Родственные заявки
В настоящей заявке испрашивается приоритет ранее поданной предварительной заявки на патент США рег.№60/141031, поданной 25 июня 1999, предварительной заявки на патент США рег. №60/142692, поданной 1 июля 1999, а также предварительной заявки на патент США рег. №60/151214, поданной 27 августа 1999. В этой заявке также испрашивается приоритет заявки на патент Германии №19930429.7, поданной 1 июля 1999, заявки на патент Германии №19931413.6, поданной 8 июля 1999, заявки на патент Германии №19931457.8, поданной 8 июля 1999, заявки на патент Германии №19931541.8, поданной 8 июля 1999, заявки на патент Германии №19932209.0, поданной 9 июля 1999, заявки на патент Германии №19932230.9, поданной 9 июля 1999, заявки на патент Германии №19932914.1, поданной 14 июля 1999, заявки на патент Германии №19940764.9, поданной 27 августа 1999, и заявки на патент Германии №19941382.7, поданной 31 августа 1999. Полное содержание всех вышеупомянутых заявок во всей своей полноте вводится в настоящее описание посредством ссылки.
Предшествующий уровень техники
Некоторые продукты и побочные продукты природных процессов метаболизма в клетках широко используются в различных областях промышленности, включая пищевую, комбикормовую, косметическую и фармацевтическую промышленность. Такими молекулами, имеющими общее название «химические продукты тонкого органического синтеза», являются органические кислоты, протеиногенные и непротеиногенные аминокислоты, нуклеотиды и нуклеозиды, липиды и жирные кислоты, диолы, углеводы, ароматические соединения, витамины и кофакторы, и ферменты. Их получение обычно осуществляют путем широкомасштабного культивирования бактерий, выращиваемых для продуцирования и секреции больших количеств конкретной нужной молекулы. Одним из ценных микроорганизмов, используемых в этих целях, является Corynebactecium glutamicum, грамположительная непатогенная бактерия. Посредством отбора штаммов был получен ряд мутантных штаммов, продуцирующих ряд нужных соединений. Однако отбор штаммов с повышенным продуцированием конкретной молекулы является длительным и трудоемким процессом.
Краткое описание изобретения
Настоящее изобретение относится к новым бактериальным молекулам нуклеиновой кислоты, имеющим широкое применение. Такими применениями являются идентификация микроорганизмов, которые могут быть использованы для продуцирования химических продуктов тонкого органического синтеза, модуляция продуцирования химических продуктов тонкого органического синтеза в С. glutamicum или родственных бактериях, типирование или идентификация С. glutamicum или родственных бактерий для использования в качестве эталонных участков для картирования генома С. glutamicum, или в качестве маркеров для трансформации. Эти новые молекулы нуклеиновой кислоты кодируют белки, называемые в данном описании белками резистентности и толерантности к стрессам (SRT).
С. glutamicum представляет собой грамположительную аэробную бактерию, которая широко используется в промышленности для широкомасштабного продуцирования ряда химических продуктов тонкого органического синтеза, а также для деградации углеводородов (например, при утечке нефти) и для окисления терпеноидов. Поэтому, молекулы нуклеиновой кислоты для SRT настоящего изобретения могут быть использованы для идентификации микроорганизмов, которые могут быть использованы для продуцирования химических продуктов тонкого органического синтеза, например, методом ферментации. Модуляция экспрессии нуклеиновых кислот, кодирующих SRT настоящего изобретения, или модификация данной последовательности молекул нуклеиновой кислоты, кодирующих SRT настоящего изобретения, могут быть осуществлены в целях модуляции продуцирования одного или нескольких химических продуктов тонкого органического синтеза из микроорганизма (например, для повышения выхода или продуцирования одного или нескольких химических продуктов тонкого органического синтеза из бактерий видов Corynebacterium glutamicum или Brevibacterium).
Нуклеиновые кислоты SRT настоящего изобретения могут быть также использованы для идентификации микроорганизма Corynebacterium glutamicum или близкородственного микроорганизма, либо для обнаружения присутствия С. glutamicum или его родственной бактерии в смешанной популяции микроорганизмов. Настоящее изобретение относится к нуклеотидным последовательностям ряда генов С. glutamicum; при этом, присутствие этого микроорганизма может быть быть установлено путем зондирования экстрагированной геномной ДНК культуры одиночной или смешанной популяции микроорганизмов в жестких условиях зондом, охватывающим область гена С. glutamicum, который является уникальным для этого микроорганизма. Хотя Corynebacterium glutamicum сам по себе является непатогенным, однако, он является родственным патогенному виду у человека, такому как Corynebacterium diphtheriae (патоген, вызывающий дифтерию); причем, детекция таких микроорганизмов имеет большое значение в медицине.
Молекулы нуклеиновых кислот, кодирующие SRT настоящего изобретения, могут также служить в качестве исходных участков для картирования генома С. glutamicum или геномов родственных микроорганизмов. Аналогичным образом, эти молекулы или их варианты или части могут служить в качестве маркеров для генетически сконструированных видов Corynebacterium или Brevibacterium.
Белки SRT, кодируемые новыми молекулами нуклеиновой кислоты настоящего изобретения, например, позволяют С. glutamicum выживать в условиях химического стресса или воздействия окружающей среды, опасных для данного микроорганизма. Если принять во внимание доступность клонирующих векторов для использования в Corynebacterium glutamicum, таких как векторы, описанные Sinskey и др. в патенте США №4649119, и способов генетической манипуляции с бактериями С. glutamicum и родственных видов Brevibacterium (например, lactofermentum} (Yoshihama et al., J.Bacteriol. 162: 591-597 (1985); Katsumata et al., J.Bacteriol. 159: 306-311 (1984) & Santamaria et al., J.Gen.Microbiol. 130: 2237-2246 (1984)), молекулы нуклеиновой кислоты настоящего изобретения можно использовать для генетического конструирования этого микроорганизма в целях получения из него лучшего или более эффективного продуцента одного или нескольких химических продуктов тонкого органического синтеза благодаря способности этих белков стимулировать рост и размножение С. glutamicum (а также непрерывное продуцирование одного или нескольких химических продуктов тонкого органического синтеза) в условиях, которые обычно препятствуют росту данного микроорганизма, таких как условия, часто встречающиеся в процессе крупномасштабного ферментативного роста. Так, например, путем сверхэкспрессии или конструирования молекулы, индуцируемой тепловым шоком протеазы, для оптимизации ее активности, может быть увеличена способность данной бактерии к разложению неправильно уложенных белков, в случае, когда данная бактерия подвергается действию высоких температур. При снижении количества неправильно уложенных (и возможно неправильно регулируемых или нефункциональных) белков, которые препятствуют осуществлению нормальных реакционных механизмов в клетке, способность этой клетки нормально функционировать в такой культуре повышается, что, в свою очередь, должно повышать ее жизнеспособность. Это общее увеличение числа клеток, обладающих большей жизнеспособностью и активностью в данной культуре, должно также приводить к увеличению выхода, продуцирования и/или эффективности продуцирования одного или нескольких нужных химических продуктов тонкого органического синтеза, что обусловлено, по крайней мере, относительно большим числом клеток, продуцирующих эти химические вещества в данной культуре.
Настоящее изобретение также относится к новым молекулам нуклеиновой кислоты, кодирующей белки SRT, которые позволяют, например, С. glutamicum выживать в условиях химического стресса или воздействия окружающей среды, опасных для данного микроорганизма. Молекулы нуклеиновых кислот, кодирующие белок SRT, называются в настоящем описании молекулами нуклеиновых кислот SRT. В предпочтительном варианте осуществления изобретения, белок SRT участвует в путях метаболизма, позволяющих С. glutamicum выживать в условиях химического стресса или воздействия окружающей среды, опасных для данного микроорганизма. Примерами таких белков являются белки, кодируемые генами, представленными в Таблице 1.
В соответствии с одним из своих аспектов, настоящее изобретение относится к изолированным молекулам нуклеиновой кислоты (например, кДНК, ДНК или РНК), содержащим нуклеотидную последовательность, кодирующую белок SRT или его биологически активные части, а также к фрагментам нуклеиновой кислоты, используемым в качестве праймеров или зондов для гибридизации в целях детекции или амплификации SRT-кодирующей нуклеиновой кислоты (например, ДНК или мРНК). В особенно предпочтительных вариантах осуществления изобретения, изолированная молекула нуклеиновой кислоты содержит одну из нуклеотидных последовательностей, представленных нечетно пронумерованными SEQ ID NOs в Списке последовательностей (например, SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7...) или кодирующую область или комплемент одной из этих нуклеотидных последовательностей. В других особенно предпочтительных вариантах осуществления настоящего изобретения, изолированная молекула нуклеиновой кислоты настоящего изобретения включает нуклеотидную последовательность, которая гибридизуется с этой молекулой или которая, по крайней мере, приблизительно на 50%, а предпочтительно, по крайней мере, приблизительно на 60%, более предпочтительно, по крайней мере, приблизительно на 70%, 80% или 90%, и даже более предпочтительно, по крайней мере, приблизительно на 95%, 96%, 97%, 98%, 99% или более является гомологичной нуклеотидной последовательности, представленной нечетно пронумерованными SEQ ID Nos в Списке последовательностей (например, SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7...) или ее части. В других предпочтительных вариантах осуществления настоящего изобретения, изолированная молекула нуклеиновой кислоты кодирует одну из аминокислотных последовательностей, представленных четно пронумерованными SEQ ID NO: в Списке последовательностей (например, SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8...). Предпочтительными белками SRT настоящего изобретения также являются белки, которые, предпочтительно, обладают, по крайней мере, одной из SRT-активностей, описанных в данной заявке.
В другом варианте осуществления изобретения, изолированная молекула нуклеиновой кислоты кодирует белок или его часть, где указанный белок или его часть включает аминокислотную последовательность, которая является достаточно гомологичной аминокислотной последовательности настоящего изобретения (например, последовательности, имеющей четно пронумерованную SEQ ID NO: в Списке последовательностей), например, достаточно гомологичной аминокислотной последовательности настоящего изобретения, так, что данный белок или его часть сохраняют SRT-активность. Предпочтительно, чтобы этот белок или его часть, кодируемые данной молекулой нуклеиновой кислоты, сохраняли способность повышать выживаемость С. glutamicum в условиях химического стресса или воздействия окружающей среды, опасных для данного микроорганизма. В одном из вариантов осуществления изобретения, белок, кодируемый данной молекулой нуклеиновой кислоты, является, по крайней мере, примерно на 50%, а предпочтительно, по крайней мере, примерно на 60%, более предпочтительно, по крайней мере, примерно на 70%, 80% или 90%, а наиболее предпочтительно, по крайней мере, примерно на 95%, 96%, 97%, 98% или 99% или более гомологичным аминокислотной последовательности настоящего изобретения (например, всей аминокислотной последовательности, выбранной из четно пронумерованных последовательностей SEQ ID NO: в Списке последовательностей). В другом предпочтительном варианте осуществления изобретения, указанным белком является полноразмерный белок С. glutamicum, который является, в основном, гомологичным всей аминокислотной последовательности настоящего изобретения (кодируемой открытой рамкой считывания, показанной соответствующими нечетно пронумерованными SEQ ID NO: в Списке последовательностей (например, SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7...).
В другом предпочтительном варианте осуществления изобретения, изолированная молекула нуклеиновой кислоты происходит от С. glutamicum и кодирует белок (например, гибридный белок SRT), включающий биологически активный домен, который является, по крайней мере, приблизительно на 50% или более гомологичным одной из аминокислотных последовательностей настоящего изобретения (например, одной из четно пронумерованных последовательностей SEQ ID NO: в Списке последовательностей) и обладает способностью повышать выживаемость С. glutamicum в условиях химического стресса или воздействия окружающей среды, опасных для данного микроорганизма, или обладает одной или более активностями, указанными в Таблице 1, и указанная изолированная молекула также включает гетерологичные последовательности нуклеиновой кислоты, кодирующие гетерологичный полипептид или регуляторные области.
В другом варианте осуществления изобретения, указанная изолированная молекула нуклеиновой кислоты имеет длину, по крайней мере, в 15 нуклеодитов и гибридизуется в жестких условиях с молекулой нуклеиновой кислоты, включающей нуклеотидную последовательность настоящего изобретения (например, нечетно пронумерованную последовательность SEQ ID NO в Списке последовательностей). Предпочтительно, чтобы указанная изолированная молекула нуклеиновой кислоты соответствовала природной молекуле нуклеиновой кислоты. Более предпочтительно, чтобы изолированная нуклеиновая кислота кодировала природный белок SRT С. glutamicum или его биологически активную часть.
В другом своем аспекте, настоящее изобретение относится к векторам, например, к рекомбинантным экспрессирующим векторам, содержащим молекулу нуклеиновой кислоты настоящего изобретения, и к клеткам-хозяевам, в которые были введены указанные векторы. В одном из вариантов осуществления изобретения, указанная клетка-хозяин используется для продуцирования белка SRT путем ее культивирования в подходящей среде. Затем, указанный белок SRT может быть выделен из данной среды или клетки-хозяина.
В еще одном своем аспекте, настоящее изобретение относится к генетически модифицированному микроорганизму, в который был введен ген SRT или в котором он был модифицирован. В одном из вариантов осуществления изобретения, геном указанного микроорганизма был модифицирован путем введения молекулы нуклеиновой кислоты настоящего изобретения, кодирующей последовательность SRT дикого типа или мутированную последовательность SRT в качестве трансгена. В другом варианте осуществления изобретения, эндогенный ген SRT в геноме данного микроорганизма был модифицирован, например, функционально разрушен посредством гомологичной рекомбинации с модифицированным геном SRT. В другом варианте осуществления изобретения, эндогенный или введенный ген SRT в микроорганизме был модифицирован с помощью одной или нескольких точковых мутаций, делеций или инверсий, но, при этом, он кодирует функциональный белок SRT. В еще одном варианте осуществления изобретения, одна или несколько регуляторных областей (например, промотор, репрессор или индуктор) гена SRT в микроорганизме были модифицированы (например, посредством делеций, усечений, инверсии или точковой мутации), так, чтобы модулировалась экспрессия гена SRT. В предпочтительном варианте осуществления изобретения, данный микроорганизм принадлежит к роду Corynebacterium или Brevibacterium, при этом, особенно предпочтительным является Cocynebacterium glutamicum. В предпочтительном варианте осуществления изобретения, указанный микроорганизм также используют для продуцирования нужного соединения, такого как аминокислота, при этом, особенно предпочтительным является лизин.
В другом своем аспекте, настоящее изобретение относится к способу идентификации присутствия или активности Corynebacterium diphtheriae у индивидуума. Этот способ предусматривает обнаружение у индивидуума одной или нескольких нуклеотидных или аминокислотных последовательностей настоящего изобретения (например, последовательностей, представленных в Списке последовательностей как SEQ ID NO:1-304), и тем самым обнаружение у данного индивидуума присутствия или активности Corynebacterium diphtheriae.
В еще одном своем аспекте, настоящее изобретение относится к изолированному белку SRT или его части, например, к его биологически активной части. В предпочтительном варианте осуществления изобретения, изолированный белок SRT или его часть обладает способностью повышать выживаемость С. glutamicum в условиях химического стресса или воздействия окружающей среды, опасных для данного микроорганизма. В другом предпочтительном варианте осуществления изобретения, изолированный белок SRT или его часть являются достаточно гомологичными аминокислотной последовательности настоящего изобретения (например, четно пронумерованной последовательности SEQ ID NO: в Списке последовательностей), при этом, указанный белок или его часть сохраняют способность повышать выживаемость С. glutamicum в условиях воздействия химических веществ или окружающей среды, опасных для данного микроорганизма.
Настоящее изобретение также относится к изолированному препарату белка SRT. В предпочтительных вариантах осуществления изобретения, белок SRT включает аминокислотную последовательность настоящего изобретения (например, одну из четно пронумерованных последовательностей SEQ ID NO в Списке последовательностей). В другом предпочтительном варианте своего осуществления, настоящее изобретение относится к изолированному полноразмерному белку, который, в основном, гомологичен полной аминокислотной последовательности настоящего изобретения (например, последовательности SEQ ID NO, имеющей четный номер в Списке последовательностей) (кодированной открытой рамкой считывания в соответствующей SEQ ID NO, имеющей нечетный номер в Списке последовательностей). В еще одном варианте осуществления изобретения, данный белок, по крайней мере, примерно на 50%, предпочтительно, по крайней мере, примерно на 60%, более предпочтительно, по крайней мере, примерно на 70%, 80% или 90%, а наиболее предпочтительно, по крайней мере, примерно на 95%, 96%, 97%, 98% или 99% или более гомологичен полной аминокислотной последовательности настоящего изобретения (например, последовательности SEQ ID NO, имеющей четный номер в Списке последовательностей). В других вариантах осуществления изобретения, изолированный белок SRT содержит аминокислотную последовательность, которая, по крайней мере, примерно на 50% или более гомологична одной из аминокислотных последовательностей настоящего изобретения (например, последовательности SEQ ID NO, имеющей четный номер в Списке последовательностей), и которая способна повышать степень выживания С. glutamicum в условиях воздействия химических веществ или окружающей среды, опасных для данного микроорганизма, или обладает одной или более активностями, указанными в Таблице 1.
Альтернативно, изолированный белок SRT включает аминокислотную последовательность, кодируемую нуклеотидной последовательностью, которая гибридизуется, например в жестких условиях, с одной из последовательностей SEQ ID NO, пронумерованных четными цифрами в Списке последовательностей, или которая, по крайней мере, примерно на 50%, предпочтительно, по крайней мере, примерно на 60%, более предпочтительно, по крайней мере, примерно на 70%, 80% или 90%, а наиболее предпочтительно, по крайней мере, примерно на 95%, 96%, 97%, 98% или 99% или более гомологична указанной последовательности. Также предпочтительно, чтобы предпочтительные формы белков SRT также имели одну или несколько биологических SRT-активностей, описанных в данной заявке.
Полипептид SRT или его биологически активная часть могут быть функционально присоединены к полипептиду, не являющемуся SRT, с образованием гибридного белка. В предпочтительных вариантах осуществления изобретения, указанный гибридный белок обладает активностью, которая отличается от активности, присущей лишь одному белку SRT. В других предпочтительных вариантах осуществления изобретения, этот гибридный белок способствует повышению выходов, продуцирования и/или эффективности продуцирования нужного химического продукта тонкого органического синтеза из С. glutamicum. В особенно предпочтительных вариантах осуществления изобретения, интеграция указанного гибридного белка в клетку-хозяина способствует модуляции продуцирования нужного соединения из данной клетки.
В другом своем аспекте, настоящее изобретение относится к способам скрининга молекул, которые модулируют активность белка SRT, либо путем взаимодействия с самим белком, либо с его субстратом, или путем связывания с партнером белка SRT, либо путем модуляции транскрипции или трансляции молекулы нуклеиновой кислоты SRT настоящего изобретения.
В другом своем аспекте, настоящее изобретение относится к способу продуцирования химических продуктов тонкого органического синтеза. Этот способ предусматривает культивирование клетки, содержащей вектор, направляющий экспрессию молекулы нуклеиновой кислоты SRT настоящего изобретения, так, чтобы продуцировались химические продукты тонкого органического синтеза. В предпочтительном варианте осуществления изобретения, указанный способ, кроме того, включает стадию получения клетки, содержащей указанный вектор, где клетку трансфецируют вектором, направляющим экспрессию нуклеиновой кислоты SRT. В другом предпочтительном варианте осуществления изобретения, указанный способ, кроме того, включает стадию выделения химического продукта тонкого органического синтеза из данной культуры. В особенно предпочтительном варианте осуществления изобретения, указанную клетку получают от бактерий рода Corynebacterium или Brevibacterium, или выбирают из штаммов, указанных в таблице 3.
В другом своем аспекте, настоящее изобретение относится к способам модуляции продуцирования молекулы из микроорганизма. Такие способы предусматривают контактирование данной клетки с агентом, который модулирует активность белка SRT или экспрессию SRT-кодирующей нуклеиновой кислоты так, чтобы клеточно-ассоциированная активность была изменена по сравнению с той же самой активностью, но в отсутствие данного агента. В предпочтительном варианте осуществления изобретения, модулируют резистентность указанной клетки к одному или более токсических химических продуктов или резистентность к одному или нескольким воздействиям окружающей среды, так, чтобы это приводило к увеличению выхода или степени продуцирования данным микроорганизмом нужного химического продукта тонкого органического синтеза. Указанным агентом, модулирующим активность белка SRT, может быть агент, который стимулирует активность белка SRT или экспрессию нуклеиновой кислоты SRT. Примерами агентов, стимулирующих активность белка SRT или экспрессию нуклеиновой кислоты, кодирующей SRT, являются небольшие молекулы, активные белки SRT и нуклеиновые кислоты, кодирующие белки SRT, которые были введены в данную клетку. Примерами агентов, ингибирующих активность или экспрессию SRT, являются небольшие молекулы и антисмысловые молекулы нуклеиновых кислот SRT.
В другом своем аспекте, настоящее изобретение относится к способам модуляции выходов нужного соединения из клетки, предусматривающим введение гена SRT дикого типа или мутантного гена SRT в клетку, либо находящегося на отдельной плазмиде, либо интегрированного в геном клетки-хозяина. Если ген интегрируется в геном, то такая интеграция может происходить беспорядочно, либо она может осуществляться путем гомологичной рекомбинации, так, чтобы нативный ген заменялся введенной копией, что приводит к продуцированию нужного соединения из модулируемой клетки. В предпочтительном варианте осуществления изобретения, указанные выходы являются повышенными. В другом предпочтительном варианте осуществления изобретения указанным химическим веществом является химический продукт тонкого органического синтеза. В особенно предпочтительном варианте осуществления изобретения, указанным химическим продуктом тонкого органического синтеза является аминокислота. В особенно предпочтительных вариантах осуществления изобретения, указанной аминокислотой является L-лизин.
Подробное описание изобретения
Настоящее изобретение относится к молекулам нуклеиновой кислоты и белка SRT, которые способствуют выживанию С. glutamicum при воздействии на этот микроорганизм химических веществ или условий окружающей среды, опасных для данного микроорганизма. Молекулы настоящего изобретения могут быть использованы для модуляции продуцирования химических продуктов тонкого органического синтеза из микроорганизмов, поскольку эти белки SRT обеспечивают непрерывный рост и размножение С. glutamicum в присутствии токсических химических условий или неблагоприятных условий окружающей среды, таких, которые могут возникать в процессе крупномасштабного ферментативного культивирования. При увеличения скорости роста или, по крайней мере, при поддержании нормального роста в плохих, если не токсических, условиях, можно увеличить выход, продуцирование и/или эффективность продуцирования одного или нескольких химических продуктов тонкого органического синтеза из такой культуры, что обусловлено, по крайней мере, относительно более высоким числом клеток, продуцирующих указанный химический продукт тонкого органического синтеза в данной культуре. Аспекты настоящего изобретения будут более подробно описаны ниже.
I. Химические продукты тонкого органического синтеза
Термин «химический продукт тонкого органического синтеза» известен специалистам и означает молекулы, продуцируемые микроорганизмом, применяемым в различных отраслях промышленности, таких как, но не ограничивающихся ими, фармацевтическая промышленность, сельское хозяйство и косметическая промышленность. Такими соединениями являются органические кислоты, такие как винная кислота, итаконовая кислота и диаминопимелиновая кислота, протеиногенные и непротеиногенные аминокислоты, пуриновые и пиримидиновые основания, нуклеозиды и нуклеотиды (как описано например, в работе Kuninaka A. (1996) Nucleotides and related compounds, p.561-612, in Biotechnology vol.6, Rehm et al., eds. VCH: Weinheim и в приведенных в ней ссылках), липиды, насыщенные и ненасыщенные жирные кислоты (например, арахидоновая кислота), диолы (например, пропандиол и бутандиол), углеводы (например, гиалуроновая кислота и трегалоза), ароматические соединения (например, ароматические амины, ванилин и индиго), витамины и кофакторы (описанные в Ullmann's Encyclopedia of Industrial Chemistry, vol.A27, «Vitamins», p.443-613 (1996) VCH: Weinheim и в имеющихся там ссылках; и Ong, A.S., Niki E. & Packer, L. (1995) «Nutrition, Lipids, Health and Disease» Proceedings of the UNESCO/Confederation of Scientific and Technological Associations in Malaysia and the Society for Free Radical Research - Asia, held Sept. 1-3, 1994 at Penang, Malaysia, AOCS Press, (1995)), ферменты, поликетиды (Cane et al. (1998) Science 282: 63-68) и другие химические соединения, описанные Gutcho (1983) Chemicals by Fermentation, Noyes Data Corporation, ISBN: 0818805086 и в имеющихся там ссылках. Метаболизм и использование некоторых из этих химических продуктов тонкого органического синтеза будут более подробно описаны ниже.
А. Метаболизм и использование аминокислот
Аминокислоты включают основные структурные звенья всех белков и, таким образом, являются необходимыми для нормального функционирования всех организмов. Термин "аминокислота" хорошо известен специалистам. Протеиногенные аминокислоты, всего 20 видов, служат в качестве структурных единиц белка, в котором они соединены пептидными связями, а непротеиногенные аминокислоты (несколько сотен из которых являются известными) обычно не встречаются в белках (см. Ullmann's Encyclopedia of Industrial Chemistry, vol.A2, p.57-97 VCH: Weinheim (1985)). Аминокислоты могут иметь оптическую D- или L-конфигурацию, хотя L-аминокислоты обычно являются единственным типом, обнаруживаемым в природных белках. Пути биосинтеза и деградации каждой из указанных 20 протеиногенных аминокислот были охарактеризованы в прокариотических и эукариотических клетках (см., например, Stryer, L. Biochemistry, 3rd edition, pages 578-590 (1988)). «Незаменимые» аминокислоты (гистидин, изолейцин, лейцин, лизин, метионин, фенилаланин, треонин, триптофан и валин), называемые так потому, что они, в основном, удовлетворяют требованиям, предъявленным к питанию, что обусловлено сложностью их биосинтеза, легко превращаются в ходе метаболизма путем простого биосинтеза в остальные 11 «заменимых» аминокислот (аланин, аргинин, аспарагин, аспартат, цистеин, глутамат, глутамин, глицин, пролин, серии и тирозин). Высшие животные сохраняют способность синтезировть некоторые из этих аминокислот, но незаменимые аминокислоты должны поступать с пищей для нормального синтеза белка в организме.
Помимо своей функции в биосинтезе белка, эти аминокислоты, сами по себе, представляют интерес как химические соединения, и многие из них, как известно, находят свое применение в пищевой, комбикормовой, химической, косметической, сельскохозяйственной и фармацевтической промышленности. Лизин является важной аминокислотой для питания не только человека, но также и животных с однокамерным желудком, таким как домашняя птица и свиньи. Глутамат является наиболее широко используемым соединением в качестве ароматизирующих добавок (моно-глутамат натрия, МГН) и широко применяется в пищевой промышленности как и аспартат, фенилаланин, глицин и цистеин. Глицин, L-метионин и триптофан используются в фармацевтической промышленности. Глутамин, валин, лейцин, изолейцин, гистидин, аргинин, пролин, серин и аланин используются как в фармацевтической, так и в косметической промышленности. Треонин, триптофан и D/L-метионин являются широко распространенными пищевыми добавками (Leuchtenberger, W. (1996) Amino aids-technical production and use, p. 466-502, Rehm et al. (eds.) Biotechnology vol.6, chapter 14a, VCH: Weimheim). Кроме того, было установлено, что указанные аминокислоты могут быть использованы в качестве предшественников для синтеза синтетических аминокислот и белков, таких как N-ацетилцистеин, S-карбоксиметил-L-цистеин, (S)-5-гидрокситриптофан и др., описанные в Ullmann's Encyclopedia of Industrial Chemistry, vol.A2, p.57-97 VCH: Weinheim, 1985.
Биосинтез этих природных аминокислот в организмах, способных к их продуцированию, таких как бактерии, был хорошо охарактеризован (обзор бактериального биосинтеза аминокислот и его регуляцию см. в Umbarger, H.E. (1978) Ann.Rev. Biochem. 47: 533-606). Глутамат синтезируется путем восстановительного аминирования α-кетоглутарата, промежуточного соединения в цикле лимонной кислоты. Каждый из глутамина, пролина и аргинина последовательно продуцируется из глутамата. Биосинтез серина представляет собой трехстадийный процесс, начинающийся с 3-фосфоглицерата (промежуточного соединения при гликолизе) и кончающийся образованием этой аминокислоты после стадий окисления, трансаминирования и гидролиза. Цистеин и глицин продуцируются из серина; при этом, первый образуется путем конденсации гомоцистеина с серином, а второй образуется путем переноса атома β-углерода боковой цепи на тетрагидрофолат в реакции, катализируемой серин-трансгидроксиметилазой. Фенилаланин и тирозин синтезируются из предшественников гликолитического и пентозо-фосфатного пути метаболизма, то есть, эритрозо-4-фосфата и фосфоенолпирувата в 9-стадийном пути биосинтеза, который отличается только последними двумя стадиями после синтеза префената. Триптофан также продуцируется из этих двух исходных молекул, но его синтез проходит в 11 стадий. Тирозин может также синтезироваться из фенилаланина в реакции, катализируемой фенилаланин-гидроксилазой. Аланин, валин и лейцин являются биосинтетическими продуктами пирувата, конечного продукта гликолиза. Аспартат образуется из оксалоацетата, промежуточного соединения цикла лимонной кислоты. Аспарагин, метионин, треонин и лизин, каждый, продуцируются путем превращения аспартата. Изолейцин образуется из треонина. Сложный 9-стадийный путь метаболизма приводит к продуцированию гистидина из 5-фосфорибозил-1-пирофосфата, активированного сахара.
При избыточном синтезе белка, аминокислоты, необходимые для данной клетки, не могут сохраняться, а вместо этого они деградируют с образованием промежуточных соединений, участвующих в главных путях метаболизма, протекающих в данной клетке (обзор см. Stryer L. Biochemistry 3rd ed. Ch.21 «Amino Acid Degradation and the Urea Cycle» p.495-516 (1988)). Хотя данная клетка обладает способностью превращать нежелательные аминокислоты в нужные промежуточные соединения метаболизма, получение аминокислот является дорогостоящим с точки зрения энергетических затрат, затрат на получение молекул-предшественников и ферментов, необходимых для их синтеза. Таким образом, не удивительно, что биосинтез аминокислоты регулируется путем ингибирования по типу обратной связи, где присутствие конкретной аминокислоты служит для замедления или полного прекращения ее собственного продуцирования (обзор механизмов обратной связи в реакциях биосинтеза аминокислот см. Stryer L. Biochemistry 3rd ed. Ch.24 «Biosynthesis of Amino Acid and Heme» p.575-600 (1988)). Таким образом, выход любой конкретной аминокислоты ограничен количеством аминокислоты, присутствующей в клетке.
В. Метаболизм витаминов, кофакторов и питательных фармацевтических продуктов и их использование
