Область техники, к которой относится изобретение
Настоящее изобретение относится к модифицированным антителам, содержащим две или более V-области Н-цепи и две или более V-области L-цепи антитела, которые проявляют ТРО-агонистическую активность путем поперечного сшивания ТРО-рецептора. Модифицированные антитела обладают активностью ТРО-агонистов, трансдуцируя сигнал в клетки путем поперечного сшивания ТРО-рецептора, и могут применяться как лекарственные средства в различных целях.
Уровень техники
Открытый в 1994 г. тромбопоэтин (ТРО) представляет собой фактор, регулирующий образование тромбоцитов, и, как известно, состоит из гликопротеина с молекулярной массой 70-80 тыс. Да и вырабатывается главным образом в печени. Тромбопоэтин - это цитокин, помогающий клеткам предшественника тромбоцитов в костном мозге выживать, размножаться, дифференцироваться и созревать, то есть он стимулирует дифференцировку и пролиферацию мегакариоцитов. Рецептор тромбопоэтина (ТРО) был идентифицирован ранее ТРО как с-Mpl, рецептор специфического фактора регуляции образования тромбоцитов (М. Souyri et al., Cell 63: 1137 (1990)). Как сообщалось, с-Mpl распространен преимущественно в клетках предшественника тромбоцитов, мегакариоцитах, и в клетках тромбоцитов, и подавление экспрессии с-Mpl избирательно тормозит образование мегакариоцитов (М. Methia et al., Blood 82: 1395 (1993)). Сообщение о том, что лигандом к с-Mpl является ТРО, основывалось на результатах анализа пролиферации клеток, специфичных к лиганду к с-Mpl, и очистки лиганда с использованием с-Mpl (F. De Sauvage et al., Nature 369: 533 (1994); TD. Bartley et al., Cell 77: 1117 (1994)). В настоящее время Mpl называют ТРО-рецептором. Как полагают, агонисты ТРО и ТРО-рецептора могут быть применены, таким образом, в качестве терапевтического средства против тромбоцитопении, например, как лекарственный препарат, снижающий тромбоцитопению, вызванную подавлением костного мозга или резекцией костного мозга при лечении больных раком.
Помимо этого, разрабатывались модифицированные антитела, в частности антитела с уменьшенными размерами молекулы, как, например, одноцепочечные Fv, для улучшения проникающей способности в ткани и опухоли за счет уменьшения размера молекул и которые можно было бы получать рекомбинантным способом. В последнее время димеры одноцепочечных Fv, особенно биспецифичные димеры, используют для наглядного сшивания клеток. Типичными примерами подобных димеров являются гетеродимеры одноцепочечных Fv, распознающие антигены раковых клеток и антигены клеток-хозяина, например, NK клеток (естественные киллеры) и нейтрофилов (Kipriyanov et al., Int. J. Cancer, 77, 9763-9772, 1998). Их получают методами конструирования из одноцепочечных Fv в виде модифицированных антител, которые более эффективны при лечении разновидностей рака путем индуцирования поперечного межклеточного сшивания. Как полагают, образование межклеточных поперечных сшивок индуцируется антителами и их фрагментами (например, Fab-фрагментом), биспецифичными модифицированными антителами и даже димерами одноцепочечных Fv, которые сами по себе моноспецифичны.
Примеры антител, о которых известно, что они способны трансдуцировать сигнал путем образования поперечных сшивок поверхностных молекул/молекулы клетки, включают такие как антитело к ЕРО-рецептору, участвующему в дифференцировке и пролиферации (JP-A 2000-958000), антитело к MuSK-рецептору (Xie et al., Nature Biotech. 15, 768-771, 1997) и др. Известно также антитело-агонист к рецептору ТРО, его фрагменты и одноцепочечные Fvs (WO 99/17364). Однако никаких сведений о том, что димеры одноцепочечных Fv и такие модифицированные антитела, как одноцепочечные бивалентные антитела, обладают агонистической активностью, не приводилось.
Обратив внимание на тот факт, что мономеры одноцепочечных Fv, происходящие от моноклональных антител (антитело MABL-1 и антитело MABL-2, полученные авторами настоящего изобретения), индуцирующих апоптоз IAP-содержащих клеток, не индуцируют апоптоз клеток, а димеры индуцируют апоптоз, авторы изобретения открыли, что димеры поперечно сшивают (димеризируют) IAP-рецептор на клеточной поверхности и таким путем сигнал трансдуцируется в клетки, что в итоге приводит к индуцированию апоптоза. Это дало основание предположить, что димеры моноспецифичных одноцепочечных Fv поперечно сшивают молекулу/молекулы на поверхности клетки (т.е. рецептор) и трандуцируют сигнал, подобно лиганду, функционируя таким образом как агонисты.
Сосредоточив свое внимание на образовании межклеточных поперечных сшивок, авторы обнаружили, что вышеуказанные димеры одноцепочечных Fv не вызывают гемагглютинации, в то время как вышеуказанные моноклональные антитела вызывают ее. Такие же результаты были получены и для одноцепочечных двухвалентных антител (одноцепочечные полипептиды, содержащие две V-области Н-цепи и две V-области L-цепи). Из этого следовало, что моноклональные антитела могут образовывать межклеточные поперечные сшивки, тогда как модифицированные антитела типа димеров одноцепочечного Fv и одноцепочечных двухвалентных антител, поперечно сшивают молекулу/молекулы поверхности клетки, однако межклеточных поперечных сшивок не образуют.
Основываясь на данных наблюдениях, авторы настоящего изобретения впервые обнаружили, что такие модифицированные антитела, как димеры одноцепочечных Fv и одноцепочечные бивалентные антитела, поперечно сшивают молекулу/молекулы на поверхности клетки или же внутриклеточную молекулу/молекулы той же клетки, помимо уже известного межклеточного поперечного сшивания, и что они приемлемы в качестве лигандов к молекуле/молекулам (в частности, в качестве лиганда, имитирующего действие естественного лиганда).
Обнаружив помимо этого, что молекулу антитела (цельный IgG) можно модифицировать в димеры одноцепочечного Fv, одноцепочечные бивалентные антитела и т.п., которые образуют поперечные сшивки молекулы/молекул поверхности клетки, уменьшая тем самым нежелательные последствия межклеточного поперечного сшивания и обеспечивая новые лекарственные средства, оказывающие только желательное воздействие на клетку, авторы таким образом завершили свое изобретение. Модифицированные антитела настоящего изобретения обладают значительно более высокой активностью, чем цельные антитела (IgG), содержащие ту же самую V-область, что и модифицированные антитела. Благодаря уменьшенным, по сравнению с молекулами антител, молекулярным размерам и благодаря отсутствию у них константных областей, они обладают улучшенной проникающей способностью в ткани.
Сущность изобретения
Целью настоящего изобретения является получение модифицированных антител-агонистов с уменьшенным размером молекулы, содержащих две или более V-области Н-цепи и две или более V-области L-цепи моноклонального антитела и проявляющих ТРО-агонистическую активность путем поперечного сшивания ТРО-рецептора.
Таким образом, настоящее изобретение относится к модифицированным антителам, содержащим две или более V-области Н-цепи и две или более V-области L-цепи, предпочтительно от 2 до 6 каждой, особенно предпочтительно от 2 до 4 каждой, наиболее предпочтительно по две каждой, и проявляющим активность как ТРО-агонисты путем поперечного сшивания ТРО-рецептора.
"Модифицированные антитела" в настоящем описании означают любое вещество, содержащее две или более V-области Н-цепи и две или более V-области L-цепи, в котором вышеозначенные V-области связаны напрямую или же через линкер с помощью ковалентной или нековалентной связи. Например, полипептиды и соединения, полученные соединением каждой V-области антитела через пептидный линкер или с помощью химического поперечносшивающего агента или т.п. Две или более V-области Н-цепи и две или более V-области L-цепи, используемые в настоящем изобретении, могут быть производными одного и того же антитела или же разных антител.
Модифицированными антителами, согласно настоящему изобретению, могут быть любые вещества, пока они сохраняют способность специфического распознавания и поперечного сшивания ТРО-рецептора и таким образом могут трансдуцировать сигнал в клетки. Сюда входят модифицированные антитела, полученные дальнейшей модификацией одной из частей аминокислотной последовательности V-области модифицированных антител.
Предпочтительными примерами модифицированных антител по настоящему изобретению являются такие мультимеры, как димеры, тримеры и тетрамеры одноцепочного Fv, содержащие V-область Н-цепи и V-область L-цепи, или одноцепочечные полипептиды, содержащие две или более V-области Н-цепи и две или более V-области L-цепи. Для случаев, когда модифицированные антитела по настоящему изобретению являются мультимерами одноцепочного Fv, например димерами, тримерами, тетрамерами и т.п., содержащими V-область Н-цепи и V-область L-цепи, предпочтительно, чтобы V-область Н-цепи и V-область L-цепи, присутствующие в одной и той же цепи, не были бы соединены таким образом, чтобы образовывать антигенсвязывающий центр.
Более предпочтительными примерами являются димеры одноцепочечного Fv, содержащего V-область Н-цепи и V-область L-цепи, или одноцепочечный полипептид, содержащий две V-области Н-цепи и две V-области L-цепи. Предпочтительно, чтобы V-область Н-цепи и V-область L-цепи в этих модифицированных антителах были соединены через линкер.
Примеры вышеуказанного мультимера одноцепочечного Fv включают мультимеры с нековалентной связью, мультимеры с ковалентной связью, полученные при помощи поперечносшивающего радикала, и мультимеры, полученные посредством поперечносшивающего реагента (антитела, фрагмента антитела или двухвалентного модифицированного антитела). В качестве поперечносшивающих радикалов для образования мультимера могут быть использованы стандартные поперечносшивающие радикалы, применяемые для поперечной сшивки пептидов. В качестве примера можно привести дисульфидную мостиковую связь по остаткам цистеина, другие поперечносшивающие радикалы, например С4-С10 алкилен (т.е. тетраметилен, пентаметилен, гексаметилен, гептаметилен и октаметилен и т.д.), или С4-С10 алкенилен (цис-/транс-3-бутенилен, цис-/транс-2-пентенилен, цис-/транс-3-пентенилен, цис-/транс-3-гексенилен и т.д.).
Кроме того, таким поперечносшивающим реагентом, который может соединяться с одноцепочечнным Fv, является, например, аминокислотная последовательность, которую можно необязательно вводить в Fv, например, антитело к FLAG-последовательности или подобной или его фрагмент или модифицированное антитело, происходящее из антитела, например одноцепочечный Fv.
"ТРО-агонистическое действие" в настоящем описании обозначает биологическое действие, протекающее в клетке/клетках, сигнал в которую/которые трансдуцируется путем поперечного сшивания ТРО-рецептора, например, пролиферация, дифференцировка или стимуляция роста мегакариоцитов или образование тромбоцитов.
ED50 ТРО-агонистического действия, согласно изобретению, определяют общепринятыми для измерения агонистического действия способами. Примеры методов такого измерения включают анализ клеточной пролиферации с применением клеточных линий, чувствительных к ТРО, типа BaF/mpl или UT7/TPO; измерение фосфорилирования белка MPL; анализ дифференциации колонии мегакариоцитов из клеток костного мозга; анализ восстановительного синтеза в тромбоцитах мышей in vivo; измерение индукции экспрессии тромбоцитарного антигена GPIIbIIIa (анти-GPIIbIIIa) с использованием линии мегакариобластных клеток лейкемии человека (СМК) или измерение индукции полиплоидии клеточной линии мегакариобластов (DAMI). ED50 - это доза, необходимая для достижения 50% реакции по отношению к максимуму активности, принятому за 100% на кривой "доза-реакция".
Предпочтительные модифицированные антитела по настоящему изобретению обладают ТРО-агонистическим действием (ED50), равным или же большим, чем у антитела с той же антигенсвязывающей областью, как у модифицированного антитела, то есть, чем у целого антитела (далее по тексту "исходное антитело"), например IgG, имеющего ту же пару V-области Н-цепи и V-области L-цепи, что и пара V-области Н-цепи и V-области L-цепи, образующая антигенсвязывающую область модифицированного антитела. Более предпочтительными являются модифицированные антитела, у которых ТРО-агонистическое действие (ED50) более чем в два раза выше, чем у исходного антитела, еще более предпочтительны модифицированные антитела с более чем 5-кратным превышением, наиболее предпочтительны с более чем 10-кратным превышением. Настоящее изобретение относится к модифицированным антителам с ТРО-агонистическим действием, содержащим V-область Н-цепи и V-область L-цепи, образующие такую же антигенсвязывающую область, как и исходное антитело, которое связывается с ТРО-рецептором, однако не обладает ТРО-агонистическим действием в отношении молекулы.
Соединения, содержащие две или более V-области Н-цепи и две или более V-области L-цепи по настоящему изобретению, - это любые соединения, которые содержат две или более V-области Н-цепи и две или более V-области L-цепи антитела и оказывают ТРО-агонистическое действие (ED50), равное или лучшее, чем у тромбопоэтина (ТРО). Предпочтительными являются такие, у которых ТРО-агонистическое действие (ED50) выше, чем у ТРО, более чем в два раза, более предпочтительны с более чем 5-кратным превышением, наиболее предпочтительны с более чем 10-кратным превышением.
Упоминаемые здесь "соединения" включают не только модифицированные антитела по изобретению, но также и любые соединения, содержащие две или более, предпочтительно от 2 до 6, более предпочтительно от 2 до 4, наиболее предпочтительно 2, антигенсвязывающие области, такие, как целые антитела или F(ab')2.
У предпочтительных модифицированных антител или соединений по настоящему изобретению, содержащих две или более V-области Н-цепи и две или более V-области L-цепи антитела, межклеточное адгезионное действие (ED50) не превышает 1/10 сравнительно с исходным антителом, у более предпочтительных сколько-нибудь существенное межклеточное адгезионное действие отсутствует.
Вышеуказанное ED50 межклеточного адгезионного действия определяется стандартными способами измерения межклеточного адгезионного действия, например измерением агломерации клеток, экспрессирующих ТРО-рецептор.
Изобретение относится к молекулам ДНК, кодирующим модифицированные антитела.
Изобретение относится к клеткам животных или микроорганизмов, которые продуцируют модифицированные антитела.
Изобретение относится к использованию модифицированного антитела в качестве ТРО-агониста.
Изобретение относится к способу трансдукции сигнала в клетки путем поперечного сшивания ТРО-рецептора с помощью модифицированного антитела и индуцирования таким образом ТРО-агонистического действия, как, например, пролиферации, дифференцировки-индукции, или стимуляции роста мегакариоцитов, образования тромбоцитов, фосфорилирования белка ТРО-рецептора и т.д.
Изобретение относится к лекарственному средству для лечения тромбоцитопении и т.п., которое содержит модифицированное антитело в качестве активного компонента.
Изобретение относится к применению модифицированного антитела в качестве лекарственного средства.
Изобретение относится к способу скрининга или количественного определения модифицированного антитела, которое содержит две или более V-области Н-цепи и две или более V-области L-цепи антитела и оказывает ТРО-агонистическое действие путем поперечного сшивания ТРО-рецептора, причем способ включает 1) получение модифицированного антитела, содержащего две или более V-области Н-цепи и две или более V-области L-цепи антитела и специфически связывающегося с ТРО-рецептором, 2) контактирование модифицированного антитела с клетками, экспрессирующими ТРО-рецептор и 3) измерение ТРО-агонистического действия, которое происходит в клетках путем образования поперечных сшивок ТРО-рецептора. Способ количественного определения применяется для контроля качества при производстве модифицированных антител по изобретению в качестве лекарственных средств и в других целях.
Модифицированные антитела могут быть как моноспецифичными модифицированными антителами, так и мультиспецифичными модифицированными антителами, например биспецифичными модифицированными антителами. Предпочтительными являются моноспецифичные модифицированные антитела.
Настоящее изобретение относится также к модифицированным антителам, у которых V-область Н-цепи и/или V-область L-цепи - это V-область Н-цепи, происходящая из человеческого антитела, и/или V-область L-цепи, происходящая из человеческого антитела. V-область Н-цепи и/или V-область L-цепи, происходящие из человеческого антитела, можно получить путем скрининга библиотеки моноклонального антитела, как это описано в WO 99/10494. V-область Н-цепи и/или V-область L-цепи, происходящие из человеческих моноклональных антител, продуцируемых трансгенными мышами и т.п., также включены в изобретение.
Кроме того, изобретение относится также к модифицированным антителам, у которых V-области Н-цепи и/или V-области L-цепи являются гуманизированными V-областями Н-цепи и/или гуманизованными V-областями L-цепи. Конкретно, гуманизированные модифицированные антитела состоят из гуманизированной V-области L-цепи, которая содержит каркасные области (FR), происходящие из V-области L-цепи человеческого моноклонального антитела, и области, определяющие комплементарность (далее по тексту "CDR"), происходящие из V-области L-цепи моноклонального антитела какого-либо млекопитающего, кроме человека (например, мышь, крыса, корова, овца, человекообразная обезьяна), и/или гуманизированной V-область Н-цепи, содержащей FR, происходящие из V-области Н-цепи человеческого моноклонального антитела, и CDR, происходящие из V-области Н-цепи моноклонального антитела какого-либо млекопитающего, кроме человека (например, мышь, крыса, корова, овца, человекообразная обезьяна). В этом случае аминокислотные последовательности CDR и FR могут быть частично изменены, например делетированы, замещены или добавлены.
V-области Н-цепи и/или V-области L-цепи модифицированных антител по изобретению могут быть V-областями Н-цепи и/или V-областями L-цепи, происходящими из моноклональных антител млекопитающих, кроме человека (например, мышь, крыса, корова, овца, человекообразная обезьяна, цыпленок и т.п.). В этом случае аминокислотные последовательности CDR и FR могут быть частично изменены, например делетированы, замещены или добавлены.
Изобретение относится также к молекулам ДНК, кодирующим различные модифицированные антитела, как это было указано выше, и к генно-инженерным методам получения рекомбинантных векторов, включающих ДНК.
Изобретение относится также к клеткам-хозяевам, трансформированным рекомбинантными векторами. Примерами клеток-хозяев могут быть человеческие клетки, мышиные и т.п. и микроорганизмы, как Е.colli. Bacillus sublilis. дрожжи и т.д.
Изобретение относится также к способу получения модифицированных антител, который включает культивирование вышеупомянутых клеток-хозяев и экстрагирование модифицированных антител из полученной культуры.
Кроме того, изобретение относится также к способу получения димера одноцепочечного Fv, который включает культивирование животных клеток-хозяев, продуцирующих одноцепочечный Fv, в бессывороточной среде для секреции одноцепочечного Fv в среду и выделение димера одноцепочечного Fv, образующегося в среде.
Изобретение относится также к применению модифицированных антител в качестве ТРО-агониста. Это значит, что оно относится к агонисту трансдукции сигнала, который включает в себя в качестве активной составляющей модифицированное антитело, полученное вышеуказанным способом.
Таким образом, фармацевтические препараты, содержащие ТРО-агонистические модифицированные антитела по изобретению в качестве активного компонента, могут быть использованы в качестве профилактических или лечебных средств при заболеваниях крови, связанных с уменьшением содержания тромбоцитов, тромбоцитопении, вызванной химиотерапией при раковых заболеваниях или лейкемии, и т.п.
Модифицированные антитела настоящего изобретения содержат две или более V-области Н-цепи и две или более V-области L-цепи, происходящие из антител. По своей структуре модифицированное антитело может быть димером одноцепочечного Fv, содержащего одну V-область Н-цепи и одну V-область L-цепи, или полипептидом, содержащим две V-области Н-цепи и две V-области L-цепи. У модифицированных антител по изобретению V-области Н-цепи и L-цепи связаны предпочтительно через пептидный линкер, который состоит из одной или более аминокислот. Получающиеся модифицированные антитела содержат вариабельные области антител и связываются с антигеном с такой же специфичностью, что и исходные моноклональные антитела.
V-область Н-цепи
Согласно настоящему изобретению, V-область Н-цепи, происходящая из антитела, распознает ТРО-рецептор и олигомеризует, например димеризует, вышеназванную молекулу путем образования поперечной сшивки, трансдуцируя таким образом сигнал в клетки. V-область Н-цепи по настоящему изобретению включает V-области Н-цепи, происходящие из какого-либо млекопитающего (например, человеческие, мышиные, крысиные, бычьи, овечьи, человекообразных обезьян и т.п.), и V-области Н-цепи, содержащие частично модифицированные аминокислотные последовательности V-областей Н-цепей. Более предпочтительной является гуманизированная V-область Н-цепи, содержащая FR V-области Н-цепи человеческого моноклонального антитела и CDR-участок V-области Н-цепи мышиного моноклонального антитела. Также предпочтительной является V-область Н-цепи, имеющая аминокислотную последовательность, происходящую из человека, которую можно получить рекомбинантным методом. V-область Н-цепи по изобретению может быть фрагментом вышеуказанной V-области Н-цепи, каковой фрагмент сохраняет способность связывать антиген.
V-область L-цепи
Согласно настоящему изобретению, V-область L-цепи, происходящая из антитела, распознает ТРО-рецептор и олигомеризует, например димеризует, вышеназванную молекулу путем образования поперечной сшивки, трансдуцируя таким образом сигнал в клетки. V-область L-цепи по настоящему изобретению включает V-области L-цепи, происходящие из млекопитающего (например, человеческие, мышиные, крысиные, бычьи, овечьи, человекообразных обезьян и т.п.), и V-области L-цепи, содержащие частично модифицированные аминокислотные последовательности V-областей L-цепи. Более предпочтительной является гуманизированная V-область L-цепи, содержащая FR V-области L-цепи человеческого моноклонального антитела и CDR V-области L-цепи мышиного моноклонального антитела. Также предпочтительной является V-область L-цепи, имеющая аминокислотную последовательность, происходящую из человека, которую можно получить рекомбинантным методом. V-область L-цепи по изобретению может быть фрагментом вышеуказанной V-области L-цепи, каковой фрагмент сохраняет способность связывать антиген.
Область, определяющая комплементарность (CDR)
Каждая V-область L-цепи и Н-цепи образует антигенсвязывающий сайт. Вариабельная область L- и Н-цепей составлена из четырех сравнительно консервативных общих каркасных областей, соединенных с тремя гипервариабельными участками, или областей, определяющих комплементарность (CDR) (Kabat E.A. et al., "Sequences of Protein of Immunological Interest", US Dept. Health and Human Services, 1983).
Главные части четырех каркасных областей (FRs) образуют β-складчатые структуры, а три CDR образуют, таким образом, петлю. В некоторых случаях CDR могут образовывать часть β-складчатой структуры. Три CDR удерживаются в пространственно близком положении друг к другу с помощью FR, который вносит вклад в формирование антигенсвязывающего сайта наряду с тремя CDR.
Эти CDR можно идентифицировать, сравнивая аминокислотную последовательность V-области полученного антитела с установленными аминокислотными последовательностями V-областей известных антител согласно эмпирическому правилу Kabat E.A. et al., "Sequences of Protein of Immunological Interest".
Одноцепочечный Fv.
Одноцепочечный Fv - это полипептитдный мономер, содержащий связанные друг с другом V-область Н-цепи и V-область L-цепи, происходящие из антител. Полученные одноцепочечные Fv включают вариабельные области исходных антител и сохраняют области, определяющие комплементарность, и, следовательно, одноцепочечные Fv связываются с антигеном с той же специфичностью, что и исходные антитела (JP-Appl. 11-63557). Часть вариабельной области и/или CDR одноцепочечного Fv по изобретению или часть ее аминокислотной последовательности можно частично изменить, например, делетировать, заместить или присоединить. Вышеуказанные V-область Н-цепи и V-область L-цепи, входящие в состав одноцепочечного Fv изобретения, можно соединять непосредственно или через линкер, предпочтительно через пептидный линкер. У одноцепочечного Fv может быть следующее строение: [V-область Н-цепи] - [V-область L-цепи] или [V-область L-цепи] - [V-область Н-цепи]. Согласно настоящему изобретению можно сделать так, чтобы одноцепочечный Fv образовывал димер, тример или тетрамер, из которых может быть образовано модифицированное антитело изобретения. Одноцепочечное модифицированное антитело
Одноцепочечные модифицированные антитела настоящего изобретения, содержащие две или более V-области Н-цепи и две или более V-области L-цепи, предпочтительно от двух до четырех каждой, особенно предпочтительно по две каждой, содержат две или более V-области Н-цепи и V-области L-цепи, как указано выше. Каждая из областей пептида должна быть организована так, чтобы модифицированное одноцепочное антитело образовывало специфическую пространственную структуру, точно имитирующую пространственную структуру, образуемую димером одноцепочечного Fv. Например, V-области можно расположить в нижеследующем порядке:
[V-область Н-цепи] - [V-область L-цепи] - [V-область Н-цепи] - [V-область L-цепи] или [V-область L-цепи] - [V-область Н-цепи] - [V-область L-цепи] - [V-область Н-цепи], где данные области соединяются соответственно через пептидный линкер.
Линкер
Согласно изобретению, линкером для соединения V-области Н-цепи с V-областью L-цепи может быть любой пептидный линкер, который может быть введен с помощью методов генной инженерии, или любой химически синтезируемый линкер. Например, согласно изобретению, можно использовать линкеры, раскрытые в литературе, например, Protein Engineering, 9(3), 299-305, 1996. Линкеры в одной и той же молекуле могут быть одинаковыми или разными. Если требуются пептидные линкеры, то ниже приводятся их примеры:
Ser
Gly-Ser
Gly-Gly-Ser
Ser-Gly-Gly
Gly-Gly-Gly-Ser
Ser-Gly-Gly-Gly
Gly-Gly-Gly-Gly-Ser
Ser-Gly-Gly-Gly-Gly
Gly-Gly-Gly-Gly-Gly-Ser
Ser-Gly-Gly-Gly-Gly-Gly
Gly-Gly-Gly-Gly-Gly-Gly-Ser
Ser-Gly-Gly-Gly-Gly-Gly-Gly
(Gly-Gly-Gly-Gly-Ser)n и
(Ser-Gly-Gly-Gly-Gly)n,
где n - целое число не менее единицы. Предпочтительная длина пептида-линкера варьирует в зависимости от того, какой рецептор будет антигеном, для одноцепочечных Fv обычно предпочтительным является диапазон 1-20 аминокислот. В случае одноцепочечных модифицированных антител, содержащих две или более V-области Н-цепи и две или более V-области L-цепи, пептидные линкеры, соединяющие области, образующие один и тот же антигенсвязывающий центр, включающий [V-область Н-цепи] - [V-область L-цепи] (или [V-область L-цепи] - [V-область Н-цепи]), имеют длину от 1-30 аминокислот, предпочтительно 1-20 аминокислот, более предпочтительно 3-18 аминокислот. Пептидные линкеры, соединяющие области, не образующие общий антигенсвязывающий центр, включающий [V-область Н-цепи] - [V-область L-цепи] (или [V-область L-цепи] - [V-область Н-цепи]), имеют длину от 1-40 аминокислот, предпочтительно 3-30 аминокислот, более предпочтительно 5-20 аминокислот. Способ введения этих линкеров будет описан ниже при объяснении конструирования ДНК, кодирующей модифицированные антитела изобретения.
Химически синтезированными линкерами, то есть химическими поперечносшивающими агентами, согласно изобретению, могут быть любые линкеры, применяемые обычно для сшивания пептидов. В число таких линкеров входят N-гидроксисукцинимид (NHS), дисукцинимидилсуберат (DSS), бис(сульфосукцинимидил)суберат (BS3), дитиобис(сукцинимидилпропионат) (DSP), дитиобис(сульфосукцинимидилпропионат) (DTSSP), этиленгликольбис(сукцинимидилсукцинат) (EGS), этиленгликольбис(сульфосукцинимидилсукцинат) (сульфо-EGS), дисукцинимидилтартрат (DST), дисульфосукцинимидилтартрат (сульфо-DST), бис[2-(сукцинимидооксикарбонилокси)этил]сульфон (BSOCOES), бис[2-(сульфосукцинимидооксикарбонилокси)этил]сульфон (сульфо-BSOCOES) и т.п. Все они являются коммерчески доступными. Предпочтительно, чтобы химически синтезированные линкеры имели длину, равную длине пептидных линкеров.
Для получения димера одноцепочечного Fv предпочтительно выбирать линкер, подходящий для димеризации в таком растворе, как культурная среда, более 20% одноцепочечных Fv, продуцируемых в клетках-хозяевах, предпочтительно более 50%, более предпочтительно более 80%, наиболее предпочтительно более 90%. Особенно предпочтительным является линкер, включающий от 2 до 12 аминокислот, предпочтительно от 3 до 10 аминокислот, или же другие линкеры, соответствующие им.
Получение модифицированных антител
Модифицированные антитела можно получать присоединением, через вышеуказанный линкер, V-области Н-цепи и V-области L-цепи, происходящих из известных или новых антител, специфически связывающихся с ТРО-рецептором. В качестве примеров одноцепочечных Fv приводятся те, что содержат V-область Н-цепи и V-область L-цепи антитела 12В5 и антитела 12Е10, раскрытых в WO 99/10494. В качестве примеров модифицированных антител по изобретению, содержащих две или более V-области Н-цепи и две или более V-области L-цепи, приводятся димер sc12B5 (линкер: 15 аминокислот), димер sc12E10 (линкер: 15 аминокислот), димер db12B5 (линкер: 5 аминокислот), димер db12E10 (линкер: 5 аминокислот), sc12B5sc(FV)2 и sc12E10sc(FV)2, которые содержат V-области Н-цепи и V-области L-цепи, происходящие из вышеуказанных моноклональных антител.
При получении модифицированных антител сигнальный пептид можно присоединять к его N-концу, если нужно, чтобы полученный полипептид был секреторным пептидом. Для эффективной очистки полипептида можно применить такую широко используемую для очистки полипептидов аминокислотную последовательность, как FLAG-последовательность. В этом случае димер можно получить с использованием анти-FLAG антитела.
Для получения модифицированного антитела по изобретению необходимо получить ДНК, то есть ДНК, кодирующую одноцепочечный Fv, или ДНК, кодирующую реконструированный одноцепочечный полипептид. Эти ДНК могут быть получены, особенно для sc12B5, db12B5, sc12E10 и/или db12E10, из ДНК, кодирующих V-области Н-цепи и V-области L-цепи, происходящие из указанных Fv. Их можно также получить методом полимеразной цепной реакции (ПЦР), используя эти ДНК в качестве матрицы и амплифицируя фрагменты ДНК, содержащиеся в них, которые кодируют требуемую аминокислотную последовательность, с помощью двух праймеров, соответствующих обоим концам последовательности.
Если нужно, чтобы каждая V-область содержала частично модифицированную аминокислотную последовательность, то такие V-области, в которых модифицированы, то есть утрачены, замещены или добавлены одна или несколько аминокислот, можно получить с помощью хорошо известного из уровня техники метода ПЦР. Для того чтобы получить модифицированное антитело, которое было бы достаточно активным в отношении специфического антигена, предпочтительно модифицировать часть аминокислотной последовательности V-области методом ПЦР, хорошо известным из уровня техники.
Для выбора праймеров для аплификации методом ПЦР, если в качестве исходного материала используется моноклональное антитело, определяют типы Н-и L-цепей методом типирования, известным из уровня техники.
Для амплификации V-областей L-цепи антитела 12В5 и антитела 12Е10 методом ПЦР 5'-концевой и 3'-концевой олигонуклеотидные праймеры выбирают, как упомянуто выше. Таким же образом выбирают 5'-концевой и 3'-концевой олигонуклеотидные праймеры для амплификации V-областей Н-цепи антитела 12В5 и антитела 12Е10.
В воплощениях изобретения используются 5'-концевые праймеры, которые содержат последовательность "GANTC", обеспечивающую сайт узнавания рестриктазы Hinf I, вблизи от их S'-конца, и 3''-концевые, которые содержат нуклеотидную последовательность "CCCGGG", обеспечивающую сайт узнавания XmaI, вблизи от их 5'-конца. Вместо этих сайтов можно использовать сайты узнавания других рестриктаз до тех пор, пока их применяют для субклонирования требуемых фрагментов ДНК в клонирующий вектор.
Специально сконструированные ПЦР-праймеры применяют для того, чтобы обеспечить соответствующие нуклеотидные последовательности на 5'-конце и 3'-конце кДНК, кодирующей V-области антител 12В5 и 12Е10, так, чтобы кДНК легко встраивалась в экспрессионный вектор и функционировала бы в эекспрессионном векторе соответствующим образом (т.е. изобретение предусматривает увеличение эффективности транскрипции за счет встраивания последовательности Козака). V-области антител 12В5 и 12Е10, полученные амплификацией методом ПЦР с использованием таких праймеров, встраивают в экспрессионный вектор HEF, содержащий желаемую С-область человека (см. WO 92/19759). Клонированные ДНК можно секвенировать любым общепринятым способом, например, с помощью автоматического секвенатора ДНК (Applied Biosystems).
Линкер, такой как, например, пептидный линкер, можно ввести в модифицированное антитело изобретения следующим образом. Конструируют праймеры, которые имеют последовательности, частично комплементарные праймерам для V-областей Н-цепей и V-областей L-цепей, описанным выше, и которые кодируют N-конец и С-конец линкера. Затем можно провести ПЦР с использованием этих праймеров для получения ДНК, кодирующей пептидный линкер, имеющий желаемые аминокислотную последовательность и длину. ДНК, кодирующие V-область Н-цепи и V-область L-цепи, можно состыковать с помощью полученной ДНК, чтобы получить ДНК, кодирующую модифицированное антитело изобретения, содержащее требуемый пептидный линкер. Как только получена ДНК, кодирующая одно из модифицированных антител, легко можно получить ДНК, кодирующие модифицированные антитела с требуемым пептидным линкером или без него, путем конструирования различных праймеров для линкера, а затем проведения ПЦР с использованием праймеров и вышеуказанной ДНК в качестве матрицы.
Каждую V-область модифицированного антитела настоящего изобретения можно гуманизировать по общепринятой методике (Sato К. et al., Cancer Res., 53, 1-6 (1993)). Как только получена ДНК, кодирующая каждый из гуманизированных Fv, легко можно получить гуманизированный одноцепочечный Fv, фрагмент гуманизированного одноцепочечного Fv, гуманизированное моноклональное антитело и фрагмент гуманизованного моноклонального антитела в соответствии с общепринятой методикой. Предпочтительно, чтобы аминокислотные последовательности их V-областей могли быть при необходимости частично модифицированы.
Далее, можно получить ДНК, происходящую от другого млекопитающего, например, ДНК, кодирующую каждую из V-областей антитела человека, таким же способом, каким была получена ДНК, кодирующая V-область Н-цепи и V-область L-цепи, происходящие из мыши, в соответствии с общепринятыми методами, упомянутыми выше. Результирующую ДНК можно использовать для получения V-области Н-цепи и V-области L-цепи других млекопитающих, в частности, происходящих из антитела человека, одноцепочечный Fv, происходящий из человека, и его фрагмент, и моноклональное антитело человеческого происхождения и его фрагмент.
В случаях, когда модифицированные антитела, соответствующие изобретению, являются биспецифическими модифицированными антителами, их получают одним из известных из уровня техники способов (например, способ, описанный в WO 9413804).
