Область техники
Настоящее изобретение относится к области иммунотерапии или диагностики опухолей, и, в частности, настоящее изобретение относится к антителу, нацеленному на ВСМА, и вариантам его применения.
Уровень техники
Множественная миелома (ММ) - распространенное гематологическое злокачественное заболевание, на долю которого приходится 2% всех смертельных исходов от рака. ММ - это гетерогенное заболевание, в основном обусловленное хромосомной транслокацией t(11; 14), t(4; 14), t(8; 14), del(13), del(17) (в числе прочего) (Drach et al., (1998) Blood 92(3): 802-809; Gertz et al., (2005) Blood 106 (8): 2837-2840; Facon et al., (2001) Blood 97 (6): 1566-1571). Основное состояние при множественной миеломе (ММ) - неконтролируемое размножение и увеличение количества плазматических клеток в костном мозге, что приводит к остеонекрозу. Пациенты с ММ могут испытывать различные симптомы заболевания, обусловленные инфильтрацией костного мозга, разрушением кости, почечной недостаточностью, иммунодефицитом и психологической нагрузкой из-за диагностированного рака. В настоящее время основными способами лечения данного заболевания являются химиотерапия и трансплантация стволовых клеток. Основными применяемыми химиотерапевтическими лекарственными средствами являются стероиды, талидомид, леналидомид, бортезомиб или комбинация различных цитотоксических агентов. У молодых пациентов можно использовать химиотерапию в высоких дозах в комбинации с трансплантацией аутологичных стволовых клеток.
ВСМА (антиген созревания В-клеток) представляет собой антиген созревания В-клеток, трансмембранный белок III типа, состоящий из 185 аминокислотных остатков, и принадлежит к надсемейству рецепторов ФНО. Лиганд ВСМА принадлежит к надсемейству ФНО, например, лиганд, индуцирующий пролиферацию (APRIL), фактор стимуляции В-лимфоцитов (BAFF). После связывания со своим лигандом ВСМА активизирует пролиферацию и выживание В-клеток. ВСМА специфически и в высокой степени экспрессируется на поверхности плазматических клеток и клеток множественной миеломы, но не экспрессируется на кроветворных стволовых клетках и других клетках здоровых тканей, поэтому ВСМА может являться идеальной мишенью для направленной терапии ММ.
В заключение можно сказать, что в данной области техники существует срочная потребность в получении антител, специфичных по отношению к ВСМА, и эффекторных иммунных клеток, нацеленных на ВСМА.
Сущность изобретения
Целью настоящего изобретения является обеспечение антител, специфичных по отношению к ВСМА и эффекторных иммунных клеток, нацеленных на ВСМА.
В первом аспекте настоящего изобретения предложено антитело, нацеленное на ВСМА, причем указанное антитело выбрано из группы, состоящей из:
(1) антитела, содержащего вариабельную область тяжелой цепи, содержащую HCDR1, представленную в SEQ ID NO: 1, 60 или 62, и/или содержащую HCDR2, представленную в SEQ ID NO: 2, 61 или 63, и/или HCDR3, представленную в любой из последовательностей SEQ ID NO: 3, SEQ ID NO: 4 или SEQ ID NO: 5;
(2) антитела, содержащего вариабельную область легкой цепи, содержащую LCDR1, представленную в SEQ ID NO: 6, и/или содержащую LCDR2, представленную в SEQ ID NO: 7, и/или содержащую LCDR3, представленную в любой из последовательностей SEQ ID NO: 8, SEQ ID NO: 9 или SEQ ID NO: 10;
(3) антитела, содержащего вариабельную область тяжелой цепи антитела согласно подпункту (1) и вариабельную область легкой цепи антитела согласно подпункту (2);
(4) антитела, являющегося вариантом антитела согласно любому из подпунктов (1) - (3) и обладающего такой же или аналогичной активностью, как и антитело согласно любому из подпунктов (1) - (3).
В конкретном варианте реализации антитело выбрано из группы, состоящей из:
(1) антитела, содержащего HCDR1, представленную в SEQ ID NO: 1, HCDR2, представленную в SEQ ID NO: 2, HCDR3, представленную в SEQ ID NO: 3, и LCDR1, представленную в SEQ ID NO: 6, LCDR2, представленную в SEQ ID NO: 7, и LCDR3, представленную в SEQ ID NO: 8;
(2) антитела, содержащего HCDR1, представленную в SEQ ID NO: 1, HCDR2, представленную в SEQ ID NO: 2, HCDR3, представленную в SEQ ID NO: 4, LCDR1, представленную в SEQ ID NO: 6, LCDR2, представленную в SEQ ID NO: 7, и LCDR3, представленную в SEQ ID NO: 9;
(3) антитела, содержащего HCDR1, представленную в SEQ ID NO: 1, HCDR2, представленную в SEQ ID NO: 2, HCDR3, представленную в SEQ ID NO: 5, LCDR1, представленную в SEQ ID NO: 6, LCDR2, представленную в SEQ ID NO: 7, и LCDR3, представленную в SEQ ID NO: 10;
(4) антитела, содержащего HCDR1, представленную в SEQ ID NO: 60, HCDR2, представленную в SEQ ID NO: 61, HCDR3, представленную в SEQ ID NO: 5, LCDR1, представленную в SEQ ID NO: 6, LCDR2, представленную в SEQ ID NO: 7, и LCDR3, представленную в SEQ ID NO: 10;
(5) антитела, содержащего HCDR1, представленную в SEQ ID NO: 62, HCDR2, представленную в SEQ ID NO: 63, HCDR3, представленную в SEQ ID NO: 5, LCDR1, представленную в SEQ ID NO: 6, LCDR2, представленную в SEQ ID NO: 7, и LCDR3, представленную в SEQ ID NO: 10;
(6) антитела, являющегося вариантом антитела согласно любому из подпунктов (1) - (5) и обладающего такой же или аналогичной активностью, как и антитело согласно любому из подпунктов (1) - (5).
В конкретном варианте реализации антитело выбрано из группы, состоящей из:
(1) антитела, где вариабельная область тяжелой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 13, аминокислотную последовательность SEQ ID NO: 17, аминокислотную последовательность SEQ ID NO: 21 или аминокислотную последовательность SEQ ID NO: 56 или аминокислотную последовательность SEQ ID NO: 58;
(2) антитела, где вариабельная область легкой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 11, аминокислотную последовательность SEQ ID NO: 15 или аминокислотную последовательность SEQ ID NO: 19;
(3) антитела, содержащего вариабельную область тяжелой цепи антитела согласно подпункту (1) и вариабельную область легкой цепи антитела согласно подпункту (2);
(4) антитела, являющегося вариантом антитела согласно любому из подпунктов (1) - (3) и обладающего такой же или аналогичной активностью, как и антитело согласно любому из подпунктов (1) - (3).
В конкретном варианте реализации антитело выбрано из группы, состоящей из:
(1) антитела, где вариабельная область тяжелой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 13, а вариабельная область легкой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 11;
(2) антитела, где вариабельная область тяжелой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 17, а вариабельная область легкой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 15;
(3) антитела, где вариабельная область тяжелой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 21, а вариабельная область легкой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 19;
(4) антитела, где вариабельная область тяжелой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 56, а вариабельная область легкой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 19;
(5) антитела, где вариабельная область тяжелой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 58, а вариабельная область легкой цепи указанного антитела содержит аминокислотную последовательность SEQ ID NO: 19;
(6) антитела, являющегося вариантом антитела согласно любому из подпунктов (1) - (5) и обладающего такой же или аналогичной активностью, как и антитело согласно любому из подпунктов (1) - (5).
Во втором аспекте настоящего изобретения предложено антитело, распознающее ту же антигенную детерминанту, что и антитело согласно первому аспекту настоящего изобретения.
В третьем аспекте настоящего изобретения предложена нуклеиновая кислота, кодирующая антитело согласно первому или второму аспекту настоящего изобретения.
В четвертом аспекте настоящего изобретения предложен экспрессирующий вектор, содержащий нуклеиновую кислоту согласно третьему аспекту настоящего изобретения.
В пятом аспекте настоящего изобретения предложена клетка-хозяин, содержащая экспрессирующий вектор согласно четвертому аспекту настоящего изобретения или нуклеиновую кислоту согласно третьему аспекту настоящего изобретения, встроенную в геном.
В шестом аспекте настоящего изобретения предложено применение антитела согласно первому или второму аспекту настоящего изобретения для получения направленного лекарственного средства, конъюгата «антитело-лекарственное средство» или многофункционального антитела, специфично нацеленное на опухолевые клетки, экспрессирующие ВСМА; или
для получения агента для диагностики опухоли, экспрессирующей ВСМА; или
для получения иммунной клетки, модифицированной химерным рецептором антигена, причем, предпочтительно, указанная иммунная клетка включает Т-лимфоцит, NK-клетку или NKT-лимфоцит.
В седьмом аспекте настоящего изобретения предложен иммуноконъюгат, содержащий:
антитело согласно первому или второму аспекту настоящего изобретения; и
функциональную молекулу, связанную с ним; причем указанную функциональную молекулу выбирают из группы, состоящей из молекулы, нацеленной на поверхностный маркер опухоли, молекулы, ингибирующей опухоли, молекулы, нацеленной на поверхностный маркер иммунной клетки, и детектируемой метки.
В конкретном варианте реализации молекула, ингибирующая опухоли, представляет собой противоопухолевый цитокин или противоопухолевый токсин. Цитокин предпочтительно включает: ИЛ-12, ИЛ-15, интерферон I типа, ФНО-альфа.
В конкретном варианте реализации молекула, нацеленная на поверхностный маркер иммунной клетки, представляет собой антитело или лиганд, связывающиеся с поверхностным маркером иммунной клетки и, предпочтительно, поверхностный маркер иммунной клетки включает: CD3, CD16, CD28, и, более предпочтительно, антитело, связывающее поверхностный маркер иммунной клетки, представляет собой антитело против CD3.
В конкретном варианте реализации молекула, нацеленная на поверхностный маркер иммунной клетки, представляет собой антитело, связывающееся с поверхностным маркером Т-клетки.
В восьмом аспекте настоящего изобретения предложена нуклеиновая кислота, кодирующая многофункциональный иммуноконъюгат согласно седьмому аспекту настоящего изобретения.
В девятом аспекте настоящего изобретения предложено применение многофункционального иммуноконъюгата согласно седьмому аспекту настоящего изобретения для получения противоопухолевого лекарственного средства или
для получения агента для диагностики опухоли, экспрессирующей ВСМА; или
для получения иммунной клетки, модифицированной химерным рецептором антигена, причем указанная иммунная клетка предпочтительно включает: Т-лимфоциты, NK-клетки или NKT-лимфоциты.
В десятом аспекте настоящего изобретения предложен химерный рецептор антигена, содержащий внеклеточный домен, трансмембранный домен и внутриклеточный сигнальный домен, причем внеклеточный домен содержит антитело согласно первому или второму аспекту настоящего изобретения, и указанное антитело предпочтительно представляет собой одноцепочечное антитело или доменное антитело.
В конкретном варианте реализации внутриклеточный домен содержит один или более из костимулирующих сигнальных доменов и/или первичных сигнальных доменов.
В конкретном варианте реализации химерный рецептор антигена дополнительно содержит шарнирный домен.
В конкретном варианте реализации трансмембранный домен выбран из группы, состоящей из альфа-, бета-, дзета-цепи TCR, трансмембранных областей CD3ε, CD3ζ, CD4, CD5, CD8α, CD9, CD16, CD22, CD27, CD28, CD33, CD37, CD45, CD64, CD80, CD86, CD134, CD137, CD152, CD154 и PD1; и/или
костимулирующий сигнальный домен выбран из группы, состоящей из внутриклеточных сигнальных областей CARD11, CD2, CD7, CD27, CD28, CD30, CD40, CD54, CD83, ОХ40, CD137, CD134, CD150, CD152, CD223, CD270, PD-L2, PD-L1, CD278, DAP10, LAT, NKD2C SLP76, TRIM, FcεRIγ, MyD88 и 41BBL; и/или
первичный сигнальный домен выбран из группы, состоящей из TCR ξ, FcR γ, FcR β, CD3 γ, CD3δ, CD3ε, CD5, CD22, CD79a, CD79b, CD278 (также называемого «ICOS»), CD66d и CD3ζ,
предпочтительно,
трансмембранный домен выбран из группы, состоящей из трансмембранных доменов CD8α, CD4, CD45, PD1, CD154 и CD28; и/или
костимулирующий сигнальный домен выбран из группы, состоящей из CD137, CD134, CD28 и ОХ40; и/или
первичный сигнальный домен выбран из CD3Cζ,
наиболее предпочтительно, трансмембранный домен выбран из CD8α или CD28, костимулирующий сигнальный домен выбран из внутриклеточного сигнального домена CD137 или CD28, а первичный сигнальный домен выбран из CD3ζ.
В конкретном варианте реализации химерный рецептор антигена содержит следующие последовательно связанные антитело, трансмембранную область и внутриклеточную сигнальную область:
антитело согласно первому или второму аспекту настоящего изобретения, трансмембранную область CD8 и CD3ζ;
антитело согласно первому или второму аспекту настоящего изобретения, трансмембранную область CD8, внутриклеточную сигнальную область CD137 и CD3ζ;
антитело согласно первому или второму аспекту настоящего изобретения, трансмембранную область CD28, внутриклеточную сигнальную область CD28 и CD3ζ; или
антитело согласно первому или второму аспекту настоящего изобретения, трансмембранную область CD28, внутриклеточную сигнальную область CD28, CD137 и CD3ζ.
В одиннадцатом аспекте настоящего изобретения предложена нуклеиновая кислота, кодирующая химерный рецептор антигена согласно десятому аспекту настоящего изобретения.
В двенадцатом аспекте настоящего изобретения предложен экспрессирующий вектор, содержащий нуклеиновую кислоту согласно одиннадцатому аспекту настоящего изобретения.
В тринадцатом аспекте настоящего изобретения предложен вирус, содержащий вектор согласно двенадцатому аспекту настоящего изобретения.
В предпочтительном варианте реализации указанный вирус является лентивирусом.
В четырнадцатом аспекте настоящего изобретения предложено применение химерного рецептора антигена согласно десятому аспекту настоящего изобретения или нуклеиновой кислоты согласно одиннадцатому аспекту настоящего изобретения или экспрессирующего вектора согласно двенадцатому аспекту настоящего изобретения или вируса согласно тринадцатому аспекту настоящего изобретения для получения генетически модифицированных иммунных клеток, нацеленных на опухолевую клетка, экспрессирующая ВСМА.
В предпочтительном варианте реализации опухоль, экспрессирующая ВСМА, представляет собой множественную миелому.
В пятнадцатом аспекте настоящего изобретения предложена генетически модифицированная иммунная клетка, трансдуцированная нуклеиновой кислотой согласно одиннадцатому аспекту настоящего изобретения или экспрессирующим вектором согласно двенадцатому аспекту настоящего изобретения или вирусом согласно тринадцатому аспекту настоящего изобретения, или экспрессирующая химерный рецептор антигена согласно десятому аспекту настоящего изобретения.
Иммунные клетки предпочтительно выбраны из Т-лимфоцитов, NK-клеток или NKT-клеток.
В конкретном варианте реализации генетически модифицированная иммунная клетка дополнительно экспрессирует последовательность, не являющуюся химерным рецептором антигена согласно десятому аспекту настоящего изобретения, и указанная другая последовательность содержит цитокин или еще один химерный рецептор антигена или рецептор хемокина или миРНК, снижающую экспрессию PD-1, или белок, блокирующий PD-L1, или TCR, или «предохранительный переключатель» (safety switch);
Цитокин предпочтительно включает ИЛ-12, ИЛ-15, ИЛ-21 или интерферон I типа;
Рецептор хемокина предпочтительно включает CCR2, CCR5, CXCR2 или CXCR4;
«Предохранительный переключатель» (safety switch) предпочтительно включает индуцибельную каспазу-9, укороченный EGFR или RQR8.
В шестнадцатом аспекте настоящего изобретения предложено применение генетически модифицированной иммунной клетки согласно пятнадцатому аспекту настоящего изобретения для получения лекарственного средства, подавляющего опухоль, причем указанная опухоль представляет собой опухоль, экспрессирующую ВСМА.
В предпочтительном варианте реализации опухоль, экспрессирующая ВСМА, представляет собой множественную миелому.
В семнадцатом аспекте настоящего изобретения предложена фармацевтическая композиция, содержащая:
антитело согласно первому или второму аспекту настоящего изобретения или нуклеиновую кислоту, кодирующую указанное антитело; или
иммуноконъюгат согласно седьмому аспекту настоящего изобретения или нуклеиновую кислоту, кодирующую указанный иммуноконъюгат; или
химерный рецептор антигена согласно десятому аспекту настоящего изобретения или нуклеиновую кислоту, кодирующую указанный химерный рецептор антигена; или
генетически модифицированную иммунную клетку согласно пятнадцатому аспекту настоящего изобретения.
Следует понимать, что различные вышеупомянутые технические признаки настоящего изобретения и различные технические признаки, конкретно описанные ниже (как в разделе «Примеры»), можно объединить друг с другом в рамках объема настоящего изобретения, получая новое или предпочтительное техническое решение, которое не будет последовательно воспроизведено в настоящем документе.
Описание чертежей
На фиг. 1 показаны ДСН-электрофореграммы ВСМА_huFc и ВСМА_muFc (восстановительные условия).
На фигуре 2 показана экспрессия ВСМА в стабильной линии клеток K562-ВСМА, обнаруженная посредством FACs.
На фигуре 3 показаны результаты твердофазного ИФА антител 7А12, 7G2 и 23F10.
На фигуре 4 показано связывание антител 7А12, 7G2 и 23F10 с K562-ВСМА и K562, обнаруженное посредством FACs.
На фигуре 5 показан анализ очищенного scFv_Fc-антитела против ВСМА посредством электрофореза в ДСН-ПААГ (восстановительные условия).
На фигуре 6 показано связывание разбавленного в градиенте очищенного scFv_Fc с K562-ВСМА, определяемое посредством FACs-анализа.
На фигуре 7 показано сродство антител 7А12, 7G2 и 23F10 к ВСМА, определяемое с помощью Biacore.
На фигуре 8 показано связывание антител 7А12, 7G2 и 23F10 с клетками линии RPMI8226, определяемое посредством FACs.
На фигуре 9 показано конкурентное связывание антител и APRIL с ВСМА, определяемое посредством твердофазного ИФА.
На фигуре 10 показан положительный показатель Т-лифоцитов, инфицированных BCMA-CAR Т-вирусом, обнаруженный посредством FACS.
На фигуре 11А показаны результаты анализа токсичности in vitro BCMA-CAR Т по отношению к ВСМА-экспрессирующим положительным и отрицательным клеткам, а на фигуре 11В показаны результаты экспериментов in vivo с BCMA-CAR Т у мышей.
На фигуре 12 показаны аминокислотные последовательности тяжелых цепей клонов 25С2, 25D2 и 23F10.
На фиг. 13А показана агрегация антитела 25С2, на фигуре 13В показана агрегация антитела 25D2, а на фигуре 13С показана агрегация антител 23F10 и 7А12.
На фигуре 14А показано связывание 25С2 и 25D2 с клетками K562-ВСМА и K562, а на фигуре 14В показана специфичность антител 23F10, 25С2 и 25D2, определяемая посредством твердофазного ИФА.
На фигуре 15 показаны результаты экспериментов по уничтожению клеток 25C2-BBZ, 25D2-BBZ, 7A12-BBZ, C11D5.3-BBZ и sPD-1-7A12-BBZ.
На фигуре 16 показаны результаты подкожной ксенотрансплантации 25C2-BBZ, 25D2-BBZ, C11D5.3-BBZ, 7A12-BBZ.
Способы выполнения изобретения
В результате обширных и интенсивных исследований авторы изобретения неожиданно обнаружили антитела, специфически связывающиеся с ВСМА, которые можно применять для получения различных таргетных противоопухолевых лекарственных средств и лекарственных средств для диагностики опухолей. Настоящее изобретение выполнено на основе вышеуказанных результатов.
Технические термины, используемые в настоящем документе, имеют общепринятые для специалистов или аналогичные значения. Некоторые термины определены ниже для понимания настоящего изобретения.
Термин «ВСМА» в настоящем документе относится к антигену созревания В-клеток, представляющему собой трансмембранный белок III типа, состоящий из 184 аминокислотных остатков (эталонная последовательность в NCBI: NP_001183.2), причем указанная аминокислотная последовательность представлена в SEQ ID No: 37. В конкретном варианте реализации ВСМА относится к ВСМА человека.
Термин «APRIL» в настоящем документе относится к лиганду А, индуцирующему пролиферацию, представляющему собой лиганд, индуцирующий пролиферацию, состоящий из 184 аминокислотных остатков (эталонная последовательность в NCBI: NP_003799.1), и принадлежащему к надсемейству ФНО.
Термин «антитело» в настоящем документе относится к антиген-связывающему иммунному белку. Термин «антитело» в настоящем документе включает интактное полноразмерное антитело, содержащее антиген-связывающую область, и любые фрагменты такого антитела, сохраняющие «антиген-связывающий фрагмент» или «антиген-связывающую область», или его одиночную цепь, например, одноцепочечный вариабельный фрагмент (scFv). Нативное антитело относится к гликопротеину, содержащему по меньшей мере две тяжелые (Н) цепи и две легкие (L) цепи или их антиген-связывающие фрагменты, связанные друг с другом дисульфидными связями. Термин «антитело» также включает все рекомбинантные формы антител, в частности, антитела, описанные в настоящем документе, например, антитела, экспрессируемые в прокариотических клетках, негликозилированные антитела и фрагменты антител, связывающиеся с антигенами и их производными, указанные здесь и далее. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно обозначаемой в настоящем документе как VH) и константной области тяжелой цепи. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначаемой в настоящем документе как VL) и константной области легкой цепи. Области VH и VL можно далее разделить на гипервариабельные области, называемые областями, определяющими комплементарность (CDR), перемежающиеся с более консервативными областями, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, выстроенных от N-конца к С-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат связывающие домены, взаимодействующие с антигеном. Константная область антитела может опосредовать связывание иммуноглобулина с тканями или факторами, включая различные иммунные клетки (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента.
Фрагменты антител включают (i) Fab-фрагменты, состоящие из VL-, VH-, CL- и CH1-доменов, в том числе Fab'- и Fab'-SH-фрагменты, (ii) Fd-фрагменты, состоящие из VH- и CH1-доменов, (iii) Fv-фрагмент, состоящий из VL- и VH-доменов одиночного антитела; (iv) dAb-фрагмент, состоящий из одиночной вариабельной области (Ward et al., 1989, Nature 341: 544-546); (v) Р(ab')2-фрагмент, двухвалентный фрагмент, содержащий два связанных Fab-фрагмента; (vi) антиген-связывающий сайт одноцепочечной молекулы Fv (Bird et al, 1988, Science 242: 423-426; Huston et al., 1988, Proc. Natl. Acad. Sci. USA 85:5879-5883); (vii) биспецифический димер одноцепочечного Fv (PCT/US92/09965); (viii) «диатело» или «триатело», поливалентные или полиспецифические фрагменты, сконструированные путем объединения генов (Tomlinson et al, 2000, Methods Enzymol. 326: 461-479; WO94/13804; Holliger et al., 1993, Proc. Natl. Acad. Sci. USA 90:6444-6448); и (ix) scFv, генетически объединенный с идентичными или отличающимися антителами (Coloma & Morrison, 1997, Nature Biotechnology 15, 159-163), но не ограничиваются указанными.
Термин «Fc» или «Fc-область» в настоящем документе включает полипептид, содержащий константную область антитела, не являющуюся первым иммуноглобулиновым доменом константной области. Таким образом, Fc относится к двум последним иммуноглобулиновым доменам константной области IgA, IgD и IgG и трем последним иммуноглобулиновым доменам константной области IgE и IgM, а также гибким шарнирам на N-конце этих доменов. Для IgA и IgM Fc может содержать J-цепь. Для IgG Fc содержит шарниры между иммуноглобулиновыми доменами Сγ2 и Сγ3, а также Cγ1 и Сγ2. Границы Fc-области могут варьировать, однако Fc-область тяжелой цепи IgG человека обычно определяют как область, содержащую остатки С226 и Р230 на С-конце, причем нумерация остатков основана на ЕС-индексе по Kabat. Для IgG1 человека в настоящем документе Fc определяют как область, содержащую остаток Р232 на С-конце, причем нумерация остатков основана на ЕС-индексе по Kabat. Fc может относиться к выделенной области или области в окружении Fc-полипептида, например, антитела. «Шарнир», как указано выше, содержит гибкий полипептид, содержащий аминокислоты между первым и вторым константными доменами антитела. Структурно СН1-домен IgG заканчивается в положении EU220, а СН2-домен IgG начинается с остатка EU237. Таким образом, для IgG1 в настоящем документе шарнир антитела определяют как область, содержащую остатки с 221 (D221 IgG1) до 231 (А231 IgG1), причем нумерация основана на ЕС-индексе по Kabat.
Термин «исходное антитело» или «исходный иммуноглобулин» в настоящем документе включает немодифицированное антитело, подлежащее модификации с получением вариантов. Исходное антитело может представлять собой природное антитело или вариант или модифицированную версию природного антитела. Термин «исходное антитело» может относиться к самому антителу, композиции, содержащей исходное антитело, или нуклеотидной последовательности, кодирующей антитело. Термин «исходное антитело» или «исходный иммуноглобулин» в настоящем документе включает антитело мыши или химерное антитело, подлежащее модификации с получением гуманизированного антитела.
