Настоящее изобретение относится к нуклеиновым кислотам, кодирующим новые химерные антигенные рецепторы (CAR), а также к самим CAR, клеткам, содержащим нуклеиновые кислоты, и их применению в терапии, в частности к способам, в которых они применяются для облегчения ответа Т-клеток на выбранную мишень.
Уровень техники изобретения
Химерные антигенные рецепторы (CAR), которые также могут обозначаться как искусственные Т-клеточные рецепторы, химерные Т-клеточные рецепторы (TCR) или химерные иммунорецепторы, являются рекомбинантными рецепторами, хорошо известными из уровня техники. Их используют прежде всего для трансформации иммунных эффекторных клеток, в частности Т-клеток, для того чтобы обеспечить эти клетки определенной специфичностью. Их в частности изучают в области иммунотерапии рака, где их можно использовать в таких методиках, как адаптивный перенос клеток. В этих способах терапии Т-клетки удаляют из пациента и модифицируют так, что они экспрессируют рецепторы, специфические в отношении антигенов, обнаруженных в определенной форме рака. Т-клетки, которые могут затем распознать и уничтожать раковые клетки, повторно вводят пациенту.
CAR первого поколения обеспечивают TCR-подобный сигнал, чаще всего использующий CD3-дзета (z) и за счет этого вызывают противоопухолевые функции. Однако вовлечение слитых рецепторов CD3z-цепи может являться недостаточным для того, чтобы вызвать существенную секрецию и/или пролиферацию IL-2 в отсутствие сопутствующего костимулирующего сигнала. В физиологических ответах Т-клеток оптимальная активация лимфоцитов требует вовлечения одного или нескольких костимулирующих рецепторов (сигнал 2), таких как CD28 или 4-1ВВ. Следовательно, Т-клетки также были сконструированы таким образом, чтобы они получали костимулирующий сигнал антиген-зависимым от опухоли способом.
Важным усовершенствованием в этом отношении было успешное конструирование "CAR второго поколения", которые трансформируют функциональный антиген-зависимый костимулирующий сигнал в первичных Т-клетках человека, обеспечивая пролиферацию Т-клеток в дополнение к противоопухолевой активности. CAR второго поколения чаще всего обеспечивают совместную стимуляцию с использованием модулей, полученных из CD28 или 4-1ВВ. Комбинированная доставка костимуляции плюс сигнал CD3-дзета делает CAR второго поколения явно превосходящими по функциональности по сравнению с их аналогами первого поколения (только сигнал CD3z). Пример CAR второго поколения приведен в патенте США №7446190.
Совсем недавно были получены так называемые "CAR третьего поколения". Они объединяют несколько сигнальных доменов, таких как CD28+4-1BB+CD3z или CD28+OX40+CD3z, для дальнейшего увеличения эффективности. В CAR 3-го поколения сигнальные домены последовательно выстраиваются в эндодомене CAR и помещаются выше CD3z.
В целом, однако, результаты, достигнутые с помощью данных CAR третьего поколения, неутешительно обеспечивали лишь незначительное улучшение по сравнению с конфигурациями 2-го поколения.
Было также предложено использование клеток, трансформированных несколькими конструкциями. Например, Kloss et al. Nature Biotechnology 2012, doi: 10.1038/nbt.2459 описывают трансдукцию T-клеток с помощью CAR, содержащих область активации сигнала (CD3-дзета-цепь), которая целенаправленно воздействует на первый антиген и химерный костимулирующий рецептор (CCR), содержащий как CD28, так и 4-1ВВ костимулирующие области, которые целенаправленно воздействуют на второй антиген. Эти две конструкции связываются с их соответствующими антигенами с различной аффинностью, что приводит к эффекту "обнаружения опухоли", который может повысить специфичность терапии с целью снижения побочных эффектов.
Желательно разработать системы, в которых Т-клетки могут поддерживаться в состоянии, в котором они могут расти, продуцировать цитокины и доставлять сигнал уничтожения через несколько повторных циклов стимуляции с помощью антиген-экспрессирующих опухолевых клеток-мишеней. Обеспечение субоптимальной костимуляции приводит к тому, что Т-клетки быстро теряют эти эффекторные функции при повторной стимуляции, входя в состояние, известное как "анергия". Если Т-клетки с CAR последовательно стимулируются повторно in vitro, они постепенно теряют эффекторные свойства (например, продукцию IL-2, способность к пролиферации) и дифференцируются, чтобы стать более эффектор-подобными, другими словами, с меньшей вероятностью проявляют эффекты костимуляции. Это является нежелательным для иммунотерапии рака, поскольку более дифференцированные клетки, как правило, обычно имеют меньшую продолжительность жизни и сниженную способность претерпевать дальнейший рост/активацию, когда их многократно стимулируют в микроокружении опухоли.
Краткое описание изобретения
Авторы настоящего изобретения обнаружили, что эффективные ответы Т-клеток могут генерироваться с использованием комбинации конструкций, в которой множественные костимулирующие области расположены в различных конструкциях.
В соответствии с первым аспектом настоящего изобретения предусматривается иммунореактивная клетка, экспрессирующая
(i) химерный антигенный рецептор второго поколения, содержащий
(a) сигнальную область;
(b) костимулирующую сигнальную область;
(c) трансмембранный домен и
(d) связывающий элемент, который специфически взаимодействует с первым эпитопом на целевом антигене; а также
(ii) химерный костимулирующий рецептор, содержащий
(e) костимулирующую сигнальную область, которая отличается от области (b);
(f) трансмембранный домен и
(g) связывающий элемент, который специфически взаимодействует со вторым эпитопом на целевом антигене.
Авторы настоящего изобретения обнаружили, что эффективность этой системы является хорошей и, в частности, может быть лучше, чем эффективность, достигнутая с использованием традиционных CAR третьего поколения, имеющих схожие элементы. Конструкции типа по настоящему изобретению можно назвать параллельными химерными активирующими рецепторами” или "pCAR".
Кроме того, пролиферация клеток, их способность поддерживать свою цитотоксическую активность и высвобождение IL-2 поддерживаются в течение многих повторных циклов стимуляции антиген-экспрессирующими опухолевыми клетками.
Не ограничиваясь теорией, конфигурация элементов в pCAR может способствовать активности. Например, по определению один из костимулирующих модулей в CAR 3-го поколения должен быть удален от своего естественного местоположения вблизи внутреннего слоя плазматической мембраны. Это может привести к тому, что он не будет нормально передавать сигнал из-за нарушения доступа к облигатным связанным с мембраной молекулам-партнерам. В качестве альтернативы, тесное сближение 2-х костимулирующих сигнальных модулей в CAR 3-го поколения может привести к возникновению стерических проблем, препятствуя полному задействованию одного или нескольких последовательных сигнальных путей. Обе эти проблемы избегаются в конфигурации по настоящему изобретению. Оба сигнальных фрагмента (b) и (е) могут быть слиты непосредственно в трансмембранный домен, гарантируя, что они оба прилегают к плазматической мембране внутри клетки. Кроме того, они могут располагаться на отдельных участках внутри клетки, чтобы они не взаимодействовали друг с другом стерически.
Подходящие иммунореактивные клетки для применения в первом аспекте настоящего изобретения включают Т-клетки, такие как цитотоксические Т-клетки, Т-клетки-хелперы или регуляторные Т-клетки и естественные клетки-киллеры (NK). В частности, иммунореактивная клетка представляет собой Т-клетку.
Подходящие элементы (а), приведенные выше, могут включать любую подходящую сигнальную область, включая любую область, содержащую иммунорецепторный тирозиновый активирующий мотив (ITAM), как описано, например, в Love et al. Cold Spring Harbor Perspect. Biol 2010 2(6)1 a002485. В конкретном варианте осуществления сигнальная область содержит внутриклеточный домен CD3-[дзета]-цепи человека, как описано, например, в патенте США №7446190, или его вариант.
В частности, она содержит домен, который охватывает аминокислотные остатки 52-163 полноразмерной CD3-дзета-цепи человека. Он имеет ряд полиморфных форм (например, Sequence ID: gb|AAF34793.1 и gb|ААА60394.1), которые показаны соответственно как SEQ ID NO: 1 и 2:
RVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO: 1)
RVKFSRSAEPPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO: 2)
Используемый в данном документе термин "вариант" относится к полипептидной последовательности, которая представляет собой встречающуюся в природе полиморфную форму основной последовательности, а также синтетические варианты, в которых одна или несколько аминокислот в пределах цепи вставлены, удалены или заменены. Однако вариант обеспечивает биологический эффект, который аналогичный эффекту основной последовательности. Например, упомянутый выше вариант будет действовать так же, как и внутриклеточный домен CD3-[дзета]-цепи человека. Аминокислотные замены можно рассматривать как "консервативные", где аминокислоту заменяют другой аминокислотой в том же классе с широко подобными свойствами. Неконсервативные замены заключаются в замене аминокислот аминокислотами другого типа или класса.
Классы аминокислот определяются следующим образом.

Как хорошо известно специалистам в данной области техники, изменение первичной структуры пептида посредством консервативной замены не может существенно изменить активность этого пептида, поскольку боковая цепь аминокислоты, которая вставлена в последовательность, может быть способна образовывать аналогичные связи и контакты также как боковая цепь аминокислоты, которая была замещена. Это происходит даже тогда, когда замена находится в области, которая является существенно важной для определения конформации пептида.
Неконсервативные замены также могут быть возможны при условии, что они не прерывают функцию полипептида, описанную выше. В широком смысле, меньше неконсервативных замен будет возможно без изменения биологической активности полипептидов.
В целом, варианты будут иметь аминокислотные последовательности, которые будут по меньшей мере на 70%, например, по меньшей мере на 71%, 75%, 79%, 81%, 84%, 87%, 90%, 93%, 95%, 96% или 98% идентичны основной последовательности, например SEQ ID NO: 1 или SEQ ID NO: 2. Идентичность в этом контексте может быть определена с использованием компьютерной программы BLASTP с SEQ ID NO: 2 или фрагмента, в частности фрагмента, описанного ниже, в качестве основной последовательности. Программное обеспечение BLAST доступно по адресу http://blast.ncbi.nlm.nih.gov/Blast.cgi (доступ от 12 марта 2009 г.).
Костимулирующая сигнальная последовательность (b) подходящим образом расположена между трансмембранным доменом (с) и сигнальной областью (а) и удалена от связывающего элемента (d). Аналогично, костимулирующая сигнальная последовательность (е) соответствующим образом расположена рядом с трансмембранным доменом (f) и удалена от связывающего элемента (g).
