Изобретение относится к биологии и медицине, конкретно - к антисенстерапии с использованием антисмысловых олигонуклеотидов.
Известно использование синтетических олигонуклеотидов с линейной структурой молекулы для блокирования ряда биологических процессов, при этом механизм действия олигонуклеотидов основан на явлении их комплементарности по отношению к своим мишеням - нуклеиновым кислотам (E. Uhlmanh. A. Peyman. Antisense oligonucleotides: A new therapeutic principle // Chemical Reviews. - 1990. - 90, 4. - Р. 543-581; Д.К. Кнорре. Ген-направленные биологически активные вещества (антисмысловые олигонуклеотиды и их производные) // Биоорганическая химия. - 1997. - 23, 1. - С. 3-17). Применение антисмысловых олигонуклеотидов описано в патенте США (4904582 от 27 февраля 1990 г.; Richard H. Tullis; Novel amphiphilic Nucleic acid conjugates).
Однако эти методы основаны на применении олигонуклеотидов, комплементарных определенным участкам геномной ДНК или отдельным участкам определенных информационных рибонуклеиновых кислот (иРНК). Так как такие участки ДНК и иРНК несут определенную информационную (смысловую) нагрузку, то комплементарные к ним олигонуклеотиды были названы антисмысловыми.
Описана возможность применения антимикробных антисмысловых олигонуклеотидов, в частности, для подавления развития микобактерий (Е. Rарарort. F. Levina, V. Metelev, P.C. Zamecnik. Antimycobacterial activities of antisense oligodeoxynucleotide phosphorothiates in drug-resistant strains // Proc. Natl. Acad. Sci. USA - 1996. - 93. - P. 709-713).
Основной недостаток применяемых антисмысловых олигонуклеотидов для подавления жизнедеятельности патогенных микроорганизмов заключается в быстрой изменчивости мишеней воздействия вследствие мутаций и появления у микроорганизмов устойчивости к препаратам.
В качестве мишеней могут быть использованы отдельные участки рибосомальной РНК (рРНК) микроорганизмов, в частности, расположенные на 3''-концевой открытой последовательности 16S рРНК. Однако эта последовательность является крайне консервативной, проявляя некоторое отличие лишь на уровне окрашивания микроорганизмов по Граму. У грамположительных бактерий она представлена последовательностью 3''-УЦЦУ (УЦЦУЦЦ) АЦУАГ-5'', а у грамотрицательных бактерий - 3''-АУ (УЦЦУЦЦ) АЦУАГ-5''. Наибольший интерес вызывает последовательность 3-УЦЦУЦЦ-5'', комплементарная так называемой последовательности Шайн-Далгарно (5''-АГГАГГ-3'') мРНК. Блокирование этой последовательности комплементарными олигонуклеотидами предотвращает присоединение к 3-концу 16S рРНК мРНК, их трансляцию на рибосомах и синтез белка. Ряд исследователей (Taniguchi Т, Weissman С.Inhibition of SBRNA 70S ribosome initiation complex formation by on oligonucleotide complementary to the 3 terminal region of E. coli 16S ribosomal RNA. - Nature, 1978, u. 275, 3. P. 770-772; Eckhardt H., Luhrrman R. Blocking of the initiation of protein biosynthesis by a pentanucleotide complementary to the 3''-end of Escherichia coli 16S rRNA // J. Biol. Chem. - 1979. - 254, 22. - P. 11183-11188) достигли при помощи комплементарных ей синтетических пентануклеогидов 70% подавления связывания иРНК с рибосомами в системе in vitro.
Известны работы, в которых с помощью комплементарных этой же oблacти олигонуклеотидов получено в бесклеточной системе почти 90% подавление синтеза белка, а в клетке - почти 50% (Jayaramen К., Mс Pariand К., Miller P., Ts"o P. Selective inhibition of Escherichia coli protein synthesis and growth by nonionic oligonucleotides complementary to the 3''-end of 16S rRNA \\ Proc. Nat. Acad. Sci. USA. - 1981. - 78. 3. - P. 1537-1541), а при использовании пентануклеотидов - полное ингибирование роста Е. coli (Rahman et al. , Antibacterial activity and inhibition of protein synthesis in Escherichia coli by antisense DNA analogs // Antisense Res Dev., 1991, Winter, - 1. 4. - P. 19-27).
