По настоящей заявке испрашивается приоритет согласно статье 35 закона США, б, в соответствии с предварительной заявкой серийный № 60/029629, поданной 25 октября 1996 года.
Область изобретения
Настоящее изобретение относится к многофункциональным химерным агонистам гемопоэтических рецепторов. Такие многофункциональные химерные агонисты гемопоэтических рецепторов проявляют один или более типов активности отдельных компонентов химерной молекулы и также могут проявлять повышенную активность по стимуляции кроветворных клеток и(или) проявлять повышенную активность, которая может включать снижение нежелательных биологических активностей, связанных с отдельными гемопоэтическими факторами роста, и(или) обладают улучшенными физическими характеристиками, которые могут включать повышенную растворимость, устойчивость и эффективность восстановления пространственной структуры.
Предпосылки изобретения
Колониестимулирующие факторы (CSFs), которые стимулируют дифференцировку и(или) пролиферацию клеток костного мозга, вызывают большой интерес ввиду их терапевтического потенциала, связанного с восстановлением подавленного уровня активности клеток, производных от кроветворных стволовых клеток.
CSFs и у человека, и у мыши уже идентифицированы и разграничены по типу их активности. Например, CSFs гранулоцитов (G-CSF) и CSF макрофагов (M-CSF) стимулируют формирование in vitro колоний нейтрофилов и макрофагов, соответственно, в то время как GM-CSF и интерлейкин-3 (IL-3) характеризуются более широким спектром активности и стимулируют формирование колоний как макрофагов нейтрофилов, так и эозинофилов. IL-3 также стимулирует формирование базофильных, мегакариоцитарных и чистых и смешанных эритроидных колоний.
В Патентах США 4877729 и 4959455 описаны кДНК IL-3 гиббона и расшифрованную последовательность ДНК IL-3 человека и последовательности белков, которые ими кодируются. Представленный hIL-3 содержит в 8-м положении белковой последовательности серин вместо пролина.
В заявке на Международный патент (РСТ) WO 88/00598 описаны IL-3, подобные IL-3 гиббона и человека. В составе hIL-3 произведена замена серина8 на пролин8. Получены подтверждения замещения цистеина серином, что приводит к разрушению дисульфидной связи и замещению одной или нескольких аминокислот по сайтам потенциального гликозилирования.
В патенте США 4810643 представлена последовательность ДНК, кодирующая G-CSF человека.
В Международном патенте WO 91/02754 представлен гибридный белок, включающий GM-CSF и IL-3, который проявляет повышенную биологическую активность, по сравнению с отдельными GM-CSF и IL-3. Также представлены негликозилированные белковые аналоги IL-3 и GM-CSF, такие как компоненты многофункционального химерного агониста гемопоэтических рецепторов.
В Международном патенте WO 92/04455 представлены гибридные белки, включающие IL-3, гибридизованный с лимфокином, выбранный из группы, состоящей из IL-3, IL-6, IL-7, IL-9, IL-11, ЕРО и G-CSF.
В Международных патентах WO 95/21197 и WO 95/21254 представлены гибридные белки, обладающие широким спектром многофункциональных кроветворных эффектов.
Патент Великобритании 2285446 относится к лиганду с-mpl (тромбопоэтин) и различными формами тромбопоэтина, для которых продемонстрировано влияние на репликацию, дифференцировку и созревание мегакариоцитов и предшественников мегакариоцитов, которые могут быть использованы в лечении тромбоцитопении.
Патент ЕР 675201 А1 относится к лиганду с-mpl (фактор роста и развития мегакариоцитов, MGDF), аллельным вариантам лиганда с-mpl и лиганду с-mpl, прикрепленному к водорастворимым полимерам, таким как полиэтиленгликоль.
В Международном патенте WO 95/21920 описаны лиганды с-mpl мыши и человека и его полипептидные фрагменты. Белки применимы для использования в терапии in vivo и ex vivo с целью стимуляции выработки тромбоцитов.
В патенте США 4703008, выданном F.-K.Lin, описана последовательность кДНК, кодирующая эритропоэтин, способы получения и использования эритропоэтина.
В Международном патенте WO 91/05867 описаны аналоги эритропоэтина человека, имеющие большее число сайтов присоединения углеводов в сравнении с эритропоэтином человека, такие как EPO(-Asn69), EPO(-Asn125, Ser127), EPO(-Thr125) и ЕРО(-Pro124, Thr125).
В Международном патенте WO 94/24160 представлены мутантные варианты эритропоэтина, которые характеризуются повышенной активностью, что связано с заменами в положениях 20, 49, 73, 140, 143, 146, 147 и 154.
В Международном патенте WO 94/28391 представлена последовательность нативного лиганда flt3 и последовательность кДНК, кодирующая лиганд flt3, способы экспрессии лиганда flt3 в клетке-хозяине, трансфицированной кДНК, и способы терапии пациентов с нарушениями кроветворения с использованием лиганда flt3.
В патенте США 5554512 представлен лиганд flt3 человека в виде выделенного белка, ДНК, кодирующего лиганд flt3, клетки-хозяина, трансформированные кДНК, кодирующей лиганд flt3, и способы терапии пациентов с использованием лиганда flt3.
В Международном патенте WO 94/26891 представлены лиганды flt3 млекопитающих, включая вариант, имеющий вставку из 29 аминокислот, и соответствующий фрагмент.
Реаранжировка белковых последовательностей
В эволюции реаранжировка последовательностей ДНК играет важную роль в возникновении многообразий структуры и функций белков. Дупликации генов и "перемешивание" экзонов представляют собой важные механизмы быстрого образования разнообразия, и, соответственно, обеспечивают организмам конкурентоспособность, особенно в случаях низкого естественного уровня мутирования (Doolittle, 1992, Protein Sci., 1, 191-200).
Разработка методик рекомбинантной ДНК сделала возможным изучение влияния изменений последовательности на пространственную структуру, организацию и функции белка. Способ создания новых последовательностей позволил установить, что в природе существуют пары белков, которые характеризуются родственной линейной реорганизацией их аминокислотных последовательностей (Cunningham et al., 1979, Proc. Natl. Acad. Sci. USA, 76, 3218-3222; Teather & Erfle, 1990, J. Bacteriol., 172, 3837-3841; Schimming et al., 1992, Eur. J. Biochem., 204, 13-19; Yamiuchi & Minamikawa, 1991, FEBS Lett., 260, 127-130; MacGregor et al., FEBS Lett., 378, 263-266). Впервые применение такого типа перестроек белков in vitro было описано Гольденбергом и Крейтоном (Goldenberg & Creighton, 1983, J. Mol. Biol., 165, 407-413). Новый N-конец был выбран в промежуточном сайте (как точка разрыва) исходной последовательности: новая последовательность характеризуется тем же порядком расположения аминокислот, как и исходная последовательность, от точки разрыва до или почти до исходного С-конца. В этой точке новую последовательность соединяют (прямо или опосредованно через добавочный фрагмент последовательности - линкер) с аминокислотой, находящейся в или рядом с исходным N-концом; новую последовательность продолжают той же последовательностью, что и оригинальная последовательность, до тех пор, пока она не достигнет самой точки или участка, соседствующего с аминокислотой, которая является N-концевой в сайте разрыва исходной последовательности - этот остаток образует новый С-конец цепи.
Такой подход был применен к белкам, размеры которых варьировались в пределах от 58 до 462 аминокислот (Goldenberg & Creighton, 1983, J. Mol. Biol., 165, 407-413; Li & Coffino, 1993, Mol. Cell. Biol., 13, 2377-2383). Исследованные белки охватывают широкий круг структурных классов, включая белки, содержащие доминирующие α-спирали, например, интерлейкин-4 (Kreitm an et al., 1995, Cytokine, 7, 311-318), β-плоскости, например, интерлейкин-1 (Horlick et al., 1992, Protein Eng., 5, 427-431), или смесь этих двух структур, например, фосфорибозилантранилатизомераза дрожжей (Luger et al., 1989, Science, 243, 206-210). Большое число категорий белковых функций представлены в перечисленных исследованиях реорганизации аминокислотных последовательностей:
 mer, 1995, J. Mol. Biol., 250, 617-626
mer, 1995, J. Mol. Biol., 250, 617-626
Результаты этих исследований в значительной степени варьируются. Во многих случаях была выявлена существенно меньшая активность, растворимость и термодинамическая стабильность (дигидрофолатредуктаза кишечной палочки, аспартат-транскарбамилаза, фосфорибозилантранилатизомераза, глицеральдегид-3-фосфатдегидрогеназа, орнитиндекарбоксилаза, оmр-А, фосфоглицератдегидрогеназа дрожжей). В других случаях белки с перестроенной последовательностью характеризуются многими свойствами, практически идентичными таковым у их естественных аналогов (основной трипсиновый ингибитор поджелудочной железы, лизоцим Т4, рибонуклеаза Т1, β-глюканаза палочки, интерлейкин-1b, домен SH3 α-спектрина, пепсиноген, интерлейкин-4). В исключительных случаях было отмечено неожиданное улучшение некоторых свойств естественной последовательности, например, растворимости и темпа восстановления пространственной структуры (у домена SH3 α-спектрина), рецепторной аффинности и противоопухолевой активности переносимой молекулы химерного белка, состоящего из интерлейкина-4 и экзотоксина псевдомонады (Kreitman et al., 1994, Proc. Natl. Acad. Sci. USA, 91, 6889-6893; Kreitman et al., 1995, Cancer Res., 55, 3357-3363).
Исходная мотивация отмеченых типов исследований была связана с изучением роли кратко- и долговременных взаимодействий складчатости белков и стабильности. Перестройки аминокислотных последовательностей такого типа представляют группу взаимодействий, которые могут превращаться из долговременных в исходной последовательности в кратковременные в перестроенной последовательности, и наоборот. Тот факт, что многие из этих перестроенных последовательностей способны поддерживать конформационное состояние по крайней мере с некоторым уровнем активности, является убедительным доказательством того, что образование складчатости белка (пространственной организации) происходит с участием разнообразных механизмов (Viguera et al., 1995, J. Mol. Biol., 247, 670-681). В случае с доменом SH3 α-спектрина подбор новых окончаний в сайтах, соответствующих оборотам β-шпилек, приводят к образованию белков с ненамного сниженной стабильностью, которые, тем не менее, способны формировать пространственную структуру.
Положения внутренних точек разрывов, используемых в перечисленных выше работах, обнаруживаются исключительно на поверхности белков и распределены вдоль первичной последовательности полипептида без отчетливого смещения по направлению к концам или середине (изменчивость относительного расстояния от исходного N-конца до точек разрывов составляла от 10 до 80% в расчете на полную длину последовательности). Длина линкеров, использовавшихся для соединения исходных N- и С-концов, составляла от 0 до 9 аминокислотных остатков. В одном случае (Yang & Schachman, 1993, Proc. Natl. Acad. Sci. USA, 90, 11980-11984) фрагмент последовательности вырезали из исходного С-концевого сегмента, а соединение осуществляли через укороченый С-конец с исходным N-концом. Гибкие гидрофильные остатки, такие как глицин и серин, часто используются в составе линкеров. Фигуероа с соавт. (Viguera et al., 1995, J. Mol. Biol., 247, 670-681) сравнивали соединение исходных N- и С-концов через 3- или 4-аминокислотные линкеры; термодинамически 3-аминокислотный линкер оказывается менее стабильным. Протасова с соавт. (Protasova et al., 1994, Protein Eng., 7, 1373-1377) использовали линкеры, состоящие из 3 или 5 остатков, при соединении исходных N-концов дигидрофолатредуктазы E.coli: только 3-аминокислотный линкер позволял получать белок с хорошим выходом.
