Область изобретения
В общих чертах настоящее изобретение относится к области молекулярной биологии бактерий-паразитов, а в частности к агентам, вызывающим заболевания типа риккетсиоза, и к бактериям Ehrlichia. Более конкретно настоящее изобретение относится к молекулярному клонированию и характеризации гена иммунореактивного белка массой 120 кДа Ehrlichia canis.
Описание уровня техники
Ehrlichia spp. представляют собой облигатные внутриклеточные грам-отрицательные бактерии, которые обитают в эндосоме гемопоэтических клеток и инфицируют различных животных-хозяев, включая человека, домашних и диких собак, оленей, лошадей, овец, крупный рогатый скот и диких грызунов. Каждый член рода Ehrlichieae обладает своим собственным тропизмом в отношении клеток-мишеней. Большинство видов Ehrlichia являются либо моноцитотройными (Е. canis, E. chaffeensis, E. sennetsu, E. risticii и E. muris), либо гранулоцитотропными (человеческая гранулоцитарная Ehrlichia, E. eqvi, E. phagocytophila и E. ewingii) за исключением Cowdria ruminatium, которая развивается в эндотелиальных клетках хозяина, и Anaplasma marginale, являющаяся эритроцитарным паразитом.
Хотя бактерии Ehrlichia уже были описаны в начале этого века, однако в Соединенных Штатах вплоть до последнего десятилетия они мало привлекали внимание исследователей, поскольку их считали патогенами, имеющими значение лишь для ветеринарии. Вновь появившийся интерес к эрлихиям обусловлен возникновением эрлихиоза, поражающего человека. В последнее десятилетие в Соединенных Штатах были открыты два новых патогена Ehrlichia (Е. chaffeensis и Е. phagocytophila-подобный микроорганизм человека) (Bakken J.S. 1994, Chen S.M., 1994, J.C.M., Fishbein D.В., 1987, Maeda, К., 1987). Ehrlichia canis, прототипный вид этого рода, представляет собой этиологический фактор эрлихиоза собак, являющийся фактически лишь одним из пяти видов Ehrlichia, который обычно инфицирует собак. Собачий эрлихиоз является заболеванием, широко распространенным во всем мире и переносимым собачьим клещом Rhipicephalus sanguineus (Groves M.G., 1975, Lewis G.E. Jr., 1977). Ehrlichia canis вызывает резкое умеренное и кратковременное повышение температуры, которое может прогрессировать с развитием тяжелого заболевания и синдрома с летальным исходом (тропическая панцитопения собак) (Buhles W.C., 1974, Greene С.Е. & J.W. Harvey, 1984, Walter, J.S. 1970). Во всем мире затраты на лечение домашних и служебных собак, инфицированных Е. canis, составляют миллионы долларов в год. Кроме того, бактерия Е. canis также ставит под угрозу здоровье людей. Ehrlichia canis или антигенно неидентифицируемый микроорганизм был недавно выделен у человека (Perez М., 1996).
Выявление генетического и антигенного состава Е. canis является главным фактором для изучения патогенеза эрлихиоза собак и разработки эффективной вакцины. По своим генетическим и антигенным свойствам бактерия Е. canis является близкородственной бактерии Е. chaffeensis (Anderson В.Е., 1991, 1992, Chen S.M.f Am. J. Trop. Med. Hyg). Поэтому собачий эрлихиоз может быть подходящей моделью для исследования патогенеза моноцитотропных видов Ehrlichia, включая Е. chaffeensis.
Предшествующие работы имеют тот недостаток, что в них ничего не сообщается о клонировании и характеризации иммунореактивного белка массой 120 кДа Ehrlichia canis. Кроме того, в предшествующих работах ничего не сообщается о рекомбинантном гене иммунореактивного белка массой 120 кДа Ehrlichia canis. Настоящее изобретение удовлетворяет этой давно назревшей необходимости и требованиям в данной области техники.
Краткое описание изобретения
В одном из вариантов своего осуществления настоящее изобретение относится к гену, кодирующему иммунореактивный белок массой 120 кДа Ehrlichia canis. Этот белок массой 120 кДа имеет предпочтительно аминокислотную последовательность SEQ ID NO:8, а его ген имеет нуклеотидную последовательность SEQ ID NO:7.
В предпочтительном варианте своего осуществления настоящее изобретение относится к экспрессирующему вектору, содержащему ген, кодирующий иммунореактивный белок массой 120 кДа Ehrlichia canis, где указанный экспрессирующий вектор при его введении в клетку способен экспрессировать этот ген.
В другом варианте своего осуществления, настоящее изобретение относится к рекомбинантному белку, содержащему аминокислотную последовательность SEQ ID NO:8. Предпочтительной является аминокислотная последовательность, кодируемая нуклеотидной последовательностью SEQ ID NO:7. Указанный рекомбинантный белок массой 120 кДа предпочтительно содержит 14 тандемных повторяющихся единиц, каждая из которых содержит 36 аминокислот. Более предпочтительно, если указанные повторяющиеся единицы являются гидрофильными. Еще более предпочтительно, если указанный рекомбинантный белок массой 120 кДа является антигеном.
В предпочтительном варианте осуществления настоящее изобретение относится к способу продуцирования рекомбинантного белка массой 120 кДа, включающему стадии получения вектора, содержащего экспрессирующую область, имеющую последовательность, кодирующую аминокислотную последовательность SEQ ID NO:8, правильно присоединенную к промотору; трансфекции указанного вектора в клетку; и культивирование этой клетки в условиях, эффективных для экспрессии этой экспрессирующей области.
Настоящее изобретение может быть описано на некоторых вариантах его осуществления, относящихся к способу ингибирования инфекции, вызываемой Ehrlichia canis у субъекта, включающему стадии идентификации субъекта, предположительно подверженного воздействию бактерии или инфицированного бактерией Ehrlichia canis; и введения композиции, содержащей антиген массой 120 кДа Ehrlichia canis в количестве, эффективном для ингибирования инфекции, вызываемой Ehrlichia canis. Такое ингибирование может быть достигнуто любыми способами, такими как, например, стимуляция у данного субъекта гуморального или клеточного иммунного ответа, либо какими-либо другими способами, такими как ингибирование нормальной функции антигена массой 120 кДа, или даже конкуренция с данным антигеном за взаимодействие с определенным агентом в организме субъекта.
Другие и дополнительные аспекты, отличительные признаки и преимущества настоящего изобретения будут очевидны из нижеследующего описания предпочтительных вариантов осуществления настоящего изобретения, приведенных в целях его описания.
Краткое описание чертежей
Для того чтобы вышеуказанные отличительные признаки, преимущества и цели настоящего изобретения и т.п. были более ясными и понятными в деталях, описание настоящего изобретения, кратко изложенного выше, может быть более конкретно проиллюстрировано на некоторых вариантах его осуществления со ссылками на прилагаемые чертежи. Эти чертежи являются частью настоящего описания. Однако при этом следует отметить, что указанные чертежи иллюстрируют предпочтительные варианты настоящего изобретения, а поэтому не должны рассматриваться как ограничение его объема.
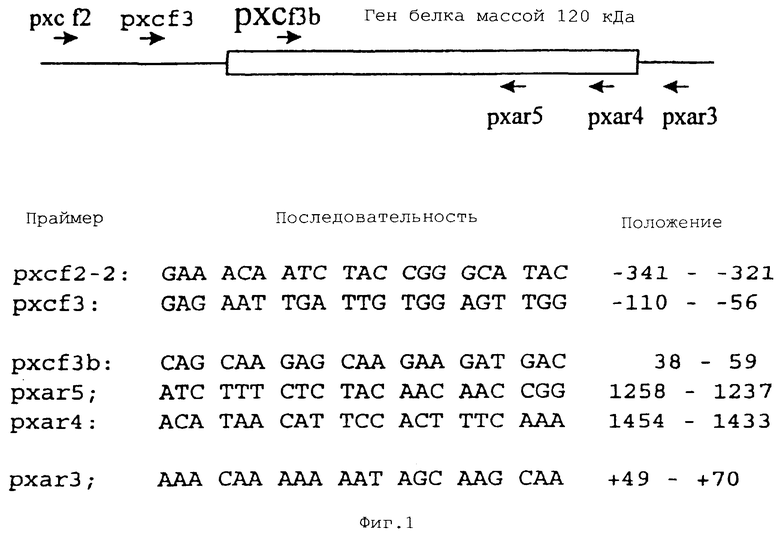
На Фиг.1 показана последовательность ДНК и положения олигонуклеотидных праймеров, происходящих от гена белка массой 120 кДа Е. chaffeensis (незаштрихованная рамка), и последовательности ДНК, фланкирующие этот ген (линия). Положения праймеров указаны как минусы и плюсы для последовательностей ДНК, расположенных выше и ниже от гена белка массой 120 кДа соответственно. Было сконструировано девять пар праймеров путем объединения каждого прямого праймера (SEQ ID NO:1-3) с каждым обратным праймером (SEQ ID NO:4-6), и эти праймеры были использованы для амплификации гена белка массой 120 кДа Е. canis с помощью ПЦР.
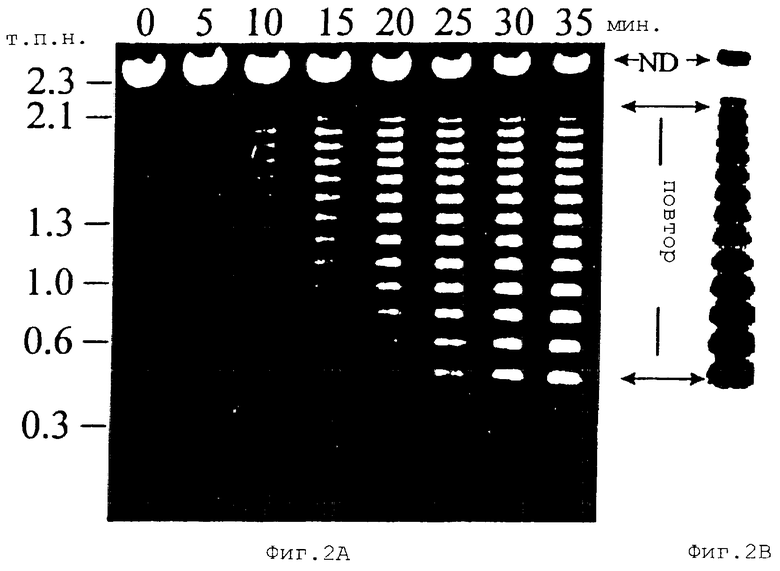
На Фиг.2А показан электрофорез в агарозном геле, проведенный для повторяющихся единиц гена белка массой 120 кДа Е. canis. Плазмида рСА120 была сначала гидролизована ферментом EcoRI для выделения вставки, а затем она была гидролизована ферментом SheI через различные интервалы времени. На Фиг.2В показано определение числа повторов методом Саузерн-блоттинга. ДНК, гидролизованную в течение 35 минут ферментом Shel, переносили из геля в панели А на найлоновые мембраны и гибридизовали с олигонуклеотидным зондом, меченным диоксигенином, который гибридизовался с последовательностями ДНК, расположенными выше от области повторов гена белка массой 120 кДа Е. canis. ND = негидролизованная ДНК.
