Настоящее изобретение касается идентификации нового подсемейства адгезивных молекул сосудов и модуляции функций таких молекул с целью лечения различных заболеваний.
В процессе эмбрионального и раннего постнатального развития эндотелиальные клетки пролиферируют и дифференцируются с образованием новых кровеносных сосудов за счет процессов васкулогенеза и ангиогенеза. У взрослых организмов эндотелий определяет гематотканевый барьер и состоит из неделящихся (покоящихся) клеток. Такие поляризованные клетки соединены друг с другом с помощью плотных контактов и контактов с помощью адгезинов с образованием непрерывного клеточного слоя. Функции эндотелиального слоя заключаются в поддержании тканевого гомеостаза, фибринолизе, свертывании, подержании тонуса сосудов и перемещении лейкоцитов. Все эти свойства находятся под контролем тонкой настройки экспрессии и функций адгезивных молекул.
Такие патологические ситуации, как воспаление, рост опухолей, образование ран или ангиогенез, приводят к временным изменениям числа и функций адгезивных молекул в эндотелии сосудов, и это обусловливает изменение гомеостаза сосудов. Как пример, опухоли приводят к возрастанию локальной концентрации ангиогенных факторов, которые индуцируют переключение неделящихся покоящихся эндотелиальных клеток в пролиферирующий эндотелий. Ангиогенное переключение индуцируется рядом факторов, включая IL-8, эпидермальный фактор роста (EGF), фактор роста сосудистого эндотелия (VEGF), растворимый VCAM-1, основной фактор роста фибробластов (bFGF) и фактор некроза опухолей (TNF). Как результат, эндотелиальные клетки существующих сосудов разрушают внеклеточный матрикс (ЕСМ) и внедряются в окружающую ткань, что приводит к васкуляризации опухолей.
В процессе ангиогенного переключения экспрессия генов эндотелия претерпевает модификации. Например, обработка эндотелиальных клеток bFGF или TNFα обусловливает четырехкратное усиление экспрессии интегрина α vβ 3 - адгезивной молекулы, участвующей в миграции эндотелиальных клеток. Кроме того, ангиогенное переключение модифицирует воспалительный ответ эндотелия, что приводит к аномальной миграции лейкоцитов по направлению к опухолям. В норме лейкоциты покидают кровяное русло путем прикрепления к эндотелию и миграции через него. Такие механизмы имеют место в многоступенчатом процессе, который вовлекает селектины, интегрины и адгезивные молекулы из суперсемейства иммуноглобулинов.
Было показано, что в эндотелии, ассоциированном с опухолями, VCAM, ICAM и селектины подвержены негативной регуляции. Негативная регуляция данных адгезивных молекул может представлять собой механизм, с помощью которого опухоли избегают инвазии цитотоксическими клетками иммунной системы.
Объектом настоящего изобретения является поиск новых адгезивных белков из суперсемейства иммуноглобулинов (Ig-Sf), которые регулируются на транскрипционном уровне в эндотелии под влиянием опухолей.
Следующим объектом настоящего изобретения является установление молекул, происходящих от новых адгезивных белков, для использования для лечения по различным показаниям, таким как, например, опухоли и воспаление.
В исследованиях, приведших к настоящему изобретению, экспериментальную мышиную модель использовали для идентификации транскриптов, регулируемых в ходе совместного культивирования линии эндотелиальных клеток с меланомными клетками. Для ограничения стратегии скрининга на адгезивные молекулы из состава Ig-Sf был разработан новый способ представления РНК, названный “направленным дифференциальным представлением”. Новое в модифицированном способе представления связано с использованием только одной группы вырожденных праймеров. Как будет показано далее в примерах, неожиданно было обнаружено, что это обеспечивает существенный уровень специфичности.
Более конкретно частично вырожденные праймеры (в данном случае уровень вырожденности находится между 2048 и 4096 разных праймеров в составе одного набора), сконструированные так, чтобы иметь мишенью консервативные последовательности, найденные в доменах С2 членов Ig-Sf, были использованы для осуществления полимеразной цепной реакции (ПЦР) в соответствии со способом направленного дифференциального представления (Samaridis & Colonna, 1997, Eur. J. Immunol., 27, 660-665).
Основываясь на этих данных, настоящее изобретение представляет способ специфичной идентификации дифференцированно экспрессированных ДНК-последовательностей, включающий использование обратнотранскриптазной ПЦР с дифференцированным представлением, в которой используют один набор частично или полностью вырожденных праймеров, специфичных в отношении гена-мишени. Одним основным ограничением стандартной стратегии представления РНК является отсутствие специфичности данного метода. С целью повышения такой специфичности авторы изобретения при проведении поиска других адгезивных молекул использовали вырожденные праймеры, направленные на последовательности, которые кодируют молекулы, включающие домены С2. Этого удалось достичь путем сопоставления доменов С2 из нескольких адгезивных молекул Ig-Sf и идентификации линейного консенсусного аминокислотного мотива, окружающего остаток цистеина, участвующего в организации структуры домена С2: Y-(RQYS)-C-х-A-S-N-х2-G. В более общем смысле такой способ также может быть использован при поиске других последовательностей, в котором обратная трансляция одной или большего числа наиболее часто встречающихся консенсусных последовательностей используется для конструирования вырожденных праймеров для дифференцированного представления.
Способ позволил идентифицировать транскрипт, негативно регулируемый в эндотелиальных клетках конфлюентностью в присутствии меланомных или карциномных клеток. кДНК кодировала новую молекулу из состава Ig-Sf с необычными структурными свойствами, которая была обозначена как CRAM-1 ("Confluency Regulated Adhesion Molecule" - "регулируемая конфлюентностью адгезивная молекула"). Недавнее описание структурно родственной молекулы JAM, вовлеченной в трансмиграцию лейкоцитов, подтвердило существование нового семейства адгезивных молекул, прототипами для которого являются JAM и CRAM-1. Сравнение с последовательностями из базы данных по маркерам EST дополнительно позволило клонировать CRAM-2 - третий член данного молекулярного семейства. На фиг.1 показаны последовательности кДНК мыши, кодирующие белки CRAM-1 и CRAM-2. В настоящей заявке обозначения JAM и JAM-1, CRAM-1 и JAM-2, а также CRAM-2 и JAM-3 использованы взаимозаменяемо.
Сравнительное тканевое распределение транскриптов, кодирующих JAM, CRAM-1 и CRAM-2, показало преобладающую экспрессию этих молекул в эндотелиальном и эпителиальном компартментах, подтверждая их роль в поддержании межклеточных контактов. Такие межклеточные взаимодействия у покоящихся эндотелиальных клеток регулируют пропускную способность сосудов, клеточный цикл и трансмиграцию лейкоцитов сквозь стенку эндотелия.
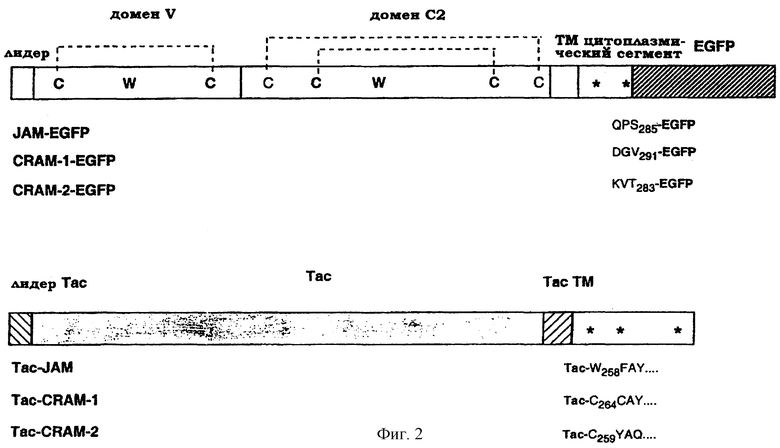
Для дальнейшего определения функции и взаимодействия данных трех молекул был применен молекулярный подход. В связи с этим были сконструированы химерные молекулы, включающие метку FLAG и последовательность усиленного зеленого флуоресцирующего белка (EGFP), образующего гибрид с растворимой или связанной с мембранной формой CRAM-1, CRAM-2 или JAM (обобщено на фиг.2). При трансфекции в клеточные линии EGFP-химерные продукты CRAM-1 и JAM оказывались в областях контактов между клетками: это подтверждает участие данных молекул в межклеточных связях. Напротив, CRAM-2 был более широко распределен по клеточной поверхности. Более того, растворимая конструкция CRAM-1 блокировала миграцию лейкоцитов через эндотелий in vitro, в то время как растворимый JAM проявил лишь незначительный подобный эффект. Взятые вместе, эти данные подтвердили главенствующую роль данного нового подсемейства адгезивных молекул в поддержании целостности сосудов и функционировании эндотелиального слоя.
Основываясь на полученных данных, настоящее изобретение представляет новое средство для противодействующих медицинских показаний, таких как хроническое воспаление и развитие опухоли, с использованием агентов, основанных на полипептидах CRAM.
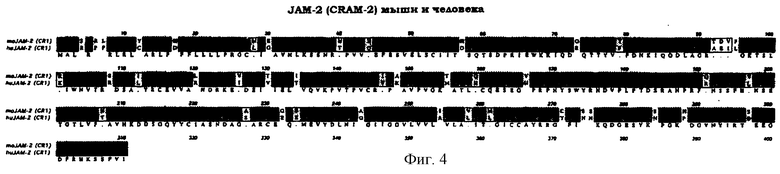
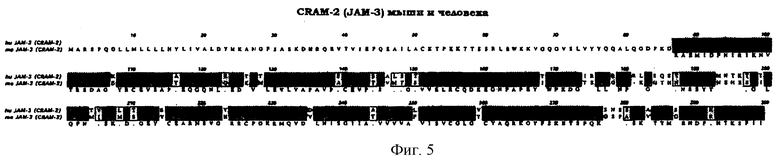
Более конкретно, настоящее изобретение касается полипептида в изолированной форме, относящегося к подсемейству суперсемейства иммуноглобулинов человека, причем данный полипептид проявляет по крайней мере 70%-ный уровень гомологии последовательности с аминокислотной последовательностью мышиных регулируемых конфлюентностью адгезивных молекул 1 или 2 (CRAM-1 или CRAM-2) в соответствии с показанным в верхнем и втором рядах на фиг.3, соответственно. На фиг.4 и 5 показано сопоставление аминокислотных последовательностей у JAM-2 (CRAM-1) и JAM-3 (CRAM-2) мыши, соответственно.
Полипептиды CRAM, найденные в организме человека или животного, являются маркерами растущих клеток. Экспрессия CRAM позитивно регулируется в клетках, являющихся растущими.
В настоящей заявке описываются два новых мышиных полипептида, которые являются членами данного семейства. Основываясь на информации о последовательности данных полипептидов, другие члены семейства могут быть идентифицированы с помощью хорошо известных методов, таких как ПЦР, перекрестная гибридизация библиотек ДНК, перекрестная реактивность антител.
Информация о последовательности может относиться либо к аминокислотной последовательности, либо к нуклеотидной последовательности, кодирующей аминокислотную последовательность.
Таким образом, изобретение, в частности, касается соответствующего полипептида человека, включающего по существу аминокислотную последовательность, показанную на фиг.6В, или аминокислотную последовательность, которая гомологична ей по крайней мере на 70%.
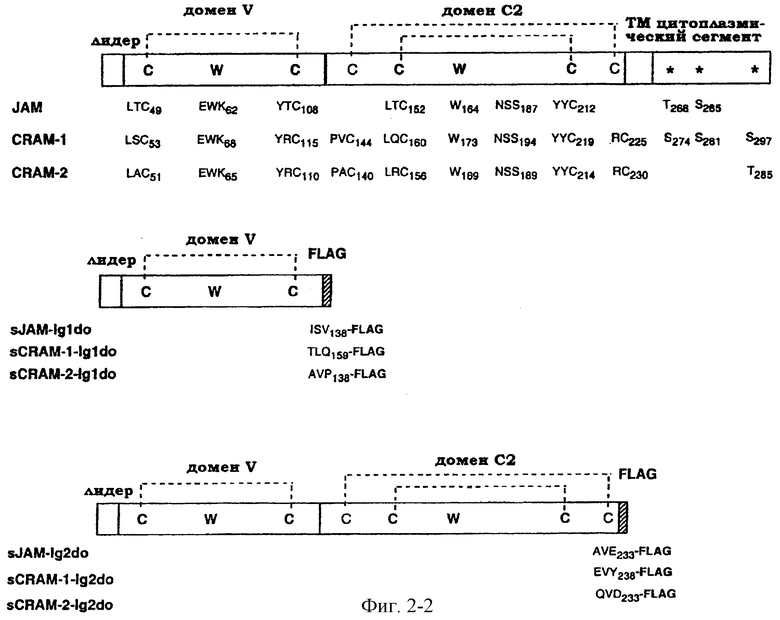
Помимо использования информации о последовательности у двух белков CRAM, описанных здесь, для идентификации других членов семейства у других видов, таких как человек, два белка и соответствующие им члены семейства также могут быть использованы для получения производных молекул, таких как антитела, специфичные в отношении (поли)пептидов по настоящему изобретению, или рекомбинантные эквиваленты белков, необязательно в растворимой форме, или пептиды, включающие по крайней мере сегмент аминокислотной последовательности данных полипептидов. Подходящими сегментами аминокислотной последовательности являются, в частности, внеклеточные домены - VC2, и проксимальная по отношению к мембране цитоплазматическая последовательность -A-[Y,Q]-[R,S]-[R,K]-G-[C,Y]-F.
