Изобретение относится к новым штаммам молочнокислых бактерий и к их использованию для повышения иммунитета.
Предпосылки создания изобретения
Потребление продуктов, содержащих молочнокислые бактерии (МКБ), ассоциируется с рядом благоприятных для здоровья факторов, включая повышение иммунитета. Существуют тысячи штаммов молочнокислых бактерий, но только некоторые из этих штаммов обладают свойствами, способствующими укреплению здоровья. Устойчивость этих бактерий к воздействию кислот и желчных солей и их способность прилипать к эпителиальным клеткам слизистой и выживать при прохождении через желудочно-кишечный тракт являются важным критерием для отбора штаммов, способствующих укреплению здоровья. До настоящего времени были идентифицированы только немногие штаммы молочнокислых бактерий, которые, как было установлено, оказывают благоприятное действие на организм.
Из патента Новой Зеландии 248057 известно, что штаммы МКБ обладают хорошей адгезивностью к эпителиальным клеткам слизистой тонкого кишечника, что тем самым придает самим этим бактериям терапевтические свойства. Микроорганизмы, описанные в этом патенте, способствуют повышению как природного иммунитета (фагоцитарной функции), так и приобретенного иммунитета (гуморальный ответ и лимфоцит-пролиферативный ответ).
Было бы желательно продуцировать другие молочнокислые бактерии (МКБ), которые способствовали бы усилению иммунных реакций широкого спектра, включая фагоцитарную функцию.
Целью настоящего изобретения является достижение этой цели тем или иным путем, или, по крайней мере, предоставление людям возможности широкого выбора молочнокислых бактерий, усиливающих иммунитет.
Описание изобретения
В соответствии с этим можно сказать, что в одном из своих аспектов настоящее изобретение в общих чертах относится к биологически чистой культуре Lactobacillus rhamnosus HN001, депонированной 18 августа 1997 г. в AGAL под номером NM97/09514.
Можно сказать, что в другом своем аспекте настоящее изобретение в общих чертах относится к биологически чистой культуре Lactobacillus rhamnosus HN067, депонированной 17 февраля 1998 г. в AGAL под номером NM97/01925.
Можно сказать, что в другом своем аспекте настоящее изобретение в общих чертах относится к композиции, содержащей биологически чистую культуру любого из штаммов Lactobacillus acidophilus HNO17, депонированного 18 августа 1997 г. в AGAL под номером NМ97/09515; Lactobacillus rhamnosus HN001, Lactobacillus rhamnosus HN067 или Bifidobacterium lactis HN019, депонированного 18 августа 1997 г. в AGAL под номером NM97/09513, в иммуностимулирующей концентрации в сочетании с физиологически приемлемым носителем или разбавителем.
В одном из вариантов настоящего изобретения указанная композиция содержит любые два или более из указанных штаммов.
Предпочтительно указанным физиологически приемлемым наполнителем или разбавителем является пищевой продукт.
Предпочтительно указанным пищевым продуктом является один из таких продуктов, как сквашенное молоко, йогурт, сыр, молочный напиток или сухое молоко.
Альтернативно указанной композицией является фармацевтическая композиция, а указанным наполнителем или разбавителем является физиологически приемлемый наполнитель или разбавитель.
Усиливающими иммунитет штаммами являются физиологически приемлемые биологически чистые штаммы гомологов или мутантов любого из следующих штаммов:
Lactobacillus acidophilus HN017,
Lactobacillus rhamnosus HN001,
Bifidobacterium lactis HN019, или
Lactobacillus rhamnosus HN067.
Можно сказать, что в другом своем варианте настоящее изобретение в общих чертах относится к способу повышения природного и приобретенного иммунитета, который предусматривает введение млекопитающему любой из вышеуказанных биологически чистых культур в иммуностимулирующей дозе.
В другом варианте осуществления настоящего изобретения в основном присутствуют биологически чистые культуры двух или трех из вышеупомянутых штаммов.
Предпочтительно указанную культуру вводят в форме композиции с физиологически приемлемым наполнителем или разбавителем.
Предпочтительно указанным физиологически приемлемым наполнителем или разбавителем является пищевой продукт.
Предпочтительно указанным пищевым продуктом является сквашенное молоко, йогурт, сыр, молочный напиток или сухое молоко.
Также можно сказать, что настоящее изобретение в общих чертах составляют части, элементы и отличительные признаки, взятые отдельно или в совокупности, имеющие отношение к описанию заявки или указанные в этом описании, а также любые или все комбинации из любых двух или более указанных частей, элементов или отличительных признаков; и в тех случаях, когда некие конкретные целые элементы, упомянутые в данном описании, имеют известные эквиваленты в той области, к которой это изобретение относится, то предполагается, что такие известные эквиваленты вводятся в настоящее описание, как если бы они были введены по отдельности.
