Область техники, к которой относится изобретение
Данное изобретение относится к модифицированным антителам, содержащим две или более V-областей Н-цепи и две или более V-областей L-цепи моноклонального антитела, которое обнаруживает агонистическую активность посредством сшивания молекулы (молекул) клеточной поверхности или внутриклеточной молекулы (молекул). Эти модифицированные антитела имеют агонистическую активность трансдукции сигнала в клетки посредством сшивания молекул (молекулы) клеточной поверхности и применимы в качестве лекарственного средства для различных целей.
Уровень техники
Патентная заявка Японии JP-A 9-295999 описывает получение специфического моноклонального антитела с использованием линии стромальных клеток селезенки в качестве сенсибилизирующего антигена, с целью получения специфических антител, которые могут узнавать вышеупомянутые стромальные клетки селезенки, и получение новых моноклональных антител, которые узнают мышиный ассоциированный с интегрином белок (мышиный IAP) в качестве антигена. Патентная заявка Японии JP-A 9-295999 описывает также, что эти моноклональные антитела способны индуцировать апоптоз миелоидных клеток.
WO 99/12973 описывает моноклональные антитела, антигеном для которых служит ассоциированный с интегрином белок человека (далее называемый IAP человека; его аминокислотная последовательность и нуклеотидная последовательность описаны в J.Cell Biol., 123, 485-496, 1993; см. также Journal of Cell Science, 108, 3419-3425, 1995), которые способны индуцировать апоптоз ядросодержащих клеток крови (гемоцитов) человека (миелоидная клетка и лимфоцит), содержащих указанный IAP человека. Эти моноклональные антитела называют антителом MABL-1 и антителом MABL-2, а гибридомы, продуцирующие эти антитела, также называют MABL-1 (FERM ВР-6100) и MABL-2 (FERM BP-6101), соответственно.
Японская патентная заявка 11-63557 описывает получение одноцепочечных Fv-областей, представляющих собой Fv-области моноклональных антител, антигеном которых является IAP человека. Одноцепочечные Fv способны индуцировать апоптоз ядросодержащих клеток крови, имеющих IAP человека.
Моноклональное антитело, узнающее IAP в качестве антигена, индуцирует апоптоз ядросодержащих клеток крови, содержащих IAP человека, но оно также вызывает гемагглютинацию in vitro. Это указывает на то, что введение большого количества моноклонального антитела, узнающего IAP в качестве антигена, может приводить к побочному эффекту, такому как гемагглютинация.
Авторы изобретения провели интенсивное исследование в отношении использования моноклональных антител против IAP человека в качестве терапевтического агента при заболеваниях крови и получили одноцепочечные Fv, имеющие одноцепочечный Fv-фрагмент, способный индуцировать апоптоз ядросодержащих клеток крови, содержащих IAP человека.
С другой стороны, были разработаны модифицированные антитела, в частности, антитела с уменьшенным молекулярным размером, например, одноцепочечные Fv, для улучшения проникновения в ткани и опухоли посредством уменьшения молекулярного размера и для получения рекомбинантым способом. Недавно димеры одноцепочечных Fv, в частности, биспецифические димеры, использовали для сшивания клеток. Типичными примерами таких димеров являются гетеродимеры одноцепочечных Fv, узнающие антигены раковых клеток и антигены клеток-хозяев, подобных NK-клеткам (естественным киллерам), и нейтрофилам (Kipriyanov et al., Int. J. Cancer, 77, 9763-9772, 1998). Они были получены способом конструирования одноцепочечных Fv в виде модифицированных антител, которые являются более эффективными в лечении раков посредством индуцирования межклеточных сшивок. Считалось, что межклеточное сшивание индуцируется антителами и их фрагментами (например, Fab-фрагментом), биспецифическими модифицированными антителами и даже димерами одноцепочечных Fv, которые являются моноспецифическими.
В качестве антител, способных передавать сигнал сшиванием молекулы (молекул) клеточной поверхности, известны антитело против ЕРО-рецептора, участвующего в дифференцировке и пролиферации клеток (JP-A 2000-95800), антитело против MuSK-рецептора (Xie et al., Nature Biotech. 15, 768-771, 1997) и другие. Однако, не было сообщений о модифицированных антителах уменьшенного молекулярного размера.
Обратив внимание на то, что одноцепочечные Fv-мономеры, полученные из антитела MABL-1 и антитела MABL-2, не индуцируют апоптоз клеток, тогда как димеры одноцепочечных Fv индуцируют апоптоз клеток, содержащих IAP, авторы данного изобретения обнаружили, что они сшивают (димеризуют) IAP-рецептор на клеточной поверхности, вследствие чего сигнал передается в эти клетки и, как результат, индуцируется апоптоз. Это предполагает, что моноспецифические одноцепочечные Fv-димеры сшивают молекулу (молекулы) клеточной поверхности (например, рецептор) и передают сигнал подобно лиганду, выполняя таким образом роль агониста.
Сосредоточившись на межклеточном сшивании, авторы изобретения обнаружили, что вышеупомянутые одноцепочечные димеры Fv не вызывают гемагглютинации, тогда как вышеупомянутые моноклональные антитела вызывают гемагглютинацию. Тот же самый результат наблюдали также с одноцепочечными бивалентными антителами (одноцепочечными полипептидами, содержащими две V-области Н-цепи и две V-области L-цепи). Это предполагает, что моноклональные антитела могут образовывать межклеточные сшивки, тогда как модифицированные антитела, подобные одноцепочечным Fv-димерам и одноцепочечным бивалентным антителам, сшивают молекулу (молекулы) клеточной поверхности, но не образуют межклеточных сшивок.
На основе этих наблюдений авторы изобретения впервые обнаружили, что модифицированные антитела, такие как одноцепочечные Fv-димеры и одноцепочечные бивалентные антитела, сшивают молекулу (молекулы) клеточной поверхности или внутриклеточную молекулу (молекулы) одной и той же клетки, наряду с известным межклеточным сшиванием, и пригодны в качестве лиганда такой молекулы (молекул) (в частности, в качестве лиганда, который имитирует действие природного лиганда).
Обнаружением дополнительно того, что молекула антитела (цельный IgG) может быть модифицирована в одноцепочечные Fv-димеры, одноцепочечные бивалентные антитела и т.п., которые сшивают молекулу (молекулы) клеточной поверхности, с уменьшением тем самым побочных действий, вызываемых межклеточным сшиванием, и обеспечением новых лекарственных средств, индуцирующих только желательное действие на клетку, авторы завершили данное изобретение. Модифицированные антитела данного изобретения имеют необыкновенно высокую активность в сравнении с природными лигандами, такими как ТРО (тромбопоэтин), ЕРО (эритропоэтин) или G-CSF (гранулоцитарный колониестимулирующий фактор), или цельными антителами (IgG), имеющими ту же самую V-область, что и модифицированные антитела. Они имеют улучшенную проницаемость в ткани вследствие уменьшенного молекулярного размера в сравнении с молекулами антител и отсутствия константных областей.
Сущность изобретения
Целью данного изобретения является обеспечение агонистических модифицированных антител малого молекулярного размера, которые содержат две или более V-областей Н-цепи и две или более V-областей L-цепи моноклональных антител и обладают агонистическим действием посредством сшивания молекулы (молекул) клеточной поверхности или внутриклеточной молекулы (молекул).
Таким образом, данное изобретение относится к модифицированным антителам, которые содержат две или более V-областей Н-цепи и две или более V-областей L-цепи, предпочтительно от 2 до 6 каждой, особенно предпочтительно от 2 до 4 каждой, наиболее предпочтительно две каждой из них и обнаруживают агонистическую активность посредством сшивания молекулы (молекул) клеточной поверхности или внутриклеточной молекулы (молекул).
Термин "модифицированные антитела" в этом описании означает любые вещества, которые содержат две или более V-областей Н-цепи и две или более V-областей L-цепи, где указанные V-области объединены непосредственно или через линкер посредством ковалентной связи или нековалентной связи, например, полипептиды и соединения, получаемые объединением каждой V-области антитела через пептидный линкер или химический сшивающий агент, и т.п. Две или более V-областей Н-цепи и две или более V-областей L-цепи, используемые в данном изобретении, могут быть получены из одного и того же антитела или из различных антител.
Предпочтительными примерами модифицированных антител данного изобретения являются мультимеры, такие как димеры, тримеры или тетрамеры одноцепочечного Fv-фрагмента, содержащего V-область Н-цепи и V-область L-цепи, или одноцепочечные полипептиды, содержащие две или более V-областей Н-цепи и две или более V-областей L-цепи. Когда модифицированные антитела данного изобретения являются мультимерами одноцепочечных Fv, такими как димеры, тримеры, тетрамеры и т.п., содержащими V-область Н-цепи и V-область L-цепи, предпочтительно, чтобы V-область Н-цепи и V-область L-цепи, находящиеся в одной и той же цепи, не ассоциировались с образованием антигенсвязывающего сайта.
Более предпочтительными примерами являются димеры одноцепочечных Fv, которые содержат V-область Н-цепи и V-область L-цепи, или одноцепочечный полипептид, содержащий две V-области Н-цепи и две V-области L-цепи. V-область Н-цепи и V-область L-цепи соединены предпочтительно через линкер в этих модифицированных антителах.
"Агонистическое действие" в этом описании означает биологическое действие, происходящее в клетке (клетках), в которые сигнал передается посредством сшивания молекулы (молекул) клеточной поверхности или внутриклеточной молекулы (молекул), например, индукция апоптоза, индукция пролиферации клеток, индукция дифференцировки клеток, индукция клеточного деления или регуляция клеточного цикла.
ED50 агонистического действия в данном изобретении определяют известными способами, используемыми для измерения агонистического действия. Примерами являются детектирование агонист-специфической гибели клеток или пролиферации клеток, детектирование экспрессии белков, специфических для дифференцировки клеток (например, специфических антигенов) или измерение киназной активности, специфической для клеточного цикла. ED50 является дозой, требующейся для достижения 50% от максимального эффекта, принятого за 100% на кривой доза-ответ.
Предпочтительные модифицированные антитела данного изобретения обладают агонистическим действием (ED50), эквивалентным или лучшим, чем агонистическое действие антитела, имеющего тот же самый антигенсвязывающий район, что и модифицированное антитело, а именно цельного антитела, такого как IgG (далее называемого "исходным антителом"), имеющего ту же самую пару V-области Н-цепи и V-области L-цепи, что и пара V-области Н-цепи и V-области L-цепи, образующая антигенсвязывающий район модифицированного антитела. Более предпочтительными являются антитела, имеющие агонистическое действие (ED50), более чем в два раза превышающее агонистическое действие исходного антитела, более предпочтительно - более чем в 5 раз, наиболее предпочтительно - более чем в 10 раз. Данное изобретение включает в себя модифицированные антитела с агонистическим действием, содержащие V-область Н-цепи и V-область L-цепи, образующие тот же самый антигенсвязывающий район, что и исходное антитело, которое связывается с молекулой-мишенью (молекулами-мишенями) клеточной поверхности или внутриклеточной молекулой (молекулами), но не обладает агонистическим действием на эту молекулу.
Соединения, содержащие две или более V-областей Н-цепи и две или более V-областей L-цепи данного изобретения могут быть любыми соединениями, которые содержат две или более V-областей Н-цепи и две или более V-областей L-цепи антитела и обнаруживают агонистическое действие (ED50), эквивлентное или лучшее, чем агонистическое действие природного лиганда, связывающегося с молекулой (молекулами) клеточной поверхности или внутриклеточной молекулой (молекулами). Предпочтительными являются соединения, имеющие агонистическое действие (ED50), более чем в два раза превышающее агонистическое действие природного лиганда, более предпочтительно - более чем в 5 раз, наиболее предпочтительно - более чем в 10 раз.
"Соединения", упоминаемые здесь, включают в себя не только модифицированные антитела данного изобретения, но также любые соединения, содержащие два или более, предпочтительно от 2 до 6, более предпочтительно от 2 до 4, наиболее предпочтительно 2 антигенсвязывающих района, такие как цельные антитела или F(ab')2.
Модифицированные антитела или соединения данного изобретения, содержащие две или более V-областей Н-цепи и две или более V-областей L-цепи антитела, предпочтительно не имеют существенного межклеточного адгезионного действия. Когда V-область Н-цепи и V-область L-цепи модифицированных антител данного изобретения происходят из одного и того же антитела, они предпочтительно имеют межклеточное адгезионное действие (ED50) не более 1/10 в сравнении с исходным антителом.
ED50 межклеточного адгезионного действия определяют в данном изобретении известными способами для измерения агонистического действия, например, измерением агломерирующего действия клеток, экспрессирующих указанную молекулу клеточной поверхности, таким как тест гемагглютинации.
Данное изобретение относится к ДНК, которые кодируют эти модифицированные антитела.
Данное изобретение относится к клеткам животных или микроорганизмам, которые продуцируют эти модифицированные антитела.
Данное изобретение относится к применению модифицированных антител в качестве агониста.
Данное изобретение относится к способу передачи сигнала в клетки посредством сшивания молекулы клеточной поверхности или внутриклеточной молекулы с использованием модифицированного антитела и тем самым индуцирования агонистического действия на клетки, такого как индукция апоптоза, индукция пролиферации клеток, индукция дифференцировки клеток, индукция клеточного деления или действие на регуляцию клеточного цикла.
Данное изобретение относится к лекарственному средству, содержащему модифицированное антитело.
Данное изобретение относится к применению модифицированного антитела в качестве лекарственного средства.
Данное изобретение относится к способу скрининга или измерения модифицированного антитела, которое содержит две или более V-областей Н-цепи и две или более V-областей L-цепи антитела и обнаруживает агонистическое действие посредством сшивания молекулы клеточной поверхности или внутриклеточной молекулы, включающему 1) получение модифицированного антитела, содержащего две или более V-областей Н-цепи и две или более V-областей L-цепи антитела и связывающего специфически указанную молекулу, 2) контактирование модифицированного антитела с клетками, экспрессирующими указанную молекулу, и 3) измерение агонистического действия, вызываемого сшиванием указанной молекулы, которое имеет место в этих клетках. Этот способ измерения применим для контроля качества при получении модифицированных антител данного изобретения в качестве лекарственного средства и для других целей.
Вышеупомянутый одноцепочечный Fv-димер включает в себя димер, образованный через нековалентную связь, димер, образованный через ковалентную связь через сшивающий радикал, и димер, образованный через сшивающий реагент (антитело, фрагмент антитела или бивалентное модифицированное антитело). Обычные сшивающие радикалы, используемые для образования поперечных связей в пептидах, могут быть использованы в качестве сшивающих радикалов для образования этих димеров. Примерами являются дисульфидные поперечные связи с участием остатка цистеина, другие сшивающие радикалы, такие как С4-С10-алкилены (например, тетраметилен, пентаметилен, гексаметилен, гептаметилен и октаметилен и т.д.) или С4-С10-алкенилены (цис/транс-3-бутенилен, цис/транс-2-пентенилен, цис/транс-3-пентенилен, цис/транс-3-гексенилен и т.д.).
Кроме того, сшивающим реагентом, который может объединяться с одноцепочечным Fv, является, например, аминокислотная последовательность, которая может быть необязательно введена в Fv, например, антитело против последовательности FLAG и т.п. или его фрагмент, или модифицированное антитело, происходящее из этого антитела, например, одноцепочечный Fv.
Данное изобретение относится также к способу индукции агонистического действия в отношении клеток путем введения первого лиганда и второго лиганда, которые объединяются с молекулой (молекулами) клеточной поверхности или внутриклеточной молекулой (молекулами), и введением вещества, которое соединяется с первым и вторым лигандами и образует поперечную связь между первым и вторым лигандами. Первый и второй лиганды могут быть любыми веществами, которые содержат сайт связывания для указанной молекулы и могут индуцировать агонистическое действие, будучи сшитыми поперечной связью. Предпочтительными примерами являются моновалентные модифицированные антитела, такие, как тот же самый или иной одноцепочечный Fv-мономер, фрагмент антитела и т.д. Веществом для сшивания вышеупомянутого лиганда могут быть любые вещества, которые индуцируют агонистическое действие в отношении клеток посредством образования поперечной связи между первым лигандом и вторым лигандом. Предпочтительными примерами являются антитела, фрагменты антител, (Fab)2 или бивалентные модифицированные антитела. Примерами бивалентных антител являются (Fab)2, димеры одноцепочечного Fv, содержащего одну V-область Н-цепи и одну V-область L-цепи, и одноцепочечные полипептиды, содержащие две V-области Н-цепи и две V-области L-цепи. Этот способ является эффективным для исследования рецепторов, которые передают сигнал в клетки посредством образования поперечных сшивок, ожидается, что он может быть использован для DDS для доставки лекарственного средства к клеткам-мишеням, и является также применимым в качестве системы доставки лекарственного средства, которая подавляет побочное действие и позволяет лекарственному средству становиться эффективным в желаемое время и в течение желаемого периода времени.
Модифицированные антитела данного изобретения могут быть любыми веществами, которые содержат V-область Н-цепи и V-область L-цепи антитела (например, антитела MABL-1, антитела MABL-2, антитела 12 В5, антитела 12Е10 и т.д.) и которые специфически узнают молекулу (молекулы) клеточной поверхности или внутриклеточную молекулу (молекулы), например, белок (рецептор или белок, участвующий в передаче сигнала) или углеводную цепь вышеупомянутого белка или белка клеточной мембраны и сшивают указанные молекулы клеточной поверхности, передавая тем самым сигнал в клетки. Включены также модифицированные антитела, в которых часть аминокислотной последовательности V-области была изменена.
В зависимости от характеристик молекулы клеточной поверхности или внутриклеточной молекулы, подлежащих объединению, например, структуры молекулы или механизма действий, модифицированные антитела могут быть моноспецифическими или полиспецифическими, например, биспецифическими. Когда модифицированное антитело соединяется с рецепторной молекулой, которая гомодимеризуется и передает сигнал в клетки (например, рецептором эритропоэтина, рецептором тромбопоэтина, G-CSF-рецептором, SCF-рецептором, EGF-рецептором, IAP (CD47) и т.п.), предпочтительным является моноспецифическое модифицированное антитело. Когда оно соединяется с рецепторной молекулой, которая гетеродимеризуется и передает сигнал в клетки (например, IL-6-рецептором, LIF-рецептором, IL-11-рецептором), предпочтительным является биспецифическое модифицированное антитело. Когда оно соединяется с рецепторной молекулой, которая гетеротримеризуется и передает сигнал в клетки (например, IL-2-рецептором, CNTF-рецептором, OSM-рецептором), предпочтительным является триспецифическое модифицированное антитело. Способ получения биспецифических одноцепочечных Fv-димеров описан в W094/13804 и т.п.
Данное изобретение относится также к модифицированным антителам, V-область Н-цепи и/или V-область L-цепи которых является V-областью Н-цепи, полученной из антитела человека, и/или V-областью L-цепи, полученной из антитела человека. V-область Н-цепи и/или V-область L-цепи, полученные из антитела человека, могут быть получены скринингом библиотеки моноклональных антител человека, как описано в W099/10494. Включены также V-область Н-цепи и V-область L-цепи, полученные из моноклональных антител человека.
Далее данное изобретение относится к модифицированным антителам, V-область Н-цепи и/или V-область L-цепи которых являются гуманизированными V-областями Н-цепи и/или гуманизированными V-областями L-цепи. Конкретно, гуманизированные модифицированные антитела состоят из гуманизированной V-области L-цепи, которая содержит каркасные области (FR), происходящие из V-области L-цепи моноклонального антитела человека, и определяющих комплементарность (гипервариабельных) участков (далее называемых "CDR"), происходящих из V-области L-цепи моноклонального антитела млекопитающего, отличного от человека (например, мыши, крысы, коровы, овцы, обезьяны), и/или гуманизированной V-области Н-цепи, которая содержит FR, происходящие из V-области Н-цепи моноклонального антитела человека, и CDR, происходящие из V-области Н-цепи моноклонального антитела млекопитающего, отличного от человека, (например, мыши, крысы, коровы, овцы, обезьяны). В этом случае аминокислотная последовательность CDR и FR может быть частично изменена, например, делетирована, заменена или дополнена.
V-области Н-цепи и/или V-области L-цепи модифицированных антител данного изобретения могут быть V-областью Н-цепи и/или V-областью L-цепи, происходящими из моноклональных антител животных, иных чем человек (таких как мышь, крыса, корова, овца, обезьяна, курица и т.п.). В этом случае аминокислотная последовательность CDR и FR может быть частично изменена, например, делетирована, заменена или дополнена.
Данное изобретение относится также к ДНК, кодирующим различные модифицированные антитела, упоминаемые выше, и способам генетической инженерии для получения рекомбинантных векторов, содержащих такие ДНК.
Данное изобретение относится также к клеткам-хозяевам, трансформированным этими рекомбинантными векторами. Примерами клеток-хозяев являются клетки животных, такие как клетки человека, клетки мыши или т.п., и микроорганизмы, такие как Е.coli. Bacillus subtilis, дрожжи или т.п.
Данное изобретение относится к способу получения модифицированных антител, включающему культивирование вышеупомянутых хозяев и экстракцию модифицированных антител из их культуры.
Данное изобретение относится далее к способу получения димера одноцепочечного Fv, включающему культивирование клеток-хозяев животного, продуцирующих одноцепочечный Fv, в бессывороточной среде для секреции одноцепочечного Fv в эту среду и выделение димера одноцепочечного Fv, образовавшегося в среде.
Данное изобретение относится также к применению модифицированных антител в качестве агониста, т.е. оно относится к агонисту передачи сигнала, который содержит в качестве активного ингредиента вышеупомянутое модифицированное антитело. Поскольку модифицированными антителами, используемыми в данном изобретении, являются модифицированные антитела, которые образуют поперечные сшивки между молекулой (молекулами) клеточной поверхности или внутриклеточной молекулой (молекулами) и индуцируют передачу сигнала, такая молекула может быть любой молекулой, которая олигомеризуется, например, димеризуется, посредством соединения с лигандом и тем самым передает сигнал в клетки.
Такая молекула клеточной поверхности включает в себя рецепторы гормонов и рецепторы цитокинов. Рецепторы гормонов включают в себя, например, рецептор эстрогена. Рецептор цитокинов и т.п. включает в себя рецептор гемопоэтического фактора, рецептор лимфокина, рецептор фактора роста, рецептор фактора регуляции дифференцировки и т.п. Примерами рецепторов цитокинов являются рецептор эритропоэтина (ЕРО), рецептор тромбопоэтина (ТРО), рецептор гранулоцитарного колониестимулирующего фактора (G-CSF), рецептор макрофагального колониестимулирующего фактора (M-CSF), рецептор гранулоцитарного-макрофагального колониестимулирующего фактора (GM-CSF), рецептор фактора некроза опухолей (TNF), рецептор интерлейкина-1 (IL-1), рецептор интерлейкина-2 (IL-2), рецептор интерлейкина-3 (IL-3), рецептор интерлейкина-4 (IL-4), рецептор интерлейкина-5 (IL-5), рецептор интерлейкина-6 (IL-6), рецептор интерлейкина-7 (IL-7), рецептор интерлейкина-9 (IL-9), рецептор интерлейкина-10 (IL-10), рецептор интерлейкина-11 (IL-11), рецептор интерлейкина-12 (IL-12), рецептор интерлейкина-13 (IL-13), рецептор интерлейкина-15 (IL-15), рецептор интерферона-альфа (IFN-альфа), рецептор интерферона-бета (IFN-бета), рецептор интерферона-гамма (IFN-гамма), рецептор гормона роста (GH), рецептор инсулина, рецептор фактора пролиферации стволовых клеток крови (SCF), рецептор фактора роста эндотелия сосудов (VEGF), рецептор эпидермального фактора роста (EGF), рецептор фактора роста нервов (NGF), рецептор фактора роста фибробластов (FGF), рецептор тромбоцитарного фактора роста (PDGF), рецептор трансформирующего фактора роста бета (TGF-бета), рецептор фактора ингибирования миграции лейкоцитов (LIF), рецептор цилиарного нейротрофического фактора (CNTF), рецептор онкостатина М (OSM), рецептор Notch-семейства (семейства больших трансмембранных рецепторов) и т.п.
Молекула внутриклеточной поверхности включает ТАК1, ТАВ1 и т.п. ТАК1 и ТАВ1 функционируют в пути передачи сигнала TGF-β, активируют МАР-киназу путем образования гетеродимера и передают ряд сигналов. Многие раковые клетки имеют мутацию рецептора TGF-β, который репрессирует рост рака и, следовательно, сигнал TGF-β не передается. Модифицированные антитела, которые могут передавать сигнал посредством образования поперечных связей между ТАК1 и ТАВ1, могут индуцировать этот сигнал TGF-β через агонистическое действие посредством соединения ТАК1/TAB1. Такие модифицированные антитела данного изобретения могут ингибировать рост TGF-β-устойчивых раковых клеток и обеспечивают новый способ для терапии рака. Другими примерами внутриклеточной молекулы являются гомодимер фактора транскрипции E2F и гетеродимер E2F/DP1, обладающие пролиферативным действием на клетки. Модифицированные антитела данного изобретения могут индуцировать агонистическое действие также на эти молекулы и, следовательно, могут быть использованы для лечения различных связанных с пролиферацией клеток (клеточно-пролиферативных) заболеваний. Модифицированные антитела данного изобретения могут индуцировать агонистическое действие посредством образования поперечных сшивок внутриклеточного фактора, участвующего в передаче сигнала, связанного с индукцией апоптоза, и, следовательно, индуцировать апоптозную гибель раковых клеток или связанных с аутоиммунным заболеванием клеток.
Для обеспечения взаимодействия модифицированных антител данного изобретения с внутриклеточной молекулой, пептиды, способные проникать через клеточную мембрану, (например, Пегелин, Пенетратин), могут быть использованы для транспорта модифицированных антител в клетки (Martine Mazel et al., Doxorubicin-peptide conjugates overcome multidrug resistance. Anti-Cancer Drugs 2001, 12, Dccrossi D. et al., The third helix of the antennapedia homeodomain translocates through biological membranes, J. Biol. Chem. 1994, 269, 10444-10450).
Таким образом, фармацевтические препараты, содержащие агонистическое модифицированное антитело в качестве активного ингредиента, применимы в качестве профилактических средств и/или лечебных средств и т.д. для различных заболеваний, таких как раки, воспаление, гормональные нарушения, заболевания крови и аутоиммунные заболевания.
Олигомеры, которые могут быть образованы рецепторными белками, могут быть гомо-олигомерами или гетеро-олигомерами и любыми олигомерами, такими как димеры, тримеры и тетрамеры. Известно, например, что рецептор эритропоэтина, рецептор тромбопоэтина, рецептор G-CSF, рецептор SCF, рецептор EGF и т.п. образуют гомодимеры, что рецептор IL-6, рецептор LIF и рецептор IL-11 образуют гетеродимеры и что рецептор IL-2, рецептор CNTF, рецептор OSM образуют гетеротримеры.
Модифицированные антитела данного изобретения содержат две или более V-областей Н-цепи и две или более V-областей L-цепи, происходящие из моноклональных антител. Структура этих модифицированных антител может быть представлена димером одноцепочечного Fv, содержащего одну V-область Н-цепи и одну V-область L-цепи, или полипептидом, содержащим две V-области Н-цепи и две V-области L-цепи. В модифицированных антителах данного изобретения V-области Н-цепи и L-цепи предпочтительно связаны через пептидный линкер, который состоит из одной или более аминокислот.Полученные модифицированные антитела содержат вариабельные области антител и связываются с антигеном с такой же специфичностью, что и специфичность исходных моноклональных антител.
V-область Н-цепи
В данном изобретении V-область Н-цепи, полученная из антитела, узнает молекулу (молекулы) клеточной поверхности или внутриклеточную молекулу (молекулы), например, белок (рецептор или связанный с передачей сигнала белок) или углеводную цепь этого белка или на клеточной мембране и олигомеризует, например, димеризует указанную молекулу через образование поперечных сшивок и тем самым трансдуцирует сигнал в клетки. V-область Н-цепи данного изобретения включает в себя V-области Н-цепи, происходящие из млекопитающего (например, человека, мыши, крысы, коровы, овцы, обезьяны и т.д.), и V-области Н-цепи, имеющие частично модифицированные аминокислотные последовательности V-областей Н-цепи. Более предпочтительной является гуманизированная V-область Н-цепи, содержащая FR V-области Н-цепи моноклонального антитела человека и CDR V-области Н-цепи моноклонального антитела мыши. Также предпочтительной является V-область Н-цепи, имеющая аминокислотную последовательность, происходящую из человека, которая может быть получена рекомбинантным способом. V-область Н-цепи данного изобретения может быть фрагментом V-области Н-цепи, который сохраняет антигенсвязывающую способность.
V-область L-цепи
В данном изобретении V-область L-цепи узнает молекулу (молекулы) клеточной поверхности или внутриклеточную молекулу (молекулы), например, белок (рецептор или связанный с трансдукцией сигнала белок) или углеводную цепь этого белка или на клеточной мембране и олигомеризует, например, димеризует указанную молекулу через образование поперечных сшивок и тем самым трансдуцирует сигнал в клетки. V-область L-цепи данного изобретения включает в себя V-области L-цепи данного изобретения, происходящие из млекопитающего (например, человека, мыши, крысы, коровы, овцы, обезьяны и т.д.), и V-области L-цепи, имеющие частично модифицированные аминокислотные последовательности V-областей L-цепи. Более предпочтительной является гуманизированная V-область L-цепи, содержащая FR V-области L-цепи моноклонального антитела человека и CDR V-области L-цепи моноклонального антитела мыши. Также предпочтительной является V-область L-цепи, имеющая аминокислотную последовательность, происходящую из антитела человека, которая может быть получена рекомбинантным способом. V-области L-цепи данного изобретения могут быть фрагментами вышеупомянутой V-области L-цепи, которые сохраняют антигенсвязывающую способность.
Определяющий комплементарность (гипервариабельный) участок (CDR)
Каждая V-область L-цепи и Н-цепи образует антигенсвязывающий сайт. Вариабельная область L- и Н-цепей состоит из сравнительно консервативных четырех общих каркасных областей, соединенных с тремя гипервариабельными участками, или определяющими комплементарность участками (CDR) (Kabat, E.A. et al., "Sequence of Protein of Immunological Interest", US Dept. Health and Human Services, 1983).
Основные части этих четырех каркасных областей (FR) образуют β-складчатые структуры и три CDR, таким образом, образуют петлю. В определенных случаях CDR могут образовывать часть этой β-складчатой структуры. Эти три CDR удерживаются в стерически близком положении относительно друг друга посредством каркасной области FR, которая вносит вклад в образование антигенсвязывающего сайта вместе с тремя CDR.
Эти CDR могут быть идентифицированы сравнением аминокислотной последовательности V-области полученного антитела с известными аминокислотными последовательностями V-областей известных антител в соответствии с эмпирическим правилом Kabat, E.A. et al., "Sequence of Protein of Immunological Interest".
Одноцепочечный Fv-фрагмент
Одноцепочечный Fv-фрагмент является полипептидным мономером, содержащим связанные друг с другом V-область Н-цепи и V-область L-цепи, которые происходят из моноклональных антител. Полученные одноцепочечные Fv содержат вариабельные области исходных ("родительских") антител и сохраняют их определяющий комплементарность район и, следовательно, одноцепочечные Fv связываются с антигеном с той же самой специфичностью, что и специфичность исходных моноклональных антител (Патентная заявка Японии JP-Appl. 11-63557). Часть вариабельной области и/или CDR одноцепочечного Fv данного изобретения может быть частично изменена, например, делетирована, заменена или дополнена. V-область Н-цепи и V-область L-цепи, составляющие одноцепочечный Fv данного изобретения, упоминаются выше и могут быть связаны непосредственно или через линкер, предпочтительно через пептидный линкер. Строение одноцепочечного Fv может быть следующим: [V-область Н-цепи]-[V-область L-цепи] или [V-область L-цепи]-[V-область Н-цепи]. В данном изобретении можно добиться того, чтобы одноцепочечный Fv образовывал димер, тример или тетрамер, из которых может быть получено модифицированное антитело данного изобретения.
Одноцепочечное модифицированное антитело
Одноцепочечные модифицированные антитела данного изобретения, содержащие две или более V-областей Н-цепи и две или более V-областей L-цепи, предпочтительно две-четыре каждой, особенно предпочтительно две каждой, содержат две или более V-областей Н-цепи и V-областей L-цепи, как упоминалось выше. Каждая область этого пептида должна быть расположена таким образом, что модифицированное одноцепочечное антитело образует специфическую стерическую структуру, конкретно имитирующую стерическую структуру, образуемую димером одноцепочечного Fv.
Например, V-области размещают в следующем порядке:
[V-область Н-цепи]-[V-область L-цепи]-[V-область Н-цепи]-[V-область L-цепи] или [V-область L-цепи]-[V-область Н-цепи]-[V-область L-цепи]-[V-область Н-цепи], где эти области соединены через пептидный линкер, соответственно.
Линкер
В данном изобретении линкеры для соединения между V-областью Н-цепи и V-областью L-цепи могут быть любым пептидным линкером, который может быть введен методом генетической инженерии, или любым химически синтезированным линкером. Например, в данном изобретении могут быть использованы линкеры, описанные в литературе, например, Protein Engineering, 9(3), 299-305, 1996. Эти линкеры могут быть одинаковыми или различными в одной и той же молекуле. Если необходимы пептидные линкеры, в качестве примеров линкеров приводятся следующие:
Ser
Gly-Ser
Gly-Gly-Ser
Ser-Gly-Gly
Gly-Gly-Gly-Ser
Ser-Gly-Gly-Gly
Gly-Gly-Gly-Gly-Ser
Ser-Gly-Gly-Gly-Gly
Gly-Gly-Gly-Gly-Gly-Ser
Ser-Gly-Gly-Gly-Gly-Gly
Gly-Gly-Gly-Gly-Gly-Gly-Ser
Ser-Gly-Gly-Gly-Gly-Gly-Gly
(Gly-Gly-Gly-Gly-Ser)n
(Ser-Gly-Gly-Gly-Gly)n
где n равно целому числу не менее 1. Предпочтительная длина линкерного пептида варьируется в зависимости от рецептора, который служит антигеном, в случае одноцепочечных Fv обычно предпочтительным является диапазон 1-20 аминокислот. В случае одноцепочечных модифицированных антител, содержащих две или более V-областей Н-цепи и две или более V-областей L-цепи, пептидные линкеры, соединяющие области, образующие один и тот же антигенсвязывающий сайт, содержащий [V-область Н-цепи]-[V-область L-цепи] (или [V-область L-цепи]-[V-область Н-цепи]), имеют длины 1-30 аминокислот, предпочтительно, 1-20 аминокислот, более предпочтительно 3-18 аминокислот. Пептидные линкеры, соединяющие области, не образующие один и тот же антигенсвязывающий сайт, содержащий [V-область Н-цепи]-[V-область L-цепи] (или [V-область L-цепи]-[V-область Н-цепи]), имеют длины 1-40 аминокислот, предпочтительно 3-30 аминокислот, более предпочтительно 5-20 аминокислот. Способ введения этих линкеров будет описан в объяснении для ДНК-конструкций, кодирующих модифицированные антитела данного изобретения.
Химически синтезированные линкеры, т.е. химические агенты, образующие поперечные связи, в соответствии с данным изобретением могут быть любыми линкерами, обычно используемыми для соединения пептидов. Примеры этих линкеров могут включать в себя N-гидроксисукцинимид (NHS), дисукцинимидилсуберат (DSS), бис(сульфосукцинимидил)суберат (BS3), дитиобис(сукцинимидилпропионат) (DSP), дитиобис(сульфосукцинимидилпропионат) (DTSSP), этиленгликольбис(сукцинимидилсукцинат) (EGS), этиленгликольбис(сульфосукцинимидилсукцинат) (сульфо-EGS), дисукцинимидилтартрат (DST), дисульфосукцинимидилтартрат (сульфо-DST), бис[2-(сукцинимидооксикарбонилокси)этил]сульфон (BSOCOES), бис[2-(сульфосукцинимидооксикарбонилокси)этил]сульфон (сульфо-BSOCOES) или т.п. Они являются коммерчески доступными. В случае химических линкеров предпочтительно, чтобы они имели длину, эквивалентную длине пептидных линкеров.
Для образования димера одноцепочечного Fv предпочтительно выбирать линкер, подходящий для димеризации в растворе, таком как культуральная среда, более чем 20%, предпочтительно более чем 50%, еще более предпочтительно более чем 80%, наиболее предпочтительно более чем 90% одноцепочечного Fv, продуцируемого в клетках-хозяевах. Конкретно, предпочтительным являются линкер, состоящий из 2-12 аминокислот, предпочтительно 3-10 аминокислот, или другие линкеры, соответствующие им.
Получение модифицированных антител
Модифицированные антитела могут быть получены соединением через вышеупомянутый линкер, V-области Н-цепи и V-области L-цепи, полученных из известных или новых моноклональных антител, специфически связывающихся с молекулой (молекулами) клеточной поверхности. В качестве примеров одноцепочечных Fv приводятся MABL1-scFv и MABL2-scFv, содержащие V-область Н-цепи и V-область L-цепи, происходящие из антитела MABL-1 и антитела MABL-2, соответственно. В качестве примеров одноцепочечных полипептидов, содержащих две V-области Н-цепи и две V-области L-цепи, приводятся MABL1-sc(Fv)2 и MABL2-sc(Fv)2, содержащие V-область Н-цепи и V-область L-цепи, происходящие из вышеупомянутых антител.
Для получения полипептида к N-концу этого полипептида может быть присоединен сигнальный пептид, если желательно, чтобы данный полипептид был секретируемым пептидом. Хорошо известная аминокислотная последовательность, пригодная для очистки полипептида, такая как FLAG-последовательность, может быть присоединена для эффективной очистки этого полипептида. В этом случае димер может быть образован с использованием анти-FLAG-антитела.
Для получения модифицированного антитела данного изобретения необходимо получить ДНК, т.е. ДНК, кодирующую одноцепочечный Fv, или ДНК, кодирующую реконструированный одноцепочечный полипептид. Эти ДНК, в частности, для MABL1-scFv, MABL2-scFv, MABLl-sc(Fv)2 и/или MABL2-sc(Fv)2, могут быть получены из ДНК, кодирующих V-область Н-цепи и V-область L-цепи, происходящие из указанного Fv. Они могут быть также получены способом полимеразной цепной реакцией (ПЦР) с использованием этих ДНК в качестве матрицы и амплификацией части содержащейся в ней ДНК, кодирующей желаемую аминокислотную последовательность, с использованием пары праймеров, соответствующих обоим концам ДНК.