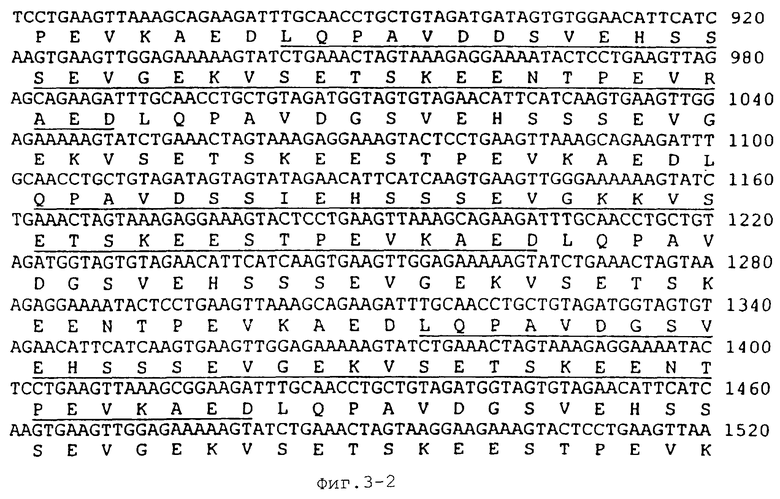
На Фиг.3 показана последовательность ДНК гена белка массой 120 кДа Е. canis (SEQ ID NO: 7) и выведенные аминокислоты (SEQ ID NO:8). Нуклеиновые кислоты повторов 1, 3, 5, 7, 9, 11 и 13 подчеркнуты.
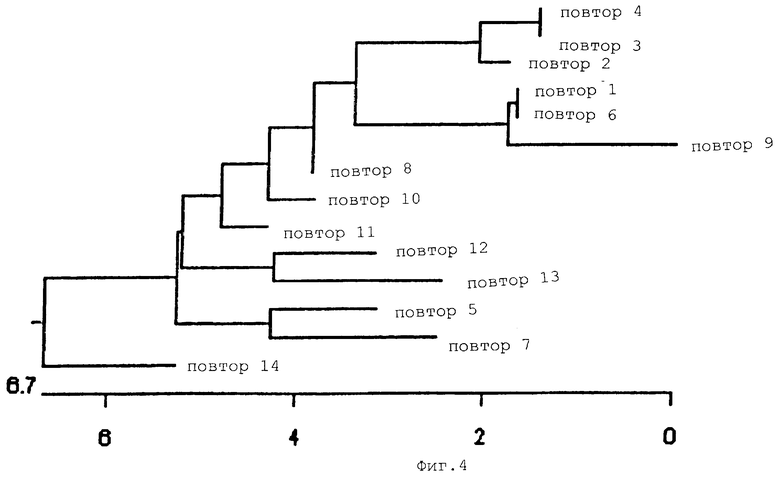
На Фиг.4 показано филогенетическое дерево повторяющихся единиц гена белка массой 120 кДа Е. canis. Шкала представляет % различий в последовательности ДНК.
На Фиг.5 проиллюстрирован сравнительный анализ первичных аминокислотных последовательностей белков массой 120 кДа Е. canis (SEQ ID NO:10) и Е. chaffeensis (SEQ ID NO:9). Черточки указывают на идентичные аминокислоты. Двоеточие указывает на консервативные замены.
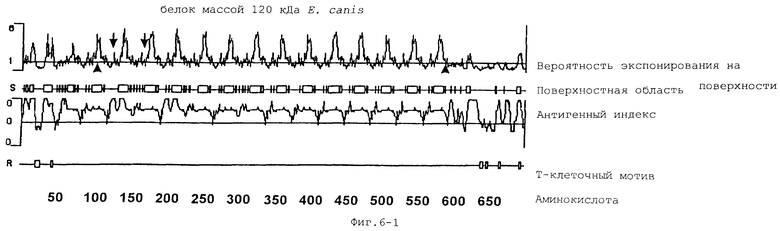
На Фиг.6 показаны вероятность экспонирования на поверхности и гидрофобность белков массой 120 кДа Е. canis E. chaffeensis. Область между клинообразными стрелками является доменом повторов. Все повторы в обоих белках являются гидрофильными и поверхностно экспонируемыми. Вторая повторяющаяся единица белка массой 120 кДа Е. canis и пик первой повторяющейся единицы белка массой 120 кДа Е. chaffeensis находятся между двумя стрелками и более крупно показаны на Фиг.7.
На Фиг.7 показано сравнение поверхностно экспонируемых аминокислот в повторяющейся единице белков массой 120 кДа Е. canis и Е. chaffeensis. На Фиг.7А показана вероятность экспонирования аминокислот на поверхности. Соответствующие области указаны двумя клинообразными стрелками на Фиг.6. Буквами жирным шрифтом показаны консервативные аминокислоты для Е. canis и Е. chaffeensis. На Фиг.7В показано сравнение первичной аминокислотной последовательности, представленной на панели А (SEQ ID NO:11-12). Черточки указывают на идентичные аминокислоты. Точками обозначены консервативные замены.
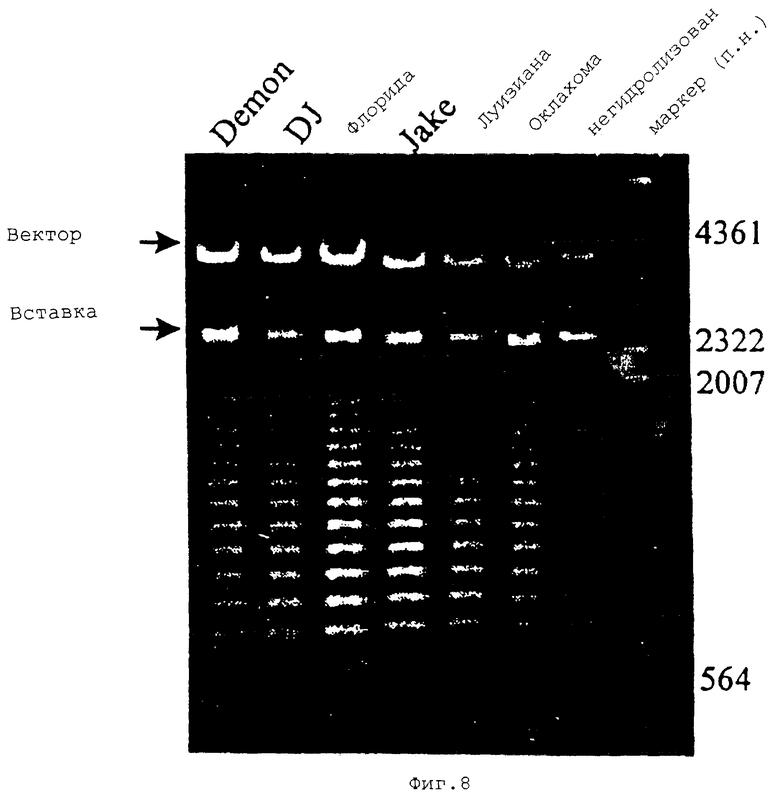
На Фиг.8 показан электрофорез в агарозном геле, проведенный для гена белка массой 120 кДа штамма Е. canis, гидролизованного SреI. Рекомбинантные плазмиды pCR2.1 были сначала гидролизованы EcoRI для выделения вставки из вектора, а затем частично гидролизованы SpeI. Негидролизованы: ДНК гена белка массой 120 кДа штамма Оклахома была гидролизована ферментом EcoRI, но не ферментом SpeI для того, чтобы показать размер вставки.
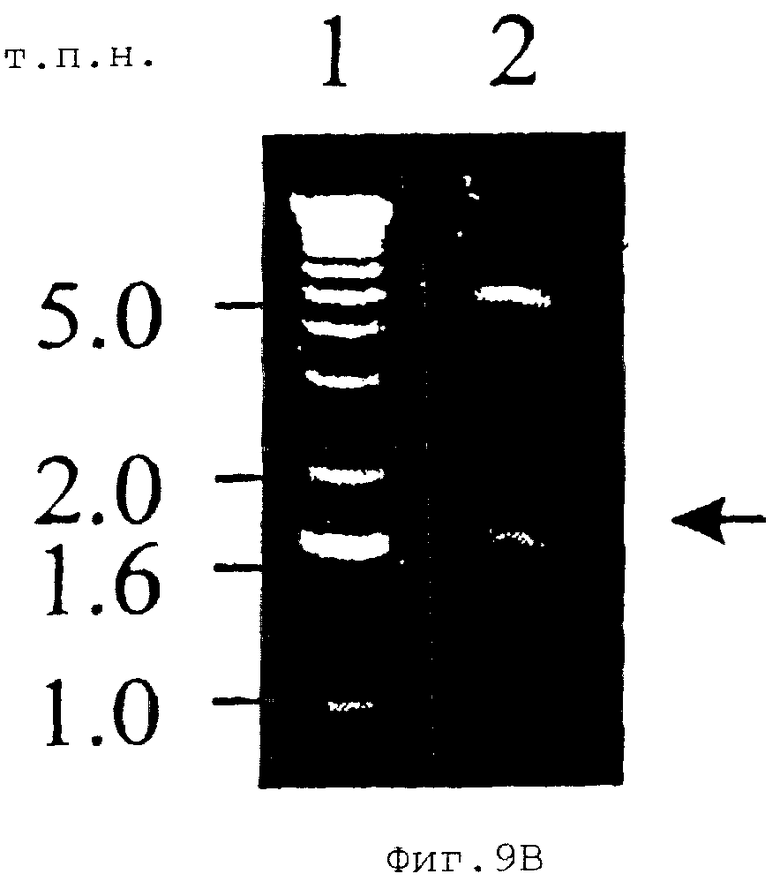
На Фиг.9А показан ПААГ-СНД-электрофорез белка массой 120 кДа Е. canis, экспрессированного в E. coli. 1, GST-гибридный белок массой 120 кДа; 2, рекомбинактный белок массой 120 кДа Е. canis, отщепленный от GST-гибридного белка тромбином. На Фиг.9В показан электрофорез в агарозном геле для плазмиды pGEX, экспрессирующей белок массой 120 кДа Е. canis. Вставка указана стрелкой.
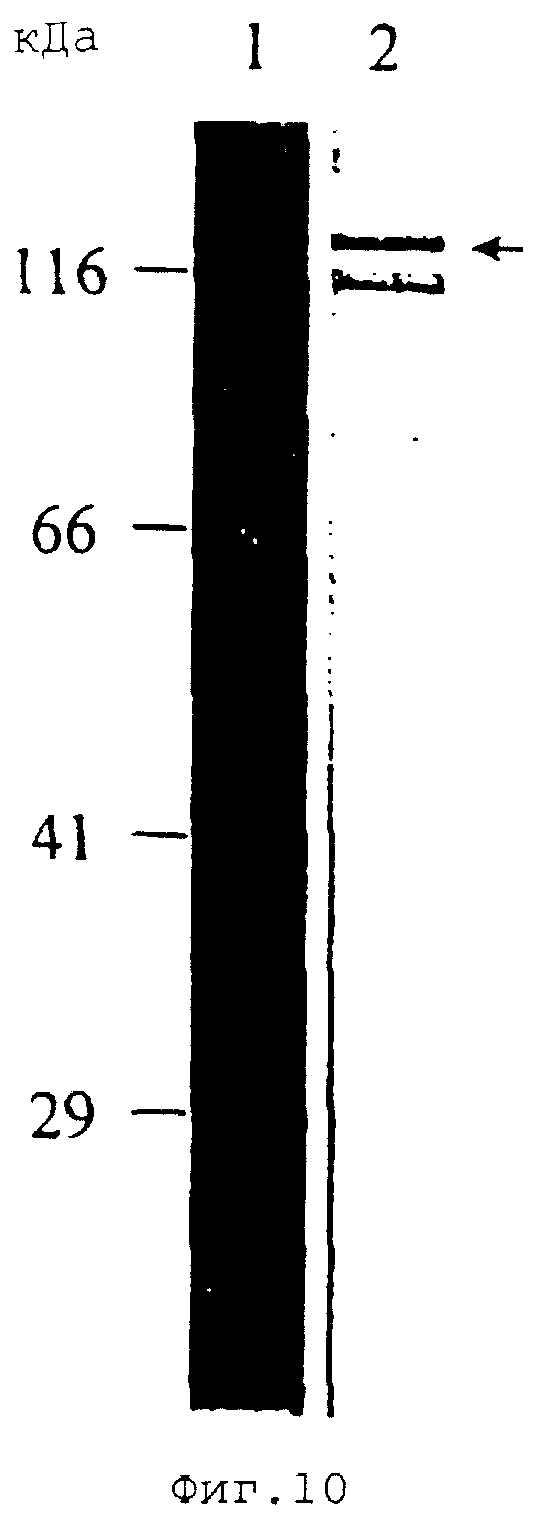
На Фиг.10 показан Вестерн-блот-анализ мышиной антисыворотки против рекомбинантного белка массой 120 кДа Е. canis, реагирующей с антигеном Е. canis (дорожка 1) и с рекомбинантным белком массой 120 кДа Е. canis (дорожка 2, стрелка).