Краткое описание чертежей
На Фиг.1 показано влияние введения мышам продукта, ферментированного L. rhamnosus HN001 или неферментированного продукта, содержащего L.rhamnosus HN001, на фагоцитарную активность лейкоцитов периферической крови, как описано в Примере 5. Мышам BALB/c давали молочный корм, содержащий 109 к.о.е. (в день) L. rhamnosus HN001, в виде либо ферментированного, либо неферментированного продукта в течение 14 дней. Фагоцитарную активность лейкоцитов периферической крови определяли с использованием проточной цитометрии и Escherichia coli, меченных флуоресцеин-изотиоцианатом. Величины представляли собой среднее значение ± стандартная ошибка. Значимые отличия (дисперсионный анализ ANOVA, программа SAS) от контроля: **Р<0,0001.
На Фиг.2 показано влияние введения мышам живого штамма L.rhamnosus HN001 или термоинактивированного штамма L.rhamnosus HN001 на фагоцитарную активность лейкоцитов периферической крови, как описано в Примере 7. Мышам ВALВ/с давали молочный корм и вводили перорально 109 к.о.е. (в день) либо живой, либо термоинактивированной бактерии L.rhamnosus HN001 в течение 14 дней. Фагоцитарную активность лейкоцитов периферической крови и перитониальных макрофагов определяли с использованием проточной цитометрии и Escherichia coli, меченной флуоресцеин-изотиоцианатом. Величины представляли собой среднее значение ± стандартная ошибка. Значимое отличие (ANOVA, программа SAS) от контроля: **Р<0,0001.
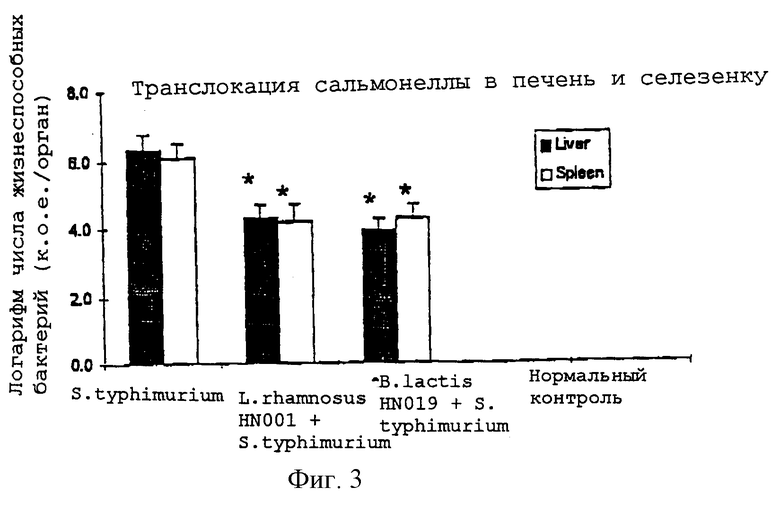
На Фиг.3 показано влияние введения мышам L.rhamnosus HN001 или B.lactis HN019 на транслокацию бактерий у мышей, экспериментально зараженных S.typhimurium, как описано в Примере 8. Мышей ВАLВ/с, которым не вводили молочнокислые бактерии, и мышей, которым вводили B.1actis HN019 или L.rhamnosus HN001, после ежедневного непрерывного введения этой бактерии перорально заражали S.typhimurium. Через шесть дней после заражения мышей подвергали гуманной эфтаназии и их печень и селезенку выделяли с последующим мониторингом транслокации бактерий. После этого суспензию тканей от выделенных органов культивировали на планшетах с агаром МасСоnkey в течение 24-48 часов, а затем подсчитывали. Полученные величины представляли собой среднее значение ± стандартная ошибка. Значимые отличия (ANOVA, программа SAS) от контроля: *Р<0,05.
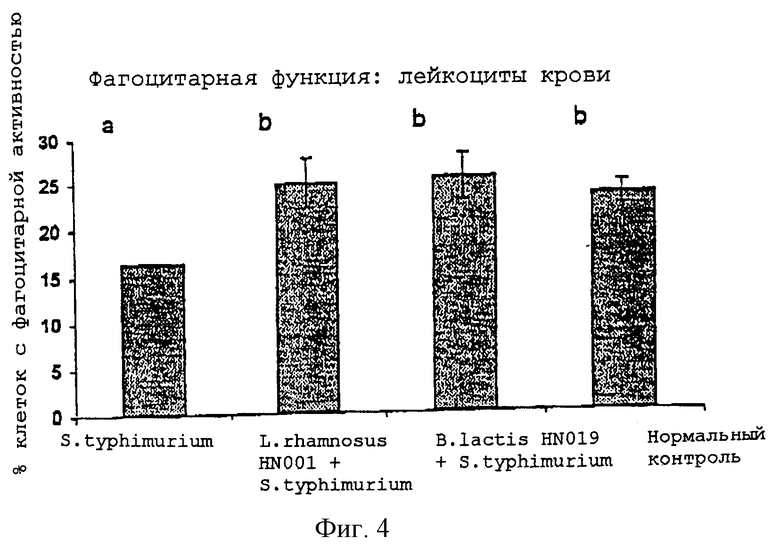
На Фиг.4 показано влияние введения мышам L.rhamnosus HN001 или B.lactis HN019 на фагоцитарную активность лейкоцитов периферической крови у мышей, экспериментально зараженных S. typhimurium, как описано в Примере 8. Мышей BALB/c, которым не вводили молочнокислые бактерий, и мышей, которым вводили B. 1actis HN039 или L.rhamnosus HN001, после ежедневного непрерывного введения этой бактерии перорально заражали S.typhimurium. Через шесть дней после заражения фагоцитарную активность лейкоцитов периферической крови определяли с использованием проточной цитометрии и Escherichia coli, меченной флуоресцеин-изотиоцианатом. Величины представляли собой среднее значение ± стандартная ошибка. Полученные величины (среднее значение ± стандартная ошибка) с различными верхними индексами являются значимо различными (ANOVA, программа SAS): Р<0,01.
