Область техники
Данное изобретение относится к молекулам цитокина человека - Фактора Некроза Опухолей α (ФНОα), которые были модифицированы так, что они оказались способны повышать содержание нейтрализующих антител к человеческому ФНОα дикого типа после введения модифицированной молекулы ФНОα человеку-реципиенту. Изобретение также относится к вакцинам против ФНОα человека на основе вышеуказанных модифицированных молекул ФНОα. Прочие аспекты изобретения станут ясны из нижеследующего описания.
Уровень техники
Физиологически иммунная система позвоночных служит в качестве защитного механизма против инвазии организма инфекционными объектами, такими как микроорганизмы. Чужеродные белки эффективно удаляются при посредстве ретикулоэндотелиальной системы с помощью высокоспецифичных циркулирующих антител, а вирусы и бактерии разрушаются при помощи сложной группы клеточных и гуморальных механизмов, включающих антитела, цитотоксические Т-лимфоциты (ЦТЛ), клетки - натуральные киллеры (НК), комплемент и т.п. Лидером в этой борьбе является лимфоцит Т-хелпер (Тх), который вместе с Антиген-Представляющими Клетками (АПК) регулирует иммунную защиту через сложную систему цитокинов.
Тх-лимфоциты распознают белковые антигены, представленные на поверхности АПК. Однако они не распознают нативные антигены как таковые. Вместо этого они, по-видимому, распознают комплексный лиганд, состоящий из двух компонентов, “обработанного” (фрагментированного) белкового антигена (так называемого эпитопа для Т-клетки) и молекулы класса II Основного Комплекса Гистосовместимости (ОКГ) (О.Werdelin et al., Imm. Rev. 106, 181 (1988)). Это распознавание, в конечном счете, позволяет Тх-лимфоцитам специфически помогать В-лимфоцитам продуцировать специфические антитела к интактному белковому антигену (Werdelin et al., см. выше). Определенная Т-клетка распознает только определенную комбинацию антигена-ОКГ и не будет распознавать тот же или другой антиген, представленный продуктом другой аллели гена ОКГ. Этот феномен называется ограниченностью ОКГ.
АПК также представляют фрагменты собственных белков, но в норме такие фрагменты игнорируются или не распознаются лимфоцитами-Т-хелперами. Это основная причина, по которой у индивидуумов в сыворотке обычно не накапливаются аутоантитела, в конечном итоге, приводящие к повреждению собственных белков индивидуума (так называемые собственные или аутобелки). Однако в редких случаях процесс идет неправильно, и иммунная система обращается против собственных компонентов индивидуума, что может приводить к аутоиммунному заболеванию.
Присутствие некоторых собственных белков невыгодно в ситуациях, в которых они при повышенных концентрациях вызывают симптомы заболевания. Так, известно, что фактор некроза опухолей α (ФНОα) может вызывать кахексию у больных раком и больных, страдающих другими хроническими заболеваниями (H.N.Langstein et al. Cancer Res. 51, 2302-2306, 1991). ФНОα также играет важную роль в воспалительном процессе (W.P. Arend et al. Arthritis Rheum. 33, 305-315, 1990), и поэтому было продемонстрировано, что нейтрализация ФНОα путем использования моноклональных антител полезна для больных с хроническими воспалительными заболеваниями, такими как ревматоидный артрит (Elliott et al., Lancet 194, 344:1105-10) и болезнь Крона (van Dullemen et al., Gastroenterology 109 (1): 129-135 (1995). Поэтому существует потребность в способе индукции образования нейтрализующих антител к таким белкам ФНОα, и данное изобретение включает вакцину против ФНОα, которая обладает этим свойством.
ФАКТОР НЕКРОЗА ОПУХОЛЕЙ
1. Основные исходные данные
Фактор некроза опухолей (ФНО) является представителем регуляторных белков семейства цитокинов (см. Walsh, G. and Headon, D.E., Protein Biotechnology, 1994, John Wiley & Sons Ltd., England, p.257-267), которое также включает интерфероны и интерлейкины. В настоящее время известны две формы ФН, ФНОα и ФНОβ соответственно. Хотя оба белка связываются с одними и теми же рецепторами и вызывают очень сходные биологические реакции, они являются различными молекулами и гомологичны менее чем на 30%. Исходный белок, названный фактором некроза опухолей, на который ссылаются как на ФНО, более правильно называть ФНОα; он также известен как кахектин. На ФНОβ также ссылаются как на лимфотоксин.
В дополнение к клеткам мозга и печени, ФНОα продуцируется многими типами клеток, в особенности активированными макрофагами, моноцитами, некоторыми Т-лимфоцитами и НК-клетками. Наиболее сильным из известных индукторов синтеза ФНОα является сложная биомолекула, называемая липополисахаридом (ЛПС). Она содержит как липидный, так и полисахаридный компоненты и также упоминается в качестве эндотоксина. Сам по себе липополисахарид лишен противоопухолевой активности. Было обнаружено, что сыворотка животных, которым вводили липополисахарид содержит фактор, токсичный для раковых клеток, и этот фактор, продуцируемый специфическими клетками в ответ на липополисахарид, был назван фактором некроза опухолей. Многие другие агенты, такие как некоторые вирусы, грибки и паразиты, также стимулируют синтез и выделение этого цитокина. Более того, ФНОα может действовать аутокринным образом, стимулируя свою собственную продукцию.
Нативный ФНОα человека является гомотримером, состоящим из трех идентичных полипептидных субъединиц, плотно соединенных вокруг общей оси симметрии, как будет дополнительно разъяснено ниже. Эта конструкция напоминает сборку белковых субъединиц во многих белках вирусных капсидов. Отдельные полипептидные субъединицы ФНОα человека не гликозилированы и состоят из 157 аминокислот. Молекула имеет молекулярную массу 17300 Да и содержит одну внутрицепочечную дисульфидную связь. ФНОα человека синтезируется вначале из молекулы-предшественника, состоящей из 233 аминокислот. Протеолитическое отщепление предпоследовательности от -76 до -1, включающей сигнальную последовательность, освобождает нативный ФНОα. ФНОα может также существовать в мембраносвязанной форме с молекулярной массой 26000 Да. Три мономерных субъединицы ФНОα соединяются нековалентно с образованием тримера, как объяснено ниже.
ФНОα вызывает свои биологические эффекты при связывании со специфическими рецепторами, присутствующими на поверхности восприимчивых клеток. Идентифицированы два различных типа рецепторов для ФНОα. Один рецептор (ФНО-R55) имеет молекулярную массу 55000 Да, тогда как второй рецептор (ФНО-R75) имеет молекулярную массу примерно 75000 Да. Эти два различных типа рецепторов обнаруживают гомологию последовательностей, не превышающую 25%. ФНО-R55 присутствует в широком диапазоне клеток, тогда как распространенность ФНО-R75 рецепторов более ограниченная. Оба рецептора являются трансмембранными гликопротеинами с экстраклеточным связывающим доменом, гидрофобным трансмембранным доменом и внутриклеточным эффекторным доменом.
Точные молекулярные механизмы, при помощи которых ФНОα индуцирует свои биологические эффекты, еще предстоит определить. Связывание ФНОα с его рецепторами, по-видимому, запускает, помимо активации аденилатциклазы, фосфолипазы А и протеинкиназ, разнообразные явления, опосредуемые G-белками. Точные биологические эффекты, вызываемые ФНОα, могут варьировать у разных типов клеток. Другие факторы, такие как присутствие других цитокинов, дополнительно модулируют наблюдаемые молекулярные эффекты, объясняемые действием ФНОα на чувствительные клетки.
Ген ФНОα был клонирован и внедрен в различные рекомбинантные системы экспрессии, как бактериальные, так и эукариотические. Доступность в результате этого больших количеств очищенного, биологически активного ФНОα облегчила клиническое исследование многих заболеваний, особенно рака. Однако многие из этих исследований с использованием ФНОα, отдельно или в комбинации с интерферонами, дали весьма противоречивые результаты. Большие количества ФНОα нельзя вводить больным из-за их токсических - если не летальных - побочных эффектов.
Как указано выше, продолжительная продукция неблагоприятно повышенных концентраций ФНОα также участвует в развитии кахексии, синдрома истощения, часто связанного с хроническими паразитарными или прочими инфекциями и с раком. ФНОα также участвует в метастазировании и росте некоторых опухолей, а также в индукции анемии. Более того, ФНОα непосредственно участвует в развитии некоторых хронических воспалительных заболеваний у людей, включая ревматоидный артрит и болезнь Крона, при которых была показана полезность введения моноклональных антител к ФНОα. ФНОα также участвует в развитии остеопороза и псориаза. Кроме того, в модельных опытах на животных было показано, что введение антител к ФНОα может снижать или предотвращать отторжение пересаженных или трансплантированных тканей.
Структура ФНОα
1. Введение
Пространственная структура фактора некроза опухолей человека (ФНОα) была установлена (см. монографию "Tumor Necrosis Factors, Structure, Function and Mechanism of Action", изданную Bharat B. Aggarwal и Jan Vilcek, 1992 Marcel Dekker, Ind., New York, Глава 5 “Кристаллическая структура ФНОα", авторы Jones, E.Y., Stuart, D.I. и Walker N.P.C.). Биологическое действие ФНОα зависит от его взаимодействия с его рецепторами. Эти взаимодействия регулируются точной конструкцией корректно сложенной третичной структуры. Таким образом, чтобы понять, как молекула ФНОα выполняет свою биологическую функцию на уровне взаимодействий аминокислот, необходимо не только знать последовательность аминокислот, но и пространственную структуру.
II. Пространственная структура
При помощи аналитического ультрацентрифугирования, рассеяния рентгеновского излучения под малым углом и гель-электрофореза было показано, что биологически активный ФНОα находится в растворе в тримерной конформации, а исследования с перекрестным связыванием показали, что это активная форма белка (Smith and Baglioni, 1987, J.Biol. Chemistry 252, 6951-6954). Опыты со связыванием показали, что тример минимум в 8 раз более активен, чем мономер. Экспериментальные доказательства свидетельствуют, что и натуральный, и рекомбинантный ФНОα при физиологических условиях существуют преимущественно в форме тримера. Анализ спектров кругового дихроизма поместил ФНОα в класс белков со слоистой структурой (см. статью Davis et al., Biochemistry 1987, 26, 1322-1326, где описаны результаты структурного анализа очищенного рекомбинантного ФНОα при помощи титрования сульфгидридных групп, гельфильтрации и кругового 20 дихроизма). Несколько различных кристаллических форм было описано для рекомбинантного ФНОα человека. Все описанные кристаллические формы обнаруживают кристаллографическую и некристаллографическую тройную симметрию, являющуюся показателем присутствия ФНОα в кристалле в виде тримера. Тримеры ФНОα лежат в свободно упакованном порядке, перфорированном каналами, заполненными растворителем, диаметром 10 нм. Только небольшая доля поверхности молекулы участвует в контактах кристаллической упаковки. Такие контакты могут слегка изменять положение некоторых боковых цепей и, возможно, даже коротких участков, по существу гибкой, основной цепи по сравнению с предпочитаемыми ими конформациями в растворе.
А. Укладка основной цепи мономера ФНОα
Общая форма отдельной субъединицы тримера ФНОα, состоящей из 157 аминокислот, напоминает клин с высотой примерно 5,5 нм и максимальной шириной в 3,5 нм вблизи основания. Топология основной цепи иллюстрируется фиг.1а-с; она по существу является структурой типа β-сандвича, образованной двумя антипараллельными β-складчатыми слоями. Укладка основной цепи соответствует классическому мотиву “рулета с начинкой” (фиг.1с) (обычному для белка вирусного капсида). Номенклатура, принятая на фиг.1 для маркировки блоков вторичной структуры, соответствует установленному порядку для вирусных структур. Присутствуют все восемь стандартных β-цепей (от В до I), но со вставкой между В и С, которая добавляет короткую цепь на краю обоих β-слоев и обрезает N-терминальную половину С, так что каждый β-складчатый слой содержит пять антипараллельных β-цепей, задний β-слой включает β-цепи В′, В, I, D и G и передний слой включает β-цепи С′, С, Н, Е и F.
N-конец очень гибок. Эта область, вплоть до остатка 10 (см. фиг.1b), является довольно независимой от остальной молекулы, и несколько первых остатков свободно принимают различные конформации в растворителе. В противоположность этому С-конец погружен в основание заднего β-слоя и образует неотъемлемую часть этого относительно плоского блока вторичной структуры. Чередование β-цепей разной длины и вставка между β-цепями В и С совместно образуют переднюю поверхность, образованную почти полностью из петель, и это та “замаскированная” сторона β-сандвича, которая в тримере обращена к растворителю. Кристаллографические данные позволили измерить относительную гибкость различных частей структуры, β-цепи образуют довольно негибкую конструкцию; в частности, задний β-слой расположен в ядре тримера и вследствие этого особенно жесток. Как и следовало ожидать, петли, которые украшают наружную, доступную для растворителя поверхность молекулы, обнаруживают высокие уровни гибкости/подвижности. В целом, происходит общее снижение ригидности, по мере того как ядро становится более свободно упакованным в верхней половине молекулы.