Наиболее близким аналогом настоящего изобретения является международная заявка WO 9814567 "Methods and Composition for inhibition bacterial growth", в которой используют 3''-последовательности 16S рРНК и предлагают комплементарные последовательности Шайн-Далгарно олигонуклеотиды как средство для подавления синтеза белков у микроорганизмов и соответственно подавления жизнедеятельности возбудителей бактериальных инфекций.
Наиболее существенным недостатком этого способа является неспецифичность действия предлагаемых композиций, используемых для подавления бактериального роста. Олигонуклеотиды, комплементарные последовательности Шайн-Далгарно. присущей всем прокариотическим микроорганизмам, вследствие консервативной природы этой последовательности скорее всего будут подавлять рост как вредной, так и полезной (резидентной) микрофлоры. Область применения таких препаратов весьма ограничена.
Задачей данного изобретения является создание олигонуклеотилов, позволяющих селективно ингибировать рост патогенных микроорганизмов.
Использование рРНК в качестве мишеней для олигонуклеотидных антимикробных препаратов является привлекательным еще и тем, что несмотря на консерватизм их молекул, в их составе есть менее консервативные (полуконсервативные и вариабельные) участки, в состав которых входят специфичные для семейств, родов, видов, или даже штаммов микроорганизмов так называемые "сигнатурные" (=подпись, рецепт) последовательности.
Сигнатурным последовательностям 16S рРНК (а, естественно, и антисигнатурным олигонуклеотидам) присущ определенный (заданный) уровень специфичности: видовой, родовой, семейный. Используя это качество, можно создать комплементарные конкретным сигнатурным последовательностям олигонуклеотиды: антивидовые, которые будут подавлять рост только одного вида микроорганизма, не принося вреда другим видам, даже если они относятся к одному роду: антиродовые, которые будут подавлять рост всех представителей данного рода; антисемейные, которые направлены против всех представителей семейства, и т.д. Такие олигонуклеотиды не будут подавлять рост микроорганизмов, если последние не находятся в определенных родственных отношениях с теми микроорганизмами, против которых данные олигонуклеотиды были разработаны.
Описанные в аналоге олигонуклеотиды не нарушают процесс самосборки рибосом, а предотвращают трансляцию информационных РНК на рибосомах и синтез белка путем блокирования прикрепления этих молекул к 3''- концевым последовательностям 16SpPHK.
Антисигнатурные олигонуклеотиды, описанные в настоящем изобретении, комплементарные (в зависимости от потребности) видо-, родо-, или семейственно-специфическим "сигнатурным" последовательностям, при взаимодействии с последними как с мишенями предотвращают посадку на рРНК рибосомальных "сердцевинных" белков, с которых начинается самосборка рибосом, чем полностью блокируют этот процесс, тотально прекращая синтез белка в микробной клетке.
Исходя из уникальных свойств сигнатурных участков рРНК мы впервые предлагаем их в качестве мишеней для селективного контроля жизнедеятельности микроорганизмов при помощи комплементарных к ним антисигнатурных синтетических олигодезоксирибонуклеотидов (АСО).
Поставленная задача решается олигонуклеотидами, предлагаемыми для подавления жизнедеятельности микроорганизмов, комплементарными сигнатурной последовательности рРНК целевого микроорганизма.
В качестве мишеней для блокирования комплементарными им олигонуклеотидами, вызывающими ингибирование синтеза белка у микроорганизма. предлагаются сигнатурные последовательности рРНК микроорганизмов.
Предлагается способ подавления жизнедеятельности целевых микроорганизмов (в том числе патогенных, в частности относящихся к классу молликутов (Mollicutes - класс мелких прокариотических микроорганизмов, не образующих клеточной стенки и являющихся паразитами и свободноживущими сапрофитами), основанный на выявлении сигнатурных последовательностей рРНК, характерных для одного или нескольких видов, семейства или рода микроорганизмов, синтезе олигонуклеотида или олигонуклеотидов, комплементарных выявленной сигнатурной последовательности рРНК микроорганизмов и обеспечении взаимодействия сигнатурной последовательности рРНК и комплементарного ему олигонуклеотида или олигонуклеотидов.
Предпочтительная форма использования олигонуклеотидов - в виде тиофосфатных и дитиофосфатных производных, устойчивых к нуклеазному расщеплению и способных к АТФ-зависимому проникновению в клетку.
