Область технического применения
Настоящее изобретение относится к слитым молекулам, обладающим активностью индуцировать невоспалительный фагоцитоз, и обеспечивает возможность использования слитых молекул для профилактики или лечения заболеваний, вызванных аномальным отложением веществ, например протеинопатии.
Предпосылки создания изобретения
Многочисленные дегенеративные заболевания характеризуются аберрантным фолдингом, полимеризацией и отложением белков. Такие протеинопатии включают различные виды амилоидоза.
Амилоидоз — это заболевание, при котором в тканях откладываются специфические белки. Амилоид — это белковые агрегаты, имеющие от 7 до 13 нм в диаметре и обладающие бета-складчатой структурой с фибриллярной морфологией, которая видна под микроскопом при окрашивании Тиофлавином Т и Конго красным. Обычно в организме нет амилоида, и в настоящее время идентифицировано 36 амилоидогенных белков (Picken, Acta Haematol. (2020), 143:322-334). К типичным примерам амилоидных заболеваний относятся болезнь Альцгеймера, болезнь Паркинсона, болезнь Гентингтона и прионная болезнь. Кроме того, существует ряд амилоидных заболеваний с различными проявлениями, зависящими от белков, вызывающих амилоидоз, и затронутых органов.
Болезнь Альцгеймера — наибольшая причина развития деменции и смертельное заболевание, при котором наблюдаются нарушения функции приобретений знаний и памяти. Ожидается, что 130 миллионов человек пострадают от болезни Альцгеймера до 2050 года по всему миру, а 1 из 9 человек в популяции старше 65 лет уже поставлен диагноз болезни Альцгеймера.
Отличительный признак болезни Альцгеймера состоит в том, что белок бета-амилоид (Aβ) продуцируется при аномальном расщеплении предшественника бета-амилоида (APP), который отлагается и скапливается около мембраны клеток головного мозга. Другой особенностью является аномальное гиперфосфорилирование микротрубочек, связанных с тау-белком.
Недавно было показано, что бета-амилоидные олигомеры и фибриллы вызывают синаптическую дисфункцию и цитотоксичность по различным путям и создают замкнутый цикл, негативно влияющий на нервные клетки путем функциональных изменений в астроцитах и микроглии, отвечающих за иммунные свойства головного мозга.
Терапевтические лекарственные препараты, одобренные FDA для болезни Альцгеймера к настоящему моменту — это препараты, ингибирующие распад ацетилхолина или ингибирующие активность NMDA-рецепторов. Эти средства предназначены для временного облегчения симптомов, а не для фундаментального лечения. Поэтому в настоящее время нет методов принципиального лечения болезни Альцгеймера и, таким образом, известно, что болезнь Альцгеймера требует наиболее дорогостоящего лечения и медицинского ухода в случае пациентов среди стареющего населения.
Для фундаментального лечения болезни Альцгеймера десятилетиями проводилась разработка лекарственных препаратов, направленных на ингибирование или элиминацию (удаление) бета-амилоида. Однако большинство лекарственных препаратов для лечения болезни Альцгеймера, направленных на ингибирование или элиминацию бета-амилоида, признаны неудачными ввиду их неэффективности на клинической стадии. Например, в случае ингибиторов бета-секретазы для снижения бета-амилоида, стратегия, направленная на предотвращение дополнительного образования бета-амилоида очень неэффективна, поскольку у пациентов с болезнью Альцгеймера с нарушениями когнитивных функций уже имеются бета-амилоидные бляшки и происходит гибель нейронных клеток.
Поскольку недавние исследования показали, что моноклональные антитела, образующие специфические связи с бета-амилоидными олигомерами и фибриллами, способствуют удалению бета-амилоида и восстановлению когнитивных функций у пациентов с болезнью Альцгеймера, то появилась надежда на лечение болезни Альцгеймера с использованием антител к бета-амилоиду.
Предполагаемый в настоящее время механизм действия бета-амилоидных моноклональных антител включает ингибирование агрегации бета-амилоидных олигомеров и фибрилл путем образования с ними связей или индуцирование микроглиального фагоцитоза бета-амилоида посредством Fc-рецепторов, которые распознают моноклональные антитела.
Однако, несмотря на разработку терапевтических лекарственных средств для болезни Альцгеймера, современная иммунотерапия с использованием анти-бета-амилоидных антител выявляет изображения связанных с амилоидом аномалий (ARIAs, сопровождающихся образованием тяжелых отеков у 55% пациентов, проходивших лечение антителами. По этой причине 35% пациентам ARIA было отменено участие в клинических испытаниях. Известно, что явление ARIA происходит в результате синаптотоксичности и цитотоксичности, вызванных воспалительным ответом, который неминуемо возникает, когда анти-бета-амилоидные антитела стимулируют Fc-рецепторы микроглиальных клеток.
Поскольку синапсы и нейроны головного мозга обладают чувствительностью к воспалительным цитокинам, лечение с использованием анти-бета-амилоидных антител имеет проблему, заключающуюся в том, что происходит неотвратимое повреждение нейронов и синапсов, даже при удалении бета-амилоида до некоторой степени. Помимо моноклональных антител, такие компании как Alector и Denali, представляют стратегии усовершенствования способности микроглии по удалению бета-амилоида путем активации таких мишеней как TREM2, регулирующих иммунологический механизм микроглии, и этим стратегиями уделено большое внимание. Однако даже при осуществлении этих стратегий при чрезмерной активации микроглии ожидается синаптическое нарушение вследствие повышения фагоцитирующей способности.
Таким образом, важное направление в лечении болезни Альцгеймера состоит в разработке метода селективного удаления только бета-амилоидных олигомеров и фибрилл без возникновения воспалительного ответа и синаптического нарушения. Таким образом, важное направление в лечении болезни Альцгеймера состоит в разработке метода. Такие лекарственные препараты внесут существенный вклад в лечение болезни Альцгеймера.
Кроме того, если метод селективного удаления направлен только на элиминацию атипичного накопления вещества в качестве мишени, например, на аномальную аккумуляцию белков, вызывающих протеинопатию, без возникновения воспалительного ответа и последующего повреждения тканей, как было сказано выше, то возможна разработка метода селективного удаления аномального накопления белков, таких как тау, альфа-синуклеин и гентингтин. Ожидается, что такие лекарственные препараты внесут существенный вклад в лечение не только неврологических заболеваний, таких как болезнь Гентингтона, но также и заболеваний, связанных с аномальным накоплением определенных веществ.
Раскрытие изобретения
Техническая задача
Настоящее изобретение относится к слитым молекулам, имеющим фагоцитоз-индуцирующую активность, и настоящее изобретение предлагает возможность применения слитых молекул для предотвращения или лечения заболеваний, вызванных аномальным накоплением таргетных веществ.
Задача, решаемая в рамках настоящего изобретения, не ограничена вышеуказанной, и другие задачи, не обозначенные в настоящем описании, будут ясно поняты специалистами в данной области техники, исходя из следующего описания.
Техническое решение
В одном аспекте настоящего изобретения предлагается слитая молекула, имеющая фагоцитоз-индуцирующую активность, причем слитая молекула включает: первый участок, способный связываться с TAM-рецептором (Tyro3, Axl и Mer тирозинкиназные рецепторы; и второй участок, который образует специфические связи с таргетным веществом.
В данном случае, TAM-рецептором может являться по крайней мере один рецептор из группы, включающей Tyro3, Axl, и MerTK, которые способны индуцировать фагоцитоз путем связывания с ламинин G-подобным доменом (или LG-доменом).
Первый участок может включать Gas6, ProS1, Tubby, Tulp1, Gal3 или их активные фрагменты. Тип или диапазон не ограничен при условии, что первый участок имеет постоянное свойство индуцировать фагоцитоз путем взаимодействия с TAM-рецептором. Первый участок может предпочтительно включать Gas6, ProS1 или их активный фрагмент.
Точнее сказать, первый участок может включать ламинин G-подобный домен Gas6, ProS1 или их активный фрагмент, содержащий ламинин G-подобный домен в качестве связанной с фагоцитозом мостиковой молекулы, которая с избытком экспрессирована в различных тканях и, таким образом, способна индуцировать фагоцитоз посредством TAM-рецептора.
При этом ламинин G-подобный домен может включать LG1-домен, LG2-домен или их комбинацию, или предпочтительно включать как LG1-домен, так и LG2-домен, которые способны индуцировать фагоцитоз при связывании с TAM-рецептором.
Первый участок может быть пептидом, включающим последовательность, выбранную по крайней мере из одной из последовательностей SEQ ID NO: 1 и SEQ ID NO: 2 или последовательность, выбранную по крайней мере из одной из последовательностей SEQ ID NO: 3 и SEQ ID NO: 4. Предпочтительно, чтобы первый участок мог быть пептидом, включающим по крайней мере одну из последовательностей SEQ ID NO: 1 и SEQ ID NO: 2, по крайней мере одну из последовательностей SEQ ID NO: 3 и SEQ ID NO: 4 и предпочтительно, чтобы первый участок мог быть пептидом, включающим последовательность, выбранную по крайней мере из одной из последовательностей SEQ ID NO: 5 или SEQ ID NO: 6. Пептид, содержащий любую из вышеупомянутых SEQ ID NOS, включает не только аминокислотную последовательность пептида, но и ее вариант. Термин «вариант последовательности» относится к белку, имеющему последовательность, в которой один или несколько аминокислотных остатков отличаются от остатков исходной аминокислотной последовательности. При условии поддержания активности слитой молекулы в конечной структуре белка возможны любые усечение, делеция, инсерция, замещение или их комбинация. Одним из примеров варианта последовательности является форма, в которой аминокислотные остатки в участках, не определяющих активность, усечены или удалены, или аминокислотные остатки в участках, которые являются важными для аутоингибирования, замещены. В некоторых случаях последовательность может быть модифицирована при помощи фосфорилирования, гликозилирования, метилирования, фарнесилирования или им подобного. Эти варианты или модификации последовательности наиболее предпочтительны, когда функционирование и/или стабильность (температурная стабильность, рН-стабильность, структурная стабильность и т.д.) и/или растворимость белка повышаются за счет мутации в аминокислотной последовательности.
Метод мутагенеза аминокислотной последовательности основан на получении молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, соответствующую аминокислотной последовательности, которая подвергается мутации при мутировании нуклеотидной последовательности, кодирующей белок, причем способ получения гена, кодирующего белок, может осуществляться in vivo или in vitro с использованием любой методики мутагенеза, хорошо известной в данной области, например, сайтнаправленного мутагенеза (Hutchinson et al., J. Biol. Chem., 253:6551, 1978; Zoller and Smith, DNA, 3:479-488, 1984; Oliphant et al., Gene, 44:177, 1986; Hutchinson et al ., Proc. Natl. Acad. Sci. U.S.A., 83:710, 1986), TAB linker (Pharmacia), PCR technique (Higuchi, 1989, "Using PCR to Engineer DNA" in PCR Technology: Principles and Applications for DNA Amplification, H. Erlich, ed., Stockton Press, Chapter 6, pp. 61-70), or the like.
Кроме того, когда первый участок включает ламинин G-подобный домен Gas6 или ProS1 или его активный фрагмент, первый участок может не содержать Gla-домен. В этом случае первый участок может не распознавать фосфатидилсерин (ФС), в то время как второй участок способен индуцировать фагоцитоз, распознавая таргетное вещество.
Кроме того, когда первый участок включает ламинин G-подобный домен Gas6 или ProS1 или его активный фрагмент, первый участок не содержит Gla-домен и EGF-домен. В этом случае возможно, что достигается не только технический эффект, связанный с отсутствием вышеуказанного Gla-домена, но также и увеличивается выход за счет подавления агрегации в процессе очистки слитой молекулы.
Таргетным веществом (веществом-мишенью) может быть вещество, накапливающееся в живых тканях, что вызывает заболевание. Например, это может быть вещество, накапливающееся в затронутых болезнью тканях пациента. Веществом, накапливающимся при заболеваниях, может быть белок. Таким заболеванием среди прочих может быть протеинопатия. Например, таргетным веществом может быть амилоид. То есть протеинопатия может быть амилоидозом. Таргетное вещество может быть одним из аномально накопившихся веществ, приведенных в таблице 1 ниже, и в этом случае аномальное накопление каждого из указанных белков может вызывать заболевание. Например, протеинопатия может быть болезнью Альцгеймера, болезнью Паркинсона, болезнью Гентингтона и прионной болезнью, и в этом случае таргетное вещество может быть аномально накопленным белком, вызывающим эти заболевания. То есть, таргетным веществом может быть бета-амиолиод, тау-белок, альфа-синуклеин, гентингтин и прионный белок, соответственно.
Таблица 1
Второй участок, который специфически связывается с таргетным веществом, может быть выбран среди прочих из антитела, его активного фрагмента, антителоподобного белка, пептида, аптамера и растворимого рецептора при условии специфической связи с таргетным веществом.
В данном случае антитело или его активный фрагмент может быть выбран из i) иммуноглобулинов, таких как IgG1, IgG2, IgG3 и IgG4; ii) фрагментов нативного антитела, таких как Fv, Fab, Fab’, F(ab’)2, VHH и VNAR и т.д.; и iii) сконструированных антител, таких как scFv, dsFv, ds-scFv, (scFv)2, диатело, триотело, тетратело, пентатело и т.д. Антитело или его активный фрагмент может быть Mab, Fab или одноцепочечным вариабельным фрагментом (scFv) на основе антитела, образующего специфическую связь с соответствующем таргетным веществом, или на основе шести участков, определяющих комплементарность (CDRs), происходящих из антитела. То есть белок или его активный фрагмент, которые специфически связываются с таргетным веществом, включают участок, необходимый для специфической связи с таргетным веществом, и их тип и диапазон не ограничены при условии, что белок или его активный фрагмент связаны с первым участком и не вызывают воспалительного ответа и синаптического нарушения. Например, таргетное вещество может быть бета-амилоидом, и в этом случае белок или его активный фрагмент, которые специфически связываются с таргетным веществом, могут включать адуканумаб или его одноцепочечный вариабельный фрагмент. Второй участок содержит Mab, Fab или одноцепочечный вариабельный фрагмент на основе шести участков, определяющих комплементарность (CDRs), происходящих из любого одного антитела из группы, в которую входят адуканумаб, семоринемаб и цинпанемаб.
Антитело или его активный фрагмент могут не содержать Fc-участка и предпочтительно могут содержать вариант Fc-участка, который не образует связи с Fc-рецептором (в особенности Fcγ-рецептором). Этот вариант Fc-участка может служить для улучшения свойств, таких как очистка.
Антителоподобный белок относится к белковому каркасу, способному образовывать специфические связи с таргетным веществом подобно антителу. Антителоподобный белок может сконструирован таким образом, чтобы иметь размер от 2 до 20 кДа, что меньше антитела (около 150 кДа в среднем), и, соответственно, он может образовывать связи с мишенью, которой антитело не может достичь. Известно, что антителоподобные белки более стабильны при высоких температурах, чем антитела, и их легче синтезировать с использованием клеток не млекопитающих, таких как вирусы и дрожжевые грибы, или легче химически синтезировать в сравнении с антителами.
В настоящем изобретении термин «аптамер» относится к одноцепочечной ДНК (оцДНК) или РНК, имеющим высокую специфичность или сродство к определенному веществу. Аптамеры имеют очень высокую аффинность к определенным веществам, стабильны, могут быть синтезированы достаточно легким способом, могут быть модифицированы различными способами для повышения аффинности связывания с такими веществами и могут целенаправленно образовывать связи с клетками, белками и даже небольшими органическими веществами. Таким образом, аптамеры характеризуются очень высокой специфичностью и стабильностью в сравнении с антителами, которые уже разработаны. Кроме того, аптамер может быть получен с использованием известного метода SELEX (Систематическая Эволюция Лигандов Экспоненциальным обогащением). Такой аптамер, который образует специфические связи, например, с бета-амиолоидом, тау-белком, альфа-синуклеином, может быть получен с использованием известного метода SELEX, а затем он может образовать связи с первым участком, образуя таким образом слитую молекулу согласно настоящему изобретению.
Аптамер в рамках настоящего исследования не ограничивается при условии, что он способен образовывать специфические связи с бета-амиолиодом, тау-белком, альфа-синуклеином, и основания для получения аптамера могут быть выбраны из аденина (А), гуанина (G), цитозина (C), урацила (U) и их дезокси-форм, если не указано иное.
Кроме того, аптамер может быть модифицирован путем сшивки по крайней мере с одним веществом, выбранным из группы, включающей полиоксиэтиленгликоль (ПЭГ), инвертированный дезокситимидин (idT), закрытую нуклеиновую кислоту (LNA), 2’-метокси-нуклеозид, 2'-аминонуклеозид, 2'F-нуклеозид, амин-линкер, тиол-линкер и холестерол, на 5’ концевом участке, промежуточном участке, 3’ концевом участке или на обоих концевых участках для повышения стабильности. Инвертированный дезокситимидин (idT) — это молекула, которая обычно используется для предотвращения нуклеазного расщепления аптамера, имеющего слабую резистентность к нуклеазе. В случае нуклеиновой кислоты 3'-OH предыдущего нуклеотида присоединен к 5'-OH следующего нуклеотида с образованием цепочки, но в случае idT 3'-OH предыдущего нуклеотида присоединен к 3'-OH следующего нуклеотида, так, что 5’-OH, а не 3’-OH, находится на поверхности. Таким образом, idT — это молекула, которая обладает эффектом ингибирования расщепления 3'-экзонуклеазой, одним типом нуклеазы.
Растворимый рецептор по настоящему изобретению включает домен, обладающий активностью в отношении образования связи с таргетным веществом, а именно, эндогенный лиганд, причем домен может происходить из эндогенного мембранного рецептора или из внутриклеточного рецептора или из их производных. В этом случае растворимый рецептор, входящий в состав второго участка слитой молекулы по настоящему изобретению, может предпочтительно быть таким, что участок, который обладает активностью, отличной от способности образовывать связи с таргетным веществом, удален из эндогенного рецептора.
В настоящем изобретении пептид, который может быть вторым участком, является веществом, отличным от антитела или его активного фрагмента, антителоподобного белка или растворимого рецептора в ряду полипептидов, имеющих аминокислоты в качестве мономеров, способных образовывать специфические связи с таргетным веществом.
Поскольку слитая молекула, согласно настоящему изобретению, индуцирует фагоцитоз путем взаимодействия с TAM-рецептором (рецепторами), то фагоцитоз может быть индуцирован в клетках, экспрессирующих TAM-рецептор(ы). Фагоцитоз в общем означает поглощение клеток или частиц, имеющих размер 0,5 мкм или более, и включает процессы прикрепления, обволакивания и расщепления клеток или частиц. В этом случае при фагоцитозе образуется фагосома, которая окружает поглощаемую клетку или частицу и расщепляет их внутри за счет фаголизоса путем слияния фагосомы и лизосомы. При фагоцитозе процесс гибели клетки в ходе апоптоза или некроза также называется эффероцитозом.
Клетки, экспрессирующие TAM-рецептор(ы), могут быть представлены по крайней мере одним типом профессиональных фагоцитов или одним типом непрофессиональных фагоцитов или комбинацией таковых. В данном случае к профессиональным фагоцитам относятся клетки, главная роль которых заключается в удалении погибших клеток и аккумуляции остатков в ходе фагоцитоза, и примером таковых являются мегалофаги, нейтрофилы, дендритные клетки и тучные клетки. Макрофаги обычно находятся в тканях, по которым могут проходить пути инфекции, и в большинстве случаев существуют различные наименования для таких клеток, включая, к примеру, макрофаги подкожной жировой ткани, моноциты костного мозга и моноциты крови, купферовские клетки печени, синусовые гистиоциты лимфатических узлов, альвеолярные макрофаги, гистиоциты соединительной ткани или гигантоциты, микроглия центральной нервной системы, плацентарные клетки Кащенко-Хофбауэра, гломерулярные мезангифальные клетки, остеокласты, эпителиальные клетки гранулемы, макрофаги красной пульпы селезёнки, перитонеальные макрофаги брюшной полости, LysoMac пейеровой бляшки и подобные им. С другой стороны непрофессиональные фагоциты относятся к клеткам, осуществляющим специфические для тканей функции, и для них фагоцитоз не является основным видом активности, но они могут осуществлять фагоцитоз по необходимости, и примерами таких клеток являются эпителиальные клетки, эндотелиальные клетки, фибробласты, мезенхимальные клетки, некоторые тканеспецифические клетки например, астроциты или олигодендроциты центральной нервной системы, ретинальные глиальные клетки Мюллера, гепатоциты, мышечные сателлитные клетки, тестикулярные клетки Сертоли и т.д., а также некоторые лимфоциты, такие как естественные киллеры, большие гранулярные лимфоциты, эозинофилы, базофилы, В-клетки и т.д. Слитая молекула согласно настоящему изобретению способна индуцировать фагоцитоз под действием фагоцитов, специфичных для тканей, из которых удаляется накопленное таргетное вещество. Например, когда атипичные белки, накапливающиеся в головном мозге, подлежат удалению, фагоцитоз может быть индуцирован в астроцитах, микроглии, олигодендроцитах или комбинациях таковых. Например, он может быть индуцирован при местном нанесении слитой молекулы согласно настоящему изобретению на эту ткань или путем манипуляции клетками в этой ткани для экспрессии и секреции слитой молекулы.
Индукция фагоцитоза может не включать воспалительный ответ. Это позволяет осуществлять удаление таргетного вещества без индукции воспалительного ответа и повреждения тканей, вызванного воспалительным ответом, который подлежит супрессии, так, что лечение тканевой дисфункции, обусловленной накоплением таргетного вещества, может быть более безопасным, чем при традиционных подходах.
Слитая молекула может содержать метку. Когда такая метка добавляется к слитой молекуле, это может быть использовано для контроля очистки, экспрессии, активности или механизма действия слитой молекулы.
Примеры меток включают среди прочего HIS-метку, T7-метку, S-метку, FLAG-метку, Strep-метку, тиоредоксин (Trx)-метку, HIS-пэтч тиоредоксиновую метку, lacZ (L-галактозидаза)-метку, хлорамфеникол-ацетилтрансфераза-метку, trpE-метку, авидин/стрептавидин/Strep-метку, гена 10 T7-метку, стафилококковый протеин A-метку, стрептококковый протеин G-метку, глутатион-S-трансфераза (GST)-метку, дегидрофолатредуктаза ДГФР-метку, целлюлозосвязывающий домен (CBDs)-метку, мальтозосвязывающий белок (MBP)-метку, галактоза-связывающий белок-метку, кальмодулин-связывающий белок (CBP)-метку, гемагглютинин вируса гриппа А-метку, HSV-метку, B-(VP7-белковый участок вируса синего языка)-метку, полицистеин-метку, полифенилаланин-метку, (Ala-Trp-Trp-Pro)n-метку, полиаспартовая кислота-метку, c-myc-метку, lac репрессор-метку и подобные. Метка может локализована на N-терминальном конце, С-терминальном конце или внутри таргетного белка.
Кроме того, слитая молекула может содержать сигнальный пептид или лидерную последовательность на N-терминальном конце. Известно, что сигнальный пептид — это короткий пептид, присутствующий на N-терминальном конце на начальной стадии синтеза белка в направлении секреторного пути, и этот пептид обеспечивает внутриклеточную локализацию соответствующего белка, мембранную топологию (в случае мембранного белка) и т.д. Сигнальный пептид может расщепляться в ходе экспрессии и экстрацеллюлярной секреции слитой молекулы.
Вышеупомянутый первый участок, второй участок, метка, сигнальный пептид или участок, имеющий минимальную функциональность (например LG1 и LG2 участки или scFv-вариабельная область тяжёлой цепи и вариабельная область легкой цепи) включены в слитую молекулу напрямую или через линкер, содержащий короткий олигопептид или полипептид. В общем случае линкер может содержать от 2 до 50 аминокислотных остатков. Длина или тип линкера особенно не ограничены при условии, что линкер способен связывать вышеуказанные участки, чтобы они имели целевую активность, формируя таким образом слитую молекулу. Примером такого линкера может быть общеизвестный олигопептидый линкер (GGGGS)n, линкер, в котором одна или несколько единиц Gly-Gly-Gly-Gly-Ser могут повторяться. Другими примерами линкера могут быть среди прочего: (GSSGGS)n, KESGSVSSEQLAQFRSLD, EGKSSGSGSESKST, GSAGSAAGSGEF, (EAAAK)n, CRRRRRREAEAC, A(EAAAK)4ALEA(EAAAK)4A, GGGGGGGG, GGGGGG, AEAAAKEAAAAKA, PAPAP, (Ala-Pro)n, VSQTSKLTRAETVFPDV, PLGLWA, TRHRQPRGWE, AGNRVRRSVG, RRRRRRRR, GFLG, и GSSGGSGSSGGSGGGDEADGSRGSQKAGVDE.
В другом аспекте настоящее изобретение предлагает молекулу нуклеиновой кислоты, кодирующую слитую молекулу и вектор экспрессии, содержащий нуклеиновую кислоту.
Как было сказано выше, последовательность молекулы нуклеиновой кислоты, кодирующей слитую молекулу, может быть мутирована путем замещения, делеции, инсерции или их комбинации одного или нескольких нуклеотидных остатков при условии, что последовательность кодирует белок, имеющий эквивалентную активность.
Последовательность молекулы нуклеиновой кислоты, кодирующий слитую молекулу, может быть выделена из природного источника или получена искусственно путем синтеза или генетической рекомбинации. Последовательность молекулы нуклеиновой кислоты, кодирующий слитую молекулу, функционально связана с вектором экспрессии, способным экспрессировать слитую молекулу.
Термин «вектор экспрессии» («экспрессирующий вектор») характеризует вектор, способный экспрессировать интересующий белок или РНК путем введения последовательности молекулы нуклеиновой кислоты, кодирующей интересующий ген, в подходящую клетку-хозяин, и относится к генетической конструкции, содержащей необходимые регуляторные элементы, функционально связанные с подлежащей экспрессии генной вставкой. Такой вектор экспрессии включает все векторы, такие как плазмидные векторы, космидные векторы, бактериофаговые векторы и вирусные векторы.
Подходящий вектор экспрессии имеет элементы контроля экспрессии, такие как промотор, стартовый кодон, стоп-кодон, сигнал полиаденилирования и энхансер. Стартовый кодон и стоп-кодон обычно являются частью последовательности нуклеиновой кислоты, кодирующей белок, и последовательность, кодирующая белок, сконструирована так, чтобы находиться в рамке считывания и функционировать в векторе. Промотор может быть конститутивным и индуцируемым. Кроме того, традиционный вектор экспрессии содержит селектируемый маркёр. Функциональная связь с вектором экспрессии может осуществляться с использованием методик генетической рекомбинации, хорошо известных в данной области, а сайт-специфичное расщепление ДНК и лигирование может быть выполнено при использовании ферментов, общеизвестных в данной области.
Вектор экспрессии может быть предпочтительно сконструирован для экспрессии слитой молекулы в клетке-хозяине для выделения и очистки слитой молекулы или для введения вектора в клетку in vivo, а соответствующая клетка может экспрессировать и секретировать слитую молекулу. С целью введения в клетку in vivo вектор может предпочтительно быть не-интегрирующимся вектором, т.е. вектором, который не интегрируется в геном клетки-хозяина.
В другом аспекте настоящего изобретения предлагается клетка, экспрессирующая слитую молекулу.
Клетки могут быть трансформированы, чтобы содержать молекулу нуклеиновой кислоты или включающий ее вектор экспрессии, и «трансформация» может быть проведена с использованием подходящих стандартных методик, выбранных в зависимости от клетки-хозяина, известных в данной области, включая среди прочего метод введения молекулы нуклеиновой кислоты в организм, клетку, ткань или орган. Такие методы в том числе включают электропорацию, слияние протопластов, осаждение фосфатом кальция (CaPO4), осаждение хлоридом кальция (CaCl2), перемешивание с использованием карбидокремниевого волокна, трансформацию при помощи агробактерий, трансформацию на основе ПЭГ, сульфат декстрана, липофектамина и трансформацию, опосредованную высушиванием/ингибированием.
Примерами клеток-хозяев среди прочего могут быть прокариотические клетки-хозяева, такие как Escherichia coli, Bacillus subtilis, Streptomyces, Pseudomonas (например, Pseudomonas putida), Proteus mirabilis или Staphylococcus (например, Staphylocus carnosus). Другие примеры клеток-хозяев включают клетки грибов, таких как Aspergillus, клетки дрожжей, включая Pichia pastoris, Saccharomyces cerevisiae, Schizosaccharomyces и Neurospora crassa, низшие эукариотические клетки или клетки, выделенные из высших эукариот, в том числе клетки насекомых, растений или млекопитающих.
После экспрессии слитой молекулы в клетках, она может быть выделена и очищена с использованием традиционных биохимических методик выделения, таких как обработка белковым преципитирующим агентом (метод высаливания), центрифугирование, обработка ультразвуком, ультрафильтрация, диализ, или с помощью различных хроматографических методов, таких как хроматография на молекулярных ситах (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография и аффинная хроматография, которые обычно используются в комбинации для выделения белков высокой чистоты (Sambrook et al., Molecular Cloning: A laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press(1989); Deuscher, M., Guide to Protein Purification Methods Enzymology, Vol. 182. Academic Press. Inc., San Diego, CA (1990)).
