Настоящее изобретение относится к улучшенным экспрессионным векторам, содержащим сайт инициации репликации системы ColE1, предназначенным для продуцирования рекомбинантных протеинов и плазмидной ДНК.
Для получения в промышленном масштабе представляющих интерес рекомбинантных протеинов или производства плазмидной ДНК широкое применение находят методы ферментации с использованием генетически модифицированных микроорганизмов (ГМО).
Основная цель оптимизации процесса ферментации заключается в получении экономичным путем максимально большого количества продукта, обладающего высоким качеством. Для достижения этой цели необходимо оптимизировать объемную продуктивность, которая представляет собой количество единиц образовавшегося продукта на единицу объема в единицу времени. Факторами, имеющими большое значение для процесса оптимизации, являются количество биомассы на единицу объема, т.е. количество клеток, способных продуцировать продукт, и количество протеина, которое может продуцировать каждая клетка. В определенной степени продуцирующая способность клетки пропорциональна количеству копий плазмиды (ККП), т.е. количеству плазмид в клетке, несущих ген, кодирующий рекомбинантный протеин. Кроме того, важное значение имеет эффективность системы транскрипции рекомбинантного протеина. В то время как некоторые промоторы являются слабыми и не позволяют в полной мере реализовать потенциальные возможности метаболизма, многие промоторы являются слишком сильными и приводят к сверхэкспрессии рекомбинантного протеина. Поскольку метаболические ресурсы должны распределяться между системами экспрессии рекомбинантного протеина и протеина организма-хозяина, то система экспрессии, которая обладает слишком большой эффективностью, быстро приводит к истощению метаболических ресурсов, что вызывает гибель клетки.
В последние годы использование плазмидной ДНК в области генной терапии послужило основой для развития целой новой отрасли индустрии. Поэтому существует потребность в получении достаточных количеств плазмидной ДНК высокого качества. В процессе получения плазмид рекомбинантный протеин не продуцируется, вместо этого клеточную «фабрику» используют для производства плазмидной ДНК. Для достижения этой цели необходимо обеспечить очень высокие скорости репликации плазмид, причем клетка-хозяин должна выполнять задачи, отличные от продуцирования рекомбинантного протеина.
Для процессов ферментации с использованием бактерий в основном применяют плазмиды ColE1 вследствие того, что при использовании такой системы можно получать большое количество копий плазмиды.
Ранее были подробно описаны плазмиды ColE1 (Chan и др., 1985), было проведено подробное исследование механизма действия сайта инициации репликации ColE1 (Cesareni и др., 1991). Процесс репликации из плазмиды ColE1 начинается с транскрипции предпраймера PHKII, расположенного на расстоянии 555 пар оснований против хода транскрипции относительно сайта инициации репликации, с помощью РНК-полимеразы хозяина (Tomizawa, 1985). // PHKII в процессе удлинения образует складки, имеющие специфическую структуру, и после полимеризации приблизительно 550 нуклеотидов начинает образовывать гибрид с ДНК-матрицей. Транскрипция предпраймера заканчивается гетерогенно и после образования гибрида предпраймер PHKII отщепляют с помощью РНКазы Н, получая активный праймер со свободным 3'-ОН-концом, который является доступным для ДНК-полимеразы I (Tomizawa, 1990; Lin-Chao и Cohen, 1991; Merlin и Polisky, 1995).
Область ColE1 содержит два промотора. PHKI представляет собой антисмысловую молекулу РНК, состоящую из 108 нуклеотидов, которая транскрибируется с помощью второго промотора на противоположной цепи и является комплементарной 5'-концу PHKII. PHKI транскрибируется от положения, находящегося на расстоянии 445 пар оснований против хода транскрипции относительно сайта инициации репликации до приблизительно того локуса, где начинается транскрипция PHKII (Merlin и Polisky, 1995; Tomizawa, 1990).
Для зрения регулирования количества копий в плазмидах ColE1 характеристики кинетики играют более важную роль, чем характеристики равновесия. Например, некоторые мутантные штаммы, имеющие мутации в PHKII, которые хотя и не затрагивают области, комплементарные PHKI, приводят к пониженному ингибированию, обусловленному PHKI. Это, по-видимому, обусловлено воздействием на время полужизни промежуточных структур РНК, уменьшением времени, в течение которого они чувствительны к действию PHKI, что приводит к увеличению количества копий плазмиды. Этот факт подтверждает важность промежуточных структур PHKII и кинетики процесса образования складчатой структуры PHKII (Gultyaev и др., 1995).
Было установлено, что истощение (недостаток) аминокислот приводит к образованию больших количеств тРНК, которые не связаны с определенными аминокислотами (ниже такие тРНК обозначены как "свободные тРНК"). Это явление можно сравнить с описанной выше ситуацией, возникающей после индукции экспрессии рекомбинантного протеина, в том случае, когда метаболические ресурсы исчерпаны.