Витамины, кофакторы и питательные фармацевтические продукты содержат молекулы другой группы, способность к синтезу которых у высших животных была утрачена и которые должны поставляться с пищей, хотя они легко синтезируются другими организмами, такими как бактерии. Эти молекулы сами по себе являются либо биологически активными веществами, либо предшественниками биологически активных веществ, которые могут служить в качестве носителей электрона или в качестве промежуточных соединений в различных путях метаболизма. Помимо их питательного значения, эти соединения также имеют важное промышленное значение как окрашивающие агенты, антиоксиданты и катализаторы или другие технологические добавки. (Обзор структуры, активности и промышленного применения этих соединений см., например, в Ullmann's Encyclopedia of Industrial Chemistry, vol. A27, «Vitamins», p.443-613 VCH: Weinheim, 1996). Термин "витамин" известен специалистам и означает питательные вещества, которые необходимы организму для нормального функционирования, но которые он сам не может синтезировать. Группа витаминов может охватывать кофакторы и питательные фармацевтические соединения. Понятие "кофактор" включает небелковые соединения, необходимые для нормальной ферментативной активности. Такими соединениями могут быть органические или неорганические соединения; при этом, молекулами кофакторов настоящего изобретения предпочтительно являются органические соединения. Термин "питательный фармацевтический препарат" включает пищевые добавки, полезные для жизнедеятельности растений и животных, а особенно для здоровья человека. Примерами таких молекул являются витамины, антиоксиданты, а также некоторые липиды (например, полиненасыщенные жирные кислоты).
Биосинтез этих молекул в организмах, способных их продуцировать, таких как бактерии, был хорошо охарактеризован (Ullman's Encyclopedia of Industrial Chemistry, vol.A27, «Vitamins», p.443-613 VCH: Weinheim, 1996; Michal, G. (1999) Biochemical Pathways: An Atlas of Biochemistry and Molecular Biology, John Wiley & Sons; Ong, A.S., Niki E. & Packer, L. (1995) «Nutrition, Lipids, Health and Disease» Proceedings of the UNESCO/Confederation of Scientific and Technological Associations in Malaysia and the Society for Free Radical Research - Asia, held Sept. 1-3, 1994 at Penang, Malaysia, AOCS Press: Champaign, IL X, 374 S).
Тиамин (витамин B1) продуцируется путем химического соединения пиримидина и тиазольных групп. Рибофлавин (витамин B2) синтезируется из гуанозин-5'-трифосфата (GTP) и рибозо-5'-фосфата. Рибофлавин, в свою очередь, используется для синтеза флавин-мононуклеотида (FMN) и флавин-аденин-динуклеотида (FAD). Семейство соединений под общим названием «витамин В6» (например, пиридоксин, пиридоксамин, пиридокса-5'-фосфат и коммерчески используемый гидрохлорид пиридоксина) представляет собой производные общей структурной единицы, 5-гидрокси-6-метилпиридина. Пантотенат (пантотеновая кислота, (R)-(+)-N-(2, 4-дигидрокси-3,3-диметил-1-оксобутил)-(β-аланин) может быть продуцирован либо путем химического синтеза, либо путем ферментации. Конечные стадии в биосинтезе пантотената состоят из АТР-регулируемой конденсации β-аланина и пантоевой кислоты. Ферменты, ответственные за стадии биосинтеза для превращения в пантоевую кислоту, в β-аланин и для конденсации в пантотеновую кислоту, известны специалистам. Метаболически активной формой пантотената является кофермент А, биосинтез которого протекает в 5 ферментативных стадий. Пантотенат, пиридоксал-5'-фосфат, цистеин и АТР являются предшественниками кофермента А. Эти ферменты не только катализируют образование пантотената, но также и продукцию (R)-пантоевой кислоты, (R)-пантолактона, (R)-пантенола (провитамина B5), пантетеина (и его производных) и кофермента А.
Биосинтез биотина из молекулы-предшественника пимелоил-СоА в микроорганизмах был досконально изучен и были идентифицированы некоторые гены, ответственные за этот синтез. Было обнаружено, что многие из соответствующих белков также вовлечены в синтез Fe-кластера и являются членами белков класса nifS. Липоевая кислота образуется из октановой кислоты и служит в качестве кофермента в энергетическом обмене, где она становится частью пируватдегидрогеназного комплекса и α-кетоглутарат-дегидрогеназного комплекса. Фолаты представляют собой группу веществ, которые являются производными фолевой кислоты, которая, в свою очередь, продуцируется из L-глутаминовой кислоты, п-аминобензойной кислоты и 6-метилптерина. Биосинтез фолевой кислоты и ее производных, начинающийся с метаболизма промежуточных соединений гуанозин-5'-трифосфата (GTP), L-глутаминовой кислоты и п-аминобензойной кислоты, был досконально изучен у некоторых микроорганизмов.
Корриноиды (такие как кобаламины и особенно витамин B12) и порфирины принадлежат к группе химических соединений, характеризующихся наличием тетрапирроловой кольцевой системы. Биосинтез витамина B12 является достаточно сложным и еще недостаточно хорошо изучен, однако, в настоящее время известны многие ферменты и субстраты, участвующие в этом биосинтезе. Никотиновая кислота (никотинат) и никотинамид являются пиридиновыми производными, которые также называют «ниацином».
Ниацин является предшественником важных коферментов NAD (никотинамид-аденин-динуклеотид) и NADP (никотинамид-аденин-динуклеотидфосфат) и их восстановленных форм.
Крупномасштабное продуцирование этих соединений основано, главным образом, на бесклеточном химическом синтезе, хотя некоторые из этих химических соединений, таких как рибофлавин, витамин В6, пантотенат и биотин, были также продуцированы с помощью крупномасштабного культивирования микроорганизмов. Лишь витамин B12 продуцируется только путем ферментации, что обусловлено сложностью его синтеза. In vitro-методики требуют значительных расходов на материалы и много времени, а поэтому часто являются дорогостоящими.
С. Метаболизм пуринов, пиримидинов, нуклеозидов и нуклеотидов и их использование.
Гены метаболизма пуринов и пиримидинов и их соответствующие белки являются главными мишенями для терапии опухолевых болезней и вирусных инфекций. Термин «пурин» или «пиримидин» означает азотистые основания, которые являются компонентами нуклеиновых кислот, коферментов и нуклеотидов. Термин «нуклеотид» включает основные структурные единицы молекул нуклеиновой кислоты, которые состоят из азотистого основания, сахара пентозы (в случае РНК, таким сахаром является рибоза; а в случае ДНК, таким сахаром является D-дезоксирибоза) и фосфорной кислоты. Термин «нуклеозид» включает молекулы, которые служат в качестве предшественников нуклеотидов, но которые не содержат молекулы фосфорной кислоты, имеющейся у нуклеотидов. Путем ингибирования биосинтеза этих молекул или их мобилизации с образованием молекул нуклеиновой кислоты можно ингибировать синтез РНК и ДНК; а путем ингибирования этой активности при направленной доставке к раковым клеткам, способность опухолевых клеток к делению и репликации может быть ингибирована. Кроме того, существуют нуклеотиды, которые не образуют молекул нуклеиновой кислоты, а вместо этого служат в качестве запасного энергетического источника (то есть, АМР) или в качестве коферментов (то есть, FAD и NAD).
В некоторых публикациях описано использование этих химических соединений в медицинских целях благодаря их влиянию на метаболизм пуринов и/или пиримидинов (например, Christopherson, R.I. & Lyons, S.D. (1990) «Potent inhibitors of de novo pyrimidine and purine biosynthesis as chemotherapeutic agents». Med. Res. Reviews 10: 505-548). Исследования ферментов, участвующих в метаболизме пуринов и пиримидинов, были направлены на разработку новых лекарственных средств, которые могут быть использованы, например, в качестве иммунодепрессантов или антипролифирирующих средств (Smith J.L., (1995) «Enzymes in nucleotide synthesis». Curr.Opin. Struct. Biol. 5: 752-757 (1995) Biochem. Soc. Transact. 23: 877-902). Однако, пуриновые и пиримидиновые основания, нуклеозиды и нуклеотиды имеют и другие применения, а именно, как промежуточные соединения в биосинтезе некоторых химических продуктов тонкого органического синтеза (например, тиамина, S-аденозилметионина, фолатов или рибофлавина), как энергоносители для данной клетки (например, АТР или GTP), и для получения самих химических соединений, обычно используемых в качестве ароматизирующих добавок (например, IMP или GMP) или для некоторых медицинских целей (см. например, Kuninaka A. (1996) Nucleotides and Related Compounds in Biotechnology, vol.6, Rehm et al., eds. VCH: Weinheim, p.561-612). Кроме того, ферменты, участвующие в метаболизме пуринов, пиримидинов, нуклеозидов или нуклеотидов, все больше служат в качестве мишеней, против которых, для защиты сельскохозяйственных культур, разрабатываются химические соединения, включая фунгициды, гербициды и инсектициды.
Метаболизм этих соединений в бактериях был охарактеризован (обзор см. например, Zaikin Н. & Dixon J.E. (1992) «de novo purine nucleotide biosynthesis», Progress in Nucleic Acid Research and Molecular Biology, vol.42. Academic Press: p. 259-287; и Michal G. (1999) «Nucieotides and Nucleosides», Chapter 8, Biochemical pathways: An Atlas of Biochemistry and Molecular Biology, Wiley: New York). Метабозизм пуринов является объектом интенсивных исследований и имеет важное значение для нормального функционирования клетки. Нарушение метаболизма пуринов у высших животных может вызывать тяжелую болезнь, такую как подагра. Пуриновые нуклеотиды синтезируются из рибозо-5-фосфата в серии стадий через промежуточное соединение инозин-5'-фосфат (IMP), в результате чего продуцируются гуанозин-5'-монофосфат (GMP) или аденозин-5'-монофосфат (АМР), из которых легко образуются трифосфатные формы, используемые как нуклеотиды. Эти соединения также используются как энергозапасающие вещества, поскольку их деградация обеспечивает энергией множество различных биохимических процессов в клетке. Биосинтез пиримидинов протекает с образованием уридин-5'-монофосфата (UMP) из рибозо-5-фосфата. UMP, в свою очередь, превращается в цитидин-5'-трифосфат (СТР). Дезоксиформы всех этих нуклеотидов образуются в одностадийной реакции восстановления из дифосфатрибозной формы нуклеотида в дифосфатдезоксирибозную форму нуклеотида. После фосфорилирования, эти молекулы способны участвовать в синтезе ДНК.
D. Метаболизм трегалозы и ее использование
Трегалоза состоит из двух глюкозных молекул, связанных друг с другом α,α-1,1-связью. Она широко используется в пищевой промышленности в качестве подсластителя, добавки для сушки или замораживания продуктов и в напитках. Однако, она также применяется в фармацевтике, косметике и биотехнологии (см, например, Nishimoto et al., (1998) патент США №5759610; Singer M.A. & Lindquist S. (1998) Trends Biotech. 16: 460-467; Paiva C.L.A. & Panek A.D. (1996) Biotech. Ann.Rev. 2: 293-314; и Shiosaka M. (1997) J. Japan 172: 97-102). Трегалоза продуцируется ферментами во многих микроорганизмах и обычно высвобождается в окружающую среду, из которой она может быть изолирована известными методами.
II. Резистентность к разрушению химическими соединениями, к воздействию окружающей среды и к антибиотикам Продуцирование химических продуктов тонкого органического синтеза обычно осуществляют крупномасштабным культивированием бактерий, разработанных для продуцирования и секреции больших количеств указанных молекул. Однако, такой тип крупномасштабной ферментации подвергает микроорганизмы стрессовым нагрузкам различных типов. Такими стрессами являются воздействие окружающей среды и химические стрессы.
А. Резистентность к воздействию окружающей среды
Примерами воздействия окружающей среды, с которым обычно сталкиваются при крупномасштабном ферментативном культивировании, являются механический стресс, тепловой шок, стресс, обусловленный недостатком кислорода, стресс, обусловленный присутствием кислородных радикалов, рН-стресс и осмотический стресс. Механизм перемешивания, используемый в большинстве реакторов для крупномасштабной ферментации для аэрации культуры, сопровождается выделением тепла, что приводит к повышению температуры культуры. Повышение температуры индуцирует хорошо охарактеризованный ответ в виде теплового шока, при котором экспрессируется ряд белков, которые не только способствуют выживанию бактерии в условиях высоких температур, но также и способствуют их выживанию в ответ на ряд других воздействий окружающей среды (см. Neidhardt, F.C., et al., eds. (1996) E.coli and Salmonella. ASM Press: Washington, D.C., p.1382-1399; Wosten M.M. (1998) FEMS Microbiology Reviews 22(3): 127-50; Bahl H. et al. (1995) FEMS Microbiology Reviews 17(3): 341-348; Zimmerman J.L., Cohill, P.R. (1991) New Biologist 3(7): 641-650; Samali A. & Orrenius, S. (1998) Cell. Stress Chaperones 3(4): 228-236, и ссылки, имеющиеся в каждой из цитируемых работ для каждого из этих случаев). Регуляция реакции теплового шока у бактерий облегчается специфическими сигма-факторами и другими клеточными регуляторами экспрессии генов (Hecker, M. Volker, U. (1998) Molecular Microbiology 29(5): 1129-1136). Одна из главных проблем, с которой сталкиваются клетки при воздействии на них высоких температур, заключается в том, что нарушается укладка белков; растущие белки имеют достаточное количество кинетической энергии в условиях высоких температур, в результате чего растущей полипептидной цепи трудно сохранять стабильную конформацию достаточно долго, чтобы правильно сложиться. Таким образом, белки, экспрессируемые во время реакции теплового шока, состоят из белков двух основных типов: шаперонов (белков, которые способствуют укладке или не-укладке других белков - см. например, Fink, A.L. (1999) Physiol. Rev. 79(2): 425-449) -, и протеаз, которые могут разрушать любые неправильно уложенные белки. Примерами шаперонов, экспрессируемых во время реакции теплового шока, являются GroEL и DNAK; а протеазами, которые, как известно, экспрессируются во время реакции теплового шока, являются Lon, FtsH и ClpB.
Помимо тепла, стрессовые нагрузки могут вызывать и другие воздействия окружающей среды. Хотя процесс перемешивания в ферментере предназначается для введения кислорода в данную культуру, кислород может оставаться в ограниченных количествах, особенно, когда данная культура продолжает расти, и потребность в кислороде тем самым возрастает; недостаточная доставка кислорода вызывает другой стресс у микроорганизмов. Клетки в культарах ферментера также подвергаются ряду осмотических стрессов, особенно, когда к культуре добавляют питательные вещества, что приводит к высокой внеклеточной и низкой внутриклеточной концентрации этих молекул. Кроме того, большие количества нужных молекул, продуцируемых этими микроорганизмами при их культивировании, могут приводить к осмотическому стрессу в бактериях. И наконец, аэробный метаболизм, такой как метаболизм, происходящий в С. glutamicum, приводит к образованию двуокиси углерода, как побочного продукта, и секреция этой молекулы может подкислять культуральную среду, что обусловлено превращением этой молекулы в карбоновую кислоту. Таким образом, при культивировании, бактерии также часто подвергаются стрессу с достижением кислотного рН. Такое превращение может быть также благоприятным, в том случае, если в культуральной среде присутствуют высокие уровни основных молекул побочных продуктов, таких как аммоний, и в этом случае бактерии в культуре могут подвергаться стрессу с достижением основного рН.
Для борьбы против таких воздействий окружающей среды, бактерии имеют тонкие системы генов, которые экспрессируются под воздействием одного или нескольких стрессов, таких как вышеупомянутая система теплового шока. Гены, экспрессируемые в ответ на осмотический шок, например, кодируют белки, способные транспортировать или синтезировать соответствующие растворимые вещества, так, чтобы осмотическое поглощение или экспорт конкретной молекулы замедлялся до контролируемых уровней. Другими примерами индуцированных стрессом бактериальных белков являются белки, участвующие в биосинтезе трегалозы; ферменты, участвующие в ppGpp-метаболизме; белки, участвующие в трансдукции сигнала, а в частности, двухкомпонентные системы, которые являются чувствительными к осмотическому давлению; и факторы транскрипции, которые ответственны за ряд факторов стресса (например, RssB-аналоги и/или сигма-факторы). Специалистам известны и многие другие гены и их белковые продукты.
В. Резистентность к химическому стрессу
Помимо стрессов, возникающих под воздействием окружающей среды, клетки могут также подвергаться ряду химических стрессов. Эти стрессы могут быть разделены на две категории. К первой категории относятся природные побочные продукты метаболизма и других клеточных процессов, секретируемые данной клеткой в окружающую среду. Ко второй категории относятся химические вещества, присутствующие во внеклеточной среде и не являющиеся продуктами данной клетки. В основном, если клетки выделяют токсические побочные продукты из концентрированной внутриклеточной цитоплазмы в относительно гораздо более разведенную внеклеточную среду, то эти продукты распределяются так, что внеклеточные уровни возможного токсичного соединения являются достаточно низкими. Однако, крупномасштабное ферментативное культивирование бактерии - это не тот случай, поскольку многие бактерии растут в относительно небольшом пространстве и при таком высоком уровне метаболизма, что побочные продукты могут накапливаться в данной среде до почти токсических уровней. Примерами таких побочных продуктов являются двуокись углерода, металлические ионы и реакционноспособные виды кислорода, такие как перекись водорода. Эти соединения могут влиять на активность или структуру молекул клеточной поверхности, либо они могут снова проникать в клетку, где они способны серьезно повреждать белки и нуклеиновые кислоты. В окружающей среде могут обнаруживаться и некоторые другие химические вещества, вредные для нормального функционирования клеток. Так, например, ионы металлов, таких как ртуть, кадмий, никель или медь, часто обнаруживаются в водных источниках и могут иметь форму тесных комплексов с клеточными ферментами, которые препятствуют нормальному функционированию этих белков.
С. Резистентность к антибиотикам
Во внеклеточной среде могут также обнаруживаться бактерицидные белки или антибиотики, которые присутствуют в ней либо в результате их занесения исследователями, либо в виде природного продукта от другого организма, используемого для повышения преимущества при конкуренции. Микроорганизмы обладают несколькими известными механизмами для своей защиты против противомикробных химических соединений. Широко применяемыми способами удаления и дезинтоксикации антибиотиков являются деградация, модификация и экспорт соединений, токсических для данной клетки. Известно, что некоторые прокариоты имеют цитоплазматические «откачивающие насосы» и обнаруживают сходство с так называемыми белками «резистентности ко множеству лекарственных средств», имеющимися у высших эукариотов (Neyfakh, A.A. et al. (1991) Proc. Natl. Acad. Sci. USA 88: 4781-4785). Примерами таких белков являются emrAB от E.coli (Lomovskaya, О. & K.Lewis (1992) Proc. Natl. Acad. Sci. USA 89: 8938-8942), ImrB от B.subtilis (Kumano M. et al (1997) Microbiology 143: 2775-2782), smr от S.aureus (Grinius, L.G. et al (1992) Piasmid 27: 119-129) или cmr от С. glutamicum (Kaidoh К. et al. (1997) Micro. Drug. Resist. 3: 345-350). Сам по себе микроорганизм С. glutamicum является непатогенным в отличие от некоторых других членов рода Corynebacterium, таких как С. diphtheriae или С. pseudotuberculosis. Известно, что некоторые патогенные коринебактерии имеют множественную резистентность к ряду антибиотиков, такие как C.jeikeium и С.urealyticum (Soriano, F. et al., (1995) Antimicrob. Agents Chemother. 39:208-214).
Линкозамиды известны как эффективные антибиотики против видов Corynebacterium (Soriano, F. et al., (1995) Antimicrob. Agents Chemother. 39: 208-214). Неожиданным результатом настоящего изобретения была идентификация гена, кодирующего белок резистентности к линкозамиду (в частности, белок резистентности к линкомицину). Белок LMRB, происходящий от С. glutamicum, обнаруживает 40%-ную гомологию с продуктом гена ImrB от В. subtilis (см. Genbank № доступа AL009126), вычисленную с использованием программы CLUSTALW версии 1.7 (Thompson, J.D., Higgins, D.G., Gibson, T.J. (1994) Nucl. Acids Res. 22: 4673-4680) с использованием стандартных параметров попарного сопоставления (PAIRWISE ALIGNMENT PARAMETERS: сопоставления медленные/точные: «штраф на пробел-пропуск» =10,0 и «штраф на пробел-удлинение» =0,10, весовая матрица белка =BLOSUM 30, весовая матрица ДНК =IUB, сопоставления быстрые/приблизительные: штраф на пробел =3, размер К-набора (слов) =1, № верхних диагоналей =5, размер окна =5, попарные сопоставления с переключением «медленно/быстро» = медленно. Параметры множественного сопоставления: «штраф на пробел-пропуск» =10/0 и «штраф на пробел-удлинение» =0,05, последовательности, расходящиеся с запаздыванием =40%, вес ДНК-транзиций =0,50, весовая матрица белка = серия BLOSUM, весовая матрица ДНК = IUB, негативная матрица использования = OFF).
Воздействие окружающей среды, химический стресс и стресс под воздействием антибиотиков или других противомикробных веществ могут повлиять на поведение микроорганизмов в процессе их культивирования и на продуцирование нужного соединения из этих микроорганизмов. Так, например, осмотический стресс микроорганизма может вызывать неприемлемое или неприемлемо быстрое поглощение одного или более соединений, что, в конечном счете, приводит к повреждению или гибели клеток под действием осмотического шока. Аналогичным образом, химические соединения, присутствующие в данной культуре, которые либо были экзогенно добавлены (например, противомикробные соединения, предназначенные для удаления нежелательных микробов), либо были генерированы самими бактериями (например, побочные продукты, такие как тяжелые металлы или кислородные радикалы, или даже противомикробные соединения), могут приводить к ингибированию продуцирования химических продуктов тонкого органического синтеза или даже к гибели микроорганизма. Гены настоящего изобретения кодируют белки С. glutamicum, которые предупреждают повреждение или гибель клеток путем специфического противодействия источнику или действию стресса, вызываемого окружающей средой или химическим веществом.
III. Элементы и способы настоящего изобретения
Настоящее изобретение основано, по крайней мере, частично, на обнаружении новых молекул, называемых здесь молекулами нуклеиновой кислоты и белка SRT, которые повышают способность С. glutamicum выживать в неблагоприятных условиях химического стресса или воздействия окружающей среды. В одном из вариантов осуществления изобретения, молекулы SRT функционируют так, что они сообщают микроорганизму С. qlutamicum резистентность к одному или нескольким стрессам, вызываемым воздействием окружающей среды или химических веществ. В предпочтительном варианте осуществления изобретения, активность молекул SRT настоящего изобретения влияет на продуцирование нужного химического продукта тонкого органического синтеза этим микроорганизмом. В особенно предпочтительном варианте осуществления изобретения, молекулы SRT настоящего изобретения имеют модулированную активность, такую, что выход, продуцирование и/или эффективность продуцирования одного или нескольких химических продуктов тонкого органического синтеза из С. glutamlcum также являются модулированными.
Термин «белок SRT» или «полипептид SRT» включает белки, которые участвуют в сообщении резистентности С. glutamicum к одному или нескольким стрессам, вызываемым окружающей средой или химическим веществом. Примерами белков SRT являются белки, кодируемые генами SRT, представленными в Таблице 1 и нечетно пронумерованными SEQ ID Nos. Термины «ген SRT» или последовательность нуклеиновой кислоты SRT" означают нуклеотидные последовательности, кодирующие белок SRT и состоящие из кодирующей области, а также из соответствующих нетранслируемых 5'- и 3'-областей последовательности. Примерами генов SRT являются гены, представленные в Таблице 1. Термины «продуцирование» или «продуктивность» известны специалистам и означают концентрацию продукта ферментации (например, нужного химического продукта тонкого органического синтеза), полученную в данное время и в данном объеме ферментации (например, кг продукта в час на литр). Термин «эффективность продуцирования» означает время, необходимое для достижения конкретного уровня продуцирования (например, время, затрачиваемое данными клетками на достижение конкретной скорости выхода химических продуктов тонкого органического синтеза). Термин «выход» или выход «продукт/углерод» хорошо известен специалистам и включает эффективность превращения углеродного источника в продукт (то есть, химический продукт тонкого органического синтеза). Выход обычно выражают, например, в кг продукта на кг углеродного источника. При увеличении выхода или продуцирования указанного соединения, увеличивается количество изолированных молекул или используемых изолированных продуктов данного соединения в данном количестве культуры по отношению к данному количеству времени. Термины «биосинтез» или «путь биосинтеза» хорошо известны специалистам и означают клеточный синтез соединения, предпочтительно, органического соединения из промежуточных соединений, который может быть многостадийным и в высокой степени регулируемым процессом. Термины «деградация» или «путь деградации» хорошо известны специалистам и означают разрушение соединения, предпочтительно, органического соединения, в клетке с получением продуктов деградации (вообще говоря, более мелких или менее сложных молекул), которая может быть многостадийным и в высокой степени регулируемым процессом. Термин «метаболизм» хорошо известен специалистам и означает все биохимические реакции, протекающие в организме. Кроме того, метаболизм конкретного соединения (например, метаболизм аминокислоты, такой как глицин) включает все пути биосинтеза, модификации и деградации в данной клетке, относящиеся к этому соединению. Термины «резистентность» и «толерантность» известны специалистам и означают способность клетки не подвергаться воздействию химических веществ или условий окружающей среды, которые должны так или иначе приводить к нарушению нормального функционирования этих микроорганизмов. Термины «стресс» или «опасность» включают факторы, которые неблагоприятно влияют на нормальное функционирование клеток, таких как С. glutamicum. Примерами стрессов являются «химический стресс», при котором клетка подвергается воздействию одного или нескольких химических соединений, негативно влияющих на эту клетку, и «стресс под действием окружающей среды», при котором клетка подвергается воздействию условий окружающей среды, которые отличаются от тех условий, к которым адаптирована данная клетка. Химические стрессы могут вызываться либо природными побочными продуктами метаболизма, такими как, но не ограничивающимися ими, реакционноспособные виды кислорода или двуокись углерода, либо химическими соединениями, так или иначе присутствующими в окружающей среде, включая, но не ограничиваясь ими, ионы тяжелых металлов или бактерицидные белки, такие как антибиотики. Стрессами, вызываемыми воздействием окружающей среды, могут быть, но не ограничиваются ими, например, температуры, выходящие за пределы нормальных температур, присутствие субоптимальных количеств кислорода, осмотическое давление или крайние значения рН.
В другом варианте осуществления изобретения, молекулы SRT способны модулировать продуцирование нужной молекулы, такой как химический продукт тонкого органического синтеза, в микроорганизме, таком как С. glutamicum. В соответствии с методами рекомбинантных ДНК, один или несколько белков SRT настоящего изобретения могут быть модифицированы так, чтобы их функция была модулированной. Изменение активности генов ответа, резистентности или толерантности к стрессу, заключающееся в повышении толерантности данной клетки к одному или нескольким стрессам, может улучшать способность данной клетки к росту и размножению в относительно стрессовых условиях при крупномасштабном культивировании в ферментере. Так, например, путем сверхэкспрессии или конструировании молекулы шаперона, индуцируемой тепловым шоком так, чтобы она имела оптимизированную активность, можно повысить способность данной бактерии к продуцированию правильно уложенных белков в температурных условиях, не являющихся оптимальными. При уменьшении количества белков с неправильной укладкой (а возможно и разрегулированных или нефункциональных), повышается способность данной клетки к нормальному функционированию в такой культуре, что, в свою очередь, приводит к увеличению ее жизнеспособности. Это общее увеличение числа клеток, обладающих большей жизнеспособностью и активностью в культуре, также должно приводить к увеличению выхода, продуцирования и/или эффективности продуцирования одного или нескольких нужных химических продуктов тонкого органического синтеза, что обусловлено, по крайней мере, относительно более высоким числом клеток, продуцирующих эти химические соединения в данной культуре.
Изолированные последовательности нуклеиновых кислот настоящего изобретения содержатся в геноме штамма Corynebacterium glutamicum, имеющегося в Американской коллекции типовых культур под номером доступа АТСС 13032. Нуклеотидная последовательность изолированной ДНК SRT С. glutamicum и предсказанные аминокислотные последовательности белков SRT С. glutamicum показаны в Списке последовательностей нечетно пронумерованными SEQ ID NO: и четно пронумерованными SEQ ID NO, соответственно.
Были проведены компьютерные анализы, которые позволили классифицировать и/или идентифицировать эти нуклеотидные последовательности как последовательности, кодирующие белки химического стресса и стресса, вызванного окружающей средой, а также белки резистентности и толерантности.
Настоящее изобретение также относится к белкам, имеющим аминокислотную последовательность, которая является, в основном, гомологичной аминокислотной последовательности настоящего изобретения (например, последовательности SEQ ID NO, имеющие четный номер в Списке последовательностей). Используемый здесь белок, имеющий аминокислотную последовательность, которая является, в основном, гомологичной выбранной аминокислотной последовательности, представляет собой белок, аминокислотная последовательность которого, по крайней мере, на 50% гомологична выбранной аминокислотной последовательности, т.е. всей выбранной аминокислотной последовательности. Белок, который имеет аминокислотную последовательность, в основном, гомологичную выбранной аминокислотной последовательности, может также представлять собой белок, аминокислотная последовательность которого, по крайней мере, примерно на 50-60%, а предпочтительно, по крайней мере, примерно на 60-70%, а более предпочтительно, по крайней мере, примерно на 70-80%, 80-90% или 90-95%, и наиболее предпочтительно, по крайней мере, примерно на 96%, 97%, 98%, 99% или более является гомологичной выбранной аминокислотной последовательности. Величины диапазонов и идентичностей, которые являются промежуточными для вышеуказанных величин (например, идентичность на 75-80%, идентичность на 85-87%, идентичность на 91-92%), также входят в объем настоящего изобретения. В настоящее изобретение, например, также входят величины диапазонов идентичностей, являющиеся комбинацией любых из вышеуказанных величин, указанных как верхний и/или нижний предел.
Белки SRT или их биологически активные части или фрагменты настоящего изобретения могут сообщать резистентность или толерантность к одному или нескольким химическим стрессам или стрессам, вызываемым воздействием окружающей среды, либо они могут обладать одной или несколькими активностями, представленными в Таблице 1.
Различные аспекты настоящего изобретения будут более подробно изложены в следующих подразделах:
А. Изолированные молекулы нуклеиновой кислоты
В одном из своих аспектов, настоящее изобретение относится к изолированным молекулам нуклеиновой кислоты, которые кодируют полипептиды SRT или его биологически активные части, а также к фрагментам нуклеиновой кислоты, достаточным для использования в качестве зондов для гибридизации или праймеров для идентификации или амплификации SRT-кодирующей нуклеиновой кислоты (например, SRT-ДНК). Используемый здесь термин «молекула нуклеиновой кислоты» включает ДНК-молекулы (например, кДНК или геномную ДНК) и РНК-молекулы (например, мРНК) и аналоги ДНК или РНК, генерированные с использованием нуклеотидных аналогов. Этот термин также охватывает нетранслируемую последовательность, расположенную у 3'- и 5'-концов кодирующей области гена: по крайней мере, примерно на расстоянии 100 нуклеотидов выше от 5'-конца кодирующей области и, по крайней мере, примерно на расстоянии 20 нуклеотидов ниже от 3'-конца кодирующей области гена. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, но предпочтительно, двухцепочечной ДНК. «Изолированной» молекулой нуклеиновой кислоты является молекула, которая отделена от других молекул нуклеиновой кислоты, присутствующих в природном источнике нуклеиновой кислоты. Предпочтительной «изолированной» нуклеиновой кислотой является молекула, не содержащая последовательности, которые обычно фланкируют нуклеиновую кислоту (то есть, последовательности, расположенные у 5' и 3'-концов нуклеиновой кислоты)в геномной ДНК данного организма, от которого происходит данная нуклеиновая кислота. Так, например, в различных вариантах осуществления изобретения, изолированная молекула нуклеиновой кислоты SRT может содержать нуклеотидные последовательности размером примерно менее, чем 5 т.п.н.-, 4 т.п.н.-, 3 т.п.н.-, 2 т.п.н.-, 1 т.п.н.-, 0,5 т.п.н.- или 0,1 т.п.н.-, которые обычно фланкируют молекулу нуклеиновой кислоты в геномной ДНК клетки, от которой происходит данная нуклеиновая кислота (например, клетка С. glutamicum}. Более того, «изолированная» молекула нуклеиновой кислоты, такая как молекула ДНК, может, в основном, не содержать другого клеточного материала или культуральной среды, если она была продуцирована рекомбинантными методами, или химических предшественников или других химических соединений, если она была продуцирована методами химического синтеза.