Подходящие костимулирующие сигнальные области для применения в качестве элементов (b) и (е), приведенных выше, также хорошо известны из уровня техники и включают членов семейства B7/CD28, таких как В7-1, В7-2, В7-Н1, В7-Н2, В7-Н3, В7-Н4, В7-Н6, В7-Н7, BTLA, CD28, CTLA-4, Gi24, ICOS, PD-1, PD-L2 или PDCD6; или белки семейства ILT/CD85, такие как LILRA3, LILRA4, LILRB1, LILRB2, LILRB3 или LILRB4; или членов суперсемейства фактора некроза опухоли (TNF), таких как 4-1ВВ, BAFF, BAFF R, CD27, CD30, CD40, DR3, GITR, HVEM, LIGHT, лимфотоксин-альфа, ОХ40, RELT, TACI, TL1A, TNF-альфа или TNF RII; или членов семейства SLAM, таких как 2В4, BLAME, CD2, CD2F-10, CD48, CD58, CD84, CD229, CRACC, NTB-A или SLAM; или членов семейства TIM, таких как TIM-1, TIM-3 или TIM-4; или другие костимулирующие молекулы, такие как CD7, CD96, CD160, CD200, CD300a, CRTAM, DAP12, Dectin-1, DPPIV, EphB6, интегрин альфа-4 бета-1, интегрин альфа-4 бета-7/LPAM-1, LAG-3 или TSLP R.
Выбор костимулирующих сигнальных областей может быть определен в зависимости от конкретного применения, предназначенного для трансформированных клеток. В частности, костимулирующие сигнальные области, выбранные для (b) и (е), приведенных выше, являются такими, которые могут работать совместно или синергически вместе. Например, костимулирующие сигнальные области для (b) и (е) могут быть выбраны из CD28, CD27, ICOS, 4-1ВВ, ОХ40, CD30, GITR, HVEM, DR3 или CD40.
В конкретном варианте осуществления один из (b) или (е) представляет собой CD28, а другой - 4-1ВВ или OX40.
В конкретном варианте осуществления (b) представляет собой CD28.
В другом конкретном варианте осуществления (е) представляет собой 4-1ВВ или ОХ40 и, в частности, представляет собой 4-1ВВ. В другом варианте осуществления (е) представляет собой CD27.
Трансмембранные домены (с) и (f), приведенных выше, могут быть одинаковыми или различными, но, в частности, являются различными, чтобы обеспечить разделение конструкций на поверхности клетки. Выбор различных трансмембранных доменов может также повысить стабильность вектора, поскольку включение прямой повторяющейся последовательности нуклеиновой кислоты в вирусном векторе делает его подверженным реаранжировке с удалением последовательностей между прямыми повторами. Если трансмембранные домены (с) и (f) являются одинаковыми, этот риск можно уменьшить путем модификации или "неоднозначности" кодонов, выбранных для кодирования одной и той же последовательности белка.
Подходящие трансмембранные домены известны из уровня техники и включают, например, трансмембранные домены CD8α, CD28, CD4 или CD3z.
Когда костимулирующая сигнальная область содержит CD28, описанный выше, трансмембранный домен CD28 представляет собой подходящий вариант. Белок с полноразмерным CD28 представляет собой белок из 220 аминокислот SEQ ID NO: 3
MLRLLLALNLFPSIQVTGNKILVKQSPMLVAYDNAVNLSCKYSYNLFSREFRASLHKGLDSAVEVCVVGNYSQQLQVYSKTGFNCDGKLGNESVTFYLQNLYVNQTDIYFCKIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAPIIPWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRS (SEQ ID NO: 3),
где трансмембранный домен показан жирным шрифтом.
В частности, одна из костимулирующих сигнальных областей основана на шарнирной области и, соответственно, трансмембранном домене и эндодомене CD28. В частности, который содержит аминокислоты 114-220 SEQ ID NO: 3, показанные ниже как SEQ ID NO: 4.
IEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRS (SEQ ID: NO: 4)
В конкретном варианте осуществления одна из костимулирующих сигнальных областей (b) или (е), приведенных выше, представляет собой модифицированную форму SEQ ID NO: 4, которая включает метку c-myc SEQ ID NO: 5.
Метка c-myc хорошо известна и представлена в SEQ ID NO: 5.
EQKLISEEDL (SEQ ID NO: 5)
Метка c-myc может быть добавлена в костимулирующую сигнальную область (b) или (е) путем введения в эктодомен или путем замены области в эктодомене, которая, следовательно, находится в области аминокислот 1-152 SEQ ID NO: 3.
В особенно предпочтительном варианте осуществления метка c-myc заменяет мотив MYPPPY в последовательности CD28. Этот мотив представляет собой потенциально опасную последовательность. Он отвечает за взаимодействие между CD28 и его природными лигандами, CD80 и CD86, таким образом, он обеспечивает возможность нецелевой токсичности, когда Т-клетки с CAR встречаются с клеткой-мишенью, которая экспрессирует любой из этих лигандов. Посредством замены этого мотива на последовательность метки, описанную выше, уменьшается потенциал для нежелательных побочных эффектов.
Таким образом, в конкретном варианте осуществления костимулирующая сигнальная область (b) конструкции представлена в SEQ ID NO: 6
IEVEQKLISEEDLLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRS (SEQ ID NO: 6)
Кроме того, включение эпитопа c-myc означает, что облегчается обнаружение Т-клеток с CAR с использованием моноклонального антитела. Это очень полезно, поскольку проточное цитометрическое обнаружение оказалось ненадежным при использовании некоторых доступных антител.
Кроме того, предоставление эпитопной метки c-myc может облегчать антиген-независимое размножение целевых Т-клеток с CAR, например, путем перекрестного сшивания CAR с использованием соответствующего моноклонального антитела либо в растворе, либо иммобилизованного на твердую фазу (например, мешок).
Более того, экспрессия эпитопа для антитела к c-myc человека, 9е10, в вариабельной области TCR, как было показано ранее, является достаточной для обеспечения опосредованной антителом и опосредованной комплементом цитотоксичности как in vitro, так и in vivo (Kieback et al. (2008) Proc. Natl. Acad. Sci. USA, 105(2) 623-8). Таким образом, предоставление таких эпитопных меток также может быть использовано в качестве "системы самоубийства", при которой антитело можно использовать для истощения Т-клеток с CAR in vivo в случае токсичности.
Связывающие элементы (d) и (g) будут различными и будут связывать одинаковые, перекрывающиеся или различные эпитопы. В конкретном варианте осуществления первый и второй эпитопы ассоциированы с одним и тем же рецептором или антигеном. Таким образом, первый и второй эпитопы, описанные выше, могут в некоторых случаях быть одинаковыми или перекрываться, так что связывающие элементы (d) и (g) будут конкурировать за их связывание. В качестве альтернативы, первый и второй эпитопы могут быть различными и быть ассоциированными с одинаковыми или различными антигенами в зависимости от конкретной предусмотренной терапии. В одном варианте осуществления антигены являются различными, но могут быть ассоциированы с одним и тем же заболеванием, таким как одна и та же специфическая форма рака.
Используемый здесь термин "антиген" относится к любому члену конкретной связывающей пары, которая будет связываться со связывающими элементами. Таким образом, этот термин включает рецепторы на клетках-мишенях.
Таким образом, подходящими связывающими элементами (d) и (g) могут являться любые элементы, которые обеспечивают pCAR возможностью распознавать мишень, представляющую интерес. Мишенью, на которую направлены pCAR по настоящему изобретению, может являться любая мишень, представляющая клинический интерес, к которой было бы желательно индуцировать ответ Т-клетки. Это включает маркеры, ассоциированные с раком различных типов, в том числе, например, один или несколько рецепторов ErbB или интегрин αvβ6 маркеры, ассоциированные с раком предстательной железы (например, с использованием связывающего элемента, который связывается с простато-специфическим мембранным антигеном(PSMA)), раком молочной железы (например, с использованием связывающего элемента, который целенаправленно воздействует на Her-2 (также известный как ErbB2)) и нейробластомами (например, с использованием связывающего элемента, который целенаправленно воздействует на GD2), меланомами, мелкоклеточной или немелкоклеточной карциномой легкого, саркомами и опухолями мозга. В конкретном варианте осуществления мишень представляет собой один или несколько димеров ErbB, описанных выше, или рецептор колониестимулирующего фактора-1 (CSF-1R) или интегрин αvβ6, все из которых вовлечены в несколько солидных опухолей.
Связывающие элементы, используемые в pCAR по настоящему изобретению, могут включать антитела, которые распознают выбранную мишень. Для удобства, антитело, используемое в качестве связывающего элемента, предпочтительно представляет собой одноцепочечное антитело (scFv) или однодоменное антитело от верблюда, человека или других видов. Одноцепочечные антитела можно клонировать из V-области генов гибридомы, специфических в отношении целевой мишени. Получение таких гибридом стало обычной процедурой, и в данном документе процедуру не будут повторять. Методика, которая может быть использована для клонирования вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL), была описана в Orlandi et al., Proc. Natl Acad. Sci. (США) 86: 3833-3837 (1989). Коротко, мРНК выделяют из линии клеток гибридомы и обратно транскрибируют в комплементарную ДНК (кДНК), например, используя набор для полимеразной цепной реакции с обратной транскриптазой (RT-PCR). Используют последовательность-специфические праймеры, соответствующие последовательности генов VH и VL. Анализ последовательности клонированных продуктов и сравнение с известной последовательностью генов VH и VL можно использовать, чтобы показать, что клонированный ген VH соответствует ожиданиям. Гены VH и VL затем соединяют друг с другом, например, с использованием олигонуклеотида, кодирующего линкер (gly4-ser)3.
В качестве альтернативы, связывающий элемент pCAR может включать лиганды, такие как пептид T1E (связывает гомо- и гетеродимеры ErbB), колониестимулирующий фактор-1 (CSF-1) или IL-34 (оба связываются с рецептором CSF-1). Пептид T1E представляет собой химерный слитый белок, состоящий из всего зрелого белка EGF человека, за исключением пяти последних N-концевых аминокислот (аминокислоты 971-975 предшественника про-эпидермального фактора роста (NP_001954.2)), которые были заменены семью последним N-концевым аминокислотам зрелого белка TGF-α человека (аминокислоты 40-46 альфа-изоформы 1 про-трансформирующего фактора роста (NP_003227.1)).