В том случае, когда желательной является каждая V-область, имеющая частично модифицированную аминокислотную последовательность, V-области, в которых одна или несколько аминокислот являются модифицированными, т.е. делетированными, замененными или добавленными, могут быть получены процедурой, известной в данной области, с использованием ПЦР. Часть аминокислотной последовательности в V-области является предпочтительно модифицированной при помощи ПЦР, известной в данной области, для получения модифицированного антитела, которое является достаточно активным против специфического антигена.
Для определения праймеров для ПЦР-амплификации необходимо определить тип Н-цепи и L-цепи желаемых антител. Однако в случае антитела MABL-1 и антитела MABL-2 сообщалось, что антитело MABL-1 имеет L-цепи к-типа и Н-цепи γ1-типа, а антитело MABL-2 имеет L-цепи к-типа и Н-цепи γ2а-типа (Патентная заявка Японии JP-Appl. 11-63557). Для ПЦР-амплификации ДНК, кодирующей Н-цепь и L-цепь антитела MABL-1 и/или антитела MABL-2, могут быть использованы праймеры, описанные Jones, S.T. et al., Bio/Technology, 9, 88-89, 1991.
Для амплификации V-областей L-цепей антитела MABL-1 и антитела MABL-2 при помощи ПЦР были выбраны 5'-концевой и 3'-концевой олигонуклеотидные праймеры, как упоминалось ранее. Таким же образом, были выбраны 5'-концевой и 3'-концевой олигонуклеотидные праймеры для амплификации V-областей Н-цепей антитела MABL-1 и антитела MABL-2.
В вариантах данного изобретения использовали 5'-концевые праймеры, которые содержат последовательность "GANTC", обеспечивающую сайт узнавания рестриктазы Hinfl вблизи их 5'-конца, и 3'-концевые праймеры, которые содержат нуклеотидную последовательность "CCCGGG", обеспечивающую сайт узнавания рестриктазы Xmal вблизи их 5'-конца. Другой сайт узнавания рестриктазы может быть использован вместо этих сайтов, пока они используются для субклонирования желаемого ДНК-фрагмента в клонирующий вектор.
Сконструированные специфическим образом ПЦР-праймеры используют для обеспечения подходящих нуклеотидных последовательностей на 5'-конце и 3'-конце кДНК, кодирующих V-области антител MABL-1 и MABL-2 таким образом, что эти кДНК легко встраиваются в экспрессионный вектор и правильно функционируют в этом экспрессионном векторе (например, данное изобретение предусматривает увеличение эффективности трансляции за счет встраивания последовательности Козака). V-области антител MABL-1 и MABL-2, полученные амплификацией при помощи ПЦР с использованием этих праймеров, встраивают в экспрессионный вектор HEF, содержащий желаемую С-область человека (см. WO 92/19759). Клонированные ДНК могут быть секвенированы с использованием общепринятого способа, например, при помощи автоматического ДНК-секвенатора (Applied Biosystems).
Линкер, такой как пептидный линкер, может быть введен в модифицированное антитело данного изобретения следующим образом. Конструируют праймеры, которые имеют частично комплементарную последовательность с праймерами для V-областей Н-цепей и V-областей L-цепей, описанных выше, и которые кодируют N-конец и С-конец линкера. Затем проводят процедуру ПЦР с использованием этих праймеров для получения ДНК, кодирующей пептидный линкер, имеющий желаемую аминокислотную последовательность и длину. ДНК, кодирующие V-область Н-цепи и V-область L-цепи, могут быть соединены через полученную ДНК для получения ДНК, кодирующей модифицированное антитело данного изобретения, которое имеет желаемый пептидный линкер. После получения ДНК, кодирующей одно из модифицированных антител, ДНК, кодирующие модифицированные антитела с желаемым пептидным линкером или без пептидного линкера, могут быть легко получены путем конструирования различных праймеров для этого линкера и затем проведения ПЦР с использованием этих праймеров и вышеупомянутой ДНК в качестве матрицы.
Каждая V-область модифицированного антитела данного изобретения может быть гуманизирована с использованием общепринятых способов (например, Sato, К. et al., Cancer Res., 53, 1-6 (1993)). После получения ДНК, кодирующей каждый из гуманизированных Fv, гуманизированный одноцепочечный Fv, фрагмент гуманизированного одноцепочечного Fv, гуманизированное моноклональное антитело и фрагмент гуманизированного моноклонального антитела могут быть легко получены в соответствии с общепринятыми способами. Предпочтительно, аминокислотные последовательности их V-областей могут быть частично модифицированными, если требуется.
Кроме того, ДНК, полученная из другого млекопитающего, например, ДНК, кодирующая V-области антитела человека, может быть получена таким же способом, как при получении ДНК, кодирующей V-область Н-цепи и V-область L-цепи, происходящие из мыши, общепринятыми способами, как упоминалось выше. Полученная ДНК может быть использована для получения V-области Н-цепи и V-области L-цепи другого млекопитающего, в частности, происходящих из антитела человека, одноцепочечного Fv, происходящего из человека, и его фрагмента, и моноклонального антитела человеческого происхождения и его фрагмента.
В том случае, когда модифицированные антитела данного изобретения являются биспецифическими модифицированными антителами, они могут быть получены известными способами (например, способом, описанным в WO 94/13804).
Как упоминалось выше, когда получают целевые ДНК, кодирующие V-области модифицированных антител и V-области гуманизированных модифицированных антител, экспрессионные вектора, содержащие их, и хозяева, трансформированные этими векторами, могут быть получены в соответствии с общепринятыми способами. Далее эти хозяева могут культивироваться в соответствии с общепринятым способом для получения реконструированного одноцепочечного Fv, реконструированного гуманизированного одноцепочечного Fv, гуманизированных моноклональных антител и их фрагментов. Они могут быть выделены из клеток или среды и могут быть очищены до гомогенной массы. Для этой цели могут быть использованы в комбинации любые способы выделения и очистки, обычно используемые для белков, например, хроматография, ультрафильтрация, высаливание и диализ, если необходимо, без ограничения.
При получении реконструированного одноцепочечного Fv данного изобретения культивированием клеток животного, таких как клетки COS7 или клетки СНО, предпочтительно клетки СНО, в бессывороточной среде, димер указанного одноцепочечного Fv, образованный в этой среде, может быть стабильно извлечен и очищен с высоким выходом. Очищенный таким образом димер может стабильно сохраняться в течение продолжительного периода. Бессывороточной средой, используемой в данном изобретении, может быть любая среда, обычно используемая для получения рекомбинантного белка, без ограничений.
Для получения модифицированных антител данного изобретения могут использоваться любые экспрессионные системы, например, эукариотические клетки, такие как клетки животных, например, адаптированные клеточные линии млекопитающих, нитчатые грибы и дрожжи, и прокариотические клетки, такие как бактериальные клетки, например, Е.coli. Предпочтительно, модифицированные антитела данного изобретения экспрессируют в клетках млекопитающих, например, клетках COS7 или клетках СНО.
В этих случаях могут быть использованы общепринятые промоторы, применимые для экспрессии в клетках млекопитающих. Предпочтительно, используют немедленно ранний промотор цитомегаловируса человека (HCMV). Экспрессионные вектора, содержащие промотор HCMV, включают в себя HCMV-VH-HCγ1, HCMV-VL-HCK и т.п., которые получены из pSV2neo (WO 92/19759).
Дополнительно, другие промоторы, которые могут быть использованы для экспрессии генов в клетках млекопитающих в данном изобретении, включают в себя вирусные промоторы, полученные из ретровируса, полиомавируса, аденовируса и обезьяньего вируса 40 (SV40), и промоторы, происходящие из млекопитающих, такие как промотор фактора-1α элонгации полипептидной цепи человека (HEF-1α). Промотор SV40 может быть легко использован в соответствии со способом Mulligan, R.C., et al. (Nature 277, 108-114 (1979)), а промотор HEF-12 может быть также использован в соответствии со способами Mizushima, S. et al. (Nucleic Acids Research, 18, 5322 (1990)).
Сайт инициации репликации (ориджин репликации) (ori), который может быть использован в данном изобретении, включает в себя ori, происходящий из SV40, полиомавируса, аденовируса, папилломавируса крупного рогатого скота (BPV) и т.п. Экспрессионный вектор может содержать в качестве селективного маркера ген II или I (пео) фосфотрансферазы АРН (3'), ген тимидинкиназы (ТК), ген ксантин-гуанинфосфорибозилтрансферазы (ECOGPT) Е.coli или ген дигидрофолатредуктазы (DHFR).
Антигенсвязывающая активность модифицированного антитела, полученного, как описано выше, может оцениваться общепринятым способом, таким как радиоиммуноанализ (РИА), твердофазный иммуноферментный анализ (ELISA) или поверхностный плазменный резонанс. Она может также оцениваться с использованием способности исходных антител к связыванию-ингибированию в качестве индекса, например, по отсутствию или присутствию зависимого от концентрации ингибирования связывания указанного моноклонального антитела с антигеном.
Более конкретно, культивируют клетки животных, трансформированные экспрессионным вектором, содержащим ДНК, кодирующую модифицированное антитело данного изобретения, например, клетки COS7 или клетки СНО. Культивированные клетки и/или супернатант среды или модифицированное антитело, очищенное из них, используют для определения связывания с антигеном. В качестве контроля используют супернатант культуральной среды, в которой культивировали клетки, трансформированные только экспрессионным вектором. В случае антигена, например, антитело MABL-1 и антитело MABL-2, исследуемый образец модифицированного антитела данного изобретения или супернатант контроля добавляют к линии клеток мышиного лейкоза, клеткам L1210, экспрессирующим IAP человека, и затем проводят анализ, такой как проточная цитометрия, для оценки антигенсвязывающей активности.
Оценку in vitro эффекта трансдукции сигнала (эффекта индукции апоптоза в случаях антител MABL-1 и антител MABL-2) проводят следующим образом: анализируемый образец модифицированного, как описано выше, антитела добавляют к клеткам, которые экспрессируют антитело, или клеткам, в которые был введен ген антитела, и оценивают изменения, вызываемые трансдукцией сигнала, например, индуцируется ли гибель клеток специфическим для IAP-антигена человека образом, с использованием общепринятых способов.
Оценку in vivo эффекта индукции апоптоза, например, в случае, когда модифицированное антитело узнает IAP человека (например, модифицированные антитела - производные антитела MABL-1 и антитела MABL-2), проводят следующим образом: получают мышиную модель миеломы человека. Мышам вводят внутривенно моноклональное антитело или модифицированное антитело данного изобретения, которое индуцирует апоптоз ядросодержащих клеток крови, содержащих IAP. Мышам контрольной группы вводят только фосфатно-солевой буферный раствор (PBS).
Индукцию апоптоза оценивают по противоопухолевому действию на основе изменения содержания IgG человека в сыворотке мышей и времени их выживания.
Как упоминалось выше, модифицированные антитела данного изобретения могут быть получены путем создания модифицированных антител, которые содержат две или более V-областей Н-цепи и две или более V-областей L-цепи и специфически связываются с молекулой-мишенью клеточной поверхности или внутриклеточной молекулой, и скрининга этих модифицированных антител с оценкой in vivo и in vitro, как описано выше.
Модифицированные антитела данного изобретения, которые содержат две или более V-областей Н-цепи и две или более V-областей L-цепи, предпочтительно две-четыре каждой, более предпочтительно две каждой, могут быть димером одноцепочечного Fv, содержащего одну V-область Н-цепи и одну V-область L-цепи, или одноцепочечным полипептидом, в котором соединены две или более V-областей Н-цепи и две или более V-областей L-цепи. Предполагается, что вследствие такой конструкции этот пептид имитирует трехмерную структуру природного лиганда и, следовательно, сохраняет превосходную антигенсвязывающую способность и агонистическую активность.
Модифицированные антитела данного изобретения имеют значительно уменьшенный молекулярный размер в сравнении с молекулой антитела (цельного IgG) и, следовательно, лучшую способность проникновения в ткани и опухоли и более высокую активность, чем исходные агонистические моноклональные антитела. Таким образом, правильный выбор исходного антитела позволяет трансдуцировать различные сигналы в клетки и индуцировать различные действия в этих клетках, такие как индукция апоптоза, индукция пролиферации клеток, индукция дифференцировки клеток, индукция клеточного деления или действие на регуляцию клеточного цикла. Фармацевтические препараты, содержащие их, применимы для лечения заболеваний, которые могут излечиваться индукцией трансдукции сигнала, например, раков, воспаления, гормональных нарушений, аутоиммунных заболеваний, а также патологического изменения крови, например, лейкоза, злокачественной лимфомы, апластической анемии, синдрома миелодисплазии и красной, или истинной полицитемии. Ожидается также, что антитело данного изобретения может быть использовано в качестве контрастного агента посредством RI-мечения. Этот эффект может быть усилен присоединением к RI-соединению или токсину.
Перечень фигур
Фиг.1 показывает результат проточной цитометрии, иллюстрирующий, что IgG-антитело человека не связывается с клетками L1210, экспрессирующими IAP человека (hIAP/L1210).
Фиг.2 показывает результат проточной цитометрии, иллюстрирующий, что химерное антитело MABL-1 специфически связывается с клетками L1210, экспрессирующими IAP человека (hIAP/L1210).
Фиг.3 показывает результат проточной цитометрии, иллюстрирующий, что химерное антитело MABL-2 специфически связывается с клетками L1210, экспрессирующими IAP человека (hIAP/L1210).
Фиг.4 схематически иллюстрирует способ получения одноцепочечного Fv в соответствии с данным изобретением.
Фиг.5 иллюстрирует структуру экспрессионной плазмиды, которая может быть использована для экспрессии ДНК, кодирующей одноцепочечный Fv данного изобретения, в Е.coli.
Фиг.6 иллюстрирует структуру экспрессионной плазмиды, которая используется для экспрессии ДНК, кодирующей одноцепочечный Fv данного изобретения, в клетках млекопитающх.
Фиг.7 показывает результат вестерн-блоттинга примера 5.4. Слева направо, маркер молекулярных масс (который соответствует 97,4, 66, 45, 31, 21,5 и 14,5 кДа сверху вниз), культуральный супернатант клеток COS7 с введенной рСНО1 и культуральный супернатант клеток COS7 с введенной рСНОМ2. Он иллюстрирует, что реконструированный одноцепочечный Fv антитела MABL-2 (показан стрелкой) содержится в культуральном супернатанте клеток с введенной рСНОМ2.
Фиг.8 показывает результат проточной цитометрии, иллюстрирующий, что антитело из культурального супернатанта клеток pCHO1/COS7 в качестве контроля не связывается с клетками pCOS1/L1210 в качестве контроля.
Фиг.9 показывает результат проточной цитометрии, иллюстрирующий, что антитело из культурального супернатанта клеток MABL2-scFv/COS7 не связывается с клетками pCOS1/L1210 в качестве контроля.
Фиг.10 показывает результат проточной цитометрии, иллюстрирующий, что антитело из культурального супернатанта клеток pCOS1/COS7 не связывается с клетками hIAP/L1210.
Фиг.11 показывает результат проточной цитометрии, иллюстрирующий, что антитело из культурального супернатанта клеток MABL2-scFv/COS7 специфически связывается с клетками hIAP/L1210.
Фиг.12 показывает результат конкурентного ELISA в примере 5.6, где связывающая активность одноцепочечного Fv данного изобретения (MABL2-scFv) в отношении антигена выявляется по ингибированию связывания мышиного моноклонального антитела MABL-2 с этим антигеном в качестве показателя, в сравнении с культуральным супернатантом клеток pCHO1/COS7 в качестве контроля.
Фиг.13 показывает результаты апоптоз-индуцирующего действия в примере 5.7, иллюстрирующие, что антитело из культурального супернатанта клеток pCHO1/COS7 в качестве контроля не индуцирует апоптоз клеток pCOS1/L1210 в качестве контроля.
Фиг.14 показывает результаты апоптоз-индуцирующего действия в примере 5.7, иллюстрирующие, что антитело из культурального супернатанта клеток MABL2-scFv/COS7 не индуцирует апоптоз клеток pCOS1/L1210 в качестве контроля.
Фиг.15 показывает результаты апоптоз-индуцирующего действия в примере 5.7, иллюстрирующие, что антитело из культурального супернатанта клеток pCHO1/COS7 в качестве контроля не индуцирует апоптоз клеток hIAP/L1210.
Фиг.16 показывает результаты апоптоз-индуцирующего действия в примере 5.7, иллюстрирующие, что антитело из культурального супернатанта клеток MABL2-scFv/COS7 специфически индуцирует апоптоз клеток hIAP/L1210.
Фиг.17 показывает результаты апоптоз-индуцирующего действия в примере 5.7, иллюстрирующие, что антитело из культурального супернатанта клеток pCHO1/COS7 в качестве контроля не индуцирует апоптоз клеток CCRF-CEM (в конечной концентрации 50%).
Фиг.18 показывает результаты апоптоз-индуцирующего действия в примере 5.7, иллюстрирующие, что антитело из культурального супернатанта клеток MABL2-scFv/COS7 специфически индуцирует апоптоз клеток CCRF-CEM (в конечной концентрации 50%).
Фиг.19 показывает хроматограмму, полученную при очистке одноцепочечного Fv, происходящего из антитела MABL-2, продуцируемого клетками СНО в примере 5.9, иллюстрирующую, что фракция А и фракция В были получены в качестве основных пиков при очистке фракции после колонки с Blue-Sepharose на гидроксиапатитной колонки.
Фиг.20 показывает результаты очистки гель-фильтрацией фракции А и фракции В, полученных в примере 5.9-(2), иллюстрирующие, что основные пики (AI и BI, соответственно) элюировались в случае фракции А при кажущейся молекулярной массе приблизительно 36 кД, а в случае фракции В при приблизительно 76 кД.
Фиг.21 является анализом электрофореза в ДСН-ПААГ фракций, полученных при очистке одноцепочечного Fv, происходящего из антитела MABL-2, продуцируемого клетками СНО, в примере 5.9, иллюстрирующим, что единственная полоса с молекулярной массой около 35 кД наблюдалась в обеих фракциях.
Фиг.22 показывает результаты анализа фракций AI и BI, полученных гель-фильтрацией при очистке одноцепочечного Fv, происходящего из антитела MABL-2, продуцируемого клетками СНО, где фракция AI содержит мономер, а фракция BI содержит димер.
Фиг.23 иллюстрирует структуру экспрессионной плазмиды, которая может быть использована для экспрессии ДНК, кодирующей одноцепочечный Fv данного изобретения, в Е.coli.
Фиг.24 показывает очистку на гель-фильтрационной колонке неочищенных продуктов одноцепочечного полипептида Fv, происходящего из антитела MABL-2, продуцируемого Е.coli, полученных в примере 5.12, где каждый пик соответствует мономеру или димеру, соответственно, одноцепочечного Fv, продуцируемого Е.coli.
Фиг.25 показывает результаты апоптоз-индуцирующего действия в примере 5.13, иллюстрирующие, что мышиное IgG-антитело в качестве контроля не индуцирует апоптоз клеток hIAP/L1210 (конечная концентрация 3 мкг/мл).
Фиг.26 показывает результаты апоптоз-индуцирующего действия в примере 5.13, иллюстрирующие, что димер MABL2-scFv, продуцируемый клетками СНО, в сильной степени индуцирует апоптоз клеток hIAP/L1210 (конечная концентрация 3 мкг/мл).
Фиг.27 показывает результаты апоптоз-индуцирующего действия в примере 5.13, иллюстрирующие, что димер MABL2-scFv, продуцируемый Е.coli, в сильной степени индуцирует апоптоз клеток hIAP/L1210 (конечная концентрация 3 мкг/мл).
Фиг.28 показывает результаты апоптоз-индуцирующего действия в примере 5.13, иллюстрирующие, что индукция апоптоза в отношении клеток hIAP/L1210 мономером MABL2-scFv, продуцируемым клетками СНО, находится на том же уровне, что и в контроле (конечная концентрация 3 мкг/мл).
Фиг.29 показывает результаты апоптоз-индуцирующего действия в примере 5.13, иллюстрирующие, что индукция апоптоза в отношении клеток hIAP/L1210 мономером MABL2-scFv, продуцируемым Е.coli, находится на том же уровне, что и в контроле (конечная концентрация 3 мкг/мл).
Фиг.30 показывает результаты апоптоз-индуцирующего действия в примере 5.13, иллюстрирующие, что мышиное IgG-антитело, используемое в качестве контроля, не индуцирует апоптоз клеток hIAP/L1210 даже при добавлении анти-FLAG-атитела (конечная концентрация 3 мкг/мл).
Фиг.31 показывает результаты апоптоз-индуцирующего действия в примере 5.13, иллюстрирующие, что мономер MABL2-scFv, продуцируемый клетками СНО, в сильной степени индуцирует апоптоз клеток hIAP/L1210 при добавлении анти-FLAG-антитела (конечная концентрация 3 мкг/мл).
Фиг.32 показывает результаты количественного измерения IgG человека в сыворотке мыши, в которую трансплантировали клеточную линию КРММ2 миеломы человека, показывающие количества IgG человека, продуцирумого клетками миеломы человека в этой мыши. Эта фигура иллюстрирует, что димер scFv/CHO в сильной степени ингибирует рост клеток КРММ2.
Фиг.33 показывает время выживания мыши после трансплантации опухоли, иллюстрируя, что время выживания в группе мышей с введенным димером scFv/CHO было значительно большим.
Фиг.34 иллюстрирует структуру экспрессионной плазмиды, которая экспрессирует модифицированное антитело [sc(Fv)2], содержащее две V-области Н-цепи и две V-области L-цепи, происходящее из антитела MABL-2.
Фиг.35 иллюстрирует структуру плазмиды, которая экспрессирует scFv (HL-типа).
где V-области соединены по типу [Н-цепь]-[L-цепь] без пептидного линкера.
Фиг.36 иллюстрирует структуру полипептида HL-типа и аминокислотные последовательности пептидных линкеров.
Фиг.37 иллюстрирует структуру плазмиды, которая экспрессирует scFv (LH-типа), где V-области соединены по типу [L-цепь]-[Н-цепь] без пептидного линкера.
Фиг.38 иллюстрирует структуру полипептида LH-типа и аминокислотные последовательности пептидных линкеров.
Фиг.39 показывает результаты вестерн-блоттинга в примере 6.4, иллюстрирующие, что экспрессируются модифицированное антитело sc(Fv)2, содержащее две V-области Н-цепи и две V-области L-цепи, и MABL2-scFv, содержащие пептидные линкеры разной длины.
Фиг.40а и 40b показывают результаты проточной цитометрии с использованием культуральных супернатантов клеток COS7, полученных в примере 6.3 (1), иллюстрирующие, что MABL2-scFv и sc(Fv)2, содержащие пептидные линкеры различной длины, имеют высокое сродство к IAP человека.
Фиг.41 а и 41b показывают результаты апоптоз-индуцирующего действия в примере 6.6, иллюстрирующие, что scFv <HL3, 4, 6, 7, LH3, 4, 6 и 7> и sc(Fv)2 в сильной степени индуцируют гибель клеток hIAP/L1210.
Фиг.42 показывает результаты оценки антигенсвязывающей способности в примере 6.10, иллюстрирующие, что димер scFv <HL5> и sc(Fv)2 имеют высокое сродство к IAP человека.
Фиг.43 показывает результаты апоптоз-индуцирующего действия in vitro в примере 6.11, иллюстрирующие, что димер scFv <HL5> и sc(Fv)2 индуцируют апоптоз клеток hIAP/L1210 и клеток CCRF-CEM зависимым от концентрации образом.
Фиг.44 показывает результаты количественного измерения М-белка, продуцируемого клеточной линией миеломы человека КРММ2, в сыворотке мыши с трансплантированными клетками миеломы человека. Эта фигура иллюстрирует, что димер scFv <HL5> и sc(Fv)2 в сильной степени ингибировали рост клеток КРММ2.
Фиг.45 показывает время выживания (дни) мышей после трансплантации опухоли, иллюстрируя, что время выживания группы мышей с введенным scFv <HL5> было значительно увеличено.
Фиг.46 показывает время выживания (дни) мышей после трансплантации опухоли, иллюстрируя, что время выживания группы мышей с введенным sc(Fv)2 было значительно увеличено.
Фиг.47 является схемой, показывающей способ конструирования ДНК-фрагмента, кодирующего реконструированный одноцепочечный Fv 12B5, содержащий линкерную последовательность, состоящую из 15 аминокислот, и его структуру.
Фиг.48 показывает результат очистки каждого одноцепочечного Fv 12B5 гель-фильтрацией, полученного в примере 7.5 (1), иллюстрирующий, что sc12B5 разделялся на два пика (фракции А и В).
Фиг.49 показывает результат анализа каждой фракции А и В электрофорезом в ДСН-ПААГ, проведенного в примере 7.5 (2).
Фиг.50 показывает результат анализа каждой фракции А и В способом хроматографии на колонке Superdex 200, проведенного в примере 7.5 (2), иллюстрирующий, что основной пик фракции А элюировался при кажущейся молекулярной массе около 44 кД, как показано в (а), и что основной пик фракции В элюировался при кажущейся молекулярной массе около 22 кД в (b).
Фиг.51 показывает результат измерения ТРО-подобной агонистической активности sc12B5 и антитела 12B5 (IgG, Fab), иллюстрирующий, что 12B5IgG и моновалентный одноцепочечный Fv (sc12B5) обнаруживали ТРО-подобную агонистическую активность, зависящую от концентрации.
Фиг.52 показывает результат измерения ТРО-подобной агонистической активности мономера и димера sc12B5, иллюстрирующий, что одноцепочечный Fv (димер sc12B5), имеющий бивалентный антигенсвязывающий сайт, имел агонистическую активность, приблизительно в 400 раз более высокую, чем моновалентный sc12B5, и что эта эффективность является эквивалентной или более высокой, чем эффективность ТРО человека.
Фиг.53 показывает результат очистки полученного одноцепочечного антитела sc12E10 гель-фильтрационной хроматографией с использованием колонки Superdex 200HR, иллюстрирующий, что sc12E10 разделялся на два пика (фракции А и В).
Фиг.54 показывает результат очистки полученного одноцепочечного антитела db12T10 гель-фильтрационной хроматографией с использованием колонки Superdex 200HR, иллюстрирующий, что db12E10 разделялся на два пика (фракции С и D).
Фиг.55 показывает электрофоретический анализ в ДСН-ПААГ фракций А и В (sc12E10) и фракций С и D (db12E10) в восстанавливающих и невосстанавливающих условиях.
Фиг.56 показывает результат анализа фракций А и В гель-фильтрационной хроматографией с использованием колонки Superdex 200HR, иллюстрирующий, что (1) основной пик фракции А элюировался с кажущейся молекулярной массой около 42 кДа, а (2) основной пик фракции В элюировался с кажущейся молекулярной массой около 20 кДа.
Фиг.57 показывает результат анализа фракций С и D гель-фильтрационной хроматографией с использованием колонки Superdex 200HR, иллюстрирующий, что (1) основной пик фракции С элюировался с кажущейся молекулярной массой около 69 кДа, а (2) главный пик фракции D элюировался с кажущейся молекулярной массой около 41 кДа.
Фиг.58 является графиком, показывающим агонистическую активность различных вариантов молекул антитела 12Е10 на MPL, иллюстрирующим, что одноцепочечные Fv (sc12E10, db12E10) обнаруживали ТРО-подобную агонистическую активность, тогда как 12Е10 IgG и 12Е10 Fab не обнаруживали.
Фиг.59 является графиком, показывающим агонистическую активность мономера и димера sc12E10 и димера и тримера db12E10 на MPL, иллюстрирующим, что димер sc12E10 и димер и тример db12E10 обнаруживали ТРО-подобную агонистическую активность, более высокую, чем активность ТРО.
Сведения, подтвержающие возможность осуществления изобретения
Данное изобретение будет иллюстрировано конкретно со ссылкой на следующие примеры, которые никоим образом не ограничивают объем данного изобретения.
Для иллюстрации способа получения модифицированных антител данного изобретения ниже приведены примеры получения одноцепочечных Fv. В этих примерах получения модифицированных антител использовали мышиные антитела против IAP человека, MABL-1 и MABL-2. Гибридомы MABL-1 и MABL-2, продуцирующие их, соответственно, депонировали в уполномоченном международном депозитарии микроорганизмов (National Institute of Bioscience and Technology, Agency of Industrial Science and Technology, Minister of International Trade and Industry (1-3 Higasi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan)) 11 сентября 1997 как PERM BP-6100 и FERM BP-6101.
Примеры
Пример 1 (Клонирование ДНК, кодирующих V-область мышиных моноклональных антител к IAP человека)
ДНК, кодирующие вариабельные районы мышиных моноклональных антител к IAP человека, MABL-1 и MABL-2, клонировали следующим образом.
1.1 Получение матричной РНК(мРНК)
мРНК гибридом MABL-1 и MABL-2 выделяли с использованием набора mRNA Purification Kit (Pharmacia Biotech).
1.2 Синтез двухцепочечной кДНК
Двухцепочечную кДНК синтезировали из приблизительно 1 мкг мРНК с использованием набора Marathon cDNA Amplification Kit (Clontech) и присоединяли к ней адаптер.
1.3 ПЦР-амплификация генов, кодирующих вариабельные районы антитела
ПЦР проводили с использованием термоциклера Thermal Cycler (Perkin Elmer).
(1) Амплификация гена, кодирующего V-область L-цепи MABL-1
Праймерами, используемыми для ПЦР-способа, являются адапторный праймер-1 (Clontech), представленный SEQ ID NO 1, который гибридизуется с частью последовательности адаптера, и МКС-праймер (Mouse Kappa Constant) (Bio/Technology, 9, 88-89, 1991), представленный SEQ ID NO 2, который гибридизуется с V-областью L-цепи типа каппа мыши.
50 мкл ПЦР-раствора содержат 5 мкл 10-кратного ПЦР-буфера II, 2 мМ MgCl2, 0,16 мМ dNTP (dATP, dGTP, dCTP и dTTP), 2,5 единиц ДНК-полимеразы AmpliTaq Gold (Perkin Elmer), 0,2 мкМ адапторный праймер SEQ ID NO 1, 0,2 мкМ МКС-праймер SEQ ID NO 2 и 0,1 мкг двухцепочечной кДНК, полученной из MABL-1. Этот раствор предварительно нагревали при начальной температуре 94°С в течение 9 минут и затем нагревали при 94°С в течение 1 минуты, при 60°С в течение 1 минуты и при 72°С в течение 1 минуты 20 секунд в указанной последовательности. Этот температурный цикл повторяли 35 раз и затем реакционную смесь дополнительно нагревали при 72°С в течение 10 минут.
(2) Амплификация кДНК, кодирующей У-область Н-цепи MABL-1
Адапторный праймер-1, представленный SEQ ID NO 1, и MHC-γ1-праймер (Mouse Heavy Constant) (Bio/Technology, 9, 88-89, 1991), представленный SEQ ID NO 3, использовали в качестве праймеров для ПЦР.
Амплификацию кДНК выполняли в соответствии со способом амплификации гена V-области L-цепи, который описан в Примере 1.3-(1), за исключением использования 0,2 мкМ MHC-γ1-праймера вместо 0,2 мкМ МКС-праймера.
(3) Амплификация кДНК, кодирующей V-область L-цепи MABL-2
Адапторный праймер-1, представленный SEQ ID NO 1, и МКС-праймер, представленный SEQ ID NO 2, использовали в качестве праймеров для ПЦР.
Амплификацию кДНК выполняли в соответствии со способом амплификации гена V-области L-цепи гена MABL-1, который описан в Примере 1.3-(1), за исключением использования 0,1 мкг двухцепочечной кДНК, полученной из MABL-2, вместо 0,1 мкг двухцепочечной кДНК из MABL-1.
(4) Амплификация кДНК, кодирующей V-область Н-цепи MABL-2
Адапторный праймер-1, представленный SEQ ID NO 1, и МНС-γ2а-праймер (Bio/Technology, 9, 88-89, 1991), представленный SEQ ID NO 4, использовали в качестве праймеров для ПЦР.
Амплификацию кДНК выполняли в соответствии со способом амплификации гена V-области L-цепи, который описан в Примере 1.3-(3), за исключением использования 0,2 мкМ МНС-γ2-праймера вместо 0,2 мкМ МКС-праймера.
1.4 Очистка ПЦР-продуктов
ДНК-фрагмент, амплифицированный при помощи ПЦР, как описано выше, очищали с использованием набора QIAquick PCR Purification Kit (Qiagen) и растворяли в 10 мМ Трис-HCl (рН 8,0), содержащем 1 мМ ЭДТА.
1.5 Лигирование и трансформация
Приблизительно 140 нг ДНК-фрагмента, содержащего ген, кодирующий мышиную V-область L-цепи типа каппа, полученный из MABL-1, как описано выше, лигировали с 50 нг вектора pGEM-T Easy (Promega) в реакционном буфере, содержащем 30 мМ Трис-HCl (рН 7,8), 10 мМ MgCl2, 10 мМ дитиотреитол, 1 мМ АТФ и 3 единицы ДНК-лигазы Т4 (Promega), при 15°С в течение 3 часов.
Затем 1 мкл этой реакционной смеси добавляли к 50 мкл компетентных клеток Е.coli DH5α (Toyobo Inc.). и клетки выдерживали на льду в течение 30 минут, инкубировали при 42°С в течение 1 минуты и опять выдерживали на льду в течение 2 минут. Добавляли 100 мкл среды SOC (Gibco BRL). Клетки Е.coli высевали на (Molecular Cloning: A Laboratory Manual, Sambrook et al., Cold Spring Harbor Press, 1989) агаризованную среду LB, содержащую 100 мкг/мл ампициллина (Sigma) и культивировали при 37°С в течение ночи для получения трансформантов Е.coli.
Трансформант культивировали в 3 мл среды LB, содержащей 50 мкг/мл ампициллина, при 37°С в течение ночи, и плазмидную ДНК выделяли из этой культуры с использованием набора QIAprep Spin Miniprep Kit (Qiagen).
Полученная плазмида, содержащая ген, кодирующий мышиную V-область L-цепи типа каппа, происходящий из гибридомы MABL-1, была обозначена pGEM-M1L.
В соответствии с описанным выше способом плазмиду, содержащую ген, кодирующий V-область Н-цепи мыши, происходящий из гибридомы MABL-1, получали с использованием очищенного ДНК-фрагмента и обозначали как pGEM-M1H.
Плазмиду, содержащую ген, кодирующий V-область L-цепи типа каппа мыши, происходящий из гибридомы MABL-2, получали с использованием очищенного ДНК-фрагмента и обозначали как pGEM-M2L.
Плазмиду, содержащую ген, кодирующий V-область Н-цепи мыши, происходящий из гибридомы MABL-2, получали с использованием очищенного ДНК-фрагмента и обозначали как pGEM-M2H.
Пример 2 (Секвенирование ДНК)
Нуклеотидную последовательность областей, кодирующих кДНК в вышеупомянутых плазмидах, определяли с использованием автоматического ДНК-секвенатора (Applied Biosystems) и набора для секвенирования ABI PRISM Dye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems) в соответствии с протоколом изготовителя.
Нуклеотидная последовательность гена, кодирующего V-область L-цепи мышиного антитела MABL-1, которая встроена в плазмиду pGEM-M1L, показана в SEQ ID NO 5.
Нуклеотидная последовательность гена, кодирующего V-область Н-цепи мышиного антитела MABL-1, которая встроена в плазмиду pGEM-M1H, показана в SEQ ID NO 6.
Нуклеотидная последовательность гена, кодирующего V-область L-цепи мышиного антитела MABL-2, которая встроена в плазмиду pGEM-M2L, показана в SEQ ID NO 7.
Нуклеотидная последовательность гена, кодирующего V-область Н-цепи мышиного антитела MABL-2, которая встроена в плазмиду pGEM-M2H, показана в SEQ ID NO 8.
Пример 3 (Определение CDR)
V-области L-цепи и Н-цепи обычно имеют сходное строение, и каждые четыре каркасные области в них связаны тремя гипервариабельными участками, т.е. определяющими комплементарность районами (CDR). Аминокислотная последовательность этой каркасной части является относительно консервативной, тогда как аминокислотная последовательность CDR имеет чрезвычайно высокую изменчивость (Kabat, E.A. et al., "Sequence of Protein of Immunological Interest", US Dept. Health and Human Services, 1983).
На основании этих фактов аминокислотные последовательности вариабельных районов мышиных моноклональных антител к IAP человека сравнивали с базой данных аминокислотных последовательностей антител, полученной Kabat et al., на предмет выявления гомологии. CDR-районы определяли на основе гомологии, как показано в таблице 1.
Пример 4 (Идентификация экспрессии клонированной кДНК. Получение химерного антитела MABL-1 и химерного антитела MABL-2)
4.1 Получение векторов, экспрессирующих химерное антитело MABL-1
кДНК-клоны, pGEM-M1L и pGEM-M1H, кодирующие V-области L-цепи и Н-цепи мышиного антитела MABL-1, соответственно, модифицировали при помощи ПЦР-способа и вводили в экспрессионный вектор HEF (WO 92/19759) для получения векторов, экспрессирующих химерное антитело MABL-1.
Прямой праймер MLS (SEQ ID NO 9) для V-области L-цепи и прямой праймер MHS (SEQ ID NO 10) для V-области Н-цепи конструировали так, чтобы они гибридизовались с ДНК, кодирующей начало лидерной последовательности каждой V-области, и содержали консенсусную последовательность Козака (J.Mol. Biol., 196, 947-950, 1987) и сайт рестриктазы HindIII. Обратный праймер MLAS (SEQ ID NO 11) для V-области L-цепи и обратный праймер MHAS (SEQ ID NO 12) для V-области Н-цепи конструировали так, чтобы они гибридизовались с ДНК, кодирующей конец J-сегмента, и содержали последовательность донорного сайта сплайсинга и сайт рестриктазы BamHI.
100 мкл ПЦР-раствора, содержащего 10 мкл 10-кратного ПЦР-буфера II, 2 мМ MgCl2, 0,16 мМ dNTP (dATP, dGTP, dCTP и dTTP), 5 единиц ДНК-полимеразы AmpliTaq Gold, 0,4 мкМ каждого из праймеров и 8 нг ДНК-матрицы (pGEM-M1L или pGEM-M1H), предварительно нагревали при начальной температуре 94°С в течение 9 минут и затем нагревали при 94°С в течение 1 минуты, при 60°С в течение 1 минуты и при 72°С в течение 1 минуты 20 секунд в указанной последовательности. Этот температурный цикл повторяли 35 раз и затем реакционную смесь дополнительно нагревали при 72°С в течение 10 минут.
