ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение относится к белку МеСР2 и его применению в белковой заместительной терапии. Более конкретно, это изобретение относится к кодон-оптимизированным последовательностям нуклеиновых кислот для экспрессии белка MeCP2, способам создания такой последовательности нуклеиновой кислоты и экспрессии такого белка, слияниям белка этого изобретения с доменом трансдукции и векторам и клеткам-хозяевам, содержащим белок этого изобретения. Кроме того, настоящее изобретение относится к применениям нуклеиновых кислот или белков настоящего изобретения в медицине, фармацевтическим композициям, содержащим последовательности нуклеиновых кислот и белки настоящего изобретения, а также способам лечения, профилактики и/или терапии нейродегенеративных или связанных с развитием нервных заболеваний, в том числе синдрома Ретта.
УРОВЕНЬ ТЕХНИКИ
Синдром Ретта (RTT) является прогрессирующим, связанным с развитием нервным нарушением. Он поражает почти исключительно женщин (Rett, 1966, Wien Med Wochenschr 116:723-6) и является одной из наиболее частых причин задержки умственного развития у женщин. RTT характеризуется динамически клиническим течением болезни с четырьмя последовательными стадиями. Во время Стадии I (возраст 6-18 месяцев) девочки прекращают приобретение новых навыков; они проявляют замедление роста головы и признаки аутизма, такие как эмоциональный уход в себя (замыкание в себе) и уменьшенный зрительный контакт. В Стадии II (возраст 1-4 года) пораженные дети теряют приобретенные навыки, такие как речь и целевое использование рук. Они развивают неравномерный характер дыхания, атаксию/апраксию туловища и походки и стереотипичные движения рук. Приблизительно у половины девочек развиваются также судороги, и некоторая стабилизация болезни имеется во время Стадии III (возраст 4-7 лет). Судороги становятся менее частыми во время Стадии IV (возраст 5-15 лет и старше), но двигательное расстройство продолжается. Гипофункция (сниженная активность), особенно среди тех, кто не может ходить, способствует частому развитию сколиоза, который может заставить девочек быть прикованными к креслу-каталке. Невропатологические признаки пациентов с синдромом Ретта включают в себя уменьшение толщины коры головного мозга, а также уменьшение размера нейронов коры головного мозга. Было описано также сильное уменьшение дендритной арборизации, но нет влияния на другие макроскопические морфологические признаки (Armstrong, 2002, Ment Retard Dev Disabil Res Rev 8:72-6). RTT представляет собой X-связанное заболевание с приближенно определенной встречаемостью 1:10000-1:15000. Amir et al. (1999) (Nat Genet 23:185-8) идентифицировали мутации в гене MECP2 в качестве причины RTT. Ген MECP2 подвержен X-инактивации. Таким образом, гетерозиготные мутантные женщины являются мозаичными в отношении недостаточности MeCP2, и это, наиболее вероятно, является одним из модулирующих факторов, влияющих на фенотип этого заболевания. Были идентифицированы мужчины, удовлетворяющие клиническим критериям синдрома Ретта, в ассоциации с кариотипом 47,XXY и из постзиготных мутаций MECP2, приводящих к соматической мозаичности (мозаицизму). В качестве ссылки, см. L. S. Weaving et al.: Journal of Medical Genetics 2005; 42:1-7; и G. Miltenberger-Miltenyi, F. Laccone: Human Mutation 2003 Volume 22, Issue 2; 107-115.
Ген MECP2 локализован на длинном плече хромосомы Х в положении Xq28 (Adler 1995, Mamm Genome 6(8):491-2). Этот ген охватывает 76 т.п.н. и состоит из четырех экзонов. Ген MECP2 кодирует белок, названный метил-СрG-связывающим белком 2 (MeCP2), причем считается, что он играет центральную роль в сайленсинге других генов. Белок MeCP2 имеет две изоформы, MeCP2 e1 и MeCP2 e2, ранее называемые MeCP2B и MeCP2A, соответственно (Mnatzakanian et al. 2004, Nat Genet 36:339-41). Изоформа e1 состоит из 498 аминокислот, а изоформа e2 имеет длину 486 аминокислот. Изоформа e1 имеет отдельный, состоящий из 21 аминокислоты, пептид на N-конце, включающий в себя участки полиаланина и полиглицина. мРНК варианта MECP2 e1 имеет в 10 раз большую экспрессию в головном мозге, чем мРНК MECP2 e2, и она является наиболее часто встречающейся изоформой белка в головном мозге мыши и человека. MeCP2 является часто встречающимся белком млекопитающих, который селективно связывает остатки 5-метилцитозина в симметрично расположенных динуклеотидах. CpG-динуклеотиды предпочтительно локализованы в промоторных районах генов. Они представляют один из элементов регуляции генов, являющихся мишенью факторов сайленсинга транскрипции после метилирования ДНК.
Еще неизвестно успешное лечение для улучшения неврологического исхода индивидуумов с синдромом Ретта. Согласно спектру мутаций MECP2 в человеке (Lam et al. 2000, J Med Genet 37(12):E41; Lee et al. 2001, Brain Dev 23:S138-43) и результатам из RTT-мыши, всеми признается, что синдром Ретта обусловлен потерей функции MeCP2. Эффективная стратегия, нацеленная на восстановление активности MeCP2, должна быть способна компенсировать потерю функции MeCP2 в недостаточных нервных клетках. В отношении ссылки, см. L. S. Weaving et al.: Journal of Medical Genetics 2005; 42:1-7; и G. Miltenberger-Miltenyi, F. Laccone: Human Mutation 2003, Volume 22, Issue 2; 107-115.
Schwarze et al. (1999, Science 285:1569-1572) сообщают, что можно доставлять биологически активные макромолекулы в живые клетки с использованием домена ТАТ (белка-трансактиватора транскрипции вируса-1 иммунодефицита человека). Они показывают получение рекомбинантного белка TAT-β-галактозидазы и его инъекцию внутрибрюшинно в мышей. Они обнаружили, что этот слитый белок распределялся во все ткани, в том числе в головной мозг, и этот слитый белок был биологически активным.
WO 00/62067 (Dowdy et al.) сообщает о применении молекул трансдукции белка (PTD), включающих в себя домены белка ТАТ для нацеливания терапевтических молекул в нервной системе.
Продуцирование белков в гетерологичных системах экспрессии (т.е. белка человека в Escherichia coli) требует соответствующего экспрессирующего вектора и кДНК-последовательности представляющего интерес гена. 64 кодона (триплетов нуклеотидов) генетического кода кодируют 20 аминокислот и три стоп-сигнала трансляции. Таким образом, генетический код является избыточным, и это означает, что некоторые аминокислоты кодируются более чем одним кодоном. Метионин и триптофан являются единственными аминокислотами, кодируемыми одним кодоном, ATG и TGG, соответственно, тогда как аргинин, лейцин и серин, каждый, кодируются шестью синонимичными кодонами. Вследствие вырожденности генетического кода, многие альтернативные последовательности нуклеиновых кислот кодируют один и тот же белок. Эти неоднозначности кодонов могут сильно влиять на экспрессию гетерологичных белков (Kane, 1995, Curr Opin Biotechnol 6(5):494-500). Частота использования кодона была идентифицирована как единственный наиболее важный фактор в экспрессии прокариотических генов (Lithwick, 2003, Genome Res 13 (12):2665-2673).
Одной целью авторов настоящего изобретения является обеспечение конструкций, кодирующих биологически активные белки MeCP2, которые способны входить в клетки нервной системы и компенсировать потерю функции MeCP2 пораженных нервных клеток.
Терапевтический подход с использованием доставки ТАТ-рекомбинантных белков в головной мозг мог бы иметь огромное преимущество возможного быстрого перехода от модели животных к пациентам. Дополнительными преимуществами могут быть применение легко контролируемой дозы такого белка, очень высокая эффективность доставки, отсутствие проблем, присущих потенциально инсерционному мутагенезу или клинических побочных эффектов вследствие иммунологической реакции против вирусных белков, которые возникают в случае подхода с использованием генной терапии. Подход с доставкой PTD-белка и его дополнительное развитие могут сделать возможным лечение нейрогенетических и изнуряющих заболеваний, таких как синдром Ретта.
Поскольку попытки экспрессии конструкций белка MeCP2 с использованием кДНК-последовательности человека в количествах, достаточных для терапевтических целей, были до сих пор безуспешными, другой целью настоящего изобретения является обеспечение экспрессионной конструкции MeCP2, которая делает возможной увеличенное продуцирование рекомбинантного белка MeCP2.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Таким образом, настоящее изобретение относится к молекуле нуклеиновой кислоты, содержащей первую последовательность нуклеиновой кислоты, кодирующую белок MeCP2 или биологически активный фрагмент или производное указанного белка или фрагмента, как определено в формуле изобретения. Кроме того, настоящее изобретение относится к полипептиду, кодируемому молекулой нуклеиновой кислоты, вектору, содержащему молекулу нуклеиновой кислоты, и клетке-хозяину, содержащему вектор, как определено в формуле изобретения. Кроме того, настоящее изобретение обеспечивает способ получения последовательности нуклеиновой кислоты, определенный в формуле изобретения, а также способ получения полипептида, определенный в формуле изобретения. Кроме того, обеспечена фармацевтическая композиция, содержащая молекулу нуклеиновой кислоты и/или полипептид, в соответствии с формулой изобретения. Настоящее изобретение относится также к способу лечения и применению этих молекул нуклеиновых кислот и/или полипептидов для применения в медицинских курсах лечения, описанных в формуле изобретения.
ПОДРОБНОЕ ОПИСАНИЕ ЭТОГО ИЗОБРЕТЕНИЯ
Настоящее изобретение решает давно поставленную задачу предоставлением средств и путей обеспечения рекомбинантных белков MeCP2, которые могут быть использованы в терапии нейродегенеративных и связанных с развитием нервных заболеваний. Авторы изобретения обнаружили, что посредством конструирования и создания оптимизированных последовательностей нуклеиновых кислот MeCP2, впервые можно обеспечивать белки MeCP2, а также фрагменты или производные таких белков или фрагментов, которые могут быть использованы для способов белковой заместительной терапии в млекопитающих.
В первом аспекте, настоящее изобретение относится к молекуле нуклеиновой кислоты, содержащей первую последовательность нуклеиновой кислоты, кодирующую белок MeCP2 или биологически активный фрагмент или производное этого белка или фрагмента, где эта последовательность нуклеиновой кислоты является кодон-оптимизированной для экспрессии в гетерологичной клетке.
“Белок MeCP2", в данном контексте, может быть изоформой е1 человека (иначе называемой изоформой B), имеющей номер доступа AAS55455, или изоформой е2 человека (иначе называемой изоформой А), имеющей номер доступа NP_004983 (базы данных последовательностей белков NCBI), соответственно. Квалифицированному в данной области специалисту будет понятно, что другие изоформы белков MeCP2 или MeCP2 человека, происходящие из других позвоночных, будут также пригодны в качестве белков MeCP2 настоящего изобретения, если эти белки или их фрагменты обнаруживают сходную биологическую активность с биологической активностью известных изоформ MeCP2 человека. Квалифицированный в данной области специалист может легко получить последовательности вышеупомянутых изоформ и их соответствующих мРНК из публично доступных баз данных.
Термин “биологически активный фрагмент”, в данном контексте, относится к полипептиду из по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150 или по меньшей мере 200 аминокислот белка MeCP2. Фрагмент белка MeCP2 содержит меньше аминокислот, чем любой из полноразмерных белков MeCP2, которые могут быть найдены под номером доступа AAS55455 или под номером доступа NP_004983, соответственно. Биологически активный фрагмент все еще проявляет биологическую активность природно-встречающегося белка, из которого он получен, хотя и необязательно в той же самой степени.
“Производное” белка MeCP2 является полипептидом, который не кодируется как таковой геномом встречающихся в природе видов. В частности, производное является полипептидом, который не является идентичным одному из полипептидов, имеющих номера доступа любой из вышеупомянутых изоформ MeCP2. Таким образом, производное содержит модификацию встречающихся в природе белков MeCP2, например, посредством замены, делеции и присоединения аминокислот, но все еще проявляет биологическую активность встречающегося в природе белка, хотя необязательно в той же самой степени. Производное включает в себя молекулы, содержащие районы, которые являются гомологичными белку MeCP2, имеющими по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 99% идентичность на протяжении аминокислотной последовательности идентичного размера, когда эти две сравниваемые последовательности сопоставляют с использованием компьютерной программы гомологии, известной в данной области, такой как, например, программа “BLAST”, которая доступна публично на http://www.ncbi.nlm.nih.gov/BLAST/. Фрагмент производного белка MeCP2 содержит меньше аминокислот, чем производное белка MeCP2, определенное выше.
“Биологически активный” фрагмент или “биологически активное” производное белка или фрагмента означает, что этот фрагмент или это производное имеет по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 100% биологической активности белка MeCP2, определенного выше. Является ли белок или фрагмент или производное MeCP2 биологически активным, можно определить, например, при помощи следующих тестов:
1. Белок MeCP2, или фрагмент, или производное считается биологически активным, если он способен связываться с метилированными цитозинами и/или индуцировать прямой рекрутинг HDAC и/или изменять состояние ацетилирования гистона H3 и гистона H4. Обычный тест включает в себя культивирование фибробластов мышей Mecp2y/- (доступных от Jackson Laboratory под B6.129P2(C)-Mecp2tml.lBird/J) и мышей дикого типа из задней конечности/кончиков хвоста, например, установленное в соответствии с протоколом, описанным в 3.1 экспериментального раздела, приведенного ниже.
Обычно, среда будет заменена и затем эти клетки будут инкубироваться в течение недели при 37°С с 10% CO2. Для тестирования на биологическую активность, например, мониторингом состояния ацетилирования гистона Н3 и гистона Н4 фибробласты мыши Mecp2y/- обрабатывают, например, 300 пмоль белка MeCP2 или фрагмента или производного белка MeCP2 или фрагмента, слитого с доменом трансдукции, таким как домен ТАТ или другие домены трансдукции, описанные ниже, и инкубируют, например, в течение 30 часов. Затем лизаты клеток иммуноблоттируют с антителом против, например, гистона Н3, ацетилированного гистона Н3 или ацетилированного H4K16, как описано ниже. Различие состояния ацетилирования между обработанными и необработанными фибробластами определяют количественно денситометрическим анализом, как представлено, например, на фиг. 5С, и, следовательно, квалифицированный в данной области специалист будет способен определить изменение в состоянии ацетилирования гистонов. Примеры 5, 6 и 8.1 в комбинации с фигурой 3 С-D и фигурой 5 ниже показывают путь выполнения и оценки таких анализов.
2. Другой возможностью тестирования на биологическую активность белка MeCP2 или фрагмента или производного белка MeCP2 или фрагмента является измерение уровня ацетилирования Н3, индуцированного экспериментами по трансдукции, проводимыми с белками, подлежащими тестированию, слитыми с доменами трансдукции белков, как описано ниже. Для этой цели могут быть использованы обычно доступные клеточные линии, такие как HeLa или NIH 3T3. При помещении этих клеток в контакт со слитыми белками, уровень ацетилирования гистона Н3 будет уменьшаться в сравнении с линией контрольных клеток, которые имеют нормальный уровень ацетилирования гистона Н3. Это уменьшение можно использовать для количественного определения биологической активности продуцируемого белка. Оценка этого теста, например, способы Вестерн-блоттинга, описана в экспериментальном разделе ниже.
3. Белки MeCP2 или фрагменты или производные таких белков могут быть определены как биологически активные по их способности связываться с метилированными цитозинами, например, при помощи анализов транскрипции in vitro, таких как описанные в Nan, X et al., Cell 88: 471-481 (1997), в Yusufzai T.M. and Wolffe, A.P., Nucl. Acids Res. 28: 4172-4179 (2000) или Yu F. et al., Nucl. Acids Res. 29(21):4493-501 (2001).
Термин “кодон-оптимизированная” последовательность нуклеиновой кислоты относится, в данном контексте, к последовательности нуклеиновой кислоты, содержащей кодоны, которые сконструированы таким образом, чтобы было возможно использование кодонов, предпочтительных для желаемых клеток-хозяев, предпочтительно для клетки-хозяина E. coli. Последовательность нуклеиновой кислоты превращают в кодон-оптимизированную последовательность нуклеиновой кислоты, имеющую идентичную транслируемую полипептидную последовательность, но с альтернативной частотой использования кодонов, в частности, с использованием наиболее часто кодонов данного организма, например, Escherichia coli. Такой способ называют «обратной трансляцией». Способ отбора и приготовления кодон-оптимизированных последовательностей нуклеиновых кислот и выполнение обратной трансляции включает, например, использование данных из гена Класса II из Henaut and Danchin: Analysis and Predictions from Escherichia coli sequences in: Escherichia coli and Salmonella, Vol. 2, Ch. 114:2047-2066, 1996, Neidhardt FC ed., ASM press, Washington, D.C. Руководство в отношении обратной трансляции конкретной белковой последовательности дается также в инструментах, доступных на http://www.prodoric.de/JCat (Grote A. et al., Nucleic Acids Research, 2005, Vol. 33, Web Server issue doi:10.1093/nar/gki376, W526-W531) или программном обеспечении Vector NTI (Invitrogen). Способ создания кодон-оптимизированной последовательности нуклеиновой кислоты белка MeCP2 включает в себя идентификацию кодонов в природно-встречающейся последовательности гена MECP2, которые обычно не ассоциированы с высокой экспрессией генов E. coli, и замену их кодонами, о которых известно, что они широко используются в экспрессии генов E. coli. Кодон-оптимизированная последовательность нуклеиновой кислоты может обнаруживать улучшенную экспрессию в сравнении с встречающейся в природе последовательностью в желаемой клетке-хозяине. Будет ли кодон-оптимизированная последовательность индуцировать улучшение в продуцировании этого белка в сравнении с неоптимизированной последовательностью, может быть испытано квалифицированным в данной области специалистом в связи с данным описанием. Пример того, как сконструировать и создать кодон-оптимизированную последовательность нуклеиновой кислоты настоящего изобретения, представлен в экспериментальном разделе ниже.
“Гетерологичной” клеткой называют в данном контексте клетку, экспрессирующую ген, который не является встречающимся в природе геном этой конкретной клетки.
В предпочтительном варианте осуществления, белок MeCP2 или биологически активный фрагмент или производное этого белка или фрагмента происходят из человека. Более конкретно, белок MeCP2 или биологически активный фрагмент или производное этого белка или фрагмента могут быть изоформой е1 MeCP2 человека или изоформой е2 MeCP2 человека или биологически активным фрагментом или производным этих изоформ или этих фрагментов. Квалифицированному в данной области специалисту будет ясно, что другие биологически активные изоформы MeCP2 человека или не из человека или их биологически активные фрагменты или производные этих изоформ или фрагментов могут быть также использованы в качестве белков MeCP2 этого изобретения.
В другом варианте осуществления, эта первая последовательность нуклеиновой кислоты, определенная в формуле изобретения, имеет по меньшей мере 60%, по меньшей мере 65%, в частности, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85% или по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 99% идентичность последовательности с ДНК-последовательностью кодон-оптимизированной изоформы е1 MeCP2 человека (SEQ ID NO:1) или кодон-оптимизированной изоформы е2 MeCP2 человека (SEQ ID NO:2). Предполагается также 100% идентичность последовательности. Молекула нуклеиновой кислоты имеет “по меньшей мере × % идентичность” c SEQ ID Nо: 1 или SEQ ID Nо: 2, если при сопоставлении рассматриваемой последовательности нуклеиновой кислоты с последовательностью наилучшего соответствия SEQ ID Nо: 1 или SEQ ID Nо: 2, идентичность последовательности между двумя сопоставленными последовательностями равна по меньшей мере × %. Такое сопоставление может быть выполнено публично доступной компьютерной программой гомологии, такой как программа “BLAST”, обеспеченная на собственной странице NCBI на http://www.ncbi.nlm.nih.gov/BLAST/. Хотя нефункциональные полипептиды MeCP2, или фрагменты, или производные могут быть применимы, например, для диагностических целей, для терапевтических целей желательными являются биологически функциональные белки или фрагменты или производные. Биологическая активность такого полипептида может быть определена вышеописанными анализами.
В дополнительном предпочтительном варианте осуществления, молекула нуклеиновой кислоты этого изобретения экспрессируется в прокариотической клетке, в частности, в грамотрицательной или грамположительной клетке, например, в клетках E. coli или Bacillus sp. Прокариотические клетки, подходящие для экспрессии молекул нуклеиновых кислот, например, грамотрицательные или грамположительные бактерии, известны в данной области. Квалифицированному в данной области специалисту будет понятно, что другие хорошо известные системы экспрессии могут быть использованы для достижения экспрессии белков из молекул нуклеиновых кислот этого изобретения. Такие подходящие клетки могут быть клетками не человека внутри или вне тела животного или клеткой человека вне тела человека. Примерами являются клетки млекопитающих, такие как клетки НЕК, клетки HeLa, клетки СНО и другие. Примерами клеток, не происходящих из млекопитающих или даже не происходящих из позвоночных, являются клетки Шнайдера дрозофилы, другие клетки насекомых, такие как клетки Sf9, клетки дрожжей, другие грибные клетки, и другие.
В другом варианте осуществления, это изобретение обеспечивает молекулу нуклеиновой кислоты этого изобретения, которая дополнительно содержит вторую последовательность нуклеиновой кислоты, кодирующую полипептид, содержащий домен трансдукции белка, где эта вторая последовательность находится в функциональной связи с первой последовательностью нуклеиновой кислоты. Конкретно, молекула нуклеиновой кислоты, содержащая первую и вторую последовательности нуклеиновой кислоты согласно этому изобретению, выбрана из любой из последовательностей SEQ ID Nо: 50 или SEQ ID Nо: 51.
Домены трансдукции белка являются районами белка с высокой плотностью положительных зарядов и способны пересекать биологические мембраны неклассическими путями. Эти районы обычно имеют длину, меньшую чем 30 аминокислот, и требуют их основного характера и, следовательно, их свойств трансдукции на основании высокого содержания аргинина и, в меньшей степени, лизина; делается ссылка, например, на Mol. Cell Proteomics. 2004, 3 (8): 746-69. Домен трансдукции может быть получен из любого белка или его части, которые могут способствовать вхождению в клетку другого белка, слитого с доменом трансдукции. Способность домена трансдукции к трансдукции белка в клетку может быть определена любым общепринятым способом мониторинга поглощения белка клетками, обычно FACS-сортингом или различными микроскопическими способами, такими как флуоресцентная микроскопия. Домены трансдукции белков, способные трансдуцировать сопряженный полипептид, могут быть получены, например, из белка-трансактиватора транскрипции вируса-1 иммунодефицита человека (ТАТ-белка, с номером доступа AAQ86751), гомеодомена Antennapedia (Derossi et al., J. Biol. Chem., 269: 10444 (1994) и HSVVP22 (Elliot and O'Hare, Cell, 88: 223 (1997)), или других доменов трансдукции белков, таких как участки полиаргининов (8-10) (в отношении обзора см. Jones S.W. et al., Br. J. Pharmacol. (2005) 145(8):1093-1102), или синтетического пептида PTD4 (описанного в Ho A. et al., Cancer Res. (2001) 15; 61(2):474-477). В предпочтительном варианте осуществления, домен трансдукции белка получен из белка ТАТ (белка-трансактиватора лентивирусов, например, ВИЧ). Еще более предпочтительно, последовательность нуклеиновой кислоты, кодирующая домен трансдукции белков, кодирует аминокислотную последовательность, которая имеет по меньшей мере 60%, по меньшей мере 65%, в частности, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или 100% идентичность последовательности с аминокислотной последовательностью домена трансдукции белка TAT (YGRKKRRQRRR, SEQ ID Nо: 54). Аминокислотная последовательность имеет “по меньшей мере × % идентичность” c SEQ ID Nо: 54, если при сопоставлении рассматриваемой аминокислотной последовательности с последовательностью наилучшего соответствия SEQ ID Nо: 54, идентичность последовательности между двумя сопоставленными последовательностями равна по меньшей мере × %. Такое сопоставление может быть выполнено публично доступной компьютерной программой гомологии, такой как программа “blastp”, обеспеченная на собственной странице NCBI на http://www.ncbi.nhn.nih.gov/BLAST/. Однако квалифицированному в данной области специалисту будет понятно, что в качестве домена трансдукции белков может быть также использован более длинный фрагмент белка ТАТ.
Термин в “функциональной связи” означает в данном контексте конфигурацию последовательностей нуклеиновых кислот настоящего изобретения, в которой одна последовательность помещена в положении относительно другой последовательности таким образом, что эти последовательности нуклеиновых кислот, после связывания, находятся в такой ориентации, что рамка трансляции кодируемых полипептидов не изменяется (т.е. эти молекулы нуклеиновых кислот связаны одна с другой «в рамке считывания»). Таким образом, полученная молекула нуклеиновой кислоты кодирует слитый белок в рамке считывания. Для достижения этого, последовательности нуклеиновых кислот настоящего изобретения могут быть организованы несколькими путями. Одна последовательность нуклеиновой кислоты может быть помещена при N-конце или при С-конце другой последовательности, либо связанной непосредственно, либо отделенной дополнительными линкерными нуклеиновыми кислотами, которые также находятся в рамке считывания с последовательностями нуклеиновых кислот этого изобретения. Предполагается также, что одна последовательность нуклеиновой кислоты является встроенной в последовательность другой нуклеиновой кислоты, при условии, что рамка считывания кодируемого полипептида остается интактной. Если желательно, чтобы последовательность, в которую помещена инсерция, все еще кодировала функциональный полипептид, биологическая активность рассматриваемого полипептида может оцениваться в соответствии с описанными в настоящем изобретении тестами.
Связывание последовательностей нуклеиновых кислот настоящего изобретения может быть достигнуто стандартными способами лигирования манипуляции ДНК, описанными в экспериментальном разделе настоящего изобретения и, например, Maniatis, T., Fritsch, E. F. & Sambrook, J. (1982) Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY). Обсуждается также другой способ лигирования, например, химическое лигирование.
В другом варианте осуществления, настоящее изобретение обеспечивает третью последовательность нуклеиновой кислоты в функциональной связи с последовательностью нуклеиновой кислоты, содержащей (i) первую последовательность нуклеиновой кислоты или (ii) первую и вторую последовательность нуклеиновой кислоты, где эта третья последовательность кодирует одну или несколько полипептидных последовательностей, подходящих для очистки белка. Конкретно, полипептидные последовательности, подходящие для очистки белка, включают в себя, но не ограничиваются ими, strep-метку и/или his-метку и/или GST-метку и/или интеиновую метку. Такие метки облегчают очистку белка аффинной хроматографией и хорошо известны в данной области (в отношении ссылки см., например, Terpe K., Appl. Microbiol. Biotechnol. (2003) 60(5):523-533). Эта третья последовательность нуклеиновой кислоты находится в функциональной связи с последовательностью нуклеиновой кислоты, содержащей первую последовательность нуклеиновой кислоты или первую и вторую последовательность нуклеиновой кислоты, т.е. помещена в положении либо на N-конце, либо на С-конце либо первой, либо второй последовательности нуклеиновой кислоты. Если (1) означает первую последовательность нуклеиновой кислоты, (2) означает вторую последовательность нуклеиновой кислоты и (3) означает третью последовательность нуклеиновой кислоты, возможные пути помещения этих последовательностей включают в себя (1)-(2)-(3), (1)-(3)-(2), (2)-(1)-(3), (2)-(3)-(1), (3)-(1)-(2) или (3)-(2)-(1). Эти последовательности нуклеиновых кислот либо могут быть лигированы друг с другом непосредственно, либо могут быть отделены друг от друга дополнительными линкерными нуклеиновыми кислотами. Лигирование с линкером или без линкера выполняют таким образом, чтобы привести все три последовательности нуклеиновых кислот в рамку считывания относительно друг друга и чтобы полученная молекула нуклеиновой кислоты кодировала слитый белок. Предполагается также, что одна последовательность нуклеиновой кислоты встроена в другую последовательность нуклеиновой кислоты, при условии, что рамка считывания кодируемого полипептида остается интактной. Если желательно, чтобы последовательность, в которую помещена инсерция, все еще кодировала функциональный полипептид, биологическая активность рассматриваемого полипептида может оцениваться в соответствии с описанными здесь тестами.
В дополнительном варианте осуществления, молекула нуклеиновой кислоты настоящего изобретения может дополнительно содержать четвертую последовательность нуклеиновой кислоты в функциональном связывании с последовательностью нуклеиновой кислоты, содержащей (i) первую последовательность нуклеиновой кислоты или (ii) первую и вторую последовательность нуклеиновой кислоты или (iii) первую и вторую и третью последовательность нуклеиновой кислоты, причем указанная четвертая последовательность кодирует один или несколько репортерных полипептидов. Подходящие репортерные полипептиды или репортерные молекулы делают возможными визуализацию и/или локализацию слитого полипептида, например, оптическими способами, такими как флуоресцентная микроскопия. Примерами подходящих репортерных полипептидов являются флуоресцентные белки, такие как GFP, CFP и YEP, или ферменты, способные образовывать детектируемую метку во вторичной реакции, такие как пероксидаза хрена, люцифераза или β-галактозидаза. Вместо репортерного полипептида, квалифицированный в данной области специалист будет также рассматривать мечение слитого белка этого изобретения другой детектируемой меткой, такой как флуоресцентные или люминесцентные органические молекулы (например, флуоресцеин, FITC или Cy5), или радиоактивной меткой.
Четвертая последовательность нуклеиновой кислоты находится в функциональной связи с последовательностью нуклеиновой кислоты, содержащей первую последовательность нуклеиновой кислоты или первую и вторую последовательность нуклеиновой кислоты или первую, вторую и третью последовательность нуклеиновой кислоты, т.е. помещена в положении либо на N-конце, либо на С-конце либо первой, либо второй, либо третьей последовательности нуклеиновой кислоты. Если (1) означает первую последовательность нуклеиновой кислоты, (2) означает вторую последовательность нуклеиновой кислоты и (3) означает третью последовательность нуклеиновой кислоты и (4) означает четвертую последовательность нуклеиновой кислоты, возможные пути помещения этих последовательностей включают в себя (1)-(2)-(3)-(4), (1)-(2)-(4)-(3), (1)-(3)-(2)-(4), (1)-(3)-(4)-(2). (1)-(4)-(2)-(3), (1)-(4)-(3)- (2), (2)-(1)-(3)-(4), (2)-(1)-(4)-(3), (2)-(3)-(1)-(4), (2)-(3)-(4)-(1), (2)-(4)-(1)-(3), (2)-(4)- (3)-(1), (3)-(1)-(2)-(4), (3)-(1)-(4)-(2), (3)-(2)-(1)-(4), (3)-(2)-(4)-(1), (3)-(4)-(1)-(2), (3)- (4)-(2)-(1), (4)-(1)-(2)-(3), (4)-(1)-(3)-(2), (4)-(2)-(1)-(3), (4)-(2)-(3)-(1), (4)-(3)-(1)-(2) или (4)-(3)-(2)-(1). Эти последовательности нуклеиновых кислот либо могут быть лигированы друг с другом непосредственно, либо могут быть отделены друг от друга дополнительными линкерными нуклеиновыми кислотами. Лигирование с линкером или без линкера выполняют таким образом, чтобы привести все четыре последовательности нуклеиновых кислот в рамку считывания относительно друг друга и чтобы полученная молекула нуклеиновой кислоты кодировала слитый белок. Предполагается также, что одна последовательность нуклеиновой кислоты встроена в другую последовательность нуклеиновой кислоты, при условии, что рамка считывания кодируемого полипептида остается интактной. Если желательно, чтобы последовательность, в которую помещена инсерция, все еще кодировала функциональный полипептид, биологическая активность рассматриваемого полипептида может оцениваться в соответствии с описанными здесь тестами.
В другом аспекте, это изобретение относится к полипептиду, кодируемому нуклеиновой кислотой этого изобретения. Способы получения полипептидов из молекул нуклеиновых кислот хорошо известны в данной области и описаны, например, Maniatis et al (supra).
Предполагается также получение слитого полипептида химическим лигированием двух или более фрагментов полипептида этого изобретения. Способы химического лигирования обеспечивают ковалентные связи между полипептидами посредством химического сшивания. Подходящими агентами сшивания являются например, бифункциональные агенты сшивания, такие как сульфо-MBS, сульфо-EMCS, сульфо-GMBS и другие агенты сшивания, доступные, например, из Pierce Chemical Company (Rockford, IL, USA).
В другом аспекте, настоящее изобретение обеспечивает вектор, содержащий молекулы нуклеиновых кислот в соответствии с изобретением. Таким вектором может быть плазмида, фагмида или космида. Например, молекула нуклеиновой кислоты может быть клонирована в прокариотический или эукариотический вектор способами, описанными в Maniatis et al. (supra). Предпочтительно, такой вектор способен экспрессировать полипептид, кодируемый молекулой нуклеиновой кислоты этого изобретения. Такие экспрессирующие векторы обычно содержат по меньшей мере один промотор и могут также содержать сигнал инициации трансляции и сигнал терминации трансляции или сигналы терминации транскрипции и полиаденилирования. Подходящими экспрессирующими векторами являются, например, вектор pET28 a+, вектор pUC18 или вектор pTRI-Ex-neo1.1, описанные в примерах ниже.
В другом аспекте, настоящее изобретение относится к клетке-хозяину, содержащей вектор по изобретению. Такой клеткой-хозяином может быть клетка не человека внутри или вне тела животного или клеткой человека внутри или вне тела человека. Другие подходящие клетки включают в себя прокариотические клетки, в частности, грамотрицательные или грамположительные бактериальные клетки. Особенно предпочтительными являются клетки E. coli или клетки Bacillus sp. Вектор может переноситься в клетку-хозяин различными способами, хорошо известными в данной области. Способы трансфекции клеток-хозяев и культивирования таких трансфицированных клеток-хозяев, а также условия продуцирования и получения полипептидов настоящего изобретения из таких трансформированных клеток-хозяев хорошо известны в данной области и описаны, например, в Maniatis et al. (supra).
В другом аспекте, настоящее изобретение относится к способу получения последовательностей нуклеиновых кислот или молекул нуклеиновых кислот настоящего изобретения, предусматривающему стадию (а) генерирования двухцепочечной молекулы нуклеиновой кислоты отжигом и удлинением первого набора подходящих олигонуклеотидных праймеров, где эти праймеры содержат последовательности нуклеиновых кислот настоящего изобретения; и (b) необязательное повторение стадии (а) со вторым, третьим или более набором подходящих олигонуклеотидных праймеров.
Подходящие олигонуклеотидные праймеры по настоящему изобретению конструируют делением представляющей интерес последовательности нуклеиновой кислоты на перекрывающиеся олигонуклеотиды, так что эти олигонуклеотиды содержат последовательности нуклеиновых кислот настоящего изобретения. Обычно диапазон перекрывания, охватываемый этими олигонуклеотидными праймерами, варьируется от 3 до 100 нуклеотидов, в частности, от 20 до 30 нуклеотидов. Второй, третий или более набор подходящих олигонуклеотидных праймеров может быть сконструирован таким образом, что каждый праймер второго, третьего или более наборов охватывает район желаемой последовательности нуклеиновой кислоты этого изобретения и может также содержать диапазон перекрывания одним или несколькими предыдущими праймерами. Предпочтительно, диапазон перекрывания варьируется между 3-100 нуклеотидами, в частности, между 20-30 нуклеотидами.
Для достижения отжига олигонуклеотидного праймера по настоящему изобретению с одним или несколькими другими олигонуклеотидными праймерами или частями последовательностей нуклеиновых кислот настоящего изобретения, должны использоваться условия и процедуры, которые хорошо известны в данной области. Квалифицированному в данной области специалисту будет понятно, что условия, которые должны быть выбраны для процедуры отжига, зависят от различных факторов, таких как длина праймера, длина перекрывающихся районов, содержание GC и т.д. После отжига нуклеотидная последовательность праймеров может быть удлинена хорошо известными способами ПЦР-реакции или другими способами, применимыми для синтеза ДНК. Можно сослаться, например, на Xiong A.S., et al. (2004), Nucl. Acids Res. 32(12):e98. Способ получения последовательности нуклеиновой кислоты по настоящему изобретению иллюстрирован, например, в примере 1 ниже.
В следующем аспекте, настоящее изобретение относится к способу получения полипептида, содержащего белок MeCP2 или биологически активный фрагмент белка MeCP2 или производное белка MeCP2 или фрагмента MeCP2, предусматривающему (a) трансформацию клетки-хозяина молекулой нуклеиновой кислоты по этому изобретению; (b) культивирование трансформированной клетки при условиях, которые делают возможной экспрессию молекулы нуклеиновой кислоты этого изобретения, с получением полипептида, содержащего белок MeCP2 или биологически активный фрагмент или производное белка MeCP2 или фрагмента; и (c) необязательное извлечение этого полипептида.
Способы трансформации клетки-хозяина молекулой нуклеиновой кислоты хорошо известны в данной области. Клетки-хозяева могут быть трансфицированы, например, такими способами, как кальций-фосфатная трансфекция, электропорация, трансфекция с использованием липофектина и т.д. Клетки-хозяева могут быть также трансформированы механическими способами, такими как микроинъекция ДНК, или с использованием таких векторов, как ретровирусы. Авторы ссылаются на Maniatis et al. (supra). Предпочтительно, клеткой-хозяином является прокариотическая клетка, более предпочтительно клетка E. coli. Условия культивирования таких трансфицированных клеток-хозяев, а также условия, позволяющие экспрессию молекул нуклеиновых кислот настоящего изобретения для получения полипептидов по настоящему изобретению, известны квалифицированному в данной области специалисту и зависят, например, от типа клетки-хозяина и типа вектора, используемых для трансформации клетки-хозяина. Примеры культивирования клеток-хозяев и выполнения экспрессии полипептидов этого изобретения приведены в разделе примеров ниже. Способы извлечения полипептида из трансформированной клетки-хозяина описаны, например, в Roe, S. (ed.), Protein Purification Applications: A Practical Approach. Oxford University Press (2001), Oxford.
В предпочтительном варианте осуществления, стадии (b) и (c) этого способа не включают в себя подвергание полипептида денатурирующим условиям. Денатурацией полипептида, в данном контексте, являются структурные изменения, вызываемые химическими или физическими действиями, которые приводят к полной или частичной потере биологической активности этого полипептида. Денатурация полипептида изменяет вторичную и третичную структуру или конформацию полипептида, который обычно сохраняет его первичную структуру. Денатурация может выполняться, например, изменениями температуры, рН, добавлением детергентов или солей, обработкой ультразвуком и т.д. В отличие от этой наиболее общей стратегии культивирования и извлечения полипептидов при денатурирующих условиях, например, для целей очистки, экстракция полипептидов при нативных условиях может предоставлять преимущество сохранения функциональной активности и уменьшения внутриклеточной деградации вследствие нарушенного фолдинга этих полипептидов. Такие нативно экстрагированные полипептиды настоящего изобретения имеют высокую эффективность трансдукции.
В предпочтительном варианте осуществления, полипептид присутствует после стадии (b) этого способа в концентрации, большей, чем 0,5, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 мг/л бактериальной культуры. Концентрация белка может быть определена количественно с использованием набора для анализа белка (Бредфорда) с использованием Кумасси (Pierce, Rockford, USA) согласно протоколу поставщика. В одном примере, концентрацию белка измеряют после элюции раствора пробы из аффинной колонки и после обессоливания. Обычно, концентрацию белка определяют против набора белков-стандартов, например, стандарта альбумина. Эти стандарты используют для калибрования фотометра, такого как Биофотометр Эппендорфа (измерение оптической плотности, например, при 595 нм). В одном примере, 100 мкл элюата белка смешивают с 5 мл «реагента Кумасси для анализа белка» (Pierce), уравновешивают при комнатной температуре в течение 10 минут и затем измеряют в спектрофотометре. Интегрированный алгоритм рассчитывает концентрацию пробы.
В другом аспекте, настоящее изобретение обеспечивает фармацевтическую композицию, содержащую молекулу нуклеиновой кислоты по настоящему изобретению, и/или полипептид по настоящему изобретению и фармацевтически приемлемый носитель. Квалифицированному в данной области специалисту будут известны подходящие фармацевтически приемлемые материалы-носители.
В другом аспекте, настоящее изобретение обеспечивает способ лечения нейродегенеративного или связанного с развитием нервного заболевания, предусматривающий введение полипептида или фармацевтической композиции по настоящему изобретению млекопитающему. Предпочтительно, этим млекопитающим является субъект-человек. Возможными способами введения являются внутривенный, внутрибрюшинный, внутриоболочечный, подкожный, ректальный, подъязычный, назальный, пероральный или трансдермальный способы.
Обычно, нейродегенеративным заболеванием является прогрессирующее нарушение, вызываемое повреждением и потерей нервных клеток. Связанное с развитием нервное заболевание вызывается патологическим отклонением пренатального и, в некоторых случаях, постнатального развития головного мозга. В частности, эти нейродегенеративные и связанные с развитием нарушения обусловлены уменьшением экспрессии MeCP2 или нарушенной функцией MeCP2. Нарушенная функция MeCP2 может быть идентифицирована по функциональным последствиям мутаций гена MECP2, как описано в Yusufzai T.M. and Wolffe A.P., Nucl. Acids Res. (2000), 28(21): 4172-4179, Kudo S. et al., Brain Dev. (2001) Suppl l:S165-73 и Ballestar E. et al., Hum. Genet. (2005) 116(1-2): 91-104. Обычно, мутации MBD (метилсвязывающего домена) изменяют способность MeCP2 связываться с ДНК или высвобождаться из ДНК и, следовательно, делают его нефункциональным. Мутации в TRD (домене репрессии транскрипции) обычно модифицируют или устраняют функцию репрессии транскрипции этого белка и приводят к уменьшению экспрессии MeCP2. В связи с предыдущим описанием, практикующий врач будет знать, получит ли пациент пользу от описанных в данном описании способов лечения и/или описанной терапии или подходит ли он для подвергания описанным способам лечения и/или описанной в данном описании терапии.
Более конкретно, связанным с развитием нервным заболеванием является синдром Ретта. Синдром Ретта является связанным с постнатальным развитием нервным заболеванием, характеризующимся прогрессирующей потерей умственных способностей, потерей тонких и более заметных двигательных навыков и способностей коммуникации. Замедление роста головы, а также развитие отличающейся картины стереотипичных движений рук являются обычными признаками синдрома Ретта. Были определены диагностические критерии, и они помогают при клинической диагностике (Hagberg B. et al. Eur. J. Paediatr. Neurol. (2002) 6(5):293-7). Инструменты для диагностики синдрома Ретта включают в себя скрининг на мутации в гене MECP2.
В другом варианте осуществления, фармацевтическую композицию вводят в подходящей дозе. Квалифицированному в данной области специалисту будут известны процедуры установления подходящей дозы для введения. Подходящая доза будет приводить к общему улучшению состояний млекопитающего, страдающего от нейродегенеративного или связанного с развитием нервного нарушения, вызываемого уменьшением экспрессии или нарушенной функцией MeCP2. Эффекты подходящей дозы могут включать в себя увеличение внимания пациента, уменьшение судорог, улучшенные двигательные навыки и симптомы, обычно ассоциированные с синдромом Ретта. Предпочтительно, эту фармацевтическую композицию вводят в дозе 0,01, 0,02, 0,03, 0,04, 0,05, 0,06, 0,07, 0,08, 0,09, 0,1, 0,2, 0,3, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 61, 68, 69, 70, 71, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100 мкг полипептида или последовательностей нуклеиновой кислоты настоящего изобретения на г массы тела млекопитающего. В другом варианте осуществления, фармацевтическую композицию настоящего изобретения вводят по меньшей мере один раз в день, по меньшей мере каждые 2, 3, 4 дня, 5, 6 или 7 дней.
В другом аспекте настоящего изобретения, обеспечено применение молекулы нуклеиновой кислоты по изобретению для получения полипептида, содержащего белок или биологически активный фрагмент MeCP2 или производное MeCP2 или фрагмента MeCP2. Примеры таких применений обеспечены в примерах ниже.
В следующем аспекте, настоящее изобретение относится к молекуле нуклеиновой кислоты или полипептиду, описанным выше, для применения в медицине и/или ветеринарии. В следующем аспекте, настоящее изобретение относится к применению молекулы нуклеиновой кислоты или полипептида настоящего изобретения для приготовления фармацевтической композиции для профилактики и/или терапии нейродегенеративного или связанного с развитием нервного заболевания, описанных выше. Более конкретно, связанное с развитием нервное заболевание обусловлено уменьшением экспрессии MeCP2 или нарушенной функцией MeCP2, как описано выше. В частности, связанным с развитием нервным заболеванием является синдром Ретта, описанный выше. Молекула нуклеиновой кислоты, полипептид или фармацевтическая композиция находится в форме, подходящей для введения млекопитающему. Предпочтительно, этим млекопитающим является субъект-человек. Возможными способами введения являются внутривенный, внутрибрюшинный, внутриоболочечный, подкожный, ректальный, подъязычный, назальный, пероральный или трансдермальный способы.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1. A. Эта фигура показывает стратегию создания синтетической последовательности, сообщенной как SEQ ID Nо: 3. Десять олигонуклеотидов были сконструированы в соответствии с кодон-оптимизированной последовательностью для Escherichia coli (стрелки). Синтез проходил посредством ПЦР-реакции в стадии 1 с образованием центрального фрагмента с использованием двух олигонуклеотидов с размерами 100 п.н., комплементарных на их конце в отношении 20 нуклеотидов (Mecp2_syn_5'_core_F и Mecp2_syn_5'_core_R). Конечный продукт имел размер 180 п.н. и представлен пунктирными продольными линиями. Затем аликвоту этого первого продукта использовали в качестве матрицы для второй ПЦР-реакции в стадии 2 с использованием двух фланкирующих олигонуклеотидов Mecp2syn_5'_F1 и Mecp2syn_5'_R1 с размерами 100 п.н., каждый. Эту реакцию продолжали сходным образом в стадии 3, 4 и 5. B. Конечный продукт этого синтеза представлен вместе с тремя сайтами рестрикции, использованными для последующих процедур клонирования. C. Схематическое представление синтетической ДНК, генерированной при помощи ПЦР с релевантными сайтами рестрикции, использованными для клонирования.
Фигура 2. Блок-схема генерирования конструкций MECP2. Эта схема показывает процедуры клонирования со всеми промежуточными стадиями, приводящими к конечным экспрессирующим векторам, содержащим синтетические гены для MeCP2e1 и MeCP2e2, слитые с доменом ТАТ.
Фигура 3. A. Приращение продуцируемого белка MeCP2e2, посредством оптимизации последовательности в соответствии с частотой использования кодонов Escherichia coli. Strep-меченые рекомбинантные слитые белки разделяли электрофорезом на 10% ДСН-ПААГ и окрашивали Кумасси синим. Количество белка, нанесенного на этот гель, соответствует равному количеству обработанной жидкой культуры бактерий. Дорожка 1, TAT-MeCP2e2-EGFP из кДНК человека; дорожка 2, TAT-hsMeCP2e2-EGFP из кДНК-последовательности MeCP2, частично оптимизированной в отношении частоты использования кодонов Escherichia coli. Дорожка 3, His-TAT-hsMeCP2 из кДНК-последовательности MeCP2, частично оптимизированной в отношении частоты использования кодонов Escherichia coli. Дорожка 4, His-TAT-sMeCP2, полученный из полного синтетического гена MECP2. Эта картина ясно показывает сильное приращение выхода белка с использованием кодон-оптимизированной кДНК-последовательности.
B.-D. Экспрессия и распределение MeCP2e2-EGFP in vitro. B. Верхний ряд: микрофотографии клеточной линии NIH3T3, стабильно экспрессирующей слитый белок TAT-MeCP2e2-EGFP. Приблизительно 40% этих клеток экспрессируют белок TAT-MeCP2e2-EGFP. Стрелки указывают на клетку (показанную при более высоком увеличении во вставках) с типичным гетерохроматическим распределением (DAPI) белка MeCP2e2. Нижний ряд: почти 100% клеток HeLa экспрессируют белок MeCP2e2. Локализация обычно в ядре без какого-либо окрашивания цитозольного белка. Вставки выявляют гетерохроматическое распределение MeCP2. C. Вестерн-блот 100 мкг белкового экстракта из нетрансфицированных контролей NHI3T3 и HeLa (wt) и соответствующих клеточных линий, стабильно экспрессирующих TAT-MeCP2e2-EGFP (st), с антителом против ацетилированного гистона Н3, поликлональным анти-MeCP2-антителом и анти-альфа-тубулин-антителом в качестве контроля нанесения. В обеих стабильных клеточных линиях этот слитый белок имеет молекулярную массу 100 кДа. D. Денситометрическое количественное определение сигналов Вестерн-блота показывает уменьшение ацетилирования гистона Н3 50-60% клеточных линий, сверхэкспрессирующих слитый белок TAT-MeCP2e2-EGFP, в сравнении с линиями дикого типа.
Фигура 4. A. Кинетика трансдукции и картина распределения TAT-MeCP2e2-EGFP в нескольких типах клеток. Клетки NIH3T3 инкубировали с 50 мкг/мл TAT-MeCP2e2-EGFP в течение 3 (a, b) или 12 часов, соответственно (c, d). Спустя 3 часа, эти клетки были только частично трансдуцированы и рекомбинантный белок был локализован в основном в цитоплазме, видимый в виде больших пятен (a, b). После 12 часов инкубации рекомбинантный белок трансдуцировал все клетки и локализовался в основном в физиологическом местоположении, ядре. e. Конфокальное изображение подтверждает, что этот белок в основном локализован в ядре. f. Рекомбинантный белок EGFP без TAT не был способен проникать в клетки при любом времени и он был виден только в виде редких внеклеточных пятен (после 12 часов инкубации). g и h. TAT-MeCP2e2-EGFP использовали также для испытания его эффективности трансдукции в первичных фибробластах пациента с синдромом Ретт с p.T158M. Появляющееся изображение указывает на то, что этот белок эффективно трансдуцировал все фибробласты. Эта локализация видна в основном в ядре, но некоторое количество белка все еще было в цитоплазме (вставка). i и j. Первичные нейроны гиппокампа из мышей дикого типа инкубировали со слитым белком TAT-MeCP2e2-EFGP. Белок MeCP2e2 является легко детектируемым в ядре.
B., C. Временная персистенция белка TAT-MeCP2e2 в трансдуцированных клетках NIH3T3. Культуры клеток NIH3T3 независимо инкубировали с одним и тем же количеством белка TAT-MeCP2e2. Затем собирали экстракты белка из этих обработанных культур клеток в разных временных точках. B. Вестерн-блот-анализ показывает присутствие этого рекомбинантного белка при 2, 24, 48, 72, 96 (едва видимый), но не при 120 часах после инкубации (дорожки 1-6, соответственно). C. Этот денситометрический анализ показывает уменьшение концентрации белка на протяжении времени. Максимальное падение приблизительно 77% наблюдали между 24 и 48 часами. Эту относительную величину рассчитывали взятием абсолютной величины в дорожке 1 за 100%. Затем величины в различных временных точках изменяли соответствующим образом.
Фигура 5. MeСР2 влияет на ацетилирование гистонов Н3 и H4K16. a. Экстракты белка из культур моноаллельных клеток, экспрессирующих MeCP2(2) дикого типа (wt) (1) и мутированный MeCP2 (2) (мутация p.T158M), анализировали на состояние ацетилирования гистона Н3 и H4K16. Независимые клеточные культуры линии моноаллельных клеток, экспрессирующих мутированный аллель, инкубировали, соответственно, с 300 пмоль рекомбинантных белков TAT-MeCP2e2 (3), MeCP2e2 (4) и TAT-EGFP (5) в течение 64 часов. Белок MeCP2 был детектируемым только в линии клеток, инкубированной с TAT-MeCP2e2 (3), но не с MeCP2e2. TAT-EGFP успешно трансдуцировал эти клетки и был детектирован анти-GFP-антителом (5). b. Нормализованный денситометрический анализ вестерн-блота с использованием тубулина в качестве стандарта для нанесения белка выявил слабое уменьшение неацетилированного гистона Н3 и гиперацетилирование гистонов H3 и H4K16 в линии моноаллельных клеток, экспрессирующей мутированный аллель (Mut), в сравнении с диким типом (WT) (белые столбцы в сравнении с черными столбцами). Инкубация с TAT-MeCP2e2 этой мутированной линии индуцировала уменьшение гиперацетилирования как H3, так и H4K16 и слабое приращение неацетилированной формы H3 (серые столбцы). Наблюдали незначимые изменения ацетилирования гистонов в независимых культурах клеток инкубацией с эквимолярной дозой MeCP2 без белков ТАТ и TAT-EGFP (наклонно и горизонтально заштрихованные столбцы). Эти результаты доказывают, что TAT-MeCP2e2 является биологически активным. c. Вестерн-блот экстрактов фибробластов wt-мыши (1), мыши Mecp2-/y (2) и MECP2-/y, инкубированных с 300 пмоль TAT-MeCP2e2 (3). Анти-MeCP2-антитело выявило эффективную трансдукцию фибробластов этим рекомбинантным белком. Тубулин использовали в качестве стандарта. d. Нормализованный денситометрический анализ с использованием тубулина в качестве стандартного белка Вестерн-блота, показанный в c., выявил уменьшение формы неацетилированного гистона Н3, отсутствие приращения ацетилированной формы гистона Н3, но приращение ацетилированной формы гистона H4K16 of Mecp2-/y (ko) в сравнении с мышами дикого типа (wt) (белые столбцы в сравнении с черными столбцами). Инкубация культуры клеток фибробластов из мышей Mecp2-/y с TAT-MeCP2e2 (ko +) индуцировала приращение неацетилированного гистона Н3 (серые столбцы) и уменьшение уровня гиперацетилирования H4K16.
Фигура 6. MeCP2 может быть нацелен на нейроны центральной нервной системы (ЦНС) in vivo. a. Иммуноблоттинг с анти-MeCP2-антителом 50 мкг лизата белка головного мозга трех мышей wt, умерщвленных спустя 20 часов после независимой внутрибрюшинной инъекции белков TAT-MeCP2e2-EGFP (дорожка 1), TAT-MeCP2e2 (дорожка 2) или MeCP2e2 без TAT (дорожка 3). В дорожке 1 можно хорошо видеть присутствие TAT-MeCP2e2-EGFP по более высокой молекулярной массе относительно эндогенного белка Mecp2. Мышь, инъецированная TAT-MeCP2e2, обнаруживала гораздо более сильный сигнал белка MeCP2e2 (дорожка 2) как в дорожке 1, так и в дорожке 3, где уровень эндогенного Mecp2 не отличается в сильной степени. b. Иммунофлуоресценция GFP в головном мозге мыши дикого типа, инъецированной белками TAT-MeCP2e2-EGFP и EGFP без TAT спустя 20 часов после инъекции. Сигнал GFP локализован в ядрах почти всех клеток в коре, мозжечковых нейронах зернистого слоя и наружных мозговых оболочках в мышах, инъецированных TAT-MeCP2e2-EGFP (1-6). EGFP является недетектируемым в головном мозге инъецированных EGFP мышей (7 и 8). c. Иммунофлуоресценция двух животных Mecp2-/y, инъецированных в каждом случае одной изоформой рекомбинантных белков, с использованием поликлонального анти-MeCP2-антитела. Первый и второй ряды изображают срезы коры мыши Mecp2-/y, обработанной в течение 20 дней рекомбинантным TAT-Mecp2e1 (1,2,3) и e2 (4,5,6). Присутствие MeCP2 в основном в ядрах этих клеток ясно видно во вставках. Срез головного мозга одной неинъецированной мыши MeCP2+/y (7,8,9) и одной мыши MeCP2-/y mice (10,11) служили в качестве контролей.
Фигура 7. A. Эксперименты по коиммунопреципитации в культурах фибробластных клеток животных Mecp2-/y, трансдуцированных TAT-MeCP2e1, показывают рекрутинг HDAC1. В верхнем ряду находится Вестерн-блот фракций иммунопреципитированных белков с использованием поликлонального анти-MeCP2-антитела ("+" преципитированная фракция в присутствии и "-" в отсутствие анти-MeCP2-антитела). В нижнем ряду находится Вестерн-блот-анализ из тех же самых преципитатов с использованием анти-HDAC1-антитела. Дорожки 1 и 2: фракция преципитированного белка из животного дикого типа. Дорожки 3,4 и дорожки 5,6: фракция преципитированного белка из фибробластов мышей Mecp2-/y, трансдуцированных TAT-MeCP2e1 и TAT MeCP2e2, соответственно. Дорожки 7 и 8: Фракции преципитированного белка из фибробластов животного Mecp2-/y. В этих фракциях иммунопреципитации присутствие обоих MeCP2, а также HDAC1 видно во фракции положительного контроля (дорожка 1), а также во фракциях клеток, трансдуцированных обеими изоформами TAT-MeCP2 (дорожки 3 и 5). Не наблюдали детектируемого Mecp2, а также белка HDAC1 в преципитированной фракции отрицательных контролей (дорожка 7). Иммунопреципитаты в отсутствие анти-MeCP2 не обнаруживали присутствия ни MeCP2, ни HDAC1.
B., C. Изоформы TAT-MeCP2 модулируют гиперацетилирование Н3 и H4K16 в головном мозге. B. Mecp2-недостаточных мышей инъецировали i.p. в течение 14 дней PBS (дорожка 2), 20 мкг/г (дорожка 3) или 30 мкг/г (дорожка 4) TAT-MeCP2e1. 50 мкг экстрактов белка головного мозга из всех инъецированных мышей (2-4) и неинъецированной мыши дикого типа (1) иммуноблоттировали с антителами против гистона Н3 (Н3), ацетилированного гистона Н3 (Н3), ацетилированного гистона H4K16 and MeCP2. Анти-альфа-тубулин-антитело использовали в качестве контроля нанесения. Мыши Mecp2-/y не обнаруживали никакого детектируемого Mecp2 или рекомбинантного белка (дорожки 2, 3 и 4). Не детектировали видимого гиперацетилирования ни Н3, ни H4K16 в инъецированных PBS MeCP2-недостаточных мышах в сравнении с мышами дикого типа. Однако инъекция рекомбинантного белка уменьшает ацетилирование гистона H4K16. Не наблюдали сильного различия для ацетилированного и неацетилированного гистона H3. C. Денситометрический анализ, показывающий зависимую от дозы биохимическую активность белка MeCP2. Аналогичные эксперименты выполняли с белком TAT-MeCP2e2 с постоянными сходными результатами. Эта относительная величина представляет денситометрические величины, выраженные в виде процента величины животных дикого типа (wt), взятой за 100%.
Фигура 8. Доставляемые в головной мозг изоформы MeCP2 продлевают время жизни и элиминируют патологию нейронов в мышах MeCP2-/y. a. Карта Каплана-Майера, изображающая кривые выживания необработанных мышей MeCP2-/y (n=11), ложнообработанных (n=6) или обработанных TAT-MeCP2el (n=5) или TAT-MeCP2e2 (n=5), соответственно. b. Двусторонний критерий Стьюдента выявил величину р<0,05 вследствие величины альфа 0,05 для обеих групп, неинъецированных/ложноинъецированных в сравнении с TAT-MeCP2e1 и в сравнении с TAT-MeCP2e2. Не было статистически значимого различия между группами TAT-MeCP2e1/TAT-MeCP2e2 (p>0,8), а также группами неинъецированных/ложноинъецированных мышей. c. Niss1-окрашивание нейронов гиппокампа CA2 выявило более мелкие и более плотно упакованные нейроны в мышах MeCP2-/y mice (1) в сравнении с мышами дикого типа. (2). В противоположность этому, обработка TAT-MeCP2el (3) обращала деструкцию нейронов в мышах Mecp2-/y. В (4) показано изображение гиппокампа мыши, обработанной TAT-MeCP2e2, который не изменяет размер нейронов. d. Количественное определение клеточного спасения в нейронах гиппокампа CA2 (n=3 для дикого типа (wt), n=3 для Mecp2-/y, n=4 для TAT-MeCP2e1-обработанных мышей и n=3 для TAT-MeCP2e2).
ПРИМЕРЫ
Следующие примеры предназначены для дополнительной иллюстрации, но не для ограничения настоящего изобретения. Эти примеры содержат технические признаки, и должно быть понятно, что это изобретение относится также к комбинациям технических признаков, представленных в этом разделе примеров.
1. Генерирование конструкций
Для установления, будет ли кодон-оптимизированная последовательность индуцировать улучшение продуцирования белка, использовали двухстадийную стратегию в планировании оптимизации кодонов. Сначала, оптимизировали только половину кодирующей последовательности MECP2, как описано ниже, отбором кодонов с частотой использования в E. coli по меньшей мере 8-10%. Затем выполняли оценку улучшенной экспрессии. Значимое приращение выхода белка с «полусинтетическим» MeCP2 подтверждало валидность этой стратегии, и во второй стадии, эту последовательность делали полностью кодон-оптимизированной, как иллюстрировано на фиг. 3А. Перекрывающийся район праймеров имел обычно длину 20 нуклеотидов с температурой плавления обычно по меньшей мере 50°С, но во многих случаях требовалось создание более длинных перекрывающихся районов для гарантии эффективного отжига во время ПЦР-опосредованного синтеза. Блок-схема для генерирования следующих конструкций приведена на фигуре 2.
1. Конструировали два олигонуклеотида, комплементарных на их 3'-конце 20 основаниям (TAT-core-F и TAT-core-R, SEQ ID Nо: 7 и SEQ ID Nо: 8). Затем двадцать пмоль каждого олигонуклеотида удлиняли с использованием полимеразы Pfu (Stratagene, La Jolla, USA) при следующих условиях: 15 мин денатурация при 96°C, с последующей одной стадией удлинения при 60°C в течение 30 минут. Затем этот продукт клонировали в вектор pBlueScript (Stratagene, La Jolla, USA) и секвенировали. Затем использовали два дополнительных праймера (TAT-new-F и TAT-new-R, SEQ ID Nо: 9 и SEQ ID Nо: 10) для амплификации предыдущего инсерта с получением продукта с сайтами рестрикции EcoRI и HindIII на 5'- и 3'-геномных концах, соответственно. Затем этот продукт, расщепленный EcoRI и HindIII, субклонировали в вектор puc18 (Invitrogen, Karlsruhe, Germany) с получением конструкции pTAT-Strep. кДНК, кодирующая TAT- и Strep-метку II, содержащиеся в конструкции pTAT-Strep, приведена в SEQ ID Nо: 5 (стадия 1, фиг. 2).
2. Клонирующую EGFP последовательность вырезали из вектора pEGFP-C1 (Clontech, CA, USA) и клонировали в конструкцию pTAT-Strep, названную puc18-TAT-EGFP-Strep (стадия 2, фиг. 2).
3. кДНК-последовательность гена MECP2 амплифицировали из клона pCR-hMeCP2 (W. Stratling, Universitatsklmik Eppendorf, Hamburg, Germany) полимеразой Pfu с использованием праймеров NotI-3G-MeCP2-F и R-MeCP2-3G-NcoI (SEQ ID NO:11 и SEQ ID NO:12). Этот ПЦР-продукт конструировали с последовательностями для сайтов рестрикции NotI и NcoI на 5'- и 3'-геномных концах и клонировали в puc18-TAT-EGFP-Strep. Эта произведенная конструкция была pTAT-MeCP2e2-EGFP-Strep (стадия 3, фиг. 2).
4. Эукариотический экспрессирующий вектор ρTri-EX-TAT-MeCP2e2-EGFP-Strep-His (стадия 4, фиг 2) генерировали амплификацией последовательности TAT-MeCP2e2-EGFP-Strep из конструкции pTAT-MeCP2e2-EGFP полимеразой Pfu с использованием праймеров BspHI-TAT-F (SEQ ID Nо: 38) и XhoI-EGFP-R (SEQ ID Nо: 39). Этот продукт ПЦР расщепляли BsphI/XhoI и клонировали в вектор pTRI-Ex-Neo.1.1 (Novagen, Darmstadt, Germany), линеаризованный NcoI/XhoI.
5. Стратегию, сходную со стратегией генерирования конструкции 1, использовали для генерирования конструкции pET28-PTD4-6his с использованием олигонуклеотидов PTD4_core_F (SEQ ID Nо: 40), PTD4_core_R (SEQ ID Nо: 41), PTD4_amp_JF (SEQ ID Nо: 42) и PTD4_amp_R (SEQ ID Nо: 43) с получением последовательности, кодирующей пептид PTD4 (Ho A et al. Cancer Res. 2001 Jan 15; 61(2):474-7.) Полученный продукт расщепляли с использованием NcoI и XhoI и клонировали в вектор pET28a(+), расщепленный аналогичным образом (Novagen, Merck, Darmstadt, Germany) с получением плазмиды pET 28-PP4-6His (стадия 5, фиг. 2). Генерированная кодирующая последовательность, содержащаяся в плазмиде pET 28-PP4-6His, приведена в SEQ ID Nо: 6.
6. Фрагмент NdeI/BspeI из pTAT-MeCP2e2-EGFP-Strep клонировали в pET28-PP4-6His, расщепленную NdeI и AgeI. Полученная конструкция была названа pET28-TAT-MeCp2e2-EGFP-6His (стадия 6, фиг. 2).
7. Фрагмент NotI/XhoI из ρTri-EX-MECP2-EGFP-Strep-6His вырезали и клонировали в pET28-TAT-MeCP2e2-EGFP-6His, расщепленную теми же самыми рестрикционными эндонуклеазами. Полученной конструкцией была ρET28-TAT-MeCP2e2-EGFP-Strep-6His.
8. Часть последовательности MeCP2 человека амплифицировали с использованием праймеров Mecp2-His-F1 (SEQ ID Nо: 44) и Mecp2_His_rev (SEQ ID Nо: 45). Этот продукт клонировали в pBluescript с получением плазмиды pBlue-6His (стадия 8, фиг. 2).
9. Фрагмент NdeI/DraI, содержащий последовательность 6 His и часть кодирующей MeCP2 последовательности, высвобождали из pBlue-фрагмента, содержащего His-метку, и клонировали в pET28-TAT-MECP2e2-EGFP-Strep-6His, расщепленную аналогичным образом. Эта генерированная плазмида была названа pET28-6HisTAT-MeCp2e2-EGFP-Strep-6His (стадия 9, фиг 2).
10. Фрагмент XhoI и DraIII из pET28-6HisTAT-MeCp2e2-EGFP-Strep-6His вырезали из этого вектора. Затем тот же самый фрагмент, но без последовательности, кодирующей последовательность 6His, амплифицировали с использованием праймеров Mecp2_pet_XhoI (SEQ ID Nо: 46) и Mecp2_pet_DraIII (SEQ ID Nо: 47). Затем этот продукт ПЦР использовали для воспроизведения этого фрагмента в pET28-6HisTAT-MECP2e2-EGFP-Strep-6His. Полученной плазмидой была pET28-6HisTAT-MECP2e2-EGFP-Strep (стадия 10, фиг. 2).
1.1 Генерирование синтетических генов:
Попытки получения рекомбинантного белка с использованием вектора ρET28-TAT-MECP2e2-EGFP-Streρ-His, содержащего кДНК MECP2 человека, были неудовлетворительными. Выход белка был равен приблизительно 0,1 мг/л бактериальной культуры. Для преодоления этой недостаточной экспрессии в Escherichia coli конструировали синтетический MeCP2, оптимизированный для этого конкретного организма для обхода ограничений неоднозначности кодонов. Аминокислотную последовательность обеих изоформ е1 и е2 MeCP2 человека обратно транслировали в соответствии с предпочтительной частотой использования кодонов Escherichia coli. GC-содержание этих кДНК-последовательностей, которое может быть также дополнительным источником ограничения продуцирования белка, также уменьшали. Затем эту искусственную последовательность создавали ПЦР-опосредованным синтезом с использованием ряда перекрывающихся нуклеотидов. Авторы изобретения синтезировали кодирующую последовательность MECP2e2 в двух отдельных реакциях с использованием ДНК-полимеразы Pfu с ультравысокой точностью (Stratagene, La Jolla) с 18 комплементарными олигонуклеотидами. В первой стадии ряд из 10 олигонуклеотидов (SEQ ID Nо: 13 - SEQ ID Nо: 22) генерировал последовательность SEQ ID Nо: 3 из 810 п.н., содержащую также последовательность, кодирующую домен ТАТ и сайты рестрикции для субклонирования. Основная стратегия для этого синтеза представлена в общих чертах на фигуре 1. В первой стадии два внутренних (core) олигонуклеотида, имеющих комплементарность 20 п.н., отжигали и удлиняли. Дополнительное удлинение этого синтетического гена проводили с использованием других комплементарных олигонуклеотидов с генерированным ПЦР-продуктом в качестве матрицы.
Компоненты реакции (для 50 мкл):
Условия термоциклера для генерирования первого фрагмента:
Первая стадия: начальная денатурация 96°C в течение 30 с; с последующими 10 циклами: 96°C в течение 20 с; 55°C в течение 30 с; 72°C в течение 1 мин;
Вторая стадия: 20 циклов: 96°C в течение 30 с; 72°C в течение 1 мин; конечное удлинение при 72°C в течение 5 мин.
Полученный продукт очищали пропусканием через колонку Montage (Millipore) PCR clean-up.
Для следующих стадий реакции 1 мкл очищенного продукта ПЦР и 10 пмоль каждой из следующих пар праймеров использовали при следующих условиях: начальная денатурация 96°C в течение 2 мин, с последующим 25 циклами 96°C в течение 20 с; 55°C в течение 30 с; 72°C в течение 1 мин. Каждую последующую стадию амплификации выполняли при одних и тех же условиях с использованием введения ДНК предыдущего генерированного фрагмента с приращением времени удлинения 10 с для дополнительных стадий амплификации (фиг. 1).
11. Генерированную последовательность клонировали в pBlueScript и проверяли на ошибки последовательности. Один клон, не содержащий ошибок, был отобран и назван pBlue-MECP2e2-syn (стадия 11, фиг. 2).
12. Фрагмент EcoRI/XcmI вырезали из pBlue-MECP2e2-syn и клонировали в pTAT-MECP2e2-EGFP-Strep, предварительно расщепленную теми же самыми рестрикционными ферментами. Полученная плазмида была названа ρTAT-MECP2e2-syn-EGFP-Strep (стадия 12, фиг 2).
13. Фрагмент NotI/SacI из ρTAT-MECP2e2-syn-EGFP-Strep вырезали и клонировали в pET28-6His-TAT-MECP2-EGFP-Strep, расщепленную аналогичным образом. Полученной плазмидой была pET28-His-TAT-MECP2e2(half)syn-EGFP-Strep (стадия 13, фиг. 2).
14. EGFP вырезали из вектора pET28-His-TAT-MECP2e2(half)syn-EGFP-Strep расщеплением SacII and BsrGI. ПЦР-продукт, генерированный с использованием обратного праймера Mecp2_BsrGI (SEQ ID Nо: 31) и прямого праймера Mecp2_1282_1306 (SEQ ID Nо: 32), регенерирующий частично делетированный MeCP2, получали и клонировали в сайты рестрикции SacII/BsrgI pET28-His-TAT-MECP2e2(half)syn-EGFP-Strep. Полученный вектор был назван pET28-His-TAT-MECP2e2(half)syn-Strep, который не кодировал EGFP (стадия 16, фиг. 2).
Второй набор из 8 олигонуклеотидов (SEQ ID Nо: 23 - SEQ ID Nо: 30) использовали для генерирования последовательности SEQ ID Nо: 4 с использованием сходной стратегии, изображенной на фигуре 1 и описанной выше.
15. Этот продукт ПЦР был клонирован в pBluescript и был назван pBlue-MECP2e2-syn_3part (стадия 14, фиг. 2).
16. XcmI/BsrGI из pBlue-MECP2e2-syn_3part вырезали из этого вектора и клонировали в ρTAT-MECP2e2-syn-EGFP-Strep с получением ρTAT-MECP2e2-syn. Этот вектор содержит полностью оптимизированную последовательность, показанную в SEQ ID Nо: 2 (стадия 15, фиг. 2).
17. Полную синтетическую последовательность MeCP2e2, содержащуюся в pTAT-MECP2e2-syn, вырезали с использованием NotI/BsrGI и клонировали в расщепленный таким же образом вектор pET28-His-TAT-MECP2e2(half)syn-Strep. Полученный фрагмент содержит полную синтетическую последовательность MeCP2 и назван pET28a-6His-TAT-MECP2e2-syn-Strep, причем он является одной из конструкций, использованных для крупномасштабного получения белка.
18. Продукт ПЦР генерировали с олигонуклеотидами с использованием MeСР2b_syn_F(SEQ ID Nо: 33), MeСР2b_syn_core (SEQ ID Nо: 34) и MeСР2b_syn_R (SEQ ID Nо: 35), содержащие последовательности изоформы MeCP2e1.
Компоненты реакции (для 50 мкл):
Условия термоциклера для генерирования первого фрагмента:
Первая стадия: начальная денатурация 96°C в течение 30 с; с последующими 10 циклами: 96°C в течение 20 с; 55°C в течение 30 с; 72°C в течение 1 мин;
Вторая стадия: 20 циклов: 96°C в течение 30 с; 72°C в течение 1 мин; конечное удлинение при 72°C в течение 5 мин.
Один мкл из генерированного ПЦР-фрагмента использовали в качестве введенной ДНК для следующей реакции:
Компоненты реакции (для 50 мкл):
Праймеры: Mecp2b_syn_F/Mecp2b_syn_R (10 пмоль/мкл) 1 мкл каждого
Один мкл ПЦР-продукта из первой ПЦР 1 мкл
Смесь dNTP (2,5 мМ каждого нуклеотида/мкл) 8 мкл
10X буфер для ДНК-полимеразы 5 мкл
ДНК-полимераза Ultra Pfu (2,5 Единиц/мкл) 2 мкл
dd Н2О 32 мкл
Начальная денатурация: 96°C в течение 1 мин.
с последующими 30 циклами: 96°C в течение 20 с; 60°C в течение 30 с; 72°C в течение 30 с;
конечная стадия удлинения: 72°C в течение 5 мин.
Этот ПЦР-фрагмент заменял фрагмент от сайта EagI до сайта PstI pET28a-6His-TAT-MECP2e2syn с генерированием pET28a-6His-TAT-MECP2e1-syn-Strep (стадия 18, фиг. 2).
19. Затем ПЦР-продукт, генерированный с использованием праймеров Mecp2-syn-NdeI_F (SEQ ID Nо: 36) и Mecp2-syn-BsrGI_R (SEQ ID Nо: 37), амплифицированный с получением полной синтетической последовательности MECP2, но без домена ТАТ, клонировали в расщепленный аналогичным образом вектор pET28-His-TAT-MECP2e2(half)syn-Strep. Полученная плазмида была названа pET28a-6His-MECP2e2-syn-Strep (стадия 19, фиг. 2).
2. Экспрессия и очистка этих белков
Решающими стадиями в этом исследовании были стадии эффективного получения рекомбинантных белков. Как описано выше, авторы изобретения создали синтетическую кДНК-последовательность MECP2 как для изоформы е1, так и для изоформы е2 в соответствии с частотой использования кодонов Escherichia coli для увеличения выхода рекомбинантных белков (фиг. 3 A). Несколько исследований in vitro, показанных ниже, демонстрировали биохимическую активность слитых изоформ TAT-MeCP2 несмотря на присутствие дополнительных доменов (см. также фиг. 5 и фиг. 7). В противоположность наиболее обычной стратегии экстракции белков, слитых с доменами трансдукции, при денатурирующих условиях, авторы настоящего изобретения решили экстрагировать рекомбинантные белки при нативных условиях для сохранения полностью их функциональной активности и для уменьшения внутриклеточной деградации вследствие неправильного фолдинга. Эти эксперименты in vitro, показанные ниже, указывали на то, что эти нативно экстрагированные белки имеют высокую эффективность трансдукции.
Для экспрессии белков штамм Escherichia coli 2 (DE3) (Novagen) трансформировали плазмидными векторами электропорацией. Единственную колонию инокулировали в 10 мл LB, дополнительной 50 мкг/мл канамицина, и выращивали в течение ночи при 37°С при встряхивании. Эту предварительную культуру разбавляли 1:25 и выращивали в 250 мл LB, дополненной 50 мкг/мл канамицина, в течение 5 часов при 37°С и затем выдерживали при 4°С в течение одного часа. Эту культуру дополнительно разбавляли 1:10 в 2,5 л обогащенной среды (2,5 г мясного экстракта на литр LB), дополненной 50 мкг/мл канамицина, и индуцировали 1 мМ IPTG в течение приблизительно 12 часов при 30°C. Эти клетки собирали центрифугированием при 7000g в течение 10 мин при 4°С. Осадок суспендировали в буфере для лизиса бактерий (15 мл/л осадка культуры) с 100 мкг/мл PMSF, 1 мкл/мл коктейля (смеси) ингибиторов протеаз (CALBIOCHEM), лизоцимом 500 мкг/мл, 1 Единицей/мл бензоназы (Merck Biosciences) и лизировали с использованием вортекса. Лизат инкубировали при комнатной температуре в течение 10 минут, затем на льду в течение 15 минут перед подверганием обработке ультразвуком (рабочие циклы 50-60 с, 5-6 выходных сигналов) 6 раз, по 1 минуте в каждом случае с интервалом 2 минуты во избежание нагревания. Лизат центрифугировали при 18000xg в течение 25 мин, 4°C, затем супернатант фильтровали с использованием фильтров 0,45 мкм (Millipore) и очищали пропусканием через аффинные колонки со стрептактином (IBA, Goettingen, Germany) с использованием протокола поставщика. Затем этот белок подвергали смене буфера с использованием обессоливающей колонки PD-10 (GE Helthcare, Freiburg, Germany). Состав буфера был следующим: 20 мМ HEPES, 300 мМ NaCl, 0,1 мМ CaCl2 и 10% глицерин. Бактериальные липополисахариды удаляли с использованием колонок EndoTrap (Profos, Regenburg, Germany) в соответствии с протоколами поставщика после уравновешивания колонок описанным ранее буфером.
3. Культивирование клеток и трансдукция слитых белков
3.1 Линии клеток
Клетки NIH3T3 (мышиный фибробласт, ATCC CCL 92) культивировали в модифицированной по способу Дульбекко среде Игла (PAN Biotech), содержащей 10% фетальную телячью сыворотку (PAN Biotech) и антибиотики пенициллин (100 Е/мл), стрептомицин (100 мкг/мл) (PAN Biotech) при 37°C с 5% CO2.
Фибробласты из кожной биопсии пациента-женщины с синдромом Ретта, несущей одну из наиболее частых мутаций гена (T158M), культивировали в среде RPMI 1640 с 15% фетальной телячьей сывороткой, пенициллином (100 Е/мл), стрептомицином (100 мкг/мл) и L-глутаматом (100 нг/мл) при 37°C с 7% CO2.
Фибробласты из мышей Mecp2-/y устанавливали в культуре из постнатального дня 7 (P7), с использованием задней конечности/кончика хвоста. Эту ткань промывали DPBS и измельчали на малые кусочки стерильным скальпелем в чашке 35 мм, содержащей среду, под током воздуха в ламинарном боксе. Среду осторожно аспирировали и заменяли новой средой, и эту чашку инкубировали в течение недели без замены среды при 37°C с 10% CO2, затем эти фибробластные клетки поддерживали в модифицированной по способу Дульбекко среде Игла, содержащей 20% фетальную телячью сыворотку и антибиотики пенициллин (100 Е/мл), стрептомицин (100 мкг/мл) при 37°C с 7% CO2.
Головной мозг детенышей постнатального дня 7 (Р7), иссеченный под ламинарным током воздуха, использовали для выделения гиппокампа и коры головного мозга для культивирования нейронов в соответствии с опубликованным протоколом (Ray, J., et al., 1993, PNAS 90:3602-6).
3.2 Установление стабильных клеточных линий, экспрессирующих белок TAT-MeCP2e2-EGFP
Для установления стабильных клеточных линий, экспрессирующих TAT-MeCP2-EGFP, авторы изобретения использовали NIH3T3 (мышиные фибробласты) и клетки HeLa (клетки карциномы шейки матки человека). Клетки HeLa культивировали в среде MEM-альфа (PAN Biotech, Aidenbach, Germany), с 10% фетальной телячьей сывороткой, пенициллином (100 Е/мл), стрептомицином (100 мкг/мл) и L-глутаматом (100 нг/мл). NIH3T3 культивировали в DMEM с 10% фетальной телячьей сывороткой, пенициллином (100 Е/мл), стрептомицином (100 мкг/мл). Клетки переносили с pTriEx-TAT-MeCP2e2-EGFP-Streρ-His с использованием реагента трансфекции «смеси генов» в соответствии с протоколом поставщиков (Novagen). Спустя два дня после трансфекции клетки собирали на среде, содержащей 500 мкг/мл неомицина (GIBCO), в течение 6 недель и после этого поддерживали на среде с 200 мкг/мл неомицина. Делали фотографии растущих клеток для одного дня в двухкамерных культуральных предметных стеклах (Falcon, BD bioscience, Heidelberg, Germany). Эти клетки промывали и монтировали препараты со средой для монтирования препаратов Vector shield mounting medium, содержащей DAPI (Vector laboratories, Burlingame, CA), и распределение флуоресценции анализировали с использованием микроскопа Olympus BX60 (Olympus Optical Co. LTD, Japan) при увеличении 200x в пяти разных полях зрения с использованием программного обеспечения SIS-анализа (Soft Imaging System GmbH, Mϋnster).
3.3 Установление линии моноаллельных первичных клеток пациента с синдромом Ретта
Фибробласты пациента трансфицировали плазмидой pCI-neo-hTERT (Counter, CM. et al., 1998, Oncogene 16:1217-22 и Lundberg, A.S. et al.,2002, Oncogene 21:4577-86), содержащей hTERT (субъединицу обратной транскриптазы теломеразы человека), кодирующую ген, с использованием реагента трансфекции SuperFect в соответствии с протоколом поставщика (Qiagen, Hilden, Germany). Трансфицированные клетки отбирали в течение 6 недель в G410 (200 мкг/мл) (Calbiochem, San Diego, USA) содержащей среде и после этого поддерживали на среде с 100 мкг/мл G418. Для установления линии моноаллельных экспрессирующих клеток, клетки трипсинизировали и разбавляли средой, отдельные клетки микропипетировали при прямой визуализации под микроскопом, отдельные клетки переносили на 24-луночные планшеты, содержащие отфильтрованную кондиционированную среду из экспоненциально растущих первичных клеток, дополненную 20 нг/мл bFGF (основного фактора роста фибробластов) (Sigma, Deisenhofen, Germany). После слияния лунки, клетки трипсинизировали и размножали. Тотальную РНК экстрагировали с использованием набора RNeasy (Qiagen, Hilden, Germany) и кДНК синтезировали с использованием обратной транскриптазы Superscript-II и праймера Oligo (dT) 12-18 (Invitrogen). Район, содержащий мутацию, анализировали при помощи ОТ-ПЦР с использованием фланкирующих праймеров (Rett_EX3-F и Rett_EX4-R, SEQ ID Nо: 48 и SEQ ID Nо: 49). Клональность подтверждали анализом последовательности продуктов ОТ-ПЦР.
4. Трансдукция слитых белков и анализ трансдуцированных клеток
Для трансдукции белков TAT-MeCP2-EGFP и EGFP, клетки выращивали до конфлюэнтности в двухкамерных культуральных предметных стеклах (Falcon, BD Bioscience, USA). Эти культуры обеспечивали свежей средой и затем обрабатывали разными концентрациями слитых белков (500-1500 пмоль/мл) в течение различных интервалов времени. Трансдукцию TAT-MeCP2e1 и TAT-MeCP2e2, а также белков MeCP2 проводили после их мечения NHS-родамином в соответствии с протоколом изготовителя (Pierce biotechnology, IL, USA), с последующей гель-фильтрацией на колонке PD-10 для удаления не включившегося родамина. Для визуализации распределения флуоресценции эти клетки промывали осторожно холодным PBS и фиксировали в свежеприготовленном 4% параформальдегиде в PBS в течение 5 мин при комнатной температуре. Клетки снова промывали PBS перед монтированием препарата со средой для монтирования Vector shield mounting medium, содержащей DAPI (Vector laboratories, Burlingame, CA), и анализировали с использованием микроскопа Olympus BX60 (Olympus Optical Co. LTD, Japan) при увеличении 200x в пяти разных полях зрения с использованием программного обеспечения SIS-анализа (Soft Imaging System GmbH, Mϋnster), а также получали конфокальные изображения с использованием лазерного сканирующего микроскопа Leica TCS SP2 AOBS.
5. Создание клеточных линий, стабильно сверхэкспрессирующих белок MeCP2e2, и успешная трансдукция первичных клеток
Авторы установили клеточные линии HeLa и NIH3T3, стабильно экспрессирующие слитый белок TAT-MeCP2e2-EGFP, меченный на С-конце аффинными метками Strep-II и 6-His, с использованием конструкции ρTri-Ex-MECP2-EGFP-Strep-His. После трансфекции клеток этой конструкцией белок был преимущественно локализован в гетерохроматических районах ядра, как особенно очевидно для линии клеток мыши NIH3T3 (фиг. 3B). В частности, в клетках NHI3T3 авторы изобретения детектировали сильное уменьшение ацетилирования гистона H3, которое, как известно, опосредуется MeCP2 (фиг. 3 C и D). Эти результаты указывают на то, что белок TAT-MeCP2e2-EGFP является функционально активным, независимо от дополнительных белковых доменов, которые были введены.
Авторы изобретения создали серию конструкций для экспрессии обеих изоформ MeCP2 с белком и без белка EGFP на С-конце и слитых на N-конце с доменом ТАТ (фиг. 2). Дополнительные конструкции содержали синтетический ген MECP2, полностью кодон-оптимизированный для Escherichia coli. Частота использования кодонов синтетической кДНК-последовательности MECP2 увеличивала продукцию этого белка в E. coli до увеличения в 200 раз (фиг. 3A). Авторы испытывали доставку ТАТ-слитых белков и обнаружили наивысшую эффективность in vitro при 500 пмоль рекомбинантных белков. После трех часов инкубации с белком TAT-MeCP2e2-EGFP, приблизительно 40% клеток NIH 3T3 обнаруживали присутствие макроскопических везикулоподобных флуоресцентных пятен. Этот белок находился в основном в цитозоле (фиг. 4 A a, b). После 12 часов, почти 100% этих клеток были флуоресцентными (фиг. 4 A c, d) и белок концентрировался в ядрах этих клеток (фиг. 4A e). Трансдукция была стабильной в течение 24 часов в трансдуцированных клетках (фиг. 4 B и C). Эксперименты с использованием белка EGFP без домена ТАТ в качестве контроля не приводили к поглощению в эти клетки (фиг. 4A f). TAT-рекомбинантные белки были также способны трансдуцировать первичные фибробласты человека (фиг. 4A g и h) и первичные нейроны из головного мозга мыши (фиг. 4A i и j).
6. MeCP2 может быть замещен в фибробластах, полученных из пациента с синдромом RTT, и в мышиных фибробластах Mecp2 -/y .
Затем авторы изобретения исследовали биологическую активность MeCP2-слитых белков после доставки в MeCP2-недостаточные клетки. Авторы детектировали гиперацетилирование гистонов Н3 и H4K16 в линиях моноклональных клеток, экспрессирующих мутированный аллель MeCP2 (T158M), в противоположность клеточной линии, экспрессирующей аллель дикого типа (фиг. 5a). Клеточную линию, экспрессирующую этот мутированный аллель, обрабатывали 300 пмоль белков TAT-MeCP2e1-TAT-MeCP2e2, MeCP2 или TAT-EGFP в течение 64 часов. Обращение гиперацетилирования гистонов Н3 и H4K16 наблюдали только после обработки обеими изоформами TAT-MeCP2 (фиг. 5a и b). Авторы изобретения получили аналогичные результаты нацеливания на фибробласты мышей Mecp2-/y (фиг. 5c). Эксперименты по коиммунопреципитации в трансдуцированных фибробластах мышей Mecp2-/y прямо демонстрировали рекрутинг гистондеацетилазы 1 (HDACl) изоформами TAT-MeCP2 (фиг. 7A). Эти результаты доказывают, что изоформы TAT-MeCP2 способны устранять биохимические изменения, вызываемые потерей функции эндогенного MeCP2.
7. Внутриклеточная стабильность белков TAT-MeCP2
Для испытания стабильности и стойкости трансдуцированного белка в клетках эти клетки инкубировали в течение 2 часов с 1 нмоль TAT-MeCP2e1 и TAT-MeCP2e2 по отдельности. Затем их промывали средой и инкубировали с новой культуральной средой для дополнительного роста. Эти клетки собирали при указанных интервалах времени и белки экстрагировали суспендированием 107 клеток в 200 мкл буфера для лизиса клеток (125 мМ Трис-HCl pH 6,8, 2% ДСН, 50 мМ NaH2PO4, 20% глицерин, 1 мМ ДTT, 1 мМ PMSF) и инкубированием на льду в течение 20 мин, осторожной обработкой ультразвуком для фрагментирования этой ДНК. Этот лизат центрифугировали при 9000×g в течение 10 мин, 4°C, и концентрацию общего белка определяли количественно по способу Бредфорда и хранили при 80°С до использования. 100 мкг проб этого белка нагревали при 75°C с 4X ДСН буфером нанесения (1 M Трис-HCl pH-6,88, 2% ДСН, 0,02% бромфеноловый синий, 5 мМ бета-2-меркаптоэтанол, 5% глицерин) и разделяли на 4-20% точных гелях для белков (Pierce Biotechnology, USA). Эти белки переносили на нитроцеллюлозную мембрану, оптимизированную для переноса белков (Amersham Biosciences, GE Healthcare). Эту мембрану обрабатывали усилителем сигнала Вестерн-блота (Pierce Biotechnology, USA) согласно протоколу поставщика. Эту мембрану блокировали 5% обезжиренным молоком в PBS с 0,05% Tween-20 (PBS-Т) в течение 1 часа. Затем эту мембрану инкубировали по отдельности с блокирующим раствором, содержащим анти-MeCP2-антитело (Upstate, USA), разбавленным 1:2000, в течение ночи при 4°C. Мышиное моноклональное анти-альфа-тубулин-антитело (Sigma, Taufkirchen, Germany), разбавленное 1:4000, использовали в качестве положительного контроля. Мембрану промывали 3 раза в течение 5 минут в каждом случае с использованием PBS-Т и затем инкубировали с тем же самым буфером, содержащим соответствующие вторичные антитела, конъюгированные со щелочной фосфатазой (Sigma), разбавленные 1:30000, в течение 1 часа. Избыток антител вымывали PBS-Т три раза и проявляли с использованием BCIP/NBT (Carl-Roth, Germany). Внутриклеточная концентрация трансдуцированного белка оставалась высокой в течение 24 часов, после чего этот уровень снижался до уровня, меньшего, чем четверть исходной концентрации, на протяжении следующих 24 часов (фиг. 4В и С).
8. Вестерн-блот-анализ
8.1 Экстракты белка культур клеток: Содержание общего белка из всех клеточных линий и клеточных культур первичных фибробластов экстрагировали суспендированием 107 клеток в 200 мкл буфера для лизиса клеток и инкубировали на льду в течение 20 мин, осторожно обрабатывали ультразвуком для фрагментирования ДНК. Этот лизат центрифугировали при 9000×g в течение 10 мин, 4°C. Вестерн-блоттинг выполняли, как описано выше. Эту мембрану инкубировали в течение ночи при 4°С с блокирующим раствором, содержащим кроличьи поликлональные антитела против гистона Н3, при разведении 1:4000, ацетилированного гистона Н3, ацетилированного гистона H4K16 (Upstate), разведенных 1:10000 и 1:500, соответственно. Мышиное моноклональное анти-альфа-тубулин-антитело (Sigma), разведенное 1:4000, использовали в качестве положительного контроля.
8.2 Экстракты белка тканей: Мышей инъецировали 100 мкг/г (массы тела) TAT-MeCP2-EGFP, TAT-MeCP2 и MeCP2. Спустя 20 часов мышей убивали. Головной мозг извлекали и промывали в холодном PBS. Затем головной мозг гомогенизировали в буфере для экстракции (62 мМ Трис-Cl pH 6,8, 2% ДСН, 5 мМ меркаптоэтанол, 0,15 мМ NaCl), 5X коктейль ингибиторов протеаз (Sigma)) и осторожно обрабатывали ультразвуком. Этот лизат осаждали при 15000×g в течение 15 мин, 4°C. Пятьдесят мкг использовали для Вестерн-блоттинга, проводимого, как описано выше, с использованием поликлонального анти-MeCP2-антитела (Upstate). В независимых экспериментах авторы изобретения инъецировали мышей 100 мкг/г (массы тела) TAT-EGFP и TAT-MeCP2e2-EGFP. Иммуноблоттинг выполняли на экстракте общего белка из головного мозга, печени, почки и сердца с использованием анти-GFP-антитела (Sigma). Плотность сигналов Вестерн-блота определяли количественно визуализирующим денситометром Olympus S-100. Соотношения полос интенсивности измеряли с использованием сигнала альфа-тубулина в качестве внутреннего контроля.