Термин «вариантное антитело» или «вариант антитела» в настоящем документе включает последовательность антитела, отличающуюся от последовательности исходного антитела по меньшей мере на одну аминокислотную модификацию по сравнению с исходным антителом. Последовательность вариантного антитела в настоящем документе обладает по меньшей мере приблизительно 80%, предпочтительно по меньшей мере приблизительно 90%, более предпочтительно по меньшей мере приблизительно 95% идентичностью аминокислотной последовательности по отношению к последовательности исходного антитела. Термин «вариант антитела» может относиться к самому антителу, композиции, содержащей исходное антитело, или нуклеотидной последовательности, кодирующей антитело.
Термин «вариант» в настоящем документе включает последовательность антитела, отличающуюся от последовательности исходного антитела по меньшей мере на одну аминокислотную модификацию по сравнению с исходным антителом. В конкретном варианте реализации последовательность вариантного антитела в настоящем документе обладает по меньшей мере приблизительно 80%), предпочтительно по меньшей мере приблизительно 90%, более предпочтительно по меньшей мере приблизительно 95%, более предпочтительно по меньшей мере приблизительно 97%, более предпочтительно по меньшей мере приблизительно 98%, наиболее предпочтительно по меньшей мере приблизительно 99% идентичностью аминокислотной последовательности по отношению к последовательности исходного антитела. Термин «вариант антитела» может относиться к самому антителу, композиции, содержащей исходное антитело, или нуклеотидной последовательности, кодирующей антитело. Термин «аминокислотная модификация» включает аминокислотную замену, добавление или делецию аминокислоты, и «аминокислотная замена» относится к замещению аминокислоты в конкретном положении последовательности исходного полипептида другой аминокислотой. Например, замена R94K означает, что аргинин в положении 94 замещен лизином, а «инсерция аминокислоты» в настоящем документе относится к добавлению аминокислоты в конкретном положении последовательности исходного полипептида. В настоящем документе «деления аминокислоты» или «делеция» относится к удалению аминокислоты в конкретном положении последовательности исходного полипептида.
Термин «консервативная модификация» или «консервативная модификация последовательности» в настоящем документе относится к аминокислотной модификации, которая не оказывает существенного влияния или не изменяет характеристики связывания антитела, содержащего аминокислотную последовательность. Такие консервативные модификации включают аминокислотные замены, инсерции и делеции. Модификации можно внедрить в антитела согласно настоящему изобретению стандартными способами, известными в данной области техники, например, посредством сайт-специфического мутагенеза и ПЦР-опосредованного мутагенеза. Консервативные аминокислотные замены представляют собой замены, при которых аминокислотные остатки заменяют аминокислотными остатками, содержащими аналогичные боковые цепи. В данной области техники существуют определения семейств аминокислотных остатков, содержащих аналогичные боковые цепи. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновую кислоту, глутаминовую кислоту), незаряженными полярными боковыми цепями (например, глицин, аспарагин, серин, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), β-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или более из аминокислотных остатков в областях CDR или каркасных областях антитела согласно настоящему изобретению можно заменить аминокислотными остатками других семейств с идентичной боковую цепью и протестировать функцию, сохранившуюся в модифицированном (вариантном) антителе.
Все положения в константной области тяжелой цепи иммуноглобулина, обсуждаемые в настоящем изобретении, нумеруют на основе ЕС-индекса по Kabat (Kabat et al., 1991, sequences of proteins of immunological interest, 5th edition, United States Public Health Service, National Institutes of Health, Bethesda; данная работа полностью включена в настоящий документ посредством ссылки). «ЕС-индекс по Kabat» относится к нумерации остатков ЕС IgG1-антитела человека, описанной в Edelman et al., 1969, Biochemistry 63: 78-85.
«Антигенная детерминанта» в настоящем документе, также называемая антигенным эпитопом, может состоять из непрерывной последовательности белка ВСМА или прерывистой трехмерной структуры последовательности белка ВСМА.
Термин «химерный рецептор антигена» или «CAR» в настоящем документе относится к полипептиду, содержащему внеклеточный домен, способный связывать антиген, трансмембранный домен и цитоплазматический сигнальный домен (т.е. внеклеточный сигнальный домен), причем внутриклеточный сигнальный домен относится к белку, передающему сигналы в клетку, продуцируя вторичный мессенджер посредством заданного сигнального пути, и тем самым регулирующему клеточную активность, или к белку, соответствующему такому мессенджеру и действующему в качестве эффектора, включая первичный сигнальный домен и функциональный сигнальный домен (т.е. костимулирующий сигнальный домен), происходящий от стимулирующей молекулы, определение которой приведено ниже. Внутриклеточный сигнальный домен генерирует сигнал, стимулирующий иммунную эффекторную функцию клеток, несущих CAR (например, CAR Т-клеток), причем примеры иммунных эффекторных функций, например, у CART-клеток, включают литическую активность клеток и хелперную активность, включая секрецию цитокинов.
Термин «первичный сигнальный домен» относится к модуляции начальной активации TCR-комплекса раздражающим образом. В одном аспекте первичный сигнальный домен активируют, например, связыванием TCR/CD3-комплекса с молекулой МНС со связанным пептидом, тем самым опосредуя Т-клеточный ответ (включая пролиферацию, активацию, дифференцировку, и т.д., но не ограничиваясь указанными). Первичный сигнальный домен, который функционирует стимулирующим образом, может содержать иммунорецепторный мотив активации тирозина или сигнальный мотив ITAM. Примеры первичных сигнальных доменов, содержащих ITAM, которые в особенности полезны для настоящего изобретения, включают последовательность, происходящую от TCR ξ, FcRγ, FcRβ, CD3γ, CD3δ, CD3ε, CD5, CD22, CD79a, CD79b, CD278 (также называемого «ICOS») и CD66d, но не ограничиваются указанными. В типичном CAR согласно настоящему изобретению, в любом одном или более из CAR согласно настоящему изобретению внутриклеточный сигнальный домен содержит внутриклеточную сигнальную последовательность, например, первичный сигнальный домен CD3ξ.
Термин «костимулирующий сигнальный домен» относится к «костимулирующей молекуле», которая является партнером по связыванию Т-клеткой, которая специфически связывается с костимулирующим лигандом, тем самым опосредуя костимулирующий ответ Т-клетки, например, пролиферацию, но не ограничиваясь ей. Костимулирующие молекулы представляют собой молекулы клеточной поверхности или их лиганды, необходимые для эффективного иммунного ответа, и неантигенные рецепторы. Костимулирующие молекулы включают молекулы МНС I класса, рецепторы лигандов BTLA и Toll, а также ОХ40, CD2, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11a/CD18) и 4-1BB (CD137), но не ограничиваются указанными.
В одном аспекте настоящего изобретения CAR содержит химерный гибридный белок, содержащий внеклеточный антиген-распознающий домен, трансмембранный домен и внутриклеточный сигнальный домен, а внутриклеточный сигнальный домен содержит функциональный сигнальный домен, происходящий от стимулирующей молекулы. В одном аспекте CAR содержит химерный гибридный белок, содержащий внеклеточный антиген-распознающий домен, трансмембранный домен и внутриклеточный сигнальный домен, а внутриклеточный сигнальный домен содержит функциональный сигнальный домен, происходящий от костимулирующей молекулы, и функциональный сигнальный домен, происходящий от стимулирующей молекулы. В одном аспекте CAR содержит химерный гибридный белок, содержащий внеклеточный антиген-распознающий домен, трансмембранный домен и внутриклеточный сигнальный домен, а внутриклеточный сигнальный домен содержит по меньшей мере два функциональных сигнальных домена, происходящих от одной или более из костимулирующих молекул, и функциональный сигнальный домен, происходящий от стимулирующей молекулы. В одном аспекте CAR включает необязательную лидерную последовательность, присоединенную к аминокислоте (ND-концевой) гибридного белка CAR. В одном аспекте CAR дополнительно содержит лидерную последовательность по N-концу внеклеточного антиген-распознающего домена, причем указанная лидерная последовательность необязательно отщепляется от антиген-распознающего домена (например, scFv) во время процессинга и локализации CAR в клеточной мембране.
Термин «CD3ξ» в настоящем документе определяют как белок, представленный как последовательность с учетным номером GenBan BAG36664.1, или эквивалентные остатки, полученные из организма, не являющегося человеком, например, мыши, грызуна, обезьяны, человекообразной обезьяны и т.п. «Домен CD3ξ» в настоящем документе определяют как аминокислотные остатки, полученные из цитоплазматического домена ξ-цепи достаточные для функциональной передачи исходного сигнала, необходимого для активации Т-клетки. В одном аспекте цитоплазматический домен ξ, содержит остатки 52-164 последовательности с учетным номером GenBan BAG36664.1, их функциональный ортолог - эквивалентные остатки, полученные из организма, не являющегося человеком, например, мыши, грызуна, обезьяны, человекообразной обезьяны и т.д.
Термин «4-1ВВ» в настоящем документе относится к члену надсемейства TNFR, содержащему аминокислотную последовательность с учетным номером GenBank ААА62478.2, или эквивалентные остатки, полученные из организма, не являющегося человеком, например, мыши, грызуна, обезьяны, человекообразной обезьяны и т.п. «Костимулирующий домен 4-1ВВ» определяют как аминокислотную последовательность 214-255 с учетным номером GenBank ААА62478.2, или эквивалентные остатки, полученные из неклассифицированного организма, например, мыши, грызуна, обезьяны, человекообразной обезьяны и т.п. В одном аспекте «костимулирующий домен 4-1ВВ» представляет собой последовательность, предоставленную в SEQ ID NO: 35, или эквивалентные остатки, полученные из организма, не являющегося человеком, например, мыши, грызуна, обезьяны, человекообразной обезьяны и т.п.
Термин «интерферон» в настоящем документе относится к полноразмерному интерферону или фрагменту интерферона (укороченному интерферону) или мутантному интерферону, в значительной степени сохраняющему биологическую активность полноразмерного интерферона дикого типа (например, сохраняющему по меньшей мере 80%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, 98% или 99% активности полноразмерного интерферона). Интерфероны включают интерфероны I типа (например, интерферон-α и интерферон-β) и интерфероны II типа (например, интерферон-γ).
Антитело согласно настоящему изобретению или его вариант можно применять для получения различных противоопухолевых таргетных лекарственных средств, а также средств для диагностики опухолей, в частности, для получения иммунных эффекторных клеток, нацеленных на ВСМА.
Антитело против ВСМА
В настоящем изобретении описаны антиген-связывающие белки, содержащие антиген-связывающую область на основе scFv, в том числе антитела. Рекомбинантный ВСМА использовали для отбора scFv из библиотеки фагового дисплея scFv человека. Эти молекулы демонстрируют прекрасную специфичность. Например, антитело распознает только клетки K562, стабильно экспрессирующие ВСМА, и не распознает клетки K562.
В некоторых вариантах реализации настоящее изобретение включает антитело, содержащее последовательность scFv, присоединенную к одной или нескольким константным областям тяжелой цепи с образованием антитела, содержащего Fc-область иммуноглобулина человека и получением двухвалентного белка, что увеличивает общее сродство и стабильность антитела. Кроме того, Fc-фрагмент обеспечивает прямое конъюгирование других молекул (включая флуоресцентные красители, цитотоксины, радиоизотопы и т.д., но не ограничиваясь указанными), например, с антителами, применяемыми в исследованиях по количественному определению антигена с целью иммобилизации антител для измерения сродства, направленной доставки терапевтических средств, применения иммунных эффекторных клеток для тестирования Fc-опосредованной цитотоксичности и многих других вариантов применения.
Результаты, представленные в настоящем документе, подчеркивают специфичность, чувствительность и практичность антител согласно настоящему изобретению при направленном воздействии на ВСМА.
Молекулы согласно настоящему изобретению основаны на одноцепочечных вариабельных фрагментах (scFv), выявленных и отобранных посредством фагового дисплея, аминокислотных последовательностях, придающих специфичность к ВСМА, и формируют основу всех антиген-связывающих белков согласно настоящему изобретению. Таким образом, scFv можно применять для проектирования различных молекул «антител», включая, например, полноразмерные антитела, их фрагменты, например, Fab и Р(ab')2-фрагменты, гибридные белки (включая scFv_Fc), поливалентные антитела, т.е. антитело, обладающее более чем одной специфичностью по отношению к тому же или другим антигенам, например, биспецифическое антитело, связывающее Т-клетки (BiTE), три-антитело и т.д. (Cuesta et al, Multivalent antibodies: when design surpasses evolution, Trends in Biotechnology 28: 355-362, 2010).
В одном варианте реализации, где антиген-связывающий белок представляет собой полноразмерное антитело, тяжелые и легкие цепи антител согласно настоящему изобретению могут быть полноразмерными цепями (наприме, антитело может содержать по меньшей мере одну, предпочтительно две интактные тяжелые цепи, и по меньшей мере одну, предпочтительно две интактные легкие цепи), и, в качестве альтернативы, может содержать антиген-связывающую группу (Fab, F(ab')2, Fv или scFv). В других вариантах реализации константную область тяжелой цепи антитела выбрана, например, из IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgD и IgE. Выбор типа антитела зависит от иммунной эффекторной функции, которую должно стимулировать сконструированное антитело. Подходящие аминокислотные последовательности константных областей различных изотипов иммуноглобулинов и способы получения широкого спектра антител известны специалистам в области конструирования рекомбинантных иммуноглобулинов.
В первом аспекте настоящего изобретения предложены антитело или его фрагмент, связывающийся с ВСМА, содержащие CDR1 тяжелой цепи, содержащий аминокислотную последовательность согласно любой из SEQ ID NO: 1, 60, 62, и/или CDR2 тяжелой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 2, 61, 63, и/или CDR3 тяжелой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 3, 4, 5. В еще одном аспекте настоящего изобретения предложены антитело или его фрагмент, связывающийся с ВСМА, содержащие CDR1 легкой цепи, содержащий аминокислотную последовательность согласно SEQ ID NO: 6, и/или CDR2 легкой цепи, содержащую аминокислотную последовательность согласно SEQ ID NO: 7, и/или CDR3 легкой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 8, 9, 10. В еще одном аспекте настоящего изобретения предложены антитело или его фрагмент, связывающийся с ВСМА, содержащие CDR1 тяжелой цепи, содержащий аминокислотную последовательность согласно любой из SEQ ID NO: 1, 60, 62, и/или CDR2 тяжелой цепи, содержащую аминокислотные последовательности согласно любой из SEQ ID NO: 2, 61, 63, и/или CDR3 тяжелой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 3, 4, 5, и/или CDR1 легкой цепи, содержащую аминокислотную последовательность согласно SEQ ID NO: 6, и/или CDR2 легкой цепи, содержащую аминокислотную последовательность согласно SEQ ID NO: 7, и/или CDR3 легкой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 8, 9, 10. ВСМА-связывающее антитело или его фрагмент предпочтительно содержит CDR1 тяжелой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 1, 60, 62, и CDR2 тяжелой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 2, 61, 63, и CDR3 тяжелой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 3, 4, 5, и/или CDR1 легкой цепи, содержащую аминокислотную последовательность согласно SEQ ID NO: 6, и CDR2 легкой цепи, содержащую аминокислотную последовательность согласно SEQ ID NO: 7, и CDR3 легкой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 8, 9, 10. ВСМА-связывающее антитело или его фрагмент более предпочтительно содержит CDR1 тяжелой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 1, 60, 62, и CDR2 тяжелой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 2, 61, 63, и CDR3 тяжелой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 2, 61, 63, и CDR3 тяжелой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 6, и CDR2 легкой цепи, содержащую аминокислотную последовательность согласно SEQ ID NO: 7, и CDR3 легкой цепи, содержащую аминокислотную последовательность согласно любой из SEQ ID NO: 8, 9, 10.
В еще одном аспекте настоящего изобретения предложены антитело или его фрагмент, связывающийся с ВСМА, содержащие последовательность вариабельной области тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 13, 17, 21, 56 и 58.
В еще одном аспекте настоящего изобретения предложены антитело или его фрагмент, связывающийся с ВСМА, содержащие последовательность вариабельной области легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 11, 15 и 19.
Каждая из последовательностей вариабельной области тяжелой и легкой цепи может связываться с ВСМА, поэтому последовательности вариабельной области тяжелой и легкой цепи можно «смешать и сопоставить» для получения молекул, связывающих ВСМА, согласно настоящему изобретению.
В еще одном аспекте настоящего изобретения предложены варианты антитела или его фрагмента, связывающегося с ВСМА. Соответственно, в настоящем изобретении предложено антитело или его фрагмент, содержащие вариабельную область тяжелой цепи и/или легкой цепи, по меньшей мере на 80% идентичную последовательности вариабельной области тяжелой или легкой цепи. Идентичность аминокислотной последовательности вариабельных областей тяжелой и/или легкой цепи предпочтительно составляет по меньшей мере 85%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, более предпочтительно 96%, более предпочтительно 97%, еще предпочтительнее 98%, наиболее предпочтительно 99%, включая, например, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% и 100%. Вариант можно получить из антитела, описанного в настоящей заявке в качестве исходного антитела, путем скрининга дрожжевой библиотеки, скрининга фаговой библиотеки, точечной мутации и т.п. Как и в способе, использованном в примере 10 настоящей заявки, антитело 23F10 использовали в качестве исходного антитела, а способ скрининга фаговой библиотеки использовали для модификации мутаций.
В еще одном аспекте настоящего изобретения предложено антитело, распознающее ту же антигенную детерминанту, что и антитело против ВСМА, описанное выше.
Свойства антитела против ВСМА
В данной области техники известны стандартные анализы для оценки связывающей способности антитела, например, антитела против ВСМА, в том числе, например, твердофазный ИФА, вестерн-блоттинг и проточно-цитометрический анализ. Подходящие способы анализа подробно описаны в примерах.
Нуклеиновые кислоты, векторы и клетки-хозяева
В настоящем изобретении также предложены выделенная нуклеиновая кислота, кодирующая антитело, связывающееся с ВСМА и его фрагментом, вектор и клетка-хозяин, содержащая указанную нуклеиновую кислоту или вектор. Нуклеиновая кислота может присутствовать в интактной клетке, в клеточном лизате или в частично очищенном или практически чистом виде.
Нуклеиновую кислоту согласно настоящему изобретению можно получить с использованием стандартных молекулярно-биологических методик, например, стандартных методик ПЦР-амплификации или клонирования кДНК, тем самым получая кДНК, кодирующую легкую и тяжелую цепи антитела или кодирующую VH- и VL-сегменты. Для антител, полученных из библиотек генов иммуноглобулинов (например, с использованием технологии фагового дисплея), из библиотеки можно получить одну или более из нуклеиновых кислот, кодирующих антитела. Способы введения чужеродных нуклеиновых кислот в клетки-хозяева в целом известны в данной области техники и могут варьироваться в зависимости от используемой клетки-хозяина.
Предпочтительные молекулы нуклеиновых кислот согласно настоящему изобретению представляют собой молекулы, выбранные из группы, состоящей из SEQ ID NO: 12, 16 и 20, кодирующих вариабельную область легкой цепи, и/или молекулы, выбранные из группы, состоящей из SEQ ID NO: 14, 18, 22, 57 и 59, кодирующих вариабельную область тяжелой цепи. Более предпочтительная молекула нуклеиновых кислот содержит последовательность SEQ ID NO: 14, кодирующую тяжелую цепь, и последовательность SEQ ID NO: 12, кодирующую легкую цепь, или содержит последовательность SEQ ID NO: 18, кодирующую тяжелую цепь, и последовательность SEQ ID NO: 16, кодирующую легкую цепь, или содержит последовательность SEQ ID NO: 22, кодирующую тяжелую цепь, и последовательность SEQ ID NO: 20, кодирующую легкую цепь, или содержит последовательность SEQ ID NO: 57, кодирующую тяжелую цепь, и последовательность SEQ ID NO: 20, кодирующую легкую цепь, или содержит последовательность SEQ ID NO: 59, кодирующую тяжелую цепь, и последовательность SEQ ID NO: 20, кодирующую легкую цепь.
Для экспрессии белка нуклеиновая кислота, кодирующая антитело согласно настоящему изобретению, может быть встроена в экспрессирующий вектор. Существуют разнообразные экспрессирующие векторы для экспрессии белка. Экспрессирующие векторы могут включать самореплицирующиеся внехромосомные векторы или векторы, встроенные в геном хозяина. Экспрессирующие векторы, используемые в настоящем изобретении, включают векторы, обеспечивающие экспрессию белка в клетках млекопитающих, бактерий, насекомых, дрожжей и системах in vitro, но не ограничиваются указанными. Как известно в данной области техники, для экспрессии антител в настоящем изобретении можно использовать различные экспрессирующие векторы, доступные для приобретения или иным образом.
Иммуноконъюгат
Кроме того, в настоящем изобретении предложен многофункциональный иммуноконъюгат, содержащий антитела, описанные в настоящем документе, и дополнительно содержащий по меньшей мере одну функциональную молекулу другого типа. Функциональную молекулу выбирают из молекулы, нацеленной на поверхностный маркер опухоли, молекулы, ингибирующей опухоль, молекулы, нацеленной на поверхностный маркер иммунной клетки, и детектируемой метки, но не ограничиваются указанными. Антитело и функциональная молекула могут образовывать конъюгат путем ковалентного присоединения, связывания, присоединения, перекрестного сшивания и т.п.
В предпочтительном варианте иммуноконъюгат может содержать антитело согласно настоящему изобретению и по меньшей мере одну молекулу, нацеленную на поверхностный маркер опухоли или молекулу, ингибирующую опухоль. Молекула, ингибирующая опухоль, может представлять собой противоопухолевые цитокины или противоопухолевые токсины. Цитокины предпочтительно включают ИЛ-2, ИЛ-7, ИЛ-12, ИЛ-15, ИФН I типа, ФНО-альфа, но не ограничиваются указанными. В конкретном варианте реализации молекула, нацеленная на поверхностный маркер опухоли, представляет собой молекулу, нацеленную на тот же поверхностный маркер опухоли, что и для антитела согласно настоящему изобретению. Например, молекула, нацеленная на поверхностный маркер опухоли, может представлять собой антитело или лиганд, связывающиеся с поверхностным маркером опухоли и, например, способные действовать синергически с антителами согласно настоящему изобретению, обеспечивая более точное направленное воздействие на опухолевые клетки.
В предпочтительном варианте иммуноконъюгат может содержать антитело в соответствии с настоящим изобретением и детектируемую метку. Такие обнаруживаемые метки включают флуоресцентные метки, хромогенные метки, например, ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминесцентные материалы, радиоактивные материалы, позитрон-излучающие металлы и ионы нерадиоактивных парамагнитных металлов, но не ограничиваются указанными. Можно задействовать более чем один маркер. Метка, используемая для мечения антитела с целью обнаружения и/или анализа и/или диагностики, зависит от используемого конкретного способа и/или методики обнаружения/анализа/диагностики, например, иммуногистохимического окрашивания образцов (ткани), проточной цитометрии и т.п. Подходящие метки для способов и/или методик обнаружения/анализа/диагностики, известных в данной области техники, хорошо известны специалистам в данной области техники.
В предпочтительном варианте иммуноконъюгат может содержать: антитело согласно настоящему изобретению и молекулу, нацеленную на поверхностный маркер иммунной клетки. Молекула, нацеленная на поверхностный маркер иммунной клетки, может представлять собой антитело или лиганд, связывающийся с поверхностным маркером иммунной клетки, способный распознавать иммунную клетку и переносить антитело согласно настоящему изобретению к иммунной клетке. Антитело по настоящему изобретению может направлять иммунную клетку на опухолевые клетки, тем самым индуцируя иммунную клетку с целью специфического уничтожения опухолей. Поверхностный маркер иммунной клетки может быть выбран из группы, состоящей из CD3, CD16, CD28, и, предпочтительно, антитело, связывающееся с поверхностным маркером иммунной клетки, представляет собой антитело против CD3. Иммунные клетки могут быть выбраны из группы, состоящей из Т-клеток, NK-клеток или NKT-клеток.
В качестве средства химического получения иммуноконъюгата путем конъюгирования, непосредственного или косвенного (например, посредством линкера), иммуноконъюгат можно получить в виде гибридного белка, содержащего антитело согласно настоящему изобретению и другие подходящие белки. Гибридный белок можно продуцировать способом, известным в данной области техники, например, рекомбинантно продуцировать путем конструирования и последующей экспрессии молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую антитело, в одной рамке считывания с нуклеотидной последовательностью, кодирующей подходящую метку.
В еще одном аспекте настоящего изобретения предложена молекула нуклеиновой кислоты, кодирующая по меньшей мере одно антитело согласно настоящему изобретению, его функциональный вариант или иммуноконъюгат. После получения соответствующей последовательности можно использовать способ рекомбинации для получения соответствующей последовательности в больших количествах. Обычно это делают путем его клонирования в вектор, переноса вектора в клетку, а затем выделения соответствующей последовательности из пролиферирующих клеток-хозяев обычными способами.
Настоящее изобретение также относится к векторам, содержащим соответствующие последовательности ДНК, описанные выше, а также соответствующим промоторам или контрольным последовательностям. Эти векторы можно использовать для трансформации подходящей клетки-хозяина с целью обеспечения возможности экспрессии белка. Клетка-хозяин может представлять собой прокариотическую клетку, например, бактериальную клетку; или клетку низших эукариот, например, дрожжевую клетку; или клетку высших эукариот, например, клетку млекопитающего.
Химерный рецептор антигена, содержащий антитело против ВСМА
В настоящем изобретении предложено множество химерных рецепторов антигенов (CAR), содержащих антитело или фрагмент антитела согласно настоящему изобретению. CAR Т-клетка обладает противоопухолевыми свойствами. В некоторых вариантах реализации клетки (например, Т-клетки) трансдуцируют вирусным вектором, кодирующим CAR. В некоторых вариантах реализации вирусный вектор представляет собой лентивирусный вектор. В некоторых вариантах реализации клетки могут стабильно экспрессировать CAR.
