ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к новым последовательностям моноклональных антител, пептидным последовательностям антигенов, с которыми связываются моноклональные антитела, а также к способам диагностики и терапии с использованием моноклональных антител и пептидов.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Совместная Международная Заявка, поданная в соответствии с РСТ (Договором о патентной кооперации), № публикации WO 95/20605, раскрывает сущность иммуностимулирующих моноклональных антител. Антитела, являющиеся предметом этой Международной Заявки, вырабатывались против лимфобластоидных В-клеток, и было показано, что они оказывают иммуностимулирующий эффект. При введении их животным-носителям опухолей было также обнаружено, что эти антитела оказывают противоопухолевый эффект.
Диагностика рака в соответствии с современными медицинскими процедурами типично является многостадийным процессом, включающим врачебный осмотр, применение различных техник визуализации, использование различных маркеров рака и т.д. В данной области техники давно существует потребность в методиках диагностики рака, которые обеспечивали бы выявление рака, а также определение типа рака, которым страдает обследуемый индивидуум.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение основано на открытии последовательностей моноклональных антител к лимфобластоидным клеткам. Настоящее изобретение, кроме того, основано на открытии того, что уровень связывания этих антител с Т-клетками больных раком является иным (более высоким или более низким), нежели уровень связывания этих антител с Т-клетками здоровых людей.
В соответствии с одним из аспектов изобретения предусмотрено моноклональное антитело, имеющее вариабельную область, выбранное из группы, состоящей из:
(а) моноклонального антитела, имеющего вариабельную область тяжелой цепи, включающую аминокислотную последовательность, изображенную на фиг.1;
(б) моноклонального антитела, имеющего вариабельную область легкой каппа-цепи, включающую аминокислотную последовательность, изображенную на фиг.2;
(в) моноклонального антитела, имеющего вариабельную область тяжелой цепи, включающую аминокислотную последовательность, изображенную на фиг.1, и вариабельную область легкой каппа-цепи, включающую аминокислотную последовательность, изображенную на фиг.2;
(г) моноклонального антитела, имеющего вариабельную область тяжелой цепи, обладающую, по меньшей мере, 70%-ной идентичностью аминокислотной последовательности, изображенной на фиг.1;
(д) моноклонального антитела, имеющего вариабельную область легкой цепи, обладающую, по меньшей мере, 70%-ной идентичностью последовательности, изображенной на фиг.2.
Согласно настоящему изобретению, термин "антитело" относится к моноклональным антителам любого из классов IgG, IgM, IgD, IgA и IgE. Термин относится к целым антителам или фрагментам антител, включающим антигенсвязывающий домен антитела, например, к антителам, у которых отсутствует Fc-фрагмент, одноцепочечным антителам, фрагментам антител, состоящим, по существу, только из вариабельного антигенсвязывающего домена антитела и т.п.
Кроме того, изобретение также относится к антителам, которые связываются с антигеном, с которым специфически связывается любое из вышеперечисленных MAT, то есть к антителам, которые обладают перекрестной реактивностью с вышеуказанными антителами.
В соответствии с одним из примеров осуществления изобретения моноклональное антитело (MAT) является химерным антителом человека и мыши, а именно - MAT с константной областью, полученной от человека, и вариабельной областью, полученной от мыши. С этой целью вариабельные области легких каппа-цепей и тяжелых цепей MAT согласно настоящему изобретению клонировали с помощью ПЦР, а их ДНК секвенировали. В соответствии с еще одним примером осуществления изобретения антитело является полностью гуманизированным антителом, то есть и вариабельная, и константная области его получены от человека.
Термин "имеющий, по меньшей мере, Х-процентную идентичность" относится к проценту остатков аминокислот, которые идентичны в двух сравниваемых последовательностях, если последовательности оптимально сопоставлены. Поэтому 70%-ная идентичность аминокислотной последовательности означает, что 70% аминокислот в двух или нескольких оптимально сопоставленных полипептидных последовательностях идентичны.
Предпочтительно, идентичность равна, по меньшей мере, 80%, более предпочтительно, по меньшей мере, 90%.
В соответствии с дополнительным аспектом изобретения обеспечены линии гибридомных клеток мыши, которые продуцируют какие-либо MAT согласно настоящему изобретению. Гибридомы могут быть получены любым из способов, известных в данной области техники (например, Kohler, G. and Milstein, С., Nature, 256:495-497 (1975)). Супернатант гибридомных клеточных линий подвергали типичному скринингу на антигенсвязывающую активность каким-либо из способов, известных в данной области техники, например, с помощью твердофазного иммуноферментного анализа (ELISA) или радиоиммунного анализа (RIA, РИА). Супернатанты подвергали скринингу на предмет продукции MAT, которые связываются с любым из пептидов согласно настоящему изобретению (что пояснено ниже) или которые связываются с клетками, с которыми связываются эти пептиды, например, клетками Дауди или Т-лимфоцитами.
Последовательности ДНК, кодирующие любую из аминокислотных последовательностей тяжелой цепи или легкой цепи вышеуказанных MAT, также включены в объем изобретения. Как, несомненно, будет ясно любому специалисту в данной области техники, из-за вырожденности генетического кода MAT согласно настоящему изобретению может кодировать множество нуклеотидных последовательностей, кроме последовательностей, изображенных на фиг.1 или 2.
Изобретение также обеспечивает экспрессирующие векторы, такие как плазмиды, включающие вышеуказанные ДНК-последовательности, а также клетки-хозяева, содержащие один или несколько этих экспрессирующих векторов.
В соответствии с другим аспектом изобретения обеспечены пептидные последовательности антигенов В-клеток, к которым могут присоединяться MAT согласно настоящему изобретению. Был проведен поиск в базе данных банка неизбыточных генов, и отдел EST определил, что эти пептидные последовательности являются новыми.
В соответствии с этим дополнительным аспектом изобретения предусмотрен пептид, выбранный из группы, состоящей из:
(а) пептида, имеющего аминокислотную последовательность, изображенную на фиг.10;
(б) пептида, имеющего аминокислотную последовательность, изображенную на фиг.11;
(в) пептида, имеющего аминокислотную последовательность, изображенную на фиг.12;
(г) пептида, обладающего, по меньшей мере, 85%-ной идентичностью любой из аминокислотных последовательностей пептидов по пунктам (а), (б) и (в), приведенным выше, и
(д) белка или пептида, включающего один или несколько пептидов по пунктам (а)-(г), приведенным выше.
Пептиды согласно настоящему изобретению могут быть использованы для различных диагностических анализов, например, таких как конкурентные иммунологические анализы, при которых определяют уровень связывания MAT согласно настоящему изобретению с их естественным антигеном, который присутствует на Т-клетках. Кроме того, пептиды можно использовать для выработки антител у иммунизированных животных, причем эти антитела затем можно использовать для любой из целей, описанных выше и ниже.
Аналоги всех вышеуказанных пептидов также образуют дополнительный аспект настоящего изобретения. Как будет ясно специалисту в данной области техники, аминокислотные последовательности пептидов согласно настоящему изобретению могут быть изменены, например, посредством добавления, делеции или консервативного или неконсервативного замещения одной или нескольких аминокислот без существенного нарушения способности пептидов связывать антитела.
Термин "консервативное замещение" относится к замещению аминокислоты одного класса аминокислотой того же класса, где класс определяется одинаковыми физико-химическими свойствами боковых цепей аминокислот и высокой частотой замещений в гомологичных белках, встречающихся в природе, что можно определить, например, с помощью стандартной матрицы частоты обмена Дейхоффа или матрицы BLOSUM. [Было охарактеризовано шесть основных классов аминокислот, которые включают: Класс I (Cys); Класс II (Ser, Thr, Pro, Ala, Gly); Класс III (Asn, Asp, Gin, Glu); Класс IV (His, Arg, Lys); Класс V (Ile, Ley, Val, Met) и Класс VI (Phe, Туr, Тrр)]. Например, замещение Asp на другой остаток Класса III, такой как Asn, Gln или Glu, является консервативным замещением. Термин "неконсервативное замещение "относится к замещению аминокислоты одного класса аминокислотой из другого класса; например, замещение Ala, остатка Класса II, остатком Класса III, таким как Asp, Asn, Glu или Gln.
Отдельные буквы, использованные выше (и ниже) для обозначения специфических аминокислот (ак), соответствуют 1-буквенным символам аминокислот, рекомендованным Комиссией по биохимической номенклатуре IUPAC-IUB.
Аналогами вышеуказанных пептидов, которые попадают в объем настоящего изобретения, являются такие пептиды, которые имеют примерно такой же уровень связывания с MAT согласно настоящему изобретению, как пептиды, изображенные на фиг.10-12. Уровень связывания можно определить любым способом, известным в данной области техники.
Пептиды и аналоги согласно настоящему изобретению можно также модифицировать химически, и такие химически модифицированные пептиды и их аналоги также образуют часть изобретения. Термин "химически модифицированный" относится к белку, в котором по меньшей мере один из его аминокислотных остатков модифицирован либо посредством естественных процессов, таких как процессинг или другие посттрансляционные модификации, либо с помощью методик химической модификации, которые хорошо известны специалистам в данной области техники. Среди многочисленных известных модификаций типичные, но не исключительные, примеры включают: ацетилирование, ацилирование, аминирование, АДФ-рибозилирование, гликозилирование, образование GPI-якорей, ковалентное присоединение жидкостей или производных липидов, метилирование, миристилирование, пегилирование, пренилирование, фосфорилирование, убихинонирование и любые сходные процессы.
Вторым открытием, на котором основано изобретение, является то, что MAT согласно настоящему изобретению могут в различной степени связываться с Т-клетками, полученными от людей, имеющих злокачественное заболевание, по сравнению со степенью связывания тех же MAT с Т-клетками здоровых людей.
Поэтому, в соответствии с еще одним аспектом настоящего изобретения, предусмотрен анализ для идентификации обследованных людей с высокой вероятностью наличия злокачественного заболевания, включающий:
(а) получение пробы жидкости организма от вышеуказанного человека;
(б) контакт вышеуказанной пробы по меньшей мере с одним MAT согласно настоящему изобретению;
(в) определение степени связывания вышеуказанных MAT с Т-клетками, содержащимися в вышеуказанной пробе, и
(г) сравнение степени связывания, определенной на этапе (в), со степенью связывания MAT согласно настоящему изобретению с Т-клетками, содержащимися в пробе, полученной от здорового человека;
достоверное различие между двумя степенями связывания указывает на то, что существует высокая вероятность наличия у обследованного человека злокачественного заболевания.
В соответствии с изобретением пробой, полученной от человека, которому необходимо провести анализ, может быть любая жидкость организма, которая содержит обнаружимое количество Т-клеток. В типичном случае проба жидкости организма является пробой крови или лимфатической жидкости. Предпочтительно, перед контактом MAT согласно настоящему изобретению с полученной пробой мононуклеарные клетки периферической крови (МКПК), содержащиеся в пробе, отделяют с помощью любого из способов, известного в данной области техники, например, с помощью центрифугирования в градиенте плотности Ficoll Hypaque, а затем отделенные клетки приводят в контакт с исследуемыми антителами.
Термин "злокачественное заболевание" в соответствии с изобретением следует понимать как любое злокачественное заболевание, известное в данной области техники, на любой его стадии.
Этот термин также охватывает злокачественные заболевания, которые находятся на ранней стадии и еще не вызывают клинических симптомов. Предпочтительно, этот термин относится к плотным опухолям.
Термин "здоровый человек" относится к человеку, у которого нет злокачественного заболевания, и может также относиться к среднему значению для нескольких человек или к значению, полученному при образовании пула из жидкостей организма, полученных от нескольких человек. Следует отметить, что, если стандартная степень связывания для здоровых людей определена, нет необходимости повторно определять этот стандарт при каждом анализе, и полученное число можно использовать постоянно. Согласно изобретению, было обнаружено, что у здоровых людей примерно 25% CD3+ Т-клеток связывают антитела согласно настоящему изобретению.
Термин "высокая вероятность" означает, что анализ согласно настоящему изобретению является анализом, выполняемым при первичном скрининге и способным выявить людей, у которых предположительно имеется злокачественное заболевание. Факт действительного наличия злокачественного заболевания у человека, выявленного с помощью способа согласно настоящему изобретению, должен быть позже подтвержден с использованием дополнительных методик, известных в данной области техники.