В другом варианте осуществления связывающий элемент pCAR включает интегрин αvβ6-специфичное связывающее средство. Интегрин αvβ6 в настоящее время рассматривается как мишень при раке, поскольку было установлено, что его экспрессия сильно повышена при многих типах рака. αvβ6 был идентифицирован как рецептор вируса ящура (FMDV) in vitro путем связывания посредством мотива RGD в вирусном капсидном белке VP1. В результате, как описано, например, в патенте США №8383593, ряд пептидов, полученных из FMDV и, в частности, пептиды, происходящие из белка VP1 из FMDV и содержащие мотив RGD, показали повышенную связывающую способность и специфичность связывания. В частности, эти пептиды содержат мотив последовательности
RGDLX5X6L (SEQ ID NO: 7) или
RGDLX5X6I (SEQ ID NO: 8),
где LX5X6L или LX5X6I содержатся в пределах альфа-спиральной структуры, где X5 и X6 представляют собой остатки, обеспечивающие образование спирали, которые имеют конформационное предпочтение, превышающее 1,0, для нахождения в середине [альфа]-спирали (из Creighton, 1993 и Расе CN и Scholtz JM (1998), Biophysical Journal, том 75, страницы 422-427). В частности, такие остатки независимо выбраны из группы, состоящей из Glu, Ala, Leu, Met, Gln, Lys, Arg, Val, Ile, Trp, Phe и Asp.
Конкретные примеры таких последовательностей включают SEQ ID NO: 9-11 или их варианты:
YTASARGDLAHLTTTHARHL (SEQ ID NO: 9)
GFTTGRRGDLATIHGMNRPF (SEQ ID NO: 10)
или
NAVPNLRGDLQVLAQKVART (SEQ ID NO: 11)
Эти пептиды могут образовывать определенную группу связывающих элементов для CAR по настоящей заявке.
Для отдельных злокачественных опухолей, таких как лимфома Ходжкина и некоторые виды рака молочной железы, двумя природными лигандами являются CSF-1 и IL-34, и они образуют особенно подходящие связывающие элементы для (d) и (g). Тем не менее, они связываются с различной аффинностью. Аффинность связывания может влиять на наблюдаемую активность. В этом случае может быть полезным обеспечить, чтобы связывающий элемент с более низкой аффинностью связывания использовали в качестве связывающего элемента (b) и чтобы элемент с более высокой аффинностью связывания использовали в качестве связывающего элемента (g). В частности, в варианте осуществления относительная аффинность CAR второго поколения (i) в отношении его родственной мишени ниже, чем у партнерского химерного костимулирующего рецептора на основе TNFR (ii). Это не исключает возможности использования фрагментов с высокой или низкой аффинностью в каждой позиции при условии сохранения этого отношения относительной аффинности. Таким образом, в случае настоящего изобретения в конкретном варианте осуществления связывающий элемент (b) представляет собой CSF-1, который имеет относительно низкую аффинность связывания, тогда как связывающий элемент (g) содержит IL-34, который имеет более высокую аффинность связывания.
Соответственно связывающий элемент ассоциирован с лидерной последовательностью, которая облегчает экспрессию на поверхности клетки. Многие лидерные последовательности известны из уровня техники, и они включают лидерную последовательность рецептора макрофагального колониестимулирующего фактора (FMS), или лидерную последовательность CD124.
В дополнительном варианте осуществления клетки, экспрессирующие pCAR, сконструированы для козкспрессии химерного рецептора цитокина, в частности 4αβ-химерного рецептора цитокина. В 4αβ эктодомен α-цепи рецептора IL-4 соединяется с трансмембранным доменом и эндодоменом β-цепи рецептора IL-2/15. Это обеспечивает селективное размножение и обогащение генетически сконструированных Т-клеток ex vivo посредством культивирования этих клеток в подходящей поддерживающей среде, которая в случае 4αβ, будет включать IL-4 в качестве единственной поддержки цитокинов. Аналогично, система может использоваться с химерным рецептором цитокина, в котором эктодомен α-цепи рецептора IL-4 соединяется с трансмембранным доменом и эндодоменом другого рецептора, который естественно связан с цитокином, который также связывается с общей γ цепью.
Как обсуждалось, эти клетки пригодны в терапии для стимулирования опосредованного Т-клетками иммунного ответа в отношении популяции клеток-мишеней. Таким образом, второй аспект настоящего изобретения предусматривает способ стимуляции опосредованного Т-клетками иммунного ответа в отношении популяции клеток-мишеней у нуждающегося в этом пациента, при этом указанный способ предусматривает введение пациенту популяции иммунореактивных клеток, описанных выше, где связывающие элементы (d) и (g) являются специфическими в отношении клетки-мишени.
В третьем аспекте настоящего изобретения предусматривается способ получения иммунореактивной клетки по любому из предыдущих пунктов, причем указанный способ предусматривает трансдуцирование клетки первой нуклеиновой кислотой, кодирующей CAR структуры (i), определенной выше; а также второй нуклеиновой кислотой, кодирующей CAR структуры (ii), определенной выше.
В частности, в этом способе лимфоциты пациента трансдуцируются нуклеиновыми кислотами, кодирующими CAR (i) и (ii). В частности, Т-клетки подвергают генетической модификации, например, с помощью ретровирус-опосредованной трансдукции, для введения нуклеиновых кислот, кодирующих CAR, в геном Т-клетки-хозяина, за счет чего обеспечивая стабильную экспрессию CAR. Затем они могут быть повторно введены пациенту необязательно после размножения, чтобы обеспечить благоприятный терапевтический эффект. Когда клетки, такие как Т-клетки, сконструированы для коэкспрессии химерного рецептора цитокина, такого как 4αβ, стадия размножения может включать стадию культивирования ex vivo в среде, которая содержит цитокин, такой как среда, содержащая IL-4 в качестве единственной поддержки цитокинов в случае 4αβ. В качестве альтернативы, химерный рецептор цитокина может включать эктодомен α-цепи рецептора IL-4, присоединенный к эндодомену, используемому общим цитокином γ с определенными свойствами, таким как IL-7. В этой установке размножение клеток в IL-4 может привести к меньшей дифференциацией клеток, используя естественную способность IL-7 достигать этого эффекта. Таким образом, можно обеспечить селективное размножение и обогащение генетически сконструированных Т-клеток с желаемым состоянием дифференциации.
В четвертом аспекте настоящего изобретения предусматривается комбинация первой нуклеиновой кислоты, кодирующей CAR (i), описанной выше, и второй нуклеиновой кислоты, кодирующей CCR (ii), описанной выше. Как указано ранее, эта комбинация упоминается как pCAR. Подходящие последовательности для нуклеиновых кислот будут очевидны для специалиста. Последовательности могут быть оптимизированы для применения в требуемой иммунореактивной клетке. Однако в некоторых случаях, как обсуждалось выше, кодоны могут отличаться от оптимума или "неоднозначного", чтобы избежать повторяющихся последовательностей. Конкретные примеры таких нуклеиновых кислот будут кодировать предпочтительные варианты осуществления, описанные выше.
Для достижения трансдукции нуклеиновые кислоты в четвертом аспекте настоящего изобретения подходящим образом вводят в вектор, такой как плазмида или ретровирусный вектор. Такие векторы, в том числе плазмидные векторы, или клеточные линии, содержащие их, образуют дополнительный аспект настоящего изобретения.
Первая и вторая нуклеиновые кислоты или векторы, содержащие их, могут быть объединены в наборе, который поставляется с целью создания иммунореактивных клеток по первому аспекту настоящего изобретения in situ.
Параллельные химерные активирующие рецепторы (pCAR), кодируемые описанными выше нуклеиновыми кислотами, образуют дополнительный аспект настоящего изобретения.
Подробное описание изобретения
Настоящее изобретение теперь будет, в частности, описано в качестве примера со ссылкой на следующие фигуры, на которых
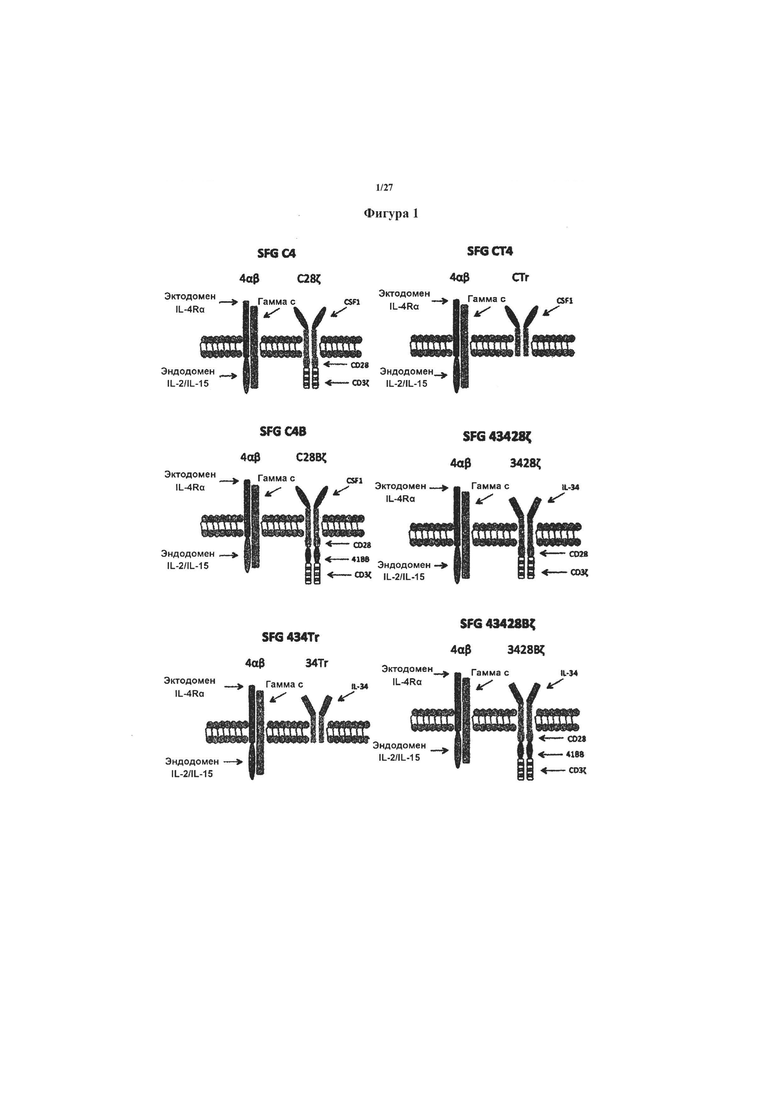
на фигуре 1 показана схематическая диаграмма, показывающая панель CAR и pCAR (называемая С34В и 34СВ), осуществляющая настоящее изобретение. Все CAR и pCAR коэкспрессировали в ретровирусном векторе SFG с 4αβ, химерном рецепторе цитокина, в котором эктодомен α-цепи рецептора IL-4 был слит с трансмембранным доменом и эндодоменом β-цепи рецептора IL-2. Применение 4αβ обеспечивает селективное обогащение и размножение генно-модифицированных Т-клеток посредством культивирования в IL-4, поскольку он вовлекает цепь гамма с (γс).