На Фиг.5 показано влияние введения мышам живого штамма L.rhamnosus HN001 или B. lactis HN019 на пролиферативный ответ лейкоцитов селезенки у мышей, экспериментально зараженных S. typhimurium, как описано в Примере 8. Мышей BALB/c, которым не вводили молочнокислые бактерии, или мышей, которым вводили B.lactis HN019 или L.rhamnosus HN001, после ежедневного непрерывного введения этой бактерии перорально заражали S.typhimurium. Через шесть дней после заражения пролиферативный ответ лейкоцитов селезенки определяли колориметрически после введения 5-бром-2'-дезоксиуридина в последние 16 часов 96-часового периода инкубирования. Полученные величины (среднее значение ± стандартная ошибка) с различными верхними индексами являются значимо различными (ANOVA, программа SAS): Р<0,01.
Модели осуществления настоящего изобретения
Лиофилизованные культуры четырех бактериальных штаммов были депонированы в Австралийской государственной аналитической лаборатории (AGAL), Региональной лаборатории Нового Южного Уэльса, 1 Suakin Street, Pymble, NSW 2073, Australia. В Таблице А приводится подробное описание депозитов.
Было обнаружено, что четыре штамма, идентифицированные в Таблице А, способствуют усилению иммунных реакций широкого спектра, включая как природный, так и приобретенный иммунный ответ.
Пример 1
Морфология и общие свойства
Для подтверждения таксонометрических характеристик штаммов проводили RAPD-анализ, секвенирование рРНК 16S и ДСН-ПААГ-анализ. Было также обнаружено, что штамм L.acidophilus HN017 генетически отличается от бактерии L.acidophilus (LC1), описанной в патенте Новой Зеландии 248057.
Для подтверждения таксонометрических характеристик L.rhamnosus HN067 проводили RAPD-анализ, секвенирование рРНК 16S и ДСН-ПААГ-анализ; причем для характеризации L.rhamnosus HN067 на молекулярном уровне использовали следующие видоспецифические праймеры Рr. 1 (прямой) 5'-CAGACTGAAAGTCTGACGG-3' и Pha. II (обратный) 5'-GCGATGCGAAXXXCTATTATT-3'.
Морфология этих штаммов и их способность к ферментации сахаров подробно описана в Таблицах 1 и 2.
Пример 2
Прилипание к клеткам кишечника
Способность пробиотических штаммов прилипать к эпителиальным клеткам тонкого кишечника человека (НТ-29 и СаСо-2) оценивали in vitro с использованием дифференцированных клеточных линий. Монослои клеток НТ-29 и СаСо-2 культивировали на покровных стеклах и помещали в многолуночные планшеты. Затем к клеточным слоям добавляли 108 к.о.е./мл МКБ в 1 мл супернатанта отработанной культуры вместе с 1 мл модифицированной по способу Дульбекко среды Игла (DMEM) и инкубировали в течение 1 часа при 37oС в атмосфере 10% СO2 - 90% воздух. Монослои промывали 4 раза PВS, фиксировали метанолом, окрашивали по Граму и определяли под микроскопом число бактерий, прилипших к эпителиальным клеткам. В среднем было просчитано 20 полей, и полученные результаты были систематизированы в Таблице 3.
Пример 3
Повышение природного и приобретенного иммунитета
Иммуностимулирующие эффекты трех штаммов L. rhamnosus HN001, L.acidophilus HN017 и B. lactis HN019 определяли путем оценки фагоцитарной функции (лейкоцитов крови и перитонеальных макрофагов) и путем количественной оценки концентраций специфических антител против белков-антигенов, использованных для имитации у мышей ответа на введение вакцины. Была использована следующая экспериментальная схема:
1. Использовались 6-7-недельные мыши BALB/c весом 20-30 г.
2. Мыши были произвольно распределены по различным группам обработки (Таблица 4).
3. Мышам в течение 10 дней вводили L.acidophilus HN017, L.rhamnosus HN001 или B.lactis HN019 (109 к.о.е./день) в 50 мкл сепарированного молока. Контрольные мыши получали только 50 мкл сухого сепарированного молока.
4. Все мыши получали корм на основе сухого сепарированного молока в течение всего эксперимента.
Лейкоциты крови и макрофаги у мышей, получавших L.acidophilus HN017, L. rhamnosus HN001 или B.lactis HN019, обнаруживали значительно более высокую фагоцитарную активность по сравнению с клетками контрольных мышей (Таблица 4). Продуцирование кислородных радикалов (окислительный стресс) лейкоцитами у мышей с пробиотическим питанием также было более высоким, чем у контрольных мышей (данные не приводятся).
