Область применения изобретения
Настоящее изобретение относится к антигенам, являющимся защитными от inter alia, инфекции Echinococcus granulosus, к вакцинам, содержащим подобные антигены, и к способам защиты хозяев, подверженных заражению паразитами Echinococcus и Taenii.
Предпосылки создания изобретения.
Эхинококкоз возникает в результате заражения личинками (метацестодная стадия) ленточных червей, принадлежащих к роду Echinococcus. Переход происходит в момент поедания хищником добычи, т.е. от травоядных или всеядных промежуточных хозяев к плотоядным окончательным хозяевам.
Из ленточных червей рода Echinococcus наибольший интерес представляет Echinococcus granulosus. Е. granulosus является ввозбудителем поликистозного эхинококкоза (ПКЭ), для которого характерен обмен между домашней собакой и домашними травоядными животными, например: овцами и крупным рогатым скотом, причем человек также может служить промежуточным хозяином для паразита, но обычно не играя важной роли в его жизненном цикле. Соответственно, Е.granulosus географически широко распространен и в его размещении наблюдается явная параллель с теми областями мира, в которых скотоводство является основным занятием. Таким образом, можно предсказать, что ПКЭ является распространенным эндемическим заболеванием на больших территориях Южной Америки, Африки, Австралии, Центральной Европы, Центральной Азии, прибрежного Средиземноморья, включая Северную Африку.
В некоторых странах в настоящее время пытаются вести борьбу с ПКЭ путем разъяснения бесед с хозяевами собак и регулярной антигельминтной обработки собак в качестве окончательных хозяев паразита. Однако, хотя подобные программы, как правило, частично успешны, полного истребления паразита сказалось гораздо труднее достигнуть.
Другой желательный подход в борьбе с ПКЭ состоит в применении вакцины, направленной против кистозной стадии жизненного цикла E. granulosus у восприимчивых к нему промежуточных хозяев. Значительное преимущество такого подхода заключается в том, что при этом предотвращается передача инфекции собаке при поедании ею содержащих кисту потрохов промежуточного хозяина. Как следствие, искоренение болезни становится реальной возможностью.
В широком смысле именно на применение вакцины и направлено настоящее изобретение.
Однако для создания промышленной вакцины существенное значение имеет идентификация характерных защитных антигенов против E. granulosus. Хотя проведенные к настоящему времени исследования (Gemmell M.A., Jmmunology 11, 325-335 (1966). Heafh и др. J.Parasitol 67, 797-799 (1981)) ясно показали, что онкосферы E. granulosus являются мощным источником таких антигенов, никаких характерных антигенов онкосфер E. granulosus, обеспечивающих защиту, идентиицировано не было.
Заявка на Международный патент РСТ/ГР91/00563 (публикация 092/01051) раскрывает иммуногенный E. granulosus пептид, а также кодирующую этот пептид ДНК-последовательность и способы, как диагностики, так и борьбы с инфекцией E. granulosus с применением данного пептида. Однако описанный E. granulosus антиген (известный под названием "антиген 5") не имеет отношения к антигенам онкосфер, исследуемых заявителями. Более того, заявителями в их опубликованном исследовании показано, что присутствие антител к антигену 5 не защищает ягнят при контрольном заражении яйцами E. granulosus (Неаth и др., Jnfl. Journal of Parasitolody 22, 1017-1021(1992)).
Краткое изложение сущности изобретения
Таким образом, цель настоящего изобретения состоит в получении особого антигена для применения в вакцине, обеспечивающей защиту от заражения E. granulosus, или хотя бы обеспечения общества правом полезного выбора.
Соответственно, одним из своих аспектов изобретением дается по существу в чистом виде антигенный полипептид с молекулярной массой в интервале 23-25 кД (при определении методом НДС-ПАГЭ), включающий аминокислоты 4-77 из приведенной на фиг. 4 последовательности, способный создавать иммунологическую реакцию на инфицирование E. granulosus у восприимчивых к заражению хозяев, или фрагмент или вариант пептида, обладающей эквивалентной защитной иммунологической активностью.
В предпочтительном воплощении изобретения полипептид включает аминокислоты 1-154 из приведенной на фиг.4 последовательности.
В еще более предпочтительном воплощении изобретения пептид представляет собой фрагмент, включающий часть или всю аминокислотную последовательность с фиг.4.
Удобно то, что полипептид или фрагмент пептида является продуктом экспрессии в клетке-хозяине, кодирующих их нуклеотидной последовательности.
Другим своим аспектом изобретение направлено на препарат, способный создавать защитную иммунологическую реакцию на инфицирование E. granulosus у восприимчивых к заражению хозяев, причем препарат включает компонент, выбранный из группы, включающей:
(а) вышеохарактеризованный полипептид,
(в) пептидный фрагмент полипептида (а), обладающий эквивалентной иммунологической активностью, и
(с) вариант (а) или (в), модифицированный вставкой, замещением или делецией одной или нескольких аминокислот и обладающих хотя бы эквивалентной иммунологической активностью.
И еще одним аспектом изобретением дается молекула ДНК, выбранная из группы, включающей:
(а) нуклеотидную последовательность, кодирующую вышеохарактеризованный антигенный полипептид,
(в) нуклеотидную последовательность, кодирующую пептидный фрагмент антигенного полипептида (а), причем фрагмент обладает эквивалентной с полипептидом (а) защитной иммунологической активностью, и
(с) нуклеотидную последовательность, кодирующую вариант полипептида (а) или пептида (в), в котором аминокислотная последовательность модифицирована вставкой, замещением или делецией одной или нескольких аминокислот, при этом вариант обладает эквивалентной с полипептдом (а) или пептидным фрагментом (в) защитной иммунологической активностью.
В рекомендуемом воплощении изобретения молекула ДНК включает хотя бы нуклеотиды 10-233 из приведенной на фиг.4 последовательности, более предпочтительно нуклеотиды 3-461 последовательности с фиг.4 и наиболее предпочтительно нуклеотиды 1-461 последовательности с фиг.4.
Другими своими аспектами изобретением даются рекомбинантные экспрессионные векторы, содержащие вышеохарактеризованную молекулу ДНК, клетки-хозяева, трансформированные такими векторами и способные экспрессировать закодированные полипептид или его пептидный фрагмент, или его вариант, а также способы продуцирования антигенного полипептида или его пептидного фрагмента, или его варианта, состоящие в культивировании вышеохарактеризованной клетки-хозяина и выделении продукта экспрессии.
Другим своим аспектом изобретением дается вакцина против инфицирования E. granulosus, содержащая вышеохарактеризованные полипептид E. granulosus, пептидный фрагмент или вариант, в смеси иммунологически приемлемым адъювантом или носителем.
И еще одним своим аспектом изобретением дается способ защиты восприимчивых хозяев от инфицирования Echinococcus или Taeniid паразитами, включающий этап введения указанному хозяину такого количества вышеохарактеризованных полипептида, пептидного фрагмента или варианта, которое обеспечивает защиту от заражения.
Полипептид, пептидный фрагмент или вариант удобно вводить указанному хозяину в виде вышеохарактеризованной вакцины.
Еще одним аспектом изобретением дается антитело, специфичное к вышеохарактеризованным антигенному полипептиду или к его пептидному фрагменту, или к его варианту.
Прочие воплощения изобретения станут очевидными из последующего описания.
Пояснения к диаграммам.
Хотя изобретение достаточно широко охарактеризовано выше, для специалиста понятно, что изобретение не ограничивается приведенной характеристикой, но включает также варианты, для которых в последующем описании приводятся примеры. В частности, лучшему пониманию настоящего изобретения способствует ссылка на прилагаемые диаграммы.
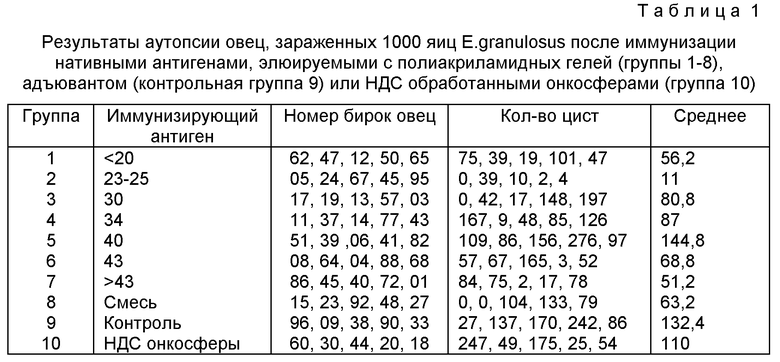
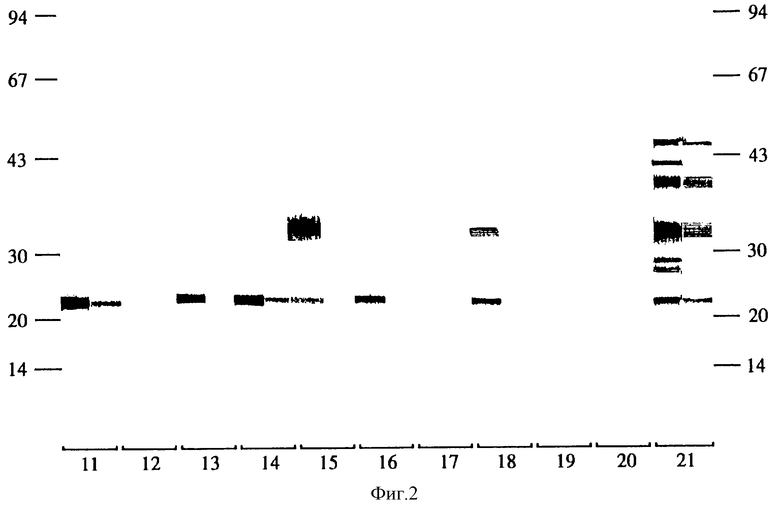
Фиг. 1 показывает IgG1 и IgG2 реакцию овечьих антител на вакцинацию полученными методом препаративного НДС-ПАГЭ фракциями онкосфер. Группы овец вакцинируют фракциями онкосфер, представляющими полученные методом НДС-ПАГЭ фракции, включающими антигены с установленной молекулярной массой. Группа 1: все молекулы имеют массу ниже 21 кД, группа 2: 23+25 кД, группа 3: 30 кД, группа 4: 34 кД, группа 5: 40 кД дублет, группа 6: 43 кД, группа 7: все молекулы имеют массу выше 43 кД, группа 8: смесь всех молекул, группа 9: только адъювант, в группе 10 овцы вакцинированы нефракционированными онкосферами, а в остальном обработаны точно также, как овцы групп 1-9, и группа 21 вакцинирована замороженным-оттаянным продуктом обработки ультразвуком активированных онкосфер E. granulosus. На фиг. 1 приведен анализ реакции антител овец в каждой из указанных групп, реагирующих с полными экстрактами онкосфер, полученными методом НДС-ПАГЭ.