Термин "степень связывания" относится к уровню связывания антитела с антигеном, предъявляемым Т-клетками обследованного человека, причем этот уровень можно определить любым из известных в данной области техники способов определения уровней связывания антител, например ELISA или вестерн-блоттингом. Степень связывания можно определить с помощью любой системы обнаружения, например с помощью иммуноглобулина к антигенам мыши или его фрагментов, соединенных с обнаружимым маркером. Примерами таких обнаружимых маркеров являются радиоактивная группа, флуоресцирующая группа, фермент, способный катализировать реакцию, продуктом которой является обнаружимый продукт (например, цветную реакцию), биотиновая группа, которую можно выявить с помощью авидина, и т.д. В предпочтительном примере осуществления изобретения степень связывания MAT согласно настоящему изобретению с Т-клетками определяют с помощью двойной маркировки, при которой антитело к Т-клетке (например, антитело к СО3+ клетке) присоединяют к одному флуоресцентному маркеру, a MAT согласно настоящему изобретению - к другому типу флуоресцентного маркера. Степень связывания затем определяют с помощью устройства для сортировки клеток, активированных флуоресцеином (FACS). Количественную оценку степени связывания производят посредством определения процента CD3+ Т-клеток (определяемых по их связыванию с антителами к CD3+ клеткам), которые также связывают MAT согласно настоящему изобретению.
В соответствии с изобретением, было обнаружено, что общее количество CD3+ клеток в пробах крови людей, имеющих злокачественные заболевания, сходно с количеством CD3+ клеток в пробах крови, полученных от здоровых людей, так что нормализация степени связывания MAT и CD3+ Т-клеток с помощью общего числа CD3+ связывающих Т-клеток у больных раком и здоровых людей правомерна. Тем не менее, процент CD3+ связывающих Т-клеток, которые связывают также и MAT согласно изобретению (далее: "CD3+ MAT клетки") у людей, имеющих злокачественные заболевания, достоверно отличается от процента CD3+ MAT клеток в крови здоровых людей. Процент CD3+ MAT клеток у человека, имеющего злокачественное заболевание, может быть достоверно более высоким или достоверно более низким, чем процент СD3+ MAT клеток в крови здоровых людей, в зависимости от типа злокачественного заболевания.
Степень связывания MAT согласно настоящему изобретению с Т-клетками, полученными от обследованного человека, будет признана "достоверно отличающейся" от степени связывания с Т-клетками, полученными от здорового человека, если различие в связывании MAT статистически достоверно при оценке с помощью любого из статистических методов, известных в данной области техники (например, с помощью t-критерия Стьюдента), которые используются для оценки результатов, полученных с помощью экспериментальных методов, указанных здесь.
Изобретение не только позволяет выявить людей с высокой вероятностью наличия у них какого-либо типа злокачественного заболевания (поскольку больные люди имеют другую степень связывания Т-клеток с MAT согласно настоящему изобретению, по сравнению со здоровыми людьми), но может также помочь идентифицировать людей с определенным типом рака посредством определения того, является ли вышеуказанная степень связывания более высокой или более низкой, чем соответствующая степень связывания у здоровых людей.
В типичном случае, процент связывания MAT согласно настоящему изобретению с Т-клетками, полученными от здоровых людей, составляет примерно 25%, то есть 25% клеток, экспрессирующих маркер CD3+ Т-клеток (определяемый по связыванию с антителом к CD3+ клеткам), также связывают и MAT согласно настоящему изобретению.
Согласно настоящему изобретению, было показано, что в пробах, полученных от больных раком предстательной железы, процент CD3+ Т-клеток, с которыми связываются MAT согласно настоящему изобретению, составляет примерно 50%.
Кроме того, было показано, что, если CD3+ Т-клетки происходят из проб, взятых у больных раком толстого кишечника или молочной железы, то процент клеток, которые также присоединяют MAT согласно настоящему изобретению, составляет примерно 7 и 10% соответственно.
Поэтому, согласно настоящему изобретению, появилась возможность определить высокую вероятность наличия специфического типа рака по образцу жидкости организма, полученному от обследованного человека, с помощью простого и одиночного анализа, основанного на степени связывания MAT согласно настоящему изобретению с CD3+ клетками, присутствующими в образце жидкости организма. Простота диагностического анализа согласно настоящему изобретению, для которого достаточно использования только одного вида MAT для идентификации человека, имеющего определенный тип рака, очень хорошо подходит для широкого скрининга населения.
Поэтому настоящее изобретение еще в одном из его аспектов предусматривает анализ для выявления обследованного человека с высокой вероятностью наличия у него специфического злокачественного заболевания, включающий:
(а) получение пробы жидкости организма от вышеуказанного человека;
(б) контакт вышеуказанной пробы с MAT согласно настоящему изобретению;
(в) определение степени связывания вышеуказанных MAT с Т-клетками, содержащимся в вышеуказанной пробе, и
(г) сравнение степени связывания, определенной на этапе (в), полученных клеток со степенью связывания MAT с Т-клетками, полученными от здорового человека; достоверное различие между степенями связывания с высокой вероятностью указывает на то, что у обследованного человека имеется злокачественное заболевание, причем то, является ли степень связывания MAT с Т-клетками, полученными от вышеуказанного человека, более высокой или более низкой, чем степень связывания MAT с Т-клетками здоровых людей, указывает на специфический тип злокачественного заболевания, которое с высокой вероятностью имеется у обследованного человека.
В частности, если степень связывания с MAT согласно настоящему изобретению достоверно выше, чем у здоровых людей, существует высокая вероятность, что у обследованного человека имеется рак предстательной железы.
Если степень связывания достоверно ниже, чем у здорового человека, существует высокая вероятность, что у обследованного человека имеется рак толстого кишечника или молочной железы.
Согласно диагностическому аспекту изобретения, составы (композиции), включающие MAT согласно настоящему изобретению, могут быть использованы для диагностики с целью выявления людей, у которых с высокой вероятностью имеется злокачественное заболевание (вообще), или для идентификации специфического злокачественного заболевания, которое, вероятно, имеется у человека. Поэтому изобретение в еще одном из его аспектов предусматривает диагностический состав, включающий MAT, относящиеся, по меньшей мере, к одному из вышеуказанных антител, совместно с подходящим носителем. Носитель может быть или растворимым носителем, например, каким-либо из физиологически приемлемых буферных растворов, известных в данной области техники (например, фосфатный буферный раствор (ФБР)), или твердым носителем, таким как, например, латексные шарики.
Настоящее изобретение также обеспечивает наборы, например, наборы для диагностических анализов, предназначенные для использования MAT согласно настоящему изобретению и выполнения диагностических анализов, сущность которых раскрыта выше. В одном из примеров осуществления изобретения диагностический набор может обычно включать по меньшей мере одно из вышеуказанных MAT в одной или нескольких емкостях, конъюгат вещества-партнера, специфически связывающегося с MAT (например, антигена или его аналога согласно настоящему изобретению), метку, способную давать обнаружимый сигнал, и инструкции по их использованию. Метка может, априорно, связываться с моноклональным антителом, или, альтернативно, метка может связываться с молекулой-носителем, которая затем специфически связывается с МАТ. Инкубации анализируемой пробы с составом, являющимся диагностическим реактивом, в течение определенного времени достаточно для связывания моноклональных антител с клетками.
В следующем аспекте изобретения предусмотрены фармацевтические композиции, включающие, в качестве активного ингредиента, одно или несколько MAT согласно настоящему изобретению. Использование вышеуказанных MAT для приготовления фармацевтических препаратов для лечения различных злокачественных заболеваний у человека также входит в объем изобретения.
Еще в одном аспекте настоящее изобретение предусматривает метод лечения злокачественных заболеваний посредством введения нуждающемуся в этом человеку терапевтически эффективного количества вышеуказанных МАТ. Терапевтически эффективное количество - это количество, способное облегчать симптомы злокачественного заболевания, снижая симптомы или полностью их устраняя.
Фармацевтические композиции, включающие пептиды согласно настоящему изобретению, также образуют аспект изобретения. Такие композиции могут использоваться, например, для активной иммунизации человека с целью получения антител, которые могут затем связываться с Т-клетками человека и вызывать иммунный ответ у человека.
Далее основные аспекты изобретения будут описаны с периодическими ссылками на прилагаемые графические материалы. В следующем ниже описании и графических материалах термин "ВАТ антитело" будет использован как равнозначный термину "MAT согласно изобретению".
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг.1 демонстрирует ДНК и пептидные последовательности вариабельной области тяжелой цепи MAT согласно настоящему изобретению.
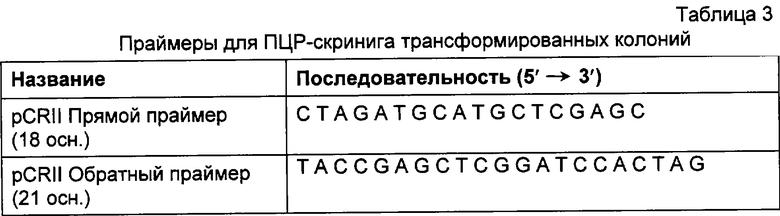
Фиг.2 демонстрирует ДНК и пептидные последовательности вариабельной области легкой каппа-цепи MAT согласно настоящему изобретению.