Для введения предлагаемых олигонуклеотидов в организм млекопитающего используют приемлемое средство доставки. На основе описанных олигонуклеотидов и фармакологически приемлемого носителя предлагается фармацевтический препарат.
Такие препараты подавляют инфекции, вызываемые в том числе резистентными к традиционным антимикробным препаратам микроорганизмами.
Изобретение иллюстрируется следующими примерами.
Пример 1. Выявление сигнатурных последовательностей 16SpPHK.
Основную проблему в данном изобретении составлял поиск именно сигнатурных последовательностей 16S рРНК. Для того чтобы идентифицировать сигнатурные последовательности, получали препараты меченой [32Р] 16S рРНК микроорганизмов. Для этого штаммы микроорганизмов выращивали на оптимальных для конкретных видов средах. Бактерии выращивали на синтетических средах бeз фосфора. Молликуты, растущие только на обогащенных органическими добавками (гидролизат дрожжей, триптический перевар мышцы бычьего сердца, сыворотка крови лошадей и т.п.) средах выращивали на специально созданной для этого класса микроорганизмов среде СМИМВ-72 (Скрипаль И.Г., Малиновская Л.П. Микробиологический журнал, 1984, - 46, 2. С. 71-75), которая в настоящее время является общепринятой среди специалистов. Среду предварительно дефосфорилировали по методу Кимбала и др. (Nucleic Acids Res., 1974, v. 1, Р. 1721-1732). Дефосфорилированную среду СМИМВ-72 для молликутов и синтетические среды для бактерий обогащали аденозином, гуанозином, цитидином, уридином (каждого нуклеозида по 20 мг на 1 л среды) и тимидином (10 мг на 1 л). Приготовленные таким способом питательные среды разливали по 300 мл в колбы, засевали соответствующим видом микроорганизмов и выращивали в условиях, оптимальных для испытуемого вида организма (температура, аэрация, освещенность). На ранней стадии лаг-фазы роста в среду вносили [32РО4] из расчета 100 мк Сi/мл (1,0 Ci=3,7•1010 беккерелей). Микробные клетки собирали при помощи центрифугирования, отмывали от среды водным раствором 0,24 M NaCl и разрушали общепринятыми способами. Меченую 16S рРНК экстрагировали фенольным методом, выделяли при помощи электрофореза в полиакриламидном геле и проводили доочистку на колонке наполненной целлюлозой CF-11 (Whatman), как это описано в руководстве (J. Sambrook, E.F. Fritsch, T. Maniatis. Molecular cloning (A laboratory manual). Second Edition, Cold Spring Harbor Laboratory Press, 1989, P. 582). Каждый полученный таким образом препарат меченной [32Р] и очищенной 16S рРНК расщепляли рибонуклеазой Т, методом, описанным в вышеупомянутом руководстве. Рибонуклеаза Т1 специфически воздействует на 3''-фосфатные группы гуанозиновых нуклеотидов и расщепляет 5''-фосфатную связь с соседним нуклеотидом. Конечными продуктами реакции являются гуанозин-3''-фосфаты и олигонуклеотиды с концевыми гуанозин-3''-фосфатными группами.
Эти продукты разделяли, как описано в выше упомянутом руководстве (Molecular cloning. - см. выше) и сиквенировали при помощи двунаправленного электрофореза по методу Сангера и соавт (J. Mol. Biol., 1965, v.13, P.373-398) в модификации Вуза и соавт. (J. Mol. Evol., 1976, v. 7, P. 197-213).