На Фиг.11 показан Вестерн-блот-анализ сыворотки выздоравливающей собаки, реагирующей с рекомбинантным белком массой 120 кДа Е. canis.
Подробное описание изобретения
В соответствии с настоящим изобретением могут быть использованы стандартные методы молекулярной биологии, микробиологии и технологии рекомбинатных ДНК, известные специалистам. Эти методы подробно описаны в литературе. Например, Maniatis, Fritish & Sambrook "Molecular Cloning. - A laboratory manual (1982); "DNA-Cloning: A Practical Approach", Volumes I and II (D.N. Glover ed. 1985); "Oligonucleotide Synthesis" (M.J.Gait ed. 1984); "Nucleic Acid Hybridization" [B.D. Hames & S.J. Higgins eds. (1985)]; "Transcription and Translation" [B.D. Hames & S.J. Higgins eds. (1984)]; "Animal Cell Culture" [R.I. Freshney, ed. (1986)]; Immobilized Cells and Enzymes" [IRL Press, (1986)]; B. Perhal, "A Practical Guide To Molecular Cloning" (1984).
Поэтому нижеследующие термины, если они встречаются, имеют значения, определенные ниже.
Описанные здесь аминокислоты имеют предпочтительно "L"-изомерную форму. Однако L-аминокислотные остатки могут быть заменены "D"-изомерными формами при условии, что нужные функциональные свойства данного полипептида в отношении связывания с иммуноглобулином останутся неизменными. NH2 означает свободную аминогруппу, присутствующую у амино-конца полипептида. СООН означает свободную карбоксигруппу, присутствующую у карбоксиконца полипептида. В соответствии со стандартной номенклатурой полипептов, J.Biol.Chem. 243:3552-59 (1969), обозначения аминокислотных остатков представлены в таблице соответствий.

Следует отметить, что все последовательности аминокислотных остатков представлены здесь формулами, левая и правая ориентация которых имеет обычное направление от амино-конца к карбокси-концу. Кроме того, следует отметить, что черточка в начале или в конце последовательности аминокислотных остатков указывает на пептидную связь с другой последовательностью одного или нескольких аминокислотных остатков. Таблица представлена для идентификации трехбуквенных и однобуквенных кодов, которые встречаются в этом описании.
Термин "репликон" означает любой генетический элемент (например, плазмиду, хромосому, вирус), который функционирует как автономная единица репликации ДНК in vivo; т.е. способен к репликации под своим собственным контролем. Термин "вектор" означает репликон, такой как плазмида, фаг или космида, к которому может быть присоединен другой сегмент ДНК, так чтобы осуществлялась репликация этого присоединенного сегмента.
Термин "молекула ДНК" означает полимерную форму дезоксирибонуклеотидов (аденина, гуанина, тимина или цитозина) либо в их одноцепочечной форме, либо в виде двухцепочечной спирали. Этот термин относится только к первичной и вторичной структуре молекулы и не ограничивается какими-либо конкретными третичными формами. Таким образом, этот термин включает двухцепочечную ДНК, обнаруженную, inter alia, в линейных молекулах ДНК (например, рестрикционных фрагментах), вирусах, плазмидах и хромосомах. При обсуждении структуры в соответствии со стандартными условными обозначениями указывается только последовательность в направлении 5’→3’ вдоль нетранскрибируемой цепи ДНК (т.е., цепи, имеющей последовательность, гомологичную последовательности мРНК).
Термин "сайт инициации репликации" означает последовательности ДНК, которые участвуют в синтезе ДНК. "Кодирующая последовательность" ДНК представляет собой двухцепочечную последовательность ДНК, которая, находясь под контролем соответствующих регуляторных последовательностей, транскрибируется и транслируется in vivo с образованием полипептида. Границы кодирующей последовательности определяются старт-кодоном у 5’ амино-конца и кодоном терминации трансляции у 3’ карбокси-конца. Кодирующей последовательностью могут быть, но не ограничиваются ими, последовательности прокариотов, кДНК от мРНК эукариотов, геномные последовательности ДНК от ДНК эукариотов (например, млекопитающего) и даже синтетические последовательности ДНК. Сигнал полиаденилирования и последовательность терминации транскрипции обычно расположены с 3’-конца по отношению к кодирующей последовательности.
Транскрипционные и трансляционные регуляторные последовательности представляют собой регуляторные последовательности ДНК, такие как промоторы, энхансеры, сигналы полиаденилирования, терминаторы и т.п., которые обеспечивают экспрессию кодирующей последовательности в клетке-хозяине.
"Промоторная последовательность" представляет собой регуляторную область ДНК, способную связываться с РНК-полимеразой в клетке и инициировать транскрипцию кодирующей последовательности в прямом направлении (→3’). В соответствии с настоящим изобретением эта промоторная последовательность ограничена у своего 3’-конца сайтом инициации транскрипции и простирается выше (в 5’-направлении) так, что она включает минимальное число оснований или элементов, необходимых для инициации транскрипции на уровнях, детектируемых выше фонового уровня. Внутри этой промоторной последовательности находится сайт инициации транскрипции, а также белок-связывающие домены (консенсусные последовательности), ответственные за связывание с РНК-полимеразой. В большинстве случаев, но не всегда эукариотические промоторы содержат "ТА-ТА"-боксы и "САТ"-боксы. Помимо консенсусных последовательностей -10 и -35 прокариотические промоторы содержат последовательности Шайна-Дальгарно.
"Последовательность регуляции экспрессии" представляет собой последовательность ДНК, которая обеспечивает контроль и регуляцию транскрипции и трансляции другой последовательности ДНК. Кодирующая последовательность "находится под контролем" транскрипционной и трансляционной регуляторных последовательностей в клетке в том случае, если РНК-полимераза транскрибирует кодирующую последовательность в мРНК, которая затем транслируется с образованием белка массой 120 кДа, кодированного этой кодирующей последовательностью.
"Сигнальная последовательность" может присутствовать возле кодирующей последовательности. Эта последовательность кодирует сигнальный пептид, который находится у N-конца по отношению к полипептиду и сообщает клетке-хозяину способность направлять этот полипептид к клеточной поверхности клетки или секретировать его в среду, и этот сигнальный пептид отщепляется в клетке-хозяине, после чего белок массой 120 кДа покидает эту клетку. Сигнальные последовательности могут присутствовать в комбинации с различными нативными белками прокариотов и эукариотов.
Используемый здесь термин "олигонуклеотид" означает зонд настоящего изобретения и представляет собой молекулу, состоящую из двух или более, а предпочтительно из более трех дезоксирибонуклеотидов. Точный его размер зависит от многих факторов, которые, в свою очередь, зависят от конечной функции и цели использования этого олигонуклеотида.
Используемый здесь термин "праймер" означает олигонуклеотид, который независимо от того, встречается ли он в природе в виде очищенного рестрикционного гидролизата или он был получен синтетически, способен действовать как участок инициации синтеза при помещении его в условия, при которых индуцируется синтез продукта удлинения праймера, являющегося комплементарным нуклеотидной цепи, то есть в присутствии нуклеотидов и индуцирующего агента, такого как полимераза ДНК, и при подходящих температуре и рН. Этот праймер может быть либо одноцепочечным, либо он должен быть достаточно длинным для того, чтобы инициировать синтез нужного продукта удлинения в присутствии индуцирующего агента. Точная длина этого праймера зависит от многих факторов, включая температуру, источник происхождения этого праймера и способ его использования. Так, например, для диагностических целей в зависимости от вариабельности целевой последовательности олигонуклеотидный праймер обычно содержит 15-25 или более нуклеотидов, хотя он может содержать и меньшее количество нуклеотидов.
Используемые здесь праймеры выбирают так, чтобы они были существенно комплементарными различным цепям конкретной целевой последовательности ДНК. Это означает, что указанные праймеры должны быть достаточно комплементарными для того, чтобы они могли гибридизоваться с их соответствующими цепями. Поэтому праймерная последовательность не должна обязательно иметь последовательность, идентичную последовательности матрицы. Так, например, некомплементарный нуклеотидный фрагмент может быть присоединен к 5’-концу праймера, а остальная праймерная последовательность должна быть комплементарна этой цепи. Альтернативно, некомплементарные основания или более длинные последовательности могут быть встроены в праймер при условии, что эта праймерная последовательность имеет достаточную комплементарность с этими последовательностями или гибридизуется с ними, образуя тем самым матрицу для синтеза продукта удлинения.
Используемые здесь термины "рестрикционные эндонуклеазы" и "рестрикционные ферменты" означают ферменты, каждый из которых разрезает двухцепочечную ДНК в специфической нуклеотидной последовательности или возле нее.
Клетка является "трансформированной" экзогенной или гетерологичной ДНК при введении этой ДНК в клетку. Трансформирующая ДНК может быть, а может и не быть, интегрированной (ковалентно присоединенной) в геном этой клетки. Например, в клетках прокариотов, дрожжей и млекопитающих эта трансформирующая ДНК может быть расположена на эписомном элементе, таком как плазмида. Что касается эукариотических клеток, то стабильно трансформированной клеткой является клетка, в которой эта трансформирующая ДНК интегрируется в хромосому так, что она наследуется дочерними клетками посредством хромосомной репликации. Эта стабильность продемонстрирована способностью эукариотической клетки создавать клеточные линии или клоны, составляющие популяцию дочерних клеток, содержащих эту трансформирующую ДНК. "Клон" представляет собой популяцию клеток, происходящих от одной клетки или ее предшественника посредством митоза. "Клеточная линия" представляет собой клон первичной клетки, которая способна стабильно расти in vitro с продуцированием множества генераций.
Две последовательности ДНК являются "в основном, гомологичными", если на протяжении определенной длины этих последовательностей ДНК имеется, по крайней мере, примерно 75%-ное (предпочтительно, по крайней мере, примерно 80%-ное, а наиболее предпочтительно, по крайней мере, примерно 90%-ное или 95%-ное) соответствие нуклеотидов. Последовательности, которые являются в основном гомологичными, могут быть идентифицированы путем сравнения этих последовательностей с использованием стандартного программного обеспечения, имеющегося в банке данных последовательностей, или в эксперименте по Саузерн-гибридизации, например в строгих условиях, определенных для этой конкретной системы. Соответствующие условия гибридизации могут быть определены каждым специалистом. Например, Maniatis et al., см. выше; DNA-Cloning, A Practical Approach", Vols. I & II, см. выше; "Nucleic Acid Hybridization", см. выше.