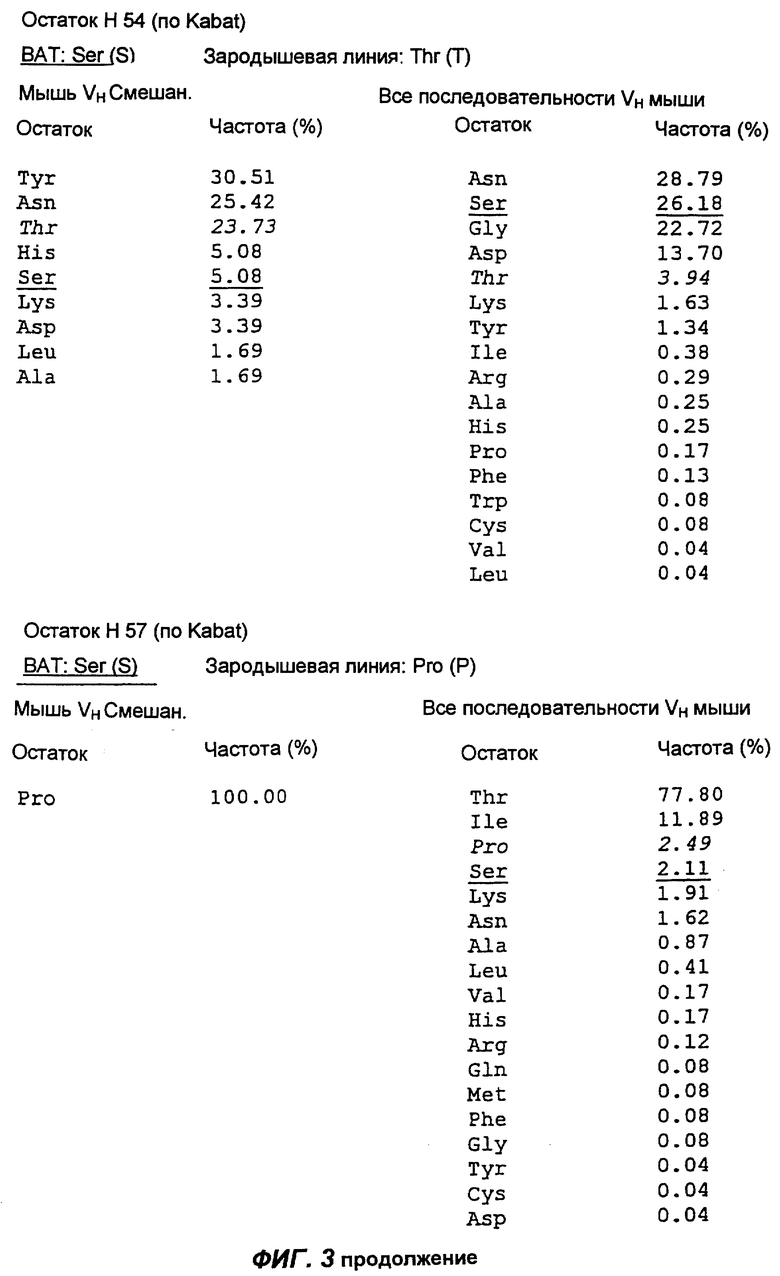
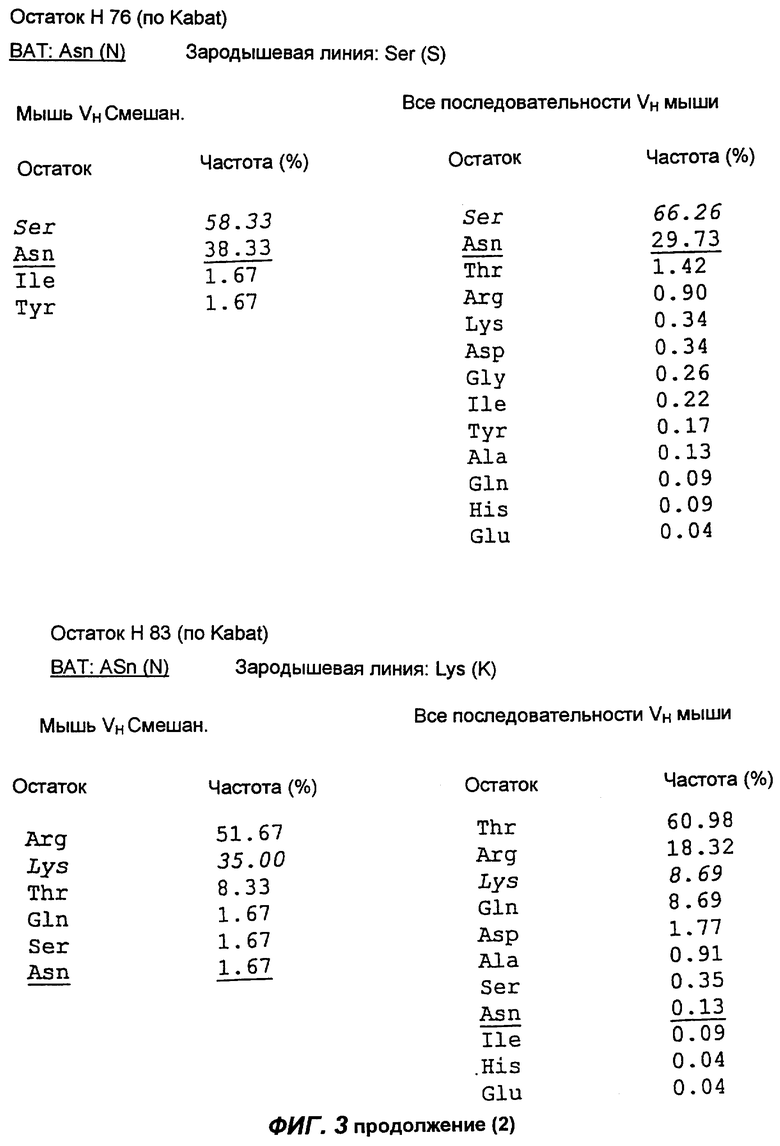
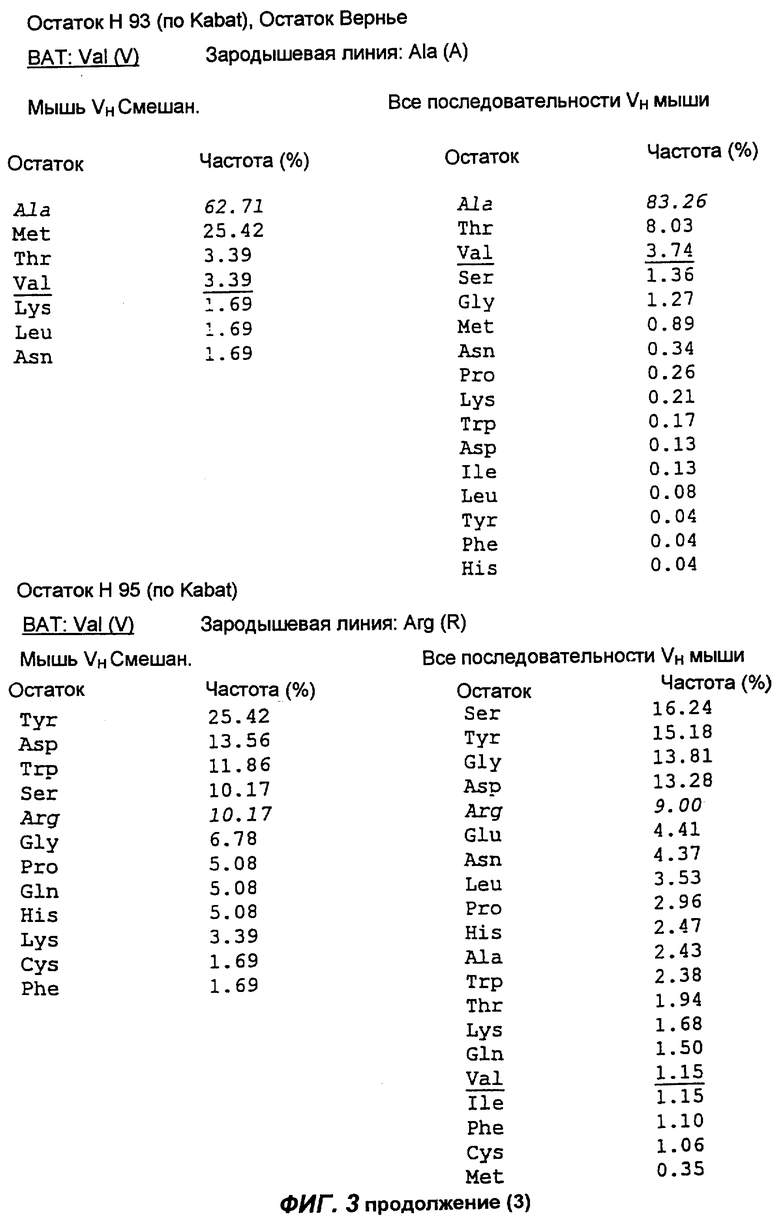
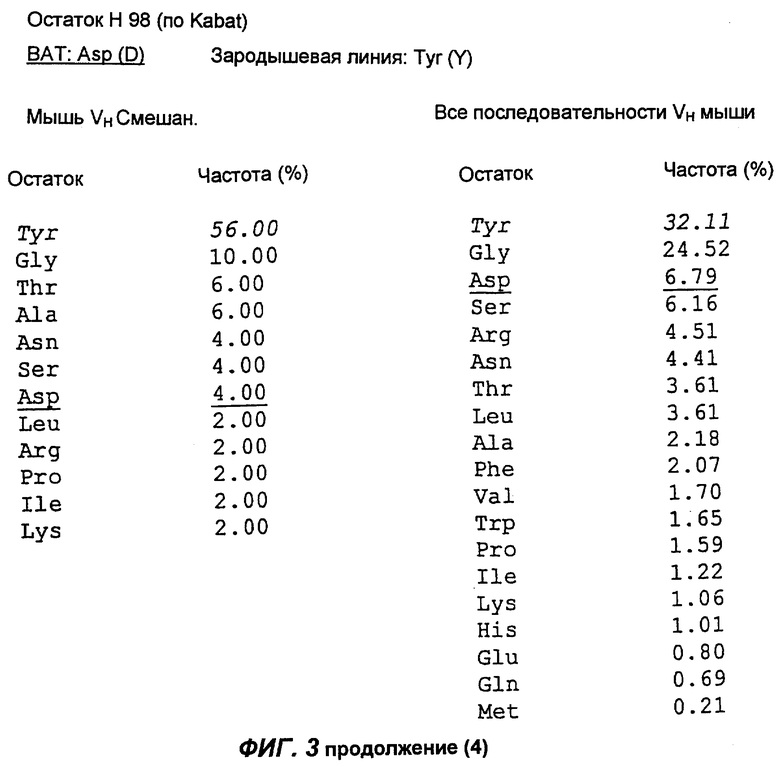
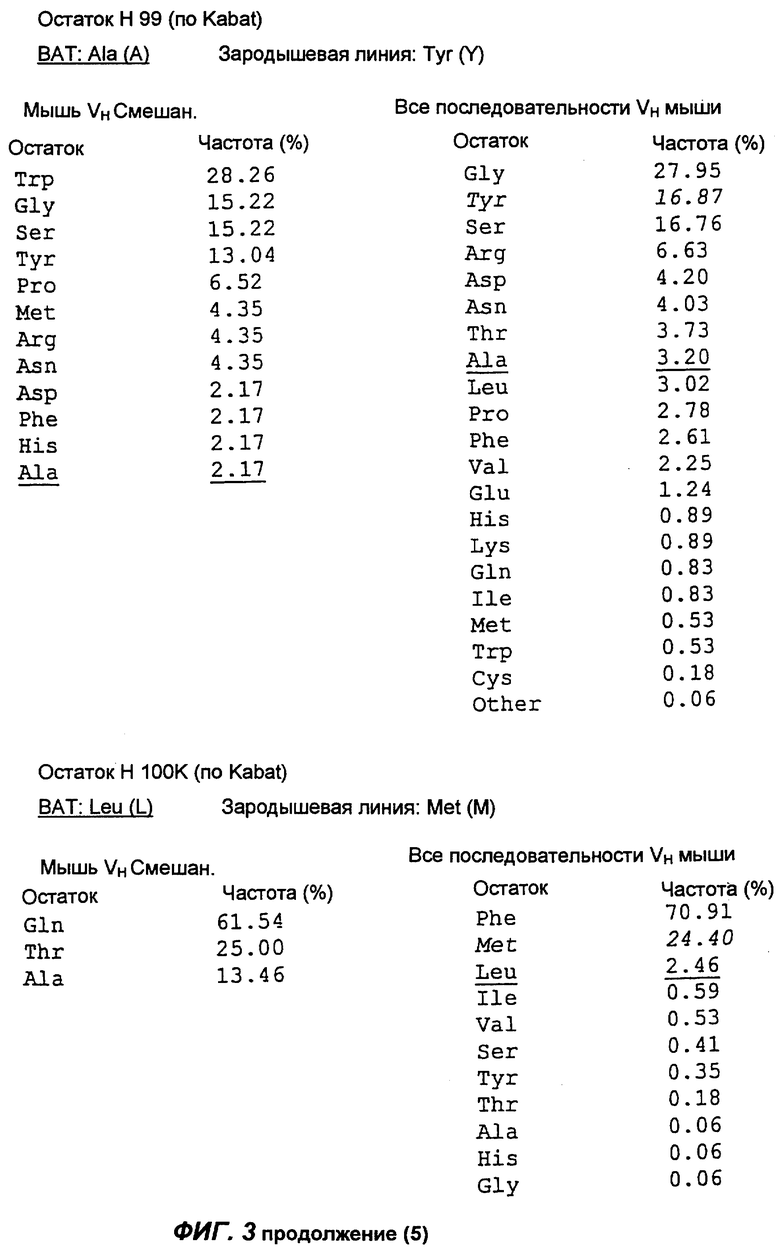
Фиг.3 демонстрирует анализ аминокислотной последовательности вариабельной области тяжелой цепи антитела согласно настоящему изобретению (обозначенной как "ВАТ"). "ВАТ" обозначает аминокислотную последовательность Vн-области ВАТ антитела, в то время как "VMS2" обозначает аминокислотную последовательность гена VMS2/VGK4 зародышевой линии клеток. Если ВАТ-последовательность и последовательность зародышевой линии идентичны, последовательность зародышевой линии обозначают точкой (.); если существуют несоответствия, указан отличающийся остаток зародышевой линии. Таблицы, приведенные ниже, и последовательности, приведенные на следующих страницах, описывают частоту, с которой определенные аминокислоты можно обнаружить в определенном положении остатка, как в смешанной подгруппе тяжелых цепей мыши (Мышь vн Смешан.) (Kabat et al., Sequences of proteins of immunological interest, (1991)), так и в большей по объему базе данных всех известных последовательностей VH мыши (все последовательности VH мыши).