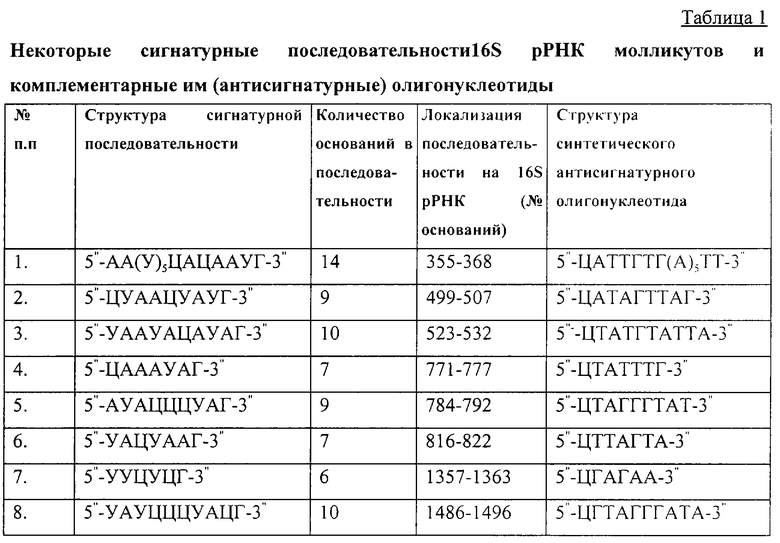
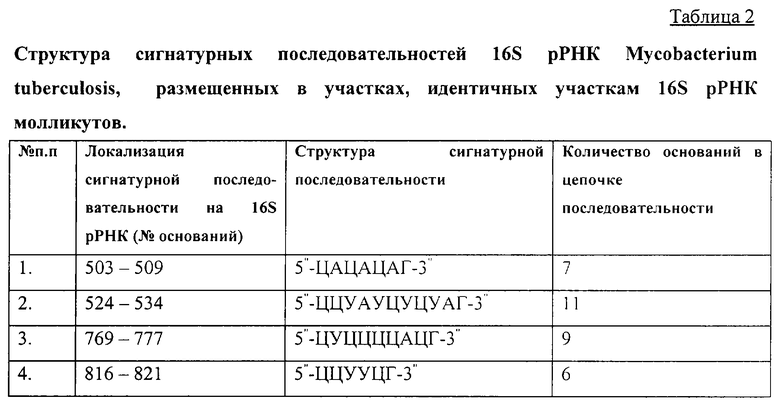
Полученные данные о составе и порядке нуклеотидов в цепи каждого из олигонуклеотидов, составляющих молекулу 16S рРНК изучаемых микроорганизмов и оканчивающихся гуанилиновой кислотой на 3''-конце, были сравнены с каталогами олигонуклеотидов 16S рРНК других микроорганизмов (банки данных Национального центра биотехнологической информации США и Европейской молекулярно-биологической лаборатории). В результате были установлены сигнатурные последовательности в составе 16S рРНК бактерий Escherichia coli. Bacillus subtilis. Agrobacterium tumefaciens и ряда штаммов разных видов молликутов (микоплазм). Наиболее значимыми для молликутов являются последовательности, перечень которых представлен в таблице 1.
Из международного банка данных (http://www.genome.ad.jp) нами была получена информация о структуре рибосомального оперона Mycobacterium tuberculosis. На основании этих данных был сформирован каталог из 97 олигонуклеотидов, 23 из которых проявили разный уровень сигнатурности (семейный, родовой или видовой). Видоспецифичные сигнатурные последовательности вида Mycobacterium tuberculosis имели уникальную структуру (таблица 2). Последовательности, размещенные в участках 16S рРНК, взаимодействующих с сердцевинными белками, имели структуру, отличающуюся от структуры аналогичных последовательностей, находящихся в этих участках 16S рРНК у других микроорганизмов, в том числе и у молликутов (таблица 1).
Из приведенных примеров видно, насколько уникальными для микроорганизмов являются сигнатурные последовательности их 16S рРНК и, естественно, настолько же специфическим, целенаправленным будет действие антисигнатурных олигонуклеотидов: только на конкретный вид микроорганизмов или группу родственных видов микроорганизмов, не нанося урона полезной и резидентной микрофлоре.
Пример 2. Синтез антисигнатурных олигонуклеотидов.
Синтез комплементарных сигнатурным последовательностям олигодезоксирибонуклеотидов (антисигнатурных олигонуклеотидов) осуществляли различными описанными в литературе методами. Антисигнатурные олигонуклеотиды были синтезированы в виде фосфодиэфиров (нормальной ДНК), тио-, и дитиофосфатных и метилофосфатных аналогов олигодезоксирибонуклеотидов.
Синтез природных олигодезоксирибонуклеотидов осуществляли твердофазным Н-фосфатным методом (Nucl. Acid. Res. - 1986. - 14, 13. - P. 5399-5407), синтез тио- и дитиофосфатных аналогов олигонуклеотидов блочным методом в растворе по модифицированной триэфирной методике (Tetrahedron lett. - 1990. - 31 (14). - Р. 1953-1956; Биоорг. химия. - 1990. - 16( 7). - С. 1574-1576; там же - 1992. - 18(4). - С. 550-553) с использованием мономеров, защищенных по межнуклеотидным фосфатам и аминогруппам экзоциклических оснований.