"Гетерологичная область" конструкции ДНК представляет собой идентифицируемый сегмент ДНК в более крупной молекуле ДНК, который не встречается в природе в ассоциации с этой более крупной молекулой. Таким образом, если эта гетерологичная область кодирует ген млекопитающего, то этот ген обычно фланкирован ДНК, которая не фланкирует геномную ДНК млекопитающего в геноме организма-источника. В другом примере кодирующая последовательность представляет собой конструкцию, где сама кодирующая последовательность не встречается в природе (например, кДНК, где геномная кодирующая последовательность содержит интроны или синтетические последовательности, имеющие кодоны, отличающиеся от нативного гена). Аллельное разнообразие или природные мутации не приводят к образованию гетерологичной области ДНК, определенной в настоящем описании.
Метками, наиболее часто используемыми в этих исследованиях, являются радиоактивные элементы, ферменты, химические вещества, которые флуоресцируют при их облучении ультрафиолетовым светом, и другие. Известен ряд флуоресцентных материалов, которые могут быть использованы в качестве меток. Такими материалами являются, например, флуоресцеин, родамин, аурамин, техасский красный, синий АМСА и люциферовый желтый. Конкретным детектирующим материалом является антикроличье антитело, вырабатываемое у коз и конъюгированное с флуоресцеином посредством изотиоцианата.
Белки могут быть также помечены радиоактивным элементом или ферментом. Радиоактивная метка может быть детектирована любыми существующими в настоящее время методами подсчета. Предпочтительный изотоп может быть выбран из 3Н, 14С, 32P, 35S, 36Cl, 51Cr, 57Co, 58Co, 59Fe, 90Y, 125I, 131I и 186Re.
Могут быть также использованы ферментные метки, которые могут быть детектированы любыми из используемых в настоящее время колориметрических, спектрофотометрических, флуороспектрофотометрических, амперометрических или газометрических методов. Этот фермент конъюгируют с выбранной частицей путем реакции с мостикобразующими молекулами, такими как карбодиимиды, диизоцианаты, глутаральдегид и т.п. Известно множество ферментов, которые могут быть использованы в этих процедурах. Предпочтительными являются пероксидаза, β-глюкуронидаза, β-D-глюкозидаза, β-D-галактозидаэа, уреаза, глюкозооксидаза + пероксидаза и щелочная фосфатаза. Описание альтернативных материалов и методов мечения можно найти, например, в патентах США №№ 3654090, 3850752 и 4016043.
Специалистами была разработана и использована конкретная аналитическая система, известная как рецепторный анализ. В рецепторном анализе анализируемый материал соответствующим образом метят, а затем некоторое количество клеточных тестируемых колоний инокулируют определенным количеством этой же метки, после чего проводят исследование на связывание для определения степени, с которой этот меченый материал связывается с клеточными рецепторами. Этим способом могут быть определены различия в аффинности данных материалов.
Специалистами используется анализ, известный как "цис/транс"-анализ. Короче говоря, в этом анализе используются две генетических конструкции, одна из которых обычно представляет собой плазмиду, непрерывно экспрессирующую нужный конкретный рецептор при ее трансфекции в соответствующую клеточную линию, а вторая представляет собой плазмиду, экспрессирующую репортер, такой как люцифераза, под контролем рецептор/лигандного комплекса. Так, например, если необходимо определить, является ли данное соединение лигандом для конкретного рецептора, то одна из этих плазмид должна быть сконструирована так, чтобы она обеспечивала экспрессию рецептора в выбранной клеточной линии, а вторая плазмида должна содержать промотор, присоединенный к гену люциферазы, в который был встроен реактивный элемент для данного конкретного рецептора. Если тестируемое соединение является агонистом для данного рецептора, то данный лиганд будет образовывать комплекс с этим рецептором и образовавшийся комплекс будет связываться с данным реактивным элементом и инициировать транскрипцию гена люциферазы. Затем индуцированную хемилюминесценцию измеряют фотометрически, строят кривые зависимости "доза-ответ" и сравнивают их с кривыми, полученными для известных лигандов. Вышеуказанный протокол подробно описан в патенте США № 4981784.
Используемый здесь термин "хозяин" означает не только прокариоты, но также и эукариоты, такие как клетки дрожжей, растений и животных. Рекомбинантная молекула ДНК или ген, которые кодируют иммунореактивный белок массой 120 кДа Ehrlichia canis настоящего изобретения, могут быть использованы для трансформации хозяина любым методом, хорошо известным каждому специалисту. Для трансформации прокариотов особенно предпочтительным является использование вектора, содержащего кодирующие последовательности гена, кодирующего иммунореактивный белок массой 120 кДа Ehrllchia canis настоящего изобретения.
Прокариотическими хозяевами могут быть E. coli, S. typhimurium, Serratia marcescenns, Mycobaсtrium vaccae и Bacillus subtllis. Эукариотическими хозяевами являются дрожжи, такие как Pichia pastoris, клетки млекопитающих и клетки насекомых.
В основном вместе с этим хозяином используются экспрессирующие векторы, содержащие промоторные последовательности, которые способствуют эффективной транскрипции встроенного фрагмента ДНК. Экспрессирующий вектор обычно содержит сайт инициации репликации, промотор(ы), терминатор(ы), а также специфические гены, способные обеспечивать фенотипический отбор в трансформированных клетках. Для достижения оптимального клеточного роста эти трансформированные хозяева могут быть ферментированы и культивированы способами, известными специалистам.
Настоящее изобретение относится в основном к чистой ДНК, кодирующей иммунореактивный белок массой 120 кДа Ehrlichia canis, цепь ДНК которой гибридизуется в условиях высокой строгости с зондом, содержащем последовательность, по крайней мере, из 15 следующих друг за другом нуклеотидов (SEQ ID NO: 6). Этот белок массой 120 кДа, кодируемый ДНК настоящего изобретения, может иметь последовательность, по крайней мере, на 80% идентичную (предпочтительно на 85%, более предпочтительно на 90%, а наиболее предпочтительно на 95%) последовательности нуклеиновых кислот, перечисленных на Фиг.3 (SEQ ID NO: 7). Более предпочтительно, чтобы эта ДНК включала кодирующую нуклеотидную последовательность SEQ ID NO: 8 или вырожденный вариант такой последовательности.
Зонд, с которым гибридизуется ДНК по настоящему изобретению, предпочтительно состоит из последовательности, по крайней мере, в 20 следующих друг за другом нуклеотидов, более предпочтительно 40 нуклеотидов, еще более предпочтительно 50 нуклеотидов, а наиболее предпочтительно в 100 нуклеотидов или более (вплоть до 100%) кодирующей последовательности нуклеотидов, перечисленных в SEQ ID NO: 8, или комплементарных нуклеотидов. Такой зонд может быть использован для детекции экспрессии гена иммунореактивного белка массой 120 кДа Ehrlichia canis в клетке человека методом, включающим стадии: а) взаимодействие мРНК, полученной из этой клетки, с меченым зондом для гибридизации; и (b) детектирования гибридизации этого зонда с мРНК.
Настоящее изобретение также относится в основном к чистой ДНК, содержащей последовательность, по крайней мере, из 15 следующих друг за другом нуклеотидов (предпочтительно 20, более предпочтительно 30, еще более предпочтительно 50, а наиболее предпочтительно, всех нуклеотидов) области, состоящей из нуклеотидов, перечисленных в SEQ ID NO: 8.
Термин "высокая жесткость" относится к гибридизации ДНК и условиям промывок, характеризующимся высокой температурой и низкой концентрацией соли, например, к условиям промывок при 65°С при концентрации соли приблизительно 0,1 × SSC или их функционального эквивалента. Так, например, условия высокой строгости могут включать гибридизацию при около 42°С в присутствии приблизительно 50% формамида; первую промывку приблизительно при 65°С примерно 2 × SSC, содержащим 1% ДСН; а затем вторую промывку приблизительно 0,1 × SSC при около 65°С.
"В основном чистая ДНК" означает ДНК, которая не является частью среды, в которой обычно присутствует эта ДНК благодаря отделению (частичной или полной очистке) некоторых или всех молекул из этой среды, или благодаря модификации последовательностей, фланкирующих эту рассматриваемую ДНК. Этот термин, кроме того, включает, например, рекомбинантную ДНК, которую вводят в вектор, в автономно реплицирующуюся плазмиду или вирус, или в геномную ДНК прокариота или эукариота; или которая существует в виде отдельной молекулы (например, кДНК или геномная ДНК или кДНК-фрагмент, продуцированный с помощью полимеразной цепной реакции (ПЦР) или расщепления эндонуклеазой) независимо от других последовательностей. Этот термин также означает рекомбинантную ДНК, которая является частью гибридного гена, кодирующего другую полилептидную последовательность, например гибридный белок. Этот термин также означает рекомбинантную ДНК, которая включает часть нуклеотидов, перечисленных в SEQ ID NO: 8, и которая кодируют альтернативный вариант сллайсинга гена, кодирующего иммунореактивный белок массой 120 кДа Ehrlichia canis.
Эта ДНК может иметь последовательность, которая, по крайней мере, примерно на 70%, предпочтительно, по крайней мере, на 75% (например, по крайней мере, на 80%), а наиболее предпочтительно, по крайней мере, на 90% идентична кодирующей последовательности нуклеотидов, перечисленных в SEQ ID NO:8. Идентичность этих двух последовательностей непосредственно зависит от числа соответствий или идентичных положений. Если положение субъединицы в обеих двух последовательностях занято аналогичной мономерной субъединицей, например, если данное положение в каждой из двух молекул ДНК занято аденином, то в этом положении они являются идентичными. Так, например, если 7 положений в последовательности из 10 нуклеотидов идентичны соответствующим положениям во второй 10-нуклеотидной последовательности, то эти две последовательности являются идентичными на 70%. Длина сравниваемых последовательностей в основном составляет, по крайней мере, 50 нуклеотидов, предпочтительно, по крайней мере, 60 нуклеотидов, более предпочтительно, по крайней мере, 75 нуклеотидов, а наиболее предпочтительно, 100 нуклеотидов. Идентичность последовательности обычно определяют с использованием программного обеспечения для анализа последовательности (например, Sequence Analysis Software Package of the Genetics Computer Group, University of Wisconsin Biotechnology Center, 1710 University Avenue, Madison, WI 53705).