Wróbel и Wegrzyn (1998) изучали стратегию избирательной индукции истощения пяти различных аминокислот. Было установлено, что существует положительная корреляция между гомологией петель антикодона тРНК, соответствующих истощению определенных аминокислот и определенных петель в структурах PHKI и PHKII. Было высказано предположение о том, что механизм трансляции захватывает большую часть аминоацил-тРНК, однако, в отличие от этого свободные тРНК имеют возможность взаимодействовать с другими молекулами, такими как PHKI и PHKII. Наиболее вероятно, что взаимодействие между тРНК и PHKI или PHKII может влиять на взаимодействие между PHKI и PHKII, приводя к более высокой частоте гибридизации PHKII-ДНК (если предположить, что взаимодействие тРНК с PHKII не изменяет каким-либо существенным образом структуру PHKII). Последнее может приводить к более высокой частоте репликации и тем самым к увеличению ККП.
Zavachev и Ivanov (1988) провели сравнение гомологии в общей сложности 21 тРНК и PHKI/PHKII. Среди них для 11 была выявлена степень гомологии либо с PHKI, либо с PHKII, превышающая 40%. Указанные тРНК разделили на три категории: а) тРНК, гомологичные PHKI: Arg, His, Leu, Lys, Phe и Thr, б) тРНК, гомологичные PHKII: f-Met (формилметионин). Try и Gly, и в) тРНК, гомологичные как PHKI, так и PHKII: Met и Val. Все тРНК имеют петли антикодонов, состоящие из 7 нуклеотидов (Hjalt и Wagner, 1992). В случае тРНК, гомологичной PHKI, наибольшая степень гомологии была выявлена в области петли 2, в то время как наименьшая степень гомологии была выявлена в области 5'-конца PHKI.
Истощение аминокислот и клеточный стресс приводят к увеличению количеств свободных тРНК, которые вступают во взаимодействие с сайтом инициации репликации плазмид ColE1. Это взаимодействие обусловлено гомологией последовательности тРНК с тремя петлеобразными структурами РНК, присутствующими в PHKI и PHKII сайта инициации репликации, и приводит к взаимодействию с контролирующим ККП механизмом системы. В результате этого происходит быстрое увеличение ККП, что приводит к нарушению процесса ферментации.
Для разрешения этих проблем в WO 89/07141 был предложен экспрессионный вектор, имеющий систему репликации ColE1 и имеющий мутацию в гене PHKII и/или в гене rop, созданную с целью усиления экспрессии. Это было достигнуто без существенного увеличения количества копий плазмиды.
Поскольку процесс ферментации с использованием бактерий эффективен только в том случае, когда систему можно поддерживать в течение продолжительного периода времени, и поскольку увеличение количества копий плазмиды является одним из основных факторов, которые приводят к коллапсу системы экспрессии, то задачей изобретения являлась разработка улучшенной системы экспрессии, отличающейся пролонгированным временем жизни (увеличенной жизнеспособностью) бактерий в процессе ферментации.
В частности, задачей изобретения была разработка системы экспрессии, в которой количество копий плазмиды после индукции системы экспрессии ограничивают по сравнению с неконтролируемой амплификацией для того, чтобы поддерживать метаболический груз ниже летальных уровней.
Следующей задачей изобретения было увеличение скоростей репликации плазмид и тем самым выхода плазмидной ДНК в процессах продуцирования плазмид.
Для решения указанных задач при создании изобретения был использован механизм репликации ColE1-типа. Конкретно это выражалось в том, что с помощью метода генной инженерии изменяли, предпочтительно уменьшали степень гомологии сайта инициации репликации ColE1 со свободными тРНК, или полностью ее ликвидировали. В альтернативном варианте создавали случайную библиотеку, предназначенную для отбора плазмид, обладающих измененной способностью к репликации, например, с большим количеством копий плазмид.
Настоящее изобретение относится к экспрессионному вектору, несущему систему репликации ColE1, в котором степень гомологии PHKI и PHKII сайта инициации репликации ColE1 со свободными тРНК модифицируют с помощью одной или нескольких мутаций в кодирующей области гена PHKI и одной или нескольких соответствующих мутаций в гене PHKII, причем указанная(ые) мутация(и) приводят к заменам одной или нескольких пар оснований в петле 1 и/или петле 2 и/или петле 3 PHKI и PHKII.
Понятие «мутация» включает как мутации, которые увеличивают, так и мутации, которые уменьшают степень гомологии PHKI и PHKII со свободными тРНК.
Для максимального сохранения вторичной структуры и температуры плавления PHKI и PHKII для того, чтобы не ухудшать способность к репликации, предпочтительно используют мутации, представляющие собой замены комплементарных оснований, т.е. мутации типа А→Т, Т→А, C→G, G→C. Могут присутствовать также и другая(ие) мутация(и) при условии, что она(и) не ухудшают механизм репликации.
