Изобретение относится к области ветеринарии, медицины и биотехнологии и может быть использовано для получения и применения противопаразитарных вакцин с целью иммунопрофилактики тканевых гельминтозов сельскохозяйственных животных и человека.
Тканевые гельминтозы наносят существенный вред сельскому хозяйству и представляют угрозу здоровью людей, проживающих в животноводческих регионах. Высокая контагиозность возбудителей, появление штаммов, устойчивых к лекарственной терапии, а также персистирование возбудителей в диких популяциях конечных хозяев поддерживают достаточно высокий инфицированности среди сельскохозяйственных животных и людей (>5,0% животных и >1000000 человек). Основным способом профилактики тканевых гельминтозов является дегельминтизация окончательных хозяев (хищников). Была предложена и вакцинация сельскохозяйственных животных личиночными стадиями паразитов (онкосферами) или же рекомбинантными антигенами паразитов, полученными в E.coli.
Так, из уровня техники известен способ иммунопрофилактики заражения тканевыми гельминтами Echinococcus granulosus и Taenia ovis с помощью вакцин на основе онкосфер данных паразитов [1]. Однако, основными недостатками коммерческого производства таких вакцин являются: опасность заражения персонала яйцами цестод, дороговизна и сложность масштабирования производства таких вакцин.
Из уровня техники известен способ получения вакцин для профилактики цистного эхинококкоза, гидатидоза, ценуроза, цистицеркоза и пр. сельскохозяйственных животных на основе рекомбинантных поверхностных антигенов онкосфер соответствующих паразитов, наработанных в E.coli [2, 3]. Однако продукция рекомбинантных белков в E.coli также обладает достаточно большим количеством недостатков. Так, E.coli способны накапливать продуцируемый белок в тельцах включения или в периплазматическом пространстве (между мембраной и клеточной стенкой), но не способны его секретировать. Также клеточная стенка E.coli в основном состоит из липополисахарида, который представляет собой мощный эндотоксин, способный при введении плохо очищенного препарата вызвать эндотоксический шок и привести даже к гибели вакцинируемого животного. Все это требует трудоемкой, тщательной и дорогостоящей очистки рекомбинантного белка. Более того, система экспрессии и посттрансляционной модификации белков бактерий лишена привычных для эукариотических организмов ферментов (шапероны, гликозилтрансферазы, дисульфидизомераза и т.д.), что зачастую приводит к синтезу белка, который сильно отличается по своей структуре от нативного. Помимо этого, системы экспрессии белков, основанные на E.coli, плохо поддаются масштабированию, что обусловлено чувствительностью E.coli к уровню насыщения культуральной среды кислородом.
Известен способ применения белков кутикулы гельминтов Dirofilaria immitis, генов, кодирующих такие белки, или эпитопов таких белков для получения антигенов с целью создания антигельминтных вакцин [4], однако, в источнике не раскрыты возможные системы экспрессии рекомбинантных антигенов, способы их культивирования, способы очистки и хранения рекомбинантных антигенов, а сам источник описывает использование кутикулярных белков нематод.
Известен способ получения вакцин, основанных на генетически модифицировании штаммах дрожжей (включая роды Saccharomyces, Pichia, Kluyveromyces, Hansenula, Yarrowia) и продуктах на их основе [5], однако, в источнике не раскрыты подходы к дизайну протективных молекул, не раскрыт способ культивирования и состав сред культивирования, способы очистки и хранения рекомбинантных антигенов. Помимо этого, источник раскрывает возможность применения таких вакцин только для иммунопрофилактики вирусных заболеваний.
Известны мультивалентные антигельминтные вакцины и способ их получения (включая, использование генноинженерных штаммов E.coli и P.pastoris) [6], однако, в источнике не раскрыты способ культивирования генноинженерных штаммов и состав сред культивирования, способы получения, очистки и хранения рекомбинантных антигенов. Помимо этого, источник раскрывает возможность применения таких вакцин только для иммунопрофилактики нематод (роды Necator, Ancylostoma, Schistosoma).
Известен способ применения лизатов рекомбинантных дрожжей, в том числе дрожжей родов Saccharomyces, Pichia, Kluyveromyces, Hansenula, Yarrowia для иммунопрофилактики широкого спектра заболеваний, в том числе гельминтозов (без конкретизации типов таких гельминтозов или видов паразитов их вызывающих) [7]. Однако, в источнике не раскрыты подходы к дизайну протективных молекул, не раскрыты способы получения генноинженерных штаммов дрожжей-продуцентов, не раскрыт способ культивирования и состав сред культивирования, способы очистки и хранения рекомбинантных антигенов, способы получения и хранения вакцин на основе рекомбинантных антигенов. Помимо этого, описанный способ не предполагает экспрессии секретируемых форм рекомбинантных антигенов, что сильно затрудняет получение инъекционных форм вакцин.
Патент US 2021/0046170 А1 [8] описывает способ получения иммуногенных композиций для иммунопрофилактики против гельминтозов, вызываемых гельминтами рода Dirofilaria и Onchocerca, в том числе на основе рекомбинантных антигенов, полученных путем ферментации дрожжей Pichia pastoris. Однако, в источнике не раскрыты подходы к дизайну протективных молекул, не раскрыты способы получения генноинженерных штаммов дрожжей-продуцентов, не раскрыт состав сред культивирования, способы очистки и хранения рекомбинантных антигенов, способы получения и хранения вакцин на основе рекомбинантных антигенов. Помимо этого, описанный способ раскрывается возможность применения такого подхода только для иммунопрофилактики гельминтозов, вызываемых гельминтами родов Dirofilaria и Onchocerca.
Известна адъювантная композиция для инъекционных вакцин против тканевых гельминтозов [9], однако в источнике не раскрыты подходы к получению антигенов, способы получения и хранения вакцин их основе. Также данный источник не содержит сведений о протективной роли таких вакцин при тканевых гельминтозах.
Патент RU 2140986 С1 от 07 мая 1993 г. [10] описывает способ получения рекомбинантных антигенов для стимулирования иммунной реакции против «гельминтных паразитов». Однако полный текст патента недоступен, что не позволяет ознакомиться с его содержанием.
Из уровня техники известен способ иммунизации овец вакциной на основе антигенов H110D, Н45, H-gal-GP и O-gal-GP, или их фрагментов, или их предшественников для иммунопрофилактики гельминтозов, вызываемых гельминтами родов Haemonchus, Ostertagia, Trichostrongylus, Chabertia, Oesophagostomum, Hyostrongylus, Nematodirus, Toxocara и Cooperia [11]. Однако в источнике не раскрыты подходы к получению антигенов, способы получения и хранения вакцин их основе. Из общедоступной информации по данному источнику нельзя понять подразумевается ли создание вакцин на основе рекомбинантных антигенов и предлагаются ли в качестве штаммов- продуцентов дрожжи родов Saccharomyces, Pichia, Kluyveromyces, Hansenula или Yarrowia.
Наиболее близким аналогом к заявляемому средству для иммунопрофилактики и технологии его получения является патент US 2020/0010515 А1 (9 января 2020 г.) [12], который описывает технологию создания вакцины против паразита Schistosoma mansoni на основе рекомбинантного антигена Sm14, полученного в дрожжах P.pastoris. Источник описывает подход к дизайну рекомбинантной молекулы Sm14, а также способ культивирования и состав сред культивирования, способы получения и очистки рекомбинантного антигена Sm14. Однако в источнике не приводится данных о составе, способе получения и хранения вакцины на основе рекомбинантного антигена Sm14, а также не приводится данных об ее иммуностимулирующих и протективных свойствах. Помимо этого, данный патент раскрывает возможность применения такого подхода только для иммунопрофилактики гельминтоза, вызываемого Schistosoma mansoni.
Технической проблемой, на решение которой направлено заявляемое изобретение, является способ получения значимых количеств рекомбинантных антигенов тканевых гельминтов, разработка и применение на их основе вакцин для иммунопрофилактики тканевых гельминтозов сельскохозяйственных животных и человека (на примере экспрессии рекомбинантного онкосферального протективного антигена EG95tr (EG95 truncated) возбудителя цистного гидатидоза Echinococcus granulosus).
Раскрытие изобретения. Техническим результатом заявляемого изобретения является высокая протективная активность полученного онкосферального антигена EG95tr (стабильная продукция антител к рекомбинантному антигену в сыворотке крови иммунизированных животных в течение не менее 4 месяцев, а также снижение инфицирования целевых животных тканевыми гельминтами не менее, чем на 78,0%) для иммунопрофилактики тканевых гельминтозов (цистного гидатидоза). Помимо этого, описанный способ получения рекомбинантных протективных антигенов генноинженерного штамма-дрожжей P.pastoris позволяет получать большие количества секретируемого рекомбинантного антигена (0.6 г рекомбинантного протективного антигена и более на л среды ферментации), что облегчает процедуру его очистки, снижает его себестоимость и делает возможным иммунизацию большого количества животных с целью профилактики распространения тканевых гельминтозов.
Полноценное сравнение с прототипом невозможно ввиду отсутствия в открытом доступе экспериментально подтвержденной информации о наличии такой активности в текстовом или графическом виде.
Эффективность заявляемого способа достигается за счет относительной легкости получения штамма-продуцента генномодифицированных дрожжей, его высокой продуктивности, простоте очистки рекомбинантного антигена и его высокой протективной активности для профилактики тканевых гельминтозов (стабильная продукция антител к рекомбинантному антигену в сыворотке крови иммунизированных животных в течение не менее 4 месяцев, а также снижение инфицирования целевых животных тканевыми гельминтами не менее чем на 78.0%).
Технический результат достигается композицией для иммунопрофилактики (вакцина) тканевых гельминтозов, характеризующаяся тем, что в ее основе используется рекомбинантный антиген гельминта, полученный путем ферментации генноинженерного микроорганизма Pichia pastoris, несущего ген искомого протективного антигена и культивируемого на среде со следующим составом: биотин - 2-4 мкг/л; пантотенат кальция - 0.4-0.8 мг/л; фолиевая кислота - 2-4 мкг/л; инозитол - 2-4 мг/л; парааминобензойная кислота - 0.2-0.4 мг/л; пиридоксин - 0.4-0.8 мг/л; рибофлавин - 0.2-0.4 мг/л; тиамин - 0.4-0.8 мг/л; Н3ВО3 - 0.5-1 мг/л; Cu2SO4 - 40-80 мкг/л; KI - 0.1-0.2 мг/л; FeCl3 - 0.2-0.4 мг/л; MnSO4 - 0.4-0.8 мг/л; Na2MoO4 - 0.2-0.4 мг/л; ZnSO4 - 0.4-0.8 мг/л; KH2PO4 - 0.1-0.2%; MgSO4 - 0.05-0.1%; NaCl - 0.01-0.02%; CaCl2*2H2O - 0.0132-0.0264%; (NH4)2SO4 -0.5-1%; СН3ОН - 1,0%; глицерол - 1,0%; гистидин - 0.004%; никотиновая кислота (10 мг/мл) - 0.004-0.008%; натрия гидроксид 10М - 0.0006-0.0012% или аналогичной (YPD, MMY, MGY, ВММ, BMG, BMGY или других сред, близких по составу) в течение 72-120 часов. При этом характер экспрессии рекомбинантного антигена может иметь конститутивный и индуцируемый характер, а сам антиген может быть секретируемым, экспрессируемым внутриклеточно или на поверхности штамма-продуцента. Экспрессируемый антиген может представлять собой любой белок любой стадии развития (яйца, личинка, имаго) тканевого гельминта, его эпитоп или его фрагмент. В качестве метки для детекции и очистки искомого антигена могут быть использованы HisTag, СВР, FLAG, GST, НА, НВН, МВР, S-метку, SUMO, ТАР, TRX, V5 или стрептавидин, прикрепленные к С-, или N-концу белка напрямую или разделенные сайтом специфического протеолиза. В качестве сигнальной последовательности, направляющей экспрессируемый антиген на путь секреции, могут быть использованы природные, модифицированные или искусственные сигнальные последовательности дрожжей родов Saccharomyces, Pichia, Kluyveromyces, Hansenula и Yarrowia. В качестве вектора для доставки ДНК в клетки может быть использован любой экспрессионный вектор, шаттл-вектор или искусственная хромосома для продукции белка в дрожжах родов Saccharomyces, Pichia, Kluyveromyces, Hansenula, Yarrowia. Экспрессия ДНК, кодирующей целевой белок, находится под контролем индуцируемого промотора (АОХ1, CUP1, FLD1, DAS, ICL1, PHO89) или конститутивного промотора (GAP, TEF1).
