 до 6 г/л. Из этого количества, опять в зависимости от рода протеина, биологически активными являются 50-95%. В случае фрагментов антител получают свыше 80% возвращенного в складки протеина. Эти значения отчетливо превышают описанные до сих пор в уровне техники сравнимые способы.
Предпочтительно, с помощью описанного способа при использовании особо сконструированных и применимых для этой цели плазмид экспрессии можно эффективно получать фрагменты антител, в особенности миниантитела. Однако согласно предлагаемому в изобретении способу предпочтительным образом возможно также получение многих других протеинов, слитых протеинов или ферментов. Примерами таких пригодных протеинов являются гибридстрептокиназа, глюкозодегидрогеназа или также протеины, которые оказывают воздействие на свертывание крови, как, например, гирудин, тромбин, гементин или теромин.
Пример 1
Для получения рекомбинантного E. coli-хозяина используют прототрофный Е. coli-K-12-штамм RV308 (1ас74-galISII::OP308strA) (Maurer и др., J. Mol. Biol. , 139, 147-161 (1980); АТСС 31608). Трансформацию с помощью пригодного для экспрессии вектора, а также все другие необходимые манипуляции с ДНК осуществляют, если не указано ничего другого, по стандартным методам. Клетки E. coli RV308 используют в качестве контроля при соответствующей ферментации в условиях высокой плотности клеток.
Вектор рНКК конструируют (фиг. 2) следующим образом: маленький MluI-фрагмент из рАК100 (Krebber и Pluckthun, 1995), который содержит "сильный" транскрипционный терминатор tHP (Nohno и др., см. выше) на участке в проксимальном направлении lac р/о, прикрепляют плазмиду pASK40 (Skerra и др., Biotechnol. , 9, 273-278 (1991)). Инсерцию hok-sok-ДНК осуществляют путем двух дальнейших стадий клонирования: арhА-ген из pKG1022 (Gerdes, 1988, см. выше) удаляют путем двукратного переваривания с помощью XhoI и EcoRI, дополняют с помощью ДНК-полимеразы (фрагмент Кленова) и религируют. Во второй стадии модифицированный BamHI-фрагмент из pKG1022 клонируют в единственном месте расщепления BamHI первого продукта клонирования. Миниантитело происходит от одноцепочечного фрагмента, в котором вариабельные домены в VH-VL-направлении связаны с гибким (изменяемым) линкером (g1у4ser)3, идущим за обогащенной пролином шарнирной областью и измененным доменом "спираль-петля-спираль" (dhlx) (Pack и др., Biotechnol., 11, 1271-1277 (1993)), ДНК-последовательности, праймеры, амплификации и клонирования легкой и тяжелой цепи мышиного/ гуманизированного МАb 425 подробно описываются в международной заявке 92/15683. Для секреции scFv425dhlx-фрагмента в периплазме N-конец VH-домена подвергают слиянию с сигнальной последовательностью ре1В. Связывающий сайт Т7g10-рибосом (Шайн-Дальгарно) с помощью методики полимеразно-цепьевой реакции (ПЦР) клонируют в XbaI- и SfiI- местах расщепления pEGl (Strittmatter и др. , 1995). Готовый полигенный экспрессирующий кластер scFv425dhlx клонируют между местами расщепления XhaI и HindIII. В результате получают вектор экспрессии рНКК согласно фиг.2.
Пример 2
Составы сред для предварительных культур в колбах Эрленмейера, основная культура в реакторе типа резервуара, снабженного мешалкой. (Biostat ED10; В. Braun; Biotech International; Melsungen, ФРГ), а также среды для подпитки FS1, FS2 и FS3 представлены в таблице. Основная культуральная среда (8 л) модифицирована по сравнению со средой, описанной Riesenberg и др., 1991 (см. выше). Для того, чтобы предотвратить осаждения, составные части добавляют в указанной последовательности согласно таблице. Глюкозу и сульфат магния добавляют в виде отдельно автоклавированных растворов. Реактор функционирует при температуре 26oС, давлении 0,15 МПа, рН-значении 6,8 и средней скорости аэрации 10 л/мин. Для установления рН-значения используют 25%-ный водный раствор аммиака. Аммиак и Ucolub N115® (Fragol Industrieschmierstoffe GmbH, Muhleim/Ruhr, ФРГ) добавляют с помощью сенсорного контроля во время ферментации для регулирования значения рН соответственно в качестве пеногасителя. FS1 получают следующим образом: 750 г глюкозы и 22,2 г MgSO4•7Н2O раздельно растворяют в 600 мл, соответственно, 50 мл воды. Растворы смешивают друг с другом после осуществленного автоклавирования. FS2 готовят путем растворения 227 г (NH4)2HPO4 и 169,5 г (NH4)H2PO4 в воде при добавлении 60 мл 25%-ного раствора аммиака, чтобы перед автоклавированием установить рН-значение = 6,8. FS3 получают из маточных (исходных) растворов в следующей последовательности: 50 мл гидратированного цитрата трехвалентного железа (6 г/л); 0,5 мл Н3ВО3 (30 г/л); 0,5 мл MnCl2•4H2O (10 г/л); 0,5 мл дигидрата этилендиаминтетрауксусной кислоты (84 г/л); 0,5 мл CuCl2•2H2O (15 г/л); 0,5 мл Na2MoO4•2Н2О (25 г/л); 0,5 мл CoCl2•2H2O (25 г/л) и 10 мл Zn(СН3СОО)2•2H2O (4 г/л).
