Настоящее изобретение относится к способу получения проинсулина человека, в частности к способу получения проинсулина человека, выраженного нижеследующей химической формулой (I):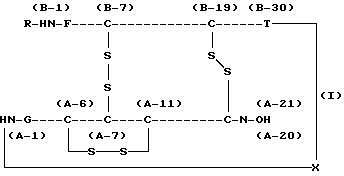
где R обозначает аминокислотный остаток или пептид, который может быть подвергнут ферментативному или химическому гидролизу, и
X обозначает связь между аминокислотной группой А-1 в цепи А инсулина и карбоксильной группой В-30 в цепи В инсулина, которая может быть отделена от цепи А или от цепи В ферментативным или химическим путем, при условии, что область от А-1 до А-21 является цепью А инсулина и область от В-1 до В-30 является цепью В инсулина.
В химической формуле (I) показано, что цепь А и цепь В имеют 3 дисульфидные связи (между А-6 и А-11, между А-7 и В-7 и между А-20 и В-19), образованные 6 остатками цистеина.
Описание предшествующего уровня техники
Предшественник инсулина человека ("проинсулин") обычно получают в процессе производства зрелого инсулина ("инсулин") методом рекомбинантных ДНК, который включает стадию введения структурного гена в плазмидную ДНК клеток Е. coli.
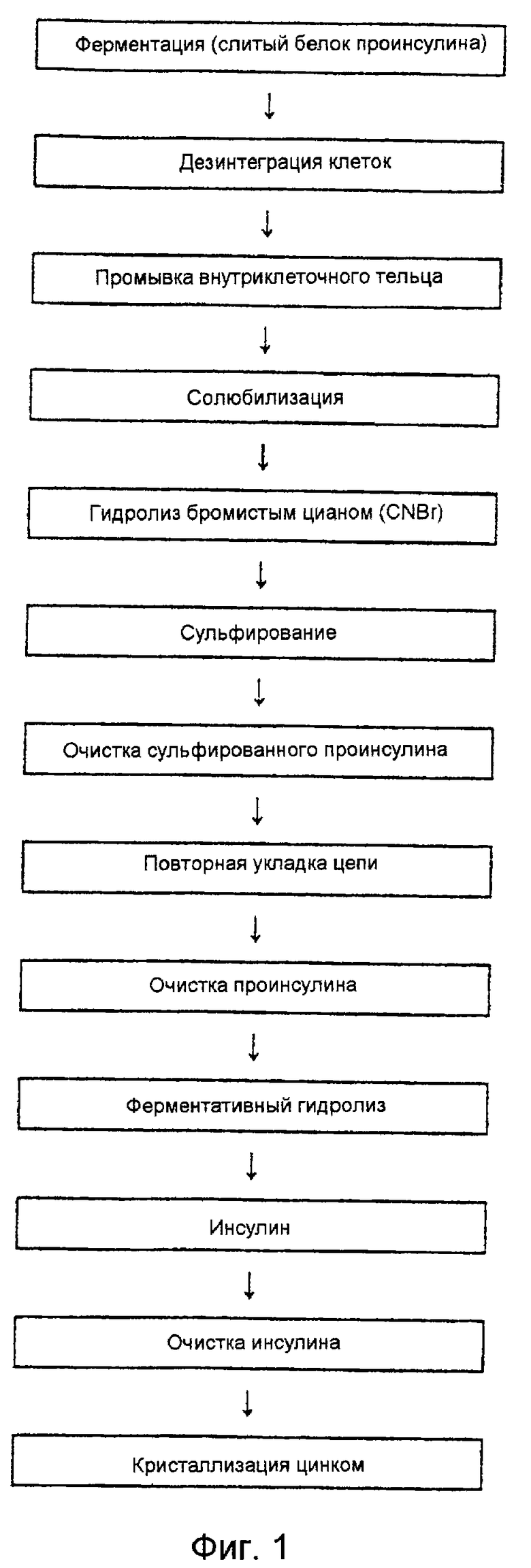
Как показано на фиг. 1, слитый белок, содержащий проинсулин, экспрессирован в виде внутриклеточных телец в клетках Е. coli; указанные внутриклеточные тельца получают центрифугированием после лизиса клеток и промывают неионным или ионным детергентом или денатурантом с низкой концентрацией. Такую обработку, сопровождаемую центрифугированием, повторяют, чтобы повысить чистоту целевого белка (см. Mukhopadhyay, A. et al. Advances in Biochemical Engineering/Biotechnology, 56, 61-108, 1997). Чтобы свести до минимума межмолекулярное гидрофобное взаимодействие и образование неправильных дисульфидных связей, промытые внутриклеточные тельца растворяют в денатуранте, таком как мочевина или раствор гуанидина•НСl, содержащем восстановитель, такой как дитиотрейтол (DTT) или 2-меркаптоэтанол, или в NaOH (см. Fischer et al., Biotechnology & Bioengineering, 41, 3-13, 1993). Растворенные внутриклеточные тельца центрифугируют с высокой скоростью, и супернатант разводят холодной водой, чтобы выделить внутриклеточные тельца в виде осадка (см. европейский патент 0055945 А2). Полученные таким образом внутриклеточные тельца содержат слитый белок проинсулина и гетеропротеин, такой как β-галактозидаза, которые связаны поперечной связью остатка метионина. Слитый белок обрабатывают бромистым цианом (CNBr) и шесть (6) групп -SH в проинсулине замещают группами -SSО3 -, что дает S-сульфонат проинсулина. Стадия сульфирования увеличивает устойчивость предшественника инсулина и эффективность последующей реакции повторной укладки цепи (см. европейский патент 0055945 А2). S-сульфонат проинсулина подвергают повторной укладке цепи для получения нативной конформации, используя восстановители, такие как 2-меркаптоэтанол, DTT и другие, или окислительно-восстановительную систему, такую как глутатион (см. Fischer et al.. Biotechnology & Bioengineering, 41, 3-13, 1993). Полученный таким образом нативный проинсулин превращают в биологически активный инсулин, для чего удаляют Х (или цепь С), связывающую цепи А и В, и производят обработку трипсином и карбоксипептидазой В (см. Kemmler W. , et al., J.B.C., 246, 6786-6790, 1971). И наконец, инсулин очищают высокоэффективной жидкостной хроматографией с обращенной фазой (RP-HPLC) и другими методами (см. Kroeff, Е. Р. , et al., J. Chromatogr., 461, 45-61, 1989) и кристаллизуют по методу Zn-кристаллизации (см. Mirsky, et al., J. Clinical Investigation, 42, 1869-1872, 1963; BraderM.L., TIBS, 16, 341-345, 1991).