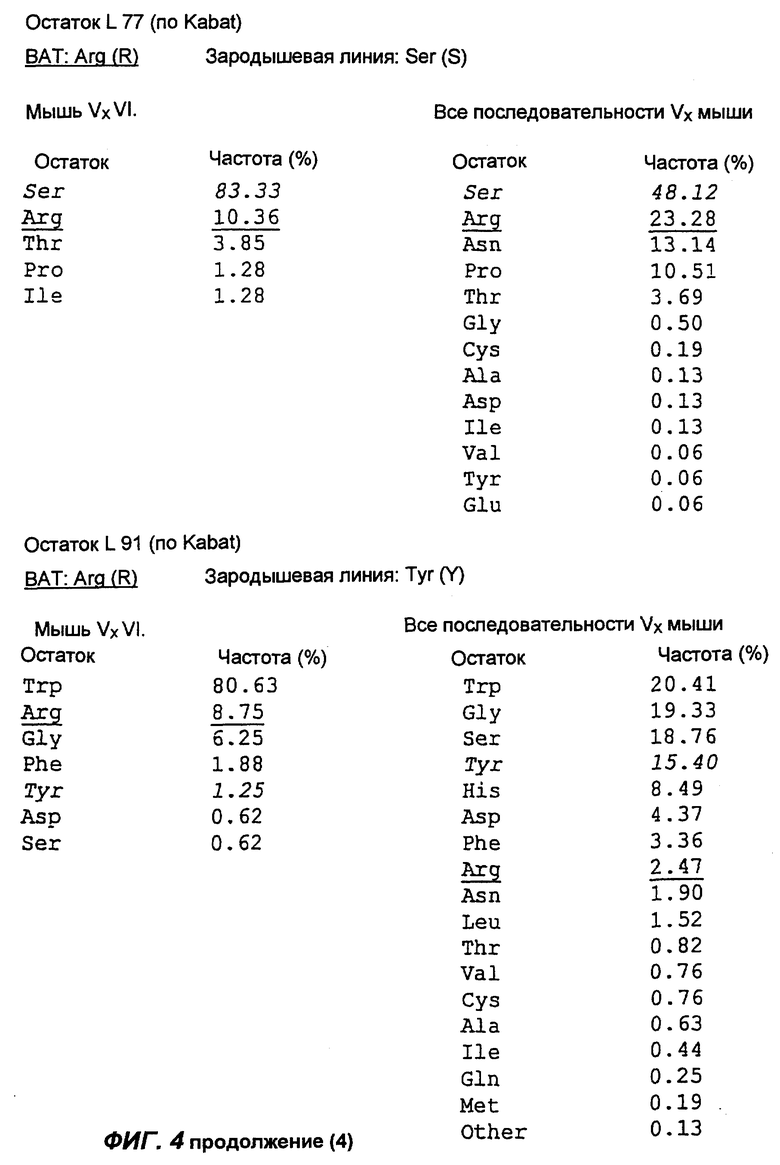
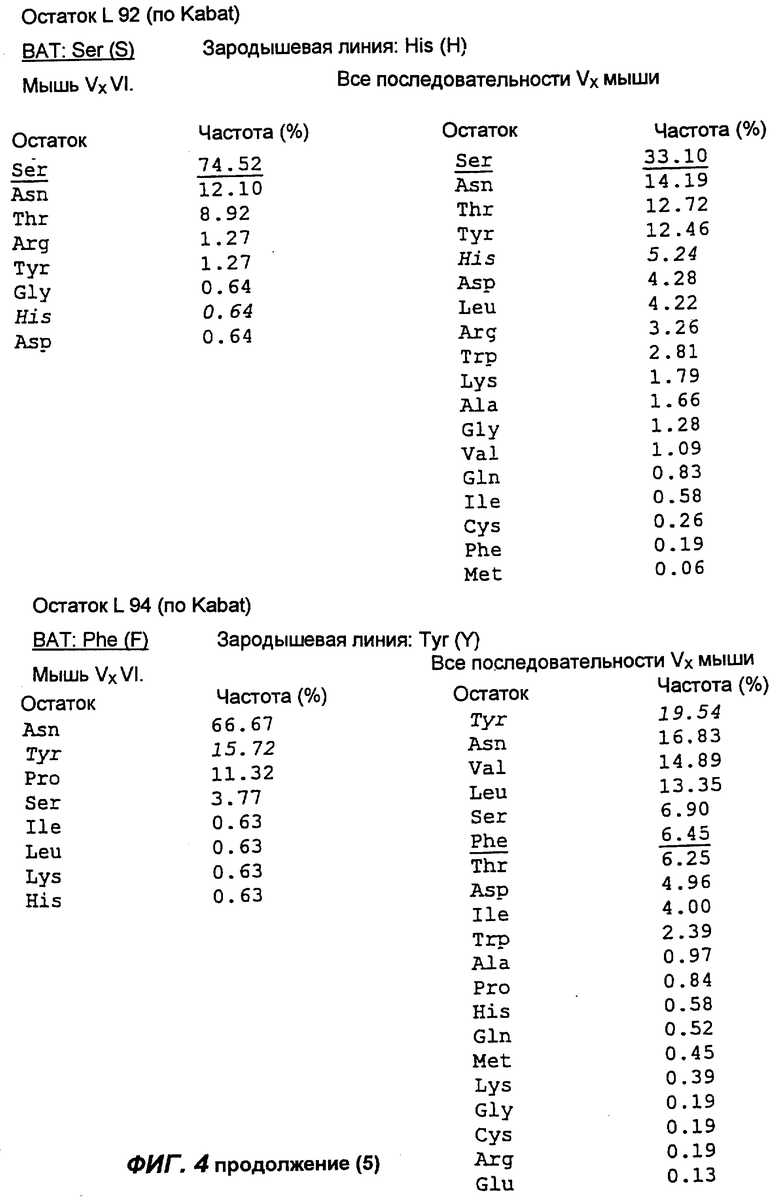
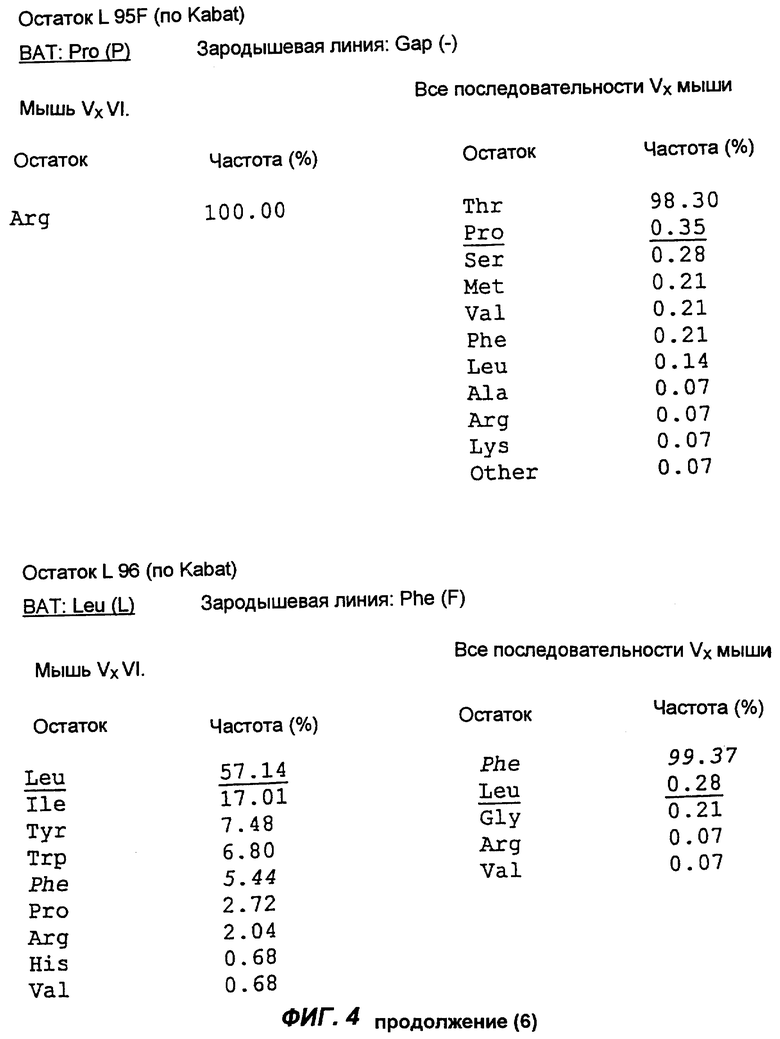
Фиг.4 демонстрирует результаты анализа аминокислотной последовательности вариабельной области легкой каппа-цепи антитела согласно настоящему изобретению (обозначенного на фиг.4 как "ВАТ"). "Мышь" обозначает аминокислотную последовательность КК области ВАТ антитела, тогда как "Зародыш" обозначает аминокислотную последовательность гена Н4 зародышевой линии клеток. Если ВАТ последовательность и последовательность зародышевой линии идентичны, последовательность зародышевой линии обозначена точкой (.); если существуют несоответствия, указан отличающийся остаток для зародышевой линии. Таблицы, приведенные ниже и на следующих страницах, описывают частоту, с которой можно обнаружить определенные аминокислоты в определенном положении остатка, как в подгруппе VI тяжелых цепей мыши согласно Kabat, так и в большей по объему базе данных всех известных последовательностей VК мыши (все последовательности VК мыши).
Фиг.5 демонстрирует ДНК и пептидные последовательности вариабельных областей легкой каппа-цепи химерного антитела согласно настоящему изобретению.
Фиг.6 демонстрирует ДНК и пептидные последовательности вариабельной области тяжелой цепи химерного антитела согласно настоящему изобретению.
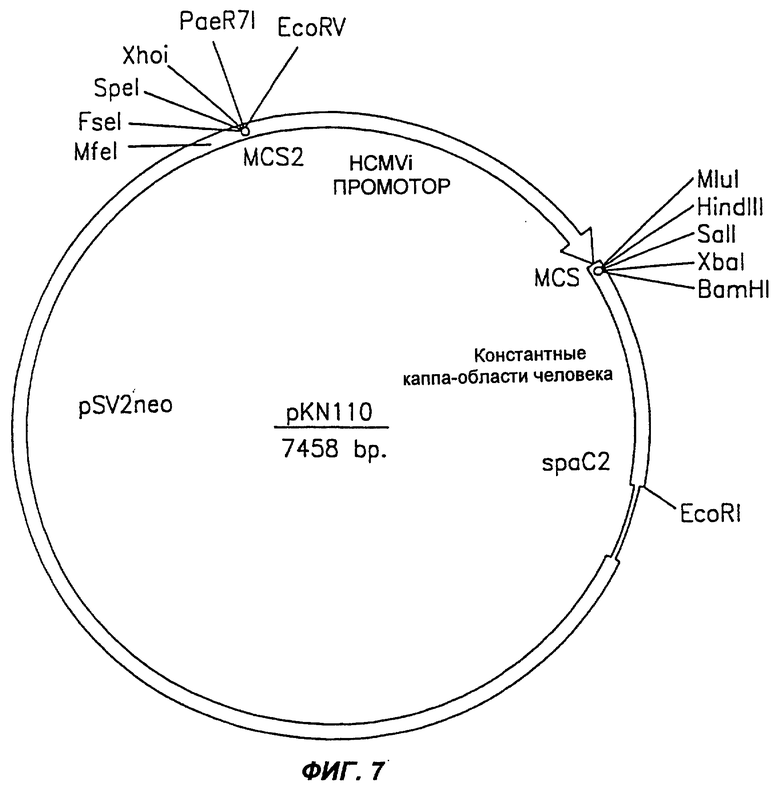
Фиг.7 демонстрирует схематическое изображение экспрессирующего вектора pKN 110 млекопитающих, используемого для экспрессии легкой каппа-цепи химерного антитела согласно настоящему изобретению.
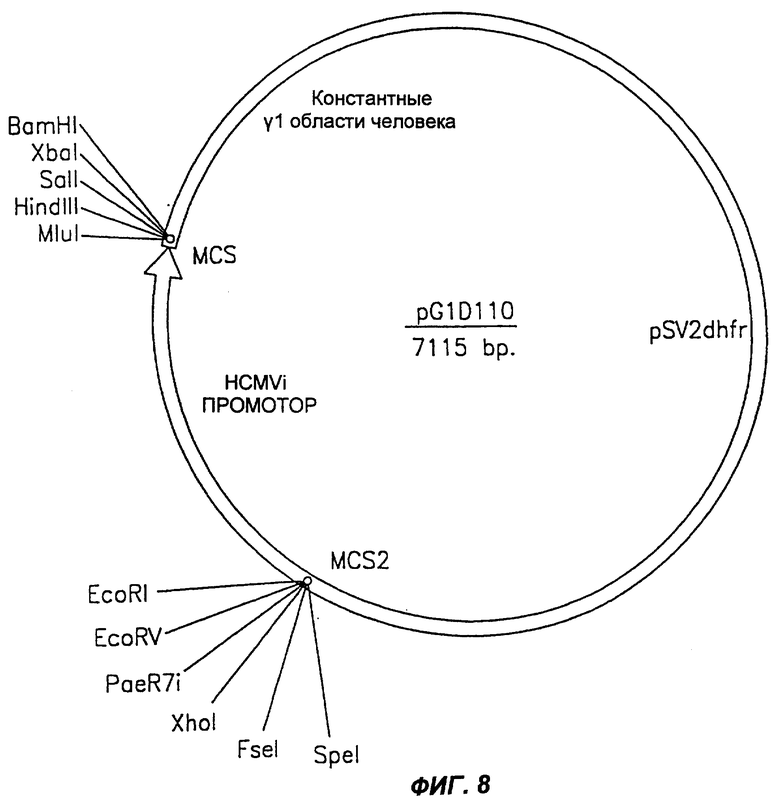
Фиг.8 демонстрирует схематическое изображение экспрессирующего вектора pG1D 110 млекопитающих, используемого для экспрессии тяжелой цепи химерного антитела согласно настоящему изобретению.