В предпочтительном варианте реализации ВСМА-связывающий фрагмент CAR представляет собой scFv-фрагмент антитела, сохраняющий эквивалентную связывающую активность, например, он связывается с тем же антигеном с сопоставимой эффективностью по сравнению с IgG-антителом, из которого он получен. Фрагмент антитела является функциональным и поэтому обеспечивает биохимическую реакцию, которая может включать активацию иммунного ответа, ингибирование инициации сигнального пути, активируемого антигеном-мишенью, ингибирование киназной активности и т.п., но не ограничивается указанными. Соответственно, в настоящем изобретении предложена BCMA-CAR, содержащий домен связывания WT1 и встроенный в Т-клетку, и способ его применения в адоптивной иммунотерапии.
В одном аспекте антиген-вязывающий домен CAR против ВСМА представляет собой scFv-фрагмент антитела, гуманизированный по сравнению с последовательностью scFv мыши, от которой он происходит.
В одном аспекте CAR согласно настоящему изобретению объединяет антиген-связывающий домен конкретного антитела с внутриклеточной сигнальной молекулой. Например, в некоторых аспектах внутриклеточные сигнальные молекулы включают CD3 ξ-цепь, сигнальные модули 4-1ВВ и CD28 и их комбинации, но не ограничиваются указанными.
В одном аспекте BCMA-CAR содержит по меньшей мере один внутриклеточный сигнальный домен, выбранный из сигнального домена CD137 (4-1ВВ), сигнального домена CD28, сигнального домена CD3ξ или любой их комбинации. В одном аспекте BCMA-CAR содержит по меньшей мере один внутриклеточный сигнальный домен, происходящий от одной или более из костимулирующих молекул, не являющихся CD137 (4-1ВВ) или CD28.
В типичном случае последовательность BCMA-CAR может представлять собой 7A12-BBZ (SEQ ID NO: 75), 25C2-BBZ (SEQ ID NO: 76), 25D2-BBZ (SEQ ID NO: 77), 7G2-BBZ (SEQ ID NO: 78), 7A12-28Z (SEQ ID NO: 79), 7A12-28BBZ (SEQ ID NO: 80), 7G2-28Z (SEQ ID NO: 81), 7G2-28BBZ (SEQ ID NO: 82), 25C2-28Z (SEQ ID NO: 83), 25C2-28BBZ (SEQ ID NO: 84), 25D2-28Z (SEQ ID NO: 85), 25D2-28BBZ (SEQ ID NO: 86). Специалист в данной области техники может выбрать обычный трансмембранный домен и внутриклеточный домен для замены трансмембранного домена и внутриклеточного домена вышеуказанной SEQ ID NO: 75-86, что входит в рамки настоящего изобретения.
Т-клетка, модифицированная химерным рецептором антигена
В настоящем изобретении также предложена иммунная клетка, содержащая химерный рецептор антигена согласно настоящему изобретению.
В еще одном аспекте Т-клетка, модифицированная химерным рецептором антигена, предложена в настоящем изобретении, дополнительно несет кодирующую последовательность чужеродного цитокина; указанный цитокин предпочтительно включает: ИЛ-12, ИЛ-15 или ИЛ-21. Иммунные клетки предпочтительно выбраны из Т-лимфоцитов, NK-клеток или NKT-клеток.
В еще одном аспекте Т-клетка, модифицированная химерным рецептором антигена, предложенная в настоящем изобретении, дополнительно содержит блокатор PD-L1 или белок, блокирующий PD-L1, например, нативный PD-1 или мутантный PD-1, способный связываться с PD-L1, или фрагмент нативного или мутантного PD-1, способного связываться с PD-L1, или антитело против PD-L1. В типичном случае блокатор PD-L1 может содержать аминокислотную последовательность, кодируемую SEQ ID NO: 70.
Фармацевтическая композиция
Антитела, иммуноконъюгаты, содержащие антитела, и генетически модифицированные иммунные клетки согласно настоящему изобретению можно применять при изготовлении фармацевтической композиции или диагностического реагента. В дополнение к эффективному количеству антитела, иммунологического конъюгата или иммунной клетки композиция может дополнительно содержать фармацевтически приемлемый носитель. Термин «фармацевтически приемлемый» означает, что при надлежащем введении молекулярных структур и композиций животным или людям они не вызывают нежелательных, аллергических или других неблагоприятных реакций.
Конкретными примерами некоторых веществ, которые можно применять в качестве фармацевтически приемлемых носителей или их компонентов, являются углеводы, например, лактоза, декстроза и сахароза; производные крахмала, например, кукурузный крахмал и картофельный крахмал; целлюлоза и ее производные, например, карбоксиметилцеллюлоза натрия, этилцеллюлоза и метилцеллюлоза; трагакантовая камедь; солод; желатин; тальк; твердые смазывающие вещества, например, стеариновая кислота и стеарат магния; сульфат кальция; растительные масла, например, арахисовое масло, хлопковое масло, кунжутное масло, оливковое масло, кукурузное масло и масло какао; многоатомные спирты, например, пропиленгликоль, глицерин, сорбит, маннит и полиэтиленгликоль; альгиновая кислота; эмульгаторы, например, Tween®; увлажнители, например, лаурилсульфат натрия; красители; вкусоароматические добавки; таблетки, стабилизаторы; антиоксиданты; консерванты; апирогенная вода; изотонические растворы хлорида натрия; и фосфатные буферы и т.п.
Композицию согласно настоящему изобретению при необходимости можно изготовить в виде различных лекарственных форм, а дозировку для пациента может определить врач в соответствии с такими факторами, как тип, возраст, масса тела и общее состояние заболевания у пациента, способ введения и т.п. Например, можно использовать инъекцию или другой вариант лечения.
Преимущества изобретения:
1. В настоящем изобретении предложены специфические антитела против ВСМА;
2. В настоящем изобретении предложены эффекторные иммунные клетки, нацеленные на ВСМА; и
3. Антитело согласно настоящему изобретению способно эффективно связываться с опухолевыми клетками, экспрессирующими ВСМА, а эффекторные иммунные клетки согласно настоящему изобретению проявляют выраженную способность уничтожать опухолевые клетки, экспрессирующие ВСМА, и, следовательно, указанное антитело и эффекторные и иммунные клетки согласно настоящему изобретению можно эффективно и безопасно применять для лечения множественной миеломы, в связи с чем они являются материальной основой для лечения множественной миеломы.
Настоящее изобретение в настоящем документе дополнительно проиллюстрировано специфическими примерами. Следует понимать, что эти примеры не предназначены для ограничения рамок изобретения. Экспериментальные способы в следующих примерах, где не указаны специфические условия, обычно выполняют в соответствии с общепринятыми условиями, например, описанными в J. Sambrook et al., Molecular Cloning Experimental Guide, Third Edition, Science Press, 2002, или в соответствии с условиями, рекомендованными изготовителем.
Пример 1. Получение рекомбинантного белка ВСМА
a. Конструирование экспрессирующей плазмиды ВСМА_huFc, ВСМА_muFc
Ген (SEQ ID NO: 39) внеклеточного сегмента ВСМА человека, Met1-Ala54 (SEQ ID NO: 38), синтезировали in vitro, вставляли в эукариотическую экспрессирующую плазмиду, содержащую Fc-фрагмент Asp104-Lys330 константной области тяжелой цепи IgG1 человека и связанную с «GS» для получения гибридного экспрессируемого белка BCMA_huFc (SEQ ID NO: 40), и последовательность соответствующего гена показана в SEQ ID NO: 41.
Ген (SEQ ID NO: 39) внеклеточного сегмента ВСМА человека вставляли в эукариотическую экспрессирующую плазмиду, содержащую Fc-фрагмент Arg100-Lys324 константной области тяжелой цепи IgG1 мыши и связанную с «GS» для получения гибридного экспрессируемого белка ВСМА_muFc (SEQ ID NO: 42), и последовательность соответствующего гена показана в SEQ ID NO: 43.
b. Экспрессия BCMA_huFc, BCMA_muFc посредством временной трансфекции
1) Через день после трансфекции 6-7 × 105 клеток 293F/мл инокулировали в 125-мл культуральные колбы;
2) В день трансфекции 3×107 клеток доводили до объема 28 мл средой для экспрессии FreeStyle™ 293;
3) Комплекс «липид-ДНК» получали последством следующих поэтапных действий:
30 мкг ДНК разбавляли Opti-MEM I до конечного объема 1 мл и тщательно перемешивали;
60 мкл 293fectinTM разбавляли Opti-MEM I до конечного объема 1 мл и тщательно перемешивали;
смесь инкубировали в течение 5 минут при комнатной температуре;
4) разбавленную ДНК разбавляли 293fectinTM и инкубировали в течение 20 минут при комнатной температуре;
5) 2 мл комплекса «ДНК-293fectin» добавляли к 28 мл клеток, культивировали при 37°С в атмосфере 8% СО2, при 125 об/мин в течение 3-4 дней и собирали надосадочную жидкость.
с. Очистка BCMA_huFc, BCMA_muFc
1) Надосадочную жидкость центрифугировали при 13000 об/мин в течение 15 мин;
2) Для аффинной очистки использовали фильтр с белком А, причем поэтапные действия перечислены ниже:
Уравновешивание: 10 объемов колонки буфера для уравновешивания использовали для фильтра с белком А.
Загрузка: загружали образец, обработанный с помощью 0,45-мкм фильтра.
Промывка: 20 объемов колонки буфера для уравновешивания использовали для удаления примесей до прекращения непрерывного потока.
Элюирование: 10 объемов колонки буфера для элюирования добавляли для элюирования исследуемого белка (в пробирку для сбора заранее добавляли 6% нейтрализующего буфера).
Составление раствора:
Буфер для уравновешивания: PBS рН 7,4
Буфер для элюирования: 0,1 М глицин, рН 2,6
Нейтрализующий буфер: 1 М трис
3) Элюат фильтровали через 0,22-мкм мембрану, концентрировали в пробирке Millipore для ультрафильтрации с пороговой молекулярной массой 10 кДа до объема 1 мл и обессоливали с использованием обессоливающей колонки PD-Midi. Собирали 1,5 мл образца. Концентрацию белка измеряли по OD280/1,47.
2 мкг забирали для электрофореза в ДСН-ПААГ, результаты показаны на фигуре 1.
Пример 2. Конструирование стабильной линии клеток K562-ВСМА
1. Конструирование плазмиды для упаковки pWPT-BCMA
Полноразмерный ген (SEQ ID NO: 37) ВСМА человека синтезировали in vitro, вводили сайты расщепления MluI, SalI (SEQ ID NO: 44) вставляемые в плазмиду pWPT для упаковки лентивируса путем двойного гидролиза.
2. Упаковка лентивирусов
a) Lenti-x 293Т гидролизовали, высевали на 10-см чашки 10 из расчета 8×106 клеток и культивировали при 37°С.
b) На следующее утро: получали смесь плазмид/ПЭИ
pWPT-BCMA, 5 мкг
psPAX.2, 7,5 мкг
pMD2.G, 2,5 мкг
добавляли к 800 мкл DMEM и инкубировали. Соответствующий объем ПЭИ составлял 45 мкл, инкубирование выполняли в течение 5 мин в 800 мкл ОМЕМ.
c) смесь плазмид добавляли по каплям в раствор ПЭИ для инкубирования, аккуратно перемешивали и инкубировали в течение 20 мин при комнатной температуре.
d) полученную смесь плазмид/ПЭИ по каплям добавляли к клеткам и перемешивали. Раствор меняли через 5 часов.
e) надосадочную жидкость вируса собирали через 72 ч, фильтровали через 0,45-мкм фильтр и временно хранили при 4°С.
3. Клетки K562, инфицированные ВСМА-вирусом
a) Середина 1 дня: хорошо выращенные клетки K562 высевали из расчета 1×105 клеток на 6-см чашку.
b) Середина 2 дня: надосадочную жидкость клеток K562 выбрасывали, добавляли 3 мл свежей полной среды и добавляли 1 мл базового раствора вируса до конечной концентрации 6 мкг/мл полибрена.
c) Утро 3 дня: надосадочную жидкость выбрасывали и добавляли 5 мл свежей полной среды.
d) Утро 6 дня: некоторое количество клеток отбирали для обнаружения в потоке.
4. Выявление смешанного клона K562-ВСМА
a) Смешанные клоны K562-ВСМА и отрицательные клетки K562 дважды промывали 1% NCS (PBS, содержащего 1% сыворотки теленка), а затем инкубировали с первичным антителом: антителом huBCMA (abcam, #17323) разбавленным 1% NCS в соотношении 1:1000 (по 50 мкл) и инкубировали в течение 50 мин при 4°С.
b) Клетки дважды промывали 1% NCS, а затем инкубировали с вторичным антителом: DyLight488-меченым антителом козы против IgG крысы (abcam, #ab98420), разбавленным 1% NCS в соотношении 1:200 (по 50 мкл) и инкубировали в течение 45 мин при 4°С.
с) Клетки трижды промывали 1% NCS, ресуспендировали в 1% NCS и обнаруживали с помощью прибора Guava easyCyte™ НТ System. Результаты показаны на фигуре 2А.
5. Посев одиночных клонов K562-ВСМА
a) Клетки смешанного клона K562-ВСМА подсчитывали и высевали одиночные клоны посредством предельного разведения.
b) Через неделю наблюдали рост клонов, среду заменяли.
c) Через две недели отбирали клетки в лунках моноклонального роста и размножали их путем культивирования.
6. Выявление одиночного клона K562-ВСМА
Способ обнаружения был таким же, как при выявлении смешанного клона, результаты эксперимента показаны на фиг. 2В. 4 из этих одиночных клонов являлись ВСМА-положительными клонами.
Пример 3. Скрининг на предмет ВСМА-специфичного scFv с использованием полной библиотеки фагового дисплея человека
Библиотека фагового дисплея, использованная в настоящем изобретении, представляла собой полную фаговую библиотеку природных scFv человека, сконструированную данной компанией и обладающей емкостью хранения 1Е + 11. scFv-фрагменты, обладающие высокой специфичностью по отношению к ВСМА, получали с помощью способов скрининга, известных специалисту. Вкратце, 10 мкг/мл антигена BCMA_huFc и Fc-фрагмента человека иммобилизовали в иммунологических пробирках, соответственно. Для снижения влияния Fc-фрагмента фаговую библиотеку добавляли в иммунологические пробирки с иммобилизованным Fc-фрагментом человека на 1 ч. Надосадочную жидкость отбирали и добавляли в иммунологические пробирки с иммобилизованным BCMA_huFc на 1,5 часа, затем неспецифический фаг отмывали, а связанный фаг элюировали и использовали для инфицирования Е. coli TG1 в логарифмической фазе роста. Элюированный фаг размножали, размноженную фаговую библиотеку очищали осаждением в ПЭГ/NaCl на следующем цикле скрининга. Пэннинг выполняли в течение 3-4 циклов с целью обогащения клонов фагов с scFv, специфически связывавшихся с ВСМА. Положительные клоны определяли стандартными способами твердофазного ИФА BCMA_huFc. Fc-фрагмент человека использовали в качестве неспецифического антигена для твердофазного ИФА с целью проверки специфичности антитела. В общей сложности выполнили скрининг 2470 клонов, причем 160 из них специфически связывались с BCMA_huFc, но не связывались с Fc-фрагментом человека при твердофазном ИФА. 76 клонов с высокими значениями сигнала выбрали для секвенирования и получили 23 одиночных последовательности. Эти 23 клона очистили и экспрессировали, получив три клона, специфически связывавшиеся с клетками K562-ВСМА (фиг. 4), и этим клонам дали названия 7G2, 7А12 и 23F10. Секвенирование позволило установить, что вариабельная область тяжелой цепи 7А12 представляла собой аминокислотную последовательность, показанную в SEQ ID NO: 13, вариабельная область легкой цепи представляла собой аминокислотную последовательность, показанную в SEQ ID NO: 11; вариабельная область тяжелой цепи 7G2 представляла собой аминокислотную последовательность, показанную в SEQ ID NO: 17, вариабельная область легкой цепи представляла собой аминокислотную последовательность, показанную в SEQ ID NO: 15; и вариабельная область тяжелой цепи 23F10 представляла собой аминокислотную последовательность, показанную в SEQ ID NO: 21, вариабельная область легкой цепи представляла собой аминокислотную последовательность, показанную в SEQ ID NO: 19.
Аминокислотная последовательность вариабельной области тяжелой цепи 7А12 (SEQ ID NO: 13):

Нуклеотидная последовательность вариабельной области тяжелой цепи 7А12 (SEQ ID NO: 14):
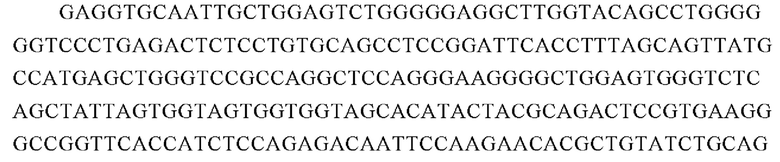
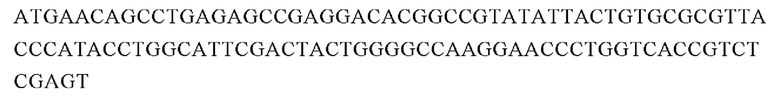
Аминокислотная последовательность вариабельной области легкой цепи 7А12 (SEQ ID NO: 11)

Нуклеотидная последовательность вариабельной области легкой цепи 7А12 (SEQ ID NO: 12):

Аминокислотная последовательность вариабельной области тяжелой цепи 7G2 (SEQ ID NO: 17):

Нуклеотидная последовательность вариабельной области тяжелой цепи 7G2 (SEQ ID NO: 17):
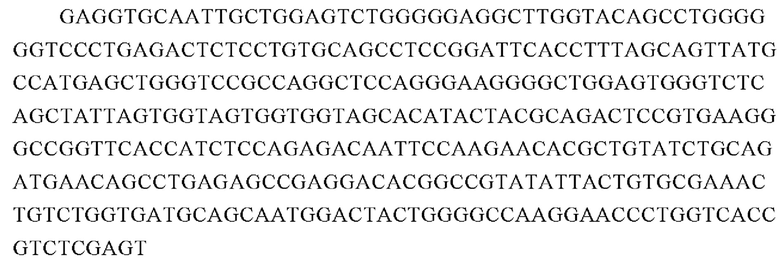
Аминокислотная последовательность вариабельной области легкой цепи 7G2 (SEQ ID NO: 15):

Нуклеотидная последовательность вариабельной области легкой цепи 7G2 (SEQ ID NO: 16):
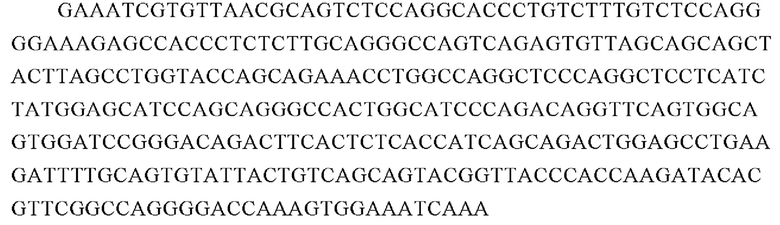
Аминокислотная последовательность вариабельной области тяжелой цепи 23F10 (SEQ ID NO: 21):

Аминокислотная последовательность вариабельной области тяжелой цепи 23F10 (SEQ ID NO: 22):
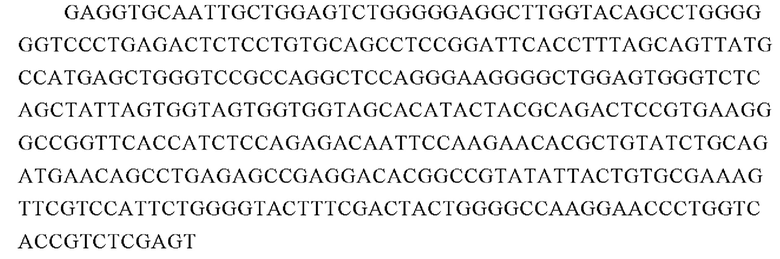
Аминокислотная последовательность вариабельной области тяжелой цепи 23F10 (SEQ ID NO: 19):

Аминокислотная последовательность вариабельной области тяжелой цепи 23F10 (SEQ ID NO: 20):
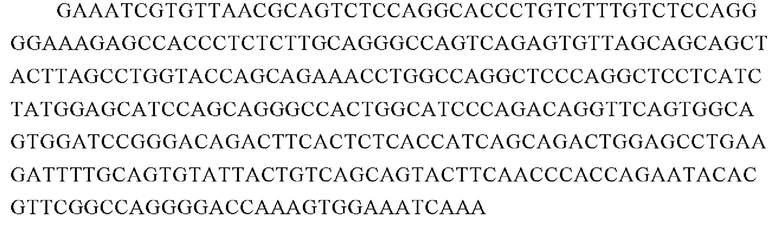
Пример 4. Конструирование гибридного scFv_Fc-антитела против ВСМА и его временная экспрессия, очистка и выявление активности в эукариотических клетках
Праймеры конструировали для VH- и VL-фрагментов 7G2, 7А12, 23F10, соответственно, и внедряли гибкий линкер, состоящий из 15 аминокислот (GGGGSGGGGSGGGGS) для получения scFv; сайт расщепления NheI и защитные основания внедряли выше VH, а сайт расщепления BamHI и защитные основания внедряли после VL. Продукт ПЦР анализировали электрофорезом в 1% агарозном геле, очищали и выделяли. После гидролиза его лигировали в эукариотический экспрессирующий вектор V152 (приобретенный в Shanghai Ruijin Biotechnology Co., Ltd.). Клетки 293F в логарифмической фазе роста временно трансфицировали с использованием реагента для трансфекции 293fectin™ (Invitrogen, 12347-019) или полиэтиленимина (ПЭИ) (Sigma-Aldrich, 408727). На 5-7 день после трансфекции собирали надосадочную жидкость и подвергали ее аффинной очистке с белком А. Полученные антитела количественно и качественно анализировали электрофорезом в ДСН-ПААГ (фиг. 5).
Связывание антитела с K562, стабильно экспрессирующими ВСМА, проверяли проточной цитометрией. Способ FACs-обнаружения осуществляли следующим образом: клетки собирали, однократно промывали ростовой средой и ресуспендировали в PBS. Концентрацию клеток доводили до 4Е+5 клеток/мл. Гибридные scFv_Fc-антитела, разбавленные в градиенте, инкубировали с клетками на льду в течение 30 минут, начальная концентрация антитела составляла 500 нМ, что представляло собой 5-кратное разбавление в общей сложности в 7 градиентах. Затем антитело инкубировали с FITC-меченым вторичным антителом против IgG мыши и после двукратной промывки обнаруживали с использованием Guava easyCyte™ НТ System. На фигуре 6 показано связывание гибридных форм scFv_Fc антител 7А12, 7G2 и 23F10 с K562-ВСМА. Все три антитела характеризовались связыванием в зависимости от концентрации, значения ЕС50 составляли 3,13 нМ, 3,42 нМ и 5,61 нМ, соответственно.
Пример 5. Определение сродства антитела с использованием поверхностного плазмонного резонанса (ППР)
Сродство различных антител к ВСМА определяли с помощью biacore Т200. Использовали следующий способ:
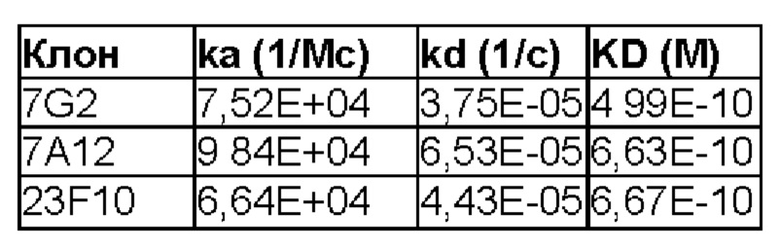
BCMA_huFc иммобилизовали на чипе СМ5 путем присоединения по аминогруппе до приблизительно 500 ЕО и через канал с иммобилизованным антигеном в качестве подвижной фазы пропускали антитело, разбавленное градиентом, со скоростью потока 30 мкл/мин. Подвижный буфер представлял собой HBS-N, температура составляла 25°С. Экспериментальные данные анализировали в BIAevaluation 3.2, кинетические кривые аппроксимировали моделью Лэнгмюра 1:1. KD 7А12 (scFv_Fc) составила 663 пМ, KD 7G2 (scFv_Fc) - 499 пМ, a KD 23F10 (scFv_Fc) - 667 пМ (см. фигуру 7). Параметры представлены в следующей таблице:
Пример 6. Определение сродства антител к опухолевым линиям клеток посредством FACs
RPMI8226 представляет собой В-лимфоциты периферической крови множественной миеломы человека. Способ FACs-обнаружения осуществляли следующим образом: клетки собирали, однократно промывали ростовой средой и ресуспендировали в PBS. Концентрацию клеток доводили до 4Е+5 клеток/мл. Гибридные scFv_Fc-антитела, разбавленные в градиенте, инкубировали с клетками на льду в течение 30 минут, начальная концентрация антитела составляла 500 нМ и в общей сложности 5-кратное разбавление в 7 градиентах. Затем антитело инкубировали с а FITC-меченым вторичным антителом против IgG мыши и после двукратной промывки обнаруживали посредством Guava easyCyte™ НТ System. На фигуре 8 показано связывание гибридных форм scFv_Fc антител 7А12, 7G2 и 23F10 в зависимости от концентрации с клетками линии RPMI8226.