Фиг.2 показывает IgG1 и IgG2 реакцию овечьих антител на вакцинацию клонированными антигенами. Группы: 11, клон S2, 12 клон S3, 13, клон 48, 14, клон 95, 15, клон 101, 16, клон 119, 17, клон 123, 18, смесь клонов, 19, клоны 33, 42 и 85, 20, GST - контроль, 21, объединенная сыворотка от 5 овец, иммунизированных замороженным - оттаянным продуктом обработки ультразвуком активированных онкосфер Е.
Методика иммунизации аналогична методике, применяемой для антигенов групп 11-20.
Фиг. 3 показывает реакцию овечьего IgG2 на вакцинацию полученными препаративными НДС-ПАГЭ фракциями антигенов онкосфер или клонированными антигенами. Группы те же, что и выше.
На фиг. 4 приведена нуклеотидная последовательность и выведенная на ее основе аминокислотная последовательность пептидного фрагмента защитного антигена изобретения. Цифрами с левой стороны указаны номера нуклеотидов от 5' к 3'-концу, цифрами с правой стороны указаны номера аминокислот. Трансляционный терминационный кодон подчеркнут.
Подробное описание изобретения.
Как указано выше, в своем основном аспекте изобретение направлено на создание антигена, способного защищать хозяина от инфицирования E. granulosus независимо от того, заражен ли хозяин червем или заражение находится на стадии цисты. К хозяевам, восприимчивым к инфицированию E. granulosus, относятся млекопитающие, включая и человека. Соответственно, примеры хозяев, для которых потенциально применимо настоящее изобретение, включают: овец, коров, свиней, коз, лошадей и человека.
В своих исследованиях заявители идентифицировали E. granulosus полипептид, способный участвовать в защите восприимчивого к инфицированию E. granulosus хозяина. Такой E. granulosus полипептид - это полипептид, молекулярная масса которого находится в интервале 23-25 кД.
Настоящее изобретение включает в свой объем антигены, происходящие из нативного E. granulosus полипептида, охарактеризованного выше, причем такие производные обладают защищающей хозяина активностью. Подобные производные обычно представляют собой пептидные фрагменты нативного полипептида, содержащими защитный эпитоп, но могут быть также и функционально эквивалентными вариантами нативного полипептида, модифицированного хорошо известными методами, например, сайтспецифичным мутагенезом (см. Adelman и др., D NA 2, 183 (1983)). К примеру, применением подобных методов можно заменить в последовательности одни аминокислоты на другие эквалентные аминокислоты. Группы аминокислот, для которых известна эквивалентность, включают:
(а) Ala Ser Thr Pro Gly
(в) Asn Asp Glu Gln
(c) His Arg Lys
(d) Met Leu Jls Val
(e) Phe Tyr Trp
В предпочтительном воплощении изобретения антиген представляет собой пептидный фрагмент, включающий часть или всю аминокислотную последовательность, приведенную на фиг.4.
Более предпочтительно, если пептидный фрагмент состоит из аминокислот 4-77 последовательности с фиг.4. Еще более предпочтительно, если пептидный фрагмент состоит из аимитнокислот 1-77 из последовательности с фиг.4. Наиболее предпочтительно, одноко, если пептидный фрагмент состоит изаминокислот 1-153 последовательности, приведенной на фиг.4.
Защитный антиген изобретения может быть получен выделением комплемента из нативных онкосфер E. granulosus использованием обычных методов очистки. Тем не менее, общепризнанно, что для получения антигена в промышленных масштабах желателен синтетический путь его получения. Подобные пути синтеза включают поэтапный твердофазный подход, описанный в работе Merryfield J. Amer. Chem. Soc. 85, 2149-2156(1963)), и получение применением методов биотехнологии. Последний путь, в частности, и был применен заявителями.
В еще одном своем аспекте изобретение, соответственно, относится к продуцированию рекомбинантными методами вышехарактеризованных антигенных полипептида или пептида.
В общем смысле продуцирование защитного антигена изобретения методами биотехнологии включает трансформирование приемлемого микроорганизма - хозяина или клетки-хозяина экспрессивным вектором, содержащим кодирующую антиген ДНК-последовательность, с последующим культивированием трансформированного хозяина и затем выделением экспрессированного антигена. Подобные методы в целом представлены в работе Sambrook и др., "Молекулярное клонирование", второе издание, Коулд Спринг Харбор Пресс (1987).
Начальный этап метода рекомбинантного продуцирования антигена включает легирование кодирующей антиген ДНК-последовательности в приемлемый экспрессионный вектор, содержащий промотор и участок связывания с рибосомой, работающими в клетке-хозяина, в которой будет трансформирована кодирующая последовательность. Наиболее обычные примеры таких экспрессионных векторов включают плазмиды, представляющие собой двуцепочечные ДНК-петли, автономно-реплицирующиеся в клетке-хозяие. Однако необходимо указать, что при осуществлении изобретения могут быть использованы и другие, отличные от плазмид приемлемые векторы.
Рекомендуется, чтобы клетка-хозяин, в которой клонируется и экспрессируется кодирующая полипептид ДНК-последовательность, была прокариотной, например, Е. сoli. Например, могут быть использованы Е.сoli Н5 (Raleigh E.A., и др. , Nucleic Acid Pеsearch 16(4) 1563-1575 (1988)), E.сoli К12 штамм 294 (АТСС 31446), E.coli В., E.coli, Х1776 (АТСС 31537), E.сoli штамм SТ9 или E. сoli JM101. Могут быть также использованы и другие прокариоты, например: бациллы, такие как Bacillus subtilis и энтеробактерии, такие как Salmonella typhimurium, Serratia marcesans или ослабленный штамм Mycobacteriuvbomis под названием Бацилла Calmette - Guerin (BCG).
В целом, если клетка-хозяина является прокариотом, применяют экспрессионные или клонирующие вектора, содержащие репликационные или контрольные последовательности, происходящие из видов, совместимых с клеткой-хозяином. Вектора кроме того могут нести маркерные последовательности, способные обеспечить фенотипный отбор трансформированных клеток. К примеру, E.сoli обычно трансформируют плазмидой рВР322, происходящей из видов E.сoli (Bolivar и др. , Gene 2, 95 (1977)). Плазмида рВР322 содержит гены устойчивости к ампициллину и тетрациклину, в силу чего обеспечивает средства легкого обнаружения трансформированных клеток.
Для применения в экспрессии плазмида, содержащая экспрессируемую ДНК, включает в себя промотор. Промоторы, наиболее часто применяемые в рекомбинантных ДНК-конструкциях, предназначены для прокариотных хозяев, включают (β-лактамазу (пенициллиназу) и лактозные промоторные системы (Chang и др. Nature 275,615 (1978), Hаkura и др. Science 198, 1056 (1977), Goedel и др., Nature 281, 544 (1979)) и триптофановую (trp) промоторную систему (Goeodel и др. , Nucleic Acld Res 8, 4057 (1980), ЕРО публ. 0036776). Хотя вышеприведенные промоторы применяются наиболее часто, были сконструированы и применены и другие микробиальные промоторы, например, Iac промотор (Amann и др., Gene 25, 167-178 (1983)), подробности, касающиеся их нуклеотидных последовательностей, опубликованы, что позволяет специалисту легировать их функционально методами генной инженерии с генами в векторах (Siebenlist и др., Cеll 20, 269 (1980)).
Помимо прокариотов могут быть также применены эукариотные микробы, например, дрожжи. Из эукариотных микроорганизмов наиболее часто применяют Saccharomyces cerevisiae или обычные пекарские дрожжи, хотя легко доступен целый ряд других штаммов. Для экспрессии в Saccharomyces наиболее часто применяют, например, плазмиду УРр7 (Stinchomb и др. Nature 282, 39 (1979), Kingsman и др. , Gene 7, 141 (1979), Tschemper и др., Gene 10, 157 (1980). Данная плазмида уже содержит trp1 ген, обеспечивающий маркер для отбора мутантного штамма дрожжей, не обладающего способностью расти в триптофане, например: АТСС 44076 или РЕР4-I (Jones, Genetics 85, 12 (1977)). Наличие trp1 повреждения в качестве характеристики генома дрожжевых клеток-хозяев обеспечивает эффективное средство обнаружения трансформации по росту в отсутствии триптофана.
Приемлемые промотирующие последовательности в дрожжевых векторах включают промоторы для 3-фосфоглицераткиназы (Hitzeman и др., J.Biol Chew 255, 2073 (0980) или других гликолитических ферментов (Hess и др., J.Aov Enzyme Reg 7, 149 (1968), Holland и др., Biochemistry 17, 4900 (1978)). К другим промоторам, обладающие дополнительным преимуществом транскрипции, регулируемой условиями роста, относятся промотирующая область для спиртовой дегидрогеназы, изоцитохром С, кислая фосфатаза, разрушающие ферменты, связанные с метаболизмом азота, и вышеупомянутая глицеральдегид -3-фосфатдегидрогеназа, а также ферменты, ответственные за усвоение мальтозы и галактозы. Приемлим любой плазмидный вектор, содержащий совместимый с дрожжами промотор, ориджин репликации и терминирующие последовательности.