Молекула нуклеиновой кислоты настоящего изобретения, например, молекула нуклеиновой кислоты, имеющая нуклеотидную последовательность SEQ ID NO, имеющая нечетный номер в Списке последовательностей, или ее часть, может быть изолирована с применением стандартных методов молекулярной биологии и с использованием информации, приведенной в данном описании. Так, например, ДНК SRT С. glutamicum может быть изолирована из библиотеки С. glutamicum с использованием всех или одной из нечетно пронумерованных последовательностей SEQ ID NO: в Списке последовательностей в качестве зонда для гибридизации, и стандартной техники гибридизации (например, как описано Sambrook, J., Fritsh, E.F. & Maniatis Т., Molecular Cloning: A Laboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989). Кроме того, молекула нуклеиновой кислоты, охватывающая всю или часть одной из последовательностей нуклеиновых кислот нуклеиновой кислоты настоящего изобретения (например, нечетно пронумерованных SEQ ID Nos), может быть изолирована с помощью полимеразной цепной реакции с использованием олигонуклеотидных праймеров, сконструированных на основе данной последовательности (например, молекула нуклеиновой кислоты, охватывающая всю или часть одной из последовательностей нуклеиновой кислоты настоящего изобретения (например, нечетно пронумерованных SEQ ID Nos: в Списке последовательностей), может быть изолирована с помощью полимеразной цепной реакции с использованием олигонуклеотидных праймеров, сконструированных на основе той же самой последовательности). Так, например, мРНК может быть изолирована из нормальных эндотелиальных клеток (например, гуанидиний-тиоизоцианатным методом экстракции Chirgwin et al. (1979) Biochemistry 18: 5294-5299, и ДНК может быть получена с использованием обратной транскриптазы (например, обратной транскриптазы вируса мышиного лейкоза Молони (MLV), поставляемого Gibco/BRL, Bethesda, MD, или обратной транскриптазы AMV, поставляемой Seikagaku America, Inc., St.Petersburg, FL). Синтетические олигонуклеотидные праймеры для полимеразной цепной реакции могут быть сконструированы на основе одной из нуклеотидных последовательностей, представленных в Списке последовательностей. Нуклеиновая кислота настоящего изобретения может быть амплифицирована с использованием кДНК, или альтернативно, геномной ДНК в качестве матрицы и соответствующих олигонуклеотидных праймеров в соответствии со стандартными методами ПЦР-амплификации. Амплифицированная таким образом нуклеиновая кислота может быть клонирована в соответствующий вектор и охарактеризована с помощью анализа ДНК-последовательности. Кроме того, олигонуклеотиды, соответствующие нуклеотидной последовательности SRT, могут быть получены стандартными методами синтеза, например, с использованием автоматического ДНК-синтезатора.
В предпочтительном варианте осуществления изобретения, изолированная молекула нуклеиновой кислоты настоящего изобретения содержит одну из нуклеотидных последовательностей, представленных в Списке последовательностей. Последовательности нуклеиновых кислот настоящего изобретения, представленные в Списке последовательностей, соответствуют SRT-ДНК Corynebacterium glutamicum настоящего изобретения. Эта ДНК включает последовательности, кодирующие белки SRT (то есть, «кодирующую область», показанную в каждой нечетно пронумерованной последовательности SEQ ID NO: в Списке последовательностей), а также 5'-нетранслирумые последовательности и 3'-нетранслируемые последовательности, также указанные в каждой нечетно пронумерованной SEQ ID NO: в Списке последовательностей). Альтернативно, молекула нуклеиновой кислоты может содержать только кодирующую область любой из указанных последовательностей нуклеиновых кислот в Списке последовательностей.
Следует отметить, что в целях настоящего изобретения, каждая из последовательностей нуклеиновых кислот и аминокислотных последовательностей, представленных в Списке последовательностей, имеет идентифицирующие номера RXA, RXN или RXS, имеющие обозначения «RXA», «RXN» или «RXS», после которых следуют 5 цифр (то есть, RXA01524, RXN00493 или RXS01027).
Каждая из указанных последовательностей нуклеиновых кислот содержит вплоть до 3 частей: область, расположенную выше от 5'-конца, кодирующую область, и нижерасположенную область. Во избежание путаницы, каждая из этих трех областей идентифицируется по тем же самым обозначениям RXA, RXN или RXS. Кроме того, выражение «одна из нечетно пронумерованных последовательностей в Списке последовательностей» относится к любым последовательностям нуклеиновых кислот в Списке последовательностей, которые могут быть также идентифицированы по их отличающимся обозначениям RXA, RXN или RXS. Кодирующая область каждой из этих последовательностей транслируется в соответствующую аминокислотную последовательность, которая также представлена в Списке последовательностей как четно пронумерованная SEQ ID NO: и которая в этом списке следует непосредственно за соответствующей последовательностью нуклеиновой кислоты. Так, например, кодирующая область RXA01524 представлена в SEQ ID NO:1, а аминокислотная последовательность, которую она кодирует, представлена как SEQ ID NO:2. Последовательности молекул нуклеиновой кислоты настоящего изобретения идентифицируются по тем же самым обозначениям RXA, RXN или RXS, как и аминокислотные молекулы, которые они кодируют, так, чтобы они могли легко коррелировать. Так, например, аминокислотная последовательность, обозначенная RXA01524, представляет собой последовательность, транслированную с кодирующей области нуклеотидной последовательности молекулы нуклеиновой кислоты RXA01524; аминокислотная последовательность, обозначенная RXN00034, представляет собой последовательность, транслированную с кодирующей области нуклеотидной последовательности молекулы нуклеиновой кислоты RXN00034; а аминокислотная последовательность, обозначенная RXS00568, представляет собой последовательность, транслированную с кодирующей области нуклеотидной последовательности молекулы нуклеиновой кислоты RXS00568. Соответствие между нуклеотидными и аминокислотными последовательностями RXA, RXN и RXS настоящего изобретения и их SEQ ID №№ представлены в таблице 1.
Некоторые гены настоящего изобретения представляют собой «F-обозначенные гены». F-обозначенными генами являются гены, представленные в таблице 1, которые имеют букву «F», стоящую перед обозначениями RXA, RXN или RXS. Так, например, SEQ ID NO:7, обозначенная, как показано в таблице 1 «F RXA00498», представляет собой F-обозначенный ген, как и SEQ ID NO:25, 33 и 37 (обозначенные в таблице 1 как «F RXA01345», «F RXA02543» и «F RXA02282», соответственно).
В одном из вариантов осуществления изобретения, молекулы нуклеиновой кислоты настоящего изобретения не включают последовательности в таблице 2. В случае гена dapD, последовательность для этого гена опубликована Wehrmann A., et al., (1998) J.Bacteriol. 180(12): 3159-3165. Однако, последовательность, полученная авторами настоящего изобретения, в основном, длиннее, чем опубликованная версия. Очевидно, что в основе опубликованной версии последовательности лежит неправильный старт-кодон, а поэтому эта последовательность представляет собой лишь фрагмент реальной кодирующей последовательности.
В другом варианте осуществления изобретения, изолированная молекула нуклеиновой кислоты настоящего изобретения включает молекулу нуклеиновой кислоты, которая является комплементарной одной из указанных нуклеотидных последовательностей настоящего изобретения (например, нечетно пронумерованной последовательности SEQ ID NO: в Списке последовательностей) или ее часть. Молекула нуклеиновой кислоты, которая является комплементарной одной из нуклеотидных последовательностей настоящего изобретения, представляет собой молекулу, которая является достаточно комплементарной одной из нуклеотидных последовательностей, представленных в Списке последовательностей (например, нечетно пронумерованной последовательности SEQ ID NO:), такая, что она может гибридизоваться с одной из нуклеотидных последовательностей настоящего изобретения, и тем самым образовывать стабильный дуплекс.
В еще одном предпочтительном варианте осуществления изобретения, изолированная молекула нуклеиновой кислоты настоящего изобретения включает нуклеотидную последовательность, которая, по крайней мере, примерно на 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59% или 60%, предпочтительно, по крайней мере, примерно на 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69% или 70%, более предпочтительно, по крайней мере, примерно, на 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79% или 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89% или 90%, или 91%, 92%, 93%, 94%, и даже более предпочтительно, по крайней мере, примерно на 95%, 96%, 97%, 98%, 99% или более гомологична нуклеотидной последовательности настоящего изобретения (например, нечетно пронумерованной последовательности SEQ ID NO: в Списке последовательностей) или ее части. В объем настоящего изобретения также входят величины интервалов и идентичностей, которые являются промежуточными для вышеуказанных диапазонов (например, 70-90%-ная идентичность или 80-95%-ная идентичность). Так, например, в объем настоящего изобретения также входят интервалы значений идентичности с использованием комбинации любых вышеупомянутых значений, указанных как верхний и/или нижний предел. В еще одном предпочтительном варианте осуществления изобретения, изолированная молекула нуклеиновой кислоты настоящего изобретения включает нуклеотидную последовательность, которая гибридизуется в жестких условиях с одной из нуклеотидных последовательностей настоящего изобретения или с ее частью.
Кроме того, молекула нуклеиновой кислоты настоящего изобретения может включать лишь часть кодирующей области данной последовательности одной из нечетно пронумерованных последовательностей SEQ ID NO: в Списке последовательностей, например, фрагмент, который может быть использован в качестве зонда или праймера, или фрагмент, кодирующий биологически активную часть белка SRT. Нуклеотидные последовательности, определенные в результате клонирования генов SRT от С. glutamicum, позволяют генерировать зонды и праймеры, сконструированные для идентификации и/или клонирования SRT-гомологов в клетках других типов и организмах, а также SRT-гомологов от других Corynebacteria или родственных видов. Зонд/праймер обычно содержат в существенной степени очищенный олигонуклеотид. Этот олигонуклеотид обычно включает область нуклеотидной последовательности, которая гибридизуется в жестких условиях, по крайней мере, примерно с 12, предпочтительно, примерно с 25, более предпочтительно, примерно с 40, 50 или 75 расположенными подряд нуклеотидами смысловой цепи одной из нуклеотидных последовательностей настоящего изобретения (например, одной из нечетно пронумерованных последовательностей SEQ ID NO: в Списке последовательностей), антисмысловой последовательностью одной из этих последовательностей или с их природными мутантами. Праймеры, полученные на основе последовательности нуклеиновой кислоты настоящего изобретения, могут быть использованы в ПЦР-реакциях для клонирования SRT-гомологов. Зонды, полученные на основе нуклеотидных последовательностей SRT, могут быть использованы для детекции транскриптов или геномных последовательностей, кодирующих те же самые или гомологичные белки. В предпочтительных вариантах осуществления изобретения, указанный зонд, кроме того, включает присоединенную к нему группу-метку, например, такой группой-меткой может быть радиоактивный изотоп, флуоресцентное соединение, фермент или кофактор фермента. Указанные зонды могут быть использованы как часть диагностического тест-набора для идентификации клеток, которые недостаточно экспрессируют белок SRT, например, путем измерения уровня SRT-кодирующей нуклеиновой кислоты в образце клетки, например, путем детекции уровней SRT-мРНК или определения, был ли ген SRT мутирован или делетирован.
В одном из вариантов осуществления изобретения, молекула нуклеиновой кислоты настоящего изобретения кодирует белок или его часть, которая включает аминокислотную последовательность, являющуюся достаточно гомологичной аминокислотной последовательности настоящего изобретения (например, четно пронумерованной последовательности SEQ ID NO: в Списке последовательностей), так, что указанный белок или его часть сохраняют способность сообщать С. glutamicum резистентность или толерантность к одному или более стрессам, вызываемым химическими соединениями или воздействием окружающей среды. Используемый здесь термин «достаточно гомологичный» относится к белкам или их частям, имеющим аминокислотные последовательности, которые включают минимальное число аминокислотных остатков (например, аминокислотный остаток, который имеет боковую цепь, аналогичную боковой цепи аминокислотного остатка в четно пронумерованной последовательности SEQ ID NO: в Списке последовательностей), идентичных или эквивалентных аминокислотной последовательности настоящего изобретения, так, что данный белок или его часть способны сообщать С. glutamicum резистентность к одному или нескольким стрессам, вызываемым химическими веществами или воздействием окружающей среды. Белки, являющиеся членами указанных путей метаболизма и описанные в настоящей заявке, способствуют увеличению резистентности или толерантности С. glutamicum к одному или более вредным факторам или стрессам, вызываемым химическими веществами или воздействием окружающей среды. Примеры такой активности также описаны в настоящей заявке. Таким образом, «функция белка SRT» заключается в сообщении С. glutamicum общей резистентности к окружающим ее элементам, которые могут неблагоприятно влиять на ее нормальный рост или функцию, и/или способствуют, прямо или опосредованно, увеличению выхода, продуцирования и/или эффективности продуцирования одного или нескольких химических продуктов тонкого органического синтеза. Примеры активностей белка SRT представлены в таблице 1.
В другом варианте осуществления изобретения, данный белок является, по крайней мере, примерно на 50-60%, предпочтительно, по крайней мере, примерно на 60-70%, более предпочтительно, по крайней мере, примерно на 70-80%, 80-90%, 90-95%, а наиболее предпочтительно, по крайней мере, примерно на 96%, 97%, 98%, 99% или более гомологичным всей аминокислотной последовательности настоящего изобретения (например, четно пронумерованной последовательности SEQ ID NO: в Списке последовательностей). Объем настоящего изобретения также включает величины интервалов и идентичностей, которые являются промежуточными для вышеуказанных диапазонов (например, 75-80%-ная идентичность или 85-87%-ная идентичность или 91-92%-ная идентичность). Так, например, в объем настоящего изобретения также входят интервалы значений идентичности с использованием комбинации любых вышеупомянутых значений, указанных как верхний и/или нижний предел.
Части белков, кодируемых молекулами нуклеиновой кислоты SRT настоящего изобретения, являются, предпочтительно, биологически активными частями одного из белков SRT. Используемый здесь термин "биологически активная часть белка SRT" означает часть, например, домен/мотив белка SRT, которая способна сообщать резистентность или толерантность к одному или более вредным факторам или стрессам, вызываемым химическими соединениями или воздействием окружающей среды, или обладает активностью, описанной в таблице 1. Для того чтобы определить, способен ли белок SRT или его биологически активная часть повышать резистентность или толерантность С. glutamicum к одному или более вредным факторам или стрессам, вызываемым химическими соединениями или воздействием окружающей среды, может быть проведен анализ на ферментативную активность. Методы осуществления таких анализов хорошо известны каждому специалисту и подробно описаны в Примере 8 описания изобретения.
Дополнительные фрагменты нуклеиновой кислоты, кодирующие биологически активные части белка SRT, могут быть получены путем изолирования части одной из аминокислотных последовательностей настоящего изобретения (например, четно пронумерованной последовательности SEQ ID NO: в Списке последовательностей), экспрессии кодируемой части белка или пептида SRT (например, путем рекомбинантной экспрессии in vitro) и оценки активности кодированной части белка или пептида SRT.
Настоящее изобретение также относится к молекулам нуклеиновой кислоты, которые отличаются от одной из нуклеотидных последовательностей настоящего изобретения (например, нечетно пронумерованной последовательности SEQ ID NO: в Списке последовательностей) (или ее части), вследствие вырожденности генетического кода, а поэтому кодируют тот же самый белок SRT, который кодируется нуклеотидными последовательностями настоящего изобретения. В другом варианте осуществления изобретения, изолированная молекула нуклеиновой кислоты настоящего изобретения имеет нуклеотидную последовательность, кодирующую белок, имеющий аминокислотную последовательность, указанную в Списке последовательностей (например, четно пронумерованную последовательность SEQ ID NO:). В еще одном варианте осуществления изобретения, молекула нуклеиновой кислоты настоящего изобретения кодирует полноразмерный белок С. glutamicum, который, в основном, гомологичен аминокислотной последовательности настоящего изобретения (кодированной открытой рамкой считывания, указанной в нечетно пронумерованной последовательности SEQ ID NO: в Списке последовательностей).
Для каждого специалиста очевидно, что в одном из вариантов осуществления изобретения, последовательности настоящего изобретения не должны включать известные последовательности, такие как последовательности, имеющиеся в Genbank, представленные в таблицах 2 или 4, которые были известны до появления настоящего изобретения. В одном из вариантов своего осуществления, настоящее изобретение относится к нуклеотидным и аминокислотным последовательностям, которые имеют большую длину, чем известная последовательность (например, последовательность, имеющаяся в Genbank) (или белок, кодированный такой последовательностью), представленная в таблице 2 или 4). Так, например, настоящее изобретение относится к нуклеотидной последовательности, которая больше и/или, по крайней мере, на 39% идентична нуклеотидной последовательности, обозначенной RXA00084 (SEQ ID NO:189), нуклеотидной последовательности, которая больше и/или, по крайней мере, на 56% идентична нуклеотидной последовательности, обозначенной RXA00605 (SEQ ID NO:11), и нуклеотидной последовательности, которая больше и/или, по крайней мере, на 50% идентична нуклеотидной последовательности, обозначенной RXA00886 (SEQ ID NO:39). Любой специалист может вычислить нижний предел процента идентичности для любой данной последовательности настоящего изобретения путем оценки GAP-вычисленных величин по шкале идентичности, представленных в Таблице 4 для каждого из трех наилучших совпадений для данной последовательности, и путем вычитания наибольшего GAP-вычисленного значения процента идентичности из 100 процентов. Для каждого специалиста также очевидно, что нуклеотидные и аминокислотные последовательности, имеющие процент идентичности, превышающий вычисленный таким образом нижний порог (например, по крайней мере, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59% или 60%, предпочтительно, по крайней мере, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69% или 70%, более предпочтительно, по крайней мере, примерно, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79% или 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89% или 90%, или 91%, 92%, 93%, 94%, и даже более предпочтительно, крайней мере, примерно 95%, 96%, 97%, 98%, 99% или более идентичны) также входят в объем настоящего изобретения.
Помимо нуклеотидных последовательностей SRT С. glutamicum, представленных в Списке последовательностей как нечетно пронумерованные SEQ ID №№, для каждого специалиста очевидно, что в данной популяции (например, в популяции С. glutamicum) может иметь место полиморфизм ДНК-последовательностей, который приводит к изменениям в аминокислотных последовательностях белков SRT. Такой генетический полиморфизм в гене SRT может присутствовать у индивидуумов данной популяции вследствие природных модификаций. Используемые здесь термины «ген» и «рекомбинантный ген» означают молекулы нуклеиновой кислоты, содержащие открытую рамку считывания, кодирующую белок SRT, предпочтительно, белок SRT С. glutamicum. Такие природные вариации могут быть, в основном, результатом 1-5%-ной вариации нуклеотидной последовательности гена SRT. Любые из таких вариаций нуклеотидной последовательности, приводящих к полиморфизму в SRT, который является следствием природных вариаций, а поэтому не приводят к изменению функциональной активности белков SRT, входят в объем настоящего изобретения.
Молекулы нуклеиновой кислоты, соответствующие природным вариантам и не происходящим от С. glutamicum-гомологам SRT-ДНК С. glutamicum настоящего изобретения, могут быть выделены исходя из их гомологии с нуклеиновой кислотой SRT С. qlutamicum, рассматриваемой в данной заявке, с использованием ДНК С. glutamicum или ее части, в качестве зонда для гибридизации в соответствии со стандартными методами гибридизации в жестких условиях гибридизации. В соответствии с этим, в другом варианте осуществления изобретения, изолированная молекула нуклеиновой кислоты настоящего изобретения имеет длину, по крайней мере, в 15 нуклеотидов и гибридизуется в жестких условиях с молекулой нуклеиновой кислоты, содержащей нуклеотидную последовательность, представленную нечетно пронумерованной последовательностью SEQ ID NO: в Списке последовательностей. В других вариантах осуществления изобретения, данная нуклеиновая кислота имеет длину, по крайней мере, в 30, 50, 100, 250 или более нуклеотидов. Используемый здесь термин «гибридизируется в жестких условиях» относится к гибридизации и промывке в описанных условиях, при которых нуклеотидные последовательности, имеющие, по крайней мере, 60%-ную гомологию, обычно продолжают гибридизоваться друг с другом. Данные условия должны быть, предпочтительно, такими, чтобы эти последовательности, имеющие гомологию, по крайней мере, примерно на 65%, более предпочтительно, по крайней мере, примерно на 70%, и даже более предпочтительно, по крайней мере, примерно, на 75% или более, в основном, продолжают гибридизоваться друг с другом. Такие жесткие условия известны каждому специалисту и их описание можно найти у Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1.-6.3.6. Предпочтительным неограничивающим примером гибридизации в жестких условиях является гибридизация в 6х хлориде натрия/цитрате натрия (SSC) примерно при 45°С с последующей одной или несколькими промывками в 0,2×SSC, 0,1% ДСН при 55-65°С. При этом, предпочтительно, чтобы изолированная молекула нуклеиновой кислоты настоящего изобретения, которая гибридизуется в жестких условиях с нуклеотидной последовательностью настоящего изобретения, соответствовала природной молекуле нуклеиновой кислоты. Используемый здесь термин «природная» молекула нуклеиновой кислоты относится к РНК- или ДНК-молекуле, имеющей нуклеотидную последовательность, встречающуюся в природе (например, кодирующую природный белок). В одном из вариантов осуществления изобретения, данная нуклеиновая кислота кодирует природный белок SRT С. glutamicum.
Помимо природных вариантов последовательности SRT, которые могут присутствовать в данной популяции, для каждого специалиста также очевидно, что в нуклеотидную последовательность настоящего изобретения могут быть внесены изменения путем мутации, которые, тем самым, приводят к изменениям в аминокислотной последовательности кодированного белка SRT, не влияя, при этом, на функциональную способность данного белка SRT. Так, например, в нуклеотидной последовательности настоящего изобретения могут быть сделаны замены нуклеотидов, приводящие к заменам в «не существенных» аминокислотных остатках. «Не существенным» аминокислотным остатком является аминокислотный остаток, который может быть заменен в последовательности дикого типа одного из белков SRT (например, в четно пронумерованной последовательности SEQ ID NO: в Списке последовательностей) без изменения активности указанного белка SRT, тогда как «существенным» аминокислотным остатком является аминокислотный остаток, который необходим для осуществления активности белка SRT. Однако, имеются и другие аминокислотные остатки (например, остатки, которые не являются консервативными или являются лишь полуконсервативными в домене, обладающем SRT-активностыо), которые могут не играть главной роли в активности белка, а поэтому они, вероятно, могут быть заменены без изменения активности SRT.
В соответствии с этим, в другом аспекте своего осуществления, настоящее изобретение относится к молекулам нуклеиновой кислоты, кодирующим белки SRT, которые содержат изменения в аминокислотных остатках, не имеющих важного значения для SRT-активности. Аминокислотная последовательность такого белка SRT отличается от аминокислотной последовательности из указанной четно пронумерованной последовательности SEQ ID NO: в Списке последовательностей, но еще сохраняет, по крайней мере, одну из описанных здесь активностей SRT. В одном из вариантов осуществления изобретения, изолированная молекула нуклеиновой кислоты имеет нуклеотидную последовательность, кодирующую белок, где этот белок включает аминокислотную последовательность, которая, по крайней мере, примерно на 50% гомологична аминокислотной последовательности настоящего изобретения, способна повышать резистентность или толерантность к одному или более вредным факторам или стрессам, вызываемым воздействием окружающей среды или химическими веществами, или имеет одну или более активностей, представленных в таблице 1. Предпочтительно, чтобы белок, кодируемый указанной молекулой нуклеиновой кислоты. был, по крайней мере, примерно на 50-60% гомологичен аминокислотной последовательности одной из нечетно пронумерованных последовательностей в Списке последовательностей, более предпочтительно, по крайней мере, примерно на 60-70%, гомологичен одной из этих последовательностей, более предпочтительно, по крайней мере, примерно на 70-80%, 80-90%, 90-95% гомологичен одной из этих последовательностей, а наиболее предпочтительно, по крайней мере, примерно на 96%, 97%, 98% или 99% гомологичен одной из аминокислотных последовательностей настоящего изобретения.
Для определения процента гомологии двух аминокислотных последовательностей (например, одной из аминокислотных последовательностей настоящего изобретения и его мутанта) или двух нуклеиновых кислот, последовательности выстраивают для их оптимального сравнения (например, в данную последовательность одного белка или нуклеиновой кислоты могут быть введены пробелы для оптимального сопоставления ее первичной последовательности с последовательностью другого белка или нуклеиновой кислоты). Затем сравнивают аминокислотные остатки или нуклеотиды в соответствующих положениях аминокислот или нуклеотидов. Если положение в одной последовательности (например, одной из аминокислотных последовательностей настоящего изобретения) занято тем же самым аминокислотным остатком или нуклеотидом, присутствующим в соответствующем положении другой последовательности (например, мутантной формы аминокислотной последовательности), то эти молекулы являются гомологичными в этом положении (то есть, используемый здесь термин "гомология" аминокислот или нуклеиновых кислот эквивалентен термину "идентичность" аминокислот или нуклеиновых кислот). Процент гомологии между этими двумя последовательностями зависит от числа идентичных положений, общих для данных последовательностей (то есть, % гомологии = число (#) идентичных положений/число всех (#) положений ×100).
Изолированная молекула нуклеиновой кислоты, кодирующая белок SRT, гомологичный последовательности белка настоящего изобретения (например, четно пронумерованной последовательности SEQ ID NO: в Списке последовательностей), может быть создана путем введения одной или нескольких нуклеотидных замен, добавлений или делеций в нуклеотидную последовательность настоящего изобретения, так, чтобы в кодированный белок были введены одна или несколько аминокислотных замен, добавлений или делеций. Мутации могут быть введены в одну из нуклеотидных последовательностей настоящего изобретения стандартными методами, такими как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Замены консервативных аминокислот, предпочтительно, осуществляют в одном или нескольких предполагаемых «не существенных» остатках аминокислот. «Замена консервативных аминокислот» означает замену присутствующего аминокислотного остатка аминокислотным остатком, имеющим аналогичную боковую цепь. Семейство аминокислотных остатков, имеющих аналогичную боковую цепь, известно специалистам. Такие семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), с кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), с незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), с неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), с бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, предполагаемый не существенный аминокислотный остаток в белке SRT, предпочтительно, заменяют другим аминокислотным остатком из семейства с той же самой боковой цепью. Альтернативно, в другом варианте осуществления изобретения, мутации могут быть введены произвольным образом во всю SRT-кодирующую последовательность или ее часть, например, путем насыщающего мутагенеза, и полученные мутанты могут быть скринированы на SRT-активность, описанную в данной заявке для идентификации мутантов, которые сохраняют SRT-активность. После мутагенеза нуклеотидной последовательности, являющейся одной из нечетно пронумерованных последовательностей SEQ ID NOs в Списке последовательностей), кодированный белок может быть рекомбинантно экспрессирован и активность этого белка может быть определена с использованием описанных здесь анализов (см. Пример 8 в Разделе «Иллюстрации примерами»).
Помимо молекул нуклеиновой кислоты, кодирующих белки SRT, описанных выше, в другом своем аспекте, настоящее изобретение относится к изолированным молекулам нуклеиновой кислоты, которые являются антисмысловыми. «Антисмысловая» нуклеиновая кислота включает нуклеотидную последовательность, комплементарную "смысловой" нуклеиновой кислоте, кодирующей белок, например, комплементарную кодирующей цепи двухцепочечной молекулы ДНК или комплементарную мРНК-последовательности. В соответствии с этим, антисмысловая нуклеиновая кислота может быть связана со смысловой нуклеиновой кислотой водородной связью. Антисмысловая нуклеиновая кислота может быть комплементарна всей SRT-кодирующей цепи, либо только ее части. В одном из вариантов осуществления изобретения, антисмысловая молекула нуклеиновой кислоты представляет собой молекулу, которая является антисмысловой по отношению к «кодирующей области» данной кодирующей цепи нуклеотидной последовательности, кодирующей белок SRT. Термин «кодирующая область» означает область нуклеотидной последовательности, содержащую кодоны, которые транслируются с образованием аминокислотных остатков (например, полная кодирующая область SEQ ID NO:120 (RXA00600), включает нуклеотиды 1-1098). В другом варианте осуществления изобретения, антисмысловая молекула нуклеиновой кислоты представляет собой молекулу, которая является антисмысловой по отношению к «некодирующей области» данной кодирующей цепи нуклеотидной последовательности, кодирующей SRT. Термин «некодирующая область» означает 5'- и 3'-последовательности, фланкирующие кодирующую область, которая не транслируется в аминокислоты (то есть, называемые также 5'- и 3'-нетранслируемыми областями).
Учитывая, что SRT-кодирующие последовательности кодирующей цепи, описанные в настоящей заявке (например, последовательности, представленные нечетно пронумерованными последовательностями SEQ ID NOs в Списке последовательностей), антисмысловые нуклеиновые кислоты настоящего изобретения могут быть сконструированы в соответствии с правилами спаривания оснований Уотсона и Крика. Антисмысловая молекула нуклеиновой кислоты может быть комплементарна всей кодирующей области SRT-мРНК, но более предпочтительно, чтобы эта молекула представляла собой олигонуклеотид, который является антисмысловым только по отношению к части, кодирующей или некодирующей области SRT-мРНК. Так, например, антисмысловой олигонуклеотид может быть комплементарен области, окружающей сайт инициации трансляции SRT-мРНК. Антисмысловой олигонуклеотид может, например, иметь длину приблизительно в 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50 нуклеотидов. Антисмысловая нуклеиновая кислота настоящего изобретения может быть сконструирована путем химического синтеза или реакций ферментативного лигирования в соответствии с известными процедурами. Так, например, антисмысловая нуклеиновая кислота (например, антисмысловой олигонуклеотид) может быть химически синтезирована с использованием природных нуклеотидов или различным образом модифицированных нуклеотидов, сконструированных для повышения биологической стабильности данных молекул или для повышения физической стабильности дуплекса, образованного между антисмысловой и смысловой нуклеиновыми кислотами, например, могут быть использованы производные фосфоротиоата и нуклеотиды, замещенные акридином. Примерами модифицированных нуклеотидов, которые могут быть использованы для генерирования данной антисмысловой нуклеиновой кислоты, являются 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-иодурацил, гипоксантин, ксантин, 4-ацетилцитозин, 5-(карбоксигидроксилметил)урацил, 5-карбоксиметиламинометил-2-тиоуридин, 5-карбоксиметиламинометилурацил, дигидроурацил, бета-D-галактозилхинозин, инозин, N6-изопентениладенин, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-аденин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2-тиоурацил, бета-D-маннозилхинозин, 5'-метоксикарбоксиметилурацил, 5-метоксиурацил, 2-метилтио-N6-изопентениладенин, урацил-5-оксиуксусная кислота (v), вибутоксозин, псевдоурацил, квеозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, метиловый эфир урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусная кислота (v), 5-метил-2-тиоурацил, 3-(3-амино-3-N-2-карбоксипропил)урацил, (acp3)w и 2,6-диаминопурин. Альтернативно, указанная антисмысловая нуклеиновая кислота может быть продуцирована биологически с использованием экспрессирующего вектора, в котором была субклонирована нуклеиновая кислота в антисмысловой ориентации (т.е., РНК, транскрибированная со вставленной нуклеиновой кислоты, будет иметь антисмысловую ориентацию по отношению к представляющей интерес целевой нуклеиновой кислоте, дополнительно описанной в нижеследующем подразделе).