Настоящее изобретение относится к вектору, включающему последовательность ДНК, которая кодирует ген, кодирующий иммунореактивный белок массой 120 кДа Ehrlichia cants, и где указанный вектор способен реплицироваться в хозяине и включает при его правильном присоединении: а) сайт инициации репликации; b) промотор; и с) последовательность ДНК, кодирующую указанный белок массой 120 кДа. Предпочтительно, чтобы вектор по настоящему изобретению содержал часть последовательности ДНК, показанной в SEQ ID NO: 8. "Вектор" может быть определен как реплицируемая нуклеотидная конструкция, например плазмида или вирусная нуклеиновая кислота. Векторы могут быть использованы для амплификации и/или экспрессии нуклеотидной кислоты, кодирующей иммунореактивный белок массой 120 кДа Ehrlichia canis. Экспрессирующий вектор представляет собой реплицируемую конструкцию, в которой нуклеотидная последовательность, кодирующая полипептид, правильно присоединена к подходящим регуляторным последовательностям, способными осуществлять экспрессию этого полипептида в клетке. Такие регуляторные последовательностях должны быть выбраны в зависимости от выбранных клеток и способа трансформации. В основном регуляторными последовательностями являются транскрипционный промотор и/или энхансер, подходящие сайты связывания с рибосомной мРНК и последовательности, которые регулируют терминацию транскрипции и трансляции. Для конструирования экспрессирующих векторов, содержащих соответствующие сигналы регуляции транскрипции и трансляции, могут быть использованы методы, хорошо известные специалистам. Например, методы, описанные Сэмбруком и др., 1989, в работе Molecular Cloning: A Laboratory Manual (2nd Ed.), Cold Spring Harbor Press, N.Y. Ген и его последовательности регуляции транскрипции считаются "правильно присоединенными" в том случае, если эти последовательности регуляции транскрипции эффективно осуществляют регуляцию транскрипции этого гена. Векторами по настоящему изобретению являются, но не ограничиваются ими, плазмидные векторы и вирусные векторы. Предпочтительными вирусными векторами по настоящему изобретению являются векторы, происходящие от ретровирусов, аденовируса, аденоассоциированного вируса, вируса SV40 или вируса герпеса.
Термин "в основном чистый белок" означает белок, который был выделен, по крайней мере, из некоторых компонентов, с которыми он обычно ассоциирован. Обычно этот белок является в основном чистым в том случае, если он, по крайней мере, на 60%, по массе не содержит белков и других природных органических молекул, с которыми он обычно ассоциирован in vivo. Предпочтительно, чтобы чистота этого препарата составляла, по крайней мере, 75%, более предпочтительно, по крайней мере, 90%, а наиболее предпочтительно, по крайней мере, на 99% по массе. В основном чистый иммунореактивный белок массой 120 кДа Ehrlichia canis может быть получен, например, путем экстракции из природного источника; путем экспрессии рекомбинантной нуклеиновой кислоты, кодирующей иммунореактивный белок массой 120 кДа Ehrlichia canis; или путем химического синтеза белка массой 120 кДа. Чистота может быть определена любым подходящим методом, например с помощью колоночной хроматографии, такой как иммуноаффинная хроматография с использованием антитела, специфичного для иммунореактивного белка массой 120 кДа Ehrlichia canis, электрофореза в полиакриламидном геле или ВЭЖХ-анализа. Белок считается в основном не содержащим ассоциированных с ним природных компонентов в том случае, если он отделен, по крайней мере, от некоторых примесей, с которыми он ассоциирован в природе. Таким образом, белок, который был химически синтезирован или продуцирован в клеточной системе, не являющейся клеткой, от которой он по своей природе происходит, будет называться, по определению, белком, в основном не содержащим природных ассоциированных с ним компонентов. В соответствии с этим в основном чистыми белками являются эукариотические белки, синтезированные в E. coli, в других прокариотах или в каком-либо другом организме, в котором они не встречаются в природе.
Помимо в основном полноразмерных белков по настоящему изобретению также относится к фрагментам (например, антигенным фрагментам) иммунореактивного белка массой 120 кДа Ehrlichia canis (SEQ ID NO: 7). Используемый здесь термин "фрагмент" относится к полипептиду, который обычно имеет, по крайней мере, 10 остатков, а в основном, по крайней мере, 20 остатков, а предпочтительно, по крайней мере, 30 (например, 50) остатков, но при этом меньшее число остатков, чем полная интактная последовательность. Фрагменты иммунореактивного белка массой 120 кДа Ehrlichia canis могут быть генерированы методами, известными специалистам, например путем ферментативного гидролиза природного или рекомбинантного иммунореактивного белка массой 120 кДа Ehrlichia canis, с помощью техники рекомбинантных ДНК с использованием экспрессирующего вектора, который кодирует определенный фрагмент иммунореактивного белка массой 120 кДа Ehrlichia canis, или методами химического синтеза. Способность фрагмента-кандидата проявлять свойства иммунореактивного белка массой 120 кДа Ehrlichia canis (например, связываться с антителом, специфичным для иммунореактивного белка массой 120 кДа Ehrlichia canis) может быть оценена описанными здесь методами. Очищенный иммунореактивный белок массой 120 кДа Ehrlichia canis или антигенные фрагменты иммунореактивного белка массой 120 кДа Ehrlichia canis могут быть использованы для генерирования новых антител или для тестирования существующих антител (например, в качестве позитивного контроля в диагностическом анализе) в соответствии со стандартными схемами, известными специалистам. Настоящее изобретение также включает поликлональные антисыворотки, продуцированные с использованием иммунореактивного белка массой 120 кДа Ehrlichia canis или фрагмента иммунореактивного белка массой 120 кДа Ehrlichia canis в качестве иммуногена, например, у кроликов. При этом используются стандартные протоколы продуцирования моноклональных и поликлональных антител, известные специалистам. Моноклональные антитела, генерированные с помощью этой процедуры, могут быть скринированы на их способность к идентификации рекомбинантных кДНК-клонов Ehrlichia canis и для их дифференциации от известных кДНК-клонов.
Кроме того, в настоящее изобретение входят фрагмент иммунореактивного белка массой 120 кДа Ehrlichia canis, которые, по крайней мере частично, кодируются частями SEQ ID NO: 7, например продукты альтернативного мРНК-сплайсинга, или альтернативных событий процессинга белка, или в которых участок этой последовательности был делегирован. Фрагмент этого белка или интактный иммунореактивный белок массой 120 кДа Ehrlichia canis может быть ковалентно связан с другим полипептидом, который, например, действует как метка, лиганд или агент, повышающий антигенность.
Термин "фармацевтически приемлемый" относится к молекулярным частицам и к композициям, которые при их введении человеку не вызывают аллергических или им подобных нежелательных реакций. Способы получения водной композиции, которая содержит белок в качестве активного ингредиента, хорошо известны специалистам. Обычно такие композиции приготавливают в виде инъекций либо в виде жидких растворов, либо в виде суспензий; при этом могут быть также получены твердые формы, которые перед инъекцией могут быть растворены или суспендрованы в жидкости. Этот препарат может быть также эмульгирован.
Белок может быть приготовлен в виде композиции в нейтральной или солевой форме. Фармацевтически приемлемыми солями являются кислотно-аддитивные соли (образованные свободными аминогруппами этого белка) и соли, образованные неорганическими кислотами, такими как, например, хлористоводородная кислота или фосфорная кислота, или органическими кислотами, такими как уксусная, щавелевая, винная, миндальная и т.п. Соли, образованные свободными карбоксильными группами, могут быть также получены из неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа (3), и из органических оснований, таких как изопропиламин, триметиламин, гистидин, прокаин и т.п. После получения композиций растворы вводят способом, подходящим для данной лекарственной композиции, и в таком количестве, которое является терапевтически эффективным. Эти композиции вводят в виде различных лекарственных форм, таких как, растворы для инъекций.
Для парентерального введения, например, в виде водного раствора, этот раствор должен быть подходящим образом забуференным, если это необходимо, а жидкому разбавителю сначала следует придать изотоничность с использованием достаточного количества физиологического раствора или глюкозы. Эти конкретные водные растворы являются особенно подходящими для внутривенного, внутримышечного, подкожного и внутрибрюшинного введения. В соответствии с этим могут быть использованы стерильные водные среды, которые будут известны специалистам из описания по настоящему изобретению. Например, одна доза может быть растворена в 1 мл изотоничного раствора NaCl и может быть либо добавлена в 1000 мл жидкости для подкожного введения либо она может быть введена в участок, предназначенный для вливания (Например, "Remingtons Pharmaceutical Sciences" 15th Edition, pages 1035-1038 and 1570-1580). При необходимости дозы могут быть несколько изменены в зависимости от состояния субъекта, подвергаемого лечению. В любом случае подходящую дозу для каждого конкретного субъекта назначает специалист, ответственный за это лечение.
Специалистам хорошо известно, что данный полипептид может различаться по своей иммуногенности. Поэтому в большинстве случаев необходимо, чтобы иммуноген (например, полипептид по настоящему изобретению) был связан с носителем. Предпочтительными примерами таких носителей являются гемоцианин лимфы улитки (KLH) и альбумин сыворотки человека. Другими носителями могут быть различные лимфокины и адъюванты, такие как IL2, IL4, IL8 и другие.
Средства для конъюгирования полипептида с белком-носителем хорошо известны специалистам и ими являются глутаральдегид, м-малеимидобензоил-N-гидроксисукцинимидоэфир, карбодиимид и бис-диазотированный бензидин. Следует также отметить, что этот пептид может быть конъюгирован с белком методами генной инженерии, хорошо известными специалистам.
Специалистам также хорошо известно, что иммуногенность конкретного иммуногена может быть повышена с использованием неспецифических стимуляторов иммунного ответа, известных как адъюванты. Предпочтительными примерами адъювантов являются полный адъювант BCG, Detox, (RIBI, Immunochem Research Inc.), ISCOMS и гидроксид алюминия (Superphos, Biosector).
Вакцинные препараты, которые содержат пептидные последовательности в качестве активных ингредиентов, в основном хорошо известны специалистам и проиллюстрированы в Патентах США № 4608251; 4601903; 4599231; 4599230; 4596792 и 4578770, каждый из которых вводится в настоящее описание посредством ссылки. Обычно такие вакцины получают в виде инъекций, либо в виде жидких растворов, либо в виде суспензий; при этом могут быть также получены твердые формы, которые перед инъекцией могут быть растворены или суспендированы в жидкости. Этот препарат может быть также эмульгирован. В большинстве случаев активный иммуногенный ингредиент смешивают с наполнителями, которые являются фармацевтически приемлемыми и совместимыми с этим активным ингредиентом. Подходящими наполнителями являются, например, вода, физиологический раствор, декстроза, глицерин, этанол или т.п. и их комбинации. Кроме того, если это необходимо, указанная вакцина может содержать небольшое количество добавок, таких как смачивающие или эмульгирующие агенты, рН-корректирующие забуферивающие агенты или адъюванты, которые повышают эффективность этих вакцин.
Белок массой 120 кДа, вероятно, представляет собой адгезин Ehrlichia spp. и дифференциально экспрессируется на поверхности клеток Е. chaffeensis. Ген Е. canis амплифицировали посредством ПЦР с использованием праймеров, происходящих от последовательности ДНК, фланкирующей ген Е. chaffeensis. Ген Е.canis клонировали, секвенировали и сверхэкспрессировали в Escherichia coli. Белок массой 120 кДа Е. canis содержит 14 тандемных повторяющихся единиц, каждая из которых имеет 36 аминокислот. Последовательности ДНК этих повторов на 94% гомологичны друг другу. Полная аминокислотная последовательность белка массой 120 кДа Е. canis на 30% гомологична белку массой 120 кДа Е. chaffeensis. Области повторов белков массой 120 кДа этих двух видов имеют общие аминокислотные последовательности, которые, как предполагается, являются поверхностно-экспонированными. Рекомбинантный белок массой 120 кДа Е. canis реагировал с сывороткой, полученной от выздоравливающих собак с эрхилиозом.
Настоящее изобретение относится к гену, кодирующему иммунореактивный белок массой 120 кДа Ehrlichia canis, и к рекомбинантному белку, кодируемому этим геном. В одном из вариантов своего осуществления настоящее изобретение относится к гену, кодирующему иммунореактивный белок массой 120 кДа Ehrlichia canis. Указанный белок массой 120 кДа предпочтительно имеет аминокислотную последовательность SEQ ID NO: 8, а указанный ген имеет нуклеотидную последовательность SEQ ID NO:7. В предпочтительном варианте осуществления настоящее изобретение относится к экспрессирующему вектору, содержащему ген, кодирующий иммунореактивный белок массой 120 кДа Ehrlichia canis, и при введении этого вектора в клетку он способен экспрессировать этот ген.
В другом варианте своего осуществления настоящее изобретение относится к рекомбинантному белку, содержащему аминокислотную последовательность SEQ ID NO:8. Эта аминокислотная последовательность предпочтительно кодируется нуклеотидной последовательностью SEQ ID NO:7. Указанный рекомбинантный белок содержит 14 тандемных повторяющихся единиц, каждая из которых имеет 36 аминокислот. Более предпочтительно, если указанные повторяющиеся единицы являются гидрофильными. Еще более предпочтительно, если указанный рекомбинантный белок является антигеном.
В предпочтительном варианте осуществления настоящее изобретение относится к способу продуцирования рекомбинантного белка массой 120 кДа, включающему стадии: получения вектора, который содержит экспрессирующую область, включающую кодирующую последовательность SEQ ID NO:8, правильно присоединенную к промотору; трансфекции этого вектора в клетку; и культивирования этой клетки в условиях, эффективных для экспрессии указанной экспрессирующей области.
Используемый здесь термин "комплемент" означает цепь нуклеиновой кислоты, которая гибридизуется с первой нуклеотидной последовательностью с образованием двухцепочечной молекулы в строгих условиях. Строгими условиями являются такие условия, при которых происходит гибридизация между двумя нуклеотидными последовательностями, имеющими высокую степень гомологии, но не происходит гибридизации случайных последовательностей. Так, например, гибридизацию при низкой температуре и/или высокой ионной силе определяют как гибридизацию в условиях низкой строгости, а гибридизацию при высокой температуре и/или низкой ионной силе определяют как гибридизацию в условиях высокой строгости. Следует отметить, что для зондов определенной длины, для последовательностей определенной длины и с определенным содержанием оснований и в случае присутствия формамида в гибридизационной смеси используются температура и ионная сила нужной строгости.
Используемый здесь термин "сконструированная" или "рекомбинантная" клетка означает клетку, в которую был введен рекомбинантный ген, такой как ген, кодирующий антиген Е. chaffeensis. Поэтому сконструированные клетки отличаются от природных клеток, которые не содержат рекомбинантно введенного гена. Таким образом, сконструированные клетки представляют собой клетки, имеющие ген или гены, искусственно введенные человеком. Рекомбинантно введенные гены присутствуют либо в форме кДНК-гена, либо в виде копии геномного гена, либо они представляют собой гены, расположенные рядом с промотором, который обычно не ассоциируется с этим введенным геном. Кроме того, этот рекомбинантный ген может быть интегрирован в геном хозяина либо он может присутствовать в векторе или в бактериальном геноме, трансфицированном в эту клетку-хозяина.
Нижеследующие примеры приводятся для иллюстрации различных вариантов осуществления изобретения и не должны рассматриваться как ограничение по настоящему изобретению.
Пример 1
Эрлихия
Штамм Оклахома Ehrlichia canis был предоставлен Dr. Jacqueline Dawson (Centers for Disease Control, Atlanta, GA). Штамм Флорида и три изолята Северной Каролины E. canis, (Demon, DJ и Jake) были предоставлены Dr. Edward В. Breitschwerdt (College of Veterinary Medicine, North Carolina State University, Raleigh, NC). Штамм Луизиана Е. canis был предоставлен Dr. R.E. Corstvet (Louisiana State University, Baton Rouge, Louisiana). Ehrlichia культивировали в клетках DH82, макрофагоподобной клеточной линии собак (Dawson J.E., 1991). Клетки DH82 собирали с помощью скребка для клеток при 100%-ном инфицировании эрлихией. Клетки центрифугировали при 17400 × g в течение 20 минут. Клеточный осадок дважды подвергали разрушению на ультразвуковом дезинтеграторе Braun-Sonic 2000 при 40 ватт в течение 30 секунд на льду. Клеточный лизат загружали на ступенчатые градиенты 42%-36%-30% ренографина, а затем центрифугировали при 80000 × g в течение 60 минут. Эрлихии в тяжелых и легких полосах собирали (Weiss E, 1975) и промывали путем центрифугирования с сахарозо-фосфатно-глутаматным буфером (SPG, 218 мМ сахарозы, 3,8 мМ КН2РО4, 7,2 мМ К2НРО4, 4,9 мМ глутамата, рН 7,0).
Пример 2
Получение ДНК
Геномную ДНК Ehrlichia canis получали из эрлихии, очищенной в градиенте плотности ренографина с использованием набора для экстракции нуклеиновой кислоты IsoQuick (ORCA Research Inc., Bothell, WA) в соответствии с инструкциями производителя. Плазмидную ДНК очищали с использованием набора для выделения плазмид с высокой степенью чистоты (Boehringer Mannheim Corp., Indianapolis, IN). ПЦР-продукт очищали с использованием набора для ПЦР-очистки QIAquick PCR Purification Kit (QIAGEN Inc., Santa Clarita, CA).
Пример 3
ПЦР-амплификация гена белка массой 120 кДа E. canis
Праймеры конструировали на основе последовательности ДНК гена белка массой 120 кДа Е. chaffeensis (SEQ ID NO:1-6, Фиг.1) (Yu, X.J. 1996). Ген белка массой 120 кДа Е. canis амплифицировали с помощью ПЦР с 30 циклами: 94°С, 30 с; 52°С 1 мин; и 72°С, 2 мин. ПЦР-амплифицированные продукты клонировали в клонирующий вектор pCR2.1 ТА (Invitrogen, Carlsbad, CA).
Пример 4
Секвенирование ДНК
ДНК секвенировали на секвенаторе ДНК ABI Prism 377 DNA Sequencer (Perkin-Elmer Applied Biosystems, Foster City, CA).
Пример 5
Однонаправленная делеция гена белка массой 120 кДа Е. canis
Область повторов делегировали с 5’-конца гена белка массой 120 кДа E.canis с использованием частичного гидролиза рестриктирующей эндонуклеазой SpeI. Плазмиду pCR120 сначала полностью гидролизовали ферментом XbaI, который имел уникальный сайт расщепления на этой плазмидной последовательности возле 5’-конца гена белка массой 120 кДа E.canis. Затем эту плазмиду частично гидролизовали ферментом SpeI. SpeI имел уникальный сайт расщепления в каждом повторе гена белка массой 120 кДа E. canis, но не имел сайта расщепления вне области повторов, включающей последовательность плазмидного вектора. Для гарантии соответствующего репрезентативного частичного гидролиза от каждой смеси для гидролиза брали аликвоту через каждые 5 минут. Гидролиз прекращали путем добавления EDTA до конечной концентрации 50 мМ и нагревания при 70°С в течение 10 минут.
После полного расщепления ферментом XbaI и частичного расщепления ферментом SреI удаляли (делегировали) различное число повторяющихся единиц, расположенных между XbaI-сайтом и каждым SpeI-сайтом рестрикции, для генерирования делегированных плазмидных ДНК с несовместимыми концами (XbaI у 3’-конца и SpeI у 5’-конца). Смесь, гидролизованную рестриктирующими ферментами, обрабатывали фрагментом Кленова для достраивания концов. Затем рестрикционную смесь разделяли путем электрофореза на агарозном геле для удаления плазмид из внутренних повторов, поскольку размеры их молекул значительно различались. Смесь делегированных плазмид экстрагировали из геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (QIAGEN Inc., Santa Clarita) и подвергали рециркуляризации с использованием лигазы Т4. Эти делегированные плазмиды грансформировали в штамм DH5α E. coli и отбирали для секвенирования в соответствии с их размерами. Альтернативно область повторов гена белка массой 120 кДа E. canis однонаправлено делегировали с 3’-конца с использованием экзонуклеазы III и системы Erase-a-Base (Promega Corp., Madison, WI) в соответствии с инструкциями производителя.
Пример 6
Определение числа повторов в гене белка массой 120 кДа E. Canis
ПЦР-амплифицированный ген белка массой 120 кДа E. canis от всех E. canis клонировали в клонирующий вектор pCR2.1 ТА (Invitrogen). Рекомбинантную плазмиду сначала расщепляли ферментом EcoRI, а затем частично расщепляли ферментом SpeI, как описано выше. Смесь для расщепления разделяли путем электрофореза в 1% агарозном геле и переносили в условиях вакуума на найлоновую мембрану. Полосы ДНК на найлоновой мембране гибридизовали с олигонуклеотидным зондом, происходящим от последовательности, расположенной выше от области повторов гена белка массой 120 кДа Е. canis. Зонды ДНК метили с использованием дигоксигенина-11-dUTP с помощью набора для присоединения DIG к олигонуклеотиду (Boehringer Mannhiem Co., Indianapolis, In) в соответствии с протоколом производителей.
Пример 7
Анализ генов
Последовательность ДНК и выведенные аминокислотные последовательности анализировали с использованием программного обеспечения Wisconsin GCG (Genetic Computer Group, Inc., Madison, WI) и программного обеспечения DNASTAR (DNASTAR, Inc., Madison, WI). Сигнальную последовательность выведенного белка массой 120 кДа анализировали с использованием программы PSORT (World Wide Web site at URL: pttp://psort. nibb. ac. jp), что позволяет с помощью методов McGeoch (D.J.McGeoch, Virus Research, 3, 271, 1985) и Von Heijne (von Heijne G. 1986, Nucl.Acids Res., 14, 4683) предполагать присутствие сигнальных последовательностей, а с помощью метода Кляйна и др. (P. Klein, M. Kanehisa & C. DeLisi, Biochem. Biophys. Acta, 815, 468, 1985) детектировать возможные трансмембранные домены.
Пример 8
Экспрессия гена белка массой 120 кДа Е. canis в E. coli
Прямое клонирование гена белка массой 120 кДа Е. canis в экспрессирующий вектор pGEX (Amersham Pharmacia Biotech, Piscataway, NJ) не происходило из-за отсутствия соответствующих сайтов расщепления рестриктирующих эндонуклеаз на обеих последовательностях ДНК гена белка массой 120 кДа Е. canis и сайта множественного клонирования вектора pGEX. Кодирующую область гена белка массой 120 кДа Е. canis от нуклеотида 175 до нуклеотида 1793 амплифицировали посредством ПЦР с использованием прямого праймера (SEQ ID NO:13) и обратного праймера (SEQ ID NO:14). ПЦР-продукты клонировали в клонирующий вектор pCR2.1 ТА (Invitrogen) с созданием EcoRI-сайта рестрикции на обеих концах вставки. Эту вставку в рекомбинантной плазмиде pCR2.1 разрезали EcoRI и выделяли из плазмидной ДНК в агарозном геле. Эту вставку экстрагировали из агарозного геля с использованием набора для экстракции из геля QIAquick Gel Extraction Kit (QIAGEN Inc., Santa Clarita) и клонировали в EcoRI-гидролизованный вектор pGEX. Белок E. canis экспрессировали в штамме BL21 E. coli в виде GST-гибридного белка. Этот GST-гибридный белок массой 120 кДа подвергали аффинной очистке с использованием гранул из глутатион-сефарозы 4В (Amersham Pharmacia Biotech, Piscataway, NJ). Рекомбинантный белок массой 120 кДа Е. canis отщепляли от GST-гибридного белка тромбином.