После удаления всех защитных групп целевой тиофосфат выделяли ионообменной хромотографией, а дитиофосфат - препаративным гель-электрофорезом в 20% ПААГ, содержащим 7М мочевину.
Синтез метилофосфатных аналогов олигонуклеотидов осуществляли согласно описанному методу (Биоорг. химия. - 1989. - 15 (2). - С. 267-276).
Пример 3. Характеристики антисигнатурных олигонуклеотидов.
Разновидности антисигнатурных синтетических олигонуклеотидов (АСО) были испытаны на следующие характеристики: устойчивость к естественной нуклеазной активности, способность к проникновению в микробную клетку и активизации в ней РНКазы Н, которая вырезает в клетке из нуклеиновых кислот гетеродуплексы (ДНК + РНК), образуемые в том числе и АСО с сигнатурными последовательностями на 16S рРНК.
Устойчивость к действию нуклеаз у АСО изучали в естественных условиях: при действии на них живых клеток молликутов, которые, как известно, обладают мощными комплексами неспецифических РНКаз (Микробиол. журн., 1991. - 53 (5). - С. 26-30; там же, 1992. - 54 (1). - С. 58-61; там же, 1993. - 55 (5). - С. 25-30) и ДНКаз (Микробиол. журн. - 1993. - 55 (1). - С. 12-19; там же, 1993. - 55 (3). - С. 17-24), которые функционируют как внутриклеточно, так и внеклеточно.
На примере АСО, комплементарных 355-368 последовательностям 16S рРНК, приведены результаты изучения устойчивости к нуклеазному расщеплению, для чего были использованы меченые АСО. Так как большинству микроорганизмов свойственна фосфатазная активность, то для защиты от фосфатаз 5''-концевого меченого фосфата АСО были синтезированы в виде 5''-бензиламидных фосфодиэфиров метилфосфатов, тио- и дитиофосфатов (Биоорган. химия. - 1986. - 12 4. - С. 475-481).
Реагент:
1. 5''-BnzNHp*CATTGTG(A)5TTp-3''
2. 5''-ВnzNНр*САТТGТG(А)5ТТрSН3-3''
3. 5''-BnzNHp*CpsApsTpsTpsGpsTpsGps(Aps)5TpsTpSCH3-3''
4. 5''-BnzNHp*CpssApssTpssTpssGpssTpssGpss(Apss)5TpssTpSCH3-3''
где р*-[32Р] - фосфат; BnzNH - CH2C6HS; С - цитозин; А - аденозин; Т - тимидин; G - гуанозин;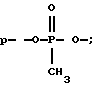
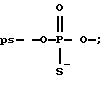
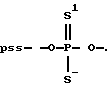
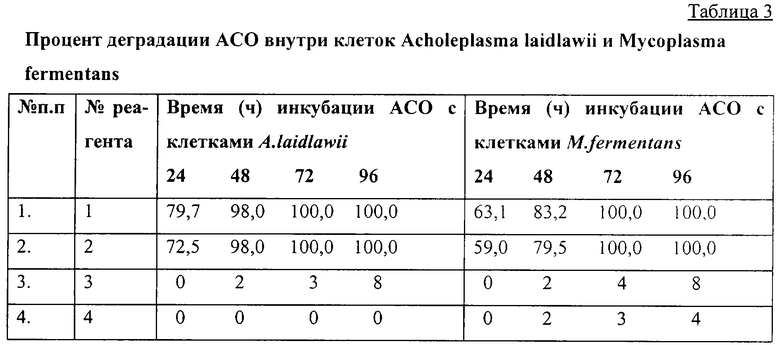
Из приведенных в таблице 3 данных видно, что тио-, дитиофосфатные аналоги АСО достаточно устойчивы к действию нуклеаз. Они оставались стабильными на протяжении 96 часов (длительность опыта).
Кроме того, как было установлено отдельными опытами, тио- и дитиофосфатные аналоги АСО, имея отрицательно заряженный рибулозофосфатный остов, целенаправленно взаимодействуют с клетками микроорганизмов и за счет активного АТФ-зависимого транспорта легко перемещались извне во внутрь их. Это не было свойственно ни фосфодиэфирам, ни метилофосфатам.
Благодаря этому механизму тио- и дитиофосфатные аналоги АСО накапливались в клетках микроорганизмов в концентрациях, в 104-105 раз превышающих их концентрацию вне клетки.