Однако известный способ получения проинсулина или инсулина является недостаточно приемлемым, так как он включает сложные стадии растворения и сульфирования, очистки, концентрирования и обессоливания; и при его осуществлении используется неэффективная реакция повторной укладки цепи, которая уменьшает выход целевого белка. Поэтому существуют все основания для разработки и создания усовершенствованного способа получения проинсулина или инсулина более простым и эффективным образом при сохранении его биологической активности.
Краткое изложение существа изобретения
Авторы настоящего изобретения поставили цель устранить недостатки, присущие известным способам получения рекомбинантного проинсулина человека, экспрессированного в виде внутриклеточного тельца, и успешно создали способ получения проинсулина человека, в котором стадии растворения и сульфирования, очистки, концентрированна и обессоливания значительно упрощены при увеличении эффективности реакции повторной укладки цепи.
Главной целью настоящего изобретения является создание простого способа получения рекомбинантного проинсулина человека с хорошей воспроизводимостью.
Краткое описание чертежей
Вышеуказанные и другие цели и отличительные особенности настоящего изобретения станут очевидными из нижеследующего описания и прилагаемых чертежей, где:
На фиг. 1 дано схематическое изображение известного способа получения инсулина человека из предшественника слитого инсулина человека, экспрессированного в рекомбинантных клетках Е. coli.
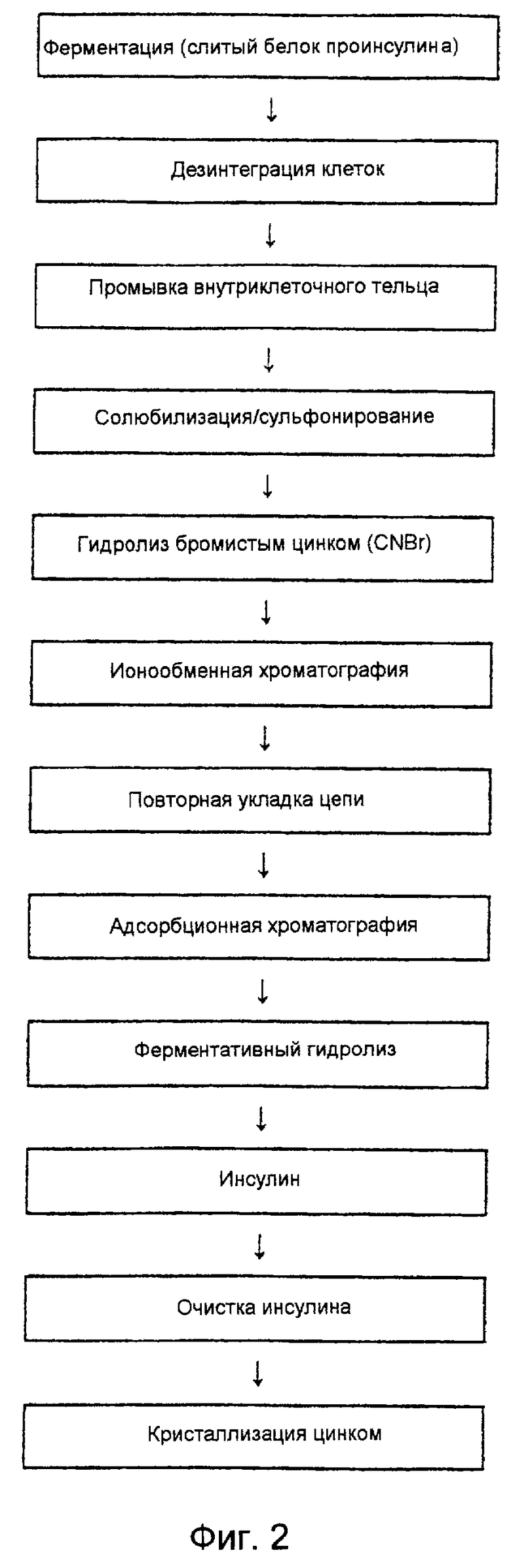
На фиг. 2 дано схематическое изображение способа получения инсулина человека из предшественника слитого инсулина человека, экспрессированного в рекомбинантных клетках Е. coli, по настоящему изобретению.