Сущность изобретения
Гемопоэтический белок, включающий аминокислотную последовательность по следующей формуле:
R1-L1-R2, R2-L1-R1 или R1-R2 или R2-R1,
при том, что R1 и R2 являются независимо отбираемыми фрагментами из группы, включающей:
(I) полипептид агониста рецептора ЕРО человека, включающий модифицированную аминокислотную последовательность ЕРО согласно формуле:
при том, что произвольно 1-6 аминокислот с N-конца и 1-5 аминокислот с С-конца могут быть делетированы из состава упомянутого полипептида рецепторного агониста;
при том, что N-конец присоединяют к С-концу напрямую или через линкер (L2), способный соединять N-конец с С-концом и имеющий новые С- и N-концы в аминокислотах;
(II) полипептид агониста рецептора фактора стволовых клеток человека, включающий модифицированную аминокислотную последовательность фактора стволовых клеток согласно формуле:

при том, что произвольно 1-23 аминокислоты могут быть делетированы от С-конца упомянутого полипептида агониста рецептора фактора стволовых клеток;
при том, что N-конец присоединяют к С-концу напрямую или через линкер (L2), способный соединять N-конец с С-концом и имеющий новые С- и N-концы в аминокислотах;
(III) полипептид агониста рецептора flt-3 человека, включающий модифицированную аминокислотную последовательность лиганда flt-3 согласно формуле:

при том, что 1-7 аминокислот произвольно делетируют с С-конца упомянутого полипептида агониста рецептора flt-3;
при том, что N-конец присоединяют к С-концу напрямую или через линкер (L2), способный соединять N-конец с С-концом и имеющий новые С- и N-концы в аминокислотах;
(IV) полипептид, включающий модифицированную аминокислотную последовательность G-CSF человека согласно формуле:

при том, что
Хаа в положении 1 - это Thr, Sеr, Аrg, Туr или Gly;
Хаа в положении 2 - это Pro или Leu;
Хаа в положении 3 - это Leu, Аrg, Tyr или Ser;
Хаа в положении 13 - это Phe, Ser, His, Thr или Pro;
Хаа в положении 16 - это Lys, Pro, Ser, Thr или His;
Хаа в положении 17 - это Cys, Ser, Gly, Ala, Ile, Tyr или Arg;
Хаа в положении 18 - это Leu, Thr, Pro, His, Ile или Cys;
Хаа в положении 22 - это Arg, Tyr, Ser, Thr или Ala;
Хаа в положении 24 - это Ile, Pro, Tyr или Leu;
Хаа в положении 27 - это Asp или Gly;
Хаа в положении 30 - это Ala, Ile, Leu или Gly;
Хаа в положении 34 - это Lys или Ser;
Хаа в положении 36 - это Cys или Ser;
Хаа в положении 42 - это Cys или Ser;
Хаа в положении 43 - это His, Thr, Gly, Val, Lys, Trp, Ala,Arg, Cys или Leu;
Хаа в положении 44 - это Pro, Gly, Arg, Asp, Val, Ala, His, Trp, Gln или Thr;
Хаа в положении 46 - это Glu, Arg, Phe, Arg, Ile или Ala;
Хаа в положении 47 - это Leu или Thr;
Хаа в положении 49 - это Leu, Phe, Arg или Ser;
Хаа в положении 50 - это Leu, Ile, His, Pro или Tyr;
Хаа в положении 54 - это Leu или His;
Хаа в положении 64 - это Cys или Ser;
Хаа в положении 67 - это Gln, Lys, Leu или Cys;
Хаа в положении 70 - это Gln, Pro, Leu, Arg или Ser;
Хаа в положении 74 - это Суs или Ser;
Хаа в положении 104 - это Asp, Gly или Val;
Хаа в положении 108 - это Leu, Ala, Val, Arg, Trp, Gln или Gly;
Хаа в положении 115 - это Thr, His, Leu или Ala;
Хаа в положении 120 - это Gln, Gly, Arg, Lys или His;
Хаа в положении 123 - это Glu, Arg, Phe или Thr;
Хаа в положении 144 - это Phe, His, Arg, Pro, Leu, Gln или Glu;
Хаа в положении 146 - это Arg или Gln;
Хаа в положении 147 - это Arg или Gln;
Хаа в положении 156 - это His, Gly или Ser;
Хаа в положении 159 - это Ser, Arg, Thr, Туг, Val или Gly;
Хаа в положении 162 - это Glu, Leu, Gly или Trp;
Хаа в положении 163 - это Val, Gly, Arg или Ala;
Хаа в положении 169 - это Arg, Ser, Leu, Arg или Cys;
Хаа в положении 170 - это His, Arg или Ser;
при том, что 1-11 аминокислот с N-конца и 1-5 аминокислот с С-конца могут произвольно делетироваться из упомянутой модифицированной аминокислотной последовательности G-CSF человека; и
при том, что N-конец присоединяют к С-концу напрямую или через линкер (L2), способный соединять N-конец с С-концом и имеющий новые С- и N-концы в аминокислотах;
(V) полипептид, включающий модифицированную аминокислотную последовательность IL-3 человека, согласно формуле:

при том, что
Хаа в положении 17 - это Ser, Lys, Gly, Asp, Met, Gln или Arg;
Хаа в положении 18 - это Asn, His, Leu, Ile, Phe, Arg или Gln;
Хаа в положении 19 - это Met, Phe, Ile, Arg, Gly, Ala или Cys;
Хаа в положении 20 - это Ile, Cys, Gln, Glu, Arg, Pro или Ala;
Хаа в положении 21 - это Asp, Phe, Lys, Arg, Ala, Gly, Glu, Gln, Asn, Thr, Ser или Val;
Хаа в положении 22 - это Glu, Trp, Pro, Ser, Ala, His, Asp, Asn, Gln, Leu, Val или Gly;
Хаа в положении 23 - это Ile, Val, Ala, Gly, Trp, Lys, Phe, Leu, Ser или Arg;
Хаа в положении 24 - это Ile, Gly, Val, Arg, Ser, Phe или Leu;
Хаа в положении 25 - это Thr, His, Gly, Gln, Arg, Pro или Ala;
Хаа в положении 26 - это His, Thr, Phe, Gly, Arg, Ala или Trp;
Хаа в положении 27 - это Leu, Gly, Arg, Thr, Ser или Ala;
Хаа в положении 28 - это Lys, Arg, Leu, Gln, Gly, Pro, Val или Trp;
Хаа в положении 29 - это Gln, Asn, Leu, Pro, Arg или Val;
Хаа в положении 30 - это Pro, His, Thr, Gly, Asp, Gln, Ser, Leu или Lys;
Хаа в положении 31 - это Pro, Asp, Gly, Ala, Arg, Leu илиGln;
Хаа в положении 32 - это Leu, Val, Arg, Gln, Asn, Gly, Ala или Glu;
Хаа в положении 33 - это Pro, Leu, Gln, Ala, Thr или Glu;
Хаа в положении 34 - это Leu, Val, Gly, Ser, Lys, Glu, Gln, Thr, Arg, Ala, Phe, Ile или Met;
Хаа в положении 35 - это Leu, Ala, Gly, Asn, Pro, Gln или Val;
Хаа в положении 36 - это Asp, Leu или Val;
Хаа в положении 37 - это Phe, Ser, Pro, Trp или Ile;
Хаа в положении 38 - это Asn или Ala;
Хаа в положении 40 - это Leu, Trp или Arg;
Хаа в положении 41 - это Asn, Cys, Arg, Leu, His, Met или Pro;
Хаа в положении 42 - это Gly, Asp, Ser, Cys, Asn, Lys, Thr, Leu, Val, Glu, Phe, Tyr, Ile, Met или Ala;
Хаа в положении 43 - это Glu, Asn, Tyr, Leu, Phe, Asp, Ala, Cys, Gln, Arg, Thr, Gly или Ser;
Хаа в положении 44 - это Asp, Ser, Leu, Arg, Lys, Thr, Met, Trp, Glu, Asn, Gln, Ala или Pro;
Хаа в положении 45 - это Gln, Pro, Phe, Val, Met, Leu, Thr, Lys, Trp, Asp, Asn, Arg, Ser, Ala, Ile, Glu или His;
Хаа в положении 46 - это Asp, Phe, Ser, Thr, Cys, Glu, Asn, Gln, Lys, His, Ala, Tyr, Ile, Val или Gly;
Хаа в положении 47 - это Ile, Gly, Val, Ser, Arg, Pro или His;
Хаа в положении 48 - это Leu, Ser, Cys, Arg, Ile, His, Phe, Glu, Lys, Thr, Ala, Met, Val или Asn;
Хаа в положении 49 - это Met, Arg, Ala, Gly, Pro, Asn, His или Asp;
Хаа в положении 50 - это Glu, Leu, Thr, Asp, Tyr, Lys, Asn, Ser, Ala, Ile, Val, His, Phe, Met или Gln;
Хаа в положении 51 - это Asn, Arg, Met, Pro, Ser, Thr или His;
Хаа в положении 52 - это Asn, His, Arg, Leu, Gly, Ser или Thr;
Хаа в положении 53 - это Leu, Thr, Ala, Gly, Glu, Pro, Lys, Ser или Met;
Хаа в положении 54 - это Arg, Asp, Ile, Ser, Val, Thr, Gln, Asn, Lys, His, Ala или Leu;
Хаа в положении 55 - это Arg, Thr, Val, Ser, Leu или Gly;
Хаа в положении 56 - это Pro, Gly, Cys, Ser, Gln, Glu, Arg, His, Thr, Ala, Tyr, Phe, Leu, Val или Lys;
Хаа в положении 57 - это Asn или Gly;
Хаа в положении 58 - это Leu, Ser, Asp, Arg, Gln, Val или Cys;
Хаа в положении 59 - это Glu, Tyr, His, Leu, Pro или Arg;
Хаа в положении 60 - это Ala, Ser, Pro, Tyr, Asn или Thr;
Хаа в положении 61 - это Phe, Asn, Glu, Pro, Lys, Arg или Ser;
Хаа в положении 62 - это Asn, His, Val, Arg, Pro, Thr, Asp или Ile;
Хаа в положении 63 - это Arg, Tyr, Trp, Lys, Ser, His, Pro или Val;
Хаа в положении 64 - это Ala, Asn, Pro, Ser или Lys;
Хаа в положении 65 - это Val, Thr, Pro, His, Leu, Phe или Ser;
Хаа в положении 66 - это Lys, Ile, Arg, Val, Asn, Glu или Ser;
Хаа в положении 67 - это Ser, Ala, Phe, Val, Gly, Asn, Ile, Pro или His;
Хаа в положении 68 - это Leu, Val, Trp, Ser, Ile, Phe, Thr или His;
Хаа в положении 69 - это Gln, Ala, Pro, Thr, Glu, Arg, Trp, Gly или Leu;
Хаа в положении 70 - это Asn, Leu, Val, Trp, Pro или Ala;
Хаа в положении 71 - это Ala, Met, Leu, Pro, Arg, Glu, Thr,Gln, Trp или Asn;
Хаа в положении 72 - это Ser, Glu, Met, Ala, His, Asn, Arg или Asp;
Хаа в положении 73 - это Ala, Glu, Asp, Leu, Ser, Gly, Thr или Arg;
Хаа в положении 74 - это Ile, Met, Thr, Pro, Arg, Gly или Ala;
Хаа в положении 75 - это Glu, Lys, Gly, Asp, Pro, Trp, Arg, Ser, Gln или Leu;
Хаа в положении 76 - это Ser, Val, Ala, Asn, Trp, Glu, Pro, Gly или Asp;
Хаа в положении 77 - это Ile, Ser, Arg, Thr или Leu;
Хаа в положении 78 - это Leu, Ala, Ser, Glu, Phe, Gly или Arg;
Хаа в положении 79 - это Lys, Thr, Asn, Met, Arg, Ile, Gly или Asp;
Хаа в положении 80 - это Asn, Trp, Val, Gly, Thr, Leu, Glu или Arg;
Хаа в положении 81 - это Leu, Gln, Gly, Ala, Thr, Arg, Val или Lys;
Хаа в положении 82 - это Leu, Gln, Lys, Trp, Arg, Asp, Glu, Asn, His, Thr, Ser, Ala, Tyr, Phe, Ile, Met или Val;
Хаа в положении 83 - это Pro, Ala, Thr, Trp, Arg или Met;
Хаа в положении 84 - это Cys, Glu, Gly, Arg, Met или Val;
Хаа в положении 85 - это Leu, Asn, Val или Gln;
Хаа в положении 86 - это Pro, Cys, Arg, Ala или Lys;
Хаа в положении 87 - это Leu, Ser, Trp или Gly;
Хаа в положении 88 - это Ala, Lys, Arg, Val или Trp;
Хаа в положении 89 - это Thr, Asp, Cys, Leu, Val, Glu, His, Asn или Ser;
Хаа в положении 90 - это Ala, Pro, Ser, Thr, Gly, Asp, Ile или Met;
Хаа в положении 91 - это Ala, Pro, Ser, Thr, Phe, Leu, Asp или His;
Хаа в положении 92 - это Pro, Phe, Arg, Ser, Lys, His, Ala, Gly, Ile или Leu;
Хаа в положении 93 - это Thr, Asp, Ser, Asn, Pro, Ala, Leu или Arg;
Хаа в положении 94 - это Arg, Ile, Ser, Glu, Leu, Val, Gln, Lys, His, Ala или Pro;
Хаа в положении 95 - это His, Gln, Pro, Arg, Val, Leu, Gly, Thr, Asn, Lys, Ser, Ala, Trp, Phe, Ile или Tyr;
Хаа в положении 96 - это Pro, Lys, Tyr, Gly, Ile или Thr;
Хаа в положении 97 - это Ile, Val, Lys, Ala или Asn;
Хаа в положении 98 - это His, Ile, Asn, Leu, Asp, Ala, Thr, Glu, Gln, Ser, Phe, Met, Val, Lys, Arg, Tyr или Pro;
Хаа в положении 99 - это Ile, Leu, Arg, Asp, Val, Pro, Gln, Gly, Ser, Phe или His;
Хаа в положении 100 - это Lys, Tyr, Leu, His, Arg, Ile, Ser, Gln или Pro;
Хаа в положении 101 - это Asp, Pro, Met, Lys, His, Thr, Val, Tyr, Glu, Asn, Ser, Ala, Gly, Ile, Leu или Gln;
Хаа в положении 102 - это Gly, Leu, Glu, Lys, Ser, Tyr или Pro;
Хаа в положении 103 - это Asp или Ser;
Хаа в положении 104 - это Trp, Val, Cys, Tyr, Thr, Met, Pro, Leu, Gln, Lys, Ala, Phe или Gly;
Хаа в положении 105 - это Asn, Pro, Ala, Phe, Ser, Trp, Gln, Tyr, Leu, Lys, Ile. Asp или His;
Хаа в положении 106 - это Glu, Ser, Ala, Lys, Thr, Ile, Gly или Pro;
Хаа в положении 108 - это Arg, Lys, Asp, Leu, Thr, Ile, Gln, His, Ser, Ala или Pro;
Хаа в положении 109 - это Arg, Thr, Pro, Glu, Tyr, Leu, Ser или Gly;
Хаа в положении 110 - это Lys, Ala, Asn, Thr, Leu, Arg, Gln, His, Glu, Ser или Trp;
Хаа в положении 111 - это Leu, Ile, Arg, Asp или Met;
Хаа в положении 112 - это Thr, Val, Gln, Tyr, Glu, His, Ser или Phe;
Хаа в положении 113 - это Phe, Ser, Cys, His, Gly, Trp, Tyr, Asp, Lys, Leu, Ile, Val или Asn;
Хаа в положении 114 - это Tyr, Cys, His, Ser, Trp, Arg или Leu;
Хаа в положении 115 - это Leu, Asn, Val, Pro, Arg, Ala, His, Thr, Trp или Met;
Хаа в положении 116 - это Lys, Leu, Pro, Thr, Met, Asp, Val, Glu, Arg, Trp, Ser, Asn, His, Ala, Tyr, Phe, Gln или Ile;
Хаа в положении 117 - это Thr, Ser, Asn, Ile, Trp, Lys или Pro;
Хаа в положении 118 - это Leu, Ser, Pro, Ala, Glu, Cys, Asp или Tyr;
Хаа в положении 119 - это Glu, Ser, Lys, Рro, Leu, Thr, Туг или Аrg;
Хаа в положении 120 - это Asn, Ala, Pro, Leu, His, Val или Gln;
Хаа в положении 121 - это Ala, Ser, Ile, Asn, Pro, Lys, Asp или Gly;
Хаа в положении 122 - это Gln, Ser, Met, Trp, Arg, Phe, Pro, His, Ile, Туr или Cys;
Хаа в положении 123 - это Ala, Met, Glu, His, Ser, Pro, Туг или Leu;