В дополнение к антителам и производным в форме (поли)пептидов настоящее изобретение также касается поли- и олигонуклеотидов, характеризующихся последовательностью, которая кодирует полный полипептид или его часть, причем этот полипептид имеет аминокислотную последовательность, которая по крайней мере на 70% гомологична аминокислотной последовательности белков CRAM-1 или CRAM-2, описанных в данном тексте. Более конкретно настоящее изобретение касается нуклеотидных последовательностей, которые по крайней мере на 70%, предпочтительно по крайней мере на 80%, более предпочтительно по крайней мере на 90%, наиболее предпочтительно по существу на 100% гомологичны последовательности ДНК CRAM-1 человека в соответствии с показанным на фиг.6.
Такие поли- или олигонуклеотиды могут являться, например, РНК или ДНК, и могут быть праймерами, зондами, антисмысловыми РНК и т.п.
Все такие молекулы могут быть использованы для модуляции функций исходных полипептидов, обнаруживаемых в организме человека или животного, или для диагностики.
С помощью антител может быть подавлен ангиогенез, например, в опухолях. Их можно использовать в качестве направляющих молекул для клеток, несущих полипептиды CRAM. Антитела могут действовать сами по себе или могут объединяться с другими молекулами, такими как токсины, радиоактивные метки, флуоресцентные метки, ферментные метки, активируемые освещением метки, а также липосомы, клетки и т.п.
Помеченные антитела, в частности, пригодны для целей диагностики с помощью антител, т.е. их можно использовать для выявления участков ангиогенеза в растущей опухоли. Кроме того, антитела, присоединенные к токсинам или радиоактивным молекулам, могут быть использованы для направленного уничтожения опухоли изнутри путем атаки, направленной на (растущие) сосуды в опухоли.
Было установлено, что молекулы типа CRAM не выявляются в нормальной сосудистой системе, за исключением венул лимфатической системы и венул с плотным эндотелием (HEV) в лимфатических органах, таких как лимфатические узлы и пейеровы бляшки. Преимущество этого состоит в том, что направленность, например, антител к CRAM, может быть высокоспецифичной в отношении, например, опухолевых клеток, что тем самым исключает нежелательные побочные эффекты.
Более того, (поли)пептиды также могут связываться с молекулой новоообразующихся сосудов, тем самым стимулируя или подавляя ангиогенез.
Растворимый (поли)пептид, обладающий по существу такой же аминокислотной последовательностью, что и полипептиды CRAM, может быть использован для лечения воспалительных реакций в эндотелии сосудов. В связи с настоящим изобретением было установлено, что миграция лейкоцитов через эндотелий может быть подавлена действием sCRAM-1-IG2Do или моноклональными антителами, специфичными в отношении CRAM-1. Такие и сходные молекулы могут быть, следовательно, использованы для подавления или стимуляции иммунологической реакции, такой как реакция, сопровождающая воспаление.
Специфичная экспрессия молекулы на сосудистых клетках HEVs in vivo, которые связаны с миграцией лимфоцитов, является свидетельством стимулирующего действия CRAM на миграцию лимфоцитов или пропускную способность сосудов. Такой эффект, следовательно, может быть обусловлен модуляцией молекул, которые в норме вовлечены в выстилку сосудистого ложа (CRAM-1, CRAM-2, JAM, РЕСАМ, VE-кадгерин). Эта информация служит основой для других применений настоящего изобретения, связанных с регуляцией межэндотелиальных связей за счет доставки рекомбинантных (поли)пептидных молекул CRAM по настоящему изобретению или моноклональных антител, специфичных для CRAM-1.
Антитела к CRAM также могут быть использованы для блокирования межклеточных взаимодействий у растущих клеток. Это приводит к дезорганизации межклеточных контактов, которые в норме являются необходимыми для выполнения кровеносными сосудами барьерной функции. Эти данные могут быть использованы для повышения пропускной способности растущих сосудов с целью улучшения доставки лекарственных средств в конкретные места, такие как растущие опухоли, постменструальная матка и т.п. Дезорганизацию межклеточных контактов можно, следовательно, использовать для блокировки развития опухолевых клеток, несущих антиген, таких как ангиомы (опухоли, происходящие из эндотелия сосудов) или некоторые быстрорастущие карциномы.
Использование для целей диагностики может быть осуществлено с помощью меченных антител, но также и меченных олигонуклеотидов, которые комплементарны ДНК или мРНК CRAM, обнаруживаемым в эндотелиальных клетках, экспрессирующих белок (белки) CRAM.
Настоящее изобретение далее будет проиллюстрировано следующим примером, в котором делаются ссылки на сопровождающие чертежи, на которых показано следующее.
Фиг.1. Последовательность кДНК мыши, кодирующей белки CRAM-1 и CRAM-2. muCRAM-1 была субклонирована в вектор pcDNA3 и секвенирована с использованием праймеров Sр6 и Т7. muCRAM-2 была получена в качестве клона IMAGE из библиотеки ЕSТ-маркеров (депозитарные №№АА690843 и W80145) и секвенирована в составе вектора pT7T3-DPac с использованием праймеров Т7 и Т3.
Фиг.2. Схематическое представление молекулярных средств, используемых в примере. Структура и важные остатки в новом семействе показаны на верхней схеме (a). Звездочки указывают на предполагаемые сайты фосфорилирования в цитоплазматическом сегменте трех молекул. Второй канонический остаток Суs домена С2 в последовательности JAM пропущен. Различные химерные молекулы представлены внизу (B) с указанием на положение и окружающие остатки сайтов слияния. На белом фоне показаны сегменты молекул, происходящих от последовательностей JAM, CRAM-1 или CRAM-2.
Фиг.3. Сопоставление аминокислотных последовательностей CRAM-1 и CRAM-2. Пробелы показаны пунктиром.
Фиг.4. Сопоставление CRAM-1 (JAM-2) мыши и человека.
Фиг.5. Сопоставление CRAM-2 (JAM-3) мыши и человека.
Фиг.6. Последовательность нуклеиновой кислоты CRAM-1 человека (верхняя схема), полная аминокислотная последовательность CRAM-1 человека (средняя схема) и частичная аминокислотная последовательность CRAM-2 человека.
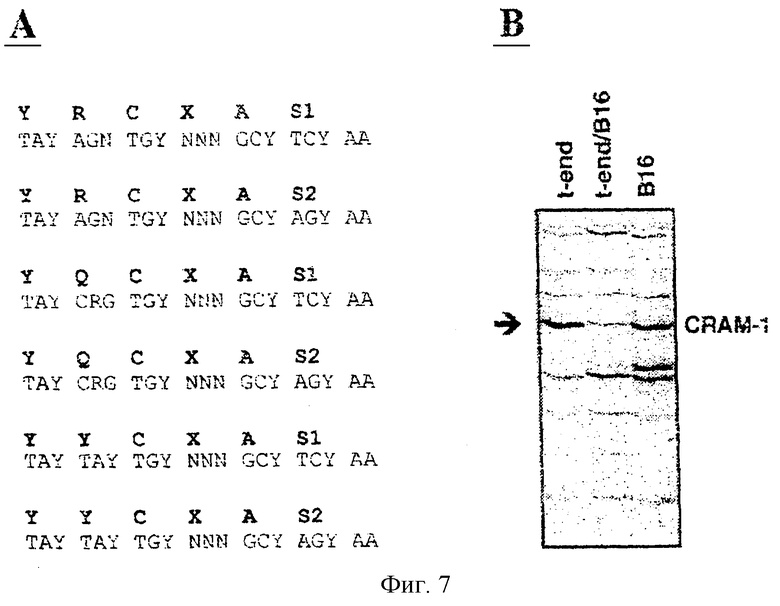
Фиг.7. Направленное дифференцированное представление с использованием вырожденных праймеров. (А): Показаны нуклеотидные последовательности праймеров для ПЦР, кодирующие последовательности, присутствующие в доменах С2 Ig. Два праймера кодируют одну и ту же последовательность благодаря кодонам, кодирующим остаток Ser. Уровень вырожденности составляет 4096 разных форм праймеров, кодирующих YRCXAS, и 2048 форм для других праймеров. (В): Показано представление радиоактивных ПЦР-продуктов, полученных с праймерами YYCXAS1. Дорожки соответствуют представлению разогнанного электрофорезом ПЦР-продукта на материале кДНК, полученной из линии эндотелиальных клеток t-end (дорожка "t-end"), линии меланомных клеток В16 (дорожка "В16") или совместной культуры этих двух клеточных линий (средняя дорожка). Стрелка указывает на интересующий ПЦР-продукт, полученный для негативно регулируемого транскрипта CRAM-1 в условиях совместного культивирования.
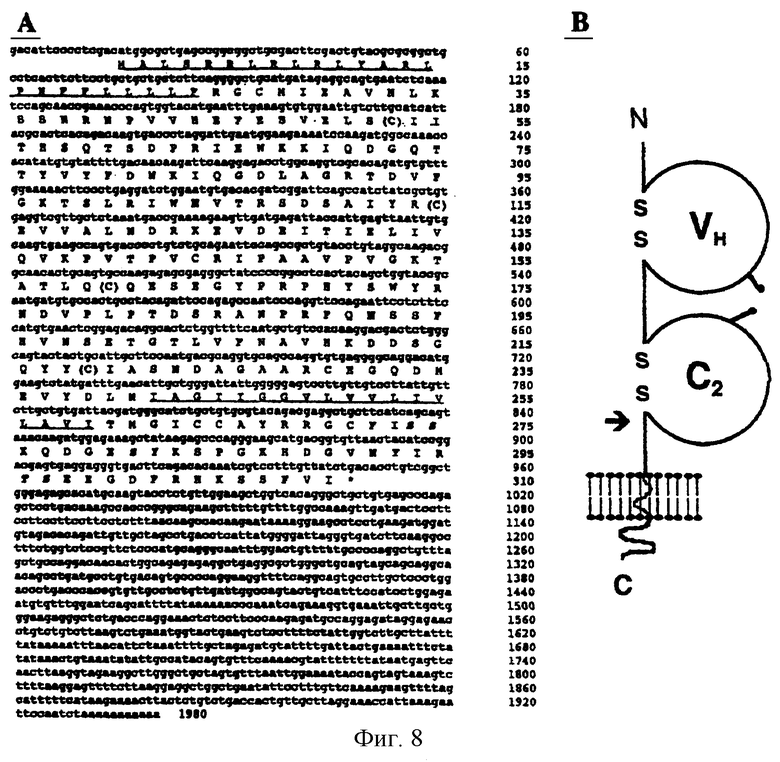
Фиг.8. (А) Нуклеотидная и расшифрованная аминокислотная последовательности кДНК регулируемой конфлюентностью адгезивной молекулы-1 (CRAM-1). Предполагаемые гидрофобные сигнальный пептид (первый) и трансмембранный сегмент (второй) подчеркнуты. Отмечены предполагаемые сайты N-гликозилирования (перечеркнуты), остатки цистеина, предположительно образующие дисульфидные связи (в скобках) и остатки Ser/Thr/Tyr в предполагаемых сайтах фосфорилирования (жирный шрифт). (В) Структурная модель белка CRAM-1 мыши. Внеклеточный сегмент показывает VH и С2-подобный иммуноглобулиновый домен с двумя предполагаемыми сайтами N-гликозилирования. Стрелка указывает на участок, являющийся мишенью для частично вырожденных праймеров (YYCXAS1), использованных для направленного дифференциального представления.
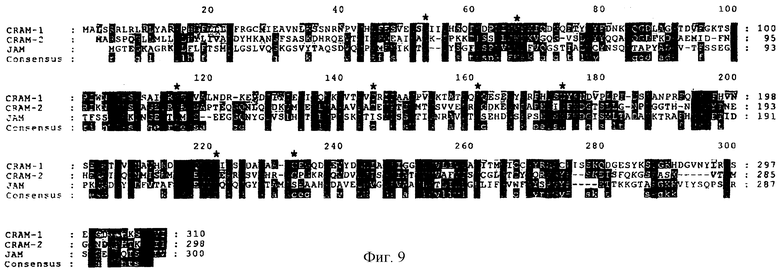
Фиг.9. Сопоставление последовательностей белков JAM, CRAM-1 и CRAM-2 мыши. Идентичные остатки взяты в прямоугольник черного цвета, а гомологичные остатки затенены серым. Общий уровень идентичности между CRAM-2 и CRAM-1 составляет 36%, между JAM и CRAM-1 - 31% и между JAM и CRAM-2 - 33%; соответствующие уровни гомологии - 52%, 52% и 49%. Пробелы в последовательностях показаны пунктиром. Консервативные канонические остатки (Суs и Тrр) в доменах V и С2 отмечены звездочками.