Мышам BALB/c перорально вводили 109 к. о.е. (в день) L. acidophilus HN017, L.rhamnosus HN001 или B.lactis HN019 в течение 10 дней. Фагоцитарную активность лейкоцитов крови и перитонеальных микрофагов определяли с использованием проточной цитометрии и Escherichia coli, помеченных флуоресцеин-изотиоцианатом. Полученные величины представляли собой среднее значение ± стандартная ошибка. Значимое отличие (t-критерий Стьюдента) от контроля: *Р<0,05, **Р<0,01.
Концентрация специфических антител IgG в сыворотке и в кишечных смывах у мышей, получавших L. acidophilus HN017, L. rhamnosus HN001 или B.lactis HN019, также была значительно выше, чем у контрольных мышей (Таблица 5).
Мышам BALB/c перорально вводили 109 к. о.е. (в день) L. acidophilus HN017, L. rhamnosus HN001 или B.lactis HN019 в течение 10 дней. Мышей иммунизировали холерным токсином (этот антиген использовали для имитации кишечной инфекции) в дни 0 и 7. Концентрацию специфических антител в сыворотке и в кишечном секрете измеряли с помощью анализа ELISA на 10-й день. Полученные величины представляли собой среднее значение ± стандартная ошибка. Значимые отличия (t-критерий Стьюдента) от контроля: *Р<0,08, **Р<0,05, ***Р<0,01.
Пример 4
Иммуностимулирующее действие молочнокислых бактерий (МКБ) после их введения в течение четырех недель
Иммуностимулирующее действие L.rhamnosus HN001, L.acidophilus HN017 и B. lactis HN019 у мышей оценивали с использованием нижеследующей экспериментальной схемы:
1. Использовались 6-7-недельные мыши BALB/c весом 20-30 г.
2. Мыши были произвольно распределены (18 на группу) по различным группам обработки.
3. После акклиматизации (7 дней) мышам в течение 28 дней (от дня 0 до дня 28) вводили 109 к.о.е. (в день) L.acidophilus HN017, L.rhamnosus HN001 или B.lactis HN019 в 50 мкл сепарированного молока. Контрольные мыши получали только 50 мкл сухого сепарированного молока (без микроорганизмов).
4. Все мыши имели свободный доступ (ad libitum) к корму, приготовленному на основе сухого сепарированного молока, и к воде во время всего эксперимента.
5. Иммуностимулирующие эффекты оценивали путем мониторинга фагоцитарной активности лейкоцитов крови и перитонеальных макрофагов, киллерной активности лимфоцитов селезенки, пролиферации лимфоцитов (клеток селезенки) в ответ на Т-клеточный митоген, СоnА (индикатор клеточно-опосредованного иммунитета) и гуморального ответа на столбнячную вакцину.
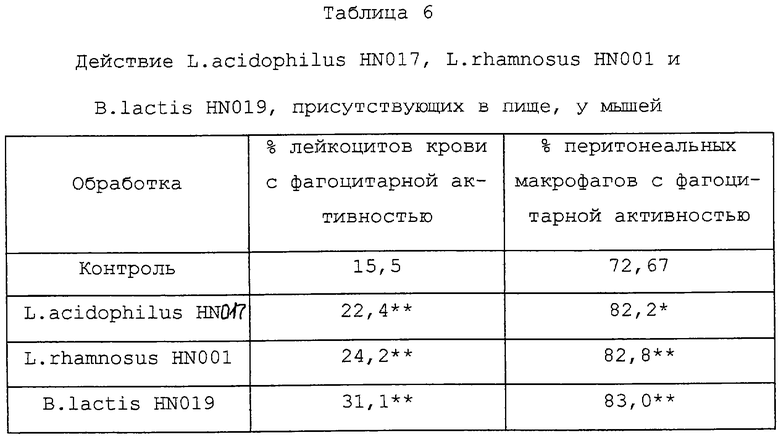
Как видно из Таблицы 6, лейкоциты (нейтрофилы, моноциты и макрофаги) у мышей, получавших L.acidophilus HN017, L.rhamnosus HN001 или B.lactis НN019, обнаруживали значительно более высокую фагоцитарную активность (как показатель природного иммунитета), чем лейкоциты у контрольных мышей.
Мыши (18 на группу) получали 109 к.о.е. (в день) L.acidophilus HN017, L. rhamnosus HN001 или B.lactis HN019 в 50 мкл сепарированного молока в течение 28 дней. Фагоцитарную активность лейкоцитов крови/перитонеальных микрофагов определяли на 28-й день с использованием проточной цитометрии и Е.coli, помеченных флуоресцеин-изотиоцианатом. Оценки представляли собой средние значения, полученные методом наименьших квадратов. Значимые отличия (анализ SAS): *Р<0,002, **Р<0,0005.