Б. Общая топология тримера ФНОα
Три мономерных субъединицы ФНОα соединяются нековалентно с образованием компактного, конической формы тримера, имеющего длину примерно 5,5 нм и максимальную ширину 5 нм. β-цепи трех отдельных β-сандвичей лежат приблизительно параллельно (наклон равен примерно 30°) общей оси тримера. Взаимодействие между субъединицами, относящееся к общей оси, осуществляется путем простой упаковки β-сандвича типа “край-к-грани”; край β-сандвича, состоящий из цепей F и G одной субъединицы, лежит поперек заднего β-слоя [GDIBB′] одной из трех родственных субъединиц (см. фиг.2). Карбоксильные концы лежат близко к общей оси. Способ упаковки “край-к-грани” обеспечивает исключительно плотное соединение субъединиц. Поэтому ядро тримера абсолютно недоступно для растворителя.
С. Распределение различных типов аминокислот
Общее распределение типов остатков в пространственной структуре ФНОα подчиняется общему правилу для белков: а именно гидрофобные остатки группируются в ядре молекулы, тогда как заряженные остатки декорируют поверхность. Поэтому ядро сандвича ФНОα, как и ожидалось, заполнено плотно упакованными крупными неполярными остатками.
Энергетика системы не благоприятствует существованию ФНОα в виде мономера. Для большой поверхности раздела, состоящей из комплементарных остатков (т.е. полярных остатков, образующих пару с полярными остатками), отсутствие поверхности, доступной для растворителя, обеспечивает значительную энергетическую выгодность образования олигомера (например, тримера). Обнажение для растворителя большой зоны высокогидрофобных остатков, в норме погруженных в нижний участок поверхности раздела тримера, также действовало бы дестабилизирующе на мономер ФНОα.
2. Иcследования структуры функции
А. Взаимодействия ФНОα/антитела
Наблюдалось, что антитела, накопленные к ФНОα у одного вида (например, человека), не вступают в перекрестную реакцию с ФНОα другого вида животных (например, мыши), несмотря на идентичность последовательностей, превышающую 80%, и способность ФНОα связываться с ФНОα-рецепторами других видов. Если степень вариации последовательности картировать на пространственной структуре, сразу же становится ясно, что зоны молекулы с наиболее вариабельными последовательностями соответствуют поверхностным петлям, доступным для антител. Области высококонсервативных остатков внутри сандвича или на поверхности раздела тримера эффективно не видимы для антител. Поэтому эпитоп для антитела к ФНОα всегда будет содержать несколько остатков, которые будут варьировать у разных видов, устраняя таким образом связывание антител. Это означает, что характеристики взаимодействия между ФНОα и его рецептором должны как-то отличаться от характеристик, необходимых для связывания антитела с ФНОα.
Б. Сайт-нaправленный мутагенез
Роль различных специфических остатков и областей молекулы ФНОα по отношению к ее биологической (цитотоксической) активности и связыванию с рецептором была исследована путем замены этих остатков на различные аминокислоты или удалением части последовательности с использованием методик целенаправленного мутагенеза (Jones et al, op. Cit. P.113-119).
Удаления до восьми остатков с N-конца без какого-либо вредного эффекта на биологическую активность достаточно для того, чтобы подчеркнуть несущественность этой области для общей стабильности молекулы. N-концевые остатки, по-видимому, оказывают непрямое, вторичное влияние на биологическую эффективность тримера ФНОα.
Неконсервативные замены в норме высококонсервативных остатков, которые образуют плотно упакованную сердцевину β-сандвича, нарушают структуру и, следовательно, уничтожают биологическую активность ФНОα (Yamagishi et al., Protein Engeneering, Vol.3, N 8, p.713-719 (1989)). Многие из таких мутантных белков (мутеинов) не могут образовывать стабильную, правильно сложенную молекулу. Некоторые консервативные замены допустимы внутри гидрофобной зоны у основания общей оси; однако, по-видимому, значительно большие вариации существуют в более свободно упакованной области вблизи вершины тримера. В частности, Цистеин 69 и Цистеин 101, которые образуют дисульфидный мостик между двумя соединительными петлями на свободно упакованной вершине молекулы, относительно нечувствительны к изменениям (см. фиг.1а). Тем не менее, в целом, чтобы сохранить определенную биологическую активность ФНОα, мутации вблизи центральной оси ФНОα должны быть высококонсервативными, сохраняющими общую форму ФНОα.
Остатки на поверхности молекулы имеют значительно большую свободу для мутаций без губительных для структуры последствий, что подтверждается увеличением числа вариаций остатков в этой категории у разных видов. Поэтому резкое снижение биологической активности ФНОα из-за замен в этой области указывает на прямое участие этих остатков в функциональном взаимодействии тримера ФНОα с его рецептором. Такими остатками, по-видимому, являются остатки, включающие Аргинин 31, Аргинин 32 и Аланин 33, расположенные в соединительной петле между В и В′-цепями заднего β-слоя, Серин 86 и Тирозин 87, расположенные в соединительной петле между Е и F нитями переднего β-слоя, и Глутаминовая кислота 146, расположенная в соединительной петле между Н-нитью переднего β-слоя и I-нитью заднего β-слоя (см. фиг.3). Они, по-видимому, приходятся на две разные “горячие зоны” на передней и задней сторонах мономера ФНОα. Распределение всех вредных мутаций безотносительно к структурной категории еще более подкрепляет эту картину. Существование таких “горячих областей” для чувствительности биологической функции к мутациям было описано Yamagishi et al., op. cit., и Goh et al. Protein Engeneering, Vol.4, № 4, p. 385-389 (1991).
Исследования мутаций полипептидов ФНОα человека были описаны во многих патентных публикациях.
Так, в документе ЕП 251037 описаны многочисленные сайт-специфически мутировавшие молекулы ФНОα, которые были растворимыми и обладали цитотоксической и противоопухолевой активностью, характерной для ФНОα человека, in vitro и/или in vivo.
Другие мутеины с активностью ФНОα описаны в документе WO 90/07579. Мутеины с более высокой аффинностью к рецептору р75-ФНО человека, чем к рецептору р55-ФНО человека, описаны в документе ЕР-А-619 372.
Ни одна из этих процитированных публикаций, включенных в данное описание посредством ссылок на них, не раскрывает сущности модификации молекулы ФНОα, имеющей целью уничтожить биологическую активность ФНОα и обеспечить способность индуцировать образование антител к человеческому ФНОα дикого типа.
Тем не менее, они дают полезную основополагающую информацию относительно ФНОα и его биологического действия. Кроме того, в них раскрыты сущность получения и экспрессии аналогов ФНОα и их формулы; описаны также анализы связывания с рецепторами.
3. Резюме
В настоящее время можно привести многообразныее данные, относящиеся к специфическому взаимоотношению структуры и функции ФНОα. Все имеющиеся доказательства указывают на важность тримера как стабильной природной единицы. Очевидно, что две “горячих” области, расположенные на разных сторонах мономера ФНОα, тесно сближены друг с другом у соседних субъединиц тримера. Поэтому функционально значимая область, состоящая из остатков 31-35, 84-87 и 143-148, по-видимому, расположена на поверхности раздела между двумя субъединицами на нижней половине тримера. Yamagishi et al. op. cit. сообщил об отсутствии способности к связыванию с рецептором и цитотоксичности при мутации с заменой Аспарагиновой кислоты в положении 143 на Тирозин, a Tsujimoto et al., J.Biochem. 101, р. 919-925 (1987) описал сходный эффект для замены Аргинина 31 и Аргинина 32 на Аспарагин и Треонин. Таким образом, этот сайт может быть непосредственно связан со связыванием с рецептором и цитотоксичностью. Интересно, что область связывания с рецептором ФНОα, по-видимому, лежит на поверхности раздела между двумя субъединицами.
В целом, подробное описание трехмерной структуры ФНОα служит для объяснения широкого спектра наблюдений относительно связывания антител, олигомеризации и целенаправленного мутагенеза. Если рассматривать структуру совместно с недавно полученными, многочисленными целенаправленными мутантами, область, имеющая биологическое значение в отношении связывания с рецептором, по-видимому, расположена на субъединицах в нижней половине тримера.
Характеристика аналогов
Согласно изобретению, раскрытому в документе WO 95/05849, авторами которого являются некоторые из авторов настоящего изобретения, предложен способ модификации собственных белков таким образом, чтобы индуцировать реакцию антител против немодифицированного собственного белка, где аналог собственного белка обеспечивается средствами молекулярной биологии. В частности, один или несколько пептидных фрагментов собственного белка замещают одним или несколькими пептидными фрагментами, содержащими иммунодоминантные, чужеродные эпитопы для Т-клеток.
До изобретения, описанного в WO 95/05849, была известна конъюгация пептидов или белков, включая собственные белки, с белками-носителями, включающими эпитоп для Т-клетки. Документ WO 92/19746 раскрывает сущность рекомбинантного полипептида, включающего последовательность LHRH, один или несколько эпитопов для Т-клеток и сайт стерилизации, который пригоден для использования в вакцинах для иммунокастрации животных.
В WO 95/05849 признано предпочтительным, чтобы иммунодоминантный эпитоп для Т-клетки был встроен так, чтобы на каждой стороне встроенного эпитопа были сохранены фланговые области исходного белка, включающие, по меньшей мере, 4 аминокислоты. Другими словами, эпитоп не должен быть конъюгирован с собственным белком как белок слияния. Предпочтительно замещение должно быть произведено так, чтобы по существу сохранить третичную структуру исходного белка. Кроме этого, не дано специфических указаний относительно внутримолекулярного положения встроенного эпитопа, оптимального для обеспечения наиболее мощной реакции антител против немодифицированного собственного белка. Предположительно, оно будет разным для разных собственных белков, но на основании общего принципа спецификации наиболее подходящее положение(я) может быть определено без чрезмерного экспериментирования путем селекции пептидов, включающих соответствующие иммунодоминантные эпитопы, замены пептидных последовательностей по существу одинаковой длины в различных частях молекулы собственного белка и оценки повышенной реакции антител при помощи подходящих методик анализа.
При использовании современных средств моделирования, вероятно, очень трудно предсказать, приведет ли замена одного или нескольких пептидных фрагментов в белке к изменению третичной структуры исходного белка. Поэтому при поиске ключевых соединений в программах разработки лекарственных средств необходимо на ранней стадии разработки принять решение о проведении стандартной процедуры скрининга, чтобы оценить, сохранил ли модифицированный собственный белок третичную структуру исходного белка. Это может быть выполнено с использованием нескольких экспериментальных методик, которые были описаны в многочисленных руководствах по определению характеристик белков, например флуоресцентной спектроскопии, кругового дихроизма в ближнем УФ-диапазоне, инфракрасной спектроскопии с преобразованием Фурье и многомерных ЯМР методик (Physical Methods to Characterize Pharmaceutical Proteins", Biotechnology Vol. 7 Eds. J.N. Herron, W. Jiskoot & D.J.A. Crommelin, Plenum Press, New York (1995)). В идеале, в процессе скрининга ключевых соединений следует комбинировать две или несколько экспериментальных методик, указанных выше, чтобы оценить, возникло или нет изменение третичной структуры.
Краткий перечень чертежей
Фиг.1(а) иллюстрирует кристаллическую структуру нативного мономера ФНОα. Данная фигура представляет собой схематическое изображение укладки субъединицы, β-цепи показаны в виде толстых стрелок в направлении от аминогруппы к карбоксигруппе, а соединительные петли изображены в виде тонких линий. Дисульфидный мостик обозначен молнией, а область высокой гибкости поперечно заштрихована. Для этой ориентации общая ось симметрии тримера будет вертикальной.
Фиг.1(b) является чертежом Сα цепи кристаллической структуры мономера ФНОα. Это подробное изображение следует использовать вместе с фиг.1(а), чтобы получить точное сопоставление аминокислотной последовательности с более понятным, но стилизованным изображением укладки субъединицы.
Фиг.1(с) изображает структуру ФНОα как мотив “рулета с начинкой”. Вставка между β-цепями В и С показана в виде пунктирной линии; соединение между В и С идет прямо поперек вершины молекулы.
Фиг.2 изображает упаковку β-сандвичей типа край-к-грани в тримере ФНО. Вид в направлении общей оси изображает узкий срез тримера с β-цепями, изображенными как ленты, идущие в страницу и из нее.
Фиг.3(а) иллюстрирует нуклеотидную последовательность ДНК, кодирующую фактор некроза опухоли (ФНОα), имеющий последовательность аминокислот, показанную на фиг.3(b).
Нуклеотидную последовательность ДНК можно получить в Банке Генов под инвентарным № М10988, SEQ ID №:339737.
Последовательность была описана Wang et al., Science 228, 149-154 (1985). Последовательность включает кодоны, кодирующие -76--1 предпоследовательность ФНОα человека.
Полная последовательность нуклеотидов гена, включающая интроны, была описана Nedwin et al., Nucleic Acids, Res. 13 (17) 6361-6373 (1985), Shirai et al., Nature 313 (6005), 803-806 (1985) и Dennica et al., Nature 312 (5996), 724-729 (1984).
Фиг.3(b) показывает аминокислотную последовательность ФНОα человека, включающую предпоследовательность -76--1.
Фиг.4(а) схематически иллюстрирует замены на иммунодоминантные эпитопы Р2 и Р30 в ФНОα дикого типа (ДТ) с образованием аналогов ФНОα от ФНO2-1 до 2-7 и от ФНО30-1 до 30-5.
Фиг.4(b) показывает точные положения замен в ДТ последовательности для отдельных аналогов ФНОα.
Фигуры 5(а) и 5(b) показывают структуру аналогов на основе фиг.1(а), где отдельные замены на Р2 и Р31 в цепях β-слоев и соединительных петлях, соответственно помечены черным цветом.