Пример 9
Иммунизация мышей
Мышей иммунизировали рекомбинантным белком массой 120 кДа Е. canis для вырабатывания антисыворотки. Рекомбинантный белок р120 смешивали с равным объемом полного адъюванта Фрейнда для первой инъекции и с неполным адъювантом Фрейнда для последующих инъекций. Мышей иммунизировали внутрибрюшинно или подкожно 50 мкг рекомбинантного гибрида белка массой 120 кДа и GST в каждом из 4 случаев.
Пример 10
Детекция гена белка массой 120 кДа Е. canis
В попытке детекции гена белка массой 120 кДа Е. canis использовали Саузерн-блоттинг. 1,2-т.п.н. фрагмент ДНК, амплифицированный из гена белка массой, 120 кДа Е. chaffeensis посредством ПЦР с использованием пары праймеров pxcf3b (SEQ ID NO:3) и pxar4 (SEQ ID NO:5, Фиг. 1), метили дигоксигенином-11-dUTP и использовали в качестве зонда для детекции гомологичного гена в Е. canis путем Саузерн-блоттинга. Саузерн-блоттинг показал, что зонд для гена белка массой 120 кДа Е. chaffeensis не гибридизовался с EcoRI-гидролизованной геномной ДНК Е. canis в условиях, при которых этот зонд давал сильную гибридизацию с геномной ДНК Е. chaffeensis. Этот результат указывает на то, что ген белка массой 120 кДа Е. canis значительно отличается от гомологичного гена белка массой 120 кДа Е. chaffeensis.
Хотя сходство между генами белка массой 120 кДа Е. canis и Е. chaffeensis является низким, однако предполагается, что они могут содержать некоторые консервативные домены, которые могут быть использованы для конструирования ПЦР-праймеров, пригодных для амплификации обоих генов белка массой 120 кДа. Поэтому этот гомологичный ген белка массой 120 кДа в Е. canis (штамм Оклахома) был дополнительно амплифицирован с помощью ПЦР. Праймеры, происходящие от гена белка массой 120 кДа Е. chaffeensis и последовательностей, фланкирующих этот ген, были использованы ранее для секвенирования гена белка массой 120 кДа Е. chaffeensis (Фиг.1). Для конструирования девяти пар праймеров использовали три прямых праймера (SEQ ID NO:1-3), каждый из которых составлял 3 пары с обратными праймерами (SEQ ID NO:4-6). ПЦР-результаты продемонстрировали, что ДНК Е. canis не была амплифицирована с использованием праймеров в кодирующей области гена белка массой 120 кДа Е. chaffeensis. 2,5 т.п.н. фрагмент ДНК амплифицировали из геномной ДНК Е. canis с помощью пары праймеров pxcf2-2 и pxar3, происходящих от некодирующих последовательностей ДНК, фланкирующих ген белка массой 120 кДа Е. chaffeensis (Фиг.1).
Пример 11
Определение числа повторяющихся единиц в гене белка массой 120 кДа E. canis
Для иллюстрации количества повторяющихся единиц, находящихся в гене белка массой 120 кДа Е. canis, использовали Саузерн-блоттинг. Рестрикционный анализ последовательности ДНК гена белка массой 120 кДа штамма Оклахома продемонстрировал, что все повторы имеют уникальный сайт расщепления эндонуклеазой SpeI. Вставка в плазмиде pCR120 была частично гидролизована SpeI. В результате неполного гидролиза ДНК гена белка массой 120 кДа Е. canis ферментом SpeI образовывалось три вида фрагментов ДНК: повторы с 5’-концевой неповторяющейся последовательностью, внутренние повторы и повторы с 3’-концевыми неповторяющимися последовательностями. После частичного SpeI-гидролиза все фрагменты ДНК с 5’-концевой неповторяющейся последовательностью гена начинались с одного и того же положения (5’-конца гена), но заканчивались в разных повторах. Эти фрагменты ДНК выделяли на агарозном геле в соответствии с их длиной, которая соответствовала числу их повторов. ДНК переносили на найлоновую мембрану и использовали для гибридизации с DIG-меченным олигонуклеотидом. Этот олигонуклеотид (SEQ ID NO:15) происходил от последовательностей ДНК, начиная с нуклеотида 38 и до нуклеотида 59, которые были расположены выше области повторов. Поэтому этот олигонуклеотид гибридизовался только с фрагментами ДНК, которые имели 5’-конец этого гена, но не с внутренними повторами и не с повторами с 3’-концом гена. Таким образом, число полос, детектированных с помощью олигонуклеотидного зонда, соответствует числу повторов. Саузерн-блоттинг выявил лэддер из 14 полос с приращением в 108 п.н. в Е. canis (Фиг.2). Эту результаты показали, что в Е. canis присутствует 14 повторов с 108 п.н. каждый.
Пример 12
Анализ последовательности ДНК гена белка массой 120 кДа Е. canis
ПЦР-амплифицированный фрагмент ДНК Е. canis клонировали в вектор pCR2.1. Полученную рекомбинантную плазмиду обозначали рСА120. Эту ДНК плазмиды рСА120 использовали в качестве матрицы для секвенирования вставки ДНК Е. canis. Сначала последовательность ДНК получали с использованием обратных праймеров Т7 и М13, которые были комплементарны векторной последовательности ДНК, фланкирующей эту вставку. Последующее секвенирование ДНК проводили путем "праймерной прогулки" по вставке и однонаправленной делеции вставки в плазмиде рСА120 путем рестрикции эндонуклеазой или экзонуклеазой III. Анализ последовательности ДНК продемонстрировал, что эта вставка ДНК содержала открытую рамку считывания (ОРС), состоящую из 2084 нуклеотидов (SEQ ID NO:7), которая кодировала 688 аминокислот (SEQ ID NO:8). Эта открытая рамка считывания была идентифицирована как ген белка массой 120 кДа Е. canis. При этом каких-либо консенсусных последовательностей ДНК промотора E. coli, расположенного возле 5’-конца этого гена, обнаружено не было. N-конец выведенных аминокислот не совпадал с консенсусной последовательностью сигнальных пептидов E. coli. В гене белка массой 120 кДа Е. canis присутствовали 14 тандемных повторов. Каждый повтор состоял из 108 нуклеотидов и кодировал 36 аминокислот (Фиг.3). Гомология аминокислотной последовательности всех повторов превышала 94% (Фиг.4). Перед первым повтором присутствовал неполный повтор, который имел делецию в 7 аминокислот (Фиг.3) и имел 70%-ную гомологию с другими повторами.
С помощью программы поиска FastA в базе данных Genbank было установлено, что ген белка массой 120 кДа Е. canis не имеет значительной гомологии с какой-либо известной последовательностью в этой базе данных. Аминокислотная гомология белков массой 120 кДа Е. canis (SEQ ID NO: 10) и Е. chaffeensis (SEQ ID NO:9) составляет 30% (Фиг.5). Белки массой 120 кДа являются более консервативными на своем N-конце и в области повторов. Аминокислотная гомология для первых 32 аминокислот на N-конце белков массой 120 кДа Е. canis и Е. chaffeensis составляет 50%. Гомология последовательности ДНК для генов белков массой 120 кДа этих двух видов составляет 58%. Была секвенирована некодирующая последовательность ДНК вышерасположенного 340 п.н.-гена белка массой 120 кДа Е. canis, и было обнаружено, что эти некодирующие области, смежные с генами белков массой 120 кДа этих двух видов Ehrlichia, имеют 84%-ную гомологию.
Пример 13
Предсказанная локализация и антигенность белка массой 120 кДа Е. canis
Выведенная аминокислотная последовательность гена белка массой 120 кДа Е. canis была проанализирована на гидрофобность, вероятность поверхностного экспонирования и антигенность с использованием программы "Protein" программного обеспечения Lasergene (DNASTRA Inc., Madison, WI). Предполагается, что все повторяющиеся единицы являются гидрофильными и экспонируются на поверхности (Фиг.6). Сравнение белков Е. canis и Е. chaffeensis продемонстрировало, что все повторяющиеся единицы в обоих белках являются поверхностно-экспонированными. Эти поверхностно-экспонированные области повторов в двух видах эрлихий (SEQ ID NO: 11-12, Фиг,7) имеют общие аминокислоты. Эти результаты дают основание предположить, что белок массой 120 кДа Е. canis является внешним мембранным белком. Гидрофильные аминокислоты в этих повторах могут представлять поверхностно-экспонированные части этого белка.
Анализ белков, проведенный методом Джеймсона-Вольфа, который позволил предсказать возможные антигенные детерминанты, показал, что белки массой 120 кДа как Е. canis, chaffeensis являются, по всей вероятности, в высокой степени антигенными (Фиг.6). Анализ белков, проведенный методом Рот-барда-Тейлора, который выявил возможную локализацию антигенных детерминант Т-лимфоцитов, продемонстрировал, что белок массой 120 кДа Е. canis имеет несколько предполагаемых Т-клеточных эпитопов и что все они локализованы на последовательностях, расположенных за пределами домена повторов. В противоположность этому каждый повтор белка массой 120 кДа Е. chaffeensis имел два Т-клеточных эпитопа.
Пример 14
Гомологичные гены в других штаммах Е. canis
Для амплификации гена белка массой 120 кДа от других штаммов Е. canis использовали ПЦР. 2,5 т.п.н. фрагмент ДНК от всех штаммов Е. canis, включая штаммы Флорида, Луизиана и три собачьих изолята Новая Каролина: Demon, DJ и Jake амплифицировали с использованием праймеров PXCf2-2 и РХАr3. Сегменты генов белка массой 120 кДа от всех штаммов Е. canis секвенировали у 5’- и 3’-концов. Анализ последовательности ДНК продемонстрировал, что последовательности ДНК, расположенные выше и ниже от области повторов, были идентичны для всех штаммов Е. canis. Полная область повторов для всех штаммов Е. canis не была секвенирована, поскольку для секвенирования необходимо, чтобы этот ген был делегирован. Был секвенирован лишь последний повтор для всех штаммов и первый повтор для штамма DJ. Последовательности первого повтора штаммов DJ и Оклахома были идентичными. Последовательности последнего повтора были идентичны для всех штаммов. Гомология гена белка массой 120 кДа от всех штаммов Е. canis, кроме того, была продемонстрирована идентичными SреI-рестрикционными физическими картами (Фиг.8).
Пример 15
Электрофорез в ПААГ с ДСН и Вестерн-блот-анализ
Ген белка массой 120 кДа Е. canis был сверхэкспрессирован в E. coli (Фиг.9). Рекомбинантный белок массой 120 кДа, кодируемый 1620 п.н. фрагментом ДНК, включающим полноразмерную область повторов гена белка массой 120 кДа, продуцировали в виде GST-гибридного белка. Молекулярная масса гибридного белка массой 120 кДа, оцененная на ДСН-геле, составляла приблизительно 140 кДа и значительно превышала предполагаемую молекулярную массу полноразмерного белка массой 120 кДа Е. canis, которая составляла лишь 73,6 кДа и была предсказана исходя из аминокислот, выведенных из последовательности ДНК (Фиг.9). Мышиные антитела против рекомбинантного белка массой 120 кДа реагировали с белком массой 120 кДа Е. canis (Фиг.10). Собачья антисыворотка против Е. canis также реагировала с рекомбинантным белком массой 120 кДа Е. canis (Фиг.11).
Пример 16
Обсуждение
Хотя Ehrlichia spp. являются облигатными внутриклеточными бактериями, однако ни ген адгезина, ни ген инвазина эрлихий не были идентифицированы. Ранее был клонирован и секвенирован ген белка массой 120 кДа Е. chaffeensis (Yu, 1997). Недавно было продемонстрировано, что белок массой 120 кДа Е. chaffeensis является внешним мембранным белком, который преимущественно экспрессируется на плотнокоровой ультраструктурной форме Е. chaffeensis, но не на ретикулярных клетках. Неинвазивный неадгезивный штамм E. coli, экспрессирующий белок массой 120 кДа, приобретал способность к адгезии и проникновению в культивированные клетки млекопитающих. Эти результаты дают основание предположить, что белок массой 120 кДа должен представлять собой адгезин или инвазии, а поэтому он может рассматриваться как кандидат на вакцину.
Ehrlichia canis и Е chaffeensis по своим генетическим и антигенным свойствам являются близкородственными бактериями. Гомология между Е. canis и Е. chaffeensis для гена 163 рРНК составляет 98%, а для гена nadA (Yu, 1997) - 89%. Поскольку очевидно, что белок массой 120 кДа играет важную роль в присоединении и проникновении Е. chaffeensis и эти два вида Ehrlichia являются близкородственными, то было высказано предположение, что в Е. canis может присутствовать аналог гена белка массой 120 кДа Е. chaffeensis, который может иметь те же самые биологические функции. Это предположение было подтверждено ПЦР-амплификацией гена белка массой 120 кДа Е. canis с использованием праймеров, происходящих от гена Е. chaffeensis. Однако ДНК гена белка массой 120 кДа Е. canis не имеет значительной гомологии с гомологичным геном Е. chaffeensis. Гомология генов белка массой 120 кДа Е. canis и Е. chaffeensis составляет лишь 30%. Низкая гомология генов этих двух видов Ehrlichia может быть объяснять отсутствие Саузерн-блот-гибридизации ДНК Е. canis с зондом для гена белка массой 120 кДа Е. chaffeensis и отсутствие ПЦР-амплификации гена Е. canis с использованием праймеров, происходящих от кодирующей области гена белка массой 120 кДа Е. chaffeensis. Было неожиданно обнаружено, что некодирующие последовательности, фланкирующие гены белка массой 120 кДа, являются более консервативными, чем кодирующие последовательности генов белка массой 120 кДа Е. canis и Е. chaffeensis. С эволюционной точки зрения, ожидается, что эта кодирующая последовательность, которая находится под давлением отбора, является более консервативной, чем некодирующая последовательность, в которой мутация не должна, как ожидается, влиять на выживаемость организма.
Хотя гены белка массой 120 кДа Е. canis и Е. chaffeensis не имеют значительной гомологии, однако очевидно, что ген Е. canis является гомологичным гену белка массой 120 кДа Е. chaffeensis, что дает основание считать, что они локализованы в аналогичных положениях в соответствующем геноме и при этом они имеют 30%-ную гомологию; а в частности, они имеют общие мотивы в области повтора. Однако оба гена белка массой 120 кДа Е. canis и Е. chaffeensis имеют различные аминокислотные последовательности и число повторов, белок массой 120 кДа Е. chaffeensis содержит 4 повтора, каждый из которых состоит из 80 аминокислот, а белок массой 120 кДа Е. canis содержит 14 повторов, каждый из которых состоит из 36 аминокислот. Однако эти повторы в обоих белках являются гидрофильными и предполагается, что они являются поверхностно-экспонированными. Несмотря на различия в числе повторов, даже общее число поверхностно-экспонированных областей в этих повторах указанных двух белков является очень близким. Повторяющиеся единицы обоих белков имеют общий мотив, состоящий из идентичных аминокислот, которые являются гидрофильными и образуют сердцевину (кор) поверхностно-экспонированных областей этих белков. Повторяющиеся единицы обоих белков богаты серином и глутаминовой кислотой. И серин и глутаминовая кислота, каждая составляет 19% от всех аминокислот повторяющейся единицы E. canis. Глутаминовая кислота и серин составляют 22 и 15% от всех аминокислот повторяющейся единицы Е. chaffeensis соответственно. Хотя с использованием программы для предполагаемой локализации белков PSORT сигнальная последовательность не была обнаружена на N-конце выведенных аминокислот белка массой 120 кДа Е. canis, однако очевидно, что белок массой 120 кДа Е. canis является поверхностным белком аналогично белку массой 120 кДа Е. chaffeensis, в котором сигнальная последовательность также отсутствует (Yu, 1997).
Аналогично гену белка массой 120 кДа Е. chaffeensis предсказанная молекулярная масса белка массой 120 кДа Е. canis значительно превышает размер молекулы, определенный по электрофоретической подвижности белка в ПААГ-ДСН. Аналогичный феномен был описан и для других белков, содержащих домены повторов, включая белки Anaplasma marginale (Allred D.R. et al., 1990), Plasmodium (Kemp D.J., 1987), Staphylococcus aureus (Hollingshead 1986, Signas 1989), и 100-кДа и 130-кДа-белки гранулоцитарной эрлихии (HGE) человека (Storey J.R., 1998). Повторяющиеся единицы 100-кДа и 130-кДа-белков HGE имеют общие последовательности с белком массой 120 кДа Е. chaffeensis (Storey, 1998). Аберантная миграция белков массой 120 кДа Е. canis и Е. chaffeensis не является следствием высокого процентного содержания некоторых аминокислот, поскольку молекулярная масса этого белка массой 120 кДа выше, чем общая молекулярная масса предсказанных аминокислот. Аберантная миграция белка массой 120 кДа, вероятно, связана с посттрансляционной модификацией этого белка массой 120 кДа, такой как гликозилирование. Посттрансляционная модификация белков массой 120 кДа Е. canis и Е. chaffeensis в настоящее время находится на стадии исследования. Поскольку белок массой 120 кДа Е. chaffeensis дифференциально экспрессируется в различных ультраструктурных формах Е. chaffeensis, то этот белок массой 120 кДа может играть определенную роль в патогенезе инфекции Е. chaffeensis. В настоящее время проводятся исследования для того, чтобы выяснить, экспрессируется ли белок массой 120 кДа Е. canis преимущественно в плотнокоровой клетке Е. canis. Хотя ген белка массой 120 кДа большинства штаммов Е. canis полностью не секвенирован, однако исходя из того факта, что известная последовательность, не включающая области повторов, а также первый и последний повторы идентичны для штаммов Е. Canis, и исходя из того факта, что все штаммы Е. canis имеют одно и то же число повторов, можно предположить, что последовательность гена белка массой 120 кДа идентична для всех штаммов Е. canis. Высокая степень гомологии последовательности ДНК и идентичное число повторов гена белка массой 120 кДа всех штаммов Е. canis свидетельствует о том, что штаммы Е. canis имеют меньшее генетическое разнообразие, чем штаммы Е. chaffeensis, у которых число повторов в гене белка массой 120 кДа сильно отличается.
Все патенты и публикации, упомянутые в настоящем описании, являются указанием на уровень техники, к которой относится настоящее изобретение. Эти патенты и публикации вводятся посредством ссылки так, как если бы каждая отдельная публикация была конкретно и отдельно введена посредством ссылки.
Для каждого специалиста очевидно, что настоящее изобретение может быть соответствующим образом адаптировано для достижения вышеупомянутых целей и получения преимуществ, присущих настоящему изобретению. Вышеприведенные примеры, а также методы, процедуры, обработки, молекулы и конкретные соединения, описанные в настоящей заявке, представляют предпочтительные варианты осуществления изобретения и носят иллюстративный характер, а поэтому они не должны рассматриваться как ограничение объема изобретения. В настоящее изобретение могут быть введены модификации и другие варианты использования по настоящему изобретению, не выходящие, однако, за рамки существа и объема изобретения, сформулированного в формуле изобретения.
Источники информации
1. Allred, D.R., et al., 1990. Proc. Natl. Аса. Sci. USA. 87:3220-4.
2. Anderson, B.E., et al., 1991. J. Clin. Microbiol. 29:2838-2842.
3. Anderson, B.E., et al., 1992. Int. J. Syst. Bacteriol. 42:299-302.
4. Bakken, J. S., et al., 1994. JAMA. 272:212-8.
5. Buhles, W.C.Jr., et al., 1974. J. Infect. Dis 130:357-367.
6. Chen, S.M., et al., 1994. J. Clin. Microbiol. 32:589-95.
7. Chen, S.M, et al., 1994. Am. J. Trop. Med. Hyg. 50:52-58.
8. Dawson J.E., et al., 1991. J. Infect. Dis. 163:564-567.
9. Fishbein, D.B., et al., 1987. JAMA. 257:3100-4.
10. Greene C.E., et al., 1984. Canine ehrlichiosis. p. 545-561. In C.E. Greene (ed). Clinical microbiology and infectious diseases of the dog and cat. The W.B. Saunders Co., Philadelphia.
11. Groves, M.G., et al., 1975. Am. J. Veterinary Research. 36:937-40.
12. Hollingshead, S.К., et aL, 1986. J. Biol. Chem. 261:1677-86.
13. Kemp, D.J., et al., 1987. Annu. Rev. Microbiol. 41:181-208.
14. Jameson, В.A., et al., 1988. CABIO, 4:181-186.
15. KIein, P., et al., 1985. Biochem. Biophys. Acta, 815: 468-76.
16. Lewis, G.E.Jr., et al., 1977. American Journal of Veterinary Research. 38:1953-5.
17. Maeda, К.N. et al., 1987. N. Engl. J. Med. 316:853-856.
18. McGeoch D. J., 1985. Virus Research 3, 271-86.
19. Perez, M., et al., 1996, J. Clin. Microbiol. 34:2133-9.
20. Rothbard, J.В., et al., 1988. The EMBO J 7:93-100.
21. Signas, С., et al., 1989. Proc. Natl. Acad. Sci. USA 86:699-703.
22. Storey, J.R., et al., 1998. Infect, Immun. 66:1356-63.
23. von Heijne G. 1986. Nucl. Acids Res. 14: 4683-90.
24. Walker, J.S., et al., 1970. J. Am. Vet. Med. Assoc. 157:43-55.
25. Yu, X-J., et al., 1996. Gene. 184:149-54.
26. Yu X-J., et al., 1997. FEMS Microbiol Letters. 154:53-8.





Изобретение относится к биотехнологии и представляет собой ген белка массой 120 кДа Ehrlichia canis, амплифицированный посредством ПЦР с использованием праймеров, происходящих от последовательностей ДНК, фланкирующих ген белка массой 120 кДа Ehrlichia chaffeensis. Рекомбинантный белок массой 120 кДа Е. canis содержит 14 тандемных повторяющихся единиц, каждая из которых состоит из 36 аминокислот. Эти повторяющиеся единицы являются гидрофильными и, как предполагается, поверхностно-экспонированными. Рекомбинантный белок массой 120 кДа Е. Canis также является антигенным и реагирует с сывороткой, продуцированной у собак, выздоравливающих от собачьего эрлихиоза. Данное изобретение позволяет ингибировать инфекцию, вызываемую Ehrlichia canis, путем введения композиции, содержащей антиген Е. canis, 5 с. и 7 з.п.ф-лы, 11 ил., 1 табл.
| REDDY G.R | |||
| et al | |||
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| Biochem Biophys Res Commun | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| Приспособление для картограмм | 1921 |
|
SU247A1 |
| WO 9412641, 09.06.1994 | |||
| Рекомбинатная плазмидная ДНК pGp 120 - 428, кодирующая гибридный белок с антигенными свойствами белка @ р 120 ВИЧ-1 | 1991 |
|
SU1789562A1 |