Антисмысловые молекулы нуклеиновой кислоты настоящего изобретения обычно вводят в клетку или генерируют in situ так, чтобы они гибридизировались или связывались с клеточной мРНК и/или геномной ДНК, кодирующей белок SRT, в целях ингибирования экспрессии данного белка, например, путем ингибирования транскрипции и/или трансляции. Такая гибридизация может происходить благодаря соответствующей комплементарности нуклеотидов с образованием стабильного дуплекса или, например, в случае антисмысловой молекулы нуклеиновой кислоты, которая связывается с ДНК-дуплексами, посредством специфических взаимодействий в главной бороздке двойной спирали. Эта антисмысловая молекула может быть модифицирована так, чтобы она специфически связывалась с рецептором или антигеном, экспрессированным на поверхности выбранной клетки, например, путем связывания антисмысловой молекулы нуклеиновой кислоты с пептидом или антителом, который(ое) связывается с рецептором или антигеном клеточной поверхности. Указанная антисмысловая молекула нуклеиновой кислоты может быть также доставлена в клетки с использованием векторов, описанных в настоящей заявке. Для достижения достаточных внутриклеточных концентраций антисмысловых молекул, предпочтительными являются векторные конструкции, в которых антисмысловая молекула нуклеиновой кислоты находится под контролем сильного прокариотического, вирусного или эукариотического промотора.
В еще одном варианте осуществления изобретения, антисмысловой молекулой нуклеиновой кислоты настоящего изобретения является α-аномерная молекула нуклеиновой кислоты, α-аномерная молекула нуклеиновой кислоты образует специфические двухцепочечные гибриды с комплементарными РНК, в которых в отличие от обычных β-единиц, цепи расположены параллельно друг другу (Gaultier et al. (1987) Nucl. Acids Res. 15: 6625-6641). Эта антисмыловая молекула нуклеиновой кислоты может также включать 2'-о-метилрибонуклеотид (Inoue et al. (1987) Nucleic. Acids Res. 15: 6131-6148) или химерный РНК-ДНК-аналог (Inoue et al. (1987) FEBS Lett. 215: 327-330).
В другом варианте осуществления изобретения, антисмысловой молекулой нуклеиновой кислоты является рибозим. Рибозимы представляют собой каталитические РНК-молекулы с рибонуклеазной активностью, которые способны расщеплять одноцепочечную нуклеиновую кислоту, такую как мРНК и с которыми они имеют комплементарную область. Таким образом, рибозимы (например, рибозимы с молоткообразной головкой (описанные Haselhoff & Gerlach (1988) Nature 334:585-591)) могут быть использованы для каталитического расщепления SRT-мРНК-транскриптов в целях ингибирования трансляции SRT-мРНК. Рибозим, обладающий специфичностью к SRT-кодирующей нуклеиновой кислоте, может быть сконструирован на основе нуклеотидной последовательности SRT-кДНК, описанной в настоящей заявке (то есть, SEQ ID NO:119 (RXA00600)). Так, например, может быть сконструировано производное РНК IVS Tetrahymena L-19, в котором нуклеотидная последовательность активного центра комплементарна нуклеотидной последовательности, расщепляемой в SRT-кодирующей мРНК. См, например, Cech et al., патент США №4987071 и Cech et al., патент США №5116742. Альтернативно, SRT-мРНК может быть использован для отбора каталитической РНК, обладающей специфической рибонуклеазной активностью, из пула РНК-молекул. См., например, Bartel, D. & Szostak, J.W. (1993) Science 261: 1411-1418.
Альтернативно, экспрессия гена SRT может быть ингибирована путем направленной доставки нуклеотидных последовательностей, комплементарных регуляторной области нуклеотидной последовательности SRT (например, промотору и/или энхансеру для гена SRT) с образованием спиральных структур из трех нитей, которые препятствуют транскрипции гена SRT в клетках-мишенях. В общих чертах, см., Helene, С (1991) Anticancer Drug Des. 6(6): 569-84; Helene, С. et al. (1992) Ann. N.Y. Acad.Sci. 660: 27-36; и Maher, L.J. (1992) Bioassays 14 (12): 807-15.
В. Рекомбинантные экспрессирующие векторы и клетки-хозяева.
В другом своем аспекте, настоящее изобретение относится к векторам, предпочтительно, к экспрессирующим векторам, содержащим нуклеиновую кислоту, кодирующую белок SRT (или его часть). Используемый здесь термин «вектор» означает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, которая была к нему присоединена. Одним типом вектора является «плазмида», которая представляет собой кольцевую двухцепочечную ДНК-петлю, в которую могут быть встроены дополнительные ДНК-сегменты. Другим типом вектора является вирусный вектор, где в данный вирусный геном могут быть встроены дополнительные ДНК-сегменты. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они были введены (например, бактериальные векторы, имеющие бактериальный сайт инициации репликации и эписомные векторы млекопитающих). Другие векторы (например, не-эписомные векторы млекопитающих), при их введении в клетку-хозяина, интегрируются в геном этой клетки-хозяина и тем самым реплицируются вместе с геномом хозяина. Кроме того, некоторые векторы способны регулировать экспрессию генов, которые были функционально встроены в них. Такие векторы в данном описании называются «экспрессирующими векторами». В основном, экспрессирующие векторы, используемые в технологии рекомбинантных ДНК, часто присутствуют в форме плазмид. В настоящем описании термины «плазмида» и «вектор» являются взаимозаменяемыми, поскольку плазмида представляет собой наиболее часто используемую форму вектора. Однако, настоящее изобретение включает и другие формы экспрессирующих векторов, таких как вирусные векторы (например, дефектные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы), которые имеют эквивалентные функции.
Рекомбинантные экспрессирующие векторы настоящего изобретения содержат нуклеиновую кислоту настоящего изобретения в форме, подходящей для экспрессии нуклеиновой кислоты в клетке-хозяине, что означает, что указанные рекомбинантные экспрессирующие векторы включают одну или более регуляторных последовательностей, которые были выбраны исходя из клеток-хозяев, используемых для экспрессии, и которые были функционально присоединены к экспрессируемой последовательности нуклеиновой кислоты. В указанном рекомбинантном экспрессирующем векторе, «функционально присоединенная» последовательность означает, что желаемая нуклеотидная последовательность присоединена к регуляторной(ым) последовательности(ям) так, чтобы происходила экспрессия данной нуклеотидной последовательности (например, в системе in vitro-транскрипции/трансляции или в клетке-хозяине после введения этого вектора в данную клетку-хозяина). Термин «регуляторная последовательность» включает промоторы, энхансеры и другие элементы, контролирующие экспрессию (например, сигналы полиаденилирования). Такие регуляторные последовательности описаны, например, у Goeddel; Gen Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990). Регуляторными последовательностями являются такие последовательности, которые ответственны за конститутивную экспрессию нуклеотидных последовательностей в клетках-хозяевах многих типов и которые ответственны за экспрессию нуклеотидной последовательности только в определенных клетках-хозяевах. Предпочтительными регуляторными последовательностями являются, например, промоторы, такие как cos-, tac-, trp-, tet-, trp-tet-, Ipp-, lac-, lpp-lac-, lacIq-, T7-, T5-, T3-, gal-, trc-, ara-, SP6-, arny, SPO2, λ-pR- или λ-РL, которые, предпочтительно, используются в бактериях. Дополнительными регуляторными последовательностями являются, например, промоторы, происходящие от дрожжей и грибов, такие как ADC1, MFα, AC, P60, CYC1, GAPDH, TEF, rp28, ADH, промоторы, происходящие от растений, такие как CAMV/35S, SSU, OCS, lib4, usp, STLS1, В33, nos или убихитиновые или фазеолиновые промоторы. Могут быть также использованы и искусственные промоторы. Для каждого специалиста очевидно, что конструирование экспрессирующего вектора может зависеть от таких факторов, как выбор клетки-хозяина, которую трансформируют и уровень экспрессии желаемого белка и т.п. Экспрессирующие векторы настоящего изобретения могут быть введены в клетки-хозяева с продуцированием белков или пептидов, включая гибридные белки или пептиды, кодируемые нуклеиновыми кислотами, описанными в настоящей заявке (например, белки SRT, мутантные формы белков SRT, гибридные белки и т.п.).
Рекомбинантные экспрессирующие векторы настоящего изобретения могут быть сконструированы для экспрессии белков SRT в прокариотических или эукариотических клетках. Так, например, гены SRT могут экспрессироваться в бактериальных клетках, таких как С. glutamicum, клетках насекомых (с использованием бакуловирусных экспрессирующих векторов), клетках дрожжей и других грибов (см. Romanes, М.А. et al. (1992) «Foreign gene expression in yeast: a review». Yeast 8: 423-488; van den Hondel, C.A.M.J.J. et al. (1991) «Heterologous gene expression in filamentous fungi»: More Gene Manipulations in Fungi, J.W.Bennet & L.L. Lasure, eds., p.396-428: Academic Press: San Diego; and van den Hondel, C.A.M.J.J. & Punt, P.J. (1991) «Gene transfer systems and vector development for filamentous fungi: Applied Molecular Genetics of Fungi», Peberdy, J.F. et al., eds., p.1-28, Cambridge University Press: Cambridge), клетках водорослей и многоклеточных растений (см. Schmidt R. & Willmitzer L. (1988) High efficiency Agrobacterium tumefaciens - mediated transformation of Arabidopsis thaliana leaf and cotyledon explants" Plant Cell Rep: 583-586) или клетках млекопитающих. Подходящие клетки-хозяева более подробно обсуждаются в работе Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990). Альтернативно, указанный рекомбинантный экспрессирующий вектор может быть транскрибирован и транслирован in vitro, например, с использованием промоторных регуляторных последовательностей Т7 и полимеразы Т7.
Экспрессию белков в прокариотах, в большинстве случаев, осуществляют с использованием векторов, содержащих конститутивные или индуцибельные промоторы, ответственные за экспрессию гибридных или негибридных белков. Гибридные векторы добавляют ряд аминокислот к белку, кодируемому в этом векторе, обычно к амино-концу данного рекомбинантного белка. Такие гибридные векторы обычно используются для трех целей: 1) для увеличения экспрессии рекомбинантного белка; 2) для увеличения растворимости рекомбинантного белка и 3) для облегчения очистки рекомбинантного белка благодаря его действию в качестве лиганда при аффинной очистке. Часто, сайт протеолитического расщепления вводят в гибридные экспрессирующие векторы на стыке между гибридной частью и рекомбинантным белком, что позволяет отделить данный рекомбинантный белок от гибридной части после очистки данного гибридного белка. Такими ферментами и их когнатными последовательностями распознавания являются фактор Ха, тромбин и энтерокиназа.
Типичными векторами для экспрессии гибридных молекул являются pGEX (Pharmacia Biotech Inc.; Smith, D.B. & Johnson, K.S. (1988) Gene 67: 31-40), pMAL (New England Biolabs, Beverly, MA) и pRIT5 (Pharmacia Piscataway, NJ), которые присоединяют глутатион-S-трансферазу (GST), связывающийся с мальтозой Е белок, или белок А, соответственно, к целевому рекомбинантному белку. В одном из вариантов осуществления изобретения, кодирующую последовательность белка SRT клонируют в экспрессирующий вектор pGEX для создания вектора, кодирующего гибридный белок, включающий, начиная от N-конца до С-конца, «GST-сайт расщепления тромбином-белок X». Этот гибридный белок может быть очищен с помощью аффинной хроматографии на глутатион-агарозной смоле. Рекомбинантный белок SRT, не присоединенный к GST, может быть выделен путем расщепления гибридного белка тромбином.
Примерами подходящих индуцибельных негибридных экспрессирующих векторов E.coli являются pTrc (Amann et al.,(1988) Gene 69: 301-315) pLG338, pACYC184, pBR322, pUC18, pUC19, Ркс30, pRep4, pHS1, pHS2, pPLc236, pMBL24, pLG200, pUR290, pIN-III113-B1, λgt11, pBdCl и Pet11d (Studier et al., Gene Expression Technology: Methods in Enzymology 185, Academic Press, San. Diego,-California (1990) 60-89; и Pouwels et al., eds. (1985) Cloning Vectors, Elsevier: New York IBSN 0444904018). Экспрессия гена-мишени из вектора pTrc основана на транскрипции с помощью РНК-полимеразы хозяина с trp-lac-гибридного промотора. Экспрессия гена-мишени из вектора рЕТ 11d основана на транскрипции с Т7 gn10-lac-гибридного промотора, опосредуемой коэкспрессированной вирусной РНК-полимеразой (Т7 gnl). Эта вирусная полимераза обеспечивается штаммами-хозяевами BL21(DE3) или HMS174(DE3) от присутствующего профага λ, содержащего ген gnl T7 под транскрипционным контролем промотора lacUV5. Для трансформации других видов бактерий, могут быть отобраны соответствующие векторы. Так, например, известно, что плазмиды pIJ101, pIJ364, pIJ702 и pIJ361 могут быть использованы для трансформации стрептомицетов, а плазмиды pUB110, pC194 или pBD214 являются подходящими для трансформации видов Bacillus. Некоторыми плазмидами, используемыми для переноса генетической информации в Corynebacterium, являются рНМ1519, pBLl, pSA77 или pAJ667 (Pouwels et al., eds. (1985) Cloning Vectors, Elsevier: New York IBSN 0444904018).
Одна из стратегий для максимизации экспрессии рекомбинантного белка заключается в экспрессии белка в бактерии-хозяине с нарушенной способностью к протеолитическому расщеплению рекомбинантного белка (Gottesman, S., Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, California (1990) 119-128). Другая стратегия заключается в изменении последовательности нуклеиновой кислоты, встраиваемой в экспрессирующий вектор, так, чтобы отдельные кодоны для каждой аминокислоты представляли собой кодоны, предпочтительно используемые в данной бактерии, выбранной для экспрессии, такой как С. glutamicum (Wada et al., (1992) Nucleic. Acids Res. 20:2111-2118). Такое изменение последовательностей нуклеиновых кислот настоящего изобретения может быть осуществлено стандартными методами синтеза ДНК.
В другом варианте осуществления изобретения, вектором, экспрессирующим белок SRT, является дрожжевой экспрессирующий вектор. Примерами векторов для экспрессии в дрожжах S. cerevisiae являются pYepSecl (Baldari et al., (1987) Embo J. 6: 229-234), 2μ, pAGI, Yep6, Yepl3, pEMBLYe23, pMFa (Kurjan & Herskowitz, (1982) Cell 30: 933-943), pJRY88 (Schultz et al., (1987) Gene 54:113-123) и pYES2 (Invitrogen Corporation, San Diego, CA). Векторами и методами для конструирования векторов, подходящих для использования в других грибах, таких как гифомицеты, являются векторы и методы, подробно описанные в работах van den Hondel, C.A.M.J.J. & Punt, P.J. (1991) «Gene transfer systems and vector development for filamentous fungi, Applied Molecular Genetics of Fungi», J.F. Peberdy, et al., eds., p.1-28, Cambridge University Press: Cambridge and Pouwels et al., eds. (1985) Cloning Vectors. Elsevier: New York (IBSN 0444904018).
Альтернативно, белки SRT настоящего изобретения могут быть экспрессированы в клетках насекомых с использованием бакуловирусных экспрессирующих векторов. Бакуловирусными векторами, подходящими для экспрессии белков в культивированных клетках насекомых (например, клетки Sf9), являются векторы серии рАс (Smith et al. (1983) Mol. Cell. Biol. 3:2156-2165) и векторы серии pVL (Lucklow & Summers (1989) Virology 170: 31-39).
В другом варианте осуществления изобретения, белки SRT настоящего изобретения могут быть экспрессированы в клетках одноклеточных растений (таких как водоросли) или в клетках высших растений (например, в сперматофитах, таких как культурные растения). Примерами растительных экспрессирующих векторов являются векторы, подробно описанные в работах Becker, D. Kemper, E., Schell, J. & Masterson, R. (1992) «New plant binary vectors with selectable markers located proximal to the left border». Plant Mol. Biol. 20:1195-1197; и Bevan M.W. (1984) «Binary Agrobacterium vectors for plant transformation», Nucl. Acid. Res. 12:8711-8721 и включают pLGV23, pGHlac+, pBIN19, pAK2004 и pDH51 (Pouwels et al., eds. (1985) Cloning Vectors. Elsevier: New York IBSN 0444904018).
В еще одном варианте осуществления изобретения, нуклеиновую кислоту настоящего изобретения экспрессируют в клетках млекопитающих с использованием экспрессирующего вектора, происходящего от млекопитающего. Примерами экспрессирующих векторов млекопитающих являются pCDM8 (Seed, В. (1987) Nature 329-840) и pMT2PC (Kaufman et al.(1987) EMBO J. 6: 187-195). При использовании в клетках млекопитающих, функции, регулируемые экспрессирующим вектором, в большинстве случаев обеспечиваются вирусными регуляторными элементами. Так, например, обычно используемые промоторы происходят от полиомавируса, аденовируса 2, цитомегаловируса и обезьяньего вируса SV40. Другие подходящие экспрессирующие системы для прокариотических и эукариотических клеток описаны в главах 16 и 17 Sambrook, J., Fritsh, E.F. & Maniatis Т., Molecular Cloning: A Laboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Laboratory Press, Cold Spring Harbor, NY, 1989.
В другом варианте осуществления изобретения, рекомбинантный экспрессирующий вектор млекопитающего обладает способностью направлять экспрессию нуклеиновой кислоты предпочтительно в клетках данного конкретного типа (например, для экспрессии нуклеиновой кислоты используется тканеспецифические регуляторные элементы). Такие тканеспецифические регуляторные элементы известны специалистам. Не ограничивающими примерами подходящих тканеспецифических промоторов являются альбуминовый промотор (печень-специфический; Pinkert et al., (1987) Genes Dev. 1: 268-277), лимфоид-специфические промоторы (Calame & Eaton (1988) Adv. Immunol, 43: 235-275), а в частности, промоторы Т-клеточных рецепторов (Winoto & Baltimore (1989) EMBO J. 8: 728-733) и иммуноглобулины (Banerji et al., (1983) Cell 33: 729-740; Queen & Baltimore (1983) Cell 33: 741-748), нейрон-специфические промоторы (например, нейрофиламентный промотор Вугпе & Ruddle (1989) PNAS 86: 5473-5477), промоторы, специфические для поджелудочной железы (Edlund et al., (1985) Science 230: 912-916), и промоторы, специфичные для молочных желез млекопитающих (например, промотор молочной сыворотки; патент США №4873316 и публикация европейской заявки №264166). В объем настоящего изобретения также входят стадиеспецифические регулируемые промоторы, например, промоторы мышиного hox (Kessel & Gruss (1990) Science 249: 374-379) и α-фетопротеиновый промотор (Campes & Tilghman (1989) Genes Dev. 3: 537-546).
Настоящее изобретение также относится к рекомбинантному экспрессирующему вектору, содержащему ДНК-молекулу настоящего изобретения, клонированную в данный экспрессирующий вектор в антисмысловой ориентации. То есть, ДНК-молекула функционально присоединена к регуляторной последовательности так, чтобы обеспечивалась экспрессия (путем транскрипции этой ДНК-молекулы) РНК-молекулы, которая является антисмысловой по отношению к SRT-мРНК. Могут быть выбраны регуляторные последовательности, функционально присоединенные к нуклеиновой кислоте, клонированной в антисмысловой ориентации, которые регулируют непрерывную экспрессию антисмысловой РНК-молекулы в клетках различных типов, например, могут быть выбраны вирусные промоторы и/или энхансеры или регуляторные последовательности, которые регулируют конститутивную тканеспецифическую или клеточноспецифическую экспрессию антисмысловой РНК. Антисмысловой экспрессирующий вектор может присутствовать в форме рекомбинантной плазмиды, фагмиды или аттенюированного вируса, в которых антисмысловые нуклеиновые кислоты продуцируются под контролем высокоэффективной регуляторной области, активность которой может быть определена типом клеток, в которые вводится данный вектор. Обсуждение регуляции экспрессии генов с использованием антисмысловых генов можно найти у Weintraub, Н. et al., Antisence RNA as a molecular tool for genetic analysis, Rewiews - Trends in Genetics., Vol. 1(1) 1986.
В другом своем аспекте, настоящее изобретение относится к клеткам-хозяевам, в которые был введен рекомбинантный экспрессирующий вектор настоящего изобретения. Термины «клетка-хозяин» и «рекомбинантная клетка-хозяин» в настоящем описании являются взаимозаменяемыми. Следует отметить, что эти термины относятся не только к конкретно рассматриваемой клетке, но также и к их потомству или их потенциальному потомству. Поскольку некоторые модификации могут присутствовать в последующих поколениях вследствие либо мутации, либо влияния окружающей среды, то такое потомство в действительности может не быть идентичным потомству родительских клеток, но, тем не менее, оно также входит в объем данного используемого здесь термина.
Клетка-хозяин может представлять собой либо прокариотическую, либо эукариотическую клетку. Так, например, белок SRT может быть экспрессирован в бактериальных клетках, таких как С. glutamicum, в клетках насекомых, в клетках дрожжей или млекопитающих (таких как клетки яичника китайского хомячка (СНО) или клетки COS). Другие подходящие клетки-хозяева известны специалистам. Микроорганизмы, родственные Corynebacterium glutamicum, которые могут быть с успехом использованы в качестве клеток-хозяев для экспрессии молекул нуклеиновой кислоты и белка настоящего изобретения, представлены в таблице 3.
Векторная ДНК может быть введена в прокариотические или эукариотические клетки стандартными методами трансформации или трансфекции. Используемые здесь термины «трансформация» и «трансфекция» относятся к ряду хорошо известных методов введения чужеродной нуклеиновой кислоты (например, линейной ДНК или РНК (например, линеаризованного вектора или генной конструкции без вектора) или нуклеиновой кислоты в форме вектора (например, плазмиды, фага, фазмиды, фагмиды, транспозона или другой ДНК)) в клетку-хозяина, включая ко-приципатацию фосфатом кальция или хлоридом кальция, трансфекцию, опосредованную DEAE-декстраном, липофекцию или электропорацию. Подходящие методы трансформации или трансфекции клеток-хозяев можно найти у Sambrook et al.,(1989). Molecular Cloning: A Laboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989 и в других лабораторных руководствах.
Известно, что для стабильной трансфекции клеток-млекопитающих, в зависимости от используемого экспрессирующего вектора и метода трансфекции, лишь небольшая часть клеток способна интегрировать в свой геном чужеродную ДНК. Для идентификации и отбора клеток с интегрированной ДНК, ген, кодирующий селективный маркер (например, ген резистентности к антибиотикам), обычно вводят в клетки-хозяева вместе с желаемым геном. Предпочтительными селективными маркерами являются маркеры, которые сообщают резистентность к лекарственным средствам, таким как G418, гигромицин и метотрексат. Нуклеиновая кислота, кодирующая селективный маркер, может быть введена в клетку-хозяина с помощью того же самого вектора, который кодирует белок SRT, либо она может быть введена с помощью отдельного вектора. Клетки, стабильно трансфецированные введенной нуклеиновой кислотой, могут быть идентифицированы путем отбора в присутствии данного лекарственного средства (например, клетки, в которые был введен селективный маркерный ген, будут выживать, а другие клетки погибнут).
Для создания гомологичного рекомбинантного микроорганизма, получают вектор, содержащий, по крайней мере, часть гена SRT, в котором были осуществлены делеция, добавление или замена в целях его модификации, например, функционального разрушения гена SRT. Таким геном SRT, предпочтительно, является ген SRT Corynebacterium glutamicum, но им может быть гомологичный ген, происходящий от родственной бактерии или даже от млекопитающего, дрожжей или насекомого. В предпочтительном варианте осуществления изобретения, вектор конструируют так, чтобы после гомологичной рекомбинации эндогенный ген SRT был функционально разрушен (т.е., чтобы он больше не кодировал функциональный белок, и такой вектор называется также «нокаут»-вектором). Альтернативно, данный вектор может быть сконструирован так, чтобы после гомологичной рекомбинации эндогенный ген SRT был мутирован или как-либо иначе модифицирован, но при этом кодировал функциональный белок (например, вышерасположенная регуляторная область может быть модифицирована с изменением экспрессии эндогенного белка SRT). В векторе для гомологичной рекомбинации, модифицированная часть гена SRT фланкируется с его 5'- и 3'-концов дополнительной нуклеиновой кислотой гена SRT, в результате чего происходит гомологичная рекомбинация между экзогенным геном SRT, содержащимся в данном векторе, и эндогенным геном SRT в микроорганизме. Дополнительная фланкирующая нуклеиновая кислота SRT имеет достаточную длину для успешной гомологичной рекомбинации с эндогенным геном. Обычно, в вектор включены несколько тысяч пар оснований фланкирующей ДНК (с 5'- и 3'-концов) (описание векторов для гомологичной рекомбинации см., например, Thomas K.R. & Capecchi M.R. (1987) Cell 51: 503). Данный вектор вводят в микроорганизм (например, путем электропорации), и клетки, в которых введенный ген SRT подвергается гомологичной рекомбинации с эндогенным геном SRT, отбирают известными методами.
В другом варианте осуществления изобретения могут быть продуцированы рекомбинантные микроорганизмы, которые содержат выбранные системы, позволяющие осуществлять регулируемую экспрессию введенного гена. Так, например, включение гена SRT в вектор под контролем lac-оперона способствует экспрессии гена SRT лишь в присутствии IPTG. Такие регуляторные системы хорошо известны специалистам.
В другом варианте осуществления изобретения, эндогенный ген SRT в клетке-хозяине разрушают (например, путем гомологичной рекомбинации или другими генетическими средствами, известными специалистам) так, чтобы не происходила экспрессия его белкового продукта. В другом варианте осуществления изобретения, эндогенный или введенный ген SRT в клетке-хозяине был модифицирован посредством одной или нескольких точковых мутаций, делеций или инверсий, но при этом, он еще кодировал функциональный белок SRT. В еще одном варианте осуществления изобретения, одна или несколько регуляторных областей (например, промотор, репрессор или индуктор) гена SRT в микроорганизме были модифицированы (например, посредством делеций, усечения, инверсии или точковой мутации) для модуляции экспрессии гена SRT. Для каждого специалиста очевидно, что клетки-хозяева, содержащие более одной модификации описанного гена и белка SRT, могут быть легко продуцированы способами настоящего изобретения и входят в объем настоящего изобретения.
Клетка-хозяин настоящего изобретения, такая как прокариотическая или эукариотическая клетка-хозяин в культуре, может быть использована для продуцирования (т.е., экспрессии) белка SRT. В соответствии с этим, настоящее изобретение, кроме того, относится к способам продуцирования белков SRT с использованием клеток-хозяев настоящего изобретения. В одном варианте осуществления изобретения, этот способ предусматривает культивирование клетки-хозяина настоящего изобретения (в которую был введен рекомбинантный экспрессирующий вектор, кодирующий белок SRT, или в геном которой был введен ген, кодирующий белок SRT дикого типа или модифицированный белок SRT) в подходящей среде до тех пор, пока не будет продуцирован белок SRT. В другом варианте осуществления изобретения, указанный способ, кроме того, предусматривает выделение белков SRT из данной среды или из клетки-хозяина.
С. Выделение белков SRT
В другом своем аспекте, настоящее изобретение относится к изолированным белкам SRT и к их биологически активным частям. «Изолированный» или «очищенный» белок или его биологически активная часть, в основном, не содержат клеточный материал при продуцировании методами рекомбинантных ДНК, либо химических предшественников или других химических соединений при химическом синтезе. Выражение "в основном, не содержит клеточного материала" относится к препаратам белка SRT, в которых данный белок был отделен от клеточных компонентов клетки, в которой он был продуцирован естественным или рекомбинантным способом. В одном из вариантов осуществления изобретения, выражение «в основном, не содержит клеточного материала» относится к препаратам белка SRT, содержащим примерно менее, чем 30% (сухой массы) белка, не являющегося SRT (также называемого «примесным белком»), более предпочтительно, примерно менее, чем 20% белка, не являющегося SRT, еще более предпочтительно, примерно менее, чем 10% белка, не являющегося SRT, a наиболее предпочтительно, примерно менее, чем 5% белка, не являющегося SRT. Если белок SRT или его биологически активная часть были продуцированы рекомбинантным методом, то предпочтительно, чтобы они также не содержали культуральную среду, то есть, культуральная среда составляет примерно менее, чем 20%, более предпочтительно, примерно менее, чем 10%, а наиболее предпочтительно, примерно менее, чем 5% по объему препарата белка. Выражение «в основном, не содержит химических предшественников или других соединений» относится к препаратам белка SRT, в которых данный белок был отделен от химических предшественников или других соединений, которые участвовали в синтезе данного белка. В одном из вариантов осуществления изобретения, выражение «в основном, не содержит химических предшественников или других соединений» относится к препаратам белка SRT, содержащим примерно менее, чем 30% (сухой массы) химических предшественников или химических соединений, не являющихся белком SRT, более предпочтительно, примерно менее, чем 20% химических предшественников или химических соединений, не являющихся белком SRT, еще более предпочтительно, примерно менее, чем 10% химических предшественников или химических соединений, не являющихся белком SRT, а наиболее предпочтительно, примерно менее, чем 5% химических предшественников или химических соединений, не являющихся белком SRT. В предпочтительных вариантах осуществления изобретения, изолированные белки или их биологически активные части не содержат примесные белки от того же самого организма, от которого происходит данный белок SRT. Обычно, указанные белки продуцируются посредством рекомбинантной экспрессии, например, белка SRT С. glutamicum в таком микроорганизме, как С. glutamicum.
Изолированный белок SRT настоящего изобретения или его часть могут сообщать резистентность или толерантность к одному или более вредным факторам или стрессам, вызываемым химическими веществами или воздействием окружающей среды, и обладает одной или более активностями, указанными в таблице 1. В предпочтительных вариантах осуществления изобретения, белок или его часть включает аминокислотную последовательность, которая значительно гомологична аминокислотной последовательности настоящего изобретения (например, четно пронумерованную последовательность SEQ ID NO: в Списке последовательностей), такую, что данный белок или его часть сохраняют способность опосредовать данную резистентность или толерантность к одному или более вредным факторам или стрессам, вызываемым химическими соединениями или воздействием окружающей среды. Указанная часть белка является, предпочтительно, биологически активной частью, описанной в настоящей заявке. В другом предпочтительном варианте осуществления изобретения, белок SRT имеет аминокислотную последовательность, кодируемую нуклеиновой кислотой, представленной четно пронумерованной последовательностью SEQ ID NO: в Списке последовательностей. В еще одном предпочтительном варианте осуществления изобретения, белок SRT имеет аминокислотную последовательность, кодируемую нуклеиновой кислотой, которая, например, гибридизуется в жестких условиях с одной из нуклеотидных последовательностей настоящего изобретения (например, с нечетно пронумерованной последовательностью SEQ ID NO: в Списке последовательностей). В еще одном предпочтительном варианте осуществления изобретения, данный белок SRT имеет аминокислотную последовательность, кодированную нуклеотидной последовательностью, которая, по крайней мере, примерно на 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59% или 60%, предпочтительно, по крайней мере, примерно на 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69% или 70%, более предпочтительно, по крайней мере, примерно на 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79% или 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89% или 90%, 91%, 92%, 93%, 94%, и даже более предпочтительно, по крайней мере, примерно на 95%, 96%, 97%, 98%, 99% или более гомологична одной из нуклеотидных последовательностей настоящего изобретения или их части. Объем настоящего изобретения также включает интервалы и величины идентичностей, которые являются промежуточными для вышеуказанных диапазонов (например, 70-90%-ная идентичность или 80-95%-ная идентичность). Так, например, в объем настоящего изобретения также входят интервалы значений идентичности с использованием комбинации любых из вышеупомянутых значений, указанных как верхний и/или нижний пределы. В еще одном предпочтительном варианте осуществления изобретения, белки SRT настоящего изобретения также обладают, по крайней мере, одной из SRT-активностей, описанных в настоящей заявке. Так, например, предпочтительный белок SRT настоящего изобретения включает аминокислотную последовательность, кодируемую нуклеотидной последовательностью, которая гибридизуется, например, гибридизуется в жестких условиях, с нуклеотидной последовательностью настоящего изобретения, и которая повышает резистентность или толерантность С. glutamicum к одному или нескольким вредным факторам или стрессам, вызываемым воздействием окружающей среды или химических веществ, или которая обладает одной или более активностями, указанными в таблице 1.
В других вариантах осуществления изобретения, белок SRT, в основном, гомологичен аминокислотной последовательности настоящего изобретения (например, четно пронумерованной последовательности SEQ ID NO: в Списке последовательностей) и сохраняет функциональную активность белка, в котором одна из аминокислотных последовательностей настоящего изобретения уже отличается по своей аминокислотной последовательности вследствие природного изменения или мутагенеза, как подробно описано выше в подразделе I. В соответствии с этим, в другом варианте осуществления изобретения, белок SRT представляет собой белок, содержащий аминокислотную последовательность, которая по крайней мере, примерно на 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59% или 60%, предпочтительно, по крайней мере, примерно на 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69% или 70%, более предпочтительно, по крайней мере, примерно на 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79% или 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89% или 90%, или 91%, 92%, 93%, 94%, и даже более предпочтительно, по крайней мере, примерно на 95%, 96%, 97%, 98%, 99% или более гомологична всей аминокислотной последовательности настоящего изобретения, и которая имеет, по крайней мере, одну из вышеописанных активностей SRT. Объем настоящего изобретения также включает интервалы и величины идентичностей, которые являются промежуточными для вышеуказанных диапазонов (например, 70-90%-ная идентичность или 80-95%-ная идентичность). Так, например, в объем настоящего изобретения также входят интервалы значений идентичности с использованием комбинации любых из вышеупомянутых значений, указанных как верхний и/или нижний пределы. В другом своем варианте, настоящее изобретение относится к полноразмерному белку С. qlutamicum, который является, в основном, гомологичным всей аминокислотной последовательности настоящего изобретения.