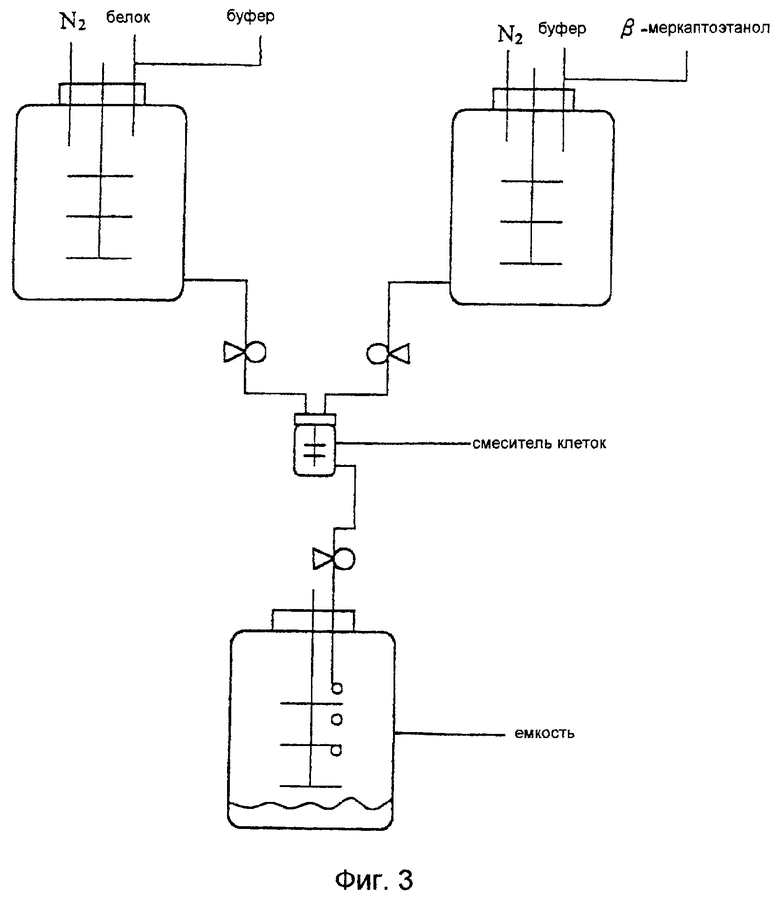
На фиг. 3 показана система повторной укладки цепи, применяемая в способе получения проинсулина человека по этому изобретению.
Подробное описание изобретения
В соответствии со способом получения проинсулина человека по настоящему изобретению предшественник инсулина человека, экспрессированный в виде внутриклеточного тельца, обрабатывают тетратионатом натрия (Na2S4O4) и сульфитом натрия (Na2SO3) на стадии растворения внутриклеточного тельца в мочевине или растворе гуанидина•НСl, в результате чего происходит замещение групп -SH в остатках цистеина предшественника инсулина группами -SSO3 -, что дает S-сульфонат проинсулина, выраженный нижеследующей химической формулой (II), который превращают в проинсулин химической формулы (I), подвергая S-сульфонат проинсулина взаимодействию с 2-меркаптоэтанолом в водной среде.
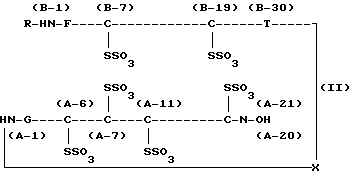
где R и Х аналогичны описанным выше.
Способ получения проинсулина человека по этому изобретению более подробно описан со ссылкой на фиг. 2 и 3. В этом способе получения проинсулина человека все стадии предпочтительно осуществляют при низкой температуре около 4oС, хотя для практического удобства их можно выполнять при комнатной температуре.
Стадия 1. Очистка внутриклеточных телец
Для получения рекомбинантного инсулина человека авторы настоящего изобретения использовали слитый белок модифицированной β-галактозидазы и проинсулина, экспрессированного в клетках Е. coli (см. корейскую патентную публикацию 94-1855). Клетки Е. coli, экспрессирующие слитый белок в виде внутриклеточного тельца, суспендируют в буферном растворе для лизиса в отношении 1: 5-10 (в отношении веса к объему) и лизируют под давлением около 630 атм (9000 фунтов на кв. дюйм). Внутриклеточные тельца центрифугируют, промывают тритоном Х-100 и дистиллированной водой и вновь центрифугируют, чтобы получить очищенные внутриклеточные тельца.
Стадия 2. Растворение и сульфирование
Очищенные внутриклеточные тельца растворяют в 0,02-0,1 М растворе трис-буфера (рН 8-10), содержащего денатурант в количестве 6-8 М мочевины или гуанидина•НСl в отношении 1:10-20 (в отношении веса к объему), более предпочтительно 1: 5-10 (в отношении веса к объему), и добавляют 0,1-0,6 М, более предпочтительно 0,2-0,5 М сульфита натрия (Nа2SО3) и 0,01-0,1 М, более предпочтительно 0,05-0,1 М тетратионата натрия (Na2S4O4). Затем смешанный раствор, содержащий внутриклеточные тельца, перемешивают, чтобы вызвать сульфирование предшественника инсулина, в результате которого происходит замещение групп -SH в предшественнике инсулина группами -SSО3 -. На этой стадии показатель рН и температуру поддерживают соответственно в пределах рН 7,0-9,5 и 4-8oС. И наконец получают слитый белок сульфированного проинсулина, в котором группы -SH в остатках цистеина проинсулина замещены группами -SSО3 -.
Стадия 3. Обработка бромистым цианом
После выполнения реакции сульфирования реакционную смесь центрифугируют со скоростью 12000 об/мин в течение 30 минут, чтобы удалить осадок. К супернатанту добавляют холодную воду в отношении 5-20:1 (в объемном отношении) и показатель рН доводят до 5-6, в результате чего образуется осадок. Осажденный белок растворяют в 70%-ной (в объемном отношении) муравьиной кислоте до концентрации 10-30 мг/мл и затем обрабатывают бромистым цианом так, чтобы молярное отношение бромистого циана к белку было равно 100:1. Это вызывает отделение S-сульфоната проинсулина от слитого белка. Полученный продукт сушат при пониженном давлении.
Стадия 4. Ионообменная хроматография
3-сульфонат проинсулина растворяют в 20 мМ трис-буфера (рН 8,0), содержащего 1 мМ EDTA и 7 М мочевины, до концентрации 30 мг/мл и вводят в колонку со смолой DEAE-сефацел, уравновешенную тем же буфером. Колонку элюируют градиентом концентрации 0-1 М NaCl, в результате чего получают S-сульфонат проинсулина в диапазоне концентраций 0,3-0,5 М NaCl.