В отличие от вектора ColE1, описанного в WO 89/07141, который имеет мутацию в гене PHKII и следовательно, благодаря ее положению относительно промотора гена PHKI, вектор по изобретению имеет мутацию в кодирующей области, более конкретно в областях петель как гена PHKI, так и гена PHKII, которые гомологичны свободным тРНК. Таким образом, в изобретении предложена новая стратегия, позволяющая направлено изменять степень гомологии и в конечном итоге регулировать скорость репликации плазмид.
В контексте настоящего описания понятие «петля» относится предпочтительно к неспаренным петлевым структурам PHKI или PHKII; однако это понятие не ограничено только областью петли и может включать также нуклеотиды, примыкающие к стволовой области, предпочтительно не более двух нуклеотидов.
Мутация(и) может(гут) представлять собой замену одного основания в петле 1, петле 2 или петле 3, или одну или любое количество замен оснований, включая замену всех оснований, в петле 1 и/или петле 2 и/или петле 3.
Предпочтительно мутация(и) присутствует(ют) в петле 2, которая представляет собой область, имеющую наиболее высокую степень гомологии со свободными тРНК.
Требуемые мутации в петле(ях) гена PHKI и PHKII можно создавать обычными методами мутагенеза и клонирования.
Согласно одному из вариантов осуществления их можно создавать следующим образом: используя ген PHKI или ген PHKII или его фрагмент в качестве матрицы, осуществляют ПЦР с использованием в качестве праймеров двух олигодезоксирибонуклеотидов, один из которых или оба несут требуемую(ые) мутацию(и). Предпочтительно ПЦР представляет собой ПЦР, которую осуществляют в две стадии. На первой стадии амплифицируют два перекрывающихся фрагмента, один из которых имеет требуемую мутацию в последовательности праймера между сайтом рестрикции, предназначенным для соединения фрагментов, и сайтом связывания праймера. Затем амплифицированные фрагменты расщепляют с помощью соответствующей рестриктазы, лигируют и используют в качестве матрицы на следующей стадии ПЦР-амплификации. На этой стадии используют те же праймеры, что и на первой стадии, которые не содержат вновь введенных сайтов рестрикции, выбранных для связывания против хода транскрипции и по ходу транскрипции с ближайшими уникальными сайтами рестрикции конкретной плазмиды.
Благодаря комплементарности генов PHKI и PHKII оба гена, используемых для получения вектора, или их фрагменты равным образом можно применять в качестве матриц при осуществлении ПЦР-амплификации. При наличии комплементарности любая(ые) мутация(и) в одной или нескольких петлях одного гена приводят к созданию соответствующих мутаций в другом, комплементарном гене; в описанном выше предпочтительном методе содержащий мутацию(и) праймер служит не только для удлинения с помощью полимеразы, но также и в качестве матрицы для ДНК-полимеразы, создавая тем самым комплементарную цепь, содержащую мутацию(и). В зависимости от того, какой ген используют в качестве матрицы: ген PHKI или ген PHKII, PHKII или PHKI автоматически будут нести комплементарную мутацию.
Предпочтительно в качестве матрицы используют плазмиду, содержащую полные гены PHKI и PHKII. В альтернативном варианте можно использовать молекулу ДНК, кодирующую полный ген PHKI или PHKII. Гены PHKI и PHKII описаны у Tomizawa и др., 1977.
В том случае, если в качестве матрицы используют фрагмент гена PHKI или PHKII, то фрагмент должен иметь длину, достаточную для того, чтобы включать все необходимые элементы, т.е. последовательность, которая должна быть подвергнута мутации, сайт связывания праймера и необязательно один или несколько сайтов рестрикции. Предпочтительно фрагмент содержит одну или несколько петель (каждая из которых состоит приблизительно из 7 нуклеотидов) и сайт связывания праймера (приблизительно 18 нуклеотидов), т.е. минимальный размер пригодного фрагмента составляет приблизительно 25-30 нуклеотидов.
Согласно предпочтительному варианту осуществления изобретения, мутации с точки зрения их расположения и количества выбирают таким образом, чтобы обеспечить существенное изменение степени гомологии с максимально возможным количеством видов свободных тРНК.
Так, в одном из вариантов осуществления изобретения модификацию PHKI и PHKII осуществляют, начиная с модификации петли 2, т.е. петли, обладающей наиболее высокой степенью гомологии, причем указанная модификация заключается в осуществлении замен в максимально возможном количестве положений. Например, как это продемонстрировано в эксперименте, описанном в примере 1, петлю 2 можно модифицировать путем замены шести из ее семи нуклеотидов, сохраняя без изменения одно основание (положение 693 в плазмиде ColE1, Genbank GI 9507253), которое тем самым может служить в качестве части нового сайта рестрикции, предпочтительно сайта Ncol. Основания заменяют соответствующими комплементарными им основаниями.