Биологически активные части белка SRT включают пептиды, содержащие аминокислотные последовательности, происходящие от аминокислотной последовательности белка SRT, например, аминокислотной последовательности SEQ ID NO, имеющей четный номер в Списке последовательностей или аминокислотной последовательности белка, гомологичной белку SRT, которая включает меньшее количество аминокислот, чем полноразмерный белок SRT, или полноразмерный белок, который является гомологичным белку SRT и обладает, по крайней мере, одной из активностей белка SRT. Обычно, биологически активные части (пептиды, например, пептиды, длина которых составляет 5, 10, 15, 20, 30, 35, 36, 37, 38, 39, 40, 50, 100 или более аминокислот) включают домен или мотив, имеющий, по крайней мере, одну из активностей белка SRT. Кроме того, другие биологически активные части, в которых были делетированы другие области данного белка, могут быть получены методами рекомбинантных ДНК и оценены на одну или несколько активностей, описанных в настоящей заявке. Предпочтительно, чтобы биологически активные части белка SRT включали один или несколько выбранных доменов/мотивов или их частей, обладающих биологической активностью.
Белки SRT, предпочтительно, продуцируют методами рекомбинантных ДНК. Так, например, молекулу нуклеиновой кислоты, кодирующую данный белок, клонируют в экспрессирующий вектор (как описано выше), этот экспрессирующий вектор вводят в клетку-хозяина (как описано выше), и белок SRT экспрессируется в данной клетке-хозяине. Затем белок SRT может быть выделен из клеток в соответствии с подходящей схемой очистки стандартными методами очистки белков. В качестве альтернативы рекомбинантной экспрессии, белок, полипептид или пептид SRT могут быть синтезированы стандартным методом химического пептидного синтеза. Кроме того, нативный белок SRT может быть выделен из клеток (например, эндотелиальных клеток), например, с использованием антитела против SRT, которое может быть продуцировано стандартными методами с использованием белка SRT настоящего изобретения или его фрагмента.
Настоящее изобретение также относится к химерным или гибридным белкам SRT. Используемый здесь термин «химерный белок» или «гибридный белок» SRT означает полипептид SRT, функционально присоединенный к не-SRT-полипептиду. Термин «полипептид SRT» означает полипептид, имеющий аминокислотную последовательность, соответствующую SRT, а термин «не-SRT-полипептид» означает полипептид, имеющий аминокислотную последовательность, соответствующую белку, который, в основном, не является гомологичным белку SRT, например, белку, который отличается от белка SRT и который происходит от того же самого или другого организма. Что касается гибридного белка, то термин «функционально присоединенный» означает, что полипептид SRT и не-SRT-полипептид присоединены друг к другу с сохранением рамки считывания. Указанный не-SRT-полипетид может быть присоединен к N-концу или к С-концу полипептида SRT. Так, например, в одном из вариантов осуществления изобретения, гибридным белком является гибридный белок GST-SRT, в котором последовательности SRT присоединены к С-концу последовательностей GST. Такие гибридные белки могут облегчать очистку рекомбинантных белков SRT. В другом варианте осуществления изобретения, указанным гибридным белком является белок SRT, содержащий гетерологичную сигнальную последовательность в его N-конце. В некоторых клетках-хозяевах (например, в клетках-хозяевах млекопитающих), экспрессия и/или секреция белка SRT может быть увеличена с использованием гетерологичной сигнальной последовательности.
Химерный или гибридный белок SRT настоящего изобретения, предпочтительно, продуцируют стандартными методами рекомбинантных ДНК. Так, например, ДНК-фрагменты, кодирующие другие полипептидные последовательности, лигируют с сохранением рамки считывания стандартными методами, например, путем использования затупленных или липких концов для лигирования, расщепления рестриктазами для получения соответствующих концов, достраивания липких концов, если это необходимо, обработки щелочной фосфатазой во избежание нежелательного присоединения, и ферментативного лигирования. В другом варианте осуществления изобретения, гибридный ген может быть синтезирован стандартными методами, включая использование автоматических ДНК-синтезаторов. Альтернативно, ПЦР-амплификация генных фрагментов может быть осуществлена с использованием якорных праймеров, которые образуют комплементарные выступающие участки между двумя последовательно расположенными генными фрагментами, которые могут быть затем подвергнуты отжигу и повторной амплификации для генерирования химерной генной последовательности (см, например, Current Protocols in Molecular Biology, eds. Ausubel et al., John Wiley & Sons, 1992). Кроме того, многие экспрессирующие векторы являются коммерчески доступными и уже кодируют часть гибрида (например, GST-полипептид).
SRT-кодирующая нуклеиновая кислота может быть клонирована в указанный экспрессирующий вектор, так, чтобы данная часть гибрида была присоединена к белку SRT с сохранением рамки считывания.
Гомологичные белки SRT могут быть генерированы путем мутагенеза, например, посредством отдельной точковой мутации или усечения белка SRT. Используемый здесь термин «гомолог» означает вариантную форму белка SRT, которая действует как агонист или антагонист активности белка SRT. Агонист белка SRT может, в основном, сохранять те же самые активности, или подсерию, биологических активностей белка SRT. Антагонист белка SRT может ингибировать одну или несколько активностей природной формы белка SRT, например, путем конкурентного связывания с ниже- или вышерасположенным членом SRT-системы, которая включает белок SRT. Так, например, белок SRT С. glutamicum и его гомологи настоящего изобретения могут повышать толерантность или резистентность С. glutamicum к одному или более стрессам, вызываемым химическими веществами или воздействием окружающей среды.
В альтернативном варианте осуществления изобретения, гомологи белка SRT могут быть идентифицированы путем скрининга комбинаторных библиотек мутантов, например, усеченных мутантов белка SRT, на агонистическую или антагонистическую активность белка SRT. В одном из вариантов осуществления изобретения, библиотеку разнообразных вариантов SRT генерируют путем комбинаторного мутагенеза на уровне нуклеиновой кислоты, где эти разнообразные варианты кодируются библиотекой разнообразных генов. Библиотека разнообразных вариантов SRT может быть создана, например, путем ферментативного лигирования смеси синтетических олигонуклеотидов в генные последовательности, так, чтобы вырожденная серия потенциальных SRT-последовательностей экспрессировалась как отдельные полипептиды, или альтернативно, как серия более крупных гибридных белков (например, для фагового отображения), содержащая серию последовательностей SRT. Существует ряд методов, которые могут быть использованы для продуцирования библиотек потенциальных SRT-гомологов из вырожденной олигонуклеотидной последовательности. Химический синтез вырожденной генной последовательности может быть осуществлен на автоматическом ДНК-синтезаторе, а затем этот синтетический ген лигируют в соответствующий экспрессирующий вектор. Использование вырожденной серии генов позволяет получить, в одной смеси, все последовательности, кодирующие желательную серию потенциальных SRT-последовательностей. Методы синтеза вырожденных олигонуклеотидов известны специалистам (см., например, Narang S.A. (1983) Tetrahedron 39: 3; Itakura et al. (1984) Annu. Rev. Biochem. 53: 323; Itakura et al. (1984) Science 198: 1056; Ike et al. (1983) Nucleic. Acid. Res. 11: 477.
Кроме того, библиотеки фрагментов кодирующего белка SRT могут быть использованы для генерирования разнообразных популяций фрагментов SRT в целях скрининга и последующего отбора гомологов белка SRT. В одном из вариантов осуществления изобретения, библиотека фрагментов кодирующей последовательности может быть создана путем обработки двухцепочечного ПЦР-фрагмента SRT-кодирующей последовательности нуклеазой в условиях, при которых происходит ник-разрыв примерно один раз на молекулу, денатурации двухцепочечной ДНК, ренатурации ДНК с образованием двухцепочечной ДНК, которая может включать смысловые/антисмысловые пары от различных продуктов ник-разрыва, удаления одноцепочечных частей из вновь образованных дуплексов путем обработки нуклеазой S1 и лигирования полученной библиотеки фрагментов в экспрессирующий вектор. Этим методом может быть получена библиотека экспрессируемых последовательностей, которая кодирует N-концевой, С-концевой и внутренние фрагменты белка SRT различных размеров.
Специалистам известны несколько методов скрининга генных продуктов комбинаторных библиотек, изготовленных посредством точковых мутаций или усечения, и скрининга кДНК-библиотек на присутствие генных продуктов, обладающих выбранными свойствами. Такие методы могут быть адаптированы для быстрого скрининга генных библиотек, генерированных путем комбинаторного мутагенеза SRT-гомологов. Наиболее широко используемые методы, которые являются подходящими для высокопроизводительного анализа, применяемого для скрининга больших библиотек генов, обычно предусматривают клонирование генной библиотеки в реплицируемые экспрессирующие векторы, трансформацию соответствующих клеток полученной библиотекой векторов и экспрессию комбинаторных генов в условиях, при которых детекция нужной активности облегчает выделение вектора, кодирующего данный ген, продукт которого был детектирован. Рекурсивный комбинированный мутагенез (REM) - новый метод, который увеличивает частоту функциональных мутантов в данных библиотеках, может быть использован в комбинации с скринирующими анализами для идентификации SRT-гомологов (Arkin & Yourvan (1992) PNAS 89: 7811-7815; Delgrave et al. (1993) Protein Engineering 6(3): 327-331).
В другом варианте осуществления изобретения, для анализа библиотеки разнообразных SRT могут быть использованы клеточные анализы с применением методов, хорошо известных специалистам.
D. Применения и способы настоящего изобретения
Молекулы нуклеиновой кислоты, белки, гомологи белка, гибридные белки, праймеры, векторы и клетки-хозяева могут быть использованы в одном или более из следующих методов: идентификации С. glutamicum и родственных организмов; картирования геномов организмов, родственных С. glutamicum; идентификации и локализации желаемых последовательностей С. glutamicum; эволюционные исследования; определения областей белка SRT, необходимых для его активности; модуляции активности белка SRT; модуляции активности SRT-пути и модуляции продуцирования клетками желаемого соединения, такого как химический продукт тонкого органического синтеза.
Молекулы нуклеиновой кислоты SRT настоящего изобретения имеют различные применения. Во-первых, они могут быть использованы для идентификации микроорганизма Corynebacterium glutamicum или близкородственного микроорганизма. Кроме того, они могут быть использованы для идентификации присутствия С. glutamicum или близкородственного микроорганизма в смешанной популяции микроорганизмов. Настоящее изобретение относится к нуклеотидным последовательностям ряда генов С. glutamicum; причем, путем зондирования экстрагированной геномной ДНК культуры отдельной или смешанной популяции микроорганизмов в жестких условиях зондом в области гена С. glutamicum, который является уникальным для этого организма, можно обнаружить присутствие указанного микроорганизма.
Хотя Corynebacterium glutamicum сам по себе является непатогенным, однако, он является родственным патогенному виду, такому как Corynebacterium diphtheriae. Corynebacterium diphtheriae является возбудителем дифтерии, быстро развивающегося, острого инфекционного заболевания с повышенной температурой, которое является как локальной, так и системной патологией. При этом заболевании, локальные поражения развиваются в верхних дыхательных путях и вызывают некротические поражения в эпителиальных клетках; указанные бациллы секретируют токсин, который распространяется через эти поражения на дистальные чувствительные ткани организма. Дегенеративные изменения, вызываемые ингибированием синтеза белка в тканях, которыми являются сердце, мышцы, ткани периферической нервной системы, надпочечники, почки, печень и селезенка, превращают это заболевание в системную патологию. Случаи заболевания дифтерией очень распространены во многих частях мира, включая Африку, Азию, Восточную Европу и ныне независимые государства бывшего Советского Союза. В настоящее время, начиная с 1990 г., эпидемия дифтерии в двух последних указанных регионах привела к гибели, по крайней мере, 5000 человек.
В одном из вариантов своего осуществления, настоящее изобретение относится к способу идентификации присутствия или активности Corynebacterium diphtheriae у данного индивидуума. Этот способ предусматривает обнаружение одной или более нуклеотидных или аминокислотных последовательностей настоящего изобретения (например, последовательностей, представленных как четно или нечетно, соответственно, пронумерованные последовательности SEQ ID NO: в Списке последовательностей) у данного индивидумма, и тем самым, обнаружение присутствия или активности Corynebacterium diphtheriae у данного индивидуума. С. glutamicum и С. diphtheriae являются родственными бактериями, и многие молекулы нуклеиновой кислоты и белка в С. glutamicum являются гомологичными нуклеиновой кислоте и белку С. diphtheriae, а поэтому они могут быть использованы для обнаружения С. diphtheriae у индивидуума.
Молекулы нуклеиновой кислоты и белка настоящего изобретения могут также служить в качестве маркеров для специфических областей данного генома. Это имеет важное значение не только для картирования генома, но также и для функциональных исследований белков С. glutamicum. Так, например, для идентификации области данного генома, с которой связывается ДНК-связывающий белок С. glutamicum, геном С. glutamicum должен быть гидролизован, а фрагменты инкубированы с ДНК-связывающим белком. Те фрагменты, которые связываются с этим белком, могут быть дополнительно зондированы молекулами нуклеиновых кислот настоящего изобретения, предпочтительно, легко детектируемыми метками; при этом, связывание такой молекулы нуклеиновой кислоты с данным фрагментом генома позволяет определить локализацию данного фрагмента на карте генома С. qlutamicum, и при многократном проведении такого зондирования с использованием различных ферментов, облегчает быструю идентификацию нуклеотидной последовательности, с которой связывается данный белок. Кроме того, молекулы нуклеиновых кислот настоящего изобретения могут быть достаточно гомологичными последовательностям родственных видов, так, что эти молекулы нуклеиновых кислот могут служить в качестве маркеров для построения геномной карты для родственных бактерий, таких как Brevibacterium lactofermentum.
Молекулы нуклеиновых кислот SRT настоящего изобретения могут быть также использованы для исследований эволюции и структуры белка. Процессы резистентности, в которых участвуют молекулы настоящего изобретения, используются клетками широкого ряда; и путем сравнения последовательностей молекул нуклеиновых кислот настоящего изобретения с последовательностями, кодирующими аналогичные ферменты от других организмов, может быть оценено эволюционное сходство этих организмов. Аналогичным образом, такое сравнение позволяет определить, какие области данной последовательности являются консервативными, а какие нет, поможет определить те области данного белка, которые являются главными для функционирования данного фермента. Этот способ определения имеет важное значение для биотехнологических исследований белков и может указывать на то, что данный белок может оказаться толерантным в отношении мутагенеза без потери своей функции.
Гены настоящего изобретения, например, ген, кодирующий LMRB (SEQ ID NO:1), или другой ген настоящего изобретения, кодирующий резистентность или толерантность к стрессам, вызываемым химическими соединениями или воздействием окружающей среды (например, резистентность к одному или более антибиотикам), может быть использован в качестве генетических маркеров для генетической трансформации (например, для переноса дополнительных генов или для разрушения предсуществующих генов) микроорганизма, такого как С. glutamicum или другого бактериального вида. Использование этих молекул нуклеиновой кислоты, которые были включены в данный трансгенный кластер (например, в плазмиду, фаг, фазмиду, фагмиду, транспозон или другой элемент нуклеиновой кислоты), позволяет проводить эффективный отбор микроорганизмов на основании признака, который позволяет данному микроорганизму выживать в условиях вредной или токсичной для него окружающей среды (например, в присутствии противомикробного соединения). Благодаря использованию одного или нескольких генов настоящего изобретения в качестве генетических маркеров можно повысить скорость и простоту конструирования и выделения тех микроорганизмов, которые имеют желаемые трансформированные признаки (например, продуцирование модулированных химических продуктов тонкого органического синтеза). Хотя использование генов настоящего изобретения для отбора трансформированных С. glutamicum и родственных бактерий является предпочтительным, однако, как описано в настоящей заявке, могут быть использованы гомологи (например, гомологи от других микроорганизмов), аллельные варианты или фрагменты гена, сохраняющие желаемую активность. Кроме того, для модуляции транскрипции данного гена, 5' и 3'-регуляторные элементы генов настоящего изобретения могут быть модифицированы как описано выше (например, путем замещения, инсерции, делеции или замены нуклеотидов более желательным генетическим элементом). Так, например, может быть применен вариант LMRB, в котором нуклеотидная последовательность в области от -1 до -200, расположенная с 5'-конца по отношению к старт-кодону, была модифицирована в целях модуляции (предпочтительно, повышения уровня) транскрипции и/или трансляции LMRB, а также могут быть применены конструкции, в которых ген настоящего изобретения (например, ген LMRB (SEQ ID NO:1) функционально присоединен к одному или нескольким регуляторным сигналам (например, к последовательностям, связывающимся с индуктором или репрессором), и которые могут быть использованы для модуляции экспрессии гена. Аналогичным образом могут быть использованы несколько копий гена настоящего изобретения (функциональных или инактивированных).
Другим применением генов настоящего изобретения (например, гена, кодирующего LMRB (SEQ ID NO:1) или другой ген резистентности к лекарственному средству или антибиотику) является разработка новых антибиотиков, которые обладают активностью, направленной против Corynebacteria и/или других бактерий. Так, например, ген настоящего изобретения может быть экспрессирован (или сверхэкспрессирован) в подходящем хозяине для генерирования микроорганизма с повышенной резистентностью к одному или нескольким лекарственным средствам или антибиотикам (в случае LMRB, в частности, линкозамидам, а особенно к линкомицину). Резистентный хозяин этого типа может быть затем использован для скрининга на химические соединения с бактериостатической и/или бактерицидной активностью, такие как новые соединения-антибиотики. Гены настоящего изобретения (например, ген LMRB) могут быть, в частности, использованы для идентификации новых антибиотиков, активность которых направлена против тех микроорганизмов, которые уже обладают резистентностью к стандартным антибиотикам.
Настоящее изобретение относится к способам скрининга молекул, которые модулируют активность белка SRT либо путем взаимодействия с самим белком или с его субстратом или партнером белка SRT по связыванию, либо путем модуляции транскрипции или трансляции молекулы нуклеиновой кислоты SRT настоящего изобретения. В указанных способах, микроорганизм, экспрессирующий один или несколько белков SRT настоящего изобретения, подвергают контакту с одним или несколькими тестируемыми соединениями, и оценивают действие каждого тестируемого соединения на активность или уровень экспрессии белка SRT.
Манипуляции с молекулами нуклеиновой кислоты SRT могут приводить к продуцированию белков SRT, функционально отличающихся от белков SRT дикого типа. Эти белки могут иметь повышенную эффективность или активность и могут присутствовать в клетке в большем количестве, чем это обычно имеет место, либо они могут иметь пониженную эффективность или активность. Целью таких манипуляций является увеличение жизнеспособности и активности клетки при воздействии на эту клетку химических стрессов и вредного воздействия окружающей среды, которыми обычно сопровождается крупномасштабное ферментативное культивирование. Таким образом, при увеличении активности или числа копий протеазы, регулируемой тепловым шоком, может быть увеличена способность данной клетки разрушать неправильно уложенные белки, которые могут так или иначе мешать нормальному функционированию клетки (например, в результате сохранения связывания с субстратами или кофакторами, несмотря на то, что этот белок не обладает соответствующей активностью по отношению к этим молекулам). То же самое относится к сверхэкспрессии или оптимизации активности одной или нескольких молекул шаперона, индуцированной тепловым или холодовым шоком. Эти белки способствуют правильной укладке растущих полипептидных цепей, и поэтому их повышенная активность или их присутствие должны способствовать увеличению процента правильно уложенных белков в данной клетке, что, в свою очередь, должно увеличивать общую метаболическую эффективность и жизнеспособность этих клеток в культуре. Сверхэкспрессия или оптимизация молекул-переносчиков, активированных осмотическим шоком, должна приводить к повышенной способности части клеток к поддержанию внутриклеточного гомеостаза, и тем самым, к увеличению своей жизнеспособности в культуре. Аналогичным образом, сверхпродуцирование или увеличение активности посредством мутагенеза белков, сообщающих клеткам резистентность к химическими стрессам различных типов (либо путем выведения вредных химических веществ из данной клетки, либо путем модификации этих химических веществ для снижения уровня вредных веществ), должно повышать приспособленность данного микроорганизма к условиям окружающей среды, содержащей вредные вещества (то есть, при крупномасштабном ферментативном культивировании), и тем самым оно может способствовать выживанию относительно большого числа клеток в такой культуре. Общим эффектом всех указанных стратегий мутагенеза является увеличение количества соединений, продуцирующих химические продукты тонкого органического синтеза в данной культуре, и тем самым, увеличение выхода, продуцирования и/или эффективности продуцирования одного или нескольких желаемых химических продуктов тонкого органического синтеза из данной культуры.
Вышеприведенный перечень стратегий мутагенеза для белков SRT в целях повышения выходов желаемого соединения не ограничивается этими стратегиями; и варианты этих стратегий мутагенеза будут очевидны для каждого специалиста. Благодаря механизмам этих стратегий мутагенеза, молекулы нуклеиновой кислоты и белка настоящего изобретения могут быть использованы для генерирования С. glutamicum или родственных штаммов бактерий, экспрессирующих мутантные молекулы нуклеиновой кислоты и белка SRT, в целях улучшения выхода, продуцирования и/или эффективности продуцирования нужного соединения. Нужное соединение может представлять собой любой природный продукт С. glutamicum, которым являются конечные продукты пути биосинтеза и промежуточные соединения природных путей метаболизма, а также молекулы, которые обычно не участвуют в метаболизме С. glutamicum, но которые продуцируются штаммом С. glutamicum настоящего изобретения.
Настоящее изобретение, кроме того, проиллюстрировано нижеследующими примерами, которые не должны рассматриваться как его ограничение. Содержание всех работ, патентных заявок, патентов, опубликованных патентных заявок, таблиц и списка последовательностей, цитируемых в настоящей заявке, вводится в настоящее описание посредством ссылки.
ИЛЛЮСТРАЦИЯ ПРИМЕРАМИ
Пример 1: Получение полной геномной ДНК Corynebacterium glutamicum ATCC 13032
Культуру Corynebacterium glutamicum (ATCC 13032) выращивали в течение ночи при 30°С при интенсивном встряхивании в среде BHI (Difco). Эти клетки собирали путем центрифугирования, супернатант отбрасывали и клетки ресуспендировали в 5 мл буфера-1 (5% исходного объема культуры; все указанные объемы были вычислены для 100 мл объема культуры). Состав буфера-1: 140,34 г/л сахарозы, 2,46 г/л MgSO4×7 Н2О, 10 мл/л раствора КН2РО4 (100 г/л, доведенного до рН 6,7 путем добавления КОН), 50 мл/л концентрата М12 (10 г/л (NH4)2SO4, 1 г/л NaCl, 2 г/л MgSO4×7 Н2О, 0,2 г/л CaCl2, 0,5 г/л дрожжевого экстракта (Difco), 10 мл/л смеси микроэлементов (200 мг/л FeSO4×Н2О, 10 мг/л ZnSO4×7 Н2О, 3 мг/л MnCl2×4 H2O, 30 мг/л Н3ВО3, 20 мг/л CoCl2×6 H2O, 1 мг/л NiCl2×6 H2O, 3 мг/л Na2MoO4×2 Н2О), 500 мг/л комплексобразующего агента (EDTA или лимонной кислоты), 100 мл/л смеси витаминов (0,2 мг/л биотина, 0,2 мг/л фолиевой кислоты, 20 мг/л п-аминобензойной кислоты, 20 мг/л рибофлавина, 40 мг/л пантотената кальция, 140 мг/л никотиновой кислоты, 40 мг/л гидрохлорида пиридоксола, 200 мг/л миоинозита). Затем к суспензии добавляли лизоцим до конечной концентрации 2,5 мг/мл. После инкубирования приблизительно в течение 4 ч при 37°С, клеточные стенки были разрушены и полученные протопласты собирали путем центрифугирования. Осадок один раз промывали 5 мл буфера-1 и один раз 5 мл ТЕ-буфера (10 мМ Трис-HCl, 1 мМ EDTA, pH 8). Затем осадок ресуспендировали в 4 мл ТЕ-буфера и добавляли 0,5 мл раствора ДСН (10%) и 0,5 мл раствора NaCI (5M). После добавления протеиназы К до конечной концентрации 200 мкг/мл, суспензию инкубировали прибл. в течение 18 ч при 37°С. ДНК очищали путем экстракции фенолом, фенолом-хлороформом-изоамиловым спиртом и хлороформом-изоамиловым спиртом с использованием стандартных процедур. Затем, ДНК осаждали путем добавления 1/50 объема ЗМ ацетата натрия и 2 объемов этанола, после чего инкубировали в течение 30 минут при -20°С и центрифугировали в течение 30 минут при 12000 об/мин в высокоскоростной центрифуге с использованием ротора SS34 (Sorvall). Затем, ДНК растворяли в 1 мл ТЕ-буфера, содержащего 20 мкг/мл РНКазы А и диализовали при 4°С против 1000 мл ТЕ-буфера, по крайней мере, в течение 3 часов. В течение этого времени, буфер меняли 3 раза. К аликвотам 0,4 мл диализованного раствора ДНК добавляли 0,4 мл 2М LiCl и 0,8 мл этанола. После инкубирования в течение 30 минут при -20°С, ДНК собирали путем центрифугирования (13000 об/мин, Biofuge Fresco, Heraeus, Hanau, Germany). Осажденную ДНК растворяли в ТЕ-буфере. ДНК, полученная в результате проведения этой процедуры, может быть использована для любых целей, включая Саузерн-блоттинг или конструирование геномных библиотек.
Пример 2: Конструирование геномных библиотек Corynebacterium glutamicum ATCC 13032 в Esherichia coli
С использованием ДНК, полученной как описано в Примере 1, библиотеки космид и плазмид были сконструированы с помощью известных и хорошо разработанных методов (см. например, Sambrook J. et al.,(1989), «Molecular Cloning: A Laboratory Manual», Cold Spring Harbor Laboratory Press, Ausubel F.M. et al. (1994) "Current Protocols in Molecular Biology", John Wiley & Sons).
Может быть использована любая плазмида или космида. В частности, были использованы плазмиды pBR322 (Sutcliffe J.G. (1979) Proc. Natl. Acad. Sci. USA, 75: 3737-3741); pACYC177 (Change & Cohen (1978) J.Bacteriol. 134:1141-1156), плазмиды серии pBS (pBSSK+, pBSSK- и другие; Stratagene, LaJolla, USA) или космиды, такие как SuperCosl (Stratagene, LaJolla, USA) или Lorist6 (Gibson, T.J., Rosenthal A. & Waterson, R.H. (1987) Gene 53: 283-286. Библиотеки генов, в частности, для использования в С. glutamicum, могут быть сконструированы с использованием плазмиды pSL109 (Lee, H.S. & A.J.Sinskey (1994) J. Microbiol. Biotechnol. 4:256-263).
Пример 3: ДНК-секвенирование и компьютерный функциональный анализ
Геномные библиотеки, описанные в Примере 2, были использованы для секвенирования ДНК в соответствии со стандартными методами, в частности, методом обрыва цепи с использованием секвенаторов АВ1377 (см., например, Fleischman, R.D. et al.(1995) «Whole-genome Random Sequencing and Assembly of Haemophilus Influenzae Rd.», Science, 269: 496-512). Были использованы секвенирующие праймеры со следующими нуклеотидными последовательностями 5'-GGAAACAGTATGACCATG-3' или 5'-GTAAAACGACGGCCAGT-3'.
Пример 4: Мутагенез in vivo
In vivo-мутагенез Corynebacterium glutamicum может быть осуществлен путем переноса плазмизной ДНК (или другого вектора) из E.coli или других микроорганизмов (например, Bacillus spp. или дрожжей, таких как Saccharomyces cerevisiae), у которых была нарушена способность к сохранению целостности своей генетической информации. Обычные штаммы-мутанты имеют мутации в генах системы репарации ДНК (например, mutHLS, mutD, mutT, и т.п. см. Rupp, W.D. (1996), DNA repair mechanisms, in: Escherichia coli and Salmonella, p.2277-2294, ASM: Washington). Эти штаммы хорошо известны каждому специалисту. Использование таких штаммов проиллюстрировано, например, Greener, A. & Callahan, M (1994) Strategies 7: 32-34.
Пример 5: Перенос ДНК от Escherichia coli в Corynebacterium glutamicum
Некоторые виды Corynebacterium и Brevibacterium содержат эндогенные плазмиды (как, например, рНМ1519 или pBLl), которые автономно реплицируются (см., например, обзор Martin, J.F. et al., (1987) Biotechnology, 5:137-146). Челночные векторы для Escherichia coli и Corynebacterium glutamicum могут быть легко сконструированы с использованием стандартных векторов для E.coli (Sambrook J. et al.,(1989). Molecular Cloning: A laboratory manual, Cold Spring Harbor Laboratory Press, Ausubel F.M. et al. (1994) «Current Protocols in Molecular Biology», John Wiley & Sons), в которые были добавлены сайт инициации репликации и подходящий маркер от Corynebacterium glutamicum. Указанные сайты инициации репликации, предпочтительно, берутся из эндогенных плазмид, выделенных из видов Corynebacterium и Brevibacterium. Конкретными используемыми маркерами трансформации для этих видов являются гены резистентности к канамицину (такие как гены, происходящие от транспозонов Тn5 или Тn903) или к хлорамфениколу (Winnacker, E.L. (1987) «From Genes to Clones - Introduction to Gene Technology», VCH, Weinheim). В литературе приводятся многочисленные примеры конструирования челночных векторов широкого ряда, которые реплицируются как в E.coli, так и в С. glutamicum и которые могут быть использованы для многих целей, включая сверхэкспрессию генов (см., например, Yoshihama et al., (1985) J.Bacteriol. 162: 591-597; Martin, J.F. et al., (1987) Biotechnology, 5: 137-146 и Eikmanns, B.J. et al. (1991) Gene, 102:93-98).
С использованием стандартных методов можно клонировать желаемый ген в любом из вышеописанных челночных векторов и вводить такой гибридный вектор в штаммы Corynebacterium glutamicum. Трансформация С. glutamicum может быть осуществлена путем трансформации протопластов (Kastsumata, R. et al.(1984), J. Bacteriol. 159: 306-311), электропорации (Liebl, Е. et al., (1989) FEMS Microbiol. Letters, 53: 399-303), а в случаях, когда используются специальные векторы, эта трансформация может быть также осуществлена путем конъюгирования (как описано, например, Schafer, A. et al.(1990) J. Bacteriol. 172: 1663-1666). Челночные векторы для С. glutamicum могут быть также перенесены в E.coli путем получения плазмидной ДНК из С. glutamicum (стандартными хорошо известными методами) и трансформации их в E.coli. Такая стадия трансформации может быть осуществлена стандартными методами, но, предпочтительно, использовать Mcr-дефицитный штамм E.coli, такой как NM522 (Gough & Murray (1983) J.Mol.Biol. 166: 1-19).
Гены могут быть сверхэкспрессированы в штаммах С. glutamicum с использованием плазмид, которые содержат pCGl (патент США №4617267) или его фрагменты, и необязательно ген резистентности к канамицину от TN903 (Grindley, N.D. & Joyce, С.М. (1980) Proc.Natl. Acad. Sci., USA, 77(12): 7176-7180). Кроме того, гены могут быть сверхэкспрессированы в штаммах С. glutamicum с использованием плазмиды pSL109 (Lee, H.-S. & A.J.Sinskey (1994) J. Microbiol. Biotechnol. 4: 256-263).
Помимо использования репликативных плазмид, сверхэкспрессия генов может быть также достигнута путем их интеграции в геном. Геномная интеграция в С. glutamicum или другие виды Corynebacterium или Brevibacterium может быть осуществлена хорошо известными методами, такими как гомологичная рекомбинация с геномной(ыми) областью(ями), интеграция, опосредованная рестриктазами (REMI) (см., например, патент DE 19823834) или путем использования транспозонов. Может быть также осуществлена модуляция активности желаемого гена путем модификации регуляторных областей (например, промотора, репрессора и/или энхансера), путем модификации, инсерции или делеции последовательностей с использованием сайт-направленных методов (таких как гомологичная рекомбинация) или методов, основанных на случайных событиях (таких как транспозонный мутагенез или REMI). Нуклеотидные последовательности, которые функционируют как терминаторы транскрипции, могут быть также встроены со стороны 3'-конца по отношению к кодирующей области одного или несколько генов настоящего изобретения; и такие терминаторы хорошо известны специалистам и описаны, например, у Winnacker, E.L. (1987) "From Genes to Clones - Introduction to Gene Technology, VCH, Weinheim.
Пример 6: Оценка экспрессии мутантного белка