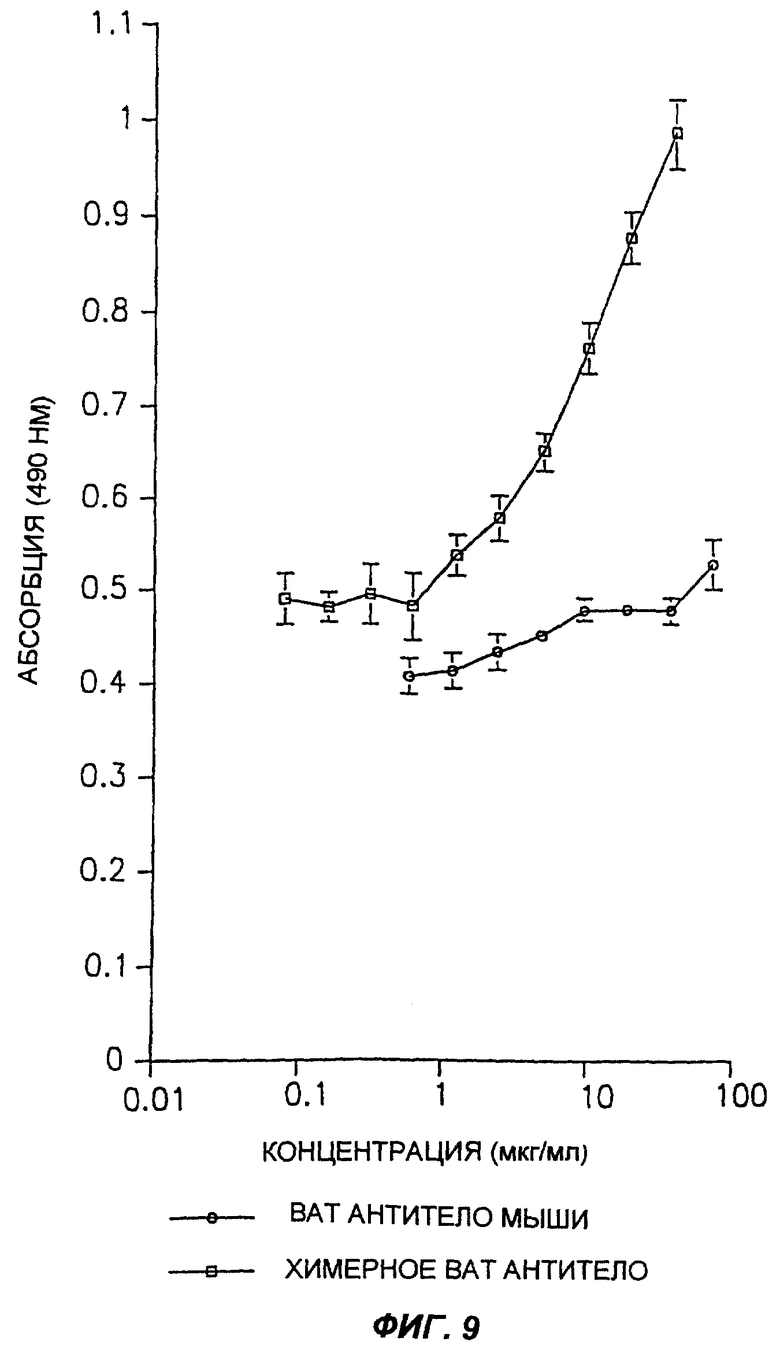
Фиг.9 демонстрирует возможности графического представления результатов анализа ELISA, измеряющего характеристики связывания химерного антитела мыши и γ1/Каппа согласно настоящему изобретению с клетками Дауди.
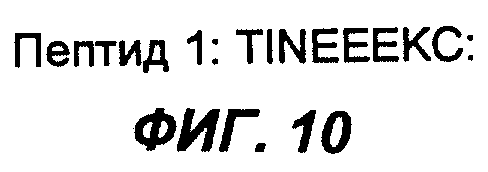
Фиг.10 демонстрирует аминокислотную последовательность пептида 1 согласно настоящему изобретению.
Фиг.11 демонстрирует аминокислотную последовательность пептида 2 согласно настоящему изобретению.
Фиг.12 демонстрирует аминокислотную последовательность пептида 3 согласно настоящему изобретению.
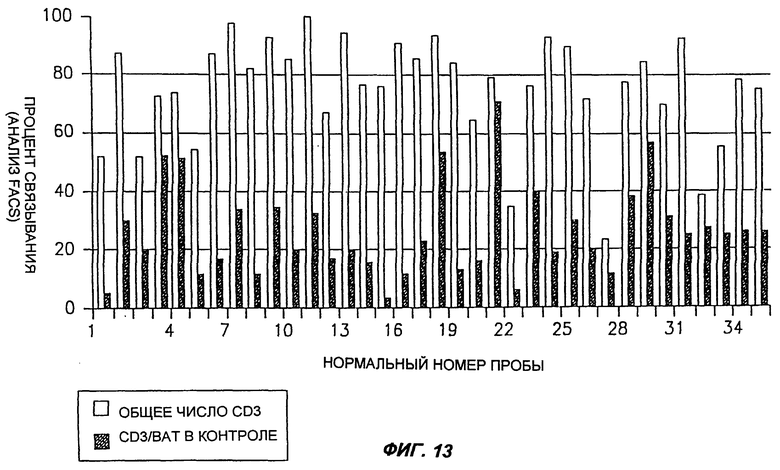
Фиг.13 является схематическим изображением, демонстрирующим процент CD3+ клеток, которые также связывают MAT согласно настоящему изобретению (обозначенные как "ВАТ"), по сравнению с общим числом CD3+ клеток в пробах крови здоровых людей, определенный с помощью анализа ФАКС.
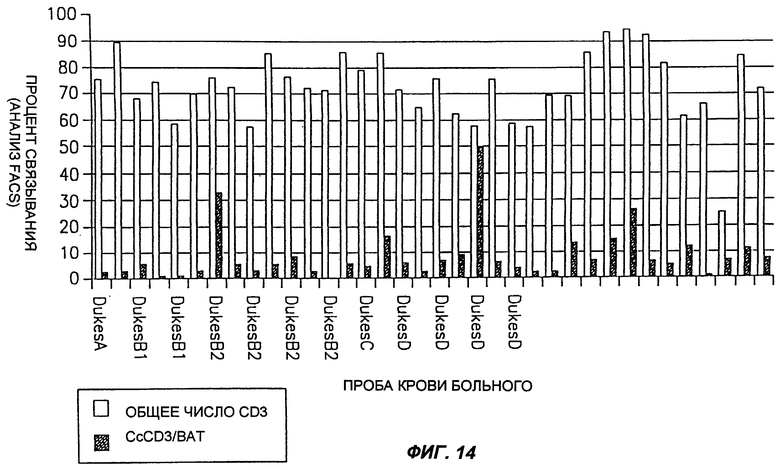
Фиг.14 демонстрирует процент CD3+ клеток, которые также связывают MAT согласно настоящему изобретению (обозначенные как "ВАТ"), по сравнению с общим числом CD3+ клеток в пробах крови, полученных от больных, имеющих карциному толстого кишечника, определенный с помощью анализа ФАКС.
Фиг.15 демонстрирует процент CD3+ клеток, которые также связывают MAT согласно настоящему изобретению (обозначенные как "ВАТ"), по сравнению с общим числом CD3+ клеток в пробах крови, полученных от больных, имеющих карциному молочной железы.
Фиг.16 демонстрирует процент CD3+ клеток, которые также связывают MAT согласно настоящему изобретению (обозначенные как "ВАТ"), по сравнению с общим числом CD3+ клеток в пробах крови, полученных от больных, имеющих карциному предстательной железы.
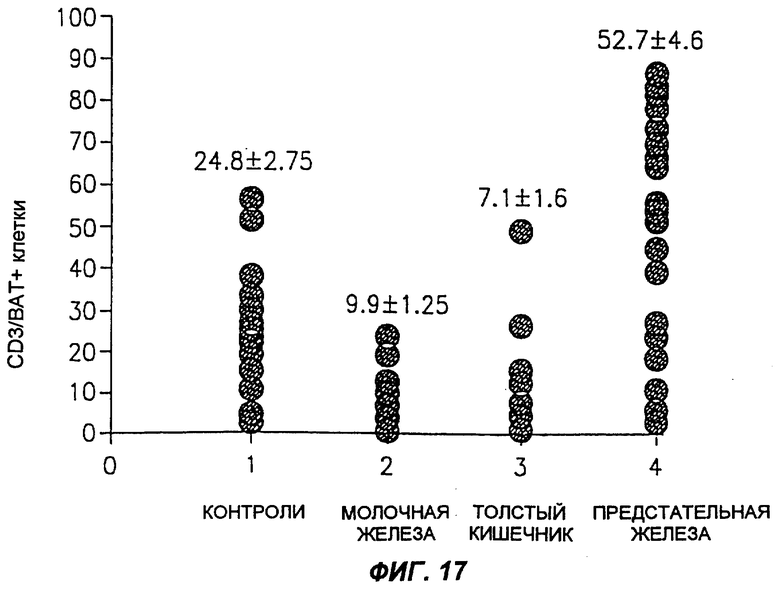
Фиг.17 является схематическим изображением, демонстрирующим средний процент CD3+ клеток, которые связывают MAT согласно настоящему изобретению (обозначенные как ВАТ), у здоровых людей, по сравнению с больными, имеющими карциному молочной железы, карциному толстого кишечника или карциному предстательной железы.
Фиг.18 является фотографией результатов вестерн-блоттинга пептидов, полученных из Т-клеток людей, имеющих рак предстательной железы, оториноларингологические карциномы (ЛОР), карциному молочной железы, или пептидов, полученных из мембран клеток Дауди. Блот инкубировали с MAT согласно настоящему изобретению и выявили повышенную концентрацию антигена в Т-клетках, полученных от больных, имеющих карциному предстательной железы, по сравнению с необнаружимым уровнем антигена в Т-клетках, полученных от больных, имеющих карциному молочной железы.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
I. СЕКВЕНИРОВАНИЕ MAT
(А) Аббревиатуры
Фетальная сыворотка телят (ФСТ); рибонуклеиновая кислота (РНК); информационная РНК (иРНК); дезоксирибонуклеиновая кислота (ДНК); копированная ДНК (кДНК); полимеразная цепная реакция (ПЦР); минута (мин); секунда (с); Трис-боратный буфер (ТББ).
(Б) Материалы
Компоненты сред и все остальные материалы для культур тканей были получены из компании Life Technologies (Великобритания). Набор для выделения РНК был получен из компании Stratagene (США), тогда как набор для синтеза 1-й нити кДНК был закуплен в компании Pharmacia (Великобритания). Все компоненты и оборудование для ПЦР-реакций, включая AmpliTaq® ДНК-полимеразу, были закуплены в компании Perkin Elmer (США). Набор ТА Cloning® был получен из компании Invitrogen (США). Агароза (UltraPure™) была получена из компании Life Technologies (Великобритания). Предварительно составленный набор для циклического секвенирования Thermo Sequences™ и аппарат для секвенирования ДНК Vistra 725 были закуплены в компании Amersham (Великобритания). Все остальные продукты для молекулярной биологии были получены из компании New England Biolabs (США).
(В) Методики эксперимента: ПЦР-клонирование и секвенирование генов вариабельной области ВАТ антитела мыши
Клеточная линия гибридомных клеток ВАТ мыши и линия клеток Дауди были успешно перенесены на MRC-CC, и обе линии клеток культивировали в суспензии с использованием RPMI (без глутамина) с добавлением 10% (объемных) ФСТ, 100 единиц/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ L-глутамина, 1 мМ пирувата натрия и 12,5 единиц/мл нистатина.
Было накоплено примерно 108 жизнеспособных клеток гибридомной клеточной линии ВАТ, и из 108 клеток была выделена общая РНК с помощью набора для выделения РНК в соответствии с инструкциями изготовителя. В наборе была использована одностадийная процедура экстракции гуанидиния тиоцианатом-фенолом-хлороформом, описанная Chromczynski and Sacchi, Anal. Biochem., 162:156, 1987. Также в соответствии с инструкциями изготовителя набор для синтеза 1-й нити кДНК был использован для получения однонитевой ДНК-копии мРНК ВАТ-гибридомы с помощью Notl-(dT)18 праймера, поставляемого в наборе. В каждом объеме конечной реакционной смеси, равном 33 мкл, использовалось примерно 5 мкг общей РНК. После завершения реакции реакционную смесь нагревали до 90°С в течение 5 мин для денатурации дуплекса РНК-кДНК и инактивации обратной транскриптазы перед охлаждением на льду.