Как было указано выше, когда получены требуемые ДНК, кодирующие V-области модифицированных антител и V-области гуманизированных модифицированных антител, то можно получить содержащие их экспрессионные векторы и хозяев, трансформированных этими векторами, стандартными способами. Кроме того, стандартными методами можно культвировать хозяев, для того чтобы получить реконструированный одноцепочечный Fv, реконструированный гуманизированный одноцепочечный Fv, гуманизированные моноклональные антитела и их фрагменты. Их можно выделить из клеток или из среды и очистить до гомогенной массы. В этих целях можно применять любые стандартные методы выделения и очистки, применяемые для белков, например хроматографию, ультрафильтрацию, высаливание и диализ, при необходимости комбинируя их без каких-либо ограничений.
Если реконструированный одноцепочечный Fv настоящего изобретения получают культивированием животной клетки, такой как, например, COS7-клетки или СНО-клетки, предпочтительно СНО-клетки, в бессывороточной среде, то можно стабильно получать и очищать с высоким выходом образующийся в данной среде димер Fv. Такой очищенный димер может стабильно храниться в течение длительного срока. В качестве бессывороточной среды, применяемой согласно изобретению, можно использовать любую среду, стандартно применяемую при получении рекомбинантных белков, без каких-либо ограничений.
Для получения модифицированных антител настоящего изобретения можно использовать любые системы экспрессии, например, такие эукариотичные клетки, как животные клетки, например, адаптированные линии клеток млекопитающих, нитчатые грибы и дрожжи, и такие прокариотичные клетки, как клетки бактерий, например, Е.coli. Модифицированные антитела по изобретению предпочтительно экспрессировать в клетках млекопитающих, например, COS7-клетках или СНО-клетках.
В таких случаях можно использовать стандартные промоторы, пригодные для экспрессии в клетках млекопитающих. Предпочтительно использовать немедленно-ранний промотор цитомегаловируса человека (HCMV). В число экспрессионных векторов, содержащих HCMV-промотор, входят HCMV-VH-HCγ 1, HCMV-VL-HCK и т.п., которые получены из pSV2neo (WO 92/19759).
Примеры других промоторов для экспрессии генов в клетках млекопитающих, которые можно использовать согласно изобретению, дополнительно включают вирусные промоторы, происходящие из ретровируса, вируса полиомы, аденовируса и обезьянего вируса 40 (SV40), и такие промоторы, происходящие из млекопитающих, как, например, фактор-1α элонгации полипептидной цепи человека (HEF-1α). Промотор SV40 легко может быть использован по способу Mulligan R.C. et al. (Nature 277, 108-114 (1979), а промотор HEF-1α - по способу Mizushima S. et al. (Nucleic Acids Research, 18, 5322 (1990).
Примеры ориджина - точки инициации репликации (ori), которые могут, согласно изобретению, быть использованы, включают ori, происходящие из SV40, вируса полиомы, аденовируса, вируса бычьей папилломы (BPV) и т.п. Экспрессионный вектор может включать в качестве селективного маркера ген фосфотрансферазы АРН (3') II или I (neo), ген тимидинкиназы (ТК), ген ксантин-гуанинфосфорибосилтрансферазы Е.coli (Ecogpt) или ген дигидрофолатредуктазы (DHFR).
Антигенсвязывающую активность полученного выше модифицированного антитела можно количественно определить стандартными методами, такими как, например, радиоиммуноанализ (РИА), ферментный иммуносорбентный анализ (ELISA) или поверхностный плазменный резонанс. Ее можно также оценить, используя в качестве показателя способность ингибировать связывание исходного антитела, например, по отсутствию или наличию зависящего от концентрации ингибирования связывания указанного моноклонального антитела с антигеном.
Если более подробно, то культивируют клетки животного происхождения, трансформированные экспрессионным вектором, содержащим ДНК, кодирующую модифицированное антитело, соответствующее изобретению, например культивируют клетки COS7 или клетки СНО. Культивируемые клетки и/или супернатант среды или очищенное модифицированное антитело используют для определения связывания с антигеном. Для контроля используют супернатант культуральной среды, в которой культивировались клетки, трансформированные одним только экспрессионным вектором. В случае антигена, например, для антитела 12В5 и антитела 12Е10 к клеткам Ba/F3, экспрессирующим MPL человека, добавляют исследуемый образец модифицированного антитела по изобретению или супернатант контрольной пробы, а затем проводят анализ, например, методом проточной цитометрии для количественного определения антигенсвязывающей активности.
В условиях in vitro количественую оценку эффекта трансдукции сигнала (например, пролиферации, дифференцировки-индукции или стимуляции роста мегакариоцитов, образования тромбоцитов или фосфорилирования белка ТРО-рецептора) выполняют следующим образом. Исследуемый образец вышеуказанного модифицированного антитела добавляют к клеткам, экспрессирующим антитело, или к клеткам, в которые введен ген данного антитела, и по изменениям, вызванным трансдукцией сигнала (например, по пролиферации, специфичной в отношении MPL-антигена человека, по измерению фосфорилирования белков или экспрессии тромбоцитоспецифического антигена), дают количественную оценку стандартными методами.
В условиях in vivo количественное определение проводят путем введения мышам моноклонального антитела, распознающего MPL, модифицированного антитела по изобретению и забуференного фосфатом физиологического раствора (PBS) в качестве контроля и путем количественного определения степени активности по изменению количества тромбоцитов в мышиной сыворотке.
Как указано выше, модифицированные антитела изобретения могут быть получены конструированием модифицированных антител, которые содержат две или более V-области Н-цепи и две или более V-области L-цепи и специфически связываются с ТРО-рецептором, и проводя скрининг модифицированных антител с использованием способов анализа in vivo или in vitro, как это описано выше.
Модифицированные антитела по изобретению, которые содержат две или более V-области Н-цепи и две или более V-области L-цепи, предпочтительно от 2 до 4 каждой, более предпочтительно по 2 каждой, могут быть димерами одноцепочного Fv, содержащего одну V-область Н-цепи и одну V-область L-цепи, или одноцепочными полипептидами, в молекуле которых соединены две или более V-области Н-цепи и V-области L-цепи. Полагают, что благодаря такому строению, пептид имитирует трехмерную структуру ТРО, а следовательно, сохраняет превосходную антигенсвязывающую способность и ТРО-агонистическую активность.
Модифицированные антитела изобретения имеют значительно меньшие молекулярные размеры по сравнению с молекулой исходного антитела (например, IgG) и поэтому отличаются высокой проникающей способностью в ткани и опухоли и более высокой активностью, чем молекулы исходного моноклонального антитела. Следовательно, модифицированные антитела изобретения способны эффективно трансдуцировать ТРО-сигнал в клетки. Фармацевтические композиции, их содержащие, могут применяться для лечения заболеваний крови, связанных со снижением количества тромбоцитов, и тромбоцитопении, вызываемой химиотерапией при раковых заболеваниях и лейкемии. Помимо этого, антитело по изобретению можно будет использовать как контрастный агент при радиоиммунном мечении. Эффект можно усилить путем присоединения к радиоиммунному соединению или токсину.
Перечень чертежей
Фиг.1 иллюстрирует результаты проточной цитометрии, показывающие, что антитело IgG человека не связывается клетками L1210, экспрессирующими IAP человека (hIAP/L1210).
Фиг.2 иллюстрирует результаты проточной цитометрии, показывающие, что химерное антитело MABL-1 специфически связывается с клетками L1210, экспрессирующими IAP человека (hIAP/L1210).
Фиг.3 иллюстрирует результаты проточной цитометрии, показывающие, что химерное антитело MABL-2 специфически связывается с клетками L1210, экспрессирующими IAP человека (hIAP/L1210).
Фиг.4 схематически иллюстрирует способ получения одноцепочечного Fv в соответствии с настоящим изобретением.
Фиг.5 отображает структуру экспрессионной плазмиды, которую можно использовать для экспрессии ДНК, кодирующей одноцепочечный Fv изобретения, в Е coli.
Фиг.6 отображает структуру экспрессионной плазмиды, которая используется для экспрессии ДНК, кодирующей одноцепочечный Fv изобретения, в клетках млекопитающих.
Фиг.7 иллюстрирует результаты Western блоттинга из Примера 5.4. Слева: маркер молекулярных масс (сверху вниз 97,4, 66, 45, 31, 21,5 и 14,5 кДа), культуральный супернатант клеток COS7, содержащих pCHO1, и культуральный супернатант клеток COS7, содержащих рСНОМ2. Здесь показано, что реконструированный одноцепочечный Fv антитела MABL-2 (стрелка) содержится в супернатанте клеток, содержащих рСНОМ2.
Фиг.8 иллюстрирует результаты проточной цитометрии, показывающие, что контрольное антитело из культурального супернатанта клеток pCHO1/COS7 не связывается с контрольными клетками pCOS1/L1210.
Фиг.9 иллюстрирует результаты проточной цитометрии, показывающие, что антитело из культурального супернатанта клеток MABL2-scFv/COS7 не связывается с контрольными клетками pCOS1/L1210.
Фиг.10 иллюстрирует результаты проточной цитометрии, показывающие, что антитело из культурального супернатанта клеток pCOS1/COS7 в качестве контроля не связывается с клетками hIAP/L1210.
Фиг.11 иллюстрирует результаты проточной цитометрии, показывающие, что антитело из культурального супернатанта клеток MABL2-scFv/COS7 специфически связывается с клетками hIAP/L1210.
Фиг.12 иллюстрирует результаты конкурентного иммуноанализа ELISA из Примера 5.6, где связывающая активность одноцепочечного Fv изобретения (MABL2-scFv) в отношении антигена, сравнительно с культуральным супернатантом контрольных клеток pCHO1/COS7, выражена в форме ингибирования связывания антигена моноклональным антителом MABL-2 мыши, принятого за показатель.
Фиг.13 иллюстрирует результаты апоптоз-индуцирующего эффекта из Примера 5.7, показывающие, что контрольное антитело из культурального супернатанта клеток pCHO1/COS7 не индуцирует апоптоз контрольных клеток pCOS1/L1210.
Фиг.14 иллюстрирует результаты апоптоз-индуцирующего эффекта из Примера 5.7, показывающие, что антитело из культурального супернатанта клеток MABL2-scFv/COS7 не индуцирует апоптоз контрольных клеток pCOS1/L1210.
Фиг.15 иллюстрирует результаты апоптоз-индуцирующего эффекта из Примера 5.7, показывающие, что контрольное антитело из культурального супернатанта клеток pCHO1/COS7 не индуцирует апоптоз клеток hIAP/L1210.
Фиг.16 иллюстрирует результаты апоптоз-индуцирующего эффекта из Примера 5.7, показывающие, что контрольное антитело из культурального супернатанта клеток MABL2-scFv/COS7 специфически индуцирует апоптоз клеток hIAP/L1210.
Фиг.17 иллюстрирует результаты апоптоз-индуцирующего эффекта из Примера 5.7, показывающие, что контрольное антитело из культурального супернатанта клеток pCHO1/COS7 не индуцирует апоптоз клеток CCRF-CEM (при 50%-ной конечной концентрации).
Фиг.18 иллюстрирует результаты апоптоз-индуцирующего эффекта из Примера 5.7, показывающие, что антитело из культурального супернатанта клеток MABL2-scFv/COS7 специфически индуцирует апоптоз клеток CCRF-CEM (при 50%-ной конечной концентрации).
Фиг.19 изображает хроматограмму, полученную при очистке одноцепочечного Fv, происходящего из антитела MABL-2, продуцируемого клетками СНО из Примера 5.9, которая показывает, что фракция А и фракция В получаются в виде основных пиков при очистке фракции с колонки с Blue-Sepharose на колонке с гидроксиапатитом.
Фиг.20 иллюстрирует результаты очистки гель-фильтрацией фракции А и фракции В, полученных в Примере 5.9-(2), показывающие, что основные пики (AI и BI соответственно) элюируются в случае фракции А с кажущимся молекулярным весом около 36 кДа и в случае фракции В - около 76 кДа.
Фиг.21 иллюстрирует SDS-PAGE-анализ фракций, полученных при очистке одноцепочечного Fv, происходящего из антитела MABL-2, продуцируемого клетками СНО в Примере 5.9, показывающий, что в обеих фракциях наблюдается единственная полоса с молекулярным весом около 35 кДа.
Фиг.22 отражает результаты анализа фракций AI и BI, полученных гель-фильтрацией при очистке одноцепочечного Fv, происходящего из антитела MABL-2, продуцируемого клетками СНО, где фракция AI включает мономер, а фракция BI включает димер.
Фиг.23 иллюстрирует структуру экспрессионной плазмиды, которую можно использовать для экспрессии ДНК, кодирующей одноцепочечный Fv изобретения, в E.coli.
Фиг.24 иллюстрирует результаты очистки на гель-фильтрационной колонке грубого препарата одноцепочечного Fv полипептида, происходящего из антитела MABL-2, продуцируемого Е.coli. полученных в Примере 5.12, где пики соответствуют мономеру или димеру одноцепочечного Fv, продуцируемого Е.coli.
Фиг.25 иллюстрирует результаты апоптоз-индуцирующего действия из Примера 5.13, показывающие, что контрольное антитело IgG мыши не индуцирует апоптоз клеток hIAP/L1210 (конечная концентрация 3 мкг/мл).
Фиг.26 иллюстрирует результаты апоптоз-индуцирующего действия из Примера 5.13, показывающие, что димер MABL2-scFv, продуцируемый клетками СНО, в значительной степени индуцирует апоптоз клеток hIAP/L1210 (конечная концентрация 3 мкг/мл).
Фиг.27 иллюстрирует результаты апоптоз-индуцирующего действия из Примера 5.13, показывающие, что димер MABL2-scFv, продуцируемый Е.coli., в значительной степени индуцирует апоптоз клеток hIAP/L1210 (конечная концентрация 3 мкг/мл).
Фиг.28 иллюстрирует результаты апоптоз-индуцирующего действия из Примера 5.13, показывающие, что индукция апоптоза клеток hIAP/L1210 мономером MABL2-scFv, продуцируемым клетками СНО, по величине такая же, что и в контроле (конечная концентрация 3 мкг/мл).
Фиг.29 иллюстрирует результаты апоптоз-индуцирующего действия из Примера 5. 13, показывающие, что индукция апоптоза клеток hIAP/L1210 мономером MABL2-scFv, продуцируемым Е.coli., по величине такая же, что и в контроле (конечная концентрация 3 мкг/мл).
Фиг.30 иллюстрирует результаты апоптоз-индуцирующего действия из Примера 5. 13, показывающие, что контрольное антитело IgG мыши не индуцирует апоптоз у клеток hIAP/L1210 даже при добавлении анти-FLAG антитела (конечная концентрация 3 мкг/мл).
Фиг.31 иллюстрирует результаты апоптоз-индуцирующего действия из Примера 5.13, показывающие, что мономер MABL2-scFv, продуцируемый клетками СНО, в значительной степени индуцирует апоптоз клеток hIAP/L1210 при условии добавления анти-FLAG антитела (конечная концентрация 3 мкг/мл).
Фиг.32 иллюстрирует результаты количественного определения IgG человека в сыворотке мыши с трансплантированной клеточной линией миеломы человека КРММ2, показывающие количество IgG человека, продуцируемого клетками миеломы человека в мыши. Видно, что димер scFv/CHO в значительной степени тормозит рост клеток КРММ2.
Фиг.33 иллюстрирует продолжительность выживания мыши после трансплантации опухоли, показывая, что у группы, которой был введен димер scFv/CHO, продолжительность выживания значительно увеличивается.
Фиг.34 иллюстрирует структуру экспрессионной плазмиды, которая экспрессирует модифицированное антитело [sc(Fv)2], содержащее две V-области Н-цепи и две V-области L-цепи, происходящие из антитела MABL-2.
Фиг.35 иллюстрирует структуру плазмиды, которая экспрессирует scFv (HL-тип) в котором V-области связаны без пептидного линкера следующим образом: [Н-цепь] - [L-цепь].
Фиг.36 иллюстрирует структуру полипептида HL-типа и аминокислотные последовательности пептидных линкеров.
Фиг.37 иллюстрирует структуру плазмиды, которая экспрессирует scFv (LH-тип), в котором V-области соединены без пептидного линкера следующим образом: [L-цепь] - [Н-цепь].
Фиг.38 иллюстрирует структуру полипептида LH-типа и аминокислотные последовательности пептидных линкеров.
Фиг.39 иллюстрирует результаты Western блоттинга из Примера 6.4, показывающие, что экспрессируются модифицированное антитело sc(Fv)2, содержащее две V-области Н-цепи и две V-области L-цепи, и MABL2-scFv, содержащие пептидные линкеры различной длины.
Фиг.40а и 40b иллюстрируют результаты проточной цитометрии с использованием культурального супернатанта клеток COS7, полученного из Примера 6.3 (1), показывающие, что MABL2-scFv и sc(Fv)2, имеющие пептидные линкеры различной длины, имеют высокое сродство к IAP человека.
Фиг.41а и 41b иллюстрируют результаты апоптоз-индуцирующего действия из Примера 6.6, показывающие, что scFv<HL3, 4, 6, 7, LH3, 4, 6, и 7> и sc(Fv)2 в значительной степени индуцируют клеточную смерть у клеток hIAP/L1210.
Фиг.42 иллюстрирует результаты определения антигенсвязывающей способности из Примера 6.10, показывающие, что димер ScFv <HL5> и sc(Fv)2 имеют высокое сродство к IAP человека.
Фиг.43 иллюстрирует результаты апоптоз-индуцирующего действия in vitro из Примера 6.11, показывающие, что димер ScFv <HL5> и sc(Fv)2 индуцируют апоптоз клеток hIAP/L1210 и клеток CCRF-CEM зависящим от концентрации образом.
Фиг.44 иллюстрирует результаты количественных измерений М-белка, продуцируемого клеточной линией миеломы человека КРММ2 в сыворотке мыши с трансплантированными клетками миеломы человека. Видно, что димер scFv <HL5> и sc(Fv)2 в значительной степени тормозят рост клеток КРММ2.
Фиг.45 иллюстрирует продолжительность выживания (в днях) мышей после трансплантации опухоли, показывая, что продолжительность выживания в группе, которой был введен scFv <HL5>, в значительной степени увеличивается.
Фиг.46 иллюстрирует продолжительность выживания (в днях) мышей после трансплантации опухоли, показывая, что продолжительность выживания в группе, которой был введен sc(Fv)2, в значительной степени увеличивается.
Фиг.47 иллюстрирует схематически способ конструирования фрагмента ДНК, кодирующего реконструированный одноцепочечный Fv 12B5, содержащий линкерную последовательность, состоящую из 15 аминокислот, и его структуру.
Фиг.48 иллюстрирует результаты очистки гель-фильтрацией каждого из одноцепочечных Fv 12B5, полученных в Примере 7.5 (1), показывающие, что sc12B5 разделяется на два пика (фракции А и В).
Фиг.49 иллюстрирует результаты анализа каждой из фракций А и В методом SDS-PAGE, проведенного в Примере 7.5 (2).
Фиг.50a и 50b иллюстрирует результаты анализа каждой из фракций А и В на колонке Superdex200, проведенного в Примере 7.5 (2), показывающие, что основной пик фракции А элюируется с кажущейся молекулярной массой около 44 кДа (а) и что основной пик фракции В элюируется с кажущейся молекулярной массой около 22 кДа (b).
Фиг.51 иллюстрирует результаты измерения ТРО-подобной агонистической активности sc12B5 и антитела 12B5 (IgG, Fab), показывающие, что 12B5IgG и моновалентный одноцепочечный Fv (sc12B5) проявляют ТРО-подобную агонистическую активность, зависящую от концентрации.
Фиг.52 иллюстрирует результаты измерения ТРО-подобной агонистической активности мономера и димера sc12B5, показывающие, что одноцепочечный Fv (димер sc12B5), имеющий бивалентный антигенсвязывающий центр, имеет агонистическую активность в 400 раз выше, чем у моновалентного sc12B5, и что его эффективность равняется или превышает эффективность ТРО человека.
Фиг.53 иллюстрирует результаты очистки полученного одноцепочечного антитела sc12E10 методом гель-фильтрационной хроматографии с использованием колонки Superdex200HR, показывающие, что db12E10 разделяется на два пика (фракции А и В).
Фиг.54 иллюстрирует результаты очистки полученного одноцепочечного антитела db12E10 методом гель-фильтрационной хроматографии с использованием колонки Superdex200HR, показывающие, что db12E10 разделяется на два пика (фракции С и D).
Фиг.55 иллюстрирует анализ методом SDS-PAGE фракций А и В (sc12E10) и фракций С и D (db12E10) в восстанавливающих и невосстанавливающих условиях.
Фиг.56 иллюстрирует результаты анализа фракций А и В методом гель-фильтрационной хроматографии с использованием колонки Superdex200HR, показывающие, что основной пик фракции А элюируется с кажущейся молекулярной массой около 42 кДа и что основной пик фракции В элюируется с кажущейся молекулярной массой около 20 кДа.
Фиг.57 иллюстрирует результаты анализа фракций С и D методом гель-фильтрационной хроматографии с использованием колонки Superdex200HR, показывающие, что основной пик фракции С элюируется с кажущейся молекулярной массой около 69 кДа и что основной пик фракции Д элюируется с кажущейся молекулярной массой около 41 кДа.
Фиг.58 иллюстрирует агонистическую активность различных молекул антитела 12Е10 к MPL, показывающую, что одноцепочечные Fv (sc12E10, db12E10) проявляют ТРО-подобную агонистическую активность, в то время как 12Е10 IgG и 12Е10 Fab не проявляют.
Фиг.59 - график, иллюстрирующий агонистическую активность мономера и димера sc12E10 и димера и тримера db12E10 в отношении MPL, показывающий, что димер sc12E10 и димер и тример db12E10 проявляют более высокую ТРО-подобную агонистическую активность, чем ТРО.
Сведения, подтверждающие возможность осуществления изобретения
Нижеследующие примеры служат лишь для иллюстрации настоящего изобретения и их ни в коем случае не следует рассматривать для ограничения объема притязаний.
Для иллюстрации способа получения модифицированных антител по изобретению ниже приводятся примеры получения одноцепочечных Fv. В приводимых примерах получения таких модифицированных антител были использованы антитела мыши к IAP человека, MABL-1 MABL-2. Гибридомы MABL-1 и MABL-2, соответственно продуцирующие их, депонированы 11-го сентября 1997 г. на международном уровне в Агентстве индустриальной науки и технологии. Министром международной торговли и промышленности (National Institute of Bioscience and Human Technology, Agency of Industrial Science and Technology, Minister of International Trade and Industry (1-3 Higasi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan) как уполномоченном депозитарии микроорганизмов под инвентарными номерами FERM ВР-6100 и FERM BP-6101.
Примеры
Пример 1 (клонирование молекул ДНК, кодирующих V-области моноклональных антител мыши к IAP человека)
Молекулы ДНК, кодирующие вариабельные области моноклональных антител мыши к IAP человека, MABL-1 и MABL-2, клонируют следующим образом.
1.1 Получение матричной РНК(мРНК)
Получают мРНК из гибридов MABL-1 и MABL-2, используя набор для очистки мРНК (mRNA Purification Kit, Pharmacia Biotech).
1.2 Синтез двухцепочечной кДНК
Синтезируют двухцепочечную кДНК из приблизительно 1 мкг мРНК, используя набор Marathon cDNA Amplification Kit (CLONTECH) с присоединением адаптора.
1.3 ПЦР-амплификация генов, кодирующих вариабельные области антитела
ПЦР проводили с помощью термоциклера (Thermal Cycler, PERKLN ELMER).
(1) Амплификация гена, кодирующего V-область L-цепи MABL-1
Праймерами, используемыми в ПЦР-методе, были Adapter Primer-1 (CLONTECH), показанный в SEQ ID NO.1, который гибридизуется с частью последовательности адаптора, и праймер МКС (Mouse Kappa Constant, Bio/Technology, 9, 88-89, 1991)), показанный в SEQ ID NO.2, который гибридизуется V-областью L-цепи мыши.
В 50 мкл ПЦР-раствора содержится 5 мкл 10-кратного ПЦР-буфера II, 2 мМ MgCl2, 0,16 мМ dNTPs (dATP, dGTP, dCTP и dTTP), 2,5 единицы ДНК-полимеразы AmpliTaq Gold (PERKIN ELMER), 0,2 мкМ адапторного праймера SEQ ID NO.1, 0,2 мкМ праймера-MKC SEQ ID NO.2 и 0,1 мкг двухцепочечной кДНК, происходящей из MABL-1. Раствор предварительно нагревают при начальной температуре 94°С в течение 9 минут, а затем в указаном порядке нагревают при 94°С в течение 1 мин, при 60°С в течение 1 мин и при 72°С - 1 мин 20 сек. Такой температурный цикл повторяют 35 раз, а затем реакционную смесь нагревают дополнительно при 72°С в течение 10 минут.
(2) Амплификация кДНК, кодирующей V-область Н-цепи MABL-1
В качестве праймеров для ПЦР используют Adapter Primer-1, показанный в SEQ ID NO.1, и праймер МНС-γ1 (Mouse Heavy Constant (Bio/Technology, 9, 88-89, 1991)), показанный в SEQ ID NO.3.
Амплификацию кДНК осуществляют так же, как амплификацию гена V-области L-цепи, которая описана в Примере 1.3-(1), с той разницей, что используют 0,2 мкМ праймера MHC-γ1, вместо 0,2 мкМ праймера МКС.
(3) Амплификация ДНК, кодирующей V-область L-цепи MABL-2
В качестве праймеров для ПЦР используют Adapter Primer-1, показанный в SEQ ID NO.1, и праймер МКС, показанный в SEQ ID NO.2.
Амплификацию кДНК проводят так же, как амплификацию гена V-области L-цепи MABL-1, которая описана в Примере 1.3-(1), с той разницей, что используют 0,1 мкг двухцепочечной кДНК, происходящей из MABL-2, вместо 0,1 мкг двухцепочечной кДНК, происходящей из MABL-1.
(4) Амплификация ДНК, кодирующей V-область Н-цепи MABL-2
В качестве праймеров для ПЦР используют Adapter Primer-1, показанный в SEQ ID NO.1, и праймер МНС-γ2а (Bio/Technology, 9, 88-89, 1991)), показанный в SEQ ID NO.4.
Амплификацию кДНК проводят так же, как амплификацию гена V-области L-цепи, которая описана в Примере 1.3-(3), с той разницей, что используют 0,2 мкМ праймера МНС-γ2а вместо 0,2 мкМ праймера МКС.
1.4 Очистка продуктов ПЦР
Фрагмент ДНК, амплифицированный методом ПЦР, очищают при помощи набора QIAquick ПЦР Purification Kit (QIAGEN) и растворяют в 10 мМ Tris-HCl (pH 8,0), содержащем 1 мМ ЭДТА.
1.5 Лигирование и трансформация
Около 140 нг фрагмента ДНК, содержащего ген, кодирующий V-область L-цепи каппа-типа мыши, происходящую из MABL-1, полученного так, как было описано выше, лигируют с 50 нг вектора pGEM-T Easy (Promega) в реакционном буфере, содержащем 30 мМ Tris-HCl (pH 7,8), 10 мМ MgCl2, 10 мМ дитиотрейтола, 1 мМ АТР и 3 единицы ДНК-лигазы Т4 (Promega), при 15°С в течение 3 часов.
Затем 1 мкл реакционной смеси добавляют к 50 мкл компетентных клеток Е.coli DH5α (Toyobo Inc.) и на 30 мин помещают на лед, инкубируют в термостате при 42°С в течение 1 мин и вновь помещают на лед на 2 мин. Добавляют 100 мкл SOC-среды (GIBCO BRL). Клетки бактерии Е.coli рассевают на агаризованную среду LB (Molecular Cloning: A Laboratory Manual, Sambrook et al., Cold Spring Harbor Laboratory Press, 1989), содержащую 100 мкг/мл ампициллина (SIGMA), и культивируют при 37°С в течение ночи для получения трансформанта Е.coli.
Трансформант культивируют в 3 мл среды LB, содержащей 50 мкг/мл ампициллина, при 37°С в течение ночи и из культуры при помощи набора QIAprep Spin Miniprep Kit (QIAGEN) выделяют плазмидную ДНК.
Полученную плазмиду, содержащую ген, кодирующий V-область L-цепи каппа-типа мыши, происходящую из гибридомы MABL-1, обозначают как pGEM-M1L.
Точно таким же образом, как это описано выше, из очищенного фрагмента ДНК получают плазмиду, содержащую ген, кодирующий V-область Н-цепи мыши, происходящую из гибридомы MABL-1, и обозначают как pGEM-M1H.
Плазмиду, содержащую ген, кодирующий V-область L-цепи каппа-типа мыши, происходящую из гибридомы MABL-2, получают из очищенного фрагмента ДНК и обозначают как pGEM-M2L.
Плазмиду, содержащую ген, кодирующий V-область Н-цепи мыши, происходящую из гибридомы MABL-2, получают из очищенного фрагмента ДНК и обозначают как pGEM-M2H.
Пример 2 (секвенирование ДНК)
Определяют нуклеотидную последовательность кДНК-кодирующей области вышеуказанных плазмид при помощи секвенатора Auto DNA Sequencer (Applied Biosystem) и набора ABI PRISM Dye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystem) в соответствии с протоколом производителя.
Нуклеотидная последовательность гена, кодирующего V-область L-цепи мышиного антитела MABL-1, включенная в плазмиду pGEM-M1L, приведена в SEQ ID NO.5.
Нуклеотидная последовательность гена, кодирующего V-область Н-цепи антитела MABL-1 мыши, входящая в плазмиду pGEM-M1H, приведена в SEQ ID NO.6.
Нуклеотидная последовательность гена, кодирующего V-область L-цепи антитела MABL-2 мыши, входящая в плазмиду pGEM-M2L, приведена в SEQ ID NO.7.
Нуклеотидная последовательность гена, кодирующего V-область Н-цепи антитела MABL-2 мыши, входящая в плазмиду pGEM-M2H, приведена в SEQ ID NO.8.
Пример 3 (определение CDR)
V-области L- и Н-цепей бывают обычно сходными по своей структуре, и в них каждые четыре каркасные области соединены тремя гипервариабельными областями, т.е. участками, определяющим комплементарность (CDR). Аминокислотная последовательность каркаса относительно консервативна, в то время как аминокислотная последовательность CDR отличается чрезвычайно большим разнообразием (Kabat E.A. et al., "Sequences of Proteins of Immunological Interest", US Dept. Health and Human Services, 1983).
С учетом этих фактов аминокислотные последовательности вариабельных областей мышных моноклональных антител к IAP человека были сопоставлены с базой данных по аминокислотным последовательностям антител, созданной Kabat et al., для выявления гомологии. Области CDR были определены на основе гомологии, как показано в Таблице 1.
Пример 4 (выявление экспрессии клонированной кДНК)
(Получение химерного антитела MABL-1 и химерного антитела MABL-2)
4.1 Получение векторов, экспрессирующих химерное антитело MABL-1
кДНК-клоны, pGEM-M1L и pGEM-M1H, кодирующие соответственно V-области L-цепи и Н-цепи антитела MABL-1 мыши, модифицируют методом ПЦР и вводят в экспрессионый вектор HEF (WO 92/19759) для получения векторов, экспрессирующих химерное антитело MABL-1.
Прямой праймер MLS (SEQ ID NO.9) для V-области L-цепи и прямой праймер MHS (SEQ ID NO.10) для V-области Н-цепи конструируют так, чтобы они гибридизовались с ДНК, кодирующей начало лидерной последовательности из каждой V-областей, и содержали консенсусную последовательность Козака (J. Mol. Biol., 196, 947-950, 1987)) и сайта для рестриктазы HindIII. Обратный праймер MLAS (SEQ ID NO.11) для V-области L-цепи и обратный праймер MHAS (SEQ ID NO.12) для V-области Н-цепи конструируют так, чтобы они гибридизовались с ДНК, кодирующей концевой участок J-области, и содержали донорную последовательность сплайсинга и сайт для рестриктазы BamHI.
100 мкл раствора, содержащего 10 мкл 10-кратного ПЦР-буфера II, 2 мМ MgCl2, 0,16 мМ dNTPs (dATP, dGTP, dCTP и dTTP), 5 единиц ДНК-полимеразы AmpliTaq Gold, по 0,4 мкМ каждого из праймеров и 8 нг ДНК-матрицы (pGEM-M1L или pGEM-M1H), нагревают предварительно при начальной температуре 94°С в течение 9 мин, а затем проводят нагревание в следующей последовательности: при 94°С в течение 1 мин, при 60°С 1 мин и при 72°С 1 мин 20 сек. Такой температурный цикл повторяют 35 раз, а затем реакционную смесь нагревают дополнительно при 72°С в течение 10 мин.
Продукт ПЦР очищают с помощью набора QIAquick ПЦР Purification Kit (QIAGEN), а затем расщепляют ферментами HindIII и BamHI. Продукт V-области L-цепи клонируют в экспрессионном векторе HEF, HEF-κ, а продукт V-области Н-цепи клонируют в экспрессионном векторе HEF, HEF-γ. После секвенирования ДНК плазмиды, содержащие ДНК-фрагменты с правильной ДНК-последовательностью, обозначают соответственно как HEF-M1L и HEF-M1H.
4.2 Получение векторов, экспрессирующих химерные антитела MABL-2
Модификацию и клонирование кДНК проводят так, как описано в Примере 4.1, с тем исключением, что используют pGEM-M2L и pGEM-M2H в качестве матричной ДНК вместо pGEM-M1L и pGEM-M1H. После секвенирования ДНК плазмиды, содержащие ДНК-фрагменты с правильной ДНК-последовательностью, обозначают соответственно как HEF-M2L и HEF-M2H.
4.3 Трансфекция в клетки COS7
Вышеупомянутые векторы экспрессии проверяют на клетках COS7 для наблюдения за временной экспрессией химерных антител MABL-1 и MABL-2.
(1) Трансфекция генами химерного антитела MABL-1
Клетки COS7 котрансформируют векторами HEF-M1L и HEF-M1H методом электропорации при помощи устройства Gene Pulser (BioRad). В кювету вносят по 10 мкг каждой ДНК и 0,8 мл PBS с 1×107 клеток/мл. Смесь обрабатывают импульсом при 1,5 кВ, при электрической емкости 25 мкФ.
После 10-минутного восстановления при комнатной температуре электропорированные клетки переносят в среду для культивирования DMEM (GIBCO BRL), содержащую 10%-ную фетальную бычью сыворотку, свободную от γ-глобулина. После 72-часового культивирования супернатант собирают, центрифугируют для удаления клеточных фрагментов и отбирают.
(2) Трансфекция генами, кодирующими химерное антитело MABL-2
Котрансфекцию клеток COS7 генами, кодирующими химерное антитело MABL-2, выполняют так же, как в Примере 4.3-(1), с тем исключением, что используют векторы HEF-M2L и HEF-M2H вместо векторов HEF-M1L и HEF-M1H. Точно таким же образом получают супернатант.
4.4 Проточная цитометрия
Для измерения связывания с антигеном проводят проточную цитометрию, используя вышеуказанный культуральный супернатант клеток COS7. Культуральный супернатант клеток COS7, экспрессирующих химерное антитело MABL-1, или клетки COS7, экспрессирующие химерное антитело MABL-2, или антитело человека IgG (SIGMA) в качестве контроля добавляют к 4×105 клеток клеточной линии лейкемии мыши L1210, экспрессирующей IAP человека, и инкубируют на льду. После отмывки туда добавляют меченное ФИТЦ антитело против IgG человека (Cappel). После инкубации и отмывания измеряют интенсивность флуоресценции при помощи аппарата FACScan (BECTON DICKINSON).
Поскольку химерные антитела MABL-1 и MABL-2 специфически связываются с клетками L1210, экспрессирующими человеческий IAP, то это подтверждает, что такие химерные антитела имеют характерную структуру V-областей моноклональных антител мыши MABL-1 и MABL-2 соответственно (Фиг.1-3).
Пример 5 (получение реконструированного одноцепочечного Fv (scFv) антитела MABL-1
и антитела MABL-2)
5.1 Получение реконструированного одноцепочечного Fv антитела MABL-1
Реконструированный одноцепочечный Fv антитела MABL-1 получают следующим образом. Методом ПЦР амплифицируют соответственно V-область Н-цепи и V-область L-цепи антитела MABL-1 и линкер, и состыковывают для получения реконструированного одноцепочечного Fv антитела MABL-1. Способ получения показан на Фиг.4. Для получения одноцепочечного Fv антитела MABL-1 используют шесть праймеров (A-F). Праймеры А, С и Е имеют смысловую последовательность, а праймеры D и F имеют антисмысловую последовательность.
Прямой праймер VHS для V-области Н-цепи (Праймер A, SEQ ID NO.13) конструируют так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области Н-цепи, содержа сайт узнавания рестриктазы Ncol. Обратный праймер VHAS для V-области Н-цепи (Праймер D, SEQ ID NO.14) конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области Н-цепи и для перекрывания линкером.
Прямой праймер LS для линкера (Праймер С, SEQ ID NO.15) конструируют так, чтобы он гибридизовался с ДНК, кодирующей N-конец линкера, и перекрывался с ДНК, кодирующей С-конец V-области Н-цепи. Прямой праймер LAS для линкера (Праймер D, SEQ ID NO.16) конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец линкера и перекрывался с ДНК, кодирующей N-конец V-области L-цепи.
Прямой праймер VLS для V-области L-цепи (Праймер Е, SEQ ID NO.17) конструировали так, чтобы он гибридизовался с ДНК, кодирующей С-конец линкера, и перекрывался с ДНК, кодирующей N-конец V-области L-цепи. Обратный праймер VLAS-FLAG для V-области L-цепи (Праймер F, SEQ ID NO.18) конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал последовательность, кодирующую FLAG-пептид (Норр Т.Р. et al., Bio/Technology, 6, 1204-1210, 1988), два стоп-кодона и сайт узнавания рестриктазы EcoRI.
На первом этапе ПЦР проводят три реакции, А-В, C-D и E-F, и очищают продукты ПЦР. Три ПЦР-продукта, полученных на первом этапе ПЦР, состыковывают в соответствии с их комплементарностью. Затем добавляют праймеры А и F и амплифицируют полноразмерную ДНК, кодирующую реконструированный одноцепочечный Fv антитела MABL-1 (вторая ПЦР). В первой ПЦР в качестве матрицы используют соответственно плазмиду pGEM-M1H, кодирующую V-область Н-цепи антитела MABL-1 (см. Пример 2), плазмиду pSC-DPI, которая содержит ДНК-последовательность, кодирующую область линкера, содержащую Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser (SEQ ID NO.19) (Huston J.S. et al., Proc. Natl. Acad. Sci. USA, 85, 5879-5883, 1988), и плазмиду pGEM-M1L, кодирующую V-область L-цепи антитела MABL-1 (см. Пример 2).
50 мкл раствора для первого этапа ПЦР содержит 5 мкл 10-кратного ПЦР-буфера II, 2 мМ MgCl2, 0,16 мМ каждого из dNTP, 2,5 единицы ДНК-полимеразы AmpliTaq Gold (PERKIN ELMER), 0,4 мкМ каждого праймера и 5 нг каждой матричной ДНК. Раствор для ПЦР предварительно нагревают при начальной температуре 94°С в течение 9 минут, а затем последовательно нагревают при 94°С в течение 1 мин, при 65°С в течение 1 мин и при 72°С 1 мин 20 сек. Такой температурный цикл повторяют 35 раз, а затем реакционную смесь нагревают дополнительно при 72°С в течение 7 минут.
ПЦР-продукты А-В (371 п.о.), C-D (63 п.о.), E-F (384 п.о.) очищают с помощью набора QIAquick ПЦР Purification Kit (QIAGEN) и состыковывают в ходе второй ПЦР. Во второй ПЦР 98 мкл ПЦР-раствора, содержащего 120 нг продукта А-В первой ПЦР, 20 нг ПЦР-продукта C-D и 120 нг ПЦР-продукта E-F, 10 мкл 10-кратного ПЦР-буфера II, 2 мМ MgCl2, 0,16 мМ каждого из dNTP, 5 единиц ДНК-полимеразы AmpliTaq Gold (PERKIN ELMER), предварительно нагревают при начальной температуре 94°С в течение 8 минут, а затем нагревают последовательно при 94°С 2 мин, при 65°С 2 мин и при 72°С 2 мин. Такой температурный цикл повторяют дважды, а затем вводят в реакцию по 0,4 мкМ соответственно каждого из праймеров А и F. Смесь предварительно нагревают при начальной температуре 94°С в течение 1 мин, а затем последовательно нагревают при 94°С 1 мин, при 65°С 1 мин и при 72°С 1 мин 20 сек. Такой температурный цикл повторяют 35 раз, а затем реакционную смесь нагревают дополнительно при 72°С в течение 7 минут.
ДНК-фрагмент размером 843 п.о., полученный во второй ПЦР, очищают и расщепляют при помощи NcoI и EcoRI. Клонируют полученный ДНК-фрагмент в вектор pSCFVT7. Экспрессионый вектор pSCFVT7 содержит pelB-сигнальную последовательность, подходящую для периплазматической системы экспрессии Е.coli (Lei S.P. et al., J. Bacteriology, 169, 4379-4383, 1987). После секвенирования ДНК плазмиду, содержащую фрагмент ДНК, кодирующий правильную аминокислотную последовательность реконструированного одноцепочечного Fv антитела MABL-1, обозначают как "pscM1" (см. Фиг.5). Нуклеотидная последовательность и аминокислотная последовательность реконструированного одноцепочечного Fv антитела MABL-1, содержащиеся в плазмиде pscM1, приведены в SEQ ID NO.20.
pscM1-вектор модифицируют методом ПЦР для получения вектора, экспрессирующего реконструированный одноцепочечный Fv антитела MABL-1 в клетках млекопитающих. Полученный ДНК-фрагмент встраивают в экспрессионный вектор pCHO1. Такой экспрессионный вектор, pCHO1, был сконструирован путем расщепления DHFR-ΔE-rvH-PM1-f (WO 92/19759) с использованием EcoRI и SmaI для элиминации гена антитела и присоединения к нему адаптера EcoRI-NotI-BamHI (Takara Shuzo).
В качестве прямого праймера для ПЦР конструируют праймер Sal-VHS, показанный в SEQ ID NO.21 так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области Н-цепи, и содержал сайт узнавания рестриктазы SalI. В качестве обратного праймера для ПЦР конструируют праймер FRH1-anti, показанный в SEQ ID NO.22 так, чтобы он гибридизовался с ДНК, кодирующей конец первой каркасной последовательности.
100 мкл ПЦР-раствора, содержащего 10 мкл 10-кратного ПЦР-буфера II, 2 мМ MgCl2, 0,16 мМ каждого из dNTP, 5 единиц ДНК-полимеразы AmpliTaq Gold, no 0,4 мкМ каждого из праймеров и 8 нг матричной ДНК (pscM1), предварительно нагревают при начальной температуре 95°С в течение 9 мин, а затем нагревают последовательно при 95°С в течение 1 мин, при 60°С 1 мин и при 72°С 1 мин 20 сек. Такой температурный цикл повторяют 35 раз, а затем реакционную смесь дополнительно нагревают при 72°С в течение 7 мин.
ПЦР-продукт очищают при помощи набора QIAquick ПЦР Purification Kit (QIAGEN), а затем расщепляют с помощью SalI и MboII для получения ДНК-фрагмента, кодирущего N-конец реконструированного одноцепочечного Fv антитела MABL-1. Вектор pscM1 расщепляют MboII и EcoRI для получения ДНК-фрагмента, кодирущего С-конец реконструированного одноцепочечного Fv антитела MABL-1. ДНК-фрагмент SalI-MboII и ДНК-фрагмент MboII-EcoRI клонируют в вектор pCHO1-Igs. После секвенирования ДНК плазмиду, содержащую требуемую ДНК-последовательность, обозначают, как "рСНОМ1" (см. Фиг.6). Экспрессионный вектор, pCHO1-Igs, содержит сигнальную последовательность IgG1 мыши, подходящую для системы секреции-экспрессии в клетках млекопитающего (NATURE, 322, 323-327, 1988). Нуклеотидная последовательность и аминокислотная последовательность реконструированного одноцепочечного Fv антитела MABL-1, содержащиеся в плазмиде pCHOM1, показаны в SEQ ID NO.23.
5.2 Получение реконструированного одноцепочечного Fv антитела MABL-2
Реконструированное одноцепочечное Fv антитела MABL-2 получают согласно вышеприведенному Примеру 5.1. Для получения плазмиды pscM2, которая содержит ДНК-фрагмент, кодирующий требуемую аминокислотную последовательность реконструированного одноцепочечного Fv антитела MABL-2, на первом этапе ПЦР используют плазмиду pGEM-M2H, кодирующую V-область Н-цепи MABL-2 (см. Пример 2), вместо pGEM-M1H и плазмиду pGEM-M2L, кодирующую V-область L-цепи MABL-2 (см. Пример 2), вместо pGEM-M1L. Нуклеотидная последовательность и аминокислотная последовательность реконструированного одноцепочечного Fv антитела MABL-2, содержащиеся в плазмиде pscM2, приведены в SEQ ID NO.24.
Вектор pscM2 модифицируют методом ПЦР для получения вектора, рСНОМ2, для экспрессии в клетках млекопитающих, который содержит ДНК-фрагмент, кодирующий правильную аминокислотную последовательность реконструированного одноцепочечного Fv антитела MABL-2. Нуклеотидная последовательность и аминокислотная последовательность реконструированного одноцепочечного Fv антитела MABL-2, содержащиеся в плазмиде рСНОМ2, показаны в SEQ ID NO.25.
5.3 Трансфекция в клетки COS7
Тестирование вектора рСНОМ2 на клетках COS7 осуществляют для того, чтобы наблюдать временную экспрессию реконструированного одноцепочечного Fv антитела MABL-2.
Клетки COS7 трансформируют вектором рСНОМ2 методом электропорации при помощи аппарата Gene Pulser (BioRad). Добавляют в кювету ДНК (10 мкг) и 0,8 мл PBS с 1×107 клеток/мл. Смесь обрабатывают импульсом при 1,5 кВ, при электрической емкости 25 мкФ.
После восстановления в течение 10 мин при комнатной температуре электропорированные клетки переносят в культуральную среду IMDM (GIBCO BRL), содержащую 10%-ную фетальную бычью сыворотку. После культивирования в течение 72 часов собирают супернатант, центрифугируют для удаления клеточных фрагментов и отбирают.
5.4 Обнаружение реконструированного одноцепочечного Fv антитела MABL-2 в культуральном супернатанте клеток COS7
Присутствие реконструированного одноцепочечного Fv антитела MABL-2 в культуральном супернатанте клеток COS7, трансфицированых вектором рСНОМ2, подтверждали методом Western-блоттинга.
Культуральный супернатант клеток COS7, трансфицированных вектором рСНОМ2, и культуральный супернатант клеток COS7, трансфицированных вектором pCHOM1, в качестве контроля подвергают электрофорезу в присутствии додецилсульфата натрия и переносят на мембрану REINFORCED NC (Schleicher & Schuell). Мембрану блокируют 5%-ным снятым молоком (Morinaga Nyu-gyo), промывают раствором 0,05% Tween 20-PBS и смешивают с антителом против FLAG (SIGMA). Мембрану инкубируют при комнатной температуре, отмывают и смешивают с антителом IgG мыши, конъюгированным с щелочной фосфатазой (Zymed). После инкубирования и отмывания при комнатной температуре добавляют раствор субстрата (Kirkegaard Perry Laboratories) для развития окраски (Фиг.7).
FLAG-пептид-специфичный белок обнаруживается только в культуральном супернатанте клеток COS7 с введенным вектором рСНОМ2, и это подтверждает, что реконструированный одноцепочечный Fv антитела MABL-2 секретировался в данном культуральном супернатанте.
5.5 Проточная цитометрия
Для измерения связывания с антигеном применяют метод проточной цитометрии, используя вышеуказанный культуральный супернатант клеток COS7. Культуральный супернатант клеток COS7, экспрессирующих реконструированный одноцепочечный Fv антитела MABL-2, или культуральный супернатант клеток COS7, трансформированных вектором pCHOM1, в качестве контроля, добавляют к 2×105 клеток клеточной линии лейкемии мыши L1210, экспрессирующих интегринассоциированный белок (IAP) человека, или клеточной линии L1210, трансформированной pCOS1, в качестве контроля. После инкубирования на льду и отмывания добавляют антитело против FLAG (SIGMA). Затем клетки инкубируют и отмывают. Затем туда добавляют меченное ФИТЦ антитело против IgG мыши (BECTON DICKINSON) и клетки еще раз инкубируют и отмывают. Потом измеряют интенсивность флуоресценции с помощью аппарата FACScan (BECTON DICKINSON).
Подтверждается, что поскольку одноцепочечный Fv антитела MABL-2 специфически связывается с клетками L1210, экспрессирующими IAP человека, то реконструированный одноцепочечный Fv антитела MABL-2 обладает сродством к интегринассоциированному белку человека (IAP) (Фиг.8-11).
5.6 Конкурентный ферментный иммуносорбентный анализ (ELISA)
Связывающую активность реконструированного одноцепочечного Fv антитела MABL-2 определяли по ингибирующей активности в отношении связывания моноклональных антител мыши с антигеном.
В каждую лунку 96-луночного планшета добавляют антитело против FLAG в концентрации 1 мкг/мл и инкубируют при 37°С в течение 2 час. После отмывания проводят блокирование с использованием 1% БСА-PBS. После инкубирования и отмывания при комнатной температуре культуральный супернатант клеток COS7, в который был введен ген человеческого IAP-антигена секреторного типа (SEQ ID NO.26), разводят PBS, до двукратного объема и вносят в каждую лунку. После инкубирования и отмывания при комнатной температуре добавляют в каждую лунку смесь из 50 мкл биотинилированного антитела MABL-2 в концентрации 100 нг/мл и 50 мкл последовательно разведенного супернатанта клеток COS7, экспрессирующих реконструированный одноцепочечный Fv антитела MABL-2. После инкубирования и отмывания при комнатной температуре добавляют в каждую лунку стрептоавидин, конъюгированный со щелочной фосфатазой (Zymed). После инкубирования и отмывания при комнатной температуре добавляют раствор субстрата (SIGMA) и при 405 нм измеряют оптическую плотность реакционной смеси в каждой лунке.
Как показывают результаты, реконструированный одноцепочечный Fv антитела MABL-2 (MABL2-scFv), по сравнению с взятым в качестве контроля культуральным супернатантом клеток COS7 с введеной РСНO1, несомненно ингибирует связывание антитела MABL-2 мыши с IAP-антигеном человека (Фиг.12), зависящим от концентрации образом. Следовательно, можно полагать, что реконструированный одноцепочечный Fv антитела MABL-2 содержит правильную структуру каждой из V-областей моноклонального антитела MABL-2 мыши.
5.7 Эффект индукции апоптоза in vitro
Апоптоз-индуцирующее действие реконструированного одноцепочечного Fv антитела MABL-2 исследуют путем окрашивания Аннексином-V (Boehringer Mannheim) с использованием клеток L1210, трансфицированных геном IAP человека, клеток L1210, трансфицированных вектором pCOS1 в качестве контроля, и клеток CCRF-CEM.
К каждому типу вышеуказанных клеток, взятых в количестве 1×105 клеток, добавляют культуральный супернатант клеток COS7, экспрессирующих реконструированный одноцепочечный Fv антитела MABL-2, или культуральный супернатант клеток COS7, трансфицированных вектором рСНО1 в качестве контроля, в конечной концентрации 50%, и смеси культивируют в течение 24 час. Затем осуществляют окрашивание Аннексином-V и измеряют интенсивность флуоресценции с помощью аппарата FACScan (BECTON DICKINSON).
Результаты окрашивания Аннексином-V показаны соответственно на Фиг.13-18. Точки в левой нижней части представляют живые клетки, точки в правой нижней части представляют клетки на ранней стадии апоптоза, точки в правой верхней части представляют клетки на поздней стадии апоптоза. Данные результаты показывают, что по сравнению с контролем реконструированный одноцепочечный Fv антитела MABL-2 (MABL2-scFv) весьма заметно индуцирует клеточную смерть клеток L1210, специфичных в отношении IAP-антигена человека (Фиг.13-16), и что реконструированный одноцепочечный Fv антитела MABL-2 также весьма заметно индуцирует клеточную смерть клеток CCRF-CEM (Фиг.17-18).
5.8 Экспрессия происходящего из MABL-2 одноцепочечного Fv в клетках СНО
Клетки СНО трансфицируют вектором рСРОМ2 для получения линии клеток СНО, которая постоянно экспрессирует одноцепочечный Fv (полипептид), происходящий из антитела MABL-2.
Клетки СНО трансформируют вектором рСРОМ2 методом электропорации при помощи прибора Gene Pulser (BioRad). В кювету вносят смесь ДНК (10 мкг) и 0,7 мл PBS с клетками СНО (1×107 клетки/мл). Смесь подвергают воздействию импульса при 1,5 кВ, при электрической емкости 25 мкФ. После восстановления при комнатной температуре в течение 10 мин электропорированные клетки переносят в α-МЕМ среду, свободную от нуклеиновой кислоты (GIBCO BRL), содержащую 10% фетальную бычью сыворотку, и культивируют. Экспрессию требуемого белка в полученных клонах подтверждают методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE) и отбирают клон с высоким уровнем экспрессии в качестве клеточной линии, продуцирующей одноцепочечный Fv, происходящей из MABL-2 антитела. Клеточную линию культивируют в бессывороточной среде CHO-S-SFM II (GIBCO BRL), содержащей 10 нМ метотрексат (SIGMA). Затем культуральнный супернатант собирают, центрифугируют для удаления фрагментов клеток и отбирают.
5.9 Очистка происходящего из MABL-2 одноцепочечного Fv, продуцируемого в клетках СНО
Культуральный супернатант клеточной линии СНО, экспрессирующей одноцепочечный Fv, полученный в Примере 5.8, концентрируют в двадцать раз, применяя для этой цели картридж для искусственного диализа (PAN130SF, ASAHI MEDICALS). Концентрированный раствор хранят при - 20°С и размораживают перед очисткой.
Очищают одноцепочечный Fv из культурального супернатанта клеток СНО при помощи трех вариантов хроматографии: на Blue-Sepharose, на гидроксиапатите и гель-фильтрацией.
(1) Колоночная хроматография на Blue-Sepharose
Концентрированный супернатант разводят в 10 раз 20 мМ ацетатным буфером (рН 6,0) и центрифугируют для удаления нерастворимых веществ (10000 об/мин, 30 мин). Супернатант наносят на колонку с Blue-Sepharose (20 мл), уравновешенную тем же буфером. После промывания колонки тем же буфером белки, адсорбированные на колонке, элюируют, используя ступенчатый градиент NaCl в том же буфере: 0,1, 0,2, 0,3, 0,5 и вплоть до 1,0 М. Фракцию, выходящую с колонки в свободном объеме, и каждую из элюированных фракций анализируют с помощью SDS-PAGE. Фракции, наличие в которых одноцепочечного Fv подтверждается (фракции, элюированные при концентрации NaCl от 0,1 до 0,3 М), объединяют и, используя CentriPrep-10 (AMICON), концентрируют приблизительно в 20 раз.
(2) Гидроксиапатит
Концентрированный раствор, полученный на этапе (1), разводят в 10 раз 10 мМ фосфатным буфером (рН 7,0) и наносят на колонку с гидроксиапатитом (20 мл, BIORAD). Колонку промывают 60 мл 10 мМ фосфатного буфера (рН 7,0). Затем белки, адсорбированные на колонке, элюируют линейным градиентом натрий-фосфатного буфера до концентрации 200 мМ (см. Фиг.19). Анализ фракций с помощью SDS-PAGE подтверждает наличие одноцепочечного Fv во фракции А и во фракции В.
(2) Гель-фильтрация
Каждую из фракций А и В с этапа (2) концентрируют отдельно с помощью CentriPrep-10 и наносят на колонку с TSKgel G3000SWG (21,5×600 мм), уравновешенную 20 мМ ацетатным буфером (рН 6,0), содержащим 0,15 М NaCl. Хроматограммы показаны на Фиг.20. Анализ фракций с помощью SDS-PAGE подтвердил, что оба главные пика (AI и BI) соответствуют желаемому одноцепочечному Fv. При анализе путем гель-фильтрации фракция А элюируется с кажущейся молекулярной массой 36 кДа, а фракция В элюируется с массой 76 кДа. Анализ очищенных одноцепочечных Fv (AI, BI) проводят в 15% полиакриламидном геле с SDS. Пробы обрабатываются в отсутствие или в присутствии восстанавливающего агента, электрофорез проводят по методу Лэммли (Laemmli). Затем окрашивают белок кумасси бриллиантовым голубым. Как показано на Фиг.21, и AI и BI дают единственную окрашенную полосу с кажущейся молекулярной массой 35 кДа, вне зависимости от отсутствия или присутствия восстанавливающего агента. На основании вышесказанного можно сделать вывод, что AI является мономером одноцепочечного Fv, a BI - связанным нековалентно димером одноцепочечного Fv. Гель-фильтрационный анализ фракций AI и BI на колонке TSKgel G3000SWG (7,5×60 мм) показал, что пик мономера обнаруживается только во фракции AI, а пик димера обнаруживается только во фракции BI (Фиг.22). Фракция димера (фракция BI) соответствует 4% общего одноцепочечного Fv. Более 90% димера во фракции димера стабильно сохраняется более месяца при 4°С.
5.10 Конструирование вектора, экспрессирующего одноцепочечный Fv, происходящий из антитела MABL-2, в клетке Е.coli
Для получения вектора, эффективно экспрессирующего одноцепочечный Fv, происходящий из антитела MABL-2, в клетках Е.coli., вектор pscM2 модифицируют методом ПЦР. Получаемый фрагмент ДНК встраивают в экспрессионный вектор pSCFVT7.
В качестве прямого праймера для ПЦР праймер Nde-VHSm02, показанный в SEQ ID NO.27, конструируют так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области Н-цепи и содержал инициирующий кодон и сайт узнавания рестриктазы NdeI. В качестве обратного праймера для ПЦР праймер VLAS, показанный в SEQ ID NO.28, конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал два инициирующих стоп-кодона и сайт узнавания рестриктазы EcoRI. Прямой праймер, Nde-VHSm02, содержит пять точечных мутаций в той части, которая гибридизуется с ДНК, кодирующей N-конец V-области Н-цепи, для эффективной экспрессии в Е.coli.
100 мкл ПЦР-раствора, содержащего 10 мкл 10-кратного ПЦР-буфера №1, 1 мМ MgCl2, 0,2 мМ каждого из dNTP, 5 единиц ДНК-полимеразы KOD (все от TOYOBO), по 1 мкМ каждого праймера и 100 нг матричной ДНК (pscM2), нагревают в следующей последовательности: при 98°С в течение 15 сек, при 65°С 2 сек и при 74°С 30 сек. Такой температурный цикл повторяют 25 раз.
ПЦР-продукт очищают с помощью набора QIAquick ПЦР Purification Kit (QIAGEN) и расщепляют ферментами NdeI и EcoRI, а затем полученный фрагмент ДНК клонируют в вектор pSCFVT7, из которого была элиминирована сигнальная последовательность ре1В при расщеплении с помощью NdeI и EcoRI. После секвенирования ДНК полученную плазмиду, содержащую ДНК-фрагмент с требуемой ДНК-последовательностью, обозначают как "pscM2DEm02"(см. Фиг.23). Нуклеотидная последовательность и аминокислотная последовательность одноцепочечного Fv, происходящего из MABL-2, содержащиеся в плазмиде pscM2DEm02, показаны в SEQ ID NO.29.
5.11 Экспрессия одноцепочечного Fv, происходящего из антитела MABL-2, в клетках Е.coli
Для получения штамма Е.coli, экспрессирующего одноцепочечный Fv, происходящий из MABL-2, Е.coli BL21 (DE3)pLysS (STRATAGENE) трансформируют вектором pscM2DEm02. С помощью SDS-PAGE полученные клоны анализируют на экспрессию требуемого белка, и в качестве штамма, продуцирующего одноцепочечный Fv, происходящий из MABL-2, отбирают клон с высоким уровнем экспрессии.
5.12 Очистка одноцепочечного Fv, происходящего из антитела MABL-2. продуцируемого в Е.coli
Единичную колонию Е.coli., полученную путем трансформации, культивируют в 3 мл среды LB при 28°С в течение 7 час, а затем в 70 мл среды LB при 28°С в течение ночи. Такую предкультуру переносят в 7 л среды LB и культивируют в ферментере (Jar-fermenter) при 28°С при перемешивании при 300 об/мин. Когда оптическая плотность среды достигает значения O.D.=1,5, бактерии индуцируют с помощью 1 мМ IPTG и затем культивируют в течение 3 час.
Культуральную среду центрифугируют (10000×g, 10 мин) и собирают осажденные бактерии. К бактериям добавляют 50 мМ Tris-HCl буфер (рН 8,0), содержащий 5 мМ ЭДТА, 0,1 М NaCl и 1%-ный Triton X-100, и бактерии разрушают ультразвуковой обработкой (выход: 4, рабочий цикл: 70%, 1 мин×10 раз). Суспензию разрушенных бактерий центрифугируют (12000×g, 10 мин) для осаждения телец включения. Полученные тельца включения смешивают с 50 мМ Tris-HCl буфером (рН 8,0), содержащим 5 мМ ЭДТА, 0,1 М NaCl и 4%-ный Triton Х-100, и еще раз подвергают ультразвуковой обработке (выход: 4, рабочий цикл: 50%, 30 сек×2 раза) и центрифугируют (12000×g, 10 мин) для того, чтобы отделить требуемый белок в виде осадка и чтобы удалить примесные белки, находящиеся в супернатанте.
Тельца включения, содержащие желаемый белок, лизируют в 50 мМ буфере Tris-HCl (рН 8,0), содержащем 6 М мочевину, 5 мМ ЭДТА и 0,1 М NaCl, наносят на гель-фильтрационную колонку Sephacryl S-300 (5×90 см, Amersharm Pharmacia), уравновешенную 50 мМ Tris-HCl буфером (рН 8,0), содержащим 4 М мочевину, 5 мМ ЭДТА, 0,1 М NaCl и 10 мМ меркаптоэтанол, при скорости подачи 5 мл/мин, для удаления ассоциированных одноцепочечных Fv с большой молекулярной массой. Полученные фракции анализируют с помощью SDS-PAGE; фракции высокой степени гомогенности белка разводят буфером, используемым при гель-фильтрации, до O.D.280=0,25. Затем для того, чтобы осуществить повторный фолдинг белка, фракции трижды диализуют против 50 мМ буфера Tris-HCl (рН 8,0), содержащего 5 мМ ЭДТА, 0,1 М NaCl, 0,5 М Arg, 2 мМ глутатион в восстановленной форме и 0,2 мМ глутатион в окисленной форме. Фракции дополнительно трижды диализуют против 20 мМ ацетатного буфера (рН 6,0), содержащего 0,15 NaCl, для того чтобы сменить буфер.
Для удаления незначительного количества белка с высокой молекулярной массой, который был поперечно сшит межмолекулярными S-S-связями, диализованный продукт наносят на гель-фильтрационную колонку Superdex 200 pg (2,6×60 см, Amersharm Pharmacia), уравновешенную 20 мМ ацетатным буфером (рН 0,6), содержащим 0, 15 М NaCl. Как показано на Фиг.24, два пика, главный и минорный, были элюированы после широких пиков, которые, как ожидалось, относятся к агрегату с высокой молекулярной массой. Анализ методом SDS-PAGE (см. Фиг.21) и позиции элюции двух пиков при гель-фильтрационном анализе дают основания полагать, что главный пик принадлежит мономеру одноцепочечного Fv, а минорный - нековалентно связанному димеру одноцепочечного Fv. 4% от всех одноцепочечных Fv составляют нековалентно связанные димеры.
5.13 Апоптоз-индуцирующая активность in vitro одноцепочечного Fv, происходящего из антитела MABL-2
Апоптоз-индуцирующее действие in vitro одноцепочечного Fv, происходящего из антитела MABL-2 (MABL2-scFv), продуцируемого клетками СНО и Е.coli., исследуют в соответствии с двумя протоколами, при помощи метода окрашивания Аннексином-V (Boehringer Mannheim), используя клетки L1210 (hIAP/L1210), в которые был введен ген LAP человека.
Согласно первому протоколу образец антител в конечной концентрации 3 мкг/мл добавляют к 5×104 клеток клеточной линии hIAP/L1210 и культивируют в течение 24 час. Анализируют образец антител, т.е. мономер и димер одноцепочечного Fv MABL-2 из клеток СНО, полученные в Примере 5.9, мономер и димер одноцепочечного Fv MABL-2 из Е.coli. полученные в Примере 5.12, и IgG-антитело мыши, взятое в качестве контроля. После культивирования проводят окрашивание Аннексином-V и измеряют интенсивность флуоресценции при помощи прибора FACScan (BECTON DICKINSON).
Согласно второму протоколу образец антител в конечной концентрации 3 мкг/мл добавляют к 5×104 клеток клеточной линии hIAP/L1210, культивируют в течение 2 час и смешивают с анти-FLAG антителом (SIGMA) в конечной концентрации 15 мкг/мл и дополнительно культивируют в течение 22 час. Анализируют образец антител мономера одноцепочечного Fv из MABL-2 из клеток СНО, полученных в Примере 5.9, и IgG-антитело мыши, взятое в качестве контроля. После культивирования проводят окрашивание Аннексином-V и измеряют интенсивность флуоресценции при помощи FACScan (BECTON DICKINSON).
Результаты анализа с помощью окрашивания Аннексином-V показаны на Фиг.25-31. Данные результаты показывают, что, в сравнении с контрольным образцом (Фиг.25), димеры одноцепочечного Fv полипептида MABL-2, продуцируемые в клетках СНО и Е.coli, заметным образом индуцируют клеточную смерть (Фиг.26, 27), в то время как у мономеров одноцепочечного Fv полипептида MABL-2, продуцируемого в клетках СНО и Е.coli, такое апоптоз-индуцирующее действие не наблюдается (Фиг.28, 29). При использовании совместно с анти-FLAG антителом, мономер одноцепочечного Fv полипептида, происходящего из MABL-2, продуцированный в клетках СНО, в значительной степени, по сравнению с контрольным образцом, индуцирует клеточную смерть (Фиг.30).
5.14 Противоопухолевый эффект мономера и димера полипептида scFv/CHO на мышиной модели человеческой миеломы
(1) Количественное определение IgG человека в мышиной сыворотке
Количественное определение IgG человека (М-белок), продуцируемого клетками человеческой миеломы и содержащегося в мышиной сыворотке, проводят методом ELISA следующим образом. Вносят в каждую лунку 96-луночного планшета (Nunc) 100 мкл козлиного антитела против IgG человека (BIOSOURCE, Lot#7902), разведенного до концентрации 1 мкг/мл 0,1%-ным бикарбонатным буфером (рН 9,6), и инкубируют при 4°С в течение ночи до иммобилизации антитела. После блокирования в каждую лунку добавляют 100 мкл ступенчато разведенной мышиной сыворотки или IgG человека (CAPPEL, Lot#00915) в качестве эталона и инкубируют в течение 2 час при комнатной температуре. После отмывания добавляют 100 мкл меченного щелочной фосфатазой антитела против IgG человека (BIOSOURCE, Lot#7902), разведенного в 5000 раз, и в течение 1 час проводят инкубацию при комнатной температуре. После отмывания добавляют раствор субстрата. После инкубации измеряют поглощение при 405 нм при помощи MICROPLATE READER Model 3550 (BioRad). Концентрацию IgG человека в сыворотке мыши определяют по калибровочной кривой, построенной по величинам поглощения для IgG человека в качестве стандарта.
(2) Получение антител для введения
Для того, чтобы приготовить образцы для введения, мономер и димер scFv/CHO полипептида разводят до концентрации 0,4 мг/мл или 0,25 мг/мл соответственно стерилизованным фильтрованием PBS(-) непосредственно в день применения.
(3) Получение мышиной модели человеческой миеломы
Мышиную модель человеческой миеломы получают следующим образом. Клетки КРММ2, пассированные in vivo (JP-Appl. 7-236475) в мышах SCID (Japan Clare), суспендировали в среде RPMI1640 (GIBCO-BRL), содержащей 10%-ную фетальную бычью сыворотку (GIBCO-BRL), и разводили до концентрации 3×107 клеток/мл. 200 мкл суспензии клеток КРММ2 (6×106 клеток/мышь) трансплантируют через хвостовую вену мыши SCID (самец, 6-недельный), которой была сделана подкожная инъекция антитела к асиало-GM1 (WAKO JUNYAKU, 1 ампула, растворенная в 5 мл) за день до трансплантации.
(4) Введение антител
Образцы антител, приготовленные в (2), мономер (250 мкл) и димер (400 мкл) вводят в мышиные модели человеческой миеломы, полученные в (3), через хвостовую вену. Начинают введение через три дня после трансплантаци клеток и КРММ2 и проводят дважды в день в течение трех дней. Контрольной группе аналогичным образом вводят 200 мкл стерилизованного фильтрованием PBS(-) дважды в день в течение трех дней в хвостовую вену. Каждая группа включает по семь мышей.
(5) Оценка противоопухолевого эффекта мономера и димера scFv/CHO полипептида на мышной модели человеческой миеломы
Противоопухолевый эффект мономера и димера scFv/CHO полипептида на мышиной модели человеческой миеломы оценивают по изменению концентрации IgG человека (М-белок) в сыворотке мыши и по времени выживания мыши. Изменение концентрации IgG человека определяют, измеряя ее в мышиной сыворотке, собираемой спустя 24 дня после трансплантации КРММ2 клеток, при помощи ELISA, как описано выше (1). Количество сывороточного IgG человека (М-протеин) в сыворотке у той группы (контрольной), которой вводили PBS(-), возросло приблизительно до 8500 мкг/мл, в то время как количество IgG человека в группе, которой вводили димер scFv/CHO, было удивительно низким, а именно оно составляло одну десятую или менее от величины в контрольной группе. Следовательно, данные результаты показывают, что димер scFv/CHO сильно подавляет рост клеток КРММ2 (Фиг.32). Как показывает Фиг.33, в группе, которой вводили димер scFv/CHO, наблюдалось значительное увеличение времени выживания по сравнению с группой, которой вводили PBS(-).
Все вышеизложенное подтверждает, что димер scFv/CHO оказывает противоопухолевый эффект на мышиной модели человеческой миеломы. Предполагается, что противоопухолевый эффект димера scFv/CHO, модифицированного антитела настоящего изобретения, является результатом апоптоз-индуцирующего действия модифицированного антитела.
5.15 Реакция гемагглютинации
Реакцию гемагглютинации и определение гемагглютинации выполняют в соответствии с "Immuno-Biochemical Investigation", Zoku-Seikagaku Jikken Koza, изданным the Biochemical Society of Japan, опубликованным Tokyo Kagaku Dojin.
Берут кровь у здорового донора, используя обработанный гепарином шприц, и трижды отмывают PBS(-), а затем готовят суспензию эритроцитов с конечной концентрацией 2% в PBS(-). Для реакции берутся следующие образцы: антитело MABL-2, мономер и димер одноцепочечного Fv полипептида, продуцированного клетками СНО, и мономер и димер одноцепочечного Fv иолипептида, продуцированного Е.coli, в качестве контроля берется IgG мыши (ZYMED). Для исследования эффекта гемагглютинации применяются 96-луночные планшеты с круглыми донцами, производимые Falcon. В лунки планшета вносят по 50 мкл на лунку вышеупомянутых образцов антител и по 50 мкл 2%-ной суспензии эритроцитов и перемешивают. После инкубирования при 37°С в течение 2 час реакционную смесь оставляют на ночь при 4°С и определяют в них гемагглютинацию. В качестве контроля используют по 50 мкл PBS(-) на лунку и аналогичным образом осуществляют постановку реакции гемагглютинации. IgG мыши и антитело MABL-2 были использованы в следующих конечных концентрациях антител: 0,01, 0,1, 1,0, 10,0 или 100,0 мкг/мл. Одноцепочечные Fv были использованы в следующих конечных концентрациях антител: 0,004, 0,04, 0,4, 4,0, 40,0 или 80,0 мкг/мл, и дополнительно 160,0 мкг/мл только для димера полипептида, продуцированного Е.coli. Результаты приводятся в Таблице 2. Гемагглютинация в случае антитела MABL-2 наблюдается при концентрации, превышающей 0,1 мкг/мл, в то время как в случаях как мономера, так и димера одноцепочечного Fv гемагглютинация не наблюдается.
Реакция гемагглютинации
Пример 6 (модифицированное антитело sc(Fv)2, ключающее две V-области Н-цепи и две V-области L-цепи, и антитело MABL-2 scFv с линкерами различной длины)
6.1 Конструирование плазмиды, экспрессирующей модифицированное антитело sc(Fv)2 MABL-2
Для получения плазмиды, экспрессирующей модифицированное антитело [sc(Fv)2], которое содержит две V-области Н-цепи и две V-области L-цепи, происходящие из антитела MABL-2, вышеупомянутую рСНОМ2, которая содержит ДНК, кодирующую scFv, происходящую из MABL-2, описанный выше, модифицируют ПЦР-методом, как описано ниже, и встраивают полученный фрагмент ДНК в рСНОМ2.
Для ПЦР используются следующие праймеры: праймер EF1 (SEQ ID NO.:30) в качестве смыслового праймера, который конструируют так, чтобы он гибридизовался с ДНК, кодирующей EF1α, и антисмысловой праймер (SEQ ID NO.:19), который конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал последовательность ДНК, кодирующую область линкера, и праймер VLLAS, содержащий сайт узнавания рестриктазы Sall (SEQ ID NO.31).
100 мкл ПЦР-раствора содержит 10 мкл 10-кратного ПЦР-буфера #1,1 мМ MgCl2, 0,2 мМ dNTP (dATP, dGTP, dCTP, dTTP), 5 единиц ДНК-полимеразы KOD (Toyobo, Inc.), no 1 мкМ каждого праймера и 100 нг матричной ДНК (рСНОМ2). ПЦР-раствор последовательно нагревают при 94°С в течение 30 сек, при 50°С в течение 30 сек и при 74°С в течение 1 мин. Данный температурный цикл повторяют 30 раз.
ПЦР-продукт очищают при помощи набора QIAquick ПЦР Purification Kit (QIAGEN), а затем расщепляют с помощью Sall. Полученный фрагмент ДНК клонируют в вектор pBluescript KS+ (Toyobo, Inc.). После секвенирования ДНК плазмиду, содержащую требуемую ДНК-последовательность, расщепляют Sall и при помощи набора Rapid DNA Ligation Kit (BOEHRINGER MANNHEIM), присоединяют полученный ДНК-фрагмент к рСНОМ2, расщепленной с помощью Sall. После секвенирования ДНК плазмиду, содержащую искомую ДНК-последовательность, обозначают как "pCHOM2(Fv)2" (см. Фиг.34). Нуклеотидная последовательность и аминокислотная последовательность области антитела sc(Fv)2 MABL-2, включенной в плазмиду pCHOM2(Fv)2, показаны в SEQ ID NO.32.
6.2. Получение плазмиды, экспрессирующей антитела scFv MABL-2, имеющие линкеры различной длины.
scFv, содержащие линкеры различной длины, и V-области, имеющие следующие структуры [Н-цепь] - [L-цепь] (далее по тексту "HL") или [L-цепь] - [Н-цепь] (далее по тексту "LH"), получают, используя в качестве матриц кДНК, кодирующие Н-цепь и L-цепь, происходящие из MABL-2, как это будет описано ниже.
Для конструирования scFv типа HL проводят ПЦР, используя в качестве матрицы рСНОМ2 (Fv)2. На этапе ПЦР применяют пару праймеров CFHL-F1(SEQ ID NO.:33) и CFHL-R2 (SEQ ID NO.:34) или пару праймеров CFHL-F2 (SEQ ID NO.:35)) и CFHL-R1(SEQ ID NO.:36) и KOD-полимеразу. Для того, чтобы получить кДНК для Н-цепи, содержащей лидерную последовательностью на 5'-конце или кДНК для L-цепи, содержащей FLAG-последовательность на 3'-конце, проводят последовательность операций ПЦР, повторяя 30 раз температурный цикл, состоящий из нагревания при 94°С в течение 30 сек, 60°С в течение 30 сек и 72°С 1 мин в указанном порядке. Полученные кДНК для Н-цепи и L-цепи перемешивают и, используя их смесь как матрицы, а также KOD-полимеразу, проводят ПЦР путем 5-кратного повторения температурного цикла, состоящего из 94°С в течение 30 сек, 60°С в течение 30 сек, а потом 72°С в течение 1 мин в указанном порядке. К реакционой смеси добавляют праймеры CFHL-F1 и CFHL-R1 и проводят ПЦР, повторяя вышеуказанный температурный цикл 30 раз, чтобы получить кДНК для HL-0 типа без линкера.
Для конструирования scFv типа LH проводят ПЦР, используя в качестве матрицы pGEM-M2L и pGEM-M2H, которые включают кДНК, кодирующие соответственно V-область L-цепи и V-область Н-цепи антитела MABL-2 (см. JP-Appl. 11-63557). Используют пару праймеров Т7 (SEQ ID NO.:37) и CFLH-R2 (SEQ ID NO.:38) или пару праймеров CFLH-F2 (SEQ ID NO.:39) и CFLH-R1 (SEQ ID NO.:40), и KOD-полимеразу (Toyobo Inc.). Для получения кДНК для L-цепи, содержащей лидерную последовательность на 5'-конце, или кДНК для Н-цепи, содержащей FLAG-последовательность на 3'-конце, проводят ПЦР, повторяя 30 раз температурный цикл, состоящий из 30 сек при 94°С, 30 сек при 60°С и 1 мин при 72°С в указанном порядке. Полученные кДНК для L-цепи и Н-цепи смешивают, проводят ПЦР, используя в качестве матрицы полученную смесь и KOD-полимеразу, путем 5-кратного повторения температурного цикла, состоящего из 30 сек при 94°С, 30 сек при 60°С и 1 мин при 72°С в указанном порядке. К реакционной смеси добавляют праймеры Т7 и CFLH-R1 и осуществляют реакцию, повторяя 30 раз вышеупомянутый температурный цикл. Для получения кДНК для LH-0 типа без линкера используют в качестве матрицы продукт реакции и проводят ПЦР с использованием пары праймеров CFLH-F4 (SEQ ID NO.:41) и CFLH-R1 путем повторения 30 раз температурного цикла, состоящего из 30 сек при 94°С, 30 сек при 60°С и 1 мин при 72°С в указанном порядке.
Полученные кДНК для LH-0 и HL-0 типов расщепляют рестриктазами EcoRI и BamHI (Takara Shuzo) и встраивают расщепленные кДНК в экспрессионую плазмиду INPEP4 для клеток млекопитающих соответственно, используя набор Ligation High (Noyobo Inc.). Компетентные клетки Е.coli JM109 (Nippon Gene) трансформируют каждой из плазмид и выделяют желаемые плазмиды из трансформированной Е.coli при помощи набора QIAGEN Plasmid Maxi Kit (QIAGEN). Таким образом получают плазмиды pCF2LH-0 и pCF2HL-0.
Для конструирования экспрессионных плазмид HL-типа, содержащих линкеры разного размера, используют в качестве матрицы pCF2HL-0, в качестве смыслового праймера CFHL-X3 (SEQ ID NO.:42), CFHL-X4 (SEQ ID NO.:43), CFHL-X5 (SEQ ID NO.:44), CFHL-X6 (SEQ ID NO.:45) или CFHL-X7 (SEQ ID NO.:46), и в качестве антисмыслового праймера BGH-1 (SEQ ID NO.:47), который комплементарен векторной последовательности. ПЦР проводят с использованием KOD-полимеразы, повторяя 30 раз температурный цикл, состоящий из 30 сек при 94°С, 30 сек при 60°С и 1 мин при 72°С в указанном порядке; продукты реакции расщепляют рестриктазами XhoI и BamHI (Takara Shuzo). Расщепленные фрагменты встраивают в pCF2HL-0 соответственно между сайтами XhoI и BamHI, используя набор Ligation High (Toyobo Inc.). Трансформируют компетентные клетки Е.coli JM109 каждой из плазмид и с помощью набора Qiagen Plasmid Maxi Kit выделяют требуемые плазмиды из трансформированной Е.coli. Таким образом получают экспрессионные плазмиды pCF2HL-3, pCF2HL-4, pCF2HL-5, pCF2HL-6 и pCF2HL-7.
Чтобы сконструировать экспрессионную плазмиду для временной экспрессии в клетках COS7, с помощью рестриктаз EcoRI и BamHI (Takara Shuzo) расщепляют плазмиды pCF2HL-0, pCF2HL-3, pCF2HL-4, pCF2HL-5, pCF2HL-6 и pCF2HL-7 и очищают полученные фрагменты размером приблизительно 800 п.о. с помощью электрофореза в агарозном геле. Полученные фрагменты встраивают, используя набор Ligation High (Toyobo Inc.), соответственно между сайтами EcoRI и BamHI экспрессионной плазмиды pCOS1 для экспрессии в клетках млекопитающих. Трансформируют каждой из плазмид компетентные клетки Е.coli DH5α (Toyobo Inc.) и выделяют из трансформированной Е.coli требуемые плазмиды, используя набор Qiagen Plasmid Maxi Kit. Таким образом получают экспрессионные плазмиды pCF2HL-0/pCOS1, pCF2HL-3/pCOS1, pCF2HL-4/pCOS1, pCF2HL-5/pCOS1, pCF2HL-6/pCOS1 и pCF2HL-7/pCOS1.
Фиг.35 иллюстрирует, в качестве типичного примера для данных плазмид, конструирование плазмиды pCF2HL-0/pCOS1, а нуклеотидная последовательность и аминокислотная последовательность MABL2-scFv <HL-0>, включенные в плазмиду, показаны в SEQ ID NO.:48. Нуклеотидные последовательности и аминокислотные последовательности линкерных областей этих плазмид показаны также на Фиг.36.
Для того чтобы сконструировать экспрессионные плазмиды LH-типа, содержащие линкеры разного размера, используют в качестве матрицы pCF2LH-0, в качестве смыслового праймера CFLH-X3 (SEQ ID NO.:49), CFLH-X4 (SEQ ID NO.:50), CFLH-X5 (SEQ ID NO.:51), CFLH-X6 (SEQ ID NO.:52) или CFLH-X7 (SEQ ID NO.:53), и праймер BGH-1 в качестве антисмыслового праймера, который комплементарен векторной последовательности. Проводят ПЦР с использованием KOD-полимеразы, повторяя 30 раз температурный цикл, состоящий из 30 сек при 94°С, 30 сек при 60°С и 1 мин при 72°С в указанном порядке; продукты реакции расщепляют рестриктазами XhoI и BamHI. Расщепленные фрагменты встраивают соответственно между сайтами XhoI и BamHI в pCF2LH-0, используя набор Ligation High. Каждой из плазмид трансформируют компетентные клетки Е.coli DH5a (Toyobo Inc.) и с помощью набора Qiagen Plasmid Maxi Kit выделяют требуемые плазмиды из трансформированной Е.coli. Таким образом получают экспрессионные плазмиды pCF2LH-3, pCF2LH-4, pCF2LH-5, pCF2LH-6 и pCF2LH-7.
Чтобы сконструировать экспрессионную плазмиду для временной экспрессии в клетках COS7, с помощью рестриктаз EcoRI и BamHI (Takara Shuzo) расщепляют плазмиды pCF2LH-0, pCF2LH-3, pCF2LH-4, pCF2LH-5, pCF2LH-6 и pCF2LH-7 и очищают полученные фрагменты размером приблизительно 800 п.о. с помощью электрофореза в агарозном геле. Полученные фрагменты встраивают, используя набор Ligation High, соответственно между сайтами XhoI и BamHI экспрессионной плазмиды pCOS1 для экспрессии в клетках млекопитающих. Каждой из плазмид трансформируют компетентные клетки Е.coli DHα (Toyobo Inc.) и выделяют из трансформированной E. coli требуемые плазмиды, используя набор Qiagen Plasmid Maxi Kit. Таким образом получают экспрессионные плазмиды CF2LH-0/pCOS1, CF2LH-3/pCOS1, CF2LH-4/pCOS1, CF2LH-5/pCOS1, CF2LH-6/pCOS1 и CF2LH-7/pCOS1.
Фиг.37 иллюстрирует, в качестве типичного примера для данных плазмид, конструирование плазмиды CF2LH-0/pCOS1, а нуклеотидная последовательность и аминокислотная последовательность MABL2-scFv <LH-0>, включенные в плазмиду, показаны в SEQ ID NO.:54. Нуклеотидные последовательности и аминокислотные последовательности линкерных областей этих плазмид показаны также на Фиг.38.
6.3 Экспрессия scFv и sc(Fv)2 в клетках COS7
(1) Получение культурального супернатанта с использованием сывороточной культуральной среды.
scFv и sc(Fv)2 HL-типа и LH-типа временно экспрессировались в клетках COS7 (JCRB9127, Japan Health Sciences Foundation). Клетки COS7 пассируют на среде DMEM (GIBCO BRL), содержащей 10%-ную фетальную бычью сыворотку (FBS) (HyClone), при 37°С в CO2-инкубаторе. Клетки COS7 трансфицируют векторами CF2HL-0, 3˜7/pCOS1 или CF2LH-0, 3˜7/pCOS1, полученными в Примере 6.2, или рСНОМ2 (Fv)2 при помощи электропорации с использованием аппарата Gene Pulser (BioRad). ДНК (10 мкг) и 0,25 мл суспензии клеток с концентрацией 2×107 клеток/мл в культуральной среде DMEM, содержащей 10%-ную FBS и 5 мМ BES (SIGMA), вносят в кювету. Дают смеси постоять 10 минут и обрабатывают импульсом при 0,17 кВ, при электрической емкости 950 мкФ. Через 10 минут после восстановления при комнатной температуре электропорированные клетки переносят в колбу объемом 75 см3 с культуральной средой DMEM (10% FBS). После культивирования в течение 72 часов культуральный супернатант собирают и удаляют центрифугированием клеточные фрагменты. Для получения культурального супернатанта (далее по тексту СМ), культуральный супернатант фильтруют с помощью насадки для фильтрования с диаметром пор 0,22 мкм (FALCON).
(2) Приготовление культурального супернатанта с использованием бессывороточной культуральной среды
Клетки, трансфицированные таким же образом, как это делается в (1), переносят в колбу объемом 75 см3 с культуральной средой DMEM (10%FBS) и культивируют в течение ночи. После культивирования супернатант отбрасывают, а клетки отмывают раствором PBS, а затем вносят в среду CHO-S-SFM II (GIBCO BRL). После культивирования в течение 72 часов культуральный супернатант собирают, центрифугируют для удаления клеточных фрагментов и фильтруют с помощью насадки для фильтрования с диаметром пор 0,22 мкм (FALCON) для получения СМ.
6.4 Обнаружение scFv и sc(Fv)2 в СМ клеток COS7
Различные MABL2-scFvs и sc(Fv)2 в СМ клеток COS7, полученные в приведенном выше Примере 6.3 (2), выявляют методом Western блоттинга.
Каждый СМ клеток COS7 подвергают SDS-PAGE и переносят на мембрану REINFORCED NC (Schleicher & Schuell). Мембрану блокируют 5%-ным снятым молоком (Morinaga Nyu-gyo) и отмывают раствором TBS. Затем туда добавляют анти-FLAG антитело (SIGMA). Мембрану инкубируют при комнатной температуре и отмывают. Добавляют меченое пероксидазой IgG-антитело мыши (Jackson Immuno Research). После инкубирования и отмывания при комнатной температуре добавляют раствор субстрата (Kirkegaard Perry Laboratories) для развития окраски (Фиг.39).
6.5 Проточная цитометрия
Для измерения связывания MABL2-scFVs и sc(Fv)2 с антигеном - интегринассоциированным белком человека (IAP) применяют метод проточной цитометрии, используя культуральные супернатанты клеток COS7, полученные в примере 6.3 (1). Добавляют тестируемые культуральные супернатанты или контрольный культуральный супернатант клеток COS7 к 2×105 клеток клеточной линии лейкемии мыши L1210, экспрессирующих человеческий IAP. После инкубирования на льду и отмывания добавляют 10 мкг/мл мышиного анти-FLAG антитела (SIGMA), затем клетки инкубируют и отмывают. Затем туда добавляют меченное ФИТЦ антитело против мышиного IgG (BECTON DICKINSON) и клетки еще раз инкубируют и отмывают. Измеряют интенсивность флуоресценции при помощи аппарата FACScan (BECTON DICKINSON). Как показывают результаты проточной цитометрии, MABL2-scFv с линкерами различной длины и sc(Fv)2 в культуральных супернатантах клеток COS7 имеют высокое сродство к человеческому LAP (см. Фиг.40а и 40b).
6.6 Апоптоз-индуцирующий эффект in vitro
Апоптоз-индуцирующее действие культуральных супернатантов COS7, полученных в Примере 6.3 (1), исследуют при помощи окрашивания Аннексином-V (Boehringer Mannheim) с использованием клеток L1210, трансфицированных геном IAP человека (hIAP/L1210).
К 5×104 клеток hIAP/L1210 добавляют культуральные супернатанты клеток COS7, трансфицированных каждым вектором, или культуральный супернатант клеток COS7 в качестве контроля 10%-ной конечной концентрации и культивируют даные смеси в течение 24 часов. Затем проводят окрашивание Аннексином-V/PI и определяют интенсивность флуоресценции при помощи аппарата FACScan (BECTON Dickinson). Как показывают результаты, scFv <HL3, 4, 6, 7, LH3, 4, 6, 7> и sc(Fv)2 в CM клеток COS7 в весьма значительной степени индуцируют клеточную смерть клеток hIAP/L1210. Эти результаты отражены на Фиг.41.
6.7 Конструирование векторов для экспрессии scFv и sc(Fv)2 в клетках СНО
Для выделения и очистки MABL2-scFv и sc(Fv)2 из культурального супернатанта конструируют экспрессионные векторы для экспрессии в клетках СНО, как описано ниже.
EcoRI-BamHI фрагменты pCF2HL-0, 3˜7, и pCF2LH-0, 3˜7, полученные в Примере 6.2, встраивают между сайтами EcoRI и BamHI экспрессионного вектора рСНО1 для клеток СНО, используя набор Ligation High. Или трансформируют компетентные клетки Е.coli DH5α. Выделяют плазмиды из трансформированной Е.coli с помощью набора QIAGEN Plasmid Midi Kit (QIAGEN) для получения экспрессионных плазмид pCHOM2HL-0, 3˜7 и pCHOM2LH-0, 3˜7.
6.8 Получение клеток СНО, экспрессирующих MABL2-scFv <HL-0, 3˜7>, MABL2-scFv <LH-0, 3˜7> и sc(Fv)2, и получение культуральных супернатантов
Для получения клеток СНО, константно экспрессирующих каждое модифицированное антитело, клетки СНО трансформируют каждой из экспрессионных плазмид pCHOM2HL-0, 3˜7, и pCHOM2LH-0, 3˜7, сконструированных в Примере 6.7, и вектором рСНОМ2 (Fv)2. В качестве типичного примера ниже приводится получение СНО-клеток, константно экспрессирующих MABL2-scFv <HL-5> или sc(Fv)2.
Экспрессионные плазмиды pCHOM2HL-5 и pCHOM2(Fv)2 линеаризируют расщеплением фермента рестриктазами Pvul и трансфицируют в клетки СНО путем электропорации при помощи аппарата Gene Pulser (BioRad). Вносят в кювету ДНК (10 мкг) и 0,75 мл PBS 1×107 клеток/мл и обрабатывают импульсом при 1,5 кВ, при электрической емкости 25 мкФ. После восстановления при комнатной температуре в течение 10 мин электропорированные клетки переносят в содержащую нуклеиновую кислоту культуральную среду α-МЕМ (GIBCO BRL), содержащую 10%-ную фетальную бычью сыворотку, и культивируют. После культивирования в течение ночи супернатант отбрасывают. Клетки отмывают раствором PBS и переносят в свободную от нуклеиновой кислоты культуральную среду α-МЕМ (GIBCO BRL), содержащую 10%-ную фетальную бычью сыворотку. После культивирования в течение 2 недель клетки культивируют на среде, содержащей 10 нМ (конечная концентрация) метотрексат (SIGMA), затем 50 нМ и 100 нМ метотрексат. Полученные клетки культивируют в бессывороточной CHO-S-SFM II среде (GIBCO BRL) в роллерном флаконе. Затем культуральный супернатант собирают, центрифугированием удаляют клеточные фрагменты и фильтруют соответственно при помощи фильтра с размером пор 0,22 мкм для получения СМ.
В соответствии с изложеным выше получают клетки СНО, которые константно экспрессируют MABL2-scFv <HL-0, -3, -4, -6, -7> и <LH-0, -3, -4, -5, -6, -7>, и их СМ.
6.9 Очистка димера MABL2-scFv <HL-5> и sc(Fv)2
MABL2-scFv <HL-5> и sc(Fv)2 очищают из CM, полученных в Примере 6.8, используют два способа очистки, приводимых ниже.
<Способ очистки 1>
HL-5 и sc(Fv)2 очищают с помощью хроматографии на колонке с иммобилизованными анти-FLAG антителами с использованием FLAG-последовательности, расположенной на С-конце полипептидов, и с помощью гель-фильтрации. Один литр СМ, полученного в 6.8, наносят на колонку (7,9 мл), заполненную гелем anti-FLAG M2 Affinity gel (SIGMA), уравновешенным 50 мМ Tris-HCl буфером (TBS, pH 7,5), содержащим 150 мМ NaCl. После промывания колонки раствором TBS, scFv элюируют буфером 0,1 М глицин-HCl, pH 3,5. Полученные фракции анализируют методом SDS-PAGE и подтверждают элюирование scFv. Смешивают фракцию scFv с Tween 20 с конечной концентрацией 0,01% и концентрируют, используя CENTRICON-10 (MILLIPORE). Концентрат наносят на колонку с TSKgel G3000SWG (7,5×600 мм), уравовешенную 20 мМ ацетатным буфером (pH 6,0), содержащим 150 мМ NaCl и 0,01% Tween 20. При скорости подачи 0,4 мл/мин scFv детектируют по поглощению при 280 нм. HL-5 элюируется в виде основной фракции в положении димера и sc(Fv)2 элюируется в положении мономера.
<Способ очистки 2>
HL-5 и sc(Fv)2 очищают в три этапа, включающие ионообменную хроматографию, хроматографию на гидроксиапатите и гель-фильтрацию. При ионообменной хроматографии используется колонка с Q-Sepharose Fast Flow (Pharmacia) для HL-5 и с SP-Sepharose Fast Flow для sc(Fv)2. На этапе 2 и после него HL-5 и sc(Fv)2 обрабатывают по единой схеме.
Первый этап для HL-5
СМ HL-5 разводят в 2 раза в 20 мМ Tris-HCl буфере (рН 9,0), содержащем 0,02% Tween 20, а затем доводят рН до 9,0 с помощью 1 М Tris. Раствор наносят на колонку Q-Sepharose Fast Flow, уравновешенную 20 мМ Tris-HCl буфером (рН 8,5), содержащим 0,02% Tween 20. Элюируют полипептид, адсорбированный на колонке линейным градиентом NaCl в том же буфере: от 0,1 до 0,55 М. Проводя мониторинг элюируемых фракций методом SDS-PAGE, отбирают фракции, содержащие HL-5, и на 2 этапе проводят хроматографию на гидроксиапатите.
Первый этап для sc(Fv)2
CM sc(Fv)2 разводят 2 раза 20 мМ ацетатным буфером (рН 5,5), содержащим 0,02% Tween 20, и доводят его рН до 5,5 с помощью 1 М уксусной кислоты. Раствор наносят на SP-Sepharose Fast Row, уравновешенную 20 мМ ацетатным буфером (рН 5,5), содержащим 0,02% Tween 20. Полипептид, адсорбированный на колонке, элюируют линейным градиентом NaCl в буфере: от 0 до 0,5 М. Проводя мониторинг элюируемых фракций методом SDS-PAGE, собирают фракции, содержащие sc(Fv)2, и на втором этапе проводят хроматографию на гидроксиапатите.
Второй этап: хроматография HL-5 и sc(Fv)2 на гидроксиапатите
Фракции HL-5 и sc(Fv)2, полученные на этапе 1, раздельно наносят на колонку с гидроксиапатитом (Type I, BIORAD), уравновешенную 10 мМ фосфатным буфером, содержащим 0,02% Tween 20, рН 7,0. После промывания колонки тем же буфером элюируют полипептиды, адсорбированные на колонке, линейным градиентом фосфатного буфера до концентрации 0,5 М. Проводя мониторинг элюируемых фракций при помощи SDS-PAGE, собирают фракции, содержащие требуемые полипептиды.
Третий этап: гель-фильтрация HL-5 и sc(Fv)2
Фракции, полученные на этапе 2, раздельно концентрируют с помощью CentriPrep-10 (MILIPORE) и наносят на колонку Superdex 200 (2,6×60 см, Pharmacia), уравновешенную 20 мМ ацетатным буфером (рН 6,0), содержащим 0,02% Tween 20 и 0,15 М NaCl. HL-5 элюируется в положении димера, a sc(Fv)HL-5 и sc(Fv)2 соответственно элюированы как основные пики в положении мономера.
Поскольку мономер HL-5 практически не обнаруживается при использовании обоих способов очистки, это доказывает, что димеры одноцепочечных Fv образуются с высоким выходом в тех случаях, когда линкер для одноцепочечного Fv содержит около 5 аминокислот. Кроме того, димеры HL-5 и sc(Fv)2 после очистки стабильно хранятся в течение месяца при 4°С.
6.10 Оценка связывающей активности очищенного димера scFv<HL-5> и sc(Fv)2 в отношении антигена
Проводят проточную цитометрию с использованием очищенного димера MABL2-scFv<HL-5> и очищенного sc(Fv)2, чтобы оценить связывание с антигеном - интегринассоциированным белком человека (IAP). 10 мкг/мл очищенного димера Маbl2-scFv<HL-5>, очищенного sc(Fv)2, антитело Mabl-2 как положительный контроль и мышиный IgG (Zymed) как отрицательный контроль добавляют к 2×105 клеток клеточной линии лейкемии мыши L1210, экспрессирующей человеческий IAP (hIAP/L1210), или клеточной линии L1210, трансформированной pCOS1 (pCOS1/L1210), в качестве контроля. После инкубирования на льду и отмывания добавляют 10 мкг/мл анти-FLAG антитела мыши (SIGMA) и затем клетки инкубируют и отмывают. К ним добавляют меченное ФИТЦ антитело против мышиного IgG (BECTON DICKINSON) и снова инкубируют клетки и отмывают. Затем измеряют интенсивность флуоресценции, используя аппарат FACScan (BECTON DICKINSON).
Поскольку очищенный димер MABL2-scFv<HL-5> и очищенный sc(Fv)2 специфически связываются с клетками hIAP/L1210, то этим подтверждается, что димер sc(Fv)<HL-5> и sc(Fv)2 имеют высокое сродство к человеческому IAP (см. Фиг.42).
6.11 Апоптоз-индуцирующая активность очищенного димера scFv<HL-5> и sc(Fv)2 in vitro
Апоптоз-индуцирующее действие очищенного димера MABL2-scFv<HL-5> и очищенного sc(Fv)2 исследуют методом окрашивания Аннексином-V (Boehringer Mannheim) с использованием клеток L1210 (hIAP/11210), в которые был введен ген человеческого LAP, и клетки клеточной линии лейкемии человека CCRF-CEM.
К 5×104 клеток клеточной линии hIAP/L1210 или к 1×105 клеток клеточной линии CCRF-CEM добавляют различные концентрации очищенного димера MABL2-scFv <HL-5>, очищенного MABL2-sc(Fv)2, антитела MABL-2 в качестве положительного контроля или IgG мыши в качестве отрицательного контроля. После культивирования в течение 24 часов проводят окрашивание Аннексином-V и при помощи прибора FACScan (BECTON DICKINSON) измеряют интенсивность флуоресценции. Как показывают результаты, димеры MABL2-scFv <HL-5> и MABL2-sc(Fv)2 в сильной степени индуцируют клеточную смерть hIAP/L1210 и CCRF-CEM зависимым от концентрации способом (см. Фиг.43). В результате показано, что димеры MABL2-scFv <HL-5> и MABL2-sc(Fv)2 значительно эффективнее индуцируют апоптоз в сравнении с исходным антителом MABL-2.
6.12 Реакция гемагглютинации с очищенным димером scFv <HL-5> и sc(Fv)2
Реакцию гемагглюцинации ставят, используя различные концентрации очищенного димера scFv <HL-5> и очищенного sc(Fv)2 в соответствии с Примером 5.15.
Гемагглюцинация наблюдалась с антителом MABL-2 в качестве положительного контроля, в то время как в случае с одноцепочечным антителом MABL2-sc(Fv)2, так и в случае MABL2-scFv <HL-5> гемагглюцинации не наблюдалось. Помимо этого, никакого существенного различия в гемагглюцинации между двумя буферами, использованными в эксперименте с антителом MABL-2, не наблюдалось. Данные результаты отражены в Таблице 3.
Реакция гемагглютинации
6.13 Противоопухолевое действие очищенных димера scFv <HL-5> и sc(Fv)2 на мышиной модели человеческой миеломы
Испытывают противоопухолевое действие димера scFv <HL-5> и sc(Fv)2, полученных и очищенных в Примерах 6.8 и 6.9. Эксперимент проводят, используя мышиную модель человеческой миеломы, полученной в Примере 5.1, и определяя количество М-белка, продуцируемого клетками человеческой миеломы в мышиной сыворотке, методом ELISA, и наблюдая за временем выживания мышей. Затем оценивают противоопухолевые эффекты димера scFv <HL-5> и sc(Fv)2 по изменению количества М-белка в мышиной сыворотке и продолжительности выживания мышей.
Для эксперимента HL-5 и sc(Fv)2 применяют в виде раствора с концентрацией 0,01, 0,1 или 1 мг/мл в растворителе, содержащем 150 мМ NaCl, 0,02% Твин и 20 мМ ацетатный буфер (рН 6,0), и вводят мышам в дозировках 0,1, 1 или 10 мг/кг. Контрольной группе мышей вводят только растворитель.
Спустя 26 дней после трансплантации клеток человеческой миеломы собирают мышиную сыворотку и измеряют количество М-белка в сыворотке методом ELISA в соответствии с Примером 5.14. Как показывают результаты, количество М-белка в сыворотке мышей обеих групп, которым были введены димер HL-5 и sc(Fv)2, уменьшалось зависящим от дозы образом (см. Фиг.44). Помимо этого, наблюдалось значительное увеличение времени выживания в обеих группах, которым были введены HL-5 (Фиг.45) и sc(Fv)2 (Фиг.46), в сравнении с контрольной группой, которой вводили только растворитель. Как показывают данные результаты, HL-5 и sc(Fv)2 по изобретению обладают отличным противоопухолевым эффектом in vivo.
Пример 7
Одноцепочечный Fv, включающий V-область Н-цепи и V-область L-цепи человеческого антитела 12В5 к человеческому MPL
ДНК, кодирующую V-области человеческого моноклонального антитела 12В5 к человеческому MPL, конструируют следующим образом.
7.1 Конструирование гена, кодирующего V-область Н-цепи 12В5
Ген, кодирующий V-область Н-цепи человеческого антитела 12В5, связывающегося с человеческим MPL, конструируют путем присоединения нуклеотидной последовательности данного гена (SEQ ID NO.:55) по 5'-концу к лидерной последовательности (SEQ ID NO.:56), происходящей из гена человеческого антитела (Eur. J. Immunol. 1996; 26; 63-69). Сконструированную нуклеотидную последовательность разбивают на 4 олигонуклеотида, содержащих частично перекрывающиеся последовательности длиной 15 п.о. каждая (12B5VH-1, 12B5VH-2, 12B5VH-3, 12B5VH-4). 12 ВН5-1 (SEQ ID NO.:57) и 12B5VH-3 (SEQ ID NO.:59) синтезированы в смысловой ориентации, а 12B5VH-2 (SEQ ID NO.:58) и 12B5VH-4 (SEQ ID NO.:60) соответственно в антисмысловой ориентации. После состыковки каждого из синтезированных олигонуклеотидов по соответствующей комплементарности добавляют внешние праймеры (12B5VH-S и 12B5VH-A) для амплификации полноразмерного гена. 12B5VH-S (SEQ ID NO.:61) в качестве прямого праймера конструируют так, чтобы он гибридизовался с 5'-концом лидерной последовательности и содержал сайт узнавания рестриктазы Hind III и последовательность Козака, а 12B5VH-A (SEQ ID NO.:62) конструируют в качестве обратного праймера так, чтобы он гибридизовался с нуклеотидной последовательностью, кодирующей С-конец V-области Н-цепи, и содержал донорную последовательность спласинга и сайт узнавания рестриктазы BamHI соответственно.
100 мкл ПЦР-раствора, содержащего 10 мкл 10-кратного буфера PCR Gold Buffer II, 1,5 мМ MgCl2, 0,08 мМ dNTP (dATP, dGTP, dCTP, dTTP), 5 единиц ДНК-полимеразы AmpliTaq Gold (все от PERIKIN ELMER) и по 2,5 пмоля каждого синтезированного олигонуклеотида (12B5VH-1 до - 4), нагревают при начальной температуре 94°С в течение 9 мин, при 94°С 2 мин, при 55°С 2 мин, при 72°С 2 мин. После двукратного повторения цикла добавляют по 100 пмолей каждого из внешних праймеров 12B5VH-S и 12B5VH-A. Смесь подвергают 35 циклам, состоящим из 30 сек при 94°С, 30 сек при 55°С, 1 мин при 72°С, после чего смесь дополнительно нагревают в течение 5 мин при 72°С.