Пример 7. Анализ конкурентного связывания антитела против ВСМА с лигандом ВСМА APRIL
1. Экспрессия очищенного рекомбинантного белка APRIL
Гибридный белок, состоящий из His115-Leu250 APRIL человека и Fc-фрагмента Asp104-Lys330 константной области тяжелой цепи IgG1 человека, соединенных «GS», подвергали рекомбинантной экспрессии. Гибридный белок APRIL_huFc (SEQ ID NO: 45), последовательность соответствующего гена представляла собой SEQ ID NO: 46. Временную трансфекцию, экспрессию и очистку выполняли, как описано в примере 1.
2. Конкурентный твердофазный ИФА
На планшете для ИФА иммобилизовали 50 нг/мл 100 мкл/пустого BCMA_muFc при 4°С в течение ночи. На следующий день планшет трижды промывали PBS, добавляли PBS, содержащий 2% сухое обезжиренное молоко и блокировали при комнатной температуре в течение 1 часа. Одновременно добавляли 40 нг/мл APRIL_huFc и антитело 7А12, 7G2 или 23F10, разбавленное градиентом (начальная концентрация 200 нМ, 3-кратное разбавление, 7 градиентов). Полученную смесь инкубировали в течение 1 ч при комнатной температуре, трижды промывали PBST и трижды - PBS. Добавляли разведение 1:1000 ПХ-меченого антитела против Fc человека, инкубировали в течение 1 ч при комнатной температуре, трижды промывали PBST и трижды - PBS. Для развития окрашивания добавляли ТМВ и считывали на микропланшетном ридере.
Результаты эксперимента показаны на фиг. 9. Все антитела - 7А12, 7G2 и 23F10 - могли в значительной степени ингибировать связывание APRIL с ВСМА, что демонстрировало способность антител согласно настоящему изобретению ингибировать связывание ВСМА с его естественным лигандом.
Пример 8. Конструирование плазмиды химерного рецептора антигена против ВСМА (CAR)
а Конструирование плазмиды химерного рецептора антигена на основе антитела 7А12 против ВСМА
Лентивирусные плазмиды, экспрессирующие химерные рецепторы антигенов на основе антитела 7А12 второго и третьего поколений конструировали с использованием PRRLSIN-cPPT.EF-1α в качестве вектора, включая PRRLSIN-cPPT.EF-1α-7A12-28Z, PRRLSFN-cPPT.EF-1α-7A12-BBZ и PRRLSFN-cPPT.EF-1α-7A12-28BBZ. Последовательность 7A12-28Z состоит из сигнального пептида CD8α (SEQ ID NO: 23), scFv 7A12 (SEQ ID NO: 47), шарнира CD8 (SEQ ID NO: 25), трансмембранной области CD28 (SEQ ID NO: 27), внутриклеточного сигнального домена (SEQ ID NO: 29) и внутриклеточного сегмента CD3ξ, (SEQ ID NO: 31) CD3; последовательность 7A12-BBZ состоит из сигнального пептида CD8α (SEQ ID NO: 23), scFv 7A12 (SEQ ID NO: 47), шарнира CD8 (SEQ ID NO: 25), трансмембранной области (SEQ ID NO: 33), внутриклеточного сигнального домена CD137 (SEQ ID NO: 35) и CD3ξ (SEQ ID NO: 31); последовательность 7A12-28BBZ состоит из сигнального пептида CD8α (SEQ ID NO: 23), 7A12-scFv (SEQ ID NO: 47), шарнира CD8 (SEQ ID NO: 25), трансмембранной области CD28 (SEQ ID NO: 27), внутриклеточного сегмента (SEQ ID NO: 29), внутриклеточного сигнального домена CD137 (SEQ ID NO: 35) и CD3ξ (SEQ ID NO: 31).
b. Конструирование плазмиды для химерного рецептора антигена на основе антитела 7G2 против ВСМА
Лентивирусные плазмиды, экспрессирующие химерные рецепторы антигенов на основе антитела 7G2 второго и третьего поколений конструировали с использованием PRRLSIN-cPPT.EF-1α в качестве вектора, включая PRRLSFN-cPPT.EF-1α-7G2-28Z, PRRLSIN-cPPT.EF-1α-7G2-BBZ и PRRLSIN-cPPT.EF-1α-7G2-28BBZ. Последовательность 7G2-28Z состоит из сигнального пептида CD8α (SEQ ID NO: 23), scFv 7G2 (SEQ ID NO: 48), шарнира CD8 (SEQ ID NO: 25), трансмембранной области CD28 (SEQ ID NO: 27), внутриклеточного сигнального домена (SEQ ID NO: 29) и внутриклеточного сегмента CD3ξ, (SEQ ID NO: 31) CD3; последовательность 7G2-BBZ состоит из сигнального пептида CD8α (SEQ ID NO: 23), scFv 7G2 (SEQ ID NO: 48)), шарнира CD8 (SEQ ID NO: 25), трансмембранной области (SEQ ID NO: 33), внутриклеточного сигнального домена CD137 (SEQ ID NO: 35) и CD3ξ, (SEQ ID NO: 31); последовательность 7G2-28BBZ состоит из сигнального пептида CD8α (SEQ ID NO: 23), 7G2-scFv (SEQ ID NO: 48), шарнира CD8 (SEQ ID NO: 25), трансмембранной области CD28 (SEQ ID NO: 27), внутриклеточного сегмента (SEQ ID NO: 29), внутриклеточного сигнального домена CD137 (SEQ ID NO: 35) и CD3ξ (SEQ ID NO: 31).
Пример 9. Получение CAR Т-клеток
1. Упаковка лентивируса, концентрация вируса и определение титра лентивирусного вектора CAR, нацеленного на ВСМА
a. Упаковка лентивируса
1) Клетки 293Т инокулировали в 10-см культуральной чашке и культивировали в течение ночи при 37°С, 5% СО2 для трансфекции; среда представляла собой DMEM, содержащую 10% фетальную телячью сыворотку (Gibico);
2) 5,4 мкг плазмиды с геном-мишенью PRRLSIN-cPPT.EF-1α-EGFP (имитация) или плазмиды с родственным CAR и 6,2 мкг плазмиды для упаковки pRsv-REV, 6,2 мкг RRE-PMDLg, 2,4 мкг Vsvg растворяли в 800 мкл холостой среды DMEM и смешивали;
3) 60 мкг ПЭИ растворяли в 800 мкл бессывороточной среды DMEM, аккуратно перемешивали (или встряхивали на вортексе при 1000 об/мин в течение 5 секунд) и инкубировали в течение 5 мин при комнатной температуре;
4) Формирование комплекса для трансфекции: смесь плазмид добавляли к смеси ПЭИ, непосредственно после добавления смесь встряхивали на вортексе или аккуратно перемешивали и инкубировали при комнатной температуре в течение 20 мин;
5) 1,6 мл комплекса для трансфекции добавляли в 10-см культуральную чашку, содержащую 11 мл среды DMEM (менять среду не нужно); через 4-5 часов меняли среду трансфицированных клеток 293Т на среду DMEM, содержащую 10% FBS, инкубировали в течение 72 ч при 37°С и собирали надосадочную жидкость вирусов.
b. Концентрация лентивирусов
1) Получение 5Х ПЭГ8000 NaCl: 8,766 г NaCl и 50 г ПЭГ8000 взвешивали и растворяли в 200 мл чистой воды Milli-Q; стерилизовали при 121°С в течение 30 мин; и хранили при 4°С;
2) Надосадочную жидкость лентивирусов фильтровали через 0,45-мкм фильтр; 7,5 мл базового раствора 5Х ПЭГ-8000 NaCl добавляли к 30 мл фильтрованного исходного раствора вируса; 3-5 раз перемешивали раз в 20-30 минут; помещали при температуре 4°С на ночь; и центрифугировали при 4°С, 4000 g в течение 20 мин;
3) Надосадочную жидкость аспирировали и выбрасывали, пробирку помещали на 1-2 минуты, остаточную жидкость аспирировали и выбрасывали; добавляли соответствующее количество раствора лентивируса для растворения осадка лентивируса; распределяли и хранили при -80°С.
с. Определение титра лентивируса
1) клетки 293Т инокулировали в 6-луночных культуральных планшетах из расчета 2×105 клеток, 1 мл/лунку; добавляли 10 мкг/мкл (начальная концентрация) раствор полибрена в 0,6 мкл/мл до конечной концентрации 6 мкг/мл; культивировали при 37°С, 5% СО2 в течение 1 часа; среда представляла собой DMEM, содержащую 10% фетальную телячью сыворотку;
2) концентрат вируса добавляли из расчета 10 мкл/лунку, при 5-кратном разбавлении в 3 градиентах, и культивировали при 37°С, 5% СО2;
3) через 72 часа после инфицирования использовали трипсин для гидролиза (30 с) клеток 293Т, добавляли 1 мл DMEM (10% FBS) для прекращения гидролиза, суспензию клеток переносили в 2-мл центрифужную пробирку (две аликвоты), центрифугировали при 5000 об/мин в течение 5 мин, надосадочную жидкость выбрасывали; клетки дважды промывали PBS (2% NBS);
4) к клеткам в контрольной группе добавляли 50 мкл PE-SA (разбавление 1:200), инкубировали на льду в течение 45 мин, дважды промывали PBS (2% NBS) и ресуспендировали в качестве контроля;
5) 50 мкл разбавленного в соотношении 1:50 Е(ab')2-антитела козы против IgG человека, меченого биотином, добавляли к клеткам в группе тестируемых клеток, инкубировали на льду в течение 45 мин и дважды промывали PBS (2% NBS); добавляли 50 мкл антитела PE-SA (разбавление 1:200) и инкубировали на льду в течение 45 мин;
6) для ресуспендирования клеток добавляли 2 мл PBS (2% NBS) и центрифугировали при 4°С, 5000 об/мин в течение 5 минут; надосадочную жидкость выбрасывали; эту процедуру повторяли дважды;
7) добавляли 500 мкл PBS (2% NBS) и переносили в проточную трубку. Обнаружение на канале РЕ выполняли посредством проточной цитометрии, количество клеток с положительным показателем 5-20% было соответствующим. Рассчитывали титр (БОЕ/мл) = количество клеток × положительный показатель / объем вируса.
2. Т-лимфоцит, трансдуцированный лентивирусом ------ Получение CAR-положительных Т-лимфоцитов
1) Активация Т-лимфоцита: лимфоциты добавляли в среду для культивирования лимфоцитов из расчета плотности приблизительно 1×106/мл, одновременно добавляли магнитные гранулы (Invitrogen) с иммобилизованными антителами против CD3 и CD28 при соотношении магнитные гранулы:клетки 2:1 и рекомбинантный ИЛ-2 человека (Shanghai Huaxin Biotech Co., Ltd.) в конечной концентрации 500 ед/мл и инкубировали в течение 48 ч;
2) Через день после инфицирования на 24-луночном планшете иммобилизовали Retronectin в конечной концентрации 5 мкг/мл и инкубировали в течение ночи при 4°С;
3) раствор Retronectin (PBS) в 24-луночном планшете выбрасывали, планшет дважды промывали 1 мл PBS;
4) К МПК-клеткам добавляли концентрированный лентивирус при MOI = 10, центрифугировали при 1000 g в течение 40 мин и переносили в клеточный инкубатор;
5) Амплификация: Инфицированные клетки пересевали через день из расчета плотности 5×105/мл, добавляя рекомбинантный ИЛ-2 человека в конечной концентрации 500 ед/мл к раствору для культивирования лимфоцитов.
3. Экспрессия химерного рецептора антигена Т-лимфоцита
1) На 7 день культивирования в центрифужную пробирку отбирали 1×106 Т-лимфоцитов, инфицированных лентивирусом;
2) Т-клетки центрифугировали при 4°С, 5000 об/мин в течение 5 мин, надосадочную жидкость выбрасывали, осадок дважды промывали PBS;
3) К тестируемым клеткам добавляли 50 мкл Р(ab')2-антитела козы против IgG человека, меченого биотином (разбавление 1:50), инкубировали на льду в течение 45 мин; дважды промывали PBS (2% NBS); и добавляли 50 мкл антитела PE-SA (разбавление 1:200) и инкубировали на льду в течение 45 мин;
4) Для ресуспендирования клеток добавляли 2 мл PBS (2% NBS) и центрифугировали при 4°С, 5000 об/мин в течение 5 минут, надосадочную жидкость выбрасывали; эту процедуру повторяли дважды;
5) Добавляли 500 мкл PBS (2% NBS) и переносили в проточную трубку. Обнаружение на канале РЕ выполняли посредством проточной цитометрии, определяя долю CAR-положительных Т-клеток.
Положительные показатели инфекции имитационных клеток, 7A12-28Z, 7A12-BBZ, 7A12-28BBZ, 7G2-28Z, 7G2-BBZ и 7G2-28BBZ Т-клетки в эксперименте по определению цитотоксичности in vitro показаны на фигуре 10 и составляли 72,8%, 60,8%, 48,7%, 57,4%, 67,5%, 68,8%, 63,6%, соответственно.
4. Анализ цитотоксичности CAR Т-клеток, нацеленных на ВСМА
Набор для анализа цитотоксичности без использования радиоактивных материалов CytoTox 96 (Promega) использовали в соответствии с инструкциями к набору для анализа цитотоксичности без использования радиоактивных материалов CytoTox 96.
Клетки-мишени: 75 мкл 2×105/мл клеток K562, K562-ВСМА и RPMI-8226 инокулировали в 96-луночные планшеты, соответственно. Эффекторные клетки: Имитационные Т-клетки и CAR Т-клетки, экспрессирующие различные химерные рецепторы антигенов, добавляли в соотношении «эффекторные клетки: мишени», равном 3:1, 1:1 или 1:3. Для каждой группы задали лунки в четырех повторностях и использовали среднее значение по 4 повторным лункам. Время обнаружения соответствовало 18 часу инкубирования клеток. Каждая экспериментальная группа и каждая контрольная группа представляли собой:
Каждая экспериментальная группа: каждая клетка-мишень + CAR Т-клетки, экспрессирующие различные химерные рецепторы антигенов;
Контрольная группа 1: максимальное высвобождение ЛДГ из клеток-мишеней;
Контрольная группа 2: спонтанное высвобождение ЛДГ из клеток-мишеней;
Контрольная группа 3: спонтанное высвобождение ЛДГ из эффекторных клеток;
Формула расчета цитотоксичности представляла собой: цитотоксичность % = [(экспериментальная группа - контроль эффекторных клеток - контроль клеток-мишеней) / (максимум для клеток-мишеней - контроль клеток-мишеней)] × 100%.
Результаты показали, что каждая из CAR Т-клеток, экспрессирующих различные химерные рецепторы антигенов, обладали значительной цитотоксической активностью in vitro против ВСМА-положительных клеток K562-ВСМА и RPMI-8226, особенно против клеток RPMI-8226, эндогенно экспрессирующих ВСМА, при почти полном отсутствии цитотоксического действия по отношению к ВСМА-отрицательным клеткам K562 (фиг. 11А).
5. Обработка NOD/ТКИН-мышей после введения В-лимфоцитов периферической крови RPMI-8226 множественной миеломы
Клетки RPMI-8226 инокулировали 40 NOD/ТКИН-мышам из расчета 8×106/мышь, соответственно. На 12 день после подкожной инокуляции опухолевых клеток средний объем опухоли составлял 75 мм3, мышей случайным образом разделили на 4 группы и вводили 1×107 CAR Т-клеток в хвостовую вену. Перед инъекцией внутрибрюшинно вводили циклофосфамид в дозировке 100 мг/кг для предварительного уничтожения остаточных Т-клеток у мышей. На 17 день после инъекции CAR Т-клеток мышей умерщвляли посредством смещения шейных позвонков.
Анализировали размер опухоли у мышей. Результаты показаны на фиг. 11В. По сравнению с группой UTD, в экспериментальных группах 7A12-28Z, 7A12-BBZ и 7A12-28BBZ отмечен значимый противоопухолевый эффект, и на 17 день после инъекции CAR Т-клеток имел место 1 случай регресса опухоли у 7 мышей в экспериментальной группе 7A12-28Z, 2 случая регресса опухоли у 7 мышей в экспериментальной группе 7A12-BBZ и 7 случаев регресса опухоли у 7 мышей в экспериментальной группе 7A12-28BBZ. Частота ингибирования опухолей составляла 84,6% (7A12-28Z), 65,4%) (7A12-BBZ) и 100% (7A12-28BBZ), соответственно.
Пример 10. Модификация антитела 23F10
В данном примере 23F10 использовали в качестве исходного антитела и модифицировали его посредством фагового дисплея. Фаговую библиотеку сконструировали на основе 23F10 с сохраненными CDR3-областями легкой и тяжелой цепи, и две фаговые библиотеки сконструировали путем случайного изменения CDR1 и CDR2 легкой цепи или CDR1 и CDR2 тяжелой цепи с использованием вырожденных праймеров, соответственно. Информация о праймерах приведена ниже:
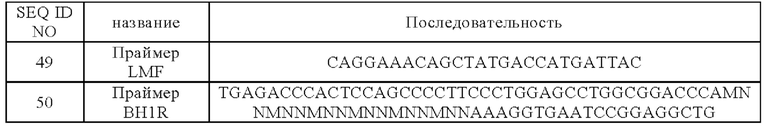
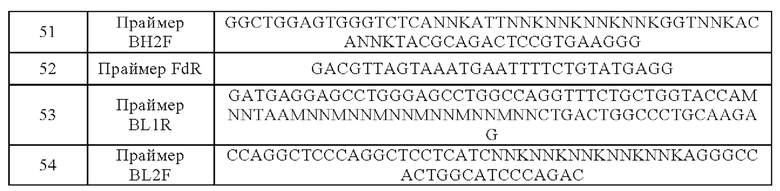
2.1 Конструирование мутанта 23F10
Матричную плазмиду вначале сконструировали на основе антитела 23F10 (scFv) (SEQ ID NO: 55). Для получения фаговых библиотек случайным образом измененных CDR1 и CDR2 легкой цепи использовали праймеры LMF и BL1R для ПЦР-амплификации фрагмента 1; праймеры BL2F и FdR использовали для ПЦР-амплификации фрагмента 2; затем фрагмент 1 и фрагмент 2 лигировали посредством мостиковой ПЦР, получая полноразмерный scFv, содержавший рандомизированную последовательность, а затем полноразмерный фрагмент гидролизовали с использованием NcoI и NotI и лигировали в идентично гидролизованную матричную плазмиду лигазой Т4. Плазмиду трансдуцировали в TG1-компетентные клетки электропорацией, емкость хранения составляла 1,50Е+9. Для получения фаговых библиотек случайным образом измененных CDR1 и CDR2 тяжелой цепи использовали праймеры LMF и BH1R для ПЦР-амплификации фрагмента 3; праймеры BH2F и FdR использовали для ПЦР-амплификации фрагмента 4; затем фрагмент 3 и фрагмент 4 лигировали посредством мостиковой ПЦР, получая полноразмерный scFv, содержавший рандомизированную последовательность, а затем полноразмерный фрагмент гидролизовали с использованием NcoI и NotI и лигировали в идентично гидролизованную матричную плазмиду лигазой Т4. Плазмиду трансдуцировали в TG1-компетентные клетки электропорацией, емкость хранения составляла 2,2Е+9.
Скрининг фаговых библиотек. В отношении способа, описанного в примере 3, начальная концентрация антигена BCMA_huFc составляла 20 нМ, и для следующего цикла скрининга выполняли 5-кратное разбавление градиентом. Пэннинг выполняли в течение 2-3 циклов с целью обогащения клонов фагов с scFv, специфически связывавшихся с BCMA_huFc. Положительные клоны определяли стандартными способами твердофазного ИФА BCMA_huFc. Fc-фрагмент человека использовали в качестве неспецифического антигена для твердофазного ИФА с целью проверки специфичности антитела. В общей сложности отобрали 80 клонов, положительных при твердофазном ИФА, константу диссоциации Kd в надосадочной жидкости определяли с использованием biacore после повторной индукции. Среди этих клонов было два клона 25С2 и 25D2, Kd которых было в 10 раз ниже, чем у исходного клона 23F10, как показано в следующей таблице:

Легкие цепи клонов 25С2 и 25D2 секвенировали и обнаружили, что они были идентичны 23F10. На фигуре 12 показано сравнение аминокислотных последовательностей тяжелой цепи клонов 25С2, 25D2 и 23F10, причем по сравнению с исходным антителом 23F10 тяжелая цепь клона 25С2 содержала 5 точечных мутаций (SEQ ID NO: 56, 57 представляют собой аминокислотную последовательность и нуклеотидную последовательность вариабельной области тяжелой цепи 25С2, соответственно), CDR1 содержала 2 точечные мутации -замену серина на глицин по 31 положению и тирозина на аспарагин по 32 положению; CDR2 содержала 2 точечные мутации - замену серина на аспарагинил по 54 положению и тирозина на фенилаланин по 59 положению, а каркасная область содержала 1 точечную мутацию - замену серина на глицин по 30 положению. По сравнению с исходным антителом 23F10 тяжелая цепь клона 25D2 содержала 4 точечные мутации (SEQ ID NO: 58, 59 представляют собой аминокислотную последовательность и нуклеотидную последовательность вариабельной области тяжелой цепи 25D2, соответственно), причем область CDR2 содержала 3 точечных мутации - замену серина на глицин по 54 положению, серина на аспарагин по 57 положению и тирозина на фенилаланин по 59 положению, а каркасная область содержала 1 точечную мутацию - замену серина на аргинин по 30 положению.
Последовательность HCDR1 25С2 приведена в SEQ ID NO: 60, а последовательность HCDR2 25С2 приведена в SEQ ID NO: 61. Последовательность HCDR1 25D2 приведена в SEQ ID NO: 62, а последовательность HCDR2 25D2 приведена в SEQ ID NO: 63. Нуклеотидная последовательность и аминокислотная последовательность scFv 25С2 приведены в SEQ ID NO: 64, 65, соответственно, а нуклеотидная последовательность и аминокислотная последовательность scFv 25D2 приведены в SEQ ID NO: 66, 67, соответственно.
2.2 Экспрессия и очистка клона 25С2, 25D2 (scFv_Fc)
В соответствии с примером 4, соответствующие сайты расщепления и защитные основания внедряли выше VH, и соответствующие сайты расщепления и защитные основания внедряли ниже VL. Продукт ПЦР анализировали электрофорезом в 1% агарозном геле, очищали и выделяли. После гидролиза его лигировали в эукариотический экспрессирующий вектор V152, содержащий Fc-фрагмент человека (приобретенный в Shanghai Ruijin Biotechnology Co., Ltd.), временно трансфицировали в клетки 293F с использованием 293Fectin и экспрессировали.
Агрегацию 25С2 и 25D2 анализировали посредством SEC. Как показано на фиг. 13А и 13В, доля антитела в мономерной форме составляла 91% и 97%, соответственно. По сравнению с исходным антителом 23F10 (30% частота мономеров) частота мономеров увеличилась на 61% и 67%), соответственно, а агрегация значительно снизилась. После концентрирования ультрафильтрацией полученные антитела количественно и качественно анализировали посредством электрофореза в ДСН-ПААГ. Выход составил 80 мкг/мл и 60 мкг/мл, соответственно (выход = масса конечного продукта/объем трансфекции).
2.3 Характеристики связывания клонов 25С2, 25D2
Использовали клетки K562 и K562 (K562-ВСМА), стабильно экспрессирующие ВСМА человека, которые собирали, промывали полной ростовой средой и высевали в титрационные микропланшеты с U-образным дном из расчета приблизительно 1-5×105 клеток/лунку. Гибридное антитело scFv_Fc, разбавленное градиентом, инкубировали с K562-ВСМА/K562 в течение 30 минут на льду, а затем инкубировали с FITC-меченым антителом против Fc человека в качестве вторичного антитела. После двухэтапной промывки выполняли анализ с использованием Guava easyCyteTM HT System, экспериментальные данные обрабатывали с использованием GraphPad Prism, получая ЕС50. На фигуре 14 показано связывание 25С2, 25D2 с клетками K562-ВСМА и K562. Результаты показали, что ЕС50 двух клонов, 25С2, 25D2 с улучшенной стабильностью и пониженной агрегацией при связывании с K562-ВСМА составляли 2,594 нМ и 1,891 нМ, соответственно, что по сравнению с 23F10 увеличилось в 3-4 раза.
2.5 Определение специфичности клонов 25С2, 25D2
Специфичность антител 23F10, 25С2 и 25D2 анализировали посредством твердофазного ИФА.
2 мкг/мл рекомбинантного BCMA_Fc человека, BCMA_Fc мыши, TACI_huFc (R&D, #174ТС), BAFF R (R&D, #1162-BR) иммобилизовали на иммунологических планшетах при 4°С в течение ночи. На следующий день добавляли 300 мкл/лунку 2% MPBS на 2 часа, затем добавляли 200 нМ очищенного антитела (в формате scFv) и инкубировали при 37°С в течение 1 часа, трижды промывали PBST (PBS, содержащим 0,05% твин-20) и трижды промывали PBS. Затем добавляли разбавленное в соотношении 1:4000 ПХ-меченого антитела против маркера Flag (sigma, #A8592-1MG), инкубировали в течение 1 ч при 37°С, трижды промывали PBST (PBS, содержащим 0,05%) твин-20) и трижды промывали PBS. Добавляли 100 мкл/лунку субстрата TMBS; окрашивание развивалось в течение 10-15 минут. Реакцию останавливали добавлением 50 мкл 2 М серной кислоты.