Фиг.6 показывает биологическую активность аналогов ФНОα в пробе L929 (Meager, A., Leung, Н., & Wooley, J. Assays for Tumor Necrosis Factor b related cytokines., J. Immunol. Meth. 116, 1-17 (1989)) по сравнению с рекомбинантным ФНОα.
Фиг.7 показывает реакцию антител к ФНОα человека у кроликов при вакцинации кроликов молекулами модифицированного ФНОα.
Фиг.8 показывает способность Р2/Р30-модифицированных молекул ФНОα человека индуцировать образование нейтрализующих антител, измеренную в пробе с клетками L929.
Фиг.9 показывает способность Р2/Р30-модифицированных молекул ФНОα человека индуцировать - при введении кроликам - образование нейтрализующих антител, измеренную в пробе с рецепторами.
Фиг.10 показывает реакцию мононуклеарных клеток периферической крови (МКПК) у трех доноров на СА и Р2 и Р30-пептиды.
Фиг.11 показывает реакцию поликлональной пролиферации у двух доноров с использованием различных Р2 и Р30-модифицированных молекул ФНО.
Фиг.12 показывает Индексы Пролиферации (ИП), рассчитанные по 34 экспериментам с Р2 и Р30-модифицированными молекулами ФНОα.
Фиг.13 показывает реакцию МКПК на Р2 и Р30-модифицрованные белки ФНОα у лиц, специфически реагирующих на Р2 и Р30 соответственно.
Фиг.14 показывает сходную реакцию МКПК у двух других доноров.
Фиг.15 показывает влияние фланговых аминокислот на распознавание Р2 и Р30 Т-клетками.
Фиг.16 иллюстрирует стратегию мутаций, использованную для получения модифицированных молекул ФНОα.
Сущность изобретения
Цель данного изобретения - обеспечить руководящие указания относительно того, как можно модифицировать в пределах общего объема изобретения, раскрытого в WO 95/05849, определенный собственный белок, а именно ФНОα человека, чтобы он был биологически активным и способным индуцировать сильную реакцию нейтрализующих антител на биологически активный ФНОα дикого типа. В контексте данного изобретения “биологически неактивный” относится к различного рода активности ФНОα дикого типа, главным образом к его цитотоксической активности.
При обсуждении третичной структуры ФНОα, приведенного выше, было показано, что биологически активный ФНОα является тримером трех своих субъединиц. Благодаря упаковке “край-к-грани”, “задний β-слой” представляет собой “скрытую” область контакта между субъединицами, которая абсолютно недоступна для растворителя. Значительные замены в этой области почти неизбежно лишают молекулу ФНОα любой биологической активности. В противоположность этому “передний β-слой” и соединительные области обеспечивают доступную площадь поверхности, которая включает области, взаимодействующие с рецепторами ФНОα. Антитела к этим областям поэтому, вероятно, будут способны нарушать связывание с рецептором и за счет этого обладать способностью к нейтрализации ФНОα.
Специалисты в данной области техники, которые хотят сконструировать детоксифицированную, но еще иммуногенную молекулу ФНОα в соответствии с изобретением по WO 95/05849, должны поэтому в первую очередь встроить иммунодоминантный эпитоп для Т-клетки в задний β-слой мономера ФНОα. Поэтому модификации этой области наиболее вероятно могут уничтожить биологическую активность ФНОα и сохранить доступный для рецепторов передний β-слой свободным для взаимодействия с антителами. Это также совпадает с обсуждением сайт-направленного мутагенеза в плотно упакованной внутренней части β-сандвича, обсуждавшегося выше.
Однако неожиданно это предположение не подтверждается. Как будет ясно из результатов тестов, приведенных ниже, результат был абсолютно противоположным, так как замены, включающие В и С-цепи заднего β-слоя, неожиданно дали аналоги ФНОα, которые были неспособны индуцировать образование нейтрализующих антител к ФНОα.
Таким образом, задача, решаемая настоящим изобретением, заключается в том, чтобы обеспечить модифицированную молекулу ФНОα, способную увеличивать количество нейтрализующих антител к человеческому ФНОα дикого типа после введения вышеуказанной модифицированной молекулы ФНОα человеку-реципиенту, в которой по меньшей мере один пептидный фрагмент молекулы ФНОα человека был заменен по меньшей мере на один пептид, про который известно, что он содержит иммунодоминантный эпитоп для Т-клетки, или усеченную форму вышеуказанной молекулы, содержащую иммунодоминантный эпитоп и одну или обе фланговых области молекулы, содержащие по меньшей мере один эпитоп ФНОα для В-клетки, причем замена приводит к существенному изменению последовательности аминокислот в переднем β-слое, в любой из соединительных петель и/или в любой из В′, I и D-цепей заднего β-слоя. Предположительно, замен в В и G-цепях заднего β-слоя следует избегать.
В контексте данного изобретения “существенное изменение” подразумевает изменение, которое больше, чем простое консервативное замещение отдельных аминокислот и точечные мутации, которые не нарушают вторичную и/или третичную структуру ФНОα дикого типа. Другими словами, эпитоп для Т-клетки, встроенный в последовательность ФНОα, должен предпочтительно встраивать последовательность, которая очень мало гомологична последовательности ФНОα дикого типа.
В соответствии с данным изобретением также обеспечивается модифицированная молекула ФНОα человека, способная увеличивать количество нейтрализующих антител к человеческому ФНОα дикого типа после введения вышеуказанной модифицированной молекулы ФНОα человеку-реципиенту, в которой по меньшей мере один пептидный фрагмент молекулы ФНОα человека был замещен по меньшей мере на один пептид, про который известно, что он содержит иммунодоминантный эпитоп для Т-клетки, или усеченная форма вышеуказанной молекулы, содержащая иммунодоминантный эпитоп и одну или обе фланговые области молекулы ФНОα человека, включающие по меньшей мере один эпитоп ФНОα для В-клетки, причем вышеуказанная модифицированная молекула ФНОα в значительной степени лишена активности ФНОα.
Термин “в значительной мере лишена активности ФНОα” в контексте настоящего изобретения предназначен обозначать, что введение человеческому существу такой модифицированной молекулы ФНОα не приводит к существенным неблагоприятным эффектам, обусловленным известными цитокинными эффектами, вызываемыми нативным ФНОα. Другими словами, модифицированные молекулы ФНОα по настоящему изобретению являются фармацевтически приемлемыми веществами.
Чтобы проверить, является ли модифицированная молекула ФНОα человека по настоящему изобретению в значительной мере лишенной активности ФНОα, может быть использована биопроба L929, описанная далее. Более того, чтобы подтвердить иммуногенный характер модифицированной молекулы ФНОα, антитела к модифицированной молекуле ФНОα, образовавшиеся у подходящего реципиента, должны достоверно ингибировать активность ФНОα дикого типа в биопробе L929, как она описана далее, и/или вышеуказанные антитела должны достоверно ингибировать связывание ФНОα дикого типа с ФНОα-рецептором 1 с молекулярной массой 55 кДа (ФНОα-R55) или ФНОα-рецептором с молекулярной массой 75 кДа (ФНОα-R75).
Этими подходящими реципиентами могут быть, например, приматы, такие как обезьяны Rhesus или Cynomuleus, грызуны, такие как крысы, мыши или морские свинки, или лагоморфы, такие как кролики.
Анализы, пригодные для оценки потенциальных возможностей модифицированных молекул по настоящему изобретению, выполняют с использованием антител или антисыворотки в концентрации, соответствующей чувствительности анализа. При этом они должны учитывать способность отображать физиологические условия относительно участвующих реагентов, или они должны обеспечивать возможность экстраполяции до физиологических условий, исходя из результатов, полученных при анализе. Другими словами, результаты анализа должны показывать, что физиологическая концентрация антител к ФНОα in vivo может снизить активность ФНОα на, по меньшей мере, 10, 15, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 100%. Специалист в данной области техники легко поймет, как определить такие условия.
Следует понимать, что подходящим “достоверным ингибированием ФНОα дикого типа” может быть такое, которое приводит к снижению или устранению неблагоприятных эффектов ФНОα дикого типа. Таким подходящим ингибированием может быть ингибирование по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или даже на 100%.
Исходя из этих предпосылок, дана характеристика модифицированных молекул ФНОα человека по данному изобретению, в которых была произведена замена в определенных областях молекулы ФНОα и которые включают нити переднего β-слоя и/или соединительные петли, так, чтобы по существу сохранить β-слоистую структуру любой из цепей заднего β-листа.
Хотя это еще не было полностью подтверждено экспериментально, следует принять, что эта особенность является общей для цепей, формирующих задний β-слой, так что предпочтительно замены следует производить в областях молекулы ФНОα, которые не содержат какой-либо полной цепи заднего β-слоя. Принимая во внимание приведенное выше обсуждение функционального значения остатков 31-35, можно сделать вывод, что соединительных петель между отдельными нитями заднего β-слоя также предпочтительно избегать.
Однако допустимо, чтобы замена производилась в тех областях молекулы ФНОα, которые содержат сегмент D-нити заднего β-слоя.
В соответствии с предпочтительным примером осуществления изобретения замена включает по меньшей мере сегмент Н-цепи переднего β-слоя и соединительной петли к 1-цепи заднего β-слоя, предпочтительно аминокислоты с 132 по 146. В соответствии с другим примером осуществления изобретения замена включает сегменты Н и I-цепей и всю соединительную петлю, предпочтительно - аминокислоты с 132 по 152. В соответствии с еще одним в настоящее время предпочтительным примером осуществления изобретения замена включает сегмент D-цепи заднего β-слоя, по меньшей мере сегмент Е-цепи переднего β-слоя и всю соединительную петлю, предпочтительно аминокислоты с 65 по 79 или с 64 по 84.
В соответствии с еще одним примером осуществления изобретения замена включает полностью С′ и С цепи переднего р-слоя и сегмент D-цепи заднего β-слоя, предпочтительно аминокислоты с 40 по 60.
В соответствии с еще одним примером осуществления изобретения замена включает по меньшей мере сегмент Е-цепи переднего β-слоя и одну или обе соединительных петли, предпочтительно аминокислоты с 76 по 90.
Встроенный эпитоп для Т-клетки должен предпочтительно быть неоднородным и известным в качестве иммуногенного для большинства типов молекул HLA (ОКГ) человека II класса. Подходящие эпитопы можно получить, например, из столбнячного анатоксина, предпочтительно эпитоп Р2 и/или Р30, Panina-Bordignon et al., Eur. J.Immunol. 19:2237-42, 1989. Также могут быть использованы эпитопы, полученные из дифтерийного анатоксина.
Предпочтительные модифицированные молекулы ФНОα человека (аналоги ФНОα), с положением замены, указанным выше, показаны в прилагаемом перечне последовательностей как SEQ ID NО:8 и SEQ ID NO:16.
Другие подходящие аналоги ФНОα перечислены как SEQ ID NO:4, 10, 14 и 16.
Изобретение также относится к усеченным аналогам вышеуказанных модифицированных аналогов ФНОα по данному изобретению. Поэтому усеченные аналоги молекул ФНОα, содержащие случайный и иммунодоминантный эпитоп для Т-клетки и одну или обе фланговых области, включающие по меньшей мере один ФНОα эпитоп для В-клетки, предпочтительно включающий по меньшей мере пять аминокислот, также могут представлять собой вероятный активный ингредиент в ФНОα-вакцине по данному изобретению. Эпитоп для Т-клетки должен вызывать пролиферацию Т-клеток, если он относится к молекулам ОКГ класса II, представляемым АПК, тогда как эпитоп для В-клетки потенциально должен распознаваться рецепторами иммуноглобулинов на В-клетках и впоследствии будет представлен молекулами I класса ОКГ на этих клетках. Это образует основу для нарастания иммунной реакции против ФНОα дикого типа, несущего эпитоп для В-клетки, в соответствии с WO 95/05849 и настоящим изобретением.
Эпитопы для В-клеток можно идентифицировать в ФНОα как теоретически, так и экспериментально. Алгоритмы идентификации потенциальных линейных эпитопов для В-клеток были опубликованы, и это образует основу экспериментального исследования природы этих потенциальных эпитопов. Антитела, накапливаемые в гетерологичной системе (например, у кроликов) в ответ на инъекции таких усеченных молекул ФНОα, включающих эпитопы для Т-клеток, можно проанализировать на способность связывать нативный ФНОα человека in vitro, предпочтительно путем нейтрализации. Группы моноклональных антител, известных в качестве нейтрализующих, можно подвергнуть скринингу in vitro в отношении способности связывать потенциальные эпитопы ФНОα для В-клеток. Оба этих способа являются стратегиями идентификации вероятных и наилучших эпитопов для В-клеток.
Вышеуказанные аналоги ФНОα, по-видимому, остаются в форме мономера, так как модификация, вероятно, разрушает свойственную дикому типу ФНОα тенденцию к тримеризации.
Тем не менее изобретение также относится к димерам, олигомерам, особенно тримерам или мультимерам заявляемых молекул ФНОα, которые лишены ФНОα-активности, а также к изолированным молекулам ДНК, которые кодируют модифицированные молекулы ФНОα согласно изобретению.
Изолированные молекулы ДНК, кодирующие предпочтительные аналоги ФНОα, имеют последовательности нуклеотидов, перечисленные как SEQ ID NO:7 и 15, а молекулы ДНК, кодирующие другие пригодные аналоги, перечислены как SEQ ID NO:3, 9, 13 и 15.
Изобретение далее включает векторы, содержащие изолированные молекулы ДНК, кодирующие аналоги, и вектора экспрессии, включающие вышеуказанные молекулы ДНК, оперативно присоединенные к подходящим последовательностям, регулирующим экспрессию.