Для ПЦР-амплификации гена вариабельной области тяжелой цепи мыши (VН гена) и гена вариабельной области легкой каппа-цепи мыши (VК гена) из линии гибридомных клеток использовали метод, описанный Jones and Bendig, Bio/Technology, 9:8, 1987. По существу, были использованы две серии вырожденных праймеров - один, предназначенный для гибридизациии лидерных последовательностей генов тяжелой цепи мыши (то есть MHV1-12; таблица 1), и второй, предназначенный для гибридизации лидерных последовательностей генов легкой каппа-цепи мыши (то есть MKV1-11; таблица 2), вместе с праймерами, предназначенными для гибридизации 5'-конца соответствующего гена константной области, для ПЦР-клонирования генов вариабельной области мышей.
Отдельные ПЦР-реакции готовили для каждого из вырожденных праймеров вместе с соответствующим ему праймером константной области в специальном помещении для ПЦР с использованием специфических протоколов, предназначенных для сведения к минимуму возможности перекрестного загрязнения. Во всех случаях для амплификации матричной кДНК использовали ДНК-полимеразу Amplitaq®. Пробирки для реакции ПЦР затем загружали в аппарат для циклического нагревания и осуществляли циклы (после первоначального плавления при 94°С в течение 1,5 мин) при 94°С в течение 1 мин и при 72°С в течение 10 мин в количестве 25 циклов. После завершения последнего цикла осуществляли последнюю стадию удлинения при 72°С в течение 10 мин, после чего реакции охлаждали до 4°С. За исключением перерыва между стадиями отжига (50°С) и удлинения (72°С), когда использовали увеличенное время линейного нарастания, равное 2,5 мин, между всеми стадиями цикла время линейного нарастания было равно 30 с.
Аликвоты объемом по 10 мкл из каждой ПЦР-реакции пропускали через гель, содержавший 1% агарозу/ФБР (рН 8,8), для того, чтобы определить, в какой реакции был образован ПЦР-продукт правильного размера. Те ПРЦ-реакции, в которых, по-видимому, были амплифицированы полномерные гены вариабельной области, повторяли для получения независимых ПЦР-клонов и таким образом минимизировали эффект ПЦР-ошибок. Аликвоты объемом 1-6 мкл этих ПЦР-продуктов правильного размера клонировали непосредственно в pCRII™ вектор, поставляемый в наборе ТА Cloning®, и трансформировали в компетентные клетки INA αF’, как описано в инструкциях изготовителей. Колонии, содержащие плазмиду с вставкой правильного размера, идентифицировали посредством ПЦР-скрининга колоний с использованием прямого (pCRII) и обратного (pCRII) олигонуклеотидных праймеров, описанных в таблице 3, согласно способу  and Clackson, Nucleic Acids Res., 17:4000, 1989.
and Clackson, Nucleic Acids Res., 17:4000, 1989.
Идентифицированные предположительно позитивные клоны были двунитевыми плазмидными ДНК, секвенированными с помощью аппарата для секвенирования ДНК Vistra и предварительно смешанного набора для циклического секвенирования Thermo Sequenase™ в соответствии с инструкциями изготовителя.
Пример 1: Клонирование и секвенирование вариабельной области тяжелой цепи ВАТ антитела
Как при любых планах гуманизации, строго следовали протоколу ПЦР-клонирования и секвенирования. Это было сделано для минимизации возможности внедрения ошибок в последовательности дикого типа генов вариабельной области VH мыши из гибридомной клеточной линии ВАТ. Только если все данные о последовательности ДНК из по меньшей мере двух различных клонов VH генов из гибридомной клеточной линии, экспрессирующей ВАТ антитело мыши, идеально совпадали, последовательности генов признавались правильными.
Три отдельных ПЦР-продукта, каждый из отдельного препарата общей РНК и последующей реакции синтеза первой цепи кДНК, были ПЦР-клонированы, и были полностью секвенированы ДНК обеих нитей. Хотя были проанализированы все двенадцать праймеров тяжелой цепи (таблица 1), только праймер MHV9 (вместе с MHCG3 - предназначенным для гибридизации СН1-домена гена γ3 тяжелой цепи мыши) был ПЦР-амплифицирован с образованием продукта, состоявшего из примерно 460 пар оснований, который затем был ТА-клонирован в клонирующий вектор pCRII™ (данные в таблице 1 не представлены).
Анализ последовательностей ДНК нескольких индивидуальных клонов каждого из трех продуктов ПЦР (каждый из отдельной реакции синтеза 1-й нити и последующей реакции ПЦР) привел к установлению последовательности вариабельной области тяжелой цепи ВАТ антитела, описанной на фиг.1. Эта последовательность была подтверждена на обеих нитях ДНК для всех трех исследованных ПЦР-клонов.
Пример 2: Кпонирование и секвенирование вариабельной области легкой каппа-цепи ВАТ антитела
Однонитевую матрицу кДНК, полученную в процессе синтеза 1-й нити, ПЦР-амплифицировали с использованием серий вырожденных праймеров легкой каппа-цепи (таблица 2). Тем не менее, это привело к амплификации нескольких ПЦР-продуктов с более чем одним вырожденным праймером, что означало, что по меньшей мере более одного гена вариабельной области было транскрибировано ВАТ гибридомной клеточной линией.
Во-первых, был обнаружен продукт ПЦР при совместном использовании МКV2-праймера (который, как и все серии MKV праймеров, гибридизирует 5'-конец последовательности ДНК сигнального пептида легкой каппа-цепи) и МКС праймера (который предназначен для гибридизации 5'-конца константной каппа-области гена мыши). Ранее проведенный авторами настоящего изобретения эксперимент показал, что MKV2 праймер будет амплифицировать в реакции ПЦР аберрантный транскрипт мРНК. Этот аберрантный псевдоген присутствовал во всех стандартных партнерах слияния, полученных из исходной линии клеток плазмацитомы МОРС-21, и был известен как МОРС-21 (Deyev, S.M., et al., Genetica, 85:45, 1991). NO-0 была линией клеток, полученной из линии МОРС-21, и эта линия была использована в качестве партнера слияния для получения ВАТ гибридомы. Вследствие этого не было неожиданным, что при использовании MKV2 праймера был обнаружен ПЦР-продукт. Этот продукт был проанализирован, и было показано, что он является нефункциональным псевдогеном (данные не приведены).
Необычным было то, что другой псевдоген, ранее идентифицированный как секретируемый родственной линией клеток NS-1 (Hamlyn, P.H., et al., Nucl. Acid Res., 9:4485, 1981) и в норме ПЦР-клонируемый при использовании MKV7 праймера вместе с МКС праймером, не был обнаружен ни в одном из ПЦР-продуктов, проанализированных к настоящему моменту. Так как линии клеток NS-1 и NS-0 были близкородственными, это не было очень неожиданным. Тем не менее, это также продемонстрировало сложную природу транскрипции легкой каппа-цепи, которая присутствовала в гибридомной клеточной линии ВАТ.
Другой ПЦР-клон, который, в конечном счете, оказался VК геном ВАТ антитела, также был успешно ПЦР-амплифицирован из ВАТ гибридомной линии клеток с использованием праймеров MKV5 и МКС. После трансформации продукта, содержавшего примерно 450 пар оснований, в INVαF’ компетентные клетки, вероятные позитивные трансформанты были идентифицированы с помощью ПЦР-скрининга, а затем была секвенирована ДНК.
При анализе последовательностей двух отдельных клонов MKV5 продукта (каждый из которых был получен из отдельной реакции синтеза 1-й цепи и последующих ПЦР-реакций) была определена последовательность ДНК гена вариабельной области легкой каппа-цепи ВАТ антитела (фиг.2). Эта последовательность также была подтверждена в обеих нитях ДНК для каждого клона.
Пример 3: Анализ последовательностей вариабельных областей ВАТ антитела мыши
Последовательность аминокислот VK и VH областей ВАТ сравнивали с консенсусными последовательностями подгрупп вариабельной области мышевидных грызунов, которые определены в базе данных Kabat (см. выше).
При этом анализе было обнаружено, что VH область ВАТ наиболее близко соответствует подгруппе VI каппа-цепи мыши. При сходном сравнении VH области ВАТ с базой данных Kabat было обнаружено, что она ближе всего соответствует консенсусной последовательности "смешанной" подгруппы тяжелых цепей мыши.
Сравнение вышеуказанных последовательностей вариабельных областей ВАТ антитела с базой данных по зародышевым линиям клеток мышевидных грызунов позволило обнаружить, что наиболее близким к VH гену ВАТ геном зародышевой линии был VMS/VGK4 (фиг.3), тогда как ближайшим к VK гену ВАТ геном зародышевой линии был Н4 (фиг.4). Как можно видеть на фиг.3, те несоответствия, которые существуют между VH геном ВАТ и ближайшим к нему геном зародышевой линий, были (что не явилось неожиданностью) преимущественно расположены в CDR2 и CDR3. Было лишь три изменения каркаса, и все они были расположены в FR3. Что касается VK гена ВАТ (фиг.4), то также не оказалось неожиданным, что большинство несоответствий были расположены в CDR. Все четыре отличия, которые были расположены в FR, были высоко консервативными изменениями, за исключением цистеина в положении 72 (нумерация по Kabat) в FR3. Его положение непосредственно рядом с важным каноническим остатком (положение 71) позволило предположить, что цистеин может играть ключевую роль в связывании антигена. Тем не менее, только путем моделирования Fv домена можно было бы подтвердить это предположение.
Тем не менее, эти анализы подтвердили, что и VH области и VK области вариабельных областей ВАТ мыши, по-видимому, являются типичными для вариабельных областей мыши.


II. КОНСТРУИРОВАНИЕ И ЭКСПРЕССИЯ ХИМЕРНОГО АНТИТЕЛА СОГЛАСНО НАСТОЯЩЕМУ ИЗОБРЕТЕНИЮ
(А) Аббревиатуры
Были использованы следующие единицы, не относящиеся к системе СИ, и другие аббревиатуры:
Полимеразная цепная реакция (ПЦР); дезоксирибонуклеиновая кислота (ДНК); копированная ДНК (кДНК); вариабельная область легкой каппа-цепи (VK); вариабельная область тяжелой цепи (VH); минута (мин); Трис-боратный буфер (ТББ); фосфатный буферный раствор (ФБР); комнатная температура (КТ); альбумин сыворотки быка (АСБ); соляная кислота (HCl); пероксидаза хрена (ПХ); молоко с низким содержанием жира (МНСЖ); час (час); процент (%); O-фенилендиамина дигидрохлорид (ОФД); сайт множественного клонирования (СМК).
(Б) Материалы
Компоненты сред и все остальные материалы для культур тканей были получены из компании Life Technologies (Великобритания). Компоненты для ПЦР-реакций, включая AmpliTaq® ДНК-полимеразу, были закуплены в компании Perkin Elmer (США). Однако набор ТА Cloning® и INVαF’ компетентные клетки были получены из компании Invitrogen (США). DH5α компетентные клетки и агароза (UltraPure™) были получены из компании Life Technologies (Великобритания).
Предварительно смешанный набор для циклического секвенирования Thermo Sequenase™ и аппарат для секвенирования ДНК Vistra 725 были закуплены в компании Amersham (Великобритания). Готовый к использованию набор реактивов для завершения циклического секвенирования (Big Dye™ Terminator Cycle Sequencing Ready Reaction Kit), используемый совместно с анализатором генов ABI Prism 310 Genetic Analyzer, был закуплен в компании РЕ Applied Biosystems (Великобритания). Все остальные описанные продукты для молекулярной биологии были приобретены либо в компании New England Biolabs (США), либо Promega (США). Иммунологические планшеты Nunc-Immuno Plate MaxiSorp™ были приобретены в компании Life Technologies (Соединенное Королевство), тогда как легко промываемые планшеты Corning для ELISA были приобретены в компании Corning Laboratory Sciences Company (Великобритания). Антитело IgG козы к антигенам человека (специфичное для Fcγ-фрагмента), конъюгат легкой каппа-цепи антитела козы к антигенам человека и пероксидазы хрена и аффинно очищенный конъюгат IgG козы к антигенам человека (специфичного для Fcγ-фрагмента) и пероксидазы хрена были получены из Jackson ImmunoResearch Laboratories Inc. (США). Субстрат K-Blue TMB и раствор Red Stop были закуплены в компании Neogen Inc. (США). Все остальные продукты для ELISA были получены из компании Sigma (Великобритания). Пакет программ Microplate Manager® для анализа данных был закуплен в компании Bio-Rad (Великобритания). Снабженная мешалкой ячейка для ультрафильтрации микрообъемов и мембранный фильтр РМ30 были получены из компании Amicon PLC (Великобритания), тогда как набор для очистки IgG Immunopure® (G) был закуплен в компании Pierce PLC (Великобритания).