На фигуре 2 показаны результаты эксперимента с применением CAR, показанных на фигуре 1. Т-клетки (1×106 клеток), экспрессирующие эти CAR и pCAR (или нетрансдуцированные (UT) в качестве контроля), совместно культивировали in vitro в течение 24 часов с опухолевыми клетками T47D, которые экспрессируют (T47D-FMS) или не имеют (T47D) родственного целевого антигена (рецептор колониестимулирующего фактора-1 (CSF-1R), кодируемый c-fms). Оставшиеся жизнеспособные опухолевые клетки затем оценивали количественно посредством МТТ-анализа.
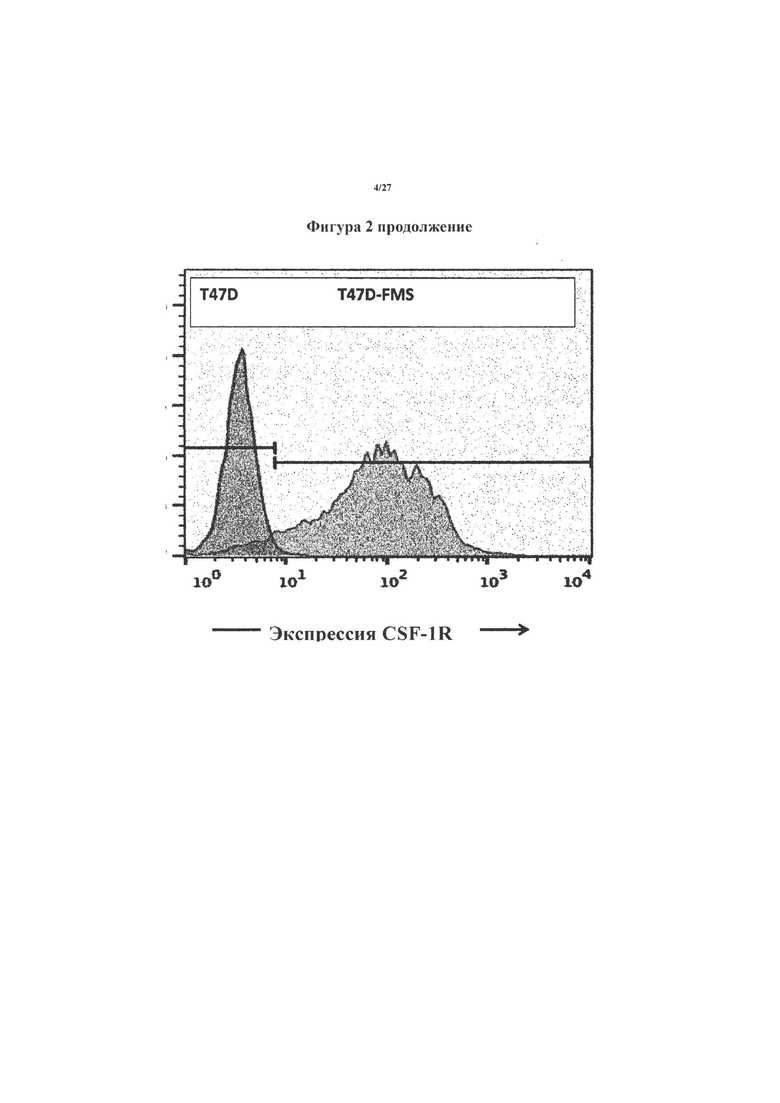
На фигуре 3 показан иллюстративный эксперимент, в котором Т-клетки, которые экспрессируют CAR и pCAR на фигуре 1 (или нетрансдуцированные Т-клетки в качестве контроля), подвергали последующим циклам стимуляции Ag в отсутствие экзогенного цитокина.
Стимуляцию обеспечивали посредством еженедельного культивирования на монослоях T47D FMS и в указанные промежутки времени подсчитывали количество Т-клеток.
На фигуре 4 показаны объединенные данные из 7 повторных экспериментов, аналогичных показанным на фигуре 3, что указывает на кратное размножение Т-клеток с CAR, которое происходило через неделю после каждого цикла стимуляции.
На фигуре 5 показаны иллюстративные анализы цитотоксичности, выполненные во время циклов стимуляции 2, 6, 9 и 12 в эксперименте, показанном на фигуре 3. Это следует из тестирования Т-клеток в отношении их способности уничтожать T47D FMS и немодифицированные монослои T47D (МТТ-анализ), через двадцать четыре часа после каждого цикла повторной стимуляции.
На фигуре 6 показаны результаты тестирования супернатанта, удаляемого из культур через один день после каждого цикла стимуляции, в отношении содержания IL-2 и IFN-γ с помощью ELISA.
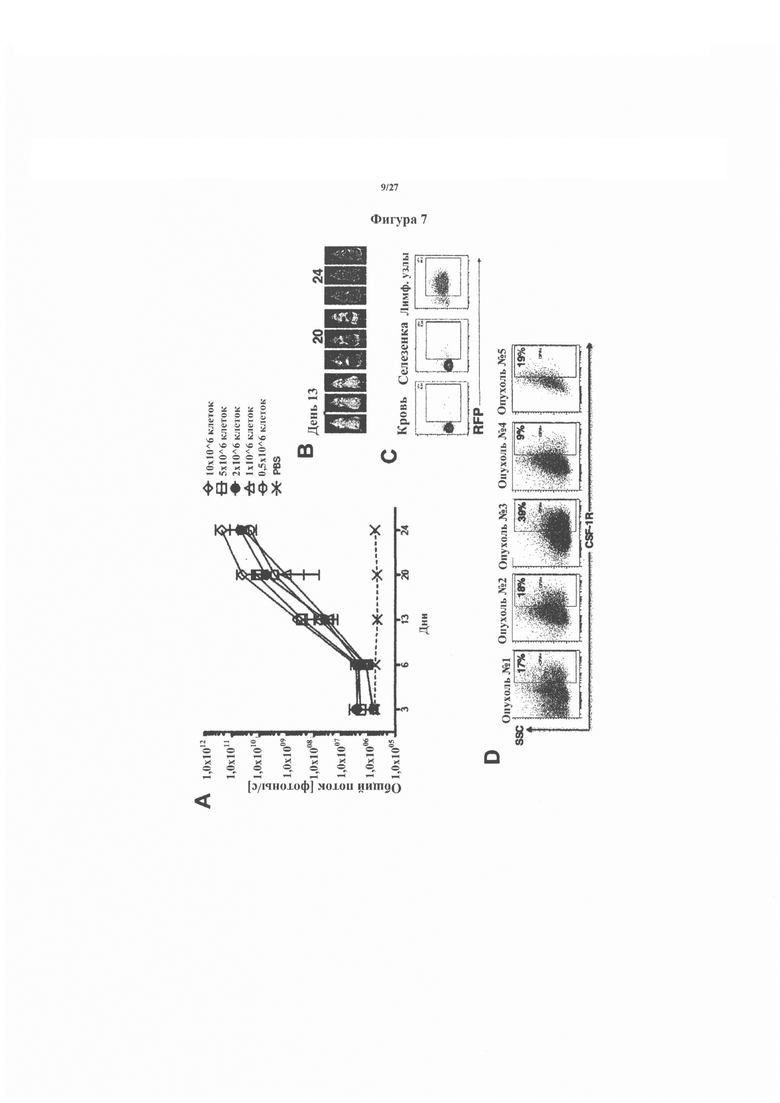
На фигуре 7 показано создание ксенотрансплантантной модели in vivo CSF-1R-экспрессирующей анапластической крупноклеточной лимфомы, которая обеспечила проведение последующего тестирования противоопухолевой активности CAR и pCAR-сконструированных Т-клеток. Модель была создана с использованием клеток K299, сконструированных для экспрессии люциферазы светлячков (luc) и красного флуоресцентного белка (RFP). На фигуре 7А показано образование опухоли после внутривенной инъекции указанных доз клеток K299 luc, количественно определяемых с помощью биолюминесцентной визуализации (BLI). Иллюстративные изображения BLI показаны на фигуре 7В у мышей, которые получили 2 миллиона опухолевых клеток. Показана экспрессия опухолевых клеток RFP+ (фигура 7С) в указанных тканях, что свидетельствует о том, что опухоли образовались только в лимфатических узлах этой модели. Экспрессия CSF-1R на пяти иллюстративных опухолях лимфатических узлов показана на фигуре 7D.
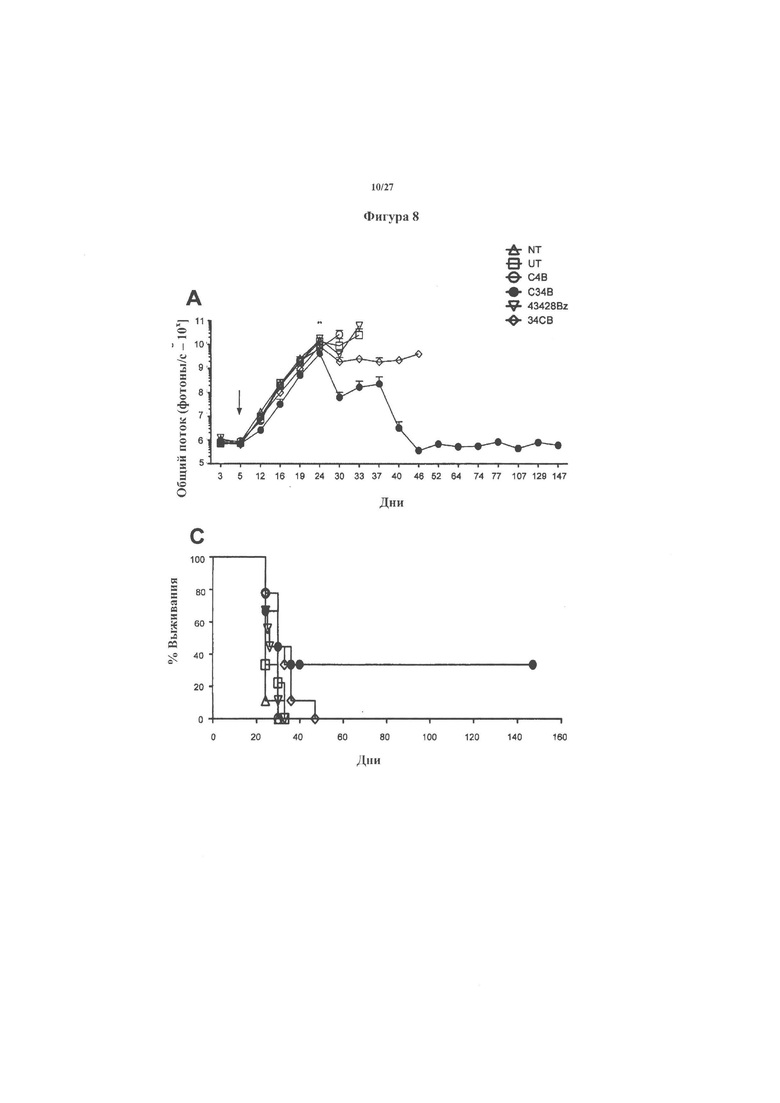
На фигуре 8 показаны результаты терапевтических исследований, в которых клетки K299 luc были введены внутривенно мышам SCID Beige (n=9 на группу, разделенных на 2 отдельных эксперимента). Через 5 дней мышей обрабатывали Т-клетками с CAR. Объединенное биолюминесцентное излучение от опухолей показана на фигуре 8А. Биолюминесцентное излучение у отдельных мышей показана на фигуре 8В, а выживаемость мышей показана на фигуре 8С.
На фигуре 9 показаны значения веса животных, используемые в терапевтическом исследовании с течением времени.
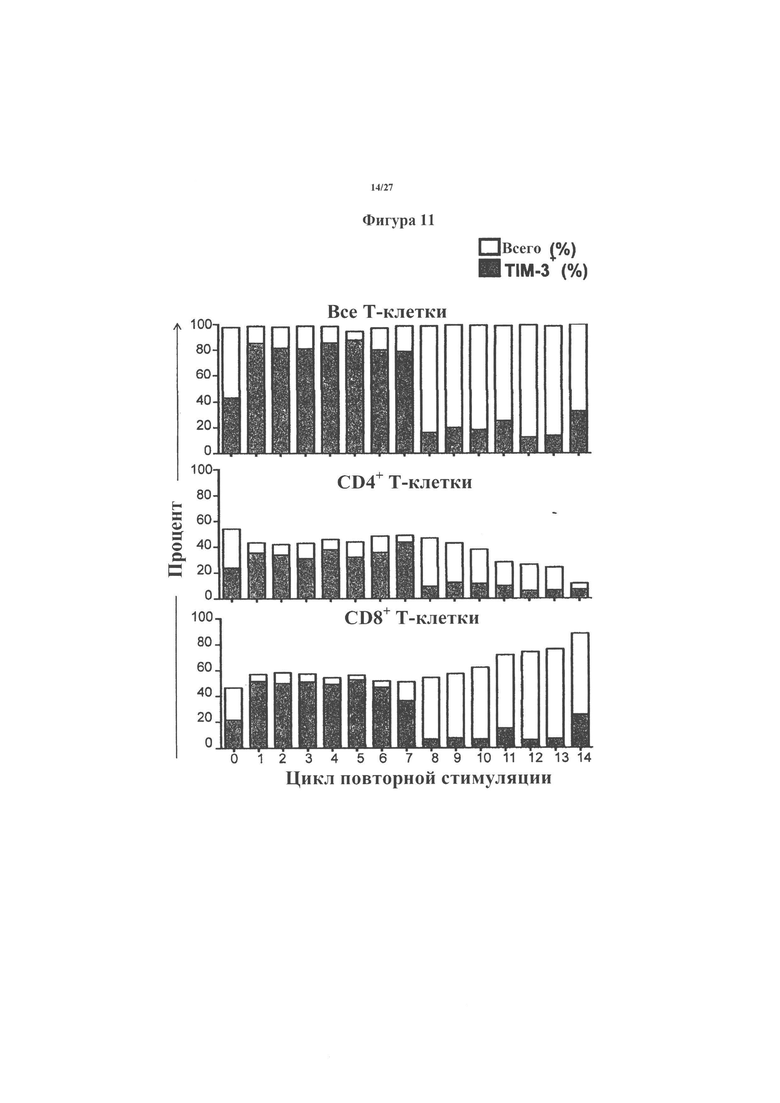
На фигурах 10-13 показаны результаты анализа экспрессии "маркеров истощения" из Т-клеток, экспрессирующих двойные CAR (С34В) по настоящему изобретению, где на фигуре 10 показаны результаты анализа PD1; на фигуре 11 показаны результаты анализа TIM3; на фигуре 12 показаны результаты анализа LAG3, а на фигуре 13 показаны результаты анализа 2В4.
На фигуре 14 показана схематическая диаграмма панели CAR и конструкций, целенаправленно воздействующих на интегрин αvβ6, которые были получены, в том числе pCAR (названный SFG TIE-41BB/A20-28z), осуществляющий настоящее изобретение. A20-28z представляет собой CAR второго поколения, который нацелен на применение пептида А20, полученного из вируса ящура. А20 связывается с высокой аффинностью с αvβ6 и с 85-1000-кратной более низкой аффинностью с другими RGD-связывающими интегринами. C20-28z представляет собой подобранный контроль, в котором ключевые элементы А20 были мутированы для отмены интегрин-связывающей активности. Все CAR были коэкспрессированы с 4αβ, как описано на фигуре 1.
На фигуре 15 представлена серия гистограмм, полученных с помощью проточной цитометрии, иллюстрирующих экспрессию интегрина в клетках А375 puro и Pane1. Клетки окрашивали антителом к β6 (Biogen Idec) с последующим вторичным антителами к РЕ мыши, антителами к αvβ3 или антителами к αvβ5 (оба конъюгированные с АРС, Bio-Techne). Рамки устанавливали на основе только вторичных антител или изотипических контролей.
На фигуре 16 представлена серия графиков, иллюстрирующих цитотоксичность CAR, в том числе pCAR по настоящему изобретению, целенаправленно воздействующих на αvβ6. Т-клетки, зкспрессирующие указанные CAR и pCAR, совместно культивировали с αvβ6-негативными (Panel и А375 puro) или αvβ6-позитивными (Вхрс3 и А375 puro β6) опухолевыми клетками. Данные показывают среднее ± SEM из 2-7 независимых экспериментов, каждое из которых выполняли в трех повторах. *р<0,05; **р<0,01; ***р<0,001.
На фигуре 17 представлена серия графиков, показывающих получение IFN-γ посредством CAR, в том числе pCAR по настоящему изобретению, целенаправленно воздействующих на αvβ6. Т-клетки, зкспрессирующие указанные CAR и pCAR, совместно культивировали с αvβ6-негативными (Panel и А375 puro) или αvβ6-позитивными (Вхрс3 и А375 puro β6) опухолевыми клетками. Данные показывают среднее ± SEM из 5-6 независимых экспериментов, каждое из которых выполняли в двух повторах. *р<0,05; **р<0,01; ***р<0,001; ns - незначимый.
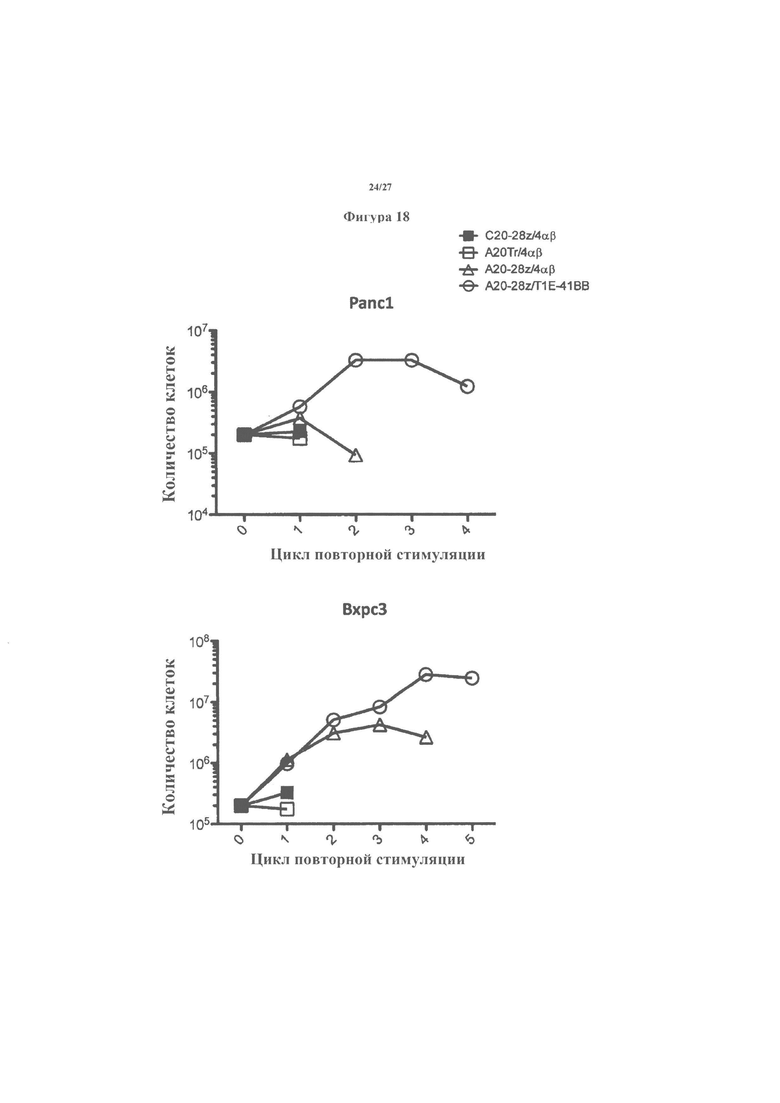
На фигуре 18 показаны результаты экспериментов по повторной стимуляции с применением CAR и pCAR-сконструированных Т-клеток, описанных выше, и указывающих способность Т-клеток A20-28z/T1E-41BB с pCAR переносить повторную стимуляцию антигеном, сопровождающуюся размножением Т-клеток и разрушением клеток-мишеней, которые экспрессируют (Вхрс3), или не экспрессируют (Pane1) интегрин αvβ6.
