NAC1 - гены (включая NAM, ATAF1, ATAF2 и CUC2) принадлежат к относительно большой семье генов, до сих пор обнаруженной только в растениях. Эти гены кодируют белки, которые находятся в их N-конце ~170 аминокислот, но являются высокодивергентными в их С-концах. Предшествующие генетические исследования петунии (Souer et al., 1996) и Arabidopsis (Aida et al., 1997) установили, что некоторые члены семьи NAC играют роль в узорообразовании меристемы побега и цветка.
Зародыши петунии, несущие мутацию nam (отсутствия верхушечной меристемы), оказываются не способными развивать верхушечную меристему побега. Редкие побеги на nam проростках несут цветки, которые развивают десять вместо пяти примордий во второй мутовке. Двойные мутанты с гомеотическим геном green petals (зеленых лепестков) демонстрируют, что nam ведет себя не зависимо от идентичности органа в мутовке 2, а также влияет на количество примордий в мутовке 3. Поразительно, мРНК nam аккумулируется в клетках в пограничных слоях меристем и примордий. Видно, что nam играет роль в установлении местоположений меристем и примордий (Souer et al., 1996).
Мутации в CUC1 и CUC2 (для чашеобразной формы семядоли), которые являются генами Arabidopsis, ведут к дефектам при разделении семядоли (зародышевых органов), чашелистиков и тычинок (органов, относящихся к цветку), так же как и в строении верхушечных меристем побегов. Эти дефекты являются наиболее наглядными в двойном мутанте. Фенотипы мутантов подсказывают общий механизм разделения смежных органов внутри одной и той же мутовки не только в зародышах, но и у цветков. Ген CUC2 был клонирован и выделен для кодирования белка, гомологичного белку NAM петунии (Aida et al., 1997).
Публикации и другие материалы, используемые здесь для освещения предпосылок изобретения или для обеспечения дополнительными практическими сведениями, объединены ссылками и для удобства собраны в списке ссылок.
Описывается новый член семьи NAC Arabidopsis, а именно NAC1. Этот ген первоначально был выделен благодаря способности его кДНК менять клеточную морфологию дрожжей S. pombe, когда он сверхэкспрессирован (Xia et al., 1996). Northern анализ показал, что NAC1 был экспессирован тканеспецифичным способом с высокими уровнями в корне и низкими уровнями в листьях. Эксперименты тотального препарата in situ показали экспрессию в активно делящемся корне и в меристемах побегов.
NAC1 разделяет высокую гомологию аминокислотной последовательности с другими членами продуктов гена NAC в N-конце. In vitro связывание ДНК рассматривает использование рекомбинантного белка GST-NAC1 и различных усеченных производных NAC1, демонстрирующих взаимодействие между зоной -90 промотора 35S и консервативной N-концевой областью белка. Интересно, что исследования дрожжей показывают, С-конец NAC1, соединенный с областью связывания ДНК GAL4, может активировать транскрипцию. Анализ различных слияний делении показал, что область трансактивации размещена в С-конце белка NAС1. В дополнение, двугибридные испытания показали, что NAC1 может гомодимеризировать, и что область димеризации расположена в консервативной N-концевой зоне белка. 21 bp предполагаемая состоящая из двух частей ядерной локализации сигнальная последовательность была обнаружена в N-концевой консервативной области NAC. Трансгенный Arabidopsis был получен используя конструкцию слияния GFP-NAC1 под контролем дексаметазона (Dex), способного индуцировать систему промотор-GRG. Анализ трансгенных линий, экспрессирующих белок слияния GFP-NAC1 при условии Dex индукции, показал ядерную локализацию химерного полипептида in vivo. Эти данные указывают, что NAC1 принадлежит к недавно идентифицированной семье факторов транскрипции.
Первый аспект изобретения - это трансгенное растение, содержащее ген, который кодирует NAC1, или функционально эквивалентный белок (означающий белок, который связывает с тем же самым связывающим участком ДНК, как это делает NAC1, и который по крайней мере на 70% является идентичным, предпочтительнее - на 80% идентичным, более предпочтительно - на 90% идентичным и наиболее предпочтительно - не менее на 95% идентичным NAC1), что приводит к большему росту трансгенного растения по сравнению с нетрансгенным растением. Растение может быть больше, поскольку оно более тяжелое, оно имеет большие листья, более толстые стебли, имеет больше корней, и /или имеет более крупные корни.
Вторым аспектом изобретения является ген, кодирующий NAC1 или функционально эквивалентный белок, и где растения, которые являются трансгенными, благодаря этому гену вырастают более крупными, чем нетрансгенные растения.
Третий аспект изобретения - это NAC1 или функционально эквивалентный белок, где если растение является трансгенным благодаря гену, кодирующему NAC1 или упомянутый функционально эквивалентный белок, упомянутое трансгенное растение будет вырастать более крупным, чем растение, которое не является трансгенным.
Другим аспектом изобретения является клетка трансгенного растения, которая содержит ген, кодирующий NAC1 или функционально эквивалентный белок, который приводит к тому, что клетка растения вырастает более крупной, чем нетрансгенная клетка растения.
Перечень чертежей
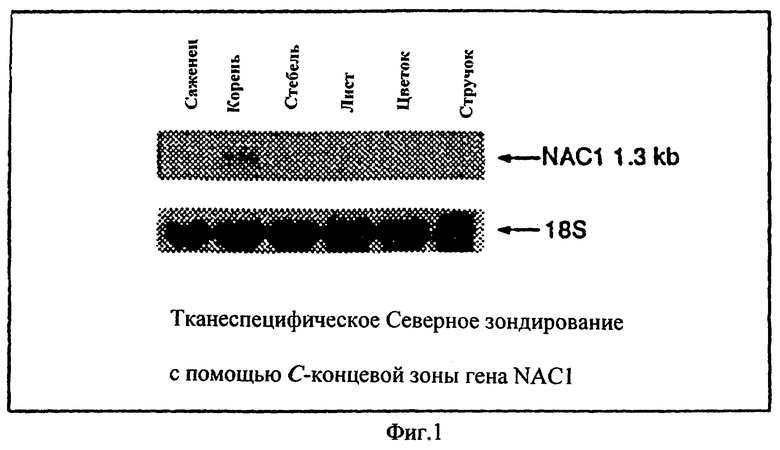
Фиг.1 - Тканеспецифичная экспрессия гена NAC1. Авторадиография пятен геля РНК, содержащих 10 μg общего РНК, полученных из различных тканей, как отмечено на бирке. Саженцы были выбраны из растений двухнедельного возраста. Стеблевые листья (лист), основной и боковые стебли (стебель), рацемозные соцветия (цветок) и стручки различных стадий были собраны с 35-40-дневных растений, растущих на почве. Корни были взяты от двухнедельных вертикальнорастущих саженцев на SM планшете. Фильтр был гибридизированн с помощью помеченного радиоактивным изотопом С-конца NAC1 и затем резондирован с помощью помеченного радиоактивным изотопом зонда рДНК 18S в качестве нагружающего стандарта (контроля).