В еще одном аспекте настоящее изобретение предлагает фармацевтическую композицию для предотвращения или лечения заболеваний, вызванных накоплением таргетного вещества в живых тканях, а также фармацевтическую композицию, содержащую слитую молекулу или вектор экспрессии. В данном случае композиция может быть местно нанесена на область, где таргетное вещество вызвало заболевание, а именно, где накапливается таргетное вещество.
В еще одном аспекте настоящее изобретение предлагает способ предотвращения или лечения протеинопатии, включающий этап введения пациенту фармацевтически эффективного количества слитой молекулы.
В другом аспекте настоящее изобретение предлагает применение слитой молекулы для производства медикамента для предотвращения или лечения протеинопатии.
Слитая молекула, являющаяся действующим веществом в фармацевтической композиции, содержится в «фармацевтически эффективном количестве». Термин «фармацевтически эффективное количество» означает количество, достаточное для достижения указанной выше эффективности или активности слитой молекулы.
Фармацевтическая композиция может вводиться перорально или парентерально, предпочтительно парентерально. Более предпочтительно, композиция может быть местно нанесена на ткань, в которой накапливается таргетное вещество, подлежащее удалению.
В настоящем описании термин «парентеральное введение» включает подкожное введение, внутривенное внутримышечное, интрастернальное введение или инфузионные методы.
Если фармацевтическая композиция приготовлена в виде инъецируемого лекарственного препарата, она может быть приготовлена традиционным методом, известным в данной области. Инъецируемый лекарственный препарат может быть в форме препарата, диспергированного в стерильной среде так, чтобы его можно было сразу вводить пациенту, или может быть в форме, которая может вводиться после диспергирования в дистиллированной воде для инъекций при соответствующей концентрации.
Если фармацевтическая композиция приготовлена для перорального введения, она может содержать один или несколько носителей, выбранных из растворителей, смазывающих веществ, связывающих веществ, разрыхлителей, подсластителей, стабилизаторов, консервантов и может содержать одну или несколько добавок ароматизаторов, витаминов и антиоксидантов.
Технологии, необходимые для приготовления фармацевтический композиции и фармацевтически приемлемые носители, добавки и т.д. широко известны специалистам в данной области техники (см., например, Handbook of Pharmaceutical Excipients, 4th edition, Rowe et al., Eds., American Pharmaceuticals Association (2003); Remington: the Science and Practice of Pharmacy, 20th edition, Gennaro, Ed., Lippincott Williams & Wilkins (2000); Remington's Pharmaceutical Sciences (19th ed., 1995)).
Подходящая доза фармацевтической композиции может варьировать в зависимости от таких факторов как способ изготовления, режим введения, возраст, вес, пол пациента, состояние здоровья, диета, время введения, метод введения, скорость экскреции и чувствительность ответа. Доза фармацевтической композиции по настоящему изобретению составляет от 0,0001 до 1 000 мкг/кг масса тела взрослого пациента.
Полезные эффекты
Настоящее изобретение относится к слитым молекулам, имеющим фагоцитоз-индуцирующую активность, которые могут решить проблему повреждения тканей, вызванного активацией воспалительного ответа, как известно из предшествующего уровня техники. Следовательно, слитая молекула способна устранять аномальное накопившиеся вещества, такие как бета-амилоид, тау-белок, альфа-синуклеин, гентингтин и прионный белок, и таким образом, может быть использована для предотвращения или лечения заболеваний, вызванных аномальным накоплением веществ, например, болезни Альцгеймера, болезни Паркинсона, болезни Гентингтона и прионной болезни. Слитая молекула может быть введена пациенту в форме очищенной слитой молекулы или вектора генной терапии, способного экспрессировать и секретировать слитую молекулу при введении в клетку.
Однако следует понимать, что эффекты настоящего изобретения не ограничены вышеуказанными эффектами и могут включать все эффекты, которые могут прогнозироваться исходя из сущности изобретения, детально раскрытого в описании или формуле.
Краткое описание чертежей
Фиг. 1 схематически показывает бета-амилоид и FITC-химерный специфический индуктор фагоцитоза на основе Gas6.
Фиг. 2 показывает результаты вестерн-блоттинга химерного индуктора фагоцитоза, содержащего FLAG-метку, полученного в соответствии с Примером разработки 1.
Фиг. 3 схематически показывает действие химерного индуктора фагоцитоза, полученного в соответствии с Примером разработки 1, на TAM-рецептор.
Фиг. 4 показывает расчетные результаты для способности αAβ-Gas6 селективно удалять бета-амилоид.
Фиг. 5 показывает расчетные результаты для способности αAβ-Gas6 селективно удалять бета-амилоид в клетках линии HMC3, полученные с помощью анализа поглощения бета-амилоида in vitro.
Фиг. 6 показывает расчетные результаты для способности αAβ-Gas6 селективно удалять бета-амилоид в клетках линии HMC3, полученные с помощью анализа поглощения бета-амилоида in vitro.
Фиг. 7 показывает результаты, указывающие на то, что способность αAβ-Gas6 селективно удалять бета-амилоид зависит от Axl ТАМ-рецепторов.
Фиг. 8 показывает результаты, указывающие на то, что способность αAβ-Gas6 селективно удалять бета-амилоид зависит от Axl ТАМ-рецепторов.
Фиг. 9 показывает результаты, указывающие на то, что способность αAβ-Gas6 селективно удалять бета-амилоид зависит от Axl ТАМ-рецепторов.
Фиг. 10 показывает результаты сравнительного анализа сигналлинга активации воспалительного ответа αAβ-Gas6 и адуканумаба с использованием THP-Axl-клеток.
Фиг. 11 показывает результаты сравнительного анализа уровней секреции провоспалительных цитокинов, вызванных αAβ-Gas6 и адуканумабом, с использованием THP-Axl-клеток.
Фиг. 12 показывает расчетные результаты противовоспалительной активности αAβ-Gas6.
Фиг. 13 показывает результаты, указывающие на то, что селективное подавление бета-амилоида микроглией значительно повышается при использовании αAβ-Gas6.
Фиг. 14 показывает результаты, указывающие на то, что подавление бета-амилоида астроцитами значительно повышается при использовании αAβ-Gas6.
Фиг. 15 показывает результаты, указывающие на то, что транскрипционные уровни провоспалительных цитокинов в астроцитах изменились при использовании αAβ-Gas6 и адуканумаба.
Фиг. 16 показывает результаты, указывающие на то, что транскрипционные уровни провоспалительных цитокинов в BV2 изменились при использовании αAβ-Gas6 и адуканумаба.
Фиг. 17 показывает расчетные результаты способности αAβ-Gas6 удалять бета-амилоидные бляшки путем введения αAβ-Gas6-белка на мышиной модели 5XFAD болезни Альцгеймера.
Фиг. 18 показывает расчетные результаты способности αAβ-Gas6 удалять бета-амилоидные бляшки путем введения αAβ-Gas6-вируса на мышиной модели 5XFAD болезни Альцгеймера.
Фиг. 19 показывает результаты, указывающие на то, что содержание бета-амилоида в лизосомах увеличивается за счет опосредованного микроглией удаления бета-амилоида при введении αAβ-Gas6-белка, полученные на мышиной модели 5XFAD болезни Альцгеймера.
Фиг. 20 показывает результаты, указывающие на то, что содержание бета-амилоида в лизосомах увеличивается за счет опосредованного астроцитами удаления бета-амилоида при введении αAβ-Gas6-белка, полученные на мышиной модели 5XFAD болезни Альцгеймера.
Фиг. 21 показывает результаты, указывающие на то, что содержание бета-амилоида в лизосомах увеличивается за счет опосредованного микроглией удаления бета-амилоида при введении αAβ-Gas6-вируса, полученные на мышиной модели 5XFAD болезни Альцгеймера.
Фиг. 22 показывает результаты, указывающие на то, что содержание бета-амилоида в лизосомах увеличивается за счет опосредованного астроцитами удаления бета-амилоида при введении αAβ-Gas6-вируса, полученные на мышиной модели 5XFAD болезни Альцгеймера.
Фиг. 23 показывает результаты, полученные на мышиной модели 5XFAD болезни Альцгеймера, указывающие на то, что опосредованное микроглией поглощение синапса, которое аномально повышалось из-за побочного эффекта адуканумаба, значительно восстановилось при введении αAβ-Gas6-вируса.
Фиг. 24 показывает результаты, полученные на мышиной модели 5XFAD модели болезни Альцгеймера, указывающие на то, что опосредованное микроглией поглощение синапса, которое аномально повышалось из-за побочного эффекта адуканумаба, восстановилось при введении αAβ-Gas6-вируса.
Фиг. 25 показывает экспериментальный протокол для оценки когнитивных способностей и памяти на мышиной модели 5XFAD болезни Альцгеймера при введении αAβ-Gas6-вируса.
Фиг. 26 показывает результаты, полученные на мышиной модели 5XFAD болезни Альцгеймера, указывающие на то, что когнитивные способности и память были в большей степени восстановлены при введении αAβ-Gas6-вируса, чем при введении адуканумаба.
Фиг. 27 показывает расчетные результаты для способности αTau-Gas6 удалять тау-белок в клетках линии HMC3, полученные с помощью анализа поглощения тау-белка in vitro.
Фиг. 28 показывает расчетные результаты для способности αTau-Gas6 удалять альфа-синуклеин в клетках линии HMC3, полученные с помощью анализа поглощения тау-белка in vitro.
Фиг. 29 показывает расчетные результаты для способности αAβ-ProS1 удалять бета-амилоид в первично культивируемых астроцитах, полученные с помощью анализа поглощения тау-белка in vitro.
Фиг. 30 показывает расчетные результаты для способности αAβ(Fab)-Gas6 удалять бета-амилоид в клетках линии HMC3, полученные с помощью анализа поглощения тау-белка in vitro.
Фиг. 31 показывает расчетные результаты для способности αAβ(Fab)-Gas6 удалять бета-амилоид в клетках линии HMC3, полученные с помощью анализа поглощения тау-белка in vitro.
Наилучший вариант осуществления изобретения
В дальнейшем в этом описании настоящее изобретение будет раскрыто более подробно со ссылками на примеры и экспериментальные примеры. Однако нижеизложенные примеры и экспериментальные примеры служат только для иллюстрации и не ограничивают объем изобретения.
Пример получения 1. Конструирование слитой молекулы на основе Gas6, обладающей способностью удалять бета-амилоид (I): участок связывания бета-амилоида в форме scFv
Для получения специфичного к бета-амилоиду (Aβ) химерного индуктора фагоцитоза на основе Gas6-белка, Gla-домен, который распознает PS (фосфатидилсерин) в апоптических клетках, был в первую очередь удален, а одноцепочечный вариабельный фрагмент (scFv) адуканумаба, антитела, специфичного к амилоиду, был введен в это положение [αAβ-Gas6(E)].
В дополнение, для эффективного образования белка, повторяющийся домен EGF, присутствующий во внутренних остатках Gas6-белка, был также удален, а scFv адуканумаба был введен в это положение с получением при этом αAβ-Gas6 (фиг. 1).
Кроме того, в качестве контроля за подтверждением специфического связывания scFv адуканумаба с бета-амилоидом, были получены αFITC-Gas6(E) и αFITC-Gas6 путем встраивания E2 scFv, который селективно распознает FITC, вместо scFv адуканумаба.
Таблица 2 показывает аминокислотные последовательности, задействованные в конструировании слитых молекул, а таблица 3 ниже показывает нуклеотидные последовательности, задействованные в конструировании слитых молекул (подчеркнутые последовательности представляют собой FLAG метки).
Таблица 2
MAPSLSPGPAALRRAPQLLLLLLAAECALADIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYSTPLTFGGGTKVEIKRGGGGSGGGGSGGGGSEVQLVESGGGVVQPGRSLRLSCAASGFAFSSYGMHWVRQAPGKGLEWVAVIWFDGTKKYYTDSVKGRFTISRDNSKNTLYLQMNTLRAEDTAVYYCARDRGIGARRGPYYMDVWGKGTTVTVSSGGGGSGGGGSCINKYGSPYTKNSGFATCVQNLPDQCTPNPCDRKGTQACQDLMGNFFCLCKAGWGGRLCDKDVNECSQENGGCLQICHNKPGSFHCSCHSGFELSSDGRTCQDIDECADSEACGEARCKNLPGSYSC
LCDEGFAYSSQEKACRDVDECLQGRCEQVCVNSPGSYTCHCDGRGGLKLSQDMDTCEDILPCVPFSVAKSVKSLYLGRMFSGTPVIRLRFKRLQPTRLVAEFDFRTFDPEGILLFAGGHQDSTWIVLALRAGRLELQLRYNGVGRVTSSGPVINHGMWQTISVEELARNLVIKVNRDAVMKIAVAGDLFQPERGLYHLNLTVGGIPFHEKDLVQPINPRLDGCMRSWNWLNGEDTTIQETVKVNTRMQCFSVTERGSFYPGSGFAFYSLDYMRTPLDVGTESTWEVEVVAHIRPAADTGVLFALWAPDLRAVPLSVALVDYHSTKKLKKQLVVLAVEHTALALMEIKVCDGQEHVVTVSLRDGEATLEVDGTRGQSEVSAAQLQERLAVLERHLRSPVLTFAGGLPDVPVTSAPVTAFYRGCMTLEVNRRLLDLDEAAYKHSDITAHSCPPVEPAAAQGSRADYKDHDG *
*
MAPSLSPGPAALRRAPQLLLLLLAAECALAQVQLVESGGNLVQPGGSLRLSCAASGFTFGSFSMSWVRQAPGGGLEWVAGLSARSSLTHYADSVKGRFTISRDNAKNSVYLQMNSLRVEDTAVYYCARRSYDSSGYWGHFYSYMDVWGQGTLVTVSGGGGSGGGGSGGGGSSVLTQPSSVSAAPGQKVTISCSGSTSNIGNNYVSWYQQHPGKAPKLMIYDVSKRPSGVPDRFSGSKSGNSASLDISGLQSEDEADYYCAAWDDSLSEFLFGTGTKLTVLGGGGGSGGGGSCINKYGSPYTKNSGFATCVQNLPDQCTPNPCDRKGTQACQDLMGNFFCLCKAGW
GGRLCDKDVNECSQENGGCLQICHNKPGSFHCSCHSGFELSSDGRTCQDIDECADSEACGEARCKNLPGSYSCLCDEGFAYSSQEKACRDVDECLQGRCEQVCVNSPGSYTCHCDGRGGLKLSQDMDTCEDILPCVPFSVAKSVKSLYLGRMFSGTPVIRLRFKRLQPTRLVAEFDFRTFDPEGILLFAGGHQDSTWIVLALRAGRLELQLRYNGVGRVTSSGPVINHGMWQTISVEELARNLVIKVNRDAVMKIAVAGDLFQPERGLYHLNLTVGGIPFHEKDLVQPINPRLDGCMRSWNWLNGEDTTIQETVKVNTRMQCFSVTERGSFYPGSGFAFYSLDYMRTPLDVGTESTWEVEVVAHIRPAADTGVLFALWAPDLRAVPLSVALVDYHSTKKLKKQLVVLAVEHTALALMEIKVCDGQEHVVTVSLRDGEATLEVDGTRGQSEVSAAQLQERLAVLERHLRSPVLTFAGGLPDVPVTSAPVTAFYRGCMTLEVNRRLLDLDEAAYKHSDITAHSCPPVEPAAAQGSRADYKDHDG*
MAPSLSPGPAALRRAPQLLLLLLAAECALADIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYSTPLTFGGGTKVEIKRGGGGSGGGGSGGGGSEVQLVESGGGVVQPGRSLRLSCAASGFAFSSYGMHWVRQAPGKGLEWVAVIWFDGTKKYYTDSVKGRFTISRDNSKNTLYLQMNTLRAEDTAVYYCARDRGIGARRGPYYMDVWGKGTTVTVSSGGGGSGGGGSDILPCVPFSVAKSVKSLYLGRMFSGTPVIRLRFKRLQPTRLVAEFDFRTFDPEGILLF
AGGHQDSTWIVLALRAGRLELQLRYNGVGRVTSSGPVINHGMWQTISVEELARNLVIKVNRDAVMKIAVAGDLFQPERGLYHLNLTVGGIPFHEKDLVQPINPRLDGCMRSWNWLNGEDTTIQETVKVNTRMQCFSVTERGSFYPGSGFAFYSLDYMRTPLDVGTESTWEVEVVAHIRPAADTGVLFALWAPDLRAVPLSVALVDYHSTKKLKKQLVVLAVEHTALALMEIKVCDGQEHVVTVSLRDGEATLEVDGTRGQSEVSAAQLQERLAVLERHLRSPVLTFAGGLPDVPVTSAPVTAFYRGCMTLEVNRRLLDLDEAAYKHSDITAHSCPPVEPAAAQGSRADYKDHDG*
MAPSLSPGPAALRRAPQLLLLLLAAECALAQVQLVESGGNLVQPGGSLRLSCAASGFTFGSFSMSWVRQAPGGGLEWVAGLSARSSLTHYADSVKGRFTISRDNAKNSVYLQMNSLRVEDTAVYYCARRSYDSSGYWGHFYSYMDVWGQGTLVTVSGGGGSGGGGSGGGGSSVLTQPSSVSAAPGQKVTISCSGSTSNIGNNYVSWYQQHPGKAPKLMIYDVSKRPSGVPDRFSGSKSGNSASLDISGLQSEDEADYYCAAWDDSLSEFLFGTGTKLTVLGGGGGSGGGGSCINKYGSPYTKNSGFATCVQNKDILPCVPFSVAKSVKSLYLGRMFSGTPVIRLRFKRLQPTRLVAEFDFRTFDPEGILLFAGGHQDSTWIVLALRAGRLELQLRYNGVGRVTSSGPVINHGMW
QTISVEELARNLVIKVNRDAVMKIAVAGDLFQPERGLYHLNLTVGGIPFHEKDLVQPINPRLDGCMRSWNWLNGEDTTIQETVKVNTRMQCFSVTERGSFYPGSGFAFYSLDYMRTPLDVGTESTWEVEVVAHIRPAADTGVLFALWAPDLRAVPLSVALVDYHSTKKLKKQLVVLAVEHTALALMEIKVCDGQEHVVTVSLRDGEATLEVDGTRGQSEVSAAQLQERLAVLERHLRSPVLTFAGGLPDVPVTSAPVTAFYRGCMTLEVNRRLLDLDEAAYKHSDITAHSCPPVEPAAAQGSRADYKDHDGDYKDHDI *
*
MAPSLSPGPAALRRAPQLLLLLLAAECALADIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYSTPLTFGGGTKVEIKRGGGGSGGGGSGGGGSEVQLVESGGGVVQPGRSLRLSCAASGFAFSSYGMHWVRQAPGKGLEWVAVIWFDGTKKYYTDSVKGRFTISRDNSKNTLYLQMNTLRAEDTAVYYCARDRGIGARRGPYYMDVWGKGTTVTVSSGGGGSGGGGSDILPCVPFSVAKSVKSLYLGRMFSGTPVIRLRFKRLQPTRLVAEFDFRTFDPEGILLF
AGGHQDSTWIVLALRAGRLELQLRYNGVGRVTSSGPVINHGMWQTISVEELARNLVIKVNRDAVMKIAVAGDLFQPERGLYHLNLTVGGIPFHEKDLVQPINPRLDGCMRSWNWLNGEDTTIQETVKVNTRMQCFSVTERGSFYPGSGFAFYSLDYMRTPLDVGTESTWEVEVVAHIRPAADTGVLFALWAPDLRAVPLSVALVDYHSTKKLKKQLVVLAVEHTALALMEIKVCDGQEHVVTVSLRDGEATLEVDGTRGQSEVSAAQLQERLAVLERHLRSPVLTFAGGLPDVPVTSAPVTAFYRGCMTLEVNRRLLDLDEAAYKHSDITAHSCPPVEPAAAGSGSGSGSGSGSYPYDVPDYA*
MGWSCIILFLVATATGDIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYSTPLTFGGGTKVEIKRKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGECRRKRGSGEGRGSLLTCGDVEENPGPMGWSCIILFLVATATGEVQLVESGGGVVQPGRSLRLSCAASGFAFSSYGMHWVRQAPGKGLEWVAVIWFDGTKKYYTDSVKGRFTISR
DNSKNTLYLQMNTLRAEDTAVYYCARDRGIGARRGPYYMDVWGKGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSSDKTHTSPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK*
MAPSLSPGPAALRRAPQLLLLLLAAECALAALLPAREATQFLRPRQRRAFQVFEEAKQGHLERECVEELCSREEAREVFENDPETDYFYPRYLDCINKYGSPYTKNSGFATCVQNLPDQCTPNPCDRKGTQACQDLMGNFFCLCKAGWGGRLCDKDVNECSQENGGCLQICHNKPGSFHCSCHSGFELSSDGRTCQDIDECADSEACGEARCKNLPGSYSCLCDEGFAYSSQEKACRDVDECLQGRCEQVCVNSPGSYTCHCDGRGGLKLSQDMDTCEDILPCVPFSVAKSVKSLYLGRMFSGTPVIRLRFKRLQPTRLVAEFDFRTFDPEGILLFAGGHQDSTW
IVLALRAGRLELQLRYNGVGRVTSSGPVINHGMWQTISVEELARNLVIKVNRDAVMKIAVAGDLFQPERGLYHLNLTVGGIPFHEKDLVQPINPRLDGCMRSWNWLNGEDTTIQETVKVNTRMQCFSVTERGSFYPGSGFAFYSLDYMRTPLDVGTESTWEVEVVAHIRPAADTGVLFALWAPDLRAVPLSVALVDYHSTKKLKKQLVVLAVEHTALALMEIKVCDGQEHVVTVSLRDGEATLEVDGTRGQSEVSAAQLQERLAVLERHLRSPVLTFAGGLPDVPVTSAPVTAFYRGCMTLEVNRRLLDLDEAAYKHSDITAHSCPPVEPAAAQGSRADYKDHDG *