при том, что аминокислоты с 1-й по 14-ю могут быть произвольно делетированы с N-конца и (или) аминокислоты с 1-й по 15-ю могут быть произвольно делетированы с С-конца упомянутой модифицированной аминокислотной последовательности IL-3 человека; при том, что аминокислоты с 0-й по 44-ю, обозначенные Хаа, отличаются от соответствующей аминокислотной последовательности нативного интерлейкина-3 (I-133) человека; и
при том, что N-конец присоединяют к С-концу напрямую или через линкер (L2), способный соединять N-конец с С-концом и имеющий новые С- и N-концы в аминокислотах;
(VI) полипептид, включающий модифицированную аминокислотную последовательность лиганда с-mpl человека, согласно формуле:

при том, что
Хаа в положении 112 делетируется или является Leu, Ala, Val, Ile, Pro, Phe, Trp или Met;
Хаа в положении 113 делетируется или является Pro, Phe, Ala, Val, Leu, Ile, Trp или Met;
Хаа в положении 114 делетируется или является Pro, Phe, Ala, Val, Leu, Ile, Trp или Met;
Хаа в положении 115 делетируется или является Gln, Gly, Ser, Thr, Туг или Asn;
и
при том, чго N-конец присоединяют к С-концу напрямую или через линкер (L2), способный соединять N-конец с С-концом и имеющий новые С- и N-концы в аминокислотах;
(VII) полипептид, включающий модифицированную аминокислотную последовательность IL-3 человека, согласно формуле:

при том, что
Хаа в положении 17 - это Ser, Lys, Gly, Asp, Met, Gln или Arg;
Хаа в положении 18 - это Asn, His, Leu, Ile, Phe, Arg или Gln;
Хаа в положении 19 - это Met, Phe, Ile, Arg, Gly, Ala или Cys;
Хаа в положении 20 - это Ile, Cys, Gln, Glu, Arg, Pro или Ala;
Хаа в положении 21 - это Asp, Phe, Lys, Arg, Ala, Gly, Glu, Gln, Asn, Thr, Ser или Val;
Хаа в положении 22 - это Glu, Trp, Pro, Ser, Ala, His, Asp, Asn, Gln, Leu, Val или Gly;
Хаа в положении 23 - это Ile, Val, Ala, Gly, Trp, Lys, Phe, Leu, Ser или Arg;
Хаа в положении 24 - это Ile, Gly, Val, Arg, Ser, Phe или Leu;
Хаа в положении 25 - это Thr, His, Gly, Gln, Arg, Pro или Ala;
Хаа в положении 26 - это His, Thr, Phe, Gly, Arg, Ala или Trp;
Хаа в положении 27 - это Leu, Gly, Arg, Thr, Ser или Ala;
Хаа в положении 28 - это Lys, Arg, Leu, Gln, Gly, Pro, Val или Trp;
Хаа в положении 29 - это Gln, Asn, Leu, Pro, Arg или Val;
Xaa в положении 30 - это Pro, His, Thr, Gly, Asp, Gln, Ser, Leu или Lys;
Xaa в положении 31 - это Pro, Asp, Gly, Ala, Arg, Leu илиGln;
Xaa в положении 32 - это Leu, Val, Arg, Gln, Asn, Gly, Ala или Glu;
Xaa в положении 33 - это Pro, Leu, Gln, Ala, Thr или Glu;
Xaa в положении 34 - это Leu, Val, Gly, Ser, Lys, Glu, Gln, Thr, Arg, Ala, Phe, Ile или Met;
Xaa в положении 35 - это Leu, Ala, Gly, Asn, Pro, Gln или Val;
Xaa в положении 36 - это Asp, Leu или Val;
Xaa в положении 37 - это Phe, Ser, Pro, Trp или Ile;
Xaa в положении 38 - это Asn или Ala;
Хаа в положении 40 - это Leu, Trp или Arg;
Xaa в положении 41 - это Asn, Cys, Arg, Leu, His, Met или Pro;
Xaa в положении 42 - это Gly, Asp, Ser, Cys, Asn, Lys, Thr, Leu, Val, Glu, Phe, Tyr, Ile, Met или Ala;
Xaa в положении 43 - это Glu, Asn, Tyr, Leu, Phe, Asp, Ala, Cys, Gln, Arg, Thr, Gly или Ser;
Xaa в положении 44 - это Asp, Ser, Leu, Arg, Lys, Thr, Met, Trp, Glu, Asn, Gln, Ala или Pro;
Xaa в положении 45 - это Gln, Pro, Phe, Val, Met, Leu, Thr, Lys, Trp, Asp, Asn, Arg, Ser, Ala, Ile, Glu или His;
Xaa в положении 46 - это Asp, Phe, Ser, Thr, Cys, Glu, Asn, Gln, Lys, His, Ala, Tyr, Ile, Val или Gly;
Xaa в положении 47 - это Ile, Gly, Val, Ser, Arg, Pro или His;
Xaa в положении 48 - это Leu, Ser, Cys, Arg, Ile. His, Phe, Glu, Lys, Thr, Ala, Met, Val или Asn;
Xaa в положении 49 - это Met, Arg, Ala, Gly, Pro, Asn, His или Asp;
Xaa в положении 50 - это Glu, Leu, Thr, Asp, Tyr, Lys, Asn, Ser, Ala, Ile, Val, His, Phe, Met или Gln;
Xaa в положении 51 - это Asn, Arg, Met, Pro, Ser, Thr или His;
Xaa в положении 52 - это Asn, His, Arg, Leu, Gly, Ser или Thr;
Xaa в положении 53 - это Leu, Thr, Ala, Gly, Glu, Pro, Lys, Ser или Met;
Xaa в положении 54 - это Arg, Asp, Ile, Ser, Val, Thr, Gln, Asn, Lys, His, Ala или Leu;
Xaa в положении 55 - это Arg, Thr, Val, Ser, Leu или Gly;
Xaa в положении 56 - это Pro, Gly, Cys, Ser, Gln, Glu, Arg, His, Thr, Ala, Tyr, Phe, Leu, Val или Lys;
Xaa в положении 57 - это Asn или Gly;
Xaa в положении 58 - это Leu, Ser, Asp, Arg, Gln, Val или Cys;
Xaa в положении 59 - это Glu, Tyr, His, Leu, Pro или Arg;
Xaa в положении 60 - это Ala, Ser, Pro, Tyr, Asn или Thr;
Xaa в положении 61 - это Phe, Asn, Glu, Pro, Lys, Arg или Ser;
Хаа в положении 62 - это Asn, His, Val, Arg, Pro, Thr, Asp или Ile;
Хаа в положении 63 - это Arg, Tyr, Trp, Lys, Ser, His, Pro или Val;
Хаа в положении 64 - это Ala, Asn, Pro, Ser или Lys;
Хаа в положении 65 - это Val, Thr, Pro, His, Leu, Phe или Ser;
Хаа в положении 66 - это Lys, Ile, Arg, Val, Asn, Glu или Ser;
Хаа в положении 67 - это Ser, Ala, Phe, Val, Gly, Asn, Ile, Pro или His;
Хаа в положении 68 - это Leu, Val, Trp, Ser, Ile, Phe, Thr или His;
Хаа в положении 69 - это Gln, Ala, Pro, Thr, Glu, Arg, Trp, Gly или Leu;
Хаа в положении 70 - это Asn, Leu, Val, Trp, Pro или Ala;
Хаа в положении 71 - это Ala, Met, Leu, Pro, Arg, Glu, Thr,Gln, Trp или Asn;
Хаа в положении 72 - это Ser, Glu, Met, Ala, His, Asn, Arg или Аsp;
Хаа в положении 73 - это Ala, Glu, Asp, Leu, Ser, Gly, Thr или Arg;
Хаа в положении 74 - это Ile, Met, Thr, Pro, Arg, Gly или Ala;
Хаа в положении 75 - это Glu, Lys, Gly, Asp, Pro, Trp, Arg, Ser, Gln или Leu;
Хаа в положении 76 - это Ser, Val, Ala, Asn, Trp, Glu, Pro, Gly или Asp;
Хаа в положении 77 - это Ile, Ser, Arg, Thr или Leu;
Хаа в положении 78 - это Leu, Ala, Ser, Glu, Phe, Gly или Arg;
Хаа в положении 79 - это Lys, Thr, Asn, Met, Arg, Ilе, Gly или Asp;
Хаа в положении 80 - это Asn, Trp, Val, Gly, Thr, Leu, Glu или Arg;
Хаа в положении 81 - это Leu, Gln, Gly, Ala, Thr, Arg, Val или Lys;
Хаа в положении 82 - это Leu, Gln, Lys, Trp, Arg, Asp, Glu, Asn, His, Thr, Ser, Ala, Tyr, Phe, Ile, Met или Val;
Хаа в положении 83 - это Pro, Ala, Thr, Trp, Arg или Met;
Хаа в положении 84 - это Cys, Glu, Gly, Arg, Met или Val;
Хаа в положении 85 - это Leu, Asn, Val или Gln;
Хаа в положении 86 - это Pro, Cys, Arg, Ala или Lys;
Хаа в положении 87 - это Leu, Ser, Trp или Gly;
Хаа в положении 88 - это Ala, Lys, Arg, Val или Trp;
Хаа в положении 89 - это Thr, Asp, Cys, Leu, Val, Glu, His, Asn или Ser;
Хаа в положении 90 - это Ala, Pro, Ser, Thr, Gly, Asp, Ile или Met;
Хаа в положении 91 - это Ala, Pro, Ser, Thr, Phe, Leu, Asp или His;
Хаа в положении 92 - это Pro, Phe, Arg, Ser, Lys, His, Ala, Gly, Ile или Leu;
Хаа в положении 93 - это Thr, Asp, Ser, Asn, Pro, Ala, Leu или Arg;
Xaa в положении 94 - это Arg, Ile, Ser, Glu, Leu, Val, Gln, Lys, His, Ala или Pro;
Xaa в положении 95 - это His, Gln, Pro, Arg, Val, Leu, Gly, Thr, Asn, Lys, Ser, Ala, Trp, Phe, Ile или Tyr;
Xaa в положении 96 - это Pro, Lys, Tyr, Gly, Ile или Thr;
Xaa в положении 97 - это Ile, Val, Lys, Ala или Asn;
Xaa в положении 98 - это His, Ile, Asn, Leu, Asp, Ala, Thr, Glu, Gln, Ser, Phe, Met, Val, Lys, Arg, Tyr или Pro;
Xaa в положении 99 - это Ile, Leu, Arg, Asp, Val, Pro, Gln, Gly, Ser, Phe или His;
Xaa в положении 100 - это Lys, Tyr, Leu, His, Arg, Ile, Ser, Gln или Pro;
Xaa в положении 101 - это Asp, Pro, Met, Lys, His, Thr, Val, Tyr, Glu, Asn, Ser, Ala, Gly, Ile, Leu или Gln;
Xaa в положении 102 - это Gly, Leu, Glu, Lys, Ser, Tyr или Pro;
Xaa в положении 103 - это Asp или Ser;
Xaa в положении 104 - это Trp, Val, Cys, Tyr, Thr, Met, Pro, Leu, Gln, Lys, Ala, Phe или Gly;
Xaa в положении 105 - это Asn, Pro, Ala, Phe, Ser, Trp, Gln, Tyr, Leu, Lys, Ile, Asp или His;
Xaa в положении 106 - это Glu, Ser, Ala, Lys, Thr, Ile, Gly или Pro;
Xaa в положении 108 - это Arg, Lys, Asp, Leu, Thr, Ile, Gln, His, Ser, Ala или Pro;
Xaa в положении 109 - это Arg, Thr, Pro, Glu, Tyr, Leu, Ser или Gly;
Xaa в положении 110 - это Lys, Ala, Asn, Thr, Leu, Arg, Gln, His, Glu, Ser или Trp;
Xaa в положении 111 - это Leu, Ile, Arg, Asp или Met;
Xaa в положении 112 - это Thr, Val, Gln, Tyr, Glu, His, Ser или Phe;
Xaa в положении 113 - это Phe, Ser, Cys, His, Gly, Trp, Tyr, Asp, Lys, Leu, Ile, Val или Asn;
Xaa в положении 114 - это Tyr, Cys, His, Ser, Trp, Arg или Leu;
Xaa в положении 115 - это Leu, Asn, Val, Pro, Arg, Ala, His, Thr, Trp или Met;
Xaa в положении 116 - это Lys, Leu, Pro, Thr, Met, Asp, Val, Glu, Arg, Trp, Ser, Asn, His, Ala, Tyr, Phe, Gln или Ile;
Xaa в положении 117 - это Thr, Ser, Asn, Ile, Trp, Lys или Pro;
Xaa в положении 118 - это Leu, Ser, Pro, Ala, Glu, Cys, Asp или Tyr;
Xaa в положении 119 - это Glu, Ser, Lys, Pro, Leu, Thr, Tyr или Arg;
Xaa в положении 120 - это Asn, Ala, Pro, Leu, His, Val или Gln;
Xaa в положении 121 - это Ala, Ser, Ile, Asn, Pro, Lys, Asp или Gly;
Xaa в положении 122 - это Gln, Ser, Met, Trp, Arg, Phe, Pro, His, Ile, Tyr или Cys;
Xaa в положении 123 - это Ala, Met, Glu, His, Ser, Pro, Tyr или Leu;
при том, что аминокислоты с 1-й по 14-ю могут быть произвольно делетированы с N-конца и (или) аминокислоты с 1-й по 15-ю могут быть произвольно делетированы с С-конца упомянутой модифицированной аминокислотной последовательности IL-3 человека; при том, что аминокислоты с 1-й по 44-ю, обозначенные Хаа, отличаются от соответствующей аминокислотной последовательности нативного (1-133) интерлейкина-3 (IL-3) человека; и
(VIII) фактор, отобранный из группы, состоящей из колониестимулирующего фактора, цитокина, лимфокина, интерлейкина;
при том, что L1 является линкером, способным связывать R1 с R2;
с оговоркой, что по крайней мере R1 или R2 отбирается из полипептидов согласно формулам (I), (II) или (III); и
произвольно упомянутый гемопоэтический белок может быть немедленно предвосхищен (метионином-1), (аланином-1) или (метионином-2, аланином-1).
Наиболее удобными точками разрыва, по которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (I), следующие: 23-24, 24-25, 25-26, 27-28, 28-29, 29-30, 30-31, 31-32, 32-33, 33-34, 34-35, 35-36, 36-37, 37-38, 38-39, 40-41, 41-42, 42-43, 52-53, 53-54, 54-55, 55-56, 77-78, 78-79, 79-80, 80-81, 81-82, 82-83, 83-84, 84-85, 85-86, 86-87, 109-110, 110-111, 111-112, 112-113, 113-114, 114-115, 115-116, 116-117, 117-118, 118-119, 119-120, 120-121, 121-122, 122-123, 123-124, 124-125, 125-126, 126-127, 127-128, 128-129, 129-130, 130-131 и 131-132.
Самыми удобными точками разрыва, по которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (I), следующие: 23-24, 24-25, 31-32, 32-33, 37-38, 38-39, 82-83, 83-84, 85-86, 86-87, 87-88, 125-126, 126-127 и 131-132.
Агонисты ЕРО-рецептора согласно настоящему изобретению могут включать аминокислотные замены, такие, какие описаны в патенте WO 94/24160, или в одном или большем числе сайтов гликозилирования (Asn24, Asn83 и Asn126) с заменами на другие аминокислоты, причем без ограничения аспарагиновой или глутаминовой кислотами, а также на делеции и (или) вставки. Также имеется в виду, что агонисты ЕРО-рецептора согласно настоящему изобретению также могут включать делеции аминокислот как в обоих N- и С-концах исходного полипептида, так и делеции в новых N- и (или) С-концах последовательности модифицированного полипептида согласно формулам, приведенным выше.
Наиболее удобными точками разрыва, по которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (II), следующие: 23-24, 24-25, 25-26, 33-34, 34-35, 35-36, 36-37, 38-39, 39-40, 40-41, 64-65, 65-66, 66-67, 67-68, 68-69, 69-70, 70-71, 89-90, 90-91, 91-92, 92-93, 93-94, 94-95, 95-96, 96-97, 97-98, 98-99, 99-100, 100-101, 101-102, 102-103, 103-104, 104-105 и 105-16, соответственно.
Самыми удобными точками разрыва, по которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (II), следующие: 64-65, 65-66, 92-93 и 93-94, соответственно.
Наиболее удобными точками разрыва, по которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (III), следующие: 36-37, 37-38, 39-40, 41-42, 42-43, 64-65, 65-66, 66-67, 86-87, 87-88, 88-89, 89-90, 90-91, 91-92, 92-93, 93-94, 94-95, 95-96, 96-97, 97-98, 98-99, 99-100 и 100-101.