(В) Методики эксперимента
В1. Создание химерного γ1/к ВАТ антитела
Предварительно выделенные ген вариабельной области легкой каппа-цепи мыши (VK) (фиг.1) и ген вариабельной области тяжелой цепи (VH) (фиг.2) были модифицированы на 5'- и 3'-концах с помощью специально сконструированных ПЦР-праймеров (таблица 1) для обеспечения экспрессии генов вариабельной области ВАТ в клетках млекопитающих как части химерного антитела мыши и человека. Для достижения этого проводили раздельные ПЦР-реакции для каждого гена вариабельной области в специальном помещении для ПЦР с использованием специфических протоколов, разработанных для минимизации возможности перекрестного загрязнения. В качестве матриц были использованы плазмиды ВАТVH-рСR2.1 и BATVK-pCR2.1, a AmpliTaq® ДНК-полимеразу использовали для амплификации этих матриц. Праймеры В8814 и В8815 (таблица 4) были использованы для ПЦР-модификации VH гена ВАТ, тогда как праймеры С0224 и С0225 (таблица 4) были использованы для ПЦР-мутации VK гена ВАТ.
Пробирки для ПЦР-реакции подвергали циклам (после начального плавления при 94°С в течение 3 мин) при 94°С в течение 50 с, 72°С в течение 1 мин 30 с в количестве 30 циклов. После завершения последнего цикла производили завершающую стадию удлинения при 72°С в течение 10 мин, после чего охлаждали реакционные смеси на льду. Затем аликвоты объемом по 5 мкл от каждой ПЦР-реакции пропускали через гель, содержавший 1,2% агарозы/ТББ (рН 8,8), чтобы определить, в какой реакции был получен ПЦР-продукт правильного размера.
Аликвоты объемом 1-2 мкл этих ПЦР-продуктов правильного размера непосредственно клонировали в вектор pCR2.1™, имеющийся в наборе ТА Cloning®, и трансформировали в компетентные клетки INVαF’, как описано в инструкциях изготовителя. Колонии, содержавшие плазмиду со вставкой правильного размера, идентифицировали посредством ПЦР-скрининга колоний с использованием олигонуклеотидных праймеров 1212 и 1233 (таблица 5) по способу Gussow and Clackson (см. выше). У идентифицированных вероятных позитивных клонов была секвенирована двунитевая плазмидная ДНК с помощью аппарата для секвенирования ДНК Vistra и анализатора генов ABI Prism 310. Предварительно смешанный набор для циклического секвенирования Thermo Sequenase™ и готовый к употреблению набор реактивов для окончания циклического секвенирования Big Dye™ использовали согласно инструкциям изготовителя с праймерами 1212 и 1233 (таблица 5).
Клоны, содержавшие корректно адаптированные VK и VH гены ВАТ (фиг.5 и 6, соответственно) субклонировали, в виде фрагментов HindIII-BamHi, в экспрессирующие векторы рКN110 (фиг.7) и pG1D110 (фиг.8), соответственно, для экспрессии химерных легких и тяжелых цепей в клетках млекопитающих. Лигированные экспрессирующие векторы (то есть рКN110-ВАТVK и pG1D110-ВАТVH) затем трансформировали в компетентные клетки DH5α. В заключение идентифицировали позитивные клоны, содержащие правильно сконструированные экспрессирующие векторы, с помощью анализа с рестрицирующими ферментами.
В2. Совместная трансфекция векторных ДНК, кодирующих химерные γ1/к ВАТ антитела, в COS клетки
Для трансфицирования экспрессирующих векторов млекопитающих в COS клетки следовали способу Kettleborough et al. Вкратце, ДНК (по 10 мкг вектора, экспрессирующего легкую каппа-цепь рG1D110-ВАТVK, и вектора, экспрессирующего тяжелую цепь PG1D110-ВАТVH) добавляли к аликвоте объемом 0,70 мл, содержавшей 1×107 клеток/мл ФБР, и подавали пульсирующее напряжение, равное 1900 В, емкость 25 мкФ, с помощью аппарата Bio-Rad Gene Pulser. После 10-минутного восстановления при КT электропорированные клетки добавляли к 8 мл DMEM, содержавшим 5% ФСТ, и инкубировали в течение 72 ч при 5%-ном содержании CO2 при 37°С. После инкубации в течение 72 ч среду собирали, центрифугировали для удаления остатков клеток и анализировали с помощью ELISA на продукцию ВАТ антитела.
В3. Количественная оценка химерного γ1/к антитела с помощью ELISA
В каждую ячейку 96-ячеечного иммунологического планшета Nunc-Immuno Plate MaxiSorp™ вначале наносили аликвоты объемом 100 мкл, содержавшие 0,4 нг/мкл IgG (со специфичным Fcγ-фрагментом) козы к антигенам человека, разводили ФБР, инкубировали в течение ночи при 4°С и удаляли перед использованием. Затем аликвоты экспериментальных проб (то есть собранных супернатантов COS клеток - центрифугированных для удаления остатков клеток) объемом по 100 мкл из каждой ячейки и пробы, разведенные в соотношении 1:2 буферным раствором для разведения проб и конъюгированных ферментов (0,1 М Трис-НСl (рН 7,0), 0,1 М NaCl, 0,02% (объем/объем) ТВИН-20 и 0,2% (вес/объем) САБ), распределяли по иммунологическому планшету. Кроме того, очищенное γ1/к антитело человека (1000 нг/мкл), которое использовали в качестве стандарта, серийно разводили в соотношении 1:2 и также наносили на иммунологический планшет. Иммунологический планшет инкубировали при 37°С в течение 1 ч, после чего три раза промывали промывочным буфером (ФБР/0,1% (объем/объем) ТВИН-20) в объеме 200 мкл/ячейку. 100 мкл конъюгата легкой каппа-цепи антитела козы к антигенам человека и пероксидазы хрена, разведенного в 5000 раз в буфере для разведения конъюгатов проб и ферментов, добавляли к каждой ячейке, затем иммунологический планшет инкубировали при 37°С в течение 1 ч, после чего его снова промывали так, как описано выше. После этого к каждой ячейке добавляли аликвоты объемом 150 мкл субстрата K-Blue, после чего иммунологический планшет инкубировали в течение 10 мин в темноте при комнатной температуре. Реакцию в конечном итоге останавливали распределением по 50 мкл Red Stop в каждую ячейку. Затем определяли оптическую плотность при длине волны 655 нм с помощью устройства для чтения микропланшетов Bio-Rad 3550 в комплексе с пакетом программного обеспечения Microplate Manager®.
В4. Очистка химерного ВАТ антитела
Химерное γ1/к ВАТ антитело выделяли из супернатантов COS клеток в два этапа. Вначале использовали микроячейку для ультрафильтрации с мешалкой и мембранным фильтром РМ30 согласно инструкциям изготовителя для уменьшения объема сырого, неочищенного супернатанта. Затем использовали набор для очистки IgG Immunopure® для аффинного выделения химерного ВАТ антитела из концентрированного супернатанта, также согласно инструкциям изготовителя.
В5. Твердофазный иммуноферментный анализ (ELISA) клеток Дауди
Твердофазный иммуноферментный анализ (ELISA) клеток был выполнен с использованием клеток Дауди, культивированных из исходного материала д-ром Hardy (Felsenstein Medical Research Center, Rabin Medical Center, Beilinson Campus, Petach Tikva, 49100, Израиль). Были произведены небольшие модификации анализа, в зависимости от того, анализировали ли ВАТ антитела мыши или химерные антитела мыши-человека. При анализе сродства ВАТ антитела мыши в качестве второго антитела использовали конъюгат IgG козы к антигенам мыши (Fab-специфичный) с пероксидазой хрена (разведенный в соотношении 1:15000). Напротив, при измерении сродства химерного ВАТ антитела использовали конъюгат аффинно очищенного IgG козы к антигенам человека (специфичного по Fcγ-фрагменту) с пероксидазой хрена (разведенный в 1000 раз).
Клетки Дауди (через два дня после пассажа) вначале наносили в количестве 105 клеток/ячейку на 96-ячеечный планшет Corning easy wash ELISA plate (легко промываемый планшет для ELISA производства компании Corning), a затем инкубировали в течение ночи при 37°С в сухом инкубаторе. На следующий день к каждой ячейке добавляли 200 мкл регидратирующего буфера (ФБР, содержащий 10% ФСТ и 0,05% азида), а затем оставляли по меньшей мере на 1 ч. Затем регидратирующий буфер декантировали, после чего к ячейкам на планшете добавляли аликвоты объемом по 50 мкл различных серийных разведений в соотношении 1:2 очищенного ВАТ антитела. Планшет снова инкубировали в течение ночи (при 4°С), два раза промывали 200 мкл/ячейку ФБР, содержащего 5% молока с низким содержанием жира (МНСЖ), и давали высохнуть. Затем добавляли 50 мкл/ячейку конъюгированного с пероксидазой хрена вторичного антитела, после чего производили серию из шести различных промывок (а именно - одной промывки ФБР, содержащим 5% МНСЖ, трех промывок тем же буфером с добавлением 0,05% ТВИН-20 и еще двумя последующими промывками ФБР/МНСЖ буфером). Затем добавляли 200 мкл/ячейку раствора субстрата ОФД в концентрации 0,4 мг/мл в 0,05 М цитратном буфере (рН 5,0) и 60 мг/мл перекиси водорода, после чего планшет для ELISA инкубировали в темноте и при комнатной температуре до развития окраски (обычно в течение примерно 30 мин). В заключение, реакцию останавливали добавлением 50 мкл/ячейку 2,5 М серной кислоты, а затем измеряли оптическую плотность при длине волны 490 нм с помощью устройства для чтения микропланшетов Bio-Rad 3550 в комплексе с пакетом программного обеспечения Microplate Manager®.
Результаты
Пример 4. Конструирование химерного γ1/к ВАТ антитела
Как и во всех программах, следовали строгому протоколу ПЦР-клонирования и секвенирования. Это делали для минимизации возможности внедрения ошибок в последовательности дикого типа генов вариабельной области мыши во время стадии ПЦР-модификации. С использованием праймеров С0224 и С0225 (таблица 1) ген ВАТVK мыши (фиг.2) был модифицирован посредством ПЦР с получением последовательности из 418 пар оснований (данные не приведены). Этот продукт ПЦР был лигирован к плазмиде pCR2.1 и трансформирован в INVαF’ компетентные клетки. Сходным образом ген VH BAT мыши (фиг.1) был мутирован посредством ПЦР с использованием праймеров В8814 и В8815 (таблица 1) с получением последовательности из 436 пар оснований (данные не приведены). Этот продукт ПЦР также был лигирован в плазмиду pCR2.1 и трансформирован в INVaF' компетентные клетки.
Затем вероятные позитивные трансформанты были выявлены с помощью ПЦР-скрининга (данные не приведены) с последующим окончательным секвенированием ДНК на ABI Prism 310 Genetic Analyzer. фиг.3 и 4 демонстрируют результаты этого анализа последовательностей ДНК химерных генов ВАТVK и ВАТVH, соответственно. Анализ был проведен как с целью подтверждения успешности мутагенеза, так и для того, чтобы продемонстрировать наличие каких-либо ошибок ПЦР, которые могли быть внедрены в гены. Только одну ПЦР-реакцию фактически проводили для каждого гена вариабельной области, и только два клона из каждой из этих ПЦР-реакций, в конечном итоге, были завершенными последовательностями ДНК.
Тем не менее, этого оказалось достаточно для выделения, по меньшей мере, одного клона для каждого модифицированного гена вариабельной области, который содержал корректно модифицированную последовательность ДНК.
Мутированные VH и VK гены затем субклонировали в соответствующие экспрессирующие векторы, в виде hindIII/BamHI фрагментов, с образованием pKN110-BATVK (7,88 тысяч оснований) и рG1D110-ВАТVK (7,55 тысяч оснований), соответственно. Правильность сконструированных экспрессирующих векторов была затем подтверждена с помощью рестрикционного ферментативного анализа (данные не приведены). Будучи трансфицированными в COS клетки, эти векторы обеспечивали временную экспрессию γ1/к варианта химерного ВАТ антитела.