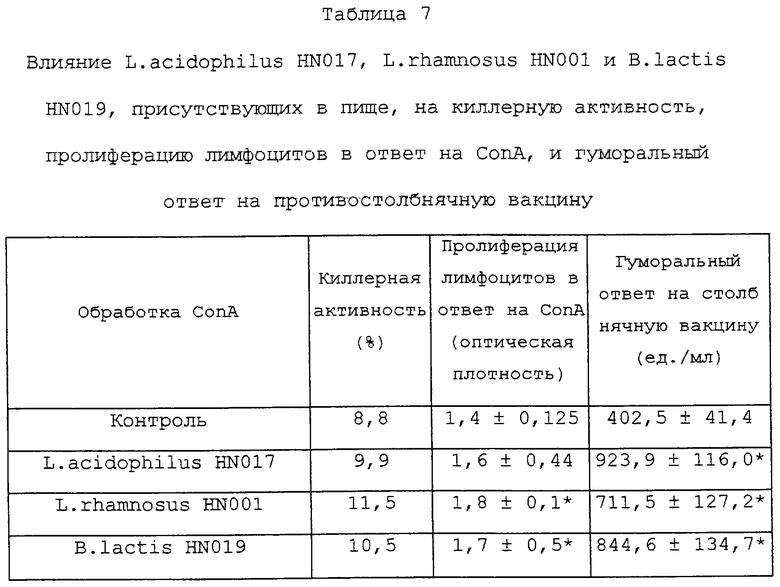
Потребление L.acidophi1us HN017, L.rhamnosus HN001 или B.lactis HN019 в течение 28 дней также приводило к повышению киллерной активности, пролиферации лимфоцитов в ответ на СоnА и гуморального ответа на столбнячную вакцину. По всем этим показателям иммунокомпетенции мыши, получавшие L.acidophilus HN017, L.rhamnosus HN001 или B.lactis HN019, давали более высокий ответ, чем контрольные мыши (Таблица 7).
Все эти результаты, взятые вместе, показали, что добавление в корм L. acidophilus HN017, L.rhamnosus HN001 или В. lactis HN019 в течение продолжительного периода времени способствует индуцированию пролонгированного усиления природного и приобретенного иммунитета по нескольким аспектам.
Мыши (18 на группу) получали 109 к.о.е. (в день) L.acidophilus HN017, L. rhamnosus HN001 или B.lactis HN019 в 50 мкл сепарированного молока в течение 28 дней (т. е. от дня 0 до 28). Киллерную активность лимфоцитов селезенки определяли на 28 день с использованием проточной цитометрии и D275-меченых клеток Yac-1. Пролиферацию лимфоцитов селезенки в ответ на СоnА оценивали на 28-й день с использованием коммерческого набора для оценки пролиферации (Boehringer Mannhiem, Germany). Для оценки гуморального ответа мышей иммунизировали столбнячной вакциной (50 мкл/дозу, CSL, Australia) в дни 7 и 21. Концентрацию специфических антител определяли с использованием ELISA; для сенсибилизации планшетов использовали антиген, поставляемый производителями вакцин (CSL, Australia). Полученные величины представляли собой среднее значение, вычисленное методом наименьших квадратов, для 18 мышей. Значимые отличия (анализ SAS): *Р<0,05.
Пример 5
Усиление природного и приобретенного иммунитета с использованием ферментированных продуктов по сравнению с неферментированными продуктами
Целью этого эксперимента является оценка иммуностимулирующего действия йогурта, приготовленного (ферментированного) с использованием пробиотического штамма L.rhamnosus HN001, по сравнению с неферментированным продуктом, содержащим штамм L. rhamnosus HN001. Иммуностимулирующий эффект определяли путем оценки фагоцитарной функции (лейкоцитов периферической крови и перитонеальных макрофагов) и пролиферации лимфоцитов в ответ на В-клеточный митоген (ЛПС).
Была использована нижеследующая экспериментальная схема:
1. Использовались 6-7-недельные мыши BALB/c весом 20-30 г.
2. Мыши били произвольно распределены по различным группам обработки.
3. Контрольные мыши получали только корм на основе цельного сухого молока на протяжении всего эксперимента.
4. Испытуемые мыши получали 2,5 г йогурта, приготовленного с использованием L. rhamnosus HN001 (109 к.о.е./день), или 2,5 г цельного молока, содержащего L.rhamnosus HN001 (109 к.о.е./день) в день, а также корм на основе цельного сухого молока в течение 14 дней.
Результаты
Мыши, получавшие йогурт с L.rhamnosus HN001 или цельное молоко, содержащее L.rhamnosus HN001, обнаруживали значительно более высокий уровень фагоцитарной активности лейкоцитов периферической крови, чем мыши, получавшие контрольный корм (Фиг.1). Это увеличение уровней активности наблюдалось независимо от того, получали ли мыши L.rhamnosus HN001 в йогурте (ферментированном бактерией L. rhamnosus HN001) или в неферментированном продукте, содержащем L.rhamnosus HN001. Каких-либо различий в уровнях фагоцитарной активности у мышей, получавших ферментированный йогурт, приготовленный с использованием L.rhamnosus HN001, и у мышей, получавших продукт на основе цельного молока (ПЦМ), содержащий L.rhamnosus HN001, не наблюдалось.