Самыми удобными точками разрыва, по которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (III), следующие: 39-40, 65-66, 89-90, 99-100 и 100-101.
Наиболее удобными точками разрыва, по которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (IV), следующие: 38-39, 39-40, 40-41, 41-42, 48-49, 53-54, 54-55, 55-56, 56-57, 57-58, 58-59, 59-60, 60-61, 61-62, 62-63, 64-65, 65-66, 66-67, 67-68, 68-69, 69-70, 96-97, 125-126, 126-127, 127-128, 128-129, 129-130, 130-131, 131-132, 132-133, 133-134, 134-135, 135-136, 136-137, 137-138, 138-139, 139-140, 140-141 и 141-142.
Самыми удобными точками разрыва, по которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (IV), следующие: 38-39, 48-49, 96-97, 125-126, 132-133 и 141-142.
Наиболее удобными точками разрыва, по которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (V), следующие: 28-29, 29-30, 30-31, 31-32, 32-33, 33-34, 34-35, 35-36, 36-37, 37-38, 38-39, 39-40, 66-67, 67-68, 68-69, 69-70, 70-71, 84-85, 85-86, 86-87, 87-88, 88-89, 89-90, 90-91, 98-99, 99-100, 100-101 и 101-102.
Самыми удобными точками разрыва, о которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (V), следующие: 34-35, 69-70 и 90-91.
Наиболее удобными точками разрыва, по которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (VI), следующие: 80-81, 81-82, 82-83, 83-84, 84-85, 85-86, 86-87, 108-109, 109-110, 110-111, 111-112, 112-113, 113-114, 114-115, 115-116, 116-117, 117-118, 118-119, 119-120, 120-121, 121-122, 122-123, 123-124, 124-125, 125-126 и 126-127.
Самыми удобными точками разрыва, по которым новые С- и N-концы могут быть созданы в приведенном выше полипептиде (VI), следующие: 81-82, 108-109, 115-116, 119-120, 122-123 и 125-126.
Многофункциональный агонист рецепторов согласно настоящему изобретению также может быть представлен следующими формулами:
(Т1)a-(L1)b-X1-(L)c-X2-(L2)d-(Т2)е
или
X1-(L)c-X2-(L)-Y1-(L)c-Y2
в которых:
X1 - пептид, состоящий из аминокислотной последовательности, соответствующей последовательности остатков n+1 через J исходного полипептида, в котором аминокислотные остатки пронумерованы последовательно от 1 через J с N-концом по остатку 1;
L - произвольный линкер;
X2 - пептид, состоящий из аминокислотной последовательности остатков от 1 до n в исходном полипептиде;
Y1 - пептид, состоящий из аминокислотной последовательности, соответствующей последовательности остатков n=1 через К исходного полипептида, включающего аминокислотные остатки, пронумерованные последовательно от 1 через К с N-концом в остатке 1;
Y2 - пептид, состоящий из аминокислотной последовательности остатков от 1 до n исходного полипептида;
L1 и L2 - произвольные пептидные спейсеры;
n - целое число от 1 до J-1;
b, c и d - независимо принимают значения 0 или 1;
а и е - также или 0, или 1 при том, что оба этих числа не могут быть одновременно 0, и
Т1 и Т2 - белки.
Многофункциональные химерные агонисты гемопоэтических рецепторов согласно настоящему изобретению могут включать аминокислотные замены, делеции и (или) вставки в отдельных полипептидных компонентах химерной молекулы. Также имеется в виду, что многофункциональные химерные агонисты гемопоэтических рецепторов согласно настоящему изобретению могут также вести делеции аминокислот как в обоих N- и С-концах исходного белка, так и делеции в новых N- и (или) С-концах последовательностей модифицированных полипептидов согласно формулам, приведенным выше.
Предпочтительное формирование в настоящем изобретении линкера (L), (L1) или (L2), упоминавшихся в приведенных выше формулах, соединяющих N-конец с С-концом, должно соответствовать полипептиду, выбираемому из группы, включающей:
Ser;
Asn;
Gly;
Thr;
GlySer;
AlaAla;
GlySerGly;
GlyGlyGly;
GlyAsnGly;
GlyAlaGly;
GlyThrGly;
AlaSerAla;
AlaAlaAla;
GlyGlyGlySer SEQ ID NO 778;
GlyGlyGlySerGlyGlyGlySer SEQ ID NO 779;
GlyGlyGlySerGlyGlyGlySerGlyGlyGlySer SEQ ID NO 780;
SerGlyGlySerGlyGlySer SEQ ID NO 781;
GluPheGlyAsnMet SEQ ID NO 782;
GluPheGlyGlyAsnMet SEQ ID NO 783;
GluPheGlyGlyAsnGlyGlyAsnMet SEQ ID NO 784;
GlyGlySerAspMetAlaGly SEQ ID NO 785;
SerGlyGlyAsnGly SEQ ID NO 786;
SerGlyGlyAsnGlySerGlyGlyAsnGly SEQ ID NO 787;
SerGlyGlyAsnGlySerGlyGlyAsnGlySerGlyGlyAsnGly SEQ ID NO 788;
SerGlyGlySerGlySerGlyGlySerGly SEQ ID NO 789;
SerGlyGlySerGlySerGlyGlySerGlySerGlyGlySerGly SEQ ID NO 790;
GlyGlyGlySerGlyGly SEQ ID NO 791;
GlyGlyGlySerGlyGlyGly SEQ ID NO 792;
GlyGlyGlySerGlyGlyGlySerGlyGly SEQ ID NO 793;
GlyGlyGlySerGlyGlyGlySerGlyGlyGlySerGly SEQ ID NO 794;
GlyGlyGlySerGlyGlyGlySerGlyGlyGlySerGlyGlyGly SEQ ID NO 795;
GlyGlyGlySerGlyGlyGlySerGlyGlyGlySerGlyGlyGlySerGlyGlyGlySerGly SEQ ID NO 796;
ProProProTrpSerProArgProLeuGlyAlaThrAlaProThrAlaGlyGlnProProLeu SEQ ID NO 797;
ProProProTrpSerProArgProLeuGlyAlaThrAlaProThr SEQ ID NO 798; и
ValGluThrValPheHisArgValSerGlnAspGlyLeuLeuThrSer SEQ ID NO 799.
Кроме того, настоящее изобретение представляет рекомбинантные экспрессирующие векторы, включающие нуклеотидные последовательности, кодирующие многофункциональные химерные агонисты гемопоэтических рецепторов, необходимые микроорганизменные экспрессионные системы и процессы продуцирования многофункциональных химерных агонистов гемопоэтических рецепторов. Изобретение также представляет фармакологические препараты, содержащие многофункциональные химерные агонисты гемопоэтических рецепторов, и способы использования многофункциональных химерных агонистов гемопоэтических рецепторов.
Вдобавок к использованию многофункциональных химерных агонистов гемопоэтических рецепторов согласно настоящему изобретению in vivo, считается, что применение In vitro могло бы основываться на способности стимулировать активацию и рост клеток костного мозга и крови перед тем, как проводить переливание крови пациентам. Другое предполагаемое использование связано с продуцированном дендритных клеток как in vivo, так и ех vivo.
Считается, что сниженная аффинность химерных белков обусловливается (по крайней мере, отчасти) неспособностью отдельных компонентов формировать свою нативную пространственную структуру в случае, когда они встраиваются в состав химерной молекулы, или возникновением пространственных помех активным сайтам в составе отдельных компонентов химерного белка. Данное изобретение преодолевает эти ограничения, предлагая новые многофункциональные химерные агонисты гемопоэтических рецепторов, которые характеризуются аффинностью по связыванию, сравнимой или даже превышающей таковую у отдельных компонентов химерной молекулы.
Краткое описание чертежей
Фиг.1 схематически иллюстрирует перестройки последовательности полипептида. N-конец (N) и С-конец (С) исходного полипептида соединены через линкер или соединены друг с другом напрямую. Белок "открыт" по точке разрыва, что создает новый N-конец (новый N) и новый С-конец (новый С): в результате образуется белок с новой первичной последовательностью аминокислот. Преобразованная молекула может быть синтезирована de novo как линейная молекула без прохождения этапов соединения исходных N-конца и С-конца и "раскрытия" полипептида в точке разрыва.
Фиг.2 показывает схематически способ I, предназначенный для создания новых белков, в которых исходные N-конец и С-конец нативного полипептида соединены с линкером, в результате чего возникают отличающиеся N-конец и С-конец. На показанном примере перестройка последовательности приводит к появлению нового гена, кодирующего белок, в котором новый N-конец создается по 97-му аминокислотному остатку исходного полипептида, исходный С-конец (остаток 174) соединяется с 11-й аминокислотой (аминокислоты 1-10 делетируются) через линкерный сегмент и новый С-конец создается по 96-й аминокислоте исходной последовательности.
Фиг.3 показывает схематически способ II, применяемый для создания новых белков, в которых исходные N-конец и С-конец нативного полипептида соединяются друг с другом без линкера с возникновением отличающихся N-конца и С-конца белка. На показанном примере перестройка последовательности приводит к образованию нового гена, кодирующего белок с новым N-концом, созданным по 97-му остатку исходного полипептида, исходным С-концом (остаток 174), присоединенным к исходному N-концу, и новым С-концом, образованным по 96-й аминокислоте исходной последовательности.
Фиг.4 схематически показывает способ III, применяемый для создания новых белков, в которых исходные N-конец и С-конец нативного полипептида соединены с помощью линкера с образованием отличающихся N-конца и С-конца полипептида. На показанном примере перестройка последовательности приводит к образованию гена, кодирующего белок с новым N-концом, образованным по 97-му остатку исходного белка, исходным С-концом (остаток 174), присоединенным к 1-й аминокислоте через линкерный сегмент, и новым С-концом, образованным в 96-й аминокислоте исходной последовательности.
Фиг.5 показывает биологическую активность многофункциональных агонистов рецепторов, включая агонисты рецептора flt3 pMON32332, pMON32333, pMON32334 и pMON32335, в сравнении с рекомбинантным нативным flt3 (Genzyme) в тесте на пролиферацию клеток MUTZ-2. МТ = имитация трансфекции.
Фиг.6 показывает последовательность ДНК, кодирующую зрелый ЕРО, основываясь на последовательности, опубликованной Lin et al. (1085, Proc. Natl. Acad. Sci. USA, 82, 7580-7584).
Фиг.7а и 7b показывают последовательность ДНК, кодирующую нативный фактор стволовых клеток, основываясь на последовательности, опубликованной Martin et al. (1990, Cell, 63, 203-211).
Фиг.8 показывает последовательность ДНК, кодирующую растворимый (цитоплазматический) фактор стволовых клеток, основываясь на последовательности, опубликованной Langley et al. (1994, Arch. Biochem. Biophys., 311, 55-61).
Фиг.9а и 9b показывают последовательность ДНК, кодирующую 209 аминокислот из состава активного лиганда flt3, согласно Lyman et al. (1995, Oncogene, 11, 1165-1172).
Фиг.10 показывает последовательность ДНК, кодирующую 134 аминокислоты растворимой (цитоплазматической) формы лиганда flt3, согласно Lyman et al., (1995, Oncogene, 11, 1165-1172).
Подробное описание изобретения
Настоящее изобретение представляет многофункциональные химерные агонисты гемопоэтических рецепторов, создаваемые путем ковалентного связывания полипептидов, каждый из которых может быть активен опосредованно через различные и специфичные клеточные рецепторы, инициируя комплементарные формы биологической активности. Кроветворение включает сложный комплекс клеточных процессов, в которых стволовые клетки непрерывно генерируют большие популяции созревающих клеток всех основных типов. В настоящее время известно по крайней мере 20 регуляторов пролиферационной активности в процессах кроветворения. Большинство этих регуляторов пролиферации могут стимулировать только тот или иной тип формирующихся колоний in vitro, при том, что детальные параметры формирования колоний, стимулируемых каждым из регуляторов, отчетливо различаются. Нет двух таких регуляторов, которые бы проявляли одинаковые параметры формирования колоний, оцениваемые по числу колоний или, что более важно, по типу дифференцирующихся клеток и параметрам их дифференцировки (созревания). Наиболее удобный способ изучения контроля пролиферации связан с использованием простых культуральных систем In vitro. Три достаточно четко различающиеся характеристики могут быть определены: изменение размера колонии, изменение числа колоний и тип клеток. Два или более факторов могут влиять на клетку-предшественник, инфицируя формирование большей величины потомства, т.е. увеличение размера колонии. Два или более факторов могут обусловливать увеличение числа клеток-предшественников, у которых инициируется пролиферация, поскольку существуют различные группы клеток-предшественников, которые реагируют исключительно на один из факторов, или потому что некоторые клетки-предшественники нуждаются в стимуляции двумя или большим числом факторов до того, как они становятся способными реагировать. Активация дополнительных рецепторов на клетке путем использования двух или более факторов может, по-видимому, усиливать митотический сигнал за счет объединения исходно дифференцированных сигнальных путей в единый итоговый сигнальный путь, достигающий ядра (Metcalf; 1989, Nature, 339, 27). Другие механизмы могут объяснять явление синергизма. Например, если один из сигнальных путей ограничен посреднической активацией дополнительного сигнального пути, опосредуемой неким вторым фактором, то в результате может достигаться "супераддитивный" ответ. В некоторых случаях активация одного типа рецепторов может индуцировать усиление экспрессии других типов рецепторов (Metcalf, 1993, Blood, 82, 3515-3523). Два или более факторов могут обеспечивать состав клеточных типов, отличающийся от такового при действии единственного фактора. Использование многофункциональных химерных агонистов гемопоэтических рецепторов может иметь существенные клинические преимущества, определяемые характером пролиферативного ответа, что невозможно при использовании отдельных аналогичных факторов.
Рецепторы гемопоэтических и других факторов роста могут быть классифицированы в два отчетливо различающихся семейства родственных белков: 1) тирозинкиназные рецепторы, включая рецепторы эпидермальных факторов роста, M-CSF (Sherr, 1990, Blood, 75, (1) и SCF (Yarden et al., 1987, EMBO J., 6, 3341); и (2) гемопоэтические рецепторы, лишенные тирозинкиназного домена, но проявляющие существенное сходство с их внеклеточным доменом (Bazan, 1990, Proc. Natl. Acad. Sci. USA, 87, 6934-6938). В эту вторую группу включают эритропоэтин (ЕРО) (D’Andrea et al., 1989, Cell, 57, 277), GM-CSF (Gearing et al., 1989, EMBO J., 8, 3667), IL-3 (Kitamura et al., 1991, Cell, 66, 1165), G-CSF (Fukunaga et al., 1990, J. Biol. Chem., 265, 14008-14015), IL-4 (Harada et al., 1990, Proc. Natl. Acad. Sci. USA, 87, 857), IL-5 (Takaki et al., 1990, EMBO'3., 9, 4367), IL-6 (Yamasaki et al., 1988, Science, 241, 825), IL-7 (Goodwin et al., 1990, Cell, 60, 941-951), LIF (Gearing et al., 1991, EMBO J., 10, 2839) и IL-2 (Cosman et al., 1986, Mol.-Immunol., 23, 935-994). У большинства рецепторов, перечисленных в составе последней группы, высокоаффинной формой является гетеродимерная. После связывания лиганда специфическая α-субъединица становится ассоциированной по крайней мере с какой-либо другой субъединицей рецептора (β-субъединицей, γ-субъединицей). Многие из перечисленных факторов включают общие рецепторные субъединицы: α-субъединицы рецепторов GM-CSF, IL-3 и IL-5 связаны с общей β-субъединицей (Kitamura et al., 1991, Cell, 66, 1165; Takaki et al., 1991, EMBO J., 10, 2833-2838), а рецепторные комплексы IL-6, LIF и IL-11 имеют общую β-субъединицу (gр130) (Таgа et al., 1989, Cell, 58, 573-581; Gearing et al., 1992, Science, 255, 1434-1437). Рецепторные комплексы IL-2, IL-4, IL-7, IL-9 и IL-15 имеют общую γ-субъединицу (Kondo et al., 1993, Science, 266, 1042-1045; Noguch et al., 1993, Science, 262, 1877; Giri et al., 1994, EMBO J., 13, 2822-2830).