В другом своем аспекте изобретение относится к реципиенту, трансформируемому вектором экспрессии вышеуказанного аналога.
Вышеуказанный реципиент может быть любым из реципиентов, обычно используемых для экспрессии, например штаммом бактерий, дрожжей, грибков или клеточных линий насекомых, млекопитающих или птиц.
Изобретение также относится к способу производства заявляемых аналогов ФНОα, согласно которому клетки-реципиенты, трансформированные вектором экспрессии аналога, культивируют при подходящих условиях, обеспечивающих продукцию аналога, а произведенный аналог извлекается.
Если это желательно, модифицированные молекулы ФНОα по настоящему изобретению могут быть экспрессированы в качестве белка слияния или образовывать часть белка слияния с подходящей усиливающей молекулой, предпочтительно иммунологически активным усилителем, таким как GM-CSF, HSP70, или интерлейкином, например интерлейкином 2.
Более специфически, модифицированные молекулы ФНОα производят путем замен на соответствующие сегменты генов, содержащих или представляющих собой иммунодоминантные эпитопы для Т-клеток, внутри гена, кодирующего нативную молекулу ФНОα человека. Впоследствии модифицированный ФНОα-ген экспрессируют в подходящем эукариотическом или прокариотическом векторе экспрессии. Экспрессированные модифицированные молекулы ФНОα очищают и повторно складывают, как описано ниже.
Хотя может быть предпочтительным встраивать целый эпитоп для Т-клетки, в некоторых случаях может оказаться полезным встроить пептидную последовательность, включающую как эпитоп, так и фланговые области белка, из которого получен эпитоп.
В соответствии с изобретением модифицированные молекулы ФНОα человека могут быть использованы в вакцинах против ФНОα. Стратегии разработки рецептур вакцин на основе очищенных белков, таких как модулированные собственные белки, в основном соответствуют стратегиям разработки рецептур любых других лекарственных средств на основе белков. Потенциальные проблемы и руководящие указания, необходимые для преодоления этих проблем, - как, например, сохранение третичной структуры - изложены в нескольких руководствах, например, "Therapeutic Peptides and Protein Formulation / Processing and Delivery Systems" Ed. A.K.Banga; Technomic Publishing AG, Basel 1995. Использование усиливающего вещества, например гидроокиси алюминия, фосфата алюминия (Адъю-Фос), фосфата кальция, аналога мурамилового дипептида или нескольких последних разработок в области усилителей действия вакцин, таких как биологически разрушаемые микрочастицы и Иском, является проблемой, хорошо знакомой научным работникам в сфере фармакологии, работающим в этой области.
Приготовление вакцин по настоящему изобретению, содержащих пептидные последовательности в качестве активных ингредиентов, в целом хорошо известно в данной области техники, что иллюстрируют патенты США № 4608251; 4601903; 4599231; 4599230; 4596792 и 4578770, включенные в данное описание посредством ссылок на них. В типичном случае такие вакцины приготавливают в виде инъецируемых вакцин, либо в виде жидких растворов, либо в виде суспензий; твердые формы, пригодные для приготовления раствора или суспензии в жидкости перед инъекцией, также могут быть приготовлены. Препарат также может быть эмульгирован. Активный иммуногенный ингредиент часто смешивают с наполнителями, которые фармацевтически приемлемы и совместимы с активным ингредиентом. Подходящими наполнителями являются, например, вода, физиологический раствор, декстроза, глицерин, этанол и тому подобные вещества и их комбинации. Кроме того, если это желательно, вакцина может содержать небольшие количества вспомогательных веществ, таких как смачивающие и эмульгирующие вещества, агенты, буферирующие рН, или усилители, которые повышают эффективность вакцин.
Вакцины традиционно вводят парентерально, путем инъекции, например подкожно или внутримышечно. Дополнительные рецептуры, которые пригодны для других способов введения, включают суппозитории и в некоторых случаях рецептуры для приема внутрь. В случае суппозиториев традиционные связующие вещества и носители могут включать, например, полиалкиленгликоли или триглицериды; такие суппозитории могут быть сформированы из микстур, содержащих активный ингредиент в диапазоне концентраций от 0,5 до 10%, предпочтительно 1-2%. Оральные рецептуры включают такие нормально используемые наполнители, как, например, фармацевтические количества маннита, лактозы, крахмала, стеарата магния, сахарина натрия, целлюлозы, карбоната магния и тому подобные. Эти композиции имеют форму растворов, суспензий, таблеток, пилюль, капсул, рецептур с отсроченным выделением или порошков и содержат 10-95% активного ингредиента, предпочтительно 25-70%.
Полипептиды могут быть введены в рецептуру вакцины в нейтральной форме или в форме соли. Фармацевтически приемлемые соли включают аддитивные соли, образующиеся при присоединении кислот (образуемые свободными аминогруппами пептида), при взаимодействии с неорганическими кислотами, такими, например, как соляная или фосфорная кислоты, или с такими органическими кислотами как уксусная, щавелевая, винная, миндальная и подобными им. Соли, образующиеся со свободными карбоксильными группами, также могут быть получены из неорганических оснований, таких, например, как гидроокиси натрия, калия, аммония, кальция или железа, и таких органических оснований, как изопропиламин, триметиламин, 2-этиламиноэтанол, гистидин, прокаин и тому подобные.
Вакцины вводят способом, совместимым с дозировочной рецептурой, и в таких количествах, которые будут терапевтически эффективными и иммуногенными. Количество, которое необходимо ввести, зависит от субъекта, подлежащего лечению, в том числе, например, от способности иммунной системы индивидуума давать иммунный ответ, и от желаемого уровня защиты, который, в свою очередь, зависит от уровня ФНОα у больного. Диапазоны подходящих дозировок - порядка нескольких сотен миллиграммов активного ингредиента на вакцинацию с предпочтительным диапазоном от 1 до 300 мкг, особенно в диапазоне от 10 до 50 мкг. Подходящие режимы начального введения и повторных иммунизаций также вариабельны, но типично представляют собой начальное введение с последующими прививками или другими введениями.
Способ применения может быть очень разным. Применимы любые традиционные методы введения вакцины. Считается, что таковые включают оральное применение на твердой физиологически приемлемой основе или в физиологически приемлемой дисперсии, парентеральное введение, введение путем инъекции и тому подобное. Дозировка вакцины будет зависеть от пути введения и будет изменяться в соответствии с возрастом лица, подлежащего вакцинации, и в меньшей степени - от массы тела лица, подлежащего вакцинации.
Некоторые модифицированные молекулы ФНОα по настоящему изобретению достаточно иммуногенны в вакцине, но для некоторых других иммунная реакция будет усилена, если вакцина дополнительно содержит повышающее иммуногенность вещество.
Различные методы достижения эффекта повышения иммуногенности для вакцины включают применение агентов, таких как гидроокись или фосфат алюминия (квасцы), обычно используемых в виде 0,05-0,1-процентного раствора в фосфатном буфере, примесь синтетических полимеров сахаров (Карбопол), используемых в виде 0,25-процентного раствора, агрегация белка в вакцине тепловой обработкой при температурах в диапазоне от 70 до 101°С в течение промежутков времени от 30 секунд до 2 минут соответственно. Также могут быть использованы агрегация путем реактивации обработанными пепсином (Fаb)-антителами к альбумину, смешивание с бактериальными клетками, такими как C.parvum, или эндотоксинами или липополисахаридными компонентами грам-отрицательных бактерий, эмульгирование в физиологически приемлемых масляных носителях, таких как маннида моноолеат (Арацель А) или эмульгирование с 20-процентным раствором перфторуглерода (Флуозоль-DA), используемым в качестве блокирующего заместителя. В соответствии с изобретением ДДА (диметилдиоктадециламмония бромид) является интересным кандидатом в вещества, повышающие иммуногенность, но QuilA и RIBI также являются интересными возможностями. Другими возможностями являются монофосфориллипид А (МФЛ) и мурамилдипептид (МДП). Другими подходящими адъювантами являются гидроокись алюминия, фосфат алюминия (Адью-Фос), фосфат кальция, аналог мурамилдипептида. Некоторые из недавних разработок в области усилителей иммуногенности вакцин, такие как биоразрушаемые микрочастицы и Иском также могут быть пригодными для применения.
Другой очень интересной (и поэтому предпочтительной) возможностью достижения эффекта усиления иммуногенности является применение методики, описанной в работе Gosselin et al., 1992 (которая включена в данное описание посредством ссылки на нее). Вкратце, представление соответствующего антигена, такого как модифицированная молекула ФНОα по настоящему изобретению может быть повышено путем конъюгации такого антигена с антителами (или антигенсвязывающими фрагментами антител) к FcγRI-рецепторам на моноцитах/макрофагах. Было продемонстрировано, что конъюгаты между антигеном и анти-FcγRI особенно повышают иммуногенность для целей вакцинации.
Другие возможности включают применение иммуномодулирующих веществ, таких как лимфокины (например, IFN-γ, ИЛ-2 и ИЛ-12) или синтетических индукторов IFN-γ, таких как поли-I:С, в сочетании с вышеуказанными адъювантами.
Во многих случаях будет необходимо производить множественные введения вакцины, обычно не более шести вакцинаций, еще чаще не более четырех вакцинаций, и предпочтительно одну или более, обычно минимум три вакцинации. Вакцинации обычно будут производиться через интервалы от двух до двенадцати недель, чаще - через интервалы от трех до пяти недель. Периодические повторные иммунизации через интервалы в 1-5 лет, обычно - через 3 года, желательны для поддержания желаемых уровней защитной иммунности.
Одна из причин для смешивания полипептидов по изобретению с веществом, повышающим иммуногенность, - эффективная активация клеточного иммунного ответа. Однако этот эффект можно также получить другими способами, например путем экспрессии эффективного антигена вакцины в непатогенном микроорганизме. Хорошо известным примером такого микроорганизма является Mycobacterium bovis BCG.
Предпочтительно, чтобы непатогенный микроорганизм был бактерией, например, выбранной из группы, состоящей из родов Mycobacterium, Salmonella, Pseudomonas и Eschericia. Особо предпочтительно, чтобы непатогенный микроорганизм был Mycobacterium bovis.
Инкорпорация одной или нескольких копий нуклеотидной последовательности, кодирующей молекулу в соответствии с данным изобретением, в микобактерию из штамма М.bovis BCG будет усиливать иммуногенный эффект штамма BCG. Ожидается, что инкорпорация более чем одной копии нуклеотидной последовательности по изобретению еще больше усилит иммунный ответ, и вследствие этого аспектом изобретения является вакцина, в которой по меньшей мере 2 копии нуклеотидной последовательности ДНК, кодирующей молекулу, инкорпорированы в геном микроорганизма, например минимум 5 копий. Копии нуклеотидных последовательностей ДНК могут либо быть идентичными, кодирующими идентичные молекулы, либо быть вариантами одной и той же нуклеотидной последовательности ДНК, кодирующей идентичные или гомологичные пептиды, либо в другом примере осуществления изобретения быть различными нуклеотидными последовательностями ДНК, кодирующими различные полипептиды, среди которых по меньшей мере один полипептид соответствует данному изобретению.
Живые вакцины по изобретению могут быть приготовлены путем культивирования трансформированной непатогенной клетки в соответствии с изобретением, и переноса этих клеток в среду для вакцины, и по выбору - добавления носителя, наполнителя и/или вещества, повышающего иммуногенность.
Кроме их использования в качестве исходных продуктов для синтеза молекулы по изобретению и для гибридизационных проб (полезны для анализов прямой гибридизации или в качестве затравок, например, в ПЦР или других методах амплификации молекул) фрагменты нуклеиновой кислоты по изобретению могут быть использованы для влияния in vivo на экспрессию антигенов, то есть фрагменты нуклеиновой кислоты могут быть использованы в так называемых ДНК-вакцинах. Недавние исследования показали, что фрагмент ДНК, клонированный в векторе, который является нерепликативным в эукариотических клетках, может быть введен в животное (включая человеческое существо) при помощи, например, внутримышечной инъекции или чрескожного введения (так называемый подход “генного ружья”). ДНК поглощают, например, мышечные клетки, и представляющий интерес ген экспрессируется промотором, который функционирует в эукариотах, например вирусным промотором, а затем продукт гена стимулирует иммунную систему. Обзор этих недавно открытых способов дан в работе Ulmer et al., 1993, которая включена в данное описание посредством ссылки на нее.
Эффективность такой “ДНК-вакцины”, по-видимому, может быть повышена путем введения гена, кодирующего продукт экспрессии, вместе с фрагментом ДНК, кодирующим полипептид, который способен модулировать иммунный ответ. Например, ген, кодирующий предшественники лимфокинов или лимфокины (например, IFN-γ, ИЛ-2 или ИЛ-12), может быть введен совместно с геном, кодирующим иммуногенный белок, либо путем введения двух отдельных фрагментов ДНК, либо путем введения обоих фрагментов ДНК, заключенных в одном и том же векторе.