При таком подходе можно при необходимости полностью ликвидировать гомологию со всеми свободными тРНК. Поэтому данный подход обеспечивает максимальную степень свободы при производстве широкого разнообразия представляющих интерес рекомбинантных протеинов независимо от того, какую аминокислотную последовательность они имеют, в частности, благодаря регулированию репликации плазмид, обеспечивающему поддержание жизнеспособности клеток при экспрессии. В этом случае амплификация плазмид регулируется только специфическим для ColE1 механизмом репликации и не зависит от флуктуации метаболизма в клетке-хозяине; при этом количество копий плазмиды остается практически постоянным в процессе ферментации.
В отличие от варианта, предусматривающего полную ликвидацию гомологии между PHKI и PHKII и свободными тРНК, указанную гомологию можно модифицировать, т.е. увеличивать или уменьшать лишь в определенной требуемой степени. Для некоторых приложений, например, в том случае, когда выход продукта оказывается недостаточным из-за того, что потенциал механизма экспрессии не реализуется в полной мере вследствие уменьшения количества копий плазмид и соответственно наличия субоптимального количества плазмид ("дозы гена"), может оказаться целесообразным повышать скорости экспрессии за счет небольшого увеличения количества копий плазмид. Это можно обеспечивать путем избирательного сохранения гомологии последовательности с определенными свободными тРНК, в частности, с редко встречающимися тРНК. В литературе описаны последовательности, гомологичные определенным свободным тРНК (Zavachev и Ivanov, 1988). Эту стратегию можно использовать также для воздействия на соотношение синтезируемых форм протеина, так что протеин может присутствовать либо в форме внутриклеточных телец или находиться в растворимой форме. Например, при определенной степени амплификации могут образовываться преимущественно внутриклеточные тельца, в то время как небольшое ее уменьшение может стимулировать образование растворимого продукта.
В некоторых случаях предпочтительно сильно увеличивать количество копий плазмид путем повышения степени гомологии последовательностей PHKI и PHKII со свободными тРНК, в частности для получения плазмидной ДНК. Мутации, необходимые для повышения степени гомологии последовательностей, также описаны в литературе (Zavachev and Ivanov, 1988) и их можно создавать на основе тех же принципов, которые описаны для уменьшения степени гомологии последовательностей.
Для конкретного случая и/или получения конкретного продукта способ можно оптимизировать путем экспериментального исследования различных мутаций. Соответствующие эксперименты можно проводить следующим образом: предназначенной(ым) для тестирования плазмидой-«кандидатом» или набором плазмид-«кандидатов», несущих мутацию(и), трансфектируют соответствующие бактериальные клетки-хозяева, выращивают в соответствующих лабораторных условиях, например, во встряхиваемых колбах, и наблюдают за процессом ферментации, регистрируя представляющие интерес параметры, в частности, рост, выход продукта и его качество, количество копий плазмид.
Согласно другому варианту осуществления, который наиболее предпочтителен для получения широкого разнообразия модификаций последовательностей, осуществляют случайным образом мутацию в одном или нескольких положениях петли 1 и/или петли 2 и/или петли 3, создавая тем самым библиотеку, которую можно использовать для конструирования экспрессионного вектора, предназначенного для реализации любого требуемого свойства экспрессионной системы. Например, плазмиду-«кандидата» можно выбирать с учетом селекционных параметров, которые наиболее важны для производства рекомбинантного протеина, таких, например, как скорость роста, продуктивность и жизнеспособность; при этом процесс осуществляют с использованием описанного выше стандартного экспериментального метода.
Кроме того, данный подход позволяет эффективно регулировать скорость экспрессии рекомбинантного протеина путем манипуляций с ККП. В том случае, если представляющий интерес ген присутствует в векторе, содержащем библиотеку, отобранная плазмида всегда будет оптимальной с точки зрения экспрессии представляющего интерес гена.
В то время, как в процессе обычной ферментации после индукции наблюдается почти десятикратное увеличение количества копий плазмид (ККП), использование способа по настоящему изобретению позволяет уменьшить или ликвидировать гомологию между тРНК и PHKII. Это приводит к увеличению доли неингибированных молекул PHKI, которые не взаимодействуют с PHKII. Таким образом, механизм репликации перестает быть связанным с большими уровнями свободных тРНК, возникающими при избыточном метаболизме вследствие экспрессии рекомбинантного протеина. Отделение механизма репликации плазмид от метаболического стресса, вызываемого экспрессией рекомбинантного протеина, позволяет получать более высокий выход рекомбинантного протеина.
Поскольку замены оснований, осуществляемые согласно настоящему изобретению, присутствуют как в PHKI, так и в PHKII, что приводит к изменению гомологии со всеми тРНК, то состав свободных тРНК (зависящий от рекомбинантного продукта) не имеет решающего значения для данного подхода.
Кроме модифицированной системы репликации ColE1 экспрессионный вектор по изобретению содержит элементы, необходимые для экспрессии протеина, т.е. контролирующие экспрессию последовательности, функционально связанные с последовательностью кДНК, кодирующей представляющий интерес протеин, включая промотор, область инициации трансляции, селектируемые маркеры (например, маркеры устойчивости к антибиотику), сайты рестрикции для встраивания ДНК, кодирующей представляющий интерес протеин, и т.д.
Предпочтительно экспрессионный вектор по изобретению получают на основе следующих векторов:
pMBl (Bolivar и др., 1977);
pBR322 (Covarrubias и др., 1981; может быть получен от MBI Fermentas по каталожному номеру №SD0041; GenBank/EMBL, добавочные номера последовательностей J01749, К00005, L08654, М10282, М10283, М10286, М10356, М10784, М10785, М10786, М33694, V01119);
pUC18 (Yanisch-Perron и др., 1985; GenBank/EMBL, добавочный номер последовательности L09137; может быть получен от MBI Fermentas по каталожному номеру №SD0061);
pUC19 (GenBank/EMBL, добавочный номер последовательности L09136, может быть получен от MBI Fermentas по каталожному номеру №SD0051);
pTZ19R (GenBank/EMBL, добавочный номер последовательности Y14835; может быть получен от MBI Fermentas по каталожному номеру №SD0141);
pTZ19U (может быть получен от MBI Fermentas по каталожному номеру №SD0161; GenBank/EMBL, добавочный номер последовательности Y14835);
pBluescriptII KS (-) (Alting-Mees и др., 1989; GenBank/EMBL, добавочный номер последовательности Х52329);
pBluescriptII KS (+) (Alting-Mees и др., 1989; GenBank/EMBL, добавочный номер последовательности Х52327);
pBluescriptII SK (-) (Alting-Mees и др., 1989; GenBank/EMBL, добавочный номер последовательности Х52330;
pBluescriptII SK (+) (Alting-Mees и др., 1989; GenBank/EMBL, добавочный номер последовательности Х52328).
С точки зрения представляющего интерес протеина на последовательность не накладывается никаких ограничений, если экспрессия плазмиды в Е. coli приводит к получению обладающего функциональной активностью протеина.
В экспериментах, проведенных при создании настоящего изобретения, использовали кДНК, кодирующую человеческую Cu-Zn-супероксид-дисмутазу. С помощью вектора, несущего эту кДНК, можно получать высокорастворимый димерный протеин, имеющий молекулярную массу 32 кДа, который состоит из 153 аминокислот и высвобождается в цитоплазму (Cserjan-Puschmann и др., 1999).
Можно использовать любую бактериальную клетку-хозяина, совместимую с плазмидами типа ColE1, предпочтительно штаммы Е. coli, в частности штамм HMS 174 (DE3) (Studier и Moffat, 1986) или штаммы Salmonella.
Другой вариант осуществления настоящего изобретения относится к клетке-хозяину, трансформированной экспрессионным вектором, несущим модифицированный сайт инициации репликации ColE1.
Для трансформации штамма-хозяина можно применять любой обычный метод, например, электропорацию или осаждение хлоридом кальция или кальцием.
Следующим объектом настоящего изобретения является способ получения представляющего интерес рекомбинантного протеина, заключающийся в том, что клетку-хозяина Е. coli трансформируют экспрессионным вектором, который имеет систему репликации ColE1, содержащую мутации в одной или нескольких петлях гена PHKI и гена PHKII, выращивают в соответствующих условиях и выделяют представляющий интерес протеин. Изобретение позволяет ускорять процесс продуцирования рекомбинантного протеина, обеспечивая механизм компенсации взаимодействия экспрессии рекомбинантного протеина и метаболизма, происходящего в клетке-хозяине. Способ по изобретению наиболее предпочтительно следует применять с использованием периодических процессов ферментации с подпиткой, т.е. процессов, в которых добавление питательных веществ связано с увеличением биомассы. Для полной реализации преимуществ периодических процессов с подпиткой, которые могут протекать в течение продолжительных периодов времени и тем самым позволяют получать более высокие выходы биомассы при большей экономичности по сравнению с обычными периодическими процессами, необходимо располагать стабильной и регулируемой системой экспрессии. Данная задача может быть успешно решена с использованием экспрессионного вектора по изобретению.
Кроме того, поскольку модификации последовательности одной или нескольких петель PHKI и PHKI ColE1 могут приводить к сильному увеличению скоростей репликации плазмид, то векторы можно успешно применять для продуцирования плазмид, например, с целью использования в генной терапии. Преимущества настоящего изобретения заключаются в возможности уменьшения стабильности так называемого «целующегося комплекса» ("kissing complex") PHKI/PHKII и тем самым повышения скоростей репликации плазмид.
Пример 1
В экспериментах использовали плазмиду pET11a (являющуюся производным плазмиды pUC19 фирмы Stratagene). Эта плазмида содержит ген бета-лактамазы, обеспечивающий устойчивость к ампициллину. Экспрессия рекомбинантного протеина в этой плазмиде контролируется эффективной РНК-полимеразой фага Т7. Оператор lac расположен между последовательностями промотора фага Т7 и сайта инициации трансляции. Это приводит к подавлению экспрессии в отсутствии индуктора ИПТГ (изопропилтиогалактозид). Плазмида pET11a-SOD несет ген кДНК, кодирующей рекомбинантный человеческий протеин Cu-Zn-супероксид дисмутазу (чСОД), представляющий собой высокорастворимый димерный протеин с молекулярной массой 32 кДа, состоящий из 153 аминокислот, который не является токсичным для клетки и высвобождается в цитоплазму (Cserjan-Puschmann и др., 1999).
Для размножения плазмиды и экспрессии СОД использовали штамм бактерии Escherichia coli HMS174 (DE3) (Studier и Moffat, 1986). Этот штамм содержит полимеразу фага, интегрированную в хромосомную ДНК. Полимераза фага Т7 имеет важное значение для экспрессии рекомбинантного протеина. Трансформанты отбирали на пластинах, содержащих ампициллин (содержащая антибиотик среда LB, 100 пкг/млампициллина) (Maniatis, и др., 1982).
Трансформацию в рассматриваемых экспериментах проводили методом электропорации с использованием устройства типа Bio-Rad Gene Pulser. Праймеры получали от фирмы Metabion (Мартинсрид, Германия) в форме полученного с помощью вакуумной сушки порошка, который растворяли в воде для получения маточных растворов с концентрацией 100 пмолей/мкл. ПЦР осуществляли в термоячейке (Т-градиент, фирма Biometra, Германия) с использованием покрытия из термостойкой полимеразы Dynazyme EXT в концентрации 1 ед./мкл (Finnzymes), 10-кратного не содержащего магния буфера с добавлением 10 мМ MgCl (поступает в продажу), 1 мМ дНТФ, ДМСО и дистиллированной воды.
Использовали праймеры pET11a-114back (SEQ ID NO:1), pEZ11a656for (SEQ ID NO:2), PHKI-Ncoback (SEQ ID NO:3) и PHKI-Ncofor (SEQ ID NO:4). Рестриктазы, лямбда-маркеры, лигазу фага Т4, фосфатазу из кишечника теленка получали от фирмы MBI-Fermentas и применяли согласно рекомендациям этой фирмы.
Для ферментации использовали ферментер вместимостью 20 л фирмы MBR Bioreactor AG (Ветцикон, Швейцария) с присоединенным к нему контроллером типа MBR IMCS-2000. Рабочий объем ферментера составлял приблизительно 12 л.
Используемая питательная среда: количество питательной среды, закачиваемой в систему во время стадии периодической подпитки, непрерывно измеряли путем взвешивания сосуда. Насос для подпитки регулировали так, чтобы он обеспечивал постоянную скорость роста μ=0,1. Добавление антивспенивателя начинали по сигналу датчика проводимости. Отсутствие контакта с окружающей средой позволяло устранить риск загрязнения ферментера. В качестве загружаемой среды использовали полусинтетическую среду, содержащую небольшие количества триптона и дрожжевого экстракта для облегчения роста после начала загрузки. Компоненты смешивали в общем объеме, составляющем приблизительно 4 л (4000 г). Для того, чтобы избежать появления осадка, химические вещества с одинаковым номером (см. номера, приведенные ниже в таблице 1) сначала по отдельности растворяли в дистиллированной воде. Раствор глюкозы дополняли дистиллированной водой до 300 г и автоклавировали отдельно. Затем все компоненты за исключением раствора глюкозы смешивали друг с другом в указанном порядке и дополняли дистиллированной водой до 3700 г.
Таблица 2:
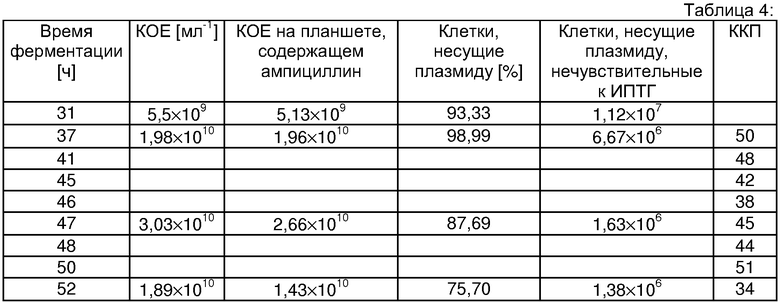
Для определения доли бактериальных клеток, несущих плазмиды, и определения того, могут ли расти клетки, несущие плазмиду, на пластинах, содержащих индуктор ИПТГ, что показывает, могут ли несущие плазмиду клетки после индукции продуцировать СОД в "обычных" количествах, осуществляли тест Коха.
Общее количество сухого вещества определяется сухой массой бактерий (СМБ).