Использование фактора кроветворения, проявляющего множественные активности, может иметь существенные преимущества за счет снижения потребностей фактор-продуцирующих клеток и их индукционных систем. В случаях наличия ограничений в способности клетки вырабатывать фактор, за счет снижения потребных концентраций каждого из требуемых факторов и за счет использования его в составе какого-либо сочетания может быть достигнуто существенное снижение необходимого числа фактор-продуцирующих клеток. Использование гемопоэтического фактора, обладающего множественными активностями, может снижать количество факторов, которые были бы необходимы, по-видимому, за счет снижения вероятности отрицательных побочных эффектов.
Новые соединения по данному изобретению представляются формулой выбора из группы, состоящей из:
R1-L1-R2, R2-L1-R1 или R1-R2 или R2-R1,
где R1 и R2 определяются так же, как это было описано выше.
Предпочтительно R2 - колониестимулирующий фактор с отличающейся, но комплементарной активностью по отношению к R1. Под комплементарной активностью понимается активность, которая усиливает или изменяет реакцию на какой-либо другой клеточный модулятор. Полипептид R1 присоединяют или напрямую, или через линекрый сегмент к полипепитиду R2. Термин "напрямую" определяет признак многофункциональных химерных агонистов гемопоэтических рецепторов, в которых полипептиды соединены друг с другом без пептидного линкера. Таким образом, L1 представляет химическую связь или полипептидный сегмент, к которому и R1, и R2 присоединены "в пределах рамки"; наиболее часто L1 - линейный пептид, к которому R1 и R2 присоединены амидными связями, соединяющими С-конец R1 с N-концом L1 и С-конец L1 с N-концом R2. Под "соединением в пределах рамки (считывания)" понимается то, что терминации трансляции или разрыва между рамками считывания (кодирующими последовательностями) в ДНК, кодирующей R1 и R2, не происходит.
Неограничивающий перечень других факторов роста, т.е. колониестимулирующих факторов (CSFs), включает: цитокины, лимфокины, интерлейкины, гемопоэтические факторы роста, которые могут быть соединены с (I), (II) или (III), включая GM-CSF, G-CSF, лиганд с-mpl (также известный как ТРО или MGDF), M-CSF, эритропоэтин (ЕРО), IL-1, IL-4, IL-2, IL-3, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL -12, IL-13, IL-15, LIF, лиганд flt3/flt2, гормон роста человека, фактор роста В-клеток, фактор дифференциации В-клеток, фактор дифференциации эозинофилов и фактор стволовых клеток (SCF), также известный как "жесткий" фактор или лиганд c-kit. Кроме того, настоящее изобретение представляет применение модифицированных молекул R1 или R2 или мутированных, или модифицированных последовательностей ДНК, кодирующих эти молекулы R1 или R2. Настоящее изобретение также представляет многофункциональные химерные агонисты гемопоэтических рецепторов, среди которых R1 или R2 могут являться вариантом hIL-3, вариантом лиганда с-mpl или вариантом G-CSF. "Вариант hIL-3" определяется как молекула hIL-3, несущая аминокислотные замены и (или) частичные делеции в соответствии с вариантами, описанными в патентах WO 94/12638, WO 94/12639 и WO 95/00646, а также другие варианты, известные специалистам в данной области техники. "Вариант лиганда с-mpl″ определяется как молекула лиганда с-mpl, который включает аминокислотные замены и (или) частичные делеции в лиганде c-mpl в соответствии с вариантами, описанными в патенте США 08/383035, а также другие варианты, известные специалистам в данной области техники. "Вариант G-CSF" определяется как молекула G-CSF, включающая аминокислотные замены и (или) частичные делеции в G-CSF согласно вариантам, описанным здесь, а также другим вариантам, известным специалистам в данной области техники. В дополнение к вышеприведенному перечню варианты IL-3 согласно патентам WO 94/12639 и WO 94/12638б, агонисты рецептора G-CSF согласно патенту WO 97/12977, агонисты рецептора с-mр1 согласно патенту WO 97/12978, агонисты рецептора IL-3 согласно патенту WO 97/12979 могут быть использованы в качестве R1 или R2 по настоящему изобретению. "Варианты IL-3" относятся к вариантам IL-3 согласно патенту WO 94/12639 и WO 94/12638. В данном тексте "слитые белки" относятся к слитым белкам согласно патенту WO 95/21197 и WO 95/21254. В данном тексте "агонисты рецептора G-CSF" соответствует агонистам рецептора G-CSF, описанным в патенте WO 97/12978. В данном тексте "агонисты рецептора с-mpl" соответствуют агонистам, описанным в патенте WO 87/12978. В данном тексте "агонисты рецептора IL-3" соответствуют агонистам рецептора IL-3, описанным в патенте WO 97/12979. В данном тексте "многофункциональные агонисты рецепторов" соответствуют многофункциональным агонистам рецепторов, описанным в патенте WO 97/12985.
Линкерный сегмент (L1) в принципе является полипептидом, включающим от 1 до 500 аминокислот. Линкеры, соединяющие две молекулы, должны обеспечивать следующее: 1) позволять двум молекулам формировать пространственную структуру и проявлять активность независимо друг от друга; 2) не проявлять склонности к формированию пространственных структур, которые могли бы интерферировать с функциональными доменами двух белков; 3) иметь минимальную гидрофобность в связи со взаимодействием функциональных доменов белков; и 4) обеспечивать пространственное разделение R1 и R2 таким образом, чтобы R1 и R2 могли бы одновременно взаимодействовать с соответствующими им рецепторами в одной и той же клетке. В целом, поверхностные аминокислоты подвижных участков белков - это глицин, аспарагин и серин. Виртуально любая пермутация аминокислотных последовательностей, содержащих эти три аминокислоты, должна предположительно удовлетворять перечисленным выше критериям линкерных последовательностей. Другие нейтральные аминокислоты, такие как треонин и аланин, также могут быть использованы в составе линкерных последовательностей. Дополнительные аминокислоты также могут быть включены в состав линкеров, что позволит добавлять уникальные сайты рестрикции в линкерную последовательность с целью облегчения конструирования многофункциональных химерных агонистов гемопоэтических рецепторов.
Предпочтительные линкеры L1 по настоящему изобретению включают последовательности, отбираемые из группы формул: (Gly3Ser)n (SEQ ID NO 861), (Gly3Ser)n (SEQ ID NO 862), (Gly5Ser)n (SEQ ID NO 863), (GlynSer)n (SEQ ID NO 864) или (AlaGlySer)n (SEQ ID NO 865).
Один из примеров высокоподвижного линкера - это богатый остатками глицина и серина спейсерный участок, присутствующий в белке pIII филаментных бактериофагов, например, бактериофагов М13 или fd (Schaller et al., 1975, Proc. Natl. Acad. Sci. USA, 72, 737-741). Этот сегмент представляет длинный подвижный спейсер, разделяющий два домена поверхностного белка pIII. Этот спейсерный участок включает такую аминокислотную последовательность:
GlyGlyGlySerGlyGlyGlySerGlyGlyGlySerGluGlyGlyGlySerGlu
GlyGlyGlySerGluGlyGlyGlySerGluGlyGlyGlySerGlyGlyGlySer
(SEQ ID NO 800).
Настоящее изобретение также представляет линкеры, в которые включается последовательность, распознаваемая эндопептидазой. Такой сайт расщепления может быть пригоден для разделения отдельных компонентов многофункционального химерного агониста гемопоэтических рецепторов с целью определения того, сформировали ли они нормальную пространственную структуру и активны in vitro. Примеры различных эндопептидаз включают (но ими не ограничиваются) плазмин, энтерокиназу, калликреин, урокиназу, тканевый активатор плазминогена, клострипаин, химозин, коллагеназу, протеазу яда гадюка Расселла, фермент постпролинового расщепления, протеазу V8, тромбин и фактор Ха.
Пептидные линкерные сегменты из участка, называемого "шарниром", в составе тяжелых цепей иммуноглобулинов IgG, IgA, IgM, IgD или IgE, обеспечивают угловые соотношения между соединенными полипептидами. Наиболее применимыми являются те шарнирные сегменты, в которых остатки цистеина замещены остатками серина. Предпочтительные линкеры по настоящему изобретению включают последовательности, производные от "шарнира"-2b γ-субъединицы IgG мыши, в которых остатки цистеина замещены остатками серина.
Эти линкеры также могут включать сайты расщепления эндопептидазами. Примеры таких линкеров включают следующие последовательности:
IleSerGluProSerGlyProIleSerThrlleAsnProSerProProSerLys
GluSerHisLysSerPro (SEQ ID NO 801) и
IleGluGlyArglleSerGluProSerGlyProIleSerThrlleAsnProSer
ProProSerLysGluSerHisLysSerPro (SEQ ID NO 802).
Настоящее изобретение, однако, не ограничается по форме, числу и размеру линкерных последовательностей и только тем требованием, чтобы его функциональность ассоциировалась с отсутствием интерференции с формированием пространственной структуры и активностью отдельных молекул, входящих в состав многофункционального химерного агониста гемопоэтических рецепторов.
Определение линкера L2
Длина аминокислотной последовательности линкера L2, предполагаемого к использованию в R1 и (или) R2, может быть выбрана эмпирическим путем или согласно шаблонам пространственной информации, или с использованием сочетания этих двух подходов.
В случае отсутствия какой-либо информации о структуре небольшие группы линкеров могут быть сформированы для тестирования с использованием схемы, в которой длина варьируется так, чтобы обеспечить интервал 0-50 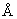 , а последовательность подбирается таким образом, чтобы соответствовать поверхностной активности (гидрофильность; Норр & Woods, 1983, Mol. Immunol., 20, 483-489; Kyte & Doolittle, J. Mol. Biol., 157, 105-132; обработанная растворителями поверхностная зона; Lee & Richards, 1971, J. Mol. Biol., 55, 379-400) и способности адаптировать необходимую конформацию без нарушения конформации R1 или R2 (конформационно подвижные; Karplus & Schulz, 1985, Naturwissenschaften, 72, 212-213). Принимая диапазон 2,0-3,8
, а последовательность подбирается таким образом, чтобы соответствовать поверхностной активности (гидрофильность; Норр & Woods, 1983, Mol. Immunol., 20, 483-489; Kyte & Doolittle, J. Mol. Biol., 157, 105-132; обработанная растворителями поверхностная зона; Lee & Richards, 1971, J. Mol. Biol., 55, 379-400) и способности адаптировать необходимую конформацию без нарушения конформации R1 или R2 (конформационно подвижные; Karplus & Schulz, 1985, Naturwissenschaften, 72, 212-213). Принимая диапазон 2,0-3,8 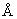 за среднюю трансляции, это может означать, что длина участка тестирования - от 0 до 30 аминокислотных остатков (при предпочтительном диапазоне 0-15 аминокислот). На примере таких эмпирических групп линкеры могут быть сконструированы с использованием кассетной последовательности, такой как "глицин-глицин-глицин-серин", повторенной n раз при том, что n составляет 1, 2, 3 или 4. Анализ известных данных позволяет отметить существование большого числа таких последовательностей, которые варьируются по длине или составу и могут служить в качестве линкеров, изначально принимая во внимание то, что они не являются ни слишком длинными, ни слишком короткими (Sandhu, 1992, Critical Rev. Biotech., 12, 437-462); если же они слишком длинны, то фактор энтропии может дестабилизировать трехмерную организацию, в частности делая пространственную укладку кинетически неэффективной; если же они слишком коротки, то они могут дестабилизировать молекулу из-за торзионного или стерического эффектов.
за среднюю трансляции, это может означать, что длина участка тестирования - от 0 до 30 аминокислотных остатков (при предпочтительном диапазоне 0-15 аминокислот). На примере таких эмпирических групп линкеры могут быть сконструированы с использованием кассетной последовательности, такой как "глицин-глицин-глицин-серин", повторенной n раз при том, что n составляет 1, 2, 3 или 4. Анализ известных данных позволяет отметить существование большого числа таких последовательностей, которые варьируются по длине или составу и могут служить в качестве линкеров, изначально принимая во внимание то, что они не являются ни слишком длинными, ни слишком короткими (Sandhu, 1992, Critical Rev. Biotech., 12, 437-462); если же они слишком длинны, то фактор энтропии может дестабилизировать трехмерную организацию, в частности делая пространственную укладку кинетически неэффективной; если же они слишком коротки, то они могут дестабилизировать молекулу из-за торзионного или стерического эффектов.
Проводившиеся исследования структуры белков позволяют установить, что расстояние между концами полипептидной цепи, определяемое как расстояние между Сα-углеродами, может быть использовано для определения длины последовательности, предполагаемой к применению, или по крайней мере для ограничения числа возможных вариантов, которые должны быть тестированы при эмпирическом отборе линкеров. Имеющиеся данные также позволяют определить, что в некоторых случаях положения концов полипептидной цепи ошибочно маркируются в структурных моделях, создаваемых с помощью анализа дифракции рентгеновских лучей или методом ядерно-магнитной резонансной спектроскопии; в случае же точных оценок такая ситуация должна учитываться в дальнейшем с целью точной оценки длины необходимого линкера. Из тех аминокислотных остатков, положение которых точно определено, выбирают два остатка, которые примыкают к концам последовательности, расстояние между их атомами Са используют для расчета приблизительной длины линкера между ними. С использованием расчетной длины как шаблона отбирают линкеры с определенным диапазоном числа остатков (рассчитываемого исходя из того, что на один остаток приходится 2-3,8 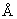 ). Такие линкеры могут быть составлены оригинальными последовательностями, по необходимости укороченными или удлиненными – при удлинении дополнительные остатки могут быть выбраны так, чтобы обеспечить подвижность и гидрофильность так, как это было описано выше; или произвольно исходная последовательность может быть заменена с целью использования группы линкеров, например, имеющих повтор "глицин-глицин-глицин-серин" согласно описанному выше подходу; или сочетание исходной последовательности и новой последовательности, имеющее подходящую общую длину, может быть произвольно использовано.