Вакцины могут быть использованы для профилактики/лечения любого из заболеваний, описанных выше, патофизиология которых характеризуется выделением ФНОα, особенно хронических воспалительных заболеваний. В качестве примеров могут быть упомянуты ревматоидный артрит и воспалительные заболевания кишечника (ВЗК). Последние включают язвенный колит и болезнь Крона, в частности колит Крона. Другие примеры - это рак, кахексия, часто связанная с раком, рассеянный склероз, диабет, псориаз, остеопороз и астма. Рак, который предпочтительно можно лечить или предупреждать в соответствии с изобретением, можно классифицировать гистологически на злокачественные эпителиальные опухоли, включающие карциномы и аденокарциномы, и на злокачественные неэпителиальные опухоли, включающие липосаркомы, фибросаркомы, хондросаркомы, остеосаркомы, лейомиосаркомы, рабдомиосаркомы, глиомы, нейробластомы, медуллобластомы, злокачественные меланомы, злокачественные менингиомы, различные лейкемии, различные миелопролиферативные заболевания, различные лимфомы (ходжкиновская лимфома и неходжкиновская лимфома), гемангиосаркому, саркому Калоши, лимфангиосаркому, злокачественную тератому, дисгерминому, семиному и хориокарциному.
Карциномы и аденокарциномы наиболее многочисленны (составляют примерно 90% смертей от рака) и поэтому являются интересными целевыми заболеваниями для лечения/профилактики согласно изобретению. Наиболее важными карциномами и аденокарциномами являются опухоли дыхательные путей (особенно бронхиального происхождения), грудной клетки, колоректальные и желудка. Однако карциномы и аднокарциномы предстательной железы, яичников, лимфоидной ткани и костного мозга, матки, поджелудочной железы, пищевода, мочевого пузыря и почек вызывают значительное количество смертей и поэтому также представляют интерес.
Предпочтительно вакцины будут назначать в качестве профилактического лечения, но ввиду хронической природы этих заболеваний и их тенденции к ремиссии и рецидивам, их можно также вводить больным, у которых диагностировано одно или несколько вышеуказанных заболеваний, и они могут служить для поддержания больного в состоянии ремиссии.
На основании ранее проведенных исследований на мышах считается, что модифицированные аналоги ФНОα по изобретению могут также вводиться как часть лечебного воздействия на вышеуказанные заболевания в острой стадии или по меньшей мере с целью перевести больного в состояние ремиссии и поддерживать в стабильном состоянии.
В настоящее время не может быть определен специфический диапазон эффективных доз, так как вакцины еще не были испытаны на людях, восприимчивых к какой-либо из болезней.
В любом случае вводимая доза будет прописана ответственным врачом.
В соответствии с одним из примеров осуществления изобретения вакцина содержит смесь двух различным образом модифицированных молекул ФНОα, содержащих различные эпитопы для Т-клеток, например Р2 и Р30, которые получены из столбнячного анатоксина. Эта смесь по выбору содержит соответствующие количества фармацевтически приемлемого вещества, усиливающего иммуногенность.
В соответствии с еще одним аспектом изобретения вакцина включает не модифицированные молекулы ФНОα человека как таковые, но предпочтительную конструкцию, включающую неинфекционную, неинтегрированную последовательность ДНК, кодирующую вышеуказанные молекулы, оперативно связанную с промоторной последовательностью, которая может регулировать экспрессию вышеуказанной последовательности нуклеотидов ДНК у людей, в количестве, достаточном, чтобы происходил захват вышеуказанной конструкции, и происходила достаточная экспрессия, чтобы индуцировать реакцию нейтрализующих антител против ФНОα.
Полезность этого типа вакцин, так называемых ДНК-вакцин, проиллюстрирована, например, в патентах США № 5589466 и 5580859, которые оба включены в данное описание посредством ссылки на них, в частности, относительно методов введения.
ДНК-вакцины могут включать вирусный вектор экспрессии, такой как ретровирусный вектор экспрессии.
В большинстве случаев вакцины по изобретению могут быть адаптированы для орального или парентерального, в частности подкожного, внутримышечного или внутрикожного введения.
Изобретение далее включает применение антител, концентрация которых увеличена за счет введения вакцины по изобретению, предпочтительно моноклональных антител, в частности диагностическое применение.
Изобретение далее относится к способу тестирования жидкостей человеческого тела на присутствие ФНОα, который включает контакт состава, содержащего модифицированный ФНОα по изобретению, с пробой жидкости человеческого тела и определение того, связываются ли вышеуказанные антитела с ФНОα в вышеуказанной пробе.
Изобретение также относится к способу диагностики связанных с ФНОα заболеваний, использующему иммунологический анализ in vitro для обнаружения ФНОα в жидкостях человеческого тела.
Вышеуказанные способы могут включать использование пробы сандвича, иммуноферментного твердофазного анализа или эквивалентных анализов, которые могут быть неамплифицированными или амплифицированными, например, с использованием авидин/биотиновой технологии.
Вакцины/рекомбинантные белки могут быть охарактеризованы с использованием следующих анализов, хорошо знакомых специалистам в данной области техники:
SDS-Page гели, окрашенные кумасси или серебром (информация о размере и чистоте),
ИЭФ (изоэлектрическая фокусировка) (информация об изоэлектрической точке),
Анализ с эндотоксином LAL (информация о чистоте),
Белок клетки реципиента (информация о чистоте),
Масс-спектроскопия (информация о молекулярной массе),
SE-жидкостная хроматография высокого давления с профилем УФ-выявления (информация о распределении по молекулярной массе),
N-концевая последовательность (информация об идентичности),
Круговой дихроизм (информация о третичной структуре),
SE-жидкостная хроматография высокого давления с выявлением malls (информация о третичной структуре при помощи рассеяния света),
Аминокислотный состав (информация об идентичности),
Иммуногенность в гетерологичных видах (мышь, крыса или кролик),
Реактивность на антитела в Вестерн-блоттинге,
ЯМР-спектроскопия,
Кристаллическая структура,
Полное определение аминокислотной последовательности.
Сведения, подтверждающие возможность осуществления изобретения
1. Введение
В предыдущей патентной заявке WO 95/05849 было показано, что эпитопы для Т-клеток модифицированных молекул ФНОα мышевидных грызунов могут создавать высокие титры антител, перекрестно реагирующих с нативным (дикого типа) ФНОα мышевидных грызунов. Эти антитела были способны мешать взаимодействию ФНОα и его рецепторов in vitro и in vivo. Положительные эффекты иммунизации против ФНОα были продемонстрированы в нескольких сериях с моделированием на животных заболеваний, вызываемых ФНОα, таких как экспериментальная кахексия, коллагеновый артрит и экспериментальный аллергический энцефаломиелит (ЭАЭ).
Эти результаты экспериментов на животных были получены, несмотря на то, что две использованные модифицированные молекулы ФНОα мышей (названные MR 103 и MR 106) не были оптимизированы в отношении иммуногенности для ОКГ класса II гаплотипов мышей DBA/1 и SJL. Эти линии мышей были использованы для моделирования коллагенового артрита и экспериментов с ЭАЭ соответственно. В другом эксперименте было показано, что различным образом модифицированная молекула ФНОα мышевидных грызунов (MR 105) вызывала более сильный иммунный ответ, чем рекомбинантный ФНОα мышей, конъюгированный с белками Е. coli с использованием формальдегида.
MR 103, MR 105 и MR 106 были молекулами мышей, и на основании предыдущей патентной заявки WO 95/05849 нельзя было сделать специфических выводов ни относительно иммуногенности для соответствующих Т-клеток модифицированного ФНОα человека, ни о потенциальной способности таких молекул индуцировать образование нейтрализующих ФНОα антител, так как все они были активными.
2. Разработка конструкций, ФНОα человека
В общих чертах, в ФНОα-вакцине для использования человеком модифицированные молекулы ФНОα человека должны удовлетворять следующим требованиям:
А. Они должны быть иммуногенными для большой доли популяции.
Б. Они должны обладать оптимальной способностью индуцировать образование нейтрализующих ФНОα антител.
В. Они не должны сохранять какой-либо остаточной биологической активности ФНОα.
Более того, в процессе отбора могут быть учтены другие практические параметры, такие как уровни рекомбинантной экспрессии, легкость очистки, растворимость и т.д.
2.1. Иммунологическая неоднoродность модифицированных молекул ФНОα
Во время разработки вакцины против ФНОα человека целью было получение модифицированных молекул ФНОα человека, которые в конечном итоге будут иммуногенными для наибольшей возможной части популяции людей, которая несомненно представляет большое число различных типов HLA (ОКГ) II класса. Поэтому вместо ОКГ-специфичных эпитопов, использованных в предыдущих экспериментах на животных, были использованы неоднородные эпитопы для Т-клеток. Из предшествующей заявки WO 95/05849 было неизвестно, как такие эпитопы могут влиять на способность этих молекул индуцировать образование нейтрализующих антител.
Были выбраны два эпитопа для Т-клеток, Р2 и Р30, полученные из столбнячного анатоксина (СА), которые были хорошо охарактеризованы в научной литературе. Известно, что эти эпитопы в СА иммунодоминантны и способны связывать по меньшей мере 80% молекул ОКГ II класса в человеческой популяции.
Более того, при использовании этих СА-эпитопов ожидалось, что будет возможно протестировать иммуногенность ФНОα-конструкций in vitro на мононуклеарных клетках периферической крови (МКПК) и линиях Т-клеток, полученных от иммунизированных СА доноров крови.
Аминокислотная последовательность эпитопа Р2 выглядит, как QYIKANSKFIGITEL, и соответствует аминокислотам СА 830-844, а последовательность Р30 эпитопа выглядит, как FNNFTVSFWLRWPKVSASHLE, и соответствует аминокислотам СА 947-967. Замена на Р2 и Р30 в двух разных молекулах ФНОα приведет к изменению примерно 10% и 15% нативной последовательности ФНОα соответственно. В случае, если оба эпитопа встроены в одну молекулу ФНОα, будет изменено примерно 25% молекулы, и можно ожидать, что это будет оказывать слишком сильное неблагоприятное влияние на оставшиеся нативными части молекулы ФНОα. Поэтому было решено разработать две молекулы ФНОα, каждая из которых содержит либо Р2, либо Р30. Можно ожидать, что вместе эти две молекулы будут иммуногенными по меньшей мере для 80% человеческой популяции. Кроме того, очень вероятно, что усеченные молекулы, состоящие частично из эпитопов Р2 или Р30 и частично из фланговых областей ФНОα, также будут вносить вклад в иммуногенность, в результате чего конструкции будут иммуногенны практически в 100% популяции.
Хотя существовала возможность индуцировать образование антител всеми конструкциями ФНОα мыши во всех линиях мышей, тестированных до сих пор (см. обсуждение MR 103, 105 и 106 выше), априорно можно было ожидать, что встраивание чужеродного эпитопа для Т клетки в определенные положения в ФНОα, будет более полезным, чем в другие положения, с точки зрения представления эпитопа Т-клеткам молекулами ОКГ II класса. Поэтому было решено получить набор различным образом модифицированных молекул ФНОα человека с Р2 и Р30 эпитопами, встроенными в молекулу в разных положениях, см. фиг.4. Впоследствии все молекулы были тестированы in vitro в анализах на основе мононуклеарных клеток периферической крови (МКПК) или Р2/Р30-специфичных линий Т-клеток, выделенных у многих здоровых иммунизированных СА доноров крови.
Однако вопреки ожидавшемуся было показано, что хотя наблюдались небольшие количественные различия, внутримолекулярное положение Р2 и Р30 эпитопов было несущественным для способности обрабатываться антигенпредставляющими клетками и впоследствии быть представленными СА-специфичным Т-клеткам, Так, Р2, встроенный в положения 132-146, 65-79 и 76-90, и Р30, встроенный в положения 40-60 и 132-152 (ФНO 2-5, 2-3, 2-7, 30-2, 30-5), были обработаны и представлены Т-клеткам. Таким образом, судя по вышеприведенному обсуждению, очень вероятно, что эти молекулы в конечном итоге будут универсально иммуногенными в человеческой популяции.
2.2. Способность модифицированных молекул ФНОα человека индуцировать образование нейтрализующих антител
Как указано выше, из предшествовавших исследований на мышах, описанных в WO 95/05849, было невозможно предсказать, какое положение будет наиболее подходящим для того, чтобы обеспечить способность индуцировать образование нейтрализующих ФНОα антител, так как все три аналога MR 103, MR 105 и MR 106 были способны индуцировать образование антител, несмотря на различные зоны замещения. Поэтому был создан набор различных молекул ФНОα человека с Р2 или Р30, встроенными в различных положениях. Замещения были случайным образом распределены по всей молекуле. Антитела, образовавшиеся у кроликов после инъекции этих молекул, впоследствии тестировали в биохимических, и биологических анализах in vitro на их способность нарушать биологическую активность ФНОα.
В неопубликованных наблюдениях авторов настоящего изобретения было показано, что в зависимости от внутримолекулярного положения встроенного эпитопа наблюдалась различная общая специфичность образовавшихся антител. Вопреки тому, что ожидалось, исходя из структурных данных о молекуле ФНОα, наблюдалось, что замещения, произведенные в переднем β-слое с использованием либо Р2, либо Р30, полностью лишали молекулы биологической активности ФНОα, но в то же время сохраняли способность модифицированных молекул индуцировать образование нейтрализующих ФНОα антител.
Было показано, что молекулы, содержащие Р2 или Р30 в положениях, указанных выше, особенно эффективны в отношении индукции образования нейтрализующих антител. Любые из этих молекул являются поэтому потенциальными кандидатами для использования в вакцинах против ФНОα человека.
2.3. Биологическая активность различных ФНОα-конструкций
Понятно, что невозможно использовать в вакцине такую токсичную молекулу, как ФНОα. Поэтому модифицированные молекулы ФНОα должны быть нетоксичными, то есть лишенными какой бы то ни было остаточной активности ФНОα.