Результаты показаны на фиг. 14В, причем антитела 7А12, 23F10, 25С2, 25D2 специфически связывались с ВСМА человека и не связывали TACI человека и BAFF R человека. Среди них связывание антител 25С2, 25D2 с ВСМА мыши было более слабым.
Пример 11. Получение 25С2, 25D2 CAR Т-клеток
В соответствии с процедурой, описанной в примере 8, сконструировали плазмиды химерного рецептора антигена на основе 25С2, 25D2, соответственно.
а. Конструирование плазмиды для химерного рецептора антигена на основе 25С2
Лентивирусную плазмиду PRRLSFN-cPPT.EF-1α-25C2-BBZ, экспрессирующую химерный рецептор антигена второго поколения на основе антитела 25С2, сконструировали с использованием PRRLSIN-cPPT.EF-1α в качестве вектора. Лентивирусную плазмиду PRRLSFN-cPPT.EF-1α-25D2-BBZ, экспрессирующую химерный рецептор антигена второго поколения на основе антитела 25D2, сконструировали с использованием PRRLSIN-cPPT.EF-1α в качестве вектора.
Последовательность 25C2-BBZ состоит из сигнального пептида CD8α (SEQ ID NO: 23), scFv 25C2 (SEQ ID NO: 64), шарнира CD8 (SEQ ID NO: 25), трансмембранной области (SEQ ID NO: 33), внутриклеточного сигнального домена CD137 (SEQ ID NO: 35) и CD3ξ (SEQ ID NO: 31).
Последовательность 25D2-BBZ состоит из сигнального пептида CD8α (SEQ ID NO: 23), scFv 25D2 (SEQ ID NO: 66), шарнира CD8 (SEQ ID NO: 25), трансмембранной области (SEQ ID NO: 33), внутриклеточного сигнального домена CD137 (SEQ ID NO: 35) и CD3ξ (SEQ ID NO: 31).
В соответствии с процедурой, описанной в примере 9, плазмиды PRRLSIN-cPPT.EF-1α-25C2-BBZ, PRRLSIN-cPPT.EF-1α-25D2-BBZ подвергали упаковке лентивируса, инфекции Т-клеток и амплификации, соответственно, получая Т-клетки 25C2-BBZ и 25D2-BBZ, модифицированные химерным рецептором антигена.
Пример 12. Получение CAR Т-клеток, экспрессирующих растворимый PD1
В данном примере получали CAR-T-клетки, экспрессирующие растворимый PD1, с использованием scFv антитела 7А12. Способ получения описан ниже:
1. Последовательность сигнального пептида PD-1 (SEQ ID NO: 68), последовательность внеклеточного сегмента PD-1 (SEQ ID NO: 69) и последовательность СН3 (SEQ ID NO: 70) синтезировали и клонировали в Т-вектор, получая плазмиду T-sPD1-Fc.
Используя плазмиду T-sPD1-Fc в качестве матрицы, верхний праймер 5'-acgcgtcctagcgctaccggtcgccaccatgcagatcccacaggcgccc-3' (SEQ ID NO: 71) и нижний праймер 5'-ctctcggggctgcccaccatacaccagggtttggaactggc-3' (SEQ ID NO: 72) использовали для ПЦР-амплификации, получая последовательность sPD1; а верхний праймер 5'-tatggtgggcagccccgagagccacag-3' (SEQ ID NO: 73) и нижний праймер 5'-aaaattcaaagtctgtttcactttacccggagacagggag-3' (SEQ ID NO: 74) использовали при амплификации, получая sPD1-СН3-фрагмент.
sPD1-СН3-фрагмент и фрагмент 7A12-BBZ (SEQ ID NO: 75) подвергали ПЦР-сплайсингу и амплифицировали, получая sPD1-CH3-7A12-BBZ, причем последовательность 7A12-BBZ состояла из сигнального пептида CD8α (SEQ ID NO: 23), scFv 7A12 (SEQ ID NO: 47), шарнира CD8 (SEQ ID NO: 25), трансмембранной области (SEQ ID NO: 33), внутриклеточного сигнального домена CD137 (SEQ ID NO: 35) и CD3ξ (SEQ ID NO: 31).
Вышеупомянутый сконструированный фрагмент sPD1-CH3-7A12-BBZ содержал сайт расщепления MluI на 5'-конце и сайт расщепления SalI на 3'-конце, которые дважды гидролизовали MluI и SalI и лигировали в идентично дважды гидролизованный вектор PRRLSIN-cPPT.EF-1α, получая плазмиду, экспрессирующую белок sPD-1-СН3 и химерный рецептор антигена, нацеленного на ВСМА.
В соответствии с процедурой, описанной в примере 9, получили Т-клетки SPD-1-7A12-BBZ, экспрессирующие sPD1 и 7A12-BBZ.
Пример 13. Эксперимент по уничтожению клеток in vitro
Эксперименты по уничтожению клеток in vitro выполняли с использованием Т-клеток 25C2-BBZ, Т-клеток 25D2-BBZ, Т-клеток 7A12-BBZ, Т-клеток C11D5.3-BBZ и Т-клеток sPD-1-7A12-BBZ в качестве эффекторных клеток, среди которых C11D5.3-BBZ (SEQ ID NO: 87) содержали CAR второго поколения, полученный с использованием антитела C11D5.3 мыши против ВСМА (см. CN 201580073309.6). Клетки-мишени представляли собой клетки миеломы человека NCI-H929 и В лимфоциты периферической крови множественной миеломы RPMI-8226.
Набор для анализа цитотоксичности без использования радиоактивных материалов CytoTox 96 (Promega) использовали в соответствии с инструкциями к набору для анализа цитотоксичности без использования радиоактивных материалов CytoTox 96.
Эффекторные клетки инокулировали в 96-луночные планшеты в соотношении «эффекторные клетки: клетки-мишени» 3:1, 1:1 или 1:3, и в соответствующие 96-луночные планшеты инокулировали 50 мкл 2×105/мл клеток NCI-H929 и RPMI-8226.
Для каждой группы задали лунки в пяти повторностях, планшеты инкубировали в инкубаторе в течение 18 ч.
Экспериментальные группы и контрольные группы задали следующим образом: экспериментальная группа: каждая клетка-мишень + Т-лимфоциты, экспрессирующие различные химерные рецепторы антигенов; контрольная группа 1: максимальное высвобождение ЛДГ из клеток-мишеней; контрольная группа 2: спонтанное высвобождение ЛДГ из клеток-мишеней; контрольная группа 3: спонтанное высвобождение ЛДГ из эффекторных клеток. Формула для расчета представляла собой: % цитотоксичности = [(экспериментальная группа - группа спонтанного высвобождения из эффекторных клеток - группа спонтанного высвобождения из клеток-мишеней) / (максимум для клеток-мишеней - спонтанное высвобождение из клеток-мишеней)] * 100.
Результаты эксперимента по уничтожению клеток показаны на фигуре 15.
Пример 14. Эксперимент по уничтожению клеток in vivo у мышей
8×106 клеток RPMI-8226 подкожно инокулировали в область правого гребня подвздошной кости мышей B-NDG, и на 18 день средний объем опухоли составлял приблизительно 243 мм3; таким образом, получили модель подкожного ксенотрансплантата у мышей B-NDG с В-лимфоцитами периферической крови множественной миеломы RPMI-8226.
Мышей с модельными подкожными ксенотрансплантатами разделили на 3 группы (по 4 в каждой группе) и вводили им 25C2-BBZ, 25D2-BBZ и нетрансфицированные Т-клетки (UTD) в дозе 5×106, соответственно. Результаты показаны в следующей таблице. На 32 день и 36 день после инокуляции опухолевых клеток у всех 4 мышей в экспериментальных группах 25C2-BBZ и 25D2-BBZ наблюдали регресс опухолей.
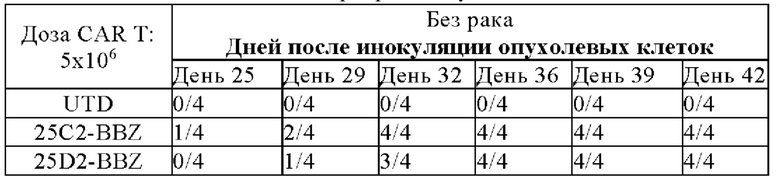
Мышей с модельными подкожными ксенотрансплантатами разделили на 3 группы (по 4 в каждой группе) и вводили им 25C2-BBZ, 25D2-BBZ, C11D5.3-BBZ, 7A12-BBZ и нетрансфицированные Т-клетки (UTD) в дозе 1×106 CAR Т-клеток на инъекцию. Регресс опухоли показан в следующей таблице и на фигуре 16.
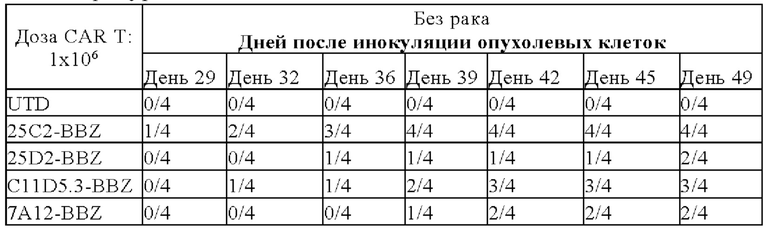
Все документы, упомянутые в настоящей заявке, полностью включены в настоящий документ посредством ссылок, как если бы каждый документ был отдельно упомянут в качестве ссылки. Кроме того, следует понимать, что специалист в данной области техники после прочтения вышеуказанной информации о настоящем изобретении может внести различные модификации и изменения, и эти эквивалентные формы также будут входить в рамки настоящего изобретения, заданные прилагаемой формулой изобретения.
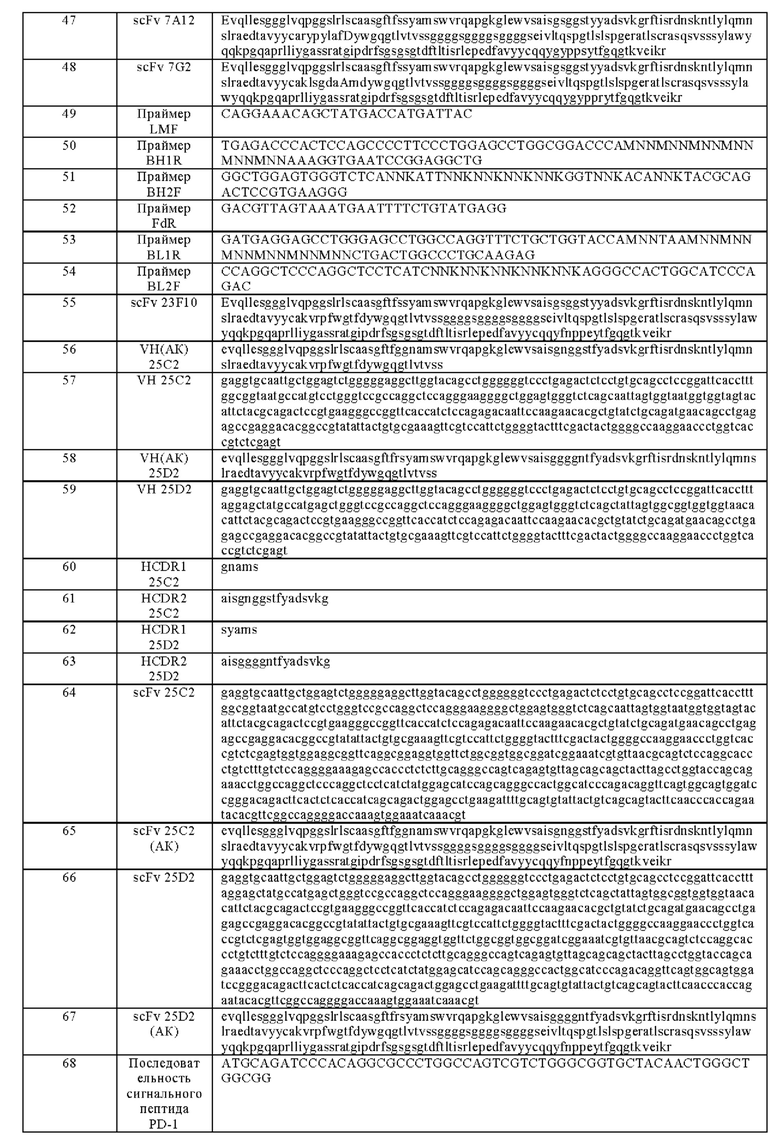
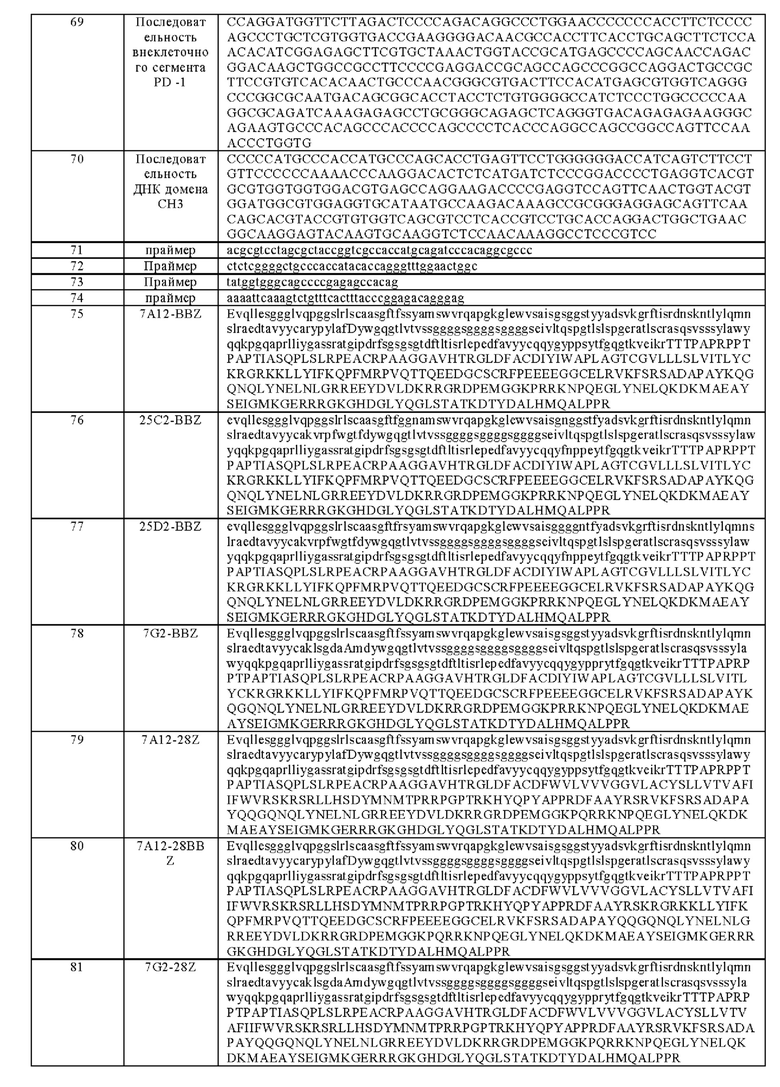
--->
Список последовательностей
<110> CARSGEN THERAPEUTICS CO., LTD.
<120> АНТИТЕЛО, НАЦЕЛЕННОЕ НА BCMA, И ЕГО ПРИМЕНЕНИЕ
<130> P2018-0137
<150> CN201710058581.8
<151> 23.01.2017
<150> CN201710920346.7
<151> 30.09.2017
<160> 87
<170> Версия PatentIn 3.5
<210> 1
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR1 7A12, 7G2, 23F10
<400> 1
Ser Tyr Ala Met Ser
1 5
<210> 2
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR1 7A12, 7G2, 23F10
<400> 2
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 3
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR3 7A12
<400> 3
Tyr Pro Tyr Leu Ala Phe Asp Tyr
1 5
<210> 4
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR3 7G2
<400> 4
Leu Ser Gly Asp Ala Ala Met Asp Tyr
1 5
<210> 5
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR3 23F10
<400> 5
Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr
1 5 10
<210> 6
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LCDR1 7A12, 7G2, 23F10
<400> 6
Arg Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala
1 5 10
<210> 7
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LCDR2 7A12, 7G2, 23F10
<400> 7
Gly Ala Ser Ser Arg Ala Thr
1 5
<210> 8
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LCDR3 7A12
<400> 8
Gln Gln Tyr Gly Tyr Pro Pro Ser Tyr
1 5
<210> 9
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LCDR3 7G2
<400> 9
Gln Gln Tyr Gly Tyr Pro Pro Arg Tyr
1 5
<210> 10
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LCDR3 23F10
<400> 10
Gln Gln Tyr Phe Asn Pro Pro Glu Tyr
1 5
<210> 11
<211> 109
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность вариабельной области легкой цепи 7A12
<400> 11
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Tyr Pro Pro
85 90 95
Ser Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 12
<211> 327
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность вариабельной области легкой цепи 7A12
<400> 12
gaaatcgtgt taacgcagtc tccaggcacc ctgtctttgt ctccagggga aagagccacc 60
ctctcttgca gggccagtca gagtgttagc agcagctact tagcctggta ccagcagaaa 120
cctggccagg ctcccaggct cctcatctat ggagcatcca gcagggccac tggcatccca 180
gacaggttca gtggcagtgg atccgggaca gacttcactc tcaccatcag cagactggag 240
cctgaagatt ttgcagtgta ttactgtcag cagtacggtt acccaccatc ttacacgttc 300
ggccagggga ccaaagtgga aatcaaa 327
<210> 13
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность вариабельной области тяжелой цепи 7A12
<400> 13
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Pro Tyr Leu Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 14
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность вариабельной области тяжелой цепи 7A12
<400> 14
gaggtgcaat tgctggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctccggatt cacctttagc agttatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcagatga acagcctgag agccgaggac acggccgtat attactgtgc gcgttaccca 300
tacctggcat tcgactactg gggccaagga accctggtca ccgtctcgag t 351
<210> 15
<211> 109
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность вариабельной области легкой цепи 7G2
<400> 15
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Tyr Pro Pro
85 90 95
Arg Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 16
<211> 327
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность вариабельной области легкой цепи 7G2
<400> 16
gaaatcgtgt taacgcagtc tccaggcacc ctgtctttgt ctccagggga aagagccacc 60
ctctcttgca gggccagtca gagtgttagc agcagctact tagcctggta ccagcagaaa 120
cctggccagg ctcccaggct cctcatctat ggagcatcca gcagggccac tggcatccca 180
gacaggttca gtggcagtgg atccgggaca gacttcactc tcaccatcag cagactggag 240
cctgaagatt ttgcagtgta ttactgtcag cagtacggtt acccaccaag atacacgttc 300
ggccagggga ccaaagtgga aatcaaa 327
<210> 17
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность вариабельной области тяжелой цепи 7G2
<400> 17
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Leu Ser Gly Asp Ala Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 18
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность вариабельной области тяжелой цепи 7G2
<400> 18
gaggtgcaat tgctggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctccggatt cacctttagc agttatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcagatga acagcctgag agccgaggac acggccgtat attactgtgc gaaactgtct 300
ggtgatgcag caatggacta ctggggccaa ggaaccctgg tcaccgtctc gagt 354
<210> 19
<211> 109
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность вариабельной области легкой цепи 23F10
<400> 19
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Phe Asn Pro Pro
85 90 95
Glu Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 20
<211> 327
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность вариабельной области легкой цепи 23F10
<400> 20
gaaatcgtgt taacgcagtc tccaggcacc ctgtctttgt ctccagggga aagagccacc 60
ctctcttgca gggccagtca gagtgttagc agcagctact tagcctggta ccagcagaaa 120
cctggccagg ctcccaggct cctcatctat ggagcatcca gcagggccac tggcatccca 180
gacaggttca gtggcagtgg atccgggaca gacttcactc tcaccatcag cagactggag 240
cctgaagatt ttgcagtgta ttactgtcag cagtacttca acccaccaga atacacgttc 300
ggccagggga ccaaagtgga aatcaaa 327
<210> 21
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность вариабельной области тяжелой цепи 23F10
<400> 21
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 22
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность вариабельной области тяжелой цепи 23F10
<400> 22
gaggtgcaat tgctggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctccggatt cacctttagc agttatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcagatga acagcctgag agccgaggac acggccgtat attactgtgc gaaagttcgt 300
ccattctggg gtactttcga ctactggggc caaggaaccc tggtcaccgt ctcgagt 357
<210> 23
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность сигнального пептида CD8-альфа
<400> 23
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro
20
<210> 24
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность сигнального пептида CD8-альфа
<400> 24
atggccttac cagtgaccgc cttgctcctg ccgctggcct tgctgctcca cgccgccagg 60
ccg 63
<210> 25
<211> 45
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность шарнира CD8
<400> 25
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp
35 40 45
<210> 26
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность шарнира CD8
<400> 26
accacgacgc cagcgccgcg accaccaaca ccggcgccca ccatcgcgtc gcagcccctg 60
tccctgcgcc cagaggcgtg ccggccagcg gcggggggcg cagtgcacac gagggggctg 120
gacttcgcct gtgat 135
<210> 27
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность трансмембранной области CD28
<400> 27
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val
20 25
<210> 28
<211> 81
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность трансмембранной области CD28
<400> 28
ttttgggtgc tggtggtggt tggtggagtc ctggcttgct atagcttgct agtaacagtg 60
gcctttatta ttttctgggt g 81
<210> 29
<211> 41
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность внутриклеточной области CD28
<400> 29
Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr
1 5 10 15
Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro
20 25 30
Pro Arg Asp Phe Ala Ala Tyr Arg Ser
35 40
<210> 30
<211> 123
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность внутриклеточной области CD28
<400> 30
aggagtaaga ggagcaggct cctgcacagt gactacatga acatgactcc ccgccgcccc 60
gggccaaccc gcaagcatta ccagccctat gccccaccac gcgacttcgc agcctatcgc 120
tcc 123
<210> 31
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность домена CD3Z
<400> 31
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Gln Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln
50 55 60
Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu
65 70 75 80
Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr
85 90 95
Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro
100 105 110
Arg
<210> 32
<211> 339
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность домена CD3Z
<400> 32
agagtgaagt tcagcaggag cgcagacgcc cccgcgtacc agcagggcca gaaccagctc 60
tataacgagc tcaatctagg acgaagagag gagtacgatg ttttggacaa gagacgtggc 120
cgggaccctg agatgggggg aaagccgcag agaaggaaga accctcagga aggcctgtac 180
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 240
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 300
acctacgacg cccttcacat gcaggccctg ccccctcgc 339
<210> 33
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность трансмембранной области CD8
<400> 33
Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu
1 5 10 15
Ser Leu Val Ile Thr
20
<210> 34
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность трансмембранной области CD8
<400> 34
atctacatct gggcgccctt ggccgggact tgtggggtcc ttctcctgtc actggttatc 60
acc 63
<210> 35
<211> 42
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность внутриклеточной области CD137
<400> 35
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
1 5 10 15
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
20 25 30
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
35 40
<210> 36
<211> 126
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность внутриклеточной области CD137
<400> 36
aaacggggca gaaagaaact cctgtatata ttcaaacaac catttatgag accagtacaa 60
actactcaag aggaagatgg ctgtagctgc cgatttccag aagaagaaga aggaggatgt 120
gaactg 126
<210> 37
<211> 184
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность BCMA
<400> 37
Met Leu Gln Met Ala Gly Gln Cys Ser Gln Asn Glu Tyr Phe Asp Ser
1 5 10 15
Leu Leu His Ala Cys Ile Pro Cys Gln Leu Arg Cys Ser Ser Asn Thr
20 25 30
Pro Pro Leu Thr Cys Gln Arg Tyr Cys Asn Ala Ser Val Thr Asn Ser
35 40 45
Val Lys Gly Thr Asn Ala Ile Leu Trp Thr Cys Leu Gly Leu Ser Leu
50 55 60
Ile Ile Ser Leu Ala Val Phe Val Leu Met Phe Leu Leu Arg Lys Ile
65 70 75 80
Asn Ser Glu Pro Leu Lys Asp Glu Phe Lys Asn Thr Gly Ser Gly Leu
85 90 95
Leu Gly Met Ala Asn Ile Asp Leu Glu Lys Ser Arg Thr Gly Asp Glu
100 105 110
Ile Ile Leu Pro Arg Gly Leu Glu Tyr Thr Val Glu Glu Cys Thr Cys
115 120 125
Glu Asp Cys Ile Lys Ser Lys Pro Lys Val Asp Ser Asp His Cys Phe
130 135 140
Pro Leu Pro Ala Met Glu Glu Gly Ala Thr Ile Leu Val Thr Thr Lys
145 150 155 160
Thr Asn Asp Tyr Cys Lys Ser Leu Pro Ala Ala Leu Ser Ala Thr Glu
165 170 175
Ile Glu Lys Ser Ile Ser Ala Arg
180
<210> 38
<211> 54
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Внеклеточный сегмент Met1-Ala54 BCMA человека
<400> 38
Met Leu Gln Met Ala Gly Gln Cys Ser Gln Asn Glu Tyr Phe Asp Ser
1 5 10 15
Leu Leu His Ala Cys Ile Pro Cys Gln Leu Arg Cys Ser Ser Asn Thr
20 25 30
Pro Pro Leu Thr Cys Gln Arg Tyr Cys Asn Ala Ser Val Thr Asn Ser
35 40 45
Val Lys Gly Thr Asn Ala
50
<210> 39
<211> 162
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность внеклеточного сегмента Met1-Ala54 BCMA
человека
<400> 39
atgctgcaga tggccggcca gtgcagccag aacgagtact tcgacagcct gctgcacgcc 60
tgcatcccct gccagctgcg gtgcagcagc aacacccccc ccctgacctg ccagcggtac 120
tgcaacgcca gcgtgaccaa cagcgtgaag ggcaccaacg cc 162
<210> 40
<211> 283
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> BCMA_huFc
<400> 40
Met Leu Gln Met Ala Gly Gln Cys Ser Gln Asn Glu Tyr Phe Asp Ser
1 5 10 15
Leu Leu His Ala Cys Ile Pro Cys Gln Leu Arg Cys Ser Ser Asn Thr
20 25 30
Pro Pro Leu Thr Cys Gln Arg Tyr Cys Asn Ala Ser Val Thr Asn Ser
35 40 45
Val Lys Gly Thr Asn Ala Gly Ser Asp Lys Thr His Thr Cys Pro Pro
50 55 60
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
65 70 75 80
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
85 90 95
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
100 105 110