Оценку активности мутированного белка в трансформированной клетке-хозяине проводят исходя из того факта, что этот мутантный белок экспресиируется так же и в таком же количестве, как и белок дикого типа. Метод, используемый для определения уровня транскрипции данного мутантного гена (индикатора количества мРНК, необходимого для трансляции с образованием генного продукта), предусматривает проведение Нозерн-блот-анализа (см. например, Ausubel et al., (1988) Current Protocols in Molecular Biology, Wiley; New York), в котором праймер, сконструированный для связывания желаемого гена, метят детектируемой меткой (обычно радиоактивной или хемилюминесцентной), так, чтобы при экстракции суммарной РНК из культуры данного микроорганизма, ее электрофорезе на геле, переносе на стабильную матрицу и инкубировании с этим зондом, связывание и количество связывающегося зонда указывало на присутствие, а также на количество мРНК данного гена. Эта информация указывает на степень транскрипции данного мутантного гена. Суммарная клеточная РНК может быть получена из Corynebacterium glutamicum несколькими методами, хорошо известными специалистам, такими как методы, описанные Bormann, E.R. et al.,(1992) Mol. Microbiol. 6: 317-326.
Для определения присутствия или относительного количества белка, транслированного из этой мРНК, могут быть использованы стандартные методы, такие как Вестерн-блот-анализ (см. например, Ausubel et al., (1988) Current Protocols in Molecular Biology, Wiley; New York). В этом способе суммарные клеточные белки экстрагируют, разделяют посредством гель-электрофореза, переносят на матрицу, такую как нитроцеллюлоза, и инкубируют с зондом, таким как антитело, которое специфически связывается с нужным белком. Этот зонд обычно метят хемилюминесцентной или колориметрической меткой, которая может быть легко детектирована. Присутствие и количество наблюдаемой метки указывает на присутствие и количество желаемого мутантного белка в данной клетке.
Пример 7: Рост генетически модифицированной Corynebacterium glutamicum - среды и условия культивирования
Генетически модифицированную бактерию Corynebacterium культивировали в синтетических или природных культуральных средах. Существует ряд различных хорошо известных и легко доступных культуральных сред для Corynebacterium (Lieb et al. (1989) Appl. Microbiol. Biotechnol., 32:205-210; von der Osten et al. (1998) Biotechnology Letters, 11:11-16; патент DE 4120867; Liebl (1992) «The Genus Corynebacterium: The Procaryotes», Volume II, Balows, A. et al., eds. Springer-Verlag). Эти среды состоят из одного или нескольких источников углерода, источников азота, неорганических солей, витаминов и микроэлементов. Предпочтительными источниками углерода являются сахара, такие как моно-, ди- или полисахариды. Так, например, превосходными источниками углерода являются глюкоза, фруктоза, манноза, галактоза, рибоза, сорбоза, рибулоза, лактоза, мальтоза, сахароза, раффиноза, крахмал или целлюлоза. Доставка сахара в эти среды может также осуществляться с помощью комплексных соединений, таких как меласса или другие побочные продукты, образованные в результате рафинирования сахара. Эти соединения могут быть также предпочтительными для доставки смесей различных источников углерода. Другими возможными источниками углерода являются спирты и органические кислоты, такие как метанол, этанол, уксусная кислота или молочная кислота. Источниками азота обычно являются органические или неорганические соединения азота или материалы, содержащие эти соединения. Примерами источников азота являются газообразный аммиак или аммиачные соли, такие как NH4Cl или (NH4)2SO4, NH4OH, нитраты, мочевина, аминокислоты или комплексные источники азота, такие как кукурузный экстракт, соевая мука, соевый белок, дрожжевой экстракт, мясной экстракт и др. Соединениями неорганических солей, которые могут быть включены в эти среды, являются хлориды, фосфаты или сульфаты кальция, магния, натрия, кобальта, молибдена, калия, марганца, цинка, меди и железа. Хелатообразующие соединения могут быть добавлены в данную среду для поддержания ионов металлов в растворе. Особенно ценными хелатообразующими соединениями являются дигидроксифенолы, такие как катехин или протокатехуат, или органические кислоты, такие как лимонная кислота. Эта среда также обычно содержит другие факторы роста, такие как витамины или стимуляторы роста, примерами которых являются биотин, рибофлавин, тиамин, фолиевая кислота, никотиновая кислота, пантотенат и пиридоксин. Факторы роста и соли часто происходят от компонентов комплексных сред, таких как дрожжевой экстракт, мелассы, кукурузный экстракт и другие. Точный состав соединений в средах в высокой степени зависит от данного конкретного эксперимента и определяется отдельно для каждого конкретного случая. Информацию относительно оптимизации сред можно найти в руководстве «Applied. Microbiol. Physiology», A Practical Approach (eds., P.M.Rhodes, P.F.Stanbury, IRL Press (1997) pp.53-73, ISBN 0 199635773). Культуральные среды могут быть также закуплены у коммерческих поставщиков, например, стандартная среда 1 (Merck) или BHI (экстракт из ядер зерна, DIFCO) или т.п.
Все компоненты сред стерилизуют, либо путем нагревания (20 минут при 1,5 бар и 121°С), либо путем стерильной фильтрации. Эти компоненты могут быть стерилизованы вместе, либо, если это необходимо, отдельно. Все компоненты сред могут присутствовать в начале культивирования, либо они могут, но необязательно, добавляться непрерывно или периодически.
Условия культивирования определяют отдельно для каждого эксперимента. Температура должна составлять в пределах от 15°С до 45°С. Эта температура может быть постоянной, либо она может меняться в процессе эксперимента. рН этой среды должен составлять в пределах от 5 до 8,5, а предпочтительно, примерно 7,0, и может поддерживаться путем добавления в данную среду буферов. Примером буфера, используемого для этих целей, является калийфосфатный буфер. Альтернативно или одновременно могут быть использованы синтетические буферы, такие как MOPS, HEPES, ACES и другие. В процессе культивирования может также поддерживаться постоянный рН культуры путем добавления NaOH или NH4OH. Если используются компоненты комплексной среды, такой как дрожжевой экстракт, то необходимость в дополнительных буферах может быть уменьшена благодаря тому, что многие комплексные соединения имеют высокую буферную емкость. Если для культивирования микроорганизмов используется ферментер, то рН может также регулироваться добавлением газообразного аммиака.
Время инкубирования обычно составляет от нескольких часов до нескольких дней. Это время выбирают так, чтобы в данном бульоне накапливалось максимальное количество продукта. Описанные здесь эксперименты по культивированию могут быть осуществлены в ряде сосудов, таких как микротитрационные планшеты, стеклянные пробирки, стеклянные колбы или стеклянные или металлические ферментеры различных размеров. Для скрининга большого числа клонов, микроорганизмы следует культивировать в микротитрационных планшетах, в стеклянных пробирках или в колбах-шейкерах, либо с перегородками, либо без них. Предпочтительно, используют 100 мл-колбы-шейкеры, наполненные на 10% (по объему) требуемой культуральной средой. Эти колбы должны встряхиваться на роторном шейкере (амплитуда 25 мм) со скоростью в пределах 100-300 об/мин. Потери от выпаривания могут быть уменьшены путем поддержания влажной атмосферы; альтернативно, должна быть осуществлена математическая коррекция потерь от выпаривания.
При тестированиии генетически модифицированных клонов должен быть также протестирован немодифицированный контрольный клон или контрольный клон, содержащий основную плазмиду без каких-либо вставок. Среду инокулируют до ОП600=0,5-1,5 с использованием клеток, выращенных на планшетах с агаром, таких как планшеты СМ (10 г/л глюкозы, 2,5 г/л NaCl, 2 г/л мочевины, 10 г/л полипептона, 5 г/л дрожжевого экстракта, 5 г/л мясного экстракта, 22 г/л агара, рН 6,8 с 2М NaOH), которые инкубируют при 30°С. Инокуляцию данной среды осуществляют либо путем введения солевой суспензии клеток С. glutamicum из планшетов СМ, либо путем добавления жидкой предварительной культуры данной бактерии.
Пример 8 - In vitro-анализ функции мутантных белков
Определение активностей и кинетических параметров ферментов является хорошо известной процедурой. Эксперименты по определению активности любого данного модифицированного фермента должны быть специально адаптированы для специфической активности фермента дикого типа, что может быть легко осуществлено любым специалистом. Общее описание ферментов, а также конкретные детали, касающиеся структур, кинетики, принципов, методов, применений и примеров определения многих ферментативных активностей, можно найти, например, в следующих работах: Dixon, M. & Webb, E.C. (1979) Enzymes. Longmans: London; Fersht (1985) Enzyme Structure and Mechanism. Freeman: New York; Walsh (1979) Enzymatic Reaction Mechanisms. Freeman: San Francisco; Price N.C. Stevens, L. (1982) Fundamentals of Enzymology. Oxford Univ. Press: Oxford; Boyer, P.D. ed. (1983) The Enzymes, 3rd ed. Academic Press: New York; Bisswanger H. (1994) Enzymkinetik, 2nd ed. VCH: Weinheim (ISBN 3527300325); Bergmeyer, H.U., Bergmeyer, J. Graβl, M., eds. (1983-1986) Methods of Enzymatic Analysis, 3rd ed. vol.I-XII, Verlag Chemie: Weinheim; and Ullmann's Encyclopedia of Industrial Chemistry (1987) vol.A9, «Enzymes» VCH: Weinheim, p.352-363.
Активность белков, связывающихся с ДНК, может быть определена несколькими хорошо разработанными методами, такими как анализы на сдвиг полосы ДНК (так называемые анализы на замедление подвижности в геле). Действие таких белков на экспрессию других молекул может быть измерено с помощью анализов на присутствие репортерного гена (такого как анализ, описанный в работе Kolmar H. et al. (1995) EMBO J. 14: 3895-3904 и в цитируемых там работах). Системы тестирования с использованием репортерного гена хорошо известны и разработаны для их применения как для прокариотических, так и для эукариотических клеток с использованием ферментов, таких как бета-галактозидаза, флуоресцентный белок, излучающий в зеленом диапазоне света, и некоторые другие ферменты.
Определение активности мембрано-транспортных белков может быть осуществлено известными методами, описанными Gennis, R.B. (1989) «Pores, Channels and Transporters», Biomembranes, Molecular Structure and Function, Springer: Heidelberg, p.85-137; 199-234 и 270-322.
Пример 9: Анализ влияния мутантного белка на продуцирование желаемого продукта
Действие генетической модификации в С. glutamicum на продуцирование желаемого соединения (такого как аминокислота) может быть оценено путем культивирования модифицированного микроорганизма в подходящих условиях (таких как описаны выше) и анализа среды и/или клеточного компонента на повышенное продуцирование желаемого продукта (то есть, аминокислоту). Такие методы анализа хорошо известны любому специалисту и ими являются спектроскопия, тонкослойная хроматография, методы окрашивания различных типов, ферментные и микробиологические методы и аналитическая хроматография, такая как высокоэффективная жидкостная хроматография (см., например, Ullmann, Encyclopedia of Industrial Chemistry (1987) vol.A2, p.89-90 and p.443-613, VCH: Weinheim, (1985); Fallon A. et al. (1987) «Applications of HPLC in Biochemistry»: Laboratory Techniques in Biochemistry and Molecular Biology, vol. 17; Rehm et al. (1993) Biotechnology, vol.3, Chapter III: «Product recovery and purification», page 469-714, VCH: Weinheim; Belter, P.A. et al. (1988) Bioseparations: downstream processing for biotechnology, John Wiley and Sons; Kennedy J.F. & Cabral J.M.S. (1992) Recovery processes for biological materials, John Wiley & Sons; Shaiewitz, J.A. & Henry, J.D. (1988) Biochemical separations, Ullmann's Encyclopedia of Industrial Chemistry, vol. B3, Chapter 11, page 1-27, VCH: Weinheim & Dechow, F.J. (1989) Separation and purification techniques in biotechnology, Noyes Publications).
Помимо оценки конечного продукта ферментации можно также проанализировать другие компоненты путей метаболизма, используемых для продуцирования желаемого соединения, такого как промежуточные соединения и побочные продукты, в целях определения полного выхода, уровня и/или эффективности продуцирования данного соединения. Методами анализа являются измерения уровней питательных веществ в данной среде (например, сахаров, углеводородов, источников азота, фосфатов и других ионов), измерения состава и роста биомассы, анализ продуцирования общих метаболитов биосинтетических путей и измерение газов, продуцируемых во время ферментации. Стандартные методы для этих измерений описаны в Applied. Microbiol. Physiology, A Practical Approach, P.M. Rhodes & P.F. Stanbury, eds., IRL Press p. 103-129, 131-163 and 165-192 (ISBN 0 199635773) и в цитируемых там работах.
Пример 10: Очистка желаемого продукта из культуры С. glutamicum
Выделение желаемого продукта из клеток С. glutamicum или супернатанта вышеописанной культуры может быть осуществлено различными методами, хорошо известными специалистам. Если желаемый продукт не секретируется из клеток, то эти клетки могут быть собраны из культуры путем центрифугирования на малой скорости, а затем они могут быть лизированы стандартными методами, такими как механическое разрушение или обработка ультразвуком. Клеточный дебрис удаляют путем центрифугирования, а фракцию супернатанта, содержащую растворимые белки, оставляют для последующего выделения желаемого соединения. Если данный продукт секретируется из клеток С. glutamicum, то эти клетки удаляют из культуры путем центрифугирования на малой скорости, а фракцию супернатанта оставляют для последующей очистки.
Фракцию супернатанта, полученную в любом методе очистки, подвергают хроматографии с использованием подходящей смолы, где либо желаемая молекула удерживается на хроматографической смоле, тогда как многие примеси в образце не удерживаются, либо где примеси удерживаются на данной смоле, а образец - нет. Такие стадии хроматографии могут быть повторены, если это необходимо, с использованием той же самой или других хроматографических смол. Любой специалист может самостоятельно выбрать смолы для хроматографии и их наиболее эффективное применение для очистки конкретной молекулы. Очищенный продукт может быть концентрирован путем фильтрации или ультрафильтрации и помещен на хранение при температуре, при которой этой продукт является максимально стабильным.
Существует широкий ряд методов очистки, известных специалистам, и эти методы не ограничиваются вышеуказанным методом. Такие методы очистки описаны, например. Bailey, J.E. & Ollis, D.F. Biochemical Engineering Fundamentals, McGraw-Hill: New York (1986).
Идентичность и чистота выделенных соединений может быть оценена стандартными методами. Такими методами является высокоэффективная жидкостная хроматография (ВЭЖХ), спектроскопические методы, методы окрашивания, тонкослойная хроматография, NIRS, ферментативный анализ или микробиологические методы. Описание этих методов анализа см. Patek et al. (1994), Appl. Environ. Microbiol. 60: 133-140; Malakhova et al. (1996) Biotekhnologiya 11: 27-32; и Schmidt et al., (1998) Bioprocess Engineer. 19: 67-70. Ullmann's Encyclopedia of Industrial Chemistry (1996) vol.A27, VCH: Weinheim, p.89-90, p.521-540, p.540-547, p.559-566, p.575-581 и р.581-587; Michal G. (1999) Biochemical pathway: An Atlas of Biochemistry and Molecular Biology, John Wiley & Sons; Fallon A. et al. (1987) «Applications of HPLC in Biochemistry»: Laboratory Techniques in Biochemistry and Molecular Biology, vol. 17.
Пример 11: Клонирование гена Corynebacterium qlutamicum, кодирующего резистентность к линкомицину, методом с использованием репортерного гена А. Идентификация гена, кодирующего белок LMRB
Плазмиду pSL 130 конструировали путем лигирования промоторной области асеВ (расеВ) С. glutamicum (Kirn H.J. et al. (1997) J. Microbiol. Biotechnol. 7: 287-292) в полининкер гибридного вектора pRS415 с Iac-опероном, у которого отсутствует промотор (Simon, R.W. et al.(1987) Gene 53: 85-96). Плазмиду pSL145 конструировали путем встраивания полученной области paceB-lac в клонирующий вектор рАСУС184 Е.coli. DH5αF' E.coli трансформировали pSL145 и полученный штамм использовали в качестве хозяина для скрининга геномной библиотеки С. glutamicum (в pSL109).
Трансформанты скринировали путем культивирования на агаровой среде, содержащей 5-бром-4-хлор-3-индолил-бета-D-галактопиранозид (X-Gal). Белую колонию, содержащую ДНК, влияющую на экспрессию lacZ, отбирали для последующего анализа. Было обнаружено, что этот клон содержит фрагмент размером 4 т.п.н. из библиотеки генов. Субклоны конструировали в pSL109 и идентифицировали субклон, который сохранял фенотип белых бактерий на чашке с X-Gal. Было обнаружено, что этот субклон содержит BamHI-XhoI-фрагмент размером 2,6 т.п.н. (плазмида pSL149-5). Этот фрагмент секвенировали и идентифицировали как ген, кодирующий мембранный белок (ген LMRB).
1442 нуклеотидов кодирующей последовательности гена LMRB кодируют полипептид из 481 аминокислотных остатков с высоким процентным содержанием гидрофобных аминокислот. Поиск в Genbank позволил установить, что белок LMRB на 40% идентичен белковому продукту гена ImrB от Bacillus subtilis (Genbank Accession AL009126, TREMBL Accession P94422), как было определено с использованием анализа CLUSTAL W (с использованием стандартных параметров).
Белок LMRB содержит мотив последовательности 158-А-Р-А-L-G-P-T-L-S-G-167 (SEQ ID N0:301), который подобен известному консенсусному мотиву белка резистентности ко множественным лекарственным средствам G-X-X-X-G-P-X-X-G-G (SEQ ID NO:302) Paulsen I.Т., & Skurray R.A. (1993) Gene 124: 1-11). Поэтому белок LMRB был классифицирован как белок резистентности к лекарственному средству.
В. In vivo-анализ функции ImrB
Ген ImrB был сверхэкспрессирован в С. glutamicum ASO19E12 (Kim H.J. et al. (1997) J. Microbiol. Biotechnol. 7: 287-292) с использованием плазмиды pSL149-5, описанной выше.
Разрушение гена LMRB осуществляли с использованием вектора PsL18-lmrB. Этот вектор конструировали следующим образом: внутренний фрагмент гена LMRB амплифицировали с помощью ПЦР в стандартных условиях с использованием праймеров 5'-CTCCAGGATTGCTCCGAAGG-3' (SEQ ID NO:303) и 5'-CACAGTGGTTGACCACTGGC-3' (SEQ ID NO:304). Полученный ПЦР-продукт обрабатывали ДНК-полимеразой Т7 и полинуклеотид-киназой Т7 и клонировали в SmaI-сайт плазмиды pSL18 (Kim Y.H. & H.-S. Lee. (1996) J.Microbiol. Biotechnol. 6: 315-320). Разрушение гена LMRB в С. glutamicum ASО19E12 осуществляли путем конъюгирования как описано ранее (Schwarzer & Punler (1991) Bio/Technology 9: 84-87).
Клетки С. glutamicum, трансформированные pSL149-5, обнаруживали такую же, как и нетрасформированные клетки, резистентность к эритромицину, пенициллину G, тетрациклину, хлорамфениколу, спектиномицину, налидиксиновой кислоте, гентамицину, стрептомицину, этидийбромиду, карбонилцианид-м-хлорфенилгидразону (СССР) и додецилсульфату натрия. Однако, при этом, наблюдались значительные различия в резистентности трансформированных и нетрасформированных клеток к линкомицину.
Было обнаружено, что клетки С. glutamicum, сверхэкспрессирующие LMRB, способны расти в присутствии 20 мкг/мл линкомицина. В противоположность этому, клетки, которые не экспрессируют избыточного LMRB (или клетки, несущие разрушение гена LMRB), не способны расти на среде с агаром, содержащим 5 мкг/мл линкомицина. Этот эффект был явно заметным в жидкой культуре. Сверхэкспрессия LMRB приводила к 9-кратному увеличению резистентности (по сравнению с диким типом) к линкомицину, а разрушение гена LMRB приводило к снижению резистентности (28% от дикого типа) к этому антибиотику.
Пример 12: Анализ последовательностей генов настоящего изобретения
Сравнение последовательностей и определение процента гомологии между двумя последовательностями являются хорошо известными методами, и они могут быть осуществлены с использованием математического алгоритма, такого как алгоритм Karlin & Altschul (1990) Proc.Natl. Acad. Sci., USA, 87: 2264-68, модифицированный как описано Karlin & Altschul (1993) Proc.Natl. Acad. Sci., USA, 90: 5873-77. Такой алгоритм вводят в программы NBLAST и XBLAST (версия 2.0) Altschul et al. (1990) J.Mol.Biol. 215: 403-10. Поиск нуклеотидов BLAST может быть осуществлен с использованием программы NBLAST, оценка =100, длина слова =12 для получения нуклеотидных последовательностей, гомологичных молекулам нуклеиновой кислоты SRT настоящего изобретения. Поиск белков BLAST может быть осуществлен с использованием программы XBLAST, оценка =50, длина слова =3 для получения аминокислотных последовательностей, гомологичных молекулам белка SRT настоящего изобретения. Для получения первичных последовательностей с «пробелами» в целях их сопоставления, может быть использована программа «Gapped BLAST», описанная Altschul et al. (1997) Nucleic. Acids Res. 25(17): 3389-3402. При использовании программ BLAST и Gapped BLAST, каждому специалисту известно, каким образом можно оптимизировать параметры данной программы (например, XBLAST и NBLAST) для анализа специфической последовательности.
Другим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм Meyers & Miller (1988) Comput.Appi.Biosci. 4: 11-17). Такой алгоритм вводят в программу ALIGN (версия 2.0), которая является частью программного пакета для сопоставления последовательности GCG. При использовании программы ALIGN для сравнения аминокислотных последовательностей, может быть использована таблица весовых остатков РАМ120, штрафа за длину пробела =12 и штрафа за пробел =4. Специалистам известны и другие алгоритмы для анализа последовательностей, и ими являются ADVANCE и ADAM, описанные Torelli & Robotti (1994) Comput. Appl. Biosci. 10: 3-5, и FASTA, описанный Pearson & Lipmann (1988) P.N.A.S. 85: 2444-8.
Процент гомологии между двумя аминокислотными последовательностями может быть также определен с использованием программы GAP, имеющейся в программном пакете GCG (доступном на сайте http:/www.gcg.com), с использованием либо матрицы Blosum 62, либо матрицы РАМ250, и веса пробела 12, 10, 8, 6 или 4, и веса длины 2, 3 или 4. Процент гомологии между двумя нуклеотидными последовательностями может быть определен с использованием программы GAP в программном пакете GCG с использованием стандартных параметров, таких как вес пробела =50, а вес длины =3.
Сравнительные анализы генных последовательностей настоящего изобретения с последовательностями, присутствующими в Genbank, осуществляли с использованием хорошо известных методов (см., например, Bexevanis & Ouellette, eds., (1998) Bioinformatics: A Practic Guide to the Analysis of Genes and Proteins. John Wiley & Sons; New York). Генные последовательности настоящего изобретения сравнивали с генами, имеющимися в Genbank, в три стадии. В первой стадии осуществляли анализ BLASTN (например, анализ для локального сравнения) для сравнения каждой последовательности настоящего изобретения с нуклеотидными последовательностями, присутствующими в Genbank, и 500 наилучших совпадений оставляли для последующего анализа. Для этих лучших 500 совпадений проводили последующий поиск с использованием FASTA (например, комбинированный анализ для локального и глобального сопоставления первичных последовательностей, в котором сопоставлялись ограниченные области последовательностей). Затем, каждую генную последовательность настоящего изобретения подвергали глобальному сравнению с каждым из наилучших трех совпадений в FASTA с использованием программы GAP в программном пакете GCG (с использованием стандартных параметров). Для получения правильных результатов, длины последовательностей, полученных из Genbank, корректировали на длину запрашиваемых последовательностей хорошо известными способами. Результаты этих анализов представлены в Таблице 4. Полученные данные являются идентичными данным, которые были бы получены, если бы (глобальный) анализ GAP был отдельно осуществлен для каждого из генов настоящего изобретения по сравнению с каждым из генов, имеющимся в Genbank, но для этого потребовалось бы значительно меньше компьютерного времени, чем для (глобального) анализа с обширной базой данных GAP. Последовательности настоящего изобретения, для каждой из которых не проведено сопоставление выше значения отсечки, можно определить в Таблице 4 по отсутствию информации. Кроме того, следует учесть, что проценты гомологии, полученные в результате сопоставления с использованием GAP и представленные в Таблице 4 под заголовком «% гомологии (GAP)», даны в Европейском формате нумерации, где «,» представляет запятую в десятичном числе. Так, например, значение «40,345» в этом столбце означает «40,345%».
Пример 13: Конструирование и операции с микроматрицами с ДНК
Последовательности настоящего изобретения могут быть также использованы для конструирования и применения микроматриц ДНК (конструирование, методика и использования ДНК-матриц хорошо известны специалистам и описаны, например, Schena М. et al. (1995) Science 270: 467-470; Wodicka L. et al. (1997) Nature Biotechnology 15: 1359-1367; DeSaizieu A. et al. (1998) Nature Biotechnology 16: 45-48; и DeRisi J.L. et al. (1997) Science 278: 680-686).
Микроматрицы с ДНК представляют собой твердые или гибкие носители, состоящие из нитроцеллюлозы, найлона, стекла, силикона или других материалов. Молекулы нуклеиновой кислоты могут быть присоединены к данной поверхности в определенном порядке. После соответствующего мечения, другие нуклеиновые кислоты или смеси нуклеиновых кислот могут быть гибридизованы с иммобилизованными молекулами нуклеиновой кислоты, а указанная метка может быть использована для мониторинга и измерения интенсивностей отдельных сигналов гибридизованных молекул в определенных областях. Эта методика позволяет одновременно определить относительное или абсолютное количество всех или отдельно выбранных нуклеиновых кислот в используемом образце нуклеиновой кислоты или смеси нуклеиновых кислот. Следовательно, микроматрицы с ДНК позволяют проводить параллельный анализ экспрессии множества (вплоть до 6800 или более) нуклеиновых кислот (см. например, Schena М. (1996) Bioassays 18(5): 427-431).
Последовательности настоящего изобретения могут быть использованы для конструирования олигонуклеотидных праймеров, способных амплифицировать определенные области одного или нескольких генов С. glutamicum путем реакции амплификации нуклеиновых кислот, такой как полимеразная цепная реакция. Выбор и конструирование 5'-или 3'-олигонуклеотидных праймеров или их соответствующих линкеров позволяет осуществлять ковалентное связывание полученных ПЦР-продуктов с поверхностью среды-носителя, описанной выше (а также описанной, например, Schena М. et al. (1995) Science 270:467-470).
Микроматрицы с нуклеиновыми кислотами могут быть также сконструированы путем in situ олигонуклеотидного синтеза, описанного Wodicka L. et al. (1997) Nature Biotechnology 15:1359-1367. При использовании фотолитографических методов, точно определенные области данной матрицы подвергают световому облучению. Защитные группы, которые были фотолобильными, активировались и присоединяли нуклеотиды, тогда как области, которые были защищены от светового облучения, не подвергались какой-либо модификации. Последовательные циклы защиты от света и активации светом позволяют осуществлять синтез различных олигонуклеотидов в определенных положениях. Конкретные небольшие области генов настоящего изобретения могут быть синтезированы на микроматрицах посредством твердофазного олигонуклеотидного синтеза.
Молекулы нуклеиновой кислоты настоящего изобретения, присутствующие в образце или в смеси нуклеотидов, могут быть гибридизованы с указанными микроматрицами. Эти молекулы нуклеиновой кислоты могут быть помечены стандартными методами. Для этого, молекулы нуклеиновой кислоты (например, молекулы мРНК или молекулы ДНК) метят путем введения меченых изотопов или флуоресцентно меченных нуклеотидов, например, во время обратной транскрипции или синтеза ДНК. Гибридизация меченых нуклеиновых кислот с микроматрицами описана, например, Schena М. et al. (1995) см. выше; Wodicka L. et al. (1997) см. выше; и DeSaizieu A. et al. (1998) (см.выше). Детекция и определение количества гибридизованных молекул связаны со специфической введенной меткой. Радиоактивные метки могут быть детектированы, например, как описано Schena М. et al. (1995) (см.выше), а флуоресцентные метки могут быть детектированы, например, как описано Shalon et al. (1996) Genome Research 6: 639-645.
Применение последовательностей настоящего изобретения в технологии с использованием микроматриц ДНК, описанных выше, позволяет проводить сравнительные анализы различных штаммов С. glutamicum или других коринебактерий. Так, например, исследования различий между штаммами исходя из профилей отдельных транскриптов и идентификация генов, которые являются важными с точки зрения специфических и/или желаемых свойств штаммов, таких как патогенность, продуктивность и толерантность к стрессам, могут быть облегчены путем применения методики с использованием матриц нуклеиновых кислот. Благодаря методике с использованием матриц нуклеиновых кислот можно также проводить сравнительные анализы профиля экспрессии генов настоящего изобретения в процессе реакции ферментации.
Пример 14: Анализ динамики клеточных популяций белков (протеомиков)
Гены, композиции и способы настоящего изобретения могут быть применены для исследования взаимодействий и динамики популяций белков, называемых «протеомиками». Популяциями этих представляющих интерес белков являются, но не ограничиваются ими, общая популяция белков С. glutamicum (например, в сравнении с популяциями белков других микроорганизмов), популяция белков, которые являются активными в специфических окружающих условиях или в условиях метаболизма (например, в процессе ферментации, при высокой или низкой температуре или при высоком или низком рН) или популяция белков, которые являются активными в процессе специфических фаз роста и развития.
Популяции белков могут быть проанализированы различными хорошо известными методами, такими как гель-электрофорез. Клеточные белки могут быть получены, например, путем лизиса или экстракции, и могут быть отделены друг от друга рядом электрофоретических методов. Электрофорез в полиакриламидном геле с додецилсульфатом натрия (ДСН-ПААГ) разделяет белки, главным образом, по их молекулярной массе. Изоэлектрическое фокусирование в геле (ИЭФ-ПААГ) разделяет белки по их изо-электрической точке (что позволяет выявить не только данную аминокислотную последовательность, но также и посттрансляционные модификации данного белка). Другим, более предпочтительным методом анализа белков является последовательная комбинация как ИЭФ-ПААГ, так и ДСН-ПААГ, известная как двумерный электрофорез в геле (описанный, например, Hermann et al. (1998) Electrophoresis 19:3217-3221; Fountoulakis et al. (1998) Electrophoresis 19: 1193-1202; Langen et al. (1997) Electrophoresis 18: 1184-1192; Antelmann et al. (1997) Electrophoresis 18: 1451-1463). Для разделения белков могут быть также использованы и другие методы разделения, такие как электрофорез в капиллярном геле; и такие методы хорошо известны специалистам.
Белки, разделенные с помощью этой методики, могут быть визуализированы стандартными методами, такими как окрашивание или мечение. Подходящие красители известны специалистам и ими являются кумасси бриллиантовый голубой, серебряный краситель или флуоресцентные красители, такие как рубиновый краситель (Sypro Ruby) (Molecular Probes). Включение радиоактивно меченных аминокислот или других предшественников белков (например, 35S-метионина, 35S-цистеина, 14С-меченых аминокислот, 15N-аминокислот, 15NO3 или 15NH4 + или 13С-меченых аминокислот) в среду С. glutamicum позволяет проводить мечение белков, происходящих от этих клеток, перед их разделением. Аналогично, могут быть использованы флуоресцентные метки. Указанные меченые белки могут быть экстрагированы, выделены и разделены ранее описанными методами.
Белки, визуализированные этими методами, могут быть, кроме того, проанализированы путем определения количества используемого красителя или метки. Количество данного белка может быть определено количественно, например, оптическими методами, и может быть проведено сравнение с количеством других белков в том же самом геле или в других гелях. Сравнение белков на гелях может быть проведено, например, с помощью оптического сравнения, путем спектроскопии, визуализирующего сканирования и анализа гелей, либо с использованием фотографических пленок и экранов. Такие методы хорошо известны специалистам.
Для определения идентичности любого данного белка может быть использовано прямое секвенирование или другие стандартные методы. Так, например, можно использовать секвенирование N- и/или С-концевых аминокислот (например, деградация по Эдману) или масс-спектрометрию (в частности, методы MALDI или ESI (см. например, Langen et al. (1997) Electrophoresis 18: 1184-1192)). В соответствии с этими методами, полученные в настоящей заявке последовательности белков могут быть использованы для идентификации белков С. glutamicum.
Данные, полученные этими способами, могут быть использованы для сравнения характера присутствия, активности или модификации белков в различных образцах при различных биологических условиях (например, в различных микроорганизмах, в определенный период времени ферментации, в определенных условиях среды или в различных биотопах и т.п.). Данные, полученные в результате проведения таких экспериментов, отдельно или в комбинации с другими методами, могут быть использованы для различных целей, например, для сравнения поведения различных организмов в данной (например, метаболической) ситуации, для повышения продуктивности штаммов, которые продуцируют химические продукты тонкого органического синтеза, или для повышения эффективности продуцирования химических продуктов тонкого органического синтеза.
Эквиваленты
Многие эквиваленты конкретных вариантов осуществления изобретения, описанных в данной заявке, являются очевидными или могут быть получены каждым специалистом путем рутинного экспериментирования. Такие эквиваленты входят в объем нижеследующей формулы изобретения.
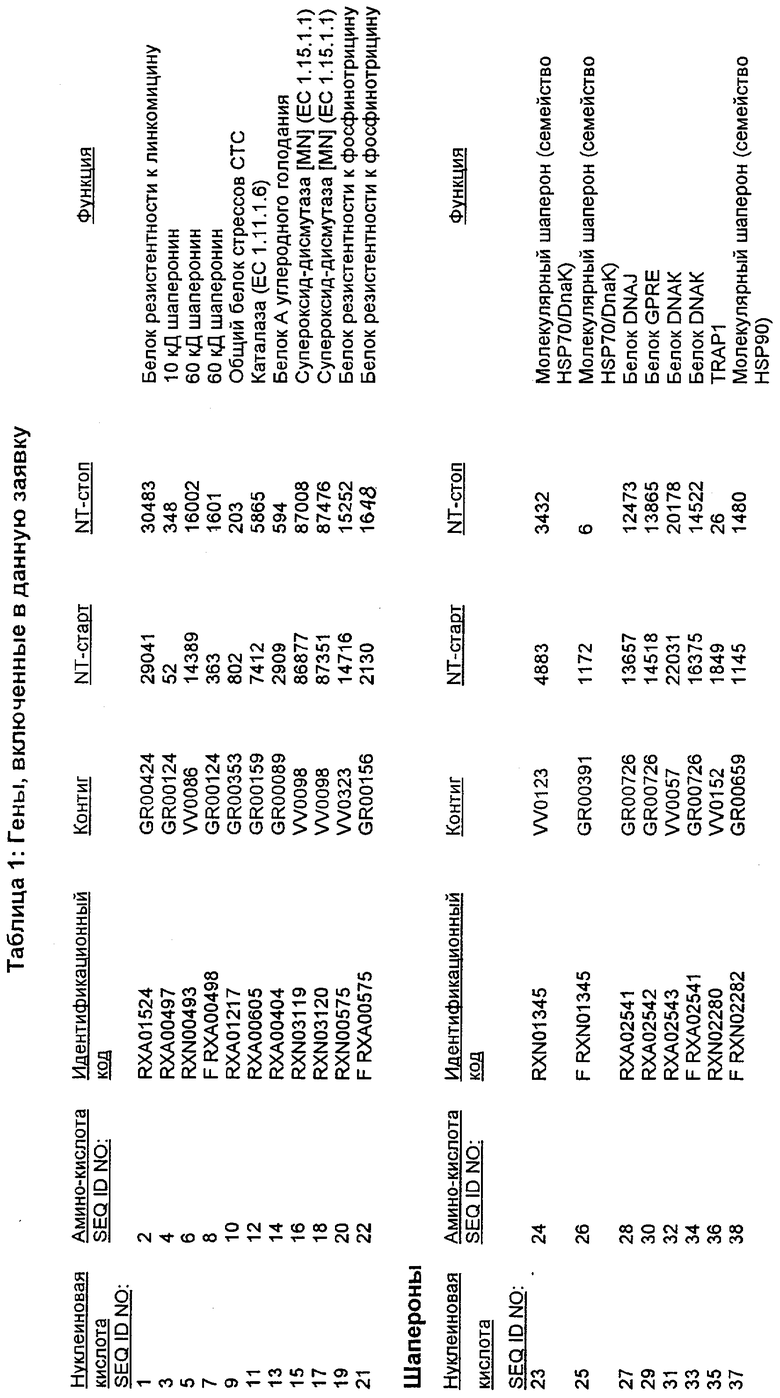
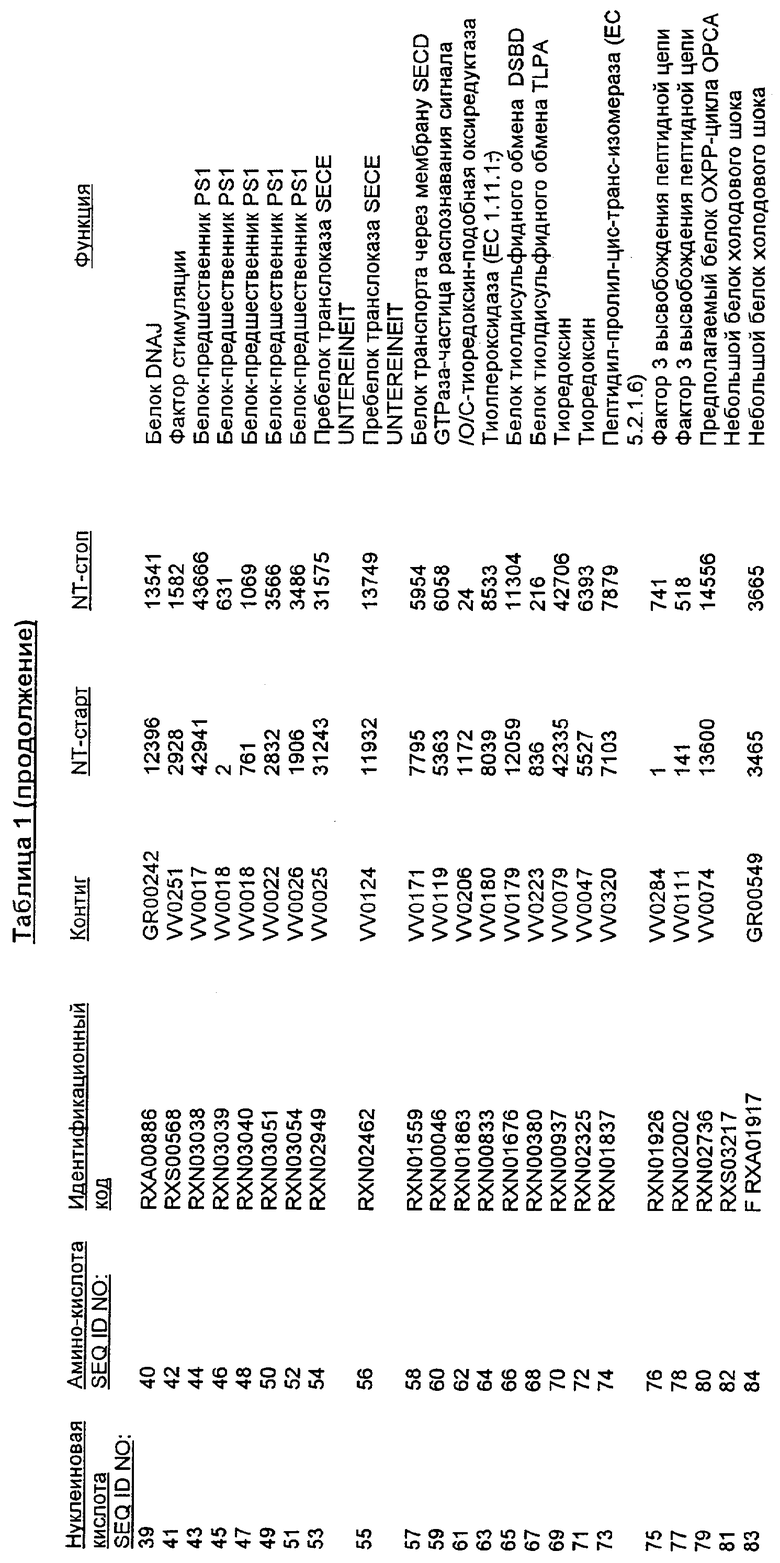