Все мутантные ФНОα-белки поэтому были тестированы in vitro в ФНОα-зависимых биопробах, а также пробах на связывание с рецепторами, чтобы проверить, являются ли они нетоксичными. Было ясно показано, что все модифицированные молекулы ФНОα человека (ФНO 2-5, 2-3, 2-7, 30-2 и 30-5) были лишены биологической активности ФНОα. Поэтому были удовлетворены все требования к этим молекулам, необходимые для того, чтобы они были частью универсальной, нетоксичной вакцины, способной индуцировать образование антител, нейтрализующих ФНОα человека.
ПРИМЕР 1
Работа по созданию генетических конструкций
Было решено получить 10 различных модифицированных молекул ФНОα человека - пять, содержащих Р2, и пять, содержащих Р30 эпитоп. Эпитопы были распределены внутри молекулы в различных положениях. Генетические конструкции были созданы с использованием различных стандартных рестрицирующих ферментов и методик мутагенеза на основе ПЦР. Генетические конструкции схематически изображены на фиг.4а и 4b. Нуклеотидные последовательности ДНК, кодирующие модифицированные молекулы ФНОα, и соответствующие аминокислотные последовательности включены как SEQ ID NO:1 - SEQ ID NO:20. Конструкции мутантного гена ФНO2-5, стратегия клонирования и мутаций и последующие экспрессия, выделение и очистка аналога ФНO2-5 объяснены ниже на примере.
Конструирование и получение ФНO2-5:
Генетическое конструирование мутантного гена ФНO2-5 человека, стратегия клонирования и мутаций
Генетическое конструирование гена, кодирующего мутантный аналог ФНO2-5 человека, было основано на традиционных методиках мутагенеза на основе ПЦР, как и для всех прочих генетических конструкций.
Нативная последовательность ДНК ФНОα человека, кодирующая растворимую часть этой молекулы, была получена традиционным ПЦР-клонированием с использованием синтетически синтезированных праймеров I и II (таблица 1 и SEQ ID NO:21 и 22) из имеющейся в продаже библиотеки кДНК человека, CLONTECH Laboratories, США (фиг.16, 1). Нативный ген был встроен в коммерческий вектор экспрессии Е.coli pET28c, который можно приобрести в фирме Novagen, США, таким образом, что ген можно было транскрибировать in frame от индуцируемого ИПТГ промотора.
Генетическое конструирование мутантного аналога ФНОα ФНO2-5 было выполнено при помощи методики ПЦР-мутагенеза, примененной к нативной последовательности ДНК. Нуклеотидная последовательность нуклеиновой кислоты, кодирующая эпитоп для Т-клетки, была включена в искусственно синтезированный 75-мерный олигонуклеотид (Праймер “mut2-5”, таблица 1 и SEQ ID NO:27) между двумя 3′- и 5′-отожженными участками гомологичной ФНО ДНК, поэтому “mut”-праймер способен отжигать последовательность нативного гена ФНОα человека в определенном сайте, выбранном для ФНO2-5 (см. фиг.16, 2а). В “mut”-олигонуклеотиде число кодонов, кодирующих эпитоп для Т-клетки, точно соответствует числу ФНО-кодонов, пропущенных между двумя 3′- и 5′-отожженными участками гомологичной ФНО ДНК. Праймер мутагенеза был использован для получения продукта ПЦР, содержащего ДНК, кодирующую эпитоп для Т-клетки и последовательность ФНОα ниже (или 3′) встроенного эпитопа (фиг.16, 2а).
Промежуток ФНОα ДНК выше (или 5′) точки внедрения эпитопа был обеспечен при помощи второго продукта ПЦР с использованием пpaймepов I и II (таблица 1, SEQ ID NО:23) (фиг.16, 26). Два продукта ПЦР в конечном итоге соединяли вместе в конечной реакции ПЦР (фиг.16, 3) с использованием двух наиболее дистальных праймеров (I, II) из двух реакций. Полную мутантную последовательность ДНК ФНО2-5 затем вводили в коммерческий вектор экспрессии Е.coli по аналогии с экспрессионным клонированном нативного гена таким образом, что ген можно было транскрибировать от ИПТГ индуцируемого промотора из трансформированных клеток.
“Мut”-праймеры, использованные для конструирования других аналогов (ФНO 2-1, 2-3, 2-4, 2-7, 30-1, 30-2, 30-3, 30-4 и 30-5), идентифицированы как SEQ ID NO:23-26 и 28-33 соответственно.

Культивация рекомбинантных бактерий, накопление и растворение инклюзионных тел.
Очистка белка ФНO2-5 аналога
Получение белка ФНO2-5 было аналогичным получению других рекомбинантных молекул ФНО
1. Инокулировать 20 мл ТВ-среды, содержащей 50 мкг/мл Карбенициллина, трансформированной линией Е.coli, несущей индуцируемый ИПТГ плазмидный вектор, включающий ген ФНО, кодирующий рекомбинантный протеин, выращивать Е.coli в течение ночи при 37°С при встряхивании.
2. Развести ночную культуру в соотношении 1:25 в 250 мл ТВ-среды с 50 мкг/мл Карбенициллина и выращивать культуру, пока OD450 не будет равно 1. Индуцировать экспрессию рекомбинантного белка путем добавления ИПТГ до конечной концентрации 1 мМ. Растить в течение ночи при энергичном встряхивании при 37°С.
3. Собрать рекомбинантные клетки со среды путем центрифугирования при 3500×g. Промыть гранулу один раз в BSB буфере. Использовать 150 мл БСБ на 50 г влажного веса бактерий.
4. Обработать ультразвуком 4 раза по 30 секунд при максимальной амплитуде до тех пор, пока бактериальная суспензия не станет абсолютно гомогенной. Обработку ультразвуком проводят с использованием MSE Soniprep 150 ультразвукового излучателя, оборудованного стандартной головкой размером 9,5 мм (Soniprep 05 38 121-1154).
5. Добавить 8 мкл ФМСФ (50 мМ) и 80 мкл раствора лизоцима (10 мг/мл) на грамм бактериальной массы. Инкубировать 30 мин при комнатной температуре.
6. Добавить 4 мг дезоксихолевой кислоты на грамм бактериальной массы, перемешать и хранить при 37°С.
7. Когда раствор станет вязким, добавить 20 мкл ДНКазы (1 мг/мл) на грамм бактериальной массы и МgСl до конечной концентрации 5 мМ, перемешать и хранить при комнатной температуре в течение 30 мин.
8. Обработать ультразвуком во льду 5 раз по 30 секунд с 30-секундными интервалами при максимальной амплитуде до тех пор, пока раствор не станет жидким и невязким.
9. Центрифугировать при 20×g в течение одного часа, сохранить супернатант для позднейшего контроля процедуры промывания, чтобы проверить, все ли инклюзионные тела были преципитированы.
10. Повторно суспензировать центрифугат в воде MiliQ (1 мл Н2O на грамм Е.coli), встряхивать в течение 1 часа.
11. Центрифугировать при 20×g в течение одного часа, сохранить супернатант, чтобы проверить, все ли инклюзионные тела были преципитированы.
12. Повторно суспензировать гранулу в 1 М растворе мочевины, добавляя 1 мл на грамм Е.coli, встряхивать в течение 1 часа.
13. Центрифугировать при 20×g в течение 1 часа, сохранить супернатант, чтобы проверить, все ли инклюзионные тела были преципитированы.
14. Повторно суспензировать гранулу в 1 М гуанидине, добавляя по 1 мл на грамм Е.coli, встряхивать в течение 1 часа.
15. Центрифугировать при 20×g в течение 1 часа, сохранить супернатант, чтобы проверить, все ли инклюзионные тела были преципитированы.
16. Повторно суспензировать гранулу в 25 мл 6 М ганидина+20 мМ Трис с рН 8,0, взбалтывать в течение ночи.
17. Центрифугировать при 20×g в течение 1 часа, сохранить супернатант, содержащий инклюзионные тела рекомбинантного белка, сохранить гранулу, чтобы проверить, все ли инклюзионные тела были растворены.
18. Белковый раствор подвергают интенсивному диализу против воды MiiliQ, а затем раствор высушивают при температуре ниже 0°С.
19. Высушенный при температуре ниже 0°С материал растворяют в 20 мМ Трисе, 6 М гуанидине, 30% 2-пропаноле (рН 8.0) в концентрации 20 мг/мл. При осторожном взбалтывании оставляют растворяться на ночь. Присутствие мономеров исследуют на колонке superdex 2000 (ХК 16, Pharmacia, диаметр: 1,6 см, высота: 750 см). Пробег в текущем буфере при 1 мл/мин. Сравнить со стандартами в том же буфере.
20. Очистку белка выполняют при помощи гель-фильтрации на колонке superdex 200 (ХК 26, Pharmacia, высота: 100 см, диаметр: 2,6 см), которая уравновешена с равновесным буфером. Пробег в уравновешивающем буфере. Используют объем пробы, равный примерно 1% от общего объема колонки.
21. Повторное складывание рекомбинантного белка выполняют путем диализа. Белок растворяют в концентрации 0,1 мг/мл в уравновешивающем буфере, и этот раствор помещают в прокипяченный мешок для диализа и диализируют против: 20 мМ Триса, 4М мочевины (рН 8.5), при трех сменах раствора, в течение ночи при комнатной температуре. Диализный мешок переносят в Трис-буфер (20 мМ Триса, 150 мМ NaCl (рН 8.0)). Сменить раствор три раза, из которых первая смена происходит при комнатной температуре. В течение ночи в холодной комнате.
Повторное складывание оценивают на колонке Superrose 12, уравновешенной с Трис-буфером (20 мМ Триса, 150 мМ NaCl (рН 8.0)). Сравнить со стандартами.
Хранение. Рекомбинантные белки хранят в высушенном при температуре ниже 0°С состоянии.
Крупномасштабное производство модифицированных молекул ФНОα может быть выполнено, как описано ниже:
Исходный материал:
500 литров ферментированной культуры Е.coli, диафильтрованной с водой до объема примерно 50 литров.
1) Промывание клеток
A) Оттаять замороженные клетки до жидкого состояния.
B) Центрифугировать клетки в течение 10 минут при 4000×g.
C) Повторно ресуспензировать гранулу в 50 мМ Трисе, 150 мМ NaCl, pH 8,0. Повторить стадии В) и С) три раза.
2) Гомогенизация клеток
Гомогенизировать клетки гомогенизатором Rannie, 5 циклов при 700 бар.
Б) Центрифугировать инклюзионные тела в течение 30 минут при 16.500* g.
B) Промыть инклюзионные тела три раза раствором, содержащим 3 М гуанидин, 1 М NaCl, 5 мМ ЭДТА, 20% сахарозы, 50 мМ триса, pH 8. Центрифугировать инклюзионные тела в течение 30 минут при 16,500* g.
3) Растворение инклюзионных тел
Ресуспензировать гранулу в 6 М гуанидине, 10 мМ ДТТ, 150 мМ NaCl, 5% этаноле, 50 мМ Трисе, pH 8 Использовать примерно 1 мл буфера/100 мг гранулы.
4) Ультрафильтрация инклюзионных тел
Удалить грубые загрязнения при помощи 0,45 мкм фильтра.
Б) Удалить высокомолекулярные компоненты фильтром на 30 КД.
B) Концентрировать инклюзионные тела фильтром на 5 КД.
5) Замена буфера
Подготовить пробу для ионообменной хроматографии. Заменить буфер на 6 М мочевину, 1 мМ ДТТ в 50 мМ Триса, pH 8 путем диафильтрации.
6) Очистка с помощью SP-Sepharose
Загрузить белком колонку с SP-Sepharose. Промыть колонку четырьмя объемами А-буфера, элюировать белок 100% Б-буфером и объединить все фракции с ФНОα.
А-буфер: 6 М мочевина, 1 мМ ДТТ, 50 мМ Трис/Cl, рН 8.
Б-буфер: 6 М мочевина, 1 М NaCl, 1 мМ ДТТ, 50 мМ Трис/Cl, рН 8.
7) Повторное складывание ФНОα человека
Развести протеиновый пул до концентрации примерно 0,1 мг ФНОα/мл в 6 М мочевине, 1 мМ ДТТ, 50 мМ NaCl, 5% этиловом спирте в 20 мМ Трис/Cl, рН 8,8, и поэтапно удалить мочевину, переходя к следующему составу буфера - путем диафильтрации.
A) 2 М мочевина, 1 мМ ДТТ, 150 мМ NaCl в 20 мМ Трис/Cl, рН 8,8 - в течение ночи при 5°С.
Б) 1 М мочевина, 1 мМ ДТТ, 150 мМ NaCl в 20 мМ Трис/Cl, рН 8,8 - в течение 8 часов при 5°С.
B) 1 мМ ДТТ, 150 мМ NaCl в 20 мМ Трис/Cl, рН 8,8 - в течение ночи при 5°С.
Г) 150 мМ NaCl в 20 мМ Трис/Cl, рН 8,8 - в течение ночи при 5°С.
8) Хранение аналогов ФНОα человека
Концентрировать белок до 1.0 мг/мл и хранить пробу при -20°С.
ТВ-среда
Растворить бульон Terrific Broth (GIBCO BRL 22711-22) в воде MilliQ в соответствии с инструкциями производителя. Автоклавировать при 121°С в течение 20 мин.
Маточный раствор карбенициллина х 100 (50 мг/мл)
Двойную натриевую соль карбенициллина (Sigma С 1389) растворяли в воде MilliQ в концентрации 50 мг/мл. Раствор стерилизовали фильтрованием через фильтр 0,2 мкм (Sterifix 0409 9206).