ПЦР-продукт очищают в 1,5%-ном геле легкоплавкой агарозы (Sigma), расщепляют рестриктазы BamHI и Hind III и клонируют в экспрессионный вектор HEF-gγl для Н-цепи человека. После определения последовательности ДНК плазмиду, содержащую правильную последовательность ДНК, обозначают как HEF-12B5VH-gγl.
Расщепляют HEF-12B5VH-gγl рестриктазы EcoRI и BamHI для получения гена, кодирующего 12B5VH, который затем клонируют в экспрессионный вектор pCOS-Fd для Fab Н-цепи человека, чтобы получить pFd-12B5H. Экспрессионный вектор для Н-цепи Fab человека конструируют, амплифицируя методом ПЦР ДНК (SEQ ID NO.:63), содержащую область интрона, расположенную между генами, кодирующими V-область Н-цепи человеческого антитела и константную область, и ген, кодирующий часть константной области человеческой Н-цепи, и встраивая ПЦР-продукт в экспрессионный вектор pCOS1 для животных клеток. Константную область Н-цепи человека для гена амплифицируют при тех же условиях, что и выше, используя HEF-gγl в качестве матрицы, G1CH1-S (SEQ ID NO.:64) в качестве прямого праймера, который конструировали так, чтобы он гибридизовался с 5'-концевой последовательностью интрона 1 и содержал сайты узнавания рестриктаз EcoRI и BamHI, и G1CH1-A (SEQ ID NO.:65) в качестве обратного праймера, который конструировали так, чтобы он гибридизовался с 5'-концом ДНК СН1-домена константной области Н-цепи человека, и содержал последовательность, кодирующую часть шарнирной области, 2 стоп-кодона и сайт узнавания рестриктазы Bgl II.
Нуклеотидная последовательность и аминокислотная последовательность реконструированной вариабельной области Н-цепи 12В5, включенные в плазмиды HEF-12B5VH-gγl и pFd-12B5H, показаны в SEQ ID NO:66.
7.2 Конструирование гена, кодирующего V-область L-цепи 12В5
Ген, кодирующий V-область L-цепи человеческого антитела 12В5, связывающийся с человеческим MPL, конструируется путем присоединения нуклеотидной последовательности гена (SEQ ID NO.:67) по 5'-концу к лидерной последовательности (SEQ ID NO.:68), происходящей из гена человеческого антитела 3D6 (Nuc. Acid Res. 1990; 18; 4927). Также, как в вышеприведенном примере, сконструированную нуклеотидную последовательность разбивают на 4 олигонуклеотида, содержащих частично перекрывающиеся последовательности длиной 15 п.о. каждая (12B5VL-1, 12B5VL-2, 12B5VL-3, 12B5VL-4) и синтезируют соответственно. 12B5VL-1 (SEQ ID NO.:69) и 12B5VL-3 (SEQ ID NO.:71) имеют смысловую ориентацию, а 12B5VL-2 (SEQ ID NO.:70) и 12B5VL-4 (SEQ ID NO.:72) имеют антисмысловую ориентацию соответственно. После состыковки каждого из синтезированных олигонуклеотидов в соответствии с комплементарностью добавляют внешние праймеры (12B5VL-S и 12B5VL-A) для амплификации полноразмерного гена. 12B5VL-S (SEQ ID NO.:73) как прямой праймер конструируют так, чтобы он гибридизовался с 5'-концом лидерной последовательности и содержал сайт узнавания рестриктазы Hind III и последовательность Козака. 12B5VL-A (SEQ ID NO.:74) как обратный праймер конструируют так, чтобы он гибридизовался с нуклеотидной последовательностью, кодирующей С-конец V-области L-цепи и содержал соответственно донорную последовательность сплайсинга и сайт узнавания рестриктазы BamHI.
По проведении ПЦР, как было упомянуто выше, продукт реакции очищают в 1,5%-ном геле легкоплавкой агарозы (Sigma), расщепляют рестриктазами BamHI и Hind III и клонируют в экспрессионный вектор HEF-gK для L-цепи человека. После определения последовательности ДНК плазмиду, содержащую правильную последовательность ДНК, обозначают как HEF-12B5VL-gK. Нуклеотидная последовательность и аминокислотная последовательность реконструированной V-области L-цепи 12В5, включенные в плазмиду HEF-12B5VL-gK, показаны в SEQ ID NO:75.
7.3 Получение реконструированного одноцепочечного Fv (scFv) 12B5
Реконструированный одноцепочный Fv антитела 12B5 конструируют так, чтобы он имел структуру 12В5УН-линкер-12В5VL и содержал FLAG-последовательность (SEQ ID NO.:76) на С-конце для облегчения детекции и очистки. Реконструированный одноцепочечный Fv 12B5 (sc12B5) конструируют, используя линкерную последовательность, состоящую из 15 аминокислот, представленных (Gly4Ser)3.
(1) Получение реконструированного одноцепочечного Fv 12B5 с использованием линкерной последовательности, состоящей из 15 аминокислот
Ген, кодирующий реконструированный одноцепочечный Fv антитела 12B5, содержащий линкерную последовательность, состоящую из 15 аминокислот, конструируют путем соединения V-области Н-цепи 12B5, линкерной области и V-области L-цепи 12B5, которые амплифицируют соответственно методом ПЦР. Этот способ схематически представлен на Фиг.47. Для получения реконструированного одноцепочечного Fv 12B5 используется шесть праймеров ПЦР (A-F). Праймеры А, С и Е имеют смысловые последовательности, а праймеры В, D и F - антисмысловые последовательности.
Прямой праймер 12B5-S (праймер A, SEQ ID NO.:77) для V-области Н-цепи конструируют так, чтобы он гибридизовался с 5'-концом лидерной последовательности Н-цепи и содержал сайт узнавания рестриктазы EcoRI. Обратный праймер HuVHJ3 (Праймер В, SEQ ID NO.:78) для V-области Н-цепи конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области Н-цепи.
Прямой праймер RHuJH3 (праймер С, SEQ ID NO.:79) для линкера конструируют так, чтобы он гибридизовался с ДНК, кодирующей N-конец линкера, и перекрывался с ДНК, кодирующей С-конец V-области Н-цепи. Обратный праймер RHuVK1 (праймер D, SEQ ID NO.:80) для линкера конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец линкера, и чтобы перекрывался с ДНК, кодирующей N-конец V-области L-цепи.
Прямой праймер HuVK1.2 (праймер Е, SEQ ID NO.:81) для V-области L-цепи конструируют так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области L-цепи. Обратный праймер 12B5F-A для V-области L-цепи (праймер F, SEQ ID NO.:82) конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал последовательность, кодирующую FLAG пептид (Норр Т.Р. et al., Bio/Technology, 6, 1204-1210, 1988), два транскрипционных стоп-кодона и сайт узнавания рестриктазы NotI.
На первом этапе ПЦР ставят три реакции, А-В, C-D и E-F, и три ПЦР-продукта, полученные на этапе 1, состыковывают в соответствии с комплементарностью. После добавления праймеров А и F амплифицируют полноразмерную ДНК, кодирующую реконструированный одноцепочечный Fv 12B5, имеющий линкер, состоящий из 15 аминокислот (вторая ПЦР). На первом этапе ПЦР используют в качестве матриц соответственно плазмиду HEF-12B5H-gγl (см. Пример 7.1), кодирующую реконструированную V-область Н-цепи 12B5, pSCFVT7-hM21 (гуманизированное антитело ONS-M21) (Ohtomo et al., Anticancer Res. 18 (1998), 4311-4316), содержащую ДНК (SEQ ID NO.:83), кодирующую линкерную область, состоящую из Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser (Huston et al., Proc. Natl. Acad. Sci. USA, 85, 5879-5883, 1988), и плазмиду HEF-12B5L-gκ (см. Пример 7.2), кодирующую реконструированную V-область L-цепи 12B5.
В 50 мкл ПЦР-раствора первого этапа содержится 5 мкл 10-кратного буфера ПЦР Gold Buffer II, 1,5 мМ MgCl2, 0,08 мМ каждого dNTP, 5 единиц ДНК-полимеразы AmpliTaq Gold (все от PERKIN ELMER) и по 100 пмоль каждого праймера и по 100 нг каждой матричной ДНК. ПЦР-раствор нагревают при начальной температуре 94°С в течение 9 мин, при 94°С 30 сек, затем при 55°С 30 сек, при 72°С 1 мин. Данный цикл повторяют 35 раз, после чего реакционную смесь дополнительно нагревают при 72°С в течение 5 мин.
ПЦР-продукты А-В, C-D и E-F состыковывают при помощи второй ПЦР. ПЦР-смесь для второго этапа объемом 98 мкл, содержащую в качестве матрицы 1 мкл продукта А-В первой ПЦР, 0,5 мкл ПЦР0-продукта C-D и 1 мкл ПЦР-продукта E-F, 10 мкл 10-кратного буфера ПЦР Gold Buffer II, 1,5 мМ MgCl2, 0,08 мМ каждого dNTP, 5 единиц ДНК-полимеразы AmpliTaq Gold (все от PERKIN ELMER), нагревают при начальной температуре 94°С в течение 9 мин, затем при 94°С 2 мин, при 65°С 2 мин, при 72°С 2 мин. После двукратного повторения данного цикла добавляют по 100 пмоль праймеров А и F. Цикл, состоящий из 94°С в течение 30 сек, 55°С в течение 30 сек, 72°С в течение 1 мин, повторяют 35 раз, после чего реакционную смесь нагревают при 72 °С в течение 5 мин.
Фрагменты ДНК, полученные в ходе второй ПЦР, очищают в 1,5%-ном геле легкоплавкой агарозы (Sigma), расщепляют ферментами EcoRI и NotI и клонируют в вектор рСНО1 и в вектор pCOS1 (Japanese Patent Application No. 8-255196). Экспрессионный вектор pCHO1 - это вектор, полученный делецией гена антитела из DHFR-ΔE-rvH-PM1-f (см. WO 92/19759) в результате расщепления SmaI и EcoRI и присоединением адаптора EcoRI-NotI-BamHI Adaptor (TAKARA SHUZO). После определения последовательности ДНК плазмиды, содержащие фрагмент ДНК, кодирующий правильную аминокислотную последовательность реконструированного одноцепочечного Fv 12B5 обозначают как pCHO-sc12B5 и pCOS-sc12B5. Нуклеотидная последовательность и аминокислотная последовательность реконструированогого одноцепочечного Fv 12B5, включенные в плазмиды pCHO-sc12B5 и pCOS-sc12B5, представлены в SEQ ID NO.:84.
7.4 Экспрессия антитела 12B5 (IgG, Fab) и одноцепочечного Fv полипептида животной клеткой
Антитело 12B5 (IgG, Fab) и одноцепочный Fv, происходящий из антитела 12B5, экспрессируют с использованием клеток COS7 или СНО.
Осуществляется временная экспрессия с использованием клеток COS7 следующим образом. Методом электропорации с использованием аппарата Gene Pulser (BioRad) осуществляют трансфекцию. Для экспрессии антитела 12B5 (IgG) добавляют по 10 мкг каждого из вышеупомянутых экспрессионных векторов, HEF-12B5H-gγl и HEF-12B5L-gκ, для экспрессии фрагмента 12B5Fab добавляют по 10 мкг pFd-12B5H и HEF-12B5L-gκ и для экспрессии одноцепочечного Fv 10 мкг pCOS-sc12B5 добавляют к клеткам COS-7 (1×107 клеток/мл), суспендированных в 0,8 мл PBS. Смесь, помещенную в кювету, обрабатывают импульсом при 1,5 кВ, при электрической емкости 25 мкФ. После 10 мин восстановления при комнатной температуре электропорированные клетки переносят в культуральную среду ДМЕМ (GIBCO BRL), содержащую 10%-ную фетальную бычью сыворотку, и культивируют. После культивирования в течение ночи клетки отмывают один раз раствором PBS, добавляют к бессывороточной среде CHO-S-SFM II и культивируют в течение 2 дней. Центрифугированием удаляют из культуральной среды клеточный дебрис и фильтруют через фильтр с размером пор 0,22 мкм для получения культурального супернатанта.
Для получения линии клеток СНО со стабильной экспрессией одноцепочечного Fv (полипептида), происходящего из антитела 12B5, экспрессионный вектор pCHO-sc12B5 вводят в клетки СНО следующим образом.
Экспрессионный вектор вводят в клетки СНО методом электропорации при помощи аппарата Gene Pulser (BioRad). Линеаризированную ДНК (100 мкг), полученную расщеплением рестриктазой Pvul, и клетки СНО (1×107 мл), суспендированные 0,8 мл PBS смешивают в кювете, дают постоять на льду в течение 10 мин и обрабатывают импульсом при 1,5 кВ, при электрической емкости 25 мкФ. После восстановления в течение 10 мин при комнатной температуре электропорированные клетки добавляют к среде CHO-S-SFM II (GIBCO BRL), содержащей10%-ную фетальную бычью сыворотку, и культивируют. После культивации в течение 2 дней культивацию продолжают в среде CHO-S-SFM II (GIBCO BRL), содержащей 5 нМ метотриксат (SIGMA) и 10%-ную фетальную бычью сыворотку. Из полученных таким образом клонов отбирают клон с высокой степенью экспрессии в качестве клеточной линии-продуцента одноцепочечного Fv 12B5. После культивации в бессывороточной среде CHO-S-SFM II (GIBCO BRL), содержащей 5 нМ метотриксат (SIGMA), получают культурный супернатант путем отделения клеточного дебриса центрифугированием.
7.5 Очистка одноцепочечного Fv, происходящего из 12B5, продуцируемого клетками СНО
Культуральный супернатант клеточной линии СНО, экспрессирующей одноцепочечный Fv 12B5, полученный в 7.4, очищают с помощью колонки с анти-FLAG антителом и гель-фильтрационной колонки.
(1) Колонка с анти-FLAG антителом
Культуральный супернатант добавляют к гемо anti-FLAG M2 Affinity Gel (SIGMA), уравновешенному PBS. После отмывания колонки тем же буфером элюируют белки, адсорбированные на колонке, с помощью 0,1 М глицин-HCl буфера (рН 3,5). Элюированные фракции сразу же нейтрализуют добавлением 1 М Tris-HCl буфера (рН 8,0). Элюированные фракции анализируют методом SDS-PAGE и концентрируют при помощи Centricon-10 (MILLIPORE) ту фракцию, в которой подтвердилось наличие одноцепочечного Fv.
(2) Гель-фильтрация
Концентрированный раствор, полученный в (1), наносят на колонку Superdex200 (10×300 мм, AMERSHAM PHARMACIA), уравновешенную PBS, содержащим 0,01%-ный Tween 20. Продукт sc12B5 элюируется двумя пиками (А, В) (см. Фиг.48). Фракции А и В анализируют, используя 14%-SDS-полиакриламидный гель. Электрофорез образца осуществляют по методу Лэммли (Laemmli) в присутствии и в отсутствие восстанавливающего агента и окрашивают с помощью Кумасси Бриллиантовым Голубым после электрофореза. Как показано на Фиг.49, фракции А и В, независимо от наличия или отсутствия восстанавливающего агента, дают единственную полосу с кажущимся молекулярным весом около 31 кДа. При анализе фракций А и В методом гель-фильтрации с использованием Superdex200 PC 3,2/30 (3,2×300 мм, AMERSHAM PHARMACIA) фракция А дает продукт, элюируемый с кажущимся молекулярным весом около 44 кДа, а фракция В с около 22 кДа (см. Фиг.50а и 50b). Как показывают результаты, фракция А является димером одноцепочечного Fv sc12B5 с нековаленетной связью, а В является мономером.
7.6 Измерение ТРО-подобной агонистической активности различных одноцепочечных Fv
ТРО-подобную активность одноцепочечного анти-MPL антитела оценивают, измеряя пролиферативную активность в отношении Ba/F3 клеток (BaF/mpl), экспрессирующих человеческий ТРО-рецептор (MPL). После двукратного отмывания BaF/Mpl клеток культуральной средой RPM11640 (GIBCO), содержащей 10%-ную фетальную бычью сыворотку (GIBCO), клетки суспендируют в культуральной среде при плотности клеток 5×105 клеток/мл. Одноцепочечное анти-MPL антитело и человеческий ТРО (R&D Systems) разводят соответственно культуральной средой. 50 мкл клеточной суспензии и 50 мкл разведенного антитела или человеческого ТРО вносят в лунки 96-луночного микропланшета (плоскодонного) (Falcon) и культивируют в CO2-инкубаторе (концентрация CO2 5%) в течение 24 часов. После инкубации добавляют 10 мкл реагента WST-8 (реагент для определения числа живых клеток SF: Nacalai Tesque) и сразу же измеряют поглощение при длине волны измерения 450 нм и опорной длине волны 620 нм с помощью флуоресцентно-абсорбционного фотометра SPECTRA Fluor (TECAN). После инкубирования в СО2-инкубаторе (концентрация CO2 5%) в течение 2 часов снова измеряют поглощение при длине волны измерения 450 нм и опорной длине волны 620 нм с помощью SPECTRA Fluor. Поскольку реагент WST-8 развивает окраску при длине волны 450 нм в зависимости от числа живых клеток, то пролиферативную активность BaF/Mpl оценивают по изменению поглощения за 2 часа с помощью ED50, который рассчитывают следующим образом. На графике пролиферации, по ординате которого откладывают поглощение, а по абсциссе - концентрацию антитела, поглощение на плато принимают за 100% скорости реакции. Пользуясь формулой линейного приближения на основании отложенных на графике значений, близких к 50% скорости реакции, рассчитывают концентрацию антитела, соответствующую 50% скорости реакции, и принимают ее за ED50.
Результаты определения агонистической активности в отношении MPL с использованием культуральных супернатантов клеток COS-7, экспрессирующих различные молекулы антитела 12В5, отражены на Фиг.51, где показано, что 12B5IgG, имеющий бивалентный антигенсвязывающий центр, увеличивает поглощение зависимым от концентрации образом и обладает ТРО-подобной агонистической активностью (ED50; 29 нМ), в то время как агонистическая активность 12B5Fab, имеющего моновалентный антигенсвязывающий центр, оказалась очень слабой (ED50; 34724 нМ). В противоположность этому одноцепочечный Fv (sc12B5), имеющий, подобно Fab, моновалентный антигенсвязывающий центр, проявляет высокую агонистическую активность с ED50, равным 75 нМ. Однако, как известно, вариабельные области Н-цепи и L-цепи одноцепочечного Fv связаны нековалентной связью и, следовательно, в растворе каждая вариабельная область диссоциирует и может связываться с вариабельной областью другой молекулы с образованием мультимеров, например димеров. При измерении молекулярного веса sc12B5, очищенного гель-фильтрацией, было установлено, что в нем присутствуют молекулы, которые были признаны мономерами и димерами (см. Фиг.48). Затем изолируют мономер sc12B5 и димер sc12B5 (см. Фиг.50) и измеряют их агонистическую активность в отношении MPL. Как показано на Фиг.51 и 52, ED50 мономера sc12B5 составляет 4438,7 нМ, что подтверждает, что его агонистическая активность оказывается более низкой по сравнению с результатами, полученными с культуральным супернатантом клеток COS-7. И наоборот, одноцепочечный Fv (димер sc12B5), имеющий бивалентный антигенсвязывающий центр, проявляет агонистическую активность (ED50; 10,1 нМ) в 400 раз более высокую, чем у моновалентного sc12B5. Помимо того, бивалентный одноцепочечный Fv проявляет агонистическую активность, эквивалентную или более высокую, чем агонистическая активность человеческого ТРО или 12B5IgG.
Пример 8
Конструирование гена, кодирующего вариабельную область человеческого антитела 12Е10 к человеческому MPL
ДНК, кодирующую вариабельную область моноклонального антитела 12Е10 человека к человеческому MPL, конструируют следующим образом.
8.1 Конструирование гена, кодирующего V-область Н-цепи 12Е10
Нуклеотидную последовательность SEQ ID NO.:86 конструируют на основе аминокислотной последовательности, описанной в WO 99/10494 (SEQ ID NO.:85), как ген, кодирующий V-область Н-цепи человеческого антитела 12Е10, связывающегося с человеческим MPL. Полноразмерную нуклеотидную последовательность получают присоединением к ее 5'-концу лидерной последовательности (SEQ ID NO.:87), происходящей из гена человеческого антитела (GenBank accession NO.AF062252). Сконструированную нуклеотидную последовательность разбивают на 4 олигонуклеотида, имеющих перекрывающиеся последовательности длиной 15 п.о. каждая (12E10VH1, 12E10VH2, 12E10VH3, 12E10VH4). 12E10VH1 (SEQ ID NO.:88) и 12E10VH3 (SEQ ID NO.:90) синтезируют в смысловой ориентации, а 12E10VH2 (SEQ ID NO.:89) и 12E10VH4 (SEQ ID NO.:91) в антисмысловой ориентации соответственно. После состыковки каждого из синтезированных олигонуклеотидов в соответствии с комплементарностью добавляют внешние праймеры (12E10VHS и 12E10VHA) для амплификации полноразмерного гена. 12E10VHS (SEQ ID NO.:92) в качестве прямого праймера конструируют так, чтобы он гибридизовался с 5'-концом лидерной последовательности и содержал сайт узнавания рестриктазы Hind III и последовательность Козака, а 12Е 10VHA (SEQ ID NO.:93) в качестве обратного праймера конструируют так, чтобы он гибридизовался с нуклеотидной последовательностью, кодирующей С-конец V-области Н-цепи и содержал донорскую последовательность сплайсинга и сайт узнавания рестриктазы BamHI соответственно.
100 мкл ПЦР-раствора, содержащего 10 мкл 10-кратного буфера ПЦР GOLD Buffer II, 1,5 мМ MgCl2, 0,08 мМ dNTP (dATP, dGTP, dCTP, dTTP), 5 единиц ДНК-полимеразы AmpliTaq Gold (все от PERKJN ELMER) и по 2,5 пмоль каждого из синтезированных олигонуклеотидов (12E10VH-1 до - 4), нагревают при начальной температуре 94°С в течение 9 мин, затем при 94°С 2 мин, при 55°С 2 мин и при 72°С 2 мин. После двукратного повторения цикла добавляют по 100 пмоль каждого из внешних праймеров 12E10VHS и 12E10VHA. Смесь подвергают циклу, включающему 94°С в течение 30 сек, 55°С в течение 30 сек, 72°С в течение 1 мин, который повторяют 35 раз, после чего дополнительно нагревают при 72°С в течение 5 мин.
ПЦР-продукт очищают в 1,5%-ном геле с легкоплавкой агарозой (Sigma), расщепляют ферментами - рестриктазами BamHI и Hind III - и клонируют в экспрессионный вектор HEF-gγl для Н-цепи человека. После определения ДНК-последовательности плазмиду, содержащую правильную ДНК-последовательность, обозначают как HEF-12E10H-gγl.
HEF-12E10H-gγl расщепляют рестриктазами EcoRI и BamHI для того, чтобы получить ген, кодирующий 12E10VH, и затем его клонируют в экспрессионный вектор для Н-цепи Fab человека, pCOS-Fd, для получения pFd-12E10H. Экспрессионный вектор для Н-цепи Fab человека получают, амплифицируя ДНК (SEQ ID NO.:63), содержащую область интрона, находящуюся между генами, кодирующими V-область Н-цепи антитела человека и константную область, и ген, кодирующий часть константной области Н-цепи человека, методом ПЦР и встраивая ПЦР-продукт в экспрессионный вектор для животных клеток pCOS1. Константную область Н-цепи человека амплифицируют для получения гена при тех же условиях, которые были описаны выше, используя в качестве матрицы HEF-gγl, в качестве прямого праймера G1CH1-S (SEQ ID NO.:64), который конструировали так, чтобы он гибридизовался с 5'-концом последовательности интрона 1 и содержал сайты узнавания рестриктазы EcoRI и BamHI, и в качестве обратного праймера - G1CH1-A (SEQ ID NO.:65), который конструировали так, чтобы он гибридизовался с 3'-концом ДНК СН1-домена константной области Н-цепи человека и содержал последовательность, кодирующую часть шарнирной области, два стоп-кодона и сайт узнавания рестриктазы Bgl II.
Нуклеотидная последовательность и аминокислотная последовательность реконструированной вариабельной области Н-цепи 12Е10, которые были включены в плазмиды HEF-12E10H-gγl и pFd-12E10H, приведены в SEQ ID NO.:94.
8.2 Конструирование гена, кодирующего V-область L-цепи 12Е10
Нуклеотидную последовательность SEQ ID NO.:96 конструируют на основе аминокислотной последовательности, описанной в WO 99/10494 (SEQ ID NO.:95) как ген, кодирующий V-область L-цепи человеческого антитела 12Е10, связывающегося с человеческим MPL. Далее его конструкция включает присоединение к его 5'-концу лидерной последовательности (SEQ ID NO.:97), происходящей из гена человеческого антитела (Mol. Immunol. 1992; 29: 1515-1518). Таким же образом, как описано выше, сконструированную нуклеотидную последовательность разбивают на 4 олигонуклеотида, имеющих перекрывающиеся последовательности по 15 п.о. каждая (12E10VL1, 12E10VL2, 12E10VL3, 12E10VL4), и синтезируют соответственно. 12E10VL1 (SEQ ID NO.:98) и 12E10VL3 (SEQ ID NO.:100) содержат смысловые последовательности, а 12E10VL2 (SEQ ID NO.:99) и 12E10VL4 (SEQ ID NO.:101) содержат соответственно антисмысловые последовательности. После состыковки каждого из синтезированных олигонуклеотидов в соответствии с комплементарностью, добавляют внешние праймеры (12E10VLS и 12Е10VЕА) для амплифицирования полноразмерного гена. 12E10VLS (SEQ ID NO.:102) как прямой праймер конструируют так, чтобы он гибридизовался с 5'-концом лидерной последовательности и содержал сайт узнавания рестриктазы EcoRI и последовательность Козака. 12E10VLA (SEQ ID NO.:103) в качестве обратного праймера конструируется так, чтобы он гибридизовался с нуклеотидной последовательностью, кодирующей С-конец V-области L-цепи и содержал сайт узнавания рестриктазы BlnI.
После проведения ПЦР, как описано выше, ПЦР-продукт очищают в 1,5%-ном геле легкоплавкой агарозы (Sigma), расщепляют рестриктазами EcoRI и BInI и клонируют в pUC19, содержащую ген константной области ламбда-цепи человека. После определения ДНК-последовательности плазмиду, содержащую правильную ДНК-последовательность, расщепляют EcoRI для получения гена, кодирующего V-область L-цепи 12Е10 и константную область лямбда-цепи человека, а затем встраивают в экспрессионный вектор pCOS1. Плазмиду, содержащую ген L-цепи 12Е10 (SEQ ID NO.:104), обозначают как pCOS-12E10L.
8.3 Получение реконструированного одноцепочечного Fv 12E10
Реконструированный одноцепочечный Fv антитела 12Е10 конструируют так, чтобы его структура была следующей: 12Е10VН-линкер-12Е10VL, и чтобы он содержал FLAG-последовательность (SEQ ID NO.:105) на С-конце для облегчения детекции и очистки. Реконструированные одноцепочечные Fv 12E10 (sc12E10 и db12E10) конструируют, используя линкерную последовательность, состоящую из 15 аминокислот, представленных (Gly4Ser)3, или из 5 аминокислот, представленных (Gly4Ser)1.
(1) Получение реконструированного одноцепочечного Fv 12E10 с использованием линкерной последовательности, состоящей из 5 аминокислот.
Ген, кодирующий реконструированный одноцепочечный Fv 12E10, содержащий линкерную последовательность, состоящую из 5 аминокислот, конструируют путем введения нуклеотидной последовательности для линкера (Gly4Ser) в 3'-концевое положение гена, кодирующего V-область Н-цепи 12E10, и в 5'-концевое положение гена, кодирующего V-область L-цепи 12E10, амплификации полученного таким образом соответствующего гена методом ПЦР и состыковки амплифицированных генов. Для получения реконструированного одноцепочечного Fv 12E10 используют четыре ПЦР праймера (А-D). Праймеры А и С имеют смысловые последовательности, а праймеры В и D антисмысловые последовательности.
В качестве прямого праймера для V-области Н-цепи используют 12E10S (праймер A, SEQ ID NO.:106). Обратный праймер DB2 (праймер В, SEQ ID NO.:107) для V-области Н-цепи конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области Н-цепи, и содержал нуклеотидную последовательность, кодирующую линкер (Gly4Ser)1, и нуклеотидную последовательность, кодирующую N-конец V-области L-цепи.
Прямой праймер DB1 (праймер С, SEQ ID NO.:108) для V-области L-цепи конструируют так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области L-цепи и содержал нуклеотидную последовательность, кодирующую линкер (Gly4Ser)1, и нуклеотидную последовательность, кодирующую С-конец V-области Н-цепи. Обратный праймер 12E10FA (праймер D, SEQ ID NO.:109) для V-области L-цепи конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал нуклеотидную последовательность, кодирующую FLAG и сайт узнавания рестриктазы NotI.
На первом этапе ПЦР проводят две реакции А-В и C-D и два ПЦР-продукта, полученных на первом этапе ПЦР, состыковывают в соответствии с комплементарностью. После добавления праймеров А и D амплифицируют полноразмерную ДНК, кодирующую реконструированный одноцепочечный Fv 12E10, содержащий линкер, состоящий из 5 аминокислот (второй этап ПЦР). На первом этапе ПЦР в качестве матриц используют соответственно плазмиду HEF-12E10H-gγl (см. Пример 8.1), кодирующую V-область Н-цепи 12E10, и pCOS-12E10L (см. Пример 8.2), кодирующую реконструированную V-область L-цепи 12E10.
50 мкл ПЦР-раствора первого этапа содержали 5 мкл 10-кратного буфера ПЦР Gold Buffer II, 1,5 мМ MgCl2, 0,08 мМ каждого из dNTP, 5 единиц ДНК-полимеразы AmpliTaq Gold (все от PERKIN ELMER), по 100 пмоль каждого праймера и по 100 нг каждой матричной ДНК. ПЦР-раствор нагревают при начальной температуре 94°С в течение 9 минут, затем при 94°С в течение 30 сек, при 55°С в течение 30 сек, при 72°С в течение 1 мин. После повторения цикла 35 разреакционную смесь дополнительно нагревают при 72°С в течение 5 мин.
Продукты ПЦР-реакции А-В (429 п.о.) и C-D (395 п.о.) состыковывают во второй ПЦР. ПЦР-смесь 2 этапа (98 мкл), содержащую по 1 мкл каждого из ПЦР-продуктов А-В и C-D первого этапа в качестве матриц, по 100 пмоль каждого из праймеров, 10 мкл 10-кратного буфера ПЦР Gold Buffer II, 1,5 мМ MgCl2, 0,08 мМ каждого dNTP, 5 единиц ДНК-полимеразы AmpliTaq Gold (все от PERKIN ELMER), подвергают реакции в тех же условиях, что были упомянуты выше.
Фрагмент ДНК (795 п.о.), полученный на втором этапе ПЦР, очищают в 1,5%-ном геле легкоплавкой агарозы, расщепляют рестриктазами EcoRI и NotI и клонируют в вектор рСНО1 или в вектор pCOS1. Экспрессионный вектор pCHO1 - это вектор, полученный делецией гена антитела из DHFR-ΔE-RVH-PM1-f (см. WO 92/19759) в результате расщепления рестриктазами EcoRI и SmaI и присоединением адаптора EcoRI-NotI-BamHI (TAKARA SHUZO). После определения ДНК-последовательности плазмиды, содержащие ДНК-фрагмент, кодирующий правильную аминокислотную последовательность реконструированного одноцепочечного Fv 12B5, обозначают как pCHO-db12E10 и pCOS-db12E10. Нуклеотидная последовательность и аминокислотная последовательность реконструированного одноцепочечного Fv 12E10, включенные в плазмиды pCHO-db12E10 и pCOS-db12E10, приведены в SEQ ID NO.:110.
(2) Получение реконструированного одноцепочечного Fv 12E10 с использованием линкерной последовательности, состоящей из 15 аминокислот
Ген, кодирующий реконструированный одноцепочечный Fv 12E10, который содержит линкерную последовательность, состоящую из 15 аминокислот, конструируют путем введения нуклеотидной последовательности линкера (Gly4Ser)3 в 3'-концевое положение гена, кодирующего V-область Н-цепи 12E10, и в 5'-концевое положение гена, кодирующего V-область L-цепи 12E10, амплификации полученных таким образом генов методом ПЦР и состыковки амплифицированных генов. Для получения реконструированного одноцепочечного Fv 12E10 используют 4 ПЦР-праймера (A-D). Праймеры А и С имеют смысловые последовательности, а праймеры В и D имеют антисмысловые последовательности.
Прямым праймером для V-области Н-цепи является 12E10S (Праймер A, SEQ ID NO.:106). Обратный праймер sc4.3 (Праймер В, SEQ ID NO.111) для V-области Н-цепи конструируют так, чтобы он гибризовался с ДНК, кодирующей С-конец V-области Н-цепи и содержал нуклеотидную последовательность, кодирующую линкер (Gly4Ser)3, и нуклеотидную последовательность, кодирующую N-конец V-области L-цепи.
Прямой праймер scl.3 (Праймер С, SEQ ID NO.:112) конструируют так, чтобы он гибридизовался с ДНК, кодирующей N-конец V-области L-цепи, и содержал нуклеотидную последовательность, кодирующую линкер (Gly4Ser)3, и нуклеотидную последовательность, кодирующую С-конец V-области Н-цепи. Обратный праймер 12E10FA (Праймер D, SEQ ID NO.:109) конструируют так, чтобы он гибридизовался с ДНК, кодирующей С-конец V-области L-цепи, и содержал нуклеотидную последовательность, кодирующую FLAG сайт узнавания рестриктазы и NotI.
На первом этапе ПЦР осуществляют две реакции А-В и C-D, и два ПЦР-продукта, полученные на первом этапе ПЦР, состыковывают в соответствии с комплементарностью. После добавления праймеров А и D амплифицируют (вторая ПЦР) полноразмерную ДНК, кодирующую реконструированный одноцепочечный Fv 12E10, содержащий линкер, состоящий из 15 аминокислот. На первом этапе ПЦР в качестве матрицы используют плазмиду pCOS-db12E10 (см. Пример 8.3 (1)), кодирующую реконструированный одноцепочечный Fv 12E10.
В 50 мкл раствора для первого этапа ПЦР содержится 5 мкл 10-кратного буфера ExTaq, 0,4 мМ каждого из dNTP, 2,5 единицы ДНК-полимеразы TaKaRa ExTaq (от TAKARA), и по 100 пмоль каждого праймера и по 10 нг каждой матричной ДНК. ПЦР-раствор нагревают при начальной температуре 94°С в течение 30 сек, затем при 94°С в течение 15 сек, при 72°С в течение 2 мин и данный цикл повторяют 5 раз. После 28-кратного повторения цикла, состоящего из нагревания при 94°С в течение 15 сек и при 70°С в течение 2 мин, реакционную смесь дополнительно нагревают при 72°С в течение 5 мин.
ПЦР-продукты А-В (477 п.о.) и C-D (447 п.о.) состыковывают во второй ПЦР. Реакционную смесь для второго этапа ПЦР (98 мкл), содержащую 1 мкл каждого из продуктов А-В и C-D первой ПЦР в качестве матриц, по 100 пмоль каждого из праймеров А и D, 5 мкл 10-кратного буфера ExTaq, 0,4 мМ каждого dNTP, 2,5 единицы ДНК полимеразы TaKaRa ExTaq (TAKARA) подвергают реакции в тех же условиях, что описаны выше.