Помимо микроорганизмов в качестве хозяев могут быть использованы культуры клеток, происходящие из многоклеточных организмов, например: млекопитающих или насекомых. В принципе, срабатывает любая такая клеточная культура, как от позвоночных, так и беспозвоночных. Однако наибольший интерес представляют клетки позвоночных, и размножение клеток позвоночных в культуре (тканевая культура) стала обычной процедурой в последние годы (Тканевые культуры, академик Пресс, под ред. Kruse и Patterson (1973). Примеры таких применимых линий клеток-хозяев включают: VERO и HELA-клетки и клетки яичников китайского хомячка (СНО клетки). Экспрессионные векторы для таких клеток включают (если необходимо): ориджин репликации, промотор, расположенный в восходящем направлении от экспрессируемого гена, а также любые необходимые участки связывания рибосомы, сайты полиаденилирования и последовательности терминации транскрипции.
При использовании в клетках млекопитающих регулирующие функции в экспрессионных векторах часто обеспечивают вирусным материалом. Например, обычно применяемые промоторы происходят из полиомы, аденовируса 2 и наиболее часто из обезьяньего вируса 40 (ОВ40). Особенно полезны ранний и поздний промоторы ОВ40, поскольку оба легко могут быть получены из вируса в виде фрагмента, содержащего также вирусный ориджин репликации ОВ40 (Tiers и др. Nafure 273, 113 (1978)). Могут быть использованы также меньшие и более крупные фрагменты ОВ40 при условии, что они включают в себя последовательности примерно в 250 п. о., простирающуюся от Hind III в направлении BglI сайта, расположенного в вирусном ориджине репликации. Кроме того, возможно также, а часто и желательно использовать промотор или регулирующие последовательности, обычно связанные с последовательностью целевого гена, при условии, что такие регулирующие последовательности совместимы с системами клетки-хозяина.
Ориджин репликации может быть обеспечен либо конструированием вектора, включающего экзогенный оридэин, происходящий, например, из ОВ40 или иного вирусного источника (напр. : полиомы, аденовируса, VSV, BPV), либо хромосомным механизмом репликации клетки-хозяина. Если вектор интегрирован в хромосому клетки-хозяина, этого часто достаточно.
После трансформации выбранного хозяина соответствующим вектором закодированные антигенные полипептид или пептид могут быть получены культивированием клетки-хозяина. В результате выделяют слитый белок.
После выделения антигенные полипептид или пептид при желании очищают. Принятая методика очистки, разумеется, будет зависеть от степени чистоты, необходимой от той области применения, для которой полипептид или пептид предназначены. Для большинства целей вакцинации достаточно отделить слитый белок от большей части остальных компонентов клеточной культуры, поскольку антиген может быть введен в вакцину в сравнительно сырой форме. Однако в случаях, если требуется более высокая степень чистоты, несущий компонент слитого белка может быть отщеплен от антигенного компонента. И вновь, как будет показано приведенными характерными примерами, этого легко достигнуть созданием соответствующего участка ферментативного расщепления между несущим компонентом и антигеном.
Если, как и рекомендуется, для продуцирования антигенного пептида применяются методы биотехнологии, первый этап состоит в получении ДНК, кодирующей целевой продукт. Такие молекулы ДНК составляют еще один аспект настоящего изобретения.
Молекула ДНК изобретения предпочтительно включает хотя бы часть или всю нуклеотидную последовательность, приведенную на фиг.4. В одном из вариантов молекула ДНК включает нуклеотиды 10-233 последовательности с фиг.4. В более предпочтительном варианте молекула ДНК включает нуклеотиды 3-233 последовательности с фиг.4. Однако наиболее предпочтительно, если молекула ДНК состоит из нуклеотидов 3-461 последовательности с фиг. 4, если не полной нуклеотидной последовательности, приведенной на фиг. 4.
ДНК молекула изобретения может быть получена в виде части, содержащейся в пределах ДНК молекулы, выделенной из соответствующего природного источника, или же может быть получена в виде безинтронной к ДНК применением обычных методов, например, приведенных в нижеследующем специальном описании. Рекомендуется кДНК.
Однако, как указано выше, настоящее изобретение включает также варианты полипептида, отличающиеся от нативных аминокислотных последовательностей вставкой, замещением или делецией одной или нескольких аминокислот. Если желателен такой вариант, изменяют нуклеотидную последовательность нативной молекулы ДНК. Подобное изменение в молекулу может быть внесено избирательным синтезом ДНК с помощью соответствующего синтезатора, например, ДНК синтезатора фирмы Эпплайд Биосистемс или, например, сайт-специфичным или кассетным мутагенезом с модификацией нативной ДНК.
После получения молекулу ДНК подготавливают для вставки вместе с выбранной регулирующей последовательностью в соответствующий блокирующий и/или экспрессионный вектор. Для этого ДНК расщепляют, наращивают и религируют по мере необходимости.
Расщепление проводят обработкой ДНК ферментами рестрикции в приемлемом буфере. Любой из большого числа промышленных ферментов рестрикции может быть использован согласно указаниям изготовителя. После расщепления нуклеиновую кислоту выделяют, например, осаждением этанолом.
Наращивание расщепленной ДНК осуществляют обычными методами. К примеру, если необходимы тупые концы, ДНК может быть образована ДНК-полимеразой I (полимераза Кленова), экстрагирована фенолом и хлороформом и осаждена этанолом.
Религация может быть проведена добавлением эквимолярных количеств целевых компонентов, соответствующим образом нарощенных для необходимого соответствия, и обработкой приемлемой лигазой (напр., Т4 ДНК-лигазой).
Помимо защитных антигенов изобретения и способа их получения настоящим изобретением дается вакцина от инфицирования E. granulosus. Такая вакцина включает в качестве необходимого компонента защищающее количество вышеуказанных E. granulosus полипептида, пептидного фрагмента или варианта в смеси с приемлемым адъювантом или носителем.
Примеры приемлемых, известных специалистам адъювантов включают: сапонины (или их производные или родственные им продукты), мурамилдипептид, димиколлят трегалозы, полный адъювант Фрейнда, неполный адъювант Фрейнда, другие эмульсии типа вода в масле, декстран, диэтиламиноэтилдекстран, алюминат калия, фосфат алюминия, гидроксид алюминия, бентонит, зимозан, полиэлектролиты, ретинол, фосфат кальция, потамин, саркозин, глицерин, сорбит, пропиленгликоль, не летучие масла и синтетические эфиры высших жирных кислот. В частности, найдено, что сапонины являются особенно эффективными адъювантами.
В других вариантах вакцина может дополнительно содержать иные лечебные для хозяина средства. Такие лечебные для хозяина средства включают противогельминтные или другие вакцины или иммуностимуляторы, например: интерфероны или интерлейкины.
Вакцина может быть введена хозяину любым известным специалисту способом. Тем не менее рекомендуется парентеральный путь введения вакцины. Термин "парентеральный" в применяемом здесь значении означает внутривенную, внутримышечную, накожную и подкожную инъекцию. Наиболее удобный путь введения подкожный.
Количество вводимой хозяину вакцины зависит от типа, размера и массы тела хозяина, а также от иммуногенности вакцины. Удобно готовить вакцину таким образом, чтобы сравнительно небольшие дозировки вакцины (1-5 мл) были достаточными для защиты.
Вакцина также может иметь вид живой рекомбинантной вирусной вакцины, включающей нуклеиновую кислоту, кодирующую полипептид, пептид или вариант. Вакцину вводят хозяину в таком виде, и уже в самом хозяине происходит экспрессия закодированных полипептида, пептидного фрагмента или варианта, создающих защищающую хозяина реакцию.
Известен ряд таких систем живых рекомбинантных вирусных вакцин. Примером подобной системы является система вируса Vaccinia (патент США 4603112 и др., Nature 354, 520 (1991)).
Еще одним своим аспектом изобретением дается способ защиты хозяина, восприимчивого к инфицированию паразитами Echinococcus и Taeniid. Способ изобретения включает в качестве необходимого этапа введение хозяину антигенного полипептида или пептидного фрагмента, или варианта как таковых, либо введение вышеописанной вакцины.
Хотя нижеследующие характерные примеры раскрывают применение антигенов изобретения только в защите от инфицирования Е.granulosus, для специалиста понятно, что применением антигенов, происходящих от одного вида паразитов, может быть достигнута защита от перекрещивающихся видом (см., например, Lightowlers, Acta Leidensia 57, 135-142 (1989)). B связи с этим понятно, что антигены изобретения являются кандидатами в защитные антигены хотя бы от следующих, отличных от Е. granulosus, Eсhinococcus или Taeniid паразитов: E. multilocularis. E. vogelii T. ovis, T. saginata, T. solium, T. multiceps и T. hyoatigena.
В качестве еще одного аспекта изобретения предлагается применение вышеописанной молекулы ДНК или ее субпоследовательности в качестве зонда. Согласно этому аспекту, молекулу ДНК применяют для идентификации путем гибридизации с ДНК Echinococcus или Taeniid паразита, например: T. saginata T. hydatigena или E. multilocularis, кодирующей иммуногенный антиген данного паразита. Этим путем могут быть выявлены дополнительные антигены паразита, пригодные для применения в вакцине.
Способ применения в качестве зонда молекулы ДНК изобретения не требует разъяснения для специалиста. К примеру, может быть применена методика, предложенная и др., "Молекулярное клонирование, лабораторное руководство", Коулд Спринг Харбор (1982).
Или же для выявления гомологичных ДНК других паразитов может быть применен способ амплификации ДНК, например, полимеразной цепной реакцией (Saiki и др. , S'cience 239, 487 (1988)) с ПЦР праймерами, основанными на нуклеотидной последовательности, приведенной на фиг.4.
Аналогично применению молекул ДНК изобретения для выявления ДНК, кодирующих соответствующий защитный антиген других паразитов, антитела-зонды со специфичностью к защитным антигенам изобретения могут быть использованы для отбора антигенов, экспрессируемых микроорганизмами, трансформированными ДНК рассматриваемого паразита. Местоположение положительного клона (клона, экспрессирующего антиген, распознаваемый антителом) позволяет идентифицировать как сам защитный антиген, так и кодирующую его ДНК. Такие антитела-зонды могут быть и поликлональными, и моноклональными, и могут быть получены любым известным способом. В частности, моноклональные антитела могут быть получены по методике Kohler и Milstein (Kohler и Milstein. C. "Непрерывные культуры слитых клеток, секретирующих антитела заданной специфичности", Nature 256, 495-497 (1975)).
Иммуногенность антигенного полипептида изобретения и пептидных фрагментов этого полипептида станет очевидной из следующих, неограничивающих изобретение примеров.
Пример 1
Испытания антигенов, полученных из онкосфер ECHIIOCOCC GRANULOSUS препаративным НДС-ПАГЕ методом
Предшествующие исследования заявителей, о которых не сообщалось, привели в индентификации ряда кандидатов в защитные антигены из E. granulosus. Эти кандидаты в защитные антигены имели следующие молекулярные массы, определенные методом НДС-ПАГЭ:
- 23-25 кД
- 30 кД
- 34 кД
- 40 кД.
Для выявления из вышеприведенных кандидатов защищающего хозяина антигена -(ов) заявителями проведены следующие эксперименты.
Материалы и методы
Препаративный гель. Применением НДС препаративной ячейки (Биорад) в камеру на 32 мм на высоту 5,5 см наливают 50 мл 15%-ного раствора полиакриламида. После отстаивания верхнюю часть заливают 8 мл концентрирующего геля и оставляют отстаиваться.
Антиген. Девять миллионов проклюнувшихся онкосфер получают следующим образом. Подсчитывают яйца E.granulosus с полностью развитыми эмбриоформами и необходимое число яиц (из расчета на 20%-ый выход активированных онкосфер) помещают в разовые пластиковые центрифужные пробирки на 15 мл (Фалькон Пластикс). В каждую пробирку помещают не более 500000. Яйца центрифугируют 2 мин при 20 g, надосадочную жидкость отбрасывают и при 37oС добавляют 10 мл искусственного желудочного сока (ИЖС), пропущенного через мембрану на 0,2 микрона (Неаth D.D. и Smyth J.D. Jn vitro культивирование Echinocoсcus granylosus, Taenia hydatigena T. ovis, T. pisiformis и T. serialis из онкосферы ди цистной личинки, Parasitology 61, 329-343 (1970). Пробирки перемешивают в роторе 1 ч при 37oС.
Яйца центрифугируют 2 мин при 200 g, надосадочную жидкость отбрасывают и заменяют при 37oС 10 мл искусственного желудочного сока (ИЖС) (Hеath и Smyth см. выше), пропущенного через мембрану на 0,2 микрона. Яйца перемешивают 30 мин при 37oС в роторе, после чего центрифугируют 2 мин при 1000 g. Надосадочную жидкость отбрасывают и заменяют 15 мл Перколля (Франция, Швеция), разбавленного асептически в отношении 9:1 (об./об.) 10x концентрированным NCTC 135 (Гибко, Н-Й) эмбриофорные блоки, активированные и неактивированные онкосферы, образующие осадок, смешивают с Перколлем инверсией. Затем пробирки центрифугируют 10 мин при 1000 g и надсадочную жидкость, содержащую онкосферы, из каждой пробирки пипеткой переносят в новые стерильные центрифужные пробирки на 15 мл. Половину надосадочной жидкости (7,5 мл) пипеткой переносят в другую стерильную центрифужную пробирку и в каждую из двух пробирок добавляют по 7,5 мл NCTC 135. Пробирки перемешивают инверсией и центрифугируют 5 мин при 1000 g. Надосадочные жидкости удаляют, а содержащие онкосферы осадки промывают 2 х по 10 мл NСТС 135, центрифугируя каждый раз 2 мин при 1000 g. Наконец осадок суспендируют в соответствующем количестве NСТС с получением в результате образцов по 10 микролитров, содержащих 40-200 онкосфер. Затем определяют число активированных и неактивированных онкосфер подсчетом всего объема 4 х 10 микролитров образцов, нанесенных на обе стороны двух слайд гемоцитометра (Импрувд Найбауэр) при 200-кратном увеличении, позволяющем отличить неактивированные онкосферы от онкосфер, лишенных своих онкосферальных мембран (активированные онкосферы).
Отделение белка онкосфер. Онкосферы (прим. 50% неактивированных и 50% активированных) кипятят в НДС образцовом буфере (Laemmli U.К. Расщепление структурных белков в ходе сборки головки бактериофага Т4, Nafure (Лондон) 277, 680-685 (1970), содержащим дитиотрейтол (ДТТ) (Сигма, 10 мг/мл) вместо меркаптоэтанола, и переносят в колонку НДС препаративной ячейки (Биорад). После того, как фронт красителя бромфенилового голубого почти достигает нижней части колонки, 24 ч отбирают фракции по 2,5 мл.
Ингибиторы ферментов. Каждый поставлен фирмой Сигма. Ингибиторы готовят в 20 мМ Трис-HCl (рН 8) таким образом, что при разбавлении (об./об.) получают следующие конечные концентрации: йодацетамид 20 мМ, апротинин 10 мкл/мл, пепстатин 2 мкг/мл, N-тозил-L-фенилаланин-хлорметилкетон (ТФХК 50 мкг/мл, Nа-п-тозил-L-лизин-хлорметилкетон (ТФХК) 50 мкг/мл, Nа-п-тозил-L-лизин-хлорметилкетон (ТЛХК) 50 мкг/мл этилендиаминтетрауксусная кислота (ЭДТК) 2 мМ, фенилметилсульфонилфторид (ФМСФ) 1 мМ). Ингибиторы добавляют во все фракции. Образцы каждой фракции прогоняют через 5-25% гель методом НДС-ПАГЭ и для выявления молекул соответствующей молекулярной массы гель окрашивают серебром. Отобранные фракции концентрируют до 10 мл в перемешиваемой ячейке на мембране Амикон UМЗ. Затем их центрифугируют 30 мин при 1oС и 2200 об/мин недостаточную жидкость отделяют от осадка НДС и диализуют 3 дня относительно 4 перемен 20 мМ Трис-буферного солевого раствора, содержащего ингибиторы ферментов (рН 7,5).
Подлежащие рассмотрению молекулярные массы были отобраны по молекулярным массам фракций следующим образом: 1. (все молекулы с молекулярной массой ниже 21 кД), 2. (23÷25 кД), 3. (30 кД), 4. (34 кД), 5. (40 кД, дублет), 6. (43 кД), 7. (все молекулы с молекулярной массой выше 43 кД), 8. (смесь всех молекул), 9. (только адъювант), 10. (образец антигена перед его загрузкой на гель. Образец обработан также центрифугированием на холоду и интенсивным диализом).
Методика иммунизации. Каждый антиген представлял собой 10 мл надосадочной жидкости. Каждая фракция представляет конкретную(ые) молекулу(ы) из 5 миллионов онкосфер, в то время как смесь состоит из всех молекул из 2 миллионов, также как и антиген перед загрузкой на гель (группа 10). Для первой инъекции 1 мл антигена гомогенизируют с 1 мл SТМ адъюванта (Bokhout и др. Veterinary Jmmunology and Immunopathology 2, 94-500 (1981), половину животных инъектируют подкожно под левые ребра, а вторую половину инъектируют в левую заднюю ногу. При втором инъектировании состав и путь введения инъекции те же за исключением того, что в каждую инъекцию включают 2 мг высушенных замораживанием Mycobacferium phlei. Инъектирование проводят с правой стороны.
Контрольное заражение яйцами. Через месяц после повторного инъектирования все ягнята получают перорально по 1000 яиц E.granulosus. Яйца хранят 6 недель при 4oС. Яйца извлечены и червей, собранных у экспериментально зарaженных собак по методике Heath D.D. и Lawrence S.B. Ежедневное продуцирование яиц собаками, инфицированными Eсhinococcus granuloss, Archivos de la Hidalidosis 30, 321-328 (1991).
Экспериментальные животные. Используют ягнят пород Ромни и Дорсет, рассеянных по всем группам. Выращивают их в условиях, исключающих заражение E. granulosus и первую инъекцию получают в возрасте 8 месяцев. Во время первого и второго инъектирования для анализа сыворотки у каждого животного с помощью вакуумного заборника отбирают по 10 мл крови, еще раз кровь отбирают после контрольного заражения. Сыворотку отделяют в асептических условиях и хранят при -20oС до момента испытания культивирования in vitro онкосфер или иммуноблоттированием.
Аутопсия. Через шесть месяцев после контрольного заражения берут еще один образец крови, затем ягнят обескровливают после оглушения пистолетом для оглушения скота. Печень и легкие животных удаляют и в них делают тонкие разрезы для определения числа и состояния присутствующих цист. В печени разрезы делают с интервалом в 2-3 мм и каждый разрез тщательно исследуют на глаз. В легких разрезы делают с интервалом в 4 мм цисты определяют пальпированием, а также осмотром на глаз.