*
Таблица 3
ATGGCCCCTTCGCTCTCGCCCGGGCCCGCCGCCCTGCGCCGCGCGCCGCAGCTGCTGCTGCTGCTGCTGGCCGCGGAGTGCGCGCTTGCCGACATTCAGATGACTCAATCTCCTAGCTCTCTGAGCGCCTCCGTTGGAGATAGAGTCACTATTACCTGCAGAGCCAGCCAATCCATCAGCTCTTATCTAAATTGGTACCAACAGAAGCCCGGCAAAGCGCCAAAGCTGCTCATCTACGCTGCAAGCTCCTTACAGAGCGGAGTACCCAGCAGATTCTCAGGCAGTGGCAGTGGGACTGACTTCACATTGACGATTAGCTCTCTGCAGCCTGAAGACTTTGCCACATACTATTGTCAGCAGAGCTATAGCACCCCGCTGACGTTTGGAGGCGGAACTAAGGTGGAAATCAAGAGAGGAGGCGGGGGCTCCGGCGGGGGTGGCTCGGGGGGAGGAGGCTCAGAGGTTCAGCTTGTCGAGTCTGGGGGGGGAG
TCGTTCAGCCAGGTAGAAGCCTCAGACTGAGCTGTGCCGCAAGTGGGTTTGCTTTTTCATCTTACGGTATGCACTGGGTGAGACAGGCTCCTGGCAAAGGACTCGAGTGGGTCGCTGTAATATGGTTCGATGGTACAAAGAAATACTATACCGATAGTGTGAAAGGAAGATTCACCATTTCACGAGACAACAGTAAAAATACCTTGTACCTTCAGATGAACACCCTGAGAGCAGAAGACACAGCCGTGTACTACTGCGCCAGAGATAGAGGTATCGGAGCAAGGCGTGGTCCCTATTATATGGATGTGTGGGGGAAGGGAACAACAGTGACTGTGAGCTCTGGCGGGGGCGGCAGCGGCGGCGGTGGCAGCTGCATCAACAAGTATGGGTCTCCGTACACCAAAAACTCAGGCTTCGCCA
CCTGCGTGCAAAACCTGCCTGACCAGTGCACGCCCAACCCCTGCGATAGGAAGGGGACCCAAGCCTGCCAGGACCTCATGGGCAACTTCTTCTGCCTGTGTAAAGCTGGCTGGGGGGGCCGGCTCTGCGACAAAGATGTCAACGAATGCAGCCAGGAGAACGGGGGCTGCCTCCAGATCTGCCACAACAAGCCGGGTAGCTTCCACTGTTCCTGCCACAGCGGCTTCGAGCTCTCCTCTGATGGCAGGACCTGCCAAGACATAGACGAGTGCGCAGACTCGGAGGCCTGCGGGGAGGCGCGCTGCAAGAACCTGCCCGGCTCCTACTCCTGCCTCTGTGACGAGGGCTTT
GCGTACAGCTCCCAGGAGAAGGCTTGCCGAGATGTGGACGAGTGTCTGCAGGGCCGCTGTGAGCAGGTCTGCGTGAACTCCCCAGGGAGCTACACCTGCCACTGTGACGGGCGTGGGGGCCTCAAGCTGTCCCAGGACATGGACACCTGTGAGGACATCTTGCCGTGCGTGCCCTTCAGCGTGGCCAAGAGTGTGAAGTCCTTGTACCTGGGCCGGATGTTCAGTGGGACCCCCGTGATCCGACTGCGCTTCAAGAGGCTGCAGCCCACCAGGCTGGTAGCTGAGTTTGACTTCCGGACCTTTGACCCCGAGGGCATCCTCCTCTTTGCCGGAGGCCACCAGGACAGCACCTGGATCGTGCTGGCCCTGAGAGCCGGCCGGCTGGAGCTGCAGCTGCGCTACAACGGTGTCGGCCGTGTC
ACCAGCAGCGGCCCGGTCATCAACCATGGCATGTGGCAGACAATCTCTGTTGAGGAGCTGGCGCGGAATCTGGTCATCAAGGTCAACAGGGATGCTGTCATGAAAATCGCGGTGGCCGGGGACTTGTTCCAACCGGAGCGAGGACTGTATCATCTGAACCTGACCGTGGGAGGTATTCCCTTCCATGAGAAGGACCTCGTGCAGCCTATAAACCCTCGTCTGGATGGCTGCATGAGGAGCTGGAACTGGCTGAACGGAGAAGACACCACCATCCAGGAAACGGTGAAAGTGAACACGAGGATGCAGTGCTTCTCGGTGACGGAGAGAGGCTCTTTCTACCCCGGGAGCGGCTTCGCCTTCTACAGCCTGGACTACATGCGGACCCCTCTGGACGTCGGGACTGAATCAACCTGGGAAGTAGAAGTCGTGGCTCACATCCGCCCAGCCGCAGACACAGGCGTGCTGTTTGCGCTCTGGGCCCCCGACCTCC
GTGCCGTGCCTCTCTCTGTGGCACTGGTAGACTATCACTCCACGAAGAAACTCAAGAAGCAGCTGGTGGTCCTGGCCGTGGAGCATACGGCCTTGGCCCTAATGGAGATCAAGGTCTGCGACGGCCAAGAGCACGTGGTCACCGTCTCGCTGAGGGACGGTGAGGCCACCCTGGAGGTGGACGGCACCAGGGGCCAGAGCGAGGTGAGCGCCGCGCAGCTGCAGGAGAGGCTGGCCGTGCTCGAGAGGCACCTGCGGAGCCCCGTGCTCACCTTTGCTGGCGGCCTGCCAGATGTGCCGGTGACTTCAGCGCCAGTCACCGCGTTCTACCGCGGCTGCATGACACTGGAGGTCAACCGGAGGCTGCTGGACCTGGACGAGGCGGCGTACAAGCACAGCGACATCACGGCCCACTCCTGCCCCCCCGTGGAGCCCGCCGCAGCCcaagGATCCCGGGCTGACTACAAAGACCATGACGGTGATTATAAAGATCATGACATCGACTACAAGGATGACGATGACAAGtga
ATGGCCCCTTCGCTCTCGCCCGGGCCCGCCGCCCTGCGCCGCGCGCCGCAGCTGCTGCTGCTGCTGCTGGCCGCGGAGTGCGCGCTTGCCCAGGTTCAGCTGGTTGAGAGCGGAGGCAATCTGGTTCAGCCCGGTGGTAGTCTGCGTCTGTCTTGTGCGGCGTCAGGGTTCACTTTCGGTAGTTTTTCAATGAGCTGGGTCCGTCAGGCACCAGGCGGTGGGCTGGAATGGGTGGCAGGTCTGTCTGCACGTAGCTCCCTGACCCACTATGCAGATAGTGTTAAAGGGCGGTTCACAATTTCACGCGACAACGCTAAGAATAGCGTCTACCTGCAAATGAACTCCCTGCGGGTCGAGGATACCGCAGTGTATTACTGCGCTCGCCGTTCTTATGACTCTAGTGGATACTGGGGCCATTTTTATAGCTACATGGATGTGTGGGGACAGGGCACTCTGGTGACCGTTTCCGGAGGCGGTGGGTCTGGAGGCGGTGGGAGTGGAGGCGGTGGGTCAAGCGTTCTGACCCAGCCGTCCTCTGTCAGCGCCGCGCCAGGCCAGAAAGTGACAATTTCCTGTTCTGGAAGTACTTCAAACATCGGCAACAATTATGTTTCCTGGTATCAGCAGCAC
CCGGGCAAAGCGCCCAAGCTGATGATTTATGATGTGTCTAAACGTCCAAGTGGTGTTCCTGACCGGTTCAGCGGTTCCAAGTCTGGGAATAGTGCCTCACTGGACATCTCAGGCCTGCAAAGCGAAGATGAGGCGGACTATTACTGCGCAGCTTGGGATGACAGCCTGTCCGAATTTCTGTTCGGCACCGGGACAAAGCTGACCGTGCTGGGCGGCGGGGGCGGCAGCGGCGGCGGTGGCAGCTGCATCAACAAGTATGGGTCTCCGTACACCAAAAACTCAGGCTTCGCCACCTGCGTGCAAAACCTGCCTGACCAGTGCACGCCCAACCCCTGCGATAGGAAGGGGACCCAAGCCTGCCAGGACCTCATGGGCAACTTCTTCTGCCTGTGTAAAGCTGGCTGGGGGGGCCGGCTCTGCGACAAAGATGTCAACGAATGCAGCCAGGAGAACGGGGGCTGCCTCCAGATCTGCCACAACAAGCCGGGTAGCTTCCACTGTTCCTGCCACAGCGGCTTCGAGCTCTCCTCTGATGGCAGGACCTGCCAAGACATAGACGAGTGCGCAGACTCGGAGGCCTGCGGGGAGGCGCGCTGCAAGAACCTGCCCGGCTCCTACTCCTGCCTCTGTGACGAGGGCTTTGCGTACAGCTCCCAGGAGAAGGCTTGCCGAGATGTGGACGAGTGTCTGCAGGGCCGCT
GTGAGCAGGTCTGCGTGAACTCCCCAGGGAGCTACACCTGCCACTGTGACGGGCGTGGGGGCCTCAAGCTGTCCCAGGACATGGACACCTGTGAGGACATCTTGCCGTGCGTGCCCTTCAGCGTGGCCAAGAGTGTGAAGTCCTTGTACCTGGGCCGGATGTTCAGTGGGACCCCCGTGATCCGACTGCGCTTCAAGAGGCTGCAGCCCACCAGGCTGGTAGCTGAGTTTGACTTCCGGACCTTTGACCCCGAGGGCATCCTCCTCTTTGCCGGAGGCCACCAGGACAGCACCTGGATCGTGCTGGCCCTGAGAGCCGGCCGGCTGGAGCTGCAGCTGCGCTACAACGGTGTCGGCCGTGTCACCAGCAGCGGCCCGGTCATCAACCATGGCATGTGGCAGACAATCTCTGTTGAGGAGCTGGCGCGGAATCTGGTCATCAAGGTCAACAGGGATGCTGTCATGAAAATCGCGGTGGCCGGGGACTTGTTCCAACCGGAGCGAGGACTGTATCATCTGAACCTGACCGTGGGAGGTATTCCCTTCCATGAGAAGGACCTC
GTGCAGCCTATAAACCCTCGTCTGGATGGCTGCATGAGGAGCTGGAACTGGCTGAACGGAGAAGACACCACCATCCAGGAAACGGTGAAAGTGAACACGAGGATGCAGTGCTTCTCGGTGACGGAGAGAGGCTCTTTCTACCCCGGGAGCGGCTTCGCCTTCTACAGCCTGGACTACATGCGGACCCCTCTGGACGTCGGGACTGAATCAACCTGGGAAGTAGAAGTCGTGGCTCACATCCGCCCAGCCGCAGACACAGGCGTGCTGTTTGCGCTCTGGGCCCCCGACCTCCGTGCCGTGCCTCTCTCTGTGGCACTGGTAGACTATCACTCCACGAAGAAACTCAAGAAGCAGCTGGTGGTCCTGGCCGTGGAGCATACGGCCTTGGCCCTAATGGAGATCAAGGTCTGCGACGGCCAAGAGCACGTGGTCACCGTCTCGCTGAGGGACGGTGAGGCCACCCTGGAGGTGGACGGCACCAGGGGCCAGAGCGAGGTGAGCGCCGCGCAGCTGCAGGAGAGGCTGGCCGTGCTCGAGAGGCACCTGCGGAGCCCCGTGCT
CACCTTTGCTGGCGGCCTGCCAGATGTGCCGGTGACTTCAGCGCCAGTCACCGCGTTCTACCGCGGCTGCATGACACTGGAGGTCAACCGGAGGCTGCTGGACCTGGACGAGGCGGCGTACAAGCACAGCGACATCACGGCCCACTCCTGCCCCCCCGTGGAGCCCGCCGCAGCCcaagGATCCCGGGCTGACTACAAAGACCATGACGGTGATTATAAAGATCATGACATCGACTACAAGGATGACGATGACAAGtga
ATGGCCCCTTCGCTCTCGCCCGGGCCCGCCGCCCTGCGCCGCGCGCCGCAGCTGCTGCTGCTGCTGCTGGCCGCGGAGTGCGCGCTTGCCGACATTCAGATGACTCAATCTCCTAGCTCTCTGAGCGCCTCCGTTGGAGATAGAGTCACTATTACCTGCAGAGCCAGCCAATCCATCAGCTCTTATCTAAATTGGTACCAACAGAAGCCCGGCAAAGCGCCAAAGCTGCTCATCTACGCTGCAAGCTCCTTACAGAGCGGAGTACCCAGCAGATTCTCAGGCAGTGGCAGTGGGACTGACTTCACATTGACGATTAGCTCTCTGCAGCCTGAAGACTTTGCCACATACTATTGTCAGCAGAGCTATAGCACCCCGCTGACGTTTGGAGGCGGAACTAAGGTGGAAATCAAGAGAGGAGGCGGGGGCTCCGGCGGGGGTGGCTCGGGGGGAGGAGGCTCAGAGGTTCAGCTTGTCGAGTCTGGGGGGGGAG
TCGTTCAGCCAGGTAGAAGCCTCAGACTGAGCTGTGCCGCAAGTGGGTTTGCTTTTTCATCTTACGGTATGCACTGGGTGAGACAGGCTCCTGGCAAAGGACTCGAGTGGGTCGCTGTAATATGGTTCGATGGTACAAAGAAATACTATACCGATAGTGTGAAAGGAAGATTCACCATTTCACGAGACAACAGTAAAAATACCTTGTACCTTCAGATGAACACCCTGAGAGCAGAAGACACAGCCGTGTACTACTGCGCCAGAGATAGAGGTATCGGAGCAAGGCGTGGTCCCTATTATATGGATGTGTGGGGGAAGGGAACAACAGTGACTGTGAGCTCTGGCGGGGGCGGCAGCGGCGGCGGTGGCAGCGACATCTTGCCGTGCGTGCCCTTCAGCGTGGCCAAGAGTGTGAAGTCCTTGTACCTGGGCCGGATGTTCAGTGGGACCCCCGTGATCCGACTGCGCTTCAAGAGGCTGCAGCCCACCAG
GCTGGTAGCTGAGTTTGACTTCCGGACCTTTGACCCCGAGGGCATCCTCCTCTTTGCCGGAGGCCACCAGGACAGCACCTGGATCGTGCTGGCCCTGAGAGCCGGCCGGCTGGAGCTGCAGCTGCGCTACAACGGTGTCGGCCGTGTCACCAGCAGCGGCCCGGTCATCAACCATGGCATGTGGCAGACAATCTCTGTTGAGGAGCTGGCGCGGAATCTGGTCATCAAGGTCAACAGGGATGCTGTCATGAAAATCGCGGTGGCCGGGGACTTGTTCCAACCGGAGCGAGGACTGTATCATCTGAACCTGACCGTGGGAGGTATTCCCTTCCATGAGAAGGACCTCGTGCAGCCTATAAACCCTCGTCTGGATGGCTGCATGAGGAGCTGGAACTGGCTGAACGGAGAAGACACCACCATCCAGGAAACGGTGAAAGTGAACACGAGGATGCAGTGCTTCTCGGTGACGGAGAGAGGCTCTTTCTACCCC
GGGAGCGGCTTCGCCTTCTACAGCCTGGACTACATGCGGACCCCTCTGGACGTCGGGACTGAATCAACCTGGGAAGTAGAAGTCGTGGCTCACATCCGCCCAGCCGCAGACACAGGCGTGCTGTTTGCGCTCTGGGCCCCCGACCTCCGTGCCGTGCCTCTCTCTGTGGCACTGGTAGACTATCACTCCACGAAGAAACTCAAGAAGCAGCTGGTGGTCCTGGCCGTGGAGCATACGGCCTTGGCCCTAATGGAGATCAAGGTCTGCGACGGCCAAGAGCACGTGGTCACCGTCTCGCTGAGGGACGGTGAGGCCACCCTGGAGGTGGACGGCACCAGGGGCCAGAGCGAGGTGAGCGCCGCGCAGCTGCAGGAGAGGCTGGCCGTGCTCGAGAGGCACCTGCGGAGCCCCGTGCTCACC
TTTGCTGGCGGCCTGCCAGATGTGCCGGTGACTTCAGCGCCAGTCACCGCGTTCTACCGCGGCTGCATGACACTGGAGGTCAACCGGAGGCTGCTGGACCTGGACGAGGCGGCGTACAAGCACAGCGACATCACGGCCCACTCCTGCCCCCCCGTGGAGCCCGCCGCAGCCcaaGGATCCCGGGCTGACTACAAAGACCATGACGGTGATTATAAAGATCATGACATCGACTACAAGGATGACGATGACAAGtga
ATGGCCCCTTCGCTCTCGCCCGGGCCCGCCGCCCTGCGCCGCGCGCCGCAGCTGCTGCTGCTGCTGCTGGCCGCGGAGTGCGCGCTTGCCCAGGTTCAGCTGGTTGAGAGCGGAGGCAATCTGGTTCAGCCCGGTGGTAGTCTGCGTCTGTCTTGTGCGGCGTCAGGGTTCACTTTCGGTAGTTTTTCAATGAGCTGGGTCCGTCAGGCACCAGGCGGTGGGCTGGAATGGGTGGCAGGTCTGTCTGCACGTAGCTCCCTGACCCACTATGCAGATAGTGTTAAAGGGCGGTTCACAATTTCACGCGACAACGCTAAGAATAGCGTCTACCTGCAAATGAACTCCCTGCGGGTCGAGGATACCGCAGTGTATTACTGCGCTCGCCGTTCTTATGACTCTAGTGGATACTGGGGCCATTTTTATAGCTACATGGATGTGTGGGGACAGGGCACTCTGGTGACCGTTTCCGGAGGCGGTGGGTCTGGAGGCGGTGGGAGTGGAGGCGGTGGGTCAAGCGTTCTGACCCAGCCGTCCTCTGTCAGCGCCGCGCCAGGCCAGAAAGTGACAATTTCCTGTTCTGGAAGTACTTCAAACATCGGCAACAATTATGTTTCCTGGTATCAGCAGCAC
CCGGGCAAAGCGCCCAAGCTGATGATTTATGATGTGTCTAAACGTCCAAGTGGTGTTCCTGACCGGTTCAGCGGTTCCAAGTCTGGGAATAGTGCCTCACTGGACATCTCAGGCCTGCAAAGCGAAGATGAGGCGGACTATTACTGCGCAGCTTGGGATGACAGCCTGTCCGAATTTCTGTTCGGCACCGGGACAAAGCTGACCGTGCTGGGCGGCGGGGGCGGCAGCGGCGGCGGTGGCAGCTGCATCAACAAGTATGGGTCTCCGTACACCAAAAACTCAGGCTTCGCCACCTGCGTGCAAAACAAAGACATCTTGCCGTGCGTGCCCTTCAGCGTGGCCAAGAGTGTGAAGTCCTTGTACCTGGGCCGGATGTTCAGTGGGACCCCCGTGATCCGACTGCGCTTCAAGAGGCTGCAGCCCACCAGGCTGGTAGCTGAGTTTGACTTCCGGACCTTTGACCCCGAGGGCATCCTCCTCTTTGCCGGAGGCCACCAGGACAGCACCTGGATCGTGCTGGCCCTGAGAGCCGGCCGGCTGGAGCTGCAGCTGCGCTACAA
CGGTGTCGGCCGTGTCACCAGCAGCGGCCCGGTCATCAACCATGGCATGTGGCAGACAATCTCTGTTGAGGAGCTGGCGCGGAATCTGGTCATCAAGGTCAACAGGGATGCTGTCATGAAAATCGCGGTGGCCGGGGACTTGTTCCAACCGGAGCGAGGACTGTATCATCTGAACCTCACCGTGGGAGGTATTCCCTTCCATGAGAAGGACCTCGTGCAGCCTATAAACCCTCGTCTGGATGGCTGTATGAGGAGCTGGAACTGGCTGAACGGAGAAGACACCACCATCCAGGAAACGGTGAAAGTGAACACGAGGATGCAGTGCTTCTCGGTGACGGAGAGAGGCTCTTTCTACCCCGGGAGCGGCTTCGCCTTCTACAGCCTGGACTACATGCGGACCCCTCTGGACGTCGGGACTGAATCAACCTGGGAAGTAGAAGTCGTGGCTCACATCCGCCCAGCCGCAGACACAGGCGTGCTGTTTGCGCTC
TGGGCCCCCGACCTCCGTGCCGTGCCTCTCTCTGTGGCACTGGTAGACTATCACTCCACGAAGAAACTCAAGAAGCAGCTGGTGGTCCTGGCCGTGGAGCATACGGCCTTGGCCCTAATGGAGATCAAGGTCTGCGACGGCCAAGAGCACGTGGTCACCGTCTCGCTGAGGGACGGTGAGGCCACCCTGGAGGTGGACGGCACCAGGGGCCAGAGCGAGGTGAGCGCCGCGCAGCTGCAGGAGAGGCTGGCCGTGCTCGAGAGGCACCTGCGGAGCCCCGTGCTCACCTTTGCCGGCGGCCTGCCAGATGTGCCGGTGACTTCAGCGCCAGTCACCGCGTTCTACCGCGGCTGCATGACACTGGAGGTCAACCGGAGGCTGCTGGACCTGGACGAGGCGGCGTACAAGCACAGCGACATCACGGCCCACTCCTGCCCCCCCGTGGAGCCCGCCGCAGCCcaaGGATCCCGGGCTGACTACAAAGACCATGACGGTGATTATAAAGATCATGACATCGACTACAAGGATGACGATGACAAGtga
ATGGCCCCTTCGCTCTCGCCCGGGCCCGCCGCCCTGCGCCGCGCGCCGCAGCTGCTGCTGCTGCTGCTGGCCGCGGAGTGCGCGCTTGCCGACATTCAGATGACTCAATCTCCTAGCTCTCTGAGCGCCTCCGTTGGAGATAGAGTCACTATTACCTGCAGAGCCAGCCAATCCATCAGCTCTTATCTAAATTGGTACCAACAGAAGCCCGGCAAAGCGCCAAAGCTGCTCATCTACGCTGCAAGCTCCTTACAGAGCGGAGTACCCAGCAGATTCTCAGGCAGTGGCAGTGGGACTGACTTCACATTGACGATTAGCTCTCTGCAGCCTGAAGACTTTGCCACATACTATTGTCAGCAGAGCTATAGCACCCCGCTGACGTTTGGAGGCGGAACTAAGGTGGAAATCAAGAGAGGAGGCGGGGGCTCCGGCGGGGGTGGCTCGGGGGGAGGAGGCTCAGAGGTTCAGCTTGTCGAGTCTGGGGGGGGAG
TCGTTCAGCCAGGTAGAAGCCTCAGACTGAGCTGTGCCGCAAGTGGGTTTGCTTTTTCATCTTACGGTATGCACTGGGTGAGACAGGCTCCTGGCAAAGGACTCGAGTGGGTCGCTGTAATATGGTTCGATGGTACAAAGAAATACTATACCGATAGTGTGAAAGGAAGATTCACCATTTCACGAGACAACAGTAAAAATACCTTGTACCTTCAGATGAACACCCTGAGAGCAGAAGACACAGCCGTGTACTACTGCGCCAGAGATAGAGGTATCGGAGCAAGGCGTGGTCCCTATTATATGGATGTGTGGGGGAAGGGAACAACAGTGACTGTGAGCTCTGGCGGGGGCGGCAGCGGCGGCGGTGGCAGCGACATCTTGCCGTGCGTGCCCTTCAGCGTGGCCAAGAGTGTGAAGTCCTTGTACCTGGGCCGGATGTTCAGTGGGACCCCCGTGATCCGACTGCGCTTCAAGAGGCTGCAGCCCACCAG
GCTGGTAGCTGAGTTTGACTTCCGGACCTTTGACCCCGAGGGCATCCTCCTCTTTGCCGGAGGCCACCAGGACAGCACCTGGATCGTGCTGGCCCTGAGAGCCGGCCGGCTGGAGCTGCAGCTGCGCTACAACGGTGTCGGCCGTGTCACCAGCAGCGGCCCGGTCATCAACCATGGCATGTGGCAGACAATCTCTGTTGAGGAGCTGGCGCGGAATCTGGTCATCAAGGTCAACAGGGATGCTGTCATGAAAATCGCGGTGGCCGGGGACTTGTTCCAACCGGAGCGAGGACTGTATCATCTGAACCTGACCGTGGGAGGTATTCCCTTCCATGAGAAGGACCTCGTGCAGCCTATAAACCCTCGTCTGGATGGCTGCATGAGGAGCTGGAACTGGCTGAACGGAGAAGACACCACCAT
CCAGGAAACGGTGAAAGTGAACACGAGGATGCAGTGCTTCTCGGTGACGGAGAGAGGCTCTTTCTACCCCGGGAGCGGCTTCGCCTTCTACAGCCTGGACTACATGCGGACCCCTCTGGACGTCGGGACTGAATCAACCTGGGAAGTAGAAGTCGTGGCTCACATCCGCCCAGCCGCAGACACAGGCGTGCTGTTTGCGCTCTGGGCCCCCGACCTCCGTGCCGTGCCTCTCTCTGTGGCACTGGTAGACTATCACTCCACGAAGAAACTCAAGAAGCAGCTGGTGGTCCTGGCCGTGGAGCATACGGCCTTGGCCCTAATGGAGATCAAGGTCTGCGACGGCCAAGAGCACGTGGTCACCGTCTCGCTGAGGGACGGTGAGGCCACCCTGGAGGTGGACGGCACCAGGGGCCAGAGCGAGGTGAGCGCCGCGCAGCTGCAGGAGAGGCTGGCCGTGCTCGAGAGGCACCTGCGGAGCCCCGTGCTCACCTTTGCTGGCGGCCTGCCAGATGTGCCGGTGACTTCAGCGCCAGTCACCGCGTTCTACCGCGGCTGCATGACACTGGAGGTCAACCGGAGGCTGCTGGACCTGGACGAGGCGGCGTACAAGCACAGCGACATCACGGCCCACTCCTGCCCCCCCGTGGAGCCCGCCGCAGCCggcagcggcagcggcagcggcagcggcagcggcagctacccatacgatgttccagattacgctTGA
GGATCCATGGGCTGGTCCTGCATCATCCTGTTCCTGGTGGCCACCGCCACCGGCGACATTCAGATGACTCAATCTCCTAGCTCTCTGAGCGCCTCCGTTGGAGATAGAGTCACTATTACCTGCAGAGCCAGCCAATCCATCAGCTCTTATCTAAATTGGTACCAACAGAAGCCCGGCAAAGCGCCAAAGCTGCTCATCTACGCTGCAAGCTCCTTACAGAGCGGAGTACCCAGCAGATTCTCAGGCAGTGGCAGTGGGACTGACTTCACATTGACGATTAGCTCTCTGCAGCCTGAAGACTTTGCCACATACTATTGTCAGCAGAGCTATAGCACCCCGCTGACGTTTGGAGGCGGAACTAAGGTGGAAATCAAGAGAAAACGAACTGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTCGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAA
GTCTACGCCTGCGAAGTCACCCATCAGGGCCTGTCCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTCGCAGAAAACGCGGAAGCGGAgagggcagaggaagtcttctaacatgcggtgacgtggaggagaatcccggccctATGGGCTGGTCCTGCATCATCCTGTTCCTGGTGGCCACCGCCACCGGCGAGGTTCAGCTTGTCGAGTCTGGGGGGGGAGTCGTTCAGCCAGGTAGAAGCCTCAGACTGAGCTGTGCCGCAAGTGGGTTTGCTTTTTCATCTTACGGTATGCACTGGGTGAGACAGGCTCCTGGCAAAGGACTCGAGTGGGTCGCTGTAATATGGTTCGATGGTACAAAGAAATACTATACCGATAGTGTGAAAGGAAGATTCACCATTTCACGAGACAACAGTAAAAATACCTTGTACCTTCAGATGAACACCCTGAGAGCAGAAGACACAGCCGTGTACTACTGCGCCAGAGATAGAGGTATCGGAGCAAGGCGTGGTCCCTATTATATGGATGTGTGGGGGAAGGGAACAACAGTGACTGTGAGCTCTGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGT
GGTGACTGTGCCCTCTAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTAGCGACAAAACTCACACAAGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCCCCGGGTAAAtga
ATGGCCCCTTCGCTCTCGCCCGGGCCCGCCGCCCTGCGCCGCGCGCCGCAGCTGCTGCTGCTGCTGCTGGCCGCGGAGTGCGCGCTTGCCGCGCTGTTGCCGGCGCGCGAGGCCACGCAGTTCCTGCGGCCCAGGCAGCGCCGCGCCTTTCAGGTCTTCGAGGAGGCCAAGCAGGGCCACCTGGAGAGGGAGTGCGTGGAGGAGCTGTGCAGCCGCGAGGAGGCGCGGGAGGTGTTCGAGAACGACCCCGAGACGGATTATTTTTACCCAAGATACTTAGACTGCATCAACAAGTATGGGTCTCCGTACACCAAAAACTCAGGCTTCGCCACCTGCGTGCAAAACCTGCCTGACCAGTGCACGCCCAACCCCTGCGATAGGAAGGGGACCCAAGCCTGCCAGGACCTCATGGGCAACTTC
TTCTGCCTGTGTAAAGCTGGCTGGGGGGGCCGGCTCTGCGACAAAGATGTCAACGAATGCAGCCAGGAGAACGGGGGCTGCCTCCAGATCTGCCACAACAAGCCGGGTAGCTTCCACTGTTCCTGCCACAGCGGCTTCGAGCTCTCCTCTGATGGCAGGACCTGCCAAGACATAGACGAGTGCGCAGACTCGGAGGCCTGCGGGGAGGCGCGCTGCAAGAACCTGCCCGGCTCCTACTCCTGCCTCTGTGACGAGGGCTTTGCGTACAGCTCCCAGGAGAAGGCTTGCCGAGATGTGGACGAGTGTCTGCAGGGCCGCTGTGAGCAGGTCTGCGTGAACTCCCCAGGGAGCTACACCTGCCACTGTGACGGGCGTGGGGGCCTCAAGCTGTCCCAGGACATGGACACCTGTGAGGACATCTTGCCGTGCGTGCCCTTCAGCGTGGCCAAGAGTGTGAAGTCCTTGTACCTGGGCCGGATGTTCAGTGGGACCCCCGTGATCCGACTGCGCTTCAAGAGGCTGCAGCCCACCAGGCTGGTAGCTGAGTTTGACTTCCGGACCTTTGACCCCGAGGGCATCCTCCTCTTTGCCGGAGGCCACCAGGACAGCACCTGGATCGTGCTGGCCCTGAGAGCCGGCCGGCTGGAGCTGCAGCTGCGCTACAACGGTGTCGGCCGTGTCACCAGCAGCGGCCCGGTCATCAACCATGGCATGTGGCAGACAATCTCTGTTGAGGAGCTGGCGCGGAATCTGGTCATCAAGGTCAACAG
GGATGCTGTCATGAAAATCGCGGTGGCCGGGGACTTGTTCCAACCGGAGCGAGGACTGTATCATCTGAACCTGACCGTGGGAGGTATTCCCTTCCATGAGAAGGACCTCGTGCAGCCTATAAACCCTCGTCTGGATGGCTGCATGAGGAGCTGGAACTGGCTGAACGGAGAAGACACCACCATCCAGGAAACGGTGAAAGTGAACACGAGGATGCAGTGCTTCTCGGTGACGGAGAGAGGCTCTTTCTACCCCGGGAGCGGCTTCGCCTTCTACAGCCTGGACTACATGCGGACCCCTCTGGACGTCGGGACTGAATCAACCTGGGAAGTAGAAGTCGTGGCTCACATCC
GCCCAGCCGCAGACACAGGCGTGCTGTTTGCGCTCTGGGCCCCCGACCTCCGTGCCGTGCCTCTCTCTGTGGCACTGGTAGACTATCACTCCACGAAGAAACTCAAGAAGCAGCTGGTGGTCCTGGCCGTGGAGCATACGGCCTTGGCCCTAATGGAGATCAAGGTCTGCGACGGCCAAGAGCACGTGGTCACCGTCTCGCTGAGGGACGGTGAGGCCACCCTGGAGGTGGACGGCACCAGGGGCCAGAGCGAGGTGAGCGCCGCGCAGCTGCAGGAGAGGCTGGCCGTGCTCGAGAGGCACCTGCGGAGCCCCGTGCTCACCTTTGCTGGCGGCCTGCCAGATGTGCCGGTGACTTCAGCGCCAGTCACCGCGTTCTACCGCGGCTGCATGACACTGGAGGTCAACCGGAGGCTGCTGGACCTGGACGAGGCGGCGTACAAGCACAGCGACATCACGGCCCACTCCTGCCCCCCCGTGGAGCCCGCCGCAGCCcaaGGATCCCGGGCTGACTACAAAGACCATGACGGTGATTATAAAGATCATGACATCGACTACAAGGATGACGATGACAAGtga
Пример получения 2. Слитая молекула на основе Gas6, направленная на тау-белок в качестве мишени
Для получения специфического к тау-белку химерного специфического индуктора фагоцитоза на основе Gas6-белка, Gla-домен и повторяющийся домен EGF были в первую очередь удалены, а одноцепочечный вариабельный фрагмент (scFv) семоринемаба, фрагмент специфичного к тау-белку антитела, был введен в это положение (αTau-Gas6). Таблица 4 показывает аминокислотную последовательность и нуклеотидную последовательность химерного индуктора фагоцитоза.