Стадия 5. Повторная укладка цепи (превращение S-сульфоната проинсулина в проинсулин)
Очищенный S-сульфонат проинсулина разводят 50 мМ раствором глицинового буфера (рН 10,6), содержащего 1 М мочевины с концентрацией 1-10 мг/мл, без обессоливания или предварительной обработки. Камеру продувают газообразным азотом, чтобы удалить кислород, и герметично закрывают. В другой камере 2-меркаптоэтанол добавляют к 50 мМ раствору глицинового буфера (рН 10,6), содержащему 1 М мочевины в эквивалентном отношении 1-3 в зависимости от количества групп -SSО3 - S-сульфоната проинсулина. Растворы белка и буфера, содержащего 2-меркаптоэтанол, быстро смешивают в отношении 1:1 (в объемном отношении), для чего обе камеры присоединяют к смесителю объемом от 0,1 мл до 10 л, после чего реакционную смесь для повторной укладки цепи, вводят, медленно перемешивая, в расположенную ниже емкость и подвергают взаимодействию в течение 15-20 часов при 4-5oС (см. фиг.3). На стадии повторной укладки цепи по крайней мере 80% S-сульфоната проинсулина можно превратить в нативный проинсулин.
Стадия 6. Адсорбционная хроматография
Для очистки и концентрирования проинсулина с повторно уложенной цепью реакционную смесь для повторной укладки цепи, содержащую проинсулин с повторно уложенной цепью, вводят в соприкосновение с полярной метакрилатной смолой HP-2MG, регулируя показатель рН этой реакционной смеси в пределах от 3 до 4, так чтобы 8 г смешанных белков, содержащих проинсулин с повторно уложенной цепью, могли соприкасаться с 1 литром смолы. Концентрацию белка в загруженном образце регулируют в диапазоне от 0,1 до 5 мг/мл в зависимости от хода реакции повторной укладки цепи проинсулина. После адсорбции смолу промывают буфером на основе уксусной кислоты (рН 3-4) и проинсулин с повторно уложенной цепью элюируют водным раствором буфера на основе уксусной кислоты (рН 3-4), содержащим 15-50% (в объемном отношении), предпочтительно 30-50% (в объемном отношении) ацетона. Проинсулин человека с повторно уложенной цепью выделяют из элюента в виде порошка для дальнейшего использования путем упаривания и сушки вымораживанием собранных активных фракций или в виде осадка, добавляя хлорид цинка к собранным активным фракциям в конечной концентрации от 0,004 до 4% (в отношении веса к объему), предпочтительно от 0,004 до 0,04% (в отношении веса к объему), для чего показатель рН элюента доводят до 5-7, предпочтительно до 5-6, добавляя NaOH, и центрифугируют.
Способ получения предшественника инсулина человека по этому изобретению обладает следующими преимуществами по сравнению с известными способами: во-первых, одновременное выполнение растворения и сульфирования позволяет упростить процесс при массовом производстве; во-вторых, успешно устраняется денатурация, такая как гелеобразование, происходящая под действием межмолекулярной полимеризации в образце с высокой концентрацией; в-третьих, полностью решается проблема снижения растворимости образца в результате образования неправильных межмолекулярных дисульфидных связей во время растворения. Хотя эту проблему можно устранить, используя такие восстановители, как 2-меркаптоэтанол или DTT, сульфирование сульфитом и тетратионатом дает более стабильный результат и обеспечивает лучшую растворимость по сравнению с обработкой восстановителями; в-четвертых, замещение групп -SH в остатках цистеина группами -SSО3 - происходит до обработки бромистым цианом, что предотвращает необратимую денатурацию остатков цистеина, которая может иметь место на стадии растворения и на последующих стадиях, например, денатурация цистеина в цистеиновую кислоту (см. патент США 4451396); в-пятых, ионообменную хроматографию или адсорбционную хроматографию выполняют сразу же после обработки бромистым цианом и упаривания без какой-либо предварительной обработки, такой как обессоливание, что предпочтительно при осуществлении непрерывного процесса в промышленном масштабе.
Отличительной особенностью настоящего изобретения является эффективное выполнение реакции повторной укладки цепи, которая является одной из наиболее существенных стадий в процессе промышленного производства инсулина. В частности, увеличение выхода и хорошая воспроизводимость в промышленном масштабе достигаются благодаря непрерывному осуществлению этой реакции в смесителе. Кроме того, можно одновременно производить очистку, концентрирование и обессоливание нативного проинсулина при его отделении от реакционной смеси, используя в промышленном процессе адсорбционную смолу после выполнения реакции повторной укладки цепи.
В известных способах получения предшественника инсулина человека, которые обычно включают реакцию повторной укладки цепи, остатки цистеина в полипептиде химической формулы (I) сульфируют, чтобы получить S-сульфонат проинсулина, который подвергают взаимодействию с 1-5 эквивалентами 2-меркаптоэтанола на одну группу -SSО3 - при нейтральном или щелочном показателе рН 7-11,5 (см. патент США 4430266). В результате выполнения такой периодической реакции выход продукта с повторно уложенной цепью составляет около 60%, хотя его можно увеличить до 80%, применяя систему рециркуляции, включающую сложные стадии доведения показателя рН до 4-6 после реакции повторной укладки цепи, что дает промежуточные вещества с неправильной укладкой цепи в агрегатном состоянии, а также повторное сульфирование и повторную укладку цепи (см. патент США 4801684). Однако в соответствии с настоящим изобретением до 80% S-сульфоната проинсулина можно превратить в нативный проинсулин в результате выполнения только одной стадии реакции повторной укладки цепи, что существенно упрощает данную стадию и увеличивает выход продукта.
Во многих исследованиях, посвященных реакции повторной укладки цепи, указывается, что основной причиной уменьшения выхода белков с повторно уложенной цепью является формирование агрегатов в результате образования нековалентных связей между доступными гидрофобными остатками промежуточных веществ во время реакции повторной укладки цепи; и что на формирование агрегатов большое влияние оказывают несколько факторов, таких как концентрация белка, объем реакционной смеси, температура, рН и т.д., в то время как концентрация белка существенно влияет на продуктивность. Таким образом, несмотря на достижение высокого выхода при осуществлении реакции повторной укладки цепи при низкой концентрации белка, это неприемлемо в промышленном масштабе. Поэтому в этой области предложены различные альтернативные способы повторной укладки цепи рекомбинантного белка. Например, предложен способ добавления ингибитора агрегации, такого как аргинин, детергента, такого как полиэтиленгликоль, или денатуранта, и способ поэтапного увеличения концентрации белка во время реакции повторной укладки цепи.
Однако указанные способы обладают недостатком, который выражается в медленном смешивании до однородного состояния таких реагентов, как S-сульфонат проинсулина и 2-меркаптоэтанол, в результате чего происходит агрегация, уменьшающая выход продукта с повторно уложенной цепью, так как для осуществления реакции повторной укладки цепи при производстве инсулина в промышленном масштабе необходим большой объем реакционной смеси. Чтобы сократить время, необходимое для достижения равновесного состояния при смешивании S-сульфоната проинсулина и 2-меркаптоэтанола, авторы настоящего изобретения предлагают выполнять реакцию повторной укладки цепи при непрерывном смешивании реагентов в смесителе небольшого объема, что в конечном счете обеспечивает высокий выход проинсулина даже при использовании S-сульфоната проинсулина с высокой концентрацией.
Для очистки проинсулина с повторно уложенной цепью после выполнения реакции повторной укладки цепи обычно используют гель-фильтрационную хроматографию, такую как хроматография на колонке Sephadex G-50 и ионообменная хроматография, которая требует применения стадии замены буферного раствора и стадии обессоливания для удаления оставшихся солей (см. патент США 4430266). Обычными методами обессоливания являются гель-фильтрация, диализ, ультрафильтрация и т.д. При осуществлении гель-фильтрации для разделения веществ в зависимости от молекулярной массы или структуры с учетом различий во времени удерживания для веществ, проходящих через гель, используют полидекстрановый гель, такой как Sephadex G-25. С другой стороны, при выполнении диализа вместо геля используют диализную мембрану и для ультрафильтрации используют картридж, такой как полое волокно и кассета, и дисковую мембрану.
Однако указанные методы обладают рядом недостатков; например, при осуществлении гель-фильтрации эффективность обработки образца зависит от объема геля, помещенного в колонку, а не от количества или концентрации образца (эффективность от 10 до 25% объема геля). Таким образом, при использовании на стадии гель-фильтрации разведенного образца размер колонки увеличивается. Кроме того, элюированный образец находится в разведенном состоянии, что создает трудности на последующих стадиях. С другой стороны, недостатком диализного метода является потеря образца, вызванная неспецифическим связыванием образца с диализной мембраной, и ограниченная эффективность. Недостатком ультрафильтрации является необходимость применения специального оборудования, потеря образца, вызванная не специфическим связыванием, загрязнение, засорение и уменьшение скорости потока, хотя этот метод характеризуется такими преимуществами как высокая эффективность и возможность эффективного концентрирования.
В соответствии с настоящим изобретением стадию обессоливания в отличие от известных способов осуществляют при помощи адсорбционной хроматографии, которая позволяет успешно решить вышеуказанные проблемы, такие как ограниченные объемы разведения образца, неспецифическое связывание и т.д. Реакционный раствор для повторной укладки цепи, содержащий активный проинсулин, доводят до кислого состояния с показателем рН 3-4 и вводят в полярную метакрилатную смолу, где удаляют практически весь проинсулин с повторно уложенной цепью, используя буферный раствор (рН 3-4), содержащий от 15 до 50% (в объемном отношении) ацетона. В этой связи следует отметить, что для адсорбции органических веществ с относительно высокой полярностью предпочтительно используют полярную метакрилатную смолу, производимую под товарным знаком HP-2MG компанией Mitsubishi Chemical Co. Стадию адсорбции/элюирования осуществляют на колонке для достижения наибольшей эффективности обессоливания, концентрирования и очистки. Однако эту стадию можно осуществлять как на колонке, так и в виде периодического процесса. На этой стадии выход может составить более 90%, и эффективные концентрации могут быть увеличены в десятки раз в зависимости от концентрации введенного образца.
То есть проинсулин с повторно уложенной цепью можно экономично обессоливать, концентрировать и очищать на одной стадии, и сам элюент можно сразу же использовать на последующих стадиях.
Настоящее изобретение далее иллюстрируется нижеследующими примерами, которые не ограничивают объем этого изобретения.
Пример 1. Очистка внутриклеточных телец
Клетки Е. coli, экспрессирующие слитые белки проинсулина в виде внутриклеточного тельца (см. корейскую патентную публикацию 94-1855), суспендируют в 0,1 М трис-буфере (рН 7,9), содержащем 50 мМ EDTA, 10% сахарозу и 0,1 мМ PMSF в отношении 1:5-10 (в отношении веса к объему), и лизируют под давлением 630 атм (9000 фунтов/кв.дюйм). Лизат центрифугируют со скоростью 5000 об/мин в течение 30 минут при 4oС, чтобы получить осадок. Осадок (300 г, мокрый вес), содержащий внутриклеточные тельца, промывают 10 объемами 2%-ного тритона X100 и дистиллированной водой и центрифугируют, с получением очищенных внутриклеточных телец.