Таким образом, тио- и дитиофосфатные аналоги АСО при применении даже в наномолярных концентрациях являются эффективным средством подавления жизнедеятельности микроорганизмов благодаря свойству активно проникать в их клетки и накапливаться там.
Пример 4. Оценка антимикоплазменной активности антисигнатурных олигонуклеотидов.
Для подавления роста микроорганизмов применяли АСО в виде тиоаналогов (без введения метки), комплементарных к последовательностям 499-507 и 524-533 на 16S рРНК молликутов:
5''-CpsApsTpsApsGpsTpsTpsApsGp-3''
5''-CpsTpsApsTpsGpsTpsApsTpsTpsAp-3''
и дитиоаналогов:
5''-CpssApssTpssApssGpssTpssTpssApssGp-3''
5''-CpssTpssApssTpssGpssTpssApssTpssTpssAp-3''
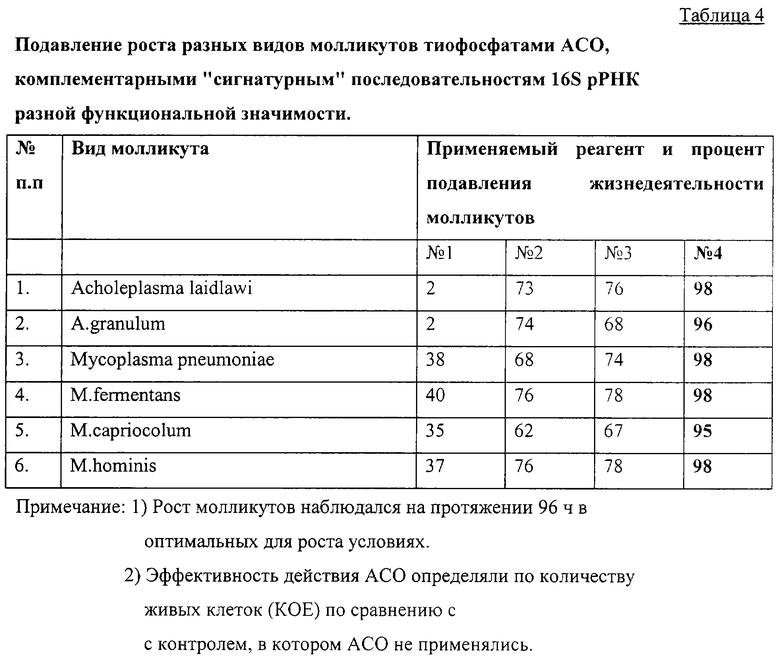
На удачный выбор АСО, применяемого для подавления роста микроорганизмов, влияет в первую очередь значимость последовательности(тей), которые они должны блокировать. Наиболее функционально значимыми последовательностями 16S рРНК являются те, с которыми взаимодействуют "сердцевинные" белки. Например, последовательности 499-507 и 523-532 (табл.1), с которыми взаимодействует белок S4 - главный из "сердцевинных" белков.
Для сравнения в таблице 4 приведены данные о подавлении роста молликутов тиоаналогами АСО, комплементарными последовательностям 499-507 и 523-532 на 16S рРНК и последовательности 355-368, с которой не взаимодействуют "сердцевинные" белки. АСО к этой последовательности тоже была синтезирована в виде тиофосфата. В таблице АСО к последовательности 355-368 приведен как реагент 1; к последовательности 499-507 - как реагент 2; к последовательности 523-532 - как реагент 3; смесь реагентов 2 и 3 (каждого по 50 нм) - как реагент 4. Концентрация каждого АСО в питательной среде составляла 100 нм.
Дитиофосфатные аналоги АСО по эффективности ингибиривания роста молликутов оказались идентичными с тиофосфатными аналогами.
То, что ни в одном случае не было достигнуто 100% подавления роста молликутов, объясняется тем, что молликуты растут только на тех средах, в состав которых входят нуклеиновые кислоты. В среде СМ ИМВ-72, на которой проводились опыты, присутствует как минимум 1 мг/мл экзогенных нуклеиновых кислот, которые, естественно, конкурируют с АСО в процессе их усвоения тестируемыми микроорганизмами, снижая эффективность этих препаратов.
Кроме того, АСО взаимодействует только с синтезированной "de novo" 16S рРНК, и эта кислота недостижима для них в уже собранных рибосомах. Клетки инокулята содержат готовые рибосомы и, естественно, 96 ч недостаточно для того, чтобы эти рибосомы перестали полностью существовать. После 96 ч живые клетки молликутов с такими рибосомами впадают в анабиоз и "оживают" лишь при высеве на свежую среду, что учитывается при подсчете колоний, которые образуются оставшимися в живых клетками (колониеобразующие единицы - КОЕ). Это объясняет невозможность достижения 100% подавления роста молликутов в таких опытах.
Совместное применение двух коротких антисигнатурных олигонуклеотидов (реагент 4), комплементарных последовательностям 499-507 и 523-532 168 рРНК обеспечивало почти 100% подавление жизнедеятельности молликутов (таблица 4).
Высокая эффективность тио- и дитиофосфатных аналогов АСО в подавлении жизнедеятельности микроорганизмов, например молликутов, определяется не только их уникальным механизмом действия, (блокируют на 16S рРНК места посадки белков и самосборку 30S субъединицы рибосомы), а и тем, что они активируют рибонуклеазу Н атакуемого микроорганизма. Последняя разрушает гетеродуплексы, образованные АСО с их сигнатурными последовательностями на 16S рРНК, и расщепляет в этих местах участки 16S рРНК, комплементарно связывающиеся с АСО. Вследствие этот 16S рРНК делится на фрагменты, которые подвергаются дальнейшему нуклеазному расщеплению, а количество собранных рибосом в клетке снижается. Освободившиеся АСО после расщепления РНК-ДНК дуплексов готовы для атаки своей комплементарной мишени на новосинтезированной 16S рРНК уже во время транскрипции рибосомного оперона.
Так как каждый из изученных олигонуклеотидов, примененных индивидуально, подавлял процесс трансляции в клетках молликутов на 100% при ингибировании жизнедеятельности этих микроорганизмов до 80%, то в условиях длительного нахождения АСО в опыте (например - культуре клеток тканей) они способны обеспечить 100% ингибирование жизнедеятельности микроорганизмов.
Из данных, свидетельствующих о том, что концентрация АСО в клетке может в 104-105 раз превышать внеклеточную, нами вычислено, что каждая из клеток молликутов поглощает примерно 500000 молекул АСО, из которых более 200000 молекул уже за 2 ч их контакта с клетками образовывали внутри последних дуплексы с комплементарными им СП на 16S рРНК. Такого количества молекул АСО внутри клеток молликутов вполне достаточно, чтобы "выключить" все находящиеся в клетке молекулы 16S рРНК из процесса самосборки 30S субъединицы рибосомы и полностью прекратить в ней процесс синтеза белка.
Молликуты от других микроорганизмов отличаются тем, что интенсивно поглощают АСО и другие нуклеиновые кислоты и объясняется это их неспособностью к самостоятельному синтезу пуринов и пиримидинов. Молликуты нуждаются в экзогенных источниках нуклеиновых кислот, которые они, обладая мощным набором внеклеточных и внутриклеточных ДНКаз и РНКаз, расщепляют до нуклеотидов для нужд синтеза собственных нуклеиновых кислот.
Это свойство молликутов облегчает решение проблемы доставки АСО в их клетки. Нами показана способность молликутов к поглощению синтетических олигодезоксирибонуклеотидов длиной в 24 основания. Сигнатурные последовательности имеют меньшую длину, соответственно молекулы АСО также будут достаточно короткими.
Но большинство бактерий в экзогенных источниках нуклеиновых кислот не нуждается и, естественно, без соответствующей модификации синтетические олигонуклеотиды в их клетки проникнуть не смогут. В настоящее время эта проблема решена путем присоединения к 3''- или 5''-концам олигонуклеотидов веществ - лигандов, по отношению к которым тот или иной микроорганизм проявляет ауксотрофность. Способы создания олигонуклеотидов с определенными лигандами по отношению к целевым микроорганизмам известны и описаны (N.V. Amirkhanow, V.V. Zarytova. Nucleosides Nycleotides, 1955, v. 14, P. 935-937; E. Rapaport et al., Proc. Natl. Acad. Sci, 1996, v. 93. P. 709-713).
Таким образом, результаты предлагаемого решения позволяют утверждать, что АСО, содержащие от 6 до 14 нуклеотидных звеньев, являются оптимальными по размеру молекулами, способными полностью подавлять развитие микроорганизмов, а для таких патогенных микроорганизмов, как молликуты, могут быть использованы в качестве антимикоплазменных препаратов для деконтаминации культур клеток.