Таблица 4
MAPSLSPGPAALRRAPQLLLLLLAAECALADDVLTQTPLSLPVTPGQPASISCRSSQSIVHSNGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSLVPWTFGQGTKVEIKGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCAASGLIFRSYGMSWVRQAPGKGLEWVATINSGGTYTYYPDSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCANSYSGAMDYWGQGTLVTVSSGGGGSGGGGSDILPCVPFSVAKSVKSLYLGRMFSGTPVIRLRFKRLQPTRLVAEFDFRTFDPEGILLFAGGHQDSTWIVLALRAGRLELQLRYNGVGRVTSSGPVINHGMWQTISVEELARNLVIKVNRDAVMKIAVAGDLFQPERGLYHLNLTVGGIPFHEKDLVQPINPRLDGCMRSWNWLNGEDTTIQETVKVNTRMQCFSVTERGSFYPGSGFAFYSLDYMRTPLDVGTESTWEVEVVAHIRPAADTGVLFALWAPDLRAVPLSVALVDYHSTKKLKKQLVVLAVEHTALALMEIKVCDGQEHVVTVSLRDGEATLEVDGTRGQSEVSAAQLQERLAVLERHLRSPVLTFAGGLPDVPVTSAPVTAFYRGCMTLEVNRRLLDLDEAAYKHSDITAHSCPPVEPAAAGSGSGSGSGSGSYPYDVPDYAEGRGSLLTCGDVEENPGPVSKGEELFTGVVPILVELDGDVNGHKFSVSGEGEGDATYGKLTLKFICTTGKLPVPWPTLVTTLTYGVQCFSRYPDHMKQHDFFKSAMPEGYVQERTIFFKDDGNYKTRAEVKFEGDTLVNRIELKGIDFKEDGNILGHKLEYNYNSHNVYIMADKQKNGIKVNFKIRHNIEDGSVQLADHYQQNTPIGDGPVLLPDNHYLSTQSALSKDPNEKRDHMVLLEFVTAAGITLGMDELYK*
ATGGCCCCTTCGCTCTCGCCCGGGCCCGCCGCCCTGCGCCGCGCGCCGCAGCTGCTGCTGCTGCTGCTGGCCGCGGAGTGCGCGCTTGCCGACGATGTATTAACACAAACTCCCCTATCATTGCCGGTGACCCCGGGCCAACCAGCTTCGATCAGCTGCCGTAGCTCTCAGAGCATCGTGCACAGCAACGGTAATACCTACCTGGAATGGTATTTGCAAAAACCGGGTCAATCCCCGCAGTTGCTGATTTATAAAGTTTCGAATCGTTTCAGCGGTGTTCCGGATCGTTTCAGCGGCTCTGGCTCCGGCACCGATTTTACGCTGAAGATCAGTCGCGTGGAAGCGGAGGACGTGGGTGTCTACTACTGCTTTCAGGGTAGTTTGGTGCCGTGGACCTTTGGTCAGGGTACTAAGGTGGAAATTAAGGGTGGTGGGGGATCAGGTGGCGGCGGCAGCGGCGGTGGCGGGAGCGAGGTACAACTAGTTGAATCAGGTGGAGGGTTGGTTCAGCCAGGTGGTTCGCTGCGTCTGAGTTGTGCGGCAAGCGGTTTGATCTTTCGCAGCTATGGTATGAGCTGGGTTCGTCAGGCGCCGGGCAAGGGTCTGGAGTGGGTGGCGACCATTAACTCT
GGCGGCACGTACACCTACTATCCCGACTCCGTGAAAGGCCGTTTCACCATCTCCCGCGACAATAGCAAAAACACCCTGTATTTGCAGATGAACTCGCTCCGCGCAGAGGACACCGCTGTGTACTACTGCGCCAATTCCTACAGCGGTGCTATGGATTATTGGGGTCAGGGCACATTGGTGACTGTAAGCAGCGGCGGGGGCGGCAGCGGCGGCGGTGGCAGCGACATCTTGCCGTGCGTGCCCTTCAGCGTGGCCAAGAGTGTGAAGTCCTTGTACCTGGGCCGGATGTTCAGTGGGACCCCCGTGATCCGACTGCGCTTCAAGAGGCTGCAGCCCACCAGGCTGGTAGCTGAGTTTGACTTCCGGACCTTTGACCCCGAGGGCATCCTCCTCTTTGCCGGAGGCCACCAGGACAGCACCTGGATCGTGCTGGCCCTGAGAGCCGGCCGGCTGGAGCTGCAGCTGCGCTACAACGGTGTCGGCCGTGTCACCAGCAGCGGCCCGGTCATCAACCATGGCATGTGGCAGACAATCTCTGTTGAGGAGCTGGCGCGGAATCTGGTCATCAAGGTCAACAGGGATGCTGTCATGAAAATCGCGGTGGCCGGGGACTTGTTCCAACCGGAGCGA
GGACTGTATCATCTGAACCTGACCGTGGGAGGTATTCCCTTCCATGAGAAGGACCTCGTGCAGCCTATAAACCCTCGTCTGGATGGCTGCATGAGGAGCTGGAACTGGCTGAACGGAGAAGACACCACCATCCAGGAAACGGTGAAAGTGAACACGAGGATGCAGTGCTTCTCGGTGACGGAGAGAGGCTCTTTCTACCCCGGGAGCGGCTTCGCCTTCTACAGCCTGGACTACATGCGGACCCCTCTGGACGTCGGGACTGAATCAACCTGGGAAGTAGAAGTCGTGGCTCACATCCGCCCAGCCGCAGACACAGGCGTGCTGTTTGCGCTCTGGGCCCCCGACCTCCGTGCCGTGCCTCTCTCTGTGGCACTGGTAGACTATCACTCCACGAAGAAACTCAAGAAGCAGCTGGTGGTCCTGGCCGTGGAGCATACGGCCTTGGCCCTAATGGAGATCAAGGTCTGCGACGGCCAAGAGCACGTGGTCACCGTCTCGCTGAGGGACGGTGAGGCCACCCTGGAGGTGGACGGCACCAGGGGCCAGAGCGAGGTGAGCGC
CGCGCAGCTGCAGGAGAGGCTGGCCGTGCTCGAGAGGCACCTGCGGAGCCCCGTGCTCACCTTTGCTGGCGGCCTGCCAGATGTGCCGGTGACTTCAGCGCCAGTCACCGCGTTCTACCGCGGCTGCATGACACTGGAGGTCAACCGGAGGCTGCTGGACCTGGACGAGGCGGCGTACAAGCACAGCGACATCACGGCCCACTCCTGCCCCCCCGTGGAGCCCGCCGCAGCCGGCAGCGGCAGCGGCAGCGGCAGCGGCAGCGGCAGCtacccatacgatgttccagattacgctGAGGGCAGAGGAAGTCTGCTAACATGCGGTGACGTCGAGGAGAATCCTGGCCCAGTGAGCAAGGGCGAGGAGCTGTTCACCGGGGTGGTGCCCATCCTGGTCGAGCTGGACGGCGACGTAAACGGCCACAAGTTCAGCGTGTCCGGCGAGGGCGAGGGCGATGCCACCTACGGCAAGCTGACCCTGAAGTTCATCTGCACCACCGGCAAGCTGCCCGTGCCCTGGCCCACCCTCGTGACCACCCTGACCTACGGCGTGCAGTGCTTCAGCCGCTACCCCGACCACATGAAGCAGCACGACTTCTTCAAGTCCGCCATGCCCGAAGGCTACGTCCA
GGAGCGCACCATCTTCTTCAAGGACGACGGCAACTACAAGACCCGCGCCGAGGTGAAGTTCGAGGGCGACACCCTGGTGAACCGCATCGAGCTGAAGGGCATCGACTTCAAGGAGGACGGCAACATCCTGGGGCACAAGCTGGAGTACAACTACAACAGCCACAACGTCTATATCATGGCCGACAAGCAGAAGAACGGCATCAAGGTGAACTTCAAGATCCGCCACAACATCGAGGACGGCAGCGTGCAGCTCGCCGACCACTACCAGCAGAACACCCCCATCGGCGACGGCCCCGTGCTGCTGCCCGACAACCACTACCTGAGCACCCAGTCCGCCCTGAGCAAAGACCCCAACGAGAAGCGCGATCACATGGTCCTGCTGGAGTTCGTGACCGCCGCCGGGATCACTCTCGGCATGGACGAGCTGTACAAGtaa
Пример получения 3. Слитая молекула на основе Gas6, направленная на альфа-синуклеин в качестве мишени
Для получения специфического к альфа-синуклеину химерного индуктора фагоцитоза на основе Gas6-белка, Gla-домен и повторяющийся домен EGF, были в первую очередь удалены, а одноцепочечный вариабельный фрагмент (scFv) синпанемаба, фрагмент специфичного к альфа-синуклеину антитела, был введен в это положение (ααSyn-Gas6). Таблица 5 показывает аминокислотную последовательность и нуклеотидную последовательность химерного индуктора фагоцитоза.
Таблица 5
MAPSLSPGPAALRRAPQLLLLLLAAECALASYELTQPPSVSVSPGQTARITCSGEALPMQFAHWYQQRPGKAPVIVVYKDSERPSGVPERFSGSSSGTTATLTITGVQAEDEADYYCQSPDSTNTYEVFGGGTKLTVLGGGGSGGGGSGGGGSEVQLVESGGGLVEPGGSLRLSCAVSGFDFEKAWMSWVRQAPGQGLQWVARIKSTADGGTTSYAAPVEGRFIISRDDSRNMLYLQMNSLKTEDTAVYYCTSAHWGQGTLVTVSSGGGGSGGGGSDILPCVPFSVAKSVKSLYLGRMFSGTPVIRLRFKRLQPTRLVAEFDFRTFDPEGILLFAGGHQDSTWIVLALRAGRLELQLRYNGVGRVTSSGPVINHGMWQTISVEELARNLVIKVNRDAVMKIAVAGDLFQPERGLYHLNLTVGGIPFHEKDLVQPINPRLDGCMRSWNWLNGEDTTIQETVKVNTRMQCFSVTERGSFYPGSGFAFYSLDYMRTPLDVGTESTWEVEVVAHIRPAADTGVLFALWAPDLRAVPLSVALVDYHSTKKLKKQLVVLAVEHTALALMEIKVCDGQEHVVTVSLRDGEATLEVDGTRGQSEVSAAQLQERLAVLERHLRSPVLTFAGGLPDVPVTSAPVTAFYRGCMTLEVNRRLLDLDEAAYKHSDITAHSCPPVEPAAAGSGSGSGSGSGSYPYDVPDYAEGRGSLLTCGDVEENPGPVSKGEELFTGVVPILVELDGDVNGHKFSVSGEGEGDATYGKLTLKFICTTGKLPVPWPTLVTTLTYGVQCFSRYPDHMKQHDFFKSAMPEGYVQERTIFFKDDGNYKTRAEVKFEGDTLVNRIELKGIDFKEDGNILGHKLEYNYNSHNVYIMADKQKNGIKVNFKIRHNIEDGSVQLADHYQQNTPIGDGPVLLPDNHYLSTQSALSKDPNEKRDHMVLLEFVTAAGITLGMDELYK*
ATGGCCCCTTCGCTCTCGCCCGGGCCCGCCGCCCTGCGCCGCGCGCCGCAGCTGCTGCTGCTGCTGCTGGCCGCGGAGTGCGCGCTTGCCTCCTATGAGCTGACTCAGCCACCCTCGGTGTCAGTGTCCCCAGGACAGACGGCCAGGATCACCTGCTCTGGAGAAGCATTGCCAATGCAATTTGCTCATTGGTACCAACAGAGGCCAGGCAAGGCCCCAGTGATAGTGGTGTACAAAGACAGTGAGAGACCGTCAGGTGTCCCTGAGCGATTCTCTGGCTCCAGCTCAGGGACAACAGCCACGTTGACCATCACTGGAGTCCAGGCAGAAGATGAGGCTGACTATTACTGCCAGTCGCCAGACAGCACTAACACTTATGAAGTCTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTGGTGGGGGATCAGGTGGCGGCGGCAGCGGCGGTGGCGGGAGCGAGGTGCAGCTGGTGGAGTCTGGGGGAGGTCTGGTCGAGCCGGGGGGGTCCCTAAGACTCTCCTGTGCAGTCTCCGGATTCGATTTCGAAAAAGCCTGGAT
GAGTTGGGTCCGCCAGGCTCCAGGGCAGGGGCTACAGTGGGTTGCCCGTATCAAGAGCACAGCTGATGGTGGGACAACAAGCTACGCCGCCCCCGTGGAAGGCAGGTTCATCATCTCAAGAGATGATTCGAGAAACATGCTTTATCTGCAAATGAACAGTCTGAAAACTGAAGACACAGCCGTCTATTATTGTACATCAGCCCACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCGGGCGGGGGCGGCAGCGGCGGCGGTGGCAGCGACATCTTGCCGTGCGTGCCCTTCAGCGTGGCCAAGAGTGTGAAGTCCTTGTACCTGGGCCGGATGTTCAGTGGGACCCCCGTGATCCGACTGCGCTTCAAGAGGCTGCAGCCCACCAGGCTGGTAGCTGAGTTTGACTTCCGGACCTTTGACCCCGAGGGCATCCTCCTCTTTGCCGGAGGCCACCAGGACAGCACCTGGATCGTGCTGGCCCTGAGAGCCGGCCGGCTGGAGCTGCAGCTGCGCTACAACGGTGTCGGCCGTGTCACCAGCAGCGGCCCGGTCATCAACC
ATGGCATGTGGCAGACAATCTCTGTTGAGGAGCTGGCGCGGAATCTGGTCATCAAGGTCAACAGGGATGCTGTCATGAAAATCGCGGTGGCCGGGGACTTGTTCCAACCGGAGCGAGGACTGTATCATCTGAACCTGACCGTGGGAGGTATTCCCTTCCATGAGAAGGACCTCGTGCAGCCTATAAACCCTCGTCTGGATGGCTGCATGAGGAGCTGGAACTGGCTGAACGGAGAAGACACCACCATCCAGGAAACGGTGAAAGTGAACACGAGGATGCAGTGCTTCTCGGTGACGGAGAGAGGCTCTTTCTACCCCGGGAGCGGCTTCGCCTTCTACAGCCTGGACTACATGCGGACCCCTCTGGACGTCGGGACTGAATCAACCTGGGAAGTAGAAGTCGTGGCTCACATCCGCCCAGCCGCAGACACAGGCGTGCTGTTTGCGCTCTGGGCCCCCGACCTCCGTGCCGTGCCTCTCTCTGTGGCACTGGTAGACTATCACTCCACGAAGAAACTCAAGAAGCAGCTGGTGGTCCTGGCCGTGGAGCATACGGCCTTGGCCCTAATGGAGATCAAGGTCTGCGACGGCCAAGAGCACGTGGTCACCGTCTCGCTGAGGGACGGTGAGGCCACCCTGGAGGTGGACGGCACCAGGGGCCAGAGCGAGGTGAGCGCCGCGCAGCTGCAGGAGAGGCTGGCCGTGCTCGAGAGGCACCTGCGGAGCCCCGTGCTCACCTTTGCTGGCGGCCTGCCAGATGTGCCGGTGACTTCAGCGCCAGTCACCGCGTTCTACCGCGGCTGCATGACACTGGAGGTCAACCGGAGGCTGCTGGACCTGGACGAGGCGGCGTACAAGCACAGCGACATCACGGCCCACTCCTGCCCCCCCGTGGAGCCCGCCGCAGCCGG
CAGCGGCAGCGGCAGCGGCAGCGGCAGCGGCAGCtacccatacgatgttccagattacgctGAGGGCAGAGGAAGTCTGCTAACATGCGGTGACGTCGAGGAGAATCCTGGCCCAGTGAGCAAGGGCGAGGAGCTGTTCACCGGGGTGGTGCCCATCCTGGTCGAGCTGGACGGCGACGTAAACGGCCACAAGTTCAGCGTGTCCGGCGAGGGCGAGGGCGATGCCACCTACGGCAAGCTGACCCTGAAGTTCATCTGCACCACCGGCAAGCTGCCCGTGCCCTGGCCCACCCTCGTGACCACCCTGACCTACGGCGTGCAGTGCTTCAGCCGCTACCCCGACCACATGA
AGCAGCACGACTTCTTCAAGTCCGCCATGCCCGAAGGCTACGTCCAGGAGCGCACCATCTTCTTCAAGGACGACGGCAACTACAAGACCCGCGCCGAGGTGAAGTTCGAGGGCGACACCCTGGTGAACCGCATCGAGCTGAAGGGCATCGACTTCAAGGAGGACGGCAACATCCTGGGGCACAAGCTGGAGTACAACTACAACAGCCACAACGTCTATATCATGGCCGACAAGCAGAAGAACGGCATCAAGGTGAACTTCAAGATCCGCCACAACATCGAGGACGGCAGCGTGCAGCTCGCCGACCACTACCAGCAGAACACCCCCATCGGCGACGGCCCCGTGCTGCTGCCCGACAACCACTACCTGAGCACCCAGTCCGCCCTGAGCAAAGACCCCAACGAGAAGCGCGATCACATGGTCCTGCTGGAGTTCGTGACCGCCGCCGGGATCACTCTCGGCATGGACGAGCTGTACAAGtaa
Пример получения 4. Слитая молекула на основе ProS1, направленная на бета-амилоид в качестве мишени
Для получения специфического к бета-амилоиду (Aβ) химерного индуктора фагоцитоза на основе ProS1-белка, Gla-домен и повторяющийся домен EGF были в первую очередь удалены, а одноцепочечный вариабельный фрагмент (scFv) адуканумаба, фрагмент специфичного к бета-амилоиду антитела, был введен в это положение (αAβ-ProS1). Таблица 6 показывает аминокислотную последовательность и нуклеотидную последовательность химерного индуктора фагоцитоза.
Таблица 6
MRVLGGRCGALLACLLLVLPVSEADIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYSTPLTFGGGTKVEIKRGGGGSGGGGSGGGGSEVQLVESGGGVVQPGRSLRLSCAASGFAFSSYGMHWVRQAPGKGLEWVAVIWFDGTKKYYTDSVKGRFTISRDNSKNTLYLQMNTLRAEDTAVYYCARDRGIGARRGPYYMDVWGKGTTVTVSSGGGGSGGGGSVVSVCLPLNLDTKYELLYLAEQFAGVVLYLKFRLPEISRFSAEFDFRTYDSEGVILYAESIDHSAWLLIALRGGKIEVQLKNEHTSKITTGGDVINNGLWNMVSVEELEHSISIKIAKEAVMDINKPGPLFKPENGLLETKVYFAGFPRKVESELIKPINPRLDGCIRSWNLMKQGASGIKEIIQEKQNKHCLVTVEKGSYYPGSGIAF
HIDYNNVSSAEGWHVNVTLNIRPSTGTGVMLALVSGNNTVPFAVSLVDSTSEKSQDILLSVENTVIYRIQALSLCSDQQSHLEFRVNRNNLELSTPLKIETISHEDLQRQLAVLDKAMKAKVATYLGGLPDVPFSATPVNAFYNGCMEVNINGVQLDLDEAISKHNDIRAHSCPSVWKKTKNSQGSRADYKDHDGDYKDHDIDYKDDDDK*ASAPLPPPPLTLLAEAAWNKAGVRLSICYFPPYCRLLAM*GPGNLALSS*RAFLGVFPLSPKECKVC*MS*RKQFLWKLLEDKQRL*RPFAGSGTPHLATGASAAKSHVYKIHLQRRHNPSATL*VG*LWKESNGSPQA
YSTRG*RMPRRYPIVWDLIWGLGTHALHVFSRG*KNV*APRTTGTWFSFEKHDDNMATTMAQSKHGLTKEMTMKYRMEGCVDGHKFVITGEGIGYPFKGKQAINLCVVEGGPLPFAEDILSAAFMYGNRVFTEYPQDIVDYFKNSCPAGYTWDRSFLFEDGAVCICNADITVSVEENCMYHESKFYGVNFPADGPVMKKMTDNWEPSCEKIIPVPKQGILKGDVSMYLLLKDGGRLRCQFDTVYKAKSVPRKMPDWHFIQHKLTREDRSDAKNQKWHLTEHAIASGSALP*
ATGAGGGTCCTGGGTGGGCGCTGCGGGGCGCTGCTGGCGTGTCTCCTCCTAGTGCTTCCCGTCTCAGAGGCAGACATTCAGATGACTCAATCTCCTAGCTCTCTGAGCGCCTCCGTTGGAGATAGAGTCACTATTACCTGCAGAGCCAGCCAATCCATCAGCTCTTATCTAAATTGGTACCAACAGAAGCCCGGCAAAGCGCCAAAGCTGCTCATCTACGCTGCAAGCTCCTTACAGAGCGGAGTACCCAGCAGATTCTCAGGCAGTGGCAGTGGGACTGACTTCACATTGACGATTAGCTCTCTGCAGCCTGAAGACTTTGCCACATACTATTGTCAGCAGAGCTATAGCACCCCGCTGACGTTTGGAGGCGGAACTAAGGTGGAAATCAAGAGAGGAGGCGGGGGCTCCGGCGGGGGT
GGCTCGGGGGGAGGAGGCTCAGAGGTTCAGCTTGTCGAGTCTGGGGGGGGAGTCGTTCAGCCAGGTAGAAGCCTCAGACTGAGCTGTGCCGCAAGTGGGTTTGCTTTTTCATCTTACGGTATGCACTGGGTGAGACAGGCTCCTGGCAAAGGACTCGAGTGGGTCGCTGTAATATGGTTCGATGGTACAAAGAAATACTATACCGATAGTGTGAAAGGAAGATTCACCATTTCACGAGACAACAGTAAAAATACCTTGTACCTTCAGATGAACACCCTGAGAGCAGAAGACACAGCCGTGTACTACTGCGCCAGAGATAGAGGTATCGGAGCAAGGCGTGGTCCCTATTATATGGATGTGTGGGGGAAGGGAACAACAGTGACTGTGAGCTCTGGCGGGGGCGGCAGCGGCGGCGGTGGCAGCGTTGTTTCAGTGTGCCTTCCCTTGAACCTTGACACAAAGTATGAATTACTTTACTTGGCGGAGCAGTTTGCAGGGGTTGTTTTATATTTAAAATTTCGTTTGCCAGAAATCAGCAGATTTTCAGCAGAATTTGATTTCCGGACATATGATTCAGAAGGCGTGATACTGTACGCAGAATCTATCGATCACTCAGCGTGGCTCCTGATTGCACTTCGTGGTGGAAAGATTGAAGTTCAGCTTAAGAATGAACATACATCCAAAATCACAACTGGAGGTGATGTTATTAATAATGGTCTATGGAATATGGTGTCTGTGGAAGAATTAGAACATAGTATTAGCATTAAAATAGCTAAAGAAGCTGTGATGGATATAAATAAACCTGGACCCCTTTTTAAGCCGGAAAATGGATTGCTGGAA
ACCAAAGTATACTTTGCAGGATTCCCTCGGAAAGTGGAAAGTGAACTCATTAAACCGATTAACCCTCGTCTAGATGGATGTATACGAAGCTGGAATTTGATGAAGCAAGGAGCTTCTGGAATAAAGGAAATTATTCAAGAAAAACAAAATAAGCATTGCCTGGTTACTGTGGAGAAGGGCTCCTACTATCCTGGTTCTGGAATTGCTCAATTTCACATAGATTATAATAATGTATCCAGTGCTGAGGGTTGGCATGTAAATGTGACCTTGAATATTCGTCCATCCACGGGCACTGGTGTTATGCTTGCCTTGGTTTCTGGTAACAACACAGTGCCCTTTGCTGTGTCCTTGGTGGACTCCACCTCTGAAAAATCACAGGATATTCTGTTATCTGTTGAAAATACTGTAATATATCGGATA
CAGGCCCTAAGTCTATGTTCCGATCAACAATCTCATCTGGAATTTAGAGTCAACAGAAACAATCTGGAGTTGTCGACACCACTTAAAATAGAAACCATCTCCCATGAAGACCTTCAAAGACAACTTGCCGTCTTGGACAAAGCAATGAAAGCAAAAGTGGCCACATACCTGGGTGGCCTTCCAGATGTTCCATTCAGTGCCACACCAGTGAATGCCTTTTATAATGGCTGCATGGAAGTGAATATTAATGGTGTACAGTTGGATCTGGATGAAGCCATTTCTAAACATAATGATATTAGAGCTCACTCATGTCCATCAGTTTGGAAAAAGACAAAGAATTCTCAAGGATCCCGGGCTGACTACAAAGACCATGACGGTGATTATAAAGATCATGACATCGACTACAAGGATGACGATGACAAGtgaGCTAGCGCCCCTCTCCCTCCCCCCCCCCTAACGTTACTGGCCGAAGCCGCTTGGAATAAGGCCGGTGTGCGTTTGTCTATATGTTATTTTCCACCATATTGCCGTCTTTTGGCAATGTGAGGGCCCGGAAACCTGGCCCTGTCTTCTTGACGAGCATTCCTAGGGGTCTTTCCCCTCTCGCCAAAGGAATGCAAGGTCTGTTGA
ACGCCAAGAACCAGAAGTGGCACCTGACCGAGCACGCCATCGCCTCCGGCTCCGCCTTGCCCtga
Пример получения 5. Слитая молекула на основе Gas6, направленная на бета-амилоид в качестве мишени (II): участки, связывающие бета-амилоид, в форме Fab или Mab
Для получения специфического к бета-амилоиду (Aβ) химерного индуктора фагоцитоза на основе Gas6-белка, Gla-домен, который распознает PS (фосфатидилсерин) в апоптических клетках, был в первую очередь удален, а антиген-связывающий фрагмент (Fab) адуканумаба или моноклональное антител (Mab) адуканумаб, специфичное к бета-амилоиду, были введены в это положение (αAβ[Fab]-Gas6 и αAβ[Mab]-Gas6). Таблицы 7 и 8 показывают аминокислотную последовательность и нуклеотидную последовательность химерного индуктора фагоцитоза.
Таблица 7
METDTLLLWVLLLWVPGSTGDEVQLVESGGGVVQPGRSLRLSCAASGFAFSSYGMHWVRQAPGKGLEWVAVIWFDGTKKYYTDSVKGRFTISRDNSKNTLYLQMNTLRAEDTAVYYCARDRGIGARRGPYYMDVWGKGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHGGGGSGGGGSDILPCVPFSVAKSVKSLYLGRMFSGTPVIRLRFKRLQPTRLVAEFDFRTFDPEGILLFAGGHQDSTWIVLALRAGRLELQLRYNGVGRVTSSGPVINHGMWQTISVEELARNLVIKVNRDAVMKIAVAGDLFQPERGLYHLNLTVGGIPFHEKDLVQPINPRLDGCMRSWNWLNGEDTTIQETVKVNTRMQCFSVTERGSFYPGSGFAFYSLDYMRTPLDVGTESTWEVEVVAHIRPAADTGVLFALWAPDLRAVPLSVALVDYHSTKKLKKQLVVLAVEHTALALMEIKVCDGQEHVVTVSLRDGEATLEVDGTRGQSEVSAAQLQERLAVLERHLRSPVLTFAGGLPDVPVTSAPVTAFYRGCMTLEVNRRLLDLDEAAYKHSDITAHSCPPVEPAAADYKDHDGDYKDHDIDYKDDDDK*
ATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACGAGGTTCAGCTTGTCGAGTCTGGGGGGGGAGTCGTTCAGCCAGGTAGAAGCCTCAGACTGAGCTGTGCCGCAAGTGGGTTTGCTTTTTCATCTTACGGTATGCACTGGGTGAGACAGGCTCCTGGCAAAGGACTCGAGTGGGTCGCTGTAATATGGTTCGATGGTACAAAGAAATACTATACCGATAGTGTGAAAGGAAGATTCACCATTTCACGAGACAACAGTAAAAATACCTTGTACCTTCAGATGAACACCCTGAGAGCAGAAGACACAGCCGTGTACTACTGCGCCAGAGATAGAGGTATCGGAGCAAGGCGTGGTCCCTATTATATGGATGTGTGGGGGAAGGGAACAACAGTGACTGTGAGCTCTGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACTGTGCCCTCTAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGC
CCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACGGCGGAGGTGGAAGCGGAGGCGGTGGAAGCGACATCTTGCCGTGCGTGCCCTTCAGCGTGGCCAAGAGTGTGAAGTCCTTGTACCTGGGCCGGATGTTCAGTGGGACCCCCGTGATCCGACTGCGCTTCAAGAGGCTGCAGCCCACCAGGCTGGTAGCTGAGTTTGACTTCCGGACCTTTGACCCCGAGGGCATCCTCCTCTTTGCCGGAGGCCACCAGGACAGCACCTGGATCGTGCTGGCCCTGAGAGCCGGCCGGCTGGAGCTGCAGCTGCGCTACAACGGTGTCGGCCGT
GTCACCAGCAGCGGCCCGGTCATCAACCATGGCATGTGGCAGACAATCTCTGTTGAGGAGCTGGCGCGGAATCTGGTCATCAAGGTCAACAGGGATGCTGTCATGAAAATCGCGGTGGCCGGGGACTTGTTCCAACCGGAGCGAGGACTGTATCATCTGAACCTCACCGTGGGAGGTATTCCCTTCCATGAGAAGGACCTCGTGCAGCCTATAAACCCTCGTCTGGATGGCTGTATGAGGAGCTGGAACTGGCTGAACGGAGAAGACACCACCATCCAGGAAACGGTGAAAGTGAACACGAGGATGCAGTGCTTCTCGGTGACGGAGAGAGGCTCTTTCTACCCCGGGAG
CGGCTTCGCCTTCTACAGCCTGGACTACATGCGGACCCCTCTGGACGTCGGGACTGAATCAACCTGGGAAGTAGAAGTCGTGGCTCACATCCGCCCAGCCGCAGACACAGGCGTGCTGTTTGCGCTCTGGGCCCCCGACCTCCGTGCCGTGCCTCTCTCTGTGGCACTGGTAGACTATCACTCCACGAAGAAACTCAAGAAGCAGCTGGTGGTCCTGGCCGTGGAGCATACGGCCTTGGCCCTAATGGAGATCAAGGTCTGCGACGGCCAAGAGCACGTGGTCACCGTCTCGCTGAGGGACGGTGAGGCCACCCTGGAGGTGGACGGCACCAGGGGCCAGAGCGAGGTGAGCGCCGCGCAGCTGCAGGAGAGGCTGGCCGTGCTCGAGAGGCACCTGCGGAGCCCCGTGCTCACCTTTGC
CGGCGGCCTGCCAGATGTGCCGGTGACTTCAGCGCCAGTCACCGCGTTCTACCGCGGCTGCATGACACTGGAGGTCAACCGGAGGCTGCTGGACCTGGACGAGGCGGCGTACAAGCACAGCGACATCACGGCCCACTCCTGCCCCCCCGTGGAGCCCGCCGCAGCCGACTACAAAGACCATGACGGTGATTATAAAGATCATGACATCGACTACAAGGATGACGATGACAAGtga
Таблица 8
METDTLLLWVLLLWVPGSTGDEVQLVESGGGVVQPGRSLRLSCAASGFAFSSYGMHWVRQAPGKGLEWVAVIWFDGTKKYYTDSVKGRFTISRDNSKNTLYLQMNTLRAEDTAVYYCARDRGIGARRGPYYMDVWGKGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKGGGGSGGGGSDILPC
VPFSVAKSVKSLYLGRMFSGTPVIRLRFKRLQPTRLVAEFDFRTFDPEGILLFAGGHQDSTWIVLALRAGRLELQLRYNGVGRVTSSGPVINHGMWQTISVEELARNLVIKVNRDAVMKIAVAGDLFQPERGLYHLNLTVGGIPFHEKDLVQPINPRLDGCMRSWNWLNGEDTTIQETVKVNTRMQCFSVTERGSFYPGSGFAFYSLDYMRTPLDVGTESTWEVEVVAHIRPAADTGVLFALWAPDLRAVPLSVALVDYHSTKKLKKQLVVLAVEHTALALMEIKVCDGQEHVVTVSLRDGEATLEVDGTRGQSEVSAAQLQERLAVLERHLRSPVLTFAGGLPDVPVTSAPVTAFYRGCMTLEVNRRLLDLDEAAYKHSDITAHSCPPVEPAAA*
ATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACGAGGTTCAGCTTGTCGAGTCTGGGGGGGGAGTCGTTCAGCCAGGTAGAAGCCTCAGACTGAGCTGTGCCGCAAGTGGGTTTGCTTTTTCATCTTACGGTATGCACTGGGTGAGACAGGCTCCTGGCAAAGGACTCGAGTGGGTCGCTGTAATATGGTTCGATGGTACAAAGAAATACTATACCGATAGTGTGAAAGGAAGATTCACCATTTCACGAGACAACAGTAAAAATACCTTGTACCTTCAGATGAACACCCTGAGAGCAGAAGACACAGCCGTGTACTACTGCGCCAGAGATAGAGGTATCGGAGCAAGGCGTGGTCCCTATTATATGGATGTGTGGGGGAAGGGAACAACAGTGACTGTGAGCTCTGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCT
CTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACTGTGCCCTCTAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCAC
GTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAA
GAGCCTCTCCCTGTCCCCGGGTAAAGGCGGAGGTGGAAGCGGAGGCGGTGGAAGCGACATCTTGCCGTGCGTGCCCTTCAGCGTGGCCAAGAGTGTGAAGTCCTTGTACCTGGGCCGGATGTTCAGTGGGACCCCCGTGATCCGACTGCGCTTCAAGAGGCTGCAGCCCACCAGGCTGGTAGCTGAGTTTGACTTCCGGACCTTTGACCCCGAGGGCATCCTCCTCTTTGCCGGAGGCCACCAGGACAGCACCTGGATCGTGCTGGCCCTGAGAGCCGGCCGGCTGGAGCTGCAGCTGCGCTACAACGGTGTCGGCCGTGTCACCAGCAGCGGCCCGGTCATCAACCATGGCATGTGGCAGACAATCTCTGTTGAGGAGCTGGCGCGGAATCTGGTCATCAAGGTCAACAGGGATGCTGTCATGAAAATCGCGGTGGCCGGGGACTTGTTCCAACCGGAGCGAGGACTGTATCATCTGAACCTCACCGTGGGAGGTATTCCCTTCCATGAGAAGGACCTCGTGCAGCCTATAAACCCTCGTCTGGATGGCTGTATGAGGAGCTGGAACTGGCTGAACGGAGAAGACACCACCATCCAGGAAACGGTGAAAGTGAACACGAGGATGCAGTG
CTTCTCGGTGACGGAGAGAGGCTCTTTCTACCCCGGGAGCGGCTTCGCCTTCTACAGCCTGGACTACATGCGGACCCCTCTGGACGTCGGGACTGAATCAACCTGGGAAGTAGAAGTCGTGGCTCACATCCGCCCAGCCGCAGACACAGGCGTGCTGTTTGCGCTCTGGGCCCCCGACCTCCGTGCCGTGCCTCTCTCTGTGGCACTGGTAGACTATCACTCCACGAAGAAACTCAAGAAGCAGCTGGTGGTCCTGGCCGTGGAGCATACGGCCTTGGCC
CTAATGGAGATCAAGGTCTGCGACGGCCAAGAGCACGTGGTCACCGTCTCGCTGAGGGACGGTGAGGCCACCCTGGAGGTGGACGGCACCAGGGGCCAGAGCGAGGTGAGCGCCGCGCAGCTGCAGGAGAGGCTGGCCGTGCTCGAGAGGCACCTGCGGAGCCCCGTGCTCACCTTTGCCGGCGGCCTGCCAGATGTGCCGGTGACTTCAGCGCCAGTCACCGCGTTCTACCGCGGCTGCATGACACTGGAGGTCAACCGGAGGCTGCTGGACCTGGACGAGGCGGCGTACAAGCACAGCGACATCACGGCCCACTCCTGCCCCCCCGTGGAGCCCGCCGCAGCCtga
Экспериментальный пример 1. Слитая молекула на основе Gas6, направленная на бета-амилоид в качестве мишени (I): домен, связывающий бета-амилоид, в форме scFv
1-1. Анализ экспрессии слитой молекулы в трансфицированных клетках
После трансфекции плазмиды в HEK293-клетки экспрессия слитой молекулы, содержащей Flag-метку согласно Примеру получения 1, была проанализирована вестерн-блоттингом с использованием Flag-метки. Результаты показаны на фиг. 2.
1-2. Анализ специфической связывающей аффинности к бета-амилоиду полученных слитых молекул
Для подтверждения того, что каждая молекула из αAβ-Gas6(E), αAβ-Gas6, αFITC-Gas6(E) и αFITC-Gas6 способна селективно распознавать бета-амилоид и FITC, культуральный бульон от трансфицированных каждой плазмидой HEK293-клеток был отобран и исследован с помощью гранул, конъюгированных с бета-амилоидным олигомером и FITC. Результаты показали, что αAβ-Gas6 (E) и αAβ-Gas6 распознают только гранулы, конъюгированные с бета-амилоидным олигомером, а αFITC-Gas6 (E) и αFITC-Gas6 распознают только FITC-гранулы, таким образом, индуцируя фагоцитоз, как показано на фиг. 4.
Несмотря на то, что αAβ-Gas6 (E) и αAβ-Gas6 показывают сходную активность, было обнаружено, что молекула αAβ-Gas6, сконструированная при дополнительном удалении домена EGF в Gas6, может быть получена в большом количестве без агрегации в ходе процесса очистки белка. Таким образом, αAβ-Gas6 была использована в последующих экспериментах.
1-3. Анализ механизма действия полученной слитой молекулы
(1) Анализ с использованием клеточной линии
Был проведен анализа поглощения бета-амилоида in vitro, при котором бета-амилоидные олигомеры были коньюгированы с индикатором рН, и таким образом, могли излучать красную флуоресценцию во внутриклеточных лизосомах, когда они были поглощены путем фагоцитоза.
В результате проведения анализа поглощения бета-амилоида in vitro с HMC3-клетками, человеческой микроглиальной клеточной линией, экспрессирующей TAM-рецепторы, было показано, что αAβ-Gas6 селективно удалял бета-амилоидные олигомеры (фиг. 5 и 6).
В частности, в эксперименте, где клетки были дополнительно обработаны антителом, которое интерферирует с функцией ТAM-рецепторов, было подтверждено, что αAβ-Gas6 удаляет бета-амилоидные олигомеры в основном посредством Axl среди Tyro3, Mertk и Axl (фиг. 7–9). В действительности, когда Axl удаляли из HMC3-клеток, активность αAβ-Gas6 существенно снижалась. Кроме того, THP-1, человеческая клеточная линия моноцитов, в которой не экспрессируются TAM-рецепторы, не показала увеличение удаления бета-амилоида под действием αAβ-Gas6, в то время как THP-Axl-клетки, сверхэкспрессирующие Axl, продемонстрировали существенно увеличенную способность удаления бета-амилоидных фибрилл зависимым от αAβ-Gas6 образом.
Далее, поскольку THP-Axl-клетки экспрессируют как Axl, так и Fc рецепторы, степень воспалительного ответа, индуцированного при поглощении бета-амилоида как αAβ-Gas6, так и адуканумабом, была проанализирована в этих клетках. С этой целью NF-kB-репортеры были вначале эквпрессированы в THP-Axl-клетках, и каждое из контрольных веществ (αAβ-Gas6 и адуканумаб) было добавлено в клетки вместе с бета-амилоидными олигомерами. В результате было подтверждено то, что при добавлении адуканумаба экспрессия NF-kB-репортера существенно повышалась, однако при добавлении αAβ-Gas6 NF-kB-репортер экспрессировался на уровне ниже или равном контрольному уровню (фиг. 10). Кроме того, в результате измерения уровней секретируемого белка IL-1b, IL-6 и TNF, которые являются тремя наиболее репрезентативными цитокинами воспаления, было показано, что при обработке THP-Ax1-клеток адуканумабом, уровни белка воспалительных цитокинов в обработанных клетках были существенно увеличены по сравнению с таковыми в контрольной группе (фиг. 11). В отличие от этого, важно, что уровни этих воспалительных цитокинов в клетках, обработанных αAβ-Gas6, не увеличивались по сравнению с таковыми в контрольной группе. Ключевым результатом является то, что гипотеза авторов предполагает, что слитый белок индуктора фагоцитоза αAβ-Gas6 не индуцирует воспалительный ответ при фагоцитировании таргетного вещества посредством TAM-рецептора, что аналогично распознаванию и эффероцитозу естественных апоптических клеток.
Кроме того, в отличие от адуканумаба, αAβ-Gas6 повышает экспрессию Twist1/2-гена, который, как известно, вовлечен в механизм подавления воспалительного ответа (фиг. 12).
(2) Анализ с использованием астроцитов и микроглии
Для исследования того, способны ли астроциты и клетки микроглии, которые являются клетками, экспрессирующими TAM-рецепторы в головном мозге, удалять бета-амилоид посредством αAβ-Gas6, первичные астроциты и микроглия были получены из головного мозга мышей, по отдельности очищены и затем культивированы. После того, как каждое из очищенных соединений αAβ-Gas6 и адуканумаба были добавлены в клетки вместе с бета-амилоидными фибриллами, была измерена в режиме реального времени степень удаления бета-амилоидных фибрилл.
Результаты показали, что молекула αAβ-Gas6 повышает способность микроглии удалять бета-амилоид в зависимости от концентрации, что было аналогично результатам, полученным в клетках клеточной линии HMC3, которые экспрессируют Axl (фиг. 13). Важно, что было показано, что при добавлении адуканумаба способность астроцитов удалять бета-амилоид не изменялась, но при добавлении αAβ-Gas6 способность астроцитов удалять бета-амилоид существенно возрастала в зависимости от концентрации (фиг. 14). Это указывает на то, что αAβ-Gas6 значительно повышает способность астроцитов удалять бета-амилоид, которая прежде была незначительной, поскольку астроциты не экспрессируют Fc-рецепторы, но экспрессируют TAM-рецепторы.
Каждое из соединений αAβ-Gas6 и адуканумаба добавляли к астроцитам и клеточной линии микроглии BV2 вместе с бета-амилоидными фибриллами для повышения удаления бета-амилоида, а затем измеряли уровни мРНК TNF, IL-1a и IL-1b в каждой клеточной линии для определения степени воспалительного ответа (фиг. 15 и 16). В результате было показано, что при обработке клеток адуканумабом уровни транскриптов и белков вышеуказанных воспалительных цитокинов в астроцитах и BV2-клетках существенно увеличивались в сравнении с таковыми в контрольной группе, но при обработке клеток молекулой αAβ-Gas6 уровни этих воспалительных цитокинов в клетках не увеличивались в сравнении с контрольной группой.
Как указывалось выше, было обнаружено, что использование слитого белка индуктора фагоцитоза αAβ-Gas6 может быть принципиально новым подходом к эффективному удалению бета-амилоидных бляшек, накапливающихся в головном мозге пациента, посредством астроцитов и микроглии без генерации воспалительного ответа, являющегося серьезным побочным эффектом терапии моноклональным антителом. Это может стать очень обнадеживающим результатом, который способен существенно усовершенствовать текущие лечебные стратегии.
1-4. Оценка эффективности in vivo
(1) Эффективность в соответствии с введением слитой молекулы или вектора экспрессии на мышиной модели 5XFAD болезни Альцгеймера.
Поскольку мыши 5XFAD одновременно экспрессируют 5 генов с мутациями, образование бета-амилоидных бляшек у этих мышей происходит раньше, а патологические симптомы, вызванные бета-амилоидными бляшками, могут исследоваться от 3 до 4-месячного возраста особи вне зависимости от старения.
Для подтверждения эффекта αAβ-Gas6 in vivo на модели 5XFAD, αAβ-Gas6 вводили в головной мозг двумя различными методами. Из предшествующих исследований известно, что адуканумаб не вводится достаточно эффективно в головной мозг с помощью внутрисосудистой или внутрибрюшинной инъекции даже особям мышиной модели болезни Альцгеймера. Таким образом, для точного сравнения и анализа эффектов αAβ-Gas6 и адуканумаба 1) было выполнено прямое канюлирование в головной мозг мышей, и каждое из очищенных соединений αAβ-Gas6 и адуканумаба вводили один раз в день в желудочек головного мозга в течение 3 недель и 2) каждое из соединений αAβ-Gas6 и адуканумаба было встроено в лентивирус (лентивирусная форма, чтобы быть экспрессированным в гиппокампе мыши, и лентивирус вводили путем стереотаксической инъекции. Важно отметить, что было обнаружено, что количество бета-амилоидных бляшек существенно понижалось как при добавлении очищенного белка αAβ-Gas6, так и при экспрессии соответствующего гена в лентивирусной форме (фиг. 17 и 18).