Технический результат достигается культивированием генноинженерного микроорганизма Pichia pastoris, несущего ген искомого протективного антигена, в среде ферментации, содержащей в качестве добавки экзогенные белки (бычий сывороточный альбумин, овальбумин и пр.), охарактеризованные или не охарактеризованные комплексы белков (куриное яйцо, мясной фарш, мясной бульон, сыворотка крови сельскохозяйственных животных и пр.) растительного, животного или микробиологического происхождения, включающие от 0.2 мг до 150 мг по тотальному белку на л конечной среды ферментации, с введением экзогенного белка или комплекса белков до автоклавирования среды ферментации или после ее автоклавирования при условии соблюдения правил асептики.
Технический результат достигается очисткой белка из среды культивирования методом последовательной ультрафильтрации через фильтры с большим размером пор (например, 50 кДа), пропускающих целевой белок, но задерживающих посторонние белки с высокой молекулярной массой, с последующей ультрафильтрацией через фильтры с малым размером пор (например, 3-5 кДа) для отмывки от пептидов и низкомолекулярных соединений (соли, в т.ч. азид натрия). Перед ультрафильтрацией среда ферментации отделяется от дрожжей P. pastoris и агрегатов денатурировавшего белка путем центрифугирования при 10000g и температуре 4°С в течение 10 минут. Для разбавления среды культивирования перед ультрафильтрацией могут быть использованы растворы 0.05-0.5% карбоната аммония и/или формиата аммония и/или цитрата аммония с азидом натрия или без него.
Технический результат также достигается очисткой искомого рекомбинантного антигена из среды культивирования методами металло-хелатной, ионообменной, аффинной и/или эксклюзионной хроматографии в одну или несколько стадий, с использованием или без использования ультрафильтрации.
Технический результат достигается сохранением очищенного рекомбинантного антигена, полученного по п. 15 или п. 18, путем моментального замораживания в жидком азоте и хранения до момента использования при температуре от минус 20 до минус 193°С или его замораживания при температуре от минус 80°С до минус 18°С с последующим лиофильным высушиванием в 6-10 стадий при давлении 0.013-0.027 кПа, температуре от минус 45°С до минус 25°С и времени лиофилизации составляет не более 24 часов при максимальной загрузке лиофильной сушки.
Технический результат достигается также средством для иммунопрофилактики тканевых гельминтозов, включающим композицию по п. 1 в терапевтически эффективном количестве и вспомогательные компоненты (терапевтически приемлемые добавки, адъюванты, наполнители, изотонические агенты, стабилизаторы и/или консерванты), которое представляет собой концентрированный раствор антигена, замороженный или лиофильно высушенный.
Технический результат достигается также способом иммунопрофилактики тканевых гельминтозов сельскохозяйственных животных или человека, включающим локальное (подкожная или внутримышечная инъекция) или пероральное введение (двукратное или большей кратности с равными интервалами не менее 14 суток) заявляемых композиции или средства, содержащие рекомбинантные антигены яиц или личинок тканевых гельминтов родов Ancylostoma, Brugia, Chabertia, Cooperia, Dictyocaulus, Dirofilaria, Dracunculus, Echinococcus, Fasciola, Haemonchus, Hyostrongylus, Hymenolepis, Loa, Mansonella, Necator, Nematodirus, Oesophagostomum, Onchocerca, Opisthorchis, Ostertagia, Schistosoma, Strongyloides, Taenia, Teladorsagia, Toxocara, Trichinella, Trichostrongylus, Trichobilharzia, Wuchereria и др. - при котором наблюдается формирование стабильной продукции антител к рекомбинантному антигену, наблюдаемых в сыворотке крови иммунизированных животных, в течение 6 месяцев, а также снижение инфицирования целевых животных тканевыми гельминтами не менее, чем на 78.0%.
Осуществление изобретения
Ниже представлено более подробное описание заявляемого изобретения. Настоящее изобретение может подвергаться различным изменениям и модификациям, понятным специалисту на основе прочтения данного описания. Такие изменения не ограничивают объем притязаний.
Тканевые гельминтозы, в частности, цистный эхинококкоз и гидатидоз, ценуроз, трихинеллез и др. являются серьезной проблемой в местах интенсивного развития животноводства и производства сельскохозяйственной продукции во многих странах мира [13]. На сегодняшний день не существует эффективных и дешевых средств профилактики данных заболеваний у сельскохозяйственных животных. Высокий уровень инфицирования сельскохозяйственных животных не только наносит экономический вред животноводству, но и неизбежно сопряжен с учащением случаев заражения человека тканевыми гельминтозами.
Основным методом борьбы с эхинококкозом до сих пор является дегельминтизация собак [13]. Однако данный подход ограничен развитием устойчивости гельминтов к применяемым препаратам. Также он не позволяет охватить популяции диких хищников (бродячие собаки, лисы, волки и т.п.), что в конечном счете не гарантирует выраженного уровня защиты для животных из группы риска (крупный рогатый скот, овцы, козы, свиньи). Одним из наиболее перспективных подходов для профилактики тканевых гельминтозов является использование у сельскохозяйственных животных противопаразитарных вакцин на основе онкосферальных или личиночных антигенов. Доказано, что антигены, приготовленные из онкосфер (яйца) или личинок паразита наиболее эффективны в стимуляции иммунитета против повторной инвазии яйцами T.ovis, T.saginata, Е.granulosus и др. На основе этих данных были предложены вакцины против Е.granulosus и T.ovis, приготовленные из онкосфер, которые были получены от дегельминтизированных собак, зараженных E.granulosus и T.ovis, соответственно [1]. Однако, основными недостатками коммерческого производства таких вакцин являются: опасность заражения персонала яйцами цестод, дороговизна и невозможность получения достаточного количества взрослых гельминтов от собак. Альтернативой таким вакцинам могут служить препараты на основе рекомбинантных антигенов, полученных методами генной инженерии. В настоящее время существует несколько вакцин для профилактики цистного эхинококкоза, гидатидоза, ценуроза, цистицеркоза и пр. сельскохозяйственных животных на основе рекомбинантных поверхностных антигенов онкосфер соответствующих паразитов, наработанных в E.coli. Однако продукция рекомбинантных белков в E.coli также обладает достаточно большим количеством недостатков. Так, E.coli способны накапливать продуцируемый белок в тельцах включения или в периплазматическом пространстве (между мембраной и клеточной стенкой), но не способны его секретировать. Также клеточная стенка E.coli в основном состоит из липополисахарида, который представляет собой мощный эндотоксин, способный при введении плохо очищенного препарата вызвать эндотоксический шок и привести даже к гибели вакцинируемого животного. Все это требует трудоемкой, тщательной и дорогостоящей очистки рекомбинантного белка. Более того, система экспрессии и посттрансляционной модификации белков бактерий лишена привычных для эукариотических организмов ферментов (шапероны, гликозилтрансферазы, дисульфидизомераза и т.д.), что зачастую приводит к синтезу белка, который сильно отличается по своей структуре от нативного. Помимо этого, системы экспрессии белков, основанные на E.coli, плохо поддаются масштабированию, что обусловлено чувствительностью E.coli к уровню насыщения культуральной среды кислородом. Системы экспрессии рекомбинантных антигенов для профилактики тканевых гельминтозов на основе E.coli являются наиболее близкими аналогами предлагаемому изобретению.
В качестве альтернативы системе экспрессии в E.coli в последнее время все больше получают развитие системы экспрессии рекомбинантных белков на основе дрожжей Pichia pastoris. Они обладают способностью осуществлять посттрансляционные модификации экспрессируемых белков, а после секретировать их. Они не содержат эндотоксина или его аналогов. Дрожжи P.pastoris способны расти на простой относительно дешевой культуральной среде, а их культуры легко поддаются масштабированию. Эти преимущества позволяют относительно дешево получить достаточно большие количества чистого рекомбинантного белка, в частности, протективного антигена. На основе системы экспрессии P.pastoris были получены высокоэффективные штаммы-продуценты альбумина и кормового белка, однако продукция антигенов для профилактики тканевых гельминтозов в данной системе экспрессии пока остается должного без внимания.
Сутью изобретения является средство на основе рекомбинантных протективных антигенов гельминтов для иммунопрофилактики тканевых гельминтозов сельскохозяйственных животных и человека (т.е. антигельминтная вакцина), способ его получения и применения.
Вакцина против тканевых гельминтозов сельскохозяйственных животных или человека представляет собой конъюгат иммуностимулирующего носителя (адъювант) и протективного (например, онкосферального) рекомбинантного антигена, полученного из среды культивирования штамма-продуцента P.pastoris. Поскольку продуцируемый белок является мажорным в среде культивирования (более 90.0%), а среда культивирования изначально содержит лишь низкомолекулярные вещества, то необходимым и достаточным для очистки антигена является пропускание среды культивирования через систему ультрафильтрации (для обогащения целевой белковой фракции и очистки ее от низкомолекулярных соединений среды культивирования).
В качестве иммуностимулирующего носителя (адъюванта) используют растворы минеральных соединений, масляные эмульсии (адъювант Фрейнда, его модификации и аналоги), компоненты бактериального происхождения (стенки бактериальных клеток, ДНК бактерий и т.п.) и адъювантно активные субстанции (PEI, CRL-8041, Твин80 и т.п.).
Установлено, что наиболее эффективной композицией вакцины являются: протективный антиген - (не менее 100 мкг) в сочетании с иммуностимулирующим носителем (адъювантом) на кг вакцинируемого животного при интраназальной двукратной иммунизации с интервалом не менее 10-14 суток. Установлено, что наиболее эффективной композицией вакцины являются: протективный антиген (не менее 40 мкг) в сочетании с иммуностимулирующим носителем (адъювантом) на кг вакцинируемого животного при инъекционной (подкожной) двукратной иммунизации с интервалом не менее 10-14 суток.
Для более наглядной иллюстрации предлагаемого способа получения рекомбинантного антигена с помощью дрожжей P.pastoris с целью создания вакцин для иммунопрофилактики тканевых цестодозов в качестве примера рассматривается получение рекомбинантного протективного онкосферального антигена EG95tr Echinococcus granulosus для иммунопрофилактики развития ларвальных цестодозов (в частности, цистного гидатидоза), однако, данный способ получения рекомбинантного протективного антигена может быть применен для создания вакцин против любых других тканевых гельминтозов (ценуроза, цистицеркоза, трихинеллеза и др.), а в качестве протективного антигена могут быть экспрессированы любые поверхностные или секретируемые антигены гельминта.
Примеры конкретного исполнения Пример 1. Получение штамма-продуцента дрожжей P.pastoris, обеспечивающего секретируемую продукцию рекомбинантного антигена для создания вакцин (на примере протективного онкосферального антигена Ecchinococcus granulosus EG95tr).