9. Иммунохимия
Мышей, инъецированных i.p. TAT-MeCP2e2-EGFP и EGFP, транскардиально перфузировали холодным 4% параформальдегидом (PFA) в буфере PBS спустя 20 часов после инъекции. Головной мозг извлекали и постфиксировали 4% PFA в течение 4 часов, криопротектировали 30% сахарозой при 4°С и хранили при -80°С. Иммуногистохимию выполняли на криосрезах с использованием кроличьего анти-GFP-антитела (Sigma). Изображения получали с использованием микроскопа Olympus BX60 (Olympus Optical Co. LTD, Japan). Аналогичные эксперименты выполняли на фиксированных формалином срезах мышей Mecp2-/y, инъецированных в течение 20 дней TAT-MeCP2e1 и e2 (20 мкг/г массы тела, каждый второй день). Эти срезы зондировали поликлональным кроличьим анти-MeCP2e2-антителом.
Это антитело было индуцировано в кролике с использованием рекомбинантного белка MeCP2e2 человека.
10. Белковая терапия in vitro и in vivo
10.1 Белковая терапия in vitro
Авторы изобретения испытывали рекомбинантные белки в линиях моноаллельных клеток, экспрессирующих мутацию T158M MeCP2. Эти клетки обнаруживают гиперацетилирование Н3 и H4K16. Инкубация как изоформы TAT-MeCP2e1, так и изоформы e2, обращала патологическое гиперацетилирование до уровня, сходного с уровнем линий клеток дикого типа (фиг. 5).
10.2 Эффективная доставка белков МеСЗ2 в головной мозг
Затем авторы изобретения исследовали вопрос доставки белка in vivo в мышей дикого типа NMRI, получающих одну единственную внутрибрюшинную инъекцию белка TAT-MeCP2e2-EGFP, TAT-EGFP или EGFP (60 мкг/г массы тела в смеси PBS/10% глицерин или в смеси 20 мМ HEPES/10% глицерин). Мыши, инъецированные TAT-EGFP и EGFP, не развивали никаких видимых побочных эффектов. Однако мыши, инъецированные изоформой TAT-MeCP2e2-EGFP, обнаруживали транзиторную гипомобильность задних конечностей, которая исчезала спустя один час-два часа после инъекции. Вестерн-блот-анализ головного мозга, печени и почки выявил поглощение белков TAT-EGFP и TAT-MeCP2-EGFP спустя 20 часов после инъекции (данные не показаны). В аналогичных экспериментах, мышей инъецировали при дозе 100 мкг/г массы тела независимо белками TAT-MeCP2e2, TAT-MeCP2e2-EGFP и MeCP2e2. Вестерн-блот-анализ выявил присутствие TAT-рекомбинантных белков в экстрактах головного мозга мышей, умерщвленных спустя 20 часов после инъекции (фиг. 6a). Иммуногистохимия срезов головного мозга с использованием анти-EGFP-антитела выявила присутствие белка TAT-MeCP2e2-EGFP в пирамидальных нейронах неокортекса, в клетках-зернах (нейронах зернистого слоя мозжечка) и в мозговых оболочках (фиг. 6b). Однако при увеличении дозы обеих изоформ TAT-MeCP2 до 130 мкг/г массы тела авторы изобретения наблюдали развитие тяжелых побочных эффектов, таких как подобные судорогам симптомы, в пределах 10-20 минут после инъекции, которые приводили в полному параличу всех четырех конечностей. Вследствие этого, две из 5 инъецированных мышей NMRI умерли в пределах 2 часов. В противоположность этому, мыши, инъецированные белками TAT-EGFP и MeCP2, не имели никаких серьезных побочных эффектов при любой испытанной дозе (таблица 1 ниже). Эти данные указывают на то, что успешная доставка белков MeCP2 в головной мозг является в принципе возможной, однако, доза должна строго контролироваться.
В следующей серии экспериментов, авторы изобретения определяли доставку слитых белков MeCP2 к головному мозгу Mecp2-недостаточных мышей, мышиной модели синдрома Ретт, созданной в лаборатории A. Bird (Chen et al. 2001, Nat Genet, 27 (3):327-31). Животных покупали из Jackson Laboratory, USA (штамм B6.129P2(C)-Mecp2tml.lBird/J), и выполняли дополнительное разведение в оборудовании для животных Института Генетики Человека (Institute of Human Genetics, Gоttingen). Контрольных мышей брали из вышеупомянутого оборудования для животных. Двух мышей Mecp2-/y инъецировали i.p. в течение 20 дней с 20 мкг/г массы тела каждый второй день TAT-MeCP2e1 и TAT-MeCP2e2, соответственно. Аналогично тому, что было получено в случае мышей дикого типа, авторы изобретения смогли детектировать белки TAT-MeCP2 с использованием иммуногистохимии на срезах головного мозга мышей Mecp2-/y (фиг. 6c). Эти эксперименты доказывают, что слитые ТАТ-белки пересекают гематоэнцефалический барьер и достигают головного мозга. При терапевтическом режиме 20 мкг/г массы тела каждый второй день авторы никогда не наблюдали никаких непосредственных или задержанных неврологических симптомов или неожиданной смерти обработанных Mecp2-/y мышей. Буфером инъекционного раствора для внутрибрюшинной доставки был 20 мМ Hepes, 100 мкМ CaCl2, 300 мМ NaCl и 10% глицерин.
10.3 Белковая терапия in vivo: терапевтические эффекты рекомбинантных изоформ TAT-MeCP2 в мышиной модели синдрома Ретта
Затем авторы изобретения испытывали, являются ли доставленные in vivo белки MeCP2 функционально активными в головном мозге MeCP2-недостаточных мышей. Авторы инъецировали один раз в день i.p. TAT-MeCP2e1 или TAT-MeCP2e2 в мышей Mecp2-/y в дозе 20 или 30 мкг/г, соответственно. При этих дозах не наблюдали клинических побочных действий. Этих животных умерщвляли после двух недель терапии и экстракты общего белка из головного мозга анализировали Вестерн-блоттингом. Инъекция 20 или 30 мкг/кг белка заметно уменьшала уровень ацетилирования H4K16 (фиг. 7B и C). Показав биохимическую активность доставляемых in vivo белков, авторы хотели оценить, может ли эта терапия исправлять фенотип MeCP2-недостаточных мышей. Авторы инъецировали мышей Mecp2-/y i.p. 20 мкг/г массы тела с использованием обеих изоформ TAT-MeCP2. Пять мышей Mecp2-/y получали TAT-MeCρ2e1 и пять мышей получали TAT-MeCP2e2. Одну мышь Mecp2-/y инъецировали PBS, одну мышь инъецировали MeCP2 и 4 мыши получали белок TAT-EGFP в той же самой дозе. Одинадцать мышей Mecp2-/y не были обработаны и служили в качестве контролей. Обработку начинали в постнатальный день 30. Среднее выживание MeCP2-инъецированных мышей Mecp2-/y (n=10) было равно 88,7 дней (92,6 для изоформы e1 и 84,8 для изоформы e2, соответственно), необработанных мышей Mecp2-/y 55,6 дней и ложноинъецированных мышей 57,0 дней (таблица 2 ниже). Это представляет увеличение продолжительности жизни почти 66% (TAT-MeCP2e1) и 53% (TAT-MeCP2e2) обработанных мышей в сравнении с необработанными мышами. Это пролонгирование выживания является статистически высоко значимым (фиг. 8a и b). Один самец мыши дикого типа, обработанный в течение трех месяцев 20 мкг/г MeCP2, не развивал никаких клинических побочных действий и был все еще здоровым при 300 днях обработки.