). Такие линкеры могут быть составлены оригинальными последовательностями, по необходимости укороченными или удлиненными – при удлинении дополнительные остатки могут быть выбраны так, чтобы обеспечить подвижность и гидрофильность так, как это было описано выше; или произвольно исходная последовательность может быть заменена с целью использования группы линкеров, например, имеющих повтор "глицин-глицин-глицин-серин" согласно описанному выше подходу; или сочетание исходной последовательности и новой последовательности, имеющее подходящую общую длину, может быть произвольно использовано.
Определение амино- и карбоксильного концов в R1 и R2
Последовательности R1 и R2, способные образовывать пространственную структуру при биологически активном статусе, могут быть сфоримированы путем отбора начальных (N-концевых) и конечных (С-концевых) положений из состава исходного полипептида и использования линкерной последовательности L2 согласно описанному выше. N- и С-концы выбирают из состава типичной последовательности, связанной с точкой разрыва, с использованием получения шаблонов так, как это было описано выше. Таким образом, новую аминокислотную последовательность создают путем отбора N- и С-концов из одного и того же участка разрыва. Во многих случаях отбор новых концов должен быть таким, чтобы исходное положение С-конца тут же предварялось N-концом. Однако данные, известные специалистам в данной области техники, указывают на то, что отбор концов в каком-либо участке сегмента может эффективно приводить либо к делециям, либо к прибавлениям остатков в N- или С-концевых положениях новой последовательности.
Центральным принципом молекулярной биологии считается то, что первичная аминокислотная последовательность белка определяет формирование пространственной трехмерной структуры, необходимой для проявления его биологических функций. Методы, известные специалистам в этой области техники, применяемые для анализа и интерпретации трехмерной организации - это тестирование дифракции рентгеновских лучей на отдельных белковых кристаллах или ядерно-магнитная резонансная спектроскопия белковых растворов. Примеры структурной информации, которой достаточно для определения точек разрывов, включают положение и характер вторичной структуры белка (спирали α и 3-10, параллельные и антипараллельные β-плоскости, развороты и обороты цепей и петли; Kabsch & Sander, 1983, Biopolymers, 22, 2577-2637), степень сольвентного воздействия на аминокислотные остатки, степень и тип взаимодействий аминокислот друг с другом (Chothia, 1984, Ann. Rev Biochem., 53, 537-572) и статическое и динамическое распределение конформационных элементов вдоль полипептидной цепи (Alber & Mathews, 1987, Methods Enzymol., 154, 511-533). В некоторых случаях известна дополнительная информация о сольвентном воздействии на аминокислотные остатки: один из примеров представляет сайт посттрансляционного прикрепления углеводорода, который является необходимым на поверхности белка. Когда информация по структуре недоступна или трудна для анализа, могут быть также использованы методы анализа первичной аминокислотной последовательности с целью предсказать вторичную и четвертичную структуры белка, сольвентную доступность и встречаемость разворотов и петель. Биохимические методы также иногда могут быть эффективны для эмпирического определения поверхностных воздействий в тех случаях, когда прямые методы структурного анализа неприменимы; например, применение идентификации сайтов разрезания цепи вследствие ограниченного протеолиза с целью определения поверхностных воздействий (Gentile & Salvatore, 1993, Eur. J. Biochem., 218, 603-621).
Таким образом, с использованием либо экспериментально полученной информации о структуре, либо с применением предсказательных методов (например, Srinivisan & Rose, 1995, Proteins: Struct., Function & Genet., 22, 81-99) исходную аминокислотную последовательность тестируют с целью классификации участков по критериям их вовлечения или невовлечения в поддержание вторичной и четвертичной структуры. Известна встречаемость последовательностей в пределах участков, для которых подтверждено участие в периодической вторичной структуре (спирали α и 3-10, параллельные и антипараллельные β-плоскости): эти участки целесообразно избегать. Сходным образом участки аминокислотной последовательности, для которых установлено или предсказывается слабая степень сольвентной чувствительности, с большой вероятностью являются частями т.н. "гидрофобной оси" белка - они также должны избегаться в ходе отбора N- и С-концов. Напротив, те участки, для которых известно или предсказано расположение на поверхности изгибов и петель, а особенно в тех сегментах, для которых известно отсутствие необходимости в обеспечении биологической активности, являются предпочтительными сайтами для локализации концевых сегментов полипептидной цепи. Протяженные сегменты аминокислотной последовательности, считающиеся предпочтительными на основании обозначенных выше критериев, определяют как "сегмент точки разрыва".
Дополнительные пептидные последовательности также могут быть добавлены с целью облегчения очистки или идентификации многофункциональных химерных агонистов гемопоэтических рецепторов (например, полигистидиновый мотив). Высокоантигенный пептид также может быть добавлен, что может обеспечить быстрое тестирование и упрощенную очистку многофункциональных химерных агонистов гемопоэтических рецепторов с применением специфичного моноклонального антитела.
Термины "мутантная аминокислотная последовательность", "мутантный белок", "вариантный белок", "мутеин" или "мутантный полипептид" обозначают полипептид, имеющий аминокислотную последовательность, которая отличается от нативной последовательности из-за аминокислотных делеций, замен или того и другого вместе, или кодируется нуклеотидной последовательностью, умышленно измененной по сравнению с нативной последовательностью. Понятие "нативная последовательность" обозначает аминокислотную или нуклеотидную последовательность, которая идентична таковой у нативной формы (формы дикого типа) гена или белка.
Гемопоэтические факторы роста могут быть охарактеризованы по их способности стимулировать формирование колоний клетками-предшественниками кроветворения у человека. Формируемые колонии включают эритроидные клетки, гранулоциты, мегакариоциты, макрофаги-гранулоциты и их комбинации. Многие гемопоэтические факторы роста обладают способностью восстанавливать функции костного мозга и популяций клеток периферической крови до терапевтически удовлетворительного уровня в исследованиях, вначале проводившихся на обезьянах, а затем на человеке. Многие или даже все эти биологические активности гемопоэтических факторов роста вовлечены в передачу сигналов и высокоаффинное связывание на рецепторах. Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению могут проявлять ценные качества, такие как проявление такой же или даже большей биологической активности в сравнении с сигнальным фактором или такие как увеличенное время полужизни, или такие как снижение числа отрицательных побочных эффектов, или сочетание такие свойств.
Многофункциональные химерные агонисты гемопоэтических рецепторов, которые имеют слабую или вообще лишены агонистической активности, могут быть использованы в качестве антагонистов, в качестве антигенов с целью получения антител для использования их в иммунологии или иммунотерапии, в качестве генетических зондов или в качестве промежуточных продуктов при конструировании других применимых мутеинов hIL-3.
Биологическая активность многофункциональных химерных агонистов гемопоэтических рецепторов по настоящему изобретению может быть определена по синтезу ДНК в фактор-зависимых клеточных линиях или путем подсчета образующихся колоний в тесте с костным мозгом in vitro.
Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению могут улучшать терапевтические показатели в сравнении с моноактивными гемопоэтическими агонистами. Например, некоторые многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению могут иметь сходную или даже большую активность в качестве фактора роста в сравнении с другими гемопоэтическими агонистами при отсутствии сходного или соответственно возрастающего уровня побочных эффектов.
Настоящее изобретение также представляет последовательности ДНК, которые кодируют белки многофункциональных химерных агонистов гемопоэтических рецепторов, последовательности ДНК, которые в существенной степени сходны и проявляют принципиально сходные функции, и последовательности ДНК, которые отличаются от ДНК, кодирующих многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению только ввиду вырожденности генетического кода. Также настоящее изобретение представляет олигонуклеотидные интермедиаты, используемые при создании мутантных ДНК и полипептидов, кодируемых этими олигонуклеотидами.
Методы генетической инженерии в настоящее время в науке стандартизованы (патент США 4935233 и Sambrook et al., 1989, "Molecular cloning: A Laboratory Manual", Cold Spring Harbor Lab.) и могут быть использованы для создания последовательностей ДНК по настоящему изобретению. Одним из таких методов является метод кассетного мутагенеза (Wells et al., 1985, Gene, 34, 315-323), в котором часть кодирующей последовательности в составе плазмиды замещается синтетическими олигонуклеотидами, которые кодируют желательные аминокислотные замены в части гена между двумя маркерными сайтами рестрикции. Пары комплементарных синтетических олигонуклеотидов, кодирующих желательный ген, могут быть сформированы с отжигом друг на друга. Последовательность ДНК в олигонуклеотиде должна кодировать последовательность аминокислот желательного гена за исключением тех, которые замещены и (или) делетированы из состава последовательности.
Плазмидная ДНК может быть обработана отобранными рестриктазами и затем лигирована на прошедшие отжиг олигонуклеотиды. Лигированные миксты могут быть использованы для трансформации компетентных клеток JM101 при маркировании резистентностью к подходящему антибиотику. Отдельные колонии могут быть отсортированы, а плазмидную ДНК можно протестировать рестриктазами или с помощью секвенирования ДНК с целью выявления плазмид, включающих желательный ген.
Клонирование последовательностей ДНК новых многофункциональных гемопоэтических агонистов при том, что по крайней мере одна из них сходна с последовательностью ДНК гена другого колониестимулирующего фактора, может быть сопровождено использованием промежуточных векторов. Как противопоставляемый подход, один из генов может быть клонирован напрямую в вектор, содержащий другой ген. Линкеры и адаптеры могут быть использованы для соединения последовательностей ДНК, равно как для замещения утрачиваемых последовательностей, где сайт рестрикции находится внутри участка, представляющего интерес. Таким образом, генетический материал (ДНК), кодирующий один полипептид, пептидный линкер и другой полипептид, оказывается внесенным в составе пригодного экспрессирующего вектора, который используют для трансформации бактерий, дрожжей, клеток насекомых и клеток млекопитающих. Трансформированный организм выращивают, а белок выделяют с применением стандартных методов. Получаемый продукт является, таким образом, новым белком, который включает колониестимулирующий фактор, соединенный через линкерный сегмент со вторым колониестимулирующим фактором.
С другой стороны, настоящее изобретение представляет плазмидный ДНК-вектор, предназначенный для использования при обеспечении экспрессии этих новых многофункциональных химерных агонистов гемопоэтических рецепторов. Эти векторы включают новые последовательности ДНК, описанные выше, которые кодируют новые полипептиды согласно настоящему изобретению. Подходящие векторы, с помощью которых трансформируют микроорганизмы, способные экспрессировать многофункциональные химерные агонисты гемопоэтических рецепторов, представлены экспрессирующими векторами, включающими нуклеотидные последовательности, кодирующие многофункциональные химерные агонисты гемопоэтических рецепторов, соединенные с последовательностями, регулирующими транскрипцию и трансляцию, которые выбирают исходя из типа используемых клеток-хозяев.
Векторы, инкорпорирующие модифицированные последовательности, как это было описано выше, включены в настоящее изобретение и использовались для получения многофункциональных химерных агонистов гемопоэтических рецепторов. Использованный вектор также включает отобранные регуляторные последовательности, находящиеся в оперативной связи с кодирующими последовательностями ДНК по настоящему изобретению, которые способны направлять репликацию и экспрессию в выбранных клетках-хозяевах.
Как другая часть настоящего изобретения, предложен способ получения новых многофункциональных химерных агонистов гемопоэтических рецепторов. Способ согласно настоящему изобретению включает культивирование подходящих клеток или клеточных линий, которые были трансформированы с использованием вектора, включающего кодирующую ДНК-последовательность, с целью экспрессии нового многофункционального химерного агониста гемопоэтических рецепторов. Подходящими клетками или клеточными линиями вполне могут быть бактериальные клетки. Например, различные штаммы E.coli хорошо известны как клетки-хозяева с точки зрения биотехнологии. Примеры таких штаммов включают штаммы E.coli JM101 (Yanish-Perron et al., 1985, Gene, 33, 103-119) и MON105 (Obukowicz et al., 1992, Appl. Environ. Microbiol., 58, 1511-1523). Также в настоящее изобретение включен способ экспрессии белка многофункционального химерного агониста гемопоэтических рецепторов с использованием хромосомного экспрессирующего вектора для E.coli, формируемого на основе бактериофага Mu (Weinberg et al., 1993, Gene, 126, 25-33). Различные штаммы сенной палочки B.subtilis также могут быть использованы в данном способе. Многие штаммы клеток дрожжей, как известно специалистам в данной области техники, также являются удобными клетками-хозяевами для экспрессии полипептидов согласно настоящему изобретению. При экспрессии в цитоплазме клеток E.coli ген, кодирующий многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению, также может быть сконструирован таким образом, чтобы с 5’-конца ген был удлинен за счет кодонов, кодирующих Met-2Ala-1 или Met-1 с N-конца кодируемого полипептида. Образующиеся в цитоплазме кишечной палочки N~-концы белков подвергаются посттрансляционному процессингу с участием метионинаминопептидазы (Ben Bassat et al., 1987, J. Bacteriol., 169, 751-757) и, вероятно, других пептидаз: соответственно даже в случае экспрессии метионин будет от N-конца отщеплен. Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению могут включать полипептиды многофункциональных химерных агонистов гемопоэтических рецепторов, имеющих с N-конца дополнительные аминокислоты Met-1, Ala-1 или Met-2Ala-1. Такие мутантные многофункциональные химерные агонисты гемопоэтических рецепторов могут также быть экспрессированы в E.coli путем слияния с сигнальным фрагментом секретируемого белка с N-конца. Этот сигнальный сегмент отщепляется от полипептида на одном из этапов процесса секреции. Дополнительные методы достижения высокоактивной экспрессии генов в клетках E.coli могут быть взяты из C.M.Savvas (1996, Microbiol. Rev., 60, 512-538).
Также согласно настоящему изобретению пригодными являются клетки млекопитающих, такие как клетки яичников китайского хомячка (СНО). Базовые методы экспрессии чужеродных генов в клетках млекопитающих обобщены в обзоре Кауфмана (R.J.Kaufman, 1987, "Genet. Engineering, Principles & Methods", Vol.9, ed. K.Setlow, Plenum Press, New York). Экспрессирующий вектор конструируют таким образом, чтобы в его составе присутствовал мощный промотор, способный проявлять функциональную активность в клетках млекопитающих и обеспечивать транскипцию сегмента, кодирующего эукариотический секреторный сигнальный сегмент, который на трансляционном уровне присоединяют к сегменту, кодирующему многофункциональные химерные агонисты гемопоэтических рецепторов. Например, могут быть использованы такие плазмиды, как pcDNA, I/Neo, pRc/RSV и pRc/CMV (производимые фирмой Invitrogen Corp., San Diego, CA). Сегмент, кодирующий эукариотический секреторный сигнальный сегмент, может быть выделен из состава гена как такового или из состава другого секретируемого белка млекопитающего (Bayne et al., 1987, Proc. Natl. Acad. Sci. USA, 84, 2638-2642). По завершении формирования вектора, включающего ген, векторной ДНК трансфицируют клетки млекопитающих. Такими клетками могут быть, например, клеточные линии COS-7, HeLa, ВНК, СНО или линия L мыши. Клетки можно культивировать, например, в среде DMEM (JRH Scientific). Секретируемый в культуральную среду полипептид может быть выделен с помощью стандартных биохимических приемов после перемежающейся экспрессии на протяжении 24-72 часов после трансфекции клеток или после формирования устойчивой клеточной линии вслед за отбором на резистентность к антибиотикам. Выбор подходящих клеток-хозяев млекопитающих и методов трансформации, культивирования, амплификации, скрининга, получения рекомбинантного продукта и его очистки хорошо известны: см., например, (Gething & Sambrook, 1981, Nature, 293, 620-625) или (Kaufman et al., 1985, Mol. Cell. Biol., 5, 1750-1759) или (Howley et al., патент США 4419446).