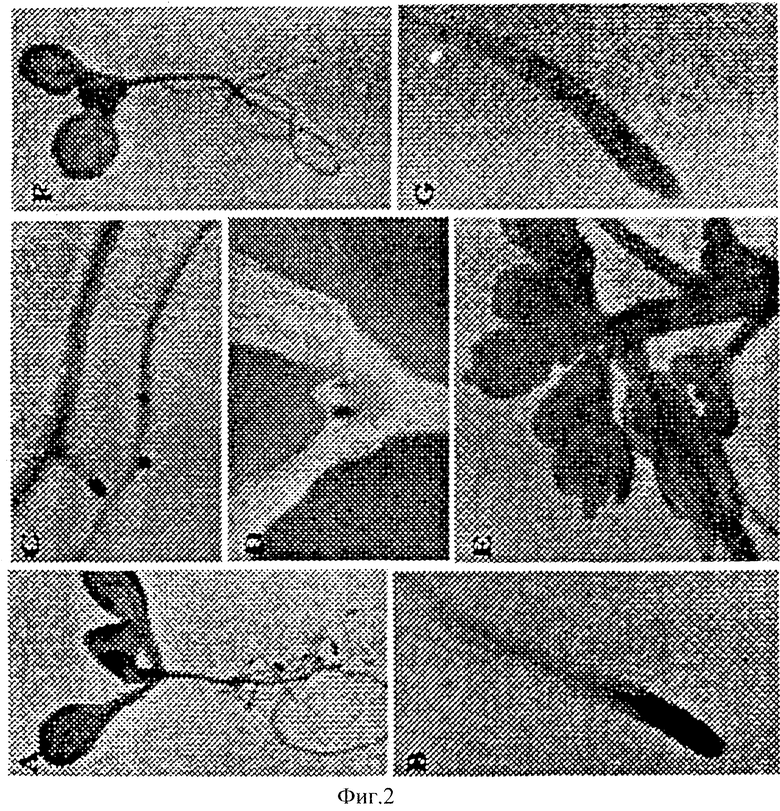
Фиг.2. - Исследование NAC1 в саженцах и цветках тотальным препаратом in situ. Гибридизация общим препаратом in situ была выполнена в саженце и цветке с помощью отмеченного дигоксигенином (DIG) С-концевого специфического антисенсного зонда (А, В, С, D и Е) и с помощью отмеченного DIG смыслового зонда (F и G). А и F являются саженцами двенадцатидневного возраста. Корневой материал образуют двенадцатидневные саженцы, а D - это семидневные саженцы. Цветки были взяты от сорокадневных растений, растущих в грунте.
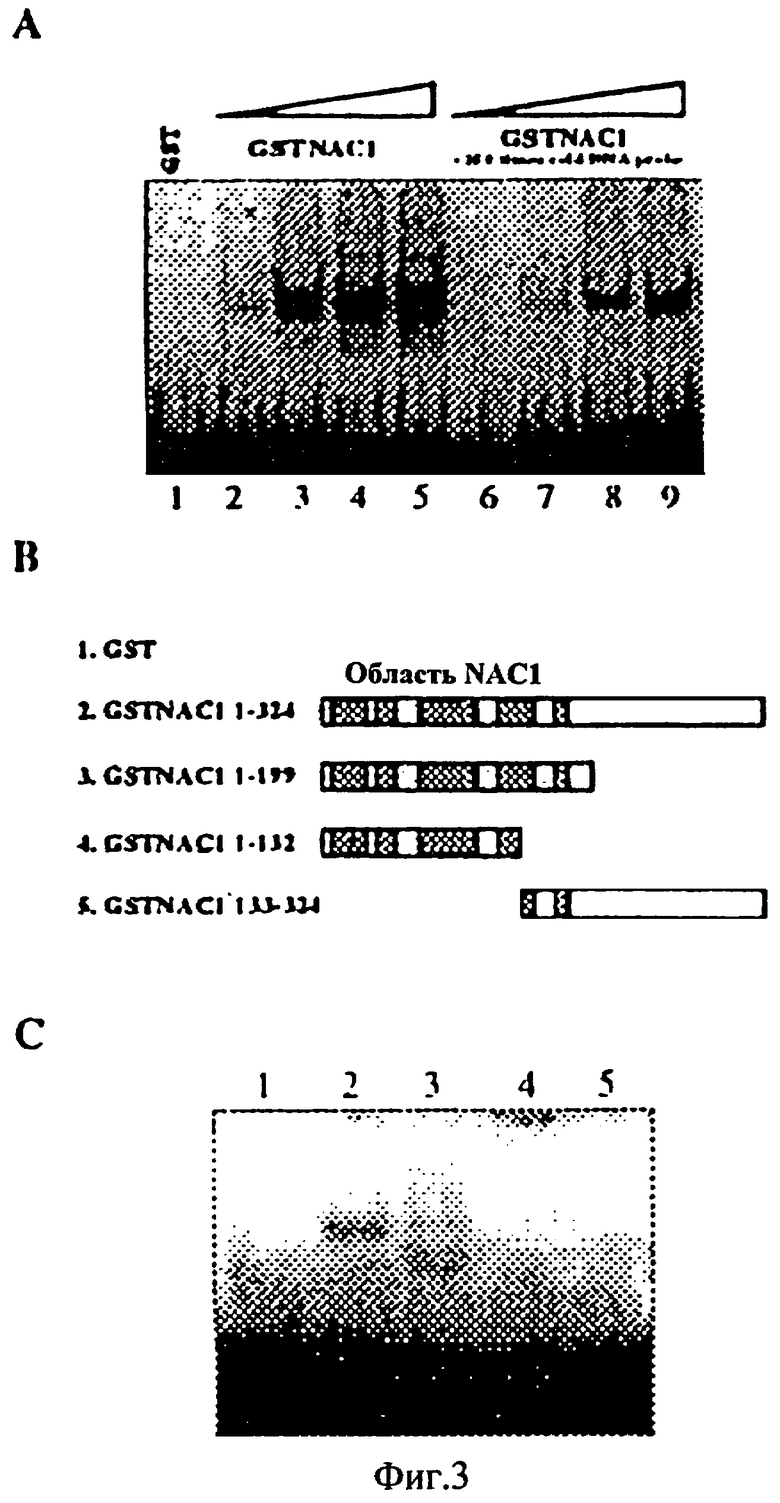
Фиг.3 - Белок слияния GST-NACI, связанный с зоной -90 промотора 35S CaMV. (А) Родство белка слияния GST-NAC1, связанного с зонами -90 промотора 35S. Количество белка как следует ниже: Полоса 1, 500 ng белка GST. Полосы от 2 до 9 - белки слияния NAС1. Полосы 2 и 6, 10 ng; полосы 3 и 7, 100 ng; полосы 4 и 8, 250 ng; полосы 5 и 9, 500 ng. В качестве несодержащего реактивных веществ конкурента был использован тот же немеченый фрагмент ДНК. (В) Карта различных делеций слияния NAC1 с GST. (С) Различные делеции белка слияния NAC1, связанного с зоной -90 промотора 35S CaMV. Было получено 20 ng каждого рекомбинантного белка.