Поскольку существует несколько штаммов паразита с отличающимися аминокислотными последовательностями протективного антигена, выбирают штамм паразита с наиболее универсальной последовательностью антигена либо штамм паразита, причиняющий наибольший экономический ущерб. В примере данного изобретения путем сравнения аминокислотных последовательностей протективного онкосферального антигена EG95 наиболее распространенного штамма E.granulosus G1 (овечий штамм) была выбрана наиболее универсальная аминокислотная последовательность [14]. Исходная аминокислотная последовательность была следующей:

В результате анализа приведенной аминокислотной последовательности в программах SignalP 4.1 Server [15] и ТМНММ Server v. 2.0 [16] было обнаружено, что последовательность MAFQLCLILFATSVLA является сигнальной (определяет компартментализацию данного белка в клетке онкосферы Е. granulosus), а последовательность MTSGSALTSAIAGFVFSCIVVVL представляет собой трансмембранный домен, заякоривающий белок EG95 в цитоплазматической мембране. Для экспрессии растворимого белка в гетерологичной системе (штамм Pichia pastoris) нативный сигнальный пептид EG95 (MAFQLCLILFATSVLA) был заменен на сигнальный пептид альфа-феромона дрожжей S.cerevisiae MRFPSIFTAVLFAASSALAAPVNTTTEDETAQIPAEAVIGYSDLEGDFDVAV LPFSNSTNNGLLFINTTIASIAAKEEGVSLEKREAEA, который направляет белок на путь секреции и при этом отщепляется протеазами Kex2 и Ste13 комплексе Гольджи штамма-продуцента, не влияя на конечную последовательность белка. Вместо трансмембранного домена к С-концу белка через адаптер GGG была прикреплена последовательность HHHHHHHHHHHH, которая представляет собой полигистидиновую метку, позволяющую детектировать белок (при помощи моноклональных антител к полигистидиновой метке) и очищать его на металло-хелатной колонке. Поскольку трансмембранный и сигнальный пептид нативного белка EG95 не содержат иммуногенных эпитопов, они были удалены из рекомбинантного белка. Итоговая аминокислотная последовательность белка EG95tr (EG95 truncated), предназначенная для экспрессии в штамме дрожжей P.pastoris представляет собой:
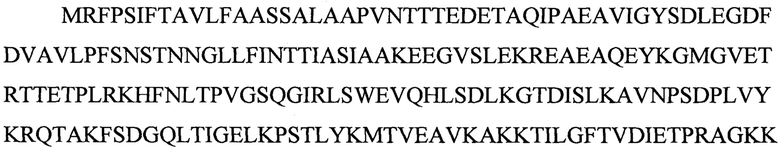
ESTVMGGGHHHHHHHHHHHH, однако в результате процессинга в комплексе Гольджи секреции подвергался белок со следующей последовательностью:

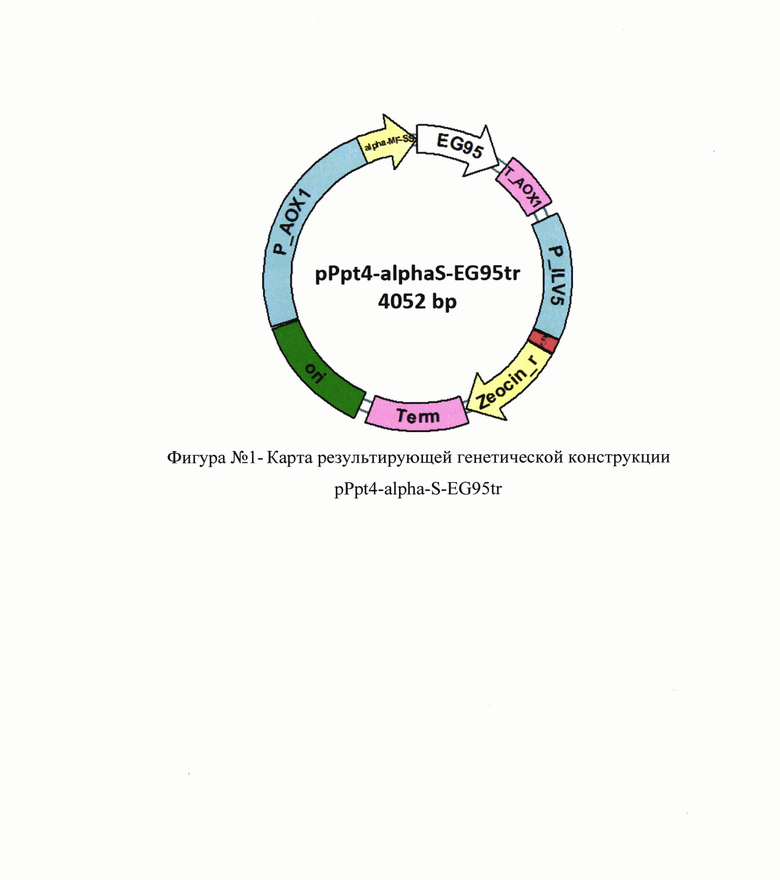
На основе полученной аминокислотной последовательности рекомбинантного антигена делали дизайн последовательности ДНК, которую клонировали в шаттл-вектор для доставки генов в клетки дрожжей. Для конверсии аминокислотной последовательности в последовательность ДНК использовали таблицу частоты распределения ко донов в геноме P.pastoris [17] при этом избегают образования нежелательных сайтов рестрикции и регуляторных элементов генома (промоторы, терминаторы, сигналы полиаденилирования, сайты сплайсинга мРНК и т.п.). Также стремились к получению ДНК, которой соответствует мРНК с максимально простой вторичной структурой. Итоговая последовательность ДНК (см. в приложении. Перечень последовательностей, SEQ ID NO: 1) была собрана из синтетических олигонуклеотидов согласно стандартному протоколу. Полученную последовательность ДНК встраивали в вектор pPpt4-alpha-S по сайтам рестрикции XhoI-NotI. В результате был получен готовый вектор, предназначенный для экспрессии протективного антигена EG95tr в культуре дрожжей P.pastoris (см. в приложении. Перечень последовательностей, SEQ ID NO: 2).
Карта результирующей генетической конструкции pPpt4-alpha-S-EG95tr приведена на фигуре №1 (прилож.). Полученную генетическую конструкцию доставляли в штамм дрожжей P.pastoris методом электропорации. Компетентные клетки для модификации получали повторными отмывками клеток ночной культуры P.pastoris ледяным раствором 1М сорбитола в комбинации с ценрифугированием. Смесь компетентных клеток и полученной генетической конструкции подвергали воздействию электрических импульсов с параметрами U=1500 В, С=25 мФ, R=200 Ом и после инкубации в течение часа в 1М сорбитоле при 30°С модицированные клетки P.pastoris высевали на чашки с агаризованной средой YPD (1,0% пептон, 1,0% D-глюкозы, 0.5% экстракта дрожжей, 1,0% агара) с разными концентрациями зеоцина (от 100 до 1000 мкг/мл). Чашки инкубируют в течение 48-72 часов при 30°С до появления колоний.
С каждой чашки с агаризованной средой YPD с зеоцином отбирали по 3 колонии, которые растирали на новые чашки с агаризованной средой YPD (клетки P.pastoris слипаются в ходе электропорации). Чашки инкубировали в течение 48-72 часов при 30°С до появления колоний.
Для отбора лучшего клона-продуцента проводили тестовую ферментацию. Для этого с каждой чашки с трансформированными P.pastoris скалывали по одной колонии и культивировали в индивидуальных пробирках со средой культивирования MGY (биотин - 4 мкг/л; пантотенат кальция - 0.8 мг/л; фолиевая кислота - 4 мкг/л; инозитол - 4 мг/л; парааминобензойная кислота - 0.4 мг/л; пиридоксин - 0.8 мг/л; рибофлавин - 0.4 мг/л; тиамин - 0.8 мг/л; Н3ВО3 - 1 мг/л; Cu2SO4 - 80 мкг/л; KI - 0.2 мг/л; FeCl3 - 0.4 мг/л; MnSO4 - 0.8 мг/л; Na2MoO4 - 0.4 мг/л; ZnSO4 - 0.8 мг/л; KH2PO4 - 0.2%; MgSO4 - 0.1%; NaCl - 0.02%; CaCl2*2H2O - 0.0264%; (NH4)2SO4 - 1,0%; глицерол - 1,0%; гистидин - 0.004%; никотиновая кислота (10 мг/мл) - 0.008%; натрия гидроксид 10М - 0.0012%) в течение 16-18 часов, затем клетки дрожжей осаждали центрифугированием, а среду меняли на среду культивирования с содержанием метанола MMY (биотин - 4 мкг/л; пантотенат кальция - 0.8 мг/л; фолиевая кислота - 4 мкг/л; инозитол - 4 мг/л; парааминобензойная кислота - 0.4 мг/л; пиридоксин - 0.8 мг/л; рибофлавин - 0.4 мг/л; тиамин - 0.8 мг/л; Н3ВО3 -1 мг/л; Cu2SO4 - 80 мкг/л; KI - 0.2 мг/л; FeCl3 - 0.4 мг/л; MnSO4 -0.8 мг/л; Na2MoO4 - 0.4 мг/л; ZnSO4 - 0.8 мг/л; KH2PO4 - 0.2%; MgSO4 - 0.1%; NaCl - 0.02%; CaCl2*2H2O - 0.0264%; (NH4)2SO4 - 1,0%; СН3ОН - 1,0%; гистидин - 0.004%; никотиновая кислота (10 мг/мл) - 0.008%; натрия гидроксид 10М - 0.0012%). Пробирки культивировали в инкубаторе-шейкере при 30°С в течение 96 часов, добавляя 1,0% метанола каждые 24 часа и восполняя испарение стерильной дистиллированной водой.
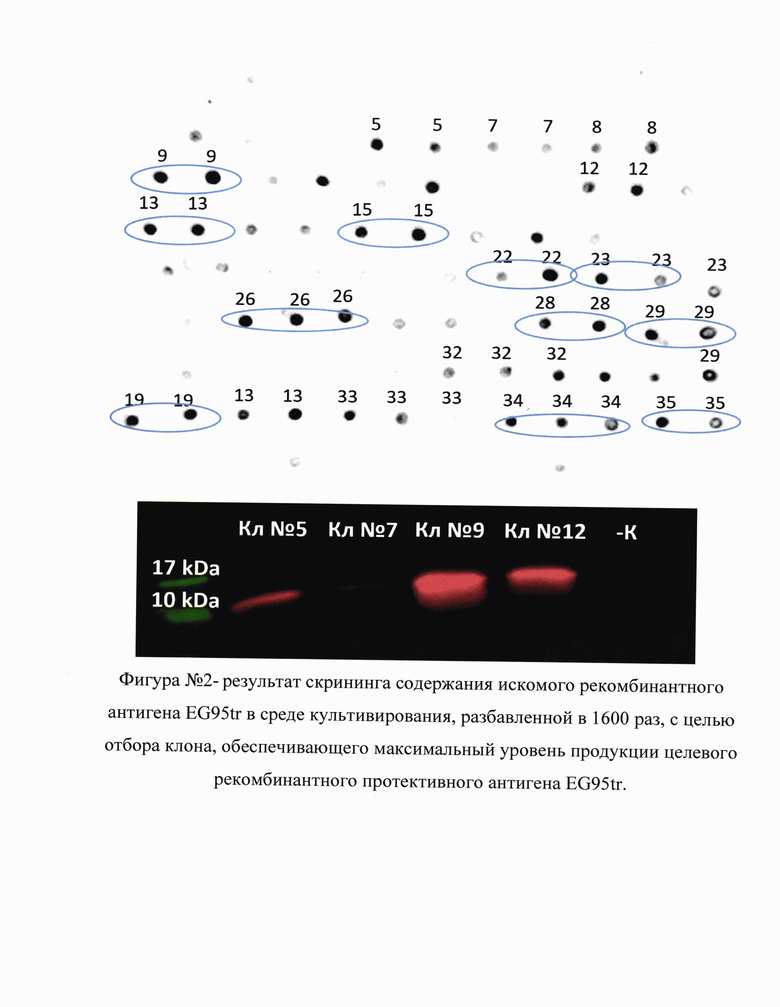
Полученную среду культивирования отделяют от осадка дрожжей центрифугированием и подвергают анализу с помощью стандартных биохимических методик Western Blot или Dot Blot. Наличие целевого рекомбинантного антигена в среде культивирования подтверждали наличием специфического сигнала на мембране, окрашенной антителами к целевому белку или метке. Первичные антитела для идентификации EG95tr и EG95 были специфичны к полигистидиновой метке (Abcam, #ab18184), поскольку коммерчески доступных антител к эпитопам EG95 на момент проведения исследования не существовало.