Другой подходящей линией клеток млекопитающих является линия клеток COS-1 зеленой мартышки. Используемой со сходной частотой линией клеток млекопитающих является клеточная линия CV-1.
При желании клетки насекомых могут быть использованы в качестве клеток-хозяев по настоящему изобретению. Например, можно обратиться к (Miller et al., 1986, Genet. Eng., 8, 277-298) и к ссылкам, приведенным здесь. К тому же, основные методы экспрессии чужеродных генов в клетках насекомых с использованием векторов на основе бакуловирусов описаны у Summers & Smith (1987, "A manual of methods for Baculovirus vectors and insect cell culture procedures, Texas Agricult. Exper. Stat. Bull No.1555). Экспрессирующий вектор формируют на основе бакуловирусного трансформационного вектора, в составе которого мощный бакуловирусный промотор (такой как промотор полиэдрина) обеспечивает транскрипцию сегмента, кодирующего эукариотический секреторный сигнальный сегмент, который на трансляционном уровне присоединяется к участку, кодирующему полипептид многофункционального химерного агониста гемопоэтических рецепторов. Например, может быть использована плазмида pVL1392 (производимая Invitrogen Corp., San Diego, СА). После конструирования вектора, несущего ген, кодирующий полипептид многофункционального химерного агониста гемопоэтических рецепторов, два микрограмма такой ДНК ко-трансфицируют с 1 микрограммом бакуло вирусной ДНК (см. Summers & Smith, 1987) в клетки насекомого линии SF9. Чистый рекомбинантный бакуловирус, несущий ген многофункционального химерного агониста гемопоэтических рецепторов, используют для заражения клеток, культивируемых, например, в не содержащей сыворотку среде Ecell-401 (JRH Bioscience, Lenexa, KS). Секретируемый в культуральную среду многофункциональный химерный агонист гемопоэтических рецепторов может быть выделен с использованием стандартных биохимических методов. Надосадочная фракция клеток млекопитающих или насекомых, экспрессирующих полипептид многофункционального химерного агониста гемопоэтических рецепторов, может быть повышена в концентрации с применением ряда стандартных концентрационных реактивов.
Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению могут быть использованы при лечении заболеваний, характеризующихся снижением уровней миелоидных, эритроидных, лимфоидных или мегакариоцитных клеток в системе кроветворения или их сочетаний. Вдобавок они могут быть использованы для активации зрелых миелоидных и (или) лимфоидных клеток. Среди состояний, восприимчивых к такому воздействию полипептидов по настоящему изобретению, лейкопения, т.е. снижение количества лейкоцитов (клеток белой крови) в циркулирующей крови. Лейкопения может индуцироваться облучением и вызываться некоторыми вирусами. Часто она оказывается побочным проявлением различных форм противоопухолевой терапии (хемотерапевтического лечения или облучения), инфекций или кровоизлияний. Терапия лейкопении с использованием многофункциональных химерных агонистов гемопоэтических рецепторов по настоящему изобретению может обеспечить исключение нежелательных побочных эффектов, возникающих при применении обозначенных выше лекарственных препаратов.
Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению могут быть использованы при терапии нейтропении и, например, при терапии таких патологий, как апластическая анемия, циклическая нейтропения, идиопатическая нейтропения, синдром Чедиак-Хигаши, системная красная волчанка (SLE), лейкоз, миелодиспластический синдром и миелофиброз.
Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению могут быть использованы в терапии или профилактике тромбоцитопении. В настоящее время единственным способом лечения тромбоцитопении является переливание тромбоцитов, что очень дорого и сопряжено с высоким риском инфекций (HIV, HBV) и аллоиммунизации. Многофункциональный химерный агонист гемопоэтических рецепторов может полностью заменить или существенно сократить потребность в переливании тромбоцитов. Тяжелая тромбоцитопения может быть следствием некоторых генетических дефектов - таких как анемия Фанкони, синдром Вискотта-Олдрича или синдром Мэй Хегглин. Приобретенная тромбоцитопения может быть результатом образования ауто- или аллоантител, например, при пурпурной иммунотромбоцитопении, системной красной волчанке, гемолитической анемии или несовместимости матери и плода. К тому же спленомегалия, рассеянная внутрисосудистая сворачиваемость крови, пурпурная тромбозная тромбоцитопения, инфекция или наличие протеза сердечного клапана могут приводить к тромбоцитопении. Тяжелая тромбоцитопения может также быть следствием хемотерапии и (или) радиотерапии опухолей. Тромбоцитопения также может быть результатом инвазий костного мозга при карциномах, лимфомах, лейкозах или фиброзах.
Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению могут быть использованы для мобилизации гемопоэтических предшественников и стволовых клеток в периферической крови. Для производных от периферической крови предшественников подтверждена эффективность в терапии пациентов с применением аутологичной пересадки костного мозга. Для гемопоэтических факторов роста, включая G-CSF и GM-CSF, было подтверждено участие в повышении числа циркулирующих предшественников и стволовых клеток в периферической крови. Это существенно упрощает процедуру сбора периферических стволовых клеток и резко снижает стоимость процедуры за счет уменьшения необходимого числа взятий проб. Многофунциональный химерный агонист гемопоэтических рецепторов может быть использован для мобилизации стволовых клеток и дальнейшего усиления эффективности трансплантации периферических стволовых клеток.
Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению также могут быть использованы для экспансии ex vivo предшественников кроветворения и стволовых клеток. Колониестимулирующие факторы (CSFs), такие как hIL-3, вводят отдельно, вместе с другими CSFs или в сочетании с трансплантатами костного мозга после высоких химиотерапевтических доз при лечении нейтропении и тромбоцитопении, которые часто являются следствием такой терапии. Однако период тяжелой нейтропении и тромбоцитопении может быть полностью не преодолен. Миелоидная линия клеток, состоящая из моноцитов (макрофагов), гранулоцитов (включая нейтрофилы) и мегакариоцитов, является критической по предотвращению инфекций и кровотечений, которые могут представлять угрозу для жизни. Нейтропения и тромбоцитопения могут быть также следствием заболевания, генетической аномалии, воздействия лекарств, ядов, облучения и многих типов терапевтических воздействий, таких как традиционная противоопухолевая терапия.
Для лечения обозначенной группы пациентов применяют пересадку костного мозга. Однако использование костномозговой ткани для возрождения системы кроветворения связано с рядом проблем: 1) число стволовых клеток в костном мозге, селезенке или периферической крови ограничено; 2) эффект "противоборства" донорской ткани и организма-реципиента, реакция "трансплантат против хозяина" (Graft vs. Host Disease); 3) отторжение трансплантата, и 4) возможная контаминация опухолевыми клетками. Стволовые клетки составляют незначительную часть среди ядерных клеток костного мозга, селезенки и периферической крови. Ясным представляется наличие зависимости от дозы - таким образом, что большее число стволовых клеток обеспечивает усиление восстановления кроветворной функции. Кроме того, экспансия стволовых клеток in vitro должна усиливать восстановление кроветворения. Для приготовления костномозгового трансплантата используют костный мозг, взятый от аллогенного донора. Однако факторы несоответствия донора и реципиента (Graft vs. Host Disease) и отторжения ткани ограничивают возможности пересадки костного мозга, причем даже в тех случаях, когда донорами являются идентичные по HLA пациенты. Альтернативой аллогенной пересадке костного мозга является аутологическая пересадка. При аутологической пересадке костномозговых трансплантатов у пациента отбирают порцию собственного костного мозга перед проведением какого-либо лечения (например, при хемотерапии миелоидных заболеваний с использованием высоких доз) и затем возвращают ее "обратно" по окончании курса лечения. Аутологическая трансплантация исключает риск Graft vs. Host Disease и отторжения ткани. Однако аутологическая пересадка костного мозга имеет свои проблемы, связанные с ограниченным числом стволовых клеток в костном мозге и с возможностью контаминации опухолевыми клетками. Фактор ограниченного количества стволовых клеток может быть преодолен путем экспансии стволовых клеток ex vivo. К тому же стволовые клетки могут быть изолированы специфическим образом (на основе присутствия специфических поверхностных антигенов, таких как CD34+), что позволяет снизить возможный риск контаминации костномозгового трансплантата опухолевыми клетками.
Перечисленные ниже патенты включают подробную информацию о разделении стволовых клеток, клетках CD34+, культивировании клеток с гемопоэтическими факторами, использовании клеток в терапии пациентов с заболеваниями кроветворения и использовании гемопоэтических факторов для экспансии клеток и генотерапии.
Патент США 5061620 представляет композиции гемопоэтиеских стволовых клеток человека, получаемые путем отделения стволовых клеток от маркированных клеток.
Патент США 5199942 описывает способ аутологической пересадки кроветворных клеток, включающий: (1) выделение клеток-предшественников кроветворения у пациента; (2) экспансия ех vivo в клетки фактора роста, выбираемого из группы, состоящей из IL-3, лиганда flt3, лиганда c-kit, GM-CSF, IL-1, химерного белка GM-CSF/IL3 или их комбинаций; (3) введение пациенту клеточного препарата.
Патент США 5240856 представляет клеточный сепаратор, являющийся приспособлением для автоматического контроля процесса разделения клеток.
Патент WO 91/16116 описывает устройства и способы селективного выделения и сепарирования клеток-мишеней из смеси клеток.
Патент WO 91/18972 описывает способы культивирования in vitro костного мозга с помощью инкубирования суспензии костномозговых клеток в биореактивных полых трубках.
Патент WO 92/18615 представляет процесс поддержания и роста числа клеток костного мозга с использованием культуральной среды, содержащей специфическую смесь цитокинов, с последующим использованием в трансплантатах.
Патент WO 93/08268 описывает способ селективного роста числа стволовых клеток, включающий этапы (а) отделения стволовых клеток, несущих антиген CD34+, от других клеток и (и) инкубирования отделенных клеток в селективной среде, что позволяет селективно увеличивать количество выбранных стволовых клеток.
Патент WO 93/18136 описывает процесс поддержания in vitro клеток млекопитающих, производных от периферической крови.
Патент WO 93/18648 представляет композицию, состоящую из клеток-предшественников нейтрофилов человека с высоким содержанием миелобластов и промиелоцитов, используемую для терапии врожденной и приобретенной нейтропении.
Патент WO 94/08039 описывает способ обогащения гемопоэтических стволовых клеток человека с помощью отбора клеток, экспресирющих белок c-kit.
Патент WO 94/11493 описывает популяцию стволовых клеток, несущих антиген CD34+ и имеющую небольшой размер, которую выделяют с применением метода противоточного отцеживания.
Патент WO 94/27698 представляет способ, сочетающий иммуноаффинное разделение и разделение центрифугированием в непрерывном потоке, предназначенный для селективного отбора популяции ядерных гемопоэтических клеток из разнородной клеточной смеси.
Патент WO 94/25848 описывает аппарат для разделения клеток, используемый для сбора заданных клеток с последующими манипуляциями ими.
Долговременное культивирование высокообогащенных несущих антиген CD34+ предшественников гемопоэтических клеток-предшественников, выделенных из костного мозга человека, в культурах, содержащих IL-1a, IL-3, IL-6 или GM-CSF, описано Брандтом с соавт. (Brandt et al., 1990, У. Clin. Invest., 86, 932-941).
Один из аспектов настоящего изобретения связан с описанием способа селективной экспансии стволовых клеток ex vivo. Термин "стволовая клетка" обозначает тотипотентные гемопоэтические клетки, а также ранние клетки-предшественники (недифференцированные клетки), которые могут быть выделены из костного мозга, селезенки или периферической крови. Термин "экспансия" обозначает весь комплекс процессов дифференциации и пролиферации клеток. Настоящее изобретение представляет способ селективной экспансии ех vivo, включающий следующие этапы: (а) отделение стволовых клеток от других клеток; (b) культивирование упомянутых сепарированных стволовых клеток в селективной среде, содержащей белок (белки) многофункционального химерного агониста гемопоэтических рецепторов, и (с) сбор упомянутых стволовых клеток. Стволовые клетки, равно как и клетки-предшественники, которым предназначено стать нейтрофилами, эритроцитами, тромбоцитами и т.п., могут быть отделены от большинства других типов клеток по присутствию-отсутствию частных антигенных маркеров, таких как CD34, которые находятся на поверхности этих клеток, а также по морфологическим признакам. Фенотип высокобогащенной фракции стволовых клеток человека описывается как CD34+, Thy-l+, lin-; однако должно быть понятно, что настоящее изобретение не ограничивается экспансией такой популяции стволовых клеток. Высокообогащенная фракция стволовых клеток человека, маркированная CD34+, может быть отделена с помощью ряда ранее описывавшихся методов, включая аффинные колонки, магнитные шарики или проточную цитометрию с использованием антител к поверхностным антигенам, таким как CD34+. Кроме того, методы физического разделения, такие как процеживание в противоположных потоках, могут быть использованы для обогащения фракции гемопоэтических предшественников. Предшественники с CD34+ гетерогенны и могут быть разделены на несколько субпопуляций, характеризующихся присутствием или отсутствием коэкспрессии различных типов молекул, ассоциирующихся клеточными поверхностями. Самые незрелые (недифференцированные) клетки-предшественники не экспрессируют никакие из известных маркеров, ассоциированных с клеточными типами, такие как HLA-DR или CD38, но они могут экспрессировать CD90 (thy-1). Другие поверхностные антигены, такие как CD33, CD38, CD41, CD71, HLA-DR или c-kit, также могут быть использованы для селективного выделения гемопоэтических предшественников. Отделенные клетки можно инкубировать в селективной среде в культуральных колбах, стерильных пакетах или полых трубках. Различные колониестимулирующие факторы могут быть использованы для контроля селективной экспансии клеток. Подходящие для контроля экспансии костного мозга ех vivo факторы включают лиганд c-kit, IL-3, G-CSF, GM-CSF, IL-1, IL-6, IL-11, лиганд flt-3 или их сочетание. Пролиферацию стволовых клеток можно контролировать с помощью подсчета числа стволовых клеток и других клеток с помощью стандартных методик (например, с помощью гемоцитометра, CFU, LTCIC) или проточной цитометрии до и после инкубации.