ИПТГ х 100 маточный раствор 100 мМ
Изопропил-бета-D-тиогалактопиранозид (ИПТГ, USB 17884). 1,19 г ИПТГ растворяли в воде MilliQ до 50 мл. Раствор стерилизовали фильтрованием через фильтр 0,2 мкм (Sterifix 0409 9206).
Буфер БСБ
Буфер для Бактериальных Суспензий
50 мМ ТРИС (Trisma base, SigmaT1503)
0,5 M NaCl (Ridel-de.Haen 31434)
5 мМ ДТТ (DL-дитиотретиол, Sigma D-0632)
рН 8,0
ФМСФ
50 мМ, фенилметилсульфонилфторид, SIGMA # P-7626, растворенный в 2-пропаноле.
Раствор лизоцима
10 мг/мл Качества III лизоцима из белка куриного яйца, (ЕС 3.2.1.17) SIGMA # L-7001
Дезоксихолевая кислота
(7-дезоксихолевая кислота) Sigma # D-6750
ДНКаза
1 мг/мл ДНКазы I, дезоксирибонуклеазы I, (ЕС. 3.1.21.1) Boehringer Cat # 1284932
МОЧЕВИНА
Мочевина (GibcoBRL 15716-020)
ГУАНИДИН
Гуанидина гидрохлорид (Sigma G4505)
2-Пропанол
Проточный буфер
20 мМ ТРИС, (Trisma base, SigmaT1503)
8 М мочевины, (GibcoBRL 15716-020)
0,1% β-меркаптоэтанола
рН 8,0
Уравновешивающий буфер
20 мМ ТРИС, (Trisma base, SigmaT1503)
8 М мочевины, (GibcoBRL 15716-020)
0,1% β-меркаптоэтанола
ПРИМЕР 2
Экспрессия, очистка и повторное складывание Р2 и Р30 модифицированных молекул ФНОα человека
Надежно определено, что рекомбинантные белки по-разному ведут себя во время экспрессии, очистки и повторного складывания. Все белки экспрессировали в Е.coli, и были получены уровни, варьировавшие от 2 до 20%. Все белки распознавали в экспериментах с Вестерн-блоттингом с использованием коммерческого поликлонального антитела кролика к ФНОα человека.
Конструкции ФНОα впоследствии экспрессировали по отдельности в 250 мл культуры в партиях объемом в 3-4 л. Все модифицированные ФНОα белки были экспрессированы в качестве инклюзионных тел, и белковые препараты с чистотой более 85% и повторно сложенные были получены так, как описано выше.
Содержание белка было определено с использованием стандартного ВСА-анализа. Все очистки белков были выполнены по меньшей мере при трех раздельных пробегах для каждой молекулы, и во всех случаях были тестированы раздельные партии белка и обнаружено сходство. Белки хранили в концентрациях 0,2-1,0 мг/мл в PBS при -20°С до использования.
Стандартные анализы для контроля качества включали SDS гель-электрофорез с окрашиванием отдельных белковых препаратов как Кумасси синим, так и серебром. Во всех случаях было обнаружено, что белки имеют ожидавшийся размер и уровень чистоты, превышающий 85%.
Молекулы с эпитопами, встроенными в положение 4 (ФНO2-4 и ФНО30-4), были экспрессированы только с относительно низкими уровнями (примерно 2%). Более того, эти молекулы было очень трудно очистить, а особенно трудным было их повторное складывание. Обе молекулы, особенно ФНО30-4, были не очень растворимы в PBS и имели тенденцию преципитировать во время хранения.
ПРИМЕР 3.
Биологическая активность различных ФНОα конструкций
Прямая биологическая активность очищенных молекул ФНОα была тестирована в биоанализе L929, который является стандартным анализом in vitro для определения биологической активности ФНОα. Как видно из фиг.6, ни одна из Р2 или Р30 модифицированных молекул ФНОα не была способна убивать клетки L929 в исследованном диапазоне концентраций (до 60 мг/мл). Имеющиеся в продаже рекомбинантные молекулы ФНОα сохраняли полную активность при концентрации примерно 0,6 нг/мл. Препарат ФНОα дикого типа также был полностью активен во всем исследованном диапазоне концентраций.
ПРИМЕР 4.
Способность модифицированных молекул ФНОα индуцировать образование нейтрализующих антител
Кролики были иммунизированы каждой из десяти конструкций, чтобы посмотреть, могут ли они индуцировать образование антител, способных нейтрализовать биологически активный ФНОα человека in vitro. Были использованы группы из трех кроликов, и каждый кролик получил 100 мг ФНОα подкожно в адъюванте полного состава Freunds Complete Adjuvant и впоследствии по 100 мг примерно каждую третью неделю в адъюванте неполного состава Freunds Incomplete Adjuvant.
В течение всего периода иммунизации, равного 18 неделям, через регулярные промежутки времени брали пробы крови, и сыворотки исследовали на анти-ФНОα активность традиционным анализом ELISA (иммуноферментным твердофазным анализом), где в качестве антигена был использован имеющийся в продаже, чистый и немодифицированный ФНОα человека.
У всех кроликов в течение периода иммунизации образовались антитела к ФНОα, и максимальный уровень или титры антител варьировали в пределах десятичного разряда в каждой группе. Средние титры антител показаны на фиг.7. ФНO2-5, ФНO2-7, ФНO2-3, ФНO2-1 и ФНО дикого типа индуцировали титр антител, равный ста, в течение 2-4 недель, тогда как ФНO2-4 дал заметно более медленную реакцию. Сходная кинетика наблюдалась для белков ФНО30, где более медленный ответ также был получен для ФНО30-4.
Способность этих сывороток нарушать взаимодействие между ФНОα человека и его рецептором была проверена в L929 анализе, а также в анализе с твердофазным рецепторным связыванием.
В анализе L929 разведенные иммуннные сыворотки и неиммунную контрольную сыворотку добавляли к имеющемуся в продаже ФНОα человека перед добавлением смеси L929 клеток. Сыворотку от всех трех кроликов в каждой группе тестировали дважды и рассчитывали средние значения. Так как нормальная сыворотка проявляла тенденцию повышать исходные уровни цветоопределяющей системы, их вычитали из всех значений, и рассчитывали относительное ингибирование в процентах. Результаты ингибирующей способности на 14 неделе программы иммунизации по всем кроликам показаны на фиг.8.
ФНO2-5, который не содержал замен в каких-либо сегментах заднего β-слоя, явно превосходит другие конструкции, и эта молекула была сопоставима с полностью токсичной молекулой ФНОα дикого типа в отношении способности индуцировать образование нейтрализующих антител (данные не показаны). ФНO2-3, который содержит небольшой замещенный сегмент в β-цепи D, и ФНO2-7, который не содержит каких-либо замененных сегментов в цепях заднего β-слоя, также были способны индуцировать образование ингибирующих антител, и титр нейтрализующих антител к ФНO2-7 достоверно возрастал в течение трех последующих недель (данные не приведены). ФНO2-4 и ФНO2-1, которые содержали G и В-цепи соответственно в заднем β-слое, были неспособны индуцировать образование нейтрализующих антител в анализе L929.
Результаты относительно ФНО30 белков были более неоднородными, хотя ФНО30-3 выглядел наилучшим на 14 неделе. ФНО30-1 и ФНО30-4, которые содержат замещения в G и В β-цепях заднего р-слоя соответственно, не индуцировали образования нейтрализующих антител в анализе L929.
В анализе с твердофазным рецепторным связыванием рекомбинантный человеческий ФНОα-рецептор 1 с молекулярной массой 55 кДа (ФНО-R55) был иммобилизован на пластинках для микротитрования, а имеющийся в продаже биотинированный ФНОα человека добавляли в соответствующем разведении. Затем определяли специфическое связывание с рецептором при помощи стрептавидина, меченного пероксидазой хрена, и хромогенного субстрата. При тестировании сыворотки от иммунизированных кроликов ее добавляли к раствору биотинированного ФНОα человека перед добавлением смеси на пластинки для микротитрования, покрытые ФНО-R55. Исследовали сыворотки от всех трех кроликов в каждой группе и рассчитывали средние значения. Сыворотку от неиммунизированного кролика использовали в качестве отрицательного контроля. Исходные значения были очень низкими, и анализ был очень чувствительным. Результаты показаны на фиг.9.
Можно видеть, что результаты, полученные при L929 анализе и анализе с твердофазным рецепторным связыванием соответственно, являются почти идентичными в отношении конструкций ФНОα 2. В твердофазном анализе разница между ФНО30-2, 30-3 и 30-5 была не столь выражена, как в анализе L929. Твердофазный анализ, однако, более воспроизводим из-за своего скорее биохимического, нежели клеточного характера, и значения для нормальной сыворотки в этом анализе не вычитались.
Относительные количества сыворотки (в процентах), при которых было получено половинное от максимального уровня ингибирование связывания ФНОα (значения IC50), были рассчитаны для ФНO2-3, ФНO2-5, ФНO2-7, ФНО30-2, ФНО30-3, ФНО30-5 и ФНОα дикого типа для каждой соответствующей антисыворотки. Принимая, что сходная форма кривой будет получена для антисывороток против ФНO2-1, ФНO2-4, ФНО30-1 и ФНО30-4, были выполнены экстраполяции, и значения IС50 были также рассчитаны для этих сывороток. Результаты показаны в таблице 2.

Можно видеть, что ФНO2-5 эквивалентен дикому типу (ДТ) ФНОα мыши в отношении способности индуцировать образование антител к ФНОα мыши у кроликов. ФНO2-1, ФНO2-4, ФНО30-1 и ФНО30-4, которые все содержат замены в В- или G-цепях заднего β-слоя, очень слабо способны или вообще не способны индуцировать образование таких антител. Это неожиданно показывает, что хотя цепи В и G заднего β-слоя не участвуют в связывании с рецептором, мутации, представленные в этой области, каким-то образом приводят к повреждению областей ФНОα человека, участвующих в связывании с рецептором ФНО30-2 и ФНО30-3, по-видимому одинаково хорошо индуцируют образование нейтрализующих антител.
ПРИМЕР 5.
Способность модифицированных молекул ФНОα стимулировать Т-клетки иммунизированных Р2 и Р30 здоровых доноров кpови
Так как ожидалось, что локализация Р2 и Р30 эпитопов в модифицированных молекулах ФНОα также будет влиять на переработку антигена и представление встроенных эпитопов - и вследствие этого на иммуногенность - все 10 молекул были тестированы в различных анализах с Т-клетками. Был использован поликлональный Молекулярный Анализ Клеток Периферической Крови (РВМС), и число Р2- и Р30-специфичных линий Т-клеток было определено с использованием обычных иммунологических методов, чтобы выявить ФНОα пептиды и рекомбинантные белки с встроенными Р2 и Р30 эпитопами.
Вначале 28 здоровых добровольцев (доноров) были тестированы в обычном пролиферационном РВМС-анализе на их способность реагировать на СА и пептиды Р2 и Р30. На основании этих результатов 19 лиц, способных достоверно реагировать на СА, Р2 и Р30 пептиды, были отобраны для дальнейших экспериментов. Хотя уровни реакций сильно варьировали, это ясно подтвердило, что Р2 и Р30 являются различными и иммунодоминантными эпитопами для Т-клеток.
Кроме 28 доноров были дополнительно отобраны еще 7 доноров. По причинам, объясненным ниже, некоторые из них были отобраны за их способность реагировать только на Р2 или на Р30. Некоторые из них были к тому же вакцинированы СА, чтобы они были способны давать сильную реакцию Т-клеток. Некоторые из второй группы доноров также были использованы для накопления Р2- или Р30 - специфичных линий Т-клеток, чтобы исследовать представление Р2 и Р30 модифицированных синтетических ФНОα пептидов и модифицированных молекул в качестве антигенов. На фиг.10 показаны репрезентативные примеры поликлональной пролиферативной РВМС реакции у трех доноров на СА, а также пептиды Р2 и Р30.
Все 10 рекомбинантных белков были исследованы по три раза, по меньшей мере в 6 различных концентрациях, начиная с примерно 100 мг/мл, и все РВМС эксперименты повторяли 2-3 раза с каждым индивидуумом. Внутриклеточное включение 3H-меченного тимидина было использовано для оценки пролиферации Т-клеток, и результаты экспериментов были получены и подсчитаны в 96-ячеечном формате. Максимальные индексы пролиферации (ИП) из каждой тирационной кривой были рассчитаны как отношение среднего числа СРМ в экспериментальных ячейках к среднему числу СРМ, полученному для ячеек без антигена (только PBS). Данные о пролиферации Т-клеток были повторены три раза, a Con А, а также Р2 и Р30 были использованы в качестве отрицательного и положительного контролей соответственно.
На фиг.11 показаны три примера из экспериментов с двумя разными донорами крови с использованием различных Р2 и Р30-модифицированных молекул ФНОα.
JH реагирует только на Р2, тогда как SR реагирует как на Р2, так и на Р30. Это также отражалось в пролиферативных реакциях на Р2- и Р30-модифицированные ФНОα белки, которые в разной степени все были способны стимулировать РВМС.
На фиг.12 показаны Индексы Пролиферации (ИП), рассчитанные по 34 экспериментам. Хотя очень трудно количественно сравнивать ИП из разных экспериментов из-за варьирования исходных значений PBS, становится ясно, что все конструкции в большей или меньшей степени способны вызывать пролиферативную реакцию.