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
115 120 125
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
130 135 140
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
145 150 155 160
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
165 170 175
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
180 185 190
Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe
195 200 205
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
210 215 220
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
225 230 235 240
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
245 250 255
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
260 265 270
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
275 280
<210> 41
<211> 849
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность BCMA_huFc
<400> 41
atgctgcaga tggccggcca gtgcagccag aacgagtact tcgacagcct gctgcacgcc 60
tgcatcccct gccagctgcg gtgcagcagc aacacccccc ccctgacctg ccagcggtac 120
tgcaacgcca gcgtgaccaa cagcgtgaag ggcaccaacg ccggatccga caaaactcac 180
acatgcccac cgtgcccagc acctgaactc ctggggggac cgtcagtctt cctcttcccc 240
ccaaaaccca aggacaccct catgatctcc cggacccctg aggtcacatg cgtggtggtg 300
gacgtgagcc acgaagaccc tgaggtcaag ttcaactggt acgtggacgg cgtggaggtg 360
cataatgcca agacaaagcc gcgggaggag cagtacaaca gcacgtaccg tgtggtcagc 420
gtcctcaccg tcctgcacca ggactggctg aatggcaagg agtacaagtg caaggtctcc 480
aacaaagccc tcccagcccc catcgagaaa accatctcca aagccaaagg gcagccccga 540
gaaccacagg tgtacaccct gcccccatcc cgggatgagc tgaccaagaa ccaggtcagc 600
ctgtggtgcc tggtcaaagg cttctatccc agcgacatcg ccgtggagtg ggagagcaat 660
gggcagccgg agaacaacta caagaccacg cctcccgtgc tggactccga cggctccttc 720
ttcctctata gcaagctcac cgtggacaag agcaggtggc agcaggggaa cgtcttctca 780
tgctccgtga tgcatgaggc tctgcacaac cactacacgc agaagagcct ctccctgtct 840
ccgggtaaa 849
<210> 42
<211> 281
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> BCMA_muFc
<400> 42
Met Leu Gln Met Ala Gly Gln Cys Ser Gln Asn Glu Tyr Phe Asp Ser
1 5 10 15
Leu Leu His Ala Cys Ile Pro Cys Gln Leu Arg Cys Ser Ser Asn Thr
20 25 30
Pro Pro Leu Thr Cys Gln Arg Tyr Cys Asn Ala Ser Val Thr Asn Ser
35 40 45
Val Lys Gly Thr Asn Ala Gly Ser Arg Asp Cys Gly Cys Lys Pro Cys
50 55 60
Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys
65 70 75 80
Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val
85 90 95
Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe
100 105 110
Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu
115 120 125
Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met His
130 135 140
Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala
145 150 155 160
Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg
165 170 175
Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met
180 185 190
Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro
195 200 205
Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn
210 215 220
Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val
225 230 235 240
Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr
245 250 255
Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn His His Thr Glu
260 265 270
Lys Ser Leu Ser His Ser Pro Gly Lys
275 280
<210> 43
<211> 843
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность BCMA_muFc
<400> 43
atgctgcaga tggccggcca gtgcagccag aacgagtact tcgacagcct gctgcacgcc 60
tgcatcccct gccagctgcg gtgcagcagc aacacccccc ccctgacctg ccagcggtac 120
tgcaacgcca gcgtgaccaa cagcgtgaag ggcaccaacg ccggatccag ggattgtggt 180
tgtaagcctt gcatatgtac agtcccagaa gtatcatctg tcttcatctt ccccccaaag 240
cccaaggatg tgctcaccat tactctgact cctaaggtca cgtgtgttgt ggtagacatc 300
agcaaggatg atcccgaggt ccagttcagc tggtttgtag atgatgtgga ggtgcacaca 360
gctcagacgc aaccccggga ggagcagttc aacagcactt tccgctcagt cagtgaactt 420
cccatcatgc accaggactg gctcaatggc aaggagttca aatgcagggt caacagtgca 480
gctttccctg cccccatcga gaaaaccatc tccaaaacca aaggcagacc gaaggctcca 540
caggtgtaca ccattccacc tcccaaggag cagatggcca aggataaagt cagtctgacc 600
tgcatgataa cagacttctt ccctgaagac attactgtgg agtggcagtg gaatgggcag 660
ccagcggaga actacaagaa cactcagccc atcatggaca cagatggctc ttacttcgtc 720
tacagcaagc tcaatgtgca gaagagcaac tgggaggcag gaaatacttt cacctgctct 780
gtgttacatg agggcctgca caaccaccat actgagaaga gcctctccca ctctcctggt 840
aaa 843
<210> 44
<211> 589
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность BCMA человека с внедренными сайтами
рестрикции MluI, SalI
<400> 44
acgcgtccta gcgctaccgg tcgccaccat gttgcagatg gctgggcagt gctcccaaaa 60
tgaatatttt gacagtttgt tgcatgcttg cataccttgt caacttcgat gttcttctaa 120
tactcctcct ctaacatgtc agcgttattg taatgcaagt gtgaccaatt cagtgaaagg 180
aacgaatgcg attctctgga cctgtttggg actgagctta ataatttctt tggcagtttt 240
cgtgctaatg tttttgctaa ggaagataaa ctctgaacca ttaaaggacg agtttaaaaa 300
cacaggatca ggtctcctgg gcatggctaa cattgacctg gaaaagagca ggactggtga 360
tgaaattatt cttccgagag gcctcgagta cacggtggaa gaatgcacct gtgaagactg 420
catcaagagc aaaccgaagg tcgactctga ccattgcttt ccactcccag ctatggagga 480
aggcgcaacc attcttgtca ccacgaaaac gaatgactat tgcaagagcc tgccagctgc 540
tttgagtgct acggagatag agaaatcaat ttctgctagg taagtcgac 589
<210> 45
<211> 365
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> APRIL_huFc
<400> 45
His Ser Val Leu His Leu Val Pro Ile Asn Ala Thr Ser Lys Asp Asp
1 5 10 15
Ser Asp Val Thr Glu Val Met Trp Gln Pro Ala Leu Arg Arg Gly Arg
20 25 30
Gly Leu Gln Ala Gln Gly Tyr Gly Val Arg Ile Gln Asp Ala Gly Val
35 40 45
Tyr Leu Leu Tyr Ser Gln Val Leu Phe Gln Asp Val Thr Phe Thr Met
50 55 60
Gly Gln Val Val Ser Arg Glu Gly Gln Gly Arg Gln Glu Thr Leu Phe
65 70 75 80
Arg Cys Ile Arg Ser Met Pro Ser His Pro Asp Arg Ala Tyr Asn Ser
85 90 95
Cys Tyr Ser Ala Gly Val Phe His Leu His Gln Gly Asp Ile Leu Ser
100 105 110
Val Ile Ile Pro Arg Ala Arg Ala Lys Leu Asn Leu Ser Pro His Gly
115 120 125
Thr Phe Leu Gly Phe Val Lys Leu Gly Ser Asp Lys Thr His Thr Cys
130 135 140
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
145 150 155 160
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
165 170 175
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
180 185 190
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
195 200 205
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
210 215 220
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
225 230 235 240
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
245 250 255
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
260 265 270
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys
275 280 285
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
290 295 300
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
305 310 315 320
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
325 330 335
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
340 345 350
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
355 360 365
<210> 46
<211> 1095
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность APRIL_huFc
<400> 46
cacagcgtgc tgcacctggt gcccatcaac gccaccagca aggacgacag cgacgtgacc 60
gaggtgatgt ggcagcccgc cctgcggcgg ggccggggcc tgcaggccca gggctacggc 120
gtgcggatcc aggacgccgg cgtgtacctg ctgtacagcc aggtgctgtt ccaggacgtg 180
accttcacca tgggccaggt ggtgagccgg gagggccagg gccggcagga gaccctgttc 240
cggtgcatcc ggagcatgcc cagccacccc gaccgggcct acaacagctg ctacagcgcc 300
ggcgtgttcc acctgcacca gggcgacatc ctgagcgtga tcatcccccg ggcccgggcc 360
aagctgaacc tgagccccca cggcaccttc ctgggcttcg tgaagctggg atccgacaaa 420
actcacacat gcccaccgtg cccagcacct gaactcctgg ggggaccgtc agtcttcctc 480
ttccccccaa aacccaagga caccctcatg atctcccgga cccctgaggt cacatgcgtg 540
gtggtggacg tgagccacga agaccctgag gtcaagttca actggtacgt ggacggcgtg 600
gaggtgcata atgccaagac aaagccgcgg gaggagcagt acaacagcac gtaccgtgtg 660
gtcagcgtcc tcaccgtcct gcaccaggac tggctgaatg gcaaggagta caagtgcaag 720
gtctccaaca aagccctccc agcccccatc gagaaaacca tctccaaagc caaagggcag 780
ccccgagaac cacaggtgta caccctgccc ccatcccggg atgagctgac caagaaccag 840
gtcagcctgt ggtgcctggt caaaggcttc tatcccagcg acatcgccgt ggagtgggag 900
agcaatgggc agccggagaa caactacaag accacgcctc ccgtgctgga ctccgacggc 960
tccttcttcc tctatagcaa gctcaccgtg gacaagagca ggtggcagca ggggaacgtc 1020
ttctcatgct ccgtgatgca tgaggctctg cacaaccact acacgcagaa gagcctctcc 1080
ctgtctccgg gtaaa 1095
<210> 47
<211> 242
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 47 7A12 scFv
<400> 47
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Pro Tyr Leu Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser
130 135 140
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser
145 150 155 160
Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala
165 170 175
Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro
180 185 190
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
195 200 205
Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr
210 215 220
Gly Tyr Pro Pro Ser Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
225 230 235 240
Lys Arg
<210> 48
<211> 243
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv 7G2
<400> 48
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Leu Ser Gly Asp Ala Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu
130 135 140
Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln
145 150 155 160
Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
165 170 175
Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile
180 185 190
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
195 200 205
Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln
210 215 220
Tyr Gly Tyr Pro Pro Arg Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu
225 230 235 240
Ile Lys Arg
<210> 49
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер LMF
<400> 49
caggaaacag ctatgaccat gattac 26
<210> 50
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер BH1R
<220>
<221> прочее
<222> (44)..(45)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (47)..(48)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (50)..(51)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (53)..(54)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (56)..(57)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (59)..(60)
<223> n представляет собой a, c, g или t
<400> 50
tgagacccac tccagcccct tccctggagc ctggcggacc camnnmnnmn nmnnmnnmnn 60
aaaggtgaat ccggaggctg 80
<210> 51
<211> 67
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер BH2F
<220>
<221> прочее
<222> (18)..(19)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (24)..(25)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (27)..(28)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (30)..(31)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (33)..(34)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (39)..(40)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (45)..(46)
<223> n представляет собой a, c, g или t
<400> 51
ggctggagtg ggtctcannk attnnknnkn nknnkggtnn kacannktac gcagactccg 60
tgaaggg 67
<210> 52
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер FdR
<400> 52
gacgttagta aatgaatttt ctgtatgagg 30
<210> 53
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер BL1R
<220>
<221> прочее
<222> (44)..(45)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (50)..(51)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (53)..(54)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (56)..(57)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (59)..(60)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (62)..(63)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (65)..(66)
<223> n представляет собой a, c, g или t
<400> 53
gatgaggagc ctgggagcct ggccaggttt ctgctggtac camnntaamn nmnnmnnmnn 60
mnnmnnctga ctggccctgc aagag 85
<210> 54
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер BL2F
<220>
<221> прочее
<222> (23)..(24)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (26)..(27)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (29)..(30)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (32)..(33)
<223> n представляет собой a, c, g или t
<220>
<221> прочее
<222> (35)..(36)
<223> n представляет собой a, c, g или t
<400> 54
ccaggctccc aggctcctca tcnnknnknn knnknnkagg gccactggca tcccagac 58
<210> 55
<211> 244
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv 23F10
<400> 55
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr
130 135 140
Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser
145 150 155 160
Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
165 170 175
Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly
180 185 190
Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
195 200 205
Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
210 215 220
Gln Tyr Phe Asn Pro Pro Glu Tyr Thr Phe Gly Gln Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Arg
<210> 56
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH(АК) 25C2
<400> 56
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Gly Asn
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Asn Gly Gly Ser Thr Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 57
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH 25C2
<400> 57
gaggtgcaat tgctggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctccggatt cacctttggc ggtaatgcca tgtcctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagca attagtggta atggtggtag tacattctac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcagatga acagcctgag agccgaggac acggccgtat attactgtgc gaaagttcgt 300
ccattctggg gtactttcga ctactggggc caaggaaccc tggtcaccgt ctcgagt 357
<210> 58
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH(АК) 25D2
<400> 58
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Gly Gly Gly Asn Thr Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 59
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH 25D2
<400> 59
gaggtgcaat tgctggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctccggatt cacctttagg agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggcg gtggtggtaa cacattctac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcagatga acagcctgag agccgaggac acggccgtat attactgtgc gaaagttcgt 300
ccattctggg gtactttcga ctactggggc caaggaaccc tggtcaccgt ctcgagt 357
<210> 60
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR1 25C2
<400> 60
Gly Asn Ala Met Ser
1 5
<210> 61
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR2 25C2
<400> 61
Ala Ile Ser Gly Asn Gly Gly Ser Thr Phe Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 62
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR1 25D2
<400> 62
Ser Tyr Ala Met Ser
1 5
<210> 63
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR2 25D2
<400> 63
Ala Ile Ser Gly Gly Gly Gly Asn Thr Phe Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 64
<211> 732
<212> ДНК
<213> Искусственная последовательность
<220>
<223> scFv 25C2
<400> 64
gaggtgcaat tgctggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctccggatt cacctttggc ggtaatgcca tgtcctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagca attagtggta atggtggtag tacattctac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcagatga acagcctgag agccgaggac acggccgtat attactgtgc gaaagttcgt 300
ccattctggg gtactttcga ctactggggc caaggaaccc tggtcaccgt ctcgagtggt 360
ggaggcggtt caggcggagg tggttctggc ggtggcggat cggaaatcgt gttaacgcag 420
tctccaggca ccctgtcttt gtctccaggg gaaagagcca ccctctcttg cagggccagt 480
cagagtgtta gcagcagcta cttagcctgg taccagcaga aacctggcca ggctcccagg 540
ctcctcatct atggagcatc cagcagggcc actggcatcc cagacaggtt cagtggcagt 600
ggatccggga cagacttcac tctcaccatc agcagactgg agcctgaaga ttttgcagtg 660
tattactgtc agcagtactt caacccacca gaatacacgt tcggccaggg gaccaaagtg 720
gaaatcaaac gt 732
<210> 65
<211> 244
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv 25C2 (АК)
<400> 65
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Gly Asn
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Asn Gly Gly Ser Thr Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr
130 135 140
Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser
145 150 155 160
Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
165 170 175
Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly
180 185 190
Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
195 200 205
Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
210 215 220
Gln Tyr Phe Asn Pro Pro Glu Tyr Thr Phe Gly Gln Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Arg
<210> 66
<211> 732
<212> ДНК
<213> Искусственная последовательность
<220>
<223> scFv 25D2
<400> 66
gaggtgcaat tgctggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctccggatt cacctttagg agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggcg gtggtggtaa cacattctac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcagatga acagcctgag agccgaggac acggccgtat attactgtgc gaaagttcgt 300
ccattctggg gtactttcga ctactggggc caaggaaccc tggtcaccgt ctcgagtggt 360
ggaggcggtt caggcggagg tggttctggc ggtggcggat cggaaatcgt gttaacgcag 420
tctccaggca ccctgtcttt gtctccaggg gaaagagcca ccctctcttg cagggccagt 480
cagagtgtta gcagcagcta cttagcctgg taccagcaga aacctggcca ggctcccagg 540
ctcctcatct atggagcatc cagcagggcc actggcatcc cagacaggtt cagtggcagt 600
ggatccggga cagacttcac tctcaccatc agcagactgg agcctgaaga ttttgcagtg 660
tattactgtc agcagtactt caacccacca gaatacacgt tcggccaggg gaccaaagtg 720
gaaatcaaac gt 732
<210> 67
<211> 244
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv 25D2 (АК)
<400> 67
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Gly Gly Gly Asn Thr Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr
130 135 140
Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser
145 150 155 160
Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
165 170 175
Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly
180 185 190
Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
195 200 205
Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
210 215 220
Gln Tyr Phe Asn Pro Pro Glu Tyr Thr Phe Gly Gln Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Arg
<210> 68
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность сигнального пептида PD-1
<400> 68
atgcagatcc cacaggcgcc ctggccagtc gtctgggcgg tgctacaact gggctggcgg 60
<210> 69
<211> 450
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность внеклеточного сегмента PD-1
<400> 69
ccaggatggt tcttagactc cccagacagg ccctggaacc cccccacctt ctccccagcc 60
ctgctcgtgg tgaccgaagg ggacaacgcc accttcacct gcagcttctc caacacatcg 120
gagagcttcg tgctaaactg gtaccgcatg agccccagca accagacgga caagctggcc 180
gccttccccg aggaccgcag ccagcccggc caggactgcc gcttccgtgt cacacaactg 240
cccaacgggc gtgacttcca catgagcgtg gtcagggccc ggcgcaatga cagcggcacc 300
tacctctgtg gggccatctc cctggccccc aaggcgcaga tcaaagagag cctgcgggca 360