Пример 2. Растворение внутриклеточных телец с использованием щелочи
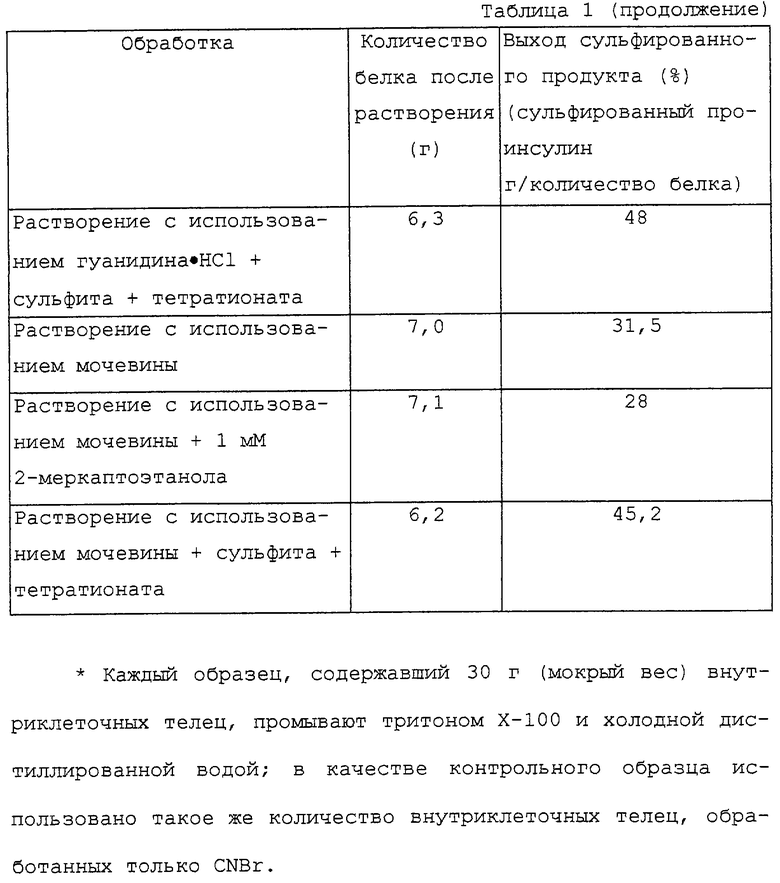
Внутриклеточные тельца, полученные в примере 1, однородно суспендируют в 20 объемах дистиллированной воды, перемешивают в течение 3 часов и центрифугируют со скоростью 12000 об/мин в течение 30 минут, чтобы удалить осадок. Показатель рН полученного супернатанта доводят до 5,5, добавляют 1 М раствор НСl и центрифугируют со скоростью 5000 об/мин в течение 30 минут, чтобы получить осадок. Осажденный белок растворяют в 70%-ной (в объемном отношении) муравьиной кислоте до концентрации 10 мг/мл. Бромистый циан добавляют в молярном отношении 100:1 в расчете на количество белка и перемешивают в течение 12 часов при 25oС. Смесь упаривают при пониженном давлении до совершенно сухого состояния и полученный таким образом белок растворяют в 20 мМ трис-буфера (рН 9,5), содержащего 7 М мочевины. Сульфит натрия и тетратионат натрия добавляют в конечной концентрации, соответственно равной 0,3 М и 0,1 М, и перемешивают в течение 6 часов. При помощи ВЭЖХ определяют концентрацию сульфированного проинсулина (см. таблицу 1).
Пример 3. Растворение внутриклеточных телец с использованием гуанидина•НСl и восстановителей
Внутриклеточные тельца, полученные в примере 1, суспендируют, как это описано ниже, в 10 объемах нескольких буферных растворов, содержащих денатурант: сначала их растворяют в 20 мМ трис-буфера (рН 9,5), содержащего 6-7 М гуанидина•НСl и 1 мМ EDTA; затем их растворяют в 20 мМ трис-буфера (рН 9,5), содержащего 6-7 М гуанидина•НСl и 1 мМ EDTA, и добавляют 1 мМ 2-меркаптоэтанола; после этого их растворяют в 20 мМ трис-буфера (рН 9,5), содержащего 6-7 М гуанидина•НСl и 1 мМ EDTA, и добавляют сульфит натрия и тетратионат натрия в конечной концентрации, соответственно равной 0,3 М и 0,1 М. Каждый раствор перемешивают в течение 12 часов при 4oС и центрифугируют со скоростью 12000 об/мин в течение 30 минут, чтобы удалить осадок. К полученному супернатанту добавляют примерно 10 объемов холодной воды и центрифугируют со скоростью 5000 об/мин в течение 30 минут, чтобы получить осадок. Осажденный белок растворяют в 70%-ной (в объемном отношении) муравьиной кислоте до концентрации 10 мг/мл. Бромистый циан добавляют в молярном отношении 100: 1 в зависимости от количества белка и перемешивают в течение 12 часов при 25oС. Смесь упаривают при пониженном давлении до совершенно сухого состояния и полученный таким образом белок растворяют в 20 мМ трис-буфера (рН 9,5), содержащего 7 М мочевины. Сульфит натрия и тетратионат натрия добавляют в конечной концентрации, соответственно равной 0,3 М и 0,1 М, и перемешивают в течение 6 часов при 25oС. При помощи ВЭЖХ определяют концентрацию сульфированного проинсулина (см. таблицу 1).
Пример 4. Растворение внутриклеточных телец с использованием мочевины и восстановителей
Внутриклеточные тельца, полученные в примере 1, суспендируют, как это описано ниже, в 10 объемах нескольких буферных растворов, содержащих денатурант: сначала их растворяют в 20 мМ трис-буфера (рН 9,5), содержащего 7-8 М мочевины и 1 мМ EDTA; затем их растворяют в 20 мМ трис-буфера (рН 9,5), содержащего 7-8 М мочевины и 1 мМ EDTA, и добавляют 1 мМ 2-меркаптоэтанола; после этого их растворяют в 20 мМ трис-буфера (рН 9,5), содержащего 7-8 М мочевины и 1 мМ EDTA, и добавляют сульфит натрия и тетратионат натрия в конечной концентрации, соответственно равной 0,3 М и 0,1 М. Каждый раствор перемешивают в течение 12 часов при 4oС и центрифугируют со скоростью 12000 об/мин в течение 30 минут, чтобы удалить осадок. К полученному супернатанту добавляют примерно 10 объемов холодной воды и центрифугируют со скоростью 5000 об/мин в течение 30 минут, чтобы получить осадок. Осажденный белок растворяют в 70%-ной (в объемном отношении) муравьиной кислоте до концентрации 10 мг/мл. Бромистый циан добавляют в молярном отношении 100:1 в зависимости от количества белка и перемешивают в течение 12 часов при 25oС. Смесь упаривают при пониженном давлении до совершенно сухого состояния и полученный таким образом белок растворяют в 20 мМ трис-буфера (рН 9/5), содержащего 7 М мочевины. Сульфит натрия и тетратионат натрия добавляют в конечной концентрации, соответственно равной 0,3 М и 0,1 М, и перемешивают в течение 6 часов при 25oС. При помощи ВЭЖХ определяют концентрацию сульфированного проинсулина (см. таблицу 1).