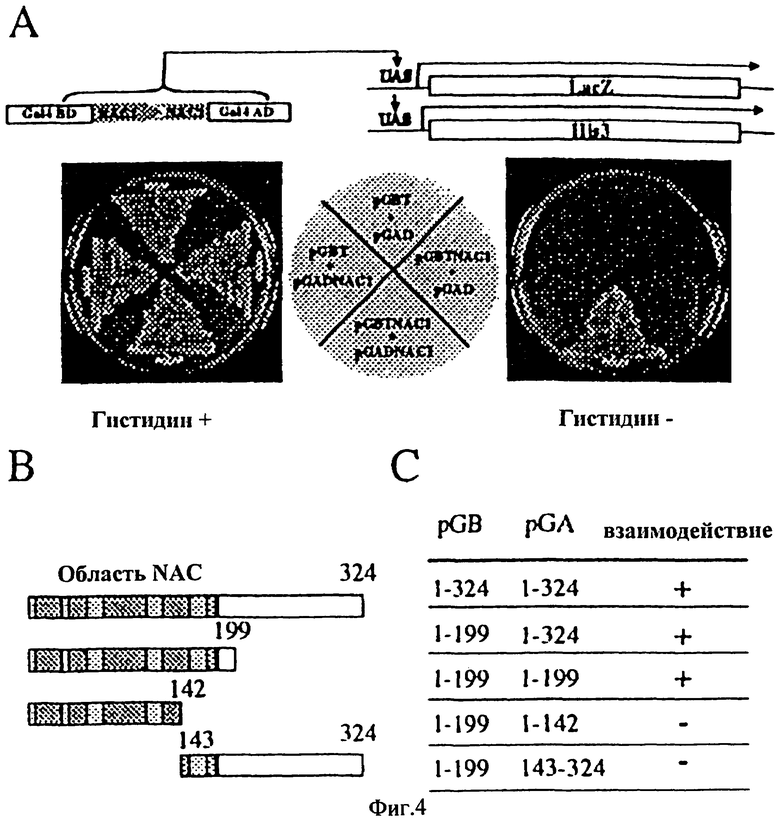
Фиг.4 - Специфическое взаимодействие между NAC1 и NAC1 в анализе двугибридной системы дрожжей. Клетки дрожжей HF7c были сотрансформированны с плазмидами, экспрессирующими одиночный GALA80 или слияние GALABD - NAC1, и с плазмидами, экспрессирующими одиночный GALAAD или GALAAD, соединенный с NAC1. (А) Клетки были посеяны штрихом на пластины с / или без гистидина плюс 10 mМ 3-АТ (Сигма) в соответствии с распределением, показанном в центре чертежа. Способность расти в отсутствие гистидина зависит от функционального восстановления активности GAL4. (В) Карта различных делеций слияния протеина NAC1 с вектором pGA. (С) Возможность взаимодействия между различными делениями вариантов NAC1 в анализе двугибридной системы.
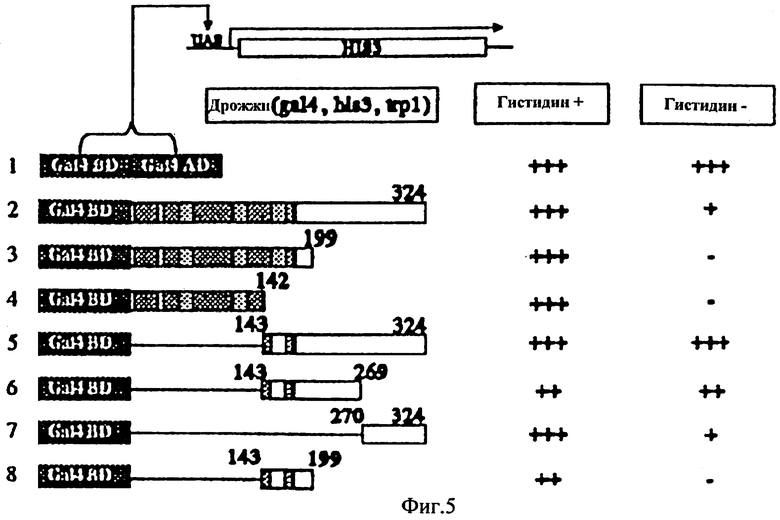
Фиг.5 - Локализация области трансактивации у белка NAC1. Различные версии делеции NACI были клонированы в вектор связывания ДНК Ga14 и трансформированы в штамм дрожжей, являющийся HF7c. Трансформируемая смесь была положена на ММ стекла с гистидином или без добавления гистидина, с необходимыми аминокислотами. Результаты были получены после 3-х дней роста со стекол.
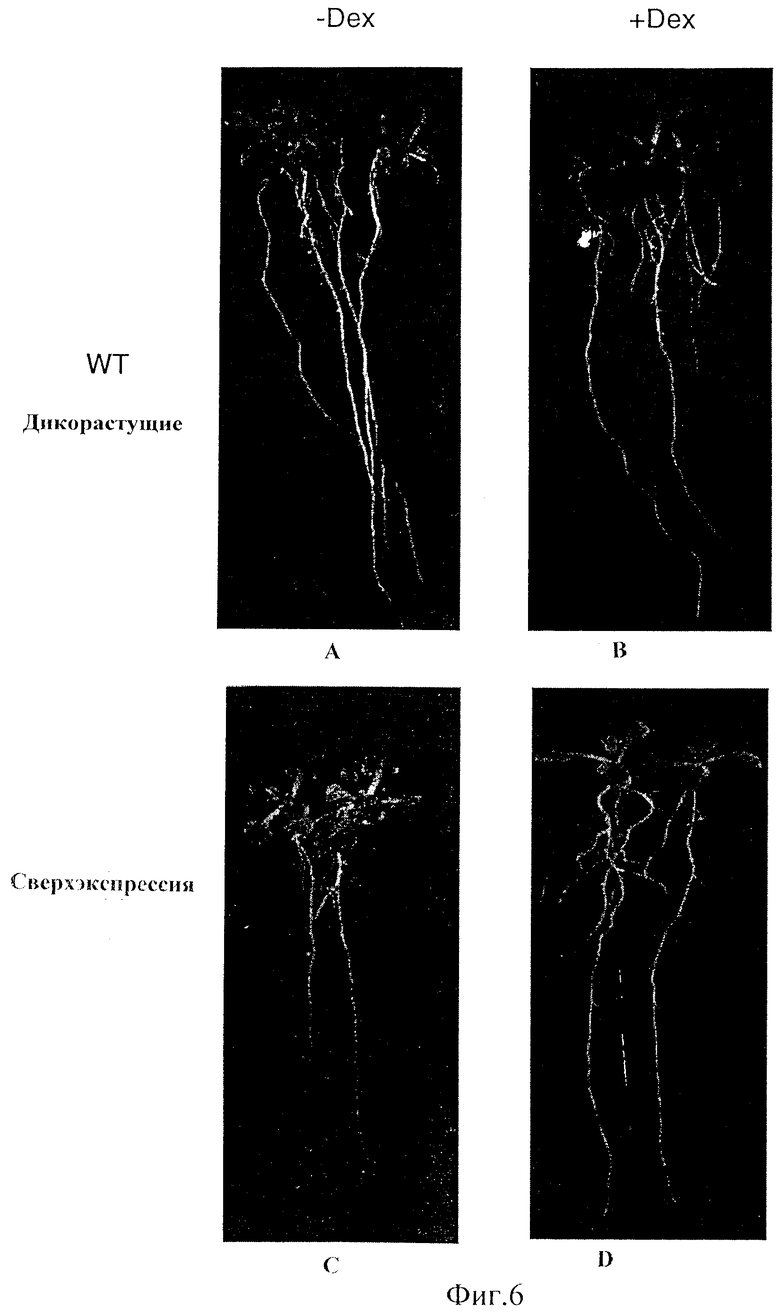
Фиг.6 - Воздействие сверхэкспрессии NAC1 в растениях. (А) Рост дикорастущих растений в отсутствие Dex. (В) Рост дикорастущих растений в присутствии Dex. (С) Рост трансгенных растений в отсутствие Dex. (D) Рост трансгенных растений в присутствии Dex.