Пример 5. Использование антисигнатурных олигонуклеотидов для деконтаминации клеточных культур.
Латентно микоплазменно инфицированная культура клеток Hela была деконтаминирована за три пересева. Каждый пересев осуществлялся на 4-й день роста культуры на среде 199, содержащей 100 нМ АСО в виде тиофосфатов (50 нМ 5''-CpsApsTpsApsGpsTpsTpsApsGps-3''+50 нМ 5''-CpsTpsApsTpsGpsTpsApsTpsTpsAps-3''). В конце третьего периода (12 день) в образцах культуры взятой для анализа при окрашивании краской 4-6-диамидино-3-фенилиндол (ДАПИ), которая взаимодействует только с прокариотической ДНК, клеток молликутов-контаминантов выявлено не было. Об этом свидетельствовало и повышение жизнедеятельности культуры: до обработки она не могла быть жизнеспособной больше 8 суток, а после обработки - 14 суток без замены среды. С заменой среды жизнеспособность деконтаминированных культур увеличивалась до 24-30 дней, а в инфицированных она не превышала 12-14 дней.
Из-за универсальности действия указанных выше тиофосфатов (они одинаково эффективно действуют на молликуты родов Acholeplasma и Mycoplasma - основные контаминанты культур клеток) они могут быть использованы для деконтаминации и других культур клеток от микоплазменной инфекции.
Еще одним позитивным качеством АСО является присущая олигонуклеотидам вообще почти полная безвредность (нетоксичность) (Morris M.С., Vidal P., Chaloin L., Heitz F., Divita G. (1997). Nucl. Acids. Res. 25, 2730-2736; Pichon C. , Freulon I., Midoux P., Mayer R., Monsigny M., Roche A.C. (1997). Antisense Nucleic Acid Drug Dev. 7, 335-343).
Описанные свойства АСО позволяют рекомендовать их в качестве действующего вещества фармацевтических препаратов для терапии ряда трудноизлечиваемых заболеваний человека и животных: микоплазмозов, хламидиозов, пневмоний, туберкулеза и пр.
При этом можно получить различные лекарственные формы, используя подходящие носители или растворители (вода, физраствор, этиловый спирт, борная кислота).
Таким образом, предлагаемые нами антисигнатурные олигонуклеотиды в отличие от известных антисуперконсервативных олигонуклеотидов являются суперспецифическими терапевтическими средствами, направленными на подавление жизнедеятельности определенных видов микроорганизмов путем адресного блокирования уникальных для каждого вида (или рода, или семейства и т.д.) последовательностей на рибосомальных РНК, вследствие чего предотвращается самосборка рибосом и полностью подавляется синтез белка в микробной клетке.
Изобретение относится к биологии и медицине, конкретно к антисенстерапии с использованием антисмысловых олигонуклеотидов. Синтезированы олигонуклеотиды для подавления жизнедеятельности патогенных микроорганизмов, в частности относящихся к классу молликутов. Олигонуклеотиды являются комплементарными сигнатурной последовательности рРНК целевого микроорганизма и включают 6 - 14 оснований в нуклеотидной последовательности. При введении в организм млекопитающего антисигнатурные олигонуклеотиды осуществляют адресное блокирование уникальных для каждого вида (рода, семейства) последовательностей на рибосомальных РНК, вследствие чего предотвращается самосборка рибосом и полностью подавляется синтез белка в микробной клетке. 2 с. и 3 з.п. ф-лы, 4 табл.
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| ОЛИГОНУКЛЕОТИДЫ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1995 |
|
RU2112766C1 |
| US 6110681 А, 29.08.2000 | |||
| US 5981280 А, 09.11.1999 | |||
| Способ определения пористости горючей массы угля | 2023 |
|
RU2811321C1 |
| РЕЗИСТЕНТНЫЕ К НУКЛЕАЗЕ ОЛИГОНУКЛЕОЗИДЫ, СПОСОБ ИХ ПОЛУЧЕНИЯ И РЕЗИСТЕНТНЫЙ К НУКЛЕАЗЕ НУКЛЕОЗИДНЫЙ ДИМЕР | 1991 |
|
RU2131436C1 |
| ОЛИГОНУКЛЕОТИДЫ И СПОСОБ ИХ ПОЛУЧЕНИЯ | 1991 |
|
RU2088588C1 |