Как показано в таблице 1, добавление сульфита натрия и тетратионата натрия после растворения с использованием мочевины или гуанидина•НСl увеличивает выход сульфированного проинсулина, который в 1,5-2 раза выше, чем у контрольного образца. С другой стороны, когда растворение осуществляют с использованием только гуанидина•НСl без какого-либо восстановителя, в процессе добавления 70%-ной (в объемном отношении) муравьиной кислоты происходит гелеобразование. Поэтому в этом случае нельзя определить количество белка после растворения и выход сульфированного продукта. Это может быть вызвано межмолекулярным гидрофобным взаимодействием или полимеризацией в результате образования дисульфидной связи. Кроме того, добавление 2-меркаптоэтанола вызывает значительное уменьшение выхода, что может быть также вызвано вышеописанными причинами.
Совершенно ясно, что межмолекулярное взаимодействие можно предотвратить введением групп -SSО3 - слитый белок сульфированного проинсулина, в результате чего вся молекула приобретает отрицательный заряд; и что растворение с использованием щелочи влияет на устойчивость белков. С другой стороны, добавление сульфита и тетратионата после растворения с использованием мочевины или гуанидина•НСl не оказывает существенного влияния на выход сульфированного проинсулина. Поэтому использование мочевины для растворения в промышленном масштабе должно способствовать значительному снижению затрат.
Кроме того, анализ ВЭЖХ образца, полученного на стадиях растворения и сульфирования, обработки бромистым цианом и растворения в 20 мМ трис-буфера, содержащего 7 М мочевины, показывает, что добавление сульфита и тетратионата после обработки бромистым цианом не увеличивает выход сульфированного продукта.
Пример 5. Растворение внутриклеточных телец с использованием мочевины и сульфирование
В соответствии с результатами, полученными в вышеописанных примерах 2-4, внутриклеточные тельца растворяют в растворе мочевины и выполняют сульфирование. То есть 110 г (мокрый вес) очищенных внутриклеточных телец растворяют в 10 объемах 20 мМ трис-буфера (рН 8,5), содержащего 8 М мочевины и 1 мМ EDTA. Сульфит натрия и тетратионат натрия добавляют в конечной концентрации, соответственно равной 0,3 М и 0,1 М, перемешивают в течение 12 часов при 4oС и центрифугируют со скоростью 12000 об/мин в течение 30 минут, чтобы удалить осадок. К полученному супернатанту добавляют примерно 10 объемов холодной воды, показатель рН раствора доводят до около 5,5, добавляя 2 н. раствор НСl, и центрифугируют со скоростью 5000 об/мин в течение 30 минут, чтобы получить осадок (250 г, мокрый вес). Количественный анализ белка показывает, что получено примерно 40 г белка.
Пример 6. Обработка бромистым цианом
Осажденный белок растворяют в 2 л 70%-ной (в объемном отношении) муравьиной кислоты. Бромистый циан (CNBr) добавляют в молярном отношении 100:1 в зависимости от количества белка и перемешивают в течение 12 часов при 25oС. Смесь упаривают при пониженном давлении до совершенно сухого состояния. Полученный белок растворяют в 20 мМ трис-буфера (рН 8,0), содержащего 7 М мочевины, и анализируют ВЭЖХ.
Пример 7. Анионообменная хроматография
Колонку (2,5 х 30) см заполняют DEAE-сефацелом со скоростью потока 1,5 объема колонки в час и уравновешивают 20 мМ трис-буфера (рН 8,0), содержащего 7 М мочевины. Затем в колон ку вводят образец, полученный в примере 6, в количестве 20 мг на 1 мл смолы и промывают колонку 1 объемом уравновешивающего буфера. Белок элюируют градиентом концентрации, используя уравновешивающий буфер, содержащий 0-1 М NaCl. Элюенты, собранные при использовании 0,35-0,45 М NaCl, анализируют ВЭЖХ, которая показывает, что чистота продукта равна 80% или выше и степень выделения равна 91%.
Пример 8. Повторная укладка цепи S-сульфоната проинсулина
S-сульфонат проинсулина (1 г), полученный сульфированием активного рекомбинантного проинсулина, очищенного ВЭЖХ с обращенной фазой, растворяют в 500 мл 50 мМ раствора глицинового буфера (рН 10,6). Раствор продувают газообразным азотом, чтобы удалить кислород, после чего камеру герметично закрывают. В другой камере 104 мкл 2-меркаптоэта-нола добавляют к 500 мл 50 мМ раствора глицинового буфера (рН 10,6), продувают газообразным азотом, чтобы удалить кислород, и камеру герметично закрывают. Оба раствора быстро вводят, перемешивая, в смеситель объемом 1 мл со скоростью потока 50 мл/час. Смешанный реакционный раствор для повторной укладки цепи вводят со скоростью потока 100 мл/час в расположенную ниже емкость, продуваемую газообразным N2, медленно перемешивают и подвергают взаимодействию в течение 18 часов при 4o С. После окончания реакции раствор подкисляют до рН 2,9 ± 0,1, добавляя 2 М раствор НСl. Анализ ВЭЖХ показывает, что выход продукта с повторно уложенной цепью равен 55%.
Пример 9. Влияние концентрации белка на повторную укладку цепи
Влияние концентрации белка на выход продукта с повторно уложенной цепью (то есть превращение S-сульфоната проинсулина в проинсулин) исследуют, выполняя ряд реакций аналогично примеру 8, кроме концентраций белка (см. таблицу 2).
Пример 10. Влияние отношения групп -SН:-SSO3 -на повторную укладку
Влияние отношения групп -SН:-SSO3 - на выход продукта с повторно уложенной цепью исследуют, выполняя ряд реакций аналогично примеру 8 с использованием разных отношений -SН:-SSO3 - (см. таблицу 3).