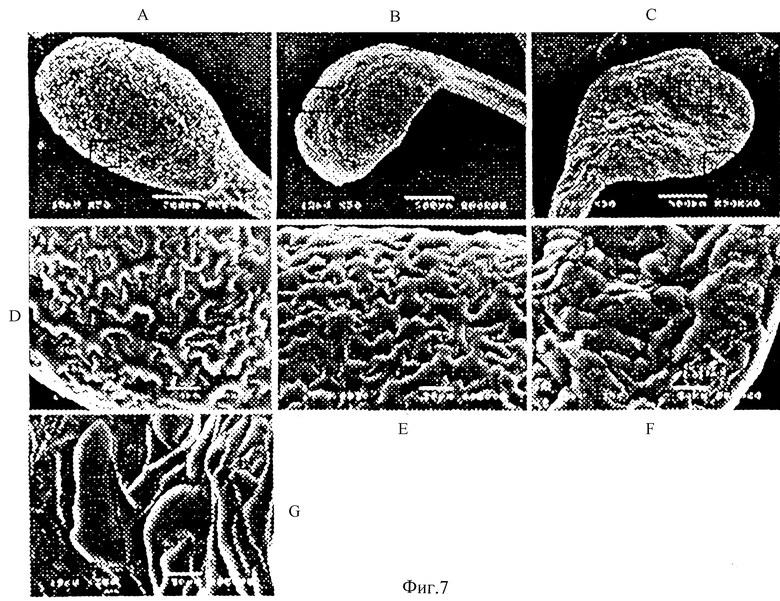
Фиг.7 - Эволюционные изменения семядоли в С-концевых специфических антисенсных растениях. Сканирующая электронная микроскопия (SEM) показывает абаксиальную поверхность семядоли. А и D (подробно) - это дикорастущие, В и Е (подробно) - это нестрогий фенотип, С и F, G (подробно) - строгий фенотип. Рамки с буквами на А, В и С показывают области, которые даются подробно.
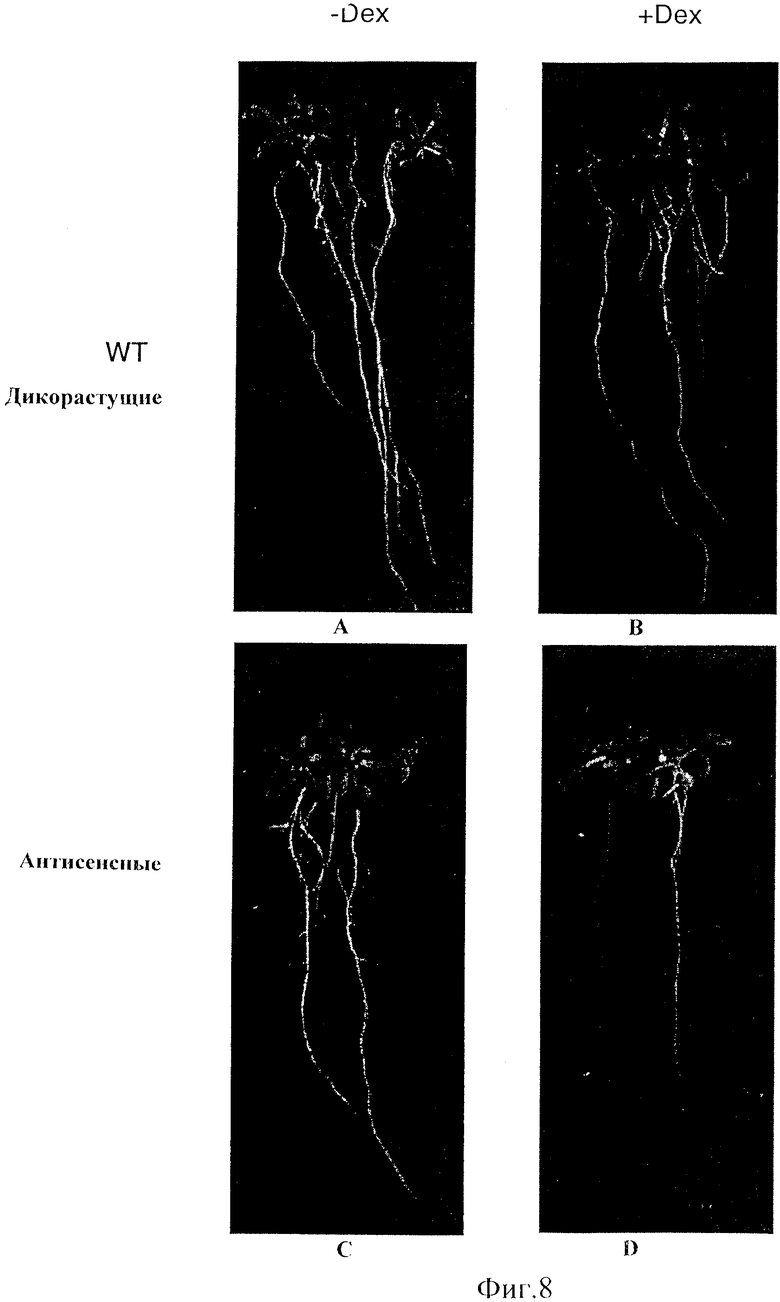
Фиг.8 - Семена дикорастущих и трансгенных растений с антисмысловым NAC1 были пророщены в MS или MS плюс Dex среде. (А) Дикорастущие в отсутствие Dex; (В) Дикорастущие, индуцированные Dex; (С) Трансгенные в отсутствие Dex; (D) Трансгенные, индуцированные Dex.
Ген nac1 был выделен в рамках программы получения цитоскелетала растения, клеточных циклическисвязанных и полярносвязанных белков, используя Schizosaccharomyces pombe. Библиотека к ДНК A. thaliana под контролем способного подавляться тиамином nmt1 промотора pREPSN была трансформирована в клетки дикорастущего S. pombe. Когда промотор был депрессирован, один трансформант показал вытянутые клетки и многоперегородчатость. Клон кДНК был выделен из этого трансформанта и ретрансформирован в S. Pombe для подтверждения изменения формы клетки, происходящего от экспрессии кДНК. Изолированный кДНК (At012) кодирует одну открытую рамку считывания (ORF) 324 аминокислот (SEQ ID No:T). Последовательность предполагаемого белка была взята из Банка генов. Поиск BLAST установил, что белок является новым. N-конец белка был обнаружен в NAC области, а область - в членах семьи генов NAC (NAM, ATAF1, ATAF2, CUC2). Эта область распространяется на первые 175 аминокислот белка. Остаток белка, кодируемого nac1, имел невысокую гомологию с другими известными последовательностями.
Гомология для полипептидов обычно измеряется, используя анализ последовательности с помощью программного обеспечения. Смотрите Пакет программ программных средств анализа последовательности University of Wisconsin Biotechnology Center, 910 University Avenue, Madison, Wisconsin 53705. Исследование протеина программными методами соответствует подобным результатам, использующим показатели гомологии определенные для различных замещений, делеций и других модификаций. Традиционные замещения обычно включают замещения внутри следующих групп: глицин, аланин, валин, изолецин, лецин; аспартиновая кислота, глутаминовая кислота; аспарагин, глутамин; серин, треонин; лизин, аргинин; фениланин и тирозин.
Семья генов NAC была определена только для растений. Эти гены кодируют белки, которые являются высоко консервативными в N-конце, но которые изменяются в остатке белка. Первые два члена ATAF1 и ATAF2 были из Arabidopsis, они были выделены с помощью их способности активизировать промотор 35S вируса мозаики цветной капусты (CaMV) в дрожжах (Н. Hirt, GenBank Accession Numbers X74755 and X74756). К-ДНК SENU5, выделенная при изучении старения листа у томатов (GenBank Accession Number Z75524; John et al., 1997), и белок NAM (Souer et al., 1996), продукт гена nam петунии, необходимы для правильного развития верхушечной меристемы побега и были предложены для определения местоположения меристемы. CUC2 (Aida et al., 1997) является членом семьи NAM и, кроме того, гомологом NAM Arabidopsis. Белок NAP Arabidopsis является также членом семьи (Sablonvski and Meyerowitz, 1998). GRAB1 (GenBank Accession Number AJ010829) и GRAB2 (GenBank Accession Number AJ010830) являются двумя недавно открытыми членами семьи NAC. Они - из пшеницы и могут взаимодействовать с белком RepA вируса карликовости пшеницы (WDV). Транс-сверхэкспрессия генов GRAB1 и GRAB1 под контролем промотора 35S может вызвать репликацию WDV в клеточной культуре пшеницы.