ПЦР-продукт очищали с использованием набора QIAquick PCR Purification Kit (Qiagen) и затем расщепляли HindIII и BamHI. Продукт из V-области L-цепи клонировали в экспрессионный вектор HEF, HEF-κ, а продукт из V-области Н-цепи клонировали в экспрессионный вектор HEF, HEF-γ. После секвенирования ДНК, плазмиды, содержащие ДНК-фрагмент с правильной ДНК-последовательностью, обозначали как HEF-M1L и HEF-M1H, соответственно.
4.2 Получение векторов, экспрессирующих химерные антитела MABL-2
Модификацию и клонирование кДНК выполняли таким же образом, как описано в примере 4.1, за исключением того, что использовали pGEM-M2L и pGEM-M2H в качестве ДНК-матрицы вместо pGEM-M1L и pGEM-M1H. После секвенирования ДНК плазмиды, содержащие ДНК-фрагменты с правильными ДНК-последовательностями, обозначали как HEF-M2L и HEF-M2H, соответственно.
4.3 Трансфекция в клетки COS7
Вышеупомянутые экспрессионные вектора тестировали в клетках COS7 для наблюдения временной экспрессии химерных антител MABL-1 и MABL-2.
(1) Трансфекция генами химерного антитела MABL-1
Клетки COS7 котрансформировали векторами HEF-M1L и HEF-M1H электропорацией с использованием устройства Gene Pulser (BioRad). Каждую ДНК (10 мкг) и 0,8 мл PBS, содержащего 1×107 клеток/мл, добавляли в кювету. Смесь обрабатывали импульсом при 1,5 кВ, при электрической емкости 25 мкФ.
После "восстановления" в течение 10 минут при комнатной температуре электропорированные клетки переносили в культуральную среду Игла в модификации Дульбекко (DMEM, Gibco BRL), содержащую 10% свободной от γ-глобулина фетальной телячьей сыворотки. После культивирования в течение 72 часов супернатант собирали, центрифугировали для удаления клеточных фрагментов и отделяли.
(2) Трансфекция генами, кодирующими химерное антитело MABL-2
Котрансфекцию клеток COS7 генами, кодирующими химерное антитело MABL-2, проводили таким же образом, как описано в примере 4.3-(1), за исключением того, что использовали вектора HEF-M2L и HEF-M2H вместо векторов HEF-M1L и HEF-M1H. Супернатант получали таким же образом.
4.4 Проточная цитометрия
Проточную цитометрию проводили с использованием вышеупомянутого культурального супернатанта клеток COS7 для измерения связывания с антигеном. Культуральный супернатант клеток COS7, экспрессирующих химерное антитело MABL-1, или клеток COS7, экспрессирующих химерное антитело MABL-2, или IgG-антитело человека (Sigma) в качестве контроля добавляли к 4×105 клеток линии клеток мышиного лейкоза L1210, экспрессирующей IAP человека, и инкубировали на льду. После промывки к ним добавляли FITC-меченное антитело против IgG человека (Cappel). После инкубации и промывки измеряли интенсивность флуоресценции с использованием прибора FACScan (Becton Dickinson).
Поскольку химерные антитела MABL-1 и MABL-2 специфически связывались с клетками L1210, экспрессирующими IAP человека, было подтверждено, что эти химерные антитела имели правильные структуры V-областей мышиных моноклональных антител MABL-1 и MABL-2, соответственно (фигуры 1-3).
Пример 5 (Получение реконструированного одноцепочечного Fv (scFv) антитела MABL-1 и антитела MABL-2)
5.1 Получение реконструированного одноцепочечного Fv антитела MABL-1
Реконструированный одноцепочечный Fv антитела MABL-1 получали следующим образом. V-область H-цепи и V-область L-цепи антитела MABL-1 и линкер соответственно амплифицировали при помощи ПЦР-способа и соединяли для получения реконструированного одноцепочечного Fv антитела MABL-1. Способ получения иллюстрируется на фиг.4. Для получения одноцепочечного Fv антитела MABL-1 использовали шесть праймеров (A-F). Праймеры А, С и Е содержат смысловую последовательность, а праймеры В, D и F содержат антисмысловую последовательность.
Прямой праймер VHS для V-области Н-цепи (Праймер A, SEQ ID NO 13) конструировали так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области Н-цепи, и содержал сайт узнавания рестриктазы NcoI. Обратный праймер VHAS для V-области Н-цепи (Праймер В, SEQ ID NO 14) конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области Н-цепи, и перекрывался с линкером.
Прямой праймер LS для линкера (Праймер С, SEQ ID NO 15) конструировали так, чтобы он гибридизовался с ДНК, кодирующей N-конец линкера, и перекрывался с ДНК, кодирующей С-конец V-области Н-цепи. Обратный праймер LAS для линкера (Праймер D, SEQ ID NO 16) конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец линкера, и перекрывался с ДНК, кодирующей N-конец V-области L-цепи.
Прямой праймер VLS для V-области L-цепи (Праймер Е, SEQ ID NO 17) конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец линкера, и перекрывался с ДНК, кодирующей N-конец V-области L-цепи. Обратный праймер VLAS-FLAG для V-области L-цепи (Праймер F, SEQ ID NO 18) конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал последовательность, кодирующую пептид FLAG (Hopp N.P. et al., Bio/Technology, 6, 1204-1210, 1988), два стоп-кодона и сайт узнавания рестриктазы EcoRI.
На первой стадии ПЦР проводили три реакции: А-В, C-D и E-F, и полученные ПЦР-продукты очищали. Три ПЦР-продукта, полученные из этой первой стадии ПЦР, состыковывали на основе их комплементарности. Затем добавляли праймеры А и F и амплифицировали полноразмерную ДНК, кодирующую реконструированный одноцепочечный Fv антитела MABL-1 (вторая ПЦР). В первой ПЦР в качестве матрицы использовали плазмиду pGEM-M1H, кодирующую V-область Н-цепи антитела MABL-1 (см. пример 2), плазмиду pSC-DPI, которая содержит ДНК-последовательность, кодирующую район линкера, содержащий: Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser (SEQ ID NO 19) (Huston, J.S., et al., Proc. Natl. Acad. Sci. USA, 85, 5879-5883, 1988), и плазмиду pGEM-MIL, кодирующую V-область L-цепи антитела MABL-1 (см. пример 2), соответственно.
50 мкл раствора для первой стадии ПЦР содержат 5 мкл 10-кратного ПЦР-буфера II, 2 мМ MgCl2, 0,16 мМ dNTP, 2,5 единицы ДНК-полимеразы AmpliTaq Gold (Perkin Elmer), 0,4 мкМ каждого из праймеров и 5 нг каждой из ДНК-матриц. ПЦР-раствор предварительно нагревали при начальной температуре 94°С в течение 9 минут и затем нагревали при 94°С в течение 1 минуты, при 65°С в течение 1 минуты и при 72°С в течение 1 минуты 20 секунд в указанной последовательности. Этот температурный цикл повторяли 35 раз и затем реакционную смесь дополнительно нагревали при 72°С в течение 7 минут.
ПЦР-продукты А-В (371 п.о.), C-D (63 п.о.) и E-F (384 п.о.) очищали с использованием набора для очистки QIAquick PCR Purification Kit (Qiagen) и проводили сборку во второй ПЦР. Во второй ПЦР 98 мкл ПЦР-раствора, содержащего 120 нг продукта первой ПЦР А-В, 20 нг ПЦР-продукта C-D и 120 нг ПЦР-продукта E-F, 10 мкл 10-кратного ПЦР-буфера II, 2 мМ MgCl2, 0,16 мМ dNTP, 5 единиц ДНК-полимеразы AmpliTaq Gold (Perkin Elmer) предварительно нагревали при начальной температуре 94°С в течение 8 минут и затем нагревали при 94°С в течение 2 минут, при 65°С в течение 2 минут и при 72°С в течение 2 минут в указанной последовательности. Этот температурный цикл повторяли дважды и затем в реакцию добавляли 0,4 мкМ каждого из праймеров А и F, соответственно. Эту смесь предварительно нагревали при начальной температуре 94°С в течение 1 минуты и затем нагревали при 94°С в течение 1 минуты, при 65°С в течение 1 минуты и при 72°С в течение 1 минуты 20 секунд в указанной последовательности. Этот температурный цикл повторяли 35 раз и затем реакционную смесь дополнительно нагревали при 72°С в течение 7 минут.
ДНК-фрагмент 843 п.о., полученный во второй ПЦР, очищали и расщепляли NcoI и EcoRI. Полученный ДНК-фрагмент клонировали в вектор pSCFVT7. Экспрессионный вектор pSCFVT7 содержит сигнальную последовательность pelB, пригодную для периплазматической экспрессионной системы Е.coli (Lei, S.P., et al., J.Bacteriology, 169, 4379-4383, 1987). После секвенирования ДНК плазмиду, содержащую ДНК-фрагмент, кодирующий правильную аминокислотную последовательность реконструированного одноцепочечного Fv антитела MABL-1, обозначали как "pscM1" (см. фиг.5). Нуклеотидная последовательность и аминокислотная последовательность этого реконструированного одноцепочечного Fv антитела MABL-1, содержащегося в плазмиде pscM1, приведены в SEQ ID NO 20.
Вектор pscM1 модифицировали при помощи ПЦР-способа для получения вектора, экспрессирующего реконструированный одноцепочечный Fv антитела MABL-1 в клетках млекопитающих. Полученный ДНК-фрагмент вводили в экспрессионный вектор pCHO1. Этот экспрессионный вектор, pCHO1, получали расщеплением DHFR-ΔE-rvH-PM1-f (WO 92/19759) EcoRI и SmaI для исключения гена антитела и присоединением к нему EcoRI-NotI-BamHI-адаптора (Takara Shuzo).
В качестве прямого праймера для ПЦР праймер Sal-VHS, показанный в SEQ ID NO 21, конструировали так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области Н-цепи, и содержал сайт узнавания рестриктазы SalI. В качестве обратного праймера для ПЦР праймер FRHlanti, показанный в SEQ ID NO 22, конструировали так, чтобы он гибридизовался с ДНК, кодирующей конец первой каркасной последовательности.
100 мкл ПЦР-раствора, содержащего 10 мкл 10-кратного ПЦР-буфера II, 2 мМ MgCl2, 0,16 мМ dNTP, 5 единиц ДНК-полимеразы AmpliTaq Gold, 0,4 мкМ каждого праймера и 8 нг ДНК-матрицы (pscMl), предварительно нагревали при начальной температуре 95°С в течение 9 минут и затем нагревали при 95°С в течение 1 минуты, при 60°С в течение 1 минуты и при 72°С в течение 1 минуты 20 секунд в указанной последовательности. Этот температурный цикл повторяли 35 раз и затем реакционную смесь дополнительно нагревали при 72°С в течение 7 минут.
ПЦР-продукт очищали с использованием набора QIAquick PCR Purification Kit (Qiagen) и затем расщепляли SaiI и MboII для получения ДНК-фрагмента, кодирующего N-конец реконструированного одноцепочечного Fv антитела MABL-1. Вектор pscMl расщепляли MboII и EcoRI с получением ДНК-фрагмента, кодирующего С-конец реконструированного одноцепочечного Fv антитела MABL-1. SalI-MboII-фрагмент ДНК и MboII-EcoRI-фрагмент ДНК клонировали в вектор pCHO1-Igs. После секвенирования ДНК плазмиду, содержащую желаемую ДНК-последовательность, обозначали как "рСНОМ1" (см. фиг.6). Экспрессионный вектор pCHO1-Igs содержит сигнальную последовательность мышиного IgG1, пригодную для системы экспрессии-секреции в клетках млекопитающих (Nature, 322, 323-327, 1988). Нуклеотидная последовательность и аминокислотная последовательность реконструированного одноцепочечного Fv антитела MABL-1, содержащегося в плазмиде pCHOMI, показаны в SEQ ID NO 23.
5.2 Получение реконструированного одноцепочечного Fv антитела MABL-2
Реконструированный одноцепочечный Fv антитела MABL-2 получали в соответствии с вышеописанным примером 5.1. На первой стадии ПЦР использовали плазмиду pGEM-M2H, кодирующую V-область Н-цепи MABL-2 (см. пример 2), вместо pGEM-M1H, и плазмиду pGEM-M2L, кодирующую V-область L-цепи MABL-2 (см. пример 2), вместо pGEM-M1L, для получения плазмиды pscM2, которая содержит ДНК-фрагмент, кодирующий желаемую аминокислотную последовательность одноцепочечного Fv антитела MABL-2. Нуклеотидная последовательность и аминокислотная последовательность реконструированного одноцепочечного Fv антитела MABL-2, содержащегося в плазмиде pscM21, показаны в SEQ ID NO 24.
Вектор pscM2 модифицировали при помощи ПЦР-способа для получения вектора, рСНОМ2, для экспрессии в клетках млекопитающих, который содержит ДНК-фрагмент, кодирующий правильную аминокислотную последовательность реконструированного одноцепочечного Fv антитела MABL-2. Нуклеотидная последовательность и аминокислотная последовательность реконструированного одноцепочечного Fv антитела MABL-2, содержащегося в плазмиде рСНОМ2, показаны в SEQ ID NO 25.
5.3 Трансфекция в клетки COS7
Вектор рСНОМ2 тестировали в клетках COS7 для наблюдения временной экспрессии реконструированного одноцепочечного Fv антитела MABL-2.
Клетки COS7 трансформировали вектором рСНОМ2 электропорацией с использованием устройства Gene Pulser (BioRad). ДНК (10 мкг) и 0,8 мл PBS, содержащего 1×107 клеток/мл, добавляли в кювету. Смесь обрабатывали импульсом при 1,5 кВ при электрической емкости 25 мкФ.
После "восстановления" в течение 10 минут при комнатной температуре электропорированные клетки переносили в культуральную среду IMDM (Gibco BRL), содержащую 10% фетальную телячью сыворотку. После культивирования в течение 72 часов супернатант собирали, центрифугировали для удаления клеточных фрагментов и отбирали.
5.4 Детектирование реконструированного одноцепочечного Fv антитела MABL-2 в культуральном супернатанте клеток COS7
Присутствие одноцепочечного Fv антитела MABL-2 в культуральном супернатанте клеток COS7, которые были трансфицированы вектором рСНОМ2, подтверждали способом Вестерн-блоттинга.
Культуральный супернатант клеток COS7, трансфицированных вектором рСНОМ2, и культуральный супернатант клеток COS7, трансфицированных pCHO1, в качестве контроля подвергали ДСН-электрофорезу и переносили на мембрану REINFORCED NC (Schleicher & Schuell). Мембрану блокировали 5% обезжиренным молоком (Morinaga Nyu-guo), отмывали 0,05% Твин 20-PBS и смешивали с анти-FLAG-антителом (Sigma). Мембрану инкубировали при комнатной температуре, промывали и добавляли конъюгированное со щелочной фосфатазой антитело против мышиного IgG (Zymed). После инкубирования и промывания при комнатной температуре добавляли раствор субстрата (Kirkegaard Perry Laboratories) для развития окраски (фиг.7).
Белок, дававший специфическую реакцию на FLAG-пептид, обнаруживали только в культуральном супернатанте клеток COS7 с введенным вектором рСНОМ2, и, следовательно, было подтверждено, что реконструированный одноцепочечный Fv антитела MABL-2 секретировался в этот культуральный супернатант.
5.5 Проточная цитометрия
Проточную цитометрию проводили с использованием вышеупомянутого культурального супернатанта клеток COS7 для измерения связывания с антигеном. Культуральный супернатант клеток COS7, экспрессирующих реконструированный одноцепочечный Fv антитела MABL-2, или культуральный супернатант клеток COS7, трансформированных вектором pCHO1 в качестве контроля, добавляли к 2×105 клеток клеточной линии мышиного лейкоза L1210, экспрессирующей интегрин-ассоциированный белок (IAP) человека, или клеточной линии L1210, трансформированной pCOSI, в качестве контроля. После инкубирования на льду и промывания добавляли мышиное анти-FLAG-антитело (Sigma). Затем клетки инкубировали и промывали. Затем к ним добавляли FITC-меченное антитело против мышиного IgG (Becton Dickinson) и клетки опять инкубировали и промывали. Затем измеряли интенсивность флуоресценции с использованием прибора FACScan (Becton Dickinson).
Поскольку одноцепочечный Fv антитела MABL-2- специфически связывался с клетками L1210, экспрессирующими IAP человека, было подтверждено, что реконструированный одноцепочечный Fv антитела MABL-2 обладает сродством к интегрин-ассоциированному белку (IAP) человека (см. фиг.8-11).
5.6 Конкурентный анализ ELISA
Связывающую активность реконструированного одноцепочечного Fv антитела MABL-2 измеряли на основе ингибирующей активности против связывания мышиных моноклональных антител с антигеном.
Анти-FLAG-антитело, доведенное до концентрации 1 мкг/мл, добавляли в каждую лунку 96-луночного планшета и инкубировали при 37°С в течение 2 часов. После промывания проводили блокирование 1% БСА-PBS. После инкубирования и промывания при комнатной температуре культуральный супернатант клеток COS7, в который был введен ген антигена IAP человека секреторного типа (SEQ ID NO 26), разбавляли PBS до двукратного объема и добавляли в каждую лунку. После инкубирования и промывания при комнатной температуре в каждую лунку добавляли смесь 50 мкл биотинилированного антитела MABL-2, доведенного до концентрации 100 нг/мл, и 50 мкл последовательно разведенного супернатанта клеток COS7, экспрессирующих реконструированный одноцепочечный Fv антитела MABL-2. После инкубирования и промывания при комнатной температуре в каждую лунку добавляли конъюгированный со щелочной фосфатазой стрептавидин (Zymed). После инкубирования и промывания при комнатной температуре добавляли раствор субстрата (Sigma) и измеряли оптическую плотность реакционной смеси в каждой лунке при 405 нм.
Результаты показали, что реконструированный одноцепочечный Fv антитела MABL-2 (MABL2-scFv) явно ингибировал зависимым от концентрации образом связывание мышиного антитела MABL-2 с антигеном IAP человека по сравнению с культуральным супернатантом клеток COS7 с введенной рСНО1 в качестве контроля (фиг.12). Таким образом, предполагается, что реконструированный одноцепочечный Fv антитела MABL-2 имеет правильную структуру каждой из V-областей мышиного моноклонального антитела MABL-2.
5.7 Индуцирующее апоптоз действие in vitro
Индуцирующее апоптоз действие реконструированного одноцепочечного Fv антитела MABL-2 исследовали методом окрашивания Аннексином-V (Boehringer Mannheim) с использованием клеток L1210, трансфицированных геном IAP человека, клеток L1210, трансфицированных вектором pCOS2, в качестве контроля, и клеток CCRF-СЕМ.
К 1×105 клеток каждой из указанных выше линий клеток добавляли культуральный супернатант клеток COS7, экспрессирующих реконструированный одноцепочечный Fv антитела MABL-2, или культуральный супернатант клеток COS7, трансфицированных вектором pCHO1, в качестве контроля в конечной концентрации 50% и эти смеси культивировали в течение 24 часов. Затем проводили окрашивание Аннексином-V и измеряли интенсивность флуоресценции при помощи прибора FACScan (Becton Dickinson).
Результаты окрашивания Аннексином-V показаны на фиг.13-18, соответственно. Точки в левом нижнем квадранте представляют живые клетки, точки в правом нижнем квадранте представляют клетки на ранней стадии апоптоза и точки в правом верхнем квадранте представляют клетки на поздней стадии апоптоза. Эти результаты показывают, что реконструированный одноцепочечный Fv антитела MABL-2 (MABL2-scFv) в значительной мере индуцировал гибель клеток L1210, специфических в отношении антигена IAP человека (фигуры 13-16), и что реконструированный одноцепочечный Fv индуцировал также значительную гибель клеток CCRF-CEM в сравнении с контролем (фигуры 17-18).
5.8 Экспрессия происходящего из MABL-2 одноцепочечного Fv в клетках СНО
Клетки СНО трансфицировали вектором рСНОМ2 для получения линии клеток СНО, которая константно экспрессирует одноцепочечный Fv (полипептид), происходящий из антитела MABL-2.
Клетки СНО трансформировали вектором рСНОМ2 электропорацией с использованием устройства Gene Pulser (BioRad). Смесь ДНК (10 мкг) и 0,7 мл PBS, содержащего клетки СНО (1×107 клеток/мл), добавляли в кювету. Смесь обрабатывали импульсом при 1,5 кВ при электрической емкости 25 мкФ. После "восстановления" в течение 10 минут при комнатной температуре электропорированные клетки переносили в свободную от нуклеиновых кислот культуральную среду α-МЕМ (Gibco BRL), содержащую 10% фетальную телячью сыворотку, и культивировали. Экспрессию желаемого белка в полученных клонах подтверждали электрофорезом в ДСН-ПААГ, и клон с высоким уровнем экспрессии отбирали в качестве клеточной линии, продуцирующей одноцепочечный Fv, происходящий из антитела MABL-2. Эту клеточную линию культивировали в бессывороточной среде CHO-S-SEM II (Gibco BRL), содержащей 10 нМ метотрексат (Sigma). Затем культуральный супернатант собирали, центрифугировали для удаления клеточных фрагментов и отбирали.
5.9 Очистка происходящего из MABL-2 одноцепочечного Fv, продуцируемого в клетках СНО
Культуральный супернатант клеточной линии СНО, экспрессирующей одноцепочечный Fv, полученный в примере 5.8, концентрировали в двадцать раз с использованием кассеты для искусственного диализа (PAN130SF, ASAHI MEDICALS). Концентрированный раствор хранили при -20°С и оттаивали перед очисткой.
Очистку одноцепочечного Fv из культурального супернатанта клеток СНО выполняли с использованием трех типов хроматографии, т.е. Blue-Sepharose, гидроксиапатита и гель-фильтрации.
(1) Колоночная хроматография на Blue-Sepharose
Концентрированный супернатант разбавляли в десять раз 20 мМ ацетатным буфером (рН 6,0) и центрифугировали для удаления нерастворимых веществ (10000 (об/мин, 30 минут). Этот супернатант наносили на колонку Blue-Sepharose (20 мл), уравновешенную тем же самым буфером. После промывания колонки тем же самым буфером белки, адсорбированные на колонке, элюировали ступенчатым градиентом NaCl в том же самом буфере, 0,1, 0,2, 0,3, 0,5 и до 1,0 М. Сошедшую в свободном объеме фракцию и каждую из элюированных фракций анализировали электрофорезом в ДСН-ПААГ. Фракции, в которых было обнаружено присутствие одноцепочечного Fv (эти фракции элюировались при 0,1-0,3 М NaCl), объединяли и концентрировали приблизительно в 20 раз с использованием CentriPrep 10 (Amicon).
(2) Гидроксиапатит
Концентрированный раствор, полученный в (1), разбавляли в 10 раз 10 мМ фосфатным буфером (рН 7,0) и наносили на гидроксиапатитную колонку (20 мл, BioRad). Колонку промывали 60 мл 10 мМ фосфатного буфера (рН 7,0). Затем белки, адсорбированные на колонке, элюировали линейным градиентом натрий-фосфатного буфера до 200 мМ (см. фиг.19). Анализ каждой фракции при помощи электрофореза в ДСН-ПААГ подтвердил присутствие одноцепочечного Fv во фракции А и фракции В.
(3) Гель-фильтрация
Каждую из фракций А и В, полученных в (2), по отдельности концентрировали с использованием CentriPrep-10 и наносили на колонку TSKgel G3000SWG (21,5×600 мм), уравновешенную 20 мМ ацетатным буфером (рН 6,0), содержащим 0,15 М NaCl. Хроматограммы показаны на фиг.20. Анализ этих фракций электрофорезом в ДСН-ПААГ подтвердил, что оба мажорных пика (AI и BI) являются желаемым одноцепочечным Fv. В этом гель-фильтрационном анализе фракция А элюировалась с кажущейся молекулярной массой 36 кДа, а фракция В элюировалась с кажущейся молекулярной массой 76 кДа. Очищенные одноцепочечные Fv (AI, BI) анализировали с использованием 15% ДСН-полиакриламидного геля. Образцы обрабатывали в отсутствие или в присутствии восстановителя, и электрофорез проводили в соответствии со способом Лэммли. Затем белок окрашивали Кумасси бриллиантовым синим. Как показано на фиг.21, как AI, так и BI давали единственную полосу с кажущейся молекулярной массой 35 кДа, независимо от отсутствия или присутствия восстановителя. Из вышеописанного сделали вывод, что AI является мономером одноцепочечного Fv, a BI является нековалентно связанным димером одноцепочечного Fv. Гель-фильтрационный анализ фракций AI и BI с колонкой TSKgel G3000SWG (7,5×60 мм) показал, что пик мономера детектируется только во фракции AI, а пик димера детектируется только во фракции BI (фиг.22). Фракция димера (фракция BI) составляла 4 процента общего количества одноцепочечных Fv. Более чем 90% этого димера во фракции димера стабильно сохранялись в течение более чем месяца при 4°С.
5.10 Конструирование вектора, экспрессирующего одноцепочечный Fv, происходящий из антитела MABL-2, в клетке Е.coli
Вектор pscM2 модифицировали ПЦР-способом для получения вектора, эффективно экспрессирующего одноцепочечный Fv антитела MABL-2 в клетках Е.coli. Полученный ДНК-фрагмент встраивали в экспрессионный вектор pSCFVT7.
В качестве прямого праймера для ПЦР праймер Nde-VHSm02, показанный в SEQ ID NO 27, конструировали так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области Н-цепи, и содержал стартовый кодон и сайт узнавания рестриктазы NdeI. В качестве обратного праймера для ПЦР, праймер VLAS, показанный в SEQ ID NO 28, конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал два стоп-кодона и сайт узнавания рестриктазы EcoRI. Прямой праймер Nde-VHSm02 содержит пять точковых мутаций в части, гибридизующейся с ДНК, кодирующей N-конец V-области Н-цепи, для эффективной экспрессии в Е.coli.
100 мкл ПЦР-раствора, содержащего 10 мкл 10-кратного ПЦР-буфера №1,1 мМ MgCl2, 0,2 мМ dNTP, 5 единиц ДНК-полимеразы KOD (все от Toyobo), 1 мкМ каждого праймера и 100 нг ДНК-матрицы (pscM2), нагревали при 98°С в течение 15 секунд, при 65°С в течение 2 секунд и при 72°С в течение 30 секунд в указанной последовательности. Этот температурный цикл повторяли 25 раз.
ПЦР-продукт очищали с использованием набора QIAquick PCR Purification Kit (Qiagen) и затем расщепляли NdeI и EcoRI и затем полученный ДНК-фрагмент клонировали в вектор pSCFVT7, из которого была удалена сигнальная последовательность pelB расщеплением NdeI и EcoRI. После секвенирования ДНК полученная плазмида, содержащая ДНК-фрагмент с желаемой ДНК-последовательностью, обозначали как "pscM2DEm02" (см. фиг.23). Нуклеотидная последовательность и аминокислотная последовательность одноцепочечного Fv, происходящего из антитела MABL-2, содержащегося в плазмиде pscM2DEm02, показаны в SEQ ID NO 29.
5.11 Экспрессия одноцепочечного Fv, происходящего из антитела MABL-2, в клетках Е.coli
Е.coli BL21(DE3)pLysS (Stratagene) трансформировали вектором pscM2DEm02 с получением штамма Е.coli, экспрессирующего одноцепочечный Fv, происходящий из антитела MABL-2. Полученные клоны анализировали на экспрессию желаемого белка методом электрофореза в ДСН-ПААГ, и клон с высоким уровнем экспрессии отбирали в качестве штамма, продуцирующего одноцепочечный Fv, происходящий из антитела MABL-2.
5.12 Очистка одноцепочечного Fv, происходящего из антитела MABL-2, продуцируемого в Е.coli
Отдельную колонию Е.coli, полученную трансформацией, культивировали в 3 мл среды LB при 28°С в течение 7 часов и затем в 70 мл среды LB при 28°С в течение ночи. Эту предварительную культуру переносили в 7 л среды LB и культивировали при 28°С при перемешивании при 300 об/мин с использованием Jar-ферментера. Когда поглощение среды достигало O.D. = 1,5, бактерии индуцировали 1 мМ IPTG и затем культивировали в течение 3 часов.
Культуральную среду центрифугировали (10000×g, 10 минут), и собирали осажденные бактерии. К этим бактериям добавляли 50 мМ Трис-HCl-буфер (рН 8,0), содержащий ЭДТА, 0,1 М NaCl и 1% Тритон Х-100 и бактерии разрушали обработкой ультразвуком (мощность: 4, рабочий цикл: 70%, 1 минута ×10 раз). Суспензию разрушенных бактерий центрифугировали (12000×g, 10 минут) для осаждения телец включения. Выделенные тельца включения смешивали с 50 мМ Трис-HCl-буфером (рН 8,0), содержащим 5 мМ ЭДТА, 0,1 М NaCl и 4% Тритон Х-100, и вновь обрабатывали ультразвуком (мощность: 4, рабочий цикл: 50%, 30 секунд × 2 раза) и центрифугировали (12000×g, 10 минут) для выделения желаемого белка в виде осадка и для удаления примесных белков, находящихся в супернатанте.
Тельца включения, содержащие желаемый белок, лизировали в 50 мМ Трис-HCl-буфере (рН 8,0), содержащем 6 М мочевину, 5 мМ ЭДТА, 0,1 М NaCl, и наносили на гель-фильтрационную колонку Sephacryl S-300 (5×90 см, Amersham Pharmacia), уравновешенную 50 мМ Трис-HCl-буфером (рН 8,0), содержащим 4 М мочевину, 5 мМ ЭДТА, 0,1 М NaCl и 10 мМ меркаптоэтанол, при скорости подачи 5 мл/мин для удаления ассоциированных одноцепочечных Fv с высокой молекулярной массой. Полученные фракции анализировали электрофорезом в ДСН-ПААГ и фракции с высокой чистотой данного белка разбавляли буфером, используемым при гель-фильтрации, до O.D.280=0,25. Затем эти фракции диализовали три раза против 50 мМ Трис-HCl-буфера (рН 8,0), содержащего 5 мМ ЭДТА, 0,1 М NaCl, 0,5 М Arg, 2 мМ глутатион в восстановленной форме и 0,2 мМ глутатион в окисленной форме, для того, чтобы произошла повторная укладка белка. Далее эту фракцию диализовали три раза против 20 мМ ацетатного буфера (рН 6,0), содержащего 0,15 М NaCl, для смены буфера.
Диализованный продукт наносили на гель-фильтрационную колонку Superdex 200 pg (2,6×60 см, Amersham Pharmacia), уравновешенную 20 мМ ацетатным буфером (рН 6,0), содержащим 0,15 NaCl, для удаления небольшого количества высокомолекулярного белка, который межмолекулярно сшивался связями S-S. Как показано на фиг.24, два пика, основной и минорный пики, элюировались после широких пиков, которые предположительно относятся к агрегату с высокой молекулярной массой. Анализ при помощи электрофореза в ДСН-ПААГ (см. фиг.21) и положения элюции этих двух пиков при гель-фильтрационном анализе предполагают, что основной пик соответствует мономеру одноцепочечного Fv, а минорный пик соответствует нековалентно связанному димеру одноцепочечного Fv. Нековалентно связанный димер составляет 4 процента общего количества одноцепочечных Fv.
5.13 Апопотоз-индуцирующая активность in vitro одноцепочечного Fv, происходящего из антитела MABL-2
Апоптоз-индуцирующую активность одноцепочечного Fv антитела MABL-2 (MABL2-scFv), продуцируемого клетками СНО и Е.coli, исследовали в соответствии с двумя протоколами методом окрашивания Аннексином-V (Boehringer Mannheim) с использованием клеток L1210 (hIAP/L1210), в которые был введен ген IAP человека.
В первом протоколе образцы антитела в конечной концентрации 3 мкг/мл добавляли к 5×104 клеткам клеточной линии hIAP/L1210 и культивировали в течение 24 часов. Анализировали образцы антитела, т.е. мономер и димер одноцепочечного Fv MABL-2 из клеток СНО, полученные в 5.9, мономер и димер одноцепочечного Fv MABL-2 из Е.coli, полученные в примере 5.12, и мышиное IgG-антитело в качестве контроля. После культивирования проводили окрашивание Аннексином-V и измеряли интенсивность флуоресценции с использованием прибора FACScan (Becton Dickinson).
Во втором протоколе образцы антитела в конечной концентрации 3 мкг/мл добавляли к 5×104 клеткам клеточной линии hIAP/L1210, культивировали в течение 2 часов и смешивали с анти-FLAG-антителом (Sigma) в конечной концентрации 15 мкг/мл и дополнительно культивировали в течение 22 часов. Анализировали образцы антитела, представленного мономером одноцепочечного Fv MABL-2 из клеток СНО, полученным в 5.9, и мышиное IgG антитело в качестве контроля. После культивирования проводили окрашивание Аннексином-V и измеряли интенсивность флуоресценции с использованием прибора FACScan.
Результаты анализа методом окрашивания на Аннексин-V показаны на фигурах 25-31. Эти результаты показывают, что димеры одноцепочечного полипептида Fv MABL-2, продуцируемые в клетках СНО и Е.coli, в сильной степени индуцировали гибель клеток (фигуры 26, 27) по сравнению с контролем (фиг.25), в то время как апоптоз-индуцирующее действие не наблюдали в случае мономеров одноцепочечного полипептида Fv MABL-2, продуцируемых в клетках СНО и Е.coli (фигуры 28, 29). Когда совместно использовали анти-FLAG-антитело, мономер одноцепочечного полипептида Fv, происходящего из антитела MABL-2, продуцируемый в клетках СНО, в сильной степени индуцировал гибель клеток (фиг.31) в сравнении с контролем (фиг.30).
5.14 Противоопухолевое действие мономера и димера полипептида scFv/CHO на мышиной модели миеломы человека
(1) Количественное определени IgG человека в мышиной сыворотке
Измерение IgG человека (М-белка), продуцируемого миеломными клетками человека и содержащегося в мышиной сыворотке, проводили при помощи ELISA следующим образом. 100 мкл козьих антител против IgG человека (Biosource, Lot#902), разбавленных до 1 мкг/мл 0,1% бикарбонатным буфером (рН 9,6), добавляли в каждую лунку 96-луночного планшета (Nunc) и инкубировали при 4°С в течение ночи, так что эти антитела иммобилизовались. После блокирования в каждую лунку добавляли 100 мкл ступенчато разведенной мышиной сыворотки или IgG человека (Cappel, Lot#00915) в качестве стандарта и инкубировали в течение 2 часов при комнатной температуре. После промывания добавляли 100 мкл меченного щелочной фосфатазой антитела против IgG человека (Biosource, Lot#6202), которое было разведено в 5000 раз, и инкубирование проводили в течение 1 часа при комнатной температуре. После промывания добавляли раствор субстрата. После инкубирования измеряли поглощение при 405 нм с использованием Microplate Reader Model 3550 (BioRad). Концентрацию IgG человека в мышиной сыворотке рассчитывали из калибровочной кривой, построенной на основании величин поглощения, полученных с IgG человека в качестве стандарта.
(2) Получение антител для введения
Мономер и димер полипептида scFv/CHO в день введения разводили, соответственно, до концентрации 0,4 мг/мл или 0,25 мг/мл стерилизованным фильтрованием PBS(-) для получения проб для введения.
(3) Получение мышиной модели миеломы человека
Мышиную модель миеломы человека получали следующим образом. Клетки КРММ2, пассированные in vivo (Заявка на патент Японии (JP-Appl. 7-236475) с использованием мышей линии SCID (Japan Clare), суспендировали в среде RPMI1640 (Gibco BRL), содержащей 10% фетальную телячью сыворотку (Gibco BRL), и доводили до 3×107 клеток/мл. 200 мкл суспензии клеток КРММ2 (6×106 клеток/мышь) трансплантировали в мышь SCID (самцы, в возрасте 6 недель) через хвостовую вену, которой за день до трансплантации инъецировали подкожно асиало-GM1-антитело (Wako Junyaku, 1 флакон, растворенный в 5 мл).
(4) Введение антител
Образцы антител, приготовленные в (2): мономер (250 мкл) и димер (400 мкл), вводили мыши-модели миеломы человека, полученной в (3), через хвостовую вену. Введение начинали спустя три дня после трансплантации клеток КРММ2 и проводили дважды в день в течение трех дней. В качестве контроля 200 мкл стерилизованного фильтрованием PBS(-) вводили таким же образом дважды в день в течение трех дней через хвостовую вену. Каждая группа состояла из семи мышей.
(5) Оценка противоопухолевого действия мономера и димера полипептида scFv/CHO с использованием мышиной модели миеломы человека
Противоопухолевое действие мономера и димера полипептида scFv/CHO с использованием мышиной модели миеломы человека оценивали по изменению концентрации IgG человека (М-белка) в мышиной сыворотке и времени выживания мыши. Изменение концентрации IgG человека определяли измерением ее в мышиной сыворотке, собранной спустя 24 дня после трансплантации клеток КРММ2, методом ELISA, описанным выше в (1). Количество сывороточного IgG человека (М-белка) в сыворотке группы (контроль), которой вводили PBS(-), увеличивалось до приблизительно 8500 мкг/мл, тогда как количество IgG человека в группе, которой вводили димер scFv/CHO, было значительно более низким, т.е. 1/10 или меньше от количества в контрольной группе. Таким образом, эти результаты показывают, что димер scFv/CHO сильно ингибирует рост клеток КРММ2 (фигура 32). Как показано на фиг.33, значительное увеличение времени выживания наблюдали в получавшей димер scFv/CHO группе в сравнении с получавшей PBS(-) группой.
Вышеописанное подтверждает, что димер scFv/CHO обладает противоопухолевым действием на мышиной модели миеломы человека. Считается, что противоопухолевое действие димера scFv/CHO, модифицированного антитела данного изобретения, является результатом апоптоз-индуцирующего действия этого модифицированного антитела.
5.15 Тест гемагглютинации
Тест гемагглютинации и определение гемагглютинации проводили в соответствии с "Immuno-Biochemical Investigation", Zoku-Seikagaku Jikken Koza, edited by the Biochemical Society of Japan, published by Tokio Kagaku Dojin.