Иммуноблоттирование. Антигены онкосферы Eсhinococcus granulosus разделяют методом НДС-ПАГЭ по опубликованной методике (Laemmli, (1970), см. выше) Hames B. D. и Rickwood D. Электрофорез белков на геле, практический подход, ИРЛ. Пресс, Оксфор (1981). Образцы солюбилизируют кипячением 3 мин в 60 мМ Трис-HCl (рН 6,8), 10% глицерина, 2% НДС, 1% ДТТ и 0,05% бромфенол голубого. Разделение проводят на градиенте геля (Т=5-25%) с применением системы вертикального электрофореза (БиоРад, Протеан II) согласно инструкциям изготовителя. Разделенные антигены переносят электрофорезом (Towbin H, Staehelin T. и Gordon J). Электрофоретический перенос белков с полиакриламидных гелей на листы нитроцеллюлозы. Методика и некоторые приложения Pioceding of Nalional Academy of Seience США, 76, 4350-4354 (1979) на нитроцеллюлозную бумагу (0,45 мкм, Шляйхер энд Шулл, Дассель, Германия) в охлаждаемой ячейке Трансблот с плоскими электродами (БиоРад, Ричмонд, США). Перенос осуществляют 2 ч при 50 вольт в 10 мМ карбонатном буфере (рН 9), содержащем 20% метанола (Dunn S. D). Влияние модификации состава буфера переноса и ренатурирования белков в гелях на распознавание белков моноклональными антителами в вестерн блотировании Analytical Biochemistry 157, 144-153 (1986). После переноса нитроцеллюлозные листы помещают в 0,5% Ропссаи, S (ВДН) в 1% уксусной кислоте (5 мин), после чего ополаскивают дистиллированной водой до момента появлении четких полос белка. Маркеры молекулярной массы (Франция) отмечают карандашом. Нитроцеллюлозу, содержащую антиген, нарезают в отдельные полоски и помещают в инкубационные подносы (Шляйхер энд Булл, Германия). Полоски блокируют 1 ч при 37oС добавлением 5% обезжиренного молочного порошка и 0,1% Твин 20 ВДН, растворенные в 20 мМ Трис-HCl (рН 7,5), 0,5% NaCl. Испытуемую сыворотку разбавляют в отношении 1:100 блокирующим буфером. Полоски выдерживают в разбавленной сыворотке в течение ночи при комнатной температуре на вращающей платформе. Затем полоски промывают 4 х 10 мин с заменой 20 мМ Трис-HCl (рН 7,5), содержащим 0,5 М NaCl и 0,1% Твин 20.
Для трансдифферинцировки класса иммуноглобулин IgC1 в IgG2) в иммуноанализ вводят дополнительный этап. После первого антитела полоски зондируют мышинными моноклональными антителами на овечий IgC1 или IgG2, полученными от Др.Кеh Bеh CSIRO МакМастер Лаборэтори, Сидней. Затем полоски зондирует козьим антимышинным IgC1 (полная молекула), меченным пероксидазой (Каппель) и разбавленным в отношении 1: 1000 блокирующим буфером. После этого полоски трижды промывают блокирующим буфером без молочного порошка или Твин 20. Активность пероксидазы определяют на глаз добавлением 3-амино-9-этилкарбазола (Сигма). Проявление прекращают на всех полосках одновременно, в результате чего может быть проведено сравнение интенсивности скрашивания, отражающей количество специфичного иммуноглобулина в каждой сыворотке.
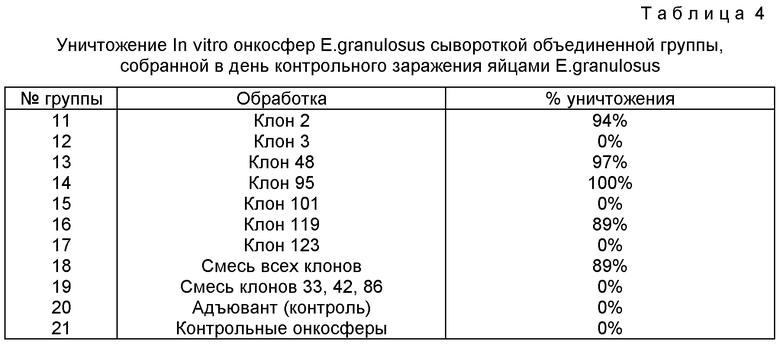
Уничтожение онкосфер in vito. Объединенную сыворотку, полученную перед контрольным заражением, от каждой из 10 групп овец испытывают на способность уничтожать онкосферы.
Для получения комплемента сыворотки плодных ягнят у близких к рождению ягнят стерильным шприцем отбирают кровь, которую сразу же переносят в стерильные полипропиленовые пробирки на 50 мл (Фалькон Пластикс). После свертывания 2 ч при комнатной температуре сгустки отделяют от стенок стерильными аппликаторными палочками в стерильном кабинете с ламинарные потоком. Затем пробирки центрифугируют 30 мин при 4oС в охлаждаемой центрифуге Бекмана. При удалении сыворотки пробирки держат на льду. Сыворотку плодных телят хранят аликвотами по 2 мл при -70oС, пока они не потребуются для культивирования.
Культивирование проводят в плоскодонных планшетах для культивирования на 96 лунок (Фалькон, Микротест III). В каждую лунку вносят 150 мкл испытуемой сыворотки (дезактивируют 30 мин при 56oС) плюс 150 мкл NCTC 135, содержащих 50 активированных онкосфер, плюс 20 мкл комплемента сыворотки плодных ягнят. Планшеты выдерживают при 37oС в инкубаторе с увлажненным СO2 и результаты определяют с помощью Лейтц Лабоверт инверсионного микроскопа спустя 24 ч и 7 дней. Там, где упомянут NСТС 135, в него добавлено 300 мг/л гидрохлорида цистеина (Сигма) и 50 мкг /мл сульфата гентамицина (Шеринг).
РЕЗУЛЬТАТЫ
Характер действия антител против антигена онкосферы, созданного в каждой группе, показан на фиг. 1. Не вызывает сомнений, что различными группами продуцируются антитела, преимущественно распознающие тот интервал молекулярных масс, которыми каждая группа была иммунизирована. В отдельных группах, например, группах I и 5 в иммунизирующих молекулах имелись эпитопы, разделяемые с зпитопами в молекулах иной молекулярной массы.
Результаты аутопсии приведены в таблице I (группы 1-10) показывают, что хотя другие кандидаты антигены и снижают число цист относительно контроля, но только антиген с молекулярной массой в 23-25 кД создает значительную степень защиты (Р=0,012, F=10,62,8 DF).
Результаты in vitro уничтожения онкосфер сывороткой групп овец, иммунизированных фракциями различной молекулярной массы, приведены в таблице 2. Полученные результаты однозначно показывают, что только антиген с молекулярной массой 23-25 кД из Е.granulosus обеспечивает защиту хозяина.
ПРИМЕР 2. АФФИННАЯ ОЧИСТКА АНТИТЕЛ-ЗОНДОВ
Аффинно очищенные антитела из иммобилизованной молекулы в 23-25 кД получают следующим образом. Ягнят 1488 и 1489 четырежды иъектируют подкожно и внутримышечно 100000 активированных онкосфер Е.granulosus. Первые две инъекции разделяет две недели, последующую проводят через 1 месяц и затем через 3 месяца. Через неделю после последней инъекции ягнятам пускают кровь и из 300 мл крови в каждом случае получают 150 мл стерильной "гипериммунной" антисыворотки. В день отбора сыворотки ягнят заражают 3000 свежесобранных яиц Е.granulosus как и двух контрольных ягнят. При аутопсии 6 месяцев спустя иммунизированные ягнята имели цисты только в месте первой инъекции, в то время как контрольные ягнята имели большое количество цист в и печени и легких.
Сыворотка ягненка 1438 использована для аффинной очистки антител, поскольку обладала более высоким титром способности уничтожать онкосферы. Антитела со специфичностью к антигену онкосферы E.granulosus, полученному замораживанием-оттаиванием и обработкой ультразвуком, антигену онкосферы полной полосы в НДС- ПАГЭ, антигенам с относительной подвижностью 0-20, 20-25, 25-30, 30-36, 36-37 кД или к антигену с относительной подвижностью 0-20, 20-30, 30-43, 43-46, 67-90, 90-180 и 180 кД подвергают аффинной очистке с полосокнитроцеллюлозы, содержащих указанные антигены. Очистку проводят по методике Bеall J. A. и M'lchell G.F. Идентификация конкретного антигена из кДНК библиотеки паразита применением антител, полученных аффинной очисткой из выбранных частей вестерн блотoв Journal of Immunological Methods' 86, 217-223 (1986). Белком-носителем служит 1% сыворотка плодного ягненка. Аффинно очищенные антитела промывают и затем применением перемешиваемой ячейки Амикон и мембраны ПМ10 концентрируют в 10 раз.
ПРИМЕР 3. КЛОНИРОВАНИЕ И ЭКСПРЕССИЯ РЕКОМБИНАНТНОЙ ДНК АНТИГЕНОВ ОНКОСФЕРЫ ЕСINOСОСС GRANOLOSOS
1. Выделение и проверка мРНК.
а) Подготовка реактивов
Все растворы, используемые при выделении и обработке мРНК, за исключением растворов, содержащих Трис/Трис (гидроксиметил)амминометан, Сигма, Сан-Луи, МО, США), обрабатывают в течение ночи 0,1% диэтилпирокарбоната (ДЭРК, Сигма) и перед употреблением выдерживают в автоклаве. Всю стеклянную посуду прокаливают в течение ночи при температуре 200oС или выше. Общие растворы Трис получают в воде, обработанной ДЭПК и в автоклаве.
(в) Онкосферы.
Зрелые яйца Е.granulosus получают и активируют проклевыванием по методике, подробно изложенной в примере 1. Примерно по 4х106 онкосфер солюбилизируют на льду в безнуклеазном 6 М гидрохлориде гуанидина (ВР ультрачистый. Бетесда Рисерч Лаборатириз, США). 0,1 М ацетата натрия с помощью тканевого гомогенизатора из стекла и тефлона. Солюбилизированные онкосферы перед выделением РНК хранят при -70oС.
(с) Выделение РНК.
Солюбилизированные онкосферы оттаивают на льду и нерастворимые вещества осаждают центрифугированием 30 минут при 4oС и 10000 об/мин в роторе типа 50Т, и ультрацентрифуге Бекман 18 (Бекман, Пало, Альто, СА, США). Надосадочную жидкость наслаивают над раствором, содержащим 4,8 М хлорида цезия (С СI БРI ультрачистый, Бетесда Рисерч Лабораториз, Гайтерсбург, МД, США) 10 мМ ЭДТК (этилендиаминтетрауксусная кислота, динатриевая соль) в количестве 6,5 мл гуанидина /5,5 мл С СI на полиалломерную пробирку для ротора Бекман SW 40. Пробирки центрифугируют 18 ч при 15oС и 3000 об/мин в SW 40 ротора, ультрацентрифуге Бекмана 18. Надосадочную жидкость отбрасывают, а основание пробирки быстро и осторожно ополаскивают 200 мкл Т10Е1 (10 мМ Трис, 2 мМ ЭДТК, рН 8). Осажденную РНК растворяют в 200 мкл Т10Е1 и вновь осаждают давлением 1: 10 объема 3 М ацетата натрия и 2,5 объема охлажденного льдом 100%-ного этанола, после чего выдерживают в течение ночи при 20oС. Осажденную РНК центрифугируют 15 минут при 4oС в микроцентрифуге, осадок ополаскивают 70%-ным этанолом, центрифугирование повторяют и конечный осадок сушат на воздухе. Затем РНК суспендируют в 50 мкл Т10Е1, и ее количество, определенное по поглощению при длине волны 260 нм, составляет 23 мкг общей РНК. Один микрограмм анализируют на 1,2% формальдегид-агарозном геле (Sambrook J. Fritsch Е. P. Maniatis. Молекулярное клонирование лабораторное руководство, Коулд Спринг Нарбор Лаборэтори Пресс (1989) и находят, что РНК включает интактную рибосомную РНК.
(о) Очистка поли -А+РНК.
Поли -А+ матричную РНК очищают на олиго(оТ)-целлюлозе типа 7 (Фармация ЛКБ Биотехнолоджи, Уппсала, Швеция) согласно инструкциям изготовителя, но особенно по методике Aviv H. Leder P. Очистка биологически активной матричной РНК глобина хроматографией на олиготимидильной кислотной целлюлозе, Proc. Nal.Acad. Sci. 69. 1408-1412 (1972). Элюируемую поли-А+ мРНК осаждают вышеприведенным способом и осадок вновь растворяют в 50 мкл Т10Е1.
(е) Трансляция in vitrо
мРНК аликвотой в 5 мкл транслируют in vitrо использованием лизата кроличьих ретикулоцитов (Amersham, Англия) согласно инструкциям изготовителя в полном реакционном объеме в 25 мкл. Продукт радиометят применением 37,5 мкС 35S-метионина (Amersham Интернейшнл, Amerham, Англия) и исследуют электрофорезом на полиакриламидном геле в 10%-ном акриламидном геле в восстановительных условиях (Laemml U.K.). Расщепление структурных белков в ходе сборки головки бактериофага Т4, Nature 227 680-685 (1971) c детектированием флюорографией с последующей пропиткой геля Амплифай (Amersham, Англия). Продукты трансляции были обильными и самыми разнооброазными по размеру, включая продукты с молекулярной массой более 94 кД.
2. Синтез и клонирование кДНК.
(а) Синтез кДНК.
Тридцать семь микролитров раствора мРНК в Т10Е1 подвергают обратной транскрипции в присутствии α32 Р-дАТФ (Amersham) с применением обратной транскриптазы вируса мышиной лейкемии Молони, а также реактивов и инструкции, прилагаемых к набору синтеза кДНК ZAP (Лот UC206, каталожный 200400, Стратаген, Джолла, США), после синтеза второй цепи аликвотное количество анализируют электрофорезом на 1% щелочном агарозном геле согласно инструкции, прилагаемой к набору синтеза кДНК ZAP, кДНК была неоднородной по размеру, и при сравнении с лямбда - Нind III ДНК маркерами (Промега Корпорейшн, Медисон, ВИ, США) включала поддающиеся обнаружению транскрипту в более, чем 2000 п.о.
(в) Клонирование кДНК
кДНК клонируют в бактериофаговый вектор Uni-ZAP XR (λ-ZAP (Стратаген) в условиях, подробно изложенных в инструкции изготовителя набора для клонирования UNI-ZAP XR (Лот 18, каталожный 237211, Стратаген), кДНК лигируют с вектором без какого-либо отбора кДНК по размеру. Образец кДНК полностью лигируют с 1 мкг векторной ДНК.
(с) Упаковка in vitro бактериофага.
Два микролитра лигированной с вектором кДНК упаковывают в инфекционный фаг применением единственной упаковочной реакции из Gigapack II Gold (Лот 1535, каталожный 200216, Стратаген) согласно инструкциям изготовителя. Полученный фаг титруют на Е.cоli PLK-F'. Остальную легированную кДНК затем упаковывают в двух равнозначных реакциях, продукты трех реакций объединяют и титруют. Полученную библиотеку кДНК клонов (кДНК библиотека) в ZAR титруют в PL K-F', и согласно анализу библиотека содержит примерно 100000 первичных клонов (методика приведена в инструкции к набору для клонирования UNI-ZAP XR, Стратаген).
3. Амплификация кДНК библиотеки.
Библиотеку кДНК целиком амплифицируют на клетках Е. coli PLK-F' при плотности 6000 бляшко-образующих единиц БОЕ на пластинку агара диаметром 150 мм. Амплифицированный фаг собирают в течение ночи в 10 мл SМ (100 мМ NaCl, 8 мМ МgSO4, 50 мМ Трис (рН 7,5), 0,01% мас./об. желатина). Амплифицированную библиотеку объединяют и титруют на клетках Е. coli X1.1-Голубой (Стратаген) в присутствии изопропилтиогалактозида(ИПТТ) и 5-бром-4-хлор-3-индоли-β-D-галактозида (XgAI) согласно инструкции к набору для клонирования IIII-АР ХR (Стратаген).
4.Иммуносрининг кДНК библиотеки
(а) Приготовление антител-зондов.
В иммуноанализе бляшек использовано два типа антител-зондов: сывороточных и аффинно очищенных антител, полученных по методике примера 2. Сывороточные образцы обедняют от "фоновой" реакционноспособности к Е.coli и/или β-галактозидазе следующим образом. Нерекомбинантный λgIII или λZAP наносят почти конфлуэнтно на приемлемый штамм E.coli (как правило, XL-I, Стратаген). Нитроциллюлозные фильтры (Шляйхер энд Шуелл, Дассель, Германия) пропитывают 10 мМ ИПТГ, сушат и после инкубирования пластинок 4 часа при 42oС поверх каждой стороны пластинки накладывают по одному фильтру. Затем пластинки выдерживают в течение ночи при 37oС и еще 30 мин при 4oС. Фильтры удаляют и ополаскивают TNT (150 мМ NaCl, 0,05% Твин 20, 10 мМ Трис, рН 8) и инкубируют в 5% обезжиренного молочного порошка (TNT (BL OTTO) 1 час. Все операции инкубирования в иммуноанализе проводят при комнатной температуре (КТo) на вращающейся платформе. Каждый применяемый в иммуноанализе образец разбавляют TNT плюс 20% сыворотки плодного теленка (FTNT) и инкубируют в чашке Петри, содержащей 20 мл разбавленной сыворотки плюс два нитроцеллюлозных фильтра, приготовленных вышеописанным способом. После инкубирования 2 ч нитроцеллюлозные фильтры удаляют, и аффинно обедненная сыворотка готова к применению в иммуноанализе бляшек.
Антитела-зонды, полученные из сыворотки аффинной очисткой (подробности см. пример 2), не подвергают аффинному обеднения с нерекомбинатными λgIII или λZAP и в иммуноанализе их применяют без разбавления.
(в) Иммуноанализ клонов кДНК библиотеки
В первичном скрининге амплифицированной кДНК библиотеки используют два источника антител. Сыворотку овцы 1488, полученную по методике примера 2, готовят вышеприведенным способом и применяют для выявления кДНК клонов, экспрессирующих антегены онкосферы E. granulosus. кДНК библиотеку наносят на пластинки и подвергают иммуноанализу по стандартным методикам, приведенным в работе Sambrооk и др. (см. выше). Вкратце. Бляшки наносят в чашках Петри диаметром 1500 мм в количестве 5000 БОЕ на пластинку LG агарозы и NZY поверхностного агара (0,9% агара). Применяют ВВ4 и XLI Голубой штаммы E.coli (Стратаген). Клоны, связывающие антитела в испытуемой свечой сыворотке, обнаруживают с помощью кроличьего антиовечьего IgG (тяжелая и легкая цепь), коньюгированного с щелочной фосфатазой (каталожный 313055003, Джексон ИммуноРисерч Лабораториз, Вест Гроув, США). Коньюгат антител разбавляют в отношении 1:2000 в BL ОТТО и инкубируют в количестве 5 мл на фильтр, каждый фильтр по отдельности по 2 ч. Затем фильтры промывают по отдельности шестью переменами TNT в течение 1 ч со вращением между переменами. Положительные бляшки обнаруживают с помощью бромхлориндолилфосфата/нитроголубого тетразолия (БХИФ\ НГТ) в качестве хромогенного субстрата по методике Harlow E., Lana D. Антитела, лабораторное руководство, Коулд Спринг Харбор лаборатори Пресс (1988). В общей сложности бляшки 129 клонов идентифицированы, как дающие предположительно положительные сигналы с сывороткой. Каждую такую бляшку снимают с пластинки агар стерильной пастеровской пипеткой и переносят в 0,5 мл SM. SM надосадочную жидкость от каждого клона, содержащую пассивно элюируемый клон бактриофага, титруют по стандартной методике (Sambrook и др., см. выше) и подвергают повторному скринингу на пластинах диаметром 75 мм и фильтрах при низкой плотности (установочно 50 бляшек на пластинку). Из 129 исходных клонов для 80 подтверждена положительная реакция, приведенная вышеописанным способом с сывороткой овцы 1488.
Аналогичным образом библиотеку подвергают скринингу с антителами, полученными аффинной очисткой по методике примера 2 от овцы 1488. Единственный положительный клон выявлен для фракции 2 сыворотки (соответствует антителам к антигенам онкосферы в 20-25 кД), который обозначен, как клон S2, и один клон собран, как реагирующий с фракцией 3 сыворотки (антигены в 25-30 кД), и обозначен клоном S3. Для обоих клонов подтверждена положительность во вторичном иммуноанализе, приведенном выше.
Каждый клон фага нанесен на единственную пластинку диаметром 150 мм с плотностью 20000 БОЕ/пластинку и общее количество фага собирают способом, приведенным выше для амплификации библиотеки.
(с) Отбор клонов
Из библиотеки клонов на основе результатов иммуноанализа клоны S2, S3, 48, 95, 101, 119 и 123 отобраны для дальнейших испытаний на предмет выявления, способны ли экспрессируемые ими антигены защитить хозяина. Клоны 33, 42 и 86, для которых предшествующими несообщенными исследованиями заявителей установлена неспособность создать статистически значимую защиту, также отобраны для использования в качестве контрольных.
5. Субклонирование и экспрессия кДНК
(а) Получение pBluescript.
Плазмида pBluescript создана фагмидным облегчением из АР общего образца, отобранного для каждого кДНК клона согласно инструкции изготовителя, прилагаемой к набору для клонирования UNI-ZAP XR (Стратаген).
(в) Субклонирование в рСЕХ-ЗЕХ
Векторной системой, выбранной для экспрессии антигенов Е.granulosus с целью испытания вакцины, служат плазмиды серии рGEХ (АМРАД Корпорейшн, Мельбурн, Австралия), экспрессирующие кодируемую паразитом часть молекулы в виде слитого белка у карбоксильного конца Schistosoma japonicum глутатион-S-трансферазы (Smith D.B. Johnson K.S. Очистка в одну стадию полипептидов, экспрессируемых в Fscherichia coli в виде слитых белков с глутатион-трансферазой, Сеnе 67, 31-40 (1988). Чтобы иметь возможность субклонировать непосредственно клонируемые кДНК из λZAP XR вектора, было необходимо модифицировать рGEX 3X вектор с целью вставки Xho I сайта рестрикции в нисходящем направлении от Есо RI сайта рестрикции в рGEX полилинкере. Это было достигнуто вставкой ренатурированных олигонуклеотидов, имеющих следующую последовательность:
5е АAТ ТСА ТАС ТСС АСТ 3'
3' СТ АТС АСС ТСА ТТА А 5'
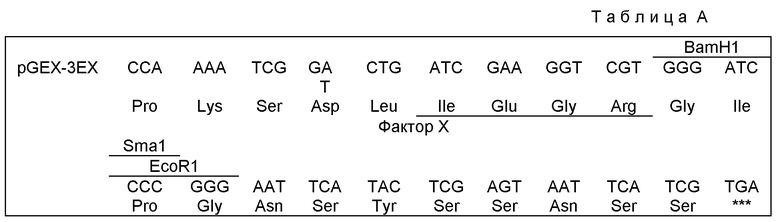
Помимо введения Хho I сайта данный конструкт сохраняет однонаправленное клонирование кДНК, созданное в λZAP XR векторе. Модифицированный вектор получил обозначение рСЕХ ЗEХ. При модификации рGEX вектора использованы обычные методики, подробно описанные у Sambrook и др. (см. выше). Если вкратце, то ренатурированные олигонуклеотиды обрабатывают Т4 Полинуклеотид-киназой (Нью Ингланд Биолабз, Беверли, КА, США), рGEX ЗX ДНК гидролизуют в присутствии Есо RI (Нью Ингланд Биолабз), фосфатизируют и лигируют с обработанными киназой ренатурированными олигонуклеотидами. Применение кальцийхлоридного метода Е. coli JM101 трансформируют лигированной плазмидой. Отбирают трансформированные плазмидой бактериальные клоны, выделяют плазмидную ДНК и модификацию подтверждают секвенированнием соответствующего участка ДНК методом дидезокси-терминации цепи после субклонирования в бактериофаг М13 ВаmHШ (положение 914)/РstI (положение 1885). pGEX 3ЕХ вектор имеет следующую ДНК-последовательность и предсказанную трансляционную аминокислотную последовательность в области полилинкера (см. табл. А).
ДНК плазмиды PВluescript очищают центрифугированием в CgCl (Sambrook и др., см.выше) и вставку кДНК вырезают следующим образом. Примерно 2 мкг каждого клона PВluescript ДНК гидролизуют в присутствии по 10 единиц каждой из EcoRI и XhoI (Амерcхам, Англия) в 20 мкл высокосолевого буфера. Вставку ДНК очищают в 1% агарозы, полосы ДНК вырезают и очищают с помощью Генеклина (БИ0101 Инк., Ла Джолла, США). После количественного определения по поглощению при 260 нм примерно 100 нг вставки ДНК лигируют (2 единицы Т4 ДНК-лигазы, Бозрингер, Маннгейм) с примерно 50 нг очищенного в CSСI, фосфатизированного продукта гидролиза рСЕХ-ЗЕХ ДНК в присутствии обеих EсoRI и ХhoI.
Лигированной рСЕХ-ЗЕХ кДНК (см. ниже) трансформируют E.соli штамм ВВ4 (Стратаген Клонинг системс, Ла Джолла, Калифорния, США каталожный 200269) электропорацией с применением генного пульсатора фирмы ВиоРад согласно инструкции изготовителя (ВиоРад, Ричмонд, CШA).
ПРИМЕР 4. ПРОДУЦИРОВАНИЕ СЛИТОГО БЕЛКА Е.СRANUL OSUS
Отобранные клоны кДНК онкосферы Е.granulosus субклонируют в экспрессионную плазмиду рСEХ-ЗEХ и трансформируют E.coli штамм ВВ4 вышеприведенным способом.
Экспрессией получают белок паразита, слитый с глутатион-S-трансферазой (ГСТ). Такие слитые белки часто растворимы, что позволяет выделять их из лизированных клеток в неденатурирующих условиях путем абсорбции шариками глутатион-агарозы.
Материалы и методы.
GST слитый белок индуцируют и очищают по существу применением методики Smith и Johnson, Gene 67, 31-40 (1988) использованием характерных стадий, подробно изложенных ниже.
1. Колонией рСЕХ трансформанта инокулируют 4 х 100 мл LВ/ампициллин среды, инкубируют в течение ночи при 37oС во встряхиваемом инкубаторе.
2. Культуры разбавляют в отношении 1:10 свежей LB/ампицилин средой и инкубируют 90 мин при 30oС.
3. Добавлением 100 мМ ИПТГ до концентрации 0,1 мМ индуцируют экспрессию.
4. Инкубирование продолжают еще 4,5 часа.
5. Клетки собирают центрифугированием при 5000 х g, 4oС, 10 мин.
6. Клеточный осадок от каждого 1 литра вновь суспендируют 15 мл холодного ФБС и хранят при -20oС/-70oС, пока не потребуется.
7. Осадок оттаивают, добавляют лизозим до концентрации 0,25 мг/мл и инкубируют 10 мин при комнатной температуре.
8. Добавляют Тритон Х-100 до концентрации 0,5%.
9. Клеточную суспензию обрабатывают на льду ультразвуком 3х12 сек импульсами с перерывами в 15 сек.
10. Нерастворимые вещества удаляют центрифугированием при 10000 х g, 4oС, 15 мин.
11. Надосадочную жидкость от 1 литра исходного продукта добавляют к 15 мл 50% шариков глутатион-агарозы и осторожно перемешивают 3 часа при комнатной температуре.
12. Шарики агарозы осаждают центрифугированием при 2000 g, 5 мин при комнатной температуре.
13. Шарики агарозы промывают 5 раз с равным объемом холодного ФБС и центрифугированием при 2000 g, 5 мин, комнатная температура.
14. Выделяют связанный с агарозой CST слитый белок.
15. Концентрацию слитого белка на шариках агарозы определяют белковым анализом Брэдфорда (Биорад).
Пример 5. ДЕМОНСТРАЦИЯ ИММУНОГЕННОСТИ Е.CRANULOSUS СЛИТОГО БЕЛКА.
Овцы во время первой вакцинации достигали возраста 8 месяцев, и вскармливались на пастбищах, свободных от яиц E.granulosus. В экспериментах применялись овцы пород Ромни и Дорсет, которые рассеивались в 9 группах из 5 вакцинированных овец и 1 контрольной группы из 8 овец. Каждому животному вводилось 50 мкг, связанного с агарозой слитого белка (полученного по методике примера 4) в смеси (1:1) солевого раствора: ТА адъюванта (Bokhout B. A. , van Gaalen C. van der Heijden Ph.J.) Подобранная эмульсия типа вода в массле. Состав и применение в качестве иммунологического адъюванта, Veterinary Immunology and Immunpathology 2, 491-500 (1981). Полный объем инъекции в 2 мл/овцу вводят половине животных подкожно, а другой половине - внутримышечно, в обоих случаях с левой стороны. Через месяц инъектирование повторяют с правой стороны. Контрольным животным вводят экспрессированную глутатион-трансферазу из рСЕХ.
Через две недели после второй инъекции у овец берут кровь и готовят сыворотку для культивирования in vitro и для иммублотирования антигена онкосферы по методике примера 1. В тот же день овец заражают 1000 яйцами E. granulosus перорально (см.пример 1).
Аутопсия.
Через шесть месяцев после заражения овец обескровливают после оглушения пистолетом для оглушения скота, легкие и печень удаляют для исследования. В печени с интервалом в 2 мм делают надрезы и тщательно на глаз определяют наличие цист, в легких надрезы делают с интервалом в 4 мм и для определения присутствия цист надрезы пальпируют. В сомнительных случаях надрезы вскрывают для подтверждения присутствия в них цист Е.granulosus.
Результаты
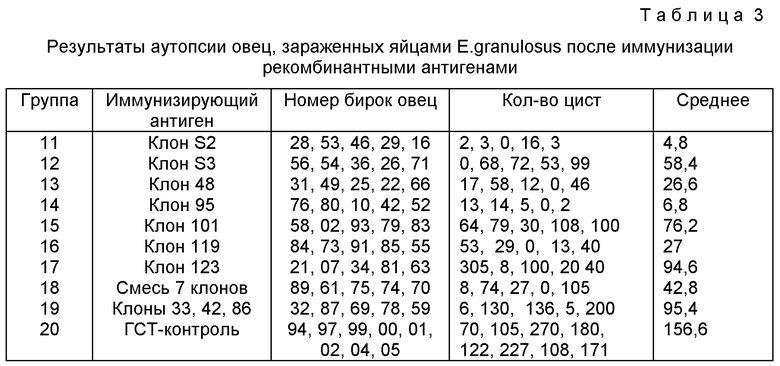
Результаты аутопсии приведены в таблице 3, в таблице 4 приведены результаты уничтожения онкосфер.
Значительную степень иммуности на контрольное заражение стимулируют клон S2 (97%, Р<0,001, F=24,04, II DГ), клон 48 (83%, Р=0,002, F=16,51, NDГ) клон 95 (96%, Р<0,001, F=23,41, NDГ), клон 119 (83%, Р=0,002, II DГ).
Уничтожение in vitro онкосфер наблюдается только в объединенной сыворотке от этих четырех клонов и в случае смеси всех клонов.
Все эти клоны продуцируют антитела, реагирующие с нативным антигеном в 23-25 кД, а два клона (101 и 123), не индуцирующие статистически значимую защиту, реагируют с нативной молекулой в 34 кД. Другой клон, не индуцирующий статистически значимую защиту (S3), реагируют с молекулой в 30 кД (фиг. 2).
Оба клона S2 и 95 индуцируют сильную IgG2 реакцию на молекулу в 23-25 кД по сравнению с менее эффективными клонами 48 и 119. Отдельные овцы в этих группах из пяти овец также усиливают гипотезу с тем, что сильная IgG2 реакция способствует в индуцировании защитной иммуности (фиг. 3).
Небольшие отклонения в результатах для овец 10 овец, иммунизированных клоном S2 или 95, указывает на то, что данная последовательность может распознаваться широким спектром фенотипов.
Пример 6. ЧАСТИЧНОЕ ДНК СЕКВЕНИРОВАНИЕ КЛОНОВ S2, 48, 95, 119
Получение pВluеscript
Фагемидная pВluеscript создана фагемидным облегчением из лямбда-Zар, общего для каждой кДНК согласно инструкции набора для клонирования UNI-ZAP XR (Стратаген).
Получение одноцепочечной ДНК.
Однацепочечная ДНК получена из плазмиды pВluеscript (Стратаген) по методике Ausabel к др. Ред. "Современные методы молекулярной биологии", Грин Паблишинг Ассошиэйт энд Вили Нтерсайенс, Нью-Йорк (1987).
Секвенивование кДНК.
кДНК частично секвенируют терминацией цепи (Sanger F., Nicklen S, Coulson A.R. Саквенирование ДНК ингибиторами терминации цепи, Proc.Nall. Acad. Sci. США 14, 5463 (1977) применением секвеназы, вариант 2 (Юнайтид Стейтс Биокемикл). Драймером служит Т4 (Адьберт Эйнштейн Колидж оф Медисайн, отдел синтеза олигонуклеотидов). Продукты реакций терминации разделяют на 6% полиакриламидном геле и оценивает визуально авторадиографией (Х-Омат, Кодак).
Результаты
Результаты представлены в таблице 5.
Как видно из таблицы 5, каждый клон, для которого показана способность экспрессировать защитные антигены против Е.granulosus включает идентичную молекулу ДНК в 224 п.о. (вышеприведенные нуклеотиды 10-233).
Пример 7. ДНК СЕВЕНИРОВАНИЕ КЛОНА 95
кДНК секвенируют с помощью Т7 Секвенсинг-набора (фармация) с применением CsCI-очищенной плазмидной ДНК и олигонуклетидных праймеров, происходящих рСЕХ последовательности в примерно 30 п.о. в восходящем и нисходящем направлении от Есо RI сайта клонирования. Олигонуклеотиды приготовлены использованием РСR-МАТЕ 391 ДНК синтезатора (Эпплайд Биосистемс, Фестер Сити, КА, США) и химикатов фирмы Эпплайд Биосистемс. Внутренняя последовательность получена применением прямых и обратных олигонукдеотидных праймеров, выявленных из внутренней кДНК-последовательности.
Полная последовательность клона 95 и предсказанная аминокислотная последовательность трансляционного белка приведены на фиг.4. кДНК клона 95 включает вставку в 715 п. о. (не включает нуклеотиды, появившиеся, как часть стратегии клонирования в λZAP). Данная кДНК содержит открытую рамку считывания в 461 п.о., кодирующую предсказанный белок с молекулярной массой 16592 Да.
Следующая ДНК-последовательность включена в полную последовательность клона 95, и ее происхождение определяется стратегией, применяемой при получении кДНК и клонирования в λZAP ХR вектор.
У 5'-конца кДНК:
5'СААТТССССАССАС 3'
у 3' - конца кДНК:
5'ААААААААААААААААААСТССАС 3'
ПОСЛЕДОВАТЕЛЬНОСТЬ с фиг.4 независимо ПОТВЕРЖДЕНА применением методики примера 6.
Промышленное применение.
Согласно настоящему изобретению дается антигенный полипептид, а также активные пептидные фрагменты и варианты такого полипептида, эффективно создающие защитную иммунологическую реакцию на инфицирование Е.granulosus у восприимчивого хозяина. Установлена, что вакцинация данным полипептидом и/или его пептидными фрагментами создает почти полную иммунность против контрольного заражения яйцами Е.granulosus. Изобретением дается биотехнологический способ экспрессирования антигена, которым тот может быть получен в промышленных количествах.
Необходимо подчеркнуть, что вышеприведенное описание дается всего лишь в качестве примера и что могут иметь место вариации, как в применяемых материалах, так и в технологии, известные специалистам.
















Изобретение относится к генной инженерии и иммунологии и может быть использовано в ветеринарии. Описан новый антигенный полипептид, а также его фрагменты и варианты, способные генерировать протективный иммунный ответ при заражении организма Echinococcus granulosus. Определены нуклеотидные последовательности, кодирующие названные формы антигенного полипептида, включение которых в соответствующие векторные системы обеспечивает возможность получения новых антигенов методом рекомбинантных ДНК, а также создание, наряду с вакцинами, содержащими готовый антигенный полипептид, рекомбинантных вирусных вакцин, включающих нуклеиновую кислоту, экспрессия которой осуществляется в организме животного. Предложенные вакцины предназначены для защиты хозяев, подверженных заражению паразитами р. Echinococcus. 5 с. и 2 з.п. ф-лы, 6 табл., 4 ил.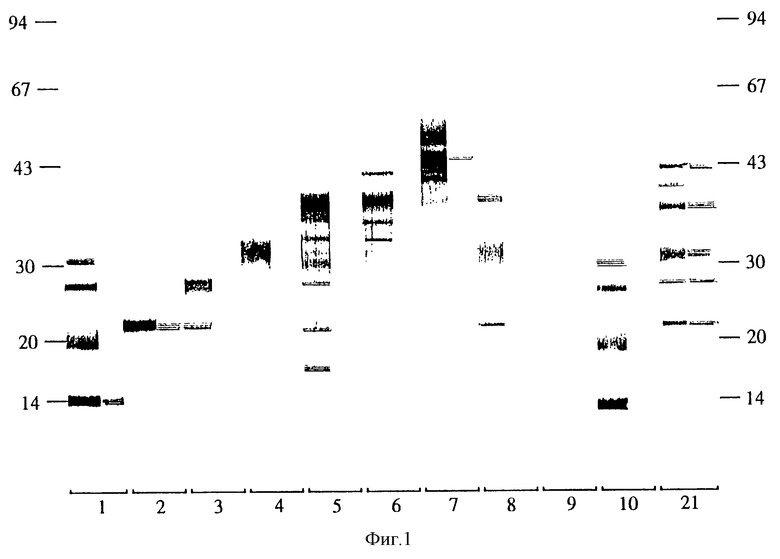
где А' представляет собой последовательность CAG TTA TGT или отсутствует;
В' представляет собой последовательность II (см. графическую часть),
или отсутствует;
или нуклеотидную последовательность, кодирующую вариант указанного полипептида, в котором аминокислотная последовательность модифицирована с помощью вставки, замены или делеции одной или нескольких аминокислот без изменения иммунологической активности.
где А представляет собой GIn-Leu-Cys или отсутствует;
В представляет собой последовательность IV (см. графическую часть),
или отсутствует;
или фрагмент, или вариант указанного полипептида, в котором аминокислотная последовательность модифицирована с помощью вставки, замены или делеции одной или нескольких аминокислот без изменения иммунологического ответа.
где А представляет собой GIn-Leu-Cys или отсутствует, В представляет собой последовательность VI (см. графическую часть),
или отсутствует;
или его фрагмент, или вариант, в котором аминокислотная последовательность модифицирована с помощью вставки, замены или делеции одной или нескольких аминокислот без изменения иммунологического ответа.
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| B | |||
| FACON et al., Molecular and Biochem | |||
| Parasitology, 45, 233-240, 1991. | |||