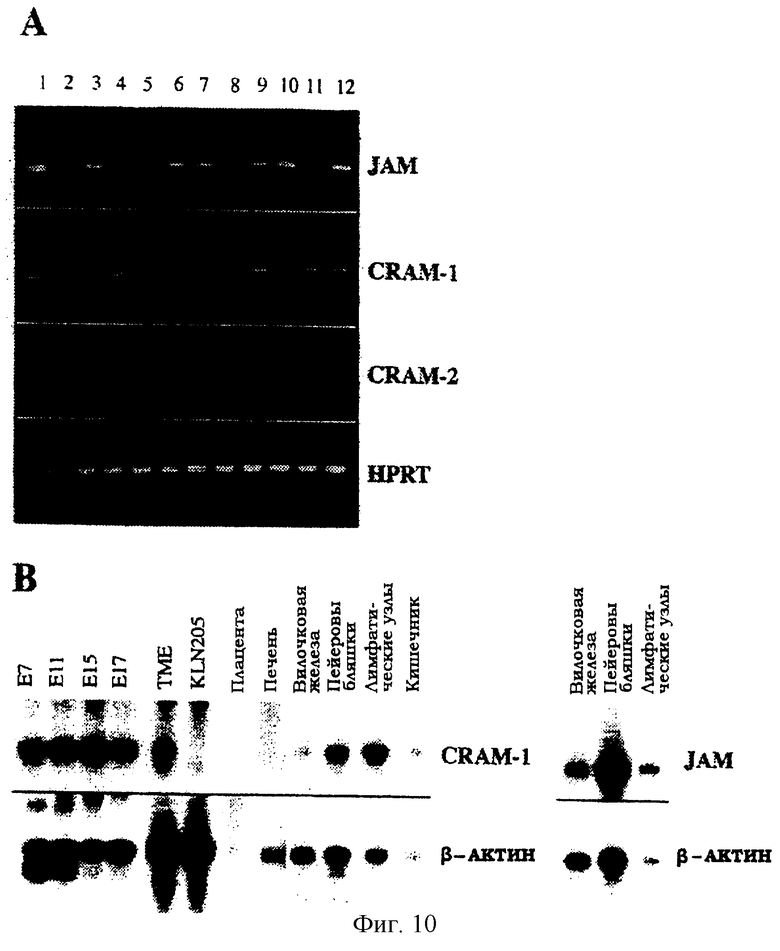
Фиг.10. Экспрессия транскриптов, кодирующих JAM, CRAM-1 и CRAM-2, выявленная методом ПЦР с ревертированием в различных линиях (А) или выявленная с помощью Нозерн-блоттинга в различных тканях (В). (А): ПЦР с ревертированием проводят на материале кДНК, происходящей от линии эндотелиальных клеток, обработанных TNF (дорожки 2 и 11 соответствуют клеткам t-end, обработанным TNF) или не обработанных (дорожки 3, 4, 6, 7, 9, 12 соответствуют b-end.5, e-end.2, t-end V++L-, t-end VlowL++, TME и t-end, соответственно). Дорожки 5 и 10 соответствуют линиям опухолевых клеток В16 (меланома) и KLN205 (карцинома). Дорожка 8 соответствует линии нетрансформированных эпителиальных клеток вилочковой железы МТЕ4-14. Дорожка 1 представляет позитивный контроль для амплификации JAM, CRAM-1 и CRAM-2 по плазмидам, включающим клонированные кДНК. (В): Авторадиограмма гибридизации 32Р-меченного зонда на Нозерн-блотте мыши. Слева показаны зонды, использованные при каждой гибридизации. Сигналы гибридизации для JAM и CRAM-1 выявляют при размере 2 тысячи пар нуклеотидов (т.п.н.).
Фиг.11. Локализация JAM-2 и JAM-1 в сформировавшихся межклеточных контактах. А: иммуноцитохимический анализ проводили на клетках TME, зафиксированных параформальдегидом, с использованием антител к JAM-2 (а) или JAM-1 (b). Стрелки указывают на конкретную локализацию белков в межклеточных контактах. Масштабная линейка 10 мкм. В: Химерные молекулы JAM-2-EGFP (а) и JAM-1-EGFP (с) были специфически локализованы в межклеточных контактах между трансфицированными клетками. Обогащение по рекомбинантным белкам EGFP не было выявлено в контактах трансфицированных и нетрансфицированных клеток (короткие стрелки). Масштабная линейка - 20 мкм. С: Иммунопреципитация JAM-2 после поверхностного биотинилирования эндотелиальных клеток ТМЕ. Антитела к РЕСАМ (дорожка 1) и к JAM-1 (дорожка 2) были использованы в качестве позитивного и негативного контролей, соответственно, для иммунопреципитации с помощью антитела CRAM-XIXH36 (дорожка 3). Справа показаны молекулярные массы. D: иммунопреципитация рекомбинантных белков EGFP из трансфицированных клеток СНО. Антитела к JAM-2 (дорожки 2, 3, 6) и к JAM-1 (дорожки 1, 4, 5) использовали для иммунопреципитации биотинилированных лизатов на материале нетрансфицированных (дорожки 1 и 2) клеток или клеток СНО, трансфицированных JAM-1-EGFP (дорожки 3 и 4) или JAM-2-EGFP (дорожки 5 и 6). Молекулярные массы указаны справа.
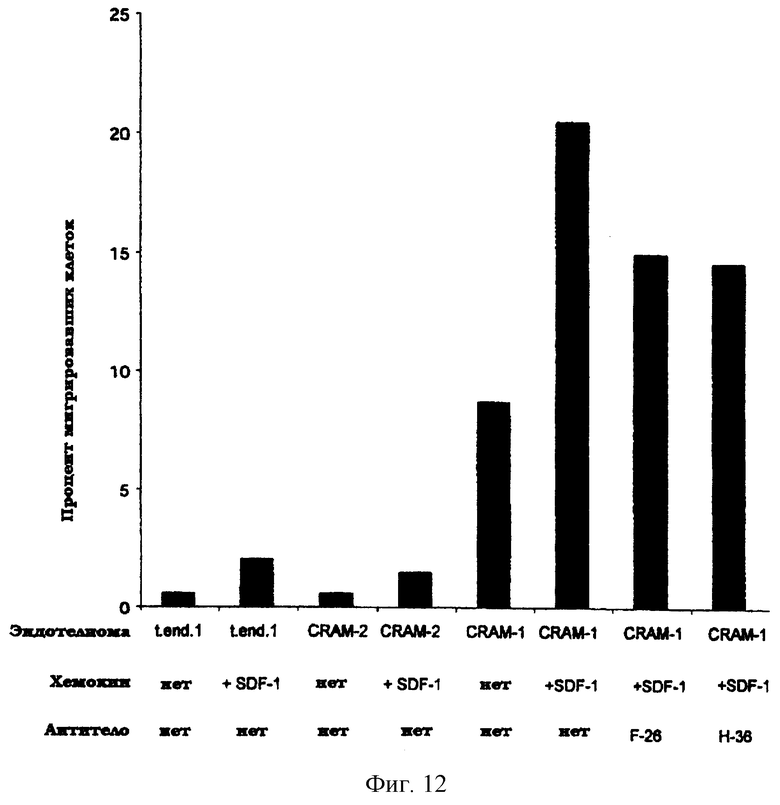
Фиг.12. Миграция спленоцитов через монослои TNF-активированных эндотелиом в присутствии или отсутствие хемокина SDF-1. Использовали три эндотелиомы: t.end.1 дикого типа или t.end.1, трансфицированная кДНК, кодирующей CRAM-1 или CRAM-2. Два моноклональных антитела были протестированы по их способности влиять на трансмиграцию - F-26 или Н-26: оба являются крысиными моноклональными антителами типа IgGI, специфичными в отношении CRAM-1 мыши.
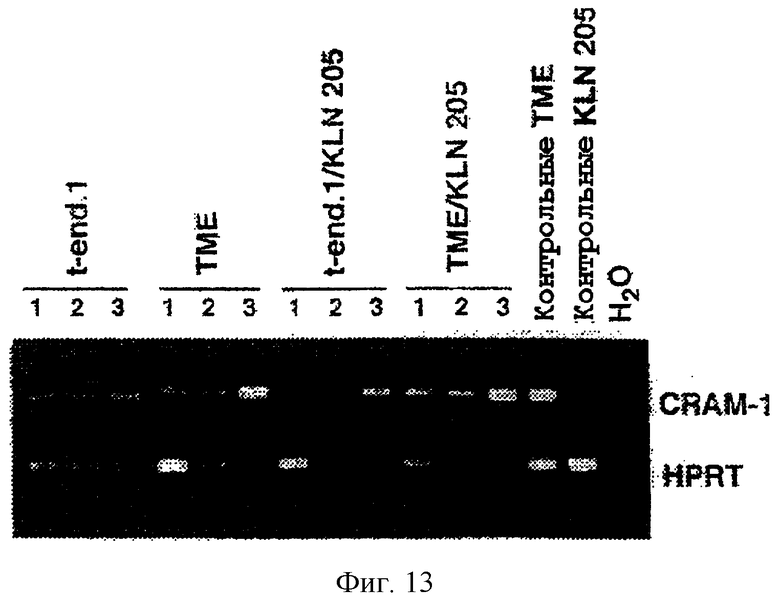
Фиг.13. Регуляция CRAM-1 как функция конфлюентности. Полуколичественную ПЦР проводили с использованием смеси праймеров, специфичных для кДНК HPRT и CRAM-1. Реакции ПЦР разгоняли в 1,2%-ном агарозном геле, который окрашивали бромистым этидием. Дорожки 1, 2 и 3 соответствуют уровням конфлюентности 100%, 50% и 10%, соответственно. Был выявлен более слабый сигнал для CRAM-1 при 100%-ной конфлюентности (дорожка 1). Указаны условия культивирования линий эндотелиальных клеток (t.end.1 и ТМЕ) самих по себе или в смеси с линией опухолевых клеток KLN-205.
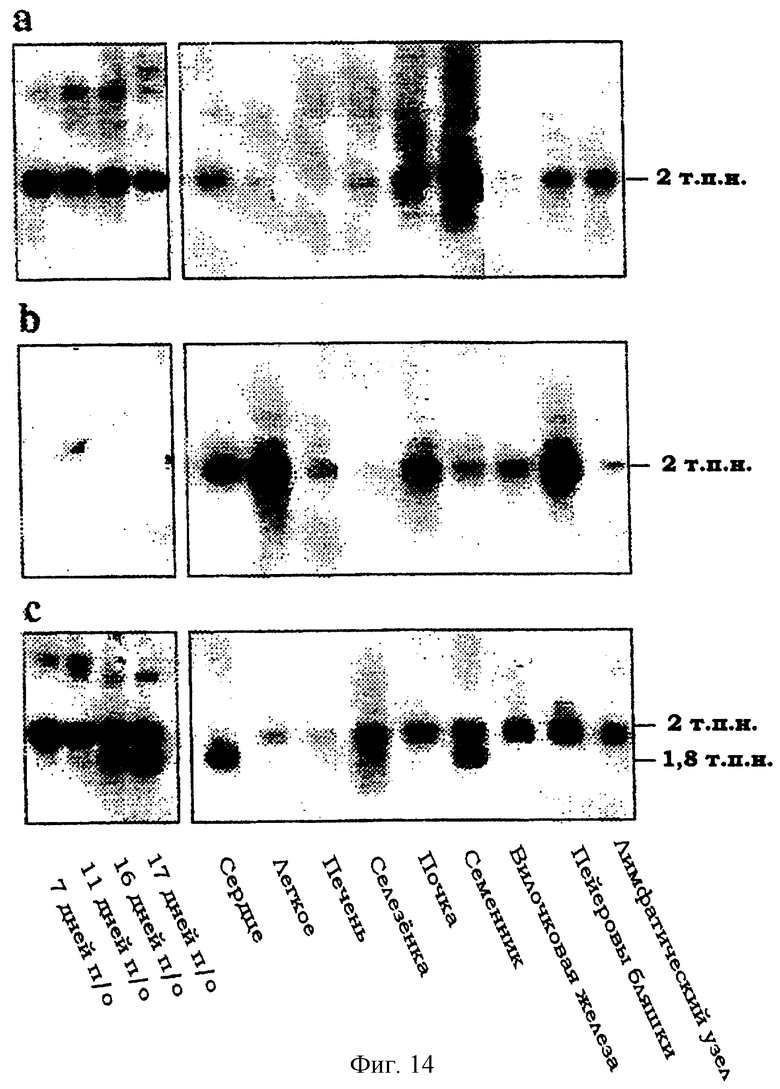
Фиг.14. Анализ методом Нозерн-блоттинга транскриптов JAM-2 (a), JAM-1 (b) или β -актина (с) в тканях мыши. Показаны параметры препаратов эмбриональных мРНК после оплодотворения (п/о) и мРНК от взрослых особей. Справа показана величина сигнала гибридизации.
Фиг.15. Иммуногистологический анализ экспрессии JAM-2, JAM-1, ZO-1 и РЕСАМ. Серийные срезы почки (a-d) или срезы брыжеечного лимфатического узла (е-l) окрашивали с помощью антител к JAM-2 (a, e, i), к JAM-1 (b, f, j), к ZO-1 (с, g, k) или РЕСАМ (d, h, l). Для каждой серии изображений (a-d, e-h и i-l) использовали идентичные установки видеосистемы CCD.