Пример 3
Несколько колоний из чашки Петри, выращенных на агаре среды Луриа при 26oС, служат для пересева в 20 мл жидкой среды Луриа. После встряхивания в течение 5 часов (200 оборотов в минуту, 26oС) 1 мл переносят в 100 мл среды для предкультивирования в колбе емкостью 500 мл и инкубируют далее. 10 мл этой предкультуры используют для пересева в 100 мл новой среды для предкультивирования. Таким образом получают 9 предкультур, которые используют все вместе для пересева в 8 л среды для основного культивирования в ферментере с первоначальной оптической плотностью, составляющей при 550 нм примерно 0,2.
Пример 4
Конструкция биореактора емкостью 10 л с оснасткой и контрольными устройствами представлена на фиг.1. Культивирование в масштабе ферментации в условиях высокой плотности клеток осуществляют с помощью цифрового измерительного и контрольного узла (DCU), мультиферментерной контрольной системы (MFCS) и контрольного устройства для газового потока. Постоянно измеряют выделение диоксида углерода и кислорода. После пересева сборник биопроб МХ-3 (New Brunswick Scientific, Watford, UK) служит для отбора асептических проб и позволяет проводить анализ данных в режиме с разделением времени (Webb и др. , Biotechnol. , 8, 926-928 (1990)). Контрольные устройства поддерживают норму входящего газового потока 10 л/мин, рН-значение равным 6,8, температуру 26oС и давление 0,15 МПа. Две контрольные петли гарантируют аэробные условия роста при рО2-значении 20%. Все важные физические величины измеряют и регистрируют во время всего процесса ферментации.
Во время порционной фазы подпитки концентрация глюкозы в культуре поддерживается равной 1,5 г/л. Для этой цели используют модифицированный анализатор инъекционного ввода потока (анализатор FIAstar 5020 с фотометром и детектирующим контрольным узлом, Tecator АВ, Швеция). Детали этой системы и способ ее функционирования описываются в уровне техники (например, Pfaff и др. , в Руководстве под ред. Munack A.,  К. "Применения компьютера в биотехнологии", Elsevier Science Ltd., Оксфорд, 1995, с.6-11).
К. "Применения компьютера в биотехнологии", Elsevier Science Ltd., Оксфорд, 1995, с.6-11).
Плотность клеток рассчитывают из измерения оптической плотности при 550 нм. Плазмидную стабильность определяют согласно Pack и др., 1993 (см. выше).
Пример 5
Количественное определение синтезированных миниантител осуществляют по методу Pack и др., 1993 (см. выше). Количество способных к функционированию миниантител определяют путем твердофазного иммуноферментного анализа, общее количество миниантител определяют путем электрофореза в 12%-ном полиакриламидном геле с добавлением додецилсульфата натрия по Лэммли (1970) путем сканирования геля. Для твердофазного иммуноферментного анализа титрационные микроплаты покрывают человеческим рецептором эпидермального фактора роста (например, известного из международной заявки 92/15683). Связанные миниантитела детектируют с помощью кроличьей сыворотки против scFv425 и конъюгированного с козьей пероксидазой противокроличьего иммуноглобулина (Jackson Immunoresearch Inc., США). Выход активных миниантител рассчитывают из рядов разбавления очищенных миниантител. При контроле установлено, что кроличья сыворотка против scFv425 не дает никакой фиксируемой перекрестной реакции с другими компонентами не содержащего плазмиду сырого экстракта E. coli RV308. Далее, добавление этого сырого экстракта к ряду разбавления такого же антитела в очищенной форме не оказывает никакого воздействия на сигналы при твердофазном иммуноферментном анализе. Для определения общего количества миниантител фотометрически детектируют гели, окрашенные кумасси-бриллиантовым синим, и концентрацию миниантител рассчитывают из ряда разбавления очищенного миниантитела, отделенного в том же самом геле. В качестве контроля служит аналогичная смесь, в которой используют клетки-хозяева E. coli, не продуцирующие никаких миниантител.
Изобретение относится к биотехнологии, касается порционной ферментации с подпиткой с особой системой вектор-хозяин E.coli для эффективного образования рекомбинантных протеинов, в особенности рекомбинантных молекул антител, предпочтительно фрагментов антител, таких как миниантитела. В данных условиях клетки E.coli растут с максимальной удельной скоростью роста до очень высоких плотностей клеток. Ограничения роста за счет субстратов или метаболических побочных продуктов не происходит. Рекомбинантный вектор содержит терминаторную последовательность в проксимальном и дистальном положении, систему lac-промотор/оператор, последовательность Шайн-Дальгарно T7gl0, сигнальную последовательность ре1В или ompA, VH- и VL-цепи мышиного гуманизированного антитела Маb425. Путем трансформации E.coli RV308 рекомбинантным вектором получают трансформированный штамм (рНКК) для экспрессии чужеродного протеина. Изобретение позволяет получать большие количества рекомбинантных протеинов. 3 с. и 10 з.п. ф-лы, 3 ил., 1 табл.
| PACK P | |||
| et al | |||
| Improved bivalent miniantibodies, with identical avidity as whole antibodies, produced by high cell density fermentation of Escherichia coli | |||
| Bio/Technology, 1993, vol.11, № 11, p.1271-1277 | |||
| KORZ D | |||
| et al | |||
| Simple fed-batch technigue for high cell density cultivation of Escherichia coli | |||
| Journal of Biotechnology, 1995, № 1, vol.39, s.59-65 | |||
| HELLMUTH K | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Journal of Biotechnology, 1994, № 3, vol.32, p.289-298 | |||
| WO 9215683 A, 17.09.1992 | |||
| GERDES K | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| J | |||
| of Biotechnology, 1988, Bd | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |

