ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Данное изобретение относится к области химии и медицины, более конкретно к новым пептидам, полезным для модуляции активности клеток иммунной системы и для подавления воспаления.
УРОВЕНЬ ТЕХНИКИ
Иммунная система представляет собой чрезвычайно сложную комбинацию клеток и композиций, которая защищает хозяина, представляющего собой млекопитающего, от широкого разнообразия патогенов, и в то же время осуществляет контроль в его теле за вредными аберрациями, такими, как новообразования. Одна ветвь иммунной системы включает клетки, которые осуществляют функции иммунной системы, (а) лимфоциты, такие, как происходящие из костного мозга В-лимфоциты и происходящие из вилочковой железы клетки Т-лимфоциты и клетки, являющиеся естественными киллерами (NK), и (б) одноядерные фагоциты, включающие как моноциты, так и макрофаги. В то время, как лимфоциты в первую очередь связаны со специфическими иммунными реакциями, благодаря их способности специфически распознавать и отличать антигенные детерминанты, одноядерные фагоциты чаще всего осуществляют общее удаление чужеродных микроорганизмов посредством фагоцитоза, а также синтез и выделение цитокинов, что индуцируется либо непосредственно самим микроорганизмом, либо в ответ на стимулированные антигеном Т-лимфоциты. Функции лимфоцитных клеток и одноядерных фагоцитов тесно связаны и имеют важное значение для правильного функционирования иммунной системы.
Одной из важных субпопуляций лимфоцитных клеток являются Т-лимфоциты, которые получили свое название из-за того факта, что они вырабатываются тимусом (вилочковой железой). Т-лимфоциты представляют собой группу клеток, включающую комплекс различных клеток, которые могут быть цитотоксическими, обладающими многочисленными механизмами, вызывающими гибель клеток, или активирующими, путем синтеза различных цитокинов, функция которых заключается в активировании других клеток. Цитотоксические Т-лимфоциты (ЦТЛ) действуют, будучи ограниченными конкретным антигеном главного комплекса гистосовместимости (ГКГ), и экспрессируют Т-клеточный рецептор клеточной поверхности, который включает как α-, так и β-цепь, и который имеет специфическое сродство к конкретному комплексу ГКГ, связанное с пептидом в желобке этого ГКГ. ЦТЛ "подогнаны" таким образом, что в норме они не действуют против тех клеток, у которых находящийся в желобке пептид является эндогенным для хозяина. Однако в том случае, когда ГКГ является чужеродным или находящийся в желобке пептид является чужеродным для хозяина, то ЦТЛ атакуют такие клетки и уничтожают их. Другие лимфоцитные клетки, которые играют важную роль в иммунной реакции, включают В-лимфоциты и NK-клетки (естественные киллеры), причем на активность обоих этих типов клеток могут влиять другие клетки иммунной системы и различные полипептиды цитокинов.
Одноядерные фагоциты составляют другую основную популяцию клеток иммунной системы и состоят из клеток, имеющих общую линию дифференцировки, главной функцией которых является фагоцитоз. Одноядерные фагоциты происходят из клеток-предшественников ствола костного мозга, и после созревания и последующей активации они могут приобретать различные морфологические формы, включая неполностью дифференцированные моноцитные клетки и макрофаги. Должное функционирование одноядерных фагоцитов зависит от способности как продуцировать различные цитокиновые белки, так и реагировать на них.
Цитокины, такие, как различные интерфероны, интерлейкины, факторы некроза опухолей, хемокины, гематопоэтические факторы роста и факторы торможения миграции клеток представляют собой группу различных белков, синтезируемых широким разнообразием различных типов клеток иммунной системы. Наиболее важным является то, что цитокины продуцируются различными лимфоцитами и одноядерными фагоцитами и/или на них реагируют в ответ на различные воздействия. В основном цитокины продуцируются во время эффекторной фазы как естественного, так специфического иммунитета и служат посредниками и регуляторами как иммунной, так и воспалительной реакций. Цитокины, как и другие полипептидные гормоны, начинают свое действие со связывания специфических рецепторов на поверхности клеток-мишеней, причем их активация часто приводит к воспалительной реакции.
Хотя активация иммунного ответа и индуцированные цитокином воспалительные реакции чрезвычайно важны для здоровья хозяина и для правильного функционирования иммунной системы, существует ряд ситуаций, в которых такая активация является нежелательной. Одна из таких конкретных ситуаций связана с трансплантацией, при которой очень редко бывает идентичная совместимость между антигенами ГКГ донора и реципиента. Вторая ситуация связана с тем, что в результате "сбоя" в части ЦТЛ они атакуют клетки, у которых как ГКГ, так и связанный с ним пептид являются эндогенными, что имеет место при аутоиммунных болезнях, таких, как инсулинзависимый сахарный диабет (ИЗСД). Кроме того, бывают случаи, когда опосредованные цитокинами функции воспалительной реакции отрицательно влияют на здоровье хозяина, например, воспалительные реакции, связанные с такими заболеваниями, как септический шок, ревматоидный артрит, болезнь Крона, колит и тому подобными. Главным подходом для разрешения ситуаций, когда активация ЦТЛ нежелательна, стала иммуносупрессия. Однако иммуносупрессанты, такие, как циклоспорин A, FK-506 и им подобные, имеют многочисленные нежелательные побочные действия. Кроме того, применялись различные подходы, направленные на контролирование или подавление воспалительных реакций, однако многие из этих подходов также имеют один или несколько нежелательных эффектов. Поэтому существует значительный интерес к идентификации новых агентов, которые могут подавлять активацию лимфоцитных клеток, в частности ЦТЛ, и в то же время обладают менее универсальным иммуноподавляющим действием на иммунную систему и имеют меньше побочных действий, чтобы оставить хозяина с существенной частью его иммунной системы для защиты от случайных инфекций. Существует также значительный интерес к идентификации новых агентов, функция которых заключается в контролировании или подавлении воспалительных реакций.
В последние несколько лет появились сообщения о том, что олигопептиды эффективны для модуляции активности иммунной системы и увеличения продолжительности жизни аллогенных трансплантатов. Эти олигопептиды основаны на α1-домене антигена-В лейкоцита человека (HLA-B) и имеют консервативную аминокислотную последовательность Аrg-Х-Х-Х-Аrg-Х-Х-Х-Х-Тyr, в которой различные аминокислоты, обозначенные как X, варьируют в пределах относительно небольшого числа аминокислот, чтобы сохранить активность (например, см. WO 95/13288). Механизм, с помощью которого эти олигопептиды воздействуют на активность иммунной системы, неясен, в частности неясно, как они взаимодействуют с субтерапевтическими дозами циклоспорина, увеличивая при этом срок жизни аллогенных трансплантатов.
Сообщается также (Manolios, NOVEL PEPTID, заявка на патент РСТ, поданная на основе Австралийских заявок PN 0589 и PN 0590 16 января 1995) о действии на опосредованное Т-клетками воспаление, оказываемое олигасахаридами формулы
A-B-C-D-E,
в которой А отсутствует или представляет собой 1 или 2 гидрофобных остатка; В представляет собой положительно заряженную аминокислоту; С представляет собой пептид, состоящий из 3 - 5 гидрофобных аминокислот; D представляет собой положительно заряженную аминокислоту и Е отсутствует или представляет собой до 8 гидрофобных аминокислот. Пептиды, которые были синтезированы, представляют собой Gly-Leu-Arg-Ile-Leu-Leu-Leu-Lys-Val; Met-Gly-Leu-Arg-Ile-Leu-Leu-Leu; Leu-Gly-Ile-Leu-Leu-Leu-Gly-Val; Leu-Asp-Ile-Leu-Leu-Leu-Gly-Val; Leu-Arg-Ile-Leu-Leu-Leu-Ile-Leu-Val и Leu-Arg-Leu-Leu-Leu-Lys-Val. Эти последовательности предсказаны по последовательности трансмембранной последовательности TCR-α. В этой заявке нет доказательств того, что эти пептиды оказывают благоприятное действие на увеличение продолжительности жизни трансплантатов.
ОТНОСЯЩАЯСЯ К НАСТОЯЩЕЙ ЗАЯВКЕ ЛИТЕРАТУРА
Buelow et al., Transplantation 59:649-654 (1995) и ссылки на литературу, цитированную в этой публикации. Manolios et al., Nature Medicine 3:84-88 (1997) описывает полученные на основе логической схемы полипептиды, которые модулируют активность Т-клеток. Публикация WO 95/13288, Clayberger et al., в которой описываются пептиды, способные модулировать активность Т-клеток. Ссылки, описывающие способы конструирования соединений с помощью компьютера с использованием взаимосвязи структуры с активностью, включая Grassy et al., J. оf Molecular Graphics 13:356-367 (1995); Haiech et al., J. оf Molecular Graphics 13: 46-48 (1995); Yasri et al., Protein Engineering 11:959-976 (1996) и Ashton et al., Drug Discovery Today 1:71-78 (1996).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Предлагаются цитомодулирующие пептиды, которые способны (1) модулировать активность различных клеток иммунной системы, в частности лимфоцитных клеток, более конкретно ЦТЛ, (2) подавлять синтез "воспалительных" цитокинов клетками, способными продуцировать такие цитокины, являясь поэтому эффективными для лечения состояний, связанных с неблагоприятными воспалительными реакциями, (3) модулировать активность гемсодержащих ферментов и/или (4) задерживать наступление инсулинзависимого сахарного диабета (ИЗСД) у хозяина, восприимчивого к ИЗСД или имеющего ИЗСД, причем пептиды основаны на схеме, полученной в результате компьютерного программирования. Примерами этих соединений являются олигопептиды, включающие последовательность В-Х-Х-Х-В-Х-Х-X-J-Tyr, в которой В представляет собой основную аминокислоту, J представляет собой Gly, В или алифатическую гидрофобную аминокислоту, содержащую от 5 до 6 атомов углерода, и Х представляет собой любую аминокислоту, иную, чем алифатическая полярная аминокислота, в которой по меньшей мере три Х представляют собой одну и ту же алифатическую неполярную аминокислоту, ее димеры и ее D-стереоизомеры, и при этом аминокислотная последовательность может представлять собой часть кольца. Эти пептиды находят применение для подавления активации лимфоцитов иммунной системы, в частности цитотоксических лимфоцитов, либо отдельно, либо совместно с другими иммуносупрессантами, в частности, для увеличения продолжительности жизни трансплантатов. Описываемые здесь пептиды также находят применение для подавления синтеза воспалительных цитокинов (например, γ-интерферона, IL-1, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12, IL-13, IL-16, MIPlα и т.п.), тем самым являясь полезными для подавления воспалительных реакций, связанных с различными заболеваниями, такими, как ревматоидный артрит, септический шок, болезнь Крона, колит, аллергические реакции, аутоиммунные заболевания и тому подобные, для подавления активности гемсодержащих ферментов, таких, как гемоксигеназа, синтаза окиси азота и др. , и задержки наступления ИЗСД у пациентов, подверженных риску ИЗСД, как in vitro, так и in vivo. Введение пептидов может осуществляться ех vivo для подлежащего пересадке органа или in vivo, любыми удобными способами, включая прямое применение или введение пептида или нуклеиновой кислоты, кодирующей желаемый пептид, в количестве, достаточном для существенного подавления активации лимфоцитов, подавления синтеза воспалительных цитокинов и связанного с ними воспалительного процесса, подавления активности основанных на геме ферментов, активности, которая была предварительно связана с воспалительными реакциями и/или для задержки наступления ИЗСД.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
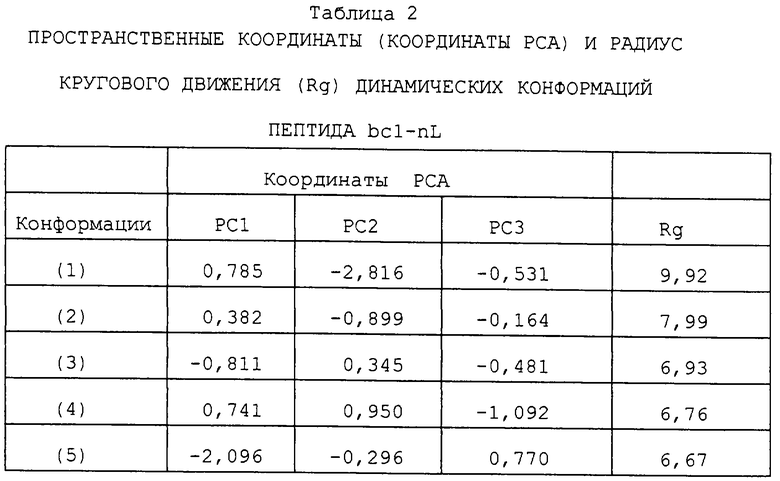
Фиг. 1 изображает конформационное пространственное образование кластеров пептида bcl-nL. Представленные конформации получены из анализа кластеров траектории пептида bcl-nL.
Фиг. 2 изображает проекции траекторий пептида в главный план эталонной траектории пептида D2.
ОПИСАНИЕ КОНКРЕТНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Предлагаются способы и композиции для модуляции активности клеток иммунной системы, в частности Т- и В-клеток и одноядерных фагоцитов, более конкретно активности клеток ЦТЛ и NK, in vitro и in vivo. Предлагаются также способы и композиции, эффективные для подавления синтеза воспалительного цитокина (цитокинов), вследствие этого находящие применение в терапевтическом лечении заболеваний, связанных с вредными воспалительными реакциями, для подавления активности различных гемсодержащих ферментов и/или для задержки наступления аутоимунных заболеваний, таких, как ИЗСД. Пептиды, обладающие специфическим действием в качестве пептидов, цитомодулирующих клетки ЦТЛ и NK, получены в соответствии с компьютерной программой, как описано в относящейся к данной заявке литературе. Следуя методике, описанной в указанной выше публикации Grassy et al., определили параметры, основанные на известных олигопептидах, для которых ранее была установлена способность подавлять активность Т-клеток. См. , например, Buelow et al., выше. Конформационное пространство, необходимое для иммуносупрессивной активности, было рассчитано так, как описано в указанной выше публикации Yasri et al.
Используя эти параметры, показали, что соединения, обладающие известной ингибиторной активностью по отношению к Т-клеткам, находятся в пределах этих параметров, и стало возможным разработать и испытать ряд новых пептидных соединений. Новые пептидные соединения оказались обладающими активностью от равной до превосходящей показатели известных соединений. Известные активные соединения включают α-домен HLA-B (антигена-В лейкоцита человека), в частности аминокислоты от 75 до 84 и вариации этой последовательности, в которых заменены не более двух аминокислот, при этом эти аминокислоты не включают R и Y, поскольку настоящее изобретение не охватывает такие известные соединения (см. , например, WO 95/13288 и цитированную выше публикацию Buelow et al). Известны также последовательности, основанные на трансмембранной области TCR-α человека, состоящей из этой последовательности и последовательностей, имеющих не более 2 мутаций в этой последовательности. Эти последовательности включают 2 основные аминокислоты, причем эти 2 основные аминокислоты отделены друг от друга четырьмя алифатическими гидрофобными аминокислотами, хотя в заявке указывается, что могут присутствовать от 3 до 5 гидрофобных аминокислот. Под мутацией подразумевается любое замещение одной аминокислоты другой аминокислотой или вставка, или делеция, каждая из которых считается за одну мутацию.
Желательно, чтобы в кор-последовательности описываемых здесь новых пептидных соединений имелись две или три основные аминокислоты, отделенные друг от друга гидрофобными аминокислотами в количестве от трех до четырех, в частности тремя гидрофобными аминокислотами, в частности, где N-конец представляет собой основную аминокислоту. Более желательно, чтобы С-концевая аминокислота представляла собой ароматическую аминокислоту, в частности тирозин. Особенно желательно, чтобы по меньшей мере одна из концевых аминокислот кор-олигопептида представляла собой аминокислоту окончания олигопептида, которая может иметь форму мономерного или олигомерного соединения.
Разработали новые выделенные пептидные соединения, включающие последовательность B-X-X-X-B-X-X-X-J-Tyr, в которой В представляет собой основную аминокислоту, а именно Lys или Аrg, в частности Аrg, по меньшей мере в одном положении, предпочтительно в обоих положениях, J представляет собой Gly, В или алифатическую гидрофобную кислоту, содержащую от 5 до 6 атомов углерода, в частности Gly или В, и Х представляет собой любую аминокислоту, иную, чем алифатическая заряженная аминокислота, предпочтительно любую аминокислоту, иную, чем полярная аминокислота, в которой по меньшей мере три Х представляют собой одну и ту же алифатическую неполярную аминокислоту, предпочтительно по меньшей мере четыре, Х представляют собой одну и ту же алифатическую неполярную аминокислоту и более предпочтительно - по меньшей мере все, кроме одного X, представляют собой одну и ту же алифатическую неполярную аминокислоту, олигомеры, в частности ее димеры и ее D-стереоизомеры, и при этом аминокислотная последовательность может представлять собой часть кольца.
Любой из N- и С-концов или оба эти конца часто могут быть увеличены, но не более чем (в сумме) на около 100, обычно не более чем на около 30, более обычно не более чем на около 20 аминокислот, часто не более чем на 9 аминокислот, причем аминокислоты имеют менее 25%, более обычно менее 20% полярных аминокислот, более конкретно менее 20% аминокислот, являющихся заряженными аминокислотами. Кроме того, концевая аминогруппа или карбоксильная группа олигопептида может быть модифицирована путем алкилирования или ацилирования с образованием эфиров, амидов или замещенных аминогрупп, причем алкильная или ацильная группа может иметь около от 1 до 30, обычно от 1 до 24, предпочтительно либо от 1 до 3, либо от 8 до 24, в частности от 12 до 18 атомов углерода.
Сюда также входят олигомеры, в частности димеры олигопептидов, которые могут быть типов "от головы к голове", "от хвоста к хвосту" или "от хвоста к голове", причем в них имеется не более шести повторов пептида. Кроме того, 1 или более аминокислот, вплоть до всего количества аминокислот, могут представлять собой D-стереоизомеры.
Кроме того, могут использоваться также структурно-ограниченные олигопептиды, такие, как циклические пептиды, содержащие примерно от 9 до 50, обычно от 12 до 36 аминокислот, причем аминокислоты, иные, чем указанные аминокислоты, могут присутствовать в качестве мостика. В некоторых случаях можно использовать мостики, представленные не аминокислотами. Имея концевые цистеины, можно сформировать дисульфидный мостик, замыкающий кольцо. Альтернативные способы образования кольца можно найти в публикациях Chen et al., Proc. Natl. Acad. Sci., USA, 89:5872-5876 (1992) и Wu et al., Protein Engineering 6:471-478 (1993).
Для целей настоящего изобретения аминокислоты (для большей части природных аминокислот или их D-стереоизомеров) разбиты на следующие категории.
1. Алифатические
(а) неполярные алифатические:
Gly, Ala, Val, nL, Ile, Leu,
(б) полярные алифатические:
(1) незаряженные
Cys, Met, Ser, Thr, Asn, Gln,
(2) заряженные:
Asp, Glu, Lys, Arg,
2. Ароматические
Phe, His, Trp, Tyr
в которых Pro может быть включен в неполярные алифатические аминокислоты, но обычно не включен в них. nL означает норлейцин, в котором неполярные алифатические аминокислоты могут быть замещены другими изомерами.
Предпочтительно, чтобы по меньшей мере 3 из шести аминокислот, обозначенных как Х в пептидной последовательности B-X-X-X-B-X-X-X-J-Tyr, представляли собой алифатические аминокислоты, содержащие от 5 до 6 атомов углерода, более предпочтительно, чтобы по меньшей мере 4 аминокислоты представляли собой алифатические аминокислоты, содержащие от 5 до 6 атомов углерода, более конкретно 6 атомов углерода. Другие аминокислоты могут представлять собой другие незаряженные алифатические аминокислоты, в частности неполярные алифатические аминокислоты или ароматические аминокислоты.
Кор-последовательность может быть увеличена в любом направлении с помощью аминокислот, которые в большинстве случаев являются липофильными, а именно алифатическими незаряженными аминокислотами, и ароматическими аминокислотами. Кроме того, как указано ранее, один или оба, а обычно один конец олигопептида может быть замещен липофильной группой, обычно алифатической или аралкильной, имеющей от 8 до 36, обычно от 8 до 24 атомов углерода и менее двух гетероатомов в алифатической цепи, причем гетероатомы обычно представляют собой, кислород, азот и серу. Цепь может быть насыщенной или ненасыщенной, желательно имеющей не более 3 сайтов, обычно не более 2 сайтов, алифатической ненасыщенности. Удобно, что можно использовать коммерчески доступные алифатические жирные кислоты, спирты и амины, такие, как лауриновая кислота, миристиловый спирт, стеариловый амин и т.п. Может быть проведена реакция липофильных групп с соответствующей функциональной группой олигопептида в соответствии с обычными способами, часто во время синтеза на носителе, в зависимости от сайта прикрепления олигопептида к носителю.
Композиции, представляющие особый интерес, имеют следующую формулу:
Arg-U-X-X-Arg-X-X-X-J-Tyr,
в которой все символы были определены выше, за исключением U, и U представляет собой незаряженную алифатическую аминокислоту или ароматическую аминокислоту, в частности неполярную алифатическую аминокислоту или ароматическую аминокислоту.
Последовательности по настоящему изобретению находят различное применение. В исследовательских целях их можно использовать для анализа физиологических путей, связанных с активацией и деактивацией ЦТЛ. Можно комбинировать ЦТЛ, в частности клеточные линии ЦТЛ, имеющие известные пептидные мишени, с пептидами по настоящему изобретению, в частности помеченными радиоактивной меткой, в присутствии или в отсутствие антигенпредставляющих клеток, по отношению к которым узко применимы эти ЦТЛ. После лизиса, осуществленного с помощью ЦТЛ, можно отделить активированные клетки ЦТЛ от покоящихся клеток ЦТЛ, с помощью маркера CD69, который отрегулирован in vitro на активацию. Разделение можно выполнить с помощью FACS (клеточного сортера с возбуждением флуоресценции) и анти-CD69, помеченного флуоресцентной меткой.
Выделив наиболее флуоресцентные клетки, например, с наиболее высоким показателем в 25%, затем лизируют эти клетки и выделяют белки, связанные с используемыми маркерами, например, с помощью хроматографии, неденатурирующего электрофореза или тому подобного. В качестве альтернативы белки разделяют с помощью электрофореза, а затем используют вестерн-блоттинг или другие методики с мечеными пептидами, чтобы идентифицировать белки, с которыми связаны пептиды по настоящему изобретению. Вместо радиоактивной метки можно использовать любой другой тип метки, обычно небольшую молекулу органического вещества, такого, как биотин, флюоресцентное вещество и т.п. Если используют биотин, то после разделения можно добавить авидин, причем авидин помечен с помощью метки, описанной выше.
Можно также сравнить Т-клетки, которые были объединены с антигенпредставляющими клетками в присутствии и в отсутствие пептидов по настоящему изобретению. В каждом случае можно получить библиотеки кДНК и использовать репрезентативный дифференциальный анализ, вычленение или тому подобные методы, чтобы обнаружить различия в экспрессии между клетками, которые были активированы в присутствии и в отсутствие пептидов по настоящему изобретению. Можно также определить, отличается ли реакция определенных субпопуляций ЦТЛ от реакции других субпопуляций на пептиды по настоящему изобретению, на основе их экспрессии или отсутствия экспрессии одного или более белков, в частности белков поверхности мембран. Таким образом можно идентифицировать ЦТЛ, которые можно удалить с помощью лейкофореза и подобных методов, чтобы свести к минимуму нежелательную атаку ткани клетками ЦТЛ.
Сообщается, что пептиды α-домена антигена-В лейкоцита человека (HLA-B) связываются с hsc70, который, как известно, служит шапероном ("спутником") и связывается с рядом последовательностей в своей роли шаперона.
В зависимости от их предполагаемого использования, в частности для введения относящимся к млекопитающим хозяевам, пептиды по настоящему изобретению можно модифицировать в широких пределах, чтобы заменить их распределение в кровотоке, уменьшить или увеличить связывание с компонентами крови, увеличить продолжительность жизни пептида в кровотоке и т.п. Пептиды по настоящему изобретению могут быть связаны с этими другими компонентами с помощью линкеров, которые являются расщепляющимися или нерасщепляющимися в физиологической среде крови. Эти пептиды могут быть присоединены в любой точке пептида, в которой присутствует функциональная группа, такая, как гидроксильная, тиольная, карбоксильная, аминогруппа или тому подобные. Желательно, чтобы связывание происходило либо в N-конце, либо в С-конце.
Пептид может быть присоединен к широкому разнообразию других олигопептидов или белков для разных целей. Например, пептиды по настоящему изобретению могут быть ковалентно связаны с иммуногеном для выработки антител по отношению к пептидам по настоящему изобретению, причем антитела могут служить для идентификации других пептидов, имеющих сравнимую конформацию. Кроме того, антитела можно использовать для получения антиидиотипических антител, которые могут конкурировать с пептидами по настоящему изобретению за связывание с сайтом-мишенью. Эти антиидиотипические антитела затем можно использовать для идентификации белков, с которыми связываются пептиды по настоящему изобретению.
В качестве альтернативы пептиды по настоящему изобретению могут экспрессироваться в соединении с другими пептидами или белками, составляя при этом часть цепи, либо внутреннюю, либо N- или С-конец. Путем обеспечения экспрессии пептидов по настоящему изобретению, можно добиться различных послеэкспрессионных модификаций. Например, посредством использования соответствующих кодирующих последовательностей можно добиться липидизации, например пренилирования или миристоилирования. В этой ситуации пептид по настоящему изобретению должен быть связан с липидной группой в конце, чтобы он был способен связываться с липидной мембраной, такой, как липосома. Для введения лекарственного средства можно использовать липосомы, в которых лекарство может быть введено в люмен липосомы, для того чтобы оно взаимодействовало с пептидами по настоящему изобретению в уменьшении активации ЦТЛ. Таким образом, иммуносупрессанты можно включить в люмен, для того чтобы иммуносупрессант и пептид по настоящему изобретению могли действовать локализованно.
Пептиды по настоящему изобретению можно ПЭГилировать, при этом полиэтиленоксигруппа обеспечивает увеличение продолжительности жизни в кровотоке. Пептиды по настоящему изобретению можно также соединять с другими белками, такими, как Fc или изотип IgG, которые могут комплементарно связываться или не связываться комплементарно, или с токсином, таким, как рицин, абрин, токсин дифтерии или тому подобными, в частности с А-цепью.
Можно получить эти композиции посредством получения гена, кодирующего определенный пептид или белок, соединенного с последовательностью ДНК, кодирующей пептид по настоящему изобретению. Этот ген можно ввести в соответствующий вектор экспрессии, множество которых коммерчески доступны, в результате чего ген затем экспрессируется в соответствующем хозяине. См. Sambrook et al., Molecular Biology: A Laboratory Manual, Cold Spring. Harbor Laboratories, Cold Spring Harbor, NY, 1989.
Пептиды по настоящему изобретению можно получить посредством химического синтеза или с помощью рекомбинантных технологий, как указано выше. В продаже имеются различные аппараты для синтеза, например автоматические синтезаторы компании Applied Biosystems Inc., Foster City, Beckman, CA, и др. При использовании синтезаторов натуральные аминокислоты можно заменить аминокислотами искусственного происхождения, в частности D-стереоизомерами, боковыми цепями, имеющими различную длину или функциональность, и тому подобным. Что касается рекомбинантных методов, можно получить последовательность нуклеиновых кислот, кодирующую множество пептидов по настоящему изобретению в тандеме, с помощью вставочной аминокислоты или последовательности, что позволяет осуществить расщепление до отдельного пептида или димера "от головы до хвоста". В случае, если отсутствует метионин, можно использовать вставочный метионин, который позволяет осуществить расщепление отдельной аминокислоты. В качестве альтернативы, можно ввести консенсусные последовательности, которые распознаются особыми протеазами для ферментативного расщепления. Конкретная последовательность и способ получения определяются имеющимися возможностями, экономическими соображениями, требуемой степенью чистоты и тому подобным.
Химическое связывание можно осуществить с различными пептидами или белками, включающими подходящие для образования химических связей функциональные группы, такие, как аминогруппы для образования амидов или замещенных аминов, например, для восстановительного аминирования, тиольные группы для образования тиоэфира или дисульфида, карбоксильные группы для образования амидов и тому подобное. Особый интерес представляют пептиды, имеющие по меньшей мере 2, более обычно 3 и не более чем около 60 лизиновых групп, в частности полилизины, имеющие примерно от 4 до 20, обычно от 6 до 18 лизиновых единиц, называемые MAP, в которых пептиды по настоящему изобретению связаны с лизиновыми аминогруппами, как правило, составляющими по меньшей мере около 20%, более обычно по меньшей мере около 50% от имеющихся аминогрупп, что обеспечивает получение мультипептидного продукта. Таким образом, можно получить молекулы, имеющие множество пептидов по настоящему изобретению, в которых ориентация пептидов по настоящему изобретению осуществляется в одном и том же направлении, и в результате получают связывающую группу, обеспечивающую ди- или олигомеризацию "от хвоста до хвоста". В качестве альтернативы можно использовать другие природные или синтетические пептиды и белки, чтобы получить скелет для прикрепления к нему пептидов по настоящему изобретению в С-конце.
В большинстве случаев используемые композиции включают по меньшей мере 20% по массе желаемого продукта, более обычно по меньшей мере 75% по массе, предпочтительно по меньшей мере около 95% по массе, а для терапевтических целей - обычно по меньшей мере около 99,5% по массе по отношению к примесям, связанным со способом получения продукта и его очистки. Обычно эти проценты основаны на общем количестве белка.
При желании в пептид можно ввести различные группы во время синтеза или во время экспрессии, что позволит осуществить связывание с другими молекулами или с поверхностью. Так, цистеины можно использовать для получения тиоэфиров, гистидины - для связывания с комплексом ионов металла, карбоксильные группы - для образования амидов или сложных эфиров, аминогруппы - для образования амидов и т.п. В качестве альтернативы можно использовать большое разнообразие меток, как описано выше, включая лиганды для связывания с антителами или естественными рецепторами, причем пептиды могут быть присоединены к носителю или, в качестве альтернативы, к другой молекуле. Как уже указывалось, пептиды по настоящему изобретению могут связываться с hsc70, что позволяет произвести выделение и очистку hsc70 от других белков, находящихся в клетке.
Пептиды по настоящему изобретению можно использовать для модуляции пролиферации и/или активации клеток ЦТЛ и/или NK. Путем соединения пептидов по настоящему изобретению с лимфоцитами модулируют пролиферацию и/или активацию ЦТЛ с помощью антигенпредставляющих клеток, как правило, по меньшей мере примерно на 20%, более обычно - по меньшей мере на 40% и предпочтительно - по меньшей мере примерно на 60%, на основе процента лизиса, как описано в экспериментальном разделе. Ингибирующая концентрация ИК50 для лизиса в основном составляет менее чем около 500 мкг/мл, как правило, менее чем около 200 мкг/мл и более чем около 0,1 мкг/мл, обычно более чем около 1 мкг/мл.
Композиции по настоящему изобретению можно использовать in vitro для подавления лизиса антигенпредставляющих клеток Т-клетками (первые являются мишенью вторых). Таким образом, в тех исследованиях, в которых желательно поддерживать смеси клеток, в которых клетки ЦТЛ были бы активированными и уничтожали антигенпредставляющие клетки, такие, как макрофаги или В-лимфоциты, или другие клетки, которые могли бы служить клетками-мишенями, например клетки новообразований, инфицированные вирусом клетки или тому подобные, лизис можно подавлять, что позволит поддерживать клеточную популяцию в течение эксперимента.
Композиции по настоящему изобретению можно также использовать ex vivo. В случае трансплантации органов, в частности твердых органов или определенных клеток, ксеногенных или аллогенных, донорный орган может быть погружен в ванну со средой, включающей пептиды по настоящему изобретению. В результате этого не допускается участие присутствующих в имплантате ЦТЛ в реакции "трансплантат против хозяина". Кроме того, в течение того периода, когда пептиды по настоящему изобретению остаются связанными с имплантатом, подавляется активация ЦТЛ реципиента. Как правило, концентрация пептида в среде варьирует в зависимости от активности пептида, уровня желаемого подавления, присутствия других соединений, отрицательно влияющих на активацию ЦТЛ и т.п. Как правило, концентрация находится в интервале примерно от 0,1 до 100 мкг/мл, более обычно - в интервале примерно от 1 до 10 мкг/мл. Другие иммуносупрессанты, которые также могут присутствовать, включают циклоспорин A, FK506, антитела для белков плазменной мембраны, связанные с отторжением трансплантата, такие, как антитела к CD4, CD8, CD2, LFA-1, ICAM-1, CD28 и тому подобные. Применяют субтерапевтические дозировки, как правило, если они присутствуют, в количестве не менее чем около 5% от обычной дозировки и не более чем около 75%, обычно в интервале примерно от 10 до 60%. Другие компоненты среды для ванны, в которую погружают трансплантаты, обычно представляют собой компоненты, обычно используемые в растворах для сохранения органов, например HBSS. Время, в течение которого орган поддерживается в среде, обычно находится в интервале от 2 до 72 часов.
Композиции по настоящему изобретению также можно применять in vivo, путем введения композиций пептидов по настоящему изобретению любым удобным способом. Композиции по настоящему изобретению можно вводить перед имплантацией, при этом введение обычно начинают не ранее чем примерно за 14 суток до имплантации и предпочтительно по меньшей мере одну дозу вводят в течение трех суток введения. Композиции по настоящему изобретению можно вводить в период времени, начинающийся примерно за 6 часов до имплантации, и можно продолжать введение в течение предусмотренного схемой введения времени после имплантации, обычно не более 30 суток после имплантации, более обычно не более 20 суток после имплантации. Однако после имплантации композиции пептидов по настоящему изобретению можно вводить по мере необходимости, в зависимости от реакции реципиента на орган или клетки. В некоторых ситуациях композиции по настоящему изобретению можно водить хронически, в течение всего времени, пока имплантат присутствует в организме хозяина.
Как правило, в случае, если пептидную композицию вводят непосредственно хозяину, то вводимый болюс с композицией по настоящему изобретению содержит в интервале примерно от около 0,1 до 50, более обычно - от около 1 до 25 мг/кг, в расчете на массу тела хозяина. Хозяин может представлять собой любое млекопитающее, включая сельскохозяйственных животных, домашних животных, лабораторных животных, приматов, в частности человека. Это количество, как правило, уточняют, в зависимости от времени полужизни пептида, причем время полужизни, как правило, составляет по меньшей мере 1 минуту, более обычно - по меньшей мере около 10 минут, желательно в интервале от около 10 минут до 12 часов. Короткие сроки полужизни приемлемы, поскольку эффективность может быть достигнута с помощью индивидуальных доз или путем продолжительного вливания или путем применения повторных доз. Дозировки из нижней части этого интервала и даже более низкие дозировки могут применяться в тех случаях, когда пептид имеет увеличенное время полужизни или если он применяется в виде депо, такого, как частицы, включающие медленно высвобождаемые композиции, введенные в матрицу, которая сохраняет пептид в течение продолжительного периода времени, например в коллагеновую матрицу, или при использовании насоса, который постоянно подает пептид в течение продолжительного времени с существенно постоянной скоростью, и т.п.
Кроме введения пептидных композиций по настоящему изобретению непосредственно в культуру клеток in vitro, подачи их к твердым органам или определенным клеткам ех vivo, или введения in vivo хозяину, представляющему собой млекопитающее, им могут вводиться или к ним подаваться также молекулы нуклеиновой кислоты (ДНК или РНК), кодирующие пептиды по настоящему изобретению, что обеспечивает эффективный источник пептидов по настоящему изобретению для желаемого применения. В большинстве случаев молекулы нуклеиновой кислоты, кодирующие пептиды по настоящему изобретению, можно клонировать в любое количество хорошо известных экспрессирующих плазмид (см. Maniatis et al., выше) и/или вирусных векторов, предпочтительно аденовирусных или ретровирусных векторов (см. , например, Jacobs et al., J. Virol., 66:2086-2095 (1992), Lowenstein, Bio/Technology, 12: 1075-1079 (1994), и Berkner, Biotechniques, 6:616-624 (1988)), под транскрипционным управлением регуляторных последовательностей, функция которых заключается в стимулировании экспрессии нуклеиновой кислоты в соответствующих условиях окружающей среды. Такие основанные на нуклеиновых кислотах носители можно подавать непосредственно к ткани трансплантата ex vivo (например, вирусное инфицирование клеток ех vivo для трансплантата) или к желаемому сайту in vivo, например, с помощью инъекции, катетера и т.п., либо, в случае основанных на вирусах носителей, можно их вводить системно. В качестве варианта можно применять тканево-специфические промоторы, с помощью которых представляющий интерес пептид экспрессируется только в определенной ткани или типе клеток по выбору. Способы получения таких основанных на нуклеиновых кислотах носителей с помощью рекомбинантных технологий хорошо известны в данной области техники, также как и способы введения основанных на нуклеиновых кислотах носителей для выработки пептидов, как in vitro, так и in vivo.
Трансплантация может включать любой орган или клетки, включая такие органы, как сердце, почки, легкое, глаза, печень, кишка, кровеносный сосуд или другой орган, а также клетки, такие, как β-инсулярные клетки, клетки костного мозга или другие клетки, причем орган или клетки могут быть аллогенными или ксеногенными, в частности, когда один или более антигенов класса I или II МНС (ГКГ, главного комплекса гистосовместимости) у донора и реципиента разные.
Пептиды по настоящему изобретению, в отдельности или в виде конъюгатов, или представляющие собой нуклеиновые кислоты носители, кодирующие такие пептиды, можно получить в виде лекарственных форм в фармацевтически пригодных средах, таких, как, например, физиологический раствор, ЗФР (забуференный фосфатом физиологический раствор), водный раствор этанола, глюкоза, пропиленгликоль или тому подобные, или в виде твердых лекарственных форм в соответствующих наполнителях, как правило, в фармакологически эффективной дозе. Концентрацию пептидов или кодирующих их нуклеиновых кислот определяют эмпирически, в соответствии с обычными методами для конкретной цели. Лекарственные формы могут включать бактерицидные агенты, стабилизаторы, буферы или тому подобное. Количество, вводимое хозяину, варьирует в зависимости от того, что именно вводится, от цели введения, такой, как профилактика или терапия, состояния хозяина, способов введения, количества введений и продолжительностью перерыва между введениями и тому подобного, и это количество может быть определено эмпирически специалистами в данной области техники. Чтобы увеличить продолжительность времени полужизни пептида по настоящему изобретению или конъюгатов пептида по настоящему изобретению, пептиды можно заключить в капсулы, ввести в люмен липосом, приготовить в виде коллоида либо можно использовать другие обычные методы, обеспечивающие увеличенную продолжительность жизни пептидов ex vivo или in vivo.
Пептиды по настоящему изобретению способны подавлять продуцирование клетками воспалительных цитокинов. Воспалительные цитокины, ингибируемые пептидами по настоящему изобретению, включают, например, фактор некроза опухолевых клеток, включая фактор-α некроза опухолевых клеток (TNF-α), интерфероны, включая γ-интерферон (INF-γ), интерлейкин IL-l, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12, IL-13, 1L-16, MIPl-α, хемокины, гематопоэтические факторы роста и тому подобные, как in vitro, так и in vivo. Поэтому пептиды по настоящему изобретению применимы как для профилактического, так и для терапевтического подавления воспалительных реакций, связанных с различными заболеваниями, такими, как септический шок, болезнь Крона, колит, ревматоидный артрит и другие аутоиммунные заболевания, аллергические реакции, атеросклероз, инфицирование, а также во многих других ситуациях, когда желательна противовоспалительная реакция.
Композиции по настоящему изобретению также способны
модулировать активность гемсодержащих ферментов как in vitro, так и in vivo. Как показано ниже, пептиды по настоящему изобретению имитируют порфириноподобные структуры, способные модулировать активность гемсодержащих ферментов, таких, как гемоксигеназа (ГО), различные изоформы синтазы окиси азота (NOS), циклооксигеназа, гуанилатциклаза и тому подобные. Поэтому композиции по настоящему изобретению могут использоваться в ситуациях, когда желательно отрегулировать экспрессию гемсодержащего фермента, например гемоксигеназы. В связи с этим известно, что гемоксигеназа участвует в путях метаболизма, иных, чем модуляция лимфоцитической активности. Поэтому при регулировке экспрессии гемоксигеназы будет оказано неблагоприятное воздействие на те пути метаболизма, в которых участвует гемоксигенеза. См., например, Willis et al., Nature Medicine, 2:87-89 (1996).
Кроме того, сообщается, что гемсодержащие ферменты, такие, как гемоксигеназа, являются фактором воспалительной реакции и могут оказывать противовоспалительное действие. Поэтому пептиды по настоящему изобретению можно использовать в культурах для оценки роли гемсодержащих ферментов в различных физиологических процессах, путем сравнения клеточных реакций в присутствии и в отсутствие пептидов по настоящему изобретению. Пептиды по настоящему изобретению можно также использовать in vivo для уменьшения воспалительной реакции, связанной с септическим шоком, болезнью Крона, колитом, включая как неспецифический язвенный колит, так и слизистый колит, ревматоидным артритом, атеросклерозом, повторной перфузией, инфекцией и тому подобным. Описание использования пептидов по настоящему изобретению для модуляции лимфоцитической активности существенно применимо для этих показаний.
Пептиды по настоящему изобретению также можно использовать для задержки наступления аутоиммунного заболевания у относящегося к млекопитающим субъекта, подверженного риску развития такого аутоиммунного заболевания. Пептиды по настоящему изобретению особенно применимы для задержки наступления инсулинзависимого сахарного диабета (ИЗСД), ревматоидного артрита или системной эритематозной волчанки у млекопитающих, подверженных риску развития этих заболеваний. Описание использования пептидов по настоящему изобретению для модуляции лимфоцитической активности существенно применимо для этих показаний.
Нижеследующие примеры предлагаются только в качестве иллюстраций, а не в качестве ограничения настоящего изобретения.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Компьютерная программа, использованная для предсказания и разработки иммуносупрессивной активности пептидов и псевдопептидов, осуществлялась следующим образом.
1. МЕТОДИКА
На основе набора первоначальных экспериментальных данных, полученных для пептидов, проявляющих или не проявляющих иммуносупрессивную активность, было получено следующее.
i. Консенсусная последовательность, содержащая аминокислоты, требующиеся для активности и позволяющие получать библиотеки новых пептидов или псевдопептидов.
ii. Набор физико-химических и топологических свойств, связанных с активностью, и преобразованный в набор ограничений с помощью методики картирования переменных (Grassy et al. , J. of Molecular Graphics, 13:356-367 (1995)).
2. КАРТИРОВАНИЕ ПЕРЕМЕННЫХ
Способ основан на физико-химических и топологических ограничениях, полученных на основе результатов первоначального набора данных.
ФИЗИКО-ХИМИЧЕСКИЕ ОГРАНИЧЕНИЯ
Способ требует определения физико-химических ограничений, определяемых как диапазоны свойств для указанной биологической активности. Способ расчета, использованный для определения набора ограничений, называется способом картирования переменных и описывается ниже.
СПОСОБ КАРТИРОВАНИЯ ПЕРЕМЕННЫХ
Этот количественный метод состоит из оценки распределения (глобального или процентного) активных и неактивных молекул как функции значений данных параметров. Суперпозиция всех графов (активность-свойство) указывает, для определенных параметров, на лимитирующие значения (низкие и/или повышенные), которые необходимы для получения активного соединения. Этот графический способ выявляет качественные нелинейные зависимости между активностью и молекулярными свойствами. Что касается свойств, связанных с взаимодействиями лиганд рецептора, то было четко установлено, что существование строгой сопряженности признаков, определяющих приспосабливаемость к рецептору, подразумевает включение определенных структурных и физико-химических свойств. Этот способ дает простые правила, которые можно использовать для прогнозирования активности неизвестных продуктов. Графическое представление, показывающее количество случаев успешного выполнения правила по отношению к числу случаев нарушения правила, позволяет сравнить распределения с показателями активности для всего изучавшегося набора молекул.
3. ФИЗИКО-ХИМИЧЕСКИЕ И ТОПОЛОГИЧЕСКИЕ ПАРАМЕТРЫ, ИСПОЛЬЗОВАННЫЕ ДЛЯ ОПРЕДЕЛЕНИЯ ОГРАНИЧЕНИЙ, СВЯЗАННЫХ С ИММУНОСУПРЕССИВНОЙ АКТИВНОСТЬЮ ПЕПТИДОВ И ПСЕВДОПЕПТИДОВ.
ЛИПОФИЛЬНОСТЬ
Липофильность пептидов выражали в виде 1оgР (где Р представляет собой коэффициент распределения определенного пептида между водой и н-октанолом). Молекулярные значения logP можно рассчитать с помощью программы TSAR 2.31, используя атомные возрастающие значения logP, определенные Гоузом и др. (Ghose et al., J. Chem. Inf. Comput., 29:163 (1989). Как показано с помощью анализа первоначального набора данных, липофильность иммуносупрессивного пептида должна быть ≥ -6,85.
ТОПОЛОГИЧЕСКИЕ ИНДЕКСЫ
ИНДЕКС БАЛАБАНА (Balaban, Chem. Phys., 89:399 (1982))
Индекс Балабана, рассчитываемый для соединенного молекулярного графа (Н подавленное), рассчитывают следующим образом: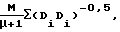
где М представляет собой количество ребер в графе, μ представляет собой цикломатическое число графа, т.е. минимальное количество ребер, которые должны быть удалены до того, как G станет ацикличным, и Di=ΣDii (с j=1) представляет собой матрицу расстояний кратчайшего пути между двумя вершинами графа.
МОЛЕКУЛЯРНЫЙ ОБЪЕМ
Молекулярный объем рассчитывают, принимая стандартное значение радиусов Ван дер Ваальса для каждого элемента. Этот расчет выполняется на расширенной конформации пептида.
ЭЛЛИПСОИДНЫЙ ОБЪЕМ
Этот объем рассчитывают после определения трех компонентов момента инерции молекулы, принимая средние значения атомных масс для составляющих соединение атомов. Этот расчет выполняется на расширенной конформации пептида.
МОЛЯРНАЯ ПРЕЛОМЛЯЮЩАЯ СПОСОБНОСТЬ
Молярную преломляющую способность рассчитывают с помощью значений атомной молярной преломляющей способности, рассчитанных Ghose et al. (цитированы выше).
ДИПОЛЬНЫЙ МОМЕНТ
Этот параметр рассчитывают на расширенной конформации пептидов. Общий дипольный момент для молекулы выражается в единицах Debye.
μ = eΣ riqi,
где ri представляет собой расстояние от атома i до начала координат, a qi представляет собой заряд атома i. Заряды атомов рассчитывают с помощью способа Charge-2 (Abraham and Smith, J. Comput. Aided Mol. Design, 3:175-187 (1989)).
ИНДЕКС КИРА Chi V 4
Этот индекс представляет собой один из индексов связности, разработанных Киром (L. B. Kier). Индекс Кира Chi V 4 рассчитывают в несколько этапов (Н включенное).
А. Определение и нумерация всех путей длины 4 на молекулярном графе пептида.
Б. Расчет каждого пути длины 4 из следующих значений:
C
для j=1,4, где δi=Zi-hi определяется для атома как разность между общим количеством валентных электронов Zi и количеством hi атомов водорода, связанных с атомом i.
В. Суммирование всех этих значений, касающихся всего набора субграфов длины 4 на графе
χ = ∑ (c
ИНДЕКС КИРА КАППА АЛЬФА
Индекс Кира Каппа альфа 1 (Kα1)
Если А представляет собой общее количество атомов молекулы (Н включенное), то Kα1 равно следующему: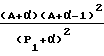
с: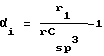
ri представляет собой ковалентный радиус атома i, а rCsp 3 ковалентный радиус углерода sp3, P1 представляет собой общее количество путей длины = 1 вдоль молекулярного графа изучаемого пептида.
Индекс Кира Каппа альфа 2 (Kα2)
Если А представляет собой общее количество атомов молекулы (Н включенное), то Kα2 равно следующему:
с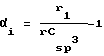
ri представляет собой ковалентный радиус атома i, а rCsp 3 ковалентный радиус углерода sp3, Р2 представляет собой общее количество путей длины = 2 вдоль молекулярного графа изучаемого пептида.
ГИБКОСТЬ Phi
Основываясь на вышеприведенных формулах, гибкость молекулы можно определить следующим образом:
Phi = (Kα1)•(Kα2)/A,
где А представляет собой общее количество атомов (Н включенное).
КОЛИЧЕСТВА АТОМОВ И ГРУПП
В качестве ограничений использовали также количество следующих типов атомов:
- общее количество атомов кислорода в пептиде,
- общее количество атомов азота в пептиде.
В качестве ограничений использовали также количество следующих групп:
- общее количество этильных групп,
- общее количество гидроксильных групп.
4. ЗНАЧЕНИЯ ОГРАНИЧЕНИЙ
СОЗДАНИЕ БИБЛИОТЕК ПЕПТИДОВ ИЛИ ПСЕВДОПЕПТИДОВ
Начиная с консенсусной последовательности Аrg-Х-Х-Х-Аrg-Х-Х-Х-Х-Тyr, в которой Х представляет собой аминокислоту, которая определена в ранее приведенной аналогичной формуле, рассчитывали вышеописанные физико-химические и топологические параметры и определяли, находятся ли эти параметры в пределах ограничений, определенных первоначальным набором данных. Например, начиная с X= Leu, nLeu, Trp, Тyr, Gly и Val, создали библиотеку, состоящую из 279936 молекул, и только 26 из них удовлетворяли условиям требуемых ограничений.
Диапазоны свойств, необходимых для получения биологической активности, суммированы в таблице 1.
8. ПОЛУЧЕНИЕ ХАРАКТЕРИСТИК КОНФОРМАЦИОННОГО ПРОСТРАНСТВА, ВОВЛЕЧЕННОГО В ИММУНОСУПРЕССИВНУЮ АКТИВНОСТЬ ПЕПТИДОВ И ПСЕВДОПЕПТИДОВ
ВЕКТОР ПРОСТРАНСТВЕННОЙ АВТОКОРРЕЛЯЦИИ СТРУКТУРЫ 3D
Концепция автокорреляционного описания молекулярной структуры была впервые введена Брото и др. (Broto et al. , Eur. J. Med. Chem., 19:66-70 (1984)). Этот вектор в основном представляет дискретизированное распределение расстояния, полученное из матрицы межатомных расстояний молекулы. Первый компонент этого вектора (А ) равен количеству атомов структур, а другие компоненты, А1. . .Аn, определяются количеством пар атомов, которые отделены расстоянием в пределах интервала, определяемого нижней границей (n-1)Di, где n представляет собой порядок элемента дискретизации вектора, а Di - приращение расстояния. Подобным же образом можно рассчитать распределение атомной характеристики Р. В этом случае взвешенный компонент автокорреляции АРn получают путем суммирования продуктов значений характеристики Р на атомах i, j, имеющих расстояния друг от друга, принадлежащие к интервалу расстояний [(n-1)Di, nDi]. Затем определяют количество компонентов вектора из уравнения nmax=(Dmax/Di)+1, в котором Dmax представляет собой наибольшее межатомное расстояние в структуре.
Вектор автокорреляции проявляет несколько полезных свойств.
- С помощью этого вектора можно существенно уменьшить объем данных конформации. Общая конформация описывается ограниченным набором n числовых значений.
- Вектор очень легко рассчитать на основе данных координат 3D. Поэтому можно рассчитать и сохранять этот вектор в течение имитации молекулярной динамики, причем в этом процессе происходит уменьшение размера хранения по сравнению с классическим сохранением набора полных матриц расстояний, что позволяет осуществлять имитацию намного дольше, чем обычно.
- Вектор автокорреляции конформации является переходно и ротационно инвариантным, а также независимым от количества атомов в молекуле.
- Этот вектор чувствителен как к малым, так и к большим изменениям в конформации: чем больше изменяется конформация, тем больше модифицируются компоненты вектора. Чувствительность зависит от приращения расстояния, выбранного для расчетов, но приращение от 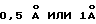 (малые молекулы) до
(малые молекулы) до 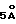 (макромолекулы) является хорошим выбором для обычных имитаций (Yasri et al., Protein Engineering, 11:959-976 (1996)).
(макромолекулы) является хорошим выбором для обычных имитаций (Yasri et al., Protein Engineering, 11:959-976 (1996)).
Можно проанализировать только часть структуры или только конкретный субнабор атомов этой структуры, например, С2 в белках, атомы N, тяжелые атомы и т.п. Вектор полностью определяется знанием структуры, поэтому можно осуществить сравнение различных структур, используя этот вектор без какого-либо эталона.
АНАЛИЗ МОЛЕКУЛЯРНОЙ ДИНАМИКИ С ПОМОЩЬЮ ВЕКТОРОВ АВТОКОРРЕЛЯЦИИ 3-D
Применительно к пептиду HLA-B2702.75-84 (аминокислотная последовательность Arg-Glu-Asn-Leu-Arg-Ile-Ala-Leu-Arg-Tyr) и к его различным активным и неактивным производным пептидам выполнили имитации молекулярной динамики с помощью программы AMBER 4.1. Имитация одной наносекунды динамики создает набор в 103 конформаций (одна конформация в пикосекунду). Для каждой конформации рассчитали вектор автокорреляции 3D с помощью программы TSAR, с приращением расстояния в 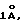 и весь набор конформаций сохранили в виде векторов автокорреляции 3D против матрицы времени (103xn).
и весь набор конформаций сохранили в виде векторов автокорреляции 3D против матрицы времени (103xn).
Целью работы было определить конформационное пространство, ответственное за иммуносупрессивную активность, путем сравнения конформационных пространств активного и неактивного пептидов, с использованием методики, описанной в ссылках, указанных в разделе "Относящаяся к настоящей заявке литература".
СТАТИСТИЧЕСКИЙ АНАЛИЗ
КЛАСТЕРНЫЙ АНАЛИЗ
Для того, чтобы сравнить различные конформации, определили матрицу расстояний между всеми этими конформациями в гиперпространстве, определяемом компонентами их невзвешенных векторов автокорреляции 3D. Чем более аналогичны структуры двух соединений, тем короче расстояние между ними. Этот способ обеспечивает возможность количественного определения жесткого молекулярного соответствия. При использовании стартовой информации в качестве эталона числовое значение этого расстояния является аналогом среднеквадратического отклонения.
АНАЛИЗ ГЛАВНЫХ КОМПОНЕНТОВ (РСА)
РСА представляет собой многомерный статистический метод анализа данных, пригодный для представления молекул в гиперпространстве их свойств (молекулярных дескрипторов). РСА можно использовать для того, чтобы свести большое число дескрипторов к меньшему числу синтетических ортогональных переменных, получаемых из линейной комбинации исходных дескрипторов. Этот способ сохраняет наибольшую часть первоначальной информации. Исходные переменные были нормализованы и была рассчитана диагонализация ковариантной матрицы с помощью классической трансформационной программы Якоби (Jacobi). Компоненты автокоррелограмного вектора 3К обеспечивают хорошее описание структуры 3К различных конформаций, но неудобны для работы, поскольку они содержат слишком много данных, что не позволяет обеспечить их легкую визуализацию. РСА может уменьшить размерность данных до представления в виде 2D или 3D, которое содержит максимально возможное количество исходной информации. При использовании РСА иммуносупрессивные пептиды проявляют хорошо определяемое общее конформационное пространство. Все пептиды, способные достигнуть этих конформационных показателей, могут проявлять иммуносупрессивную активность.
КООРДИНАТЫ КОНФОРМАЦИОННОГО ПРОСТРАНСТВА БИОАКТИВНОЙ КОНФОРМАЦИЙ ПЕПТИДА bcl-nL
На фиг. 1 показано двумерное конформационное пространство и связанные с ним конформации пептида bcl-nL, в котором пептид bcl-nL имеет аминокислотную последовательность Arg-nL-nL-nL-Arg-nL-nL-nL-Gly-Tyr и в котором nL представляет собой норлейцин (см. ниже). Показанные структуры были получены посредством применения кластерного анализа на всей траектории пептида bcl-nL.
ГЛАВНЫЕ КОНФОРМАЦИИ ПЕПТИДА bcl-nL
Структурные свойства главных исследованных конформаций пептида bcl-nL (фиг. 1, (1), (2), (3), (4) и (5)) и его конформационное пространство суммированы в таблице 2. Эти свойства касаются координат в трехмерном пространстве, определяемых тремя первыми главными компонентами (координатами РСА), а также радиуса кругового движения (Rg).
КОНФОРМАЦИОННОЕ ПРОСТРАНСТВО АКТИВНЫХ ПЕПТИДОВ
Траекторию пептида D2 (аминокислотная последовательность Arg-Val-Asn-Leu-Arg-Ile-Ala-Leu-Arg-Tyr) описали с помощью метода автокорреляции 3D и данные проанализировали с помощью анализа главных компонентов. Это дало возможность получить главный план, определяемый 2 первыми главными компонентами, который содержит все конформации, исследованные в течение траектории. Траекторию пептида D2 использовали в качестве эталона траектории и все рассчитанные траектории спроектировали в этот главный план (фиг. 2).
Иммуносупрессивные пептиды проявляют хорошо определяемое общее конформационное пространство, обладающее следующими характеристиками.
Размеры РСА:
PC1: минимум = -2,0; максимум = 2,0
РС2: минимум = -2,0; максимум = 1,0
РС3: минимум = -1,0; максимум = 1,0
Пептиды, полученные в виде композиций, даны в таблице 3.
ПРИМЕР 1. АНТИПРОЛИФЕРАТИВНАЯ АКТИВНОСТЬ bс-ПЕПТИДОВ
Взрослые 6-8-недельные самцы мышей линий C57BL6/J (В6, Н-2d), Ва1b/с(Н-2d) и CBA/JH-2k) были приобретены в Jackson Laboratory, Bar Harbor, ME. Их сохраняли и поддерживали в лаборатории для содержания животных в SangStat Medical Corporation согласно указаниям Национального Института здоровья (США) и в соответствии с правилами Департамента здравоохранения.
Пептиды синтезировали в synt:em (Nimes, Франция) с помощью синтезатора пептидов и методов Fmoc-химии. Все пептиды синтезировали в виде амидов, а затем преобразовывали в ацетатные соли. Пептиды очищали с помощью препаративной ВЭЖХ с обращенной фазой и с помощью аналитической ВЭЖХ с обращенной фазой показали, что они являются более чем на 95% гомогенными. Аминокислотное содержание подтверждали с помощью анализа аминокислот. Перед использованием пептиды сначала растворяли в 1 объеме ДМСО (Sigma), а затем добавляли 99 объемов культуральной среды. Окончательная концентрация ДМСО в культуре составляла не более 0,25%.
Суспензии клеток селезенки получали после лизиса красных кровяных телец с помощью гипотонического шока. Затем клетки промывали в культуральной среде и, наконец, ресуспендировали в среде RPMI-1640 с 10% FBS (сыворотки плода коровы)(среда R-10) или в среде без сыворотки AIM-V (Gibco, Grand Island, NY).
Клетки селезенки, выделенные из мышей линии СВА, затем подвергали стимуляции (по 2•105 на лунку) моноклональным антителом анти-СD3 (Pharmagen, Сан Диего), при окончательной концентрации от 0,1 до 1 мкг/мл в 96-луночных круглодонных микрокультуральных планшетах (Nunc, Дания). В начале культивирования добавляли bс-пептиды в различных концентрациях. Клетки инкубировали в течение 3 суток при 37oС и 5% СО2. За 24 часа до сбора выросших в культуре клеток в каждую лунку добавляли по 1 мк Ci[3H]-TdR (Amersham, Arlington Heights, IL). Затем клетки собирали с помощью харвестера (приспособления для сбора клеток) марки Фильтермейт 196 Харвестер (Packard, Downers Grove, IL) и измеряли степень включения тимидина с помощью сцинтилляционного счетчика для микропланшетов марки ТопКаунт (Packard).
Результаты, полученные в этих исследованиях, показали, что раствор ЗФР/ДМСО без пептидов и контрольный пептид 2705 (аминокислотная последовательность Arg-Glu-Asp-Leu-Arg-Thr-Leu-Leu-Arg-Tyr) не оказывали влияния на пролиферацию Т-клеток, а bс-пептиды ингибировали пролиферацию Т-клеток, при этом величина ингибирования составляла между 35 и 75%. Таким образом, эти данные показывают, что bс-пептиды проявляют заметную способность ингибировать пролиферацию Т-клеток.
ПРИМЕР 2. ВЛИЯНИЕ bс-ПЕПТИДОВ НА ЦИТОТОКСИЧЕСКУЮ АКТИВНОСТЬ Т-КЛЕТОК
Чтобы проанализировать влияние пептидов на цитотоксическую активность Т-клеток, получали эффекторные клетки СВА к В6, после 6 суток культивирования 4•106 клеток селезенки СВА с 5•106 обработанных митомицином клеток селезенки В6, в лунках 24-луночного планшета (Nuncion Delta, Nunc, Дания) в среде RPMI-1640 с 10% FBS. Затем эффекторные клетки собирали и промывали. В качестве клеток-мишеней использовали EL4(Н-2b), клетки лимфомы мышей, индуцированные в C57BL/6N. Культивируемые клетки EL4 поддерживали и пассировали каждые три дня (51Сr) в 20 мкл в течение 1 часа при 37oС. Затем эффекторные клетки (Е) и клетки-мишени (Т) помещали в V-образные культуральные чашки (Nunc, Дания) при соотношениях Е:Т, составляющих 3:1, 10:1, 30:1 и 100:1 соответственно. Пептиды разбавляли средой R-10 до рабочих концентраций и добавляли в начале 4-часового периода инкубации. Для определения максимального выхода в отдельные лунки добавляли 1% Тритон Х-100. Затем чашки центрифугировали в течение 2 минут, чтобы увеличить клеточный контакт перед 4-часовым периодом инкубации. После инкубации брали по 75 мкл надосадочной жидкости из каждой лунки и подсчитывали количество 51Сr с помощью сцинтилляционного счетчика для микропланшетов марки ТопКаунт. Степень лизиса клеток рассчитывали с помощью нижеприведенной формулы:
где СРМ - количество отсчетов в минуту.
Результаты, полученные из этих анализов, показали, что контрольный пептид 2705 при концентрациях вплоть до 100 мкг/мл не оказывал никакого влияния на опосредованный Т-клетками лизис клеток-мишеней, в то время, как bс-пептиды ингибировали опосредованный клетками лизис, причем степень ингибирования зависела от дозы. Полумаксимальное ингибирование активности клеток ЦТЛ, вызванное bс-петидами, наблюдалось при концентрации около 0,5 мкг/мл.
ПРИМЕР 3. ВЛИЯНИЕ bс-ПЕПТИДОВ НА АЛЛОГЕННЫЕ ТРАНСПЛАНТАТЫ in vivo
Иммуносупрессивную активность bс-пептидов оценивали на модели васкуляризованного полностью несовместимого аллотрансплантата сердца мыши. Конкретно, абдоминальную гетеротопическую пересадку сердца осуществляли, как было ранее описано в работе Ono and Lindsey, J. Thoracic Cardiovasc. Surg., 7:225 (1969). Мышам линии СВА, являвшимся реципиентами сердец С57В1/6, ежедневно вводили различные дозы пептида, после пересадки органа. Пептиды растворяли в ДМСО и разбавляли в ЗФР (окончательная концентрация ДМСО составляла 10%) перед внутрибрюшинным введением. Животным вводили пептиды, начиная со дня трансплантации и до пятого или девятого дня. Приживание трансплантата отслеживали ежедневно путем прямой пальпации и отторжение определяли по прекращению пальпируемых сокращений сердца. Статистическую значимость продолжительности выживания сердечного аллотрансплантата рассчитывали с помощью теста Манна-Уитни.
Результаты этих анализов показали, что контрольный пептид 2702.75-84 (аминокислотная последовательность Аrg-Glu-Asn-Leu-Arg-Lle-Ala-Leu-Arg-Tyr), введенный в дозе 80 мг/кг•сутки (сутки 0-9), способствовал продлению выживания сердечного аллотрансплантата до 10,7±2,6 суток, по сравнению с 8±1,4 суток у контрольных животных, которые получали ЗФР/ДМСО (р<0,01). Введение контрольного пептида 2702.75-84 в дозе 40 мг/кг•сутки не оказывало заметного эффекта на выживание трансплантата по сравнению с контрольным вариантом. В противоположность этому, однако, введение bс-пептидов в такой низкой дозе, как 1 мг/кг•сутки, привело к значительному увеличению продолжительности выживания сердечного аллотрансплантата, причем 50% трансплантатов оставались живыми в течение более 28 суток. Таким образом, эти результаты показывают, что bс-пептиды обладают иммуносупрессивной активностью, достаточной для увеличения продолжительности выживания трансплантатов у млекопитающих.
ПРИМЕР 4. СПОСОБНОСТЬ bс-ПЕПТИДОВ СВЯЗЫВАТЬСЯ С hsc70
Для того чтобы определить, способны ли bс-пептиды связываться с hsc70, выполняли анализы на связывание белков. Конкретно, синтезировали пептид 2702.75-84 в биотинилированной форме, с присоединенным к N-концу биотином, через состоящий из шести атомов углерода спейсер. На поверхность планшетов для ИФА (твердофазного иммуноферментного анализа) (Nunc Maxisorb, Nunc, США) наносили 100 нг/мл рекомбинантного hsc70 (Stressgen, Victoria, Canada) в 100 мМ Na-цитратного буфера с рН 4,0 и оставляли до следующего дня при температуре 4oС. После этого оставшиеся сайты связывания блокировали путем инкубации чашек с ЗФР/0,1% Твин 20 (PBS/Tween, Sigma) в течение 2 часов при комнатной температуре. Несвязанный материал удаляли путем троекратной промывки чашек смесью ЗФР/Твин. После добавки биотинилированного 2702.75-84 в смеси ЗФР/Твин/1%ДМСО чашки инкубировали в течение 2 часов при комнатной температуре, трижды промывали, затем инкубировали с 0,1 мкг/мл стрептавидина, конъюгированного с пероксидазой хрена (стрептавидин-HRP) (Jackson ImmunoResearch Laboratories, West Grove, PA) и снова промывали. Связанный стрептавидин-HRP обнаруживали с помощью 3 мг/мл о-фенилендиамина (OPD, Sigma) в субстратном буфере (SangStat, Menlo Park, CA). Реакцию останавливали добавлением 1М НСl и замеряли поглощение (оптическая плотность ОП490-ОП605), с помощью аппарата для прочтения планшетов ИФА. Связывание немеченого пептида замеряли в конкурентном анализе. Биотинилированный 2702.75-84 (3 мкМ) смешивали с различными количествами немеченого bс-пептида и затем инкубировали в планшетах с нанесенным на них hsc70 в течение 3 часов при комнатной температуре. Затем определяли связанный биотинилированный 2702.75-84, как описано выше.
Результаты этих экспериментов показали, что аффинная хроматография с использованием пептида 2702.84-75-75-84 (инвертированного димера) привела к выделению hsc70 и hsp70 (хитшокового белка). После инкубации планшетов для ИФА, на поверхность которых был нанесен hsc70 с биотинилированным пептидом 2702.75-84, авторы настоящего изобретения наблюдали зависящее от дозы связывание этого пептида с hsc70. Связывание биотинилированного пептида 2702.75-84 с hsc70 может ингибироваться добавкой увеличивающихся концентраций пептида 2702.75-84. Полумаксимальное ингибирование (ИК50) наблюдалось при 7,0±3,0 мкМ, подобное ИК50 наблюдалось для bс-пептидов (ИК50=2,5-10 мкМ), что свидетельствует о том, что bс-пептиды эффективно связываются с hsc70.
ПРИМЕР 5. ВЛИЯНИЕ bс-ПЕПТИДОВ НА ГЕМОКСИГЕНАЗУ И ДРУГИЕ ГЕМСОДЕРЖАЩИЕ ФЕРМЕНТЫ
Влияние bс-пептидов на hsc32 (гемоксигеназный) оценивали путем измерения активности гемоксигеназы (ГО) в присутствии и в отсутствие bс-пептидов. Конкретно, образцы мышиной селезенки гомогенизировали на льду в лизирующем буфере Трис-С1 (рН 7,4), содержащем 0,5% Тритона Х-100 и ингибиторы протеазы. Образцы замораживали небольшими аликвотами и так хранили до использования. Гомогенаты селезенки использовали в качестве источника ГО для всех измерений активности. Полученную из печени крыс биливердинредуктазу очищали способом, описанным в публикации Kutty and Maines, J. Biol. Chem., 256:3956 (1981). Активность ГО измеряли путем смешивания 100 мкл гомогената селезенки с 0,8 мкМ NADPH, 0,8 мМ глюкоза-6-фосфата, 1,0 единицы G-6-Р-дегидро-геназы, 1 мМ МgС12 и 10 мкл биливердинредуктазы при 4oС. Реакцию инициировали добавлением гемина (20 мкл 2,5 мМ раствора). Реакционную смесь инкубировали при 37oС в темноте в течение 30 минут. В конце инкубационного периода все нерастворимые материалы подвергали центрифугированию и анализировали надосадочную жидкость на концентрацию билирубина с помощью модифицированного метода Хилмана и Бейера (Hillman and Beyer, Z. Klin. Chem., 5:92 (1981) (с помощью диагностического набора 552 компании Sigma). Контрольные варианты включали образцы селезенки в отсутствие NADPH-генерирующей системы и все компоненты реакционной смеси в отсутствие гомогенатов селезенки.
Результаты этих экспериментов показали, что по сравнению с неактивным контрольным пептидом 2705.75-84, bc-пептиды (100 мкг/мл) ингибировали активность ГО более, чем на 50%. Таким образом выяснилось, что, по-видимому, bс-пептиды способны ингибировать активность гемоксигеназы.
После того, как было установлено, что bс-пептиды эффективно ингибируют активность гомоксигеназы, определяли действие bс-пептидов на другие гемсодержащие ферменты, такие, как синтаза окиси азота (NOS). Конкретно, bс-пептиды и контрольный пептид 2702 исследовали в анализах активности ферментов на их способность ингибировать три различных изоформы NOS (нейронную NOS, эндотелиальную NOS и индуцируемую цитокином NOS) in vitro. Результаты этих экспериментов показали, что bс-пептиды способны ингибировать NOS со значительно меньшим показателем ИК50, чем показатель контрольного пептида 2702. Таким образом выяснилось, что, повидимому, bс-пептиды имитируют порфириноподобные структуры, которые особенно влияют на активность гемсодержащих ферментов, и это было подтверждено с помощью компьютерного моделирования.
ПРИМЕР 6. ОПОСРЕДОВАННОЕ bс-ПЕПТИДАМИ ИНГИБИРОВАНИЕ СИНТЕЗА ВОСПАЛИТЕЛЬНЫХ ЦИТОКИНОВ
Для того чтобы определить влияние bс-пептидов на синтез воспалительных цитокинов, на клетки макрофага RAW264.7 в культуре с 10 мкг/мл бактериального липополисахарида (ЛПС) воздействовали таким образом, чтобы стимулировать продуцирование ими воспалительного цитокина фактора-α некроза опухолевых клеток (TNF-α) (см. Alleva et al., J. Immunol., 153:1674 (1994) и Tonetti et al., Biochem. Biophys. Res. Comm., 230:636-640 (1997)), либо в отсутствие, либо в присутствии (от 1 до 100 мкМ) испытуемого пептида. После 24-часовой инкубации определяли количество TNF-α в культуральных надосадочных жидкостях с помощью ИФА. Без добавки ЛПС клетки RAW264.7 не продуцировали обнаруживаемых количеств TNF-α.
Результаты, полученные в этих экспериментах, показали, что в то время как контрольный пептид D2RP (аминокислотная последовательность Arg-Val-Asn-Leu-Pro-Lle-Ala-Leu-Arg-Tyr) не проявил способности ингибировать продуцирование TNF-α клетками макрофага RAW264.7, bс-пептиды ингибировали продуцирование TNF-α, причем степень ингибирования зависела от дозы. Таким образом, bс-пептиды найдут применение в подавлении синтеза воспалительных цитокинов, тем самым являясь ценным средством для лечения воспалительных и связанных с воспалениями заболеваний.
ПРИМЕР 7. ДЕЙСТВИЕ bс-ПЕПТИДОВ ПРИ ПРИМЕНЕНИИ ИХ НА МОДЕЛИ СЕПТИЧЕСКОГО ШОКА НА ЖИВОТНЫХ
Введение ЛПС мышам представляет собой приемлемую модель септического шока на животных (см. Otterbein et al., Amer. J. Physiol., 272 (2):1 (1997), Albrecht et al., Hepatology 26:1553-1559 (1997), Haziot et al., J. Immunol. 154: 6529-6532 (1995) и Otterbein et al., Amer. J. Respir. Cell Mol. Biol., 13: 595-601 (1995)). Для того чтобы получить еще больше доказательств полезности bс-пептидов для лечения воспалительных реакций и воспалительных состояний (таких, как септический шок), авторы испытывали влияние bс-пептидов на выживание мышей после введения им ЛПС. Конкретно, мышам вводили 20 мг/кг контрольного пептида 2705 или bс-пептида в маннитоле или только маннитол. Через 16 часов после введения пептида в маннитоле или только маннитола мышам вводили с помощью инъекции 100 мг/кг ЛПС и два раза в день отслеживали выживание мышей.
Результаты этих экспериментов показали, что в то время как все мыши, которым вводили контрольный пептид 2705 в маннитоле или только маннитол, погибли на следующий день после введения ЛПС, более 50% мышей, получавших bс-пептид, были живы на второй день после введения ЛПС, и более 25% мышей, получавших bс-пептид, были живы на третий день после введения ЛПС. Таким образом, эти данные показывают, что bс-петиды эффективны для лечения воспалительных состояний, таких, как септический шок.
ПРИМЕР 8. ВВЕДЕНИЕ bс-ПЕПТИДОВ ПОСРЕДСТВОМ ПЕРЕНОСА ГЕНОВ in vivo ПОВЫШАЕТ ВЫЖИВАЕМОСТЬ МЫШИНЫХ ГЕТЕРОТОПИЧЕСКИХ СЕРДЕЧНЫХ ТРАНСПЛАНТАТОВ
Далее авторы задались целью определить, может ли локальная доставка bс-пептидов путем опосредованного плазмидами переноса генов повысить выживаемость трансплантатов in vivo на мышиной модели гетеротопического сердечного трансплантата. Конкретно, неонатальные сердца донора C57BL/6 подкожно трансплантировали в наружное ухо мышей-реципиентов линии CBA/J. Во время трансплантации непосредственно в аллотрансплантат инъецировали bс-пептид или 20 мкг плазмидной ДНК, кодирующей представляющий интерес пептид. Выживание аллотрансплантатов определяли путем отслеживания электрокардиограмм и отторжение определяли на основе прекращения электрической активности сердца. Выживание трансплантатов выражали в сутках (среднее ± стандартная ошибка). Статистическую значимость определяли с помощью непарного t-теста Стьюдента.
Прямая инъекция 1 мкг контрольного пептида 2702 в аллотрансплантат не продлила срока его выживания (13,3±0,75, против 13,9±0,9 в контроле без инъекции пептида), а инъекция 400 мкг пептида 2702 продлила срок выживания (22,0±0,58). Инъекция 20 мкг плазмидной ДНК, кодирующей контрольный пептид 2702, еще более продлила срок выживания трансплантата - до 30,3±1,03. Подобные же результаты получили и для другой плазмиды, кодирующей пептид bcl (29,0±4,08), в то время как не наблюдалось значимого увеличения срока выживания при использовании плазмиды, кодирующей контрольный пептид 2705, не обладающий иммуномодуляторной активностью in vitro или in vivo (16,5±0,96).
ПРИМЕР 9. ВЛИЯНИЕ bс-ПЕПТИДОВ НА ЗАДЕРЖКУ НАСТУПЛЕНИЯ ИНСУЛИНЗАВИСИМОГО САХАРНОГО ДИАБЕТА (ИЗСД)
Поскольку в настоящем описании показано, что пептиды по настоящему изобретению являются иммуномодулирующими, авторы изобретения далее поставили цель определить, являются ли bс-пептиды эффективными для подавления наступления аутоиммунных болезней in vivo. В качестве модели аутоиммунного заболевания вообще авторы определяли способность bс-пептидов задерживать или подавлять наступление ИЗСД in vivo. Конкретно, 20 мг/кг bс-пептида вводили внутрибрюшинно 6-недельным самкам мышей, не страдающих диабетом (НСД), в то время как контрольным мышам либо ничего не вводили, либо вводили неактивное пептидное соединение. Вышеуказанные варианты повторяли каждую неделю. У подопытных животных один раз в неделю определяли содержание глюкозы в крови. Наступление ИЗСД у подопытных животных определяли по уровню глюкозы в крови, превышающему 200 мг/дл.
Результаты вышеописанных экспериментов показали, что у 70 - 80% контрольных НСД мышей, не получавших пептид, ИЗСД развился к возрасту 22 недель. Никакой разницы не наблюдалось между группами контрольных животных, которые либо не получали пептид, либо получали неактивный контрольный пептид. Однако для животных, которым вводили bс-пептиды, только у одного животного развился ИЗСД к возрасту 16 недель, а у всех остальных подопытных животных ИЗСД не развился к возрасту 24 недель. Таким образом, эти результаты показывают, что введение bс-пептидов эффективно для задержки наступления ИЗСД in vivo.
Из вышеприведенных результатов очевидно, что композиции и методы по настоящему изобретению обеспечивают существенное ингибирование цитотоксичности ЦТЛ. При этом выяснилось, что пептиды по настоящему изобретению обеспечивают существенно повышенную эффективность ингибирования лизиса по сравнению с ранее применявшимися олигопептидами. Использование композиций по настоящему изобретению обеспечивает существенные преимущества, заключающиеся в том, что при этом требуются меньшие количества олигопептидов для достижения терапевтического уровня, а также в подавлении гемоксигеназной активности.
Все публикации и заявки на патенты, упоминаемые в настоящем описании, включены в него в качестве ссылок в такой же самой степени, как если бы для каждой отдельной публикации или заявки на патент было бы конкретно и индивидуально указано, что они включены в качестве ссылки.
Теперь, когда изобретение полностью описано, специалистам обычной квалификации в данной области техники очевидно, что можно осуществить много изменений и модификаций данного изобретения, которые не выходят за пределы существа или объема прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СЛИТЫЙ БЕЛОК PD1-4-1BBL И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2018 |
|
RU2769513C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ НА CD127 | 2015 |
|
RU2734076C2 |
| Терапевтические средства | 2016 |
|
RU2747733C1 |
| СЛИТЫЙ БЕЛОК SIRPα-4-1BBL И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2018 |
|
RU2769769C2 |
| ПЕПТИД, ПОЛУЧЕННЫЙ ИЗ DEPDC1, И СОДЕРЖАЩАЯ ЕГО ВАКЦИНА | 2016 |
|
RU2765574C2 |
| АНТИТЕЛА И ПОЛИПЕПТИДЫ, НАПРАВЛЕННЫЕ ПРОТИВ CD127 | 2017 |
|
RU2769352C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ДИАГНОСТИКИ И ЛЕЧЕНИЯ ОПУХОЛИ | 2010 |
|
RU2563359C2 |
| АНТИТЕЛО, НАЦЕЛЕННОЕ НА ВСМА, И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2799655C2 |
| СЛИТАЯ МОЛЕКУЛА, СПОСОБНАЯ ИНДУЦИРОВАТЬ НЕВОСПАЛИТЕЛЬНЫЙ ФАГОЦИТОЗ | 2022 |
|
RU2823919C1 |
| ИММУНОМОДУЛИРУЮЩИЕ ОЛИГОПЕПТИДЫ | 2007 |
|
RU2503684C2 |

Изобретение относится к олигопептиднам, включающим аминокислотную последовательность B-X-X-X-B-X-X-X-J-Tyr, в которой В - Lys или Arg; X представляет собой любую аминокислоту, иную, чем заряженная алифатическая аминокислота, или ее D-изомер; J - Gly, Lys или Arg; указанные аминокислоты представляют собой существующие в природе L-изомеры, их D-изомеры и норлейцин, и где указанный олигопептид включает иные, чем (а) встречающаяся в природе последовательность α-домена антигена-В лейкоцита человека (HLA-B) 75-84, (б) встречающаяся в природе последовательность трансмембранной последовательности α-цепи Т-клеточного рецептора человека и (в) последовательность, являющаяся мутантной по отношению либо к (а), либо к (б), имеющая не более двух мутаций в качестве активной части указанного олигопептида; способу подавления активации лимфоцитных клеток, способу трансплантации донорных органов или клеток млекопитающих, способу ингибирования синтеза белка воспалительного цитокина клетками, способными продуцировать указанный белок воспалительного цитокина, способу подавления воспалительной реакции у млекопитающих, способу модуляции активности гемсодержащего фермента и способу задержки наступления аутоиммунной болезни у млекопитающего. 7 с. и 29 з.п. ф-лы, 3 табл., 2 ил.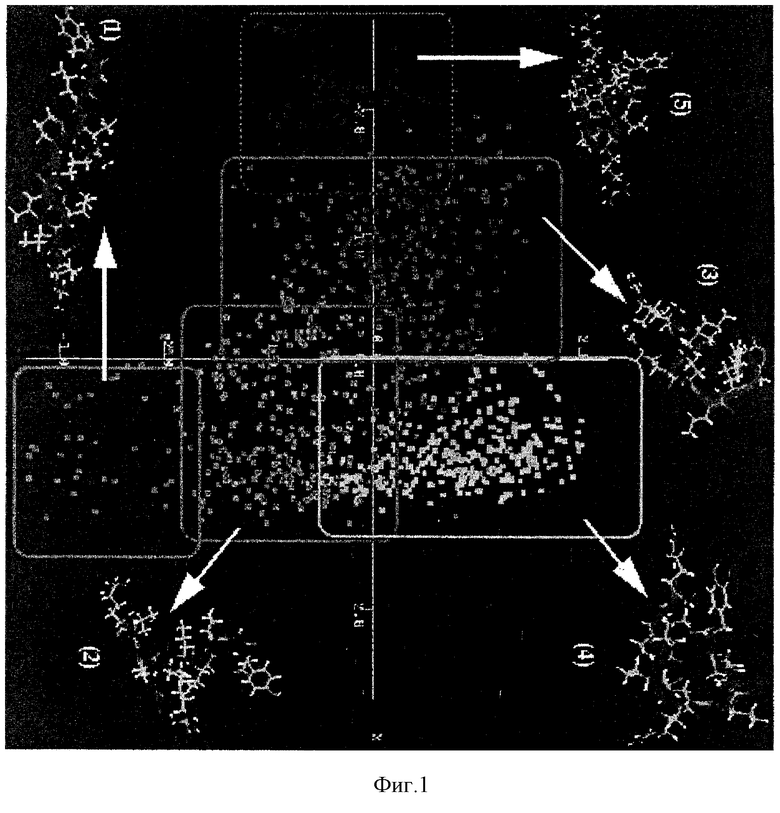
B-X-X-X-B-X-X-X-J-Tyr,
в которой В - Lys или Arg;
X представляет собой любую аминокислоту, иную, чем заряженная алифатическая аминокислота, или ее D-изомер;
J - Gly, Lys или Arg, где указанные аминокислоты представляют собой существующие в природе L-изомеры, их D-изомеры и норлейцин и где указанный олигопептид включает иные, чем (а) встречающаяся в природе последовательность α1- домена антигена-В лейкоцита человека (HLA-B) 75-84, (б) встречающаяся в природе последовательность трансмембранной последовательности α- цепи Т-клеточного рецептора человека и (в) последовательность, являющаяся мутантной по отношению либо к (а), либо к (б), имеющая не более двух мутаций в качестве активной части указанного олигопептида.
(a) Arg-Leu-Leu-Leu-Arg-Leu-Leu-Leu-Gly-Tyr;
(b) Arg-Val-Leu-Leu-Arg-Leu-Leu-Leu-Gly-Tyr;
(c) Arg-Ile-Leu-Leu-Arg-Leu-Leu-Leu-Gly-Tyr;
(d) Arg-Leu-Val-Leu-Arg-Leu-Leu-Leu-Gly-Tyr;
(e) Arg-Leu-Ile-Leu-Arg-Leu-Leu-Leu-Gly-Tyr;
(f) Arg-Leu-Leu-Val-Arg-Leu-Leu-Leu-Gly-Tyr;
(g) Arg-Leu-Leu-Ile-Arg-Leu-Leu-Leu-Gly-Tyr;
(h) Arg-Leu-Leu-Leu-Arg-Val-Leu-Leu-Gly-Tyr;
(i) Arg-Leu-Leu-Leu-Arg-Ile-Leu-Leu-Gly-Tyr;
(j) Arg-Leu-Leu-Leu-Arg-Leu-Val-Leu-Gly-Tyr;
(k) Arg-Leu-Leu-Leu-Arg-Leu-Ile-Leu-Gly-Tyr;
(l) Arg-Leu-Leu-Leu-Arg-Leu-Leu-Val-GIy-Tyr;
(m) Arg-Leu-Leu-Leu-Arg-Leu-Leu-Ile-Gly-Tyr;
(n) Arg-Trp-Leu-Leu-Arg-Leu-Leu-Leu-Gly-Tyr;
(о) Arg-Leu-Trp-Leu-Arg-Leu-Leu-Leu-GIy-Tyr;
(p) Arg-Leu-Leu-Trp-Arg-Leu-Leu-Leu-Gly-Tyr;
(q) Arg-Leu-Leu-Leu-Arg-Trp-Leu-Leu-Gly-Tyr;
(r) Arg-Leu-Leu-Leu-Arg-Leu-Trp-Leu-Gly-Tyr;
(s) Arg-Leu-Leu-Leu-Arg-Leu-Leu-Trp-Gly-Tyr;
(t) Arg-Tyr-Leu-Leu-Arg-Leu-Leu-Leu-Gly-Tyr;
(u) Arg-Leu-Tyr-Leu-Arg-Leu-Leu-Leu-Gly-Tyr;
(v) Arg-Leu-Leu-Tyr-Arg-Leu-Leu-Leu-Gly-Tyr;
(w) Arg-Leu-Leu-Leu-Arg-Tyr-Leu-Leu-Gly-Tyr;
(x) Arg-Leu-Leu-Leu-Arg-Leu-Tyr-Leu-Gly-Tyr;
(y) Arg-Leu-Leu-Leu-Arg-Leu-Leu-Tyr-Gly-Tyr;
(z) Arg-Nle-Nle-Nle-Arg-Nle-Nle-Nle-Gly-Tyr.
23. Способ по п.20, в котором указанная стадия соединения включает осуществление контакта указанных клеток с молекулой нуклеиновой кислоты, кодирующей указанный олигопептид; причем указанный олигопептид экспрессируется указанной молекулой нуклеиновой кислоты.
| RU 95122726 A, 27.02.1998 | |||
| RU 93058563 A, 27.12.1996 | |||
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |