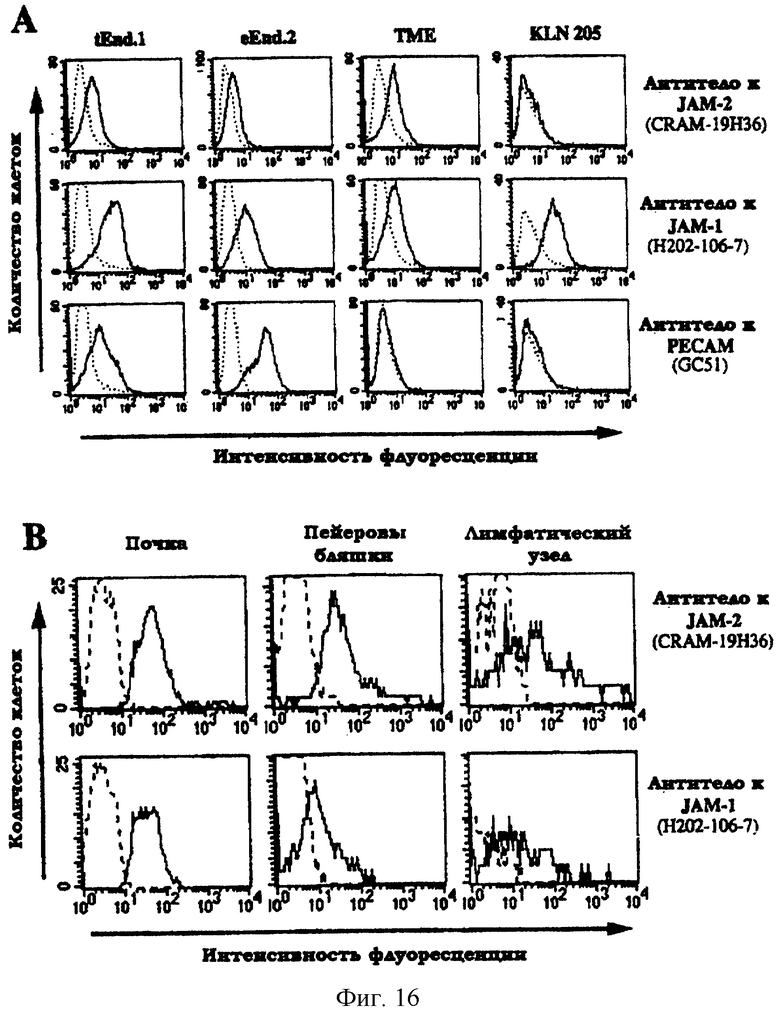
Фиг.16. Экспрессия JAM-2 в эндотелиальных клетках. А: цитофлуорометрический анализ экспрессии JAM-2, JAM-1 и РЕСАМ в линиях эндотелиальных клеток (tEnd.1, eEnd.2 и ТМЕ) или линии плоскоклеточной карциномы (KLN-205). Показанные пунктиром графики представляют негативные контроли, полученные с использованием антитела, специфичного для CD4. В: цитофлуорометрический анализ JAM-2 на свежевыделенных эндотелиальных клетках. Указанные органы диспергировали путем обработки коллагеназой/диспазой, окрашивали DilAc-LDL, CD31, анти-JAM-2 или анти-JAM-1 в соответствии с указанным. Гистограммы были получены путем изъятия популяции эндотелиальных клеток, позитивных по DilAc-LDL (FL-2) и CD31 (FL-3). Негативные контроли были получены путем исключения первичных моноклональных антител к JAM-1 или JAM-2.
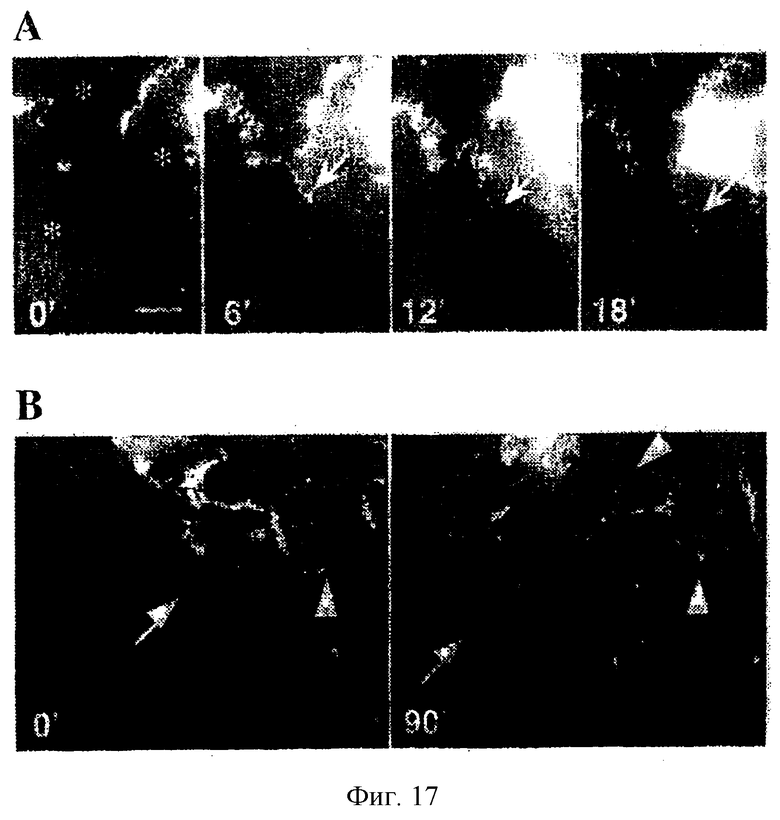
Фиг.17. Локализация JAM-2-EGFP в процессе формирования межклеточного контакта. Отдельные изображения флуоресценции получали каждые 3 минуты в течение 1 часа в процессе формирования монослоя клеток СНО, трансфицированных JAM-2-EGFP. Показаны изображения, полученные в первые 18 минут. Во время 0 звездочки указывают на три клетки, присутствующие в поле зрения. Во время 6, 12 и 18 минут стрелки указывают на перемещение JAM-2-EGFP в область новообразующегося межклеточного контакта. (В): локализация JAM-2-EGFP после повреждения. Стрелки указывают на поврежденную сторону, а короткие стрелки указывают на мембранные отростки, богатые JAM-2-EGFP. На изображениях указано прошедшее время. Масштабная линейка 10 мкм.
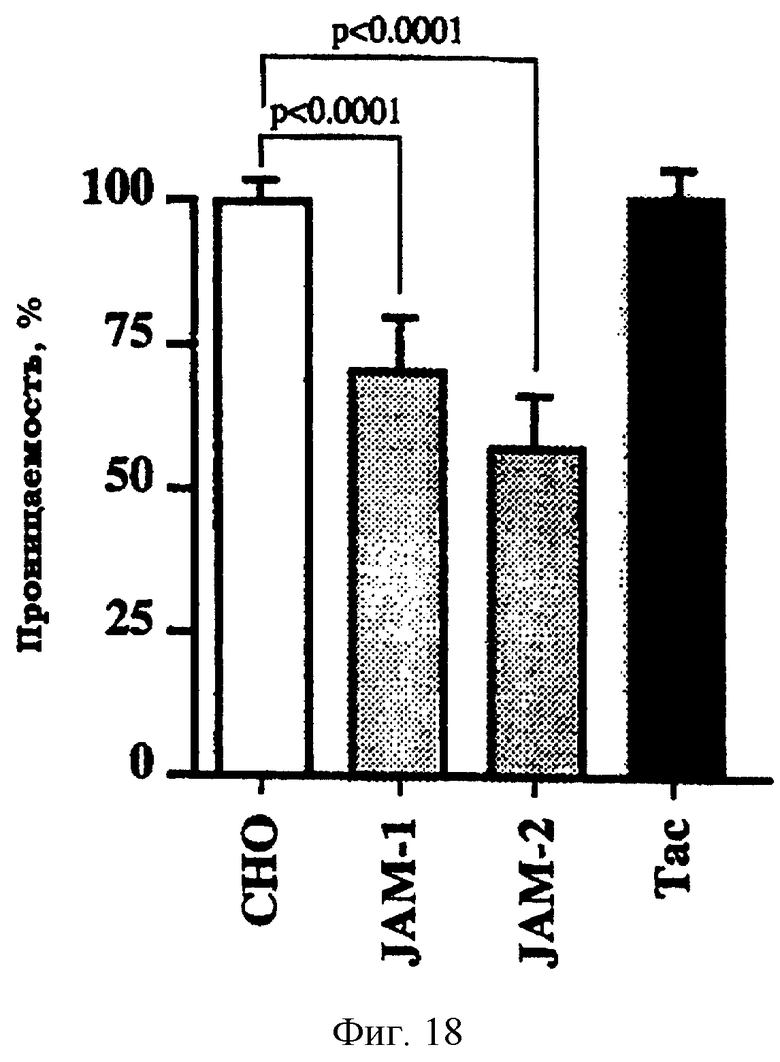
Фиг.18. Экспрессия JAM-2 снижает параклеточную проницаемость. (А): Параклеточную проницаемость оценивали по диффузии FITC-помеченного декстрана через монослои нетрансфицированных клеток СНО, через клетки СНО, трансфицированные Тас (huIL2Rα ) или с использованием указанного EGFP-химерного белка (с JAM-1 или JAM-2). Трансфекция клеток СНО конструкциями JAM-2-EGFP или JAM-1-EGFP приводила к существенному снижению параклеточной проницаемости (соответственно, 57,8±4,9% и 70,8±3,6%: р<0,0001), в то время как трансфекция Тас не влияла сколько-нибудь существенно на параклеточную проницаемость (100,4±4,4%: р=0,9872). Полученные величины нормализованы по нетрансфицированным клеткам СНО.
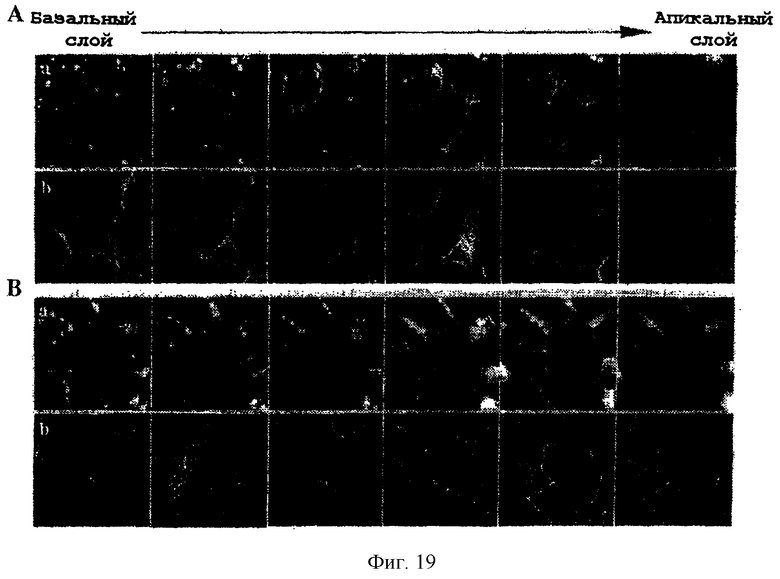
Фиг.19. Направление JAM-2-EGFP (А) и JAM-1-EGFP (В) в уже существующие плотные контакты. Конфлюирующие клетки MDCK, трансфицированные по стабильному типу JAM-2-EGFP (А) или JAM-1-EGFP (В), окрашивали анти-окклудином и антикроличьим-техасским красным. Серии изображений через каждый 0,9 мкм от базального до апикального слоев показаны для EGFP-флуоресценции (а) или окклудинового окрашивания (b). Базальный уровень (слева) был произвольным образом определен так, что в серии изображений уровень плотного контакта приходился на срезы +3,6 мкм и +4,5 мкм (четвертое и пятое изображение слева направо).
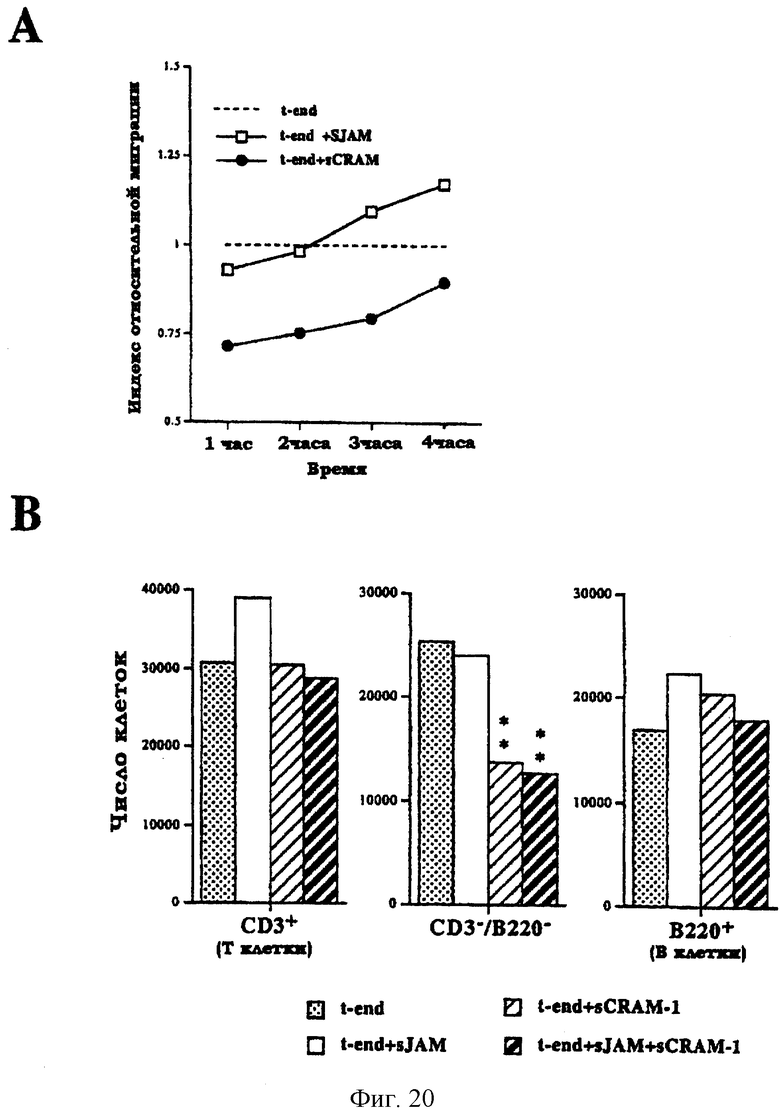
Фиг.20. Влияние растворимых рекомбинантных молекул на миграцию лейкоцитов через эндотелий. (А): трансмиграция выражена как относительный индекс и нормализована по величинам, полученным для необработанной линии клеток t-end (пунктирная линия индекса 1). Показаны результаты, полученные в присутствии 1 мкг sJAM-Ig2do (незакрашенные квадратики) или в присутствии 1 мкг sCRAM-1-Ig2do (закрашенные кружки). Индекс подсчитывают как среднее по пяти независимым экспериментам по трансмиграции. (В): Фенотип трансмигрировавших клеток выражен как число клеток, подсчитанных на основании процентов, определенных с помощью FACS-анализа после окрашивания антителами анти-СD3-FITS и анти-В220-РЕ. Звездочки указывают на стадии эксперимента, характеризовавшиеся достоверным отличием от контроля.
В примерах обозначения JAM и JAM-1, CRAM-1 и JAM-2, равно как CRAM-2 и JAM-3 используются взаимозаменяемо.
ПРИМЕР
МАТЕРИАЛЫ И МЕТОДЫ
Клеточные линии
Линии клеток эндотелиомы вилочковой железы (tEnd.1) и эмбриональной эндотелиомы (eEnd.2) (Williams et al., 1989, Cell, 57, 1053-1063) были предоставлены д-ром W.Risau и д-ром B.Engelhardt (Max Planck Institute, Bad-Nauheim, Германия). Трансформированная вирусом SV40 линия ТМЕ эндотелиальных клеток лимфатического узла была предоставлена д-ром A.Hamann (Harder et al., 1991, Exp. Cell Res., 197, 259-267). Клетки плоскоклеточной карциномы KLN-205, клетки СНО, клетки MDCK и линия миеломных клеток Sp2/0 были получены из Американской коллекции типовых культур (АТСС). Все клетки, за исключением СНО, культивировали в среде DMEM (Gibco BRL, Paisley, Шотландия), дополненной 10% ПТС (РАА Laboratories, Linz, Австрия), 2 мМ глутамина, 100 ед./мл пенициллина и 100 ед./мл стрептомицина (все от Gibco BRL). Клетки СНО культивировали в среде Nut.Mix.F-12 (НАМ), дополненной тем же, что указано выше. Прикрепленные клетки отделяли путем промывки в ФСБ с 0,15 мМ ЭДТА с последующей 5-минутной инкубацией с трипсином-ЭДТА при 37° С.
Представление, клонирование и секвенирование
В эксперименте по совместному культивированию 5× 105 клеток t.End.1 культивировали вместе с 2,5× 104 меланомных клеток В16 F10 в течение 64 часов в чашках Петри диаметром 10 см. В качестве контроля 5× 105 клеток t.End.1 и 2,5× l04 клеток В16 F10 культивировали раздельно при тех же условиях, в результате чего конфлюирующие монослои получали через 64 часа. Тотальную РНК экстрагировали непосредственно на чашках Петри с помощью реагента Trizol в соответствии с инструкциями производителя (Gibco BRL, Paisley, Шотландия). кДНК получали на материале 5 мкг тотальной РНК, используя олиго-дТ-праймер (16-нуклеотидный) и обратную транскриптазу Superscript (Gibco BRL, Paisley, Шотландия). Количество и качество кДНК проверяли путем проведения 27 циклов ПЦР на 1 мкл кДНК, разведенной 1:5, используя праймеры, специфичные для кДНК гена "домашнего хозяйства" HPRT. Затем дифференциальную ПЦР проводили со следующими вырожденными праймерами: 5'-TAYAGNTGYNNNGCYTCYAA-3', 5'-TAYCRGTGYNNNGCYTCYAA-3' и 5'-TAYTAYTGYNNNGCYTCYAA-3’, - кодирующими наиболее часто встречающиеся аминокислотные последовательности С2-доменов - YRCXAS, YQCXAS и YYCXAS. Использованными условиями ПЦР было следующее: 2 мкл разведенной кДНК; 2,5 мкл 10× буфера для ПУР Goldstar PCR; 2 мкл MgCl2; 2 мкл 0,3 мМ вырожденных праймеров; 0,5 мкл 0,1 мМ dNTP; 0,1 мкл α -33Р-dАТР, 10 мКи/мл (Amersham Pharmacia Biotech,  Швейцария); 15,65 мкл Н2О; 0,25 мкл ДНК-полимеразы Taq Goldstar (Eurogentech, Seraing, Бельгия).
Швейцария); 15,65 мкл Н2О; 0,25 мкл ДНК-полимеразы Taq Goldstar (Eurogentech, Seraing, Бельгия).
Параметры ПЦР были следующими: 45 секунд при 94° С, 90 секунд при 50° С и 45 секунд при 72° С в 40 циклах. Добавляли загрузочный буфер с формамидом/ЭДТА и образцы денатурировали в течение 2 минут при 94° С. Затем ПЦР-продукты разделяли в 6%-ном полиакриламидном геле и анализировали авторадиографически с использованием пленки Kodak OM-Mat. Сравнивали интенсивность полос.
Дифференцированно экспрессированные полосы вырезали из высушенного полиакриламидного геля и фрагменты извлекали кипячением и этанольной преципитацией в соответствии с описанным ранее (Liang & Pardee, 1992, Science, 257, 967-970). Затем ПЦР-продукты повторно амплифицировали с использованием повышенных концентраций dNTPs (0,2 мМ вместо 2 мкМ) без 33P-dATP. Продукты повторной амплификации клонировали в вектор pGem-T Easy (Promega Corp., Wallisellen, Швейцария) в соответствии с описанным ранее (Sambrook, Fritsch & Maniatis, "Molecular Cloning", 2d ed., Cold Spring Harbor Laboratory Press, 1989).
Последовательности нуклеиновых кислот в двух независимых клонах определяли с использованием набора для термосеквенирования с флуоресцентно помеченными праймерами Thermo Sequence Fluorescent Labeled Primer Cycle Sequencing kit (Amersham Pharmacia Biotech, D bendorf, Швейцария) и ДНК-анализаторной системы LI-COR (MWG-Biotech GmbH, Ebersberg, Германия).
bendorf, Швейцария) и ДНК-анализаторной системы LI-COR (MWG-Biotech GmbH, Ebersberg, Германия).
Идентификация JAM-3
Анализ и сравнение последовательностей были осуществлены с помощью средств, доступных из Интернета с сервера ExPASy Molecular Biology Server, а именно Blast, Prosite, Swiss-Prot. Были идентифицированы три различных EST, гомологичных CRAM-1 (депозитарные №№АА726206, АА052463 и АА175925). Ни один из них не кодирует полноразмерный транскрипт и не включает старт-кодон ATG. С учетом этого 5'-кодирующая последовательность была получена с использованием ПЦР-системы 5'-RACE для "быстрой амплификации концов кДНК", версия 2,0, в соответствии с инструкциями изготовителя (Gibco BRL, Paisley, Шотландия).
Три использованных праймера были сконструированы на основе EST-последовательностей следующим образом:
5'-GAGGTACTTGCATGTGCT-3' для синтеза первой цепи, 5'-CGACAGGTGTCAGATAACA-3’ и 5'-CACCCTCCTCACTCGT-3' для двух ПЦР "гнездового" типа. ПЦР-продукт 5'-RACE клонировали в состав вектора pGem-T. Для получения полноразмерной кодирующей последовательности CRAM-1 клонированный ПЦР-продукт 5'-RACE и EST-маркер (депозитарный №АА726206) расщепляли рестриктазами HpaI и NotI и лигировали в вектор pGem-t. Клонирование полноразмерного CRAM-2 было основано на такой же стратегии сравнения последовательностей и методе 5'-RACE. Полноразмерная кДНК, кодирующая CRAM-2, была в конечном счете получена на материале ЕSТ-маркеров депозитарных номеров АА690843 и W80145. Два указанных клона различаются по длине 3'-нетранслируемого участка.
Нозерн-блоттинг
Тотальную мРНК из клеток или тканей экстрагировали с использованием Trizol (Life Technologies AG, Basel, Швейцария) в соответствии с инструкциями изготовителя. Полиаденилированную мРНК экстрагировали из 250 мкг тотального пула РНК с помощью набора Oligotex mRNA Purification kit (Qiagen,  Швейцария). Нозерн-блотт эмбриональной поли-А был приобретен у Clontech (P.H. Stehelin & Cie AG, Basel, Швейцария). Рибозонды были сформированы на материале вектора pcDNA3 (Invitrogen, Leek, Нидерланды): они включали последовательности, кодирующие иммуноглобулиновые домены JAM-1 и JAM-2, или полноразмерную кодирующую последовательность β -актина. Гибридизацию проводили при 62° С в буфере, содержащем 50% формамида. Затем блоты дважды промывали (0,5× SSC, 0,1% ДСН, 67° С) и авторадиографировали на пленку Kodak X-Omat при -80° С.
Швейцария). Нозерн-блотт эмбриональной поли-А был приобретен у Clontech (P.H. Stehelin & Cie AG, Basel, Швейцария). Рибозонды были сформированы на материале вектора pcDNA3 (Invitrogen, Leek, Нидерланды): они включали последовательности, кодирующие иммуноглобулиновые домены JAM-1 и JAM-2, или полноразмерную кодирующую последовательность β -актина. Гибридизацию проводили при 62° С в буфере, содержащем 50% формамида. Затем блоты дважды промывали (0,5× SSC, 0,1% ДСН, 67° С) и авторадиографировали на пленку Kodak X-Omat при -80° С.
Эксперимент по конфлюентности
Исследовали влияние конфлюентности эндотелиальных клеток на уровни мРНК JAM-2. Эндотелиальные клетки ТМЕ в количестве 2× 105 культивировали на культуральных чашках диаметром 6, 10 и 15 см до достижения различных уровней конфлюентности спустя 64 часа, варьировавшихся в диапазоне от 10 до 100%. Число клеток через 64 часа, проверяемое методом исключения трипанового синего с последующим подсчетом, было одинаковым во всех случаях и не было связано с площадью поверхности чашки Петри.
Полуколичественную реакцию ПЦР или Нозерн-блоттинг использовали для определения относительного количества транскрипта в различных условиях. Для выявления транскрипта JAM-2 использовали пару праймеров 5'-GACTCACAGACAAGTGAC-3' и 5’-CACCCTCCTCACTCGT-3’, в результате чего получили амплификационный продукт длиной 750 п.н. В качестве внутреннего контроля следующие праймеры, специфичные для кДНК Hprt, были использованы для амплификации фрагмента длиной 350 п.н.: 5'-СТТССАТАСАССССАСАСТТТСТТС-3' и 5'-GAGGGTAGGCTGGCCTATAGGCT-3'.
Конструирование экспрессирующих векторов
Последовательность, кодирующая EGFP, была субклонирована из вектора pEGFP-1 (Clontech, P.H.Stehelin & Cie AG, Basel, Швейцария) в плазмиду pcDNA3 с использованием сайтов HindIII и NotI, которая в результате получила наименование pcDNA3/EGFP. Рестрикционные сайты с 3'-конца, узнаваемые рестриктазами HpaI и ScaI, обнаруженные в последовательности, кодирующей цитоплазматический домен в составе, соответственно, JAM-2 и JAM-1, использовали для химеризации этих двух последовательностей по N-концу EGFP в векторе pcDNA3 (Invitrogen, Leek, Нидерланды). Вставки, кодирующие JAM-2 или JAM-1, вырезали из pGemt или pRc/CMV с использованием расщепления рестриктазами SacII/HpaI или HindIII/ScaI, соответственно.
Затем кодирующие последовательности клонировали в вектор pcDNA3/EGFP, который расщепляли рестриктазой AgeI, затупляли по концам путем заполнения и далее расщепляли их рестриктазами HindIII или SacII. В результате образуются сайты химеризации по аминокислотным положениям DGV291 для JAM-2 и QPS285 для JAM-1. Трансфекцию клеток СНО осуществляли в соответствии с описанным ранее (Ballestrem et al., 1998, J. Cell Sci., 111, 1649-1658).
Стабильные трансфектанты, использовавшиеся для тестов на проницаемость, были отобраны с помощью культивирования трансфицированных клеток СНО в течение двух недель в культуральной среде, содержащей 1 мг/мл антибиотика G418. Резистентные колонии выделяли и проверяли на интенсивность флуоресценции EGFP методом проточной цитометрии (прибор FACScalibur, Becton Dickinson, Mountain View, CA) и картирования флуоресценции с помощью микроскопа (Axiovert, Zeiss, Oberkochen, Германия).
Микроскопию с покадровым видеоизображением проводили с использованием флуоресцентного микроскопа Axiovert и программы Openlab для сбора и анализа изображения.
Экспрессирующий вектор pcDNA3 для клеток млекопитающих (Invitrogen, Leek, Нидерланды) был модифицирован путем внесения последовательности, кодирующей метку FLAG (G.Wiedle, Dep. Pathology, CMU, Geneva). Конструкции с меткой FLAG, включающие последовательности, кодирующие растворимые формы белков JAM, CRAM-1 и CRAM-2, получали с помощью ПЦР. Во всех случаях прямые праймеры были сформированы для внесения старт-кодона ATG. Обратные праймеры были сконструированы по последовательностям, кодирующим шарнирный сегмент одной из растворимых форм Ig, или по последовательности, кодирующей участок между С2 и трансмембранным доменами у двух растворимых молекул с Ig-доменами. Все обратные праймеры имели 3'-"выступы", включающие XbaI-рестрикционный сайт, необходимый для прямого внутрирамочного клонирования в модифицированный вектор с меткой FLAG. ДНК-полимеразу Pfu использовали в ПЦР с целью исключения частых мутаций (Stratagene, La Jolla, СА, США). ПЦР-фрагменты были затем расщеплены рестриктазой XbaI и клонированы в состав вектора pcDNA3 с меткой FLAG, который расщепляли рестриктазой EcoRI, заполняли "липкие" концы фрагментом Кленова и затем разрезали рестриктазой XbaI.
Реагенты и иммунофлуоресцентный анализ
Использовали следующие моноклональные антитела: анти-PECAM (GC51, IgG2a крысы; EA-3, IgG1 крысы) и анти-JAM (Н202.106.7.4, IgG1 крысы) (Malerque et al., 1998, Mol. Immunol., 35, 1111-1119; Piali et al., 1993, Eur. J. Immunol., 23, 2464-2471).
Панель CRAM-антител, специфичных в отношении JAM-2, была сформирована в лаборатории с использованием стандартных методов и рекомбинантной растворимой молекулы в качестве иммуногена (Aurrand-Lions et al., 1996, Immunity, 5, 391-405). Отобранные гибридомы подвергали скринингу методом ТИФА на выработку антител, специфически распознающих рекомбинантную растворимую молекулу JAM-2. Позитивные клоны дополнительно тестировали на клетках СНО, трансфицированных кДНК JAM-2 (данные не показаны).
Все CRAM-антитела относятся к изотипу IgG1 или IgG2a, за исключением CRAM-25F24, которое представляет подкласс IgG2b. Антитела были очищены на сефарозных колонках с белком-G (Pharmacia Biotech Europe,  , Швейцария) в соответствии с инструкциями изготовителя. Моноклональное антитело CRAM-19H36 использовали для иммунопреципитации, в то время как CRAM-18F26 являлось реагентом, использованным для иммуногистохимического анализа. Сходные результаты были получены с моноклональными антителами CRAM-4H31 и CRAM-17D33.
, Швейцария) в соответствии с инструкциями изготовителя. Моноклональное антитело CRAM-19H36 использовали для иммунопреципитации, в то время как CRAM-18F26 являлось реагентом, использованным для иммуногистохимического анализа. Сходные результаты были получены с моноклональными антителами CRAM-4H31 и CRAM-17D33.
Иммунофлуоресцентный анализ был проведен с использованием вторичных реагентов, соединенных с FITC или техасским красным (Jackson Immunoresearch, Milan AG, La Roche, Швейцария), для цитофлуорометрического и иммуногистохимического анализа, соответственно.
Для иммуногистохимического анализа образцы были зафиксированы в течение 5 минут холодным (-20° С) метанолом. Образцы регидратировали в ФСБ, 0,2% желатине, 0,05% Твин-20, инкубировали в течение ночи с первичными антителами с последующей промывкой и проявляли с помощью подходящих вторичных реагентов, соединенных с техасским красным. Для анализа свежих эндотелиальных клеток диспергирование свежевыделенных тканей осуществляли с использованием обработки коллагеназой/диспазой в соответствии с разработанными процедурами (Kramer et al., 1984, J.Ceil Biol., 99, 692-698). Диспергированные клетки окрашивали в течение 2 часов при 37° С с помощью Dil-ацетилированного LDL (Molecular Probe Europe BV, Leiden, Нидерланды) с последующим окрашиванием анти-СКАМ-19 и козьим антикрысиным FITC-зондом. После трехкратной промывки клетки окрашивали биотинилированным анти-СD31 (Pharmingen) и стрептавидиновым красным-670 (Life Technologies AG, Basel, Швейцария).
Экспрессию JAM-1 или JAM-2 анализировали на материале клеток, позитивных по двум маркерам эндотелиальных клеток: ацетилированного LDL (FL-2) и CD31 (FL-3). Негативные контроли были получены путем исключения первичного антитела.
Иммунопреципитация
Иммунопреципитацию проводили в соответствии с описанным ранее (Aurrand-Lions et al., 1997, Cellular Immunology, 176, 173-179) с использованием 10 мМ буфера Трис-HCl - рН=7,4, 150 мМ NaCI, 0,5% Тритон Х-100, 0,5% NP40, смеси протеазных ингибиторов (Roche Diagnostics Ltd., Rotkreuz, Швейцария) для лизиса. После иммунопреципитации, электрофореза в ДСН-ПААГ и переноса на нитроцеллюлозную мембрану биотинилированные белки были проявлены с использованием стрептавидина, соединенного с пероксидазой (Jackson Immunoresearch), и ECL (Amersham Pharmacia Biotech, Великобритания).
Тесты на проницаемость
Проницаемость определяли с использованием камер Transwell (диаметр 6,5 мм, РС-фильтры, размер пор 0,4 мкм: Costar Corp.). Вкратце, 1× 104 трансфицированных или нетрансфицированных клеток СНО культивировали до конфлюентности на фильтрах, предварительно покрытых в течение 30 минут 0,2%-ным желатином. Через 5 дней культуральную среду заменяли на предварительно подогретую культуральную среду Nut/F12, не содержащую ПТС (500 мкл в нижнем отсеке камеры и 200 мкл в верхнем отсеке). FITC-декстран (молекулярная масса 38900: Sigma Chemical Co.) добавляли в верхний отсек в конечной концентрации 1 мг/мл.
Через 1 час камеры вынимали, и уровень флуоресценции считывали непосредственно в нижнем отсеке с использованием Cytofluor II. Среднюю интенсивность флуоресценции в пяти независимых камерах подсчитывали и анализировали с использованием программы Statview и сравнения с помощью критерия Стьюдента. Для нормализации экспериментов величину средней интенсивности флуоресценции, полученную для клеток СНО дикого типа, принимали за 100%.
Трансфекция и очистка растворимых молекул
Трансфекция по непостоянному типу клеток 293-Т, Воsс-23 или стабильная Трансфекция клеток СНО растворимыми конструкциями Ig1Do и Ig2DO Flagtag/pcDNA-3 была осуществлена с использованием реагента липофектамина в соответствии с инструкциями изготовителя (Gibco BRL, Paisley, Шотландия). Образованные после трансфекции надосадочные фракции собирали каждые два дня в течение 10-дневного периода. Гранулы М2 (Kodak, New Haven, США), ковалентно соединенные с анти-FLAG антителом, промывали дважды в ФСБ, содержащем смесь для ингибирования протеаз (Boehringer Mannheim, Германия). Затем гранулы инкубировали при 4° С в течение 3 часов с надосадочной фракцией, полученной от трансфицированных клеток. После 5-кратной промывки в ФСБ, содержащем протеазные ингибиторы, колонку заполняли гранулами, и рекомбинантные молекулы элюировали 10 мМ глициновым буфером, рН=3,4, в соответствии с инструкциями изготовителя. Элюированные фракции, содержащие рекомбинантные белки, затем концентрировали на фильтре Centricon-10 (Millipore) и диализовали против ФСБ.
Окончательную концентрацию белка определяли с использованием теста Micro BCA (Biorad). Очищенные продукты затем подвергли электрофорезу в ДСН-полиакриламидном геле с последующим окрашиванием кумасси голубым для анализа их чистоты.
Тесты на трансмиграцию
Трансмиграция лейкоцитов сквозь эндотелиальные клетки была осуществлена в соответствии с описанным ранее (Wheerasinghe et al., 1998, J. Cell Biol., 142, 595-607). Вкратце, 1× 105 клеток t-end культивировали в течение 2 дней в камерах Transwell (поликарбонатные фильтры, размер пор 5 мкм: Costar) в присутствие 1 мкг растворимых рекомбинантных Ig-молекул: sJAM 2do или sCRAM-1 2do. Через 2 дня 1× 106 лейкоцитов, полученных из лимфатических узлов и пейеровых бляшек, добавляли в верхний отсек камеры, а число трансмигрировавших клеток контролировали в ходе эксперимента каждый час. Через 4 часа трансмигрировавшие клетки, полученные в пяти независимых камерах, объединяли и подвергали цитофлуорометрическому анализу на В- и Т-клеточные маркеры В220 и CD3. Результаты были получены с использованием устройства FACScalibur и анализаторной программы Cell-Quest (Becton-Dickinson).
Для теста на трансмиграцию спленоцитов 3× 105 эндотелиальных клеток высевали в камеры Transwell (поликарбонатные фильтры, размер пор 8 мкм: Costar), предоставляя возможность клеткам образовывать монослой за 18 часов. Культуральную среду из верхнего отсека камеры удаляли, и 1× 106 лейкоцитов в 100 мкл, свежевыделенных из селезенки с помощью центрифугирования в Ficoll, добавляли в верхний отсек камеры. SDF-1 (в конечной концентрации 40 нМ) добавляли в культуральную среду нижнего отсека камеры для установления градиента хемокинов между нижним и верхним отсеками. В эксперименте с антителами очищенные антитела 18-F26 или 19-Н36 добавляли в концентрации 10 мкг/мл в верхний отсек камеры, содержащий спленоциты. Через 4 часа трансмигрировавшие лейкоциты (в нижний отсек) собирали и подсчитывали. Полученные результаты выражены в процентах от числа внесенных клеток.
Результаты
Направленное дифференцированное представление
Регуляция генов в эндотелиальных клетках зависит от окружающей их среды. Настоящее изобретение было направлено на идентификацию генов, которые регулируются в процессе контакта эндотелия с опухолевыми клетками. Для этой цели был разработан тест in vitro, в котором использовали совместное культивирование меланомных опухолевых клеток (В16) и линии эндотелиомных клеток (t-end). Тотальная РНК, экстрагированная из этой клеточной смеси, была использована в качестве матрицы для синтеза кДНК, которую подвергали дифференцированному ПЦР-скринингу. В качестве контролей использовали кДНК, полученные на материале эндотелиальных или меланомных клеток, культивировавшихся отдельно друг от друга. Для идентификации транскриптов, регулируемых фактом совместного культивирования, сравнивали три различных типа. Для ограничения анализа теми последовательностями, которые кодируют поверхностно-клеточные молекулы, относящиеся к суперсемейству Ig, были использованы частично вырожденные праймеры, мишенями для которых являются последовательности, окружающие С-концевой остаток цистеина в доменах С2 молекул суперсемейства Ig. Наиболее воспроизводимый тип ПЦР-продуктов был получен при использовании праймеров, которые кодируют последовательность YYCxASI (фиг.7А). Данный улучшенный способ представления РНК был назван "направленным дифференцированным представлением" (TDD).
В повторных экспериментах со способом TDD было выявлено 16 дифференцированно экспрессирующихся генов. В результате последующих клонирования и анализа нуклеотидной и аминокислотной последовательностей три из 16 ПЦР-продуктов были определены как вероятные "кандидаты", кодирующие неизвестные члены Ig-суперсемейства. Один из трех "кандидатов" (CRAM-1) был выбран для дальнейшего исследования. При раздельном культивировании эндотелиальные клетки t-end.1 и меланомные клетки В16 экспрессировали высокие уровни транскрипта CRAM-1. Однако при условиях совместного культивирования уровень экспрессии CRAM-1 регулировался негативно, т.е. снижался (фиг.7В). Трансляция фрагмента длиной 350 п.н., соответствующего CRAM-1, показала присутствие аминокислотной последовательности YYCxAS, указывающий на концы иммуноглобулинового С2-домена с последующей открытой рамкой считывания (ORF), кодирующей гидрофобный мотив из 18 аминокислот, который определен как трансмембранный сегмент.
CRAM-1 - член иммуноглобулинового суперсемейства
Сравнение последовательностей между ПЦР-продуктом и базами данных по нуклеотидным последовательностям указало на гомологичные и идентичные последовательности в базах данных EST-маркеров мыши. Присутствие EST указывало на то, что ПЦР-продукт соответствовал последовательности, экспрессируемой in vivo. Было установлено, что в трех EST на их 5'-конце имеется последовательность длиной 300 п.н., которая идентична 300 п.н. в составе продукта TDD. У каждого EST 3'-концы включали полиаденилированный "хвост". В целом EST имели длину 1270 п.н. и соответствовали 3'-концу транскрипта CRAM. Поскольку 5'-конец транскрипта отсутствовал в составе клона кДНК EST, то он был получен с помощью ПЦР 5'-RACE. Полученная в результате полноразмерная последовательность длиной 1980 нуклеотидов у постулированной кДНК CRAM-1 показана на фиг.8А. Отчетливый консенсусный сайт (GACATGG) для инициации трансляции имеется в 16 п.н. вниз от 5'-конца, после которого расположена единственная ORF, для которой предсказывается кодирование белка из 310 аминокислот. Состоящий из 31 аминокислоты участок, следующий за предполагаемым инициирующим метионином, обладал признаками сигнального сегмента. Сайт расщепления был предсказан между остатками Ala-31 и Val-32.
Предполагаемая структура мышиного белка CRAM-1 показана на фиг.8В и состоит из внеклеточного сегмента, включающего вариабельный домен тяжелой цепи и иммуноглобулиновый С2-подобный домен (Pfam, The Sanger Centre and Blast) с двумя сайтами потенциального N-гликозилирования (аминокислоты 104 и 192). Анализ гидрофобности (Tmpred, ISREC) предсказал наличие трансмембранного участка между положениями 242-260. Предполагаемый цитоплазматический домен состоял из 49 аминокислот и включал ряд высококонсервативных сайтов фосфорилирования по остаткам Ser/Thr и Туг (фиг.8А: остатки выделены курсивом). Поиск известных параметров с помощью программы Prosite позволил идентифицировать мотивы SSK/SYK как протеинкиназные-С, SKQD/TSEE как СК2 и KQDGESY/KHDGVNY как мотивы тирозинкиназного фосфорилирования.
JAM, CRAM-1 и CRAM-2 определяют новое подсемейство
Некоторые белки проявили высокий уровень гомологии с CRAM-1. Два члена суперсемейства Ig: антиген АЗЗ человека и часть адгезивной молекулы нервных клеток мыши N-CAM, как было установлено, характеризуются уровнем гомологии с CRAM-1 41% и 46%, соответственно. JAM, как другой член Ig-Sf, имел сходную с CRAM-1 структуру при уровне идентичности 34% и гомологии 54% аминокислотной последовательности. Значительный уровень идентичности между JAM и CRAM-1 использовали для поиска третьей близкородственной последовательности в базах данных EST, а именно СRAM-2. Уровень идентичности трех молекул предполагает существование нового подсемейства молекул в составе Ig-суперсемейства (фиг.9). Гомология затрагивала не только общую структуру доменов V и С2 (С54-С118 и С147-С235 на фиг.9), но также и последовательности в пределах цитоплазматических доменов. Интересно, что наиболее различающиеся участки у трех молекул были обнаружены в начале домена V (положения 40-60) и в проксимальном цитоплазматическом сегменте (положения 280-300). Функции двух указанных участков соответствуют последовательностям, вовлеченным в связывание лиганда и передачу сигнала у других членов Ig-суперсемейства: это предполагает роль JAM, CRAM-1 и CRAM-1 в межклеточных связях.
Тканевое распределение мРНК JAM, CRAM-1 и CRAM-2
Экспрессия трех транскриптов в клетках различного происхождения была выявлена с использованием метода ОТ-ПЦР. Все протестированные эндотелиальные и эпителиальные клетки и большинство исследованных опухолевых клеток были позитивными, хотя и в разной степени у разных транскриптов (фиг.10А). Наивысший уровень экспрессии CRAM-1 был выявлен в SV40-трансформированной HEV-клеточной линии ТМЕ (дорожка 9) и в линии эмбриональных эндотелиальных клеток e-end.2 (дорожка 4). Транскрипты CRAM-2 и JAM показали более ограниченное распределение: они были обнаружены в линиях эндотелиальных клеток взрослых особей вместе с транскриптом CRAM-1 (дорожки 3, 7, 9 и 12). Нужно отметить, что транскрипты JAM и CRAM-2 были подвержены сильной негативной регуляции при действии TNF на эндотелиальные клетки, в то время как уровень транскрипта CRAM-1 оставался неизменным (дорожки 2 и 11). Интересно, что линия эмбриональных эндотелиальных клеток (дорожка 4) или линия эндотелиальных клеток взрослых особей, представляющие ангиогенный вариант t-end (дорожка 6), не были способны экспрессировать JAM или CRAM-2.
Тканевое распределение транскрипта JAM-2 анализировали с помощью Нозерн-блоттинга и сравнивали с JAM-1 (фиг.14). Длина транскрипта JAM-2 составляла 2 т.п.н., он был интенсивно экспрессирован в эмбриональной ткани и пейеровых бляшках, лимфатических узлах, почках и семенниках взрослых животных. В семенниках был выявлен предполагаемый сплайсинговый вариант длиной 1,8 т.п.н. Экспрессия транскрипта JAM-2 была низкой в легких, печени, селезенке и вилочковой железе. Относительное количество JAM-1 и JAM-2 сравнивали в процессе эмбриогенеза: мРНК, кодирующая JAM-2, становилась выявляемой по крайней мере через 7,5 дней после оплодотворения, в то время как мРНК JAM-1 в ходе эмбриогенеза не была выявлена вовсе.
Полученные результаты предполагают, что CRAM-1 широко экспрессирован в ходе эмбриогенеза и проявляет экспрессию, ограниченную эпителиальными или эндотелиальными компонентами взрослых тканей. Это согласуется с идеей, согласно которой этот белок участвует в обеспечении и поддержании поляризованной организации клеток.
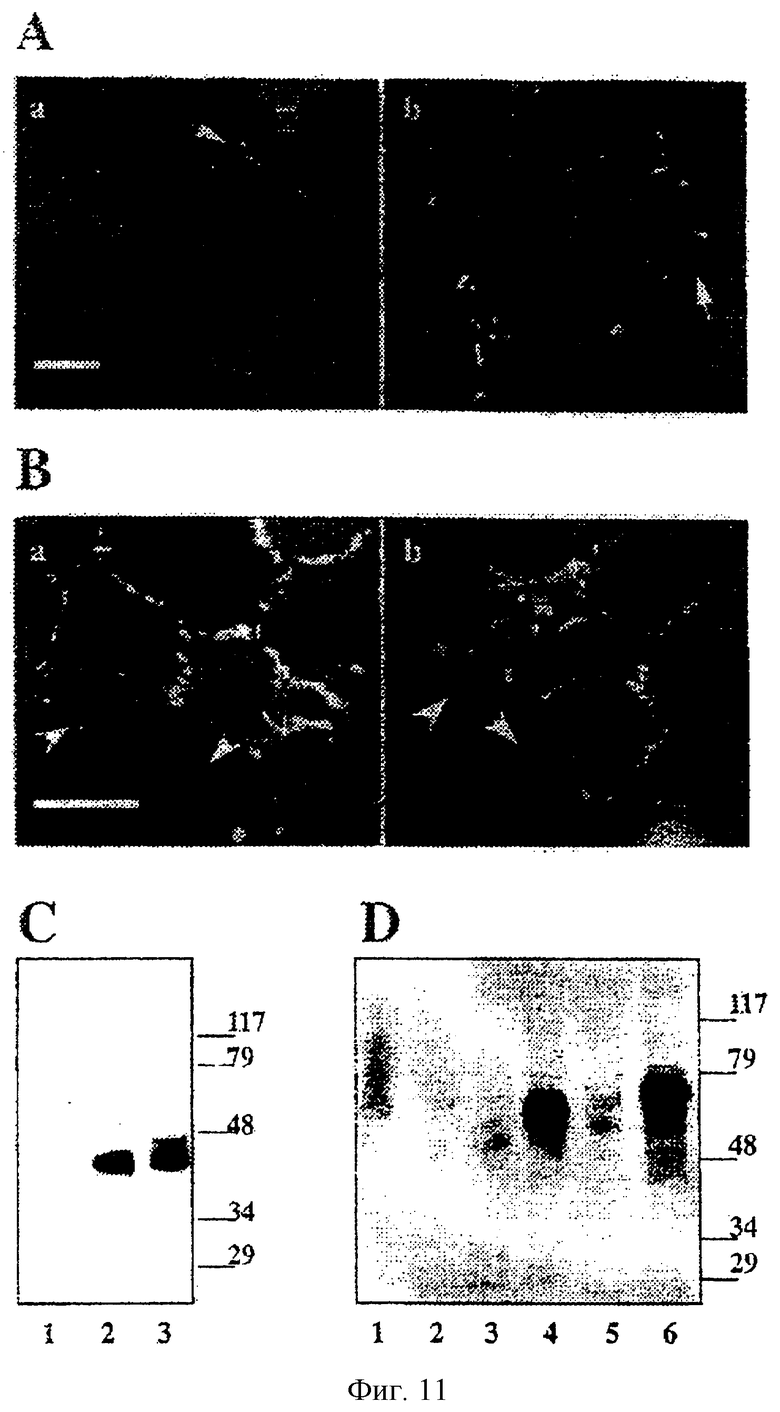
JAM-2 - белок с Мг 45-кД, локализация которого в межклеточных контактах зависит от гомофильных взаимодействий
Поскольку HEV-производная клеточная линия ТМЕ экспрессировала наивысший уровень JAM-2, эту линию эндотелиальных клеток использовали для дальнейшего анализа субклеточной локализации JAM-2 со сравнением с таковым у JAM-1. Локализация белка JAM-2 на поверхности эндотелиальных клеток была ограничена межклеточными контактами (фиг.11А: а). Окрашивание на JAM-2 было слабее по сравнению с таковым у JAM-1 и менее выраженным на выростах мембран между клетками.
Затем исследовали, взаимодействует ли присутствующий при межклеточных контактах белок JAM-2 с JAM-2 по гомофильному типу или он взаимодействует по гетерофильному типу с другой молекулой на соседней клетке. Для этой цели создавали гибрид белка JAM-2 с зеленым флуоресцирующим белком (JAM-2-EGFP), и этой конструкцией трансфицировали клетки СНО. Когда клетки СНО, трансфицированные кДНК JAM-2-EGFP, достигали стадии конфлюентности, JAM-2 выявлялся исключительно в межклеточных контактах, где обе контактирующие клетки экспрессировали этот белок (фиг.11В), в то время как контакты между экспрессирующими и неэкспрессирующими клетками были свободны от JAM-2 (фиг.11В: а; отмечено короткими стрелками). Такой же результат был получен в случае, когда клетки были трансфицированы химерной молекулой JAM-1-EGFP (фиг.11В: b). Это указывает на то, что и JAM-2, и JAM-1 требовал гомофильного взаимодействия для того, чтобы локализоваться в межклеточных контактах.
Для биохимической характеристики JAM-2 была проведена иммунопреципитация химерных белков JAM-2 или EGFP, экспрессированных клеточной линией ТМЕ или трансфицированными клетками СНО, соответственно (фиг.11С и D). Антитело, специфичное для JAM-2 - CRAM-19H36-иммунопреципитировало единственную полосу с Мг 45 кД из лизата клеток ТМЕ (фиг.11С, дорожка 3).
Установленная молекулярная масса была идентична при использовании восстанавливающих и невосстанавливающих условий (данные не показаны) и соответствовала молекулярной массе, предсказанной для расшифрованной аминокислотной последовательности JAM-2, если для каждого сайта N-гликозилирования прибавляли по 5 кД. Иммунопреципитация JAM-1 с использованием антитела Н202-106, специфичного в отношении JAM, дала единственную полосу с меньшей молекулярной массой (примерно 42 кД: дорожка 2), что исключает возможную перекрестную реактивность между антителами к JAM-2 и к JAM-1.
Иммунопреципитация рекомбинантных белков JAM-1-EGFP или JAM-2-EGFP после биотинилирования поверхности клеток дала единственные широкие полосы с Мг 70 кД и 73 кД, соответственно: это указывает на то, что молекулы экспрессировались на поверхности трансфицированных клеток СНО (фиг.11D: дорожки 4 и 6). Эти молекулярные массы были ожидаемыми с учетом того, что EGFP характеризуется молекулярной массой 28 кД.
Плотность и миграция лейкоцитов
Для того, чтобы понять, как молекулы влияют на целостность монослоя эндотелиальных клеток и как они регулируют функцию сосудистого эндотелия, тесты на миграцию лейкоцитов через эндотелий были проведены в присутствии рекомбинантных растворимых JAM или CRAM-1. Эндотелиальные клетки культивировали в течение двух дней в присутствии sJAM-Ig2Do или sCRAM-1-Ig2Do. Целостность монослоя в течение этого периода не нарушалась, что, по-видимому, объясняется молекулярной избыточностью механизма образования межклеточных контактов. Тест на трансмиграцию проводили в присутствии 1 мкг рекомбинантных растворимых молекул. Как показано на фиг.20А, на число трансмигрировавших клеток в незначительной степени влияло присутствие sJAM-Ig2Do (незакрашенные квадратики) в течение первых трех часов. Через 4 часа число трансмигрировавших клеток возрастало по сравнению с контролем (пунктирная линия). Напротив, в присутствии sCRAM-1-Ig2Do (закрашенные кружки) количество трансмигрировавших клеток резко снижалось.
Поскольку популяции лейкоцитов являлись гетерогенными, оценивали то, действует ли sCRAM-1-Ig2Do на конкретную субпопуляцию лейкоцитов или же трансмиграция блокируется неспецифичным образом. Для этой цели трансмигрировавшие клетки помечали по лимфоцитарным маркерам CD3 и В220 (фиг.20В).
Было отмечено, что конструкция sCRAM-1-Ig2Do специфически блокировала трансмиграцию лейкоцитов нелимфоидного типа, а именно клетки миелоидной линии (центральный рисунок, заштрихованные столбцы). Напротив, sJAM-Ig2Do незначительно повышала число трансмигрировавших Т-клеток (левый рисунок, незакрашенный столбец) без какого-либо влияния на другие клеточные субпопуляции.
Более того, когда эндотелиомные клетки, трансфицированные CRAM-1, использовали в тесте на трансмиграцию, было выявлено усиление трансмиграции (фиг.12), в то время как трансфекция CRAM-2 не проявляла какого-либо влияния на трансмиграцию. Когда в данном тесте добавляли SDF-1, миграция лейкоцитов через эндотелий достигала 20%. Это частично блокировалось моноклональными антителами, специфичными для CRAM-1.
Полученные данные указывают на то, что вовлечение CRAM-1 в контакт между эндотелиальными клетками может регулировать функцию эндотелиального слоя. Можно предположить, что молекулы данного семейства станут барьером, если эндотелиальные клетки достигнут стадии конфлюентности. В связи с этим регуляция транскриптов CRAM-1 была исследована в эндотелиальных клетках при разных условиях культивирования.
CRAM негативно регулируется высокой конфлюентностью
Поскольку транскрипт, который кодирует CRAM-1, не регулируется с участием TNF, но негативно регулируется совместным культивированием эндотелия с опухолевыми клетками, тест на конфлюентность был использован для дальнейшего анализа этой регуляции. При низкой конфлюентности клетки активно делились, и взаимодействия с участием CRAM-1 не происходили, в то время как при высокой конфлюентности клетки делились менее активно, причем на фоне привлечения CRAM-1. Уровень экспрессии мРНК CRAM-1 определили при различной плотности клеток с помощью полуколичественной ОТ-ПЦР. Как показано на фиг.13, уровень экспрессии транскриптов CRAM-1 снижался при достижении конфлюентности (дорожки 1, 2, 3 на фиг.13 соответствуют конфлюентности 100, 50 и 10%, соответственно). Данный эффект отчетливо выявлялся у клеток t-end, но был еще более выражен в клеточной линии ТМЕ, которая интенсивно экспрессировала CRAM-1. Негативная регуляция CRAM-1 в эндотелии также усиливалась в случае, когда эндотелиальные клетки культивировали вместе с карциномными клетками KLN-205, которые сами по себе не экспрессируют CRAM-1. Это подтвердило наличие связи между экспрессией CRAM-1 и клеточным циклом, поскольку для опухолевых клеток было описано увеличение темпа роста эндотелиальных клеток в случае контакта с ними. Заслуживает внимания то, что результаты, полученные для карциномных клеток KLN-205, оказались идентичны тем, которые были получены авторами изобретения в исходном скрининге меланомной опухоли В16. Это указывает на базовый механизм, по которому опухоли влияют на поведение эндотелия.
JAM-2 интенсивно экспрессируется во время эмбриогенеза и ограничен HEV и субпопуляциями эндотелиальных клеток в тканях взрослых особей
Для более точного установления тканевого распределения JAM-2 иммуногистологический анализ был проведен на срезах почки и брыжеечного лимфатического узла (фиг.15), которые экспрессировали наивысшие уровни транскрипта JAM-2. В кортикольном слое почек специфическое окрашивание интертубулярных структур было выявлено с использованием моноклональных антител к РЕСАМ (GC51) или анти-JAM-2 (CRAM-XVIIIF26), в то время как анти-ZO-1 или анти-JAM-1 преобладающим образом окрашивали эпителиальные клетки канальцев (данные не показаны).
Следовательно, заявители сконцентрировали внимание на сосудистых структурах, которые погружаются в мозговое вещество и соответствуют эндотелиальным структурам радиальных вен или vasa recta. Для этого были изготовлены серийные срезы и сосудистые структуры идентифицировали с помощью окрашивания антителом к РЕСАМ (фиг.15d). На эквивалентном участке соседних срезов линейные прокрашенные зоны между эндотелиальными слоями были выявлены с помощью антител к JAM-2, JAM-1 или ZO-1 (фиг.15а, b и с, соответственно). На срезах брыжеечного лимфатического узла типичное окрашивание венул с плотным эндотелием (HEV) было получено с помощью mAb анти-JAM-2 (фиг.15е). Также было обнаружено, что HEV экспрессируют JAM-1, ZO-1 или РЕСАМ (фиг.15f, g и h) при слабых различиях в субклеточной локализации окрашенных зон (фиг.15e-h: вставки). В кортикольном слое брыжеечных лимфатических узлов типичное окрашивание субкапсулярных пазух было получено при использовании всех антител (фиг.15i-l), что соответствует окрашиванию афферентных лимфатических сосудов. Таким образом, окрашивание моноклональным антителом CRAM-18F26, специфичным в отношении JAM-2, ограничено некоторыми эндотелиальными клетками или структурами, тесно связанными с сосудистой системой.
Для уточнения того, отображается ли на изображениях, показанных на фиг.15, окрашивание эндотелиальных клеток, цитофлуорометрический анализ экспрессии JAM-2 был проведен на материале различных клеточных линий или свежевыделенных эндотелиальных клеток из диспергированных тканей. Линии эндотелиальных клеток (tEnd.1, eEnd.2 и ТМЕ) экспрессировали низкие уровни JAM-2 на клеточной поверхности и различные уровни JAM-1 (фиг.16А).
Цитометрический анализ свежевыделенных эндотелиальных клеток проводили путем троекратного окрашивания клеточных суспензий, полученных после диспергирования органа действием коллагеназы/диспазы. Эндотелиальные клетки идентифицировали путем отбора клеток, окрашенных и PECAM/CD31, и ацетилированным LDL (Voyta et al., 1984, J. Cell Biol., 99, 2034). Окрашивание на JAM-2 или JAM-1 на такой дважды позитивной клеточной популяции показано на фиг.16В. В почках и пейеровых бляшках все изолированные клетки, позитивные по CD31 и ацетилированным LDL, также окрашивались на JAM-2: это означает, что по крайней мере в этих органах эндотелиальные клетки экспрессируют JAM-2 in vivo. Когда окрашивание применяли к клеткам, выделенным из лимфатического узла, экспрессия JAM-2 обнаруживалась только в клеточной субпопуляции, что отражает возможную гетерогенность фенотипов эндотелиальных клеток в составе такой ткани.
Взятые вместе, полученные результаты цитометрического и иммуногистохимического анализа показывают, что JAM-2 коэкспрессируется с JAM-1 в эндотелиальных клетках почек, пейеровых бляшек и лимфатических узлов.
Динамическая локализация JAM-2 в межклеточных контактах
Для выяснения механизма, по которому JAM-2 специфически локализуется в межклеточных контактах, был применен метод повременной (покадровой) видеомикроскопии. Клетки СНО, стабильно трансфицированные флуоресцирующей химерной молекулой, обрабатывали трипсином и помещали на предметные стекла для анализа изображения. После распределения клеток поверхностная экспрессия JAM-2-EGFP не была однородной, но была кластеризована в областях межклеточных контактов (фиг.17А: клетки отмечены звездочками). В процессе образования новых межклеточных контактов было выявлено перемещение JAM-2-EGFP к местам соединения клеток, и интенсивный сигнал флуоресценции был выявлен в точке нового взаимодействия между клетками, образующими новый межклеточный контакт (стрелки). Химерный белок характеризовался обогащением в мембранных выростах между контактирующими клетками, что выглядело как структура типа застежки-молнии через 12 или 18 минут. Интересно, что локализация JAM-2 в области первичных межклеточных контактов не утрачивалась в процессе образования нового контакта мембран (см. клеточные контакты в верхнем левом углу). Эти данные указывали на то, что JAM-2-EGFP специфическим образом менял локализацию в сторону нового клеточного контакта и что после этого такая локализация оказывалась стабильной.
Для дальнейшего определения требований к локализации JAM-2 повременная видеомикроскопия была проведена после повреждения клеточного монослоя (фиг.17В). Клетки со стороны поврежденного края сохраняли JAM-2 в своих неповрежденных контактных сайгах (короткие стрелки), но утрачивали локализацию JAM-2 в поврежденном сайте (стрелки): это указывает на то, что вовлечение JAM-2 лигандом противоположной клетки является необходимым условием для поддержания его мембранной локализации. За 90-минутный период после повреждения клетки, граничащие с повреждением, начинали перемещаться в поврежденную область. Интересно, что такие клетки сохраняли контакты с соседними клетками за счет мембранных выростов, которые проявляли яркую флуоресценцию, т.е. были позитивными по JAM-2 (короткие стрелки). Полученные данные подтвердили гипотезу, согласно которой гомофильные взаимодействия JAM-2 могут участвовать в установлении или поддержании межклеточных контактов.
JAM-2 повышает плотность монослоя и участвует в плотных соединительных комплексах
Поскольку ряд молекул, участвующих в межклеточных связях, как было показано, регулирует параклеточную проницаемость клеточных монослоев, исследовали, может ли JAM-2 также влиять на подобную функцию. Трансфекция JAM-2-EGFP снижала параклеточную проницаемость FITC-декстрана и улучшала скрепление монослоев клеток СНО на 42,5%, в то время как трансфекция посторонней молекулой Тас (IL2R-α ) не снижала на статистически значимом уровне параклеточную проницаемость трансфицированных клеток СНО (фиг.18). Трансфекция JAM-1-EGFP также снижала параклеточную проницаемость монослоя трансфицированных клеток СНО.
Полученные результаты подняли вопрос о том, способны ли химерные молекулы участвовать в столь специализированной субклеточной структуре, какую представляют плотные контакты поляризованных эпителиальных клеток.
Для ответа на этот вопрос химерные белки EGFP были использованы для трансфекции клеток MDCK, и их субклеточную локализацию сравнивали с такой у маркера плотных контактов - окклудина. Как показано на фиг.19А, когда серийные изображения через каждые 0,9 мкм анализировали на EGFP-флуоресценцию и сравнивали с окрашиванием на окклудин, специфичное обогащение по JAM-2-EGFP было выявлено в межклеточных контактах на уровне плотного соединения. На базовом уровне (слева) были выявлены межклеточные пятна EGFP-флуоресценции. Сходный анализ (фиг.19В) клеток MDCK, трансфицированных JAM-1-EGFP, показал сходную колокализацию с окклудином. Тем не менее распределение флуоресценции JAM-1-EGFP не было столь однородным, чем то, которое выявляется для JAM-2-EGFP на уровне плотных соединений.
Обсуждение
Данный пример описывает применение новой стратегии скрининга для идентификации регулируемых транскриптов, кодирующих члены иммуноглобулинового суперсемейства адгезивных молекул. Описываемым здесь является клонирование с помощью такого способа новой молекулы CRAM-1 в качестве регулируемого транскрипта. Регуляция, выявленная в эндотелиальных клетках, культивируемых в присутствие опухолей, подтверждена с помощью полуколичественной ОТ-ПЦР, и было показано, что она зависит от стадии роста клеток. Вследствие дифференцированной экспрессии в условиях разной конфлюентности клеток было предложено название "регулируемая конфлюентностью адгезивная молекула-1" (CRAM-1).
Также описываемым в данном тексте является близкородственная CRAM-1 последовательность, обозначенная как CRAM-2. CRAM-1 и -2 представляют собой прототипы нового подсемейства адгезивных молекул, которое также включает недавно описанную молекулу JAM (Chretien et al., 1998, Eur. J.Immunol., 28, 4094-4104; Malergue et al., 1998, Mol. Immunol., 35, 1111-1119).
CRAM-1 и JAM по преимуществу экспрессируются эндотелиальной и эпителиальной тканью в области межклеточных контактов и придают специфические свойства поляризованным клеточным слоям. Влияние рекомбинантных растворимых молекул в тесте на миграцию через эндотелий и регуляция JAM, CRAM-1 и CRAM-2 указывают на, что эти три молекулы играют важную роль в поддержании физиологических параметров сосудов.
Новая стратегия скрининга, названная "направленным дифференцированным представлением" ("Targeted Differential Display" - TDD), как было подтверждено, является эффективным способом селективной амплификации представляющей интерес кДНК. В способе TDD успешно используются частично вырожденные праймеры, обеспечивающие избирательную направленность на консервативный участок - Y(Y/Q/R)CXAS - в составе С2-подобных Ig-доменов. Повторные эксперименты приводят к воспроизводимым типам представления. Из 16 дифференцированно экспрессированных транскриптов три соответствовали генам, обладающим существенным уровнем гомологии с консервативными Ig-последовательностями. Такое повышение специфичности позволяет преодолеть основные трудности, имеющиеся в уже известных методах классического фингерпринтинга и дифференцированного представления РНК. Фингерпринтинг РНК в течение долгого времени использовался для идентификации дифференцированно экспрессируемых генов. Однако из-за использования специфичных по последовательности праймеров данный способ выявляет транскрипты только отобранных и уже известных белков. С другой стороны, в методе представления РНК используют случайные праймеры и проводят неспецифическую амплификацию транскриптов. В этом случае целью является точный учет любых различий в уровнях мРНК между двумя биологическими системами, сравнение которых проводят. Способ TDD является преимущественным способом скрининга, который сочетает специфичность РНК-фингерпринтинга с вырожденностью дифференцированного представления РНК, что обеспечивает избирательность. Благодаря направленности на родственные транскрипты, данный способ в существенной степени уменьшил время, необходимое для скрининга. Следовательно, становится возможной идентификация новых членов конкретных семейств белков. Это является существенным улучшением известных стратегий неспецифического скрининга.
Данные общие свойства были использованы для конструирования рекомбинантных белков с целью изучения функций JAM, CRAM-1 и CRAM-2. В настоящем примере влияние sJAM-Ig2Do и sCRAM-1-Ig2Do описывается для трансмиграционного теста in vitro. Специфические блокирующие эффекты в отношении миграции миелоидных клеток могли быть выявлены с использованием sCRAM-1-Ig2Do, в то время как sJAM-Ig2Do проявил только незначительное влияние на лимфоциты.
Транскрипты JAM и CRAM-2 проявили сходные параметры тканевого распределения и регуляции экспрессии под действием TNF: это указывает на то, что они действуют по сходным физиологическим механизмам. Напротив, транскрипты CRAM-1 не регулируются действием TNF, но регулируются скоростью пролиферации или плотностью эндотелиальных клеток. Действительно, сверхэкспрессия транскриптов CRAM-1 в делящихся клетках и их негативная регуляция в покоящихся клетках указывают на то, что данная молекула участвует в формировании непрерывного монослоя. Ее функцией в конфлюирующих клеточных монослоях является поддержание слоя эндотелиальных клеток и связанных с этим свойств. Поскольку различные популяции лейкоцитов должны мигрировать в область формирования иммунного ответа, то предполагается, что нелимфоидные клетки перемещаются по зависящему от CRAM-1 механизму, в то время как лимфоидные клетки мигрируют с участием JAM или CRAM-2. В этом случае иммунный ответ может быть модулирован с использованием комбинаций различных рекомбинантных растворимых молекул.













Изобретение относится к биотехнологии и может быть использовано в медицине для лечения различных заболеваний, в частности воспалительных реакций. В изобретении описан новый полипептид в изолированной форме, относящийся к подсемейству суперсемейства иммуноглобулинов человека (Ig-Sf). Данный полипептид проявляет по крайней мере 70%-ный уровень гомологии с аминокислотной последовательностью мышиных регулируемых конфлюентностью адгезивных молекул CRAM-1 или CRAM-2 (фиг.3, 6 представлены в заявке). Изобретение касается также антител, специфичных по отношению к полипептиду. Показана возможность антител и растворимого полипептида для лечения воспаления и опухолей. В изобретении описан поли- или олигонуклеотид, кодирующий полноразмерный полипептид или его часть; является праймером, зондом, антисмысловой РНК, имеет нуклеотидную последовательность (НП), по существу идентичную НП CRAM-1 человека. Изобретение позволяет получить новые адгезивные белки из суперсемейства Ig-Sf, которые регулируются на транскрипционном уровне в эндотелии под влиянием опухолей. 5 н. и 14 з.п. ф-лы, 21 ил.

| WO 9824897 А, 11.06.1998 | |||
| WO 9842739 А, 01.10.1998. |