На фигуре 19 показаны результаты экспериментов по повторной стимуляции с использованием pCAR-сконструированных Т-клеток, в которых A20-28Z коэкспрессировали с Т1Е-41ВВ, T1E-CD27 или T1E-CD40, что обеспечило проведение сравнительной оценки совместной стимуляции дополнительными членами семейства рецепторов TNF. Контрольные Т-клетки являлись нетранедуцированными (NT), в то время как CAR содержали усеченные (tr) эндодомены. Т-клетки повторно стимулировали на клетках-мишенях, которые экспрессируют (Вхрс3), или не экспрессируют (Panc1) интегрин αvβ6, проводя сравнение с нестимулированными Т-клетками. В случае клеток Вхрс3 превосходное размножение (фигура 19А), сопровождающееся устойчивой цитотоксической активностью (фигура 19В), наблюдали с Т-клетками A20-28z/T1E-CD27. Напротив, с клетками Panel превосходное размножение (фигура 19А), сопровождающееся устойчивой цитотоксической активностью (фигура 19В), наблюдали с Т-клетками A20-28z/T1E-CD27. Эти данные показывают, что дополнительные члены семейства рецепторов TNF также могут оказывать совместную стимуляцию с применением формата pCAR.
Пример 1
Получали панели CAR, целенаправленно воздействующих на рецептор CSF-1 (кодируемую c-FMS), который является сверхэкспрессированым при лимфоме Ходжкина, анапластической крупноклеточной лимфоме и некоторых солидных опухолях, таких как трижды негативный рак молочной железы, и схематически проиллюстрированы на фигуре 1. Панель CAR включала CAR как второго, так и третьего поколения с любым из двух природных лигандов, CSF-1 или IL-34, в качестве целевых фрагментов. Хотя оба CSF-1 и IL-34 связываются с рецептором CSF-1, IL-34 связывается с гораздо более высокой аффинностью (в 34 раза выше, чем CSF-1).
Конструкции SFG 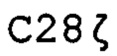 и SFG CTr клонировали в ретровирусном векторе SFG в виде фрагментов NcoI/XhoI, обеспечивая то, что их стартовые кодоны находятся на участке встречающегося в природе сайта NcoI, ранее занятого удаленным геном env. Экспрессия гена достигается из длинного концевого повтора (LTR) вируса мышиного лейкоза Молони (MoMLV), который характеризуется промоторной активностью, и вирусная упаковка РНК обеспечивается сигналом упаковки MoML ψ, который фланкирован участком донора сплайсинга и участком акцептора сплайсинга.
и SFG CTr клонировали в ретровирусном векторе SFG в виде фрагментов NcoI/XhoI, обеспечивая то, что их стартовые кодоны находятся на участке встречающегося в природе сайта NcoI, ранее занятого удаленным геном env. Экспрессия гена достигается из длинного концевого повтора (LTR) вируса мышиного лейкоза Молони (MoMLV), который характеризуется промоторной активностью, и вирусная упаковка РНК обеспечивается сигналом упаковки MoML ψ, который фланкирован участком донора сплайсинга и участком акцептора сплайсинга.
Все остальные конструкции сконструировали и клонировали с использованием способа клонирования с использованием полимеразы и удлинением неполностью комплементарных праймеров (PIPE). Способ клонирования PIPE является альтернативой, основанной на ПЦР, традиционным способам клонирования, зависимым от рестрикционного фермента и лигирования. Это устраняет необходимость включения сайтов рестрикции, которые могут кодировать дополнительные нежелательные остатки в экспрессированные белки. Способ PIPE основан на неэффективности процесса амплификации в заключительных циклах реакции ПЦР, возможно, из-за снижения доступности dNTP, что приводит к получению частично одноцепочечных (PIPE) ПЦР-продуктов с выступающими 5'-концами. Для векторной линеаризации ПЦР использовали набор вектор-специфических праймеров и другой набор праймеров с перекрывающимися последовательностями 5'-векторного конца, которые затем использовали для вставки амплификации, образуя неполные продукты удлинения с помощью PIPE. На следующей стадии продукты PIPE смешивали и одноцепочечные перекрывающиеся последовательности отжигали и собирали в виде полной конструкции SFG CAR. Успешное клонирование подтверждали диагностическим рестрикционным расщеплением. Секвенирование ДНК выполняли на всех конструкциях для подтверждения того, что предсказываемая кодирующая последовательность присутствовала без каких-либо ПЦР-индуцированных мутаций (Source Bioscience, Великобритания).
Панель включала два химерных активирующих рецептора (pCAR) ''двойного нацеливания'', в которых CSF-1 или IL-34 соединены с 28z и 4-1ВВ, или наоборот. Комбинации pCAR двойного нацеливания затем стехиометрически коэкспрессировали в одной и той же популяции Т-клеток с использованием Thosea Asigna (Т) 2А-содержащего ретровирусного вектора. Один из этих CAR обозначали как ''С34В'' (CSF1-28Z плюс IL34-41BB), а другой называли ''34СВ'' (IL34-28z плюс CSF1-41BB).
В этих Т-клетках с CAR двойного нацеливания оба костимулирующих мотива (CD28/4-1BB) помещали в их естественное месторасположение вблизи мембраны, физически отделяли друг от друга, и коэкспрессировали в одной и той же Т-клетке.
Все CAR коэкспрессировали с IL-4-реактивным 4αβ рецептором с использованием дополнительного Т2А элемента в векторе. Это позволяет обогащать/размножать Т-клетки с использованием IL-4, что облегчает сравнение функции этих разнообразных популяций клеток после отбора.
Основное внимание в экспериментах заключалось в проверке поведения Т-клеток при повторной стимуляции с опухолевыми клетками-мишенями, которые либо экспрессируют, либо не имеют целевой рецептор FMS/CSF-1. В каждом цикле 1 миллион указанных размноженный с помощью IL-4 Т-клеток с CAR суспендировали в RPMI + человеческой АВ-сывороткой и культивировали с конфлюентным монослоем (24-луночная планшет) антиген-экспрессирующей мишени (T47D FMS) или мишени без антигена (T47D).
После этого, если Т-клетки с CAR выживали и уничтожали монослой, 1 миллион Т-клеток удаляли и повторно стимулировали аналогичным образом каждую неделю. Общее количество клеток было экстраполировано в каждый момент времени в зависимости от размножения Т-клеток, которое происходило в каждом недельном цикле.
Во всех этих экспериментах Т-клетки культивировали в отсутствие какого-либо экзогенного цитокина, такого как IL-2 или IL-4, поэтому им пришлось продуцировать свои собственные цитокины, чтобы выживать и размножаться. Продукцию цитокинов (IFN-γ и IL-2) измеряли с помощью ELISA в супернатантах, собранных из совместных культур Т-клеток/опухолевых клеток, обеспечивая второй маркер эффективной костимуляции.
Было обнаружено (фигура 2), что при их первом воздействии на монослой опухоли, который экспрессирует мишень (FMS-кодированный CSF-1-рецептор), все CAR, которые, как предполагается, будут уничтожать, делают это (объединенные данные из 12 экспериментов). Контролем являются UT (нетрансдуцированные), Р4 (целенаправленно воздействует на нерелевантный антиген, PSMA) и СТ4, в котором эндодомен усечен. Как и ожидалось, ни одна из Т-клеток с CAR не уничтожает опухолевые клетки, у которых отсутствует рецептор CSF-1 (T47D).
Иллюстративный эксперимент повторной стимуляции показан на фигуре 3. Объединенные данные повторной стимуляции из 7 экспериментов показаны на фигуре 4. В этом случае пролиферация в первом цикле была подобной для большинства конструкций, хотя целевые конструкции второго и третьего поколений, целенаправленно воздействующие на IL-34, продемонстрировали худший результат. Это может быть связано с тем, что аффинность в отношении фрагмента, целенаправленно воздействующего на IL-34, является слишком большой.
Однако в более поздних циклах комбинация двойного pCAR С34В (28z CAR второго поколения, целенаправленно воздействующий на CSF-1, коэкспрессируемый с костимулирующим мотивом 4-1ВВ, целенаправленно воздействующим на IL-34) последовательно становилась очевидно превосходящей.
В эксперименте, показанном на фигуре 3, супернатант собирали через 24 часа после каждого цикла повторной стимуляции и анализировали в отношении содержания цитокинов (IFN-γ и IL-2) с помощью ELISA. Процент жизнеспособности оставшихся опухолевых клеток измеряли посредством МТТ-анализа (иллюстративные примеры, показанные на фигуре 5). Результаты продукции цитокинов показаны на фигуре 6. Было обнаружено, что только Т-клетки с С34В CAR сохраняют способность продуцировать IL-2 на протяжении каждого цикла стимуляции. Это утрачивалось всеми другими комбинациями CAR после первого цикла. Устойчивое сохранение способности вырабатывать IL-2 через рекурсивную повторную стимуляцию обычно не наблюдается с Т-клетками с CAR, и это предполагает, что эта доставка двойной костимуляции в корне меняет дифференциацию этих клеток in vitro, задерживая начало анергии.
Также контролировали количество жизнеспособных Т-клеток после разрушения монослоя в последовательных циклах Ag-стимуляции, и результаты показаны на фигуре 5. После второго цикла повторной стимуляции все CAR, за исключением С34В, начинали терять способность достигать CSF-1R-зависимого уничтожения опухолевых клеток. Напротив, Т-клетки, которые экспрессируют С34В, сохраняют антиген-зависимую активность в этом цитотоксическом анализе до 13 повторяющихся циклов повторной стимуляции, но никогда не вызывают цитотоксичность против немодифицированных клеток T47D.
Кроме того, на этих Т-клетках с помощью проточной цитометрии также измеряли так называемые "маркеры истощения'' (PD1, TIM3, 2В4 и LAG3). Результаты показаны на фигурах 10-13. Как и ожидалось, процент Т-клеток, которые экспрессировали различные маркеры истощения, постепенно увеличивался на повторно стимулированных Т-клетках, но это не замедляло пролиферацию, разрушение опухолевых клеток или выделение цитокинов клетками С34В при стимуляции антигенами. Это говорит о том, что превосходящая функция С34В не является результатом отсроченной регуляции маркеров истощения.
Таким образом, подход pCAR по настоящему изобретению, по-видимому, поддерживает клетки в состоянии, благодаря которому они сохраняют реактивность к антигену через большее количество циклов повторной стимуляции. Имеются свидетельства того, что он может замедлить дифференциацию за пределами контролируемого состояния памяти и, как представляется, задерживает начало анергии, сохраняя при этом способность клеток продуцировать IL-2 после активации.
Пример 2
Анализ эффектов in vivo
Панель CAR, используемую в примере 1, описанном выше, тестировали в отношении противоопухолевой активности с использованием высокоагрессивной ксенотрансплантной модели in vivo, в которой рецептор-мишень CSF-1 экспрессируется на низких уровнях и в которой заболевание распространяется по всем лимфатическим узлам (фигура 7). Опухолевые клетки были помечены люциферазой светлячков, обеспечивая неинвазивный контроль тяжести заболевания.
Мышей SCID/Beige рандомизировали на 6 групп (9 животных на группу объединены в двух независимых экспериментах) и инокулировали внутривенно (IV) опухолевыми клетками 2×106 К299, повторно суспендированными в 200 мкл PBS. На 5-й день группы обрабатывали с помощью одного из терапевтических режимов, указанных ниже.
• Группа С4В: 20×106 Т-клеток с С4В IV
• Группа С34В: 20×106 Т-клеток с С34В IV
• 43428Bz: 20×106 Т-клеток с 43428Bz IV
• Группа 34СВ: 20×106 Т-клеток с 34СВ IV
• Группа UT (нетрансдуцированные): 20×106 нетрансдуцированных Т-клеток IV
• Группа NT (необработанные): 200 мкл PBS IV
Рост опухоли контролировали с использованием биолюминесцентной визуализации (BLI) в соответствующие моменты времени на протяжении всего исследования.
Результаты показаны на фигуре 8. Опять же, наиболее эффективной системой была система pCAR, С34В, обозначенная более низким средним излучением BLI (фигуры 8А-В), задержкой развития опухоли или регрессии опухоли, что приводит к длительному выживанию мышей (фигура 8С).
Животных взвешивали на протяжении всего эксперимента, и значительная токсичность не отмечалась (фигура 9).
Пример 3
Выбор целевых фрагментов для конструирования pCAR, которые вызывают активацию Т-клеток αvβ6-зависимым образом.
Получали панели CAR, которые целенаправленно воздействуют на интегрин αvβ6 отдельно или вместе с расширенным семейством ErbB и схематично показаны на фигуре 14. Связывающим элементом, используемым в этом случае, являлся пептид А20 (SEQ ID NO: 11), полученный из GH-петли капсидного белка VP1 вируса ящура (серотип 01 BFS) (США 8927501). Его размещали ниже сигнального пептида CD124 и сливали с эндодоменами CD28 и  с образованием
с образованием  CAR 2-го поколения. Получали контроль
CAR 2-го поколения. Получали контроль  содержащий аналогичную конструкцию, но с рандомизированным нацеливающим пептидом (названным С20), в котором ключевой мотив RGDL был заменен на АААА. Второй контроль содержал А20, слитый с усеченным эндодоменом CD28 (А20-Tr).
содержащий аналогичную конструкцию, но с рандомизированным нацеливающим пептидом (названным С20), в котором ключевой мотив RGDL был заменен на АААА. Второй контроль содержал А20, слитый с усеченным эндодоменом CD28 (А20-Tr).
Для создания pCAR по настоящему изобретению (названный TIE-41BB/A20-28z), A20-28z был коэкспрессирован с химерным костимулирующим рецептором, содержащим целевой пептид pan-ErbB (T1E), слитый с CD8α-трансмембранным доменом и 41ВВ эндодоменом.
Там, где это указано, CAR коэкспрессировали с 4αβ химерным рецептором цитокинов, чтобы обеспечить опосредованное IL-4 обогащение in vitro. Эквимолярная коэкспрессия IL-4-реактивного 4αβ химерного рецептора цитокина, в котором α-эктодомен рецептора IL-4 слит с трансмембранным доменом и эндодоменом β-цепи общего рецептора IL-2/15, была достигнута с использованием Thosea Asigna (Т) 2А рибосомального пропускающего пептида. Эти химерные молекулы экспрессировали в Т-клетках человека посредством ретровирусного переноса гена.
Паттерн экспрессии интегрина в клеточных линиях рака А375 оценивали с использованием проточной цитометрии (фигура 15) и их разделяли на αvβ6-отрицательные (Panel и А375 puro) или αvβ6-положительные (Вхрс3 и А375 puro β6) опухолевые клетки. Эти клетки совместно культивировали с Т-клетками с CAR при соотношении эффектор : мишень 1:1 в течение либо 24, либо 28, либо 72 часов, после чего цитотоксичность оценивали с помощью МТТ-анализа и экспрессировали относительно необработанных опухолевых клеток. Результаты показаны на фигуре 16.
Эти данные показывают, что Т-клетки с CAR A20-28z уничтожают все клетки-мишени, которые экспрессируют интегрин αvβ6 (Вхрс3 и А375 β6 puro), но сохраняют мишени, у которых отсутствует этот интегрин (Panel и А375 puro). Во-вторых, контрольные CAR C20-28z и А20-Tr являются неактивными в этих анализах. В-третьих, Т-клетки, которые экспрессируют pCAR T1E-41BB/A20-28z, вызывают эффективное уничтожение клеток-мишеней, которые экспрессируют интегрин αvβ6 (Вхрс3 и А375 β6 puro). Все эти результаты соответствуют ожиданиям. Примечательно, однако, что Т-клетки, которые экспрессируют pCAR Т1Е-41BB/A20-28z, также вызывают уничтожение клеток-мишеней, у которых отсутствует αvβ6 (Panel и А375 puro). Это указывает на то, что в конфигурации pCAR способность пептида А20 связывать интегрины без αvβ6 с низкой аффинностью является достаточной для активации этих сконструированных Т-клеток.
Затем оценивали продукцию IFN-γ в pCAR и контрольных сконструированных Т-клетках. Опухолевые клетки, у которых отсутствовали αvβ36 (Panel и A375 puro) или которые экспрессировали αvβ6 (Вхрс3 и А375 puro β6), совместно культивировали с генетически сконструированными Т-клетками при соотношении эффектор : мишень 1:1 и супернатант собирали после 24, 48 или 72 часов. Уровни IFN-γ определяли количественно с помощью ELISA (eBioscience). Результаты показаны на фигуре 17. Как и ожидалось, контроли не образовывали значительных количеств IFN-γ, в то время как Т-клетки с CAR AB20-28z высвобождали IFN-γ при культивировании с αvβ6-положительными (Вхрс3 и А375 puro β6) опухолевыми клетками. Примечательно, что Т-клетки, которые экспрессируют pCAR по настоящему изобретению, TIE-41BB/A20z, продуцируют больше IFN-γ, чем Т-клетки A20-28z при культивировании с αvβ6-положительными (Вхрс3) опухолевыми клетками. Кроме того, Т-клетки TIE-41BB/A20z+ продуцировали IFN-γ при культивировании с αvβ6-негативными (Pane1 и А375 puro) опухолевыми клетками. Еще раз это демонстрирует, что в конфигурации pCAR низкоаффинное связывание пептида А20 с интегринами без αvβ6 является достаточным для инициации активации этих сконструированных Т-клеток.
Затем популяции Т-клеток с CAR повторно стимулировали раз в две недели в отсутствие поддержки IL-2 на опухолевых клетках Panel (αvβ6-отрицательные) или Вхрс3 (αvβ6-положительные). Опухолевые клетки совместно культивировали с Т-клетками-CAR, полученными от пациента с аденокарциномой протоков поджелудочной железы (PDAC) при соотношении эффектор : мишень 1:1 (фигура 18). Т-клетки первоначально добавляли при 2×105 клеток/лунка и подсчитывали через 72 часа после совместного культивирования для оценки размножения (верхние панели). Цитотоксичность оценивали через 72 часа после добавления Т-клеток с помощью МТТ-анализа (нижние панели). Если было достаточное количество Т-клеток (2×105), Т-клетки повторно стимулировали на свежем монослое опухоли, и процесс повторяли позже еще через 72 часа.
Результаты показаны на фигуре 18. Это иллюстрирует, что Т-клетки A20-28z/T1E-41BB+ подвергали нескольким циклам размножения, сопровождаемым высвобождением IL-2 (данные не показаны) и разрушением клеток αvβ6+ Вхрс3. И снова их также подвергли нескольким циклам размножения, сопровождаемым высвобождением IL-2 и разрушением опухолевых клеток Panel.
В целом результаты ясно показали, что pCAR, содержащий A20-28z/T1E-41ВВ, проявляет улучшенную функциональность in vitro по сравнению с CAR 2-го поколения, целенаправленно воздействующим на αvβ6. Кроме того, Т-клетки A20-28z/T1E-41BB+ также подвергали активации с помощью клеток Panel или А375 puro, которые экспрессируют от минимального до невыявляемого уровни этого интегрина. Принимая во внимание полученные данные с использованием С34В pCAR (примеры 1 и 2), это указывает на то, что конфигурация pCAR обеспечивает осуществление активации Т-клетки при последовательной повторной стимуляции, когда взаимодействие с высокой аффинностью связывания происходит с 41ВВ CCR, тогда как с более низкой аффинностью взаимодействия происходит с 28z CAR 2-го поколения.
Пример 4
Использование альтернативного члена семейства рецепторов TNF, CD27 для конструирования функционального pCAR.
Используя pCAR A20-28z/T1E-41BB в качестве исходного материала, сконструировали дополнительные pCAR, в которых модуль 41ВВ был заменен альтернативными членами семейства рецепторов TNF, а именно CD27 или CD40. Сконструировали контрольные pCAR, в которых эндодомены были усечены (tr). Клетки-мишени, зкспрессирующие (Вхрс3) или не имеющие (Panel) αvβ6, высевали при плотности 5×104 клеток на лунку 24-луночного планшета. Через 24 часа 5×104 Т-клеток с pCAR добавляли к клеткам-мишеням или в пустые лунки ("нестимулированные") без поддержки экзогенных цитокинов. Спустя еще 72 часа Т-клетки собирали из лунок и подсчитывали (фигура 19А). Проводили МТТ-анализ для определения процентной жизнеспособности оставшихся клеток-мишеней, сравнивая с контрольными клетками-мишенями, которые были высеяны без добавления Т-клеток (фигура 19В). Если Т-клетки пролиферировали после каждого цикла стимуляции, их повторно стимулировали на свежих клетках-мишенях, точно так же, как описано выше. Пролиферацию Т-клеток с pCAR (фигура 19А) и МТТ-анализ (фигура 19В) проводили через 72 часа, как и раньше. Повторяющуюся повторную стимуляцию Т-клеток с pCAR и оценку уничтожения клеток-мишеней продолжали таким образом, пока Т-клетки больше не пролиферировали в течение каждого 72-часового цикла.