Кроме того, при количественном определении уровней бета-амилоида в лизосомах микроглии и астроцитов после введения αAβ-Gas6 в головной мозг в форме белка или вируса было показано, что способность удалять бета-амилоид существенно повышалась для обоих видов клеток (фиг. 19–22).
Это позволяет предположить, что поскольку TAM-рецепторы экспрессированы как в микроглии, так и в астроцитах, то микроглия и астроциты могут распознавать и удалять бета-амилоид при введении αAβ-Gas6, что аналогично результатам исследований in vitro.
(2) Сравнение эффектов терапии антителом и слитой молекулой по настоящему изобретению
Известно, что при болезни Альцгеймера синапсы беспорядочно удаляются микроглией, что приводит к уменьшению количества синапсов. Неожиданно, это явление усугублялось при введении адуканумаба в опытах на мышиной модели болезнью Альцгеймера, но при экспрессии αAβ-Gas6 в вирусной форме, аномальное удаление синапсов микроглией было восстановлено до нормального уровня (фиг. 23 и 24).
Кроме того, результаты оценки когнитивных способностей и памяти для запоминания формы или расположения нового объекта, полученные на мышиной модели болезни Альцгеймера согласно протоколу, приведенному на фиг. 25, подтверждают, что экспрессия αAβ-Gas6 приводит к исключительно высоким восстановительным эффекты когнитивной функции и памяти в сравнении с адуканумабом (фиг. 26).
Кроме того, для подтверждения того, способен ли индуцирующий фагоцитоз химерный белок по настоящему изобретению, эффективно удалять различные таргетные вещества в дополнение к бета-амилоиду, были получены белки-индукторы фагоцитоза, специфичные к тау-белку и альфа-синуклеину (αSyn), как раскрыто в Примерах получения 2 и 3, а эффект удаления ими таргетных веществ был протестирован в ходе следующих протоколов в Экспериментальных примерах 2 и 3.
Экспериментальный пример 2. Слитая молекула на основе Gas6, направленная на тау-белок в качестве мишени
Был проведен анализа поглощения тау-белка in vitro, при котором тау олигомеры были коньюгированы с индикатором рН, и таким образом, могли излучать красную флуоресценцию во внутриклеточных лизосомах, когда они были поглощены посредством фагоцитоза. HMC3-клетки, представляющие собой человеческую микроглиальную линию клеток, экспрессирующую TAM-рецепторы, обрабатывали культурной средой, содержащей белок-индуктор фагоцитоза [αTau-Gas6] согласно Примеру получения 2, и проводили анализ поглощения тау-белка in vitro. Как показывают результаты на фиг. 27, было подтверждено, что тау олигомеры селективно удалялись при использовании αTau-Gas6.
Экспериментальный пример 3. Слитая молекула на основе Gas6, направленная на альфа-синуклеин в качестве мишени
Был проведен анализа поглощения альфа-синуклеина in vitro, при котором олигомеры альфа-синуклеина были коньюгированы с индикатором рН, и таким образом, могли излучать красную флуоресценцию во внутриклеточных лизосомах, когда они были поглощены посредством фагоцитоза. HMC3-клетки, представляющие собой человеческую микроглиальную клеточную линию, экспрессирующую TAM-рецепторы, обрабатывали культурной средой, содержащей белок-индуктор фагоцитоза [ααSyn-Gas6], согласно Примеру получения 3, и проводили анализ поглощения альфа-синуклеина in vitro. Как показывают результаты на фиг. 27, было подтверждено, что олигомеры альфа-синуклеина селективно удалялись при использовании ααSyn-Gas6.
Экспериментальный пример 4. Слитая молекула на основе ProS1, направленная на бета-амилоид в качестве мишени
Далее, для подтверждения того, является ли химерный белок, индуцирующий фагоцитоз, полученный при использовании лиганда ТАМ-рецептора, отличного от Gas6, таким же эффективным, был сконструирован белок αAβ-ProS1, как раскрыто в Примере получения 4, с использованием ProS1-лиганда, и была проведена оценка его эффективности. С этой целью первично культивируемые астроциты мыши, экспрессирующие TAM-рецепторы, были обработаны культуральной средой, содержащей αAβ-ProS1, и проводили анализ поглощения Aβ in vitro согласно Экспериментальным примерам 1-3. Как показывают результаты на фиг. 29, было подтверждено, что олигомеры бета-амилоида селективно удалялись при использовании αAβ-ProS1.
Экспериментальный пример 5. Слитая молекула основе Gas6, направленная на бета-амилоид в качестве мишени (II): участки связывания бета-амилоида в форме Fab или Mab
Далее, для подтверждения того, могут ли применяться различные связывающие участки таргетных веществ, отличных от scFv, в качестве доменов, связывающих таргетные белки, были сконструированы белки-индукторы фагоцитоза согласно Примеру разработки 5 с использованием антиген-связывающего фрагмента (Fab) или полноразмерной формы моноклонального антитела (Mab) вместо scFv, которые исследовались экспериментально (αAβ[Fab]-Gas6 и αAβ[Mab]-Gas6). С этой целью HMC3-клетки, представляющие собой человеческую микроглиальную клеточную линию, экспрессирующую TAM-рецепторы, были обработаны культурной средой, содержащей каждое из соединений αAβ[Fab]-Gas6 и αAβ[Mab]-Gas6, и проводили анализ поглощения Aβ in vitro согласно Экспериментальным примерам 1-3. Как показывают результаты на фиг. 30 и 31, было подтверждено, что олигомеры бета-амилоида селективно удалялись при использовании каждого из αAβ[Fab]-Gas6 и αAβ[Mab]-Gas6.
Объем данного изобретения определяется прилагаемой формулой изобретения, и все изменения или модификации, выведенные из смыслового содержания и объема формулы изобретения, и эквивалентные таковым, также считаются входящими в объем настоящего изобретения.
Принцип изобретения
Принцип изобретения также был описан в наилучшем варианте осуществления изобретения выше.
Промышленная применимость
Слитые молекулы, обладающие активностью индуцировать фагоцитоз, в соответствии с примерами осуществления настоящего изобретения, могут решить проблему повреждения тканей, вызванных активацией воспалительного ответа, как это известно из предшествующего уровня техники. Следовательно, слитые молекулы способны эффективно удалять аномально накопившиеся вещества, такие как бета-амилоид, тау-белок, альфа-синуклеин, гентингтин и прионный белок, и таким образом, могут быть использованы для предотвращения или лечения заболеваний, вызванных аномальным накоплением веществ, например, болезни Альцгеймера, болезни Паркинсона, болезни Гентингтона и прионной болезни. Поэтому слитые молекулы могут использоваться в терапевтической отрасли для лечения вышеуказанных болезней.
--->
<110> KOREA ADVANCED INSTITUTE OF SCIENCE AND TECHNOLOGY
<120> Fusion molecules that induce non-inflammatory phagocytosis
<130> P20220004KR
<150> KR 10/2021/0013045
<151> 2021-01-29
<160> 6
<170> KoPatentIn 3.0
<210> 1
<211> 173
<212> PRT
<213> Artificial Sequence
<220>
<223> amino acid sequence
<400> 1
Gly Arg Met Phe Ser Gly Thr Pro Val Ile Arg Leu Arg Phe Lys Arg
1 5 10 15
Leu Gln Pro Thr Arg Leu Val Ala Glu Phe Asp Phe Arg Thr Phe Asp
20 25 30
Pro Glu Gly Ile Leu Leu Phe Ala Gly Gly His Gln Asp Ser Thr Trp
35 40 45
Ile Val Leu Ala Leu Arg Ala Gly Arg Leu Glu Leu Gln Leu Arg Tyr