Работа, описанная здесь, подробно рассказывает об области NAC в пяти блоках, основываясь на сохранении и отслеживании каждого блока. Выстроенная последовательность белка NAC1 вместе со всеми этими белками показывает, что NAC1 имеет похожие характерные особенности с другими членами семьи NAС, которые относятся к недавно открытой области NAС.
Геномная структура nac1 была исследована посредством сравнения геномной последовательности с последовательностью кДНК. Ген NAC1 размещен на хромосоме 1. Он включает два интрона в N-концевой зоне кодирования. Первый интрон -это 1215 пар основания, и встречается в кодоне для 67 аминокислоты. Второй интрон - это 102 пары основания, размещен между кодонами для 160 и 161 аминокислот. Поиски области были выполнены и привели к определению предполагаемого состоящего из двух частей ядерной локализации сигнала в области NAC NAC1. Эта 21 аминокислотная сигнальная последовательность размещена со 117 по 137 аминокислоту и была впервые обнаружена у члена семьи NAС. Выполнение таких же исследований других описанных членов семьи NAC для обнаружения подобного сигнала терпели неудачу. Гомологи NAC были обнаружены в рисе базы данных EST, но не было замечено гомологов в других организмах, таких как млекопитающие, Дрозофила и дрожжи.
Эволюционное развитие гена NAC1
Для изучения роли гена NAC1 во время развития растения, его паттерн экспрессии был изучен посредством Northern анализа, с использованием специфического зонда nac1. Поскольку члены семьи NAC являются очень консервативными в N-концах даже на уровне ДНК, был использован С-концевой зонд. 10 μg общего РНК из каждой ткани были использованы для гибридизации. Относительно высокий уровень экспрессии РНК nac1 был замечен в корнях (Фиг.1). Более низкая экспрессия наблюдалась в двухнедельных саженцах, которые содержали всю растительную ткань. Очень низкая экспрессия была отмечена в материале, полученном из стеблей и листьев. Отсутствие экспрессии было отмечено в смешанных стадиях стручков и смешанных созревших цветках и соцветиях, даже после долгого выдерживания.
Исследования тотальным препаратом in situ растений возраста от 7 до 12 дней были выполнены, используя как смысловой, так и антисмысловой зонды в С-концевой зоне кодирования гена. Экспрессия nac1 была замечена в активно делящихся корнях и верхушечных меристемах побега (Фиг.2). Экспрессия была особенно высокой в делящейся зоне конца корня (Фиг.2А и 2В) и зоне образования бокового корня (Фиг.2С). Более низкая экспрессия была отмечена в семядоле и молодых листьях. Все эти результаты соотносятся с результатами ученых из северных штатов США. В цветках было отмечено отсутствие чисто специфической гибридизации (Фиг.2Е). Также, отсутствие сигнала было отмечено в стручках и стеблях (данные не показаны). Отсутствие специфического сигнала было замечено в саженцах, гибридизированных с помощью смыслового зонда РНК (Фиг.2.F и 2G).
Связывание ДНК и области связывания ДНК
Первые два гена семьи NAC, ATAF1 и ATAF1, были клонированы благодаря их способности активировать конструкцию промотора 35S CaMV (вируса мозаики цветной капусты) в дрожжах. Эти два белка, так же как другие члены семьи NAC, хранятся в N-концевой области NAC. Белки GRAB1 и GRAB2 из пшеницы также связывают зону -500 промотора 35S. Промотор 35S CaMV содержит несколько известных элементов связывания ДНК. Число факторов транскрипции растения было определено как связывание с различными элементами в регуляторной зоне промотора.
Для определения может ли белок NAC1 связываться с промотором 35S, зона -90 промотора 35S была впервые использована как зонд. nac1 был клонирован в вектор pGEX-4-2Т GST-слияния и трансформирован в Е. coli. Рекомбинантный белок GST-слияния был исследован. Исследование было выполнено для тестирования специфического связывания с ДНК. После того как белок слияния GST был исследован, сам GST был использован в качестве стандарта. GST не был способен связывать зону -90 промотора 35S, даже когда было использовано 500 ng очищенного белка (см. Фиг.3А).
Были испытаны различные количества очищенного белка GST NAC1. Связывание было зарегистрировано с помощью такого немногого количества, как 10 ng белка, и только с помощью пленки 4-часовой экспозиции (Фиг.3А). Необходимо заметить, что очищенный белок GST, содержал только около 10% формы интактного GST слияния. Большинство белка разрушилось во время бактериального роста и очищения (от бактерий). Это произошло потому, что когда был добавлен IPTG для индуцировния экспрессии белка, бактерия не могла хорошо расти, и белок начал разрушаться до очистки. Добавление до избытка не содержащего реактивных веществ конкурента ДНК промотора 35S уничтожило образование комплекса ДНК-белок зависимым от концентрации образом (Фиг.3А). Эти результаты показывают, что белок NAC1 специфично связывает зону -90 промотора 35S.
Было также тестировано связывание NAC1 с элементом AS-1 и мутантной формой элемента, от -83 до -63 промотора 35S. Факторы zip транскрипции растения были определены как связывание с этой зоной. Связывающее родство было уменьшено, и неясное различие было обнаружено между оригинальным элементом AS-1 и мутирующей формой. Этот результат показывает, что NAC1 имел сайт связывания ДНК, отличный от факторов транскрипции bZIP, а последовательности вокруг элемента AS-1 являются важными для специфического связывания NAC1.
Для тестирования области связывания ДНК NAC1 различные формы делеций nac1 были клонированы в вектор GST слияния, и белки слияния были выделены для тестирования способности связывания ДНК с зоной -90 промотора 35S. В этом исследовании были использованы 20 ng каждого белка. Результаты показаны на Фиг.3С. Усеченная форма GST NAC1 содержала первые 199 аминокислот, интактную область NAC. Это поддерживало подобное специфическое связывание с зондом ДНК. Сокращенная версия, содержащая только 3 первых интактных блока области NAC и часть IV-го блока, утратила способность связывания ДНК. Подобно этому комплементация этой формы делеций, остаток белка, содержащий часть IV-го блока и интактный V-й блок, не связываются с зондом ДНК. Вместе эти результаты указывают, что область связывания ДНК размещена в области NAC, и что IV-и блок является важным для связывания ДНК, а V-й блок недостаточен для связывания ДНК.
Гомодимеры NAC1 и область димеризации
Факторы транскрипции обычно формируют димеры, когда они связываются с ДНК. Для определения, формирует ли NAC1 димеры, мы использовали двугибридный метод. Метод был эффективно использован для тестирования димеризации и исследования взаимодействия белок-белок, включая димеризацию. Интактный NAC1 был соединен с областью активизации ДНК Ga 14 и с областью связывания ДНК, затем совместно трансформирован в дрожжи репортер. Чистое взаимодействие было замечено, когда NAC1 был выражен обоими векторами. Дрожжи были способны вырастать в три дня, даже при использовании 10 mМ 3-АТ, чтобы блокировать неспецифические взаимодействия (Фиг.4А). Необходимо отметить, что когда NAC1 был выражен только вектором связывания ДНК, он мог с низким уровнем самоактивировать систему так, что дрожжи могли расти очень медленно (Фиг.4А).
Для определения зоны NAC1, требуемой для димеризации, мы построили серии делеций и изучили их способность формировать комплексы в дрожжах. Первая делеция С-конца была выполнена от 200-ой аминокистлоты до конца белка. Она отделилась от белка первых 199 аминокислот. Этот фрагмент белка 199 аминокислот содержит интактную область NAC, и он был способен создавать димеры с интактным NAC1. Этот вариант 199 аминокислот дал отсутствие фонового сигнала подобно некультивированному полной длины варианту NAC1. Вследствие этого вариант 199 аминокислот был использован в качестве фиксированного партнера для тестирования других делеций. Были подготовлены разновидности различных делеций в векторе области активации (Фиг.4В). Когда такая делеция (без последовательности за 199-ой аминокислотой) была препарирована в pGA, взаимодействие было таким же сильным, как с интактным белком. Но когда делеция повредила IV-й блок области NAC, а структура содержала только первые три интактных блока области NAC, формация димеров была разрушена (Фиг.4С). Делеция N-конца, переносящая только последнюю половину IV-го блока и интактный V-й блок, имела результатом неспособность белка формировать димеры. Данные поддержали заключение, что область NAM NAC1 содержит область димеризации. IV-й блок является важным для формирования димера, но V-й блок сам по себе недостаточен для формирования димера. Белок NAC1 может формировать гомодимеры, и это было подтверждено фильтрацией геля. Когда бактерия экпрессировала белок первых 199 аминокислот NAC1, очищенный пептид, включая полную область NAC, дважды увеличился в размере в нативном геле, как в денатурированном геле.
Трансактивация и область трансактивации
При двугибридном исследовании дрожжей мы обнаружили, что NAC1 может самоактивировать систему к уменьшению уровня, а С-концевая делеция не может. Поэтому мы использовали модифицированный одногибридный метод с помощью фактора транскрипции гибрида для определения области активации гена NAC1. Вектор pGBT8, который содержит только область связывания ДНК Ga14, был использован кроме области отсутствия активации. Различные делеции гена NAC1 были клонированы в эту плазмиду в слиянии с областью связывания ДНК Ga14, и полученные в результате плазмиды были трансформированы в дрожжи, содержащие ген репортер UASgal1-CYC1-HIS3. Отбор был выполнен на лишенных гистидина планшетах. Если часть белка слияния содержала активность активации, транскрипция Ga14 могла быть восстановлена и изменена на промоторе HIS3. С помощью синтеза гистидина дрожжи могут расти на среде без гистидина. Область активации Ga14 была использована в качестве позитивного стандарта, и это дало полную активность, как показано на Фиг.5. Как упоминалось выше, полный белок NAC1 дал только низкий уровень активности активации. N-концевое слияние, содержащее первые 132 аминокислоты или первые 199 аминокислот, дало отсутствие активности. Подобно этому слияние с фрагментом пептида аминокислот 143-199 дало отсутствие активности. По контрасту, слияние, содержащее аминокислоты от 133 до карбокси-границы, полностью активизировало ген репортер, дававший сигнал такой же определенный, как и фактор транскрипции Ga14 позитивного стандарта. Использование слияния, содержащего только аминокислоты 143-269, также дало полную активность. Это является высоко кислотной зоной аминокислоты. С-концевое слияние, содержащее только последние 54 аминокислоты, привело к низкой активности. Результаты показывают, что область активации NAC1 размещена в С-окончании.
Поскольку семья NAC была обнаружена только в растениях, а не в других организмах, важно контролировать активность трансактивации в растениях. Эксперимент in vitro был устроен для контролирования области активации. Были созданы две плазмиды. Первая плазмида переносила кассету Ga14 связывания ДНК-MCS-Nos окончания, помеченную промотором 35S CaMV. Тестирующий ген мог быть клонирован в MCS в рамке с помощью области связывания ДНК Ga14. Другая плазмида содержит структуру гена-репортера 6xUAS-TATA-Luc, которая может эксперессировать люциферазу, если некоторые белки могут связаться с элементом 6xUAS для активации промотора. Совместная бомбардировка двумя плазмидами вместе в различных комбинациях демонстрировала подходящую систему, способную к репродукции. Мы использовали область активации из вируса VP16 в качестве позитивного стандарта, и это дало очень высокую активность. По контрасту ни одна вторая плазмида (pTALuc), ни она плюс вектор pGa1, не дали существенной активности люциферазы. Подобный результат был получен из дрожжевой системы. Полная активность трансактивации была установлена в пептиде аминокислот 143-269. Последние 54 аминокислоты также дали низкую активность. N-конец NAC1 показал как отсутствие активности, так и векторного самоконтроля. Обе системы - в растении и в дрожжах, продемонстрировали, что NAC1 содержит область трансактивации, и зона, соответствующая этой активности, находится между 142-269 аминокислотами.
Трансгенные растения Arabidopsis, которые сверхэкспрессируют и недоэксперссируют NAC1
В этом исследовании была использована трансгенная система, способная индуцироваться глюкокортикоидом, как ранее описанная (Aoyama and Chua, 1997). Фактор транскрипции, контролируемый глюкокортикоидом, кодируется с помощью GVG, чья транскрипция контролировалась промотором 35S CaMV. Транскрипция трансгена контролируется активизированным глюкокортикоидом промотором (6XUASga14). 1287 пара основания полного NAC1 кодирующего кДНК (SEQ ID NO:1), была клонирована в двойной вектор трансформации растения рТА7002 в обоих направлениях, и отдельно трансформировано в экотип Arabidopsis thaliana lansberg с помощью метода корневой трансформации. Трансгенные линии сверхэкспрессии и недоэкспрессии были отобраны из среды, содержащей гигромицин. Были отобраны шесть независимых трансгенных линий сверхэкспрессии и четыре независимых линии недоэкспрессии. Все эти линии расщепили для одного локуса Т-ДНК согласно коэффициентам устойчивости расщепления гигромицина.
Шесть трансгенных растений сверхэкспрессии были трех различных фенотипов. Четыре линии были одного фенотипа, а оставшиеся две линии, каждая, показали отличный фенотип. Эти последние два фенотипа являются подобными антисмысловым растениям, как будет описано далее. Изучение первых четырех линий сходного фенотипа показали, что эти растения росли быстрее и были более крупными, чем нетрансгенные растения при in vitro, так и в условиях роста в почве. Большее количество боковых корней было получено, когда были использованы воздействующие условия, такие как на планшетной вертикали или в "plantcon growths". Данные представлены для условий роста in vitro. Саженцы различного возраста были пересажены из среды MS не только в среду MS, но и MS плюс 10 μМ дексаметазона (Dex), затем проверенны после однонедельного роста. 10 растений из каждой линии гомозиготов были взвешены после очищения агара или земли с растений. Однонедельные саженцы, пересаженные в среду Dex, были от 1,1 до 1,3 раз тяжелее, чем неиндуцированные растения, 15-дневные саженцы были тяжелее от 1,3 до 1,8 раз, а 25-дневные саженцы были тяжелее от 1,4 до 2,6 раз. Увеличенный вес индуцированных растений был результатом больших листьев, более толстых стеблей и большего количества корней, но главным вкладом был размер листа. Эпидермальные клетки листа от различных частей растений были исследованы с помощью SEM (сканирующей электронной микроскопии). SEM показала, что размер клетки старых листьев был больше, чем контрольный размер клетки, тогда как ясного отличия размера клетки новых растущих листьев не было. Корни вертикального роста представленной линии показаны на Фиг.6. Большее количество более длинных боковых корней были получены у индуцированных Dex линий. Общая РНК была приготовлена из индуцированных 48 часов растений и ложных стандартов и была измерена посредством Northern пятен, зондирующих С-концевым специфическим зондом nac1. По результатам все четыре трансгенных линии показали, что трансген экспрессирован. Размер mРНК трансгена больше, чем размер природного mРНК и является легко различимым.
Две оставшихся сверхэксперессирующих линии так же хорошо, как все четыре отобранных линии антисмысловых трансгенных растений, показали некоторое разделение различных комбинаций фенотипов. Одна из двух сверхэкспрессирующих линий и одна из антисмысловых линий показали фенотип выгнутой семядоли на этапе проростка. Нечеткие фенотипы были отмечены в более поздней стадии вегетативного развития соцветий. Две антисмысловых линии показали очень типичные фенотипы на стадии проростков, так же как мутант cuc2. В этом случае отдельная серцевидная и слитая семядоля и розетка проростков были отмечены невысоким процентом прорастания, около 7-10%. Большинство этих проростков, которые не могли произвести верхушечную меристему побега, позже погибли. Одна из антисмысловых линий и одна сверхэкспрессирующая линия не показали каких-либо отличий от неиндуцированных растений в течение всех стадий вегетативного роста и прорастания. Но на стадии развития соцветий органы цветков были поражены, особенно лепестки и тычинки. Фенотипы были обнаружены в первых нескольких цветках соцветия. Эти цветки имели короткие лепестки и тычинки, и там, где был более строгий фенотип, цветки не смогли открыться полностью. Некоторые из них смогли открыться, но были видны чистые короткие тычинки. Эти цветки были мужскими и стерильными женскими. Этот тип фенотипа перекрывает сверхэкспрессию другого члена семьи NAC Arabidopsis - гена NAP.
Трансгенное растение Arabidopsis, недоэкспрессирующее
С-концевой специфический ген NAC1
Так как члены семьи гена NAC являются высоко консервативными в N-конце, не только на уровне аминокислоты, но также на уровне последовательности нуклеотида, антисмысловые нуклеиновые кислоты до этой зоны будут поражать гомологичные гены, как показано на примере петунии. Кроме того, С-концевые специфические антисмысловые конструкции были приготовлены, используя вектор рТА7002, и трансформированы в Arabidopsis, как описано выше. Для этой конструкции мы использовали 0,6 kb BamHI-NotI С-концевой фрагмент. Northern пятна обнаружили только одну полосу, соответствующую гену NAC1 во всех тканях, используя этот фрагмент как зонд для четырех из шести гомозиготных линий, которые мы отобрали для исследований. Все показали сходные фенотипы, главным образом влияли семядоля и развитие корня. Нечеткие фенотипы были замечены в течение всех других стадий роста растения. Одна представленная линия С-специфических антисмысловых растений описывается здесь. Семядоли диких или неиндуцированных проростков показали слегка вогнутую и гладкую поверхность. На первой неделе прорастания нечеткий фенотип был замечен в развитии семядоли, но 10-дневные саженцы от 20 до 25% показали строгий фенотип с выгнутой семядолей. Незначительное число также показало вогнутый фенотип. Феномен мог быть легко замечен под световой микроскопией или даже увиден непосредственно глазом. На более поздней стадии черные точки могли быть увидены на поверхности семядолей. Абаксиальные эпидермальные клетки были проверены посредством SEM, и ненормальное размножение клеток было обнаружено над всеми поверхностями семядоли, хотя уровни ненормального размножения были различными (Фиг.7). Эти клетки теряли нормальную подобную головоломке-пазл структуру и становились приведенными в беспорядок из-за различных размеров и форм клеток (Фиг.7С, 7F и 7G). Семядоли этих от 75 до 80% саженцев, которые показали отсутствие строгого или ясно выгнутого фенотипа под световой микроскопией, также были проверены посредством SEM. Результаты SEM указали, что поверхность клеток этих семядолей была также ненормальной. Клетки выглядели раздутыми и сильно расширенными.
Фенотипы корней были легко определены на саженцах вертикального роста. Семена были высеяны на MS или MS плюс Dex среду. После одной недели развития саженцы были перенесены в ту же свежую среду и непрерывно росли одну неделю, а затем были проверены. Небольшое количество корней было получено у саженцев, которые имели С-концевую антисмысловую экспрессию. Три линии имели небольшие корни или немного боковых корней, и одна линия почти не имела боковых корней. Эти саженцы, которые имели небольшие корни или немного боковых корней, были проверены с помощью микроскопии, и она показала, что появление боковых корней могло быть вызвано, но они не могли вытягиваться. Кроме того, саженцы выглядели так, как если бы они содержали меньше корней по сравнению с неиндуцированными саженцами (Фиг.8). Похожие результаты были получены от С-концевых антисмысловых растений, выращенных в жидкости. Однонедельным саженцам были добавлены в жидкость среда MS или MS плюс Dex, и после 12-дневного роста неиндуцированные корни были большими, чем индуцированные корни саженцев.
Представленное изобретение далее подробно раскрывается в предложенных примерах, которые даны для иллюстрации и никоим образом не ограничивают изобретение. Используются стандартные технологии, известные специалистам, или технологии, описываемые ниже.
Пример 1. Растительные материалы и условия роста
Для экспериментов был использован экотип Arabidopsis thaliana lansberg. Поверхность семян была стерилизована с помощью 20% отбеливающего вещества плюс 0,01% Triton X-100, и три раза промыты стерильной водой. После последнего промывания в семена была добавлена 0,15% агароза, и они были высеяны в чашки с MS или MS плюс дексаметазон (Dex) в различных концентрациях с 3%-ной сахарозой. Чашки были выдержаны при 4°С два дня и перенесены в помещение для тканевой культуры с условиями 22°С и долготе дня (16 часов света /8 часов темноты). Затем от двух до трех недель саженцы были высажены в горшки с почвой и росли в камере роста с фотопериодом 16 часов света /8 часов темноты при 22° и 75% влажности.
Для Dex обработки дексаметазон (Sigma) был разбавлен в DMSO (димексид), для получения 100 mМ запасного раствора дексаметазона, который хранился при -20%. Для роста растений in vitro Dex был добавлен в чашки в количестве 0,1, 1 и 10 μМ. Dex был непрерывно добавляем в среду один раз в неделю. Был использован светоконтроль и контроль растений. Dex был сначала разбавлен в стерильной воде при 1/25 объема среды и затем добавлялся на поверхность среды. Для обработки растений, растущих в почве, Dex добавлялся в стерильную воду, содержащую 0,01% Triton Х-100 при 30 μM, затем разбрызгивался так, чтобы покрыть все поверхности растения, один раз в два дня. Для ложных стадартов стерильной водой, содержащей 0,01% Triton X-100, обрызгивалось такое же количество растений.
Пример 2. Конструкции трансформации и трансформация рстений
Дважды трансформированная плазмида рТА7002, содержащая полную двукомпонентную способную индуцироваться глюкокортикоидом систему (Aoyama and Chua, 1997), была использована в качестве вектора. Для сверхэкспрессии и полных антисмысловых конструкций у фрагмента 1287 пары основания Sal1-Not1, содержащего полную кДНК кодирования NAC1, был "затуплен конец" и он был клонирован в Xho1, сожженый и обработанный щелочной фосфотазой внутренностей теленка вектор рТА7002. Для приготовления С-концевой специфической антисмысловой конструкции у фрагмента Ваm H1-Not1 605 спаренного основания, содержащего С-концевую специфическую зону NAC1 кодирования кДНК, был "затуплен конец" и он был клонирован в вектор рТА7002, как упоминалось выше. Для трансгенных конструкций, содержащих кассету слияния GFP-NAC1, фрагмент Nae1-Sal1, генерированный PCR, содержащий только зону кодирования NAC1, был клонирован в промежуточный вектор pGFP2(GA)5II слияния GFP, усвоенный Nae1 и Sal1 для получения плазмиды pGFPNAC1. Затем слияние GFPNAC1 было вырезано Хbа1 плюс Рас1 и клонировано в вектор рТА7002 с помощью тех же операций, как было описано выше. Как растительный материал для трансформации были использованы корни Arabidopsis thaliana lansberg (Valvekens et al., 1988). Семена Т2 были пророщены на планшетах SM, содержащих 20 μg/mL гигромицина В для отбора устойчивых растений и перенесены в почву для выращивания и получения гомозиготных семян 73. Две независимых линии гомозиготных растений T4 с одной вставкой из каждой конструкции были использованы для подробных анализов.