Среди второй группы доноров некоторые индивидуумы были вакцинированы СА за 1-2 месяца до экспериментов. Все ИП, полученные у этих лиц, находились среди наивысших значений ИП, наблюдавшихся для всех модифицированных ФНОα конструкций, и результаты было легко оценивать. Три других индивидуума (DL, ID и LS) были вакцинированы более 5 лет тому назад, и все они дали уровни ИП ниже среднего. Это подтверждает, что полученные ИП были антиген-специфичными. Для первой группы доноров не было данных относительно сроков последней вакцинации СА, и их иммунизационный статус был явно очень разнообразным.
Невозможно выбрать предпочтительный ФНОα 2 или ФНОα 30 белок на основании только среднего уровня значений ИП. Он может быть обусловлен различным состоянием вакцинации исследованных индивидуумов и различной природой РВМС-анализов, полученных от лиц с различным статусом реактивности. В любом случае было неожиданно, что Р2 и Р30, встроенные в ФНОα в любых положениях, по-видимому, способны перерабатываться и представляться Т-клеткам. Чтобы исключить возможность того, что, несмотря на повторную аффинную хроматографию, гель-фильтрацию и диализ, препараты антигенов все еще могли содержать значительные концентрации неспецифических митогенов, были выполнены дальнейшие эксперименты.
Доноры из второй группы, про которых было известно, что они не реагируют на Р2 и реагируют на Р30, а также доноры с противоположным паттерном реакции были тестированы в РВМС-анализах. На фиг. 13 реакции на Р2, Р30 и белки ФНОα 2 и ФНОα 30 показаны для DL и ID.
Можно видеть, что специфические реакции были получены на соответствующие эпитопы для Т-клеток и соответственно модифицированные молекулы ФНОα. Однако не наблюдалось достоверных пролиферативных реакций у DL на белки ФНОα 30 (верхняя полоса) и достоверных пролиферативных реакций у ID на белки ФНОα 2 (нижняя полоса), что подтверждает, что в очищенных препаратах ФНОα отсутствуют неспецифические митогены.
Эта возможность была исследована далее с использованием Р2 и Р30-специфичных линий Т-клеток, выделенных из второй группы доноров, которые культивировали в течение по меньшей мере шести недель с по меньшей мере тремя раундами стимуляции соответствующими синтетическими Р2 и Р30 пептидами. На фиг.14 показаны результаты двух таких экспериментов.
Можно видеть, что Р2- и Р30-специфичные линии Т-клеток стимулировались только соответствующими Р2 и Р30 белками. Более того, можно опять видеть, что все ФНОα конструкции были способны индуцировать пролиферацию Т-клеток. Это указывает на то, что хотя переработка антигена может быть количественно важной для представления Р2 и Р30, она не представляется существенным качественным лимитирующим фактором для представления антигена.
В литературе сообщалось, что фланговые области эпитопов для Т-клеток могут влиять на связывание антигенных пептидов с молекулами МНС класса II. Так как ни одно из положений в ФНОα, которые были выбраны для встраивания Р2 или Р30, по-видимому, не является запрещающим представление антигена, было исследовано, могут ли различные фланговые по отношению к встроенным эпитопам последовательности ФНОα влиять на реакцию Т-клеток на Р2 и Р30 эпитопы в результате различного связывания с молекулами HLA класса II человека. Поэтому были синтезированы пептиды, показанные в таблице 3, которые представляют встроенные эпитопы и фланговые аминокислоты ФНОα человека. Они обозначены как РР2-5, РР30-3 и т.д. Аминокислотные последовательности показаны в списке последовательностей как последовательности от SEQ ID NO:34 до SEQ ID NO:42, обозначенные от Рер2-1 до Рер30-5.

Эти пептиды были использованы для стимуляции Р2- и Р30-специфичных линий Т-клеток. Результаты стимуляции Р30-линий показаны на фиг.15. Можно видеть, что Р2-специфичные линии Т-клеток от MR и KG обнаруживают параллельные паттерны стимуляции, будучи стимулированы либо пептидом, либо соответствующим ФНОα-белком. Ясно, что ФНO2-5 является хорошим потенциальным антигеном, и эти данные дополнительно подтверждают, что наблюдавшаяся пролиферация Т-клеток является антиген-специфичной. В целом не было качественных различий в паттерне стимуляции, если Р30-специфичные линии Т-клеток от MR и KG стимулировали пептидами, или белками (данные не приведены). Р30-специфичная линия Т-клеток от НС предпочтительно распознавала ФНО30-3 и в меньшей степени реагировала с ФНО30-2.
ЗАКЛЮЧЕНИЕ
Было создано и охарактеризовано десять различным образом модифицированных белков ФНОα человека. Они были сконструированы так, чтобы включать два хорошо известных различных Р2 и Р30 эпитопа для Т-клеток, чтобы быть потенциально иммуногенными по меньшей мере для 85% населения.
Все белки можно было экспрессировать и очистить, хотя ФНO2-4 и ФНО30-4 с низкими уровнями. Мутации в этом положении в ФНОα также, по-видимому, нарушают повторное складывание, что приводит к белкам с низкой растворимостью. Ни у одной модифицированной молекулы ФНОα невозможно было обнаружить биологической активности ФНОα.
Кроликов иммунизировали всеми десятью белками и нативным ФНОα. Через 2-3 месяца после иммунизации во всех сыворотках было возможно обнаружить высокие титры сильно перекрестно-реактивных антител к немодифицированному ФНОα человека.
Способность этих антител нарушать биологическую активность нативного ФНОα была исследована в двух различных анализах In vitro - биологическая проба L929 и твердофазное связывание с рецептором. Оба анализа показали в общих чертах одно и то же. ФНO2-1, ФНO2-4, ФНО30-1 и ФНО30-4 не были способны индуцировать образование значительного количества нейтрализующих антител, тогда как ФНО2-5 превосходил все остальные конструкции. ФНО30-2 и ФНО30-3 были примерно одинаково эффективны в отношении индукции образования нейтрализующих антител и примерно в два раза лучше ФНО30-5, который был достаточно хорошим.
Несколько неожиданно оказалось невозможным обнаружить достоверные различия между способностями молекул стимулировать пролиферацию РВМС. Можно было продемонстрировать, что это не было обусловлено присутствием митогенов в препаратах антигенов, и на основании этих данных был сделан вывод, что все положения, выбранные для Р2 и Р30, позволяют представление соответствующих эпитопов. Специфичность реакции далее была документирована при использовании тестирования модифицированных ФНОα белков с использованием эпитоп-специфичных линий Т-клеток, а также синтетических пептидов, содержащих встроенные эпитопы, а также фланговые последовательности ФНОα. В этих экспериментах было ясно продемонстрировано, что ФНO2-5, ФНО30-2 и ФНО30-3 (среди прочих конструкций) были наиболее сильными потенциальными иммуногенами.




































































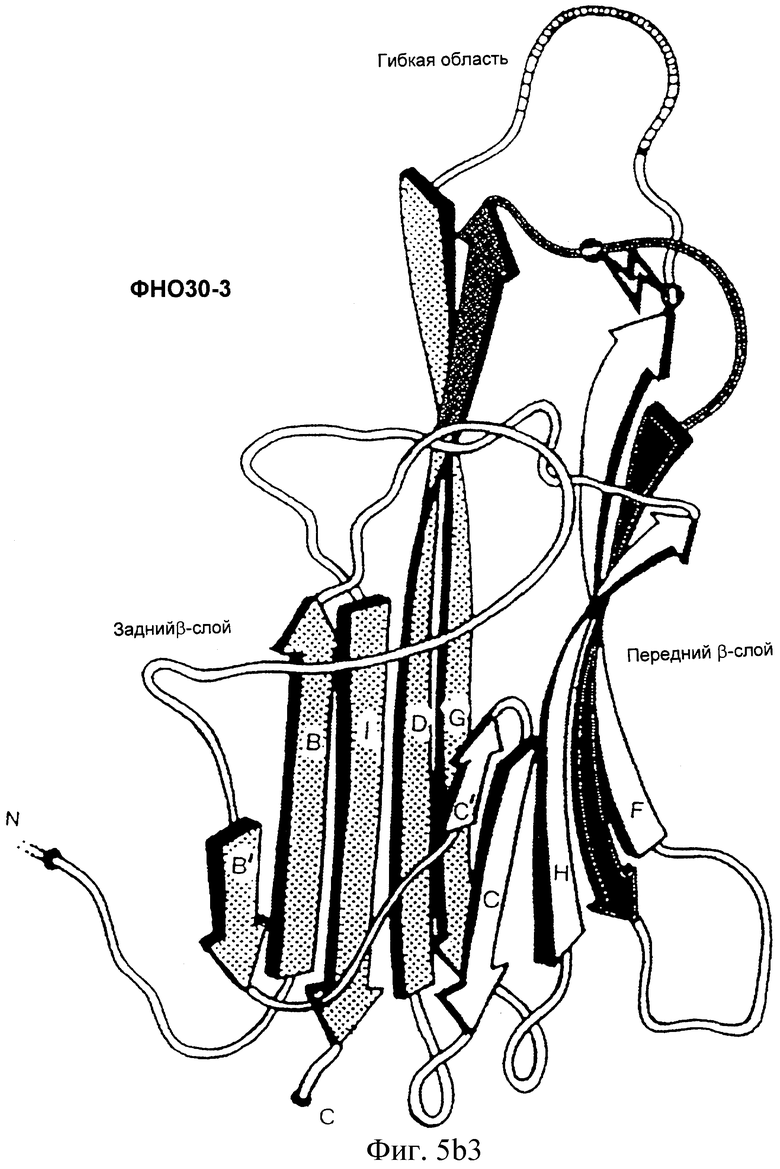




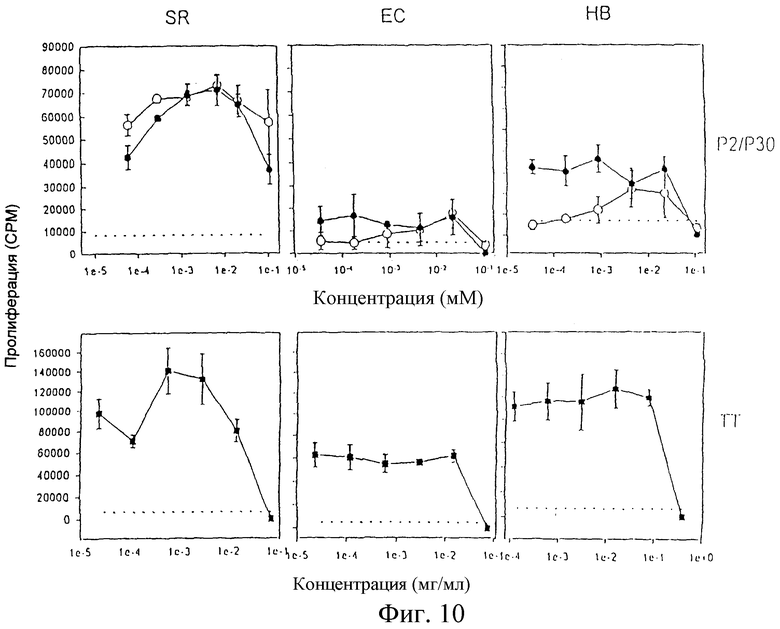




Изобретение относится к биотехнологии, генной инженерии и медицине и может быть использовано для получения модифицированной молекулы ФНОα человека, способной индуцировать образование нейтрализующих антител. Модифицированную молекулу ФНОα человека, способную увеличивать количество нейтрализующих антител к человеческому ФНОα дикого типа после введения человеку-реципиенту модифицированной молекулы ФНОα, получают путем замены по меньшей мере одного пептидного фрагмента молекулы ФНОα человека на по меньшей мере один пептид, содержащий иммунодоминантный эпитоп для Т-клетки, или создания усеченной формы вышеуказанной молекулы, содержащей иммунодоминантный эпитоп и одну или обе фланговых области молекулы ФНОα человека, содержащие по меньшей мере один ФНОα эпитоп для В-клетки, причем замена приводит к существенному изменению аминокислотной последовательности переднего β-слоя в любой из соединительных петель и/или в любой из В′, I или D цепей заднего β-слоя. Модифицированные молекулы ФНОα человека или ДНК, кодирующие их, используют в рецептурах в качестве вакцин против ФНОα по выбору с фармацевтически приемлемыми веществами, повышающими иммуногенность антител, для предотвращения или лечения хронических воспалительных заболеваний, таких как ревматоидный артрит и воспалительные заболевания кишечника, рак, рассеянный склероз, диабет, псориаз, остеопороз или астма. Жидкости тела человека исследуют на присутствие ФНОα при контакте с составом, содержащим модифицированный ФНОα. Изобретение позволяет разрабатывать средства для лечения хронических воспалительных заболеваний. 12 с. и 25 з.п. ф-лы, 16 ил., 3 табл.
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| ТЕРМОЭЛЕКТРИЧЕСКИЙ МАТЕРИАЛ НА ОСНОЗ СУЛЬФИДА ЦЕРИЯ;€л ПЛТЕНТРО-ВСЬСиШЗНАЯ" пхничса-АйБИБЛИОТЕКА | 0 |
|
SU251037A1 |
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| Гидравлическая система управления гидромеханической передачей транспортного средства | 1975 |
|
SU619372A1 |