Кроме того, в качестве дополнительного компонента программы гуманизации ВАТ антитела, был также субклонирован ВАТVH ген, в виде HindIII/BamH1 фрагмента, в оба вектора экспрессии тяжелой цепи pG3D110 и pG4D1100. Эти векторы были идентичны pG1D110, за исключением замещения кДНК копии генов константной области γ1 человека, либо кДНК копией генов константной области 3γ (в случае pG3D110), либо кДНК генов константной области γ3 (в случае pG3D110), либо кДНК генов константной области γ4 (в случае pG3D110). Конструкция этих векторов (то есть, PG3D110-BATVK, обоих γ3/к и γ4/к вариантов химерного ВАТ антитела в COS клетки.
Пример 5. Временная экспрессия химерного γ1/к ВАТ антитела
Два вектора рКN110-ВАТVK и pG1D110-BATVH совместно трансфицировали в COS клетки в серии повторных экспериментов с временной экспрессией. После экспрессии в течение 72 ч химерное ВАТ γ1/к антитело мыши-человека было обнаружено в супернатантах совместно трансфицированных COS клеток посредством анализа ELISA на γ1/к. Исходя из результатов этих анализов, было рассчитано, что средняя концентрация химерного ВАТ γ1/к антитела, обнаруженного в среде, была равна 509±272 нг/мл.
Интересно, что γ3/к и γ4/к варианты химерного ВАТ антитела, по-видимому, продуцировали достоверно большие количества антитела после их экспрессии COS клетками. А именно, если в COS клетки были совместно трансфицированы рС3D110-ВАТVн и pKN110-BATVК, при начальном анализе супернатанта (с использованием способа ELISA, описанного в Разделе В3, и антитела lg3/каппа человека в качестве стандарта) были измерены уровни экспрессии химерного γ3/к ВАТ антитела, равные 6,7 мкг/мл. Более того, если в COS клетках экспрессировались рG4D110-ВАТVH и рКN110-ВАТVK, в тех же анализах ELISA (с использованием антитела lg4/каппа человека в качестве стандарта) были измерены уровни экспрессии химерного γ4/к ВАТ антитела, равные 8,2 мкг/мл.
Пример 6. Очистка химерного γ1/к ВАТ антитела
Накапливая примерно по 8 мл от каждой совместной трансфекции, выполняли серию трансфекции до тех пор, пока не было собрано 200 мл супернатанта COS клеток. Объем этого супернатанта затем уменьшали до 15 мл путем пропускания супернатанта через микроячейку для ультрафильтрации с мешалкой и мембранным фильтром РМ30, имеющим молекулярный вес отсечки, равный 30 кДа.
Набор для выделения IgG Immunopure® (G) фактически включал колонку на 2 мл с иммобилизованным Белком G. Антитело элюировали из колонки 6 мл элюирующего буфера, элюат собирали фракциями по 1 мл. Концентрацию химерного γ1/к ВАТ антитела в каждой фракции затем анализировали с помощью способа ELISA, описанного в Разделе В3. При этом анализе было обнаружено, что химерное антитело присутствовало во Фракции 3 (42,05 мкг/мл) и Фракции 4 (20,05 мкг/мл), что соответствовало общему обратному выделению 62,1 мкг химерного γ1/к ВАТ антитела. Его хранили при -20°С до последующего переноса на Curetech для дальнейшего анализа.
Пример 7. Анализ связывания клеток Дауди с химерным γ1/к ВАТ антителом
С использованием ELISA анализа клеток Дауди было четко показано, что очищенное химерное γ1/к ВАТ антитело связывается с клетками Дауди. Фиг.9 демонстрирует типичный пример эксперимента. Однако менее убедительным было связывание сходных концентраций ВАТ антитела мыши, в тех же анализах ELISA, которое, по-видимому, было ниже, чем связывание химерного антитела. Тем не менее, поскольку конъюгированное вторичное антитело, использованное для выявления связывания антител с клетками Дауди, отличалось от всех рекомбинантных антител, невозможно было обоснованно произвести прямое сравнение связывания двух вариантов.


IV. АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ 3 ПЕПТИДОВ.
ПОЛУЧЕННЫХ ИЗ АНТИГЕНА ЛИНИИ ЛИМФОБЛАСТОИДНЫХ В-КЛЕТОК ДАУДИ, С КОТОРЫМ СВЯЗЫВАЮТСЯ MAT СОГЛАСНО НАСТОЯЩЕМУ ИЗОБРЕТЕНИЮ
Три пептида, содержащиеся в антигенном эпитопе лимфобластоидных В-клеток Дауди, с которыми связываются MAT согласно настоящему изобретению, были секвенированы. Их последовательности приведены на фиг.10, 11 и 12.
Поиск, проведенный в базе данных банка неизбыточных генов и Отделе EST, не выявил совпадений при пропускании этих трех пептидов в виде запроса с использованием алгоритма TBLASTN (Версия 2) с уровнем ОЖИДАНИЯ, равным 10, и матрицы BLOSUM 62.
Тем не менее, поскольку эти пептиды были мелкими пептидами, их предъявили еще раз при более высоком уровне ОЖИДАНИЯ, чтобы сделать поиск менее строгим. Кроме того, фильтр не был маскирован от низкой сложности, что могло бы исключить потенциальные смешанные совмещения (например, совпадения областей, богатых пролином, или проли-А концов) из отчетов, оставив только области, статистика которых отражает специфичность их попарного совмещения. Для трех пептидов согласно настоящему изобретению не было обнаружено совпадений при работе с базами данных банка генов и отдела EST даже при очень низкой строгости.
Поэтому, в соответствии с вышеприведенными результатами, три вышеуказанных пептида представляются новыми пептидами.
V. ДИАГНОСТИКА ЗЛОКАЧЕСТВЕННЫХ ЗАБОЛЕВАНИЙ У БОЛЬНЫХ С ПОМОЩЬЮ MAT СОГЛАСНО НАСТОЯЩЕМУ ИЗОБРЕТЕНИЮ
Лимфоциты периферической крови, полученные от обследованных людей, были дважды мечеными с помощью антитела к CD3 и одного из MAT согласно настоящему изобретению. Определяли процент СD3+ клеток, которые связывали MAT согласно настоящему изобретению. Было показано, что число CD3+ MAT+ клеток у людей, имевших злокачественные заболевания, отличается от процента этих клеток в пробах крови, полученных от здоровых людей. Тот факт, что существует достоверное отличие процентного содержания CD3+ клеток у людей, имеющих злокачественное заболевание, и что это процентное содержание выше или ниже процентного содержания CD3+ МАТ+ клеток в пробах, полученных от здоровых людей, позволяет с высокой вероятностью определить, есть ли у человека злокачественное заболевание, а также специфический вид злокачественного заболевания, которое может быть у обследованного человека.
Обычно лимфоциты периферической крови человека получали из 20 мл крови, взятой либо у здорового человека, либо у больного раком, посредством центрифугирования в градиенте плотности Ficoll Hypaque. Клетки промывали и суспендировали в ФБР, содержавшем 0,5% САБ и 0,05% кислоты. Пробы, содержавшие 0,5×106 клеток, использовали для анализа FACS. Вначале клетки инкубировали с насыщающим количеством MAT согласно настоящему изобретению в течение 45 мин при 0°С, с последующей инкубацией их с MAT к антигенам мыши, конъюгированным с FITZ, в течение 30 мин на льду. После двух промывок и центрифугирования при 1200 об/мин клетки инкубировали с антителами к CD3 человека, конъюгированными с РЕ, в течение 30 мин, на льду. После этой инкубации клетки два раза промывали, и пробу анализировали с помощью FACS сканирования (Bectan Dickinson). Результаты приведены на фиг.13-17.
Как можно видеть на фиг.13, а также на фиг.17, процент CD3+ ВАТ+ клеток (по сравнению с общим числом CD3+ клеток) в пробах крови, полученных от здоровых людей, лежит в диапазоне около 25%. Как показано на фиг.14, процент CD3+ ВАТ+ клеток в пробах крови, полученных от больных раком толстого кишечника, значительно ниже, по сравнению со здоровыми людьми, и лежит в диапазоне около 7%. Сходным образом, процент CD3+ ВАТ+ клеток в пробах крови, полученных от больных раком молочной железы, лежал в диапазоне около 10% (фиг.15). Эти результаты отчетливо показывают, что карциномы толстого кишечника и молочной железы можно идентифицировать по тому факту, что процент CD3+ ВАТ+ клеток ниже, по сравнению со здоровыми людьми.
Процент CD3+ ВАТ+ клеток в пробах крови, полученных от больных раком предстательной железы, достоверно выше процентного содержания этих клеток в пробах крови от здоровых людей, как показано на фиг.16, и лежит в диапазоне около 50%. Эти результаты четко показывают, что карциному предстательной железы можно идентифицировать по тому факту, что процент CD3+ ВАТ+ клеток выше по сравнению со здоровыми людьми. Как показано на фиг.18, количество антигена, к которому присоединяются MAT согласно настоящему изобретению, обнаруженного на Т-клетках и выделенного у больных карциномой предстательной железы, очень велико, тогда как в Т-клетках, полученных от больных карциномой молочной железы, антиген невозможно обнаружить.
Вышеприведенные результаты показывают, что MAT согласно настоящему изобретению можно использовать для того, чтобы определить, что отдельный человек страдает определенным родом злокачественного заболевания. Поэтому, если пробу крови получают от обследуемого человека, и степень связывания MAT согласно настоящему изобретению достоверно высока (в диапазоне около 50%), существует очень высокая вероятность того, что обследуемый человек страдает раком предстательной железы. Напротив, если процент CD3+ клеток в пробе достоверно ниже, по сравнению со здоровыми людьми (в диапазоне между примерно 7 и 10%), существует очень высокая вероятность того, что обследуемый человек страдает карциномой молочной железы или толстого кишечника. Очевидно, что, если обследованный человек является мужчиной, существует очень высокая вероятность, что он страдает карциномой толстого кишечника.
Вышеприведенные примеры не являются ограничивающими, и дополнительные корреляции между процентами CD3+ клеток, которые связывают MAT согласно настоящему изобретению, и другие злокачественные заболевания также находятся в пределах объема изобретения.





| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ ОХ40 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2562874C1 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К CD22 ЧЕЛОВЕКА, И ИХ ПРИМЕНЕНИЯ В ТЕРАПИИ И ДИАГНОСТИКЕ | 2003 |
|
RU2342401C2 |
| АНТИТЕЛА | 2005 |
|
RU2482131C2 |
| АНТИТЕЛО ПРОТИВ PD-L1 | 2017 |
|
RU2744862C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2694412C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К КЛАУДИНУ 6 (CLDN6) | 2010 |
|
RU2675997C2 |
| ЛЕЧЕНИЕ РАКА ПРИ ПОМОЩИ НАПРАВЛЕННЫХ НА МИШЕНЬ АНТИТЕЛ IN VIVO | 2011 |
|
RU2642305C2 |
| АНТИТЕЛО ПРОТИВ ILT17 | 2012 |
|
RU2599450C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К СD52 | 2012 |
|
RU2605307C2 |
| ГУМАНИЗИРОВАННОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СПЕЦИФИЧНОЕ К СИНДЕКАНУ-1 | 2013 |
|
RU2551235C2 |







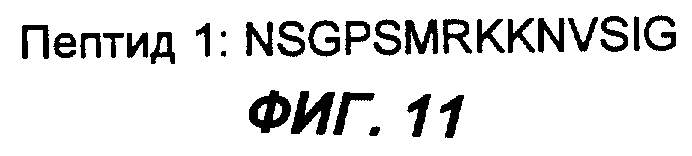




Изобретение относится к области биотехнологии и медицины, а именно, к новым последовательностям нуклеотидов ДНК и последовательностям аминокислот моноклональных антител (MAT), вырабатываемых против лимфобластоидных клеток, и к пептидам, с которыми связываются МАТ. Изобретение также относится к диагностическим анализам, в которых вышеуказанные антитела или пептиды используются для выявления индивидуумов с высокой вероятностью наличия злокачественных заболеваний и, иногда, для выявления индивидуума, имеющего специфическое злокачественное заболевание. Кроме того, изобретение относится к фармацевтическим композициям, включающим MAT или пептиды согласно настоящему изобретению и предназначенным для применения при лечении различных злокачественных заболеваний, а также к способам лечения злокачественных заболеваний с использованием MAT или пептидов согласно настоящему изобретению. Использование изобретения позволит проводить дифференциальную диагностику рака. 7 н. и 23 з.п. ф-лы, 18 ил., 5 табл.

| WO 9721104 А, 12.06.1997 | |||
| US 4708930 A, 24.11.1987. |