50 55 60
Asn Gly Val Gly Arg Val Thr Ser Ser Gly Pro Val Ile Asn His Gly
65 70 75 80
Met Trp Gln Thr Ile Ser Val Glu Glu Leu Ala Arg Asn Leu Val Ile
85 90 95
Lys Val Asn Arg Asp Ala Val Met Lys Ile Ala Val Ala Gly Asp Leu
100 105 110
Phe Gln Pro Glu Arg Gly Leu Tyr His Leu Asn Leu Thr Val Gly Gly
115 120 125
Ile Pro Phe His Glu Lys Asp Leu Val Gln Pro Ile Asn Pro Arg Leu
130 135 140
Asp Gly Cys Met Arg Ser Trp Asn Trp Leu Asn Gly Glu Asp Thr Thr
145 150 155 160
Ile Gln Glu Thr Val Lys Val Asn Thr Arg Met Gln Cys
165 170
<210> 2
<211> 194
<212> PRT
<213> Artificial Sequence
<220>
<223> Gas6 derived LG2
<400> 2
Gly Ser Phe Tyr Pro Gly Ser Gly Phe Ala Phe Tyr Ser Leu Asp Tyr
1 5 10 15
Met Arg Thr Pro Leu Asp Val Gly Thr Glu Ser Thr Trp Glu Val Glu
20 25 30
Val Val Ala His Ile Arg Pro Ala Ala Asp Thr Gly Val Leu Phe Ala
35 40 45
Leu Trp Ala Pro Asp Leu Arg Ala Val Pro Leu Ser Val Ala Leu Val
50 55 60
Asp Tyr His Ser Thr Lys Lys Leu Lys Lys Gln Leu Val Val Leu Ala
65 70 75 80
Val Glu His Thr Ala Leu Ala Leu Met Glu Ile Lys Val Cys Asp Gly
85 90 95
Gln Glu His Val Val Thr Val Ser Leu Arg Asp Gly Glu Ala Thr Leu
100 105 110
Glu Val Asp Gly Thr Arg Gly Gln Ser Glu Val Ser Ala Ala Gln Leu
115 120 125
Gln Glu Arg Leu Ala Val Leu Glu Arg His Leu Arg Ser Pro Val Leu
130 135 140
Thr Phe Ala Gly Gly Leu Pro Asp Val Pro Val Thr Ser Ala Pro Val
145 150 155 160
Thr Ala Phe Tyr Arg Gly Cys Met Thr Leu Glu Val Asn Arg Arg Leu
165 170 175
Leu Asp Leu Asp Glu Ala Ala Tyr Lys His Ser Asp Ile Thr Ala His
180 185 190
Ser Cys
<210> 3
<211> 177
<212> PRT
<213> Artificial Sequence
<220>
<223> amino acid sequence
<400> 3
Leu Leu Tyr Leu Ala Glu Gln Phe Ala Gly Val Val Leu Tyr Leu Lys
1 5 10 15
Phe Arg Leu Pro Glu Ile Ser Arg Phe Ser Ala Glu Phe Asp Phe Arg
20 25 30
Thr Tyr Asp Ser Glu Gly Val Ile Leu Tyr Ala Glu Ser Ile Asp His
35 40 45
Ser Ala Trp Leu Leu Ile Ala Leu Arg Gly Gly Lys Ile Glu Val Gln
50 55 60
Leu Lys Asn Glu His Thr Ser Lys Ile Thr Thr Gly Gly Asp Val Ile
65 70 75 80
Asn Asn Gly Leu Trp Asn Met Val Ser Val Glu Glu Leu Glu His Ser
85 90 95
Ile Ser Ile Lys Ile Ala Lys Glu Ala Val Met Asp Ile Asn Lys Pro
100 105 110
Gly Pro Leu Phe Lys Pro Glu Asn Gly Leu Leu Glu Thr Lys Val Tyr
115 120 125
Phe Ala Gly Phe Pro Arg Lys Val Glu Ser Glu Leu Ile Lys Pro Ile
130 135 140
Asn Pro Arg Leu Asp Gly Cys Ile Arg Ser Trp Asn Leu Met Lys Gln
145 150 155 160
Gly Ala Ser Gly Ile Lys Glu Ile Ile Gln Glu Lys Gln Asn Lys His
165 170 175
Cys
<210> 4
<211> 183
<212> PRT
<213> Artificial Sequence
<220>
<223> amino acid sequence
<400> 4
Tyr Tyr Pro Gly Ser Gly Ile Ala Gln Phe His Ile Asp Tyr Asn Asn
1 5 10 15
Val Ser Ser Ala Glu Gly Trp His Val Asn Val Thr Leu Asn Ile Arg
20 25 30
Pro Ser Thr Gly Thr Gly Val Met Leu Ala Leu Val Ser Gly Asn Asn
35 40 45
Thr Val Pro Phe Ala Val Ser Leu Val Asp Ser Thr Ser Glu Lys Ser
50 55 60
Gln Asp Ile Leu Leu Ser Val Glu Asn Thr Val Ile Tyr Arg Ile Gln
65 70 75 80
Ala Leu Ser Leu Cys Ser Asp Gln Gln Ser His Leu Glu Phe Arg Val
85 90 95
Asn Arg Asn Asn Leu Glu Leu Ser Thr Pro Leu Lys Ile Glu Thr Ile
100 105 110
Ser His Glu Asp Leu Gln Arg Gln Leu Ala Val Leu Asp Lys Ala Met
115 120 125
Lys Ala Lys Val Ala Thr Tyr Leu Gly Gly Leu Pro Asp Val Pro Phe
130 135 140
Ser Ala Thr Pro Val Asn Ala Phe Tyr Asn Gly Cys Met Glu Val Asn
145 150 155 160
Ile Asn Gly Val Gln Leu Asp Leu Asp Glu Ala Ile Ser Lys His Asn
165 170 175
Asp Ile Arg Ala His Ser Cys
180
<210> 5
<211> 400
<212> PRT
<213> Artificial Sequence
<220>
<223> amino acid sequence
<400> 5
Asp Ile Leu Pro Cys Val Pro Phe Ser Val Ala Lys Ser Val Lys Ser
1 5 10 15
Leu Tyr Leu Gly Arg Met Phe Ser Gly Thr Pro Val Ile Arg Leu Arg
20 25 30
Phe Lys Arg Leu Gln Pro Thr Arg Leu Val Ala Glu Phe Asp Phe Arg
35 40 45
Thr Phe Asp Pro Glu Gly Ile Leu Leu Phe Ala Gly Gly His Gln Asp
50 55 60
Ser Thr Trp Ile Val Leu Ala Leu Arg Ala Gly Arg Leu Glu Leu Gln
65 70 75 80
Leu Arg Tyr Asn Gly Val Gly Arg Val Thr Ser Ser Gly Pro Val Ile
85 90 95
Asn His Gly Met Trp Gln Thr Ile Ser Val Glu Glu Leu Ala Arg Asn
100 105 110
Leu Val Ile Lys Val Asn Arg Asp Ala Val Met Lys Ile Ala Val Ala
115 120 125
Gly Asp Leu Phe Gln Pro Glu Arg Gly Leu Tyr His Leu Asn Leu Thr
130 135 140
Val Gly Gly Ile Pro Phe His Glu Lys Asp Leu Val Gln Pro Ile Asn
145 150 155 160
Pro Arg Leu Asp Gly Cys Met Arg Ser Trp Asn Trp Leu Asn Gly Glu
165 170 175
Asp Thr Thr Ile Gln Glu Thr Val Lys Val Asn Thr Arg Met Gln Cys
180 185 190
Phe Ser Val Thr Glu Arg Gly Ser Phe Tyr Pro Gly Ser Gly Phe Ala
195 200 205
Phe Tyr Ser Leu Asp Tyr Met Arg Thr Pro Leu Asp Val Gly Thr Glu
210 215 220
Ser Thr Trp Glu Val Glu Val Val Ala His Ile Arg Pro Ala Ala Asp
225 230 235 240
Thr Gly Val Leu Phe Ala Leu Trp Ala Pro Asp Leu Arg Ala Val Pro
245 250 255
Leu Ser Val Ala Leu Val Asp Tyr His Ser Thr Lys Lys Leu Lys Lys
260 265 270
Gln Leu Val Val Leu Ala Val Glu His Thr Ala Leu Ala Leu Met Glu
275 280 285
Ile Lys Val Cys Asp Gly Gln Glu His Val Val Thr Val Ser Leu Arg
290 295 300
Asp Gly Glu Ala Thr Leu Glu Val Asp Gly Thr Arg Gly Gln Ser Glu
305 310 315 320
Val Ser Ala Ala Gln Leu Gln Glu Arg Leu Ala Val Leu Glu Arg His
325 330 335
Leu Arg Ser Pro Val Leu Thr Phe Ala Gly Gly Leu Pro Asp Val Pro
340 345 350
Val Thr Ser Ala Pro Val Thr Ala Phe Tyr Arg Gly Cys Met Thr Leu
355 360 365
Glu Val Asn Arg Arg Leu Leu Asp Leu Asp Glu Ala Ala Tyr Lys His
370 375 380
Ser Asp Ile Thr Ala His Ser Cys Pro Pro Val Glu Pro Ala Ala Ala
385 390 395 400
<210> 6
<211> 393
<212> PRT
<213> Artificial Sequence
<220>
<223> amino acid sequence
<400> 6
Val Val Ser Val Cys Leu Pro Leu Asn Leu Asp Thr Lys Tyr Glu Leu
1 5 10 15
Leu Tyr Leu Ala Glu Gln Phe Ala Gly Val Val Leu Tyr Leu Lys Phe
20 25 30
Arg Leu Pro Glu Ile Ser Arg Phe Ser Ala Glu Phe Asp Phe Arg Thr
35 40 45
Tyr Asp Ser Glu Gly Val Ile Leu Tyr Ala Glu Ser Ile Asp His Ser
50 55 60
Ala Trp Leu Leu Ile Ala Leu Arg Gly Gly Lys Ile Glu Val Gln Leu
65 70 75 80
Lys Asn Glu His Thr Ser Lys Ile Thr Thr Gly Gly Asp Val Ile Asn
85 90 95
Asn Gly Leu Trp Asn Met Val Ser Val Glu Glu Leu Glu His Ser Ile
100 105 110
Ser Ile Lys Ile Ala Lys Glu Ala Val Met Asp Ile Asn Lys Pro Gly
115 120 125
Pro Leu Phe Lys Pro Glu Asn Gly Leu Leu Glu Thr Lys Val Tyr Phe
130 135 140
Ala Gly Phe Pro Arg Lys Val Glu Ser Glu Leu Ile Lys Pro Ile Asn
145 150 155 160
Pro Arg Leu Asp Gly Cys Ile Arg Ser Trp Asn Leu Met Lys Gln Gly
165 170 175
Ala Ser Gly Ile Lys Glu Ile Ile Gln Glu Lys Gln Asn Lys His Cys
180 185 190
Leu Val Thr Val Glu Lys Gly Ser Tyr Tyr Pro Gly Ser Gly Ile Ala
195 200 205
Gln Phe His Ile Asp Tyr Asn Asn Val Ser Ser Ala Glu Gly Trp His
210 215 220
Val Asn Val Thr Leu Asn Ile Arg Pro Ser Thr Gly Thr Gly Val Met
225 230 235 240
Leu Ala Leu Val Ser Gly Asn Asn Thr Val Pro Phe Ala Val Ser Leu
245 250 255
Val Asp Ser Thr Ser Glu Lys Ser Gln Asp Ile Leu Leu Ser Val Glu
260 265 270
Asn Thr Val Ile Tyr Arg Ile Gln Ala Leu Ser Leu Cys Ser Asp Gln
275 280 285
Gln Ser His Leu Glu Phe Arg Val Asn Arg Asn Asn Leu Glu Leu Ser
290 295 300
Thr Pro Leu Lys Ile Glu Thr Ile Ser His Glu Asp Leu Gln Arg Gln
305 310 315 320
Leu Ala Val Leu Asp Lys Ala Met Lys Ala Lys Val Ala Thr Tyr Leu
325 330 335
Gly Gly Leu Pro Asp Val Pro Phe Ser Ala Thr Pro Val Asn Ala Phe
340 345 350
Tyr Asn Gly Cys Met Glu Val Asn Ile Asn Gly Val Gln Leu Asp Leu
355 360 365
Asp Glu Ala Ile Ser Lys His Asn Asp Ile Arg Ala His Ser Cys Pro
370 375 380
Ser Val Trp Lys Lys Thr Lys Asn Ser
385 390