С помощью методов Dot Blot и Western Blot отбирали клоны с наиболее интенсивным сигналом - такие клоны обладали наибольшей продукцией целевого белка-антигена. На фигуре №2 (прилож.) приведен пример результатов скрининга содержания искомого рекомбинантного антигена EG95tr в среде культивирования, разбавленной в 1600 раз, с целью отбора клона, обеспечивающего максимальный уровень продукции целевого рекомбинантного протективного антигена EG95tr. Верхняя панель - Dot Blot анализ среды культивирования отдельных клонов с целью установления таковых с наибольшим уровнем продукции EG95tr. Нижняя панель - Western Blot анализ среды культивирования клонов P.pastoris, отобранных по результатам Dot Blot анализа, для подтверждения идентичности полученного рекомбинантного белка EG95tr. Первичные антитела специфичны к полигистидиновой метке (Abcam, #ab18184). Перспективные клоны-продуценты сохраняли в YPD с 15-20,0% глицерола.
Пример 2. Подтверждение идентичности полученного белка искомому онкосферальному рекомбинантному антигену Echinococcus granulosus EG95tr и его наработка в препаративных количествах
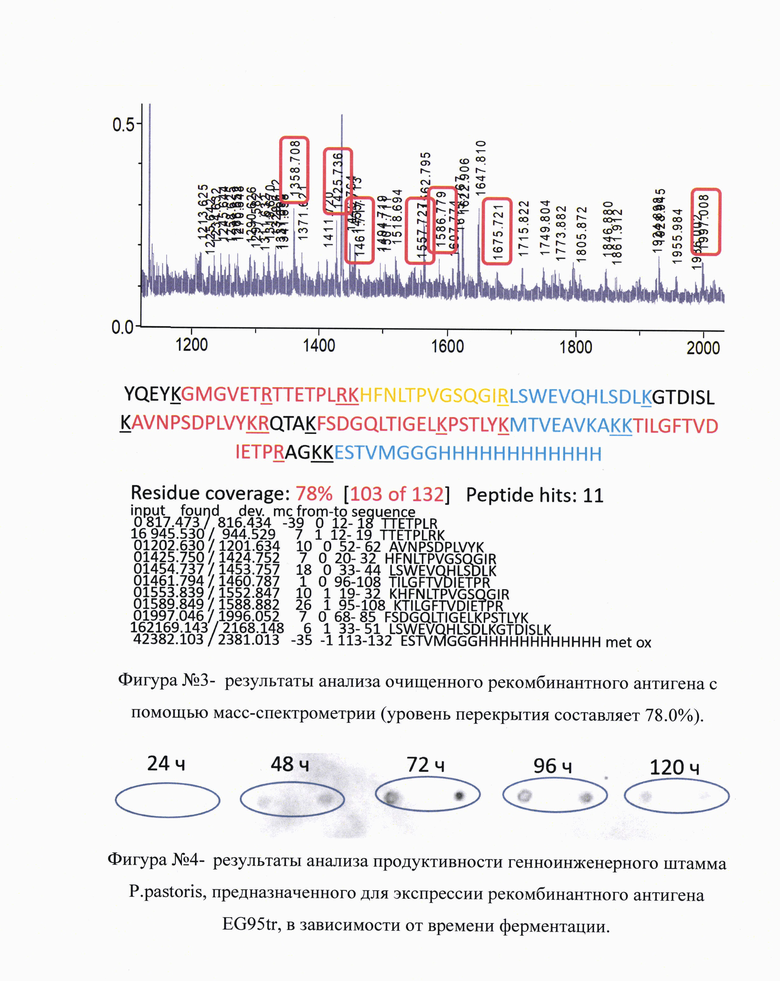
Идентичность полученного рекомбинантного антигена искомому белку дополнительно подтверждали с помощью масс-спектрометрии. Для этого проводили денатурирующий электрофорез белка в полиакриламидном геле. Полученный гель окрашивали в соответствии со стандартным протоколом по Кумасси и вырезали окрашенный фрагмент геля с белком соответствующей молекулярной массы. Полученный фрагмент геля подвергали воздействию трипсина в течение 24 часов при 37°С. Получившиеся пептиды очищали из геля и подвергали анализу на масс-спектрометре. Удовлетворительным результатом считали перекрытие детектированных фрагментов с теоретически предсказанными на 50,0% и более, на фигуре №3 (прилож.) приведены результаты анализа очищенного рекомбинантного антигена с помощью масс-спектрометрии (уровень перекрытия составляет 78.0%). Приведены последовательности идентифицированных пептидов, их пики и локализация в молекуле рекомбинантного антигена EG95tr.
Для получения препаративного количества полученного рекомбинантного антигена EG95tr проводили ферментацию в ферментере. Небольшое количество замороженного образца перспективного клона высаживали на чашку с агаризованной средой MGY и культивировали в течение 48 часов при 30°С. Полученную биомассу переносили в жидкую среду MGY объемом, составляющим 10,0% от объема ферментера, и культивировали в шейкере-инкубаторе в течение 16-24 часов при 30°С.
Полученной культурой инокулировали ферментер со средой MGY, а через 24 часа ферментации среду меняли на среду культивирования MMY. После этого ферментацию осуществляли в течение 96-120 часов при интенсивных аэрации и перемешивании, периодически добавляя метанол и концентрат питательной среды. Каждые 24 часа отбирали пробы для последующего определения оптимального времени ферментации методами Western Blot или Dot Blot. При продукции протективного онкосферального антигена Ecchinococcus granulosus EG95tr оптимальным временем ферментации оказался интервал в диапазоне 72-96 часов. На фигуре №4 (прилож.) приведены результаты анализа продуктивности генноинженерного штамма P.pastoris, предназначенного для экспрессии рекомбинантного антигена EG95tr, в зависимости от времени ферментации. Экспериментально установлено, что оптимальное время ферментации - 72-96 часов (каждая временная точка проанализирована в 2 повторах). Первичные антитела специфичны к полигистидиновой метке (Abcam, #ab18184). Дальнейшая ферментация не приводила к увеличению концентрации белка EG95tr в среде культивирования.
Пример 3. Очистка и консервация протективного онкосферального антигена Echinococcus granulosus EG95tr из среды культивирования
Очистку протективного онкосферального антигена Echinococcus granulosus EG95tr из среды культивирования осуществляли путем ультрафильтрации. Перед ультрафильтрацией среду ферментации отделяли от дрожжей P.pastoris и агрегатов денатурировавшего белка путем центрифугирования при 10000g и температуре 4°С в течение 10 минут. После этого ферментационную среду, содержащую целевой белок, разбавляли в 10 раз холодным раствором, содержащим 0.05-0.5% ацетата аммония и 0.1-0.2% азида натрия, и фильтровали через фильтр, размер пор которого свободно пропускает целевой белок, но задерживает высокомолекулярные белки, продуцируемые P.pastoris в среду культивирования. Для очистки EG95tr (17-18 кДа) используют фильтр Sartorius™ Vivaflow™ 50 с размером пор 50 кДа (Sartorius, #VFS502) или аналогичный.
Полученный фильтрат пропускали через фильтр, размер пор которого задерживает целевой белок EG95tr (17-18 кДа), но пропускает низкомолекулярные белки, продуцируемые P.pastoris, пептиды и низкомолекулярные соединения (соли, в т.ч. азид натрия). Для концентрирования EG95tr (17-18 кДа) и отмывки от солей среды культивирования и азида натрия используют фильтр Vivaflow® TFF Cassette, PES с размером пор 3 кДа (Sartorius, #VF20P1) или аналогичный. Результаты анализа полученных фракций свидетельствуют о том, что искомый рекомбинантный проходит через фильтр с размером пор 50 кДа, но не проходит через фильтр с размером пор 3 кДа и концентрируется во фракции 3-50 кДа.
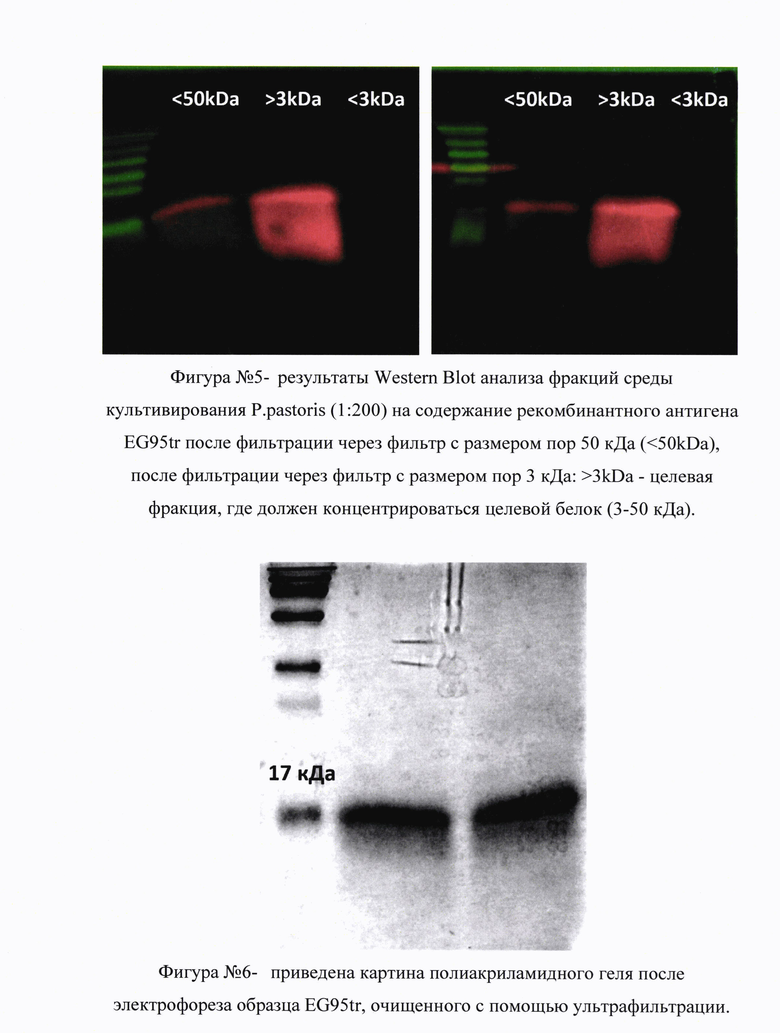
На фигуре №5 (прилож.) приведены результаты Western Blot анализа фракций среды культивирования P.pastoris (1:200) на содержание рекомбинантного антигена EG95tr после фильтрации через фильтр с размером пор 50 кДа (<50kDa), после фильтрации через фильтр с размером пор 3 кДа:>3kDa - целевая фракция, где должен концентрироваться целевой белок (3-50 кДа); <3kDa - проскок, прошедший через фильтр. Полученные результаты свидетельствуют о том, что искомый рекомбинантный проходит через фильтр с размером пор 50 кДа, но не проходит через фильтр с размером пор 3 кДа и концентрируется во фракции 3-50 кДа. Первичные антитела специфичны к полигистидиновой метке (Abcam, #ab18184). Электрофорез полученного белкового образца в полиакриламидном геле с последующей окраской геля Coomassie brilliant blue g-250 показал, что искомый протективный антиген EG95tr представляет собой мажорный белок во фракции 3-50 кДа после очистки с помощью ультрафильтрации (не визуализируются другие белки в данной фракции). Полученный образец белка был вырезан, трипсинизирован, выделен из геля и отправлен на повторный масс-спектрометрический анализ, который подтвердил его идентичность искомому EG95tr антигену.
На фигуре №6 (прилож.) приведена картина полиакриламидного геля после электрофореза образца EG95tr, очищенного с помощью ультрафильтрации. Гель окрашен Coomassie brilliant blue g-250, который неспецифически окрашивает все белки. Полученные результаты свидетельствуют о том, что искомый протективный антиген EG95tr представляет собой мажорный белок во фракции 3-50 кДа после очистки с помощью ультрафильтрации. Полученный образец белка был вырезан, трипсинизирован, выделен из геля и отправлен на масс-спектрометрический анализ. Очищенный (обогащенный) антиген EG95tr замораживали в жидком азоте и лиофилизировали. В результате ферментации и очистки методом ультрафильтрации на каждый литр исходной среды ферментации удалось получить 0.6 г лиофилизированного белкового препарата, содержащего не менее 90,0% рекомбинантного белка EG95tr.
Пример 4. Оценка стабильности полученного рекомбинантного онкосферального антигена Echinococcus granulosus EG95tr в зависимости от условий хранения
Была изучена стабильность полученного рекомбинантного онкосферального антигена Echinococcus granulosus EG95tr в зависимости от условий хранения. В результате исследования было установлено, что повторные циклы замораживания-оттаивания приводят к снижению содержания белка в растворе.
На фигуре №7 (прилож.) приведены результаты анализа стабильности рекомбинантного протективного антигена EG95tr после двух циклов замораживания-оттаивания его раствора с помощью метода Western Blot (первичные антитела специфичны к полигистидиновой метке - Abcam, #ab18184). Видно, что с каждым циклом замораживания-оттаивания уменьшается интенсивность специфичного сигнала, соответствующего белку EG95.
Даже один цикл замораживания-оттаивания приводит к значительной деградации EG95tr (около 50,0%): образец «после замораживания» дает более слабый сигнал при окрашивании мембраны (метод Western Blot) специфическими антителами по сравнению с образцом «до замораживания», что отражает меньшее количество искомого белка EG95tr.
Лиофилизация образца, напротив, обеспечивает его высокую сохранность в течение минимум года, даже в условиях хранения при 4°С.
На фигуре №8 (прилож.) приведены результаты Western Blot анализа стабильности рекомбинантного протективного антигена EG95tr после одного цикла лиофилизации-восстановления через год после лиофилизации. Первичные антитела специфичны к полигистидиновой метке (Abcam, #ab18184).
Пример 5. Увеличение продуктивности ферментации штамма-продуцента P.pastoris путем введения в среду культивирования экзогенных белков, охарактеризованных или неохарактеризованных белковых препаратов
Одной из наиболее важных задач с целью увеличения общего выхода секретируемого белка в культуре P.pastoris является его стабилизация в среде культивирования после секреции. Разрушение целевого белка в среде культивирования может быть обусловлено как нестабильностью самого белка, так и действием протеаз, секретируемых клетками P.pastoris с целью обеспечения растущей культуры аминокислотами. Для увеличения времени жизни целевого белка в среде культивирования было предложено культивирование при пониженных значениях кислотности среды питательной среды (рН), при которых основные протеазы, продуцируемые P.pastoris, малоактивны [18]. Также существуют штаммы P.pastoris с выключенной экспрессией некоторых из протеаз (SMD1168, SMD1169). Однако было показано, что продуктивность в штаммах SMD1168 и SMD1169 зачастую не выше, а ниже, чем в исходном штамме [19]. Предположительно это обусловлено недостаточным снабжением растущей культуры аминокислотами. Понижение же рН питательной среды может негативно сказаться на стабильности и растворимости целевого белка. Еще одним подходом, направленным одновременно на улучшение питания культуры аминокислотами и увеличение стабильности секретируемого белка, является добавление в среду культивирования так называемых Кислот Касамино [18]. Однако данная добавка к среде культивирования является дорогостоящей, что снижает коммерческую эффективность биотехнологического производства при ее использовании.
Нами было установлено, что введение экзогенного белка, охарактеризованного или неохарактеризованного белкового препарата в среду культивирования, многократно повышает эффективность ферментации и на 1-2 порядка (в 10-100 раз) увеличивает содержание в среде культивирования целевого рекомбинантного протективного антигена. Эффективность данного подхода была опробована нами на примере продукции рекомбинантного протективного антигена Echinococcus granulosus EG95tr.
В качестве источников экзогенных белков, вводимых в среду ферментации, были использованы:
1. среда культивирования клеток млекопитающих DMEM (Dulbecco's Modified Eagle Medium), содержащая 10,0% фетальной бычьей сыворотки (50,0% об./об. от конечного объема среды ферментации);
2. бычья фетальная сыворотка (0.33%, 1,0%, 2.5%, 5,0%, 10,0% об./об. от конечного объема среды ферментации);
3. бычий сывороточный альбумин (60 мг/л, 150 мг/л, 300 мг/л);
4. сырое куриное яйцо (1 шт. на 50 л среды ферментации);
5. фарш, полученный из сырого или вареного нежирного мяса (1 г на 10-15 л среды ферментации);
6. несоленый мясной бульон, полученный в результате варки нежирного куска мяса (60 мг, 150 мг по тотальному белку на литр стартовой среды ферментации).
В результате ферментации в присутствии экзогенных белков наблюдали многократное увеличение содержания в среде культивирования искомого рекомбинантного протективного антигена Echinococcus granulosus EG95tr [фигуры №9 и 10 (прилож.)]. В то время, как в группах контроля (MMY, MMY + DMEM без сыворотки, MMY +дистиллированная вода и пр.) увеличения количества рекомбинантного белка EG95tr в среде ферментации не наблюдали. При добавлении фетальной бычьей сыворотки до автоклавирования среды ферментации в количестве 1.0%, 2.5% и 5.0% от конечного объема среды ферментации наблюдали наиболее выраженный рост продуктивности культуры P.pastoris, в то время, как добавление 0.33% или 10.0% фетальной бычьей сыворотки к среде ферментации приводило к значительно менее выраженному увеличению продукции целевого белка EG95tr, хотя оно и превышало уровень экспрессии EG95tr в группе контроля (без добавления экзогенного белка). Аналогичное увеличение уровня целевого протективного антигена EG95tr в среде культивирования наблюдали при добавлении до автоклавирования 60-150 мг бычьего сывороточного альбумина на литр среды ферментации.
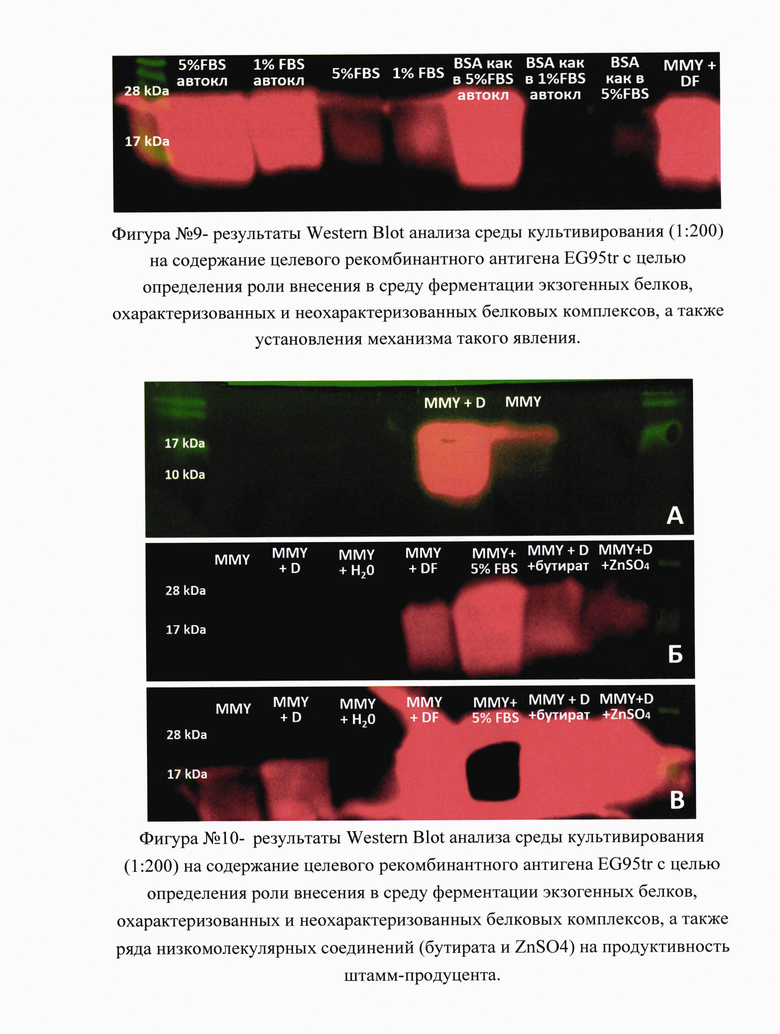
На фигуре №9 (прилож.) приведены результаты Western Blot анализа среды культивирования (1:200) на содержание целевого рекомбинантного антигена EG95tr с целью определения роли внесения в среду ферментации экзогенных белков, охарактеризованных и неохарактеризованных белковых комплексов, а также установления механизма такого явления. Видно, что данный эффект является дозозависимым и не зависит от типа белка (комплекс белков бычьей фетальной сыворотки с равным успехом может быть заменен эквивалентным количеством бычьего сывороточного альбумина) и денатурация белка при автоклавировании лишь увеличивает продуктивность штамма-продуцента. Это свидетельствует о том, что наблюдаемый эффект на продуктивность штамма дрожжей носит неспецифический характер, а вносимый белковый препарат, по-видимому, служит источником необходимых аминокислот и ограничивает влияние протеаз, секретируемых штаммом P.pastoris, на целевой белковый продукт. Первичные антитела специфичны к полигистидиновой метке (Abcam, #ab18184). MMY + DF - MMY с добавлением (1:1 об./об.) среды культивирования клеток млекопитающих DMEM, содержащей 10,0% FBS (фетальной бычьей сыворотки).
Из полученных результатов видно, что данный эффект является дозозависимым и не зависит от типа белка (комплекс белков бычьей фетальной сыворотки с равным успехом может быть заменен эквивалентным количеством бычьего сывороточного альбумина) и денатурация белка при автоклавировании лишь увеличивает продуктивность штамма-продуцента. Это свидетельствует о том, что наблюдаемый эффект на продуктивность штамма дрожжей носит неспецифический характер и не зависит от типа и природы вносимого белка, а вносимый белковый препарат, по-видимому, служит источником необходимых аминокислот и ограничивает влияние протеаз, секретируемых штаммом P.pastoris, на целевой белковый продукт.
Полученные результаты позволяют утверждать, что эффективность ферментации P.pastoris может быть многократно повышена путем введения в среду культивирования экзогенного рекомбинантного белка, охарактеризованного или не охарактеризованного белкового препарата, причем белковый препарат может содержать как нативные, так и денатурированные белки.
В рамках данного исследования также оценивали влияние низкомолекулярных добавок (бутират, ZnSO4) на продуктивность штамма P.pastoris в ходе ферментации, на фигуре №10 (прилож.) приведены результаты Western Blot анализа среды культивирования (1:200) на содержание целевого рекомбинантного антигена EG95tr с целью определения роли внесения в среду ферментации экзогенных белков, охарактеризованных и не охарактеризованных белковых комплексов, а также ряда низкомолекулярных соединений (бутирата и ZnSO4) на продуктивность штамм-продуцента. А - добавление в среду ферментации MMY среды культивирования клеток млекопитающих DMEM с 10,0% фетальной бычьей сывороткой привело к многократному увеличению (>10 раз) продукции целевого антигена EG95tr в среде культивирования; Б - добавление в среду ферментации MMY 5.0% фетальной бычьей сыворотки и других добавок (бутират, ZnSO4). В - то же, что и Б, но увеличено время экспозиции. Видно, что в обычной среде ферментации MMY количество искомого рекомбинантного антигена многократно (>100 раз) меньше, чем в группе, где в среду роста была добавлена 5,0% фетальная бычья сыворотка. Первичные антитела специфичны к полигистидиновой метке (Abcam, #ab18184). MMY -стандартная среда ферментация дрожжей Pichia pastoris (Methanlo-Minimal- Yeast medium); MMY + D - MMY с добавлением (1:1 об./об.) пустой среды культивирования клеток млекопитающих DMEM (Dulbecco's Modified Eagle Medium); MMY + H2O - MMY с добавлением (1:1 об./об.) дистиллированной воды; MMY + DF - MMY с добавлением (1:1 об./об.) среды культивирования клеток млекопитающих DMEM, содержащей 10.0% FBS (фетальной бычьей сыворотки); MMY + 5.0% FBS - MMY с добавлением FBS (фетальной бычьей сыворотки) - 5.0% от конечного объема. Однако, достоверного их влияния на эффективность ферментации выявлено не было.
Пример 6. Получение штамма-продуцента дрожжей P.pastoris, обеспечивающего продукцию рекомбинантного антигена на поверхности своих клеток, с целью иммунопрофилактики тканевых гельминтозов - т.н., живая вакцина (на примере протективного онкосферального антигена Echinococcus granulosus EG95)
Биоинформатический анализ, синтез и клонирование кДНК протективного онкосферального антигена EG95, а также трансформацию и культивирование штамма-продуцента дрожжей P. pastoris осуществляли, как это описано в примере 1, однако, в качестве аминокислотной последовательности для экспрессии была выбрана последовательность
НННН, т.е. нативный сигнальный пептид EG95 (MAFQLCLILFATSVLA) был заменен на сигнальный пептид альфа-феромона дрожжей S.cerevisiae MRFPSIFTAVLFAASSALAAPVNTTTEDETAQIPAEAVIGYSDLEGDFDVAV LPFSNSTNNGLLFINTTIASIAAKEEGVSLEKREAEA, но последовательность MTSGSALTSAIAGFVFSCIVVVL, заякоривающая белок EG95 в цитоплазматической мембране дрожжей продуцентов была сохранена, а к ее С-концу для детекции целевого белка была присоединена последовательность полигистидиновой метки НННННН. Результирующая нуклеотидная последовательность искомой кДНК (см. прилож. Перечень последовательностей, SEQ ID NO: 3), была собрана из синтетических олигонуклеотидов и клонирована в вектор pPpt4-alpha-S по сайтам рестрикции XhoI-NotI, как это описано в примере 1.
В результате трансформации компетентных клеток P.pastoris созданной генетической конструкцией и последующего скрининга клонов был отобран клон P.pastoris, экспрессирующий на своей поверхности рекомбинантный протективный антиген Echinococcus granulosus EG95, что подтверждается результатами Western Blot анализа осадков полученного штамма-продуцента P.pastoris.
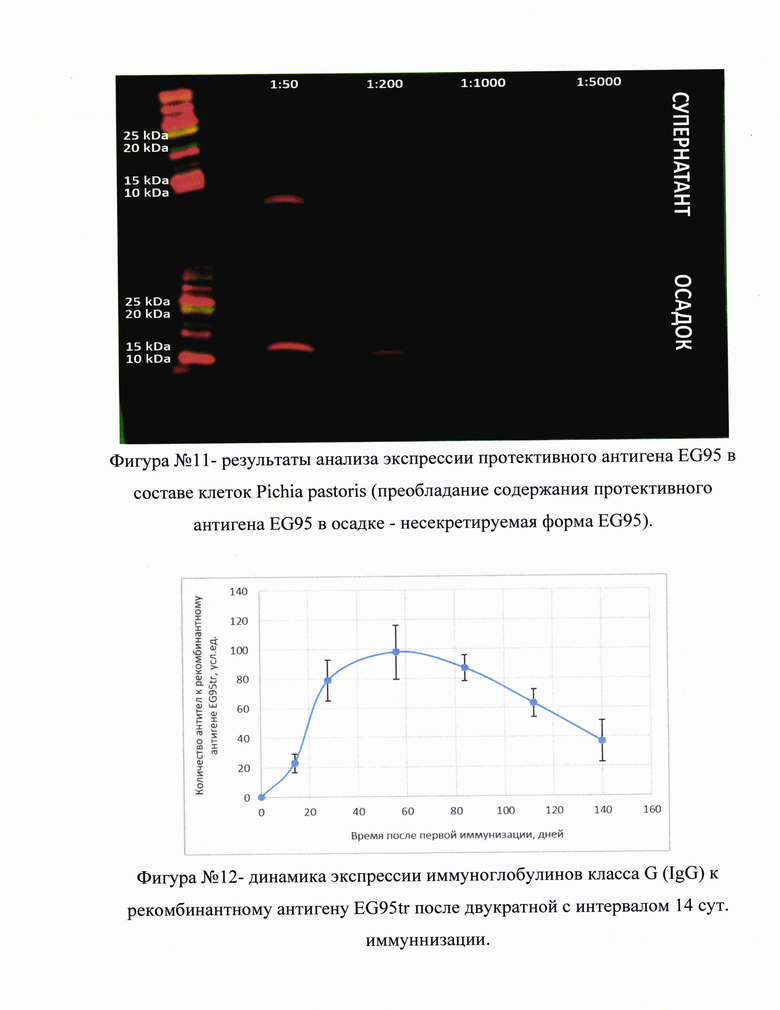
На фигуре №11 (прилож.) приведены результаты анализа экспрессии протективного антигена EG95 в составе клеток Pichia pastoris (преобладание содержания протективного антигена EG95 в осадке - несекретируемая форма EG95).
Пример 7. Оценка способности вакцины на основе протективного онкосферального антигена Echinococcus granulosus EG95tr препятствовать заражению сельскохозяйственных животных
Иммунизацию животных осуществляли путем введения рекомбинантного антигена EG95tr в сочетании с адъювантом интраназально или подкожно с интервалом не менее 10-14 суток. Эффективность протективного антигена при интраназальном пути введения оценивали на мышах линии С57/Black (самцы и самки, 21-25 г), в дозах 50, 100, 150 и 200 мкг/кг в сочетании с иммуностимулирующим носителем (адъювантом) PEI (10 мкг/мл). Эффективность протективного антигена при подкожном пути введения оценивали на ягнятах Романовской породы (барашки и ярочки, 3.5-4.5 кг), в дозах 20, 40 и 100 мкг/кг в сочетании с иммуностимулирующим носителем (адъювантом) полный адъювант Фрейнда, содержащий суспензию термически инактивированных Mycobacterium tuberculosis.
Протективную активность предлагаемой композиции оценивали по содержанию антител к рекомбинантному антигену EG95tr в сыворотке крови экспериментальных животных в течение 6 месяцев после первой иммунизации и по устойчивости к заражению тканевыми гельминтами {Echinococcus granulosus) на различной стадии развития (онкосферы, протосколексы) через 28 суток после первой иммунизации.
Для оценки содержания в сыворотке крови антител/иммуноглобулинов класса G (IgG) к рекомбинантному антигену EG95tr у мышей забирали кровь перед иммунизацией, а также спустя 14, 28, 56, 84, 112, 140 и 168 суток после первой иммунизации (2, 4, 8, 12, 16, 20 и 24 недель, соответственно), для этого на каждый срок передозировкой наркоза убивали по 6 мышей. Сыворотку крови получали из свернувшейся крови путем отделения кровяного сгустка центрифугированием свернувшейся крови в полипропиленовых пробирках соответствующего объема. Содержание IgG к рекомбинантному антигену EG95tr в сыворотке крови мышей оценивали с помощью иммуноферментного анализа (ИФА) в 96-луночном планшете с иммобилизацией рекомбинантного антигена EG95tr на подложке.
Оценку способности предлагаемой композиции снижать степень заражения тканевыми гельминтами (на примере Echinococcus granulosus) проводили путем забивания и вскрытия экспериментальных животных (через 6 месяцев после первой иммунизации) с последующим изучением и оценкой наличия, количества и размеров ларвоцист Echinococcus granulosus в брюшной полости и внутренних органах экспериментальных животных.
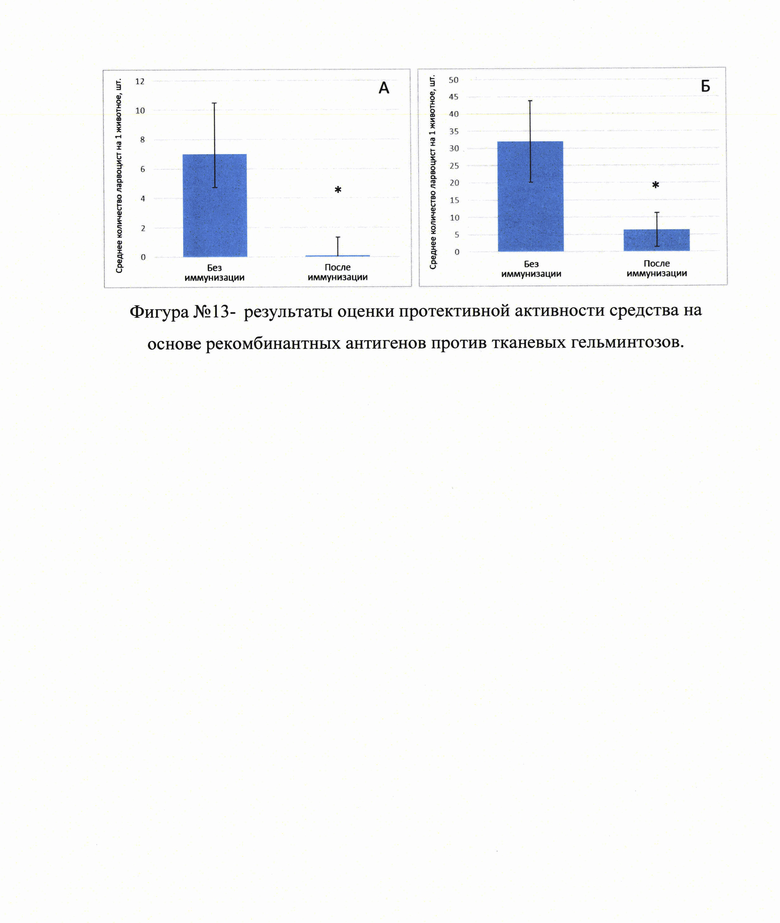
В результате проведенных исследований было установлено, что наиболее эффективной композицией вакцины для интраназальной иммунизации являются: протективный антиген - (не менее 100 мкг) в сочетании с иммуностимулирующим носителем (адъювантом) на кг вакцинируемого при условии не менее, чем двукратной иммунизации с интервалом не менее 10-14 сут. Данная схема применения рекомбинантного антигена EG95tr обеспечивала стабильную продукцию антител к рекомбинантному антигену в течение не менее 6 месяцев, на фигуре №12 (прилож.) приведена динамика экспрессии иммуноглобулинов класса G (IgG) к рекомбинантному антигену EG95tr после двукратной с интервалом 14 сут. иммуннизации. Продукция IgG к рекомбинантному антигену EG95tr в сыворотке крови иммунизированных животных составила не менее 4 месяцев (n=3), а также снижала степень заражения мышей протосколексами Echinococcus granulosus на 84.5%, на фигуре №13А (прилож.)] приведены результаты оценки протективной активности средства на основе рекомбинантных антигенов против тканевых гельминтозов: А - иммунизация мышей протективным онкосферальным антигеном EG95tr для защиты от инфицирования протосколексами Echinococcus granulosus (* - р<0.05 по сравнению с группой без иммунизации, n≥12);
У 75.0% мышей из экспериментальной группы не было обнаружено ни одной цисты, в то время, как в группе контроля в каждом животном при обследовании были обнаружены цисты Echinococcus granulosus.
В результате проведенных исследований было установлено, что наиболее эффективной композицией вакцины для подкожной иммунизации являются: протективный антиген - (не менее 40 мкг) в сочетании с иммуностимулирующим носителем (адъювантом) на кг вакцинируемого при условии не менее, чем двукратной иммунизации с интервалом не менее 10-14 сут. Данная схема применения рекомбинантного антигена EG95tr снижала степень заражения ягнят онкосферами Echinococcus granulosus на 78.0%, на фигуре №13Б (прилож.)] иммунизация ягнят протективным онкосферальным антигеном EG95tr для защиты от инфицирования онкосферами Echinococcus granulosus (* - р<0.05 по сравнению с группой без иммунизации, n≥6). Степень инфицирования животных различными стадиями тканевых гельминтов после иммунизации протективным онкосферальным антигеном EG95tr уменьшилась не меньше, чем на 78.0%, что свидетельствует об эффективности предлагаемого средства.
Список литературы:
1. Gemmell MA - Hydatidosis and cysticercosis. Induced resistance to the larval phase. Aust. Vet. J. (1970) 46: 366-369; Osborn PJ, Heath DD, Parmeter SN -Immunization of lambs against infection with Taenia ovis using an extract of Taenia ovis eggs. Research in Veterinary Science (1981) 31: 90-92.
2. Gauci CG, Jenkins DJ, Lightowlers MW. Protection against cystic echinococcosis in sheep using an Escherichia coli-expressed recombinant antigen (EG95) as a bacterin. Parasitology. 2022; 150(1):l-3. doi: 10.1017/S0031182022001457.
3. Gauci C, Jayashi C, Lightowlers MW. Vaccine development against the Taenia solium parasite: the role of recombinant protein expression in Escherichia coli. Bioengineered. 2013; 4(5):343-7. doi: 10.4161/bioe.23003.
4. US 2008/0280368 13 ноября 2008 г., Parasitic helminth cuticlin proteins and uses thereof, https://patentcenter.uspto.gov/applications/12143153, Дата последнего обращения: 06.08.2023.
5. US 7736642 B2 15 июня 2010 г., Yeast-based vaccine for inducing an immune response, https://patentcenter.uspto.gov/applications/11670902, Дата последнего обращения: 06.08.2023.
6. US 2011/0200640 A1 от 18 августа 2011 г., Multivalent antihelminthic vaccine, https://patentcenter.uspto.gov/applications/12999132, Дата последнего обращения: 06.08.2023.
7. US 2021/0154250 A1 от 13 октября 2020 г., Lysates of recombinant yeast for inducing cellular immune responses, https://patentcenter.uspto.gov/applications/17047134, Дата последнего обращения: 06.08.2023.
8. US 2021/0046170 A1 от 21 октября 2020 г., Biomarkers and immunogenic compositions for filarial parasites, https://patentcenter.uspto.gov/applications/17076616, Дата последнего обращения: 06.08.2023.
9. RU 2348427 C2 от 26 февраля 2007 г., Адъювантная композиция для инъекционных вакцин против тканевых гельминтозов, https://www.fips.ru/iiss/document.xhtml?faces- redirect=true&id=91ba944bd9db3fd4428ae8886a7163fa, Дата последнего обращения: 06.08.2023.
10. RU 2140986 С1 от 07 мая 1993 г., Молекула нуклеиновой кислоты (варианты), вектор экспрессии или клонирования, синтетический полипептид, способ получения синтетического полипептида, композиция вакцины для стимулирования иммунной реакции против гельминтных паразитов у человека или животных, способ стимуляции иммунной реакции против гельминтных паразитов у человека или животных, олигосахарид, https://www.fips.m/iiss/document.xhtml?facesredirect=true&id=d6449638f93fb135c74b84fced5bf342, Дата последнего обращения: 06.08.2023.
11. RU 95118396 А от 23 августа 1995 г., Применение скрытого антигена гельминтов для приготовления вакцинной композиции и способ стимулирования иммунного ответа против паразитических гельминтов животных, https://www.fips.ru/iiss/document.xhtml?faces-redirect=true&id=883643ca362a33311b1f726a66005741, Дата последнего обращения: 06.08.2023.
12. US 2020/0010515 A1 от 9 января 2020 г., Recombinant vaccine against helminths in pichia pastoris and methods for producing and purifying proteins for use as vaccines against helminthes, https://patentcenter.uspto.gov/applications/16478521, Дата последнего обращения: 06.08.2023.
13. Бессонов А.С. Цистный эхинококкоз и гидатидоз / Всерос .ин-т гельминтологии им. К.И. Скрябина. - М., 2007.
14. http://www.uniprot.org/uniprot/Q9GND7, Дата последнего обращения: 06.08.2023.
15. http://www.cbs.dtu.dk/services/SignalP/, Дата последнего обращения: 06.08.2023.
16. http://www.cbs.dtu.dk/services/TMHMM/, Дата последнего обращения: 06.08.2023.
17. http://www.kazusa.or.jp/codon/cgi-bin/showcodon.cgi?species=4922, Дата последнего обращения: 06.08.2023.
18. Macauley-Patrick S, Fazenda ML, McNeil В, Harvey LM. Heterologous protein production using the Pichia pastoris expression system. Yeast (2005), 22: 249-270.
19. Van den Hazel HB, Kielland-Brandt MC, Winther JR. Biosynthesis and function of yeast vacuolar proteases. Yeast (1996) 12: 1-1.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="СРЕДСТВО ДЛЯ
ИММУНОПРОФИЛАКТИКИ ТКАНЕВЫХ ГЕЛЬМИНТОЗОВ СЕЛЬСКОХОЗЯЙСТВЕННЫХ
ЖИВОТНЫХ И ЧЕЛОВЕКА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ.xml"
softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2023-09-14">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023120863/20(045475)</ApplicationNumberText>
<FilingDate>2023-08-09</FilingDate>
</ApplicationIdentification>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное научное учреждение "Федеральный научный центр -
Всероссийский научно-исследовательский институт экспериментальной
ветеринарии им. К.И. Скрябина и Я.Р. Коваленко Российской академии
наук" (ФГБНУ ФНЦ ВИЭВ РАН)</ApplicantName>
<ApplicantNameLatin>All Russian Scientific Research Institute for
Fundamental and Applied Parasitology of Animals and Plant, a branch
of the Federal State Budget Scientific Institution Federal Scientific
Centre VIEV</ApplicantNameLatin>
<InventionTitle languageCode="ru">СРЕДСТВО ДЛЯ ИММУНОПРОФИЛАКТИКИ
ТКАНЕВЫХ ГЕЛЬМИНТОЗОВ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ И ЧЕЛОВЕКА,
СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ</InventionTitle>
<SequenceTotalQuantity>3</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>434</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..434</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ctcgagaaaagagaggccgaagctcaggaatacaagggaatgggtgttg
aaaccagaactactgaaactccacttagaaaacatttcaatctaacacccgtcggttctcaaggtatcag
gttgagctgggaggttcagcacttgtcggacctgaagggtacagatatatcattaaaggcagtgaaccca
tctgacccattggtctataaacgtcaaacagccaaattctccgatggtcaattgactattggagagctga
aaccttccaccctctacaaaatgactgttgaagctgtaaaggctaaaaagacgattcttggctttactgt
ggatatcgagacacctagagcaggaaaaaaggagagtacagttatgggtggaggtcatcaccaccaccat
catcatcatcaccatcaccattaaggggcggccgc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>4210</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..4210</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>aacatccaaagacgaaaggttgaatgaaacctttttgccatccgacatc
cacaggtccattctcacacataagtgccaaacgcaacaggaggggatacactagcagcagaccgttgcaa
acgcaggacctccactcctcttctcctcaacacccacttttgccatcgaaaaaccagcccagttattggg
cttgattggagctcgctcattccaattccttctattaggctactaacaccatgactttattagcctgtct
atcctggcccccctggcgaggttcatgtttgtttatttccgaatgcaacaagctccgcattacacccgaa
catcactccagatgagggctttctgagtgtggggtcaaatagtttcatgttccccaaatggcccaaaact
gacagtttaaacgctgtcttggaacctaatatgacaaaagcgtgatctcatccaagatgaactaagtttg
gttcgttgaaatgctaacggccagttggtcaaaaagaaacttccaaaagtcggcataccgtttgtcttgt
ttggtattgattgacgaatgctcaaaaataatctcattaatgcttagcgcagtctctctatcgcttctga
accccggtgcacctgtgccgaaacgcaaatggggaaacacccgctttttggatgattatgcattgtctcc
acattgtatgcttccaagattctggtgggaatactgctgatagcctaacgttcatgatcaaaatttaact
gttctaacccctacttgacagcaatatataaacagaaggaagctgccctgtcttaaacctttttttttat
catcattattagcttactttcataattgcgactggttccaattgacaagcttttgattttaacgactttt
aacgacaacttgagaagatcaaaaaacaactaattattgaaagaattccgaaacgatgagattcccatct
attttcaccgctgtcttgttcgctgcctcctctgcattggctgcccctgttaacactaccactgaagacg
agactgctcaaattccagctgaagcagttatcggttactctgaccttgagggtgatttcgacgtcgctgt
tttgcctttctctaactccactaacaacggtttgttgttcattaacaccactatcgcttccattgctgct
aaggaagagggtgtctctctcgagaaaagagaggccgaagctcaggaatacaagggaatgggtgttgaaa
ccagaactactgaaactccacttagaaaacatttcaatctaacacccgtcggttctcaaggtatcaggtt
gagctgggaggttcagcacttgtcggacctgaagggtacagatatatcattaaaggcagtgaacccatct
gacccattggtctataaacgtcaaacagccaaattctccgatggtcaattgactattggagagctgaaac
cttccaccctctacaaaatgactgttgaagctgtaaaggctaaaaagacgattcttggctttactgtgga
tatcgagacacctagagcaggaaaaaaggagagtacagttatgggtggaggtcatcaccaccaccatcat
catcatcaccatcaccattaaggggcggccgctcaagaggatgtcagaatgccatttgcctgagagatgc
aggcttcatttttgatacttttttatttgtaacctatatagtataggattttttttgtcattttgtttct
tctcgtacgagcttgctcctgatcagcctatctcgcagcagatgaatatcttgtggtaggggtttgggaa
aatcattcgagtttgatgtttttcttggtatttcccactcctcttcagagtacagaagattaagtgaaac
cttcgtttgtgcggatccttcagtaatgtcttgtttcttttgttgcagtggtgagccattttgacttcgt
gaaagtttctttagaatagttgtttccagaggccaaacattccacccgtagtaaagtgcaagcgtaggaa
gaccaagactggcataaatcaggtataagtgtcgagcactggcaggtgatcttctgaaagtttctactag
cagataagatccagtagtcatgcatatggcaacaatgtaccgtgtggatctaagaacgcgtcctactaac
cttcgcattcgttggtccagtttgttgttatcgatcaacgtgacaaggttgtcgattccgcgtaagcatg
catacccaaggacgcctgttgcaattccaagtgagccagttccaacaatctttgtaatattagagcactt
cattgtgttgcgcttgaaagtaaaatgcgaacaaattaagagataatctcgaaaccgcgacttcaaacgc
caatatgatgtgcggcacacaataagcgttcatatccgctgggtgactttctcgctttaaaaaattatcc
gaaaaaattttctagagtgttgttactttatacttccggctcgtataatacgacaaggtgtaaggaggac
taaaccatggctaaactcacctctgctgttccagtcctgactgctcgtgatgttgctggtgctgttgagt
tctggactgatagactcggtttctcccgtgacttcgtagaggacgactttgccggtgttgtacgtgacga
cgttaccctgttcatctccgcagttcaggaccaggttgtgccagacaacactctggcatgggtatgggtt
cgtggtctggacgaactgtacgctgagtggtctgaggtcgtgtctaccaacttccgtgatgcatctggtc
cagctatgaccgagatcggtgaacagccctggggtcgtgagtttgcactgcgtgatccagctggtaactg
cgtgcatttcgtcgcagaagagcaggactaacaattgacaccttacgattatttagagagtatttattag
ttttattgtatgtatacggatgttttattatctatttatgcccttatattctgtaactatccaaaagtcc
tatcttatcaagccagcaatctatgtccgcgaacgtcaactaaaaataagctttttatgctcttctctct
ttttttcccttcggtataattataccttgcatccacagattctcctgccaaattttgcataatcctttac
aacatggctatatgggagcacttagcgccctccaaaacccatattgcctacgcatgtataggtgtttttt
ccacaatattttctctgtgctctctttttattaaagagaagctctatatcggagaagcttctgtggccgt
tatattcggccttatcgtgggaccacattgcctgaattggtttgccccggaagattggggaaacttggat
ctgattaccttagctgcaggtaccactgagcgtcagaccccgtagaaaagatcaaaggatcttcttgaga
tcctttttttctgcgcgtaatctgctgcttgcaaacaaaaaaaccaccgctaccagcggtggtttgtttg
ccggatcaagagctaccaactctttttccgaaggtaactggcttcagcagagcgcagataccaaatactg
ttcttctagtgtagccgtagttaggccaccacttcaagaactctgtagcaccgcctacatacctcgctct
gctaatcctgttaccagtggctgctgccagtggcgataagtcgtgtcttaccgggttggactcaagacga
tagttaccggataaggcgcagcggtcgggctgaacggggggttcgtgcacacagcccagcttggagcgaa
cgacctacaccgaactgagatacctacagcgtgagctatgagaaagcgccacgcttcccgaagggagaaa
ggcggacaggtatccggtaagcggcagggtcggaacaggagagcgcacgagggagcttccagggggaaac
gcctggtatctttatagtcctgtcgggtttcgccacctctgacttgagcgtcgatttttgtgatgctcgt
caggggggcggagcctatggaaaaacgccagcaacgcggcctttttacggttcctggccttttgctggcc
ttttgctcacatgttctttcctgcggtacccagatccaattcccgctttgactgcctgaaatctccatcg
cctacaatgatgacatttggatttggttgactcatgttggtattgtgaaatagacgcagatcgggaacac
tgaaaaatacacagttattattcatttaaat</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>476</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..476</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ctcgagaaaagagaggccgaagctcaggaatacaagggaatgggtgttg
aaaccagaactactgaaactccacttagaaaacatttcaatctaacacccgtcggttctcaaggtatcag
gttgagctgggaggttcagcacttgtcggacctgaagggtacagatatatcattaaaggcagtgaaccca
tctgacccattggtctataaacgtcaaacagccaaattctccgatggtcaattgactattggagagctga
aaccttccaccctctacaaaatgactgttgaagctgtaaaggctaaaaagacgattcttggctttactgt
ggatatcgagacacctagagcaggaaaaaaggagagtacagttatgacttcaggttccgcattgacatct
gccatcgctggatttgtcttcagttgtattgttgtagtgttgacccatcatcaccatcaccattaagggg
cggccgc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АДЪЮВАНТНАЯ КОМПОЗИЦИЯ ДЛЯ ИНЪЕКЦИОННЫХ ВАКЦИН ПРОТИВ ТКАНЕВЫХ ГЕЛЬМИНТОЗОВ | 2007 |
|
RU2348427C2 |
| ОЧИЩЕННЫЙ АНТИГЕННЫЙ ПОЛИПЕПТИД, ПРОТЕКТИВНЫЙ ПРОТИВ ИНФЕКЦИИ ECHINOCOCCUS GRANULOSUS, МОЛЕКУЛА ДНК, КОДИРУЮЩАЯ УКАЗАННЫЙ АНТИГЕННЫЙ ПОЛИПЕПТИД, ПРЕПАРАТ, СОДЕРЖАЩИЙ УКАЗАННЫЙ АНТИГЕННЫЙ ПОЛИПЕПТИД, ВАКЦИНА, СОДЕРЖАЩАЯ УКАЗАННЫЙ АНТИГЕННЫЙ ПОЛИПЕПТИД ИЛИ УКАЗАННУЮ МОЛЕКУЛУ ДНК | 1993 |
|
RU2205875C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОГО АНТИГЕНА Trichinella spiralis | 2005 |
|
RU2287342C1 |
| СУППОЗИТОРИИ ДЛЯ ИММУНОПРОФИЛАКТИКИ ВИРУСНЫХ ИНФЕКЦИЙ | 2004 |
|
RU2296560C2 |
| КОМБИНИРОВАННАЯ ВАКЦИНА ДЛЯ ИММУНОПРОФИЛАКТИКИ КОКЛЮША, ДИФТЕРИИ, СТОЛБНЯКА И ВИРУСНОГО ГЕПАТИТА В И Д | 2003 |
|
RU2233673C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА Е У ЖИВОТНЫХ | 2012 |
|
RU2501568C1 |
| КОМБИНИРОВАННАЯ ВАКЦИНА ДЛЯ ИММУНОПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА В И Д, СТОЛБНЯКА И ДИФТЕРИИ | 2003 |
|
RU2233672C1 |
| Способ профилактики ларвальной стадии альвеолярного эхинококкоза | 2016 |
|
RU2609858C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА В (ВАРИАНТЫ) | 2003 |
|
RU2238105C1 |
| ВАКЦИНАЦИЯ С ПОМОЩЬЮ РЕКОМБИНАНТНЫХ ДРОЖЖЕЙ С ФОРМИРОВАНИЕМ ЗАЩИТНОГО ГУМОРАЛЬНОГО ИММУННОГО ОТВЕТА ПРОТИВ ОПРЕДЕЛЕННЫХ АНТИГЕНОВ | 2012 |
|
RU2630620C2 |

Изобретение относится к области ветеринарии, медицины и биотехнологии. Предложена вакцина для иммунопрофилактики тканевых форм эхинококкозов сельскохозяйственных животных и человека, содержащая рекомбинантный онкосферальный протективный антиген эхинококка EG95tr, кодируемый нуклеотидной последовательностью SEQ ID NO: 1 или SEQ ID NO: 3, и вспомогательные компоненты. Предложен способ получения вакцины для иммунопрофилактики тканевых форм эхинококкозов сельскохозяйственных животных и человека. Способ предусматривает получение ДНК, кодирующей рекомбинантный антиген эхинококка EG95, имеющей нуклеотидную последовательность SEQ ID NO: 1 или SEQ ID NO: 3, клонирование указанной ДНК в шаттл-вектор и доставку генетической конструкции в клетки дрожжей Pichia pastoris с получением генноинженерного микроорганизма Pichia pastoris, несущего ген протективного антигена EG95, ферментацию на средах YPD, MMY, MGY, ВММ, BMG, BMGY, дефинитивных средах, очистку полученного рекомбинантного антигена эхинококка EG95tr из среды культивирования центрифугированием или декантированием среды ферментации с последующим разбавлением среды культивирования, с последующей ультрафильтрацией через фильтры с размером пор 50 кДа и последующей ультрафильтрацией через фильтры с размером пор 3-5 кДа, с последующей очисткой искомого рекомбинантного антигена из среды культивирования или фильтрата, консервацию очищенного рекомбинантного антигена путем моментального замораживания в жидком азоте или лиофильного высушивания, добавление вспомогательных компонентов, в качестве которых используют адъюванты, наполнители, изотонические агенты, стабилизаторы и/или консерванты. Предложено применение вакцины для иммунопрофилактики тканевых форм эхинококкозов сельскохозяйственных животных и человека в терапевтических дозах из расчета на протективный антиген для подкожной 40 мкг/кг и 100 мкг/кг для интраназальной иммунизации, в сочетании с иммуностимулирующим носителем, не менее двух раз с интервалом 10-14 суток для иммунопрофилактики тканевых форм эхинококкозов сельскохозяйственных животных и человека. Онкосферальный антиген EG95tr обладает высокой протективной активностью, что обеспечивает стабильную продукцию антител к рекомбинантному антигену в сыворотке крови иммунизированных животных в течение не менее 4 месяцев, а также снижение инфицирования целевых животных тканевыми гельминтами не менее чем на 78,0% для иммунопрофилактики цистного гидатидоза. 3 н.п. ф-лы, 13 ил., 7 пр.
1. Вакцина для иммунопрофилактики тканевых форм эхинококкозов сельскохозяйственных животных и человека, содержащая рекомбинантный онкосферальный протективный антиген эхинококка EG95tr, кодируемый нуклеотидной последовательностью SEQ ID NO: 1 или SEQ ID NO: 3, и вспомогательные компоненты.
2. Способ получения вакцины для иммунопрофилактики тканевых форм эхинококкозов сельскохозяйственных животных и человека по п. 1, предусматривающий:
- получение ДНК, кодирующей рекомбинантный антиген эхинококка EG95, имеющей нуклеотидную последовательность SEQ ID NO: 1 или SEQ ID NO:3, клонирование указанной ДНК в шаттл-вектор и доставку генетической конструкции в клетки дрожжей Pichia pastoris с получением генноинженерного микроорганизма Pichia pastoris, несущего ген протективного антигена EG95, ферментацию на средах YPD, MMY, MGY, ВММ, BMG, BMGY, дефинитивных средах со следующим составом: биотин - 2-4 мкг/л; пантотенат кальция - 0.4-0.8 мг/л; фолиевая кислота - 2-4 мкг/л; инозитол - 2-4 мг/л; парааминобензойная кислота - 0.2-0.4 мг/л; пиридоксин -0.4-0.8 мг/л; рибофлавин - 0.2-0.4 мг/л; тиамин - 0.4-0.8 мг/л; Н3ВО3 - 0.5-1 мг/л; Cu2SO4 - 40-80 мкг/л; KI - 0.1-0.2 мг/л; FeC13 - 0.2-0.4 мг/л; MnSO4 -0.4-0.8 мг/л; Na2MoO4 - 0.2-0.4 мг/л; ZnSO4 - 0.4-0.8 мг/л; KH2PO4 - 0.1-0.2%; MgSO4 - 0.05-0.1%; NaCl - 0.01-0.02%; СаС12*2H2O - 0.0132-0.0264%; (NH4)2SO4 - 0.5-1,0%; СН3ОН - 1,0%; глицерол - 1,0%; гистидин - 0.004%; никотиновая кислота 10 мг/мл - 0.004-0.008%; натрия гидроксид 10М - 0.0006-0.0012%, с внесением до автоклавирования среды ферментации или после ее автоклавирования при соблюдении правил асептики 0.2-150 мг на один литр среды экзогенного белка или белковой композиции, культивирование в течение 72-120 часов,
- очистку полученного рекомбинантного антигена эхинококка EG95tr из среды культивирования центрифугированием или декантированием среды ферментации с последующим разбавлением среды культивирования в 2-100 раз растворами 0.05-0.5% карбоната аммония, и/или формиата аммония, и/или цитрата аммония, и/или 10-150 мМ фосфатным буфером, и/или раствором, содержащим 10-100 мМ Tris и/или 100-500 мМ хлорида натрия, с азидом натрия или без него, с добавлением антибиотика или без него, с последующей ультрафильтрацией через фильтры с размером пор 50 кДа и последующей ультрафильтрацией через фильтры с размером пор 3-5 кДа, с последующей очисткой искомого рекомбинантного антигена из среды культивирования или фильтрата методами металло-хелатной, ионнообменной, аффинной и/или эксклюзионной хроматографии в одну или несколько стадий, в сочетании с ультрафильтрацией или без нее,
- консервацию очищенного рекомбинантного антигена путем моментального замораживания в жидком азоте или лиофильного высушивания в 6-10 стадий при давлении 0.013-0.027 кПа, температуре от минус 45°С до минус 25°С и времени лиофилизации не более 24 часов при максимальной загрузке лиофильной сушки,
- добавление вспомогательных компонентов, в качестве которых используют адъюванты, наполнители, изотонические агенты, стабилизаторы и/или консерванты.
3. Применение вакцины по п. 1 в терапевтических дозах из расчета на протективный антиген для подкожной 40 мкг/кг и 100 мкг/кг для интраназальной иммунизации, в сочетании с иммуностимулирующим носителем, не менее двух раз с интервалом 10-14 суток для иммунопрофилактики тканевых форм эхинококкозов сельскохозяйственных животных и человека.
| Способ восстановления спиралей из вольфрамовой проволоки для электрических ламп накаливания, наполненных газом | 1924 |
|
SU2020A1 |
| WO 9323542 A1, 25.11.1993 | |||
| Liang Wang et al | |||
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Mohammad Mostafa Poursei et al., A novel B- | |||