Пример 11. Влияние концентрации мочевины на повторную укладку цепи
Влияние концентрации мочевины на выход продукта с повторно уложенной цепью исследуют, выполняя ряд реакций аналогично примеру 8, кроме концентраций мочевины (см. таблицу 4).
Пример 12. Повторная укладка цепи S-сульфоната проинсулина, очищенного ионообменной хроматографией
Элюент, содержащий 10 г S-сульфоната проинсулина, полученного в примере 1, разводят 50 мМ раствором глицинового буфера (рН 10,6), содержащим 1 М мочевины, до конечного объема, равного 5 л. Смесь продувают газообразным азотом, чтобы удалить кислород, после чего камеру герметично закрывают. В другой камере 781 мкл 2-меркаптоэтанола добавляют к 5 л 50 мМ раствора глицинового буфера, содержащего 1 М мочевины, продувают смесь газообразным азотом, чтобы удалить кислород, и камеру герметично закрывают. Оба раствора быстро вводят, перемешивая, в смеситель объемом 1 мл со скоростью потока 500 мл/час. Реакционную смесь для повторной укладки цепи вводят со скоростью потока 1 л/час в емкость, деаэрируемую газообразным N2, медленно перемешивают и подвергают взаимодействию в течение 18 часов при 4oС. После окончания реакции раствор подкисляют до рН 2,9 ± 0,1, добавляя 2 М раствор НСl. Анализ ВЭЖХ показывает, что выход продукта с повторно уложенной цепью равен 81%.
Пример 13. Очистка рекомбинантного проинсулина человека адсорбционной хроматографией
Смолу HP-2MG (Mitsubishi Chemical Co., Japan), полярную метакрилатную смолу, оставляют набухать в количестве 1 г смолы на 5 мл ацетона в течение 6 часов при комнатной температуре. Затем смолу обильно промывают в указанном порядке 0,1 н. раствором NaOH, дистиллированной водой, 0,1 н. раствором НСl, дистиллированной водой и 20 мМ уксусной кислоты (рН 3,2 ± 0,2) и вводят в колонку. Колонку уравновешивают 3 объемами уравновешивающего буфера (20 мМ уксусной кислоты, рН 3,2 ± 0,2) со скоростью потока, равной 1 объему колонки в час. Реакционный раствор, содержащий проинсулин с повторно уложенной цепью, полученный в примере 12, вводят в колонку в количестве 8 г белка на 1 л смолы и промывают колонку 1 объемом буфера на основе 20 мМ уксусной кислоты (рН 3,2 ± 0,2). Проинсулин с повторно уложенной цепью элюируют таким же буфером, содержащим 30% ацетона. В результате этого получают 92% или больше проинсулина с повторно уложенной цепью, не содержащего загрязняющих примесей, таких как глицин и мочевина. ВЭЖХ и количественный анализ белка показывают, что при увеличении концентрации белка примерно в 10 раз чистота продукта возрастает примерно в 1,3 раза. Элюент, содержащий активный проинсулин, упаривают, чтобы удалить ацетон, и сушат вымораживанием, либо показатель рН элюента доводят до 5,4, добавляя 1 н. раствор NaOH и хлорид цинка в конечной концентрации 0,04% (в отношении веса к объему), что дает проинсулин с повторно уложенной цепью.
Как описано выше и проиллюстрировано примерами, настоящее изобретение относится к способу получения проинсулина человека, в котором значительно упрощены стадии растворения и сульфирования, концентрирования, обессоливания и очистки при увеличении выхода продукта реакции с повторно уложенной цепью. В соответствии с настоящим изобретением предшественник рекомбинантного инсулина человека можно получать с хорошей воспроизводимостью.



Изобретение относится к области биотехнологии и касается способа получения проинсулина человека. Сущность изобретения включает получение предшественника рекомбинантного инсулина посредством суспендирования клеток Е. coli, экспрессирующих слитые белки проинсулина в форме телец включения, лизиса, суспендирования телец включения в растворе, содержащем мочевину или гуанидин HCl, сульфирования слитых белков проинсулина сульфонатом натрия и тетратионатом натрия, центрифугирования слитого белка S-сульфоната проинсулина, его очистки посредством анионообменной хроматографии, его разведения в буферном растворе и продувки газообразного азота, смешивании с буферным раствором, содержащим 2-меркаптоэтанол и перемешивание с получением реакционной рефолдинг-смеси, очистки адсорбционной и элюиций проинсулина человека с реконструированной цепью. Преимущество изобретения заключается в увеличении выхода продукта. 16 з.п.ф-лы, 4 табл., 3 ил.
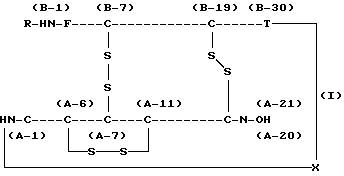
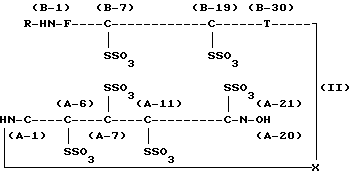
где R обозначает аминокислотный остаток или пептид, который может быть подвергнут ферментативному или химическому гидролизу;
Х обозначает связь между аминокислотной группой А-1 в цепи А инсулина и карбоксильной группой В-30 в цепи В инсулина, которая может быть отделена от цепи А или от цепи В ферментативным или химическим путем, при условии, что область от А-1 до А-21 является цепью А инсулина и область от В-1 до В-30 является цепью В инсулина.
| СПОСОБ ВЫДЕЛЕНИЯ РЕКОМБИНАНТНОГО ПРОИНСУЛИНА | 1991 |
|
RU2049793C1 |
| СПОСОБ ВЫДЕЛЕНИЯ И ОЧИСТКИ ИНСУЛИНА ИЛИ ЕГО БИОТЕХНОЛОГИЧЕСКИХ ПРЕДШЕСТВЕННИКОВ | 1994 |
|
RU2081122C1 |
| Обзор информации ММБП "Современные тенденции развития биотехнологии за рубежом" | |||
| - М., 1988, в.8, стр.5. | |||