Эти данные еще раз подтверждают превосходную функциональность А20-28z/T1E-41BB pCAR, когда Т-клетки стимулируются клетками-мишенями Panel, что указывает на устойчивую пролиферацию Т-клеток и уничтожение опухолевых клеток. Это обеспечивает дальнейшее подтверждение того, что связывание с низкой аффинностью пептида А20 с интегринами без αvβ6 является достаточным для инициации активации этих сконструированных Т-клеток. Примечательно, однако, что А20-28z/T1E-CD27 pCAR достигла наибольшего уровня пролиферации (фигура 19А) и продолжительного уничтожения опухолевых клеток (фигура 19В) при повторной стимуляции на αvβ6-экспрессирующих клетках Вхрс3. Напротив, pCAR на основе CD40 демонстрировали умеренную функцию в этих анализах. Вместе эти данные показывают, что ряд членов семейства рецепторов TNF можно использовать для разработки pCAR, которые демонстрируют превосходную функциональность, примером которой является CD27 или 41ВВ.
--->
SEQUENCE LISTING
<110> King's College London
<120> Therapeutic Agents
<130> P3052/WO
<150> GB1513540.3
<151> 2015-07-31
<160> 11
<170> BiSSAP 1.3.6
<210> 1
<211> 112
<212> PRT
<213> Homo sapiens
<400> 1
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 2
<211> 112
<212> PRT
<213> Homo sapiens
<400> 2
Arg Val Lys Phe Ser Arg Ser Ala Glu Pro Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 3
<211> 220
<212> PRT
<213> Homo sapiens
<400> 3
Met Leu Arg Leu Leu Leu Ala Leu Asn Leu Phe Pro Ser Ile Gln Val
1 5 10 15
Thr Gly Asn Lys Ile Leu Val Lys Gln Ser Pro Met Leu Val Ala Tyr
20 25 30
Asp Asn Ala Val Asn Leu Ser Cys Lys Tyr Ser Tyr Asn Leu Phe Ser
35 40 45
Arg Glu Phe Arg Ala Ser Leu His Lys Gly Leu Asp Ser Ala Val Glu
50 55 60
Val Cys Val Val Tyr Gly Asn Tyr Ser Gln Gln Leu Gln Val Tyr Ser
65 70 75 80
Lys Thr Gly Phe Asn Cys Asp Gly Lys Leu Gly Asn Glu Ser Val Thr
85 90 95
Phe Tyr Leu Gln Asn Leu Tyr Val Asn Gln Thr Asp Ile Tyr Phe Cys
100 105 110
Lys Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu Lys Ser
115 120 125
Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro Ser Pro
130 135 140
Leu Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val Val Gly
145 150 155 160
Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile
165 170 175
Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met
180 185 190
Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro
195 200 205
Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser
210 215 220
<210> 4
<211> 107
<212> PRT
<213> Homo sapiens
<400> 4
Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu Lys Ser Asn
1 5 10 15
Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro Ser Pro Leu
20 25 30
Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val Val Gly Gly
35 40 45
Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe
50 55 60
Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn
65 70 75 80
Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr
85 90 95
Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser
100 105
<210> 5
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> tag
<400> 5
Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu
1 5 10
<210> 6
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> costimulatory signalling region
<400> 6
Ile Glu Val Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Leu Asp Asn
1 5 10 15
Glu Lys Ser Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys
20 25 30
Pro Ser Pro Leu Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val
35 40 45
Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala
50 55 60
Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser
65 70 75 80
Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His
85 90 95
Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser
100 105 110
<210> 7
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic peptide
<220>
<221> SITE
<222> 5,6
<223> Xaa is any amino acid residue
<400> 7
Arg Gly Asp Leu Xaa Xaa Leu
1 5
<210> 8
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic peptide
<220>
<221> SITE
<222> 5,6
<223> Xaa is any residue
<400> 8
Arg Gly Asp Leu Xaa Xaa Ile
1 5
<210> 9
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic peptide
<400> 9
Tyr Thr Ala Ser Ala Arg Gly Asp Leu Ala His Leu Thr Thr Thr His
1 5 10 15
Ala Arg His Leu
20
<210> 10
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic peptide
<400> 10
Gly Phe Thr Thr Gly Arg Arg Gly Asp Leu Ala Thr Ile His Gly Met
1 5 10 15
Asn Arg Pro Phe
20
<210> 11
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic peptide
<400> 11
Asn Ala Val Pro Asn Leu Arg Gly Asp Leu Gln Val Leu Ala Gln Lys
1 5 10 15
Val Ala Arg Thr
20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ И СПОСОБЫ ИММУНОТЕРАПИИ | 2014 |
|
RU2729118C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНОВ | 2015 |
|
RU2745705C2 |
| АНТИТЕЛО К IL-13RA2 И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2756623C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИММУНОТЕРАПИИ | 2013 |
|
RU2832516C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИММУНОТЕРАПИИ | 2013 |
|
RU2729401C2 |
| МЕЗОТЕЛИНОВЫЕ CAR-РЕЦЕПТОРЫ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2822193C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА АНТИГЕН СОЗРЕВАНИЯ В-КЛЕТОК, И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2761632C2 |
| НЕ РЕСТРИКТИРОВАННЫЕ ПО HLA T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812917C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИММУНОТЕРАПИИ | 2014 |
|
RU2798348C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ К КАППА-АНТИГЕНУ МИЕЛОМЫ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2743188C2 |




















Настоящее изобретение относится к области иммунологии. Предложена иммунореактивная клетка для лечения рака, экспрессирующая химерный антигенный рецептор второго поколения и химерный костимулирующий рецептор. Кроме того, рассмотрен способ получения иммунореактивной клетки, комбинация нуклеиновых кислот, вектор, комбинация векторов, способ стимуляции иммунного ответа. Данное изобретение может найти дальнейшее применение в терапии различных видов рака. 6 н. и 14 з.п. ф-лы, 25 ил., 4 пр.
1. Иммунореактивная клетка для лечения рака, экспрессирующая
(i) химерный антигенный рецептор второго поколения, содержащий
(a) сигнальную область;
(b) костимулирующую сигнальную область;
(с) трансмембранный домен и
(d) связывающий элемент, который специфически взаимодействует с первым эпитопом на целевом раковом антигене; а также
(ii) химерный костимулирующий рецептор, содержащий
(e) костимулирующую сигнальную область, которая отличается от области (b);
(f) трансмембранный домен и
(g) связывающий элемент, который специфически взаимодействует со вторым эпитопом на целевом раковом антигене,
где аффинность связывания связывающего элемента (d) не выше, чем аффинность связывания связывающего элемента (g); и
где иммунореактивная клетка выбрана из цитотоксической T-клетки, Т-клетки-хелпера и естественной клетки-киллера (NK).
2. Иммунореактивная клетка по п. 1, где сигнальная область (а) содержит внутриклеточный домен CD3-[дзета]-цепи человека или его вариант.
3. Иммунореактивная клетка по п. 1 или 2, где костимулирующие сигнальные области (b) и (e) выбраны из CD28, CD27, ICOS, 4-1BB, OX40, CD30, GITR, HVEM, DR3 или CD40.
4. Иммунореактивная клетка по п. 3, где один из (b) или (e) представляет собой CD28, а другой представляет собой 4-1BB или OX40.
5. Иммунореактивная клетка по п. 4, где (b) представляет собой CD28.
6. Иммунореактивная клетка по п. 3, где (b) представляет собой CD28 и (e) представляет собой 4-1BB или CD27.
7. Иммунореактивная клетка по любому из пп. 1-6, где трансмембранные домены (с) и (f) выбраны из трансмембранных доменов CD8α и CD28.
8. Иммунореактивная клетка по любому из пп. 1-7, где первый и второй эпитопы ассоциированы с одним и тем же раковым антигеном.
9. Иммунореактивная клетка по любому из пп. 1-8, которая коэкспрессирует химерный рецептор цитокина.
10. Иммунореактивная клетка по п. 9, где химерный рецептор цитокина представляет собой 4αβ.
11. Иммунореактивная клетка по любому из пп. 1-10, где по меньшей мере один из связывающего элемента (d) или связывающего элемента (g) является лигандом для димера ErbB, рецептора для колониестимулирующего фактора-1 (CSF-1R) или связывающим средством, специфическим в отношении интегрина αvβ6.
12. Иммунореактивная клетка по любому из пп. 1-11, где связывающий элемент (d) содержит CSF-1 и связывающий элемент (g) содержит IL-34.
13. Иммунореактивная клетка по любому из пп. 1-7 и 9-11, где связывающий элемент (d) представляет собой связывающее средство, специфическое в отношении интегрина αvβ6, которое является пептидом, содержащим мотив последовательности
RGDLX5X6L (SEQ ID NO: 7) или
RGDLX5X6I (SEQ ID NO: 8),
где LX5X6L или LX5X6I содержатся в пределах альфа-спиральной структуры, где X5 и X6 представляют собой остатки, обеспечивающие образование спирали, и связывающий элемент (g) представляет собой пептид TIE.
14. Иммунореактивная клетка по любому из пп. 1-13, где аффинность связывания связывающего элемента (d) ниже, чем связывающего элемента (g).
15. Способ получения иммунореактивной клетки по любому из пп. 1-14, при этом указанный способ предусматривает трансдуцирование клетки первой нуклеиновой кислотой, кодирующей CAR структуры (i), определенный в п. 1, а также второй нуклеиновой кислотой, кодирующей CCR структуры (ii), определенный в п. 1.
16. Способ по п. 15, где иммунореактивная клетка содержит химерный рецептор цитокина и где стадию размножения осуществляют в присутствии указанного цитокина.
17. Комбинация первой нуклеиновой кислоты, кодирующей CAR (i), определенный в п. 1, и второй нуклеиновой кислоты, кодирующей CCR (ii), определенный в п. 1, для получения иммунореактивной клетки по любому из пп. 1-14.
18. Вектор, содержащий комбинацию по п. 17, для получения иммунореактивной клетки по любому из пп. 1-14.
19. Комбинация векторов, содержащая комбинацию по п. 17, для получения иммунореактивной клетки по любому из пп. 1-14.
20. Способ стимуляции опосредованного Т-клетками иммунного ответа в отношении популяции клеток-мишеней у нуждающегося в этом пациента, при этом указанный способ предусматривает введение пациенту иммунореактивной клетки по любому из пп. 1-14, описанной выше, где связывающие элементы (d) и (g) являются специфическими в отношении клетки-мишени.
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| KLOSS CC et | |||
| al | |||
| "Combinatorial antigen recognition with balanced signaling promotes selective tumor eradication by engineered T cells", Nature Biotechnology, Vol | |||
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Контрольный стрелочный замок | 1920 |
|
SU71A1 |
| LANITIS E et | |||
| al | |||
| "Chimeric Antigen Receptor T Cells with Dissociated Signaling Domains Exhibit Focused Antitumor Activity | |||