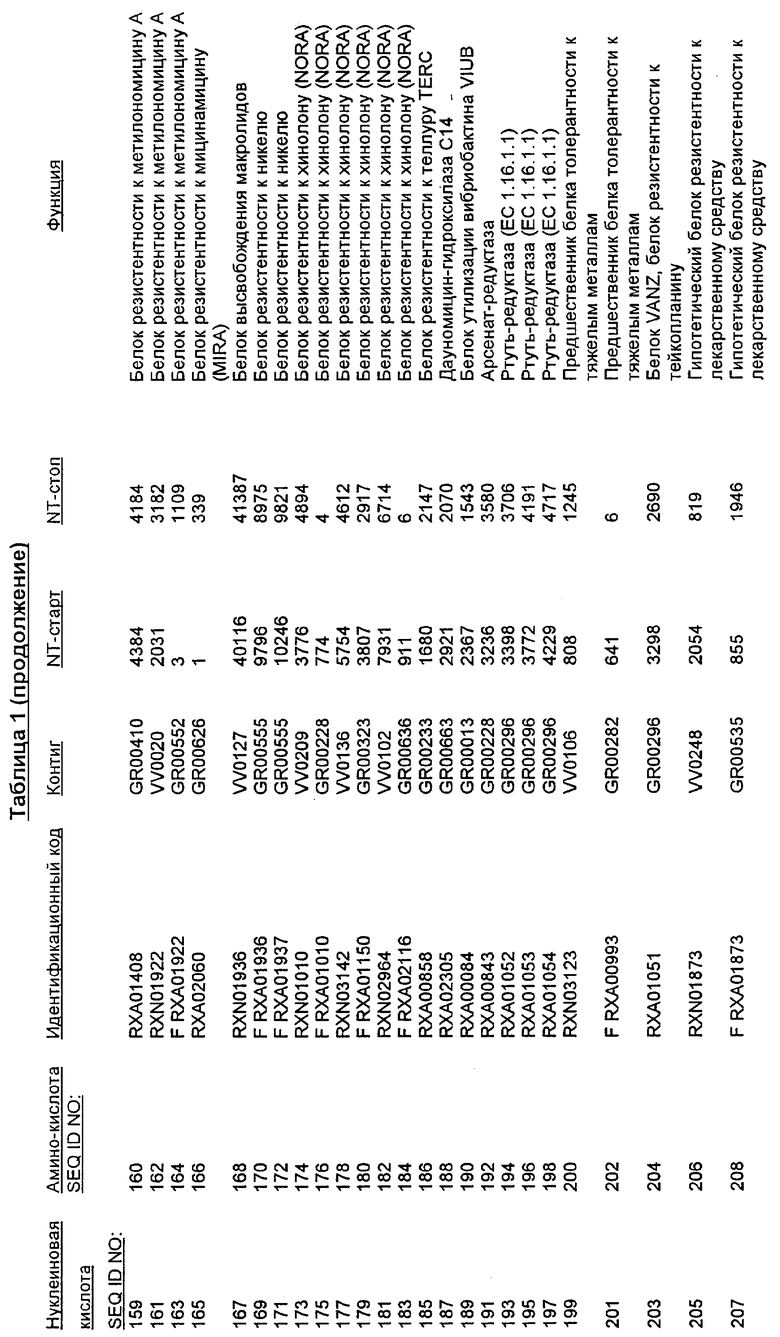
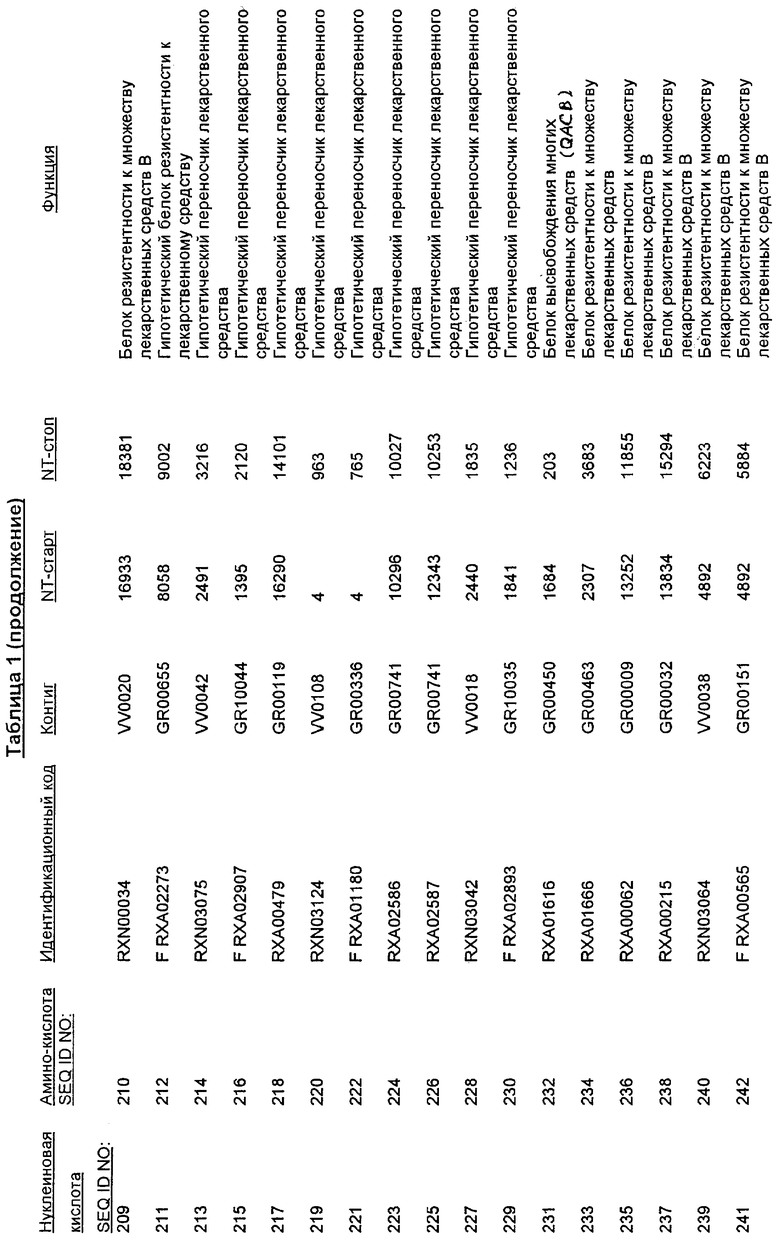
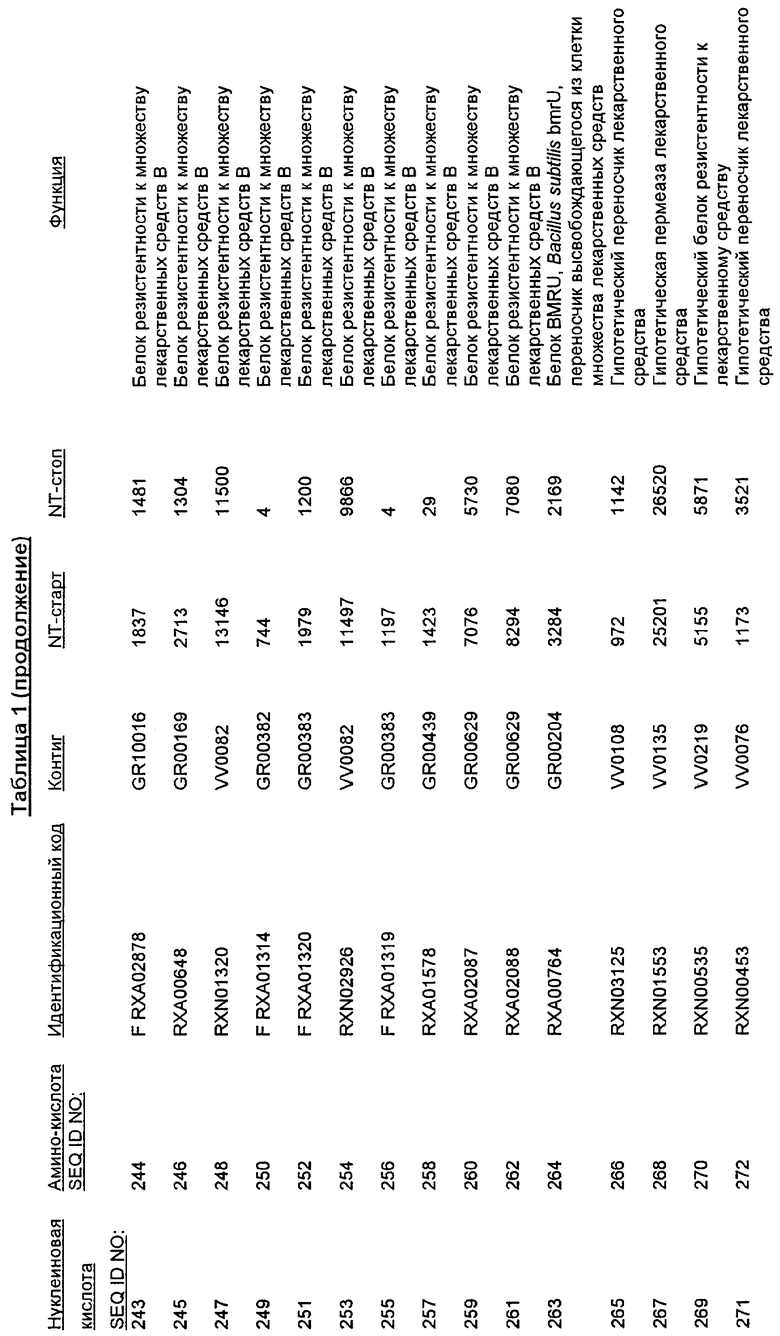
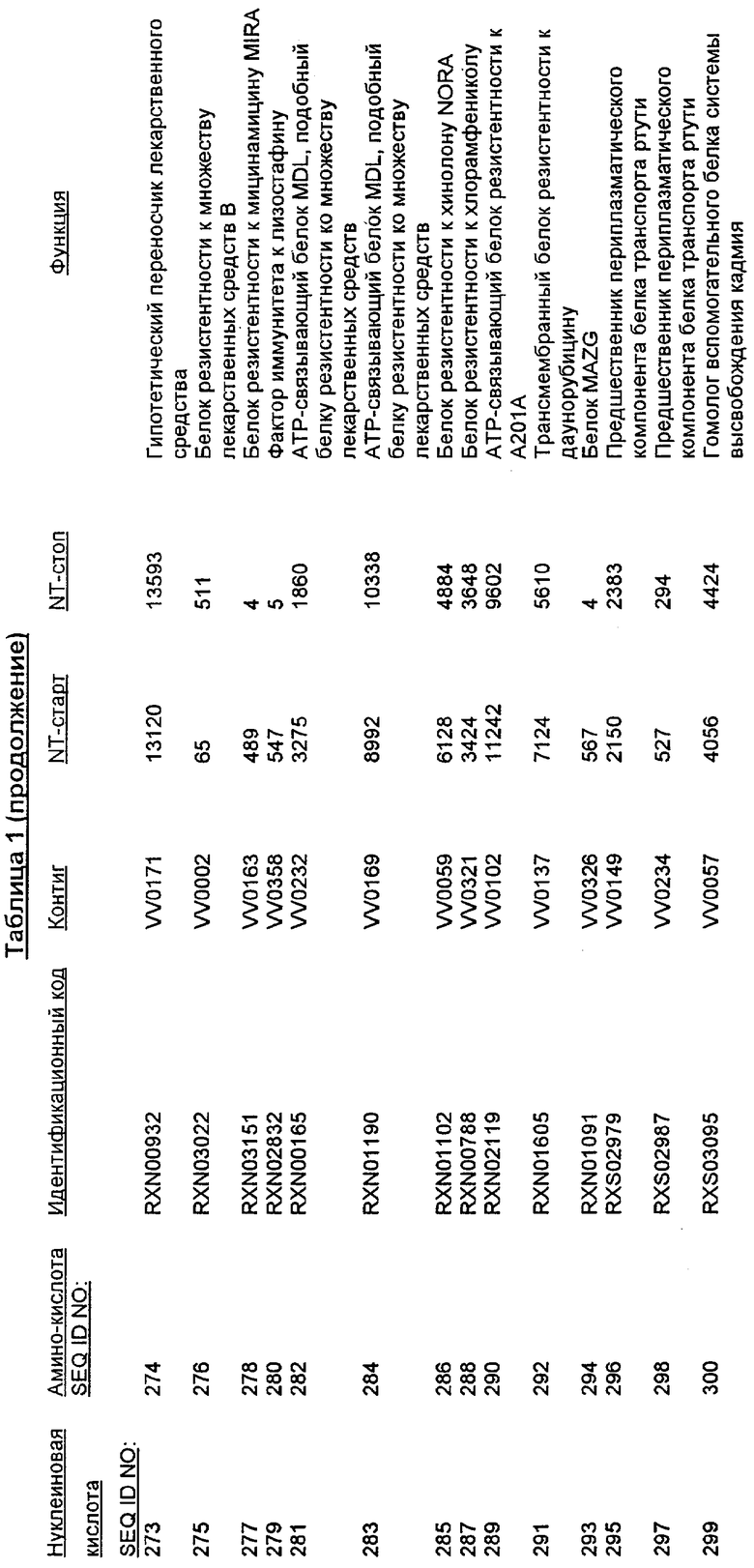
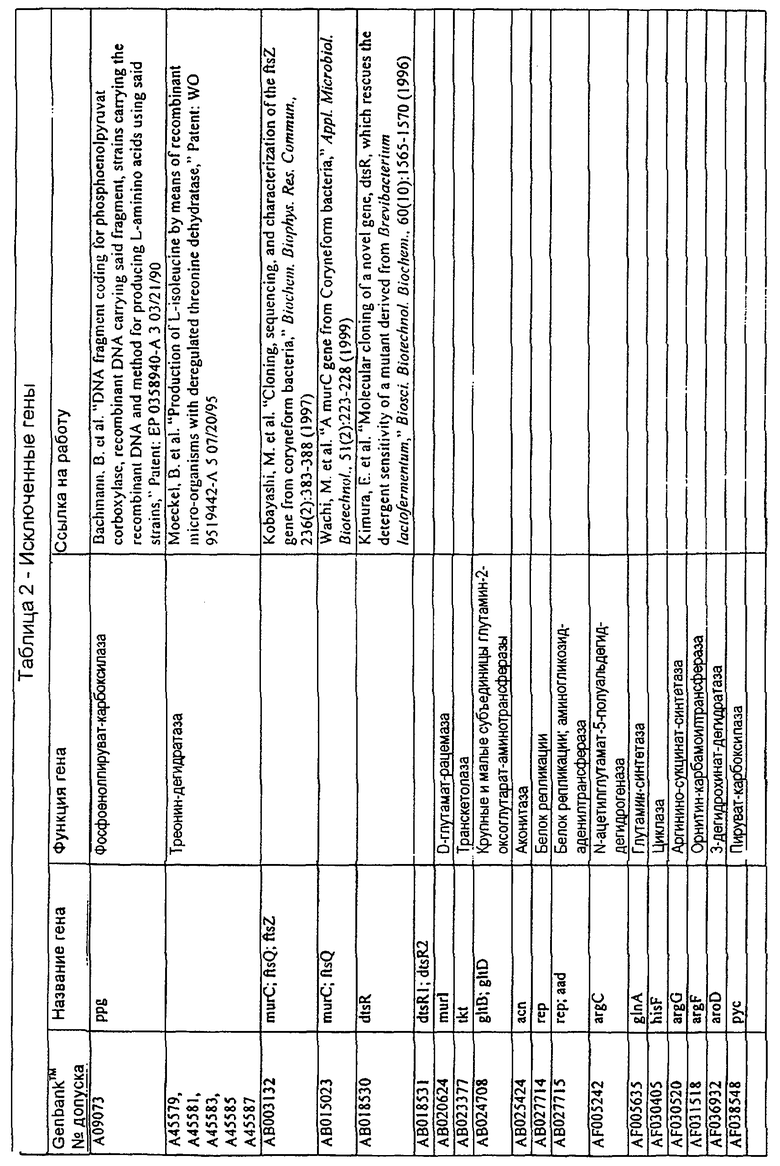
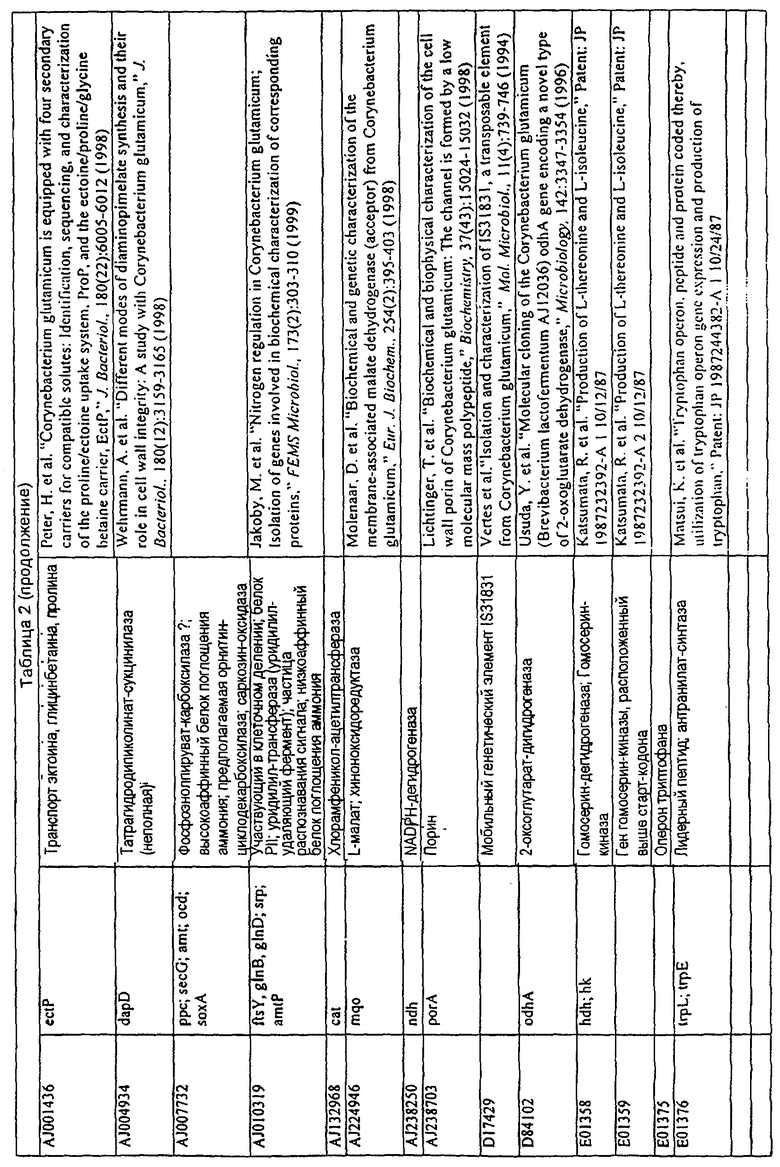
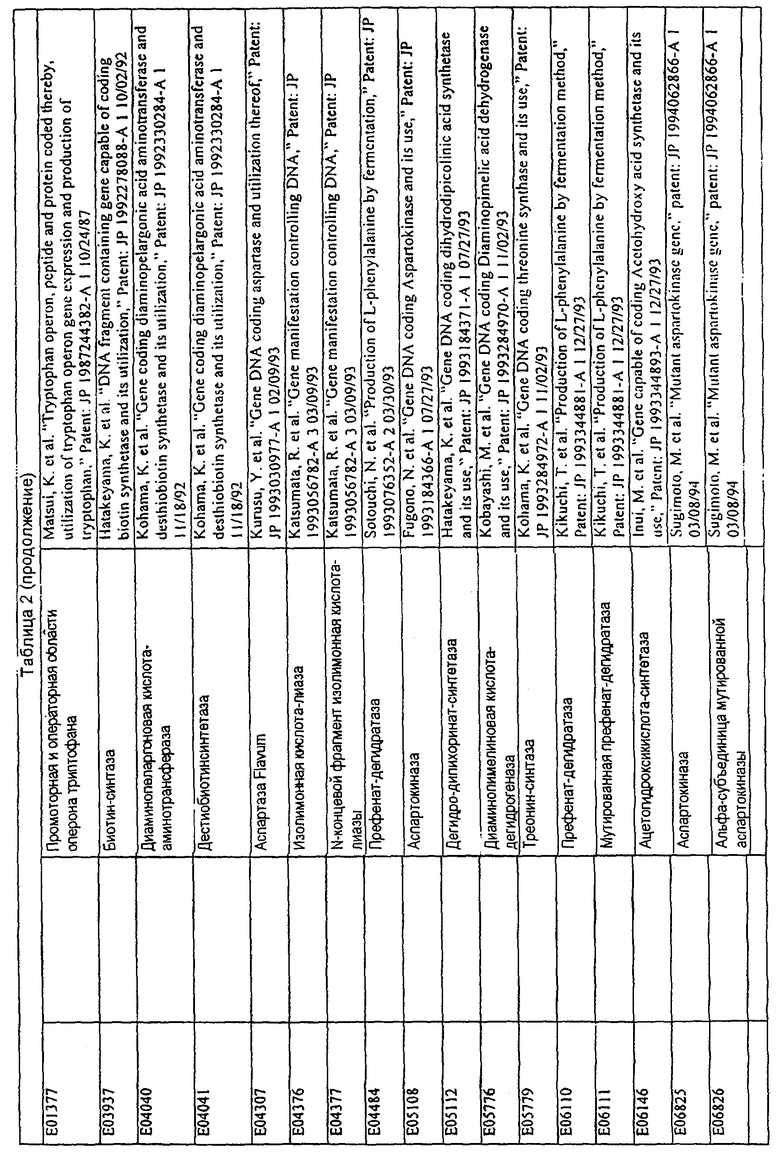
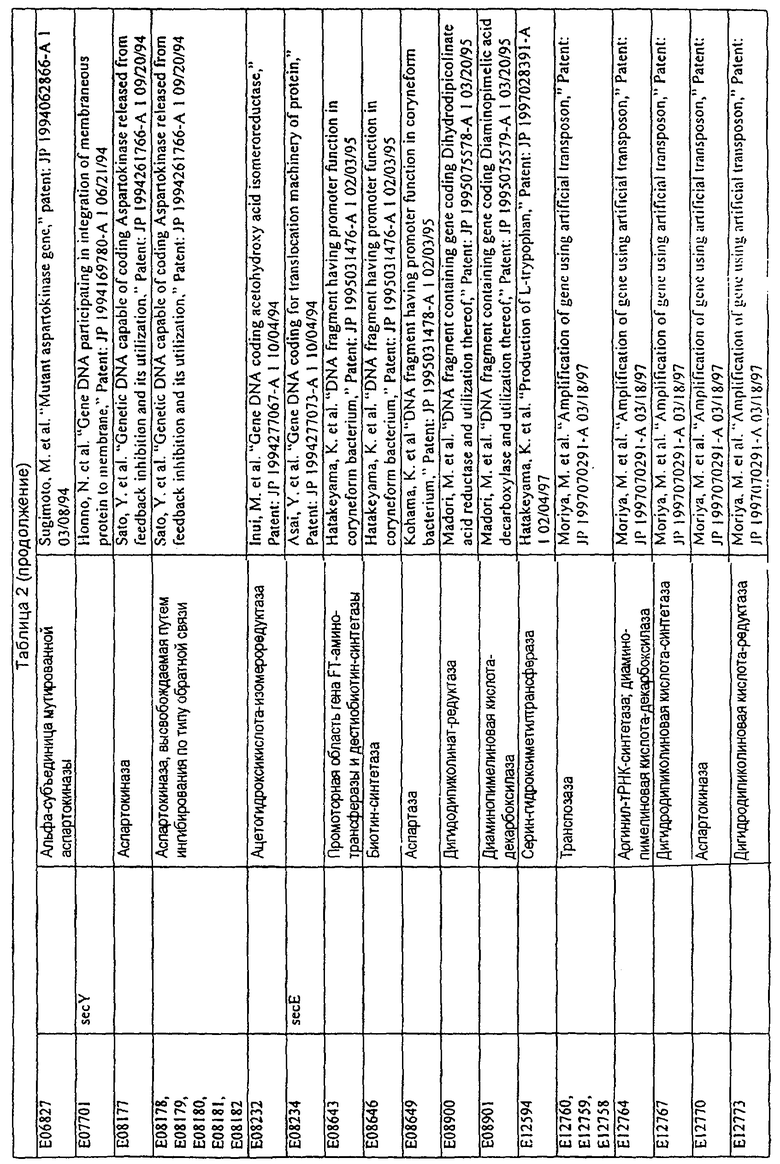
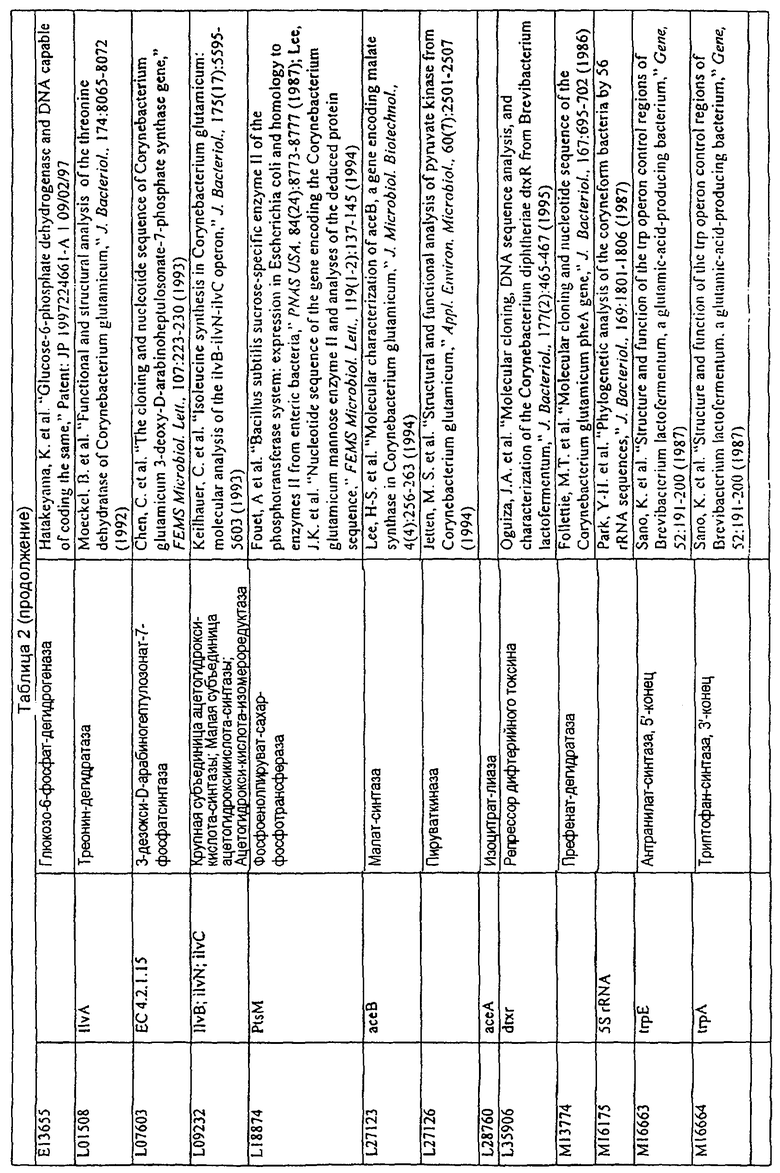
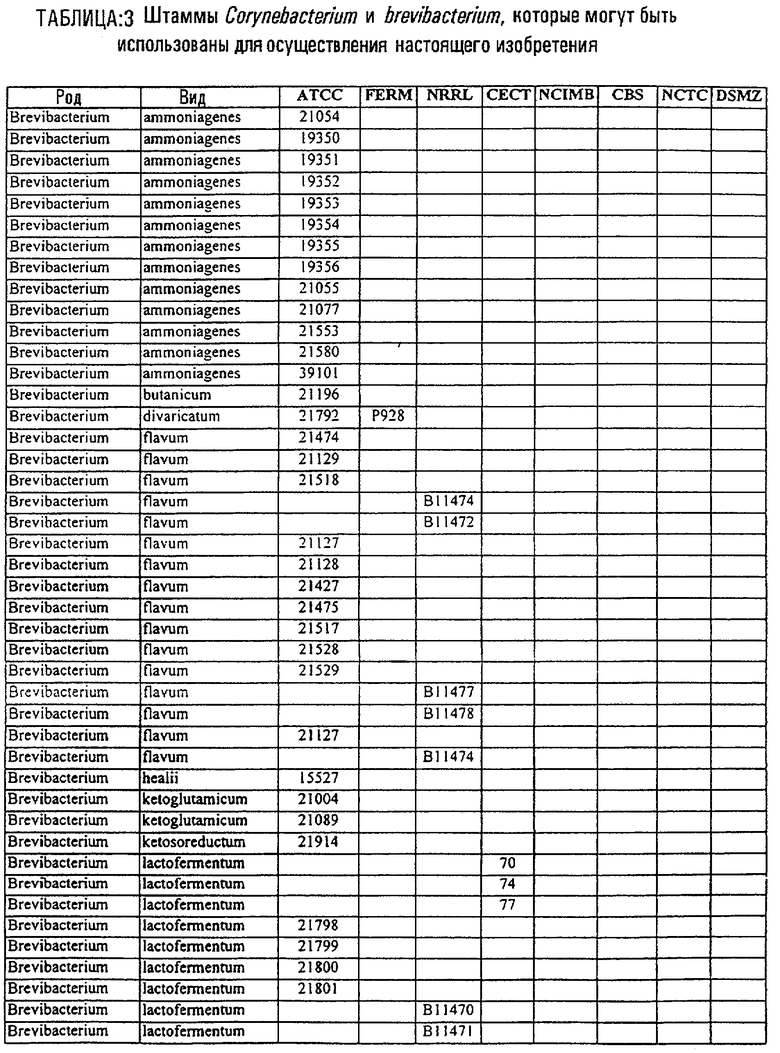
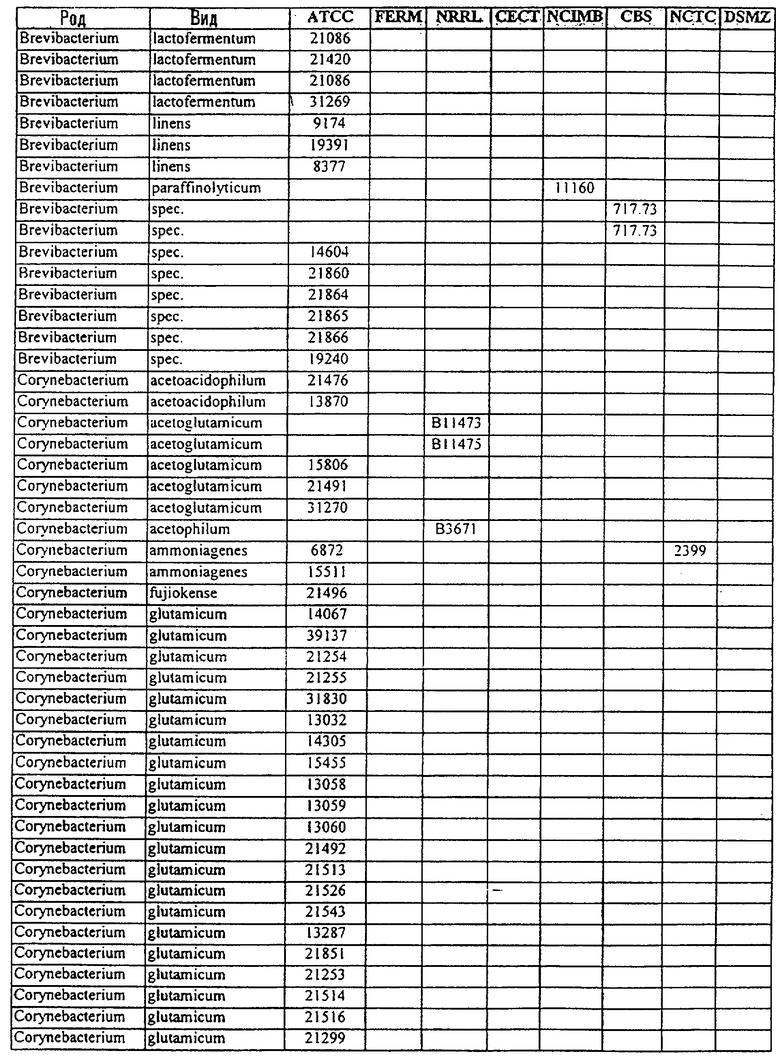
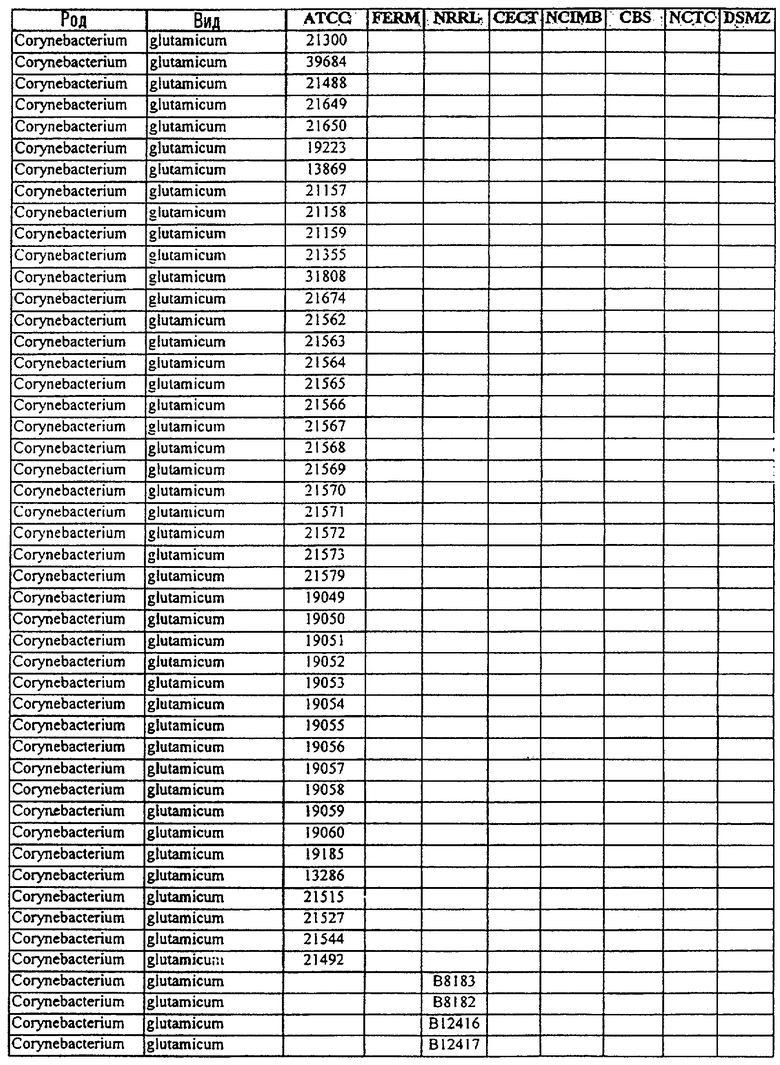
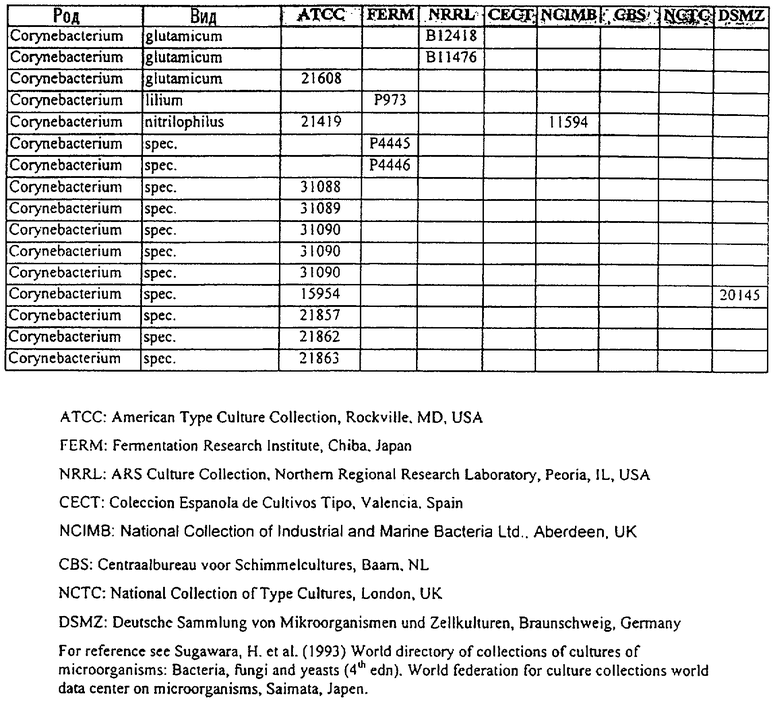




