Для каждого образца стеклянный стакан сушили в течение ночи при 105°С, охлаждали в эксикаторе и затем взвешивали на аналитических весах.
ККП можно рассчитывать на основе корреляции размеров (количества пар оснований) геномной ДНК и плазмидной ДНК.
Для получения плазмидной ДНК клеточный дебрис, полученный из препарата образца, ресуспендировали в 150 мкл раствора I (50 мМ глюкоза, 10 мМ ЭДТК, 25 мМ Трис-HCl, рН 8,0). Добавляли 200 мкл ДСН (0,5%-ный раствор ДСН (фирма ICN Biochemicals), добавляли 50 мкл лизоцима (фирма Sigma), препараты смешивали и инкубировали в течение 10 мин при 37°С, раствор гомогенизировали путем вращения. Образцы хранили на льду до проведения измерений с помощью спектрофлуорометра (типа Hitachi F-2000).
Для определения количества плазмидной ДНК проводили очистку ДНК, содержащейся в клеточном дебрисе, с помощью набора типа GFX (фирма MBI, Fermentas) согласно инструкциям поставщика со следующими модификациями: после стадии лизиса добавляли определенное количество (˜2 мкг) pUC19 в качестве внутреннего стандарта. После элюирования ДНК в 50 мкл воды осуществляли линеаризацию плазмиды с помощью Hind III в течение 1 ч при 37°С. Для осуществления капиллярного электрофореза образец, содержащий расщепленную рестриктазой ДНК, переносили в сосуд для образца, избегая появления воздушных пузырьков. Образцы загружали в автосэмплер. Калибровку капилляра проводили, промывая буфером в течение 15-20 мин. Измерение абсорбции проводили при 260 нм и 280 нм с помощью диодной матрицы. После анализа капилляр промывали буфером и хранили при 4°С. Количество хромосомной ДНК рассчитывали путем вычитания количества плазмидной ДНК/мг СМБ из общего количества ДНК/мг СМБ. Поскольку количество добавленного внутреннего стандарта известно, то ККП можно рассчитывать согласно методу, описанному у Breuer и др. (1998), по следующим формулам:

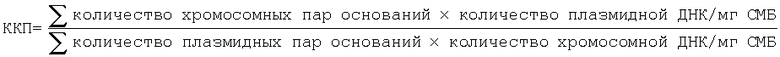
Для определения количества СОД обладающие способностью к связыванию антитела (моноклональное антитело к СОД, клон 30F11, который получали от фирмы Novocastra Laboratories Ltd., Великобритания) разводили в соотношении 1:100 буфером для сенсибилизации поверхностей (200 мкг/мл). 100 мкл разведенного раствора антитела вносили в каждую лунку титрационного микропланшета, инкубировали при 4°С в течение ночи или при комнатной температуре в течение по меньшей мере 2 ч. Планшет трижды промывали буфером для отмывки, после чего буфер удаляли путем осторожного встряхивания планшета. Образец и стандарт в планшетах для разведения разводили последовательно в соотношении 1:2 с помощью автоматического устройства для пипетирования. По 50 мкл каждого индивидуального разведения вносили (с помощью автоматического устройства) в сенсибилизированный антителом планшет и инкубировали в течение 1 ч при комнатной температуре.
Планшет промывали буфером для отмывки. Конъюгированное антитело разводили в соотношении 1:500 буфером для разведения (Porstmann и др., 1988).
В таблице 3 представлены мутации в сайте инициации репликации (в таблице перечислены положения, в которых произведены замены, номера соответствуют полной последовательности ColE1, хранящейся под номером GI="9507253" в Genbank):
Процесс ферментации с использованием плазмиды pET11achSOD в штамме Е. coli HMS174 (DE3) проиллюстрирован на чертеже. Представлены данные об общем и удельном производстве рекомбинантного протеина (СОД), а также о скорости производства, qp. Приведены также величины общей сухой массы бактерий (СМБ) и количества копий плазмиды (ККП). В отличие от стандартных процессов (например, описанных у Cserjan-Puschmann, 1999) установлено, что ККП остается практически постоянным даже после индукции, осуществляемой на 45-м часу ферментации.
Результаты исследований, полученных с использованием теста Коха, и количества копий плазмиды представлены в таблице 4
Пример 2
В экспериментах использовали плазмиду pET11a-SOD, описанную в примере 1.
Для размножения плазмиды и экспрессии СОД использовали штамм бактерии Escherichia coli HMS174 (DE3), описанный в примере 1. Все манипуляции с бактериями и плазмидной ДНК осуществляли согласно методу, описанному в примере 1. Олигонуклеотиды и ферменты получали из тех же источников, которые описаны в примере 1.
Использовали праймеры pet11a-Sca-I-for (SEQ ID NO:5), pet11a-AlwN-I (SEQ ID NO:6), pet11a-Xba-I-back (SEQ ID NO:7) и PHK-I-randomXba-I-back (SEQ ID NO:8).