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ БЕТА-АМИЛОИДА, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2777844C1 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ БЕТА-АМИЛОИДУ/ЧЕЛОВЕЧЕСКОМУ РЕЦЕПТОРУ ТРАНСФЕРРИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2730682C1 |
| СЛИТЫЙ БЕЛОК, СОДЕРЖАЩИЙ ВАРИАНТЫ ЧЕЛОВЕЧЕСКОГО БЕЛКА LEFTY А, И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2793631C2 |
| HER-2-НАПРАВЛЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ 4-1BBL | 2019 |
|
RU2815451C2 |
| ИСКУССТВЕННЫЕ АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2763798C1 |
| ВЫСОКОГЛИКОЗИЛИРОВАННЫЙ СЛИТЫЙ БЕЛОК НА ОСНОВЕ ФАКТОРА СВЕРТЫВАНИЯ КРОВИ ЧЕЛОВЕКА VIII, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2722374C1 |
| МОЛЕКУЛЫ IMMTAC, СВЯЗЫВАЮЩИЕ РЕСТРИКТИРОВАННЫЙ ПО HLA-A*02 ПЕПТИД, С УВЕЛИЧЕННЫМ ПЕРИОДОМ ПОЛУВЫВЕДЕНИЯ | 2020 |
|
RU2833530C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ КОМПЛЕКСА ОПУХОЛЕВЫЙ АНТИГЕН NY-ESO-1/HLA-A*02 | 2016 |
|
RU2775394C2 |
| СВЯЗЫВАНИЕ СЛИТОГО БЕЛКА С БЕЛКОМ CD47 И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2787521C2 |
| СЛИТЫЙ БЕЛОК, СОДЕРЖАЩИЙ РЕЦЕПТОР TGF-БЕТА, И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2776204C1 |
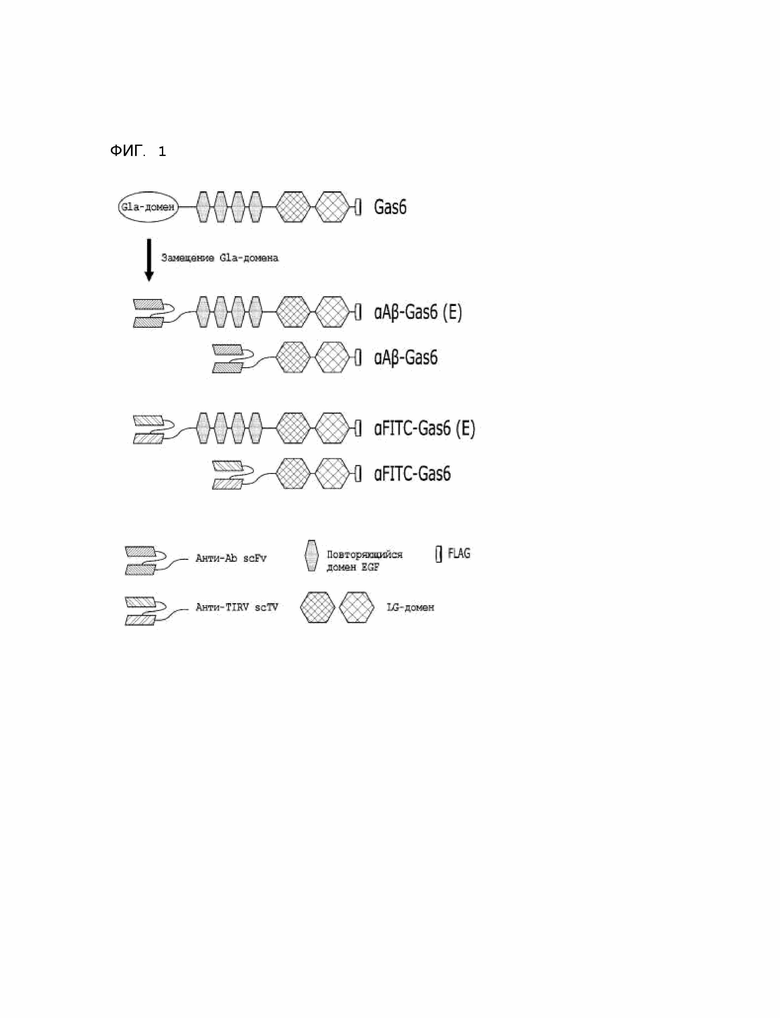
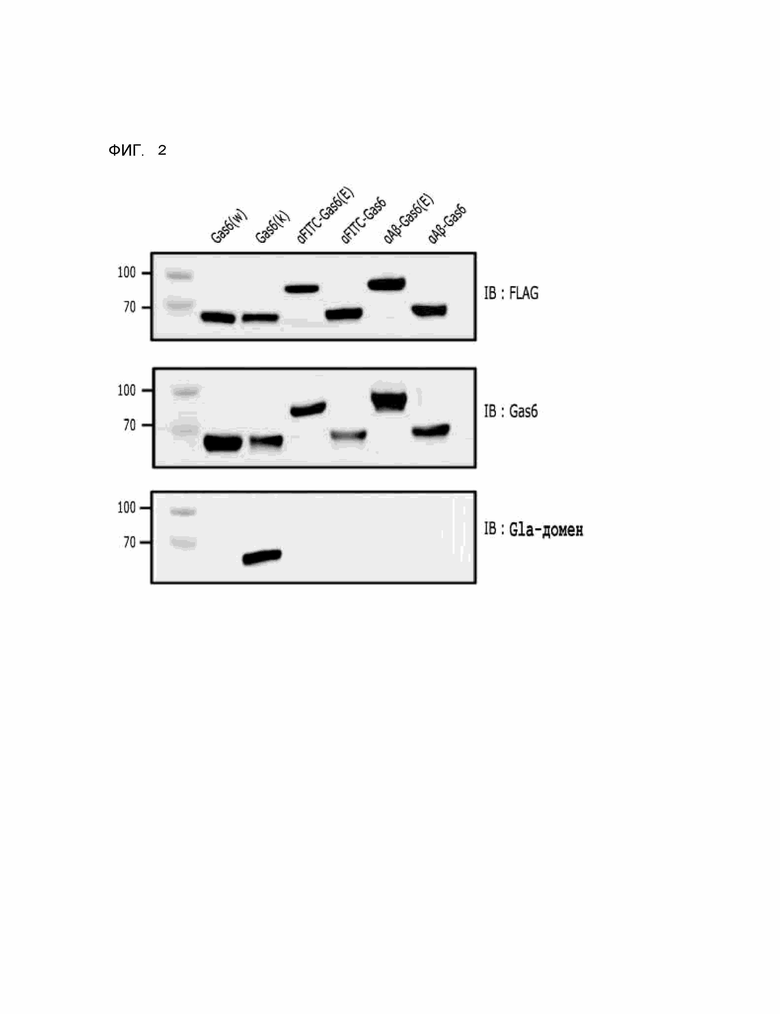
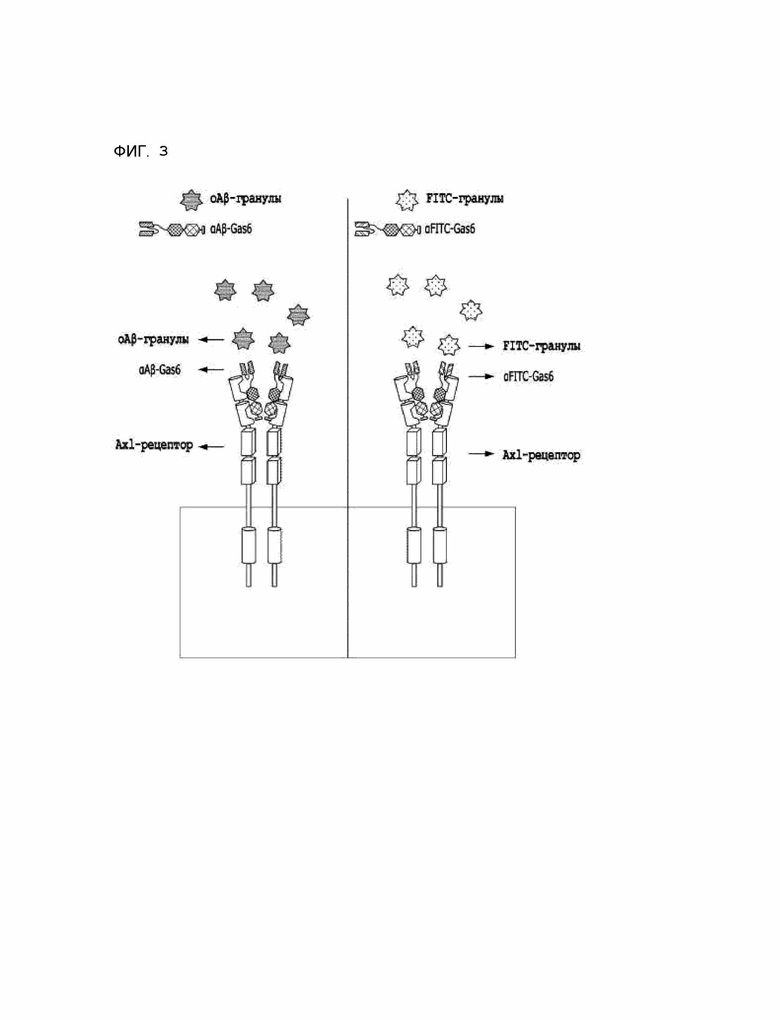
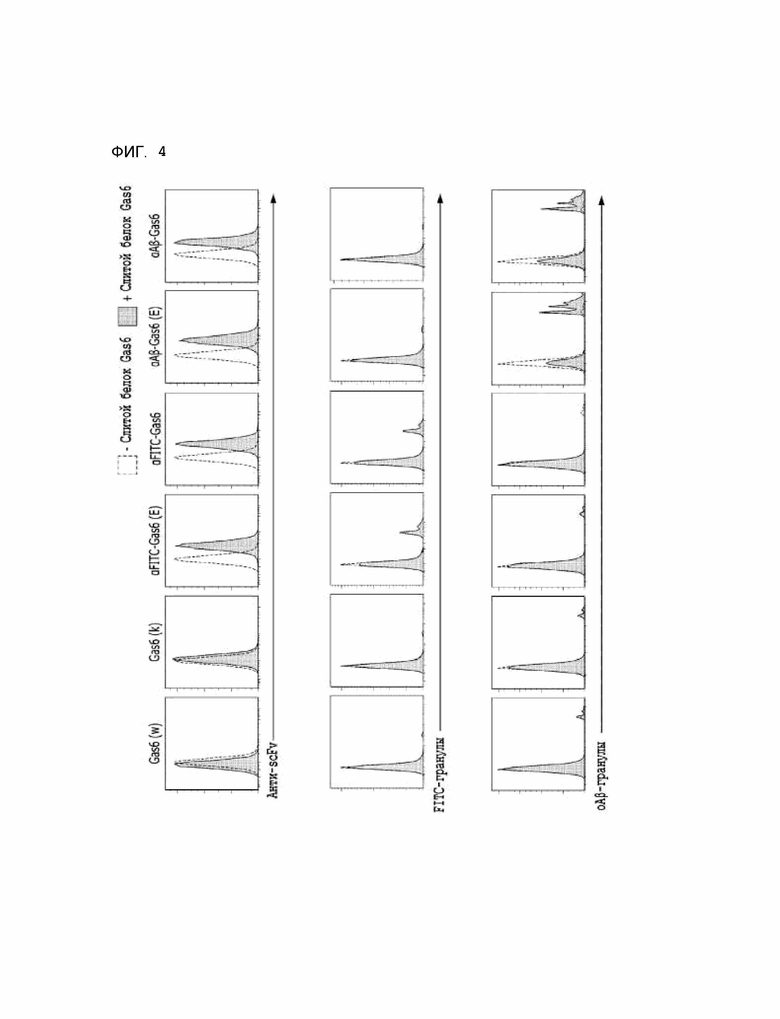
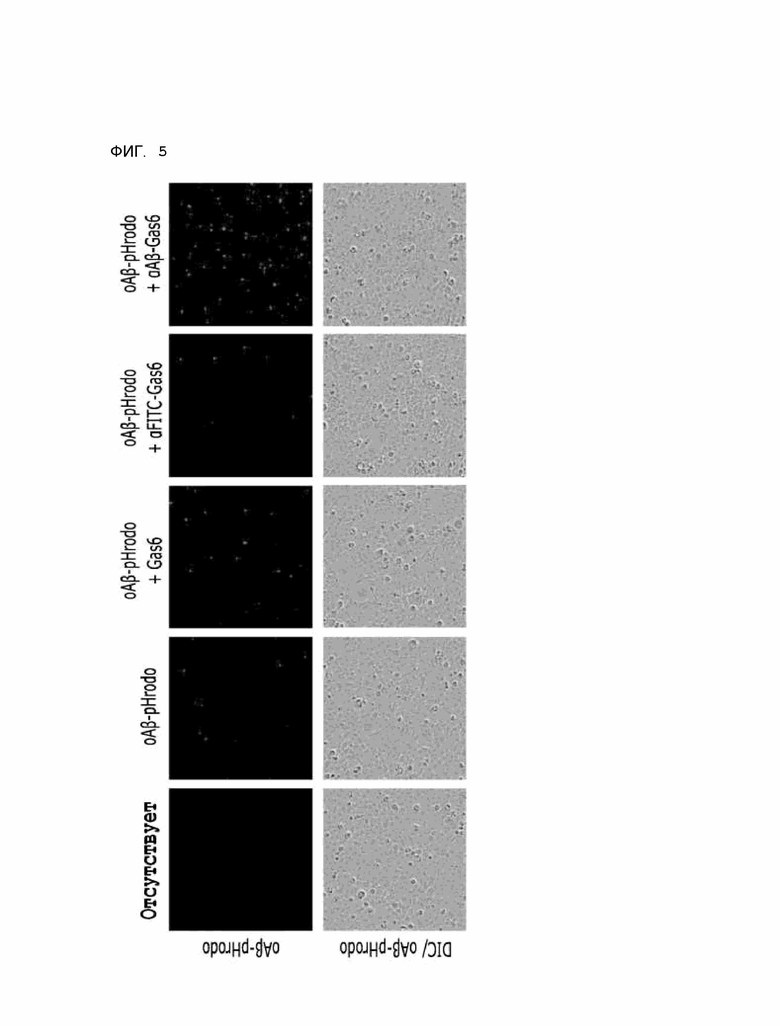
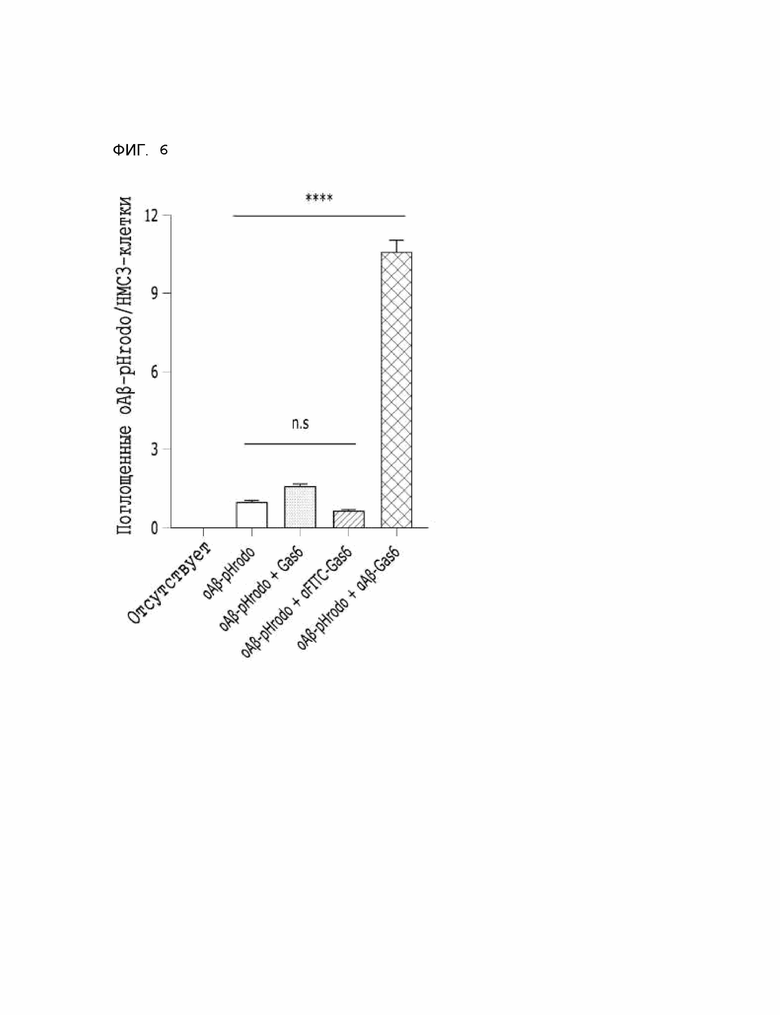
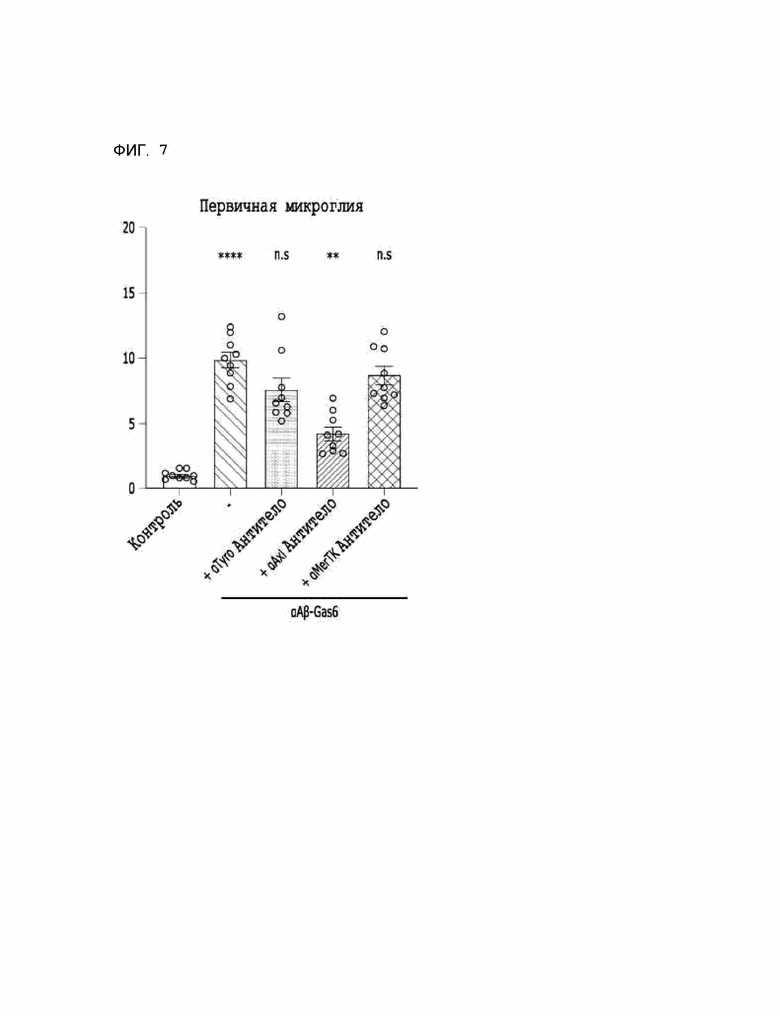
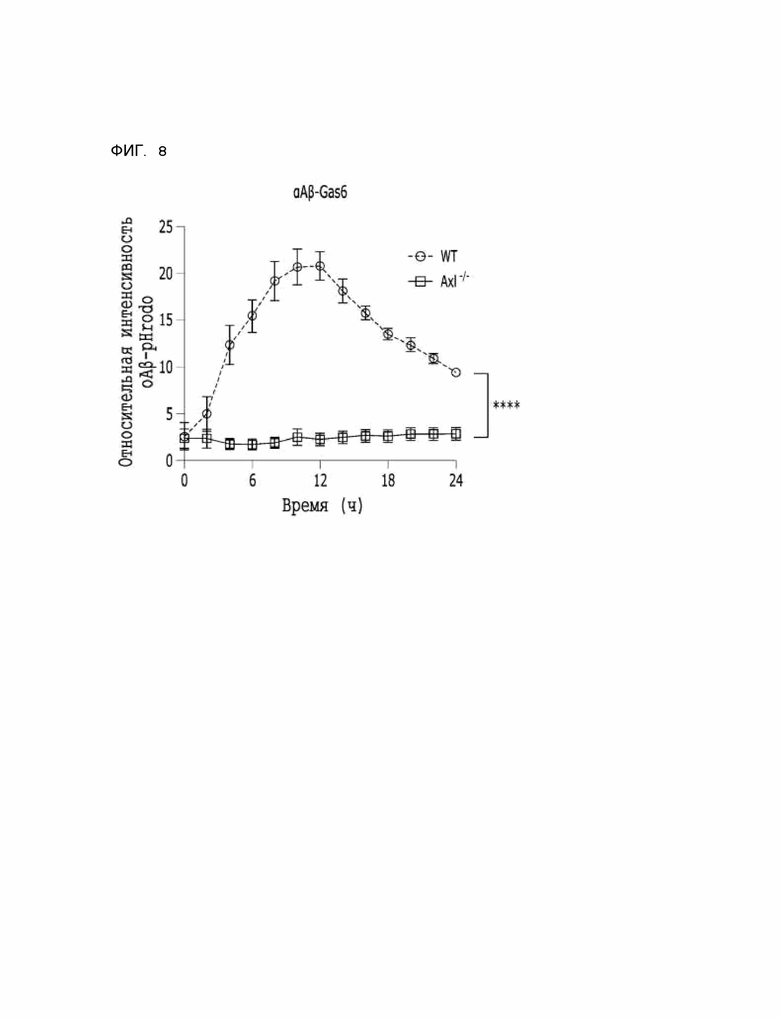
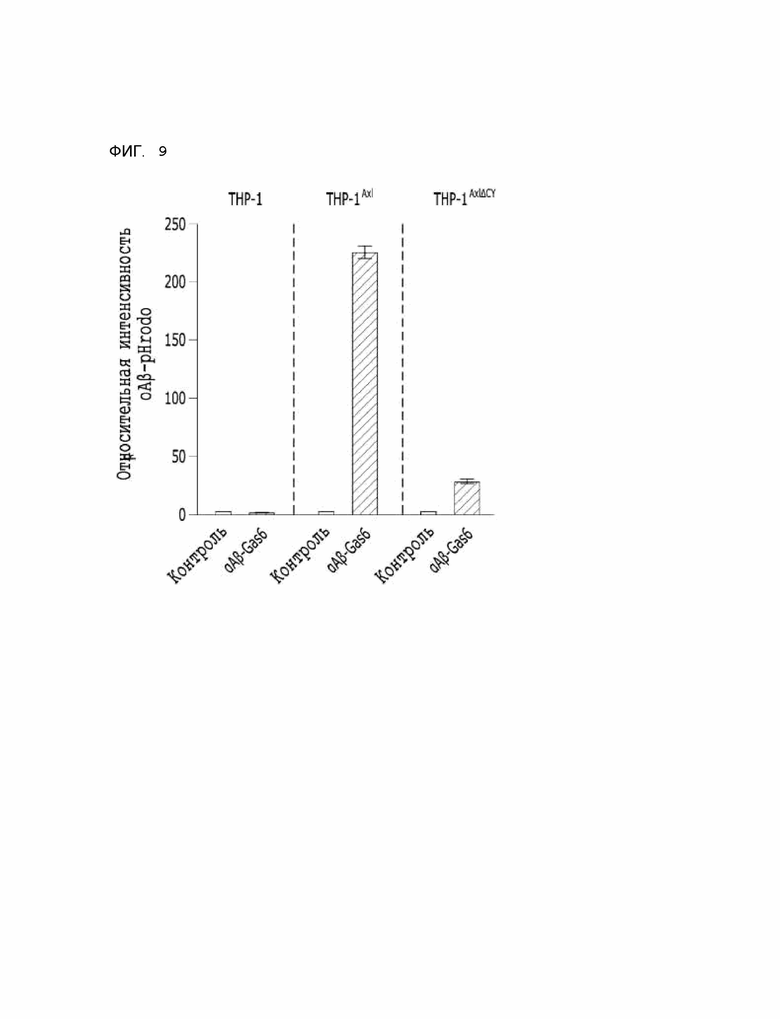
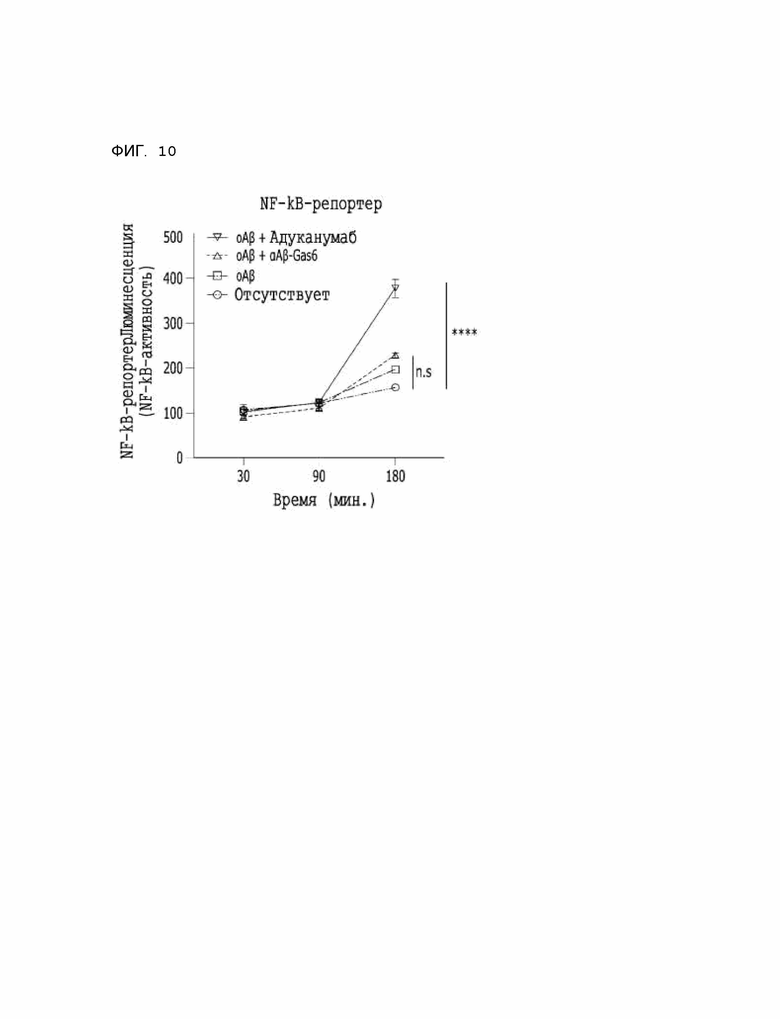
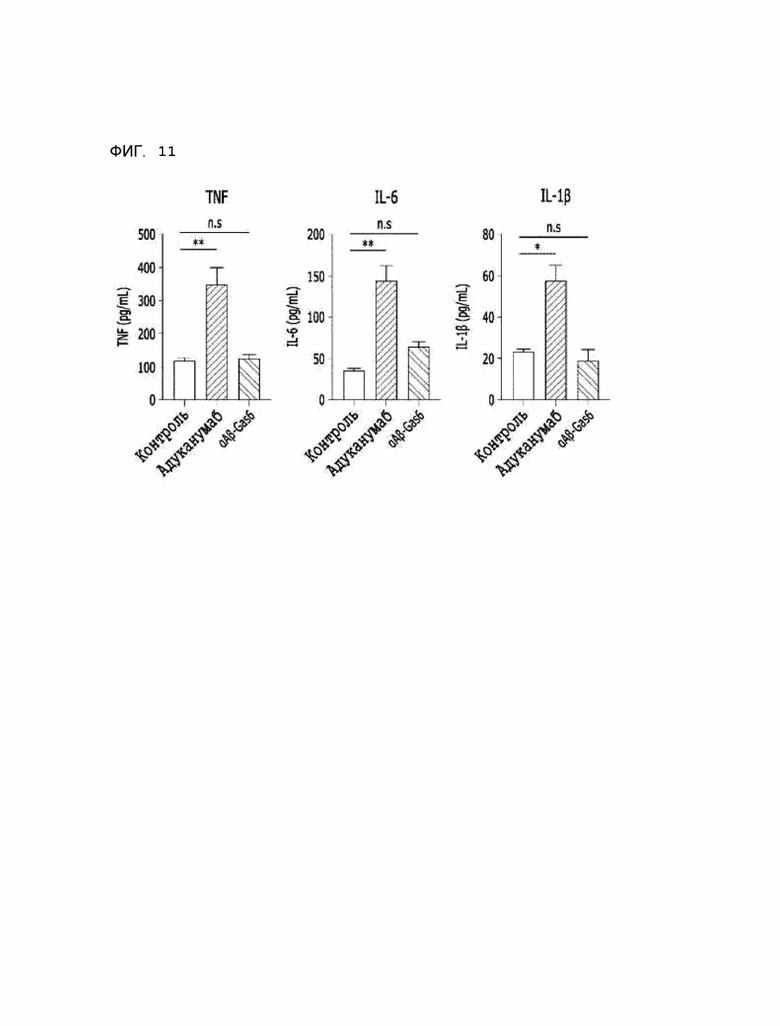
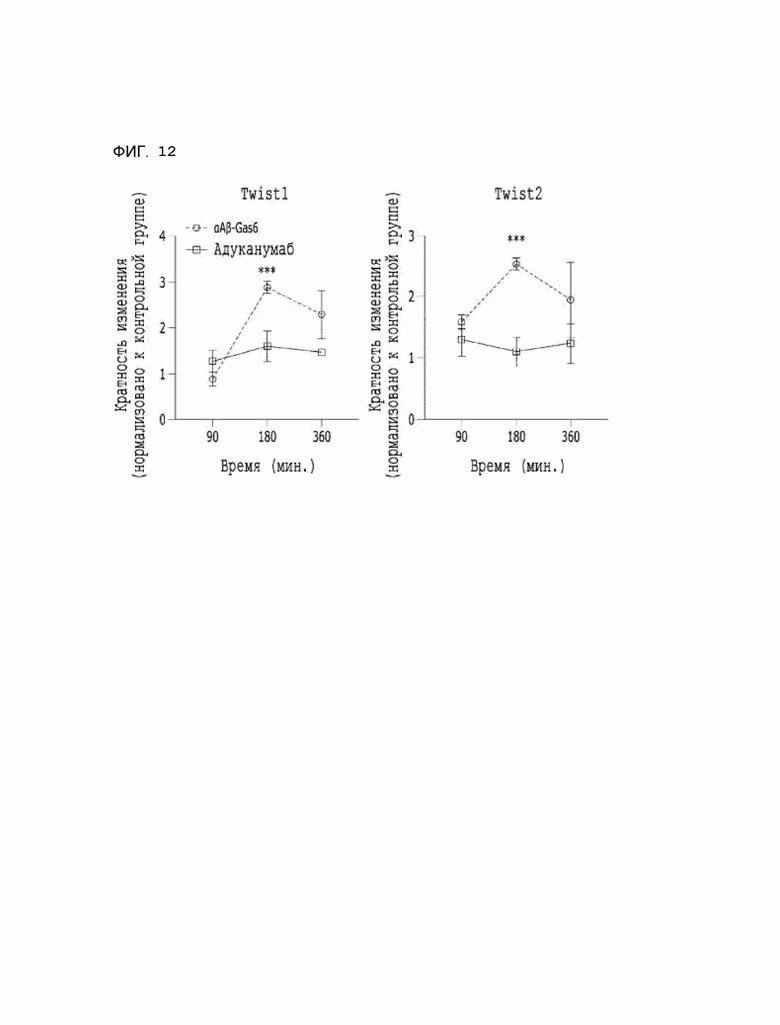
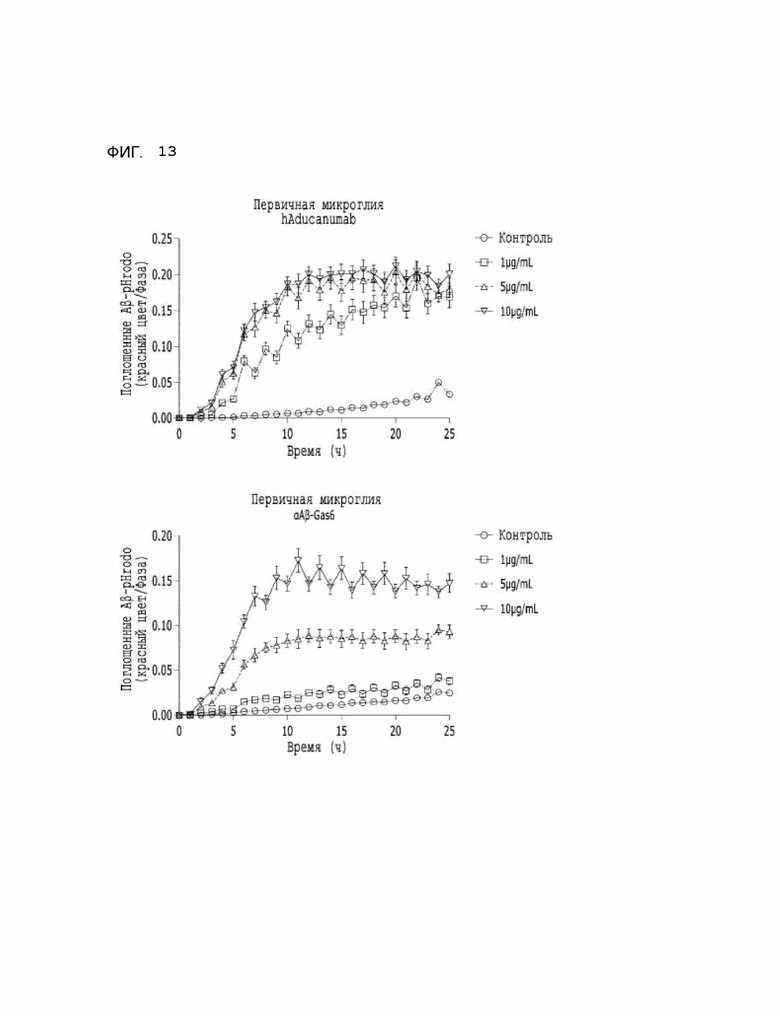
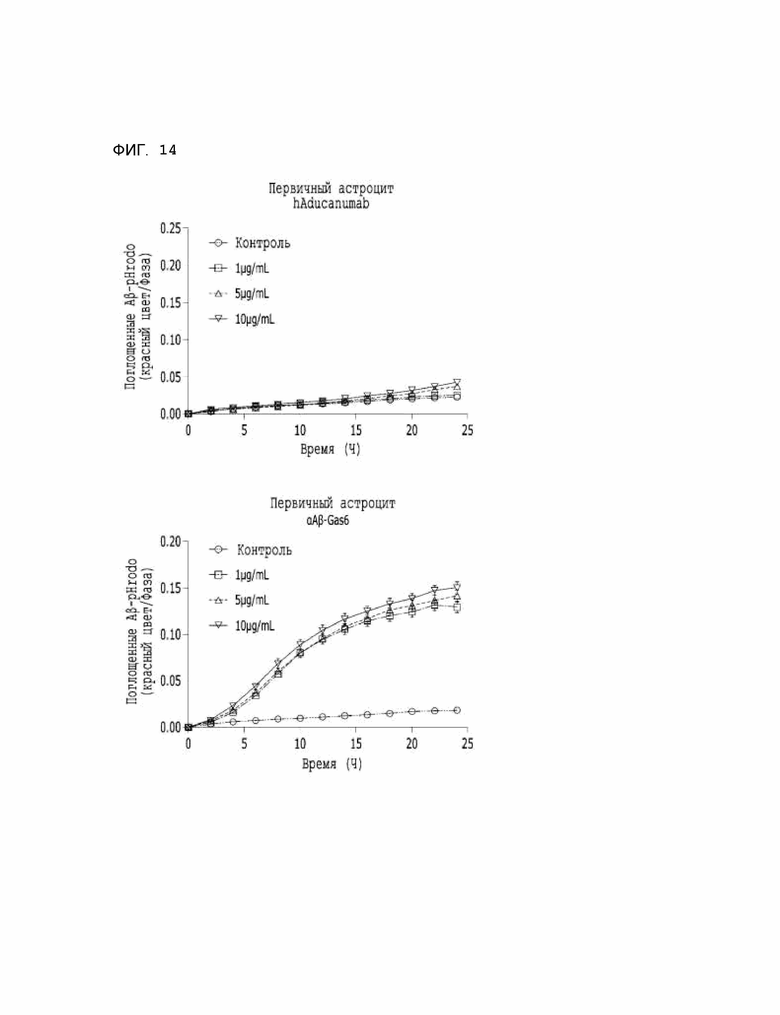
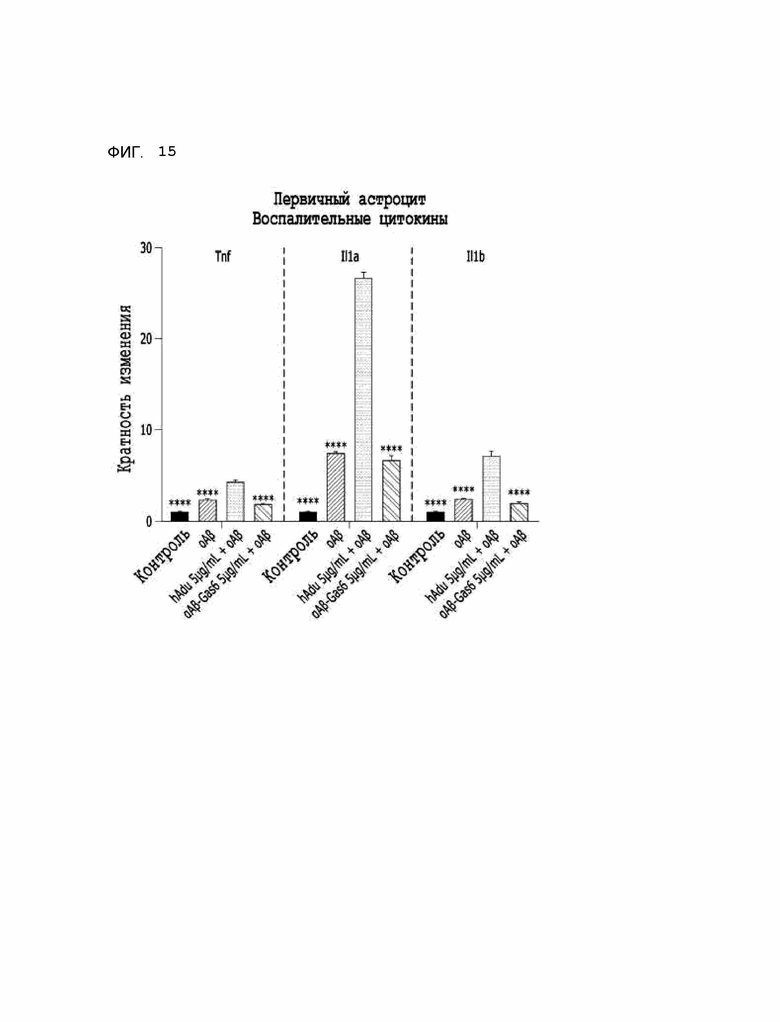
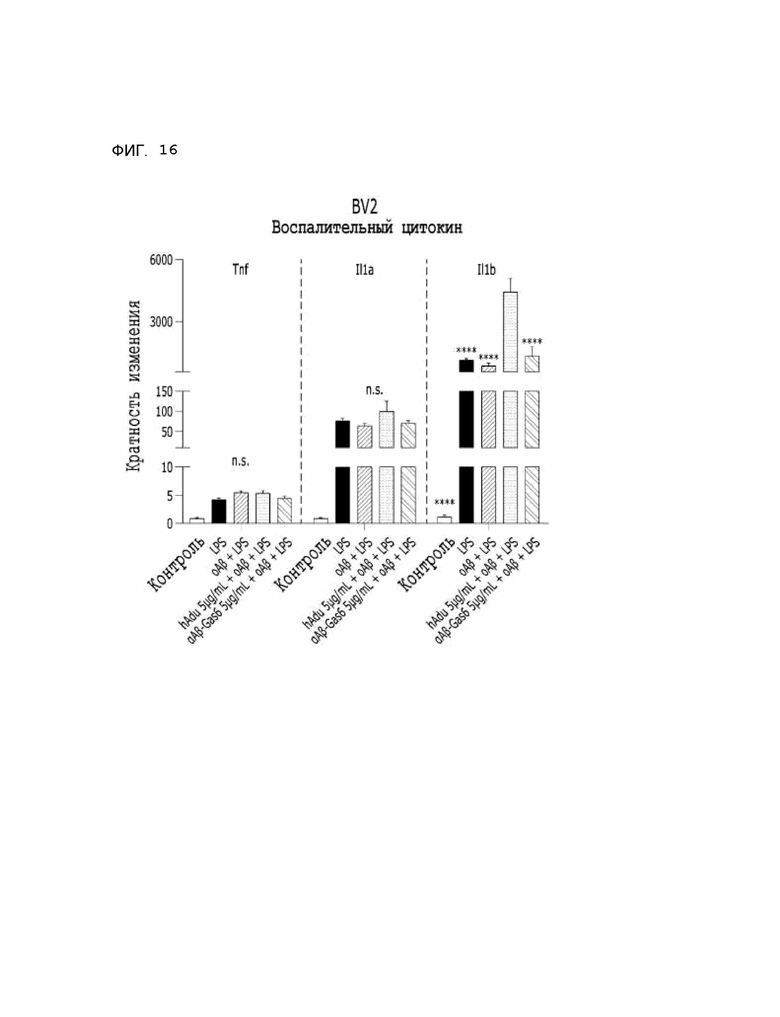
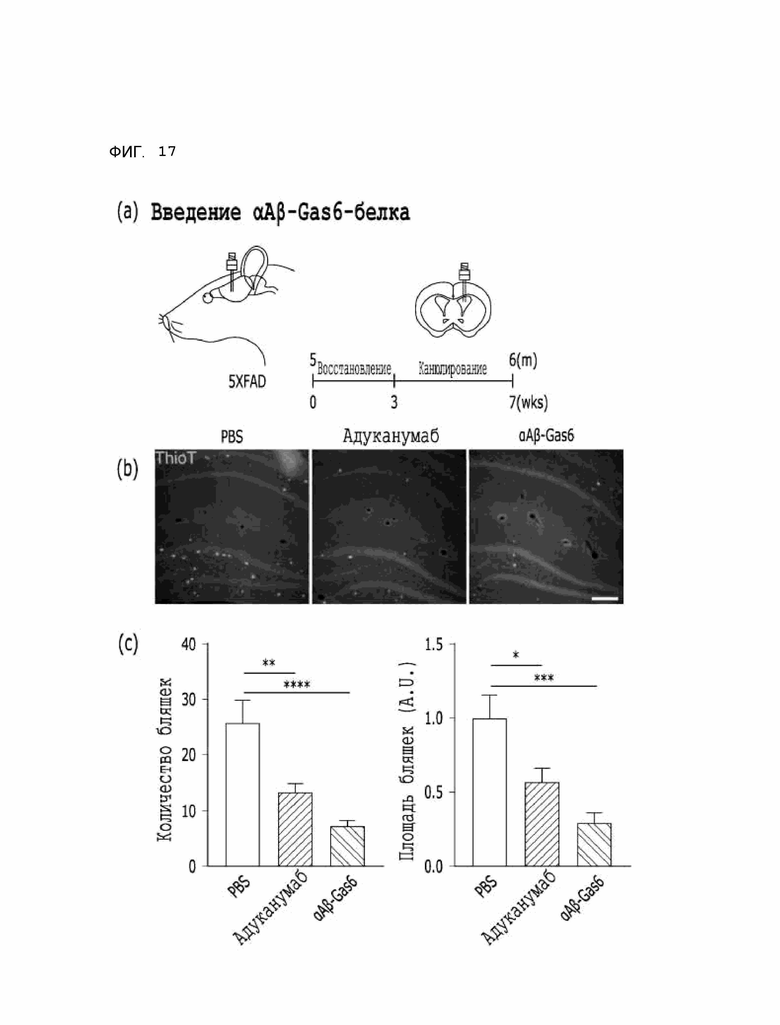
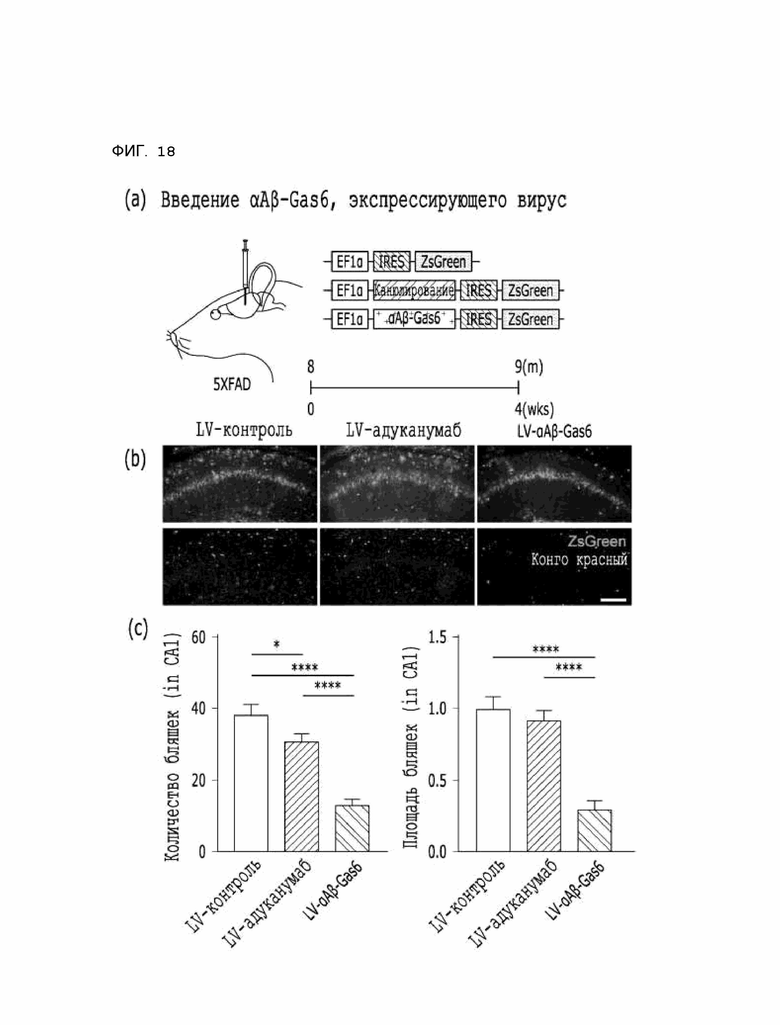
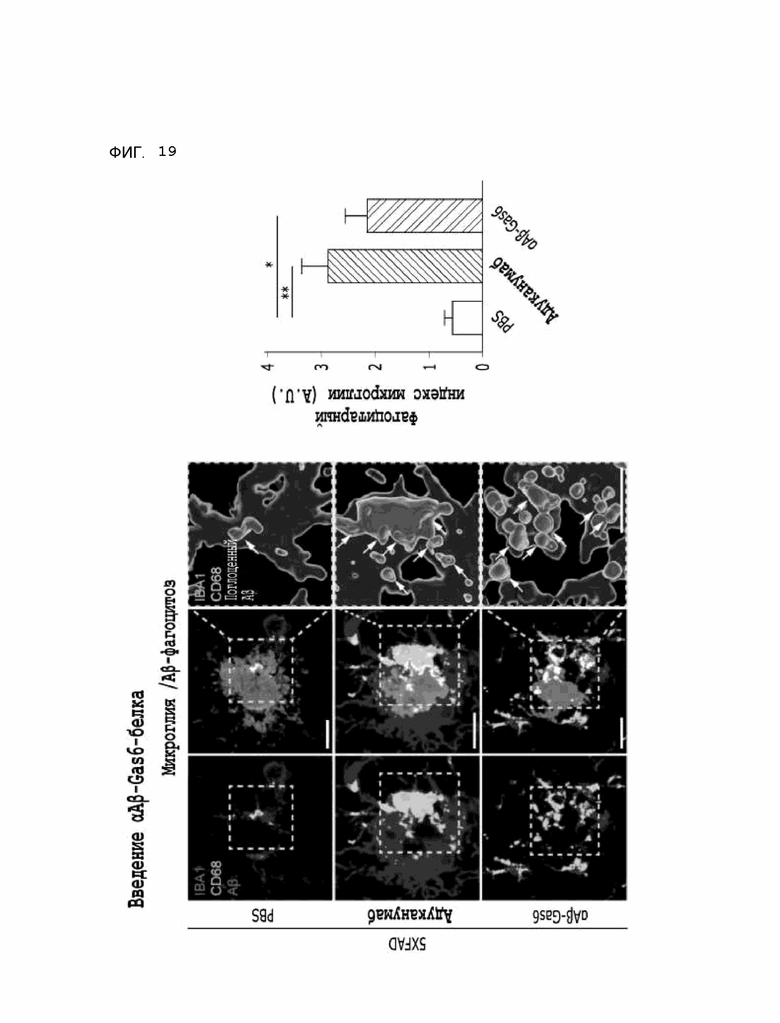
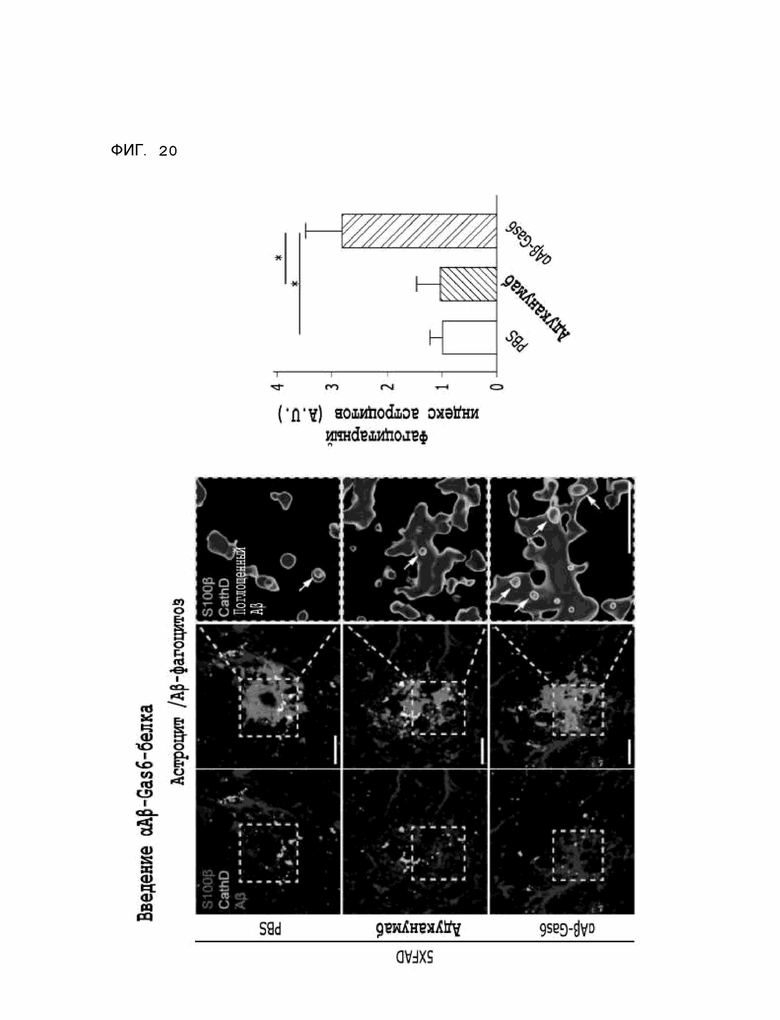
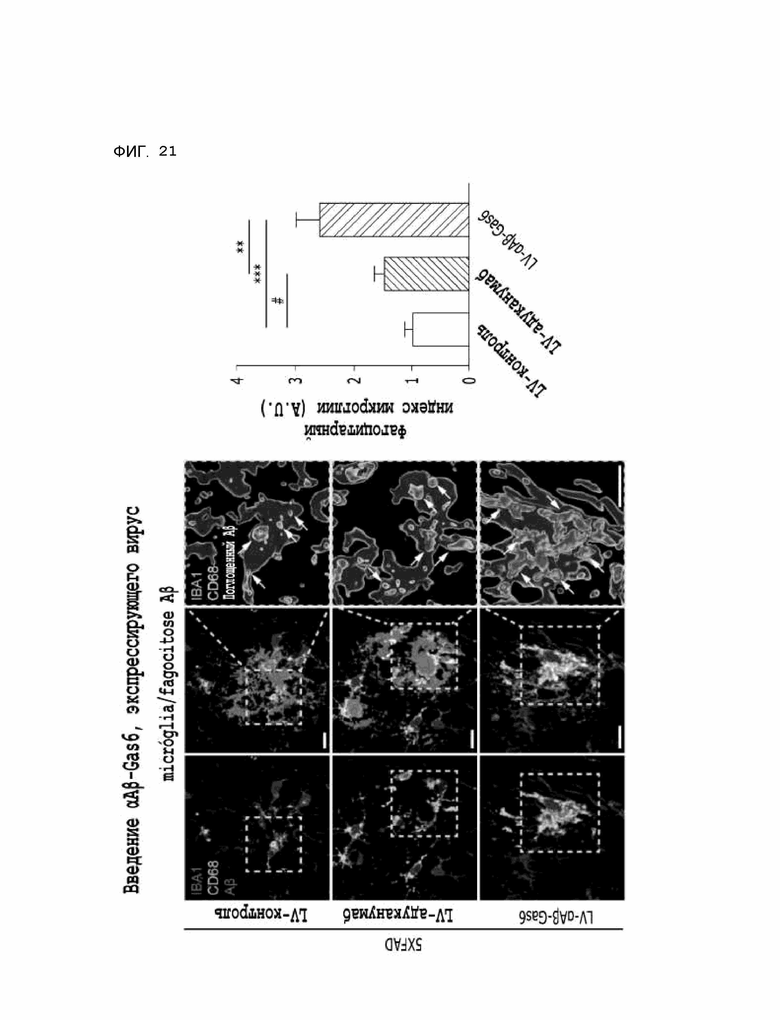
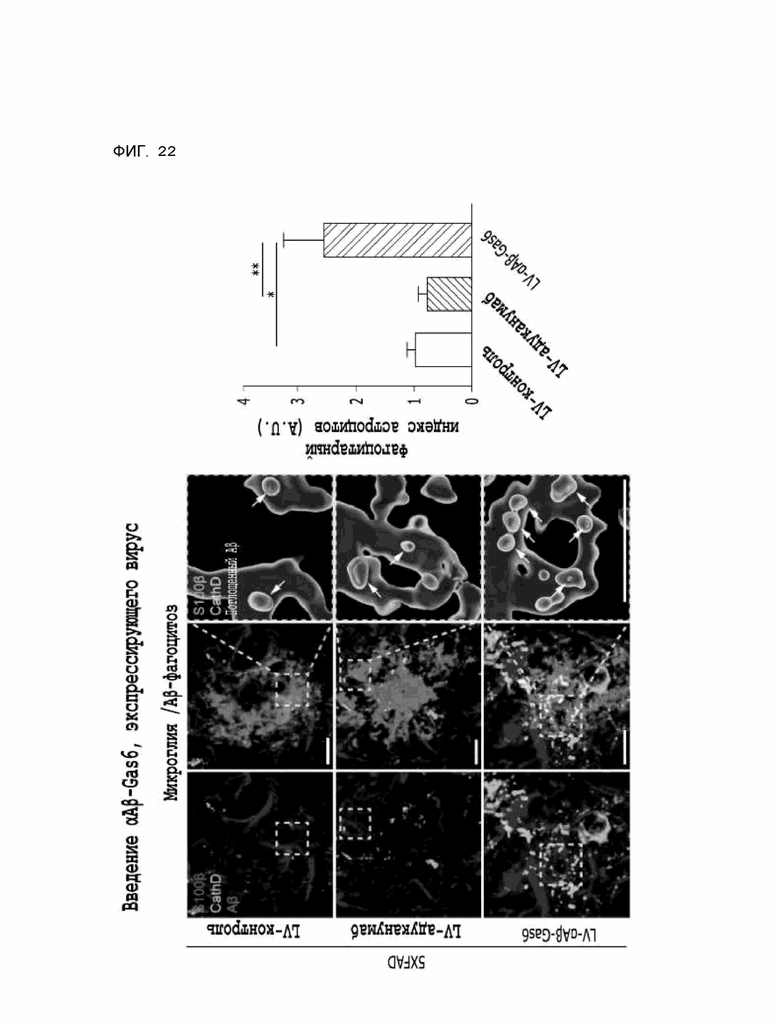
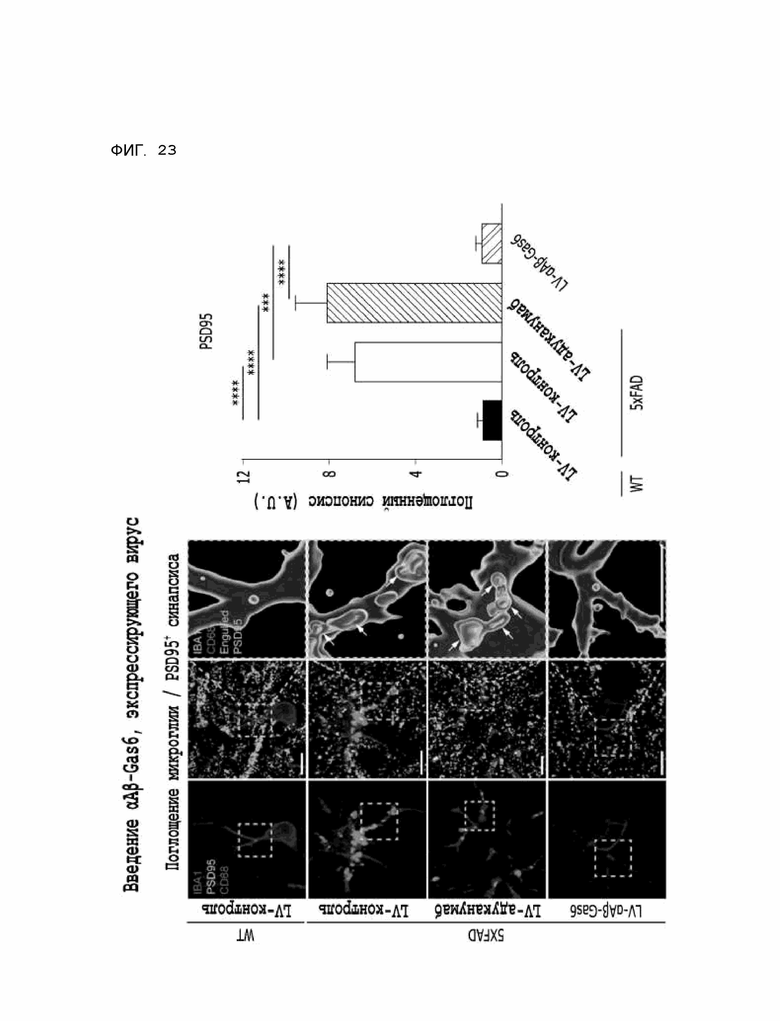
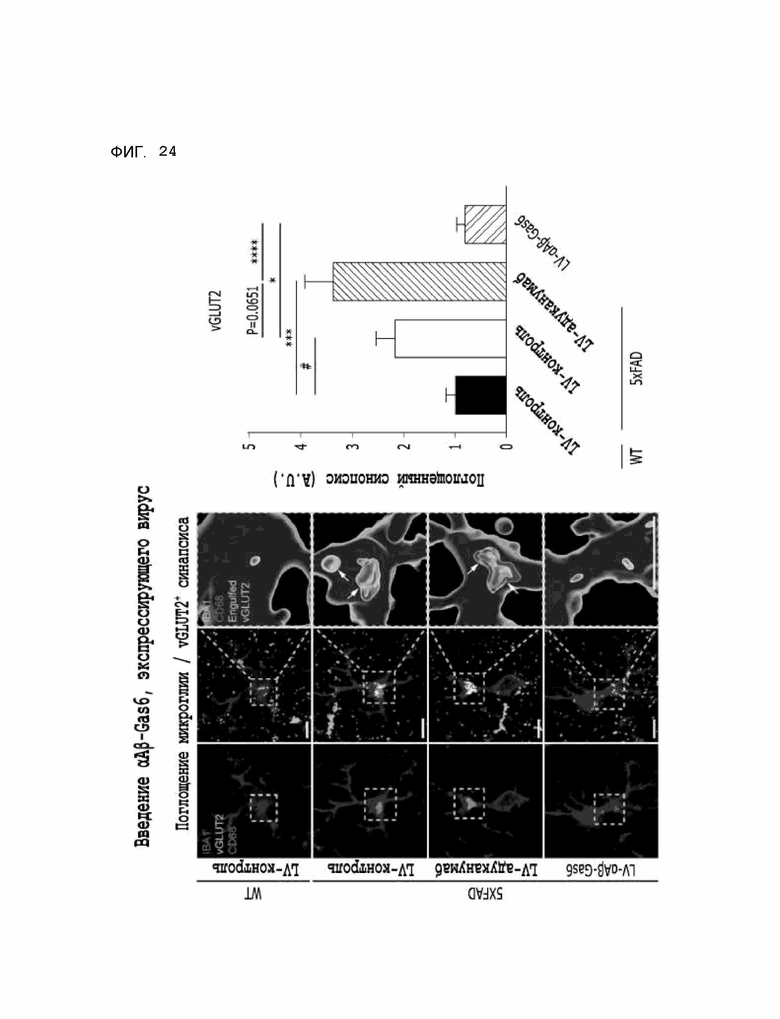
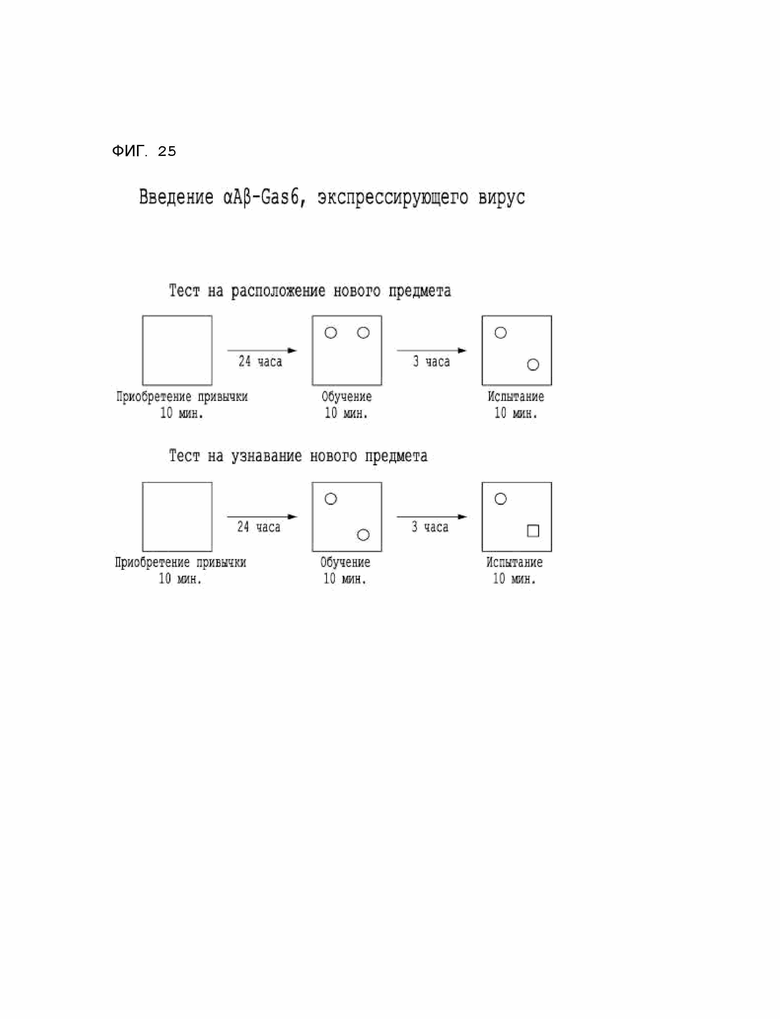
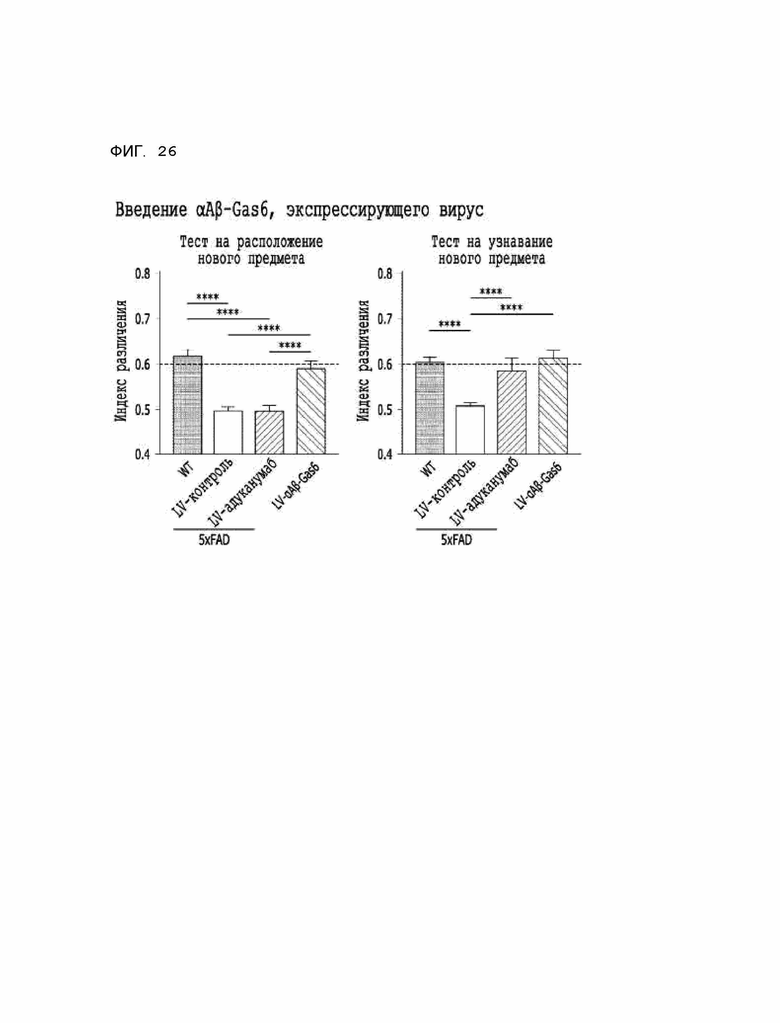
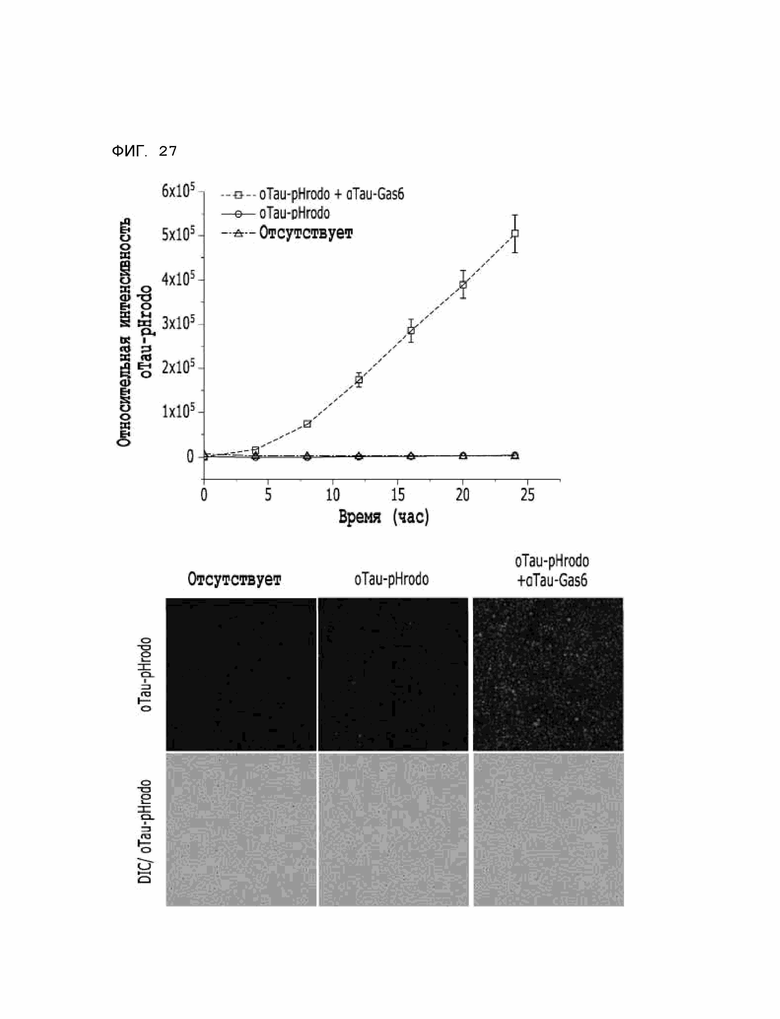
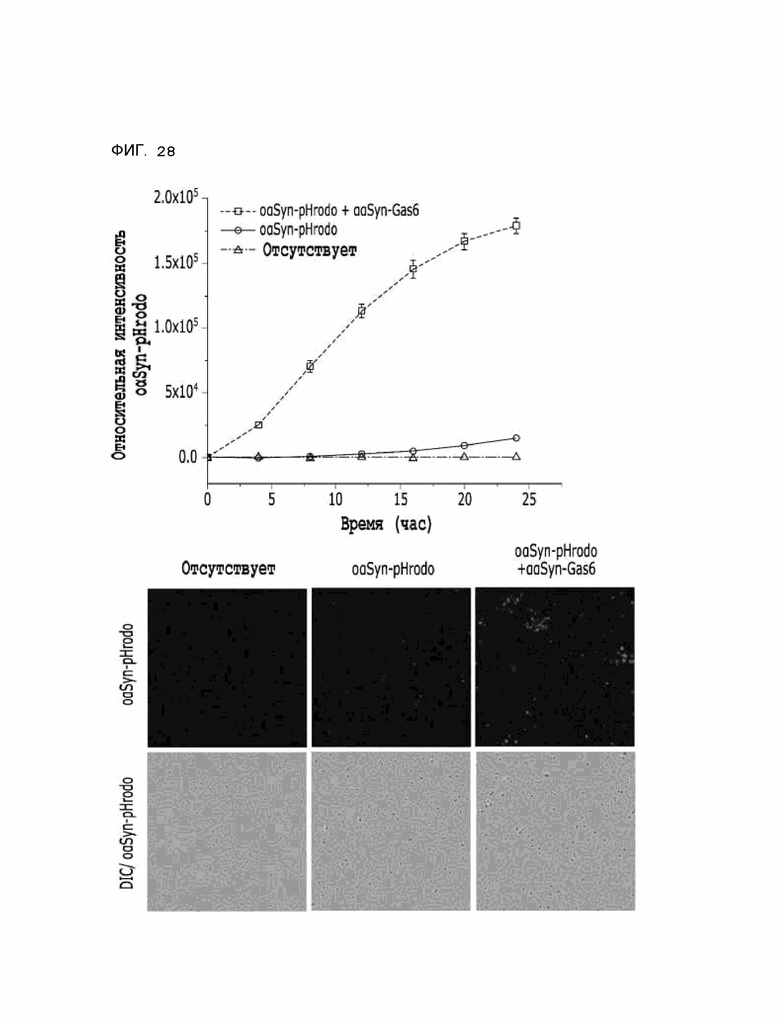
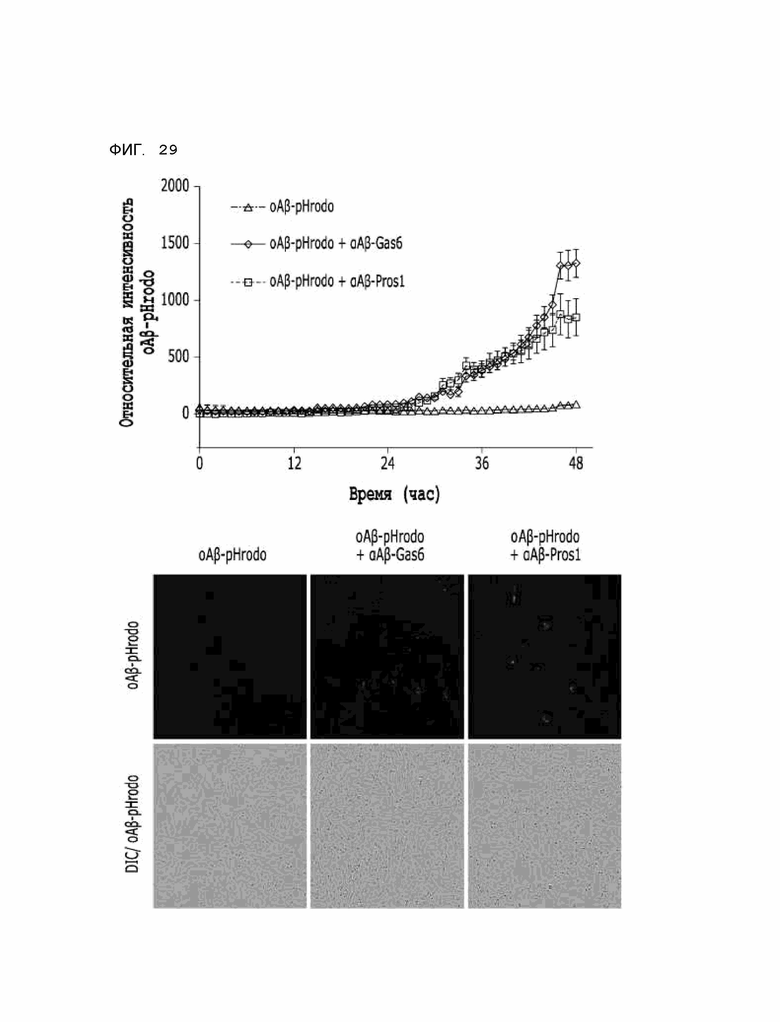
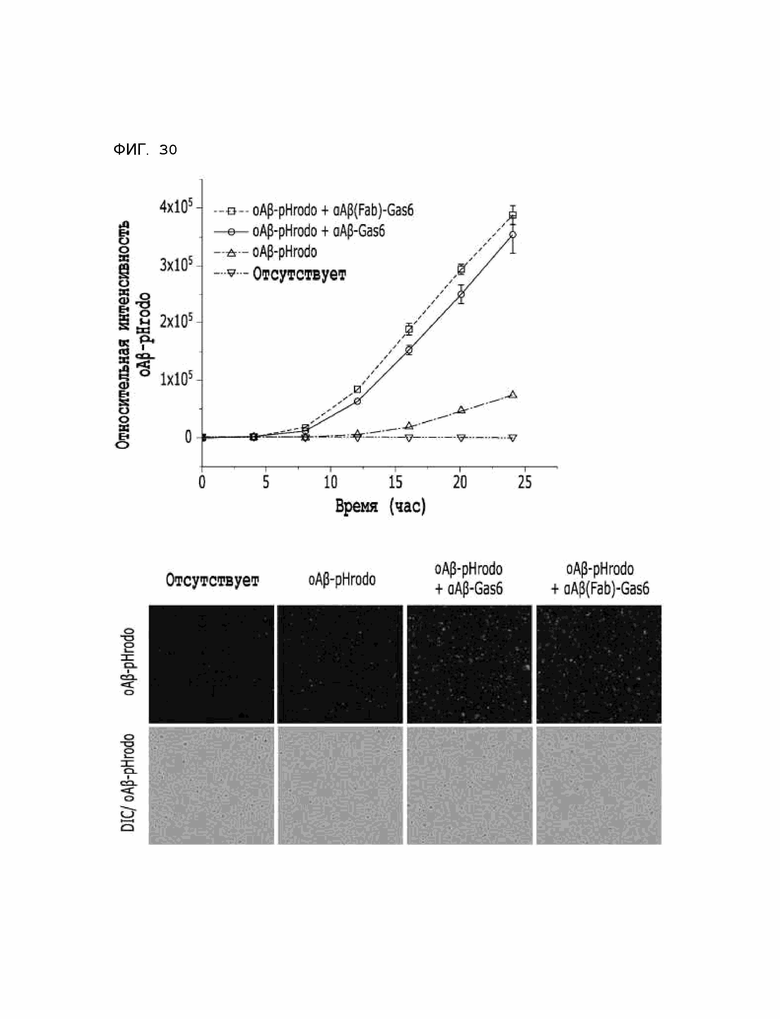
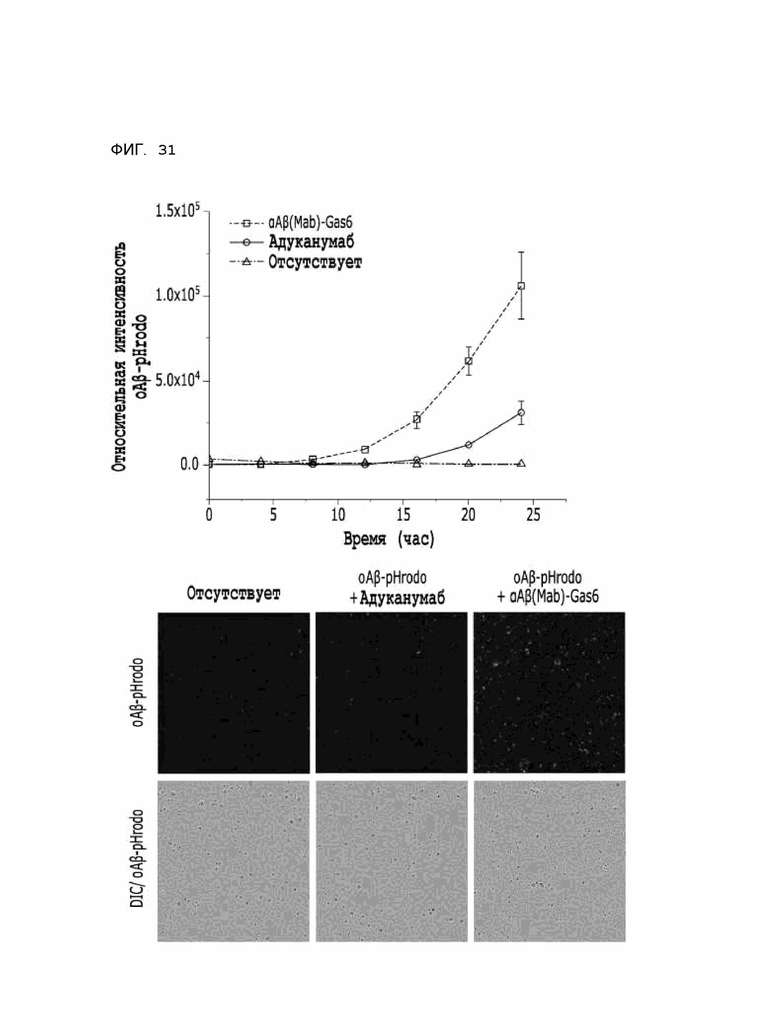
Настоящее изобретение относится к области биотехнологии, в частности к слитой молекуле, обладающей активностью индуцировать невоспалительный фагоцитоз, причем слитая молекула содержит: первый участок, способный связываться с TAM-рецептором, и второй участок, который специфически связывается с таргетным веществом, причем первый участок содержит ламинин-G-подобный домен Gas6 или ProS1 или их активный фрагмент. Также раскрыты нуклеиновая кислота, кодирующая слитую молекулу, вектор экспрессии, клетка, экспрессирующая слитую молекулу, и фармацевтическая композиция, содержащая слитую молекулу или вектор экспрессии. Представленное изобретение эффективно для индукции невоспалительного фагоцитоза белковых веществ, аномальное накопление которых вызывает развитие заболеваний. 5 н. и 11 з.п. ф-лы, 31 ил., 8 табл., 5 пр.
1. Слитая молекула, обладающая активностью индуцировать фагоцитоз, причем слитая молекула содержит: первый участок, способный связываться с TAM-рецептором, и второй участок, который специфически связывается с таргетным веществом, причем первый участок содержит ламинин-G-подобный домен Gas6 или ProS1 или их активный фрагмент.
2. Слитая молекула по п. 1, причем TAM-рецептор представляет собой по крайней мере один рецептор, выбранный из группы, включающей Tyro3, Axl и MerTK.
3. Слитая молекула по п. 1, причем первый участок содержит Gas6, ProS1, Tubby, Tulp1 или Gal3 или их активные фрагменты.
4. Слитая молекула по п. 1, причем первый участок представляет собой ламинин-G-подобный домен, содержащий последовательность SEQ ID NO: 1 и 2, или ламинин-G-подобный домен, содержащий последовательность SEQ ID NO: 3 и 4.
5. Слитая молекула по п. 1, причем таргетное вещество представляет собой вещество, накапливающееся в живых тканях, и это накопление вызывает заболевание.
6. Слитая молекула по п. 5, причем таргетным веществом является амилоид.
7. Слитая молекула по п. 5, причем заболеванием является амилоидоз.
8. Слитая молекула по п. 1, причем таргетное вещество выбрано из бета-амиолиода, тау, альфа-синуклеина, белка гентингтина, прионного белка, амилоидного предшественника бета-амилоида белкового происхождения, PrPSc ассоциированного с микротрубочками тау, экзона 1 гентингтина, TAR ДНК-связывающего белка 43, супероксиддисмутазы 1 (SOD1), ABri-пептида, ADan-пептида, фрагмента легкой цепи иммуноглобулина, фрагмента тяжелой цепи иммуноглобулина, N-концевого фрагмента белка сывороточного амилоида А, транстиретина, бета-2-микроглобулина, N-концевого фрагмента аполипопротеина AI, удлиненного с С-конца аполипопротеина AII, N-концевого фрагмента аполипопротеина AIV, аполипопротеина C-II, аполипопротеина C-III, фрагмента гельсолина, лизоцима, фрагмента альфа цепи фибриногена, усеченного по N-концу цистатина С, амилина, кальцитонина, предсердного натрийуретического фактора, пролактина, инсулина, лактадгерина, медина, лактотрансферрина, лактоферрина, ODAM (одонтогенного амелобласт-ассоциированного белка), SP-C белка (легочного сурфактант-ассоциированного белка С), LECT-2 (хемотаксина-2 лейкоцитарного происхождения), галектина-7, корнеодесмозина, С-концевого фрагмента TGFBI (или кератоэпителина), SGI (семеногелина-1), S100-белка (A8 или A9) и энфувиртида.
9. Слитая молекула по п. 1, причем второй участок, специфически связывающийся с таргетным веществом, выбран из антитела, его активного фрагмента, антителоподобного белка, пептида, аптамера и растворимого рецептора, которые способны специфически связываться с таргетным веществом.
10. Слитая молекула по п. 1, причем фагоцитоз индуцирован в клетках, экспрессирующих TAM-рецептор.
11. Слитая молекула по п. 10, причем клетки, экспрессирующие TAM-рецептор, представляют собой по крайней мере один тип профессиональных фагоцитов, по крайней мере один тип непрофессиональных фагоцитов или их комбинацию.
12. Слитая молекула по п. 1, причем индукция фагоцитоза не включает воспалительный ответ.
13. Молекула нуклеиновой кислоты, кодирующая слитую молекулу по п. 1, причем указанная слитая молекула содержит: первый участок, способный связываться с TAM-рецептором, и второй участок, который специфически связывается с таргетным веществом, причем первый участок содержит ламинин-G-подобный домен Gas6 или ProS1 или их активный фрагмент.
14. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п. 13.
15. Клетка, экспрессирующая слитую молекулу по п. 1, содержащая молекулу нуклеиновой кислоты по п. 13 или вектор экспрессии по п. 14.
16. Фармацевтическая композиция для предотвращения или лечения заболеваний, вызванных накоплением таргетного белкового вещества в живых тканях, причем фармацевтическая композиция содержит слитую молекулу по п. 1 или вектор экспрессии по п. 14 и фармацевтически приемлемый носитель.
| WO 2017083700 A1, 18.05.2017 | |||
| US 2018186855 A1, 05.07.2018 | |||
| CA 3098093 A1, 07.11.2019 | |||
| CN 112194726 A, 08.01.2021 | |||
| ВОРОБЬЕВА Н.В | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Счетный сектор | 1919 |
|
SU107A1 |