gagctcaggg tgacagagag aagggcagaa gtgcccacag cccaccccag cccctcaccc 420
aggccagccg gccagttcca aaccctggtg 450
<210> 70
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК-последовательность домена CH3
<400> 70
cccccatgcc caccatgccc agcacctgag ttcctggggg gaccatcagt cttcctgttc 60
cccccaaaac ccaaggacac tctcatgatc tcccggaccc ctgaggtcac gtgcgtggtg 120
gtggacgtga gccaggaaga ccccgaggtc cagttcaact ggtacgtgga tggcgtggag 180
gtgcataatg ccaagacaaa gccgcgggag gagcagttca acagcacgta ccgtgtggtc 240
agcgtcctca ccgtcctgca ccaggactgg ctgaacggca aggagtacaa gtgcaaggtc 300
tccaacaaag gcctcccgtc c 321
<210> 71
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 71
acgcgtccta gcgctaccgg tcgccaccat gcagatccca caggcgccc 49
<210> 72
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 72
ctctcggggc tgcccaccat acaccagggt ttggaactgg c 41
<210> 73
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 73
tatggtgggc agccccgaga gccacag 27
<210> 74
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 74
aaaattcaaa gtctgtttca ctttacccgg agacagggag 40
<210> 75
<211> 465
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 7A12-BBZ
<400> 75
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Pro Tyr Leu Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser
130 135 140
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser
145 150 155 160
Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala
165 170 175
Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro
180 185 190
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
195 200 205
Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr
210 215 220
Gly Tyr Pro Pro Ser Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
225 230 235 240
Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr
245 250 255
Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala
260 265 270
Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile
275 280 285
Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu Ser
290 295 300
Leu Val Ile Thr Leu Tyr Cys Lys Arg Gly Arg Lys Lys Leu Leu Tyr
305 310 315 320
Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu
325 330 335
Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu
340 345 350
Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln
355 360 365
Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu
370 375 380
Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly
385 390 395 400
Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln
405 410 415
Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu
420 425 430
Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr
435 440 445
Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro
450 455 460
Arg
465
<210> 76
<211> 467
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 25C2-BBZ
<400> 76
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Gly Asn
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Asn Gly Gly Ser Thr Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr
130 135 140
Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser
145 150 155 160
Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
165 170 175
Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly
180 185 190
Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
195 200 205
Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
210 215 220
Gln Tyr Phe Asn Pro Pro Glu Tyr Thr Phe Gly Gln Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala
245 250 255
Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg
260 265 270
Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys
275 280 285
Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu
290 295 300
Leu Ser Leu Val Ile Thr Leu Tyr Cys Lys Arg Gly Arg Lys Lys Leu
305 310 315 320
Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln
325 330 335
Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly
340 345 350
Cys Glu Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr
355 360 365
Lys Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg
370 375 380
Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met
385 390 395 400
Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu
405 410 415
Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys
420 425 430
Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu
435 440 445
Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu
450 455 460
Pro Pro Arg
465
<210> 77
<211> 467
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 25D2-BBZ
<400> 77
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Gly Gly Gly Asn Thr Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr
130 135 140
Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser
145 150 155 160
Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
165 170 175
Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly
180 185 190
Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
195 200 205
Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
210 215 220
Gln Tyr Phe Asn Pro Pro Glu Tyr Thr Phe Gly Gln Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala
245 250 255
Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg
260 265 270
Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys
275 280 285
Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu
290 295 300
Leu Ser Leu Val Ile Thr Leu Tyr Cys Lys Arg Gly Arg Lys Lys Leu
305 310 315 320
Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln
325 330 335
Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly
340 345 350
Cys Glu Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr
355 360 365
Lys Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg
370 375 380
Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met
385 390 395 400
Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu
405 410 415
Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys
420 425 430
Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu
435 440 445
Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu
450 455 460
Pro Pro Arg
465
<210> 78
<211> 466
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 7G2-BBZ
<400> 78
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Leu Ser Gly Asp Ala Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu
130 135 140
Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln
145 150 155 160
Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
165 170 175
Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile
180 185 190
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
195 200 205
Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln
210 215 220
Tyr Gly Tyr Pro Pro Arg Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu
225 230 235 240
Ile Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro
245 250 255
Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro
260 265 270
Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp
275 280 285
Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu
290 295 300
Ser Leu Val Ile Thr Leu Tyr Cys Lys Arg Gly Arg Lys Lys Leu Leu
305 310 315 320
Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu
325 330 335
Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys
340 345 350
Glu Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys
355 360 365
Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu
370 375 380
Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly
385 390 395 400
Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu
405 410 415
Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly
420 425 430
Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser
435 440 445
Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro
450 455 460
Pro Arg
465
<210> 79
<211> 468
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 7A12-28Z
<400> 79
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Pro Tyr Leu Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser
130 135 140
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser
145 150 155 160
Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala
165 170 175
Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro
180 185 190
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
195 200 205
Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr
210 215 220
Gly Tyr Pro Pro Ser Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
225 230 235 240
Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr
245 250 255
Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala
260 265 270
Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Phe
275 280 285
Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu
290 295 300
Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg
305 310 315 320
Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro
325 330 335
Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala
340 345 350
Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr
355 360 365
Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg
370 375 380
Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met
385 390 395 400
Gly Gly Lys Pro Gln Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
405 410 415
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
420 425 430
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
435 440 445
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala
450 455 460
Leu Pro Pro Arg
465
<210> 80
<211> 510
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 7A12-28BBZ
<400> 80
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Pro Tyr Leu Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser
130 135 140
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser
145 150 155 160
Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala
165 170 175
Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro
180 185 190
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
195 200 205
Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr
210 215 220
Gly Tyr Pro Pro Ser Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
225 230 235 240
Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr
245 250 255
Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala
260 265 270
Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Phe
275 280 285
Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu
290 295 300
Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg
305 310 315 320
Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro
325 330 335
Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala
340 345 350
Tyr Arg Ser Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln
355 360 365
Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser
370 375 380
Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys
385 390 395 400
Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln
405 410 415
Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu
420 425 430
Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Gln Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 81
<211> 469
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 7G2-28Z
<400> 81
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Leu Ser Gly Asp Ala Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu
130 135 140
Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln
145 150 155 160
Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
165 170 175
Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile
180 185 190
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
195 200 205
Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln
210 215 220
Tyr Gly Tyr Pro Pro Arg Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu
225 230 235 240
Ile Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro
245 250 255
Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro
260 265 270
Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp
275 280 285
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
290 295 300
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser
305 310 315 320
Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly
325 330 335
Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala
340 345 350
Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala
355 360 365
Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg
370 375 380
Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu
385 390 395 400
Met Gly Gly Lys Pro Gln Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr
405 410 415
Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly
420 425 430
Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln
435 440 445
Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln
450 455 460
Ala Leu Pro Pro Arg
465
<210> 82
<211> 511
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 7G2-28BBZ
<400> 82
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Leu Ser Gly Asp Ala Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu
130 135 140
Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln
145 150 155 160
Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
165 170 175
Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile
180 185 190
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
195 200 205
Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln
210 215 220
Tyr Gly Tyr Pro Pro Arg Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu
225 230 235 240
Ile Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro
245 250 255
Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro
260 265 270
Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp
275 280 285
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
290 295 300
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser
305 310 315 320
Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly
325 330 335
Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala
340 345 350
Ala Tyr Arg Ser Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys
355 360 365
Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys
370 375 380
Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Gln
435 440 445
Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp
450 455 460
Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg
465 470 475 480
Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr
485 490 495
Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 83
<211> 470
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 25C2-28Z
<400> 83
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr
130 135 140
Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser
145 150 155 160
Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
165 170 175
Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly
180 185 190
Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
195 200 205
Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
210 215 220
Gln Tyr Phe Asn Pro Pro Glu Tyr Thr Phe Gly Gln Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala
245 250 255
Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg
260 265 270
Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys
275 280 285
Asp Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser
290 295 300
Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg
305 310 315 320
Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro
325 330 335
Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe
340 345 350
Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro
355 360 365
Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly
370 375 380
Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro
385 390 395 400
Glu Met Gly Gly Lys Pro Gln Arg Arg Lys Asn Pro Gln Glu Gly Leu
405 410 415
Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile
420 425 430
Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr
435 440 445
Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met
450 455 460
Gln Ala Leu Pro Pro Arg
465 470
<210> 84
<211> 512
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 25C2-28BBZ
<400> 84
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr
130 135 140
Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser
145 150 155 160
Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
165 170 175
Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly
180 185 190
Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
195 200 205
Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
210 215 220
Gln Tyr Phe Asn Pro Pro Glu Tyr Thr Phe Gly Gln Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala
245 250 255
Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg
260 265 270
Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys
275 280 285
Asp Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser
290 295 300
Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg
305 310 315 320
Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro
325 330 335
Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe
340 345 350
Ala Ala Tyr Arg Ser Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe
355 360 365
Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly
370 375 380
Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg
385 390 395 400
Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln
405 410 415
Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp
420 425 430
Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro
435 440 445
Gln Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
450 455 460
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
465 470 475 480
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
485 490 495
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 85
<211> 470
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 25D2-28Z
<400> 85
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Gly Gly Gly Asn Thr Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr
130 135 140
Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser
145 150 155 160
Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
165 170 175
Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly
180 185 190
Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
195 200 205
Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
210 215 220
Gln Tyr Phe Asn Pro Pro Glu Tyr Thr Phe Gly Gln Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala
245 250 255
Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg
260 265 270
Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys
275 280 285
Asp Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser
290 295 300
Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg
305 310 315 320
Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro
325 330 335
Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe
340 345 350
Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro
355 360 365
Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly
370 375 380
Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro
385 390 395 400
Glu Met Gly Gly Lys Pro Gln Arg Arg Lys Asn Pro Gln Glu Gly Leu
405 410 415
Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile
420 425 430
Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr
435 440 445
Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met
450 455 460
Gln Ala Leu Pro Pro Arg
465 470
<210> 86
<211> 512
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 25D2-28BBZ
<400> 86
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Gly Gly Gly Asn Thr Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Arg Pro Phe Trp Gly Thr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr
130 135 140
Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser
145 150 155 160
Gln Ser Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
165 170 175
Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly
180 185 190
Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
195 200 205
Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
210 215 220
Gln Tyr Phe Asn Pro Pro Glu Tyr Thr Phe Gly Gln Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala
245 250 255
Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg
260 265 270
Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys
275 280 285
Asp Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser
290 295 300
Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg
305 310 315 320
Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro
325 330 335
Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe
340 345 350
Ala Ala Tyr Arg Ser Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe
355 360 365
Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly
370 375 380
Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg
385 390 395 400
Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln
405 410 415
Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp
420 425 430
Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro
435 440 445
Gln Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
450 455 460
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
465 470 475 480
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
485 490 495
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 87
<211> 412
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> C11D5.3-BBZ
<400> 87
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ser Ile Asn Trp Val Lys Arg Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro Ala Tyr Ala Tyr Asp Phe
50 55 60
Arg Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Tyr Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Leu Asp Tyr Ser Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser
100 105 110
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Asp Ile Val Leu Thr Gln Ser Pro Pro Ser Leu Ala
130 135 140
Met Ser Leu Gly Lys Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser
145 150 155 160
Val Thr Ile Leu Gly Ser His Leu Ile His Trp Tyr Gln Gln Lys Pro
165 170 175
Gly Gln Pro Pro Thr Leu Leu Ile Gln Leu Ala Ser Asn Thr Thr Thr
180 185 190
Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro
195 200 205
Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val
210 215 220
His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro
225 230 235 240
Leu Ala Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu
245 250 255
Tyr Cys Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro
260 265 270
Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys
275 280 285
Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe
290 295 300
Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu
305 310 315 320
Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp
325 330 335
Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys
340 345 350
Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala
355 360 365
Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys
370 375 380
Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr
385 390 395 400
Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
405 410
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-BCMA АНТИТЕЛА, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЁЛУЮ ЦЕПЬ | 2017 |
|
RU2781301C2 |
| Т-КЛЕТКИ, МОДИФИЦИРОВАННЫЕ ХИМЕРНЫМ РЕЦЕПТОРОМ АНТИГЕНА, НАЦЕЛЕННЫМ НА CS1, ДЛЯ ЛЕЧЕНИЯ АМИЛОИДОЗА AL | 2018 |
|
RU2774895C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ CD3E/BCMA И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800164C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА АНТИГЕН СОЗРЕВАНИЯ В-КЛЕТОК, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2829097C2 |
| Моноклональное антитело или его антигенсвязывающий фрагмент, которое специфически связывается с BCMA, и его применение | 2022 |
|
RU2820350C2 |
| Антитело к BCMA, его антигенсвязывающий фрагмент и их медицинское применение | 2020 |
|
RU2819660C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ CD3 | 2017 |
|
RU2779489C2 |
| АНТИТЕЛО, СПОСОБНОЕ СВЯЗЫВАТЬСЯ С ТИМИЧЕСКИМ СТРОМАЛЬНЫМ ЛИМФОПОЭТИНОМ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2825304C2 |
| АНТИТЕЛА К C10ORF54 И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714232C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ И СВЯЗЫВАЮЩИЕ АГЕНТЫ, НАЦЕЛЕННЫЕ НА DLL3 | 2020 |
|
RU2822366C2 |
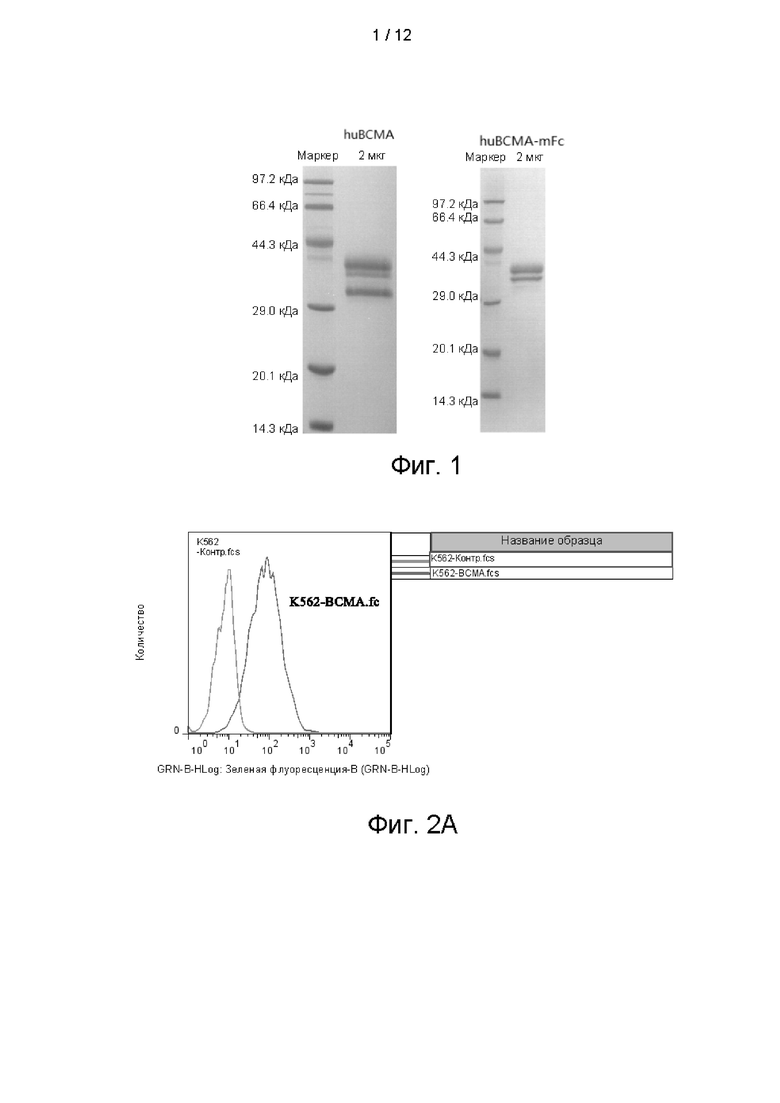
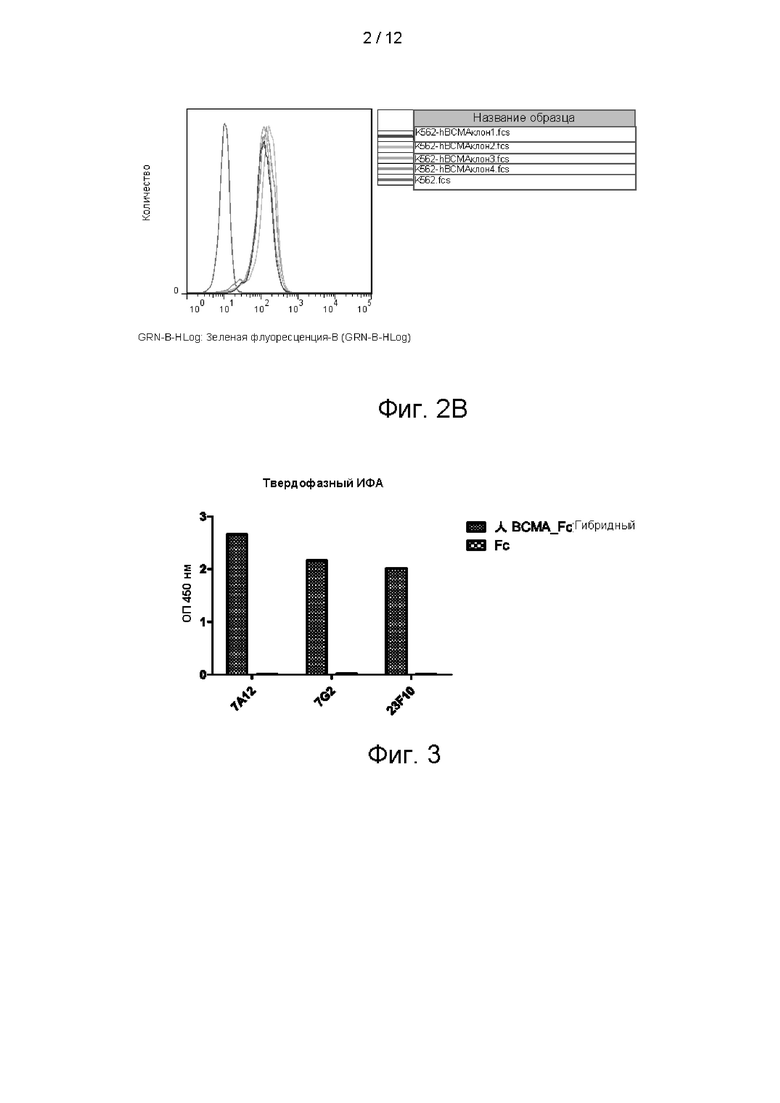
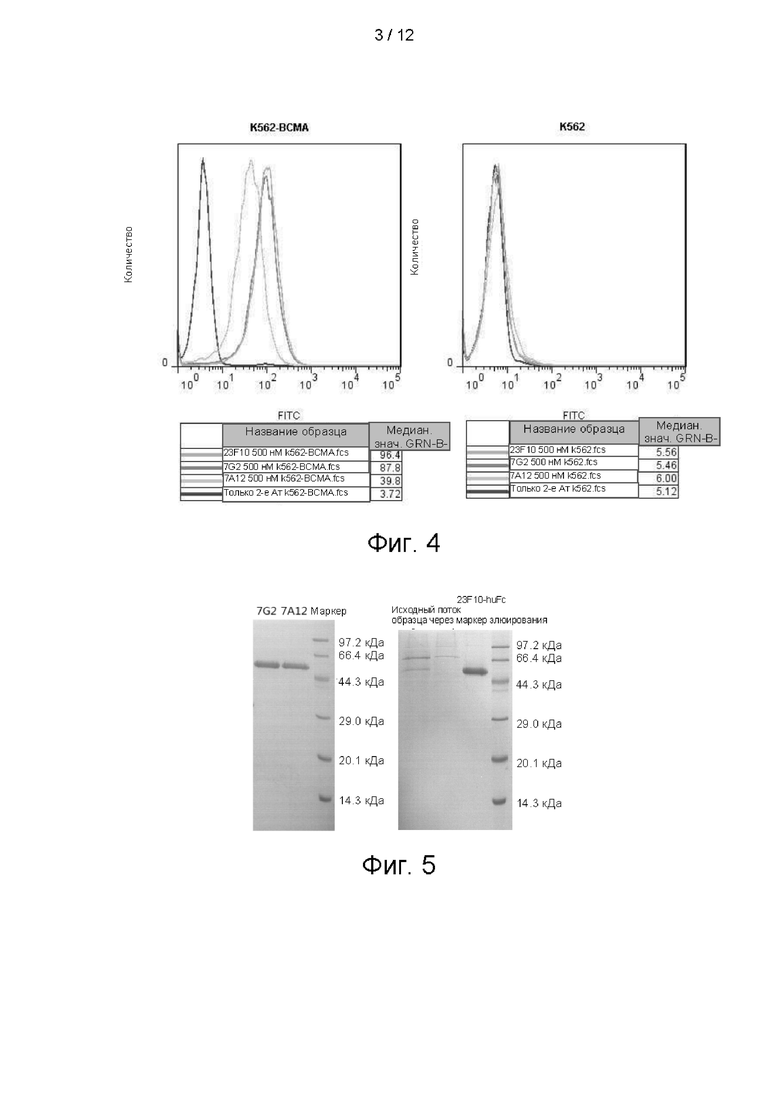
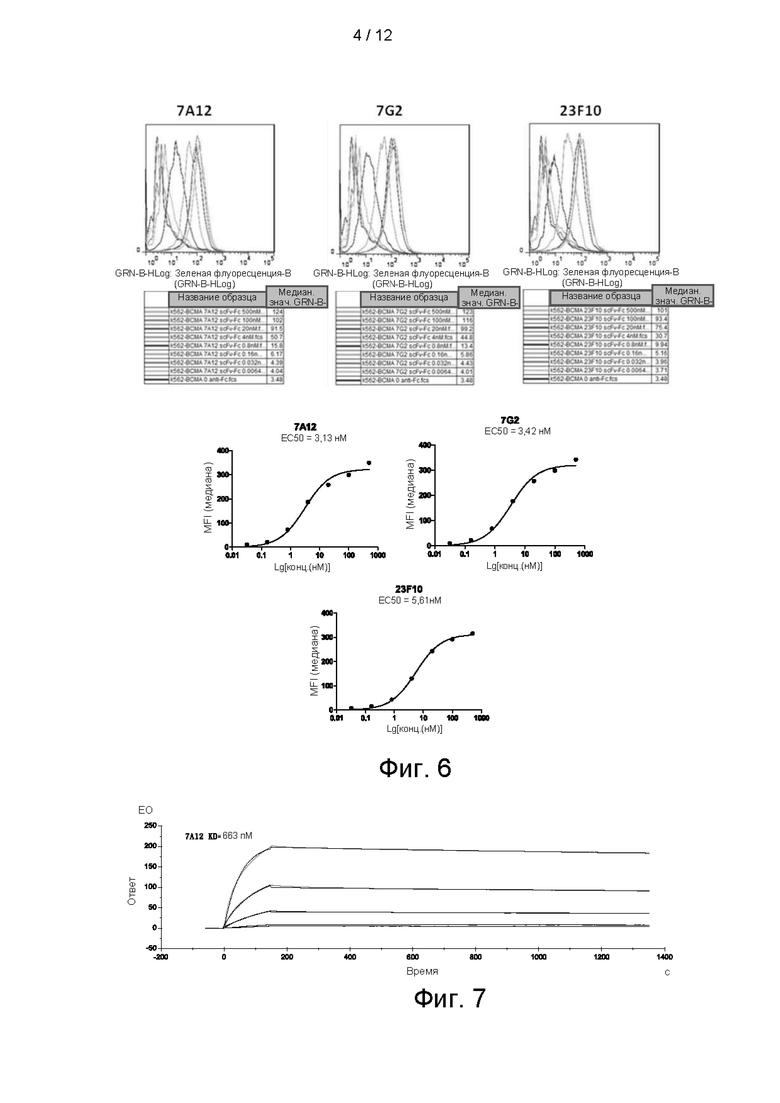
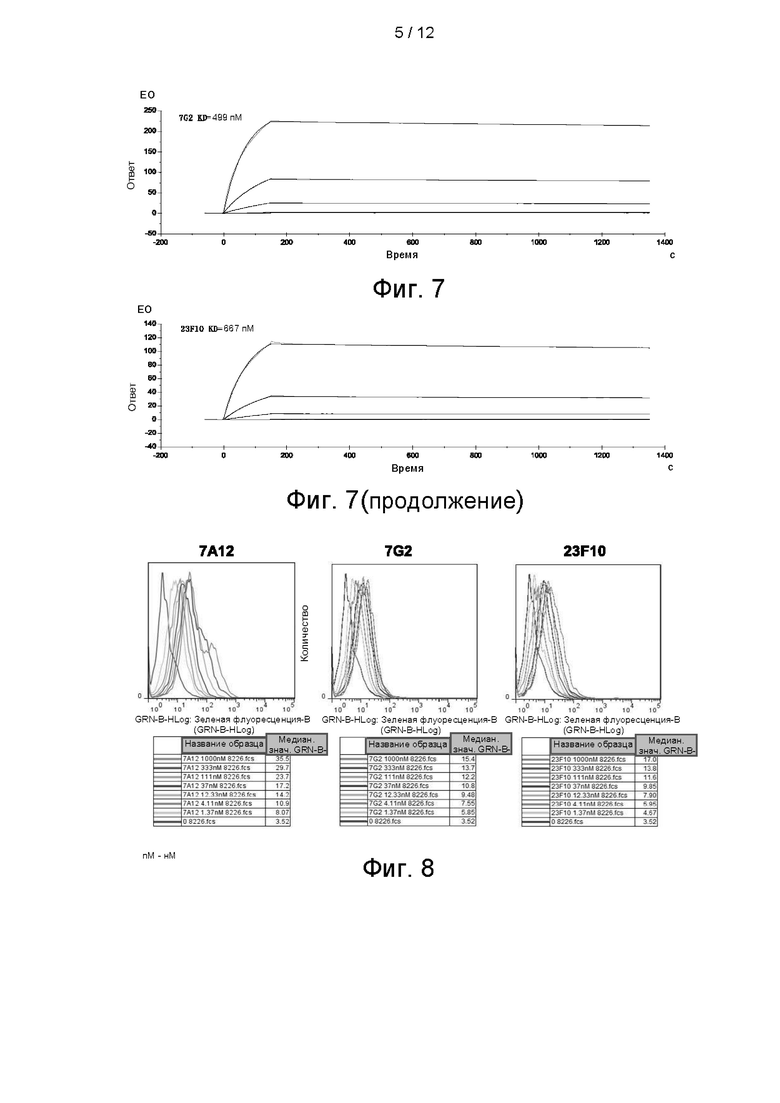
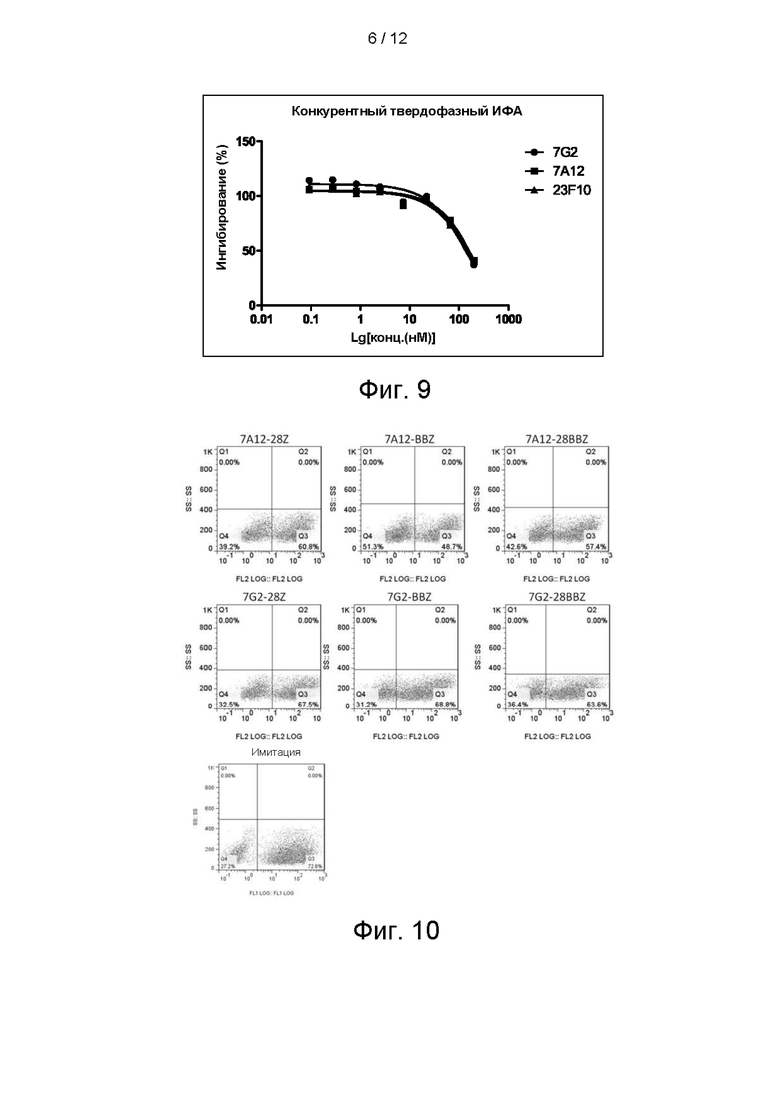
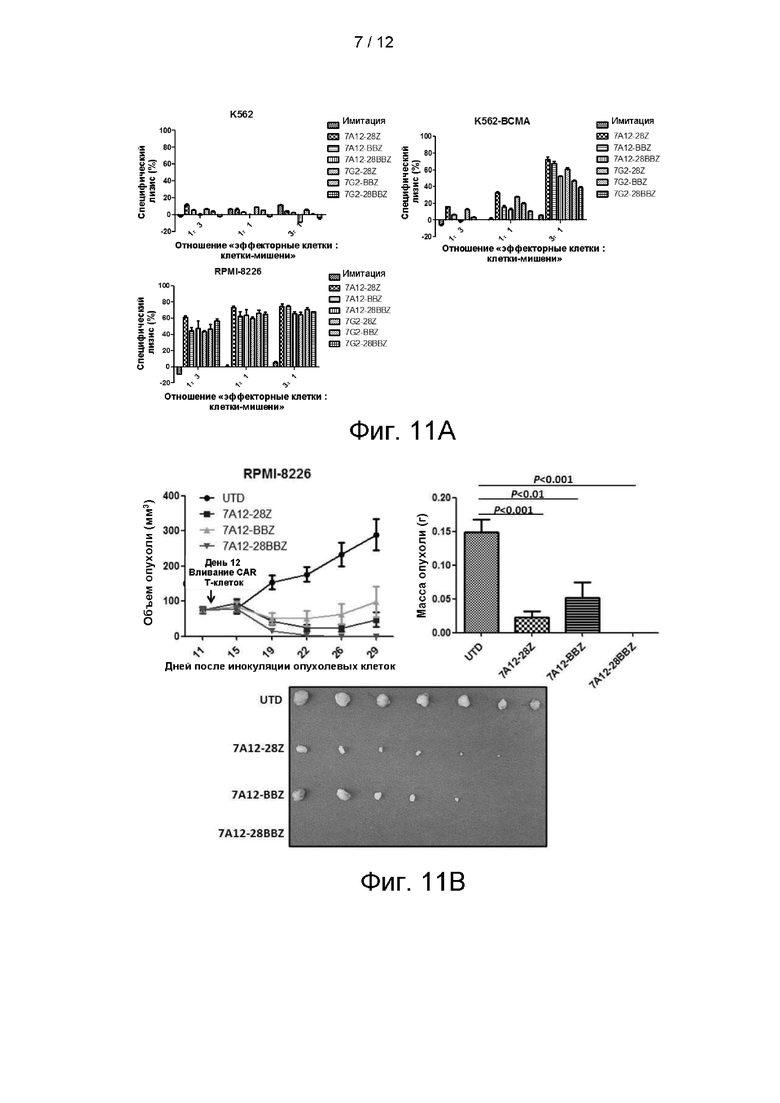
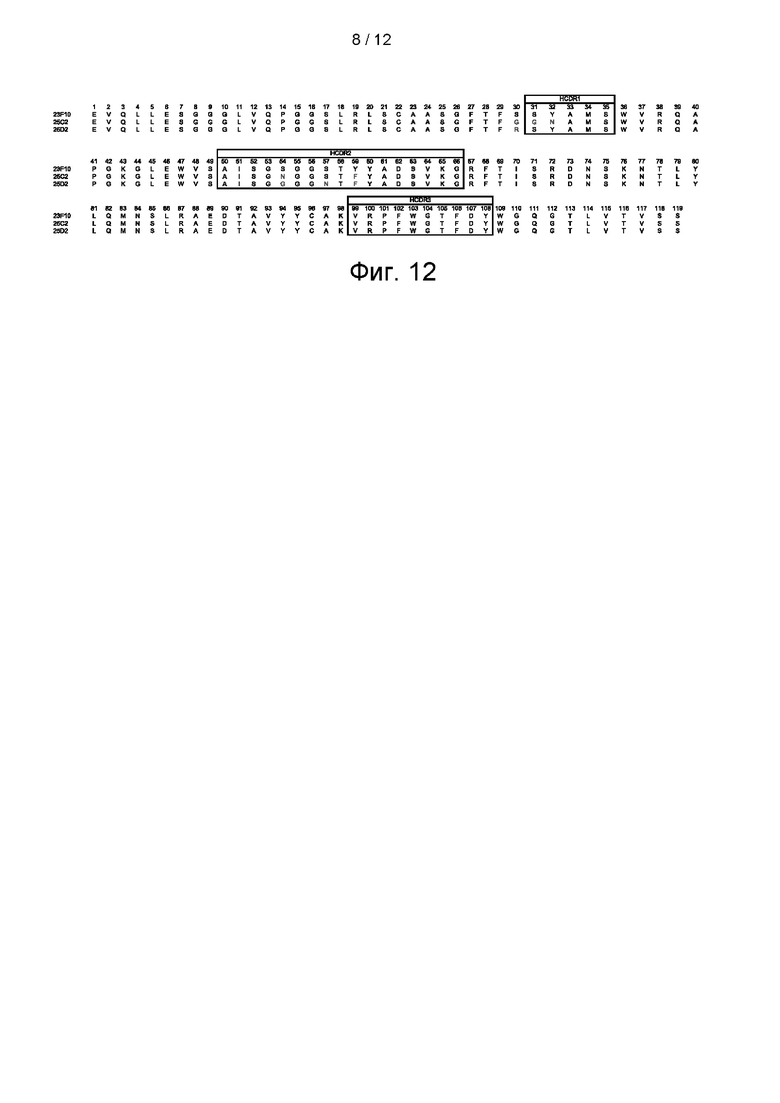
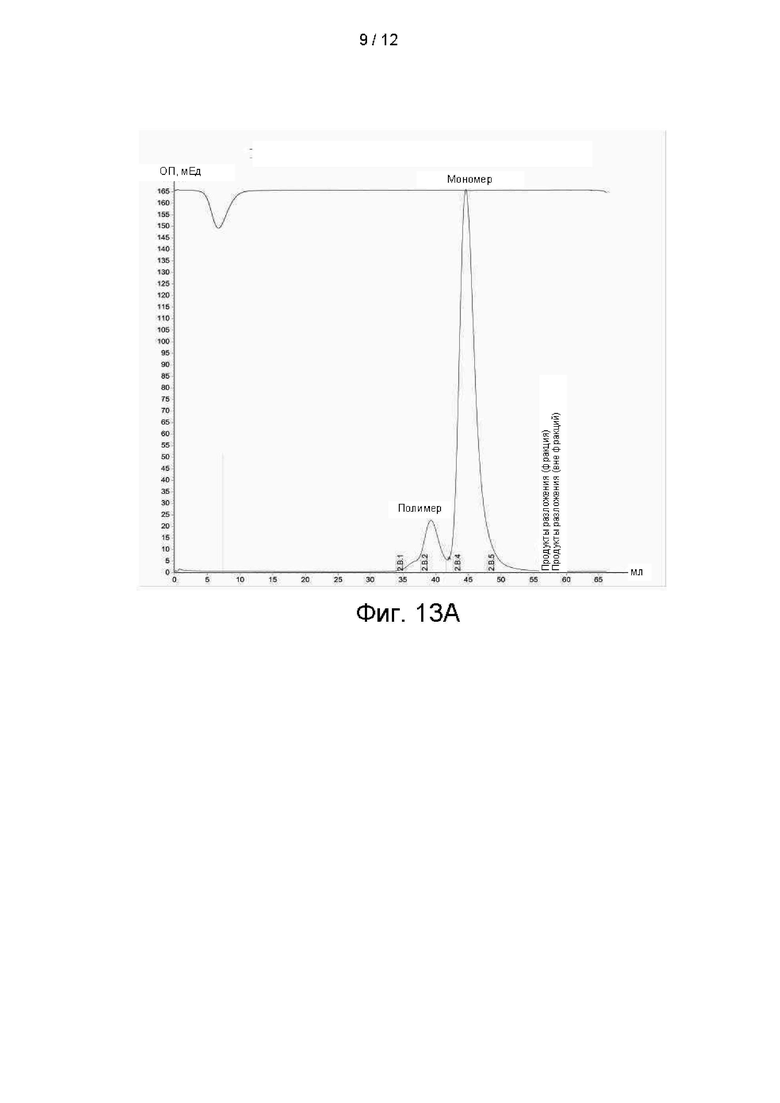
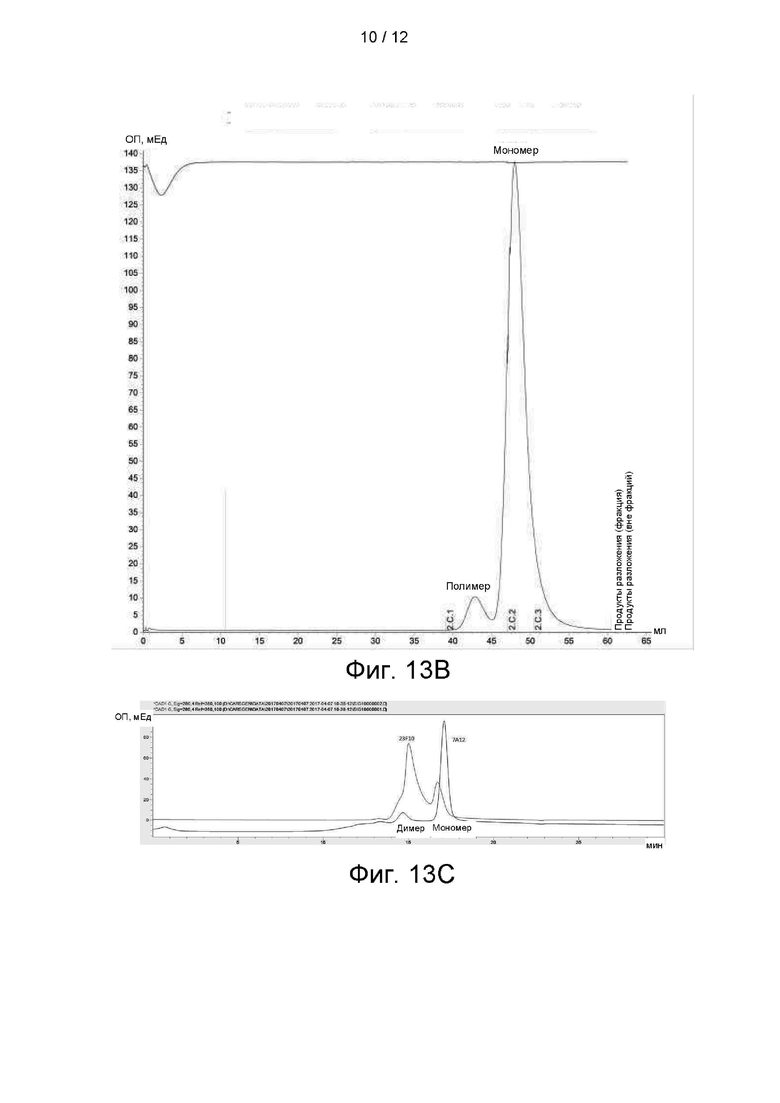
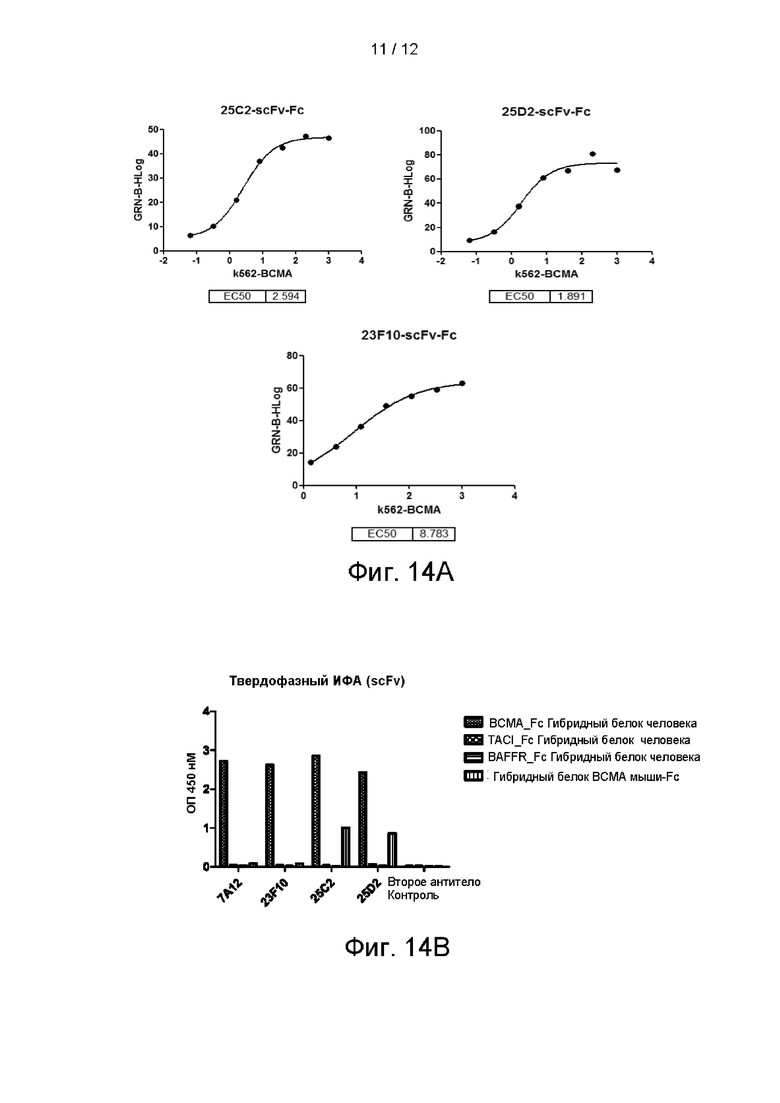
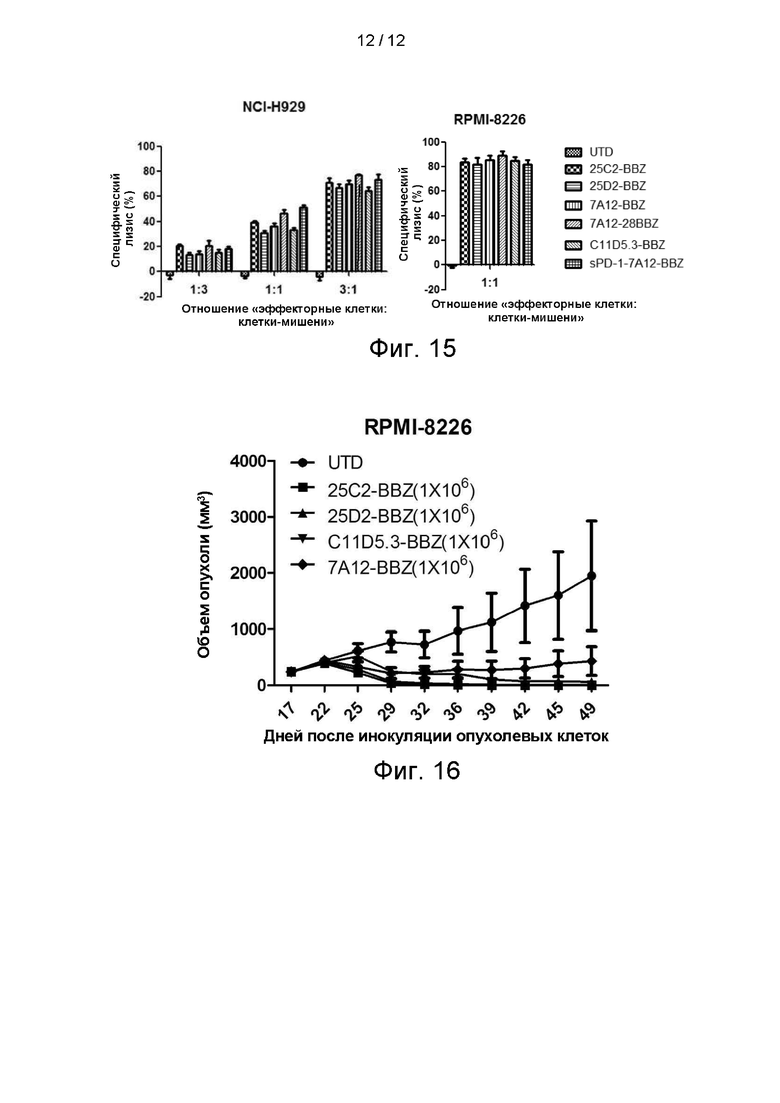
Изобретение относится к области биохимии, в частности к антителу, которое специфически связывается с ВСМА. Также раскрыты нуклеиновая кислота, кодирующая указанное антитело; вектор и клетка-хозяин, экспрессирующие указанное антитело; иммуноконъюгат и химерный рецептор антигена, содержащие указанное антитело; нуклеиновая кислота, кодирующая указанный химерный рецептор; вектор, содержащий указанную нуклеиновую кислоту; вирус, содержащий указанный вектор; иммунная клетка, содержащая указанные нуклеиновую кислоту, вектор или вирус. Раскрыты применение указанного антитела для получения лекарственного средства, конъюгата, многофункционального антитела, агента для диагностики опухоли, иммунной клетки; применение указанного иммуноконъюгата для получения лекарственного средства, агента для диагностики опухоли, иммунной клетки; применение указанной иммунной клетки для получения лекарственного средства. Изобретение позволяет эффективно лечить заболевания, ассоциированные с экспрессией BCMA. 23 н. и 20 з.п. ф-лы, 16 ил., 4 табл.
1. Антитело, специфически связывающееся с BCMA, причем указанное антитело выбрано из группы, состоящей из:
(1) антитела, содержащего HCDR1, представленную в SEQ ID NO: 1, HCDR2, представленную в SEQ ID NO: 2, HCDR3, представленную в SEQ ID NO: 3, и LCDR1, представленную в SEQ ID NO: 6, LCDR2, представленную в SEQ ID NO: 7, и LCDR3, представленную в SEQ ID NO: 8;
(2) антитела, содержащего HCDR1, представленную в SEQ ID NO: 1, HCDR2, представленную в SEQ ID NO: 2, HCDR3, представленную в SEQ ID NO: 4, LCDR1, представленную в SEQ ID NO: 6, LCDR2, представленную в SEQ ID NO: 7, и LCDR3, представленную в SEQ ID NO: 9;
(3) антитела, содержащего HCDR1, представленную в SEQ ID NO: 1, HCDR2, представленную в SEQ ID NO: 2, HCDR3, представленную в SEQ ID NO: 5, LCDR1, представленную в SEQ ID NO: 6, LCDR2, представленную в SEQ ID NO: 7, и LCDR3, представленную в SEQ ID NO: 10;
(4) антитела, содержащего HCDR1, представленную в SEQ ID NO: 60, HCDR2, представленную в SEQ ID NO: 61, HCDR3, представленную в SEQ ID NO: 5, LCDR1, представленную в SEQ ID NO: 6, LCDR2, представленную в SEQ ID NO: 7, и LCDR3, представленную в SEQ ID NO: 10;
(5) антитела, содержащего HCDR1, представленную в SEQ ID NO: 62, HCDR2, представленную в SEQ ID NO: 63, HCDR3, представленную в SEQ ID NO: 5, LCDR1, представленную в SEQ ID NO: 6, LCDR2, представленную в SEQ ID NO: 7, и LCDR3, представленную в SEQ ID NO: 10.
2. Антитело по п. 1, отличающееся тем, что указанное антитело выбрано из группы, состоящей из:
(1) антитела, где вариабельная область тяжелой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 13, аминокислотную последовательность SEQ ID NO: 17, аминокислотную последовательность SEQ ID NO: 21 или аминокислотную последовательность SEQ ID NO: 56, или аминокислотную последовательность SEQ ID NO: 58;
(2) антитела, где вариабельная область легкой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 11, аминокислотную последовательность SEQ ID NO: 15 или аминокислотную последовательность SEQ ID NO: 19;
(3) антитела, содержащего вариабельную область тяжелой цепи антитела согласно подпункту (1) и вариабельную область легкой цепи антитела согласно подпункту (2).
3. Антитело по п. 2, отличающееся тем, что указанное антитело выбрано из группы, состоящей из:
(1) антитела, где вариабельная область тяжелой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 13, а вариабельная область легкой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 11;
(2) антитела, где вариабельная область тяжелой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 17, а вариабельная область легкой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 15;
(3) антитела, где вариабельная область тяжелой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 21, а вариабельная область легкой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 19;
(4) антитела, где вариабельная область тяжелой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 56, а вариабельная область легкой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 19;
(5) антитела, где вариабельная область тяжелой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 58, а вариабельная область легкой цепи указанного антитела имеет аминокислотную последовательность SEQ ID NO: 19.
4. Нуклеиновая кислота, кодирующая антитело по любому из пп. 1-3.
5. Экспрессирующий вектор, содержащий нуклеиновую кислоту по п. 4.
6. Клетка-хозяин для экспрессии антитела, которое специфически связывается с BCMA, содержащая экспрессирующий вектор по п. 5 или нуклеиновую кислоту по п. 4, встроенную в геном.
7. Применение антитела по любому из пп. 1-3 для получения лекарственного средства нацеленного действия, которое специфически нацеливается на опухолевые клетки, экспрессирующие BCMA.
8. Применение антитела по любому из пп. 1-3 для получения конъюгата «антитело-лекарственное средство», которое специфически нацеливается на опухолевые клетки, экспрессирующие BCMA.
9. Применение антитела по любому из пп. 1-3 для получения многофункционального антитела, которое специфически нацеливается на опухолевые клетки, экспрессирующие BCMA.
10. Применение антитела по любому из пп. 1-3 для получения агента для диагностики опухоли, экспрессирующей BCMA.
11. Применение антитела по любому из пп. 1-3 для получения иммунной клетки, модифицированной химерным рецептором антигена.
12. Применение по п. 11, отличающееся тем, что указанная иммунная клетка включает Т-лимфоцит, NK-клетку или NKT-лимфоцит.
13. Иммуноконъюгат для распознавания BCMA, содержащий
антитело по любому из пп. 1-3; и
функциональную молекулу, связанную с ним; причем указанная функциональная молекула представляет собой молекулу, нацеленную на поверхностный маркер опухоли.
14. Иммуноконъюгат для распознавания BCMA, содержащий
антитело по любому из пп. 1-3; и
функциональную молекулу, связанную с ним; причем указанная функциональная молекула представляет собой молекулу, ингибирующую опухоли.
15. Иммуноконъюгат для распознавания BCMA, содержащий
антитело по любому из пп. 1-3; и
функциональную молекулу, связанную с ним; причем указанная функциональная молекула представляет собой молекулу, нацеленную на поверхностный маркер иммунной клетки.
16. Иммуноконъюгат для распознавания BCMA, содержащий
антитело по любому из пп. 1-3; и
функциональную молекулу, связанную с ним; причем указанная функциональная молекула представляет собой детектируемую метку.
17. Иммуноконъюгат по п. 14, отличающийся тем, что указанная молекула, ингибирующая опухоли, представляет собой противоопухолевый цитокин или противоопухолевый токсин.
18. Иммуноконъюгат по п. 17, отличающийся тем, что указанный цитокин включает: ИЛ-12, ИЛ-15, интерферон I типа, ФНО-альфа.
19. Иммуноконъюгат по п. 15, отличающийся тем, что молекула, нацеленная на поверхностный маркер иммунной клетки, представляет собой антитело или лиганд, связывающиеся с поверхностным маркером иммунной клетки.
20. Иммуноконъюгат по п. 19, отличающийся тем, что указанный поверхностный маркер иммунной клетки включает: CD3, CD16, CD28.
21. Иммуноконъюгат по п. 20, отличающийся тем, что указанное антитело, связывающее поверхностный маркер иммунной клетки, представляет собой антитело против CD3.
22. Иммуноконъюгат по п. 19, отличающийся тем, что молекула, нацеленная на поверхностный маркер иммунной клетки, представляет собой антитело, связывающееся с поверхностным маркером Т-клетки.
23. Применение иммуноконъюгата по любому из пп. 15-21 для получения противоопухолевого лекарственного средства против опухолей, экспрессирующих BCMA.
24. Применение иммуноконъюгата по любому из пп. 15-21 для получения агента для диагностики опухоли, экспрессирующей BCMA.
25. Применение иммуноконъюгата по любому из пп. 15-21 для получения иммунной клетки, модифицированной химерным рецептором антигена.
26. Применение по п. 25, отличающееся тем, что указанная иммунная клетка включает: Т-лимфоциты, NK-клетки или NKT-лимфоциты.
27. Химерный рецептор антигена для распознавания BCMA, содержащий внеклеточный домен, трансмембранный домен и внутриклеточный сигнальный домен, причем внеклеточный домен содержит антитело по любому из пп. 1-3.
28. Химерный рецептор антигена по п. 27, отличающийся тем, что указанное антитело представляет собой одноцепочечное антитело или доменное антитело.
29. Химерный рецептор антигена по п. 27, отличающийся тем, что внутриклеточный сигнальный домен содержит один или более из костимулирующих сигнальных доменов и/или первичных сигнальных доменов.
30. Химерный рецептор антигена по п. 27, отличающийся тем, что указанный химерный рецептор антигена дополнительно содержит шарнирный домен.
31. Химерный рецептор антигена по п. 29, отличающийся тем, что трансмембранный домен выбран из группы, состоящей из альфа-, бета-, дзета-цепи TCR, трансмембранных областей CD3ε, CD3ζ, CD4, CD5, CD8α, CD9, CD16, CD22, CD27, CD28, CD33, CD37, CD45, CD64, CD80, CD86, CD134, CD137, CD152, CD154 и PD1; и/или
костимулирующий сигнальный домен выбран из группы, состоящей из внутриклеточных сигнальных областей CARD11, CD2, CD7, CD27, CD28, CD30, CD40, CD54, CD83, OX40, CD137, CD134, CD150, CD152, CD223, CD270, PD-L2, PD-L1, CD278, DAP10, LAT, NKD2C SLP76, TRIM, FcεRIγ, MyD88 и 41BBL; и/или
первичный сигнальный домен выбран из группы, состоящей из TCR ξ, FcR γ, FcR β, CD3 γ, CD3δ, CD3ε, CD5, CD22, CD79a, CD79b, CD278 (также называемого «ICOS»), CD66d и CD3ζ.
32. Химерный рецептор антигена по п. 31, отличающийся тем, что трансмембранный домен выбран из группы, состоящей из трансмембранных доменов CD8α, CD4, CD45, PD1, CD154 и CD28; и/или
костимулирующий сигнальный домен выбран из группы, состоящей из CD137, CD134, CD28 и OX40; и/или
первичный сигнальный домен выбран из CD3ζ.
33. Химерный рецептор антигена по п. 32, отличающийся тем, что трансмембранный домен выбран из CD8α или CD28, костимулирующий сигнальный домен выбран из внутриклеточного сигнального домена CD137 или CD28, а первичный сигнальный домен выбран из CD3ζ.
34. Химерный рецептор антигена по п. 27, отличающийся тем, что указанный химерный рецептор антигена содержит следующие последовательно связанные антитело, трансмембранную область и внутриклеточную сигнальную область:
антитело по любому из пп. 1-3, трансмембранную область CD8 и CD3ζ;
антитело по любому из пп. 1-3, трансмембранную область CD8, внутриклеточную сигнальную область CD137 и CD3ζ;
антитело по любому из пп. 1-3, трансмембранную область CD28, внутриклеточную сигнальную область CD28 и CD3ζ; или
антитело по любому из пп. 1-3, трансмембранную область CD28, внутриклеточную сигнальную область CD28, CD137 и CD3ζ.
35. Нуклеиновая кислота, кодирующая химерный рецептор антигена по любому из пп. 27-34.
36. Экспрессирующий вектор, содержащий нуклеиновую кислоту по п. 35.
37. Вирус для доставки генетической информации, содержащий вектор по п. 36.
38. Применение химерного рецептора антигена по любому из пп. 27-34 или нуклеиновой кислоты по п. 35, или экспрессирующего вектора по п. 36, или вируса по п. 37 для получения генетически модифицированных иммунных клеток, нацеленных на опухолевую клетку, экспрессирующую BCMA.
39. Генетически модифицированная иммунная клетка, трансдуцированная нуклеиновой кислотой по п. 35 или экспрессирующим вектором по п. 36, или вирусом по п. 37; и экспрессирующая химерный рецептор антигена по любому из пп. 27-34.
40. Генетически модифицированная иммунная клетка по п. 39, отличающаяся тем, что указанная иммунная клетка выбрана из T-лимфоцитов, NK-клеток или NKT-клеток.
41. Генетически модифицированная иммунная клетка по п. 39 или 40, отличающаяся тем, что она дополнительно экспрессирует последовательность, не являющуюся химерным рецептором антигена по любому из пп. 27-34, и указанная другая последовательность содержит цитокин или еще один химерный рецептор антигена или рецептор хемокина, или миРНК, снижающую экспрессию PD-1, или белок, блокирующий PD-L1, или TCR, или «предохранительный переключатель» (safety switch).
42. Генетически модифицированная иммунная клетка по п. 41, отличающаяся тем, что указанный цитокин включает ИЛ-12, ИЛ-15, ИЛ-21 или интерферон I типа;
рецептор хемокина включает CCR2, CCR5, CXCR2 или CXCR4; и
«предохранительный переключатель» включает индуцибельную каспазу-9, укороченный EGFR или RQR8.
43. Применение генетически модифицированной иммунной клетки по любому из пп. 39-42 для получения лекарственного средства, подавляющего опухоль, причем указанная опухоль представляет собой опухоль, экспрессирующую BCMA.
| АНТИ-GM-CSF АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2006 |
|
RU2447085C2 |
| CN 103562225 A, 05.02.2014 | |||
| WO 2014122143 A1, 14.08.2014. | |||