Кровь брали из здорового донора с использованием обработанных гепарином шприцов и промывали PBS(-) три раза и затем готовили суспензию эритроцитов с конечной концентрацией 2% в PBS(-). Опытными образцами были антитело MABL-2, мономер и димер одноцепочечного полипептида Fv, продуцируемого клетками СНО, и мономер и димер одноцепочечного полипептида Fv, продуцируемого Е.coli, и контролем был IgG мыши (Zymed). Для исследования эффекта гемагглютинации использовали круглодонные 96-луночные планшеты, доступные от Falcon. Добавляли 50 мкл на лунку вышеупомянутых образцов антител и 50 мкл 2% суспензии эритроцитов и смешивали в лунках. После инкубирования в течение 2 часов при 37°С реакционные смеси выдерживали при 4°С в течение ночи и определяли в них гемагглютинацию. В качестве контроля использовали 50 мкл на лунку PBS(-) и тест гемагглютинации проводили таким же образом. Мышиный IgG и антитело MABL-2 использовали в конечной концентрации антител 0,01, 0,1, 1,0, 10,0 или 100,0 мкг/мл. Одноцепочечные Fv использовали в конечной концентрации 0,004, 0,04, 0,4, 4,0, 40,0 или 80,0 мкг/мл и дополнительно 160 мкг/мл только в случае димера этого полипептида, продуцируемого Е.coli. Результаты показаны в таблице 2. В случае антитела MABL-2 гемагглютинацию наблюдали при концентрации более, чем 0,1 мкг/мл, тогда как гемагглютинации не наблюдали как для мономера, так и для димера одноцепочечного Fv.

Пример 6 Модифицированное антитело sc(Fv)2, содержащее две V-области Н-цепи и две V-области L-цепи, и scFv антитела MABL-2, имеющие линкеры различной длины
6.1 Конструирование плазмиды, экспрессирующей sc(Fv)2 антитела MABL-2
Для получения плазмиды, экспрессирующей модифицированное антитело [sc(Fv)2], которое содержит две V-области Н-цепи и две V-области L-цепи, происходящие из антитела MABL-2, вышеупомянутую плазмиду рСНОМ2, которая содержит ДНК, кодирующую scFv, происходящий из MABL-2, описанный выше, модифицировали способом ПЦР, как описывается ниже, и полученный ДНК-фрагмент встраивали в рСНОМ2.
Праймерами, используемыми для ПЦР, являются праймер EF1 (SEQ ID NO 30) в качестве смыслового праймера, который конструируют так, чтобы он гибридизовался с ДНК, кодирующей EF1α, и антисмысловой праймер (SEQ ID NO 19), который конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал ДНК-последовательность, кодирующую линкерный участок, и праймер VLLAS, содержащий сайт узнавания рестриктазы SaIl (SEQ ID NO 31).
100 мкл ПЦР-раствора содержат 10 мкл 10-кратного ПЦР-буфера №1, 1 мМ MgCl2, 0,2 мМ dNTP (dATP, dGTP, dCTP и dTTP), 5 единиц ДНК-полимеразы KOD (Toyobo, Inc.), 1 мкМ каждого из праймеров и 100 нг ДНК-матрицы (рСНОМ2). ПЦР-раствор нагревали при 94°С в течение 30 секунд, при 50°С в течение 30 секунд и при 74°С в течение 1 минуты в указанном порядке. Этот температурный цикл повторяли 30 раз.
ПЦР-продукт очищали с использованием набора QIAquick PCR Purification Kit (Qiagen) и расщепляли SalI. Полученный ДНК-фрагмент клонировали в вектор pBlueScript KS+(Toyobo, Inc.). После секвенирования ДНК плазмиду, содержащую желаемую ДНК-последовательность, расщепляли SalI и полученный ДНК-фрагмент соединяли с использованием набора Rapid DNA Ligation Kit (Boehringer Mannheim) с рСНОМ2, расщепленной SalI. После ДНК-секвенирования ДНК плазмиду, содержащую желаемую ДНК-последовательность, обозначали как "pCHOM2(Fv)2" (см. фиг.34). Нуклеотидная последовательность и аминокислотная последовательность района sc(Fv)2 антитела MABL-2, содержащегося в плазмиде pCHOM2(Fv)2, показаны в SEQ ID NO 32.
6.2 Получение плазмиды, экспрессирующей scFv антитела MABL-2, имеющие линкеры разной длины
Полипептиды scFv, содержащие линкеры различной длины и V-области, которые расположены в следующем порядке [Н-цепь]-[L-цепь] (далее называемые "HL") или [L-цепь]-[Н-цепь] (далее называемые "LH"), получали с использованием в качестве матрицы кДНК, кодирующих Н-цепь и L-цепь, происходящие из MABL-2, как упоминалось выше.
Для конструирования scFv HL-типа ПЦР-процедуру проводили с использованием pCHOM2(Fv)2 в качестве матрицы. На ПЦР-стадии использовали пару праймеров CFHL-Fl (SEQ ID NO 33) и CFHL-R2 (SEQ ID NO 34) или пару праймеров CFHL-F2 (SEQ ID NO 35) и CFHL-R1 (SEQ ID NO 36) и ДНК-полимеразу KOD. ПЦР-процедуру проводили путем повторения 30 раз температурного цикла, состоящего из 94°С в течение 30 секунд, 60°С в течение 30 секунд и 72°С в течение 1 минуты в указанном порядке для получения кДНК для Н-цепи, содержащей лидерную последовательность на 5'-конце, или кДНК для L-цепи, содержащей FLAG-последовательность на ее 3'-конце. Полученные кДНК для Н-цепи и L-цепи смешивали и ПЦР проводили путем повторения 5 раз температурного цикла, состоящего из 94°С в течение 30 секунд, 60°С в течение 30 секунд и 72°С в течение 1 минуты в указанном порядке с использованием этой смеси в качестве матрицы и полимеразы KOD. К этой реакционной смеси добавляли праймеры CFHL-F1 и CFHL-R1 и затем проводили ПЦР-реакцию путем повторения 30 раз вышеупомянутого температурного цикла для получения кДНК для HL-0-типа без линкера.
Для конструирования scFv LH-типа ПЦР-реакцию проводили с использованием в качестве матрицы pGEM-M2L и pGEM-M2H, которые содержат кДНК, кодирующие V-область L-цепи и V-область Н-цепи антитела MABL-2, соответственно (см. Заявку на патент Японии JP-Appl. 11-63557). Использовали пару праймеров Т7 (SEQ ID NO 37) и CFLH-R2 (SEQ ID NO 38) или пару праймеров CFLH-F2 (SEQ ID NO 39) и CFLH-R1 (SEQ ID NO 40) и полимеразу KOD (Toyobo, Inc.). ПЦР-реакцию выполняли повторением 30 раз температурного цикла, состоящего из 94°С в течение 30 секунд, 60°С в течение 30 секунд и 72°С в течение 1 минуты в указанном порядке для получения кДНК L-цепи, содержащей лидерную последовательность на 5'-конце, или кДНК Н-цепи, содержащей FLAG-последовательность на ее 3'-конце. Полученные кДНК L-цепи и Н-цепи смешивали и ПЦР проводили с использованием этой смеси в качестве матрицы и полимеразы KOD повторением 5 раз температурного цикла, состоящего из 94°С в течение 30 секунд, 60°С в течение 30 секунд и 72°С в течение 1 минуты в указанном порядке. К этой реакционной смеси добавляли праймеры Т7 и CFHL-R1 и проводили реакцию повторением 30 раз вышеупомянутого температурного цикла. Продукт реакции использовали в качестве матрицы и ПЦР проводили с использованием пары праймеров CFLH-F4 (SEQ ID NO 41) и CFLH-R1 повторением 30 раз температурного цикла, состоящего из 94°С в течение 30 секунд, 60°С в течение 30 секунд и 72°С в течение 1 минуты в указанном порядке для получения кДНК LH-0-типа без линкера.
Полученные кДНК типов LH-0 и HL-0 расщепляли рестриктазами EcoRI и BamHI (Takara Shuzo) и расщепленные кДНК встраивали в экспрессионную плазмиду INPEP4 для клеток млекопитающих с использованием набора Ligation High (Toyobo Inc.), соответственно. Компетентные клетки Е.coli JM109 (Nippon Gene) трансформировали каждой плазмидой и желаемые плазмиды выделяли из трансформированных Е.coli с использованием набора QIAGEN Plasmid Maxi Kit (Qiagen). Таким образом получали плазмиды pCF2LH-0 и pCF2HL-0.
Для конструирования экспрессионных плазмид HL-типа, содержащих линкеры различного размера, использовали pCF2LH-0 в качестве матрицы и CFHL-X3 (SEQ ID NO 42), CFHL-X4 (SEQ ID NO 43), CFHL-X5 (SEQ ID NO 44), CFHL-X6 (SEQ ID NO 45) или CFHL-X7 (SEQ ID NO 46) в качестве смыслового праймера и праймер BGH-1 (SEQ ID NO 47) в качестве антисмыслового праймера, который комплементарен векторной последовательности. ПЦР-реакции проводили с использованием полимеразы KOD повторением 30 раз температурного цикла, состоящего из 94°С в течение 30 секунд, 60°С в течение 30 секунд и 72°С в течение 1 минуты в указанном порядке и продукты реакции расщепляли рестриктазами XhoI и BamHI (Takara Shuzo). Расщепленные фрагменты встраивали между сайтами XhoI и BamHI в pCF2HL-0 с использованием набора Ligation High (Toyobo Inc.), соответственно. Компетентные клетки Е.coli JM109 трансформировали каждой плазмидой и желаемые плазмиды выделяли из трансформированных Е.coli с использованием набора Qiagen Plasmid Maxi Kit. Таким образом получали экспрессионные плазмиды pCF2HL-3, pCF2HL-4, pCF2HL-5, pCF2HL-6 и pCF2HL-7.
Для конструирования экспрессионной плазмиды для временной экспрессии в клетках COS7 плазмиды pCF2HL-0, pCF2HL-3, pCF2HL-4, pCF2HL-5, pCF2HL-6 и pCF2HL-7 расщепляли рестриктазами EcoRI и BamHI (Takara Shuzo) и полученные фрагменты размером около 800 п.о. очищали электрофорезом в агарозном геле. Полученные фрагменты встраивали между сайтами EcoRI и BamHI в экспрессионной плазмиде pCOSI для экспрессии в клетках млекопитающих с использованием набора Ligation High (Toyobo Inc.), соответственно. Компетентные клетки Е.coli DH5α (Toyobo Inc.) трансформировали каждой плазмидой и желаемые плазмиды выделяли из трансформированных Е.coli с использованием набора Qiagen Plasmid Maxi Kit. Таким образом получали плазмиды CF2HL-0/pCOS1, CF2HL-3/pCOS1, CF2HL-4/pCOS1, CF2HL-5/pCOS1, CF2HL-6/pCOS1 и CF2HL-7/pCOS1.
В качестве типичного примера этих плазмид конструкция плазмиды CF2HL-0/pCOS1 показана на фиг.35, а нуклеотидная последовательность и аминокислотная последовательность MABL2-scFv <HL-0>, содержащегося в этой плазмиде, показаны в SEQ ID NO 48. Нуклеотидные последовательности и аминокислотные последовательности линкерных участков в этих плазмидах также показаны на фиг.36.
Для конструирования экспрессионных плазмид LH-типа, содержащих линкеры различного размера, использовали pCF2LH-0 в качестве матрицы и CFLH-X3 (SEQ ID NO 49), CFLH-X4 (SEQ ID NO 50), CFLH-X5 (SEQ ID NO 51), CFLH-X6 (SEQ ID NO 52) или CFLH-X7 (SEQ ID NO 53) в качестве смыслового праймера и праймер BGH-1 в качестве антисмыслового праймера, который комплементарен векторной последовательности. ПЦР-реакцию проводили с использованием полимеразы KOD повторением 30 раз температурного цикла, состоящего из 94°С в течение 30 секунд, 60°С в течение 30 секунд и 72°С в течение 1 минуты в указанном порядке и продукты реакции расщепляли рестриктазами XhoI и BamHI. Расщепленные фрагменты встраивали между сайтами XhoI и BamHI в pCF2LH-0 с использованием набора Ligation High, соответственно. Компетентные клетки Е.coli DH5α (Toyobo Inc.) трансформировали каждой плазмидой и желаемые плазмиды выделяли из трансформированных Е.coli с использованием набора Qiagen Plasmid Maxi Kit. Таким образом получали плазмиды pCF2LH-3, pCF2LH-4, pCF2LH-5, pCF2LH-6 и pCF2LH-7.
Для конструирования экспрессионной плазмиды для временной экспрессии в клетках COS7 плазмиды pCF2LH-0, pCF2LH-3, pCF2LH-4, pCF2LH-5, pCF2LH-6 и pCF2LH-7 расщепляли рестриктазами EcoRI и BamHI (Takara Shuzo) и полученные фрагменты размером около 800 п.о. очищали электрофорезом в агарозном геле. Полученные фрагменты вводили между сайтами EcoRI и BamHI в экспрессионной плазмиде pCOS1 для экспрессии в клетках млекопитающих с использованием набора Ligation High, соответственно. Компетентные клетки Е.coli DH5α (Toyobo Inc.) трансформировали каждой плазмидой и желаемые плазмиды выделяли из трансформированных Е.coli с использованием набора Qiagen Plasmid Maxi Kit. Таким образом получали плазмиды CF2LH-0/pCOS1, CF2LH-3/pCOS1, CF2LH-4/pCOS1, CF2LH-5/pCOS1, CF2LH-6/pCOS1 и CF2LH-7/pCOS1.
В качестве типичного примера этих плазмид конструкция плазмиды CF2LH-0/pCOS1 показана на фиг.37, а нуклеотидная последовательность и аминокислотная последовательность MABL2-scFv <LH-0>, содержащегося в этой плазмиде, показаны в SEQ ID NO 54. Нуклеотидные последовательности и аминокислотные последовательности линкерных участков в этих плазмидах также показаны на фиг.38.
6.3 Экспрессия scFv и sc(Fv)2 в клетках COS7
(1) Получение культурального супернатанта с использованием сывороточной культуральной среды
scFv и sc(Fv)2 HL-типа и LH-типа временно экспрессировали в клетках COS7 (JCRB9127, Japan Health Sciences Foundation). Клетки COS7 субкультивировали в среде DMEM (Gibco BRL), содержащей 10% фетальную телячью сыворотку (HyClone), при 37°С в СО2-инкубаторе. Клетки COS7 трансфицировали CF2HL-0, 3˜7/pCOS1, или CF2LH-0, 3˜7/pCOS1, полученными в примере 6.2, или вектором pCHOM2(Fv)2 электропорацией с использованием прибора Gene Pulser (BioRad). ДНК (10 мкг) и 0,25 мл суспензии 2×107 клеток/мл в культуральной среде DMEM, содержащей 10% фетальную телячью сыворотку (ФТС) и 5 мМ BES (Sigma), добавляли в кювету. После выдерживания в течение 10 минут эти смеси обрабатывали импульсом при 0,17 кВ, при электрической емкости 950 мкФ. После "восстановления" в течение 10 минут при комнатной температуре электропорированные клетки переносили в культуральную среду DMEM (10% ФТС) в колбе на 75 мл. После культивирования в течение 72 часов культуральный супернатант собирали и центрифугировали для удаления клеточных фрагментов. Культуральный супернатант подвергали фильтрованию с использованием надеваемого на горло колбы фильтра с размером пор 0,22 мкм (Falcon) с получением культурального супернатанта (далее называемого "СМ").
(2) Получение культурального супернатанта с использованием бессывороточной культуральной среды
Клетки, трансфицированные так же, как в (1), переносили в среду DMEM (10% ФТС) в колбе на 75 мл и культивировали в течение ночи. После культивирования супернатант выбрасывали и клетки промывали PBS и затем добавляли к среде CHO-S-SFM II (Gibco BRL). После культивирования в течение 72 часов культуральный супернатант собирали, центрифугировали для удаления клеточных фрагментов и фильтровали с использованием надеваемого на колбу фильтра с размером пор 0,22 мкм (Falcon) с получением СМ.
6.4 Детектирование scFv и sc(Fv)2 в СМ клеток COS7
Различные MABL2-scFv и sc(Fv)2 в СМ клеток COS7, полученные в вышеупомянутом примере 6.3 (2), детектировали методом Вестерн-блоттинга.
Каждый СМ (супернатант) COS7 подвергали электрофорезу в ДСН-ПААГ и переносили на мембрану REINFORCED NC (Schleicher & Schuell). Мембрану блокировали 5% обезжиренным молоком (Morinaga Nyu-guo) и промывали забуференным Трис солевым раствором (TBS). Затем к ней добавляли анти-FLAG-антитело (Sigma). Мембрану инкубировали при комнатной температуре и промывали. Добавляли меченное пероксидазой мышиное IgG-антитело (Jackson ImmunoResearch). После инкубирования и промывания при комнатной температуре добавляли раствор субстрата (Kirkegaard Perry Laboratories) для развития окраски (фиг.39).
6.5 Проточная цитометрия
Проточную цитометрию проводили с использованием культуральных супернатантов клеток COS7, полученных в примере 6.3 (1), для измерения связывания MABL2-scFv и sc(Fv)2 с IAP-антигеном (интегрин-ассоциированным белком) человека. Культуральные супернатанты, подлежащие тестированию, или культуральный супернатант клеток COS7 в качестве контроля добавляли к 2×105 клеток клеточной линии мышиного лейкоза L1210, экспрессирующей интегрин-ассоциированный белок (IAP) человека. После инкубирования на льду и промывания к ним добавляли 10 мкг/мл мышиного анти-FLAG-антитела (Sigma), и затем эти клетки инкубировали и промывали.
Затем к ним добавляли FITC-меченное антитело против мышиного IgG (Becton Dickinson), и клетки опять инкубировали и промывали. Затем измеряли интенсивность флуоресценции с использованием прибора FACScan (Becton Dickinson). Результаты проточной цитометрии показывают, что MABL2-scFv, имеющие линкеры различной длины, и sc(Fv)2 в культуральных супернатантах клеток COS7 имеют высокое сродство к IAP человека (см. фиг.40а и 40b).
6.6 Апоптоз-индуцирующее действие in vitro
Апоптоз-индуцирующее действие культуральных супернатантов COS7, полученных в примере 6.3 (1), исследовали методом окрашивания Аннексином-V (Boehringer Mannheim) с использованием клеток L1210, трансфицированных геном IAP человека (hIAP/L 1210).
К 5×104 клеток hIAP/L1210 добавляли культуральные супернатанты клеток COS7, трансфицированных каждым из векторов, или культуральный супернатант клеток COS7 в качестве контроля в конечной концентрации 10% и смеси культивировали в течение 24 часов. Затем проводили окрашивание Аннексином-V/PI и измеряли интенсивность флуоресценции с использованием прибора FACScan (Becton Dickinson). Результаты показали, что scFv <HL3, 4, 6, 7, LH3, 4, 6, 7> и sc(Fv)2 в СМ клеток COS7 индуцировали в сильной степени гибель клеток hIAP/L1210. Эти результаты показаны на фиг.41.
6.7 Конструирование векторов для экспрессии scFv и sc(Fv)2 в клетках СНО
Для выделения и очистки MABL2-scFv и sc(Fv)2 из культурального супернатанта конструировали экспрессионные вектора для экспрессии в клетках СНО, как описано ниже.
EcoRI-BamHI-фрагменты pCF2HL-0, 3˜7 и pCF2LH-0, 3˜7, полученные в примере 6.2, встраивали между сайтами EcoRI и BamHI в экспрессионный вектор рСНО1 для клеток СНО с использованием набора Ligation High. Компетентные клетки Е.coli DH5α трансформировали каждой плазмидой. Плазмиды выделяли из трансформированной Е.coli с использованием набора Qiagen Plasmid Midi kit (Qiagen) с получением экспрессионных плазмид pCHOM2HL-0, 3˜7 и pCHOM2LH-0, 3˜7.
6.8 Получение клеток СНО, экспрессирующих MABL2-scFv <HL-0, 3˜7>, MABL2-scFv <LH-0, 3˜7> и sc(Fv)2, и получение их культуральных супернатантов
Клетки СНО трансформировали каждой из экспрессионных плазмид pCHOM2HL-0, 3˜7 и pCHOM2LH-0, 3˜7, сконструированных в примере 6.7, и вектором pCHOM2(Fv)2 с получением клеток СНО, стабильно экспрессирующих каждое модифицированное антитело. В качестве типичного примера получение клеток СНО, стабильно экспрессирующих MABL2-scFv <HL-5> или sc(Fv)2, иллюстрируется далее.
Экспрессионные плазмиды pCHOM2HL-5 и pCHOM2(Fv)2 линеаризовали расщеплением рестриктазой PvuI и подвергали трансфекции в клетки СНО электропорацией с использованием прибора Gene Pulser (BioRad). ДНК (10 мкг) и 0,75 мл PBS, содержащего 1×107 клеток/мл, добавляли в кювету и обрабатывали импульсом при 1,5 кВ, при электрической емкости 25 мкФ. После «восстановления» в течение 10 минут при комнатной температуре электропорированные клетки переносили в содержащую нуклеиновую кислоту культуральную среду α-МЕМ (Gibco BRL), содержащую 10% фетальную телячью сыворотку, и культивировали. После культивирования в течение ночи супернатант отбрасывали. Клетки промывали PBS и добавляли в свободную от нуклеиновых кислот культуральную среду α-МЕМ (Gibco BRL), содержащую 10% фетальную телячью сыворотку. После культивирования в течение двух недель клетки культивировали в среде, содержащей 10 нМ (конечная концентрация) метотрексат (Sigma), затем 50 нМ и 100 нМ метотрексат. Полученные клетки культивировали в бессывороточной среде CHO-S-SFM II (Gibco BRL) в роллерных флаконах. Культуральный супернатант собирали, центрифугировали для удаления клеточных фрагментов и фильтровали с использованием фильтра с размером пор 0,22 мкм с получением СМ, соответственно.
Согласно вышеописанному, получали клетки СНО, которые стабильно экспрессируют MABL2-scFv<HL-0, -3, -4, -6, -7, и <LH-0, -3, -4, -5, -6, -7>, и СМ из них.
6.9 Очистка димера MABL2-scFv <HL-5> и sc(Fv)2
MABL2-scFv <HL-5> и sc(Fv)2 очищали из CM, полученных в примере 6.8, с использованием двух способов очистки, описанных ниже.
<Способ очистки 1>
HL-5 и sc(Fv)2 очищали аффинной хроматографией на колонке с анти-FLAG-антителами с использованием FLAG-последовательности, локализованной на С-конце полипептидов, и гель-фильтрацией. Один литр СМ, полученного в 6.8, наносили на колонку (7,9 мл), заполненную анти-FLAG М2-аффинным гелем (Sigma), уравновешенную 50 мМ Трис-HCl-буфером (TBS, рН 7,5), содержащим 150 мМ NaCl. После промывания колонки TBS scFv элюировали 0,1 М глицин-HCl-буфером, рН 3,5. Полученные фракции анализировали электрофорезом в ДСН-ПААГ, и была подтверждена элюция scFv. Фракцию scFv смешивали с Твином 20 до конечной концентрации 0,01% и концентрировали с использованием Centricon-10 (Millipore). Концентрат наносили на колонку TSKgel G3000SWG (7,5×600 мм), уравновешенную 20 мМ ацетатным буфером (рН 6,0), содержащим 150 мМ NaCl и 0,01% Твин 20. При скорости подачи 0,4 мл/мин scFv детектировали по поглощению при 280 нм. HL-5 элюировался в виде основной фракции в положении, соответствующем димеру, a sc(Fv)2 элюировался в положении, соответствующем мономеру.
<Способ очистки 2>
HL-5 и sc(Fv)2 очищали с использованием трех стадий, включающих ионообменную хроматографию, хроматографию на гидроксиапатите и гель-фильтрацию. При ионообменной хроматографии использовали колонку Q sepharose fast flow (Pharmacia) для HL-5, а колонку SP-sepharose fast flow использовали для sc(Fv)2. Начиная со второй стадии, HL-5 и sc(Fv)2 обрабатывали с использованием одной и той же процедуры.
Первая стадия для HL-5
СМ (супернатант) HL-5 разбавляли в два раза 20 мМ Трис-HCl-буфером (рН 9,0), содержащим 0,02% Твин 20, и затем рН доводили до 9,0 1 М Трис. Раствор наносили на колонку Q sepharose fast flow, уравновешенную 20 мМ Трис-HCl-буфером (рН 8,5), содержащим 0,02% Твин 20. Полипептид, адсорбированный на колонке, элюировали линейным градиентом NaCl в том же самом буфере, от 0,1 до 0,55 М. Анализируя элюированные фракции при помощи электрофореза в ДСН-ПААГ, собирали фракции, содержащие HL-5, и подвергали хроматографии с гидроксиапатитом на второй стадии.
Первая стадия для sc(Fv)2
CM sc(Fv)2 разбавляли в два раза 20 мМ ацетатным буфером (рН 5,5), содержащим 0,02% Твин 20, и затем рН доводили до 5,5 1 М уксусной кислотой. Раствор наносили на колонку SP-Sepharose fast flow, уравновешенную 20 мМ ацетатным буфером (рН 5,5), содержащим 0,02% Твин 20. Полипептид, адсорбированный на колонке, элюировали линейным градиентом NaCl в том же самом буфере, от 0 до 0,5 М. Анализируя элюированные фракции при помощи электрофореза в ДСН-ПААГ, собирали фракции, содержащие sc(Fv)2, и подвергали хроматографии с гидроксиапатитом на второй стадии.
Вторая стадия: Хроматография HL-5 и sc(Fv)2 на гидроксиапатите
Фракции HL-5 и sc(Fv)2, полученные на первой стадии, по отдельности наносили на гидроксиапатитную колонку (Типа I, BioRad), уравновешенную 10 мМ фосфатным буфером, содержащим 0,02% Твин 20, рН 7,0. После промывания колонки тем же самым буфером полипептиды, адсорбированные на колонке, элюировали линейным градиентом фосфатного буфера до 0,5 М. Анализируя элюированные фракции при помощи электрофореза в ДСН-ПААГ, собирали фракции, содержащие желаемые полипептиды.
Третья стадия: Гель-фильтрация HL-5 и sc(Fv)2
Каждую фракцию, полученную на второй стадии, отдельно концентрировали с использованием CentriPrep-10 (Millipore) и наносили наносили на колонку Superdex 200 (2,6×60 см, Pharmacia), уравновешенную 20 мМ ацетатным буфером (рН 6,0), содержащим 0,02% Твин 20 и 0,15 М NaCl. HL-5 элюировался в положении, соответствующем димеру, a sc(Fv)HL-5 и sc(Fv)2 элюировались в виде основного пика в положении, соответствующем мономеру, соответственно.
Поскольку мономер HL-5 почти не детектировался ни при одном из двух способов очистки, это показало, что димеры одноцепочечных Fv образовывались с высокими выходами, когда линкер для одноцепочечного Fv содержит около 5 аминокислот. Кроме того, димер HL-5 и sc(Fv)2 стабильно сохранялись в течение месяца при 4°С после очистки.
6.10 Оценка связывающей активности очищенных димера scFv<HL-5> и sc(Fv)2 в отношении антигена
Проточную цитометрию проводили с использованием очищенного димера MABL2-scFv <HL-5> и очищенного sc(Fv)2 для оценки связывания с IAP-антигеном (интегрин-ассоциированным белком) человека. 10 мкг/мл очищенного димера MABL2-scFv <HL-5>, очищенного sc(Fv)2, антитела MABL-2 в качестве положительного контроля или мышиного IgG (Zymed) в качестве отрицательного контроля добавляли к 2×105 клеток мышиной клеточной линии лейкоза L1210, экспрессирующей IAP человека (hIAP/L1210), или клеточной линии L1210, трансформированной pCOS1 (pCOS1/L1210) в качестве контроля. После инкубирования на льду и промывания добавляли 10 мкг/мл мышиного анти-FLAG-антитела (Sigma) и затем клетки инкубировали и промывали. Добавляли FITC-меченое антитело против мышиного IgG (Becton Dickinson) и клетки опять инкубировали и промывали. Затем измеряли интенсивность флуоресценции с использованием прибора FACScan (Becton Dickinson).
Поскольку очищенный димер MABL2-scFv <HL-5> и очищенный sc(Fv)2 специфически связывались с клетками hIAP/L1210, было подтверждено, что димер scFv <HL-5> и sc(Fv)2 обладают высоким сродством к IAP человека (см. фиг.42).
6.11 Апоптоз-индуцирующая активность очищенного димера scFv <HL-5> и sc(Fv)2 in vitro
Апоптоз-индуцирующее действие очищенного димера MABL2-scFv <HL-5> и очищенного sc(Fv)2 испытывали по окрашиванию Аннексином-V (Boehringer Mannheim) с использованием клеток L1210 (hIAP/L1210), в которые был введен ген IAP человека, и клеток клеточной линии лейкоза человека CCRF-CEM.
Различные концентрации очищенного димера MABL2-scFv <HL-5>, очищенного MABL2-sc(Fv)2, антитела MABL-2 в качестве положительного контроля или мышиного IgG в качестве отрицательного контроля добавляли к 5×104 клеток клеточной линии hIAP/L1210 или 1×105 клеток клеточной линии CCRF-CEM. После культивирования в течение 24 часов проводили окрашивание Аннексином-V и интенсивность флуоресценции измеряли с использованием прибора FACScan (Becton Dickinson). В результате димер MABL2-scFv <HL-5> и MABL2-sc(Fv)2 в сильной степени индуцировали гибель клеток hIAP/L1210 и CCRF-CEM зависимым от концентрации образом (см. фиг.43). В результате было показано, что димер MABL2-scFv <HL-5> и MABL2-sc(Fv)2 с лучшей эффективностью индуцировали апоптоз в сравнении с исходным антителом MABL-2.
6.12 Тест гемагглютинации с использованием очищенных димера scFv <HL-5> и sc(Fv)2
Тест гемагглютинации проводили с использованием различных концентраций очищенного димера scFv <HL-5> и очищенного sc(Fv)2 в соответствии с примером 5.15.
Гемагглютинацию наблюдали с антителом MABL-2 в качестве положительного контроля, тогда как гемагглютинацию не наблюдали как с одноцепочечным антителом MABL2-sc(Fv)2, так и с MABL2-scFv <HL-5>. Кроме того, не было существенного различия в гемагглютинации между двумя буферами, используемыми с антителом MABL-2. Эти результаты показаны в Таблице 3.
Тест гемагглютинации Разбавитель: PBS (мкг/мл)
6.13 Противоопухолевое действие очищенных димера scFv <HL-5> и sc(Fv)2 на мышиной модели миеломы человека
Противоопухолевые эффекты тестировали для димера scFv <HL-5> и sc(Fv)2, полученных и очищенных в примерах 6.8 и 6.9. Тест проводили с использованием мышиной модели миеломы человека, полученной в примере 5.1, путем определения количества М-белка, продуцируемого клетками миеломы человека, в мышиной сыворотке при помощи ELISA и изучения времени выживания мышей. Затем противоопухолевые эффекты димера scFv <HL-5> и sc(Fv)2 оценивали по изменению количества М-белка в мышиной сыворотке и времени выживания мышей.
В этом тесте HL-5 и sc(Fv)2 использовали в виде раствора с концентрацией 0,01, 0,1 или 1 мг/мл в растворителе, содержащем 150 мМ NaCl, 0,02% Твин и 20 мМ ацетатный буфер, рН 6,0, и вводили мышам в дозе 0,1, 1 или 10 мг/кг. Контрольной группе мышей вводили только растворитель.
Сыворотку мышей собирали спустя 26 дней после трансплантации миеломных клеток человека и количество М-белка в сыворотке измеряли при помощи ELISA в соответствии с примером 5.14. В результате количество М-белка в сыворотке обеих групп мышей, которым вводили HL-5, димер и sc(Fv)2, уменьшалось зависимым от дозы образом (см. фиг.44). Кроме того, наблюдали значительное удлинение времени выживания в обеих группах, которым вводили HL-5 (фиг.45) и sc(Fv)2 (фиг.46), в сравнении с контрольной группой, которой вводили растворитель. Эти результаты показывают, что HL-5 и sc(Fv)2 данного изобретения проявляют превосходное противоопухолевое действие in vivo.
Пример 7
Одноцепочечный Fv, содержащий V-область Н-цепи и V-область L-цепи человеческого антитела 12В5 против MPL человека
ДНК, кодирующую V-области человеческого моноклонального антитела 12В5 против MPL человека, конструировали следующим образом:
7.1 Конструирование гена, кодирующего V-область Н-цепи 12В5
Ген, кодирующий V-область Н-цепи человеческого антитела12В5, связывающегося с MPL человека, конструировали присоединением нуклеотидной последовательности этого гена (SEQ ID NO 55) по 5'-концу к лидерной последовательности (SEQ ID NO 56), происходящей из гена человеческого антитела (Eur. J. Immunol. 1996; 26: 63-69). Сконструированную нуклеотидную последовательность разделяли на четыре олигонуклеотида, имеющих перекрывающиеся последовательности из 15 п.о. каждый (12B5VH-1, 12B5VH-2, 12B5VH-3, 12B5VH-4). 12B5VH-1 (SEQ ID NO 57) и 12B5VH-3 (SEQ ID NO 59) синтезировали в смысловой ориентации, а 12B5VH-2 (SEQ ID NO 58) и 12B5VH-4 (SEQ ID NO 60) - в антисмысловой ориентации, соответственно. После сборки синтезированных олигонуклеотидов на основе взаимной комплементарности, добавляли внешние праймеры 12B5VH-S и 12B5VH-A) для амплификации полноразмерного гена. 12B5VH-S (SEQ ID NO 61) конструировали так, чтобы он как прямой праймер гибридизовался с 5'-концом лидерной последовательности и содержал сайт узнавания рестриктазы HindIII и последовательность Козака, а 12B5VH-A (SEQ ID NO 62) конструировали так, чтобы он как обратный праймер гибридизовался с нуклеотидной последовательностью, кодирующей С-конец V-области Н-цепи, и содержал донорный сайт сплайсинга и сайт узнавания рестриктазы BamHI, соответственно.
100 мкл ПЦР-раствора, содержащего 10 мкл 10-кратного ПЦР-буфера Gold II, 1,5 мМ MgCl2, 0,08 мМ dNTP (dATP, dGTP, dCTP и dTTP), 5 единиц ДНК-полимеразы AmpliTaq Gold (все от Perkin Elmer) и по 2,5 пмоль каждого из синтезированных олигонуклеотидов 12B5VH-1 - 4), нагревали при начальной температуре 94°С в течение 9 минут, при 94°С в течение 2 минут, при 55°С в течение 2 минут и при 72°С в течение 2 минут. После повторения этого цикла два раза добавляли 100 пмоль внешних праймеров 12B5VH-S и 12B5VH-A. Смесь подвергали 35 циклам, состоящим из нагревания при 94°С в течение 30 секунд, при 55°С в течение 30 секунд и 72°С в течение 1 минуты, и нагревали при 72°С в течение дополнительных 5 минут.
ПЦР-продукт очищали с использованием 1,5% геля легкоплавкой агарозы (Sigma), расщепляли рестриктазами BamHI и HindIII и клонировали в экспрессионный вектор HEF-gγ1 для Н-цепи человека. После определения последовательности ДНК плазмида, содержащая правильную ДНК-последовательность, была обозначена как HEF-12B5H-gγ1.
HEF-12B5H-gγ1 расщепляли рестриктазами EcoRI и BamHI с получением гена, кодирующего 12B5VH, который затем клонировали в экспрессионный вектор pCOS-Fd для Fab Н-цепи человека с получением pFd-12B5H. Экспрессионный вектор для Fab Н-цепи человека конструировали путем амплификациии ДНК (SEQ ID NO 63), содержащей участок интрона, находящийся между генами, кодирующими V-область Н-цепи и константную область антитела человека, и ген, кодирующий часть константной области Н-цепи человека, при помощи ПЦР и встраивания этого ПЦР-продукта в экспрессионный вектор для клеток животных pCOS1. Константную область Н-цепи человека амплифицировали для этого гена в условиях, описанных выше, с использованием в качестве матрицы HEF-gγ1, в качестве прямого праймера G1CH1-S (SEQ ID NO 64), который конструировали так, чтобы он гибридизовался с 5'-концевой последовательностью интрона 1 и содержал сайт узнавания рестриктаз EcoRI и BamHI, и в качестве обратного праймера G1CH1-A (SEQ ID NO 65), который конструировали таким образом, чтобы он гибридизовался с 3'-концом ДНК домена CHl константной области Н-цепи человека и содержал последовательность, кодирующую часть шарнирной области, два стоп-кодона и сайт узнавания рестриктазы BglII.
Нуклеотидная последовательность и аминокислотная последовательность реконструированной вариабельной области Н-цепи 12В5, которая была встроена в плазмиды HEF-12B5H-gγ1 и pFd-12B5H, показаны в SEQ ID NO 66.
7.2 Конструирование гена, кодирующего V-область L-цепи 12В5
Ген, кодирующий V-область L-цепи человеческого антитела 12В5, связывающегося с MPL человека, конструировали присоединением нуклеотидной последовательности этого гена (SEQ ID NO 67) по 5'-концу к лидерной последовательности (SEQ ID NO 68), происходящей из гена человеческого антитела 3D6 (Nuc. Acid Res. 1990; 18: 4927). Таким же образом, как описано выше, эту сконструированную нуклеотидную последовательность разделяли на четыре олигонуклеотида, имеющих перекрывающиеся последовательности из 15 п.о. каждый (12B5VL-1, 12B5VL-2, 12B5VL-3, 12B5VL-4), и синтезировали соответственно. 12B5VL-1 (SEQ ID NO 69) и 12B5VL-3 (SEQ ID NO 71) имели смысловую ориентацию, а 12B5VL-2 (SEQ ID NO 70) и 12B5VL-4 (SEQ ID NO 72) имели антисмысловую ориентацию, соответственно. Синтезированные олигонуклеотиды состыковывали на основе взаимной комплементарности и смешивали с внешними праймерами 12B5VL-S и 12B5VL-A) для амплификации полноразмерного гена. 12B5VL-S (SEQ ID NO 73) конструировали так, чтобы он как прямой праймер гибридизовался с 5'-концом лидерной последовательности и содержал сайт узнавания рестриктазы HindIII и последовательность Козака. 12B5VL-A (SEQ ID NO 74) конструировали так, чтобы он как обратный праймер гибридизовался с нуклеотидной последовательностью, кодирующей С-конец V-области L-цепи, и содержал донорный сайт сплайсинга и сайт узнавания рестриктазы BamHI.
После проведения ПЦР, как описано выше, ПЦР-продукт очищали с использованием 1,5% геля легкоплавкой агарозы (Sigma), расщепляли рестриктазами BamHI и HindIII и клонировали в экспрессионный вектор HEF-gκ для L-цепи человека. После определения последовательности ДНК плазмида, содержащая правильную ДНК-последовательность, была обозначена как HEF-12B5L-gκ. Нуклеотидная последовательность и аминокислотная последовательность реконструированной V-области L-цепи 12В5, которая была встроена в плазмиду HEF-12B5L-gκ, показаны в SEQ ID NO 75.
7.3 Получение реконструированного одноцепочечного Fv 12B5 (scFv)
Реконструированный одноцепочечный Fv антитела 12B5 конструировали так, чтобы он имел структуру 12В5VН-линкер-12В5VL и содержал FLAG-последовательность (SEQ ID NO 76) на С-конце для облегчения детекции и очистки. Реконструированный одноцепочечный Fv 12B5 (sc12B5) конструировали с использованием линкерной последовательности, состоящей из 15 аминокислот, представленной (Gly4Ser)3.
(1) Получение реконструированного одноцепочечного Fv 12B5 с использованием линкерной последовательности, состоящей из 15 аминокислот
Ген, кодирующий реконструированный одноцепочечный Fv антитела 12B5, который содержал линкерную последовательность, состоящую из 15 аминокислот, конструировали соединением V-области Н-цепи 12B5, линкерного участка и V-области L-цепи 12B5, с амплификацией этого продукта при помощи ПЦР, соответственно. Этот способ изображен схематически на фиг.47. Для получения этого реконструированного одноцепочечного Fv 12B5 использовали шесть праймеров (A-F). Праймеры А, С и Е имели смысловые последовательности, а праймеры В, D и F имели антисмысловые последовательности.
Прямой праймер 12B5-S (Праймер A, SEQ ID NO 77) для V-области Н-цепи конструировали так, чтобы он гибридизовался с 5'-концом лидерной последовательности Н-цепи и содержал сайт узнавания рестриктазы EcoRI. Обратный праймер HuVHJ3 (Праймер В, SEQ ID NO 78) для V-области Н-цепи конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области Н-цепи.
Прямой праймер RHuJH3 (Праймер С, SEQ ID NO 79) для линкера конструировали так, чтобы он гибридизовался с ДНК, кодирующей N-конец линкера, и перекрывался с ДНК, кодирующей С-конец V-области Н-цепи. Обратный праймер RHuVK1 (Праймер D, SEQ ID NO 80) для линкера конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец линкера, и перекрывался с ДНК, кодирующей N-конец V-области L-цепи.
Прямой праймер HuVK-1.2 (Праймер Е, SEQ ID NO 81) для V-области L-цепи конструировали так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области L-цепи. Обратный праймер 12B5F-A для V-области L-цепи (Праймер F, SEQ ID NO 82) конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал последовательность, кодирующую пептид FLAG (Hopp N.P. et al., Bio/Technology, 6, 1204-1210, 1988), два транскрипционных стоп-кодона и сайт узнавания рестриктазы NotI.
На первой стадии ПЦР проводили три реакции А-В, C-D и E-F, и три ПЦР-продукта, полученные на первой стадии ПЦР, состыковывали на основе взаимной комплементарности. После добавления праймеров А и F амплифицировали полноразмерную ДНК, кодирующую реконструированный одноцепочечный Fv 12B5, содержащий линкер, состоящий из 15 аминокислот (вторая ПЦР). В ПЦР первой стадии в качестве матриц использовали плазмиду HEF-12B5H-gγ1 (см. пример 7.1), кодирующую реконструированную V-область Н-цепи 12B5, pSCFVT-hM21 (гуманизированное антитело ONS-M21) (Ohtomo et al., Anticancer Res. 18 (1998), 4311-4316), содержащую ДНК (SEQ ID NO 83), кодирующую линкерный участок, состоящий из Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser (Huston et al., Proc. Natl. Acad. Sci. USA, 85, 5879-5883, 1988), и плазмиду HEF-12B5L-gκ (см. пример 7.2), кодирующую реконструированную V-область L-цепи 12B5, соответственно.
50 мкг ПЦР-раствора для первой стадии ПЦР содержали 5 мкл 10-кратного ПЦР-буфера Gold II, 1,5 мМ MgCl2, 0,08 мМ dNTP, 5 единиц ДНК-полимеразы AmpliTaq Gold (все от Perkin Elmer), no 100 пмоль каждого праймера и 100 нг каждой ДНК-матрицы. ПЦР-раствор нагревали при начальной температуре 94°С в течение 9 минут, затем при 94°С в течение 30 секунд, при 55°С в течение 30 секунд и при 72°С в течение 1 минуты. После повторения этого цикла 35 раз реакционную смесь дополнительно нагревали при 72°С в течение 7 минут.
ПЦР-продукты А-В, C-D и E-F состыковывали при помощи второй ПЦР. Раствор ПЦР-смеси для второй стадии в объеме 98 мкл, содержащий в качестве матрицы 1 мкл ПЦР-продукта А-В первой стадии, 0,5 мкл ПЦР-продукта C-D и 1 мкл ПЦР-продукта C-D и 1 мкл ПЦР-продукта E-F, 10 мкл 10-кратного ПЦР-буфера Gold II, 1,5 мМ MgCl2, 0,08 мМ dNTP, 5 единиц ДНК-полимеразы AmpliTaq Gold (все от Perkin Elmer), нагревали при начальной температуре 94°С в течение 9 минут, затем при 94°С в течение 2 минут, при 65°С в течение 2 минут и при 72°С в течение 2 минут.После повторения этого цикла два раза добавляли по 100 пмоль каждого из праймеров А и F. После повторения цикла, состоящего из нагревания при 94°С в течение 30 секунд, при 55°С в течение 30 секунд и при 72°С в течение 1 минуты, 35 раз реакционную смесь нагревали при 72°С в течение 5 минут.
ДНК-фрагменты, полученные в результате второй ПЦР, очищали с использованием 1,5% геля легкоплавкой агарозы, расщепляли EcoRI и NotI и клонировали в вектор рСНО1 и вектор pCOS1 (Патентная заявка Японии №8-255196). Экспрессионный вектор pCHO1 представлял собой вектор, полученный делецией гена антитела из DHFR-ΔE-rv-PM1-f (см. WO 92/19759) в результате расщепления EcoRI и SmaI и присоединением EcoRI-NotI-BamHI-адаптора (Takara Shuzo). После определения последовательности ДНК плазмиды, содержащие ДНК-фрагмент, кодирующий правильную аминокислотную последовательность реконструированного одноцепочечного Fv 12B5, были обозначены как pCHO-sc12B5 и pCOS-sc12B5. Нуклеотидная последовательность и аминокислотная последовательность реконструированного одноцепочечного Fv антитела 12B5, включенного в плазмиды pCHO-sc12B5 и pCOS-sc12B5, показаны в SEQ ID NO 84.
7.4 Экспрессия антитела 12B5 (IgG, Fab) и одноцепочечного полипептида Fv клеткой животного
Антитело 12B5 (IgG, Fab) и одноцепочечный Fv, полученный из антитела 12B5, экспрессировали с использованием клеток COS7 или клеток СНО.
Временную экспрессию с использованием клеток COS7 осуществляли следующим образом. Трансфекцию проводили способом электропорации с применением устройства Gene Pulser (BioRad). Для экспрессии антитела 12В5 (IgG) добавляли по 10 мкг вышеупомянутых экспрессионных векторов HEF-12B5H-gγ1 и HEF-12B5L-gκ, для экспрессии 12В5 Fab-фрагмента добавляли по 10 мкг pFd-12B5H и HEF-12B5L-gκ и для экспрессии одноцепочечного Fv 10 мкг pCOS-sc12B5 добавляли к клеткам COS7 (1×107 клеткам/мл), суспендированным в 0,8 мл PBS. Смесь, находящуюся в кювете, обрабатывали импульсом 1,5 кВ, при электрической емкости 25 мкФ. После "восстановления" в течение 10 минут при комнатной температуре электропорированные клетки добавляли в культуральную среду DMEM (Gibco BRL), содержащую 10% фетальную телячью сыворотку, и культивировали. После культивирования в течение ночи клетки промывали один раз PBS, добавляли в бессывороточную среду CHO-S-SFM II и культивировали в течение 2 дней. Культуральную среду центрифугировали для удаления клеточного дебриса (остатков клеток) и фильтровали через фильтр с размером пор 0,22 мкм для получения культурального супернатанта.
Для получения клеточной линии СНО, стабильно экспрессирующей одноцепочечный Fv (полипептид), происходящий из антитела 12В5, экспрессионный вектор pCHO-sc12B5 вводили в клетки СНО следующим образом.
Экспрессионный вектор вводили в клетки СНО электропорацией с использованием прибора Gene Pulser (BioRad). Линеаризованную ДНК (100 мкг), полученную расщеплением рестриктазой Pvul, и клетки СНО (1×107 клеток/мл), суспендированные в 0,8 мл PBS, смешивали в кювете, оставляли без перемешивания на льду на 10 минут и обрабатывали импульсом 1,5 кВ, при электрической емкости 25 мкФ. После "восстановления" в течение 10 минут при комнатной температуре электропорированные клетки добавляли в среду CHO-S-SFM II (Gibco BRL), содержащую 10% фетальную телячью сыворотку, и культивировали. После культивирования в течение 2 дней культивирование продолжали в CHO-S-SFM II (Gibco BRL), содержащей 5 нМ метотрексат (Sigma) и 10% фетальную телячью сыворотку. Из полученных таким образом клонов отбирали клон с высокой скоростью экспрессии в качестве клеточной линии-продуцента одноцепочечного Fv 12B5. После культивирования в бессывороточной среде CHO-S-SFM II (Gibco BRL), содержащей 5 нМ метотрексат (Sigma), получали культуральный супернатант центрифугированием для отделения клеточного дебриса.
7.5 Очистка одноцепочечного Fv, происходящего из 12B5, продуцируемого клетками СНО
Культуральный супернатант клеточной линии СНО, экспрессирующей одноцепочечный Fv 12B5, полученный в 7.4, очищали с использованием колонки с анти-FLAG-антителом и гель-фильтрационной колонки.
(1) Колонка с анти-FLAG-антителом
Культуральный супернатант добавляли к анти-FLAG М2-аффинному гелю (Sigma), уравновешенному PBS. После промывания этой колонки тем же самым буфером белки, адсорбированные на колонке, элюировали 0,1 М глицин-HCl-буфером (рН 3,5). Элюированные фракции незамедлительно нейтрализовали добавлением 1 М буфера Трис-HCl (рН 8,0). Элюированные фракции анализировали электрофорезом в ДСН-ПААГ, и фракцию, которая, как было подтверждено, содержала одноцепочечный Fv, концентрировали с использованием Centricon-10 (Millipore).
(2) Гель-фильтрация
Концентрированный раствор, полученный в (1), наносили на колонку Superdex 200 (10×300 мм, Amersham Pharmacia), уравновешенную PBS, содержащим 0,01% Твин 20. Продукт sc12B5 элюировался в виде двух пиков (А, В) (см. фиг.48). Фракции А и В анализировали при помощи электрофореза в 14% ДСН-полиакриламидном геле. Электрофорез образцов проводили в присутствии и в отсутствие восстанавливающего агента согласно способу Лэммли и окрашивали Кумасси бриллиантовым синим после электрофореза. Как показано на фиг.49, фракции А и В, независимо от присутствия восстанавливающего агента, давали единственную полосу, имеющую кажущуюся молекулярную массу около 31 кДа. При анализе фракций А и В гель-фильтрацией с использованием Superdex 200 PC 3.2/30 (3,2×300 мм, Amersham Pharmacia), фракция А давала элюированный продукт с кажущейся молекулярной массой около 44 кДа, а фракция В давала продукт с кажущейся молекулярной массой 22 кДа (см. фиг.50а и b). Эти результаты показывают, что фракция А является связанным нековалентно димером одноцепочечного Fv sc12B5, а В является мономером.
7.6 Измерение ТРО-подобной агонистической активности различных одноцепочечных Fv
ТРО-подобную активность анти-MPL-одноцепочечного антитела оценивали путем определения пролиферативного действия на клетки Ba/F3 (BaF/MpI), экспрессирующие ТРО-рецептор человека (MPL). После промывания клеток BaF/MpI два раза культуральной средой RPMI1640 (Gibco), содержащей 10% фетальную телячью сыворотку (Gibco), клетки суспендировали в культуральной среде при плотности клеток 5×105 клеток/мл. Анти-MPL-одноцепочечное антитело и ТРО человека (R & D System) разбавляли этой культуральной средой, соответственно. 50 мкл клеточной суспензии и 50 мкл разведенного антитела или ТРО человека добавляли в 96-луночный планшет (плоскодонный) (Falcon) и культивировали в СО2-инкубаторе (концентрация CO2: 5%) в течение 24 часов. После инкубирования добавляли 10 мкл реагента WST-8 (реагента для определения числа живых клеток SF: Nacalai Tesque) и поглощение немедленно измеряли при длине волны измерения 450 нм и при опорной длине волны 620 нм с использованием флуоресцентно-абсорбционного фотометра SPECTRA Fluor (Tecan). После инкубирования в СО2-инкубаторе (концентрация CO2: 5%) в течение 2 часов оптическую плотность при длине волны измерения 450 нм и опорной длине волны 620 нм опять измеряли при помощи SPECTRA Fluor. Поскольку реагент WST-8 развивал окраску при длине волны 450 нм в зависимости от числа живых клеток, пролиферативную активность BaF/Mp1 на основе изменения оптической плотности через 2 часа оценивали по ED50, рассчитываемой следующим образом. На кривой реакции пролиферации, где оптическую плотность откладывали по оси ординат против концентрации антитела по оси абсцисс, оптическая плотность на плато принималась за 100% скорость реакции. Получив формулу аппроксимации по способу линейной аппроксимации на основе отложенных величин, близких к 50% скорости реакции, рассчитывали концентрацию антител, соответствующую 50% скорости реакции, и принимали ее за ED50.
Результаты агонистической активности в отношении MPL, измеренные с использованием культуральных супернатантов клеток COS7, экспрессирующих различные варианты молекулы антитела 12В5, обнаружили, как иллюстрируется на фиг.51, что антитело 12B5IgG, имеющее бивалентный антигенсвязывающий сайт, увеличивало оптическую плотность зависимым от концентрации образом и имело ТРО-подобную агонистическую активность (ED50; 29 нМ), тогда как агонистическая активность антитела 12B5Fab, имеющего моновалентный антигенсвязывающий сайт, была очень слабой (ED50; 34724 нМ). В противоположность этому, одноцепочечный Fv (sc12D5), имеющий моновалентный антигенсвязывающий сайт, как и Fab, обнаруживал сильную агонистическую активность при величине ED50 75 нМ. Однако было известно, что вариабельные области Н-цепи и L-цепи одноцепочечного Fv связаны через нековалентную связь, и, следовательно, каждая вариабельная область диссоциирована в растворе и может быть ассоциирована с вариабельной областью другой молекулы с образованием мультимеров, таких как димеры. При измерении молекулярной массы sc12B5, очищенного гель-фильтрацией, было подтверждено существование молекул, которые были признаны как мономер и димер (см. фиг.48). Затем выделяли мономер sc12B5 и димер sc12B5 (см. фиг.50) и измеряли их агонистическую активность в отношении MPL. Как показано на фиг.51 и 52, ED50 мономера sc12B5 была 4438,7 нМ, что подтверждало, что агонистическая активность была меньше в сравнении с результатом, полученным с использованием культурального супернатанта клеток COS7. В противоположность этому одноцепочечный Fv (димер sc12B5), имеющий бивалентный антигенсвязывающий сайт, показал приблизительно в 400 раз более сильную агонистическую активность (ED50; 10,1 нМ) в сравнении с моновалентным sc12B5. Кроме того, бивалентный одноцепочечный Fv обнаружил агонистическую активность, эквивалентную или более высокую, чем агонистическая активность ТРО и 12B5IgG человека.
Пример 8
Конструирование гена, кодирующего вариабельную область человеческого антитела 12Е10 против MPL человека
ДНК, кодирующую вариабельную область человеческого моноклонального антитела 12Е10 против MPL человека, конструировали следующим образом.
8.1 Конструирование гена, кодирующего V-область Н-цепи 12Е10
Нуклеотидную последовательность SEQ ID NO 86 конструировали как ген, кодирующий V-область Н-цепи человеческого антитела 12Е10, связывающегося с MPL человека, на основе аминокислотной последовательности, описанной в WO 99/10494 (SEQ ID NO 85). Полноразмерную нуклеотидную последовательность конструировали присоединением к ее 5'-концу лидерной последовательности (SEQ ID NO 87), происходящей из гена человеческого антитела (номер доступа GenBank AF062252). Сконструированную нуклеотидную последовательность разделяли на четыре олигонуклеотида, имеющих перекрывающиеся последовательности из 15 п.о. каждая (12E10VH-1, 12E10VH-2, 12E10VH-3, 12E10VH-4). 12E10VH-1 (SEQ ID NO 88) и 12E10VH-3 (SEQ ID NO 90) синтезировали в смысловой ориентации, а 12E10VH-2 (SEQ ID NO 89) и 12E10VH-4 (SEQ ID NO 91) в антисмысловой ориентации, соответственно. После состыковки синтезированных олигонуклеотидов на основе взаимной комплементарности, внешние праймеры 12E10VH-S и 12E10VH-A) добавляли для амплификации полноразмерного гена. 12E10VH-S (SEQ ID NO 92) конструировали так, чтобы он как прямой праймер гибридизовался с 5'-концом лидерной последовательности и содержал сайт узнавания рестриктазы HindIII и последовательность Козака, a 12E10VH-А (SEQ ID NO 93) конструировали так, чтобы он как обратный праймер гибридизовался с нуклеотидной последовательностью, кодирующей С-конец V-области Н-цепи, и содержал донорноый сайт сплайсинга и сайт узнавания рестриктазы BamHI, соответственно.
100 мкл ПЦР-раствора, содержащего 10 мкл 10-кратного ПЦР-буфера Gold II, 1,5 мМ MgCl2 0,08 мМ dNTP (dATP, dGTP, dCTP и dTTP), 5 единиц ДНК-полимеразы AmpliTaq Gold (все от Perkin Elmer) и по 2,5 пмоль каждого из синтезированных олигонуклеотидов 12E10VH-1-4), нагревали при начальной температуре 94°С в течение 9 минут, затем при 94°С в течение 2 минут, при 55°С в течение 2 минут и при 72°С в течение 2 минут. После повторения этого цикла два раза добавляли 100 пмоль внешних праймеров 12E10VH-S и 12E10VH-A. Смесь подвергали циклу, состоящему из нагревания при 94°С в течение 30 секунд, при 55°С в течение 30 секунд и 72°С в течение 1 минуты, 35 раз и нагревали при 72°С в течение дополнительных 5 минут.
ПЦР-продукт очищали с использованием 1,5% геля легкоплавкой агарозы (Sigma), расщепляли рестриктазами BamHI и HindIII и клонировали в экспрессионный вектор HEF-gγ1 для Н-цепи человека. После определения ДНК-последовательности плазмида, содержащая правильную ДНК-последовательность, была названа HEF-12E10H-gγ1.
HEF-12E10H-gγ1 расщепляли рестриктазами EcoRI и BamHI с получением гена, кодирующего 12E10VH, который затем клонировали в экспрессионный вектор pCOS-Fd для Fab Н-цепи человека для получения pFd-12E10H. Экспрессионный вектор для Fab Н-цепи человека конструировали путем амплификации ДНК (SEQ ID NO 63), содержащей участок интрона, находящийся между генами, кодирующими V-область Н-цепи и константную область антитела человека, и ген, кодирующий часть константной области Н-цепи человека, при помощи ПЦР и встраивания этого ПЦР-продукта в экспрессионный вектор для клеток животного pCOS1. Константную область Н-цепи человека амплифицировали для этого гена в условиях, описанных выше, с использованием в качестве матрицы HEF-gγ1, в качестве прямого праймера G1CH1-S (SEQ ID NO 64), который конструировали так, чтобы он гибридизовался с 5'-концевой последовательностью интрона 1 и содержал сайты узнавания рестриктаз EcoRI и BamHI, и в качестве обратного праймера G1CH1-A (SEQ ID NO 65), который конструировали так, чтобы он гибридизовался с 3'-концом ДНК домена СН1 константной области Н-цепи человека и содержал последовательность, кодирующую часть шарнирной области, два стоп-кодона и сайт узнавания рестриктазы BgIII.
Нуклеотидная последовательность и аминокислотная последовательность реконструированной вариабельной области Н-цепи 12Е10Н, которая была включена в плазмиды HEF-12E10H-gγ1 и pFd-12E10H, показаны в SEQ ID NO 94.
8.2 Конструирование гена, кодирующего V-область L-цепи 12Е10
Нуклеотидную последовательность SEQ ID NO 96 конструировали как ген, кодирующий V-область L-цепи человеческого антитела 12Е10, связывающегося с MPL человека, на основе аминокислотной последовательности, описанной в WO 99/10494 (SEQ ID NO 95). Дополнительно конструкцией было предусмотрено присоединение к ее 5'-концу лидерной последовательности (SEQ ID NO 97), происходящей из гена человеческого антитела (Mol. Immunol. 1992; 29: 1515-1518). Таким же образом, как описано выше, эту сконструированную нуклеотидную последовательность разделяли на четыре олигонуклеотида, имеющих перекрывающиеся последовательности из 15 п.о. каждая (12E10VL1, 12E10VL2, 12E10VL3, 12E10VL4), и синтезировали соответственно. 12E10VL1 (SEQ ID NO 98) и 12E10VL3 (SEQ ID NO 100) имели смысловую ориентацию, а 12E10VL2 (SEQ ID NO 99) и 12E10VL4 (SEQ ID NO 101) имели антисмысловую ориентацию, соответственно. Каждый из синтезированных олигонуклеотидов состыковывали на основе взаимной комплементарности и смешивали с внешними праймерами (12E10VLS и 12E10VLA) для амплификации полноразмерного гена. 12E10VLS (SEQ ID NO 102) конструировали так, чтобы он как прямой праймер гибридизовался с 5'-концом лидерной последовательности и содержал сайт узнавания рестриктазы EcoRI и последовательность Козака. 12E10VLA (SEQ ID NO 103) конструировали так, чтобы он как обратный праймер гибридизовался с нуклеотидной последовательностью, кодирующей С-конец V-области L-цепи, и содержал донорный сайт сплайсинга и сайт узнавания рестриктазы BlnI.
После проведения ПЦР, как описано выше, ПЦР-продукт очищали с использованием 1,5% геля легкоплавкой агарозы (Sigma), расщепляли рестриктазами EcoRI и BlnI и клонировали в вектор pUC19, содержащий ген для константной области лямбда-цепи человека. После определения ДНК-последовательности плазмиду, содержащую правильную ДНК-последовательность, расщепляли EcoRI для получения гена, кодирующего V-область L-цепи 12Е10 и константную область лямбда-цепи человека, и затем встраивали в экспрессионный вектор pCOS1. Плазмида, содержащая ген L-цепи 12Е10 (SEQ ID NO 104), была обозначена как pCOS-12E10L.
8.3 Получение реконструированного одноцепочечного Fv 12E10
Реконструированный одноцепочечный Fv антитела 12E10 конструировали так, чтобы его структура была представлена 12Е10VН-линкер-12Е10VL и содержала FLAG-последовательность (SEQ ID NO 105) на С-конце для облегчения детекции и очистки. Реконструированные одноцепочечные Fv 12E10 (sc 12E10 и db 12E10) конструировали с использованием линкерных последовательностей, состоящих из 15 аминокислот, представленных (Gly4Ser)3, или 5 аминокислот, представленных (Gly4Ser)1.
(1) Получение реконструированного одноцепочечного Fv 12E10 с использованием линкерной последовательности, состоящей из 5 аминокислот
Ген, кодирующий реконструированный одноцепочечный Fv 12E10, который содержал линкерную последовательность, состоящую из 5 аминокислот, конструировали введением нуклеотидной последовательности для этого линкера (Gly4Ser)1 с 3'-конца гена, кодирующего V-область Н-цепи 12Е10, и с 5'-конца гена, кодирующего V-область L-цепи 12E10, амплификацией полученного таким образом гена при помощи ПЦР и состыковкой амплифицированных генов. Для получения реконструированного одноцепочечного Fv 12Е10 использовали четыре праймера (A-D). Праймеры А и С имели смысловые последовательности, а праймеры В и D имели антисмысловые последовательности.
Прямым праймером для V-области Н-цепи был 12E10S (Праймер A, SEQ ID NO 106). Обратный праймер DB2 (Праймер В, SEQ ID NO 107) для V-области Н-цепи конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области Н-цепи, и содержал нуклеотидную последовательность, кодирующую линкер (Gly4Ser)1, и нуклеотидную последовательность, кодирующую N-конец V-области L-цепи.
Прямой праймер DB1 (Праймер С, SEQ ID NO 108) для V-области L-цепи конструировали так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области L-цепи, и содержал нуклеотидную последовательность, кодирующую линкер (Gly4Ser)1, и нуклеотидную последовательность, кодирующую С-конец V-области Н-цепи. Обратный праймер 12E10FA для V-области L-цепи (Праймер D, SEQ ID NO 109) конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал последовательность, кодирующую FLAG, и сайт узнавания рестриктазы NotI.
На первой стадии ПЦР проводили две реакции А-В и C-D, и два ПЦР-продукта, полученные на первой стадии ПЦР, состыковывали на основе взаимной комплементарности. После добавления праймеров А и D амплифицировали полноразмерную ДНК, кодирующую реконструированный одноцепочечный Fv 12E10, содержащий линкер, состоящий из 5 аминокислот (вторая ПЦР). В ПЦР первой стадии в качестве матрицы использовали плазмиды HEF-12E10H-gγ1 (см. пример 8.1), кодирующую реконструированную V-область Н-цепи 12E10, и pCOS-12E10L (см. пример 8.2), кодирующую реконструированную V-область L-цепи 12E10, соответственно.
50 мкл ПЦР-раствора для первой стадии содержали 5 мкл 10-кратного ПЦР-буфера Gold II, 1,5 мМ MgCl2, 0,08 мМ dNTP, 5 единиц ДНК-полимеразы AmpliTaq Gold (все от Perkin Elmer), no 100 пмоль каждого праймера и 100 нг каждой ДНК-матрицы. ПЦР-раствор нагревали при начальной температуре 94°С в течение 9 минут, затем при 94°С в течение 30 секунд, при 55°С в течение 30 секунд и при 72°С в течение 1 минуты. После повторения этого цикла 35 раз реакционную смесь нагревали при 72°С дополнительно в течение 5 минут.
ПЦР-продукты А-В (429 п.о.) и C-D (395 п.н.) состыковывали при помощи второй ПЦР. Реакцию в ПЦР-растворе для второй стадии (98 мкл), содержавшем в качестве матрицы 1 мкл ПЦР-продукта первой стадии А-В и C-D, 100 пмоль каждого из праймеров, 10 мкл 10-кратного ПЦР-буфера Gold II, 1,5 мМ MgCl2, 0,08 мМ dNTP и 5 единиц ДНК-полимеразы AmpliTaq Gold (все от Perkin Elmer), проводили в тех же самых условиях, которые описаны выше.
ДНК-фрагмент 795 п.о., полученный во второй ПЦР, очищали с использованием 1,5% геля легкоплавкой агарозы, расщепляли EcoRI и NotI и клонировали в вектор рСНО1 или вектор pCOS1. Экспрессионный вектор pCHO1 представлял собой вектор, сконструированный путем делеции гена антитела из DHFR-ΔE-RVH-PM1-f (см. WO 92/19759) в результате расщепления EcoRI и SmaI и присоединения к EcoRI-NotI-BamHI-адаптору (Takara Shuzo). После определения ДНК-последовательности плазмиды, содержащие ДНК-фрагмент, кодирующий правильную аминокислотную последовательность реконструированного одноцепочечного Fv 12E10, были обозначены как pCHO-db 12E10 и pCOS-db 12E10. Нуклеотидная последовательность и аминокислотная последовательность реконструированного одноцепочечного Fv антитела 12E10, включенного в плазмиды pCHO-db12E10 и pCOS-db12E10, показаны в SEQ ID NO 110.
(2) Получение реконструированного одноцепочечного Fv 12E10 с использованием линкерной последовательности, состоящей из 15 аминокислот
Ген, кодирующий реконструированный одноцепочечный Fv антитела 12Е10, который содержал линкерную последовательность, состоящую из 15 аминокислот, конструировали путем введения нуклеотидной последовательности для линкера (Gly4Ser)3 с 3'-конца гена, кодирующего V-область Н-цепи 12Е10, и с 5'-конца гена, кодирующего V-область L-цепи 12Е10, амплификации полученного таким образом гена при помощи ПЦР и состыковки амплифицированных генов. Для получения этого реконструированного одноцепочечного Fv 12E10 использовали четыре праймера (A-D). Праймеры А и С имели смысловые последовательности, а праймеры В и D имели антисмысловые последовательности.
Прямым праймером для V-области Н-цепи был 12E10S (Праймер A, SEQ ID NO 106). Обратный праймер sc4.3 (Праймер В, SEQ ID NO 111) для V-области Н-цепи конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области Н-цепи, и содержал нуклеотидную последовательность, кодирующую линкер (Gly4SeR)3, и нуклеотидную последовательность, кодирующую N-конец V-области L-цепи.
Прямой праймер scl.3 (Праймер С, SEQ ID NO 112) для V-области L-цепи конструировали так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области L-цепи, и содержал нуклеотидную последовательность, кодирующую линкер (Gly4Ser)3, и нуклеотидную последовательность, кодирующую С-конец V-области Н-цепи. Обратный праймер 12E10FA для V-области L-цепи (Праймер D, SEQ ID NO 109) конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал последовательность, кодирующую FLAG, и сайт узнавания рестриктазы NotI.
На первой стадии ПЦР проводили две реакции А-В и C-D, и два ПЦР-продукта, полученные на первой стадии ПЦР, состыковывали на основе взаимной комплементарности. После добавления праймеров А и D амплифицировали полноразмерную ДНК, кодирующую реконструированный одноцепочечный Fv 12E10, содержащий линкер, состоящий из 15 аминокислот (вторая ПЦР). В ПЦР первой стадии в качестве матрицы использовали плазмиду pCOS-db12E10 (см. пример 8. 3(1)), кодирующую реконструированный одноцепочечный Fv 12E10.
50 мкг ПЦР-раствора для первой стадии ПЦР содержали 5 мкл 10-кратного ExTaq-буфера II, 0,4 мМ dNTP, 2,5 единиц ДНК-полимеразы TaKaRa ExTaq (TAKARA), по 100 пмоль каждого праймера и 10 нг каждой ДНК-матрицы. ПЦР-раствор нагревали при начальной температуре 94°С в течение 30 секунд, затем при 94°С в течение 15 секунд и 72°С в течение 2 минут и этот цикл повторяли 5 раз. После повторения 28 раз цикла нагревания при 94°С в течение 15 секунд и при 70°С в течение 2 минут реакционную смесь дополнительно нагревали при 72°С в течение 5 минут.
ПЦР-продукты А-В (477 п.о.) и C-D (447 п.о.) состыковывали при помощи второй ПЦР. В ПЦР-смеси для второй стадии (98 мкл), содержавшей в качестве матрицы 1 мкл ПЦР-продуктов первой стадии А-В и C-D, 100 пмоль каждого из праймеров А и D, 5 мкл 10-кратного ExTaq-буфера, 0,4 мМ dNTP, 2,5 единиц ДНК-полимеразы TaKaRa ExTaq (TAKARA), реакцию проводили при тех же самых условиях, которые описаны выше.
ДНК-фрагмент 825 п.о., полученный во второй ПЦР, очищали с использованием 1,0% геля легкоплавкой агарозы, расщепляли EcoRI и NotI. Полученный таким образом ДНК-фрагмент клонировали в вектор рСНО1 или вектор pCOS1. После определения ДНК-последовательности плазмиды, содержащие ДНК-фрагмент, кодирующий правильную аминокислотную последовательность реконструированного одноцепочечного Fv 12E10, были обозначены как pCHO-sc12E10 и pCOS-sc12E10. Нуклеотидная последовательность и аминокислотная последовательность этого реконструированного одноцепочечного Fv антитела 12E10, включенного в плазмиды pCHO-sc12E10 и pCOS-sc12E10, показаны в SEQ ID NO 113.
8.4 Экспрессия антитела 12E10 (IgG, Fab) и одноцепочечного полипептида Fv клеткой животного
Антитело 12Е10 (IgG, Fab) и одноцепочечный Fv, происхоящий из антитела 12Е10 (линкерная последовательность 5 аминокислот, 15 аминокислот), экспрессировали с использованием клеток COS7 или клеток СНО.
Временную экспрессию с использованием клеток COS7 выполняли следующим образом. Трансфекцию проводили способом электропорации с применением устройства Gene Pulser (BioRad). Для экспрессии антитела 12Е10 (IgG) добавляли по 10 мкг вышеупомянутого экспрессионного вектора HEF-12E10H-gγ1 и pCOS-12E10L, для экспрессии Fab-фрагмента 12Е10 добавляли по 10 мкг pFd-12E10H и pCOS-12E10L и для экспрессии одноцепочечного Fv 10 мкг pCOS-sc12E10 или 10 мкг pCOS-db12E10 добавляли к клеткам COS7 (1×107 клеток/мл), суспендированным в 0,8 мл PBS. Смесь, находящуюся в кювете, обрабатывали импульсом 1,5 кВ, при электрической емкости 25 мкФ. После "восстановления" в течение 10 минут при комнатной температуре электропорированные клетки добавляли в культуральную среду DMEM (Gibco BRL), содержащую 10% фетальную телячью сыворотку, и культивировали. После культивирования в течение ночи клетки промывали один раз PBS, добавляли в бессывороточную среду CHO-S-SFM II (Gibco BRL) и культивировали в течение 3 дней. Культуральную среду центрифугировали для удаления клеточного дебриса и фильтровали через фильтр с размером пор 0,22 мкм.
Для получения клеточной линии СНО, стабильно экспрессирующей одноцепочечный Fv (полипептид), происходящий из антитела 12Е10, экспрессионные вектора pCHO-sc12E10 или pCOS-ds12E10 вводили в клетки СНО, соответственно.
Каждый экспрессионный вектор вводили в клетки СНО электропорацией с использованием прибора Gene Pulser (BioRad). Линеаризованную ДНК (100 мкг), полученную расщеплением рестриктазой Pvul, и клетки СНО (1×107 клеток/мл), суспендированные в 0,8 мл PBS, смешивали в кювете, оставляли без перемешивания на льду на 10 минут и обрабатывали импульсом 1,5 кВ, при электрической емкости 25 мкФ. После "восстановления" в течение 10 минут при комнатной температуре электропорированные клетки добавляли в среду CHO-S-SFM II (Gibco BRL), содержащую 10% диализованную фетальную телячью сыворотку и нуклеиновые кислоты, и культивировали. После культивирования в течение 2 дней культивирование продолжали в свободной от нуклеиновых кислот среде CHO-S-SFM II (Gibco BRL), содержащей 10% диализованную фетальную телячью сыворотку. Из полученных таким образом клонов отбирали клон с высоким уровнем экспрессии в качестве клеточной линии-продуцента одноцепочечного Fv 12E10. После культивирования в бессывороточной среде CHO-S-SFM II (Gibco BRL), культуральный супернатант центрифугировали для отделения клеточного дебриса (остатков клеток) и фильтровали через фильтр с размером пор 0,22 мкм.
8.5 Очистка одноцепочечного Fv, происходящего из 12E10, продуцируемого клетками СНО
Культуральные супернатанты, продуцируемые клеточными линиями СНО, экспрессирующими одноцепочечный Fv 12E10 (sc 12E10, db 12E10), полученные в примере 8.4, очищали с использованием колонки с анти-FLAG-антителом и гель-фильтрационной колонки, соответственно, для получения очищенных одноцепочечных Fv.
(1) Очистка с использованием колонки с анти-FLAG-антителом
Каждый культуральный супернатант (sc 12E10, db 12E10) наносили на колонку с анти-FLAG М2-аффинным гелем (Sigma), уравновешенным 50 мМ Трис-HCl-буфером (рН 7,4), содержащим 150 мМ NaCl. После промывания этой колонки тем же самым буфером белки, адсорбированные на колонке, элюировали 100 мМ глицин-HCl-буфером (рН 3,5). Элюированные фракции незамедлительно нейтрализовали добавлением 1 М Трис-HCl-буфера (рН 8,0) и анализировали электрофорезом в ДСН-ПААГ. Фракции, которые, как было подтверждено, содержали одноцепочечный Fv, объединяли и концентрировали приблизительно в 20 раз с использованием Centricon-10 (Millipore).
(2) Гель-фильтрация
Концентрированный раствор, полученный в (1), добавляли на колонку Superdex 200 HR (10×300 мм, Amersham Pharmacia), уравновешенную PBS, содержащим 0,01% Твин 20. Хроматограммы представлены на фиг.53 и 54. Продукт sc12E10 элюировался в виде двух пиков (А, В) (см. фиг.53). Продукт db12E10 элюировался в виде двух пиков (С, D) (см. фиг.54). Фракцию каждого пика собирали, обрабатывали в присутствии и в отсутствие восстанавливающего агента, подвергали электрофорезу согласно способу Лэммли и окрашивали Кумасси бриллиантовым синим после электрофореза. Как показано на фиг.55, все фракции А, В, С и D, независимо от присутствия или отсутствия восстанавливающего агента, давали единственную полосу, имеющую кажущуюся молекулярную массу около 31 кДа. При анализе этих фракций гель-фильтрацией с использованием Superdex 200 HR фракция А давала продукт, элюируемый с кажущейся молекулярной массой около 42 кДа, фракция В - 20 кДа, фракция С - 69 кДа и фракция D - 41 кДа (см. фиг.57). Эти результаты предполагают, что происходящая из sc12E10 фракция А является нековалентно связанным димером одноцепочечного Fv sc12E10 и фракция В является мономером одноцепочечного Fv, а происходящая из db12E10 фракция С является нековалентно связанным тримером одноцепочечного Fv и D является нековалентно связанным димером одноцепочечного Fv.
8.6 Измерение ТРО-подобной агонистической активности различных одноцепочечных Fv
ТРО-подобную активность одноцепочечного анти-MPL-антитела оценивали измерением пролиферативной активности в отношении клеток Ba/F3 (BaF/mpl), экспрессирующих ТРО-рецептор человека (MPL).
После промывания клеток BaF/mpI два раза культуральной средой RPMI1640 (Gibco), содержащей 1% фетальную телячью сыворотку (Gibco), эти клетки суспендировали в среде при плотности клеток 5×105 клеток/мл. Одноцепочечное анти-MPL-антитело и ТРО человека (R & D Systems) разбавляли этой культуральной средой, соответственно. 50 мкл клеточной суспензии и 50 мкл разведенного антитела или ТРО человека добавляли в 96-луночный микропланшет (плоскодонный) (Corning) и культивировали в СО2-инкубаторе (концентрация СО2: 5%) в течение 24 часов. После инкубирования добавляли 10 мкл реагента WST-8 (реагента для измерения числа живых клеток SF: Nacalai Tesque) и поглощение немедленно измеряли при длине волны измерения 450 нм и при опорной длине волны 655 нм с использованием флуоресцентно-абсорбционного фотометра Benchmark Plus (BioRad). После инкубирования в CO2-инкубаторе (концентрация СО2: 5%) в течение 2 часов вновь измеряли оптическую плотность при длине волны измерения 450 нм и опорной длине волны 655 нм при помощи Benchmark Plus. Поскольку реагент WST-8 развивал окраску при длине волны 450 нм в зависимости от числа живых клеток, пролиферативную активность BaF/rnpl оценивали по изменению оптической плотности за 2 часа.
Агонистическая активность в отношении MPL, измеренная с использованием культуральных супернатантов клеток COS-7, экспрессирующих различные варианты молекул антитела 12Е10, показана на фиг, 58. Одноцепочечные Fv, имеющие линкер из 5 аминокислот (ds12E10) и линкер из 15 аминокислот (sc12E10), увеличивали оптическую плотность зависимым от концентрации образом, показывая ТРО-подобную агонистическую активность (ED50; 9 пМ и 51 пМ, соответственно), тогда как 12E10IgG и 12E10Fab не имели активности.
Было известно, что Н-цепь и L-цепь одноцепочечного Fv ассоциируют не только внутри молекулы, но также между молекулами с образованием мультимеров, таких как димеры. Культуральные супернатанты клеток СНО, экспрессирующих одноцепочечные Fv антитела 12Е10, подвергали гель-фильтрации и испытывали на агонистическую активность в отношении MPL. Эти результаты показаны на фиг.59. Димер, который содержался в sc12E10 в небольшом количестве, показал приблизительно в 5000 раз более сильную ТРО-подобную агонистическую активность (димер sc12E10, ED50; 1,9 пМ) в сравнении с мономером (мономер sc12E10, ED50; >10 нМ). Активность была более высокой, чем активность ТРО (ED50; 27 пМ). Димер db12E10 (ED50; 2,0 пМ) показал сильную активность, сравнимую с активностью димера sc12E10. Тример db12E10 (ED50; 7,4 пМ), который, как предполагалось, был тримером исходя из его молекулярной массы, полученной при гель-фильтьрации, показал высокую активность, которая является более низкой, чем активность димера db12E10. Эти результаты предполагают, что для активности агонистического антитела 12Е10 важным является то, что антигенсвязывающий сайт является бивалентным (димер). С учетом того факта, что 12E10-IgG не имел активности, предполагается, что важными являются и другие факторы, кроме бивалентности, такие как местоположение антигенсвязывающего сайта, расстояние или угол.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
Модифицированные антитела данного изобретения обладают агонистическим действием, способным трансдуцировать сигнал в клетки посредством образования поперечных связей между молекулой (молекулами) клеточной поверхности, и обладают преимуществами, благодаря тому, что их проницаемость в ткани и опухоли является высокой вследствие меньшего молекулярного размера в сравнении с исходной молекулой антитела (цельным IgG). Данное изобретение обеспечивает модифицированные антитела, которые имеют необыкновенно высокую агонистическую активность в сравнении с природными лигандами, такими как ТРО, и исходное антитело (цельный IgG). Даже в том случае, если исходное антитело не имеет агонистической активности, может быть получено модифицированное антитело с более высокой агонистической активностью в сравнении с природными лигандами. Это связано с тем, что модифицированные антитела имеют форму, более близкую к лиганду в сравнении с исходными антителами. Таким образом, модифицированные антитела могут быть использованы в качестве трансдуцирующих сигнал агонистов для достижения индукции апоптоза, индукции пролиферации клеток, индукции дифференцировки, индукции клеточного деления или регуляторного действия на клеточный цикл. Модификация молекулы антитела с образованием модифицированного антитела в соответствии с данным изобретением приводит к уменьшению побочных эффектов, вызываемых образованием межклеточных сшивок, и обеспечивает новые лекарственные средства, вызывающие только требуемый эффект путем сшивания молекулы (молекул) клеточной поверхности. Медицинские препараты, содержащие в качестве активного ингредиента модифицированное антитело данного изобретения, применимы в качестве профилактических средств и/или лечебных средств при раках, воспалениях, гормональных нарушениях, аутоиммунных заболеваниях и заболеваниях крови, например, лейкозе, злокачественной лимфоме, апластической анемии, синдроме миелоплазии и красной, или истинной, полицитемии.

