Пример 3. Northern анализ и анализ тотальным премаратом in situ
Общий РНК был выделен из различных тканей при помощи набора Qiaren для приготовления РНК. Northern блот-анализ был выполнен согласно Nagy et al., (1988). Для зондов фрагмент PCR, покрывающий аминокислоты от 200 до 324, был помечен α-32P dCTP, используя Redi - служащий затравкой набор для мечения (Amersham Internhational). Целые растения или цветки были зафиксированы для гибридизаций тотальным препаратом in situ. Антисмысловые РНК из С-концевой специфической зоны NAC1 были помечены дигоксигенином UTP (Boehringer Biochemica, Mannheim, Germany). Гибридизация была обнаружена с помощью Fab-фрагмента анти-дигоксигенина, конъюктивировавшего со щелочной фосфотазой. В качестве контрольных зондов были приготовлены смысловые транскрипты из того же клона, используя 73 транскриптазу.
Пример 4. Двугибридный анализ дрожжей
Для анализа димеризациии был использован двугибридный анализ дрожжей (Bartel et al., 1993; Fields and Song, 1989; Chevray and Nathans, 1992; Lee et al., 1995). Был использован дрожжевой штамм HF7c (MATa urа3-52 his3-200 ade2-101 lys2-801 trp1-901 leu2-3,112 gal4-542 gal 80-538 LYS2::GAL1UAS-GAL1TATA-HIS3 URA3::GAL41-mers(x3)-CyC1TATA-LacZ, который содержит два гена репортера LacZ и HIS3. Совместная трансформация дрожжей с плазмидами, переносящими особую область связывания ДНК GA14 и слияние области активации GA14, была выполнена с помощью способов, известных специалистам в этой области. Смотрите Burke and Olson, 1986; Rudolph et al., 1985; Sakai et al., 1984. Другие условия были подробно описаны в другом месте и хорошо известны специалистам. Для подтверждения взаимодействия между двумя белками слияния активность β-галактозидазы была исследована с помощью анализа фильтра реплики.
Две версии NACI были клонированы в область связывания GA14 ДНК вектора pGBT8, производного pGBT9 (Clontech), который содержит более подвижный полилинкер. Плазмида pGB NAC11-324, содержит полную открытую рамку считывания для NAC1, соединенного с областью связывания ДНК GA 14. Плазмида pGB NAC11-199 содержит только нуклеиновую кислоту, кодирующую первые 199 аминокислот в том же векторе. Различные усеченные версии NAC1 были слиты в область активации GA14 вектора pGAD424. Плазмида pGA NAC11-324 содержит полную открытую рамку считывания для NACI. Плазмида pGA NAC11-199 содержит нуклеиновую кислоту, кодирующую первые 199 аминокислот NAC1. Плазмида pGA NAС11-142 содержит нуклеиновую кислоту, кодирующую первые 142 аминокислоты NAC1, а плазмида pGA NAC1143-324 содержит нуклеиновую кислоту, кодирующую аминокислоты от 143 до С-конца пептида.
Пример 5. Трансфекция эпидермальных клеток с помощью бомбардировки частицами
Лепестки были разрезаны на кусочки 2×2 см и помещены на впитывающий фильтр с жидкой MS средой. 2 μg каждого сочетания плазмид было использовано при каждой бомбардировке в вакууме при 27 дюймах ртутного столба, используя воздействие гелия 1100 psi. После бомбардировки планшеты были выдержаны одну ночь (16 часов) в комнате для роста растений, и 50 mМ люциферина (Promega) было разбрызгано на материал. Результаты были зафиксированы спустя 10 минут.
Пример 6. Получение белков GST-слияния и связывание ДНК
Продукт PCR, содержащий полную зону кодирования гена NAC1, был клонирован в pGEX-A-2T для получения конструкции слияния pGST-NAC1.
GST-NAC1 1-199 была произведена сжиганием pGST-NAC1 с ВаmHI и SalI, обработана Klenow и религирована для получения pGST-NAC1 1-199, которая содержит только первые 199 аминокислот белка NAC1. GST-NAC1 1-132 была получена разрезанием GST-NAC1 XhoI плюс SalI, и было религировано.
GST-NAC1 133-324 была построена с помощью клонирования фрагмента XhoI-NotI клона оригинальной кДНК в pGEX-4-2T. Плазмиды были трансформированы в Е. coli BL21(DE3). Трансформанты выращивались ОD600 от 0,6 до 0,9 и затем были индуцированы для проявления активности белка слияния при 30° в течение 4 часов при помощи добавления до 0,4 mМ IPTG. Клетки промыли PBS один раз и ресуспендировали в буфере GST (GCB: 50 mМ Tris-HC1 при рН8.0, 200 mM NaC1, 1mM EDTA, 1% Triton X-100, 10 mМ β-меркаптоэтанол, по 5 μg/mL лейпептина, пепстатина и алротинина, и 1 mM PMSF). Клетки были лизированы обработкой ультразвуком на льду (5×30 сек), и лизаты были очищены центрифугированием (15 мин при 12,000 rpm). Белок слияния был восстановлен аффинной хромотографией на гранулах глутатион-Сефароза (Pharmacies). Реакция связывания ДНК содержала 2×105 срm зонда ДНК, 5 μL 4х буфера реакции (100 mM NaC1, 40 mМ HEPES, pH 7.5, 2mM EDTA, 20% глицерол, 140 mM β-меркаптоэтанол), 0,5 μg poly dIdC (Pharmacia), 1 μL 1% Нонидет Р40 и различное количество белков в общем объеме 20 μl. 10 μL реакции были использованы на 4-6% 1/2 ТВЕ нативного геля. Фрагмент BamHI 100 пары основания, содержащий зону -90 35S был помечен α-32 PdCTP в реакции мечения Klenow.
Хотя изобретение было раскрыто в этой патентной заявке ссылками к подробностям представленной реализации изобретения, очевидно, что отдается предпочтение иллюстративному, а не ограниченному в понимании раскрытию изобретения. Можно ожидать, что специалистам в этой области будут часто встречаться изменения в духе изобретения и в пределах заявленных формул.
Описание включает листинг или распечатку последовательности, описывающей нуклеиновая кислоту, включающую основания 89-1060 согласно разделу листинга последовательности SEQ ID NO:1, и изолированный белок, состоящий из аминокислотной последовательности, показанной в разделе листинга последовательности SEQ ID NO:2. На данный листинг последовательности содержатся ссылки в пунктах 1 и 2 формулы изобретения, данный листинг последовательности является неотъемлемой частью описания и формулы заявки.




Изобретение относится к генной инженерии растений. Введение гена nac1, выделенного из Arabidopsis, в растение влияет на регуляцию развития семядоли и бокового корня. Сверхэкспрессия этого гена может привести к получению более крупного растения, растения с более крупными корнями и большим количеством боковых корней, чем у дикорастущих аналогов. 7 с. и 9 з.п. ф-лы, 8 ил.
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| Plant journal | |||
| v | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| RU 95122394 A, 10.11.1997. | |||