ДНК-фрагмент (825 п.о.), полученный во второй ПЦР, очищают в 1,0%-ном геле легкоплавкой агарозы, расщепляют рестриктазами EcoRI и NotI. Полученный таким образом ДНК-фрагмент клонируют в вектор рСНО1 или в вектор pCOS1. После определения последовательности ДНК плазмиды, содержащие ДНК-фрагмент, кодирующий правильную аминокислотную последовательность реконструированного одноцепочечного Fv 12E10, обозначают как pCHO-sc12E10 и pCOS-sc12E10. Нуклеотидная последовательность и аминокислотная последовательность реконструированного одноцепочечного Fv 12E10, включенные в плазмиды pCHO-sc12E10 и pCOS-sc12E10, приведены в SEQ ID NO:113.
8.4 Экспрессия антитела 12E10 (IgG, Fab) и одноцепочечного Fv-полипептида в животных клетках
Антитело 12E10 (IgG, Fab) и одноцепочечный Fv, происходящий из антитела 12E10 (линкерная последовательность из 5 аминокислот, из 15 аминокислот), экспрессируют с использованием клеток COS-7 или СНО.
Временная экспрессия с применением клеток COS-7 осуществляется следующим образом. Методом электропорации с использованием аппарата Gene Pulser II (BioRad) осуществляют трансфекцию. Для экспрессии антитела 12E10 (IgG) добавляют по 10 мкг каждого из вышеупомянутых экспрессионых векторов, HEF-12E10H-gγl и pCOS-12E10L, для экспрессии фрагмента 12E10Fab добавляют по 10 мкг pFd-12E10H и pCOS-12E10L и для экспрессии одноцепочечного Fv pCOS-sc12E10 (10 мкг) или pCOS-db 12E10 (10 мкг) добавляют к клеткам COS-7 (1×107 клеток/мл), суспендированных в 0,8 мл PBS. Смесь, помещенную в кювету, обрабатывают импульсом при 1,5 кВ, при электрической емкости 25 мкФ. После 10 мин восстановления при комнатной температуре электропорированные клетки помещают в среду DMEM (GIBCO BRL), содержащую 10%-ную фетальную бычью сыворотку, и культивируют. После культивирования в течение ночи клетки отмывают один раз раствором PBS, добавляют к бессывороточной среде CHO-S-SFM II (GIBCO BRL) и культивируют в течение 3 дней. Центрифугированием удаляют из культурального супернатанта клеточный дебрис и фильтруют через фильтр с диаметром пор 0,22 мкм.
Для получения клеточной линии СНО, стабильно экспрессирующей одноцепочечный Fv (полипептид), происходящий из антитела 12Е10, экспрессионный вектор pCHO-sc12E10 или pCHO-dsl2E10 вводят в клетки СНО следующим образом.
Каждый экспрессионный вектор вводят в клетки СНО методом электропорации при помощи аппарата Gene Pulser II (BioRad). Линеаризированную ДНК (100 мкг), полученную расщеплением рестриктазой Pvul, и клетки СНО (1×107 клеток/мл), суспендированных в виде взвеси в 0,8 мл PBS, смешивают в кювете, дают постоять на льду в течение 10 мин и обрабатывают импульсом при 1,5 кВ, при электрической емкости 25 мкФ. После восстановления в течение 10 мин при комнатной температуре электропорированные клетки добавляют к среде CHO-S-SFM II (GIBCO BRL), содержащей 10%-ную фетальную бычью сыворотку и нуклеиновую кислоту, и культивируют. После культивирования в течение 2 дней культивирование продолжают в свободной от нуклеиновой кислоты среде CHO-S-SFM II (GIBCO BRL), содержащей 10%-ную диализированную фетальную бычью сыворотку. Из полученных таким образом клонов отбирают клон с высокой степенью экспрессии в качестве клеточной линии-продуцента одноцепочечного Fv 12E10. После культивирования в бессывороточной среде CHO-S-SFM II (GIBCO BRL), культурный супернатант центрифугируют для удаления клеточного дебриса и фильтруют через фильтр с диаметром пор 0,22 мкм.
8.5 Очистка одноцепочечного Fv, происходящего из 12Е10, продуцируемого клетками СНО
Культуральные супернатанты, продуцируемые линиями клеток СНО, экспрессирующими одноцепочечные Fv 12E10 (sc12E10, db12E10), полученные в Примере 8.4, очищают с помощью колонки с анти-FLAG антителом и гель-фильтрационной колонки соответственно, для того чтобы получить очищенные одноцепочечные Fv.
(1) Очистка с помощью колонки с анти-FLAG антителом
Каждый культуральный супернатант (sc12E10, db12E10) наносили на колонку с anti-FLAG M2 Affinity gel (SIGMA), уравновешенную 50 мМ Tris-HCl буфером (рН 7,4), содержащим 150 мМ NaCl. После отмывания колонки тем же буфером, элюируют белки, адсорбированные на колонке, с помощью 100 мМ глицинового буфера (рН 3,5). Элюированные фракции сразу же нейтрализуют добавлением 1 М Tris-HCl буфера (рН 8,0) и анализируют методом SDS-PAGE. Те фракции, в которых подтвердилось наличие одноцепочечного Fv, объединяют и концентрируют в 20 раз при помощи Centricon-10 (AMICON).
(2) Гель-фильтрация
Концентрированный раствор, полученный в (1), наносят на колонку Superdex200 HR (10×300 мм, AMERSHAM PHARMACIA), уравновешенную PBS, содержащим 0,01%-ный Tween 20. Хроматограммы приведены на Фиг.53 и 54. Продукт sc12E10 элюируется двумя пиками (А, В) (см. Фиг.53). Продукт db12E10 элюируется двумя пиками (С, D) (см. Фиг.54). Фракцию каждого пика собирают, обрабатывают в присутствии и в отсутствие восстанавливающего агента и проводят электрофорез по методу Лэммли и после электрофореза окрашивают Кумасси Бриллиантовым Голубым. Как показано на Фиг.55, все фракции А, В, С и D, независимо от наличия или отсутствия восстанавливающего агента, дают единственную полосу, имеющую кажущийся молекулярный вес около 31 кДа. При анализе этих фракций методом гель-фильтрации с использованием Superdex200 HR, фракция А дает продукт, элюируемый с кажущимся молекулярным весом около 42 кДа, фракция В 20 кДа (см. Фиг.56), фракция С 69 кДа и фракция D 41 кДа (см. Фиг.57). Как показывают результаты, происходящая из sc12E10 фракция является димером одноцепочечного Fv с нековалентной связью, и фракция В - мономером одноцепочечного Fv, а происходящая из db12E10 фракция С является тримером одноцепочечного Fv с нековалентной связью и фракция D - димером одноцепочечного Fv с нековалентной связью.