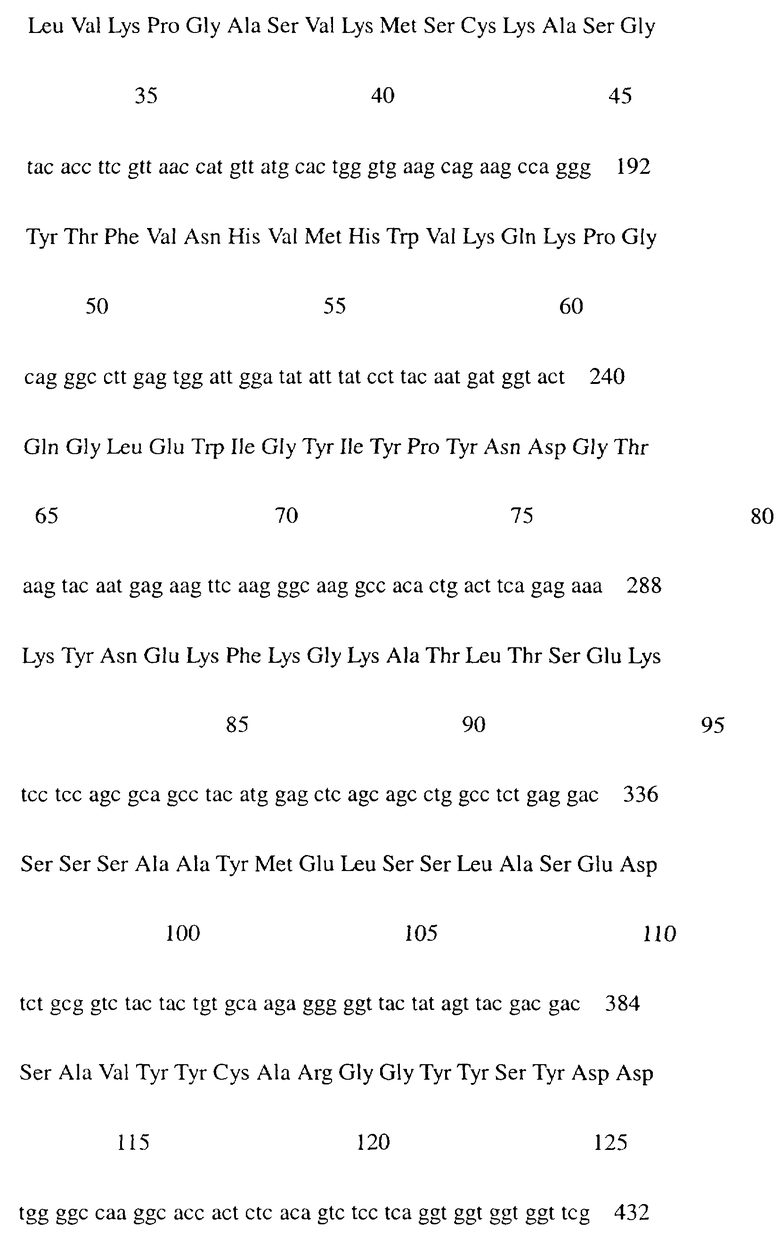





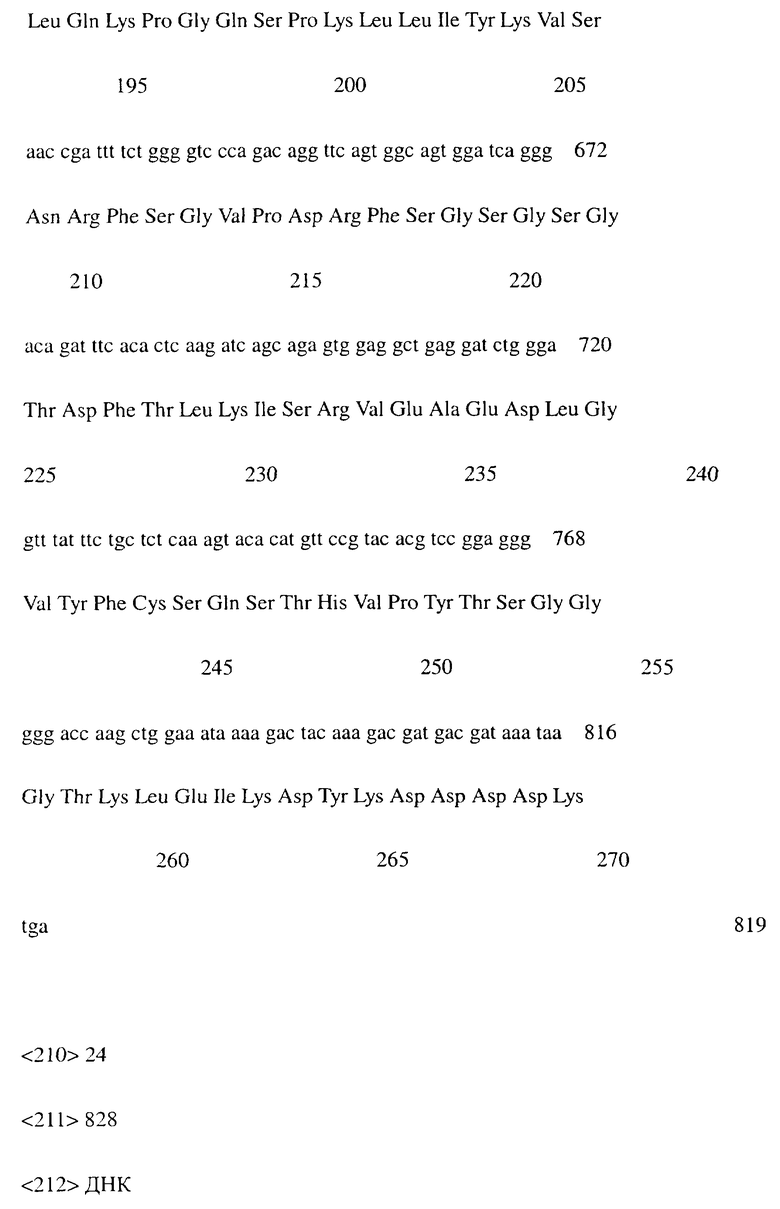























































































| название | год | авторы | номер документа |
|---|---|---|---|
| ДЕГРАДИРОВАННОЕ АНТИТЕЛО, ЯВЛЯЮЩЕЕСЯ АГОНИСТОМ TPO | 2001 |
|
RU2287534C2 |
| СПОСОБ И НАБОР ДЛЯ ВЫЯВЛЕНИЯ ГЕНА, КОДИРУЮЩЕГО МЕМБРАНОСВЯЗАННЫЙ БЕЛОК, ВЕКТОР (ВАРИАНТЫ) | 1999 |
|
RU2246538C2 |
| ОПУХОЛЕСПЕЦИФИЧНОЕ АНТИТЕЛО ПРОТИВ EGFR И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2730605C2 |
| Рекомбинантный Fab-scFv на основе нейтрализующего антитела против интерферона бета-1а человека и антитела против рецептора ErbB2 человека | 2019 |
|
RU2748953C1 |
| МОНОМОЛЕКУЛЯРНЫЙ ХИМЕРНЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР К РАКОВОМУ АНТИГЕНУ СА125 | 2018 |
|
RU2747095C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ МОДИФИКАЦИИ ЗАДАННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВОЙ КИСЛОТЫ-МИШЕНИ | 2012 |
|
RU2663354C2 |
| СКОНСТРУИРОВАННЫЕ ВЫСОКОАФФИННЫЕ T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ ЧЕЛОВЕКА | 2014 |
|
RU2740648C2 |
| АНТИ-ГЛИПИКАН 3-АНТИТЕЛО, ИМЕЮЩЕЕ МОДИФИЦИРОВАННУЮ САХАРНУЮ ЦЕПЬ | 2005 |
|
RU2451030C2 |
| СПОСОБ СЕКРЕТОРНОЙ ПРОДУКЦИИ БЕЛКА | 2012 |
|
RU2588439C2 |
| АНТИТЕЛО ПРОТИВ МОЛЕКУЛЫ КЛЕТОЧНОЙ АДГЕЗИИ L1 ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И СОДЕРЖАЩИЙ ЕГО ХИМЕРНЫЙ РЕЦЕПТОР АНТИГЕНА | 2019 |
|
RU2789360C2 |
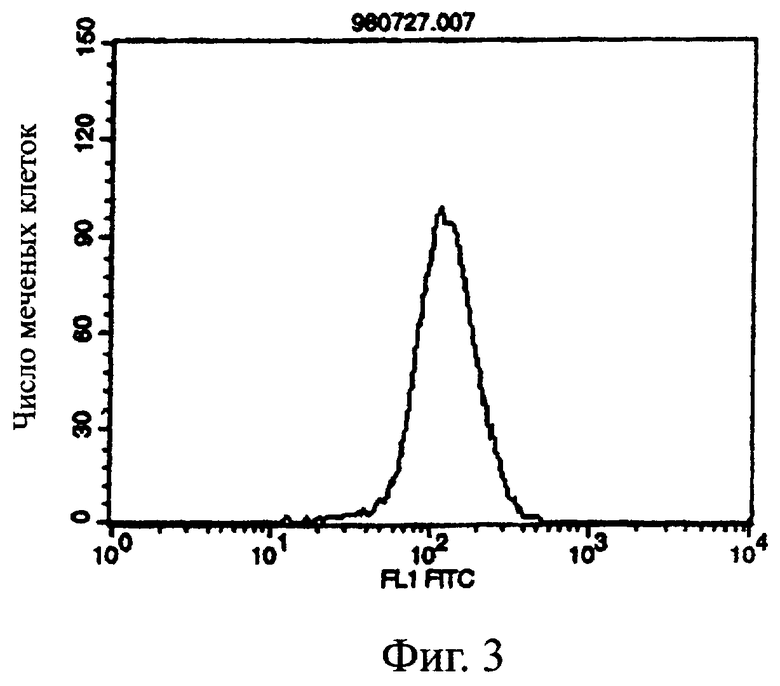
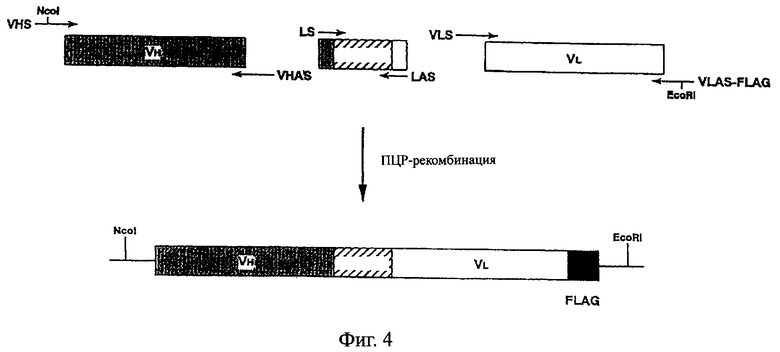
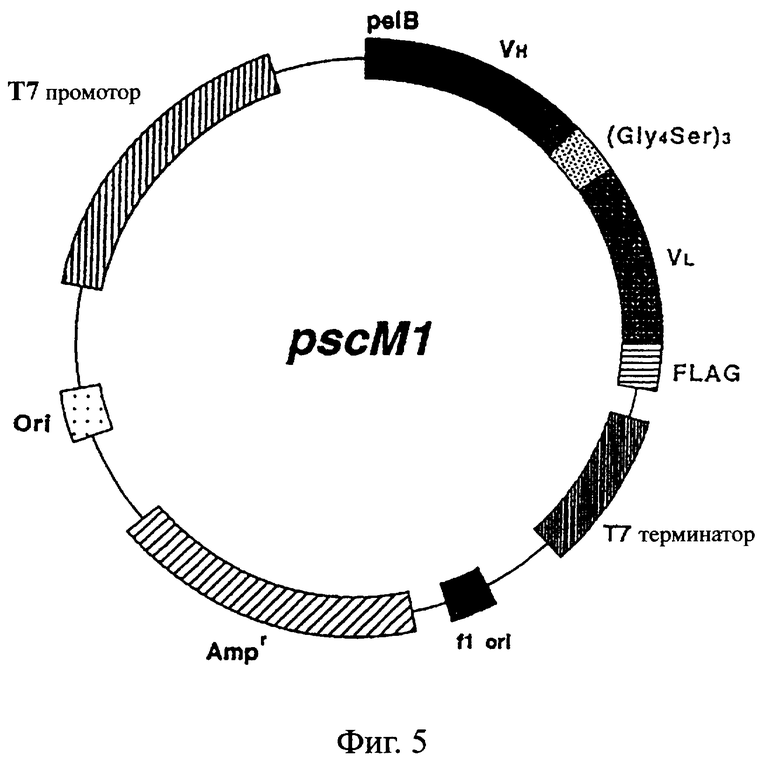
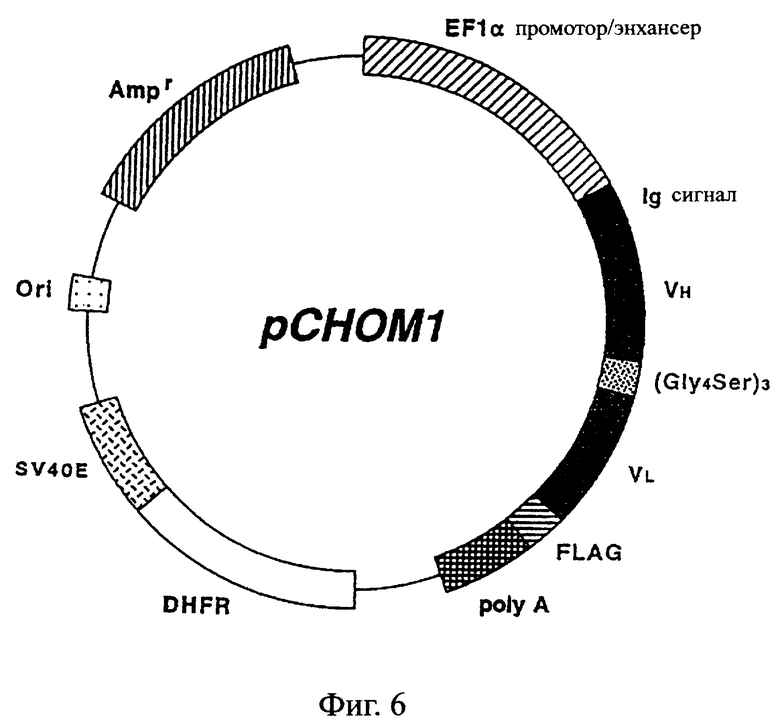
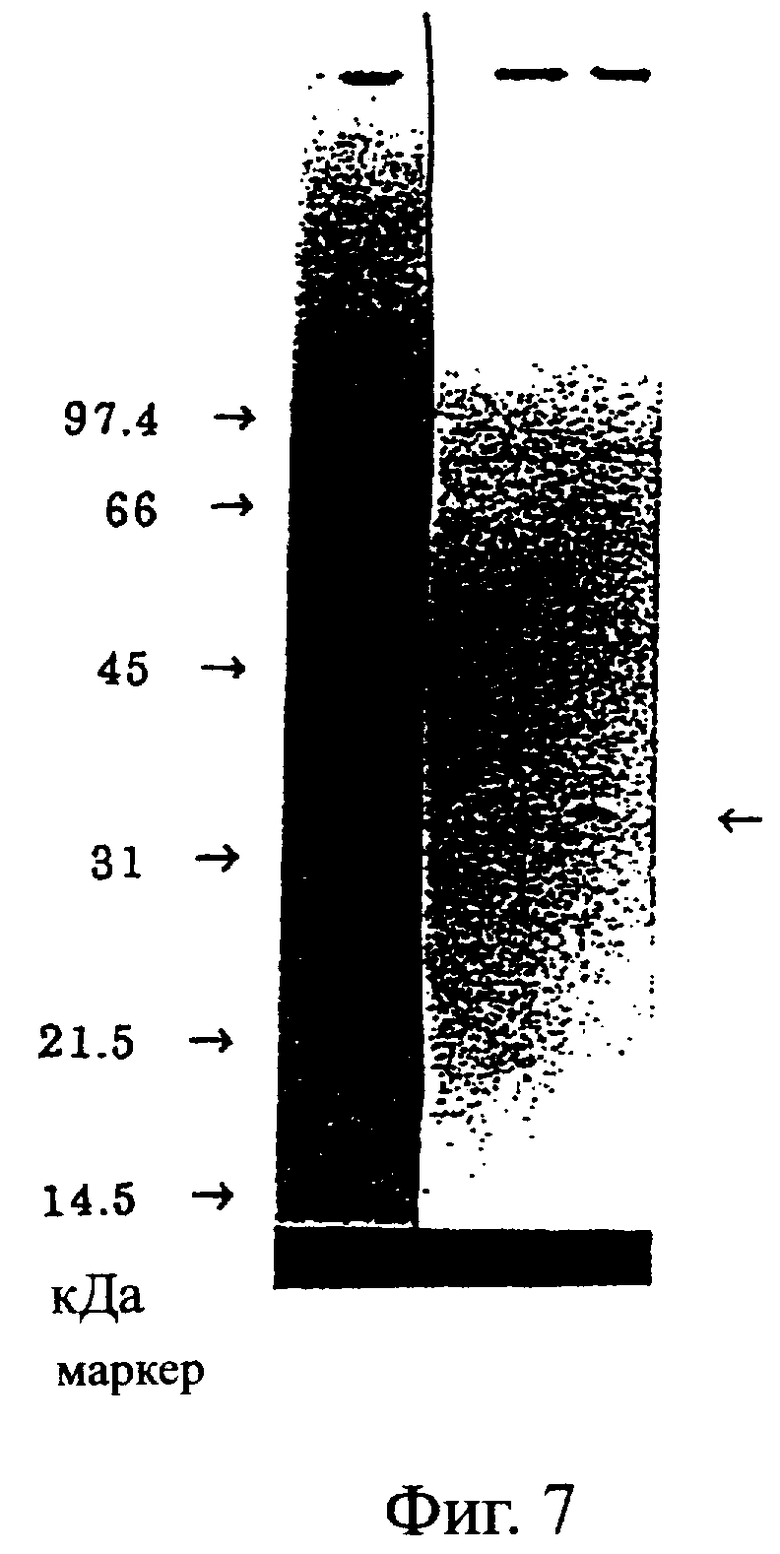
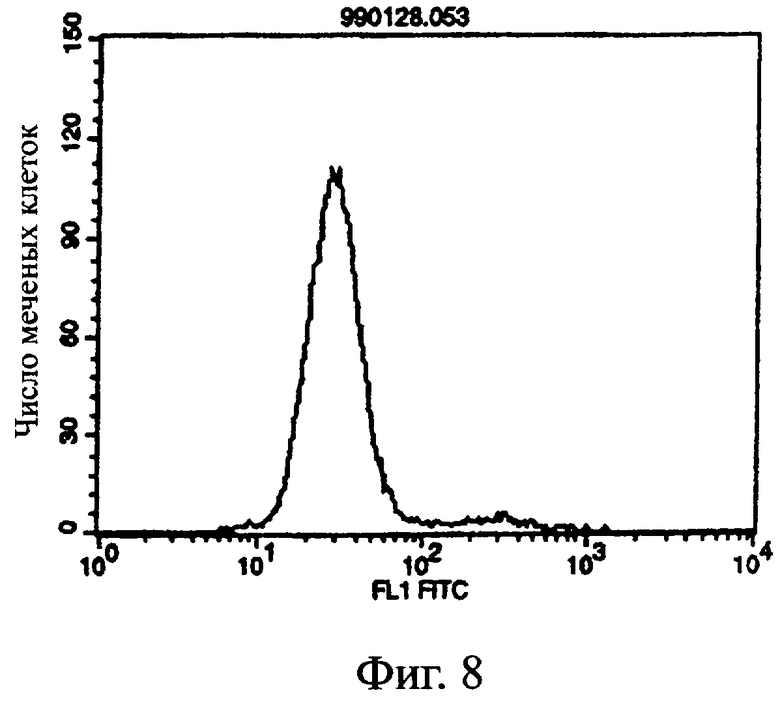
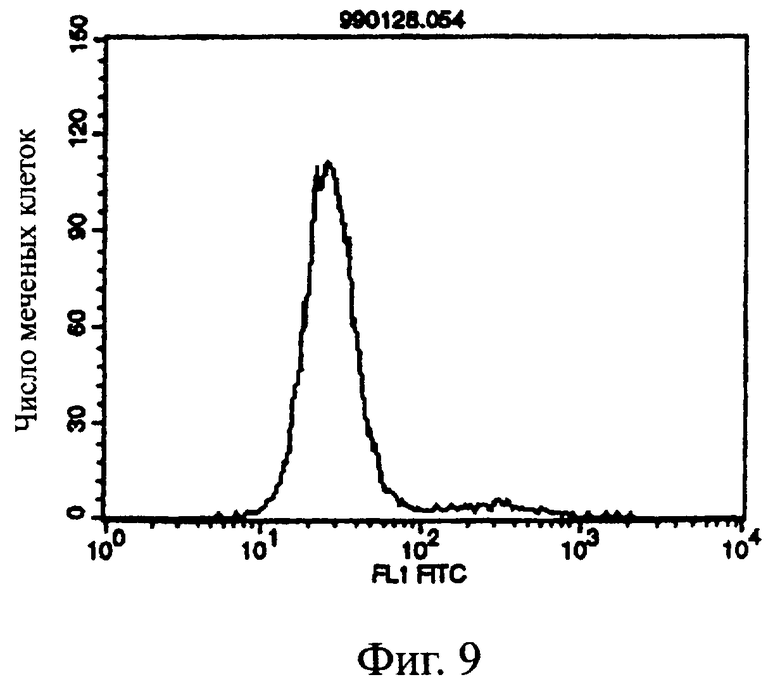
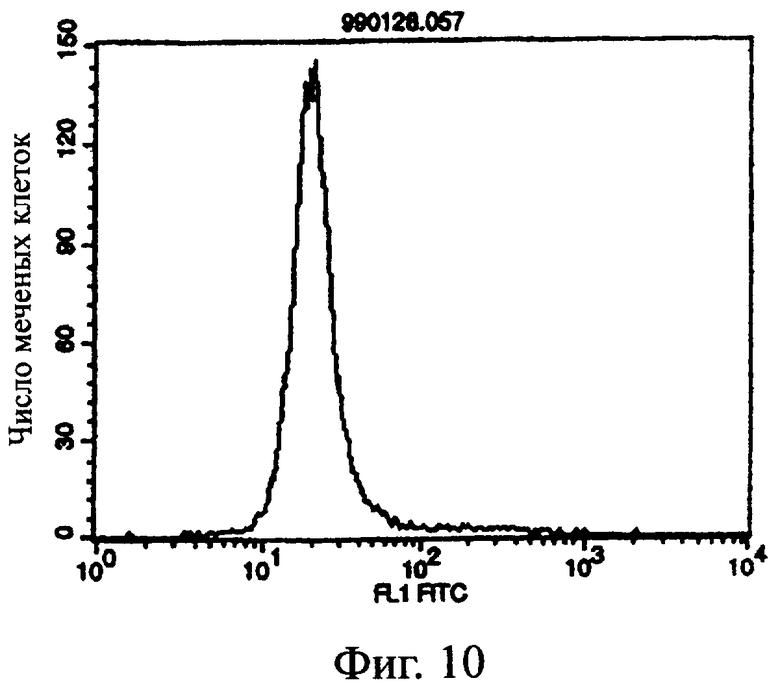
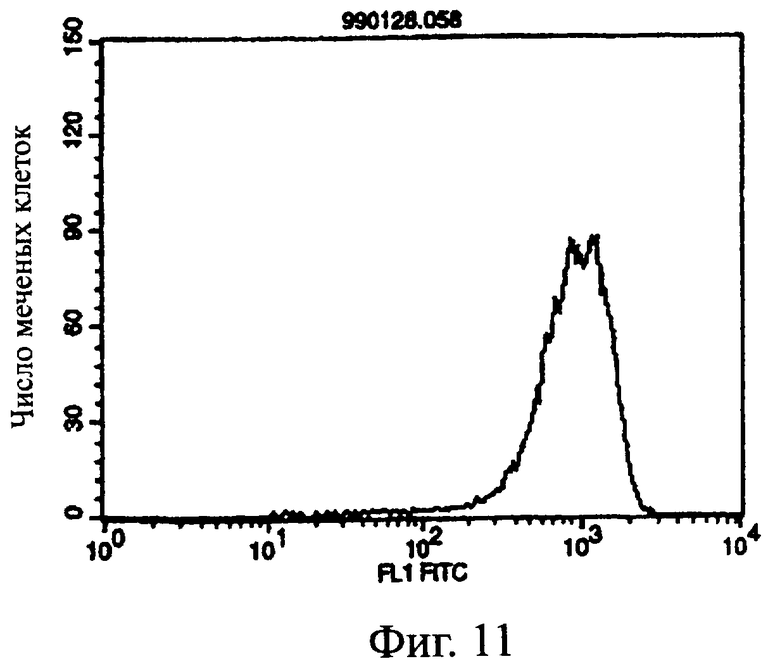
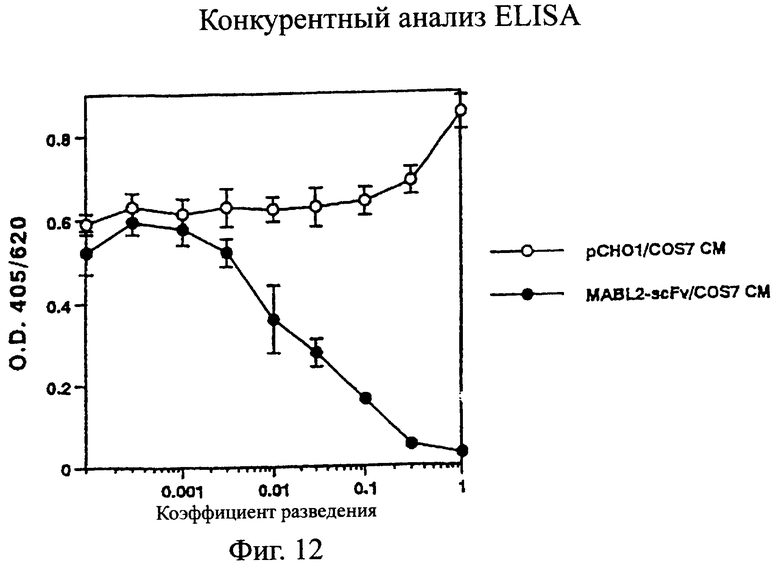
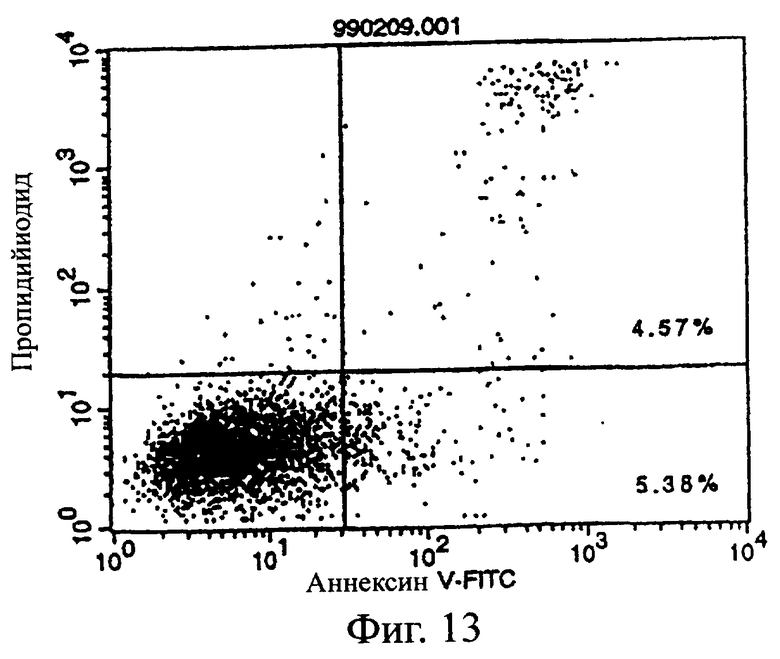
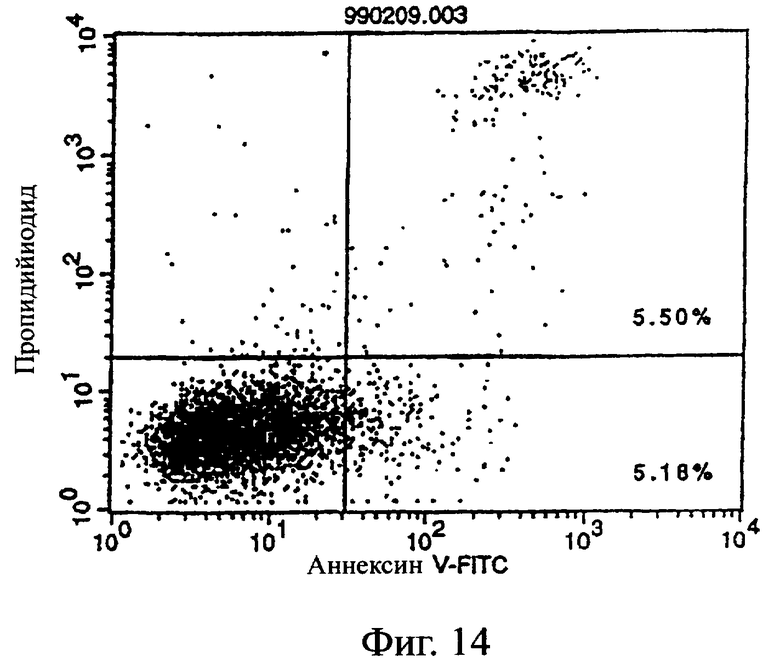
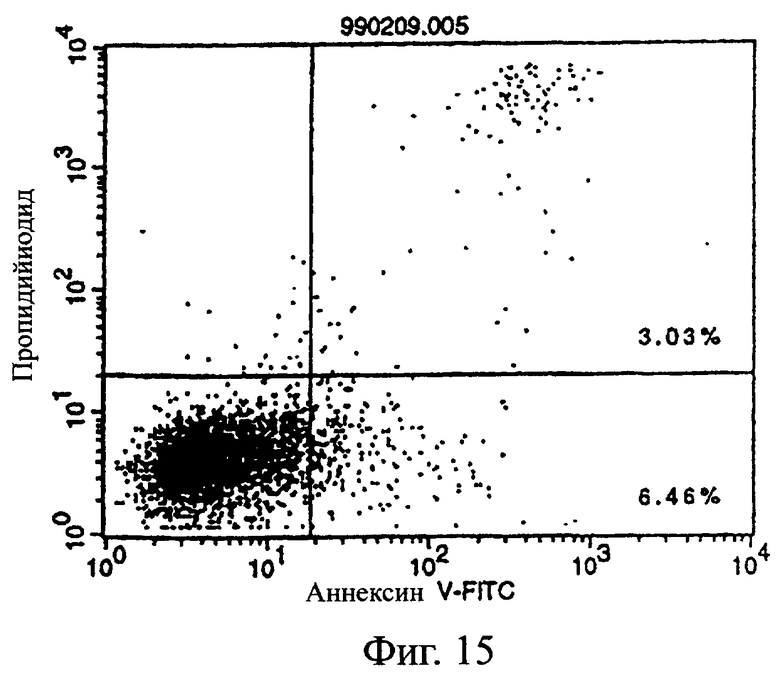
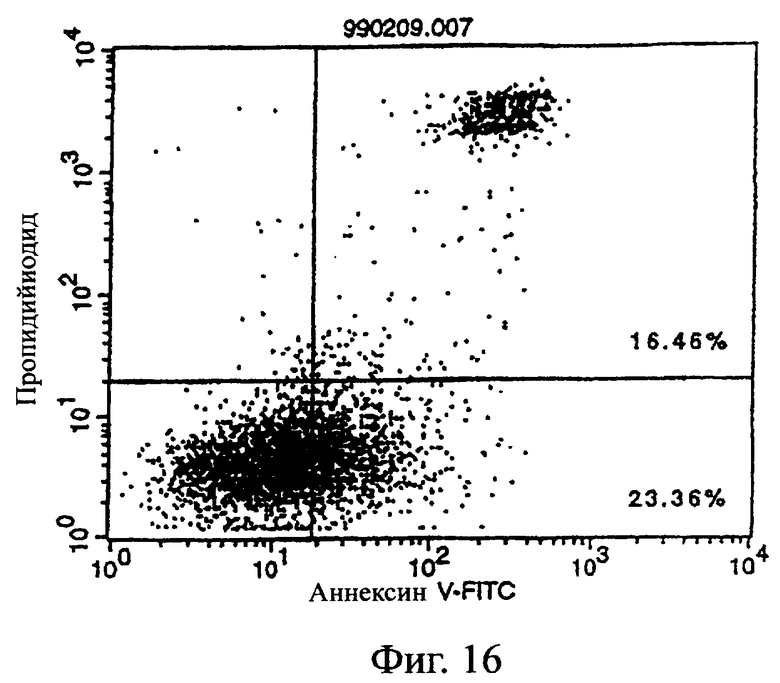
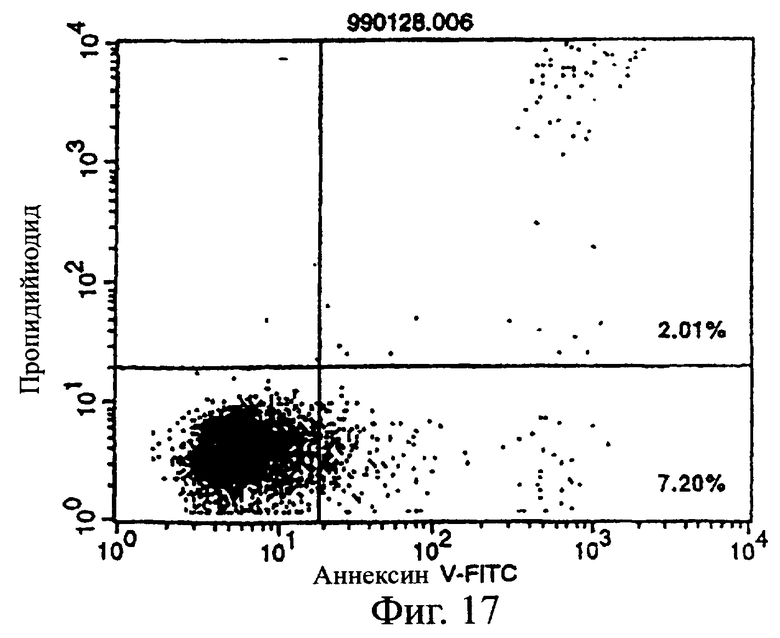
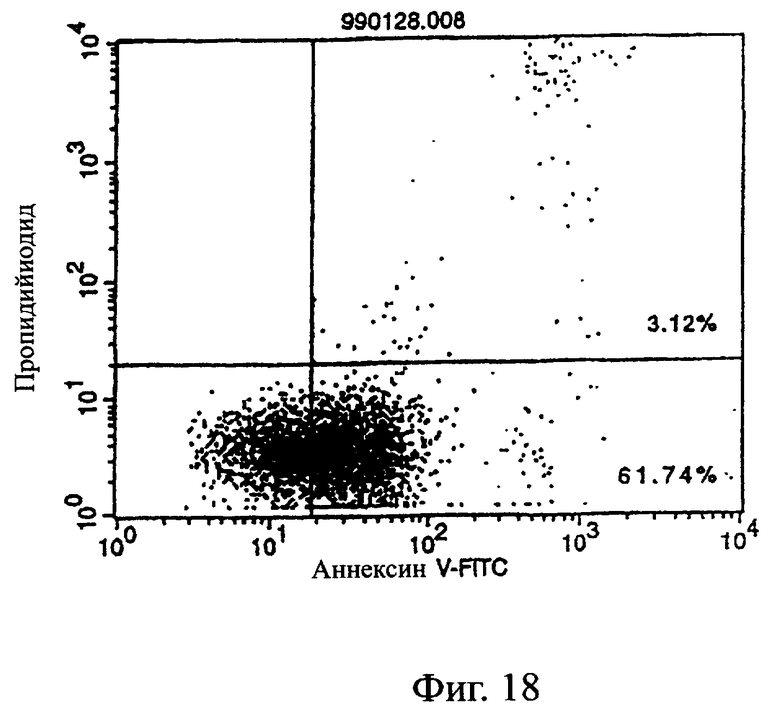
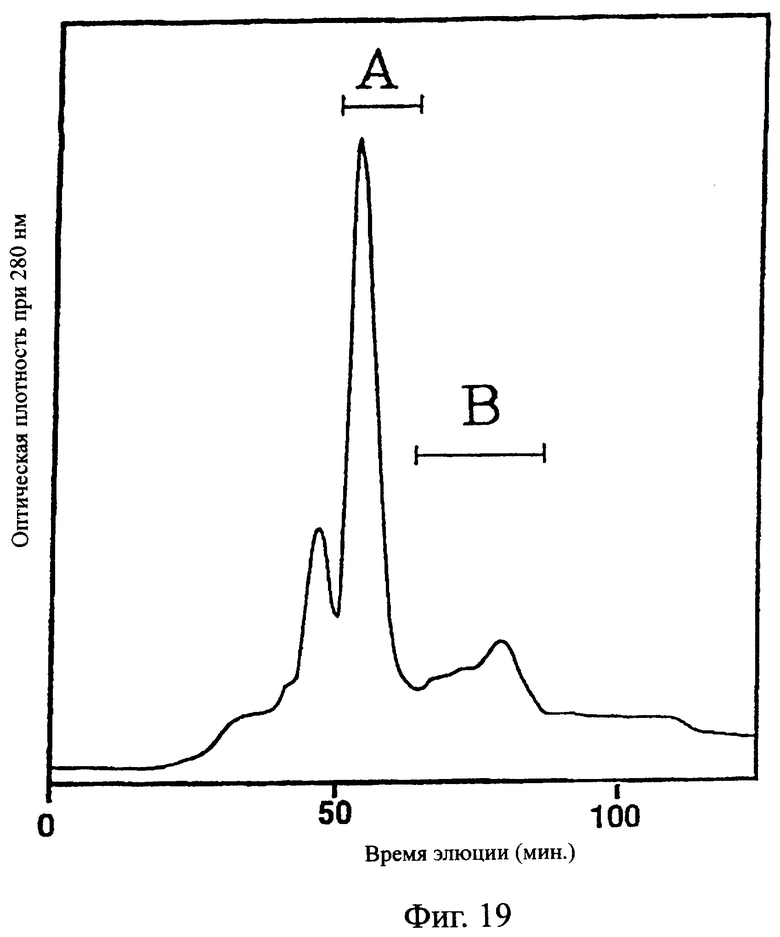
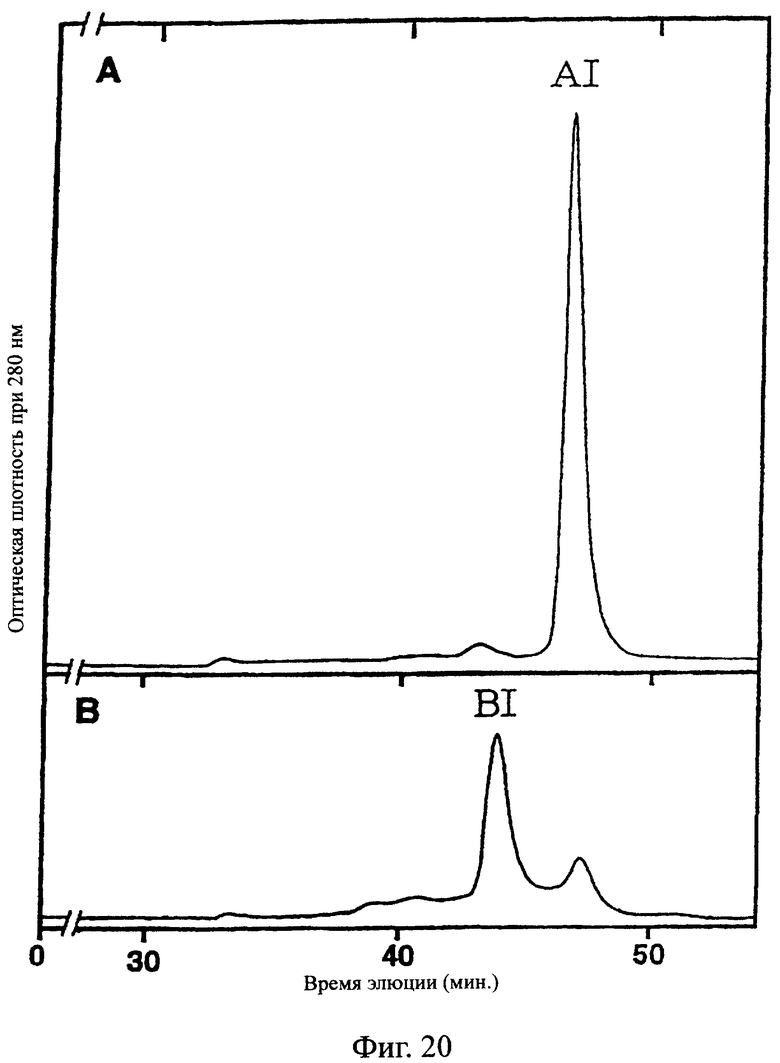
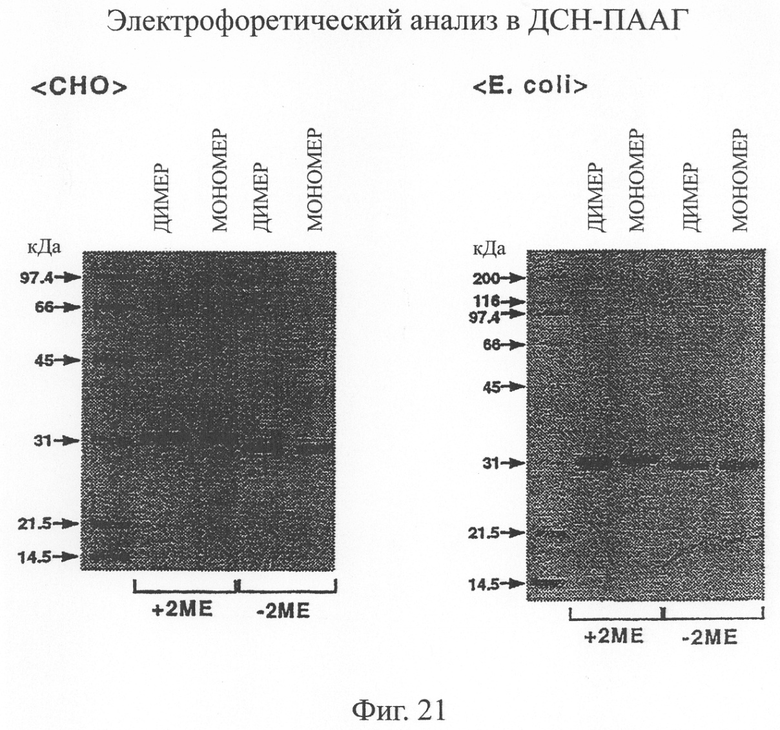
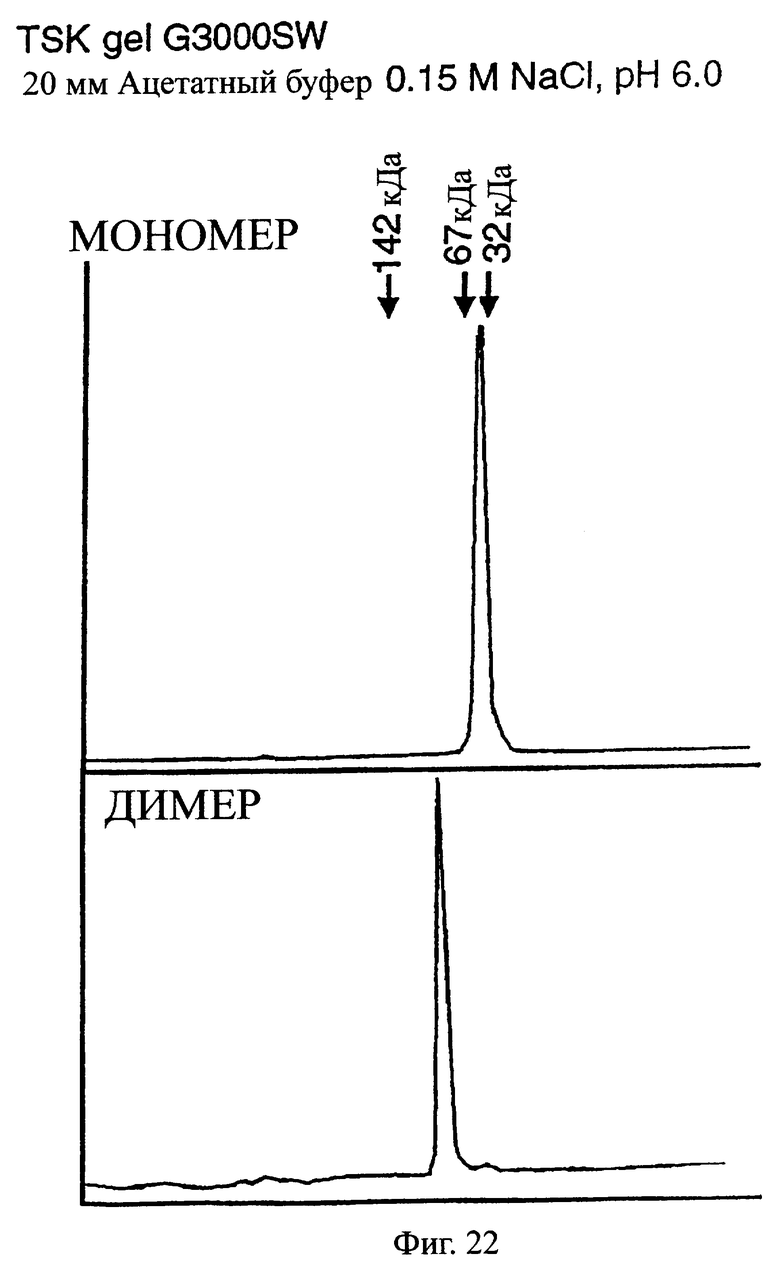
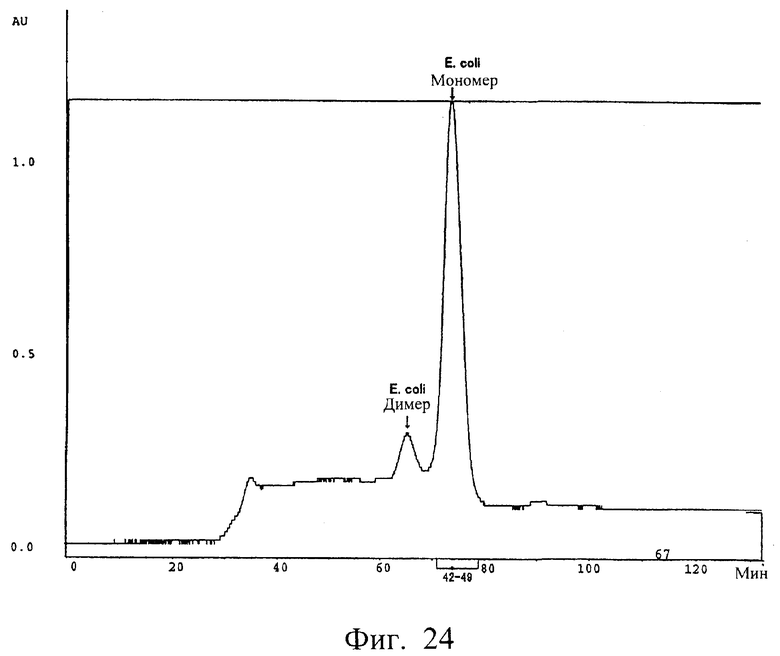
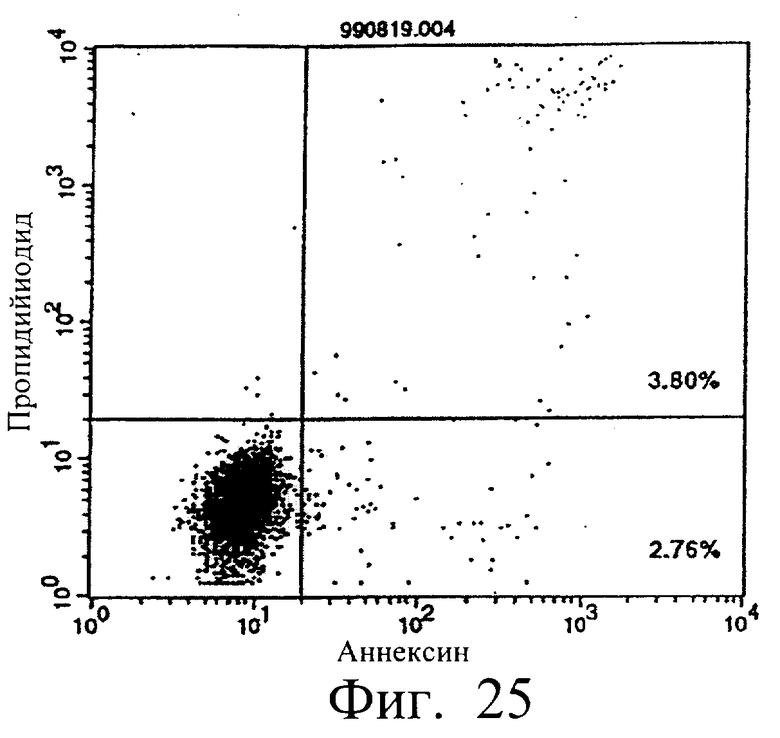
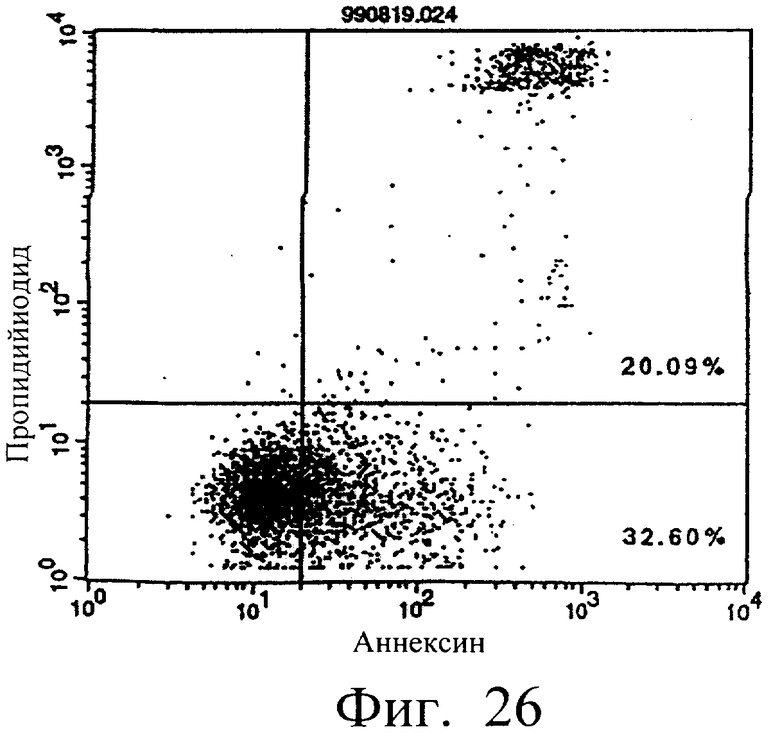
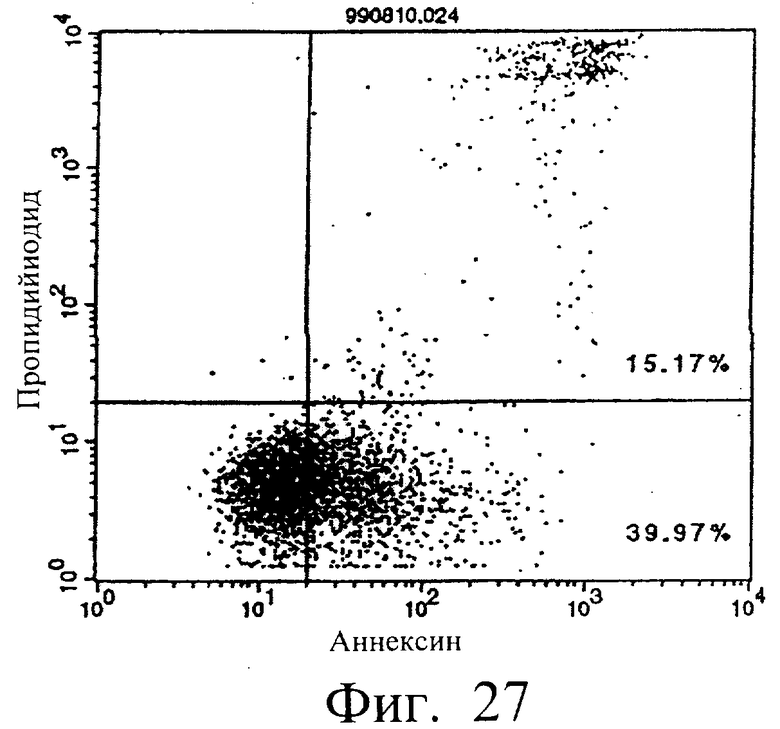
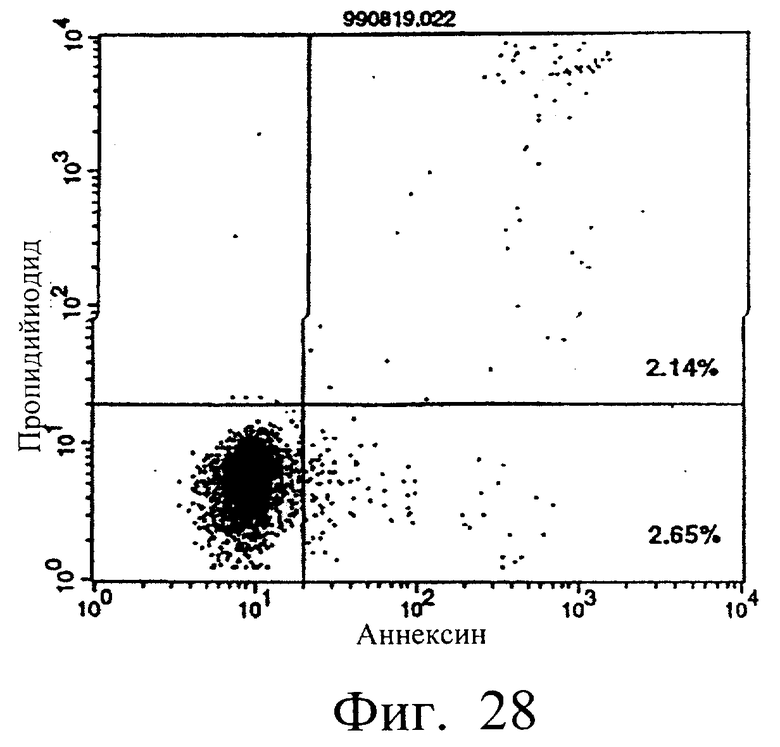
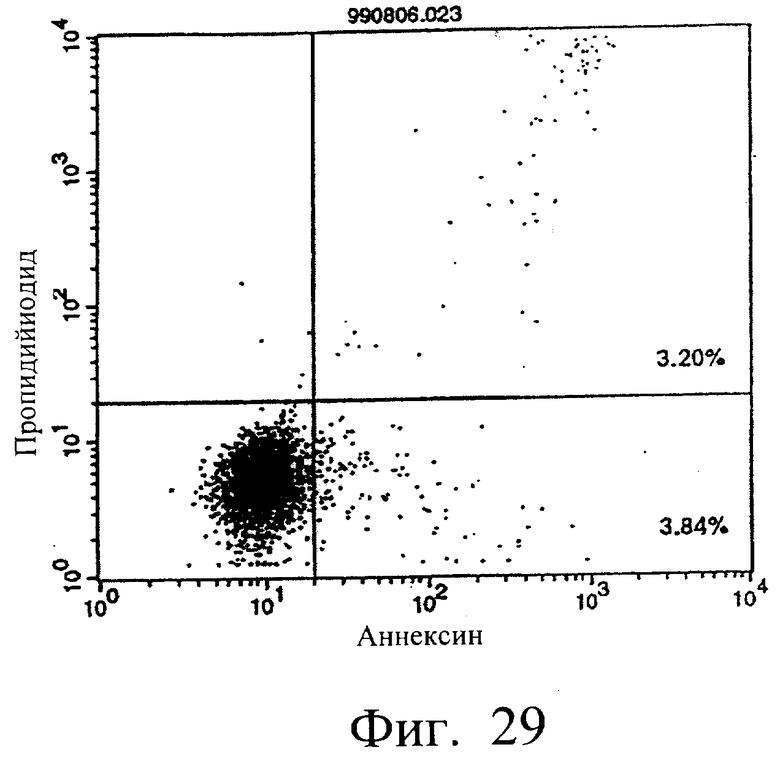
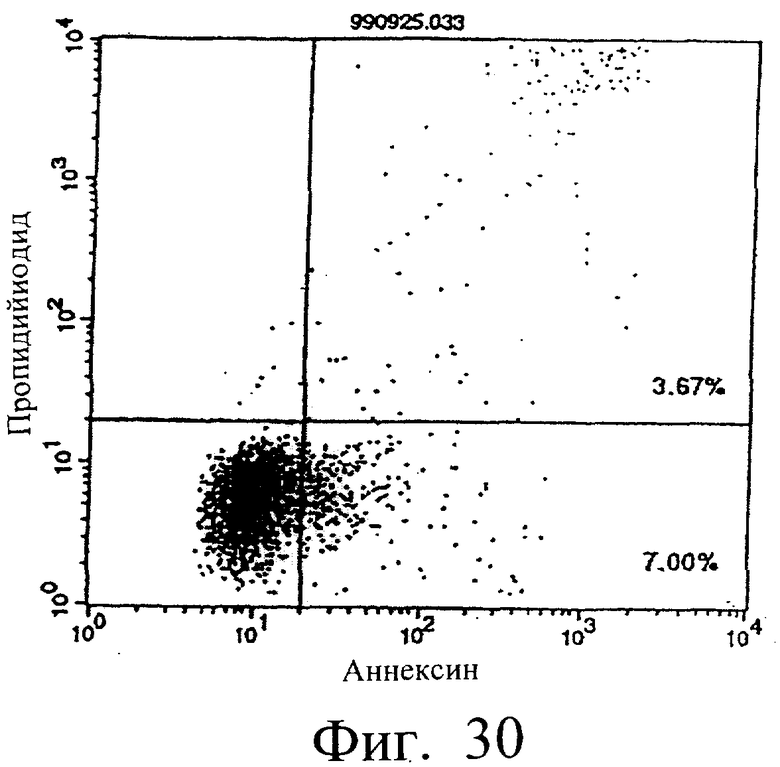
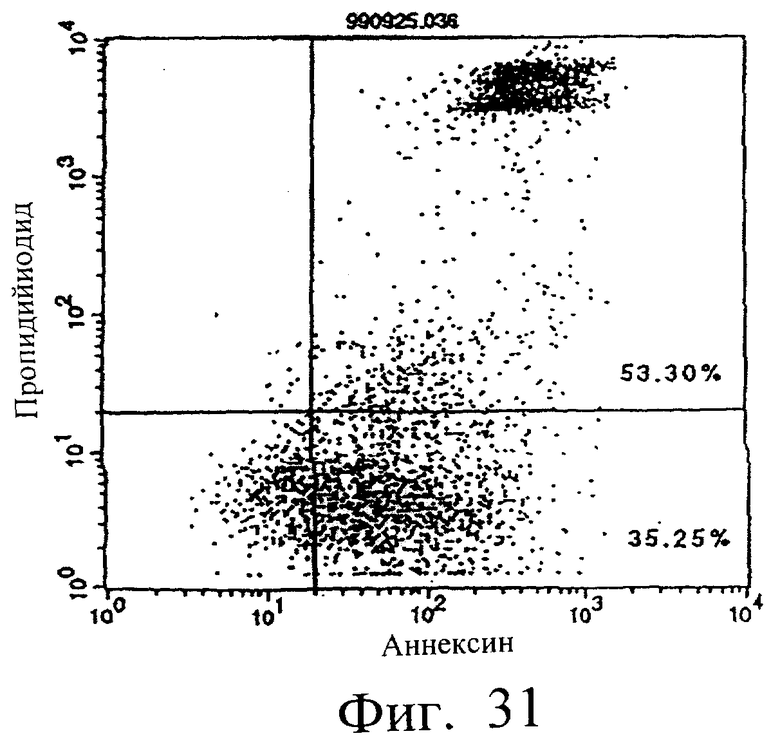
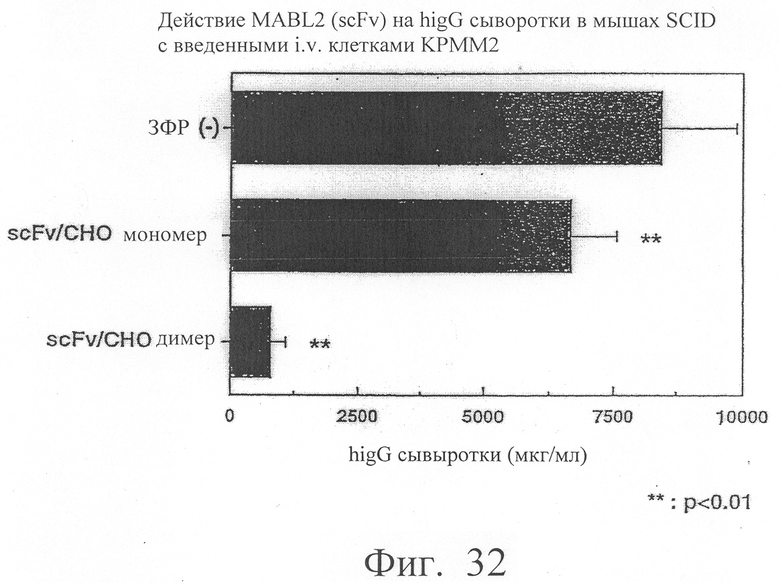
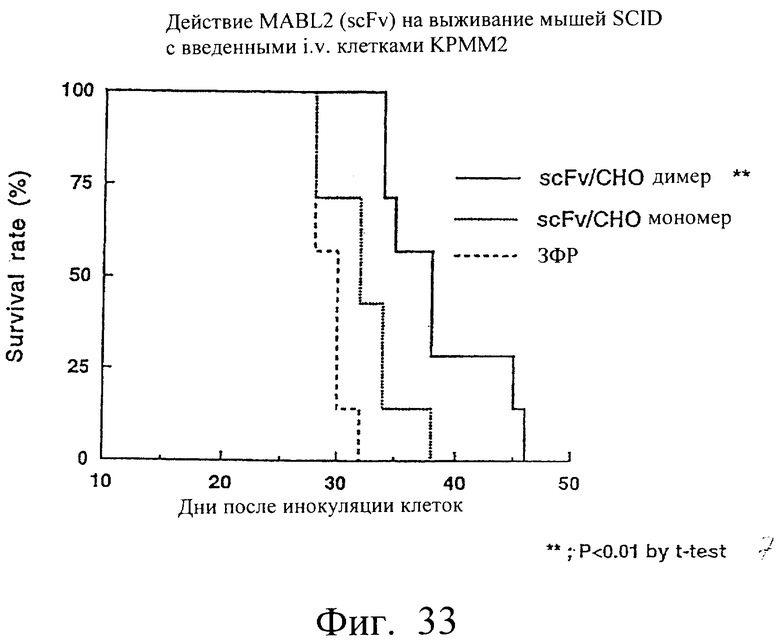
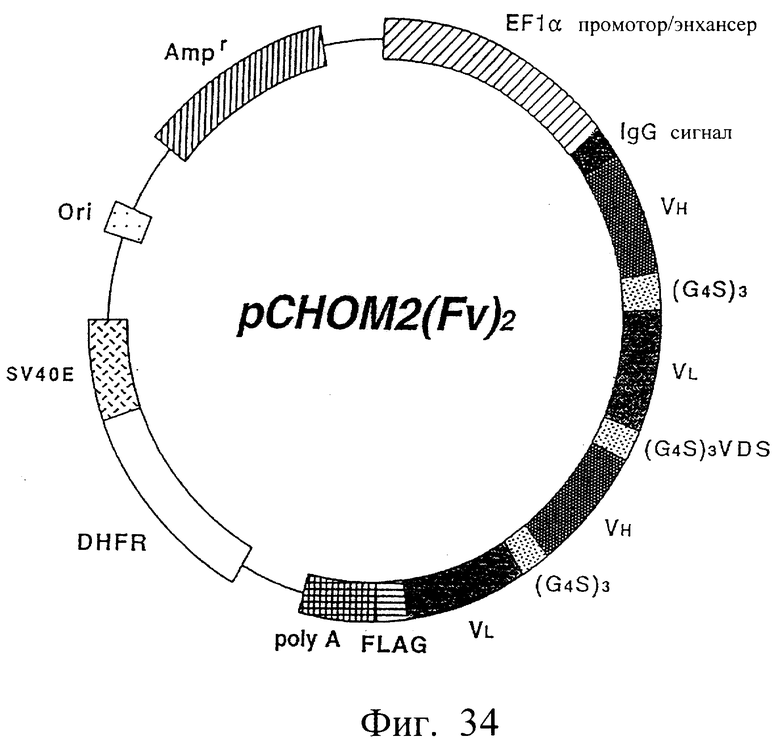
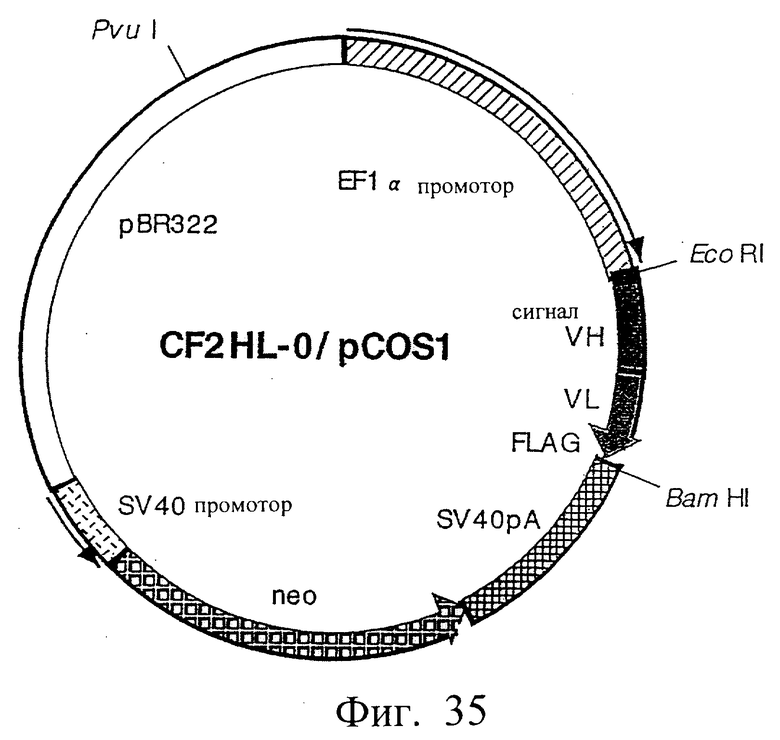

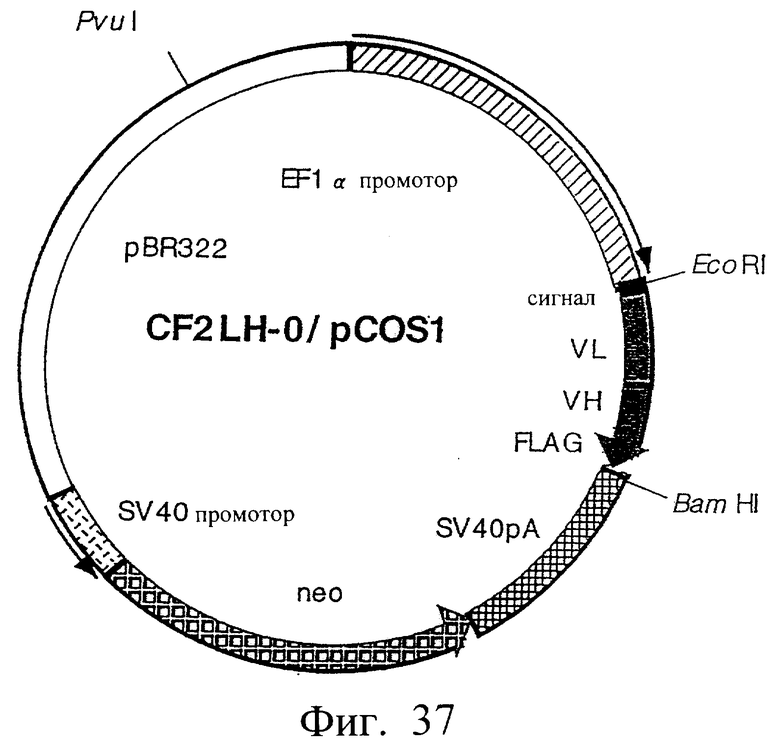
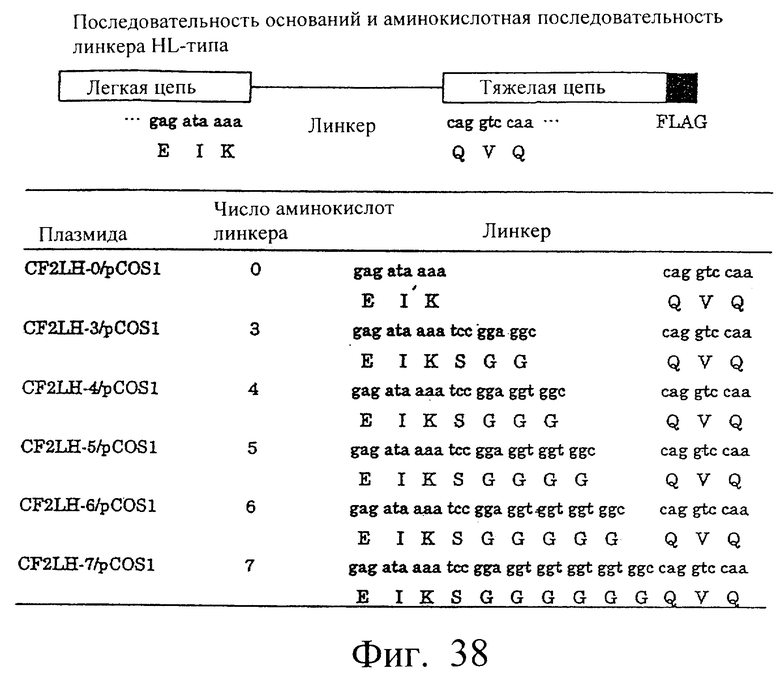
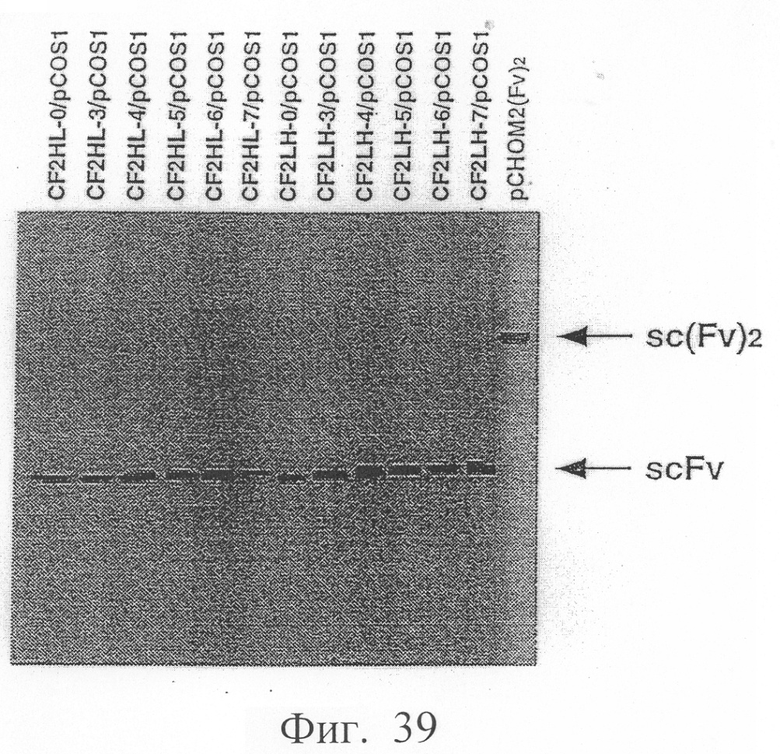
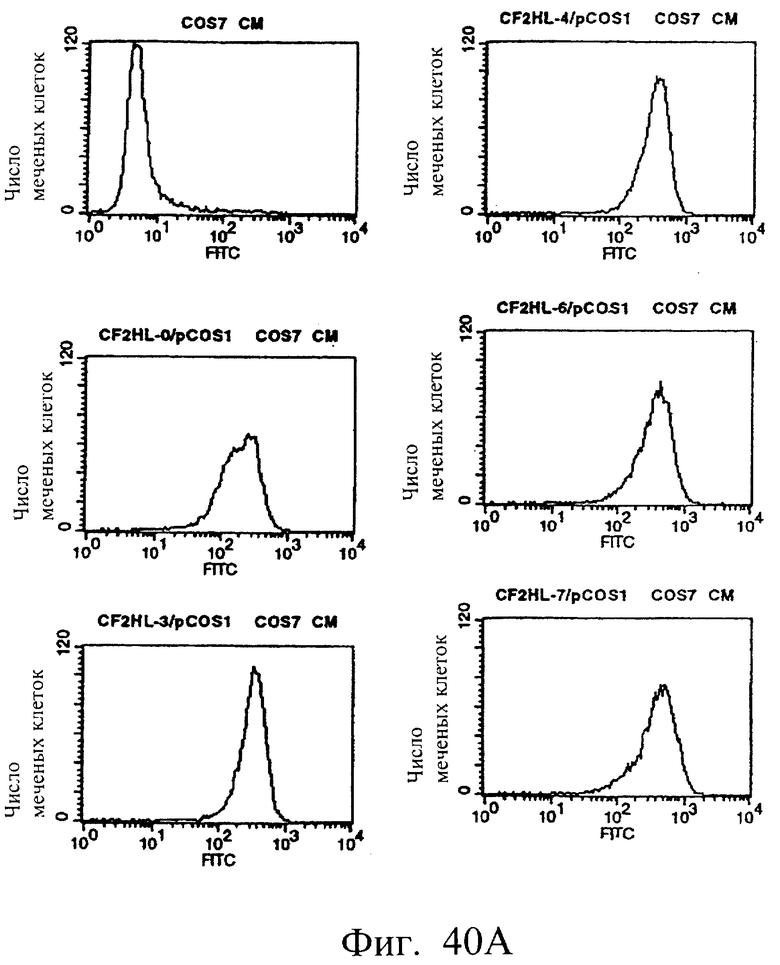
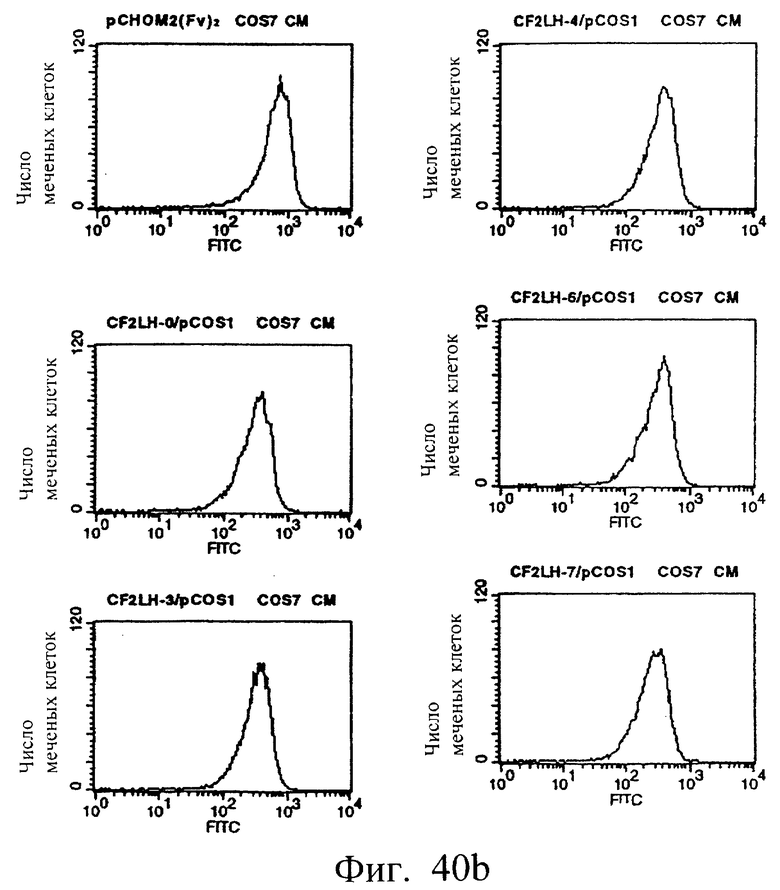
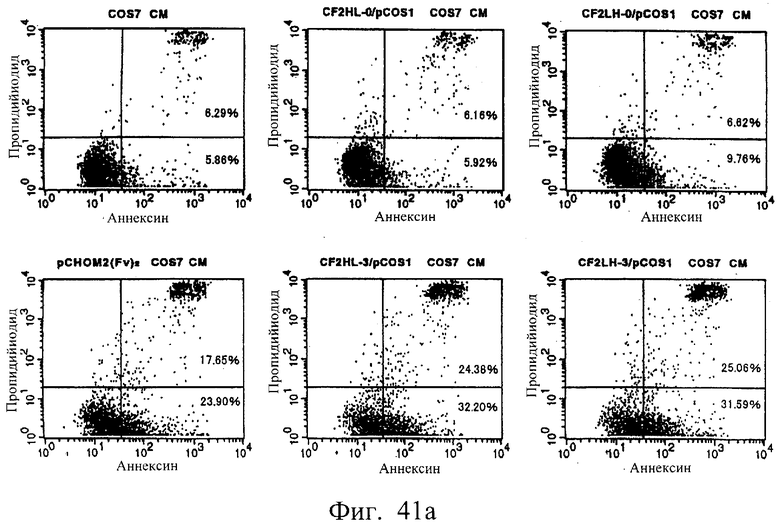
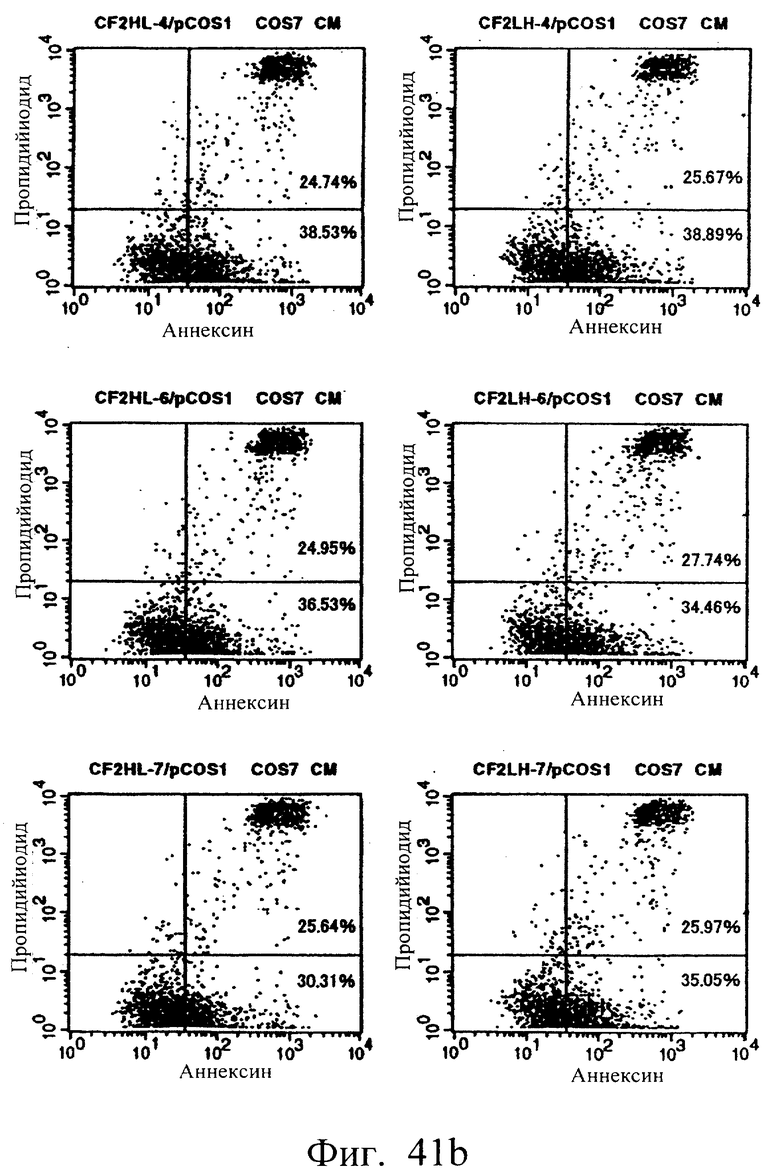
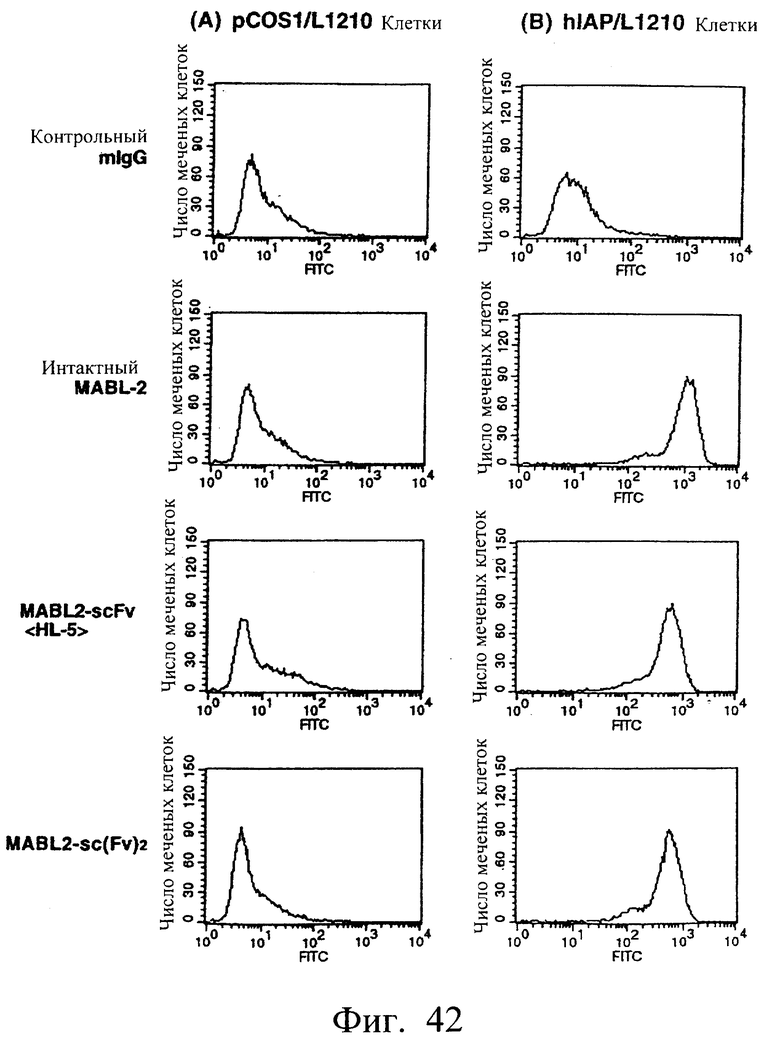
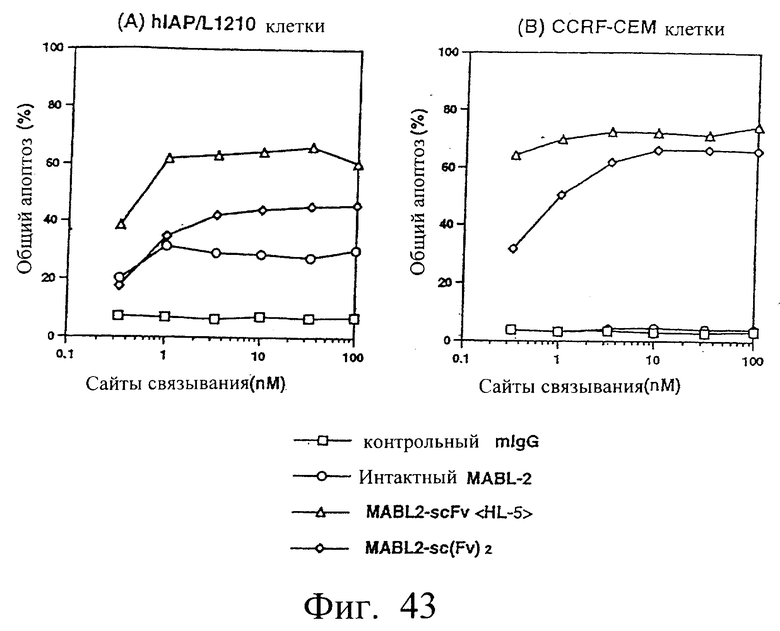
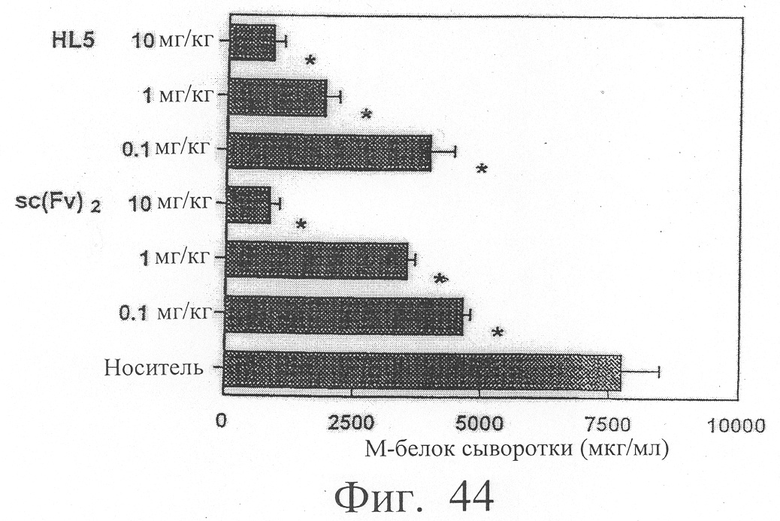
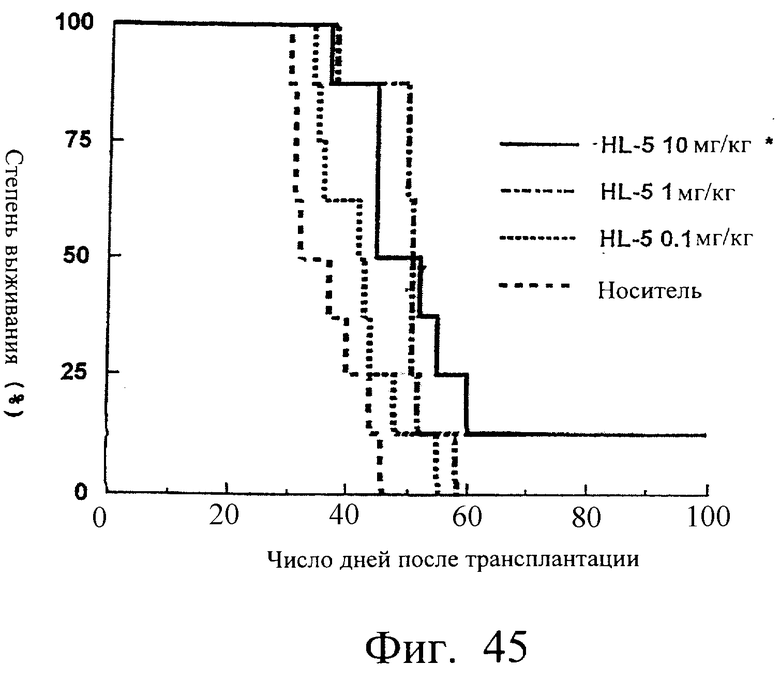