Для отбора наиболее перспективных «кандидатов» из пула различных клонов применяли два подхода:
1. Отбор клеток, содержащих плазмиды с наибольшим количеством копии.
Клетки с большим ККП должны обладать повышенной устойчивостью к ампициллину. Пул бактерий высевали на чашку Петри с LB-агаром, содержащим либо 0,1, либо 10 мг/мл ампициллина. Отбирали 10 колоний из планшета с LB-средой, содержащей 10 мг/мл ампициллина, и проводили анализ последовательности. В результате было выявлено 7 клонов (см. таблицу 5), последовательности которых отличались от последовательностей петли 2 PHKI и PHKII.
Вторым критерием отбора была стабильность плазмид. Бактерии, содержащие пул плазмид, культивировали во встряхиваемых колбах при 37°С до достижения оптической плотности ОП=2 с использованием синтетической среды, не содержащей ампициллин. После осуществления трех пересевов (что соответствует приблизительно 20 поколениям) бактерии, полученные после последнего пересева, вносили в планшет с LB-средой, содержащей ампициллин, для отбора тех бактерий, которые еще содержали плазмиду, при этом одиночные колонии отбирали для анализа последовательности.
Результат этого отбора представлен в таблице 6:
При осуществлении этого отбора применяли тот же ферментер и устройства, которые были описаны в примере 1. Использовали ту же питательную среду и скорость роста, которые указаны в примере 1.
ККП определяли и рассчитывали согласно методу, описанному в примере 1 Характеристики клонов-«кандидатов» с точки зрения влияния на их ККПТ оценивали в опытах с использованием периодического процесса культивирования с подпиткой. Результаты для перспективных «кандидатов» представлены в таблице 7; ККП различных клонов представлены для неиндуцированного и индуцированного состояния.
Клон ColE1Mut9 (и ColE1Mut1, который имеет ту же самую последовательность) оказался очень перспективным «кандидатом» в отношении получения плазмид. Количество копий плазмиды, равное 750, приблизительно в 14 раз превышает соответствующую величину, характерную для плазмиды ColE1 дикого типа.
ККП для клона ColE1Mut22 также оказалось существенно более высоким (в 2,5 раза).
Более низкие ККП для ColE1Mut54 могут быть благоприятными с точки зрения производства рекомбинантного протеина благодаря меньшей метаболической нагрузке.
Изобретение относится к биотехнологии. Предложен экспрессионный вектор, содержащий систему репликации ColE1, в котором гомологию PHKI и PHKII сайта инициации репликации ColE1 со свободными тРНК модифицируют посредством одной или нескольких мутаций в кодирующей области гена PHKI или PHKII. Мутации приводят к замене одного или нескольких оснований в петле 1, 2, 3, PHKI и/или PHKII. Описана бактериальная клетка, трансформированная указанным вектором, и способ получения представляющего интерес протеина из указанной бактериальной клетки, включающий выращивание клетки, выделение белка и его очистку. Использование изобретения обеспечивает регулирование количества копий плазмиды. Повышение степени гомологии PHKI или PHKII со свободными тРНК приводит к увеличению количества копий плазмиды. Уменьшение степени гомологии PHKI или PHKII со свободными тРНК приводит к уменьшению метаболической нагрузки клетки, что благоприятно для получения рекомбинантного протеина. 3 н. и 9 з.п. ф-лы, 1 ил., 7 табл.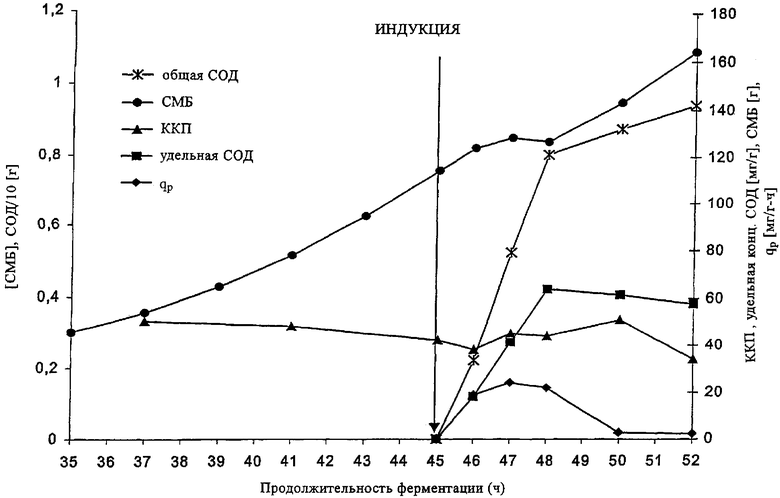
| WO 8907141 А, 10.08.1989 | |||
| RAZMIC S | |||
| et al | |||
| Determinants of RNA hairpin loop-loop complex stability | |||
| - Journal of molecular biology, vol.248, no.5, 1995, с.968-984 | |||
| WROBEL et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