Мыши, получавшие неферментированный продукт, как и мыши, получавшие продукт, ферментированный бактерией L.rhamnosus HN001, обнаруживали более высокую степень пролиферации лимфоцитов в ответ на ЛПС, чем контрольные мыши (Таблица 8). При этом каких-либо различий в уровнях фагоцитарной активности у мышей, получавших неферментированный продукт, содержащий L.rhamnosus HN001, и у мышей, получавших продукт, ферментированный L.rhamnosus HN001, не наблюдалось.
Мыши BALB/c получали молочный корм, содержащий 109 к.о.е. (в день) L. rhamnosus HN001 либо в виде неферментированного продукта, либо в виде йогурта, ферментированного штаммом L. rhamnosus HN001 (ферментированный продукт) в течение 14 дней. Контрольные мыши получали молочный корм, не содержащий каких-либо МКБ. Пролиферативный ответ определяли колориметрически после введения 5-бром-2'-дезоксиуридина в последние 16 часов 96-часового периода инкубирования. Полученные величины представляли собой среднее значение ± стандартная ошибка. Значимое отличие (t-критерий Стьюдента) от контроля: *Р=0,05.
Эти результаты в целом дают основание предположить, что добавление в пищу L.rhamnosus HN001 приводит к усилению ряда иммунных функций, включая фагоцитарную активность и пролиферацию лимфоцитов. Штамм L.rhamnosus HN001, присутствующий либо в ферментированном, либо в неферментированном продукте, способствует эффективному усилению иммунных функций; причем для некоторых функций большую реакцию дает ферментированный продукт, а для других функций усиление дает неферментированный продукт.
Пример 6
Усиление природного и приобретенного иммунитета с использованием L. rhamnosus HN067
Эксперимент 1
Иммуностимулирующее действие L.rhamnosus HN067 оценивали путем мониторинга фагоцитарной способности лейкоцитов периферической крови и перитонеальных макрофагов (показатель неспецифического иммунитета) и путем количественной оценки концентраций специфических антител, продуцированных в ответ на иммунизацию антигеном, холерным токсином (используемым для имитации иммунного ответа на кишечную вакцину) у мышей.
Была использована нижеследующая экспериментальная схема:
1. Использовались 6-7-недельные мыши BALB/c весом 20-30 г. В течение всего эксперимента мышам давали корм на основе сепарированного молока.
2. Мышам в тестируемой группе (n=6) перорально вводили L.rhamnosus HN067 (109 к.о.е./в день) в 50 мкл сепарированного молока в течение 10 дней. Контрольные мыши (n=6) получали только 50 мкл сепарированного сухого молока (без каких-либо МКБ).
Результаты
У мышей, получавших L.rhamnosus HN067, лейкоциты крови и перитонеальные макрофаги обнаруживали значительно более высокую фагоцитарную активность (повышенную фагоцитарную функцию) по сравнению с клетками контрольных мышей. Результаты представлены ниже в Таблице 9.
Мышам BALB/c (6 на группу) в течение 10 дней давали молочный корм, содержащий или не содержащий перорально вводимый штамм L.rhamnosus НN067 (109 к. о.е./день). Фагоцитарную активность лейкоцитов крови и перитонеальных микрофагов определяли с использованием проточной цитометрии и бактерий Е. coli, помеченных флуоресцеин-изотиоцианатом. Данные представляли собой среднее значение, полученное методом наименьших квадратов, ± стандартная ошибка (LSM). Значимые отличия (программа SAS) от контроля: *Р=0,0005, **Р=0,0001.
Концентрация специфических антител против холерного токсина, то есть против антигена, использованного для пероральной иммунизации, в сыворотке и кишечных смывах у мышей, получавших L.rhamnosus HN067, также была значительно выше, чем у контрольных мышей (Таблица 10).
Мышам BALB/c в течение 10 дней давали молочный корм, содержащий или не содержащий перорально вводимый штамм L.rhamnosus HN067 (109 к.о.е./день). Мышей перорально иммунизировали холерным токсином (10 мкг/дозу), т.е. антигеном, используемым для имитации кишечной инфекции, в дни 0 и 7. Концентрацию антител в сыворотке и в кишечном секрете измеряли с помощью анализа ELISA на 10-й день. Данные представляли собой среднее значение, полученное методом наименьших квадратов, ± стандартная ошибка (LSM). Значимые отличия (программа SAS) от контроля: *Р=0,02, **Р=0,0039.
Эксперимент 2
Иммуностимулирующее действие L.rhamnosus HN067 у мышей оценивали с использованием нижеследующей экспериментальной схемы:
1. Использовали 6-7-недельных мышей BALB/c весом 20-30 г. Эти мыши получали корм на основе сухого сепарированного молока и воду ad libitum в течение всего эксперимента.
2. После акклиматизации в течение 7 дней мышам первой группы (n=20) перорально вводили 109 к.о.е. (в день) L.rhamnosus HN067 в 50 мкл сепарированного молока (группа 1, n=20) в течение 14 дней. Контрольные мыши (группа 2, n=20) получали сепарированное молоко, не содержащее каких-либо микроорганизмов.
3. Иммуностимулирующие эффекты оценивали путем мониторинга фагоцитарной активности лейкоцитов крови и перитонеальных макрофагов и пролиферации лимфоцитов селезенки в ответ на фитогемагглютинин (ФГА) и липополисахариды (ЛПС) (Т- и В-клеточные митогены соответственно).
Результаты
У мышей, получавших L.rhamnosus HN067, лейкоциты крови и перитонеальные макрофаги обнаруживали значительно более высокую фагоцитарную активность (показатель природного иммунитета) по сравнению с лейкоцитами и макрофагами контрольных мышей (Таблица 11).
Мыши BALB/c в течение 14 дней получали молочный корм, содержащий или не содержащий перорально вводимый штамм L. rhamnosus HN067 (109 к.о.е./день). Фагоцитарную активность лейкоцитов крови/перитонеальных микрофагов определяли на 14-й день с использованием проточной цитометрии и бактерий Е. coli, помеченных флуоресцеин-изотиоцианатом. Данные представляли собой среднее значение, полученное методом наименьших квадратов, ± стандартная ошибка (LSM). Значимые отличия (программа SAS): *Р=0,002, **Р=0,0001.
Мыши, получавшие L.rhamnosus НN067 в течение 14 дней, также обнаруживали более высокий уровень пролиферации лимфоцитов в ответ на ФГА и ЛПС по сравнению с контрольными мышами (Таблица 12).
Мыши BALB/c в течение 14 дней получали молочный корм, содержащий или не содержащий перорально вводимый штамм L.rhamnosus HN067 (109 к.о.е./день). Пролиферацию лимфоцитов селезенки в ответ на ФГА и ЛПС оценивали на 14-й день с использованием коммерческого набора для оценки пролиферации (Boehringer Mannhiem, Germany). Данные представляли собой среднее значение, полученное методом наименьших квадратов, ± стандартная ошибка (LSM). Значимые отличия (программа SAS): *Р<0,08, **Р<0,01.
В целом мыши, получавшие L.rhamnosus HN067, обнаруживали значительно более высокое усиление ряда функций иммунного ответа хозяина, включая фагоцитарную функцию лейкоцитов, гуморальный ответ на пероральную иммунизацию и пролиферацию лимфоцитов в ответ на Т- и В-клеточные митогены. Лейкоциты крови (нейтрофилы и моноциты) и макрофаги являются основными эффекторами природного иммунитета и играют важную роль в защите организма от микробных инфекций. Была также зарегистрирована четкая корреляция между in vitrо-пролиферацией лимфоцитов в ответ на митогены (Т-клеточные и В-клеточные митогены) и иммунокомпетентностью организма. Поэтому полученные результаты дают основание предположить, что введение L.rhanmоsus HN067 может способствовать усилению некоторых функций природного и приобретенного иммунитета.
Пример 7
Усиление природного и приобретенного иммунитета с использованием живой и термоинактивированной бактерии L.rhamnosus HN001
Целью настоящего исследования является оценка иммуностимулирующего действия пробиотического штамма L. rhamnosus HN001 в зависимости от того, присутствует ли он в форме живого микроорганизма или в термоинактивированной форме. Действие на иммунную функцию оценивали путем определения фагоцитарной активности лейкоцитов периферической крови. Действие живого и термоинактивированного микроорганизма L. rhamnosus HN001 на гуморальный иммунитет оценивали путем иммунизации мышей холерным токсином с последующим определением концентраций продуцированных специфических антител.
Была использована нижеследующая экспериментальная схема:
1. Использовали 6-7-недельных мышей BALB/c весом 20-30 г.
2. Мыши были произвольно распределены по различным группам обработки.
3. Контрольные мыши получали корм на основе сухого сепарированного молока на протяжении всего эксперимента.
4. Испытуемые мыши получали либо 109 к.о.е./день живых бактерий L.rhamnosus HN001, либо 109 к.о.е./день термоинактивированных бактерий L.rhamnosus HN001 в день, а также корм на основе сухого сепарированного молока в течение 14 дней.
5. Мышей перорально иммунизировали холерным токсином в день 0 и в день 7 приема пищи.
Результаты
Мыши, получавшие L.rhamnоsus HN001, обнаруживали значительно более высокий уровень фагоцитарной активности лейкоцитов периферической крови по сравнению с мышами, получавшими контрольный корм (Фиг.2). Это повышение уровней активности наблюдалось независимо от того, получали ли мыши живую бактерию L. rhamnosus HN001 или ее термоинактивированную форму. Каких-либо различий в уровнях фагоцитарной активности у мышей, получавших живую L.rhamnosus HN001, и у мышей, получавших термоинактивированную L.rhamnosus HN001, не наблюдалось.
Мыши, получавшие живую L.rhamnosus HN001, как и мыши, получавшие термоинактивированную L. rhamnosus HN001, обнаруживали более высокий уровень продуцирования антител в сыворотке и в слизистой, чем контрольные мыши. Однако уровень ответа у мышей, получавших живую L.rhamnosus HN001, был значимо выше (Таблица 13).
Мышам BALB/c давали молочный корм и перорально вводили L.rhamnosus HN001 (109 к. о.е./день) либо в форме живой бактерии, либо в форме термоинактивированной бактерии в течение 14 дней. Контрольные мыши не получали МКБ. Мышей перорально иммунизировали холерным токсином в дни 0 и 7. Продуцирование антител (в сыворотке и в кишечном секрете) определяли с помощью анализа ELISA на 14-й день. Полученные величины представляли собой среднее значение ± стандартная ошибка. Значимые отличия (t-критерий Стьюдента) от контроля: *Р= 0,05, **Р=0,0005.
Полученные результаты дают основание предположить, что как живые, так и термоинактивированные бактерии L.rhamnosus HN001 способны усиливать определенные функции природного и приобретенного иммунитета у мышей.
Пример 8
Противоинфекционные свойства B.1actis HN019 и L.rhamnosus HN001
Целями настоящего исследования являлись:
1. Оценка эффективной защиты, обеспечиваемой бактериями B.1actis HN019 и L. rhаmnosus HN001, против желудочно-кишечного патогена Salmonella typhimurium.
2. Определение роли иммуностимуляции, индуцированной В. lactis HN019 и L.rhamnosus HN001, в защите от инфекции S. typhimurium у мышей.
Противоинфекционные свойства оценивали путем измерения бактериальной транслокации в печень и селезенку. Иммуностимулирующие эффекты оценивали путем определения фагоцитарной функции (лейкоцитов периферической крови и перитонеальных макрофагов) и пролиферации лимфоцитов в ответ на Т-клеточный митоген (ФГА).
Была использована нижеследующая экспериментальная схема:
1. Использовали 6-7-недельных мышей BALB/c весом 20-30 г.
2. Мыши были произвольно распределены по 4 различным группам обработки и были помещены в отдельные клетки.
3. Все мыши получали корм на основе сухого сепарированного молока на протяжении всего эксперимента.
4. Испытуемым мышам за 7 дней до экспериментального заражения начинали ежедневно вводить B.1actis HN019 или L. rhamnosus HN001 (109 к.о.е./день) и это введение продолжали в течение всего испытания.
5. Мышей, которым вводили B. lactis HN019 или L.rhamnosus HN001, и контрольную группу (которым не вводили МКБ) перорально заражали Salmonella typhimurium (ATCC 1772), 8•105 к.о.е./день, в течение 5 дней, начиная со дня 7.
6. Группу неинфицированного контроля не подвергали заражению S.typhimurium.
7. На 6-й день после заражения мышей использовали для оценки транслокации бактерий в печень и селезенку и для оценки иммунной функции.
Результаты
Мыши, получавшие B.lactis HN019 и L.rhamnosus HN001, обнаруживали значительно более низкие уровни транслокации бактерий в печень и селезенку, чем мыши, которые получали лишь S.typhimurium (Фиг.3).
Экспериментальное инфицирование приводило к значительному подавлению фагоцитарной функции (Фиг.4); причем фагоцитарная активность у контрольных мышей, зараженных S. typhimurium, была значительно ниже, чем у неинфицированных мышей. Однако инфицирование бактерией S. typhimurium мышей, получавших B.lactis HN019 или L.rhamnosus HN001, не оказывало какого-либо влияния на фагоцитарную способность лейкоцитов периферической крови. На это указывают аналогичные уровни фагоцитарной активности у мышей, получавших B.lactis HN019 или L. rhamnosus HN001 и зараженных S. typhimurium, и у нормальных неинфицированных контрольных мышей.
Мыши, получавшие B. lactis HN019, как и мыши, получавшие L.rhamnosus HN001, обнаруживали более высокую степень пролиферации лимфоцитов в ответ на ФГА, чем контрольные мыши, зараженные S.typhimurium (Фиг.5). При этом какого-либо значимого отличия в ответах у мышей, получавших B.lactis HN019 или L.rhamnosus HN001, и у неинфицированных контрольных мышей не наблюдалось.
Эти результаты в целом позволяют предположить, что введение B.lactis HN019 или L.rhamnosus HN001 способно обеспечивать защиту организма от кишечных патогенов, таких как Salmonella typhimurium. Повышенная резистентность к инфекции сопровождается усилением иммунных функций.














Изобретение относится к биотехнологии и медицине. Штамм Lactobacillus rhamnosus HN001 и штамм Lactobacillus rhamnosus HN067 обладают иммуностимулирующим действием. Композиция содержит один или более штаммов молочнокислых бактерий в иммуностимулирующей концентрации и физиологически приемлемый наполнитель или разбавитель. Физиологически приемлемым наполнителем или разбавителем могут являться сквашенное молоко, йогурт, сыр, молочный напиток или сухое молоко. Способ предусматривает введение млекопитающему штаммов молочнокислых бактерий или композиций, содержащих эти штаммы бактерий. Изобретение позволяет повысить иммунитет млекопитающих. 10 с. и 3 з.п.ф-лы, 14 табл., 5 ил.
| СПОСОБ ВЫРАЩИВАНИЯ ХЛЕБОПЕКАРНЫХ ДРОЖЖЕЙ | 0 |
|
SU295794A1 |
| Экономайзер | 0 |
|
SU94A1 |
| СМЕСЬ ДЛЯ ПРИГОТОВЛЕНИЯ МОРОЖЕНОГО | 1997 |
|
RU2116035C1 |