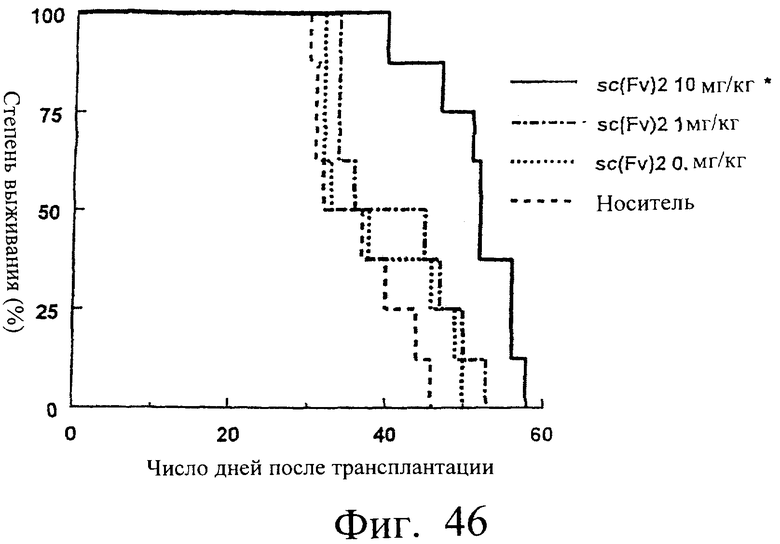
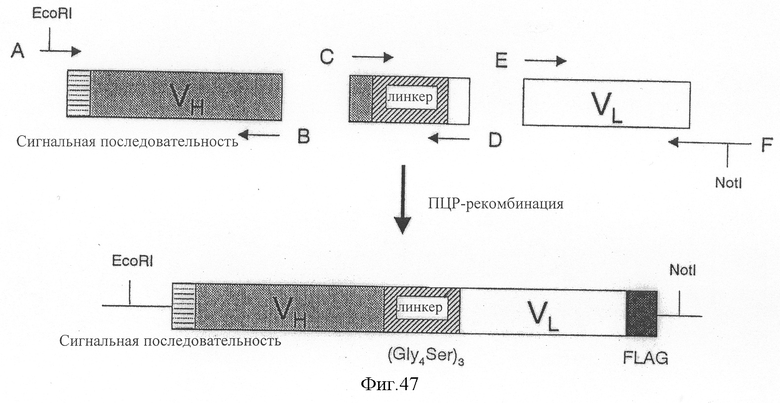
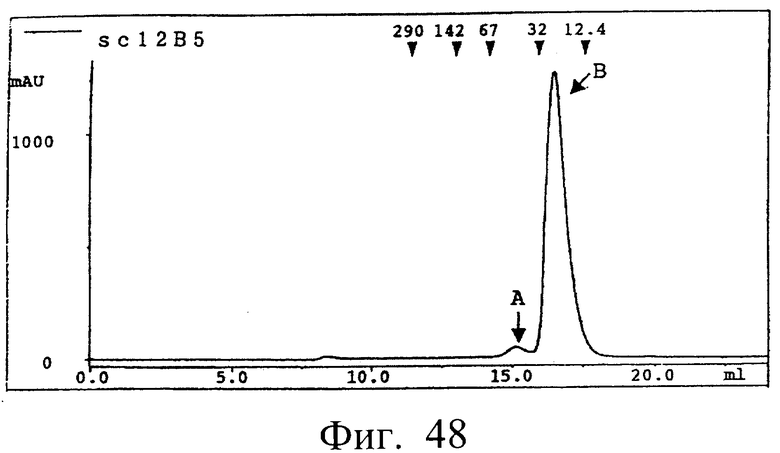
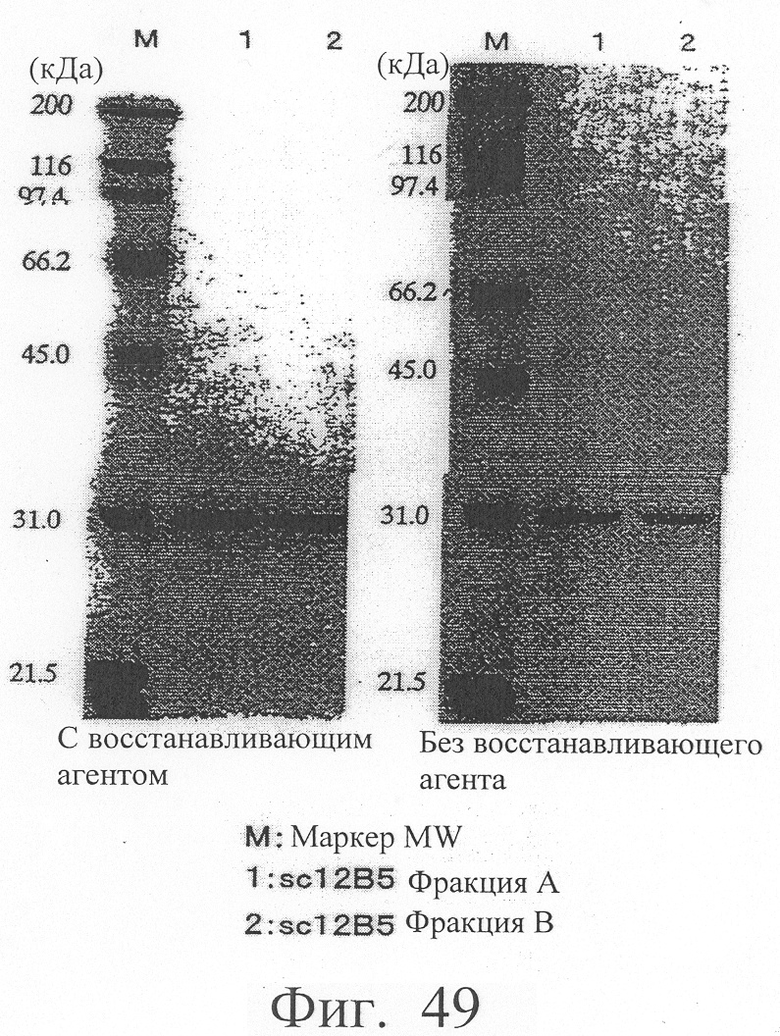
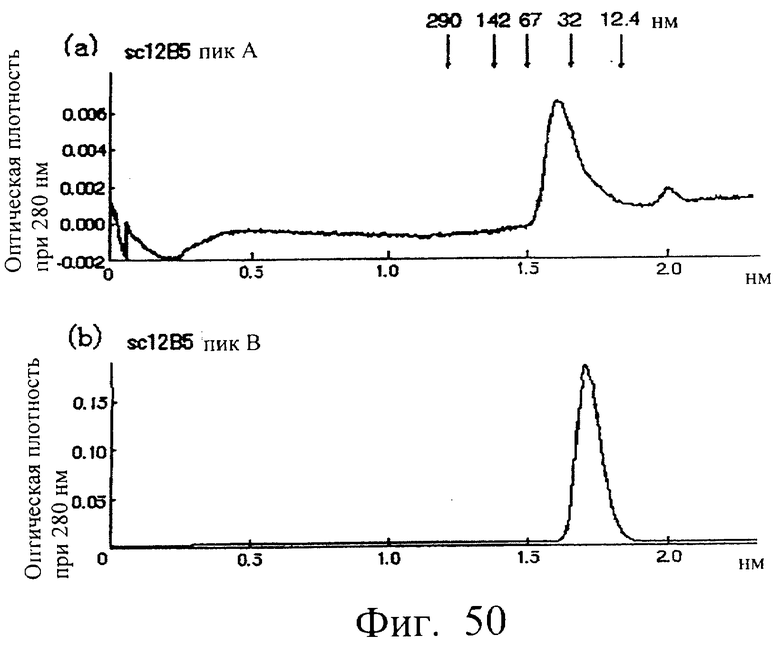
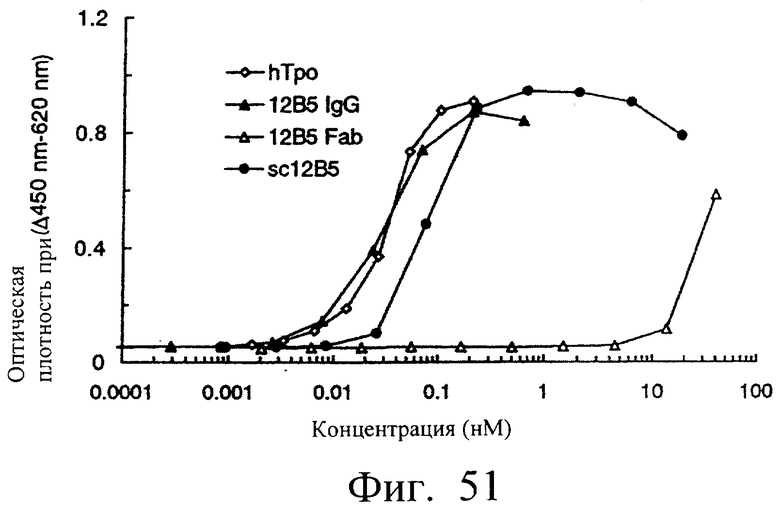
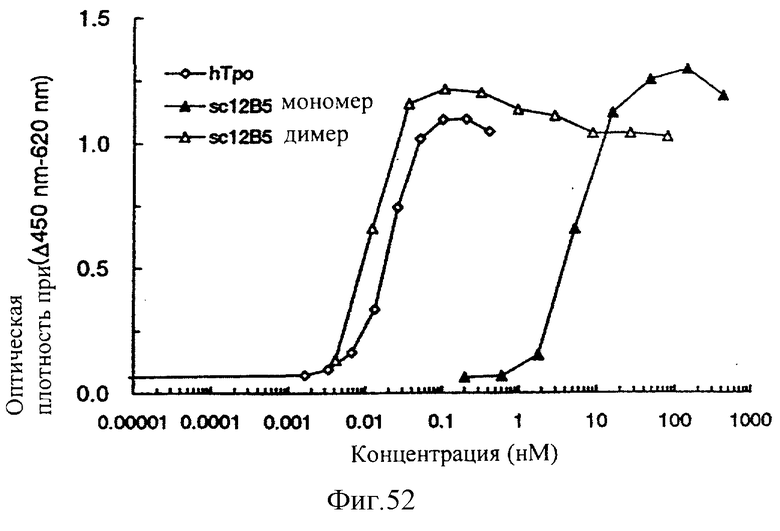
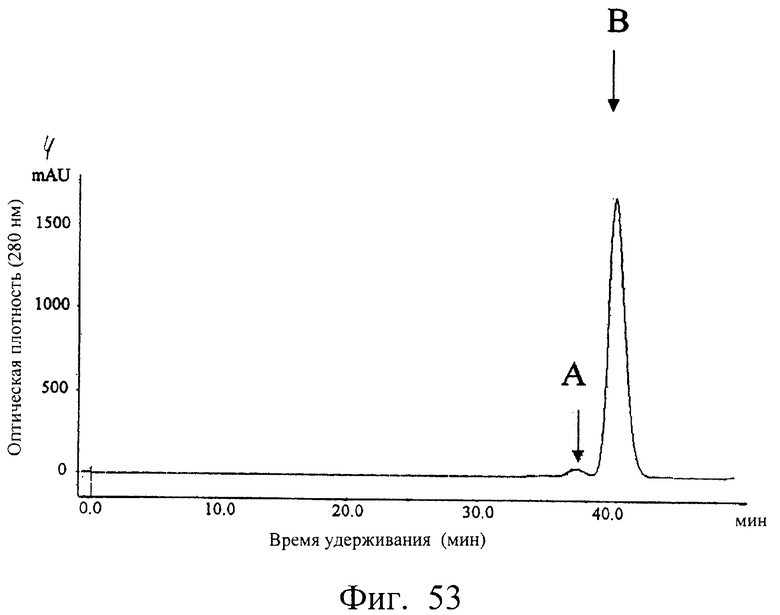
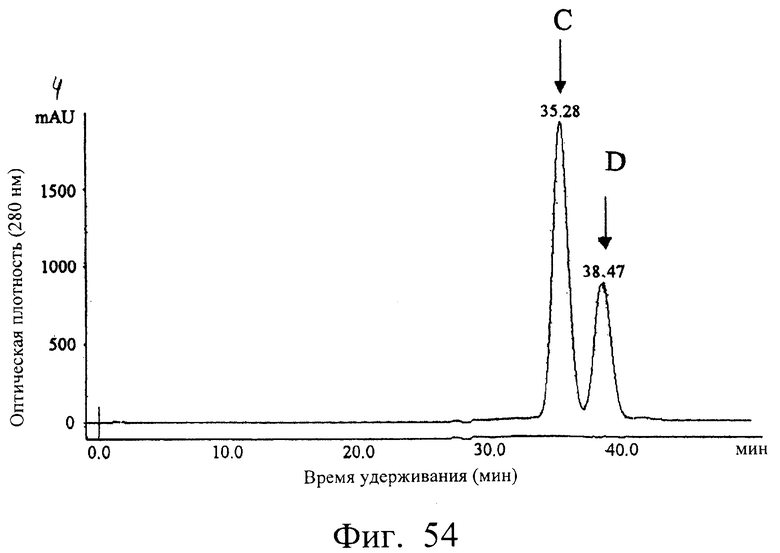

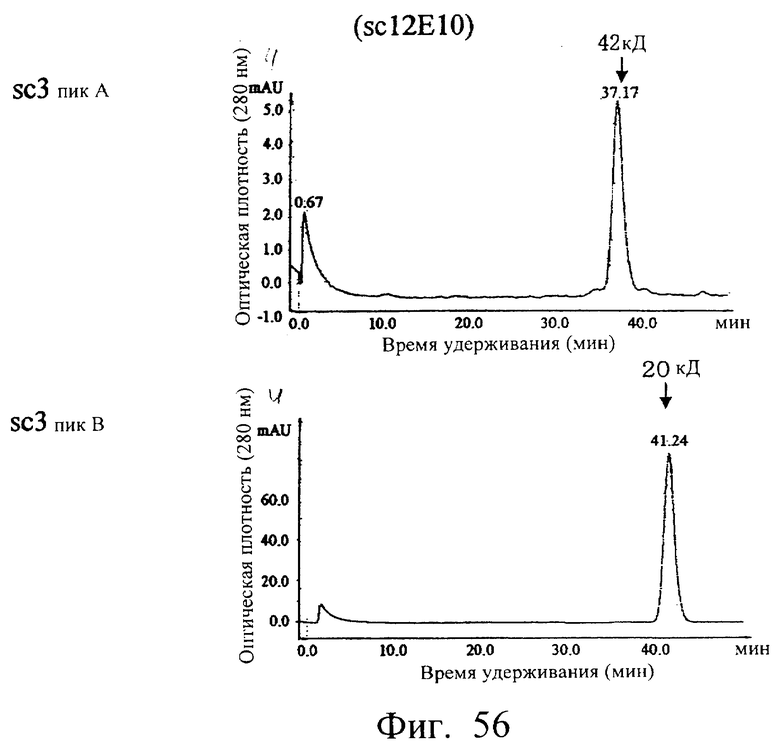
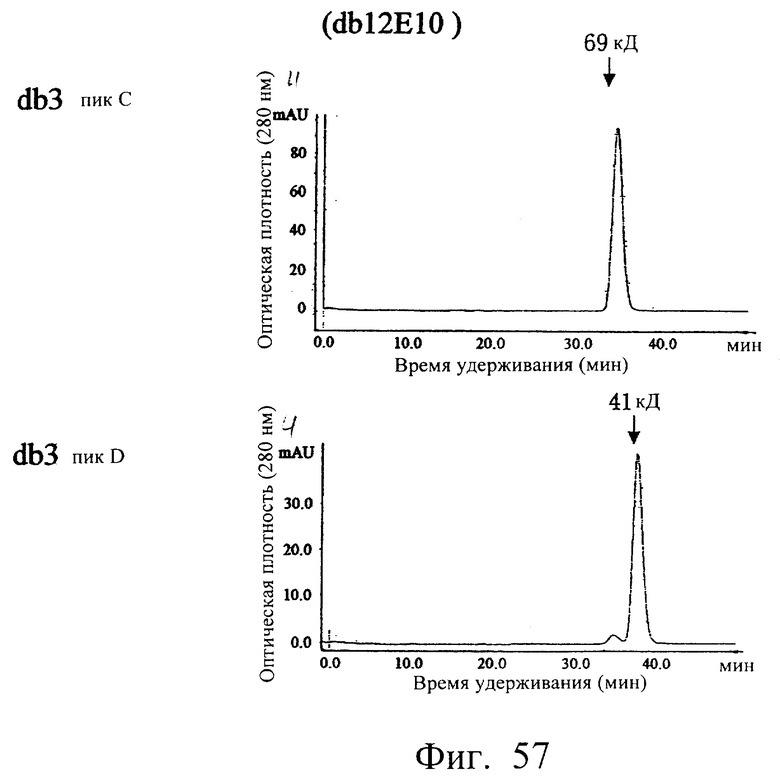
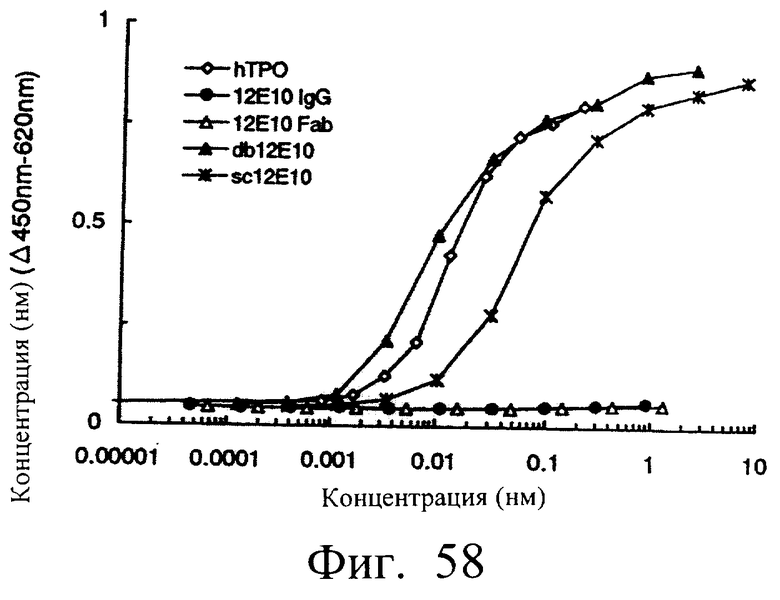
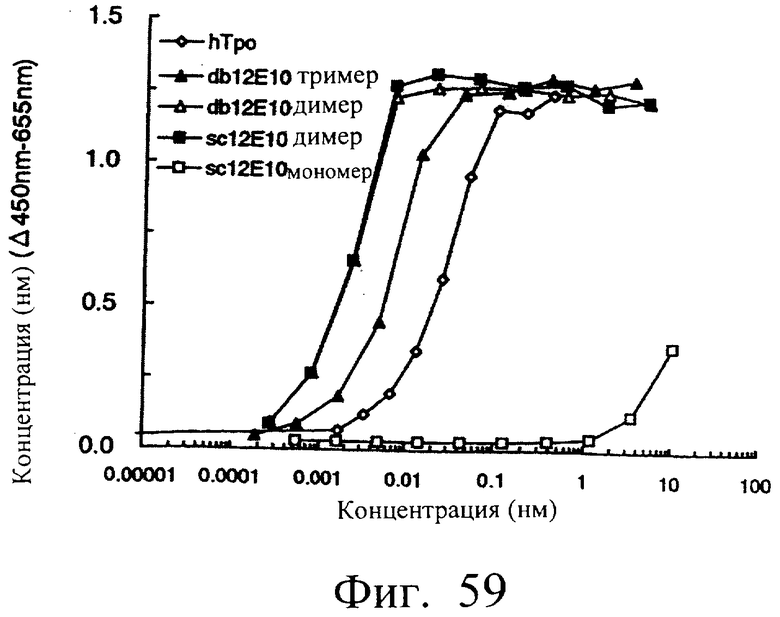
Изобретение относится к иммунобиотехнологии, в частности к модифицированному генно-инженерными методами антителу. Полученное антитело содержит две или более V-областей Н-цепи и две или более V-областей L-цепи моноклонального антитела и может трансдуцировать сигнал в клетки посредством образования поперечных сшивок молекулы (молекул) клеточной поверхности, выполняя посредством этого роль агониста. Модифицированное антитело может быть использовано в качестве агониста трансдукции сигнала и, следовательно, применимо в качестве профилактического средства и/или лечебного средства при различных заболеваниях, таких как рак, воспаление, гормональные нарушения и заболевания крови. Модифицированное антитело может быть использовано для индуцирования агонистического действия в отношении клеток. Изобретение охватывает также способ обнаружения указанного антитела, а также способ измерения агонистического действия антитела в отношении клеток. Изобретение позволяет получать и использовать антитела малого молекулярного размера, обладающего агонистическим действием посредством сшивания молекулы клеточной поверхности или внутриклеточной молекулы. 7 н. и 24 з.п. ф-лы, 59 ил., 3 табл.
(i) мультимер одноцепочечного Fv, содержащего V-область Н-цепи и V-область L-цепи;
или
(ii) одноцепочечный полипептид, содержащий две или более V-областей Н-цепи и две или более V-областей L-цепи.
(i) мультимер одноцепочечного Fv, содержащего V-область Н-цепи и V-область L-цепи
или
(ii) одноцепочечный полипептид, содержащий две или более V-областей Н-цепи и две или более V-областей L-цепи,
включающий стадии:
(1) получения модифицированного антитела, содержащего две или более V-областей Н-цепи и две или более V-областей L-цепи антитела и связывающегося специфически с молекулой (молекулами) клеточной поверхности или внутриклеточной молекулой (молекулами);
(2) взаимодействия клеток, экспрессирующих указанные молекулу (молекулы) клеточной поверхности или внутриклеточную молекулу (молекулы), с этим модифицированным антителом и
(3) измерения агонистического действия в клетках, вызываемого образованием поперечных сшивок указанной молекулы (молекул) клеточной поверхности или внутриклеточной молекулы (молекул).
(1) получения модифицированного антитела, содержащего две или более V-областей Н-цепи и две или более V-областей L-цепи антитела и связывающегося специфически с молекулой (молекулами) клеточной поверхности или внутриклеточной молекулой (молекулами),
(2) взаимодействия клеток, экспрессирующих указанные молекулу (молекулы) клеточной поверхности или внутриклеточную молекулу (молекулы), с этим модифицированным антителом и
(3) измерения агонистического действия в клетках, вызываемого образованием поперечных сшивок указанной молекулы (молекул) клеточной поверхности или внутриклеточной молекулы (молекул).
| US 5885574 А, 23.03.1999 | |||
| WO 9912973 A1, 18.03.1999 | |||
| WO 9838216 A1, 03.09.1998 | |||
| Комбинированная машина для обработки почвы и посева на лесосеках | 1937 |
|
SU53634A1 |