* В 2 экспериментах мыши умирали. Доза 30 мкг/г рекомбинантных белков не оказывала никакого конкретного действия на мышей, независимо от того, какой белок инъецировали.
b. Инъецированные MeCP2
c. Инъецированные TAT-EGFP
d. Это животное было все еще подвижным и активным, но его умерщвляли вследствие очень тяжелой и неизлечимой инфекции кожи, вызвавшей калечащее поведение самокусание.
11. Доставка белка TAT-MeCP2 избавляет от повреждения нейронов in vivo
Было описано, что делеция MeCP2 в постнатальных нейронах ЦНС приводит к тяжелым невропатологическим изменениям, таким как изменения дендритных отростков и сморщиванию нейронов (Armstrong DD. 2002, Ment Retard Dev Disabil ResRev 8:72-6). Для исследования, защищает ли также обработка белком TAT-MeCP2 нейроны от морфологических изменений, нейроны из гиппокампа и мозжечка исследовали гистологически. Размер нейронов оценивали количественно в окрашенных Nissl срезах (фиг. 8c) в районе CA2 гиппокампа или на окрашенных кальбиндином клетках Пуркинье мозжечка. Мутантные нейроны в гиппокампе были меньшими по размеру и были более плотно упакованы, чем нейроны дикого типа (фиг. 8с, 1 и 2). Важно, что MeCP2-обработанные животные обнаруживали более крупные нейроны без каких-либо видимых морфологических изменений (фиг. 8c 3). Количественное определение выявило, что размер нейронов был неотличимым в обработанных TAT-MeCP2e1 нейронах гиппокампа в сравнении с диким типом, тогда как изоформа MeCP2e2 не оказывала действия на размер нейронов (фиг. 8c 4). Измерения пирамидальных кортикальных нейронов слоя V в MeCP2-недостаточных мышах 50-дневного возраста не обнаруживали различий относительно ситуации дикого типа. Кроме того, мозжечковые клетки Пуркинье cохранялись в мышах MeCP-/y и не обнаруживали макроскопических морфологических отклонений от нормы после обработки TAT-MeCP2 (данные не показаны). Взятое вместе, уменьшение размера нейронов в гиппокампе могло бы обращаться применением MeCP2e1.

























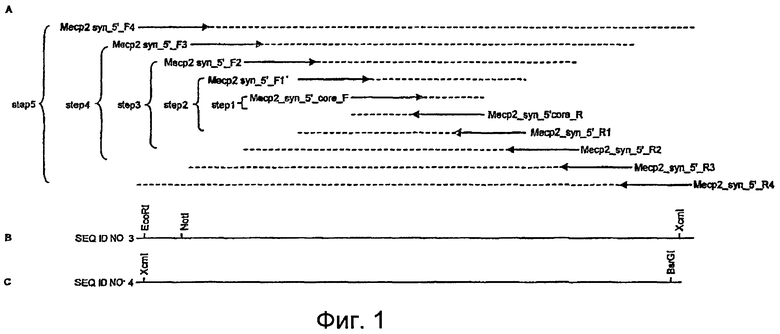
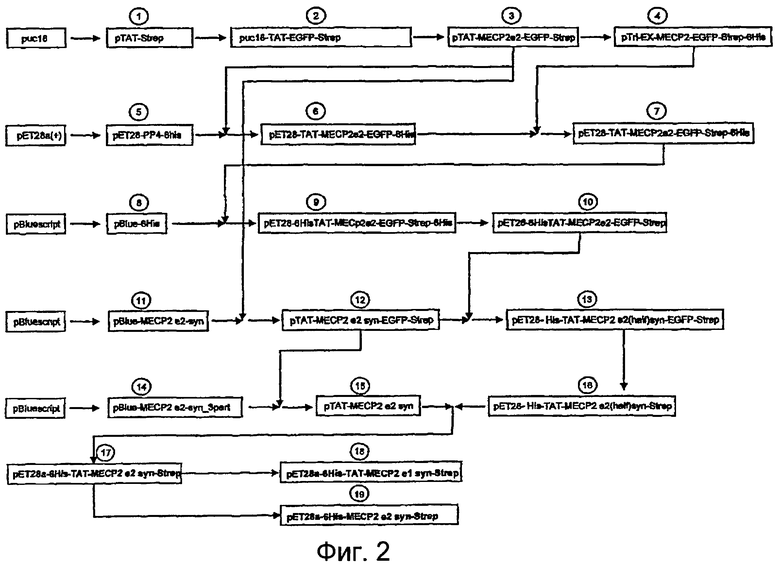
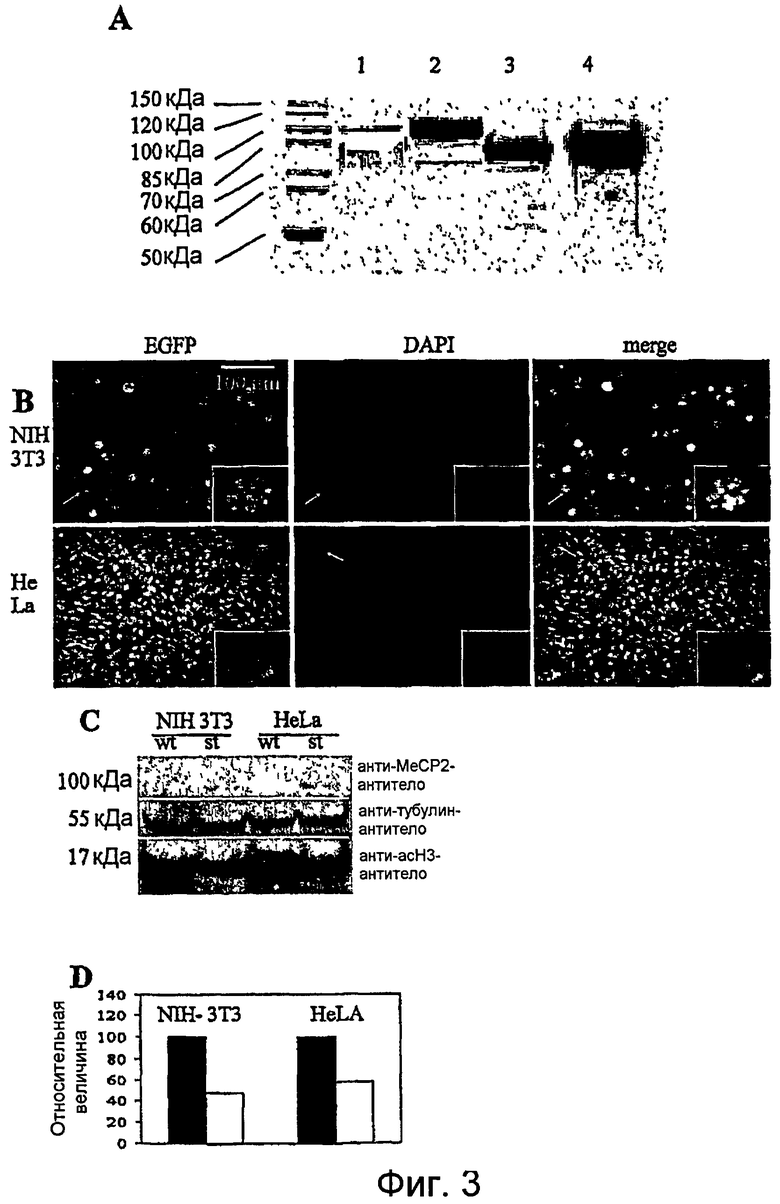

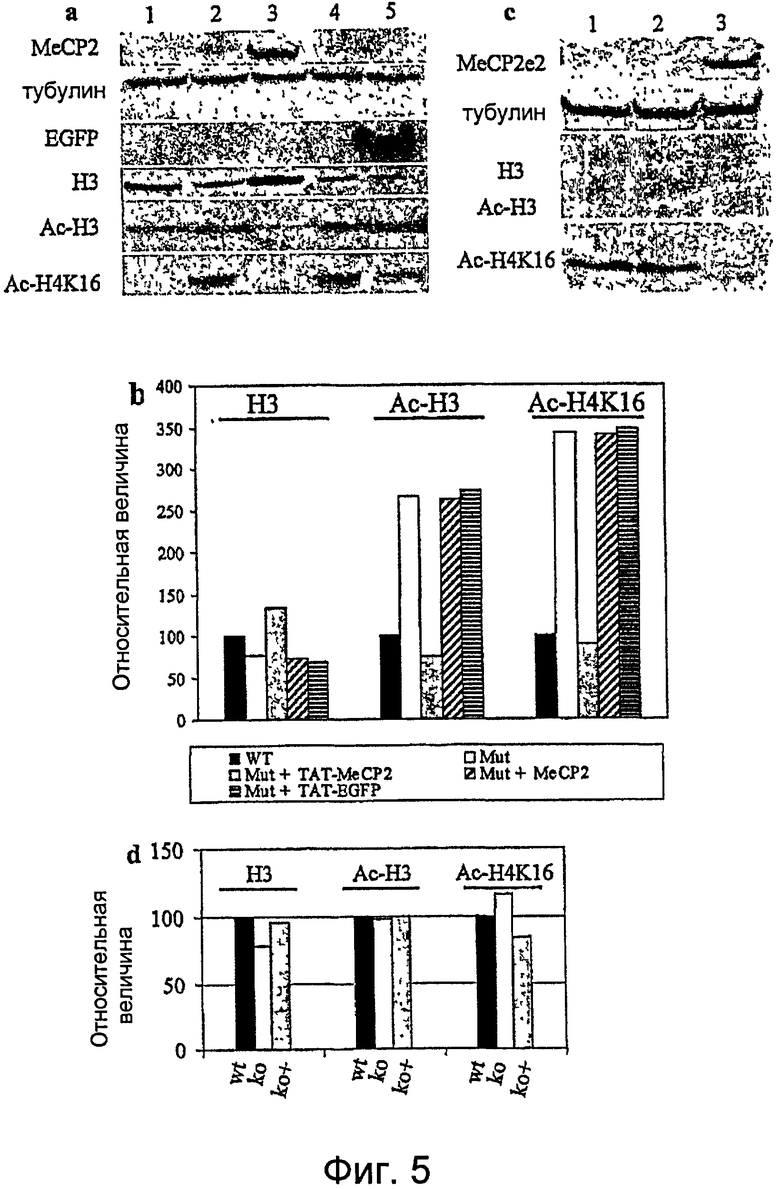
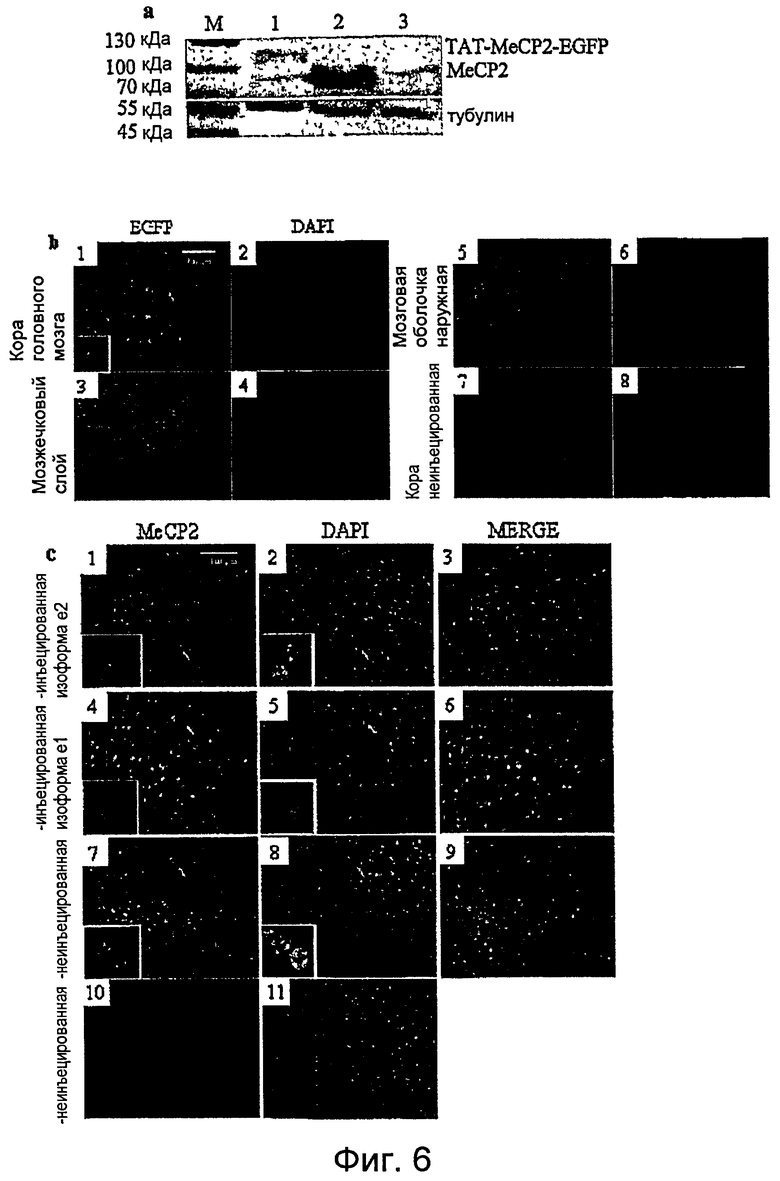
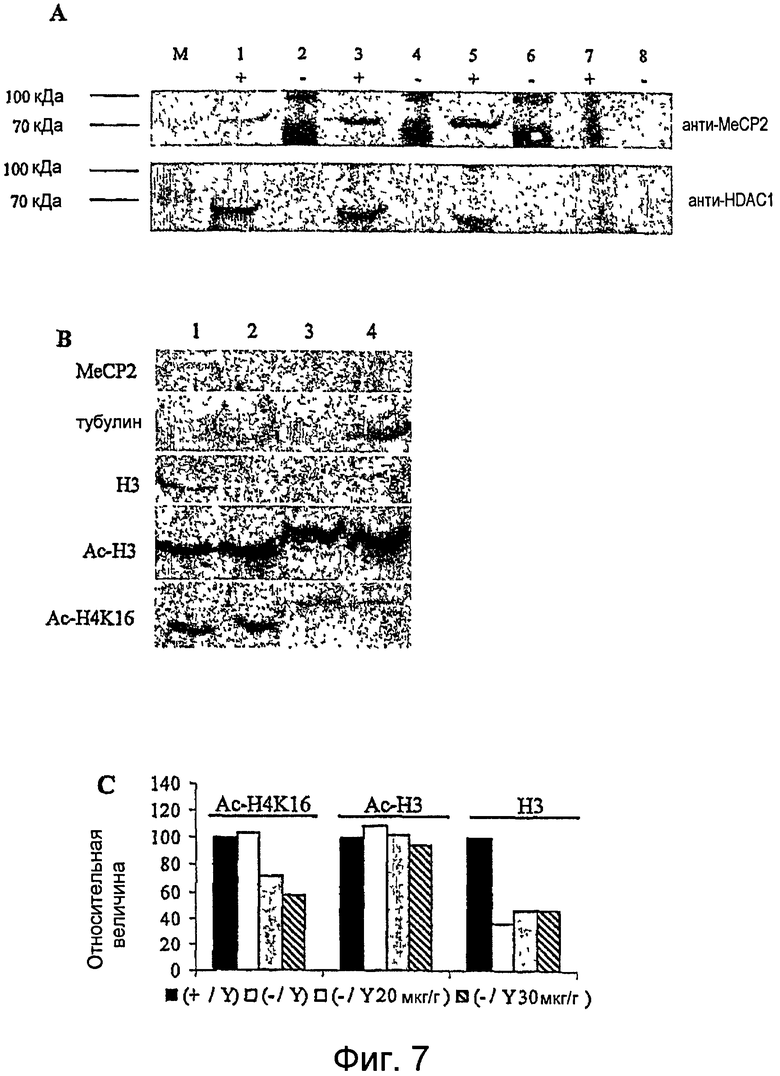
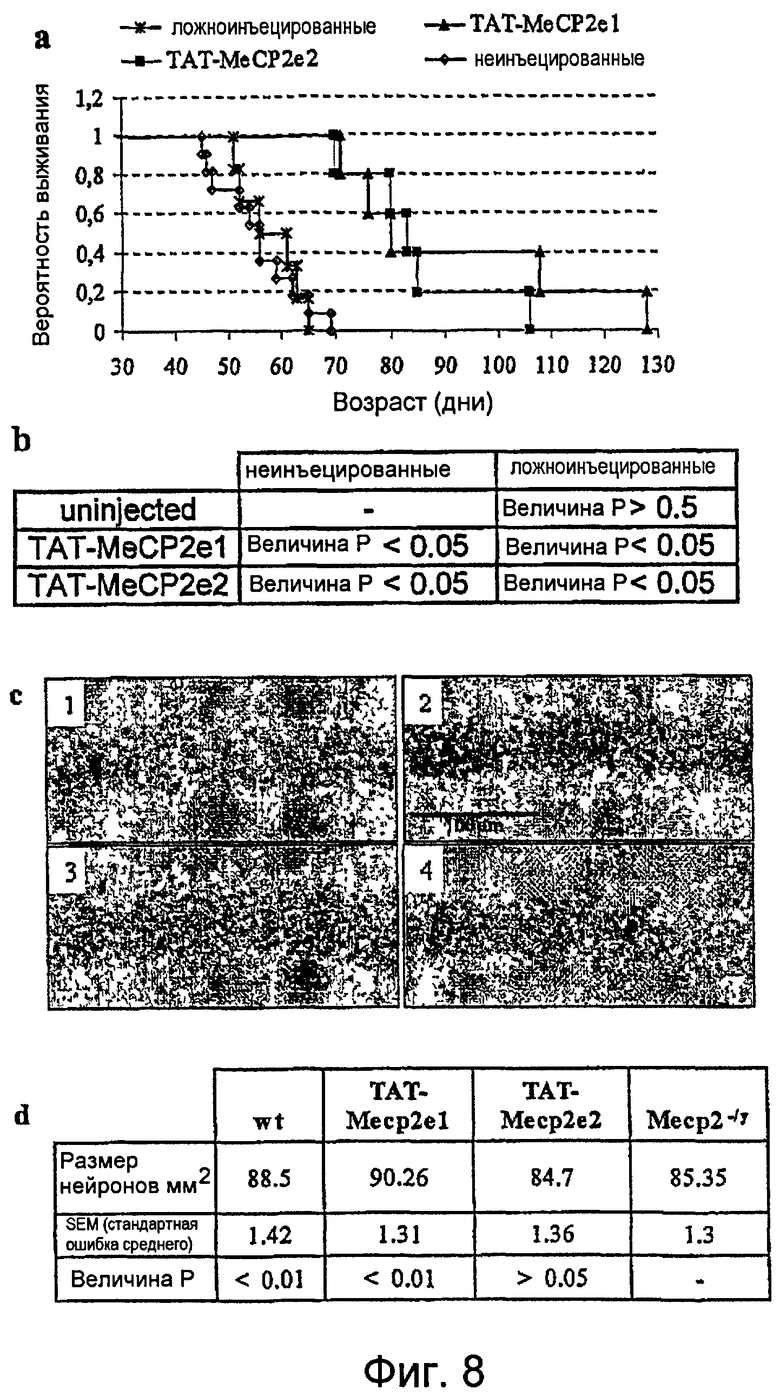
Изобретение относится к области биотехнологии и касается применения слитого полипептида, содержащего (i) белок МеСР2 и (ii) домен трансдукции белка для изготовления лекарственного средства для лечения и/или профилактики связанного с развитием нервного заболевания, а также к фармацевтическим композициям, содержащим такой полипептид, и способу лечения связанного с развитием нервного заболевания. Настоящее изобретение позволяет компенсировать потерю функции МеСР2 пораженных нервных клеток. 3 н. и 8 з.п. ф-лы, 8 ил., 2 табл.
1. Применение полипептида, содержащего (i) белок МеСР2 или биологически активный фрагмент или функциональное производное указанного белка или фрагмента и (ii) домен трансдукции белка, где этот белок МеСР2 слит с доменом трансдукции белка, для изготовления лекарственного средства для лечения и/или профилактики связанного с развитием нервного заболевания, где указанный фрагмент содержит по меньшей мере 150 аминокислот белка МеСР2, и где указанное производное указанного белка или фрагмента имеет по меньшей мере 60% идентичности к указанному белку или его фрагменту, и где указанный фрагмент или указанное производное имеет по меньшей мере 60% указанной биологической активности, определенной при помощи анализа транскрипции in vitro, при этом связанное с развитием нервное заболевание обусловлено уменьшением экспрессии МеСР2 или нарушенной функцией МеСР2.
2. Применение по п.1, где связанное с развитием нервное заболевание является синдромом Ретта.
3. Применение по п.1, где домен трансдукции белка произведен из белка-трансактиватора транскрипции вируса-1 иммунодефицита человека (белка ТАТ).
4. Применение по п.1, где белок МеСР2 или биологически активный фрагмент или их производное происходит из человека.
5. Применение по п.1, где белок МеСР2 или биологически активный фрагмент или их производное являются выбранными из группы, включающей изоформу е1 МеСР2 человека или изоформу е2 МеСР2 человека или их биологически активный фрагмент или производное указанных изоформ или указанных фрагментов.
6. Применение по п.1, где указанный домен трансдукции белка имеет по меньшей мере 60% идентичности с аминокислотной последовательностью домена трансдукции белка TAT (SEQ ID No:54).
7. Применение по п.1, где указанный домен трансдукции белка имеет по меньшей мере 70% идентичности с аминокислотной последовательностью домена трансдукции белка TAT (SEQ ID No:54).
8. Фармацевтическая композиция для лечения и/или профилактики связанного с развитием нервного заболевания, содержащая эффективное количество полипептида, который содержит (i) белок МеСР2 или биологически активный фрагмент или функциональное производное указанного белка или фрагмента и (ii) домен трансдукции белка, где этот белок МеСР2 слит с доменом трансдукции белка МеСР2, и где указанное производное указанного белка или фрагмента имеет по меньшей мере 60% идентичности к указанному белку или его фрагменту, и где указанный фрагмент или указанное производное имеет по меньшей мере 60% указанной биологической активности, определенной при помощи анализа транскрипции in vitro, и фармацевтически приемлемый носитель, при этом связанное с развитием нервное заболевание обусловлено уменьшением экспрессии МеСР2 или нарушенной функцией МеСР2.
9. Способ лечения и/или профилактики связанного с развитием нервного заболевания, включающий введение фармацевтической композиции по п.8 млекопитающему, при этом связанное с развитием нервное заболевание обусловлено уменьшением экспрессии МеСР2 или нарушенной функцией МеСР2.
10. Способ по п.9, где указанную фармацевтическую композицию вводят в дозе из расчета 0,1-200 мкг полипептида на г массы тела млекопитающего.
11. Способ по п.9, где указанную фармацевтическую композицию вводят по меньшей мере один раз в день или каждый второй день.
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| US 20040126877 A1, 01.07.2004. | |||