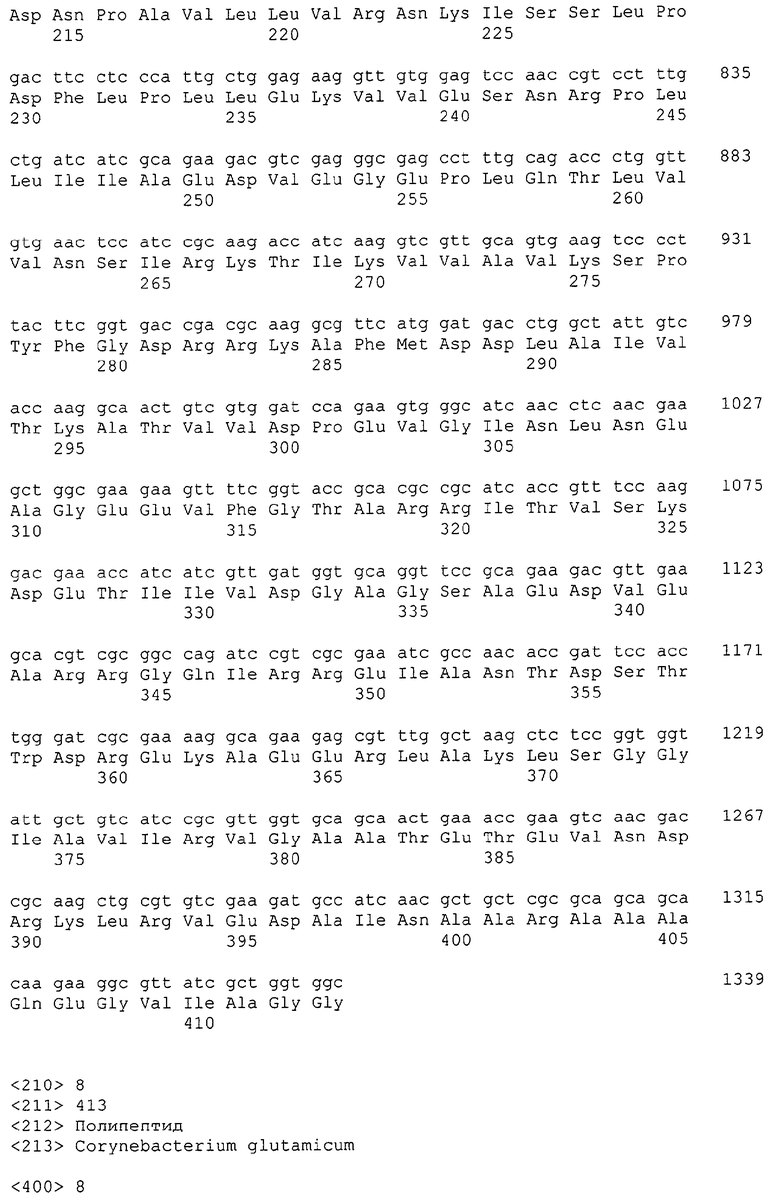










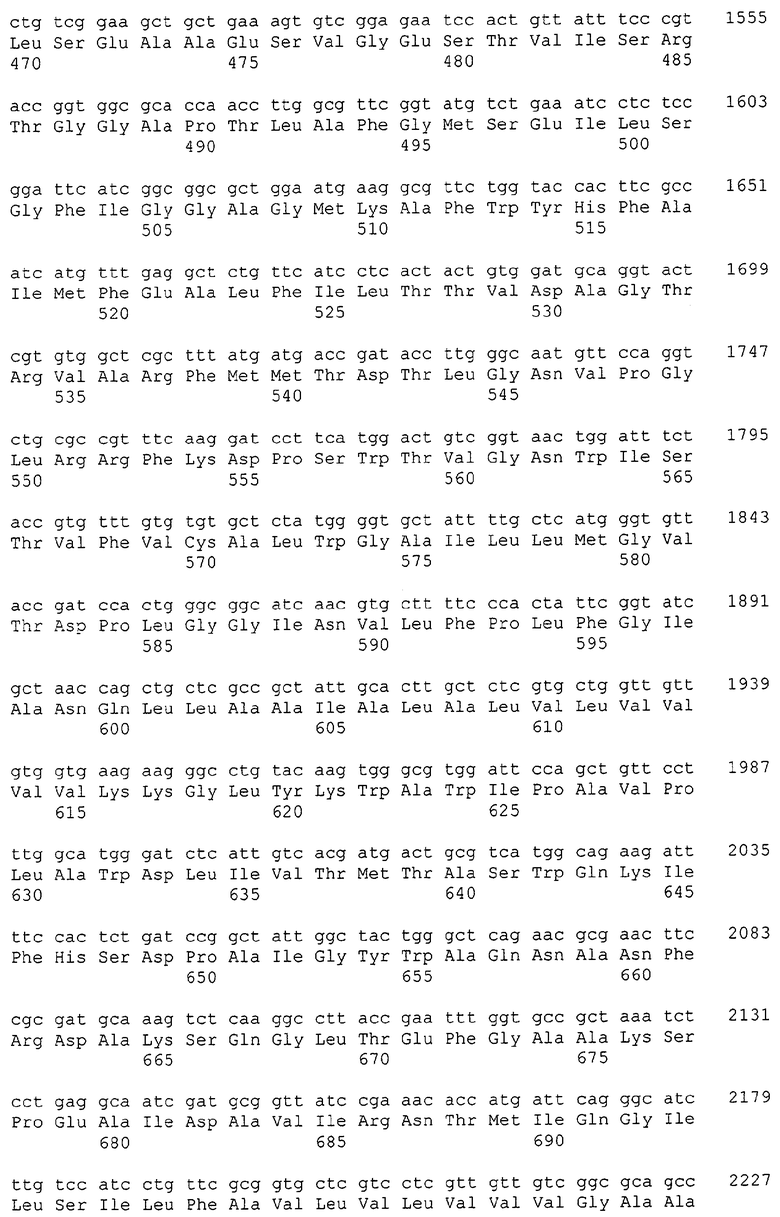




















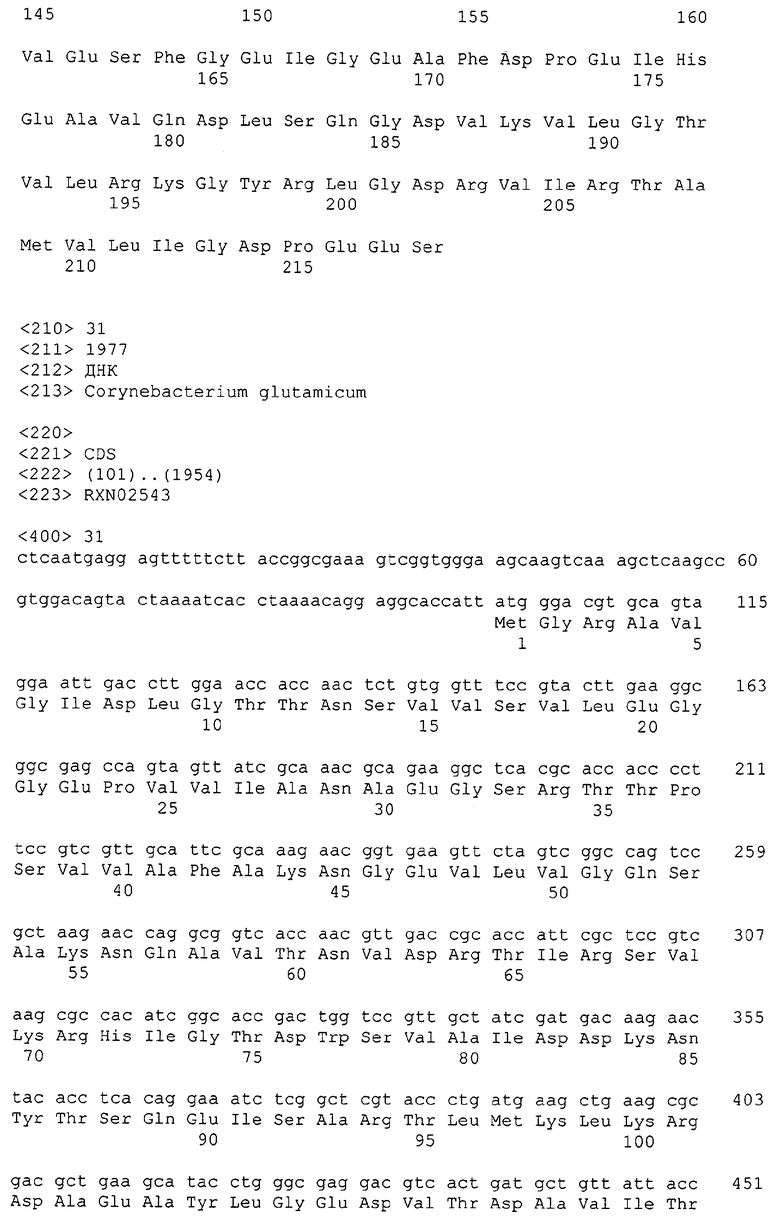







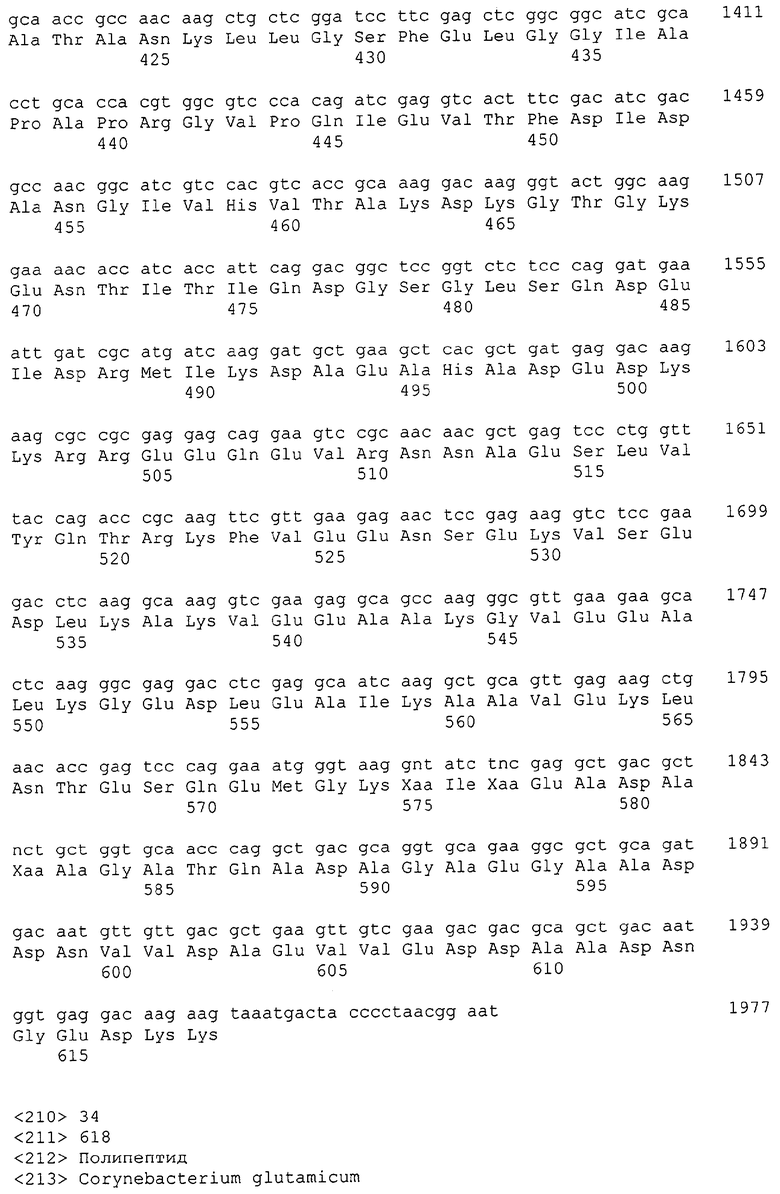













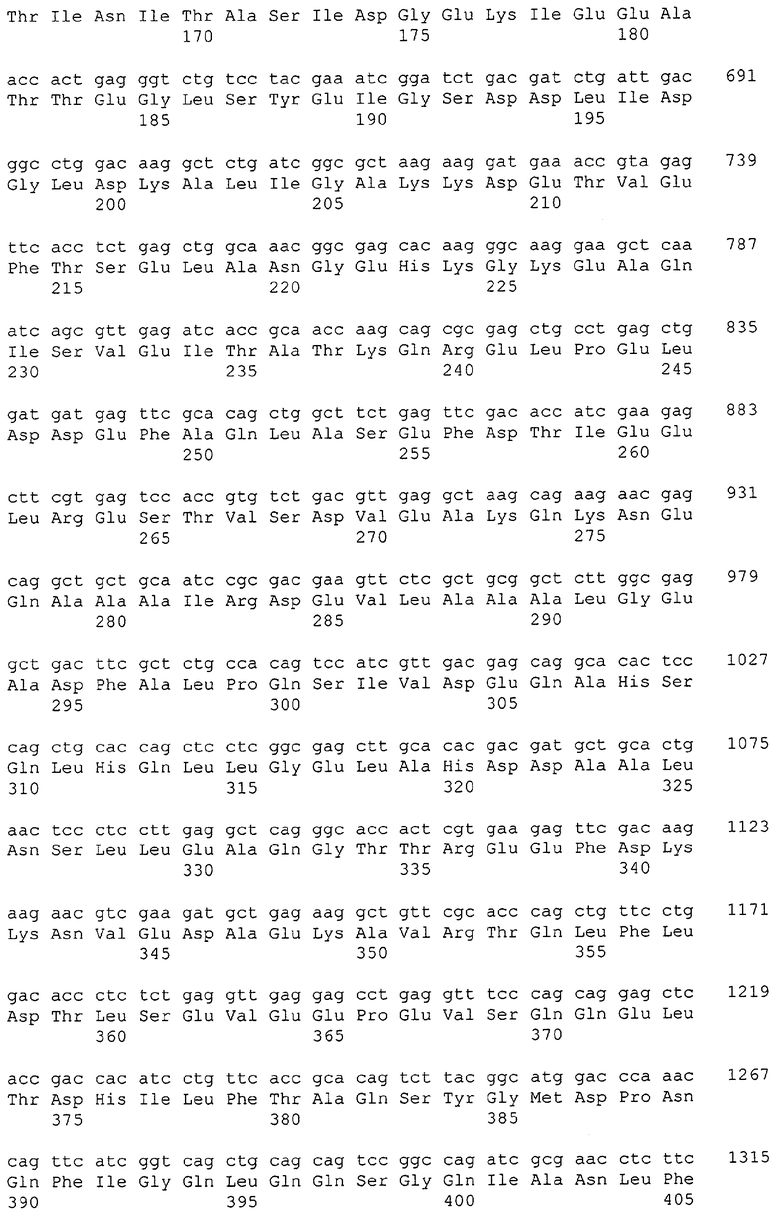









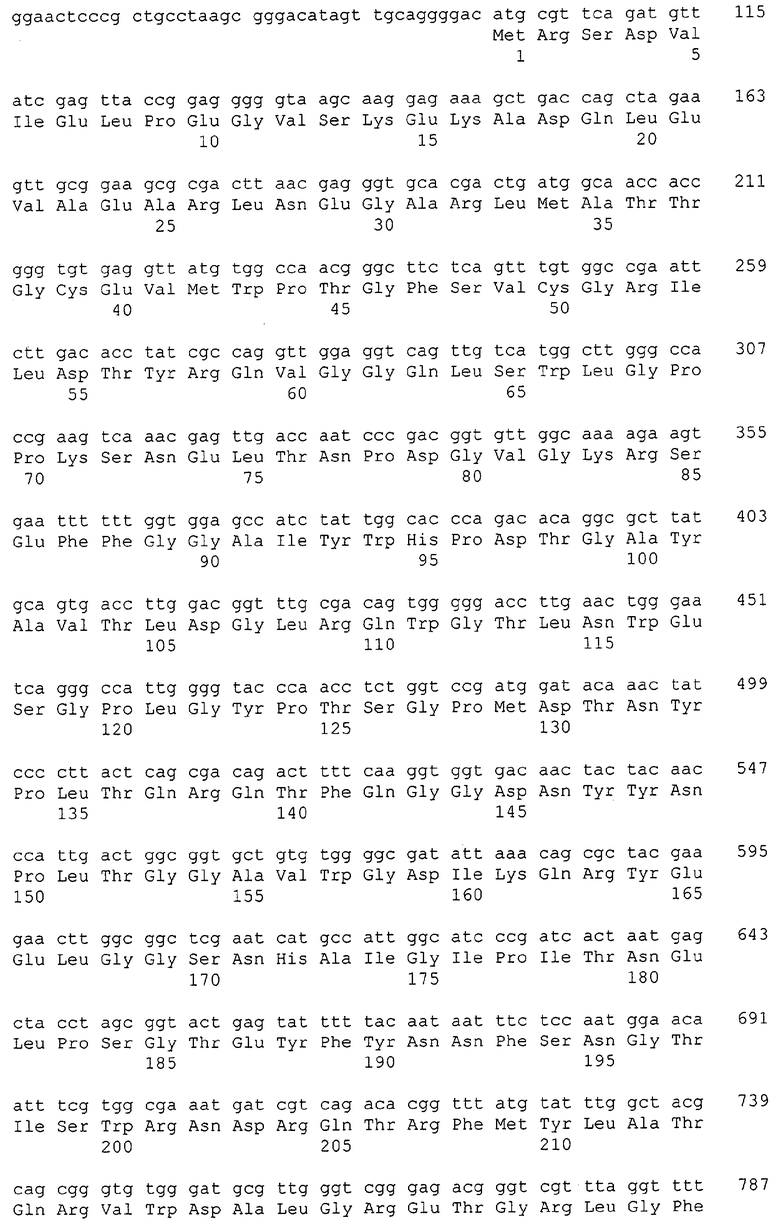















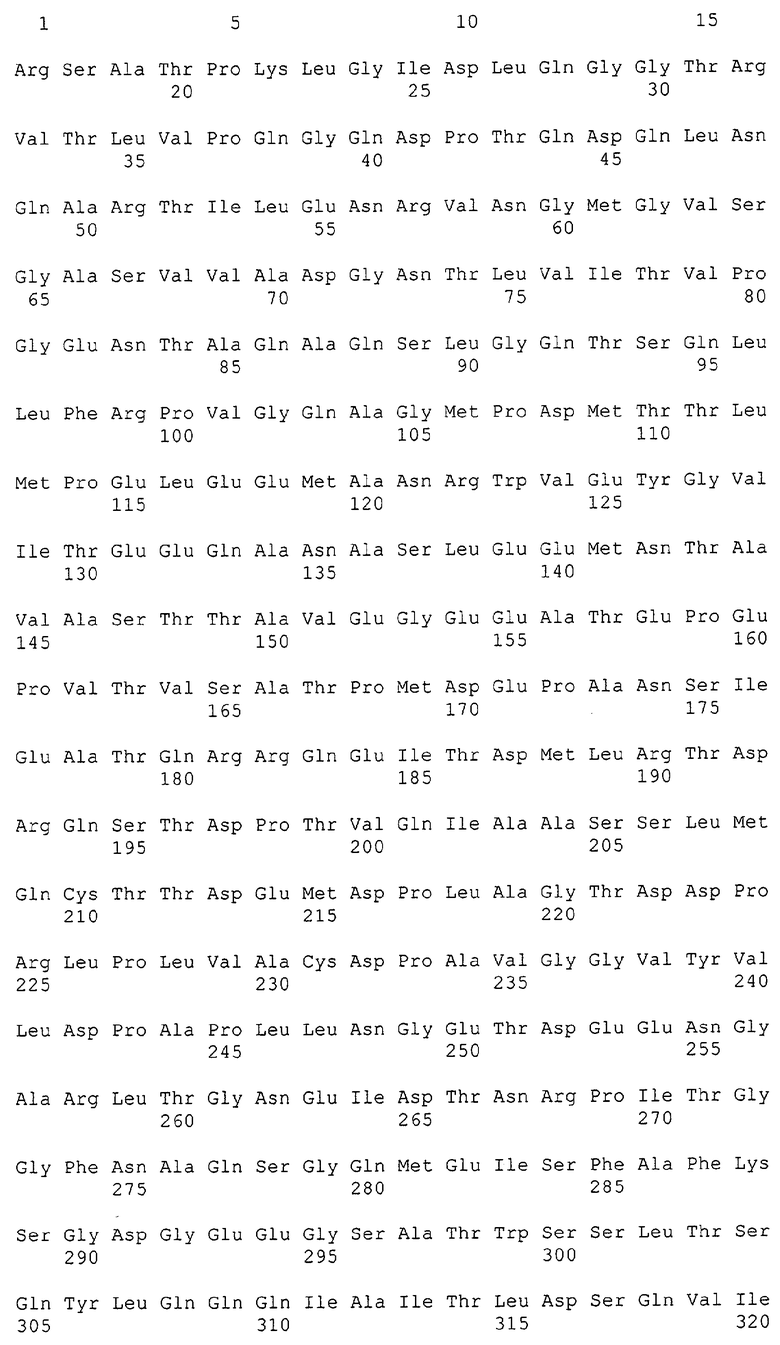














































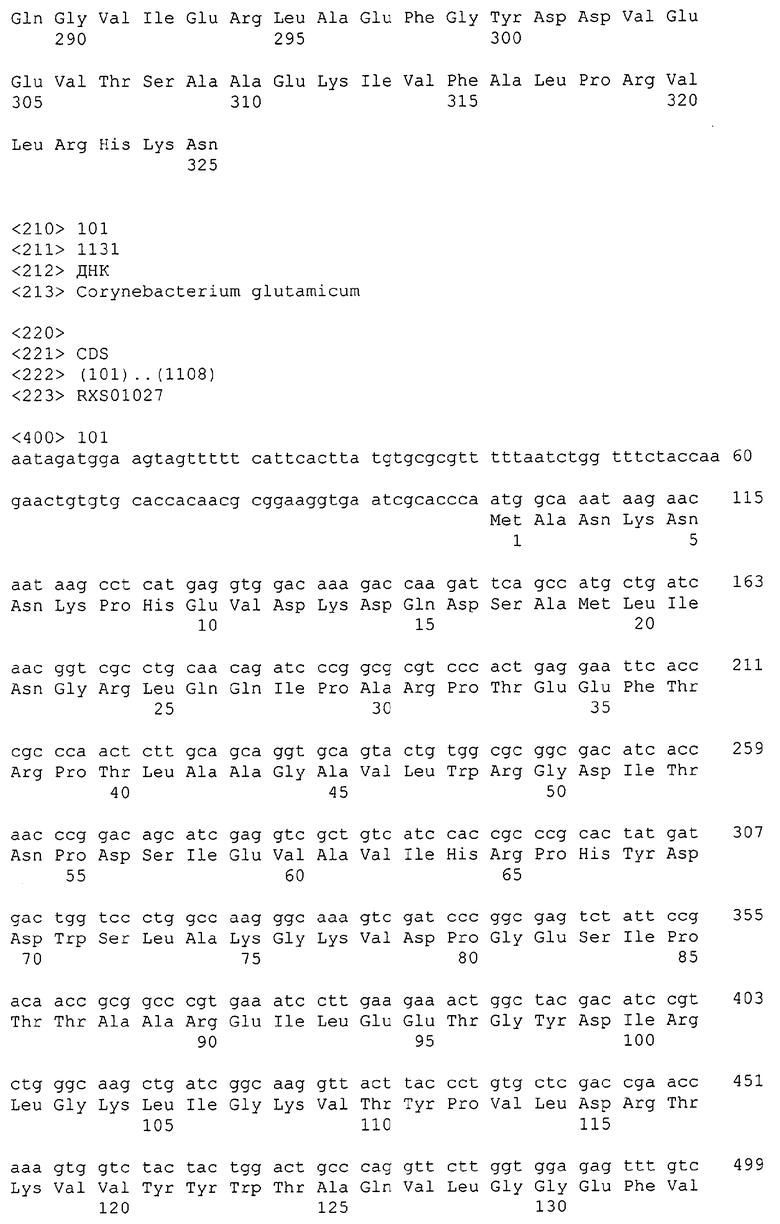
















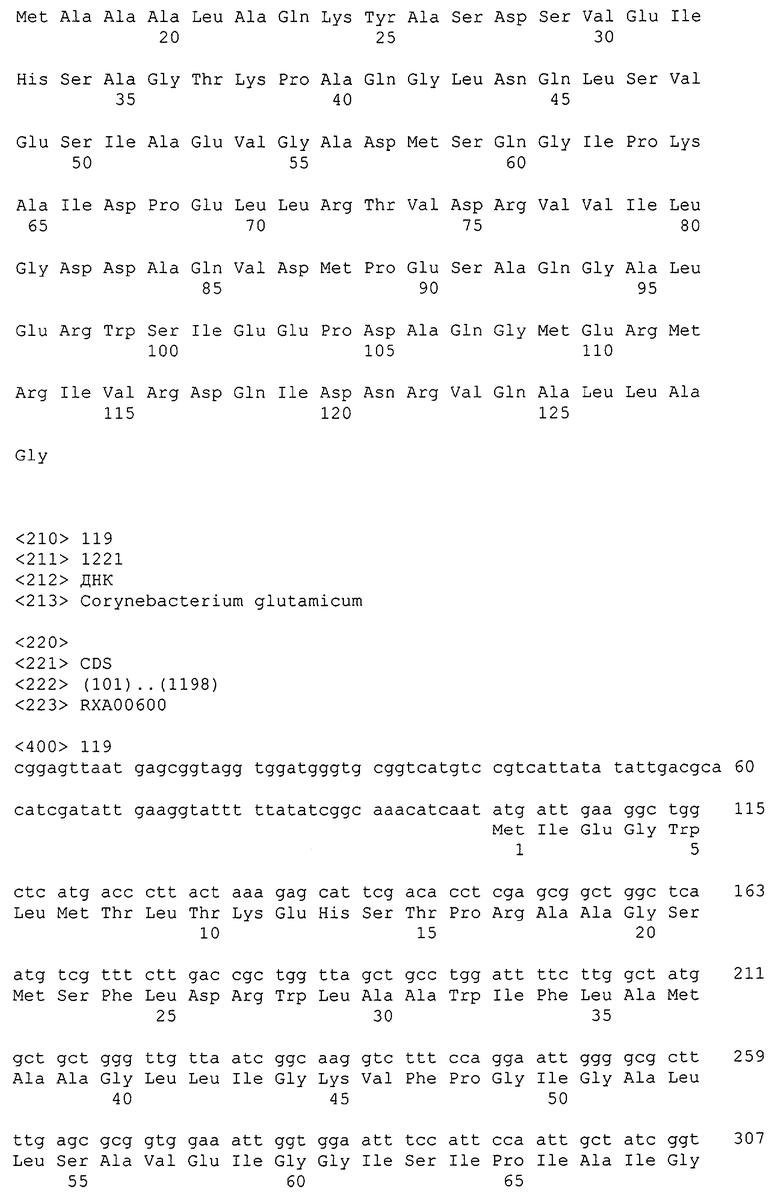






























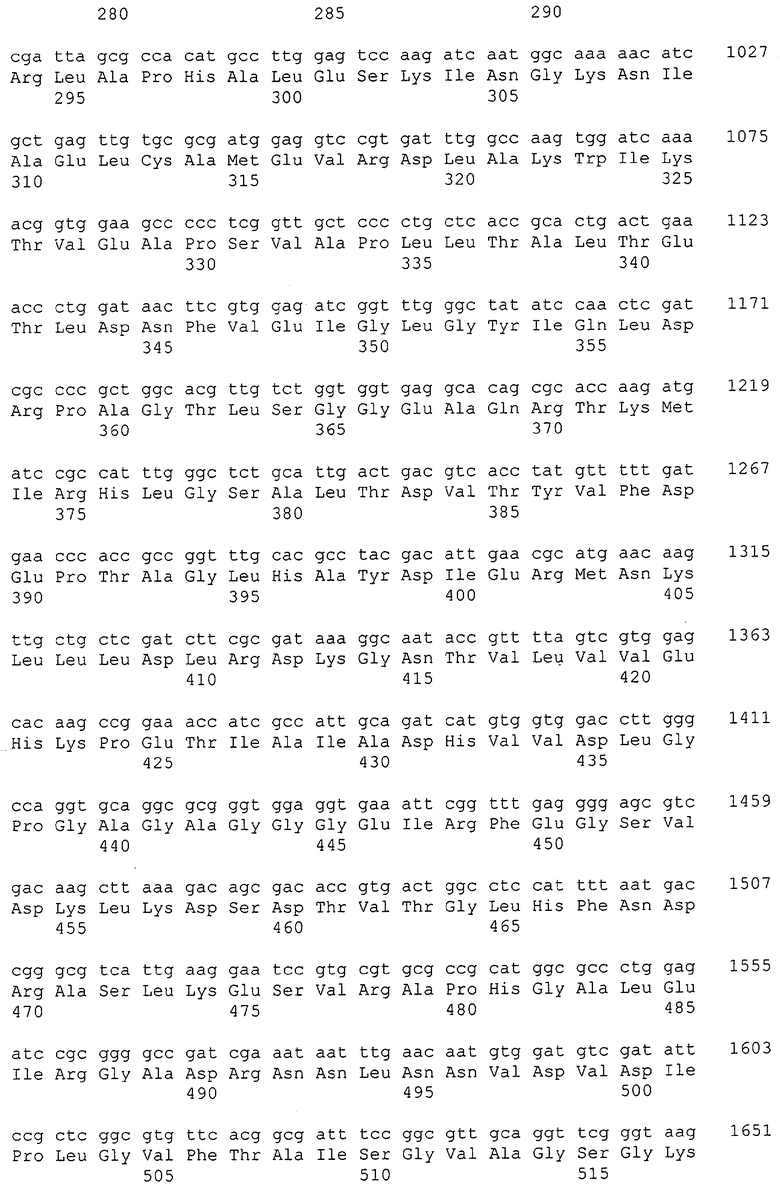
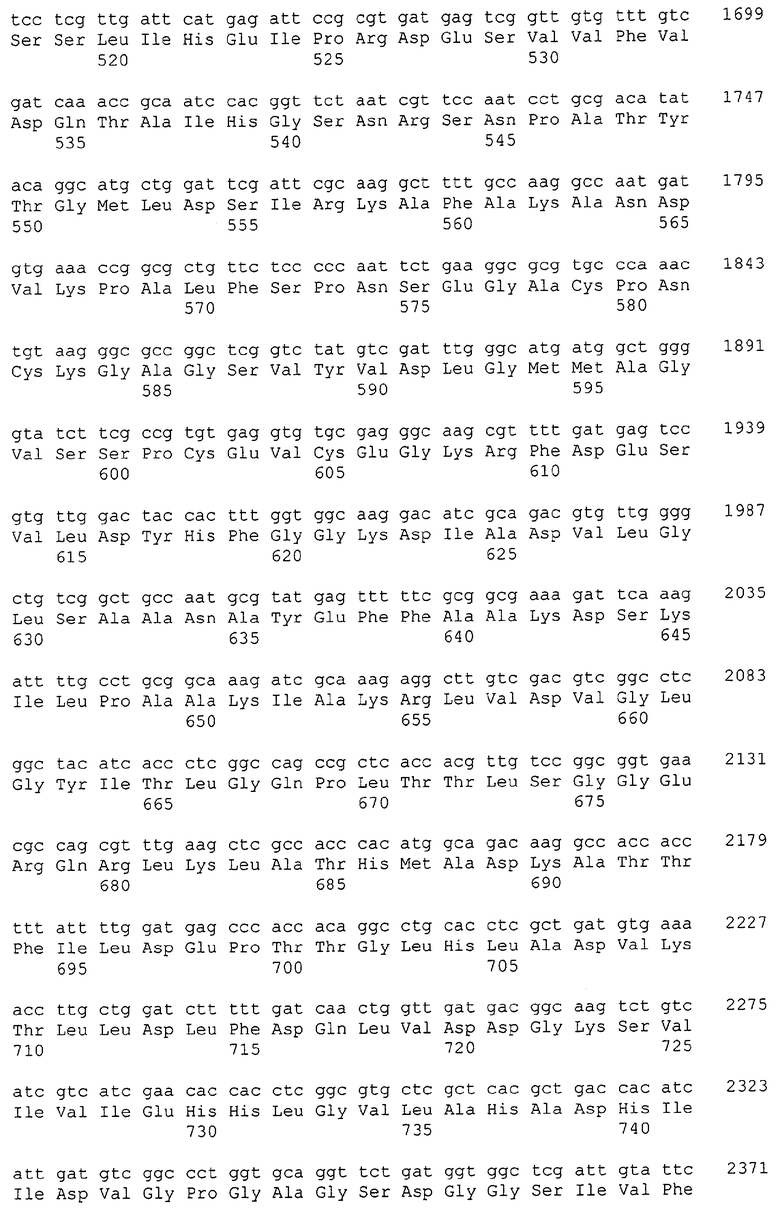





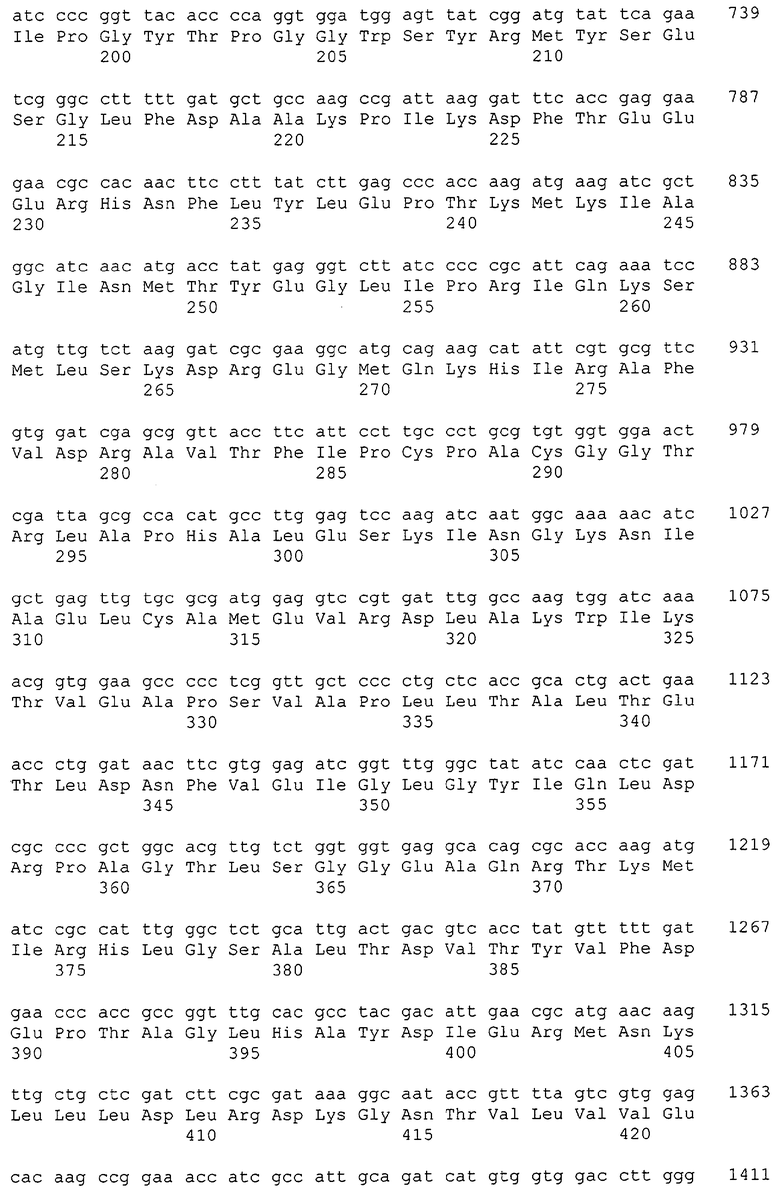




























































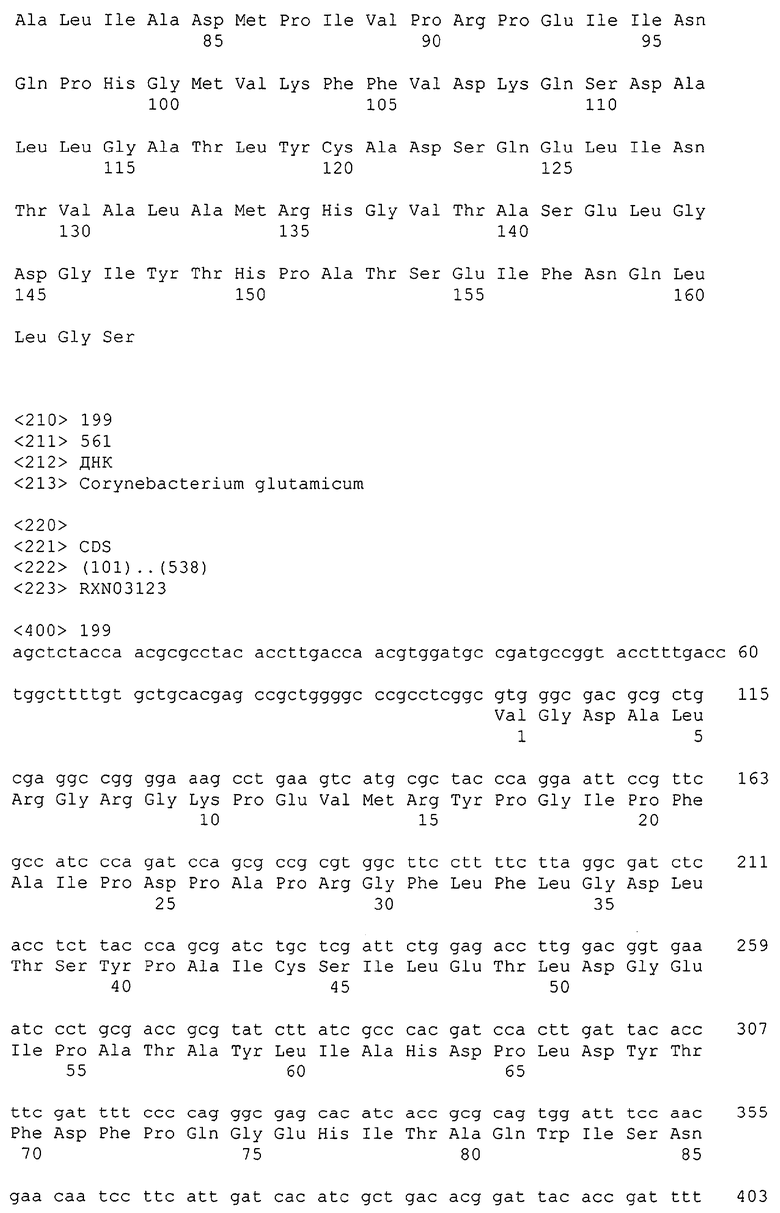



























































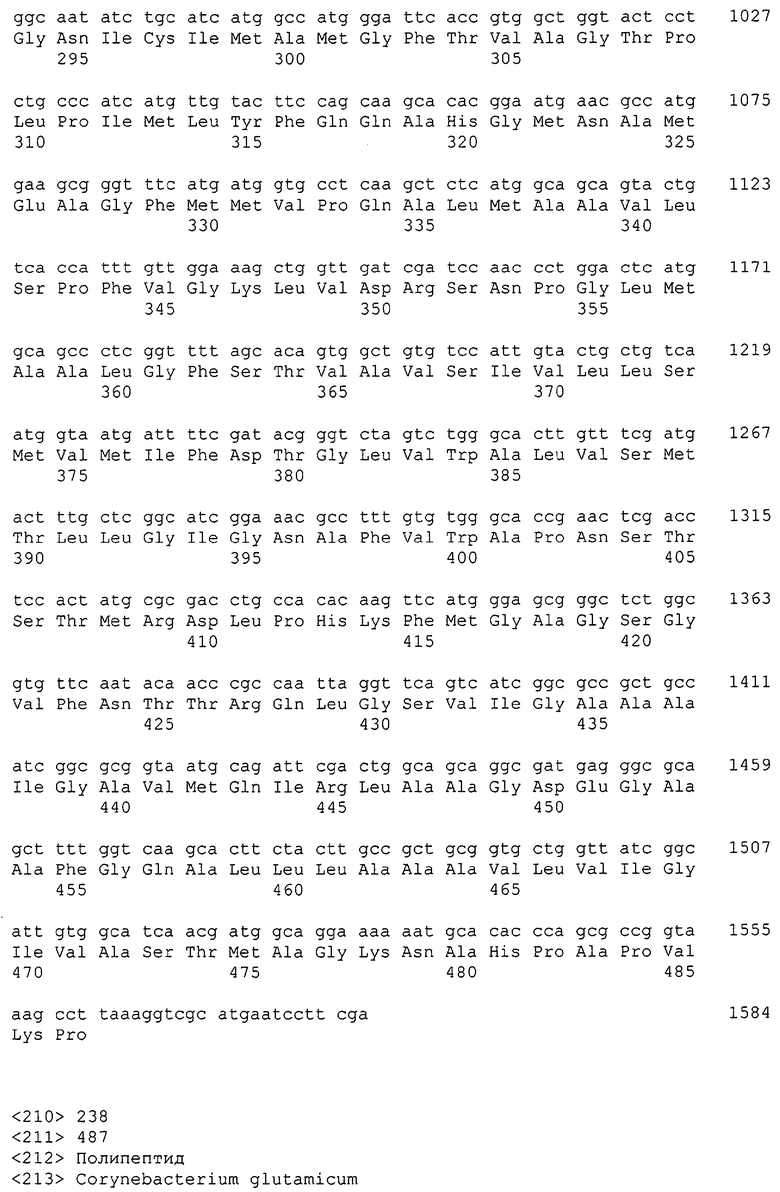



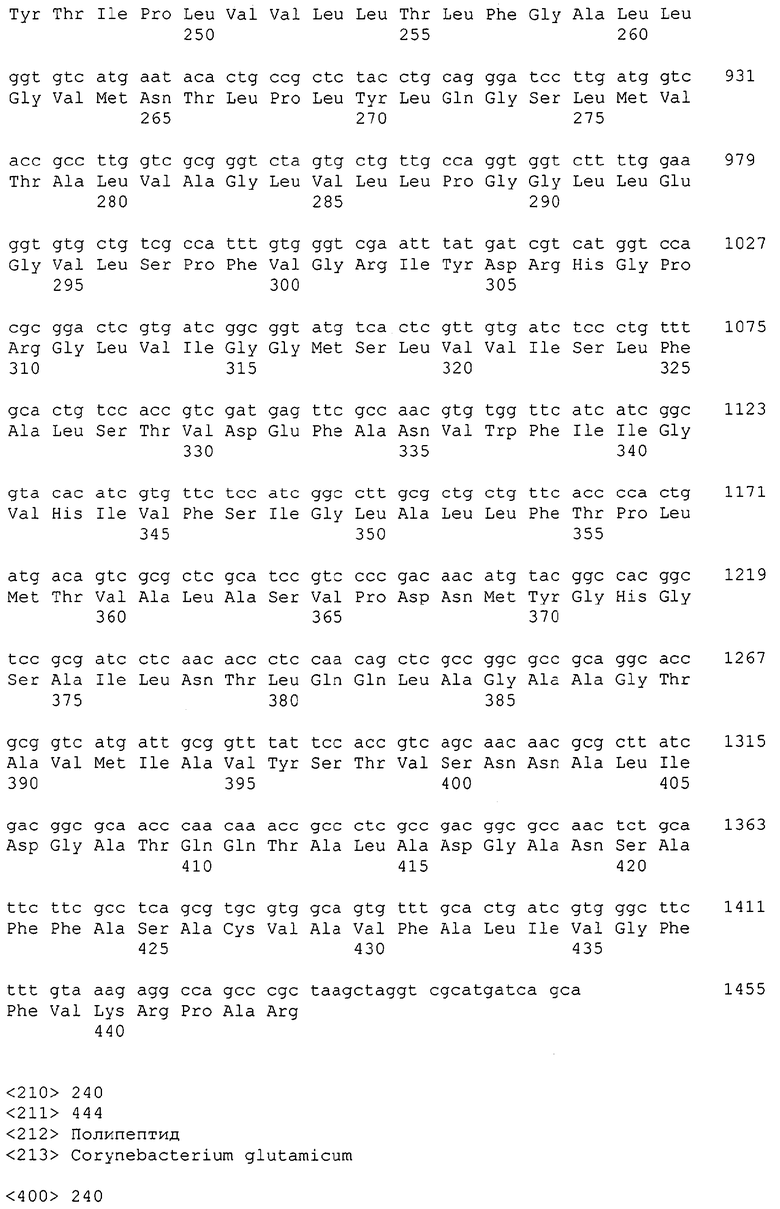



















































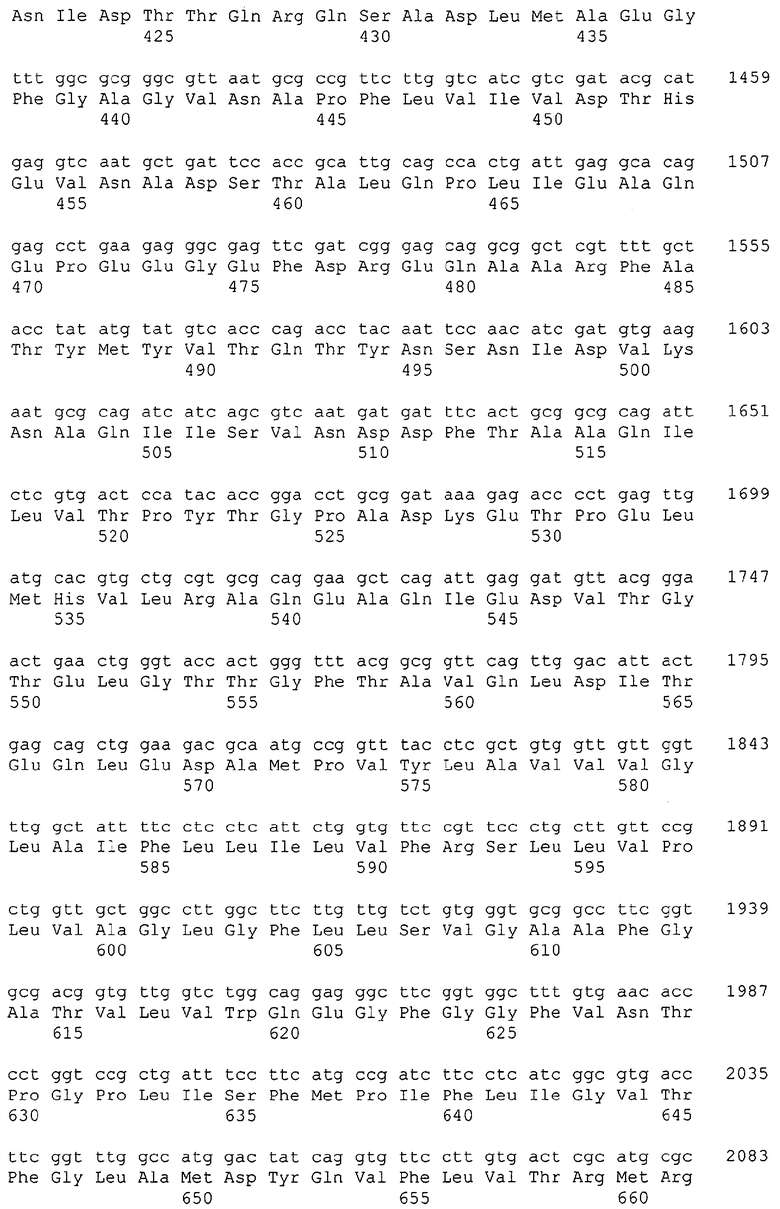






































Изобретение относится к биотехнологии и представляет собой выделенные молекулы нуклеиновой кислоты Corynebacterium glutamicum, которые кодируют полипептид с активностью белка резистентности к линкомицину. Изобретение относится к также рекомбинантным экспрессирующим векторам, содержащим такие молекулы нуклеиновых кислот, и клеткам-хозяевам, в которые были введены эти экспрессирующие векторы. Данное изобретение касается также способа получения аминокислот при помощи культивирования указанных клеток. Изобретение позволяет расширить ассортимент ферментов и получать аминокислоты с высокой степенью эффективности. 24 н. и 12 з.п. ф-лы, 4 табл.
Приоритет по пунктам:
| Способ изготовления носителей информации на полосковых магнитных доменах | 1978 |
|
SU752472A1 |
| PETER H | |||
| et al | |||
| Corynebacterium glutamicum is equipped with four secondary carriers for compatible solutes: identification, sequencing, and characterization of the proline/ectoine uptake system, ProP, and the ectoine/proline/glycine betaine carrier, EctP | |||
| J Bacteriol | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| WO 9902692 A, 21.01.1999 | |||
| ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ ПОЛИПЕПТИД С АКТИВНОСТЬЮ НИТРИЛГИДРАТАЗЫ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ НИТРИЛГИДРАТАЗЫ, ШТАММ БАКТЕРИЙ ESHCERICHIA COLI - ПРОДУЦЕНТ ПОЛИПЕПТИДА СО СВОЙСТВАМИ НИТРИЛГИДРАТАЗЫ, СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДА СО СВОЙСТВАМИ НИТРИЛГИДРАТАЗЫ | 1991 |
|
RU2081173C1 |