8.6 Измерение ТРО-подобной агонистической активности различных одноцепочечных Fv
ТРО-подобную активность одноцепочечного антитела к mpl оценивают, измеряя пролиферативную активность в отношении клеток Ba/F3 (BaF/mpl), экспрессирующих человеческий ТРО-рецептор (MPL).
После двукратного отмывания клеток BaF/mpl средой RPMI1640 (GIBCO), содержащей 1%-ную фетальную бычью сыворотку (GIBCO), клетки суспендируют в среде при плотности клеток 5×105 клеток/мл. Одноцепочечное к MPL антитело или ТРО человека (R&D Systems) соответственно разводят в среде. 50 мкл клеточной суспензии и 50 мкл разведенного антитела или человеческого ТРО вносят в лунки 96-луночного микропланшета (с плоским дном) (Corning) и культивируют в CO2-инкубаторе (концентрация CO2 5%) в течение 24 часов. После инкубирования добавляют 10 мкл реагента WST-8 (реагент для измерения количества живых клеток SF: Nacalai Tesque) и сразу же измеряют поглощение при длине волны измерения 450 нм и опорной длине волны 655 нм при помощи абсорбционного фотометра Benchmark Plus (BioRad). После инкубирования в СО2-инкубаторе (концентрация СО2 5%) в течение двух часов снова при помощи Benchmark Plus измеряют поглощение при длине волны измерения 450 нм и опорной длине волны 655 нм. Поскольку реагент WST-8 развивает окраску при длине волны 450 нм в зависимости от числа живых клеток, то пролиферативная активность BaF/mpl оценивается на основании изменения поглощения за 2 часа.
Агонистическая активность в отношении MPL, измеренная с использованием культуральных супернатантов клеток COS-7, экспрессирующих различные молекулы антитела 12Е10, представлена на Фиг.58. Одноцепочечные Fv, имеющие линкер из 5 аминокислот (dsl2E10) и линкер из 15 аминокислот (sc12E10), увеличивали поглощение, зависимым от концентрации образом, демонстрируя ТРО-подобную агонистическую активность (ED50; 9 пМ и 51 пМ соответственно), в то время, как 12E10IgG и 12E10Fab не проявляли активности.
Как известно, Н-цепь и L-цепь одноцепочечного Fv образуют связи не только внутри молекулы, но также и между молекулами с образованием мультимеров, например димеров. Затем культуральный супернатант клеток СНО, экспрессирующих одноцепочечные Fv 12E10, подвергают гель-фильрации и исследуют на агонистическую активность в отношении MPL. Результаты показаны на Фиг.59. Димер, содержащийся в sc12E10 в малом количестве, проявил ТРО-подобную агонистическую активность (димер sc12E10, ED50; 1,9 пМ, в 5000 раз превышающую активность мономера (мономер sc12E10, ED50; >10 нМ). Эта активность выше, чем у ТРО (ED50; 27 пМ). Димер db12E10 (димер db 12E10, ED50; 2,0 пМ) продемонстрировал более высокую активность сравнительно с димером sc12E10. Тример db12E10 (ED50; 7,4 пМ), который предположительно являлся тримером на основании молекулярного веса, определенного гель-фильтрацией, обладает высокой активностью, которая оказалась ниже, чем активность димера db12E10. На основе полученных результатов можно полагать, что для активности антитела-агониста 12E10 важно, чтобы антигенсвязывающий центр был бивалентным (димер). Принимая во внимание, что IgG 12E10 не продемонстрировал никакой активности, можно предположить, что и другие факторы, помимо бивалентности, имеют значения, например, такие как расположение антигенсвязыващего центра, удаленность или угол.
Промышленное применение
Модифицированные антитела изобретения обладают агонистической активностью - способностью трансдуцировать сигнал в клетки путем образования поперечных сшивок молекул (молекулы) поверхности клетки и обладает тем преимуществом, что их способность проникновения в ткани и опухоли за счет уменьшенных в сравнении с молекулой антитела (целый IgG) молекулярных размеров высока. Данное изобретение обеспечивает модифицированные антитела со значительно более высокой агонистической активностью, чем у ТРО или исходных антител (целый IgG). В частности, даже не обладающие агонистической активностью исходные антитела могут быть преобразованы в модифицированные антитела с агонистической активностью более высокой, чем у ТРО. Таким образом, модифицированные антитела могут применяться в качестве агонистов трансдукции сигнала. Модификация молекулы антитела приводит к уменьшению побочных эффектов, вызываемых образованием межклеточных поперечных сшивок, и обеспечивает новые лекарственные средства, вызывающие только желательные последствия при поперечной сшивке молекулы или молекул клеточной поверхности. Медицинские препараты, содержащие в качестве активного компонента модифицированные антитела изобретения, могут применяться как профилактические и/или лекарственные средства при заболеваниях крови, связанных с тромбоцитами, при тромбоцитопении, вызванной химиотерапией при лечении разных видов рака или лейкемии и т.п.







































































| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННОЕ АГОНИСТИЧЕСКОЕ АНТИТЕЛО | 2001 |
|
RU2295537C2 |
| СПОСОБ И НАБОР ДЛЯ ВЫЯВЛЕНИЯ ГЕНА, КОДИРУЮЩЕГО МЕМБРАНОСВЯЗАННЫЙ БЕЛОК, ВЕКТОР (ВАРИАНТЫ) | 1999 |
|
RU2246538C2 |
| ОПУХОЛЕСПЕЦИФИЧНОЕ АНТИТЕЛО ПРОТИВ EGFR И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2730605C2 |
| Рекомбинантный Fab-scFv на основе нейтрализующего антитела против интерферона бета-1а человека и антитела против рецептора ErbB2 человека | 2019 |
|
RU2748953C1 |
| АНТИ-ГЛИПИКАН 3-АНТИТЕЛО, ИМЕЮЩЕЕ МОДИФИЦИРОВАННУЮ САХАРНУЮ ЦЕПЬ | 2005 |
|
RU2451030C2 |
| МОНОМОЛЕКУЛЯРНЫЙ ХИМЕРНЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР К РАКОВОМУ АНТИГЕНУ СА125 | 2018 |
|
RU2747095C2 |
| СПОСОБ СЕКРЕТОРНОЙ ПРОДУКЦИИ БЕЛКА | 2012 |
|
RU2588439C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ МОДИФИКАЦИИ ЗАДАННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВОЙ КИСЛОТЫ-МИШЕНИ | 2012 |
|
RU2663354C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ С УСТОЙЧИВОСТЬЮ К РАСТВОРИМОМУ СЕА | 2010 |
|
RU2573893C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ С УСТОЙЧИВОСТЬЮ К РАСТВОРИМОМУ СЕА | 2006 |
|
RU2426743C2 |

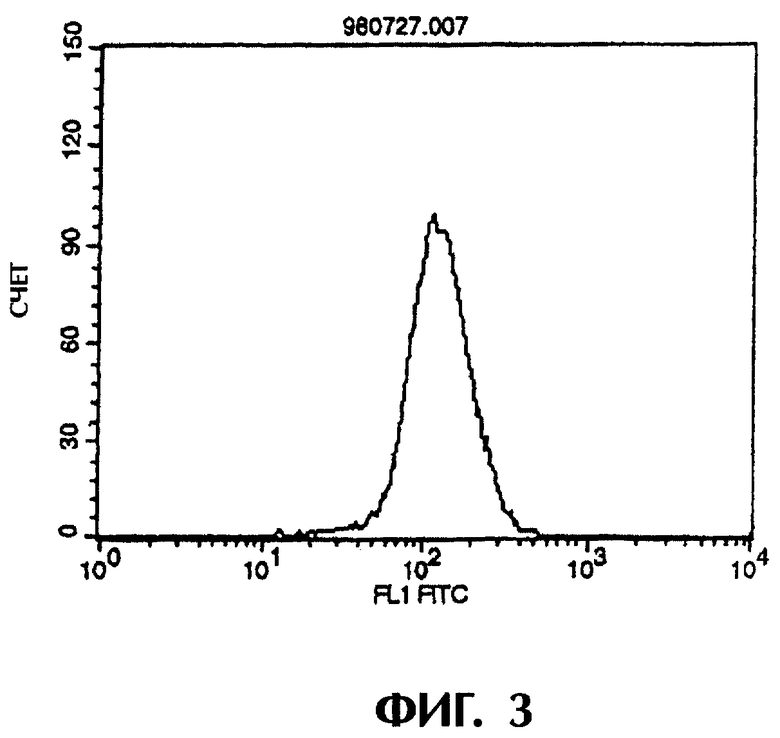
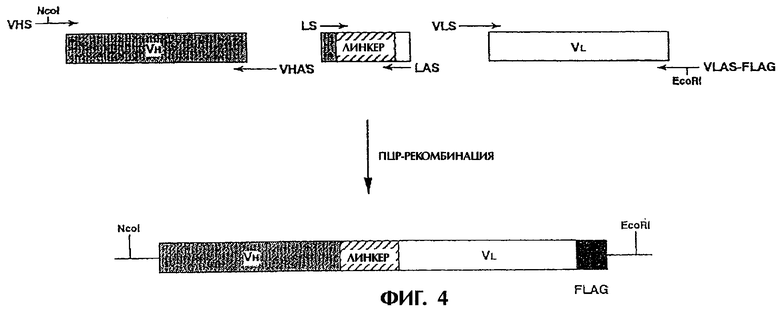
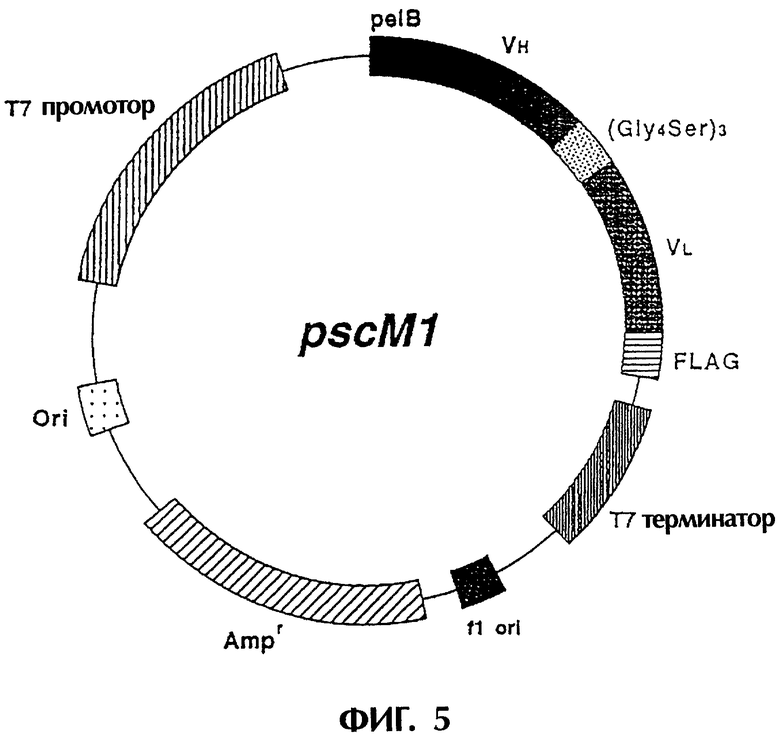
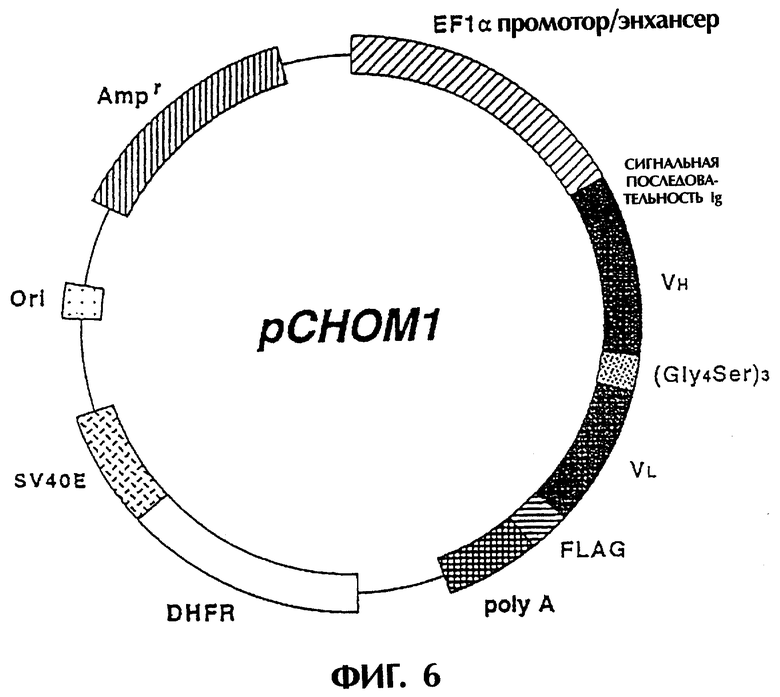
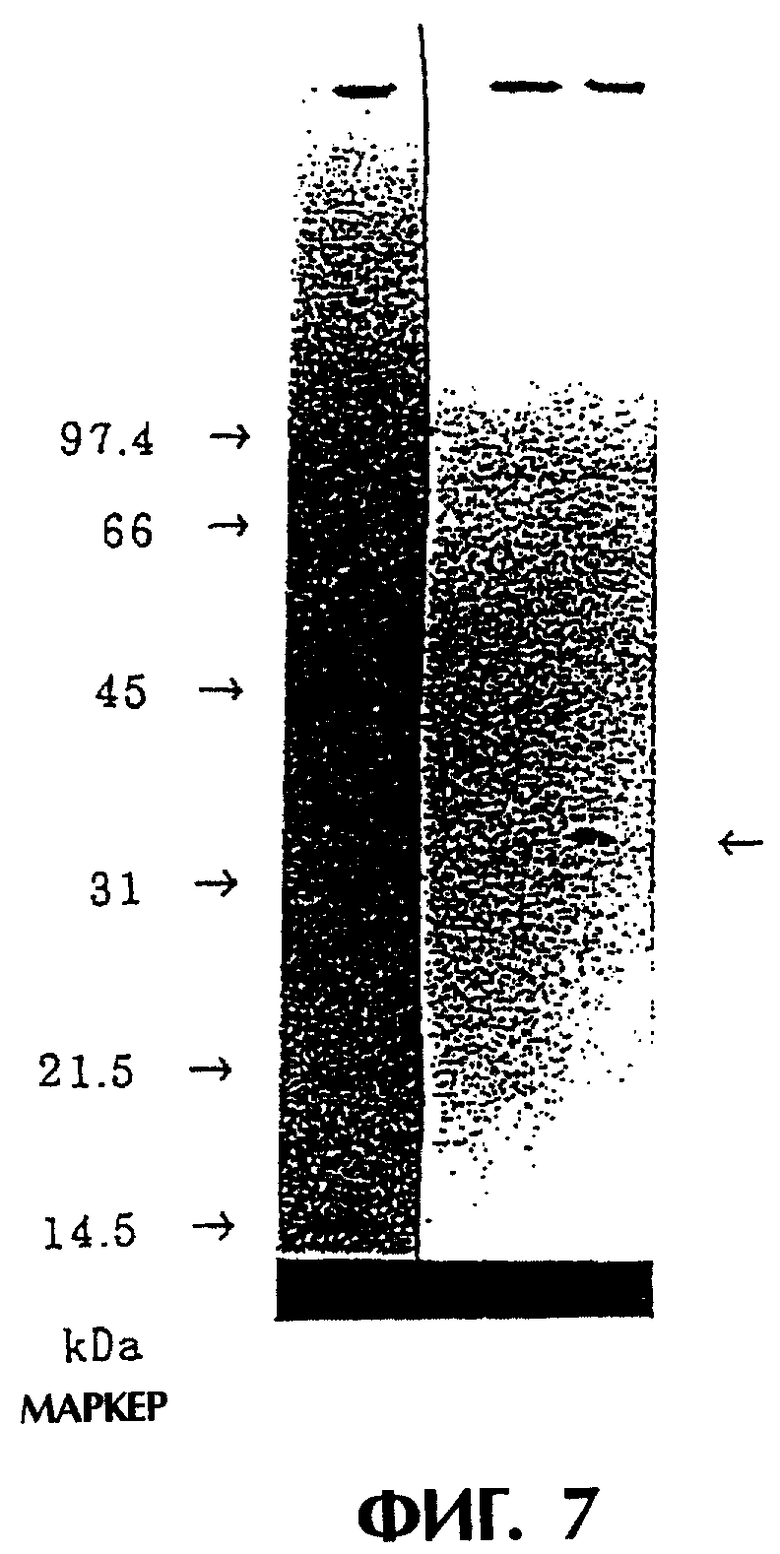
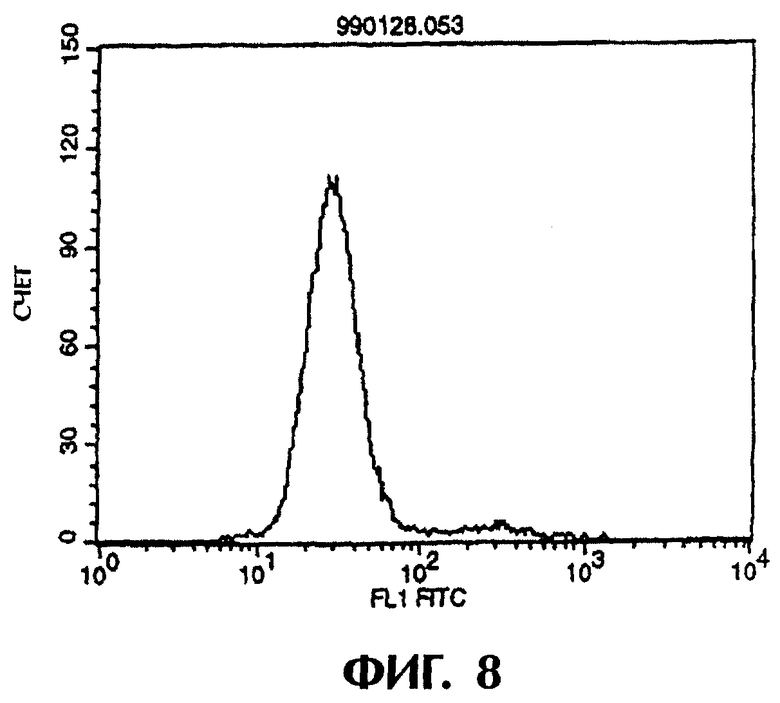

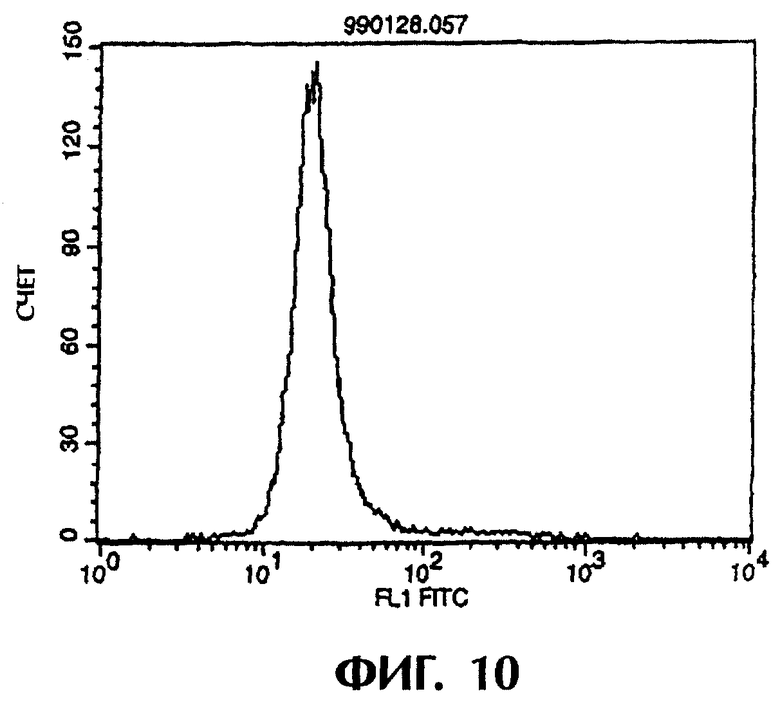
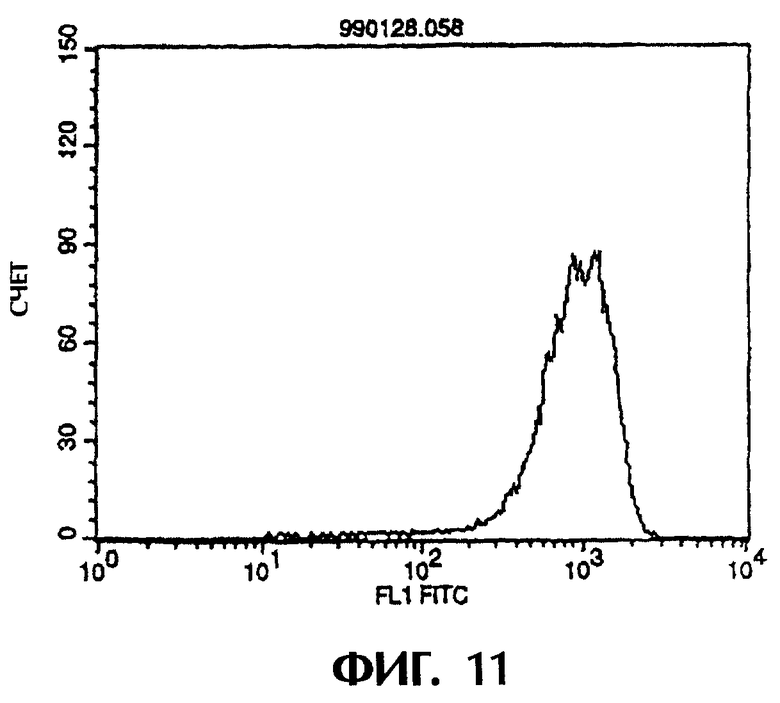
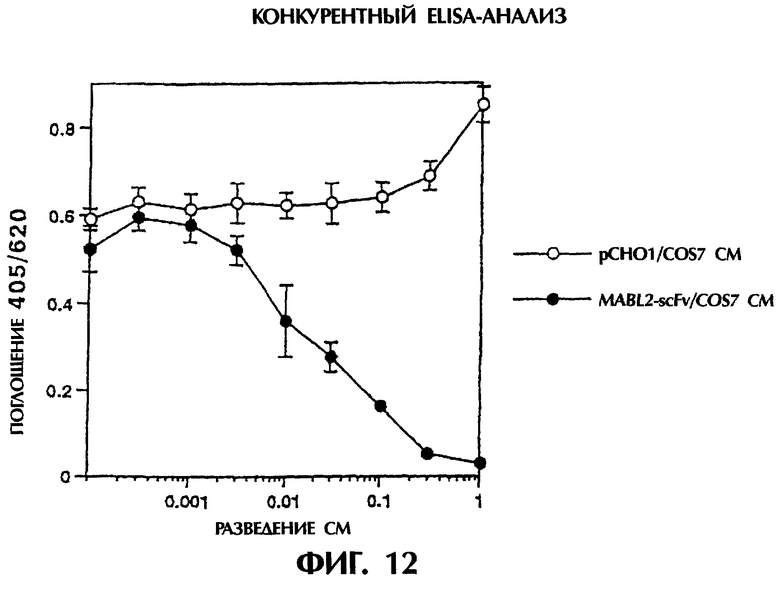
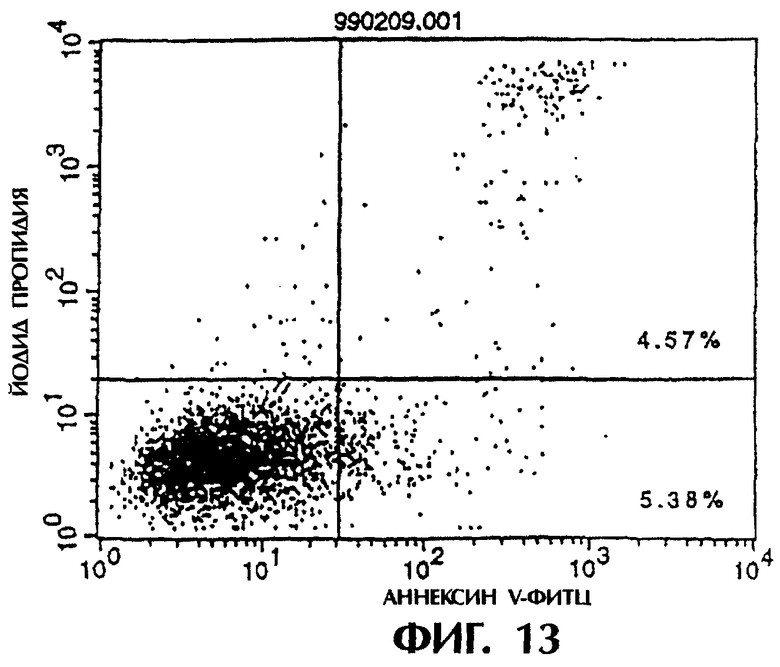
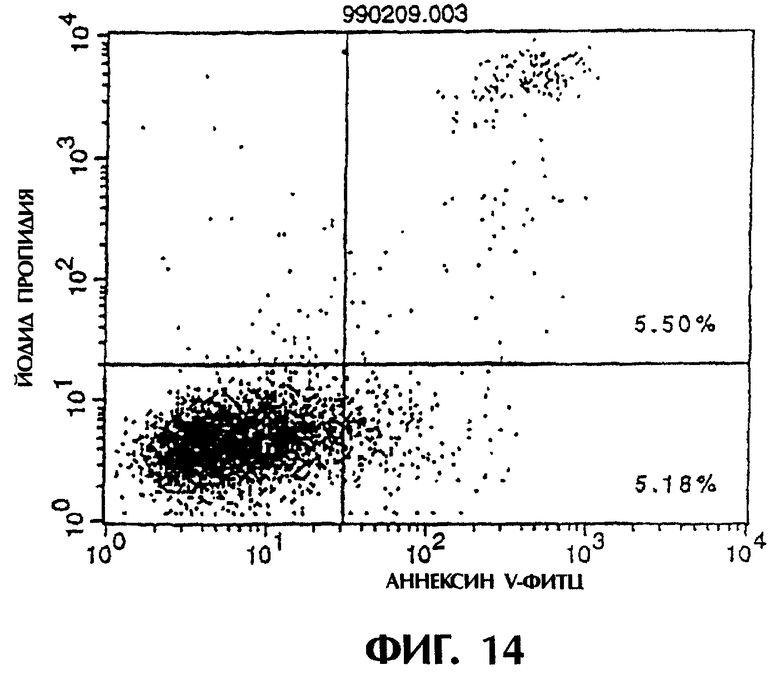

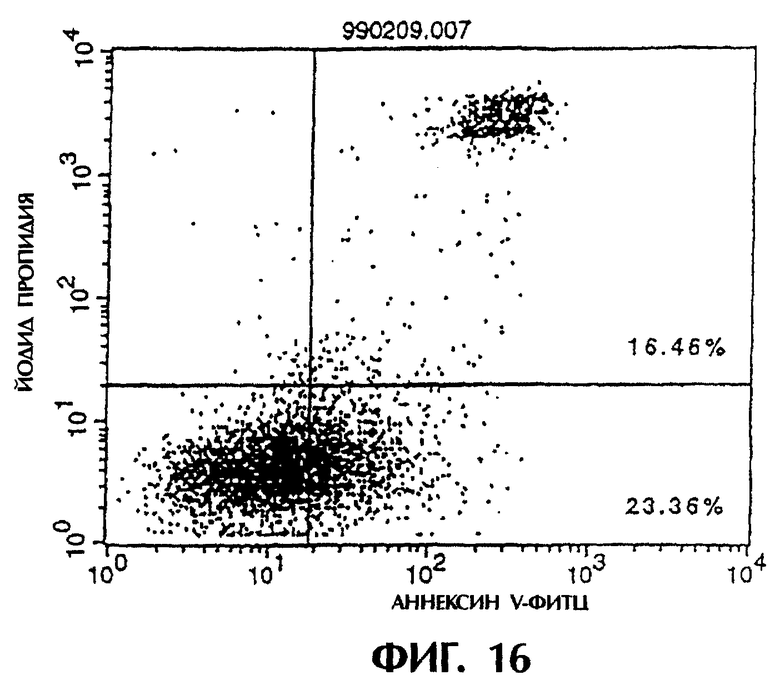
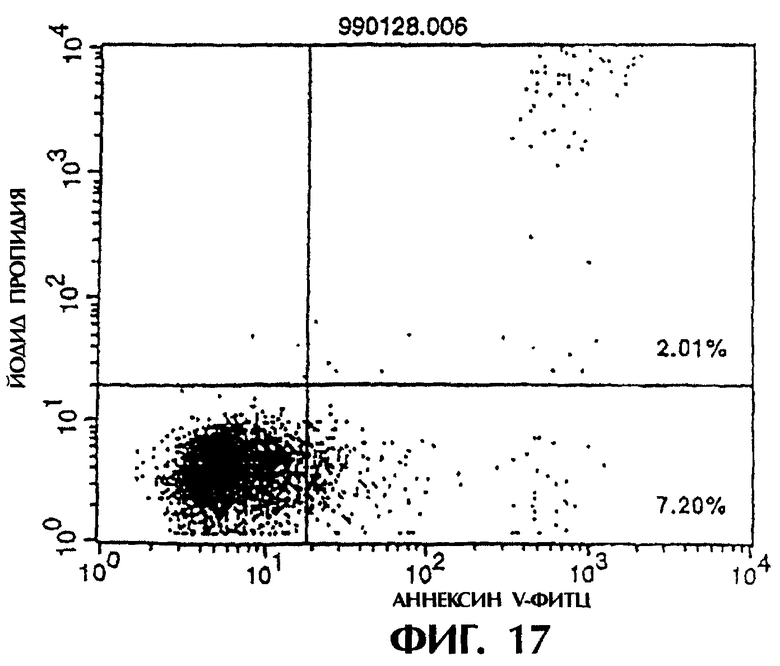
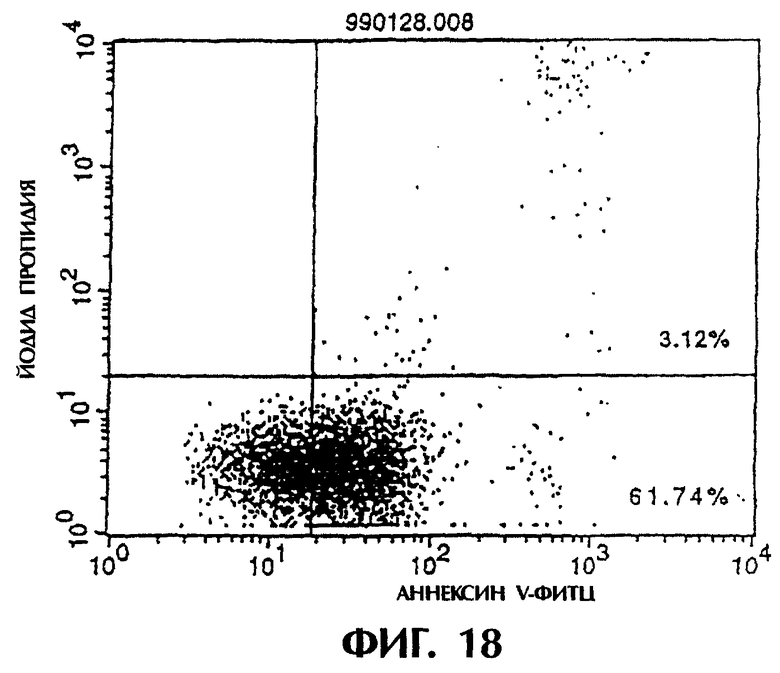
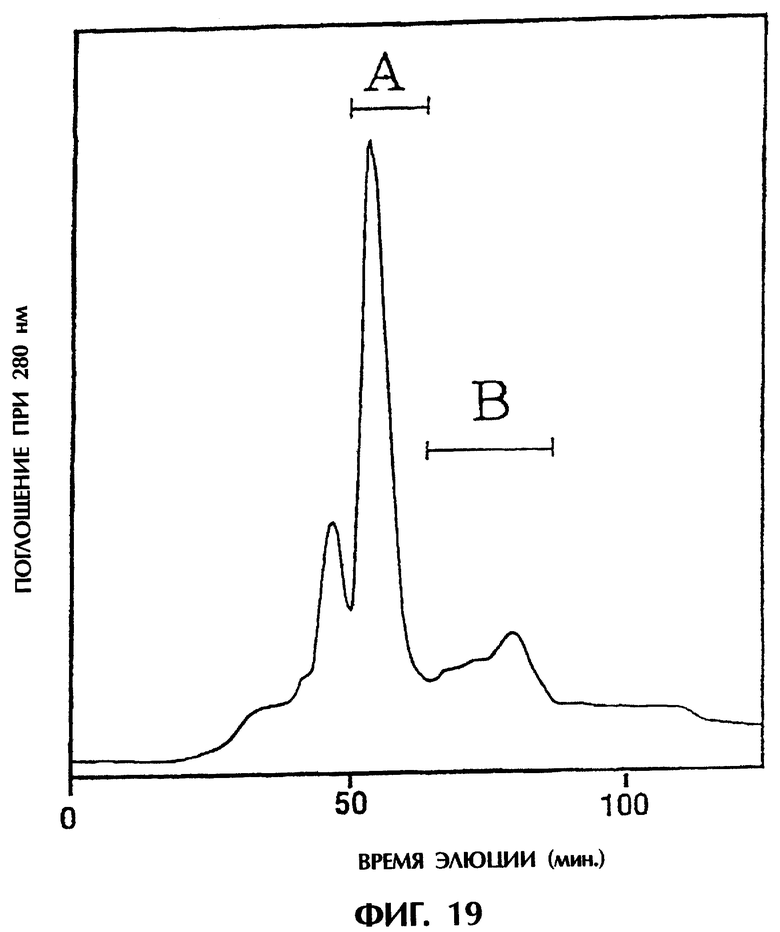
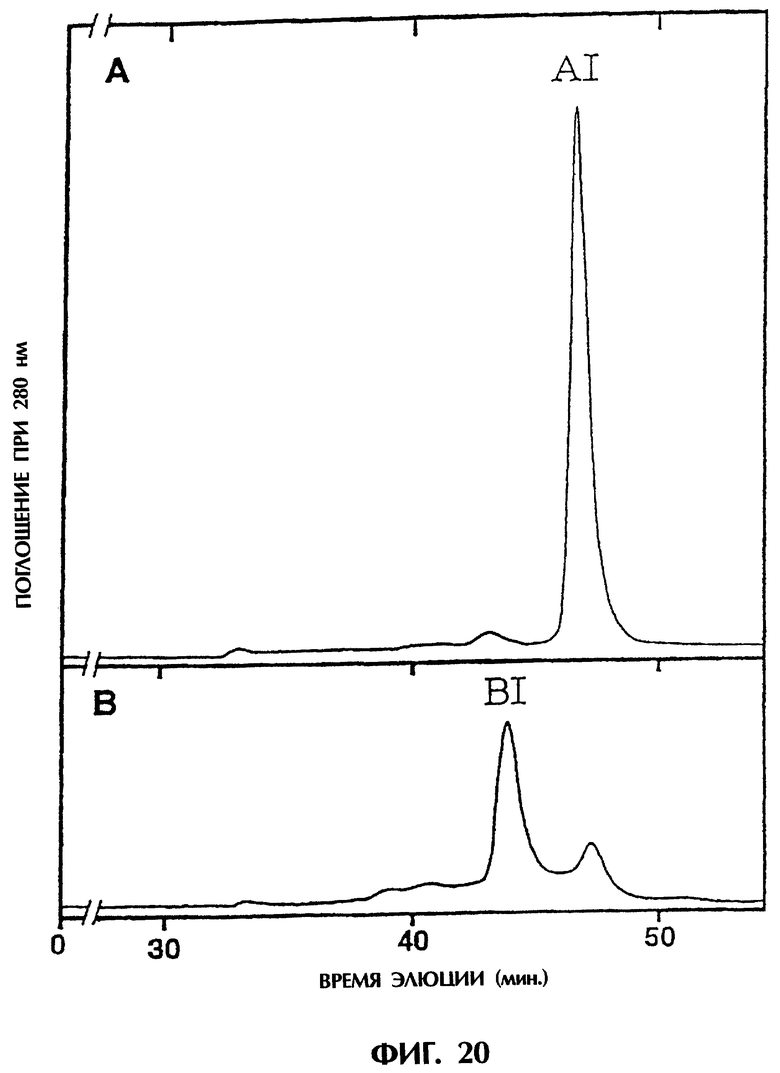
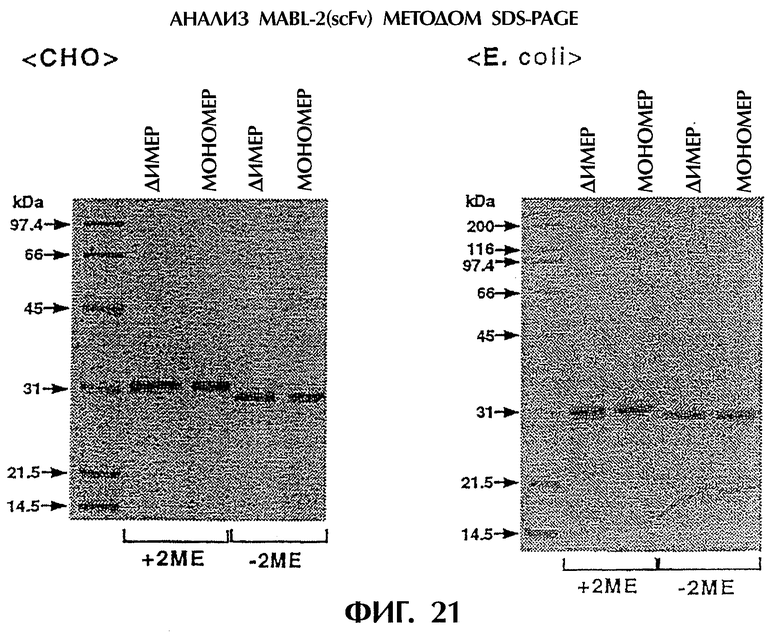
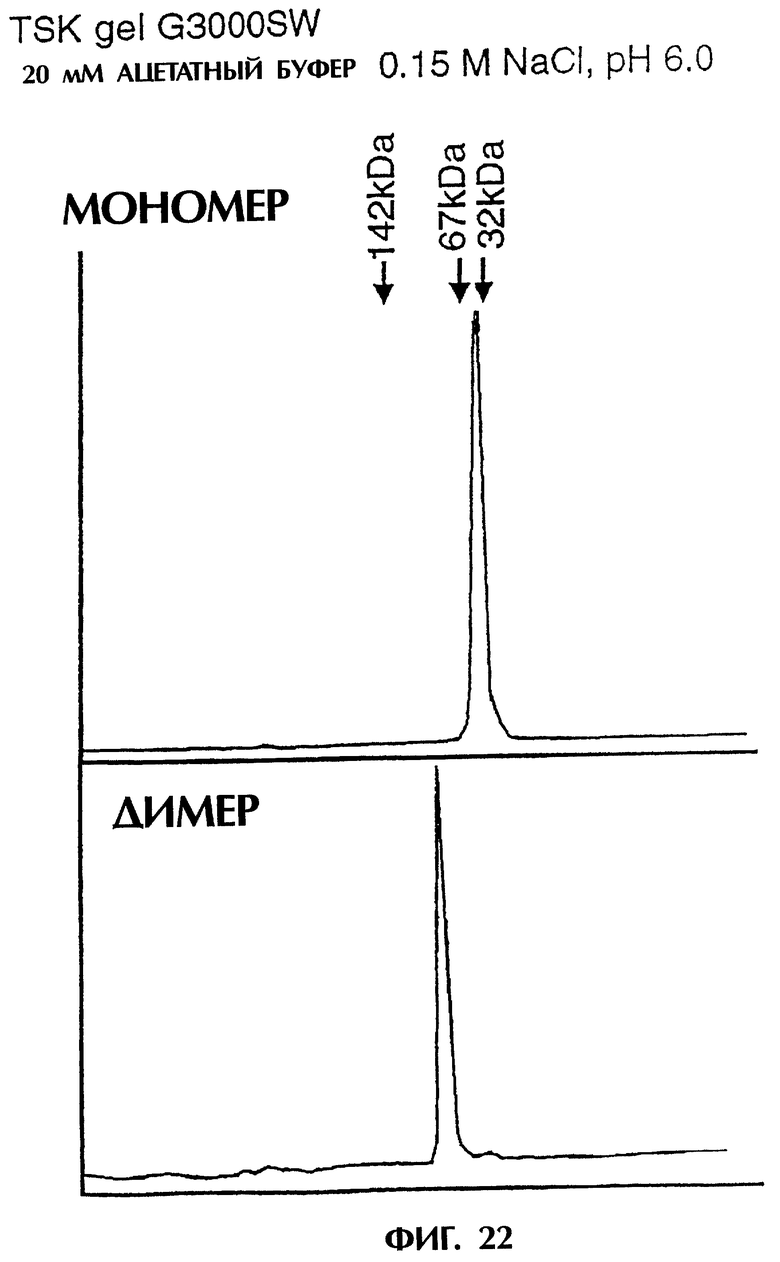
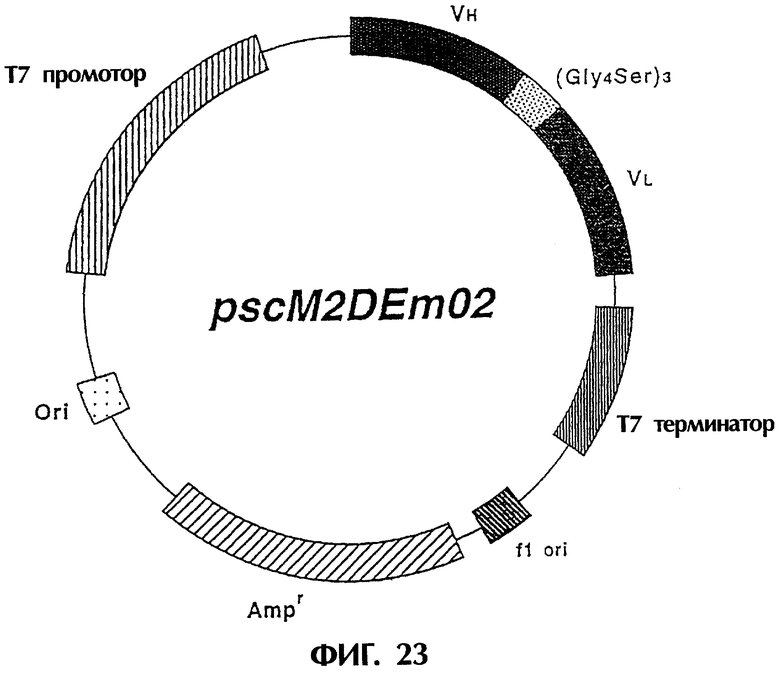
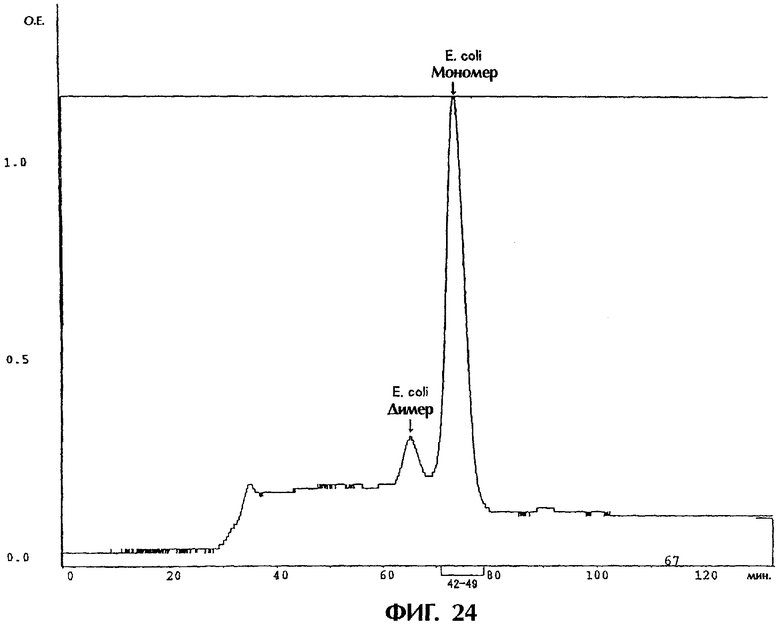
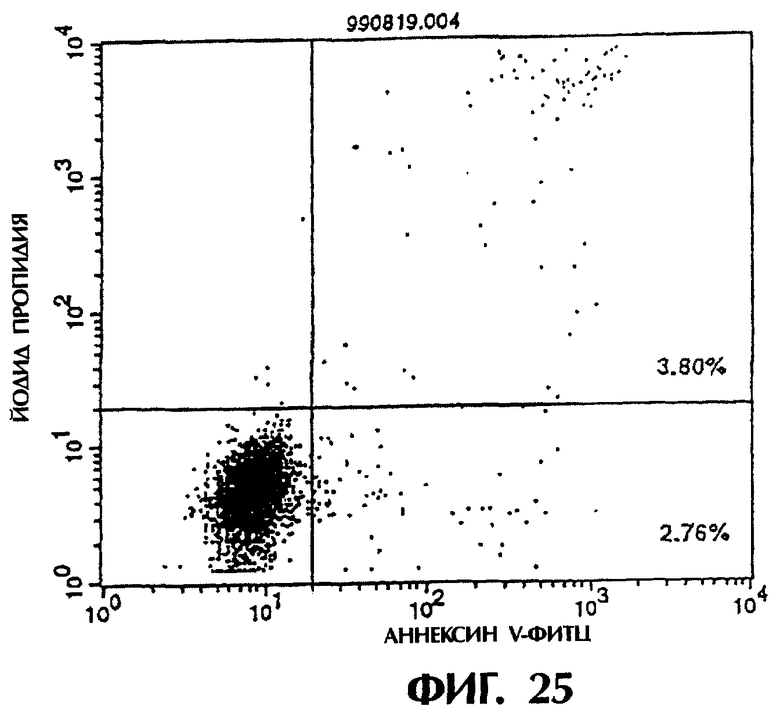
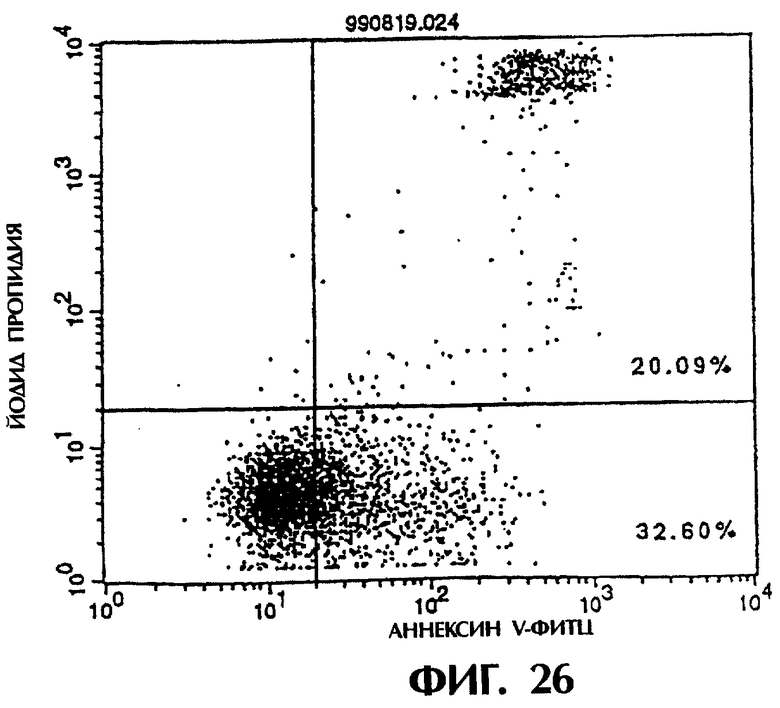

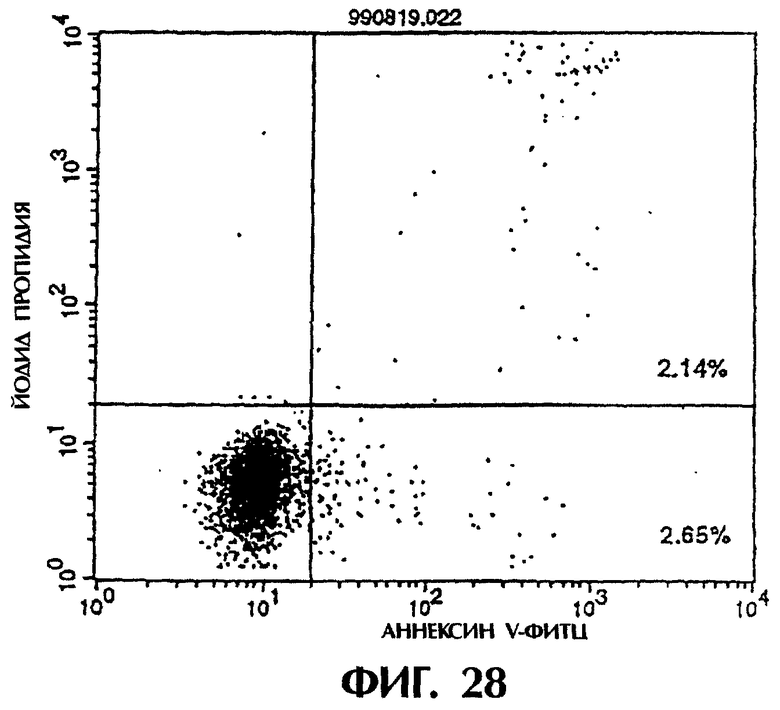
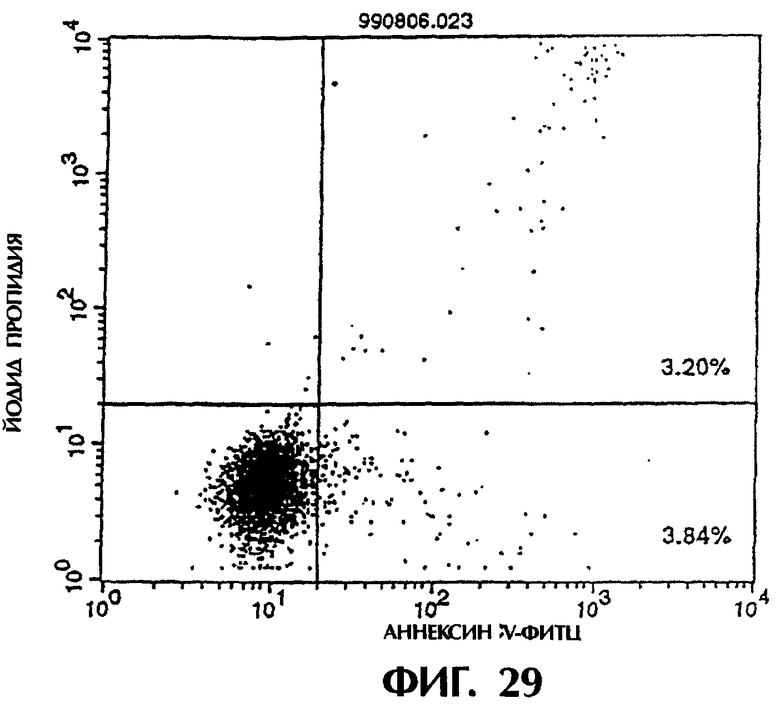
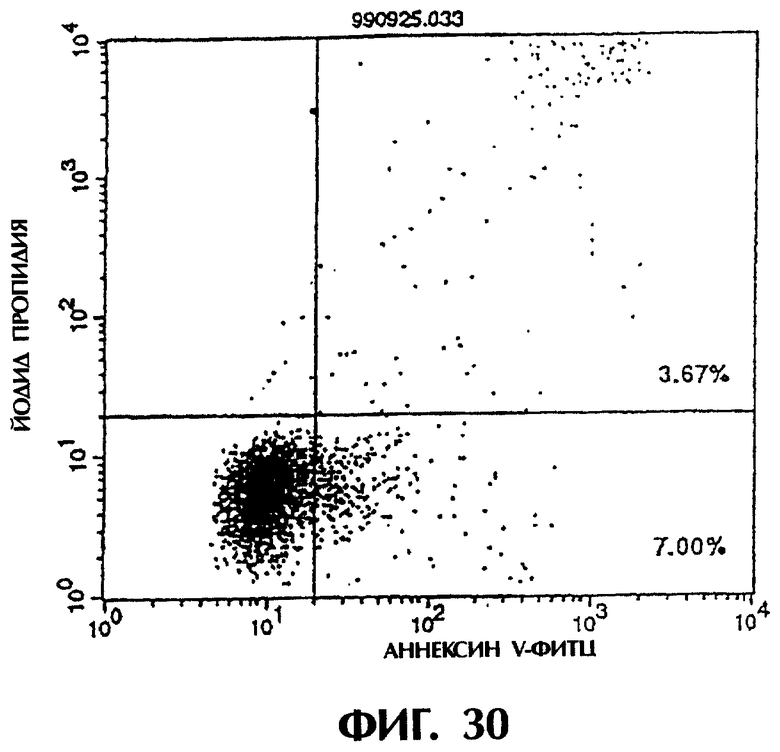
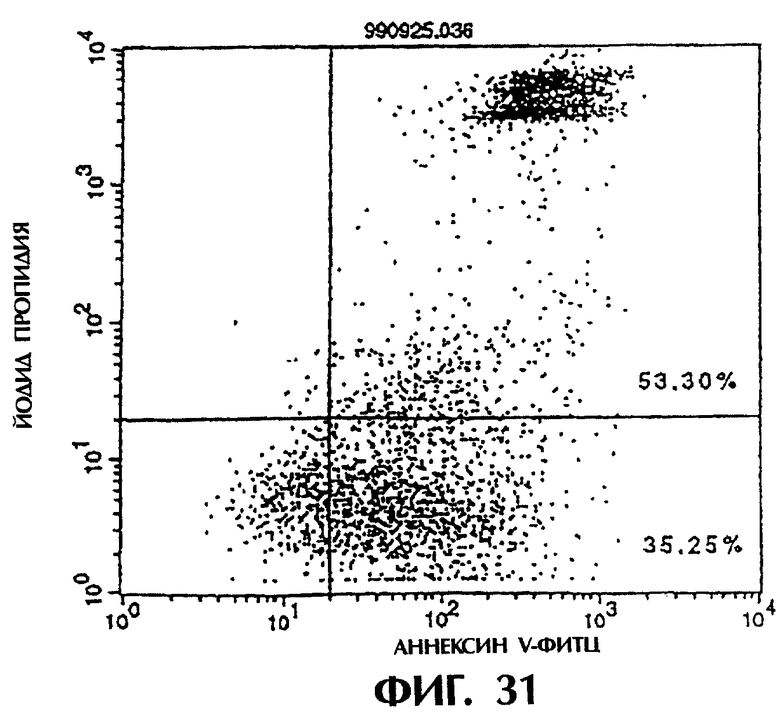
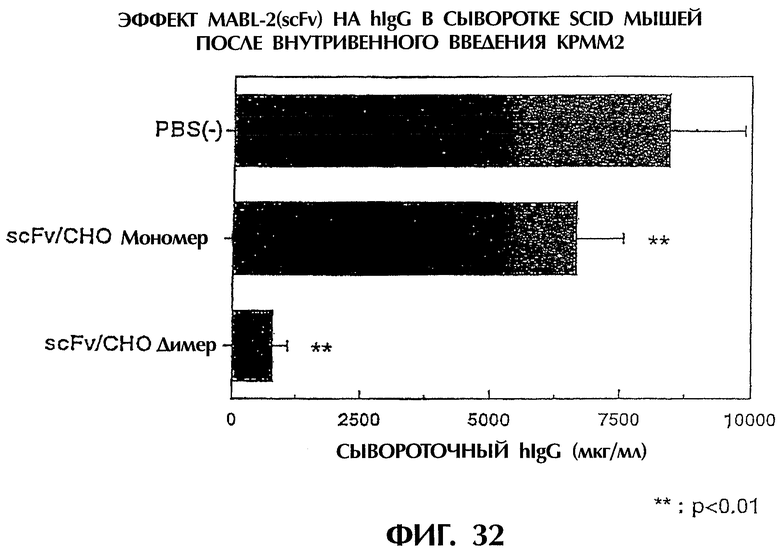
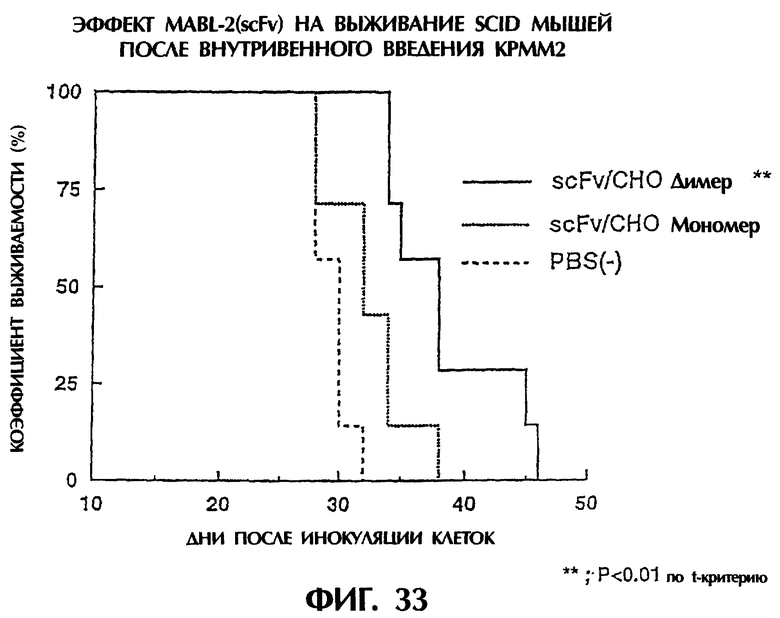
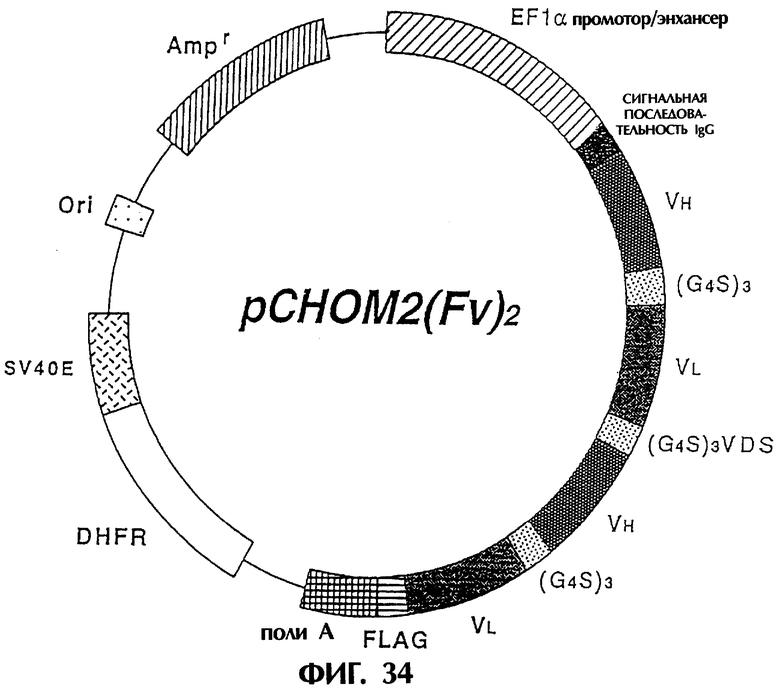
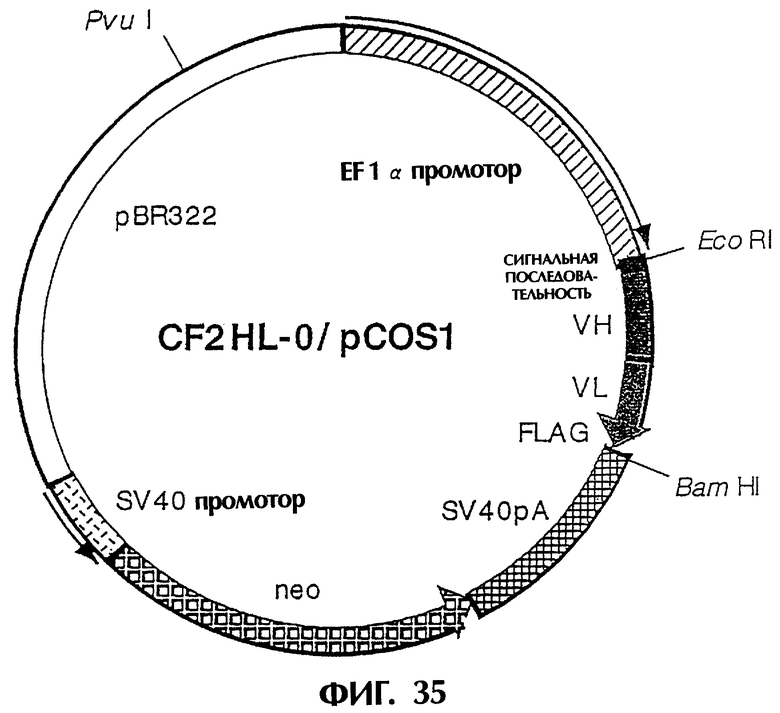
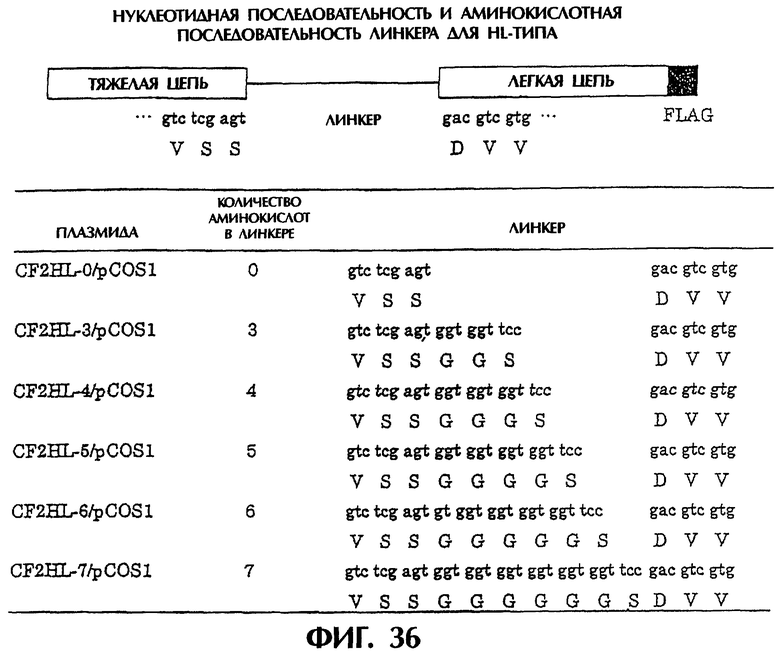
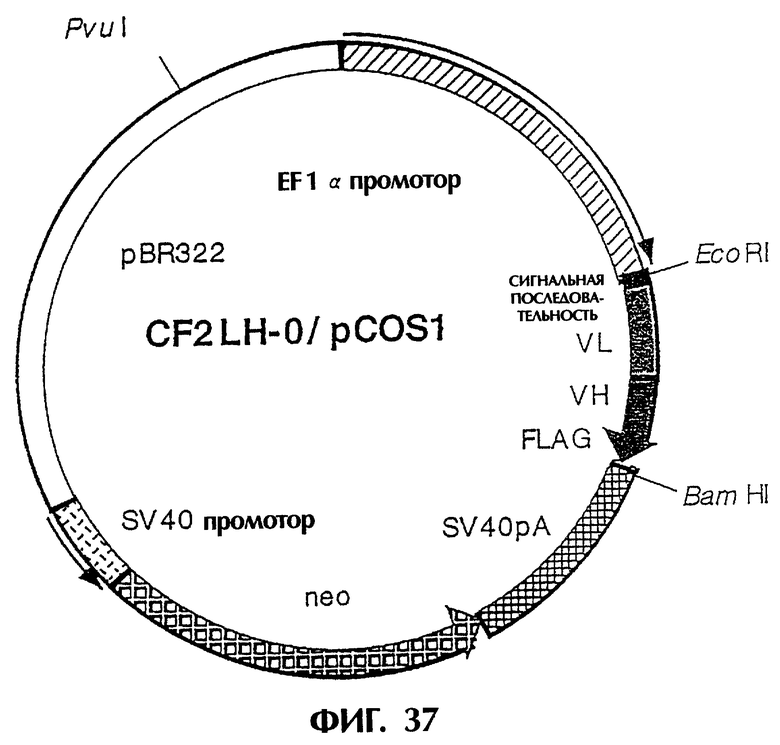
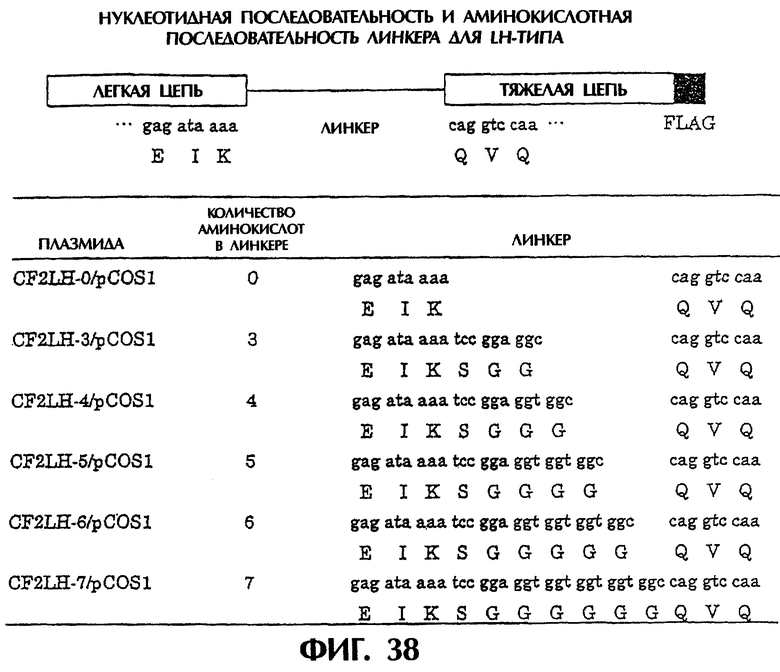
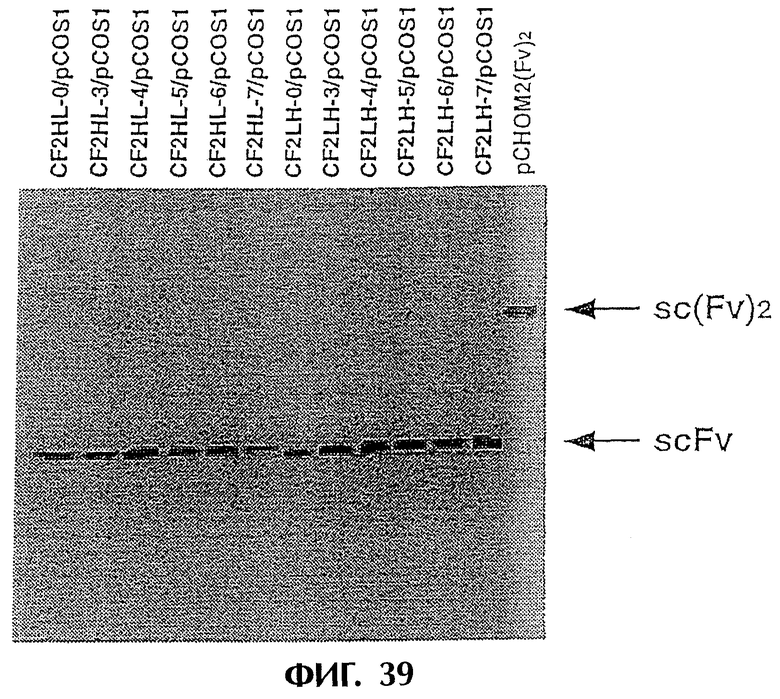
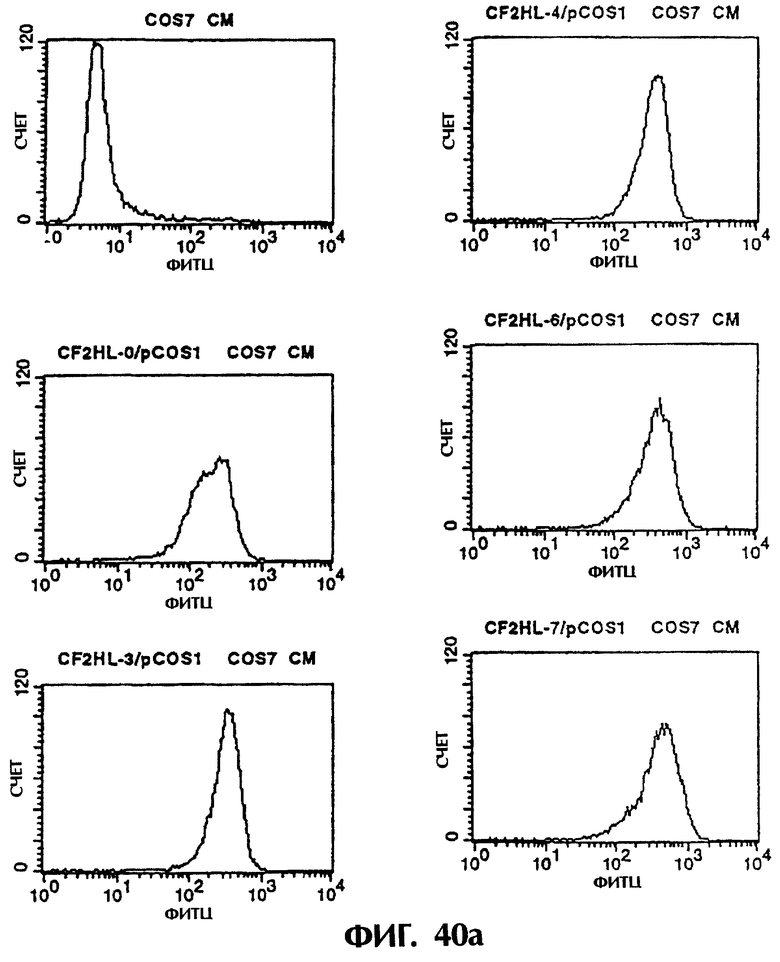
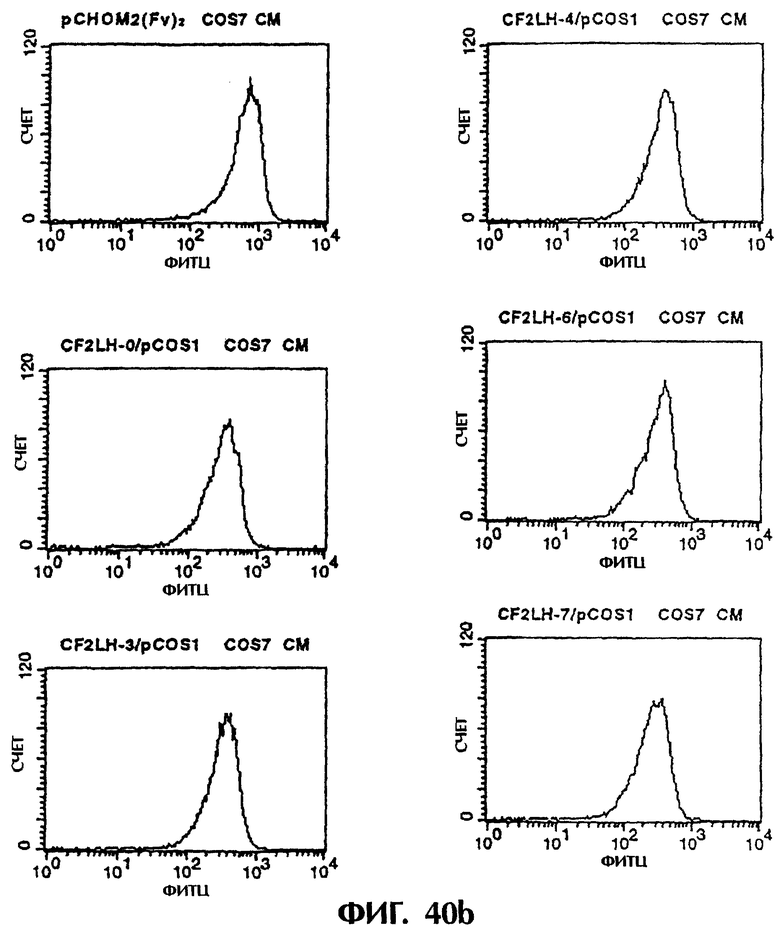
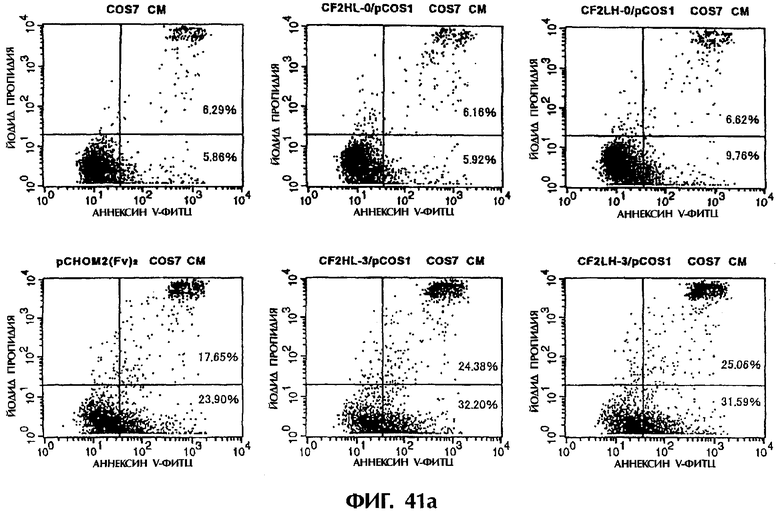
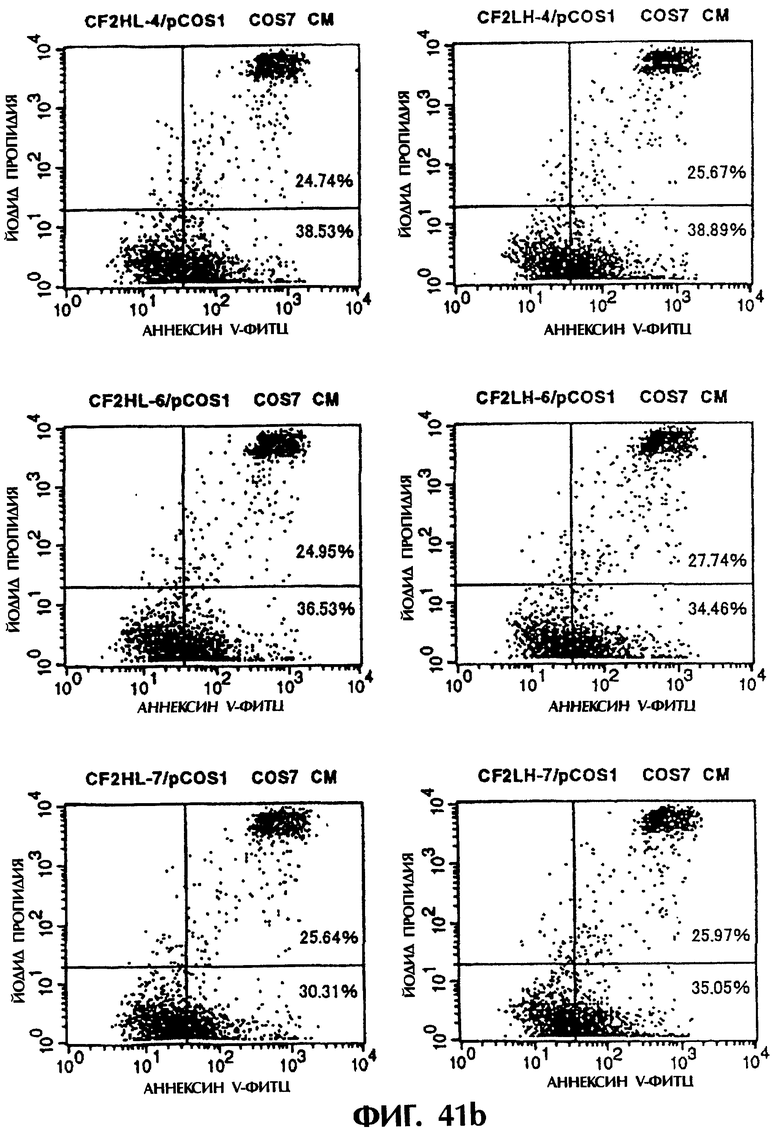

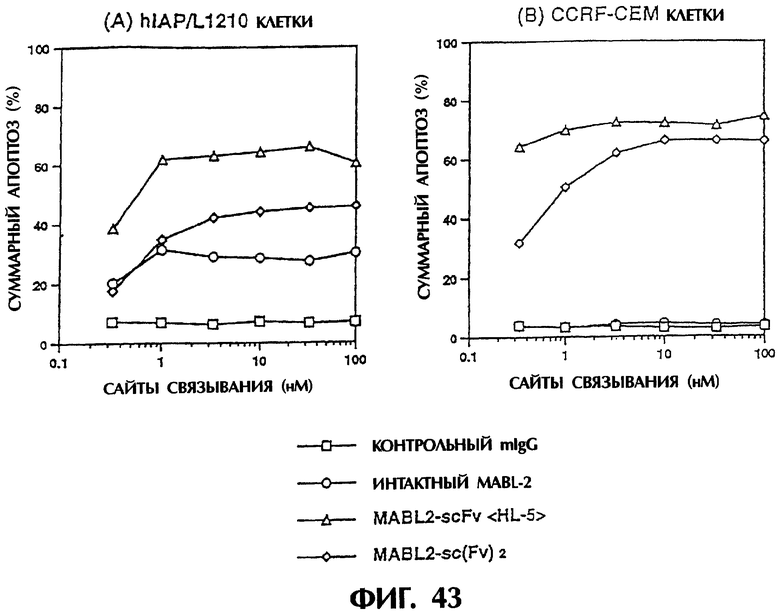
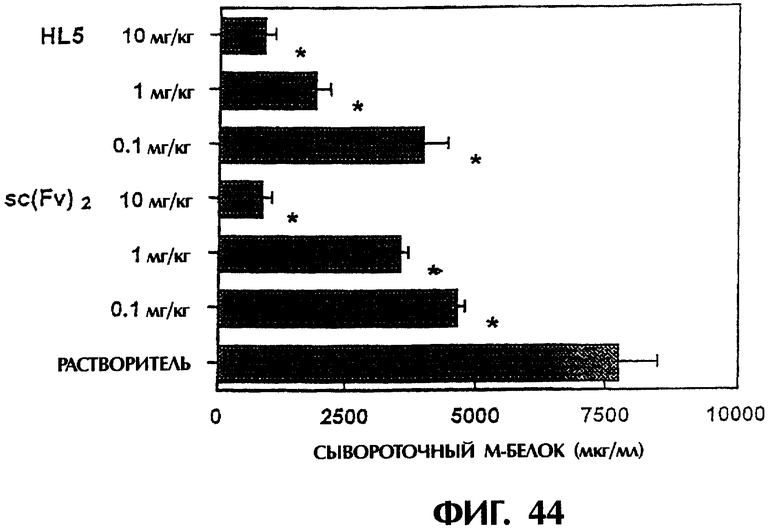
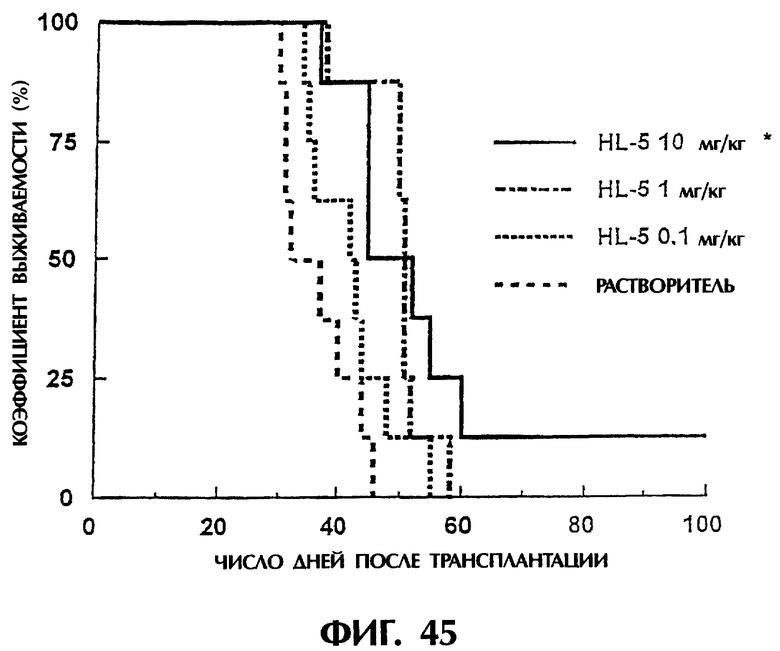
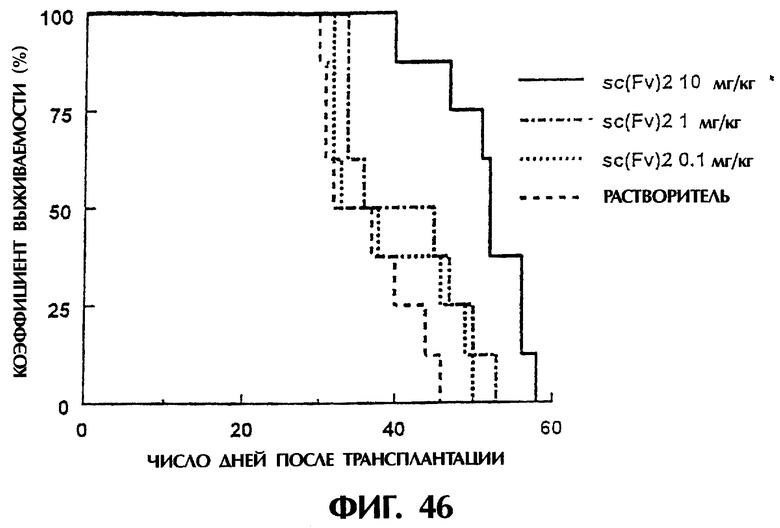
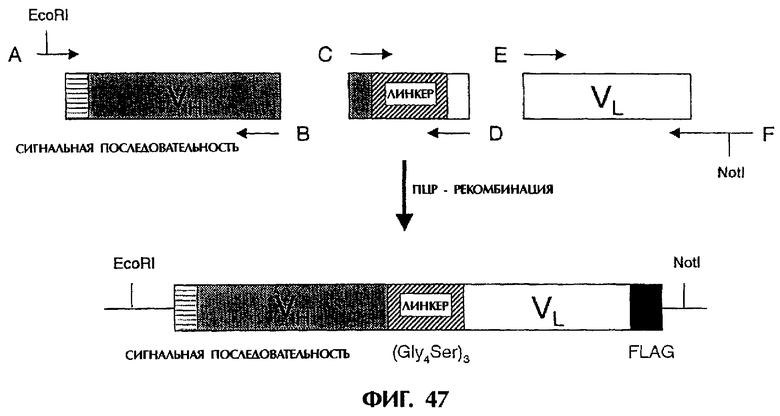
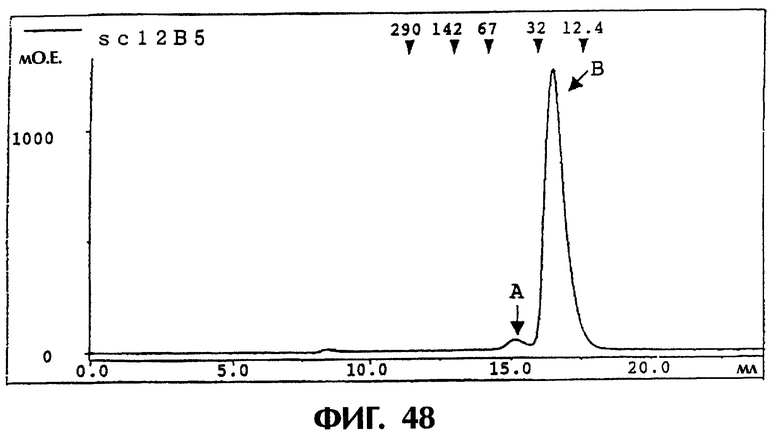


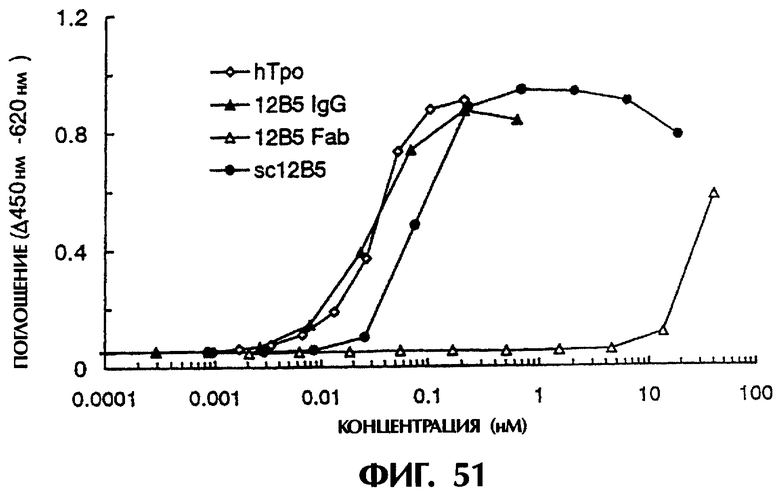
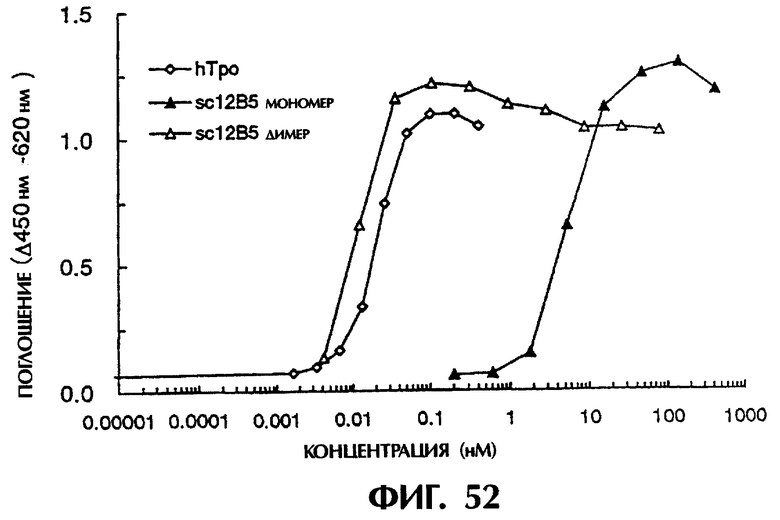
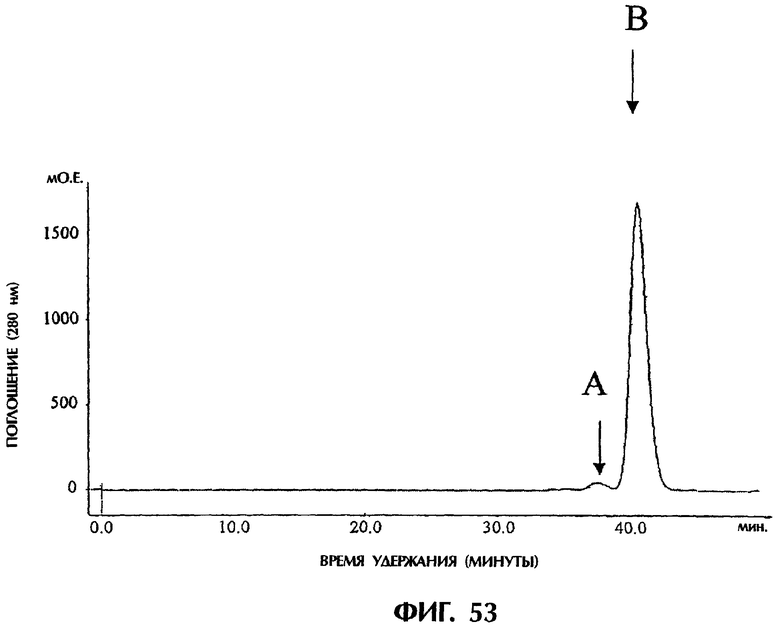

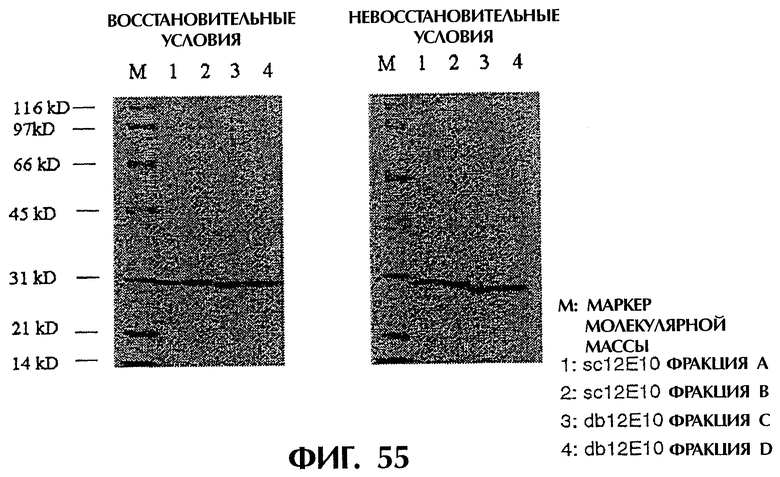
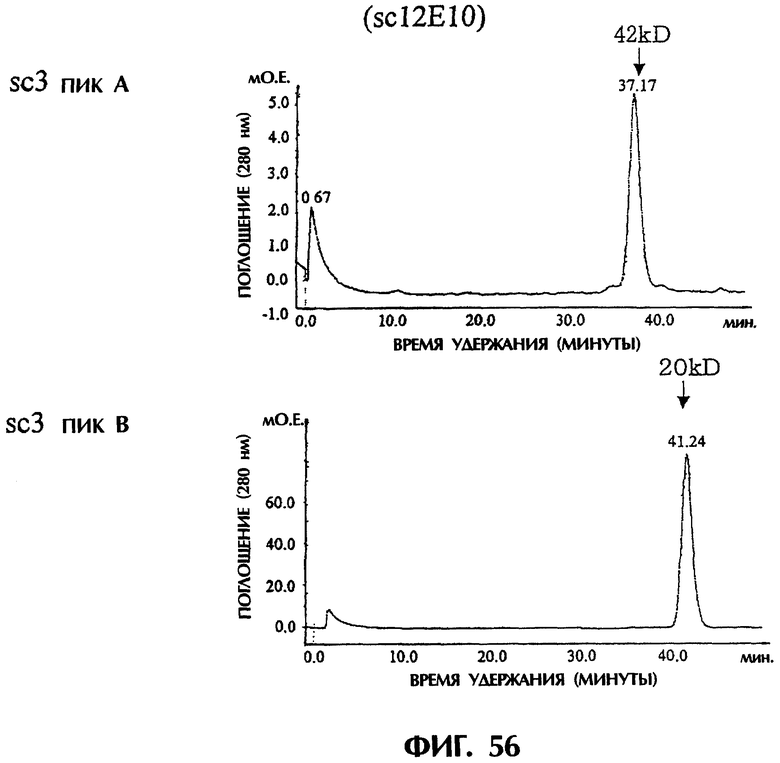
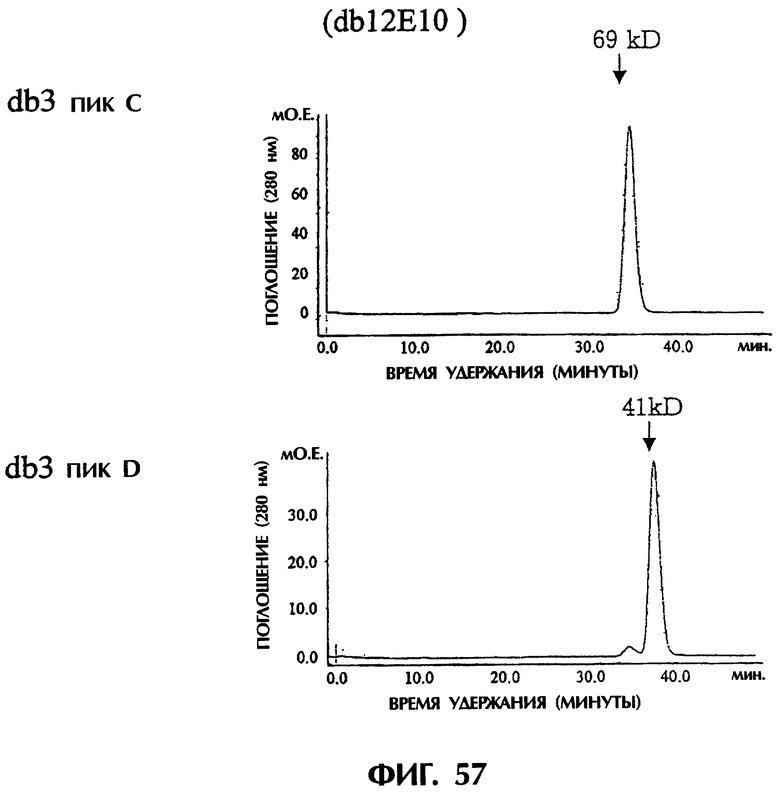
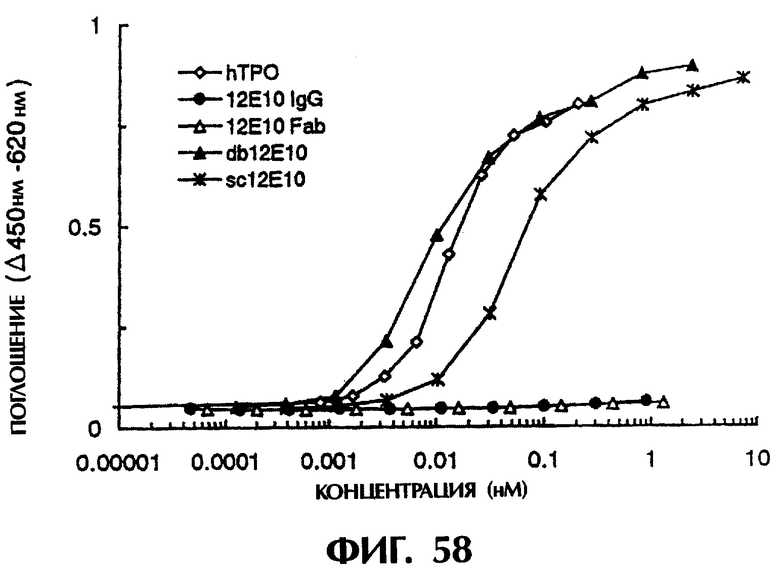

Изобретение относится к биотехнологии и может быть использовано в медицинской практике. Модифицированное антитело по изобретению содержит две или более V-области Н-цепи и две или более V-области L-цепи антитела, соединенные напрямую или через линкер ковалентной или нековалентной связью. Антитело имеет меньший размер по сравнению с исходным антителом, обладающим или по существу не обладающим агонистическим действием к рецептору ТРО. Антитело по изобретению является агонистом ТРО, способно специфически распознавать и поперечно сшивать рецептор ТРО. Для индукции агностического действия в клетках, экспрессирующих рецептор ТРО, клетки контактируют с модифицированным антителом. Измерение ТРО-агонистического действия антитела осуществляют по поперечной сшивке рецептора ТРО. Модифицированное антитело может быть использовано в качестве лекарственного средства для лечения тромбоцитопении. Модифицированные антитела обладают более высокой активностью, чем цельные антитела (IgG), и благодаря уменьшенным молекулярным размерам и отсутствию константных областей они обладают улучшенной проникающей способностью в ткани. 5 н. и 33 з.п. ф-лы, 59 ил., 3 табл.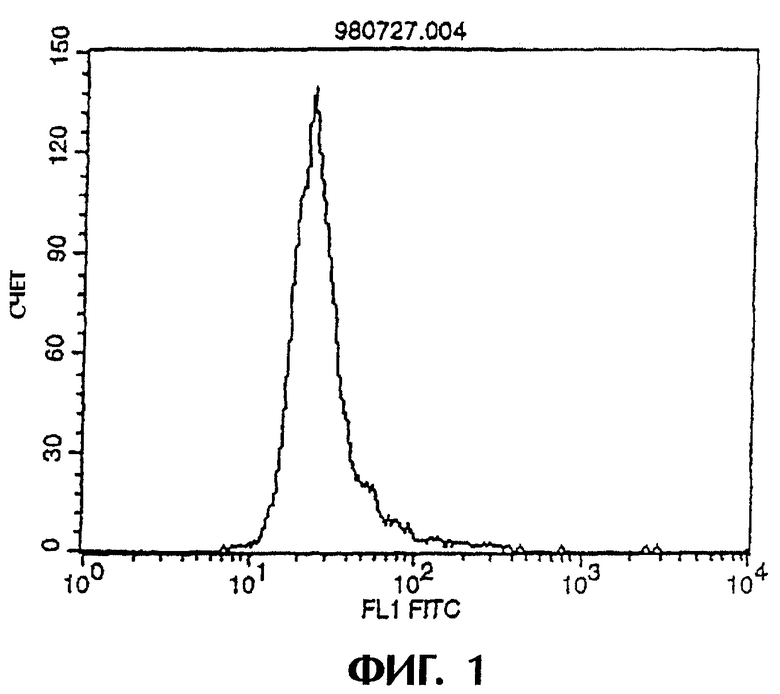
(1) получение модифицированного антитела, включающего две или более V-области Н-цепи и две или более V-области L-цепи антитела и связывающегося специфически с рецептором ТРО,
(2) осуществление реакции взаимодействия клеток, экспрессирующих указанный рецептор ТРО, с указанным модифицированным антителом и
(3) измерение ТРО-агонистического действия в клетках, вызванного поперечной сшивкой рецептора ТРО.
| US 5885574 А, 23.03.1999 | |||
| WO 00/53634 A, 14.09.2000 | |||
| Устройство для нагнетания раствора в скважину | 1982 |
|
SU1035132A1 |