Ряд методов экспансии стволовых клеток ex vivo были описаны и основываются на группе методов отбора и экспансии, использующих различные колониестимулирующие факторы, включая лиганд c-kit (Brandt et al., 1994, Blood, 83, 1507-1514; McKenna et al., 1995, Blood, 86, 3413-3420), IL-3 (Brandt et al., 1994, Blood, 83, 1507-1514; Sato et al., 1993, Blood, 82, 3600-3609), G-CSF (Sato et al., 1993, Blood, 82, 3600-3609), GM-CSF (Sato et al., 1993, Blood, 82, 3600-3609), IL-1 (Muench et al., 1993, Blood, 81, 3463-3473), IL-6 (Sato et al., 1993, Blood, 82, 3600-3609), IL-11 (Lemoli et al., 1993, Exper. Hematol., 21, 1668-1672; Sato et al., 1993, Blood, 82, 3600-3609), лиганд flt3 (McKenna et al., 1995, Blood, 86, 3413-3420) и (или) их сочетание (Brandt et al., 1994, Blood, 83, 1507-1514; Haylock et al., 1992, Blood, 80, 1405-1412; Koller et al., 1993, Biotechnology, 11, 358-363; Lemoli et al., 1993, Exper. Heamtol., 21, 1668-1672; McKenna et al., 1995, Blood, 86, 3413-3420; Muench et al., 1993, Blood, 81, 3463-3473; Patchen et al., 1994, Biotherapy, 7, 13-26; Sato et al., 1993, Blood, 82, 3600-3609; Smith et al., 1993, Exper. Hematol., 21, 870-877; Steen et al. Stem Cells 12: 214 [1994], Tsujino et al., 1993, Exper. Hematol., 21, 1379-1386). Среди отдельных колониестимулирующих факторов самым патентным с точки зрения экспансии клеток периферической крови с антигеном CD34+ оказывается hIL-3 (Sato et al., 1993, Blood, 82, 3600-3609; Kobayashi et al., 1989, Blood, 73, 1836-1841). Однако ни один отдельный фактор не может быть столь же эффективным, как комбинация нескольких факторов. Настоящее изобретение представляет способы контроля экспансии ex vivo, в которых используются многофункциональные химерные агонисты гемопоэтических рецепторов, которые являются гораздо более эффективными в сравнении с отдельными факторами.
Также настоящее изобретение представляет способы контроля устойчивости (непрерывности) и (или) экспансии гемопоэтических клеток-предшественников, которые основаны на инокуляции клеток в культивируемый сосуд в культуральной среде, состояние которой установлено путем отработки с культурой линии клеток стромы, такой как HS-5 (патент WO 96/02662; Roecklein & Torok-Strob, 1995, Blood, 85, 997-1105), и в которую добавляют многофункциональный химерный агонист гемопоэтических рецепторов по настоящему изобретению.
Также важно отметить, что многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению могут быть использованы при передерживании (хранении) крови, когда агонисты эритропоэтиновых рецепторов вводят пациенту с целью увеличения числа клеток крови, а кровяные продукты должны быть выведены у пациента перед какими-либо лечебными процедурами. Кровь сохраняют и вводят обратно пациенту по окончании этих процедур. К тому же можно подчеркнуть возможность использования многофункциональных химерных агонистов гемопоэтических рецепторов для введения их донорам до взятия крови с целью увеличения числа кровяных клеток, давая таким образом возможность донору безопасно сдать больше крови.
Другим предполагаемым клиническим применением факторов роста является активация in vitro гемопоэтических клеток-предшественников и стволовых клеток в генотерапии. С учетом того, что клетки-предшественники кроветворения характеризуются значительной продолжительностью жизни, а их дочерние клетки распределяются практически по всему организму, гемопоэтические клетки-предшественниками являются перспективными объектами для трансгенеза ex vivo. Для того, чтобы обеспечить работу необходимого гена, встраиваемого в геном стволовой клетки или клетки-предшественника, нужно стимулировать клеточные деления и репликацию ДНК. Цикличность гемопоэтических стволовых клеток характеризуется низкой частотой: значит, факторы роста могут быть эффективны с точки зрения контроля трансдукции гена и соответствующего повышения клинических перспектив генотерапевтического подхода. Потенциальные аспекты генотерапии (см. обзор Crystal, 1995, Science, 270, 404-410) включают: 1) лечение многих врожденных нарушений метаболизма и иммунодефицита (Кау & Woo, 1994, Trends Genet., 10, 253-257), 2) лечение нейропатологий (Friedmann, 1994, Trends Genet., 10, 210-214), 3) лечение рака (Culver & Blaese, 1994, Trends Genet., 10, 174-178) и 4) лечение инфекционных заболеваний (Gilboa 8& Smith, 1994, Trends Genet., 10, 139-144).
Существует широкий круг известных специалистам в данной области техники способов внесения генетического материала в клетку-хозяина (реципиентную клетку). Для этой цели созданы различные векторы вирусной и невирусной природы, позволяющие вносить в первичные клетки гены, имеющие медицинское значение. Векторы вирусной природы: 1) рекомбинантный, дефицитный по репликации ретровирус (Boris-Lawrie & Temin, 1993, Curr. Opinion Genet. Develop., 3, 102-109; Boris-Lawrie & Temin, 1994, Ann. New York Acad. Sci., 716, 59-71; Miller, 1992, Current Topics Microbiol. Immunol., 1992, 158, 1-24) и 2) рекомбинантный, дефицитный по репликации аденовирус (Berkner, 1988, Bio-Techn., 6, 616-629; Berkner, 1992, Current Topics Microbiol. Immunol., 158, 39-66; Brody & Crystal, 1994, Ann. New York Acad. Sci., 716, 90-103). Векторы невирусной природы включают ДНК-белковые комплексы (Cristiano et al., 1993, Proc. Natl. Acad. Sci. USA, 90, 2122-2126; Curiel et al., 1991, Proc. Natl. Acad. Sci. USA, 88, 8850-8854; Curiel, 1994, Ann. New York Acad. Sci., 716, 36-58), электропорацию и перенос, опосредованный липосомами, например, катионными липосомами (Farhood et al., 1994, Ann. New York Acad. Sci., 716, 23-35).
Настоящее изобретение предлагает усовершенствование существующих способов экспансии гемопоэтических клеток, в которые может быть внедрен новый генетический материал, в частности предлагая использование многофункциональных химерных агонистов гемопоэтических рецепторов, обладающих повышенной биологической активностью, включая активности, которые не могут проявлять отдельные колониестимулирующие факторы.
Многие лекарственные препараты могут вызывать подавление работы костного мозга или дефицит гемопоэза. Примерами таких лекарств являются азатимидин, DDI, алкилирующие агенты и антиметаболиты, применяемые в химиотерапии, антибиотики (такие как хлорамфеникол, пенициллин, ганикловир, дауномицин), сульфопрепараты, фенотиазоны, транквилизаторы (такие как мепробамат), анальгетики (такие как аминопирин и дипирон), антиконвульсивные препараты (такие как фенитоин или карбамазепин), препараты против функций щитовидной железы (такие как пропилтиоурацил и метимазол) и диуретики. Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению могут быть использованы для профилактики или лечения подавленной функции костного мозга или дефицита гемопоэза, которые часто отмечаются у пациентов при применении перечисленных выше лекарственных препаратов.
Дефицит гемопоэза также часто встречается вследствие вирусных, микробных и паразитарных инфекций, при ожогах и в результате диализа, применяемого при лечении почечных заболеваний или дисфункций почек. Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению могут быть использованы для лечения таких случаев дефицита гемопоэза.
Лечение дефицита гемопоэза может включать введение пациенту фармацевтического препарата, включающего многофункциональные химерные агонисты гемопоэтических рецепторов. Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению также могут быть использованы для активации и амплификации клеток-предшественников кроветворения путем обработки этих клеток in vitro белками многофункциональных химерных агонистов гемопоэтических рецепторов по настоящему изобретению перед инъецированием таких клеток пациенту.
При лечении различных форм иммунодефицита, например, по Т- и (или) В-лимфоцитам, или форм иммунных заболеваний (например, ревматического артрита) также могут достигаться существенные успехи при применении многофункциональных химерных агонистов гемопоэтических рецепторов по настоящему изобретению. Формы иммунодефицита могут быть следствием вирусных инфекций (такими вирусами как HTLVI, HTLVII, HTLVIII), сильного облучения, противоопухолевой терапии или результатом других клинических воздействий. Многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению также могут быть использованы (отдельно или в сочетании с другими колониестимулирующими факторами) для терапии других типов дефицита клеток крови, включая тромбоцитопению (дефицит тромбоцитов) или анемию. Другие возможности применения новых полипептидов - терапия пациентов In vivo и ex vivo при восстановлении трансплантатов костного мозга, а также продуцирование моно- и поликлональных антител, осуществляемое стандартными методами для диагностических или терапевтических целей.
Также настоящее изобретение представляет способы и терапевтические рецептуры для терапии описанных выше заболеваний и состояний. Такие рецептуры включают терапевтически эффективные количества одного или нескольких многофункциональных химерных агонистов гемопоэтических рецепторов по настоящему изобретению при смешении с фармацевтически пригодным носителем. Этот состав может быть введен любым из перечисленных способов - парентерально, внутривенно или подкожно. При введении терапевтического состава по настоящему изобретению предпочтительно его приготовление в виде лишенных пирогенов (компонентов, способных повышать температуру) водных растворов, пригодных для введения парентерально. Приготовление пригодных для парентерального введения белковых растворов с учетом потребных параметров рН, изотоничности, стабильности и др., хорошо известно специалистам в данной области техники.
Еще одним способом применения многофункциональных химерных агонистов гемопоэтических рецепторов по настоящему изобретению является их вовлечение в формирование большего числа дендритных клеток из соответствующих предшественников с последующим их использованием в качестве адъювантов при иммунизации. Дендритные клетки играют важную роль в иммунной системе. Они являются специфическими антиген-представляющими клетками и наиболее эффективны в активации покоящихся Т-клеток, а также являются антиген-представляющими клетками, активирующими неспециализированные Т-лимфоциты in vivo, таким образом инициируя первичный иммунный ответ.
Они эффективно интернализируют, процессируют и представляют растворимые опухоль-специфичные антигены (Аg). Дендритные клетки обладают уникальной способностью кластеризовать неспециализированные Т-клетки и реагировать на накопление Аg быстрой позитивной регуляцией экспрессии МНС (главного комплекса гистосовместимости) и ко-стимуляторных молекул, а также продуцированием цитокинов и миграцией к лимфатическим органам. С учетом того, что дендритные клетки играют ключевую роль в сенсибилизации организма к новому антигену при формировании СD4-зависимого иммунного ответа, они могут также играть принципиальную роль в генерировании и регуляции опухолевого иммунитета.
Дендритные клетки происходят от костномозговых предшественников с CD34+, родственных гранулоцитам и макрофагам: в частности, у человека подтверждено существование отдельной колоние-формирующей единицы дендритных клеток (CFU-DC), что обеспечивает формирование "чистых" колоний дендритных клеток. К тому же прошедшие стадию CFU СР14+-интермедиаты характеризуются потенциалом к дифференциации в сторону дендритных клеток или макрофагов в условиях присутствия различающихся цитокинов. Такой бипотентный предшественник присутствует в костном мозге, пуповинной крови и периферической крови. Дендритные клетки могут быть выделены в ходе анализа созревания культивируемых дендритных клеток на основе специфичности поверхностно-клеточных маркеров, таких как CD1a+, CD3-, CD4-, CD20-, CD40+, CD80+ и CD83+.
Основанные на использовании дендритных клеток подходы поддерживают способ усиления иммунного ответа против опухолей и инфекционных возбудителей. СПИД является заболеванием, в терапии которого могут быть использованы дендритные клетки, поскольку дендритные клетки могут играть важную роль в поддержании репликации HIV-1. Для иммунотерапии является необходимым генерирование дендритных клеток от раковых больных, их обработка in vitro опухолевыми антигенами, выделенными из хирургически удаленных опухолевых масс ткани, и реинъекция этих клеток раковым больным. Относительно сырые мембранные фракции опухолевых клеток являются достаточными для использования в качестве источника опухолевого антигена, что позволяет исключить необходимость молекулярной идентификации опухолевого антигена. Опухолевый антиген может также быть синтетическим полипептидом, углеводородом или последовательностью нуклеиновой кислоты. К тому же согласованное введение цитокинов, таких как многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению, может также обеспечить индукцию противоопухолевого иммунитета. Можно предвидеть, что иммунотерапия может быть осуществлена in vivo, при том, что многофункциональные химерные агонисты гемопоэтических рецепторов по настоящему изобретению вводят раковому больному отдельно или вместе с другими гемопоэтическими факторами роста с целью увеличения количества дендритных клеток и содержания эндогенного опухолевого антигена, презентируемого дендритными клетками. Также следует отметить, что иммунотерапия in vivo может быть проведена с использованием экзогенного антигена. Также можно отметить, что иммунотерапевтические мероприятия могут включать мобилизацию предшественников дендритных клеток или зрелых дендритных клеток путем введения пациенту многофункциональных химерных агонистов гемопоэтических рецепторов по настоящему изобретению отдельно или в сочетании с другими гемопоэтическими факторами роста, последующего отбора предшественников дендритных клеток и зрелых дендритных клеток у пациента, обработки дендритных клеток антигеном и возвращения дендритных клеток в организм пациента. Далее, дендритные клетки, которые выделяются из организма, культивируются ех vivo в присутствии многофункциональных химерных агонистов гемопоэтических рецепторов по настоящему изобретению отдельно или вместе с другими гемопоэтическими факторами роста с целью увеличения числа дендритных клеток до этапа воздействия антигена. Подходы, основанные на использовании дендритных клеток, также позволяют представить способ снижения иммунного ответа при аутоиммунных заболеваниях.
Исследования дендритных клеток в значительной степени затруднены из-за сложности приготовления клеток в достаточном количестве и в достаточно "чистой" их форме. В методике экспансии клеток ех vivo колониестимулирующий фактор гранулоцитов-макрофагов (GM-CSF) и фактор-α некроза опухолей (TNF-α) кооперируются по генерированию ех vivo дендритных клеток из гемопоэтических предшественников (СD34+-клеток), находящихся в костном мозге, пуповинной крови или периферической крови, а лиганд flk-2/flt-3 и лиганд c-kit (фактор стволовых клеток - SCF) проявляют синергизм по усилению GM-CSF- и TNF-α-индуцируемому генерированию дендритных клеток (Siena et al., 1995, Exper. Hematol., 23, 1463-1471). Также представляется способ экспансии предшественников дендритных клеток или зрелых дендритных клеток ех vivo с использованием многофункциональных химерных агонистов гемопоэтических рецепторов по настоящему изобретению с целью накопления достаточного количества дендритных клеток для иммунотерапевтических методов.
Дозовый режим, входящий в терапию перечисленных выше заболеваний и состояний, должен быть определен с учетом физических кондиций, определяемых различными факторами, которые могут модифицировать активность лекарственных препаратов - таких как индивидуальное состояние, вес тела, пол, предпочитаемый рацион, тяжесть инфекции, время введения препарата и другие клинические показатели. В целом, дневной режим рекомендуется в пределах 0,2-150 мкг белка многофункциональных химерных агонистов гемопоэтических рецепторов на 1 кг веса тела. Дозы должны приспосабливаться с учетом активности конкретного многофункционального химерного агониста гемопоэтических рецепторов; при этом есть смысл отметить, что дозовый режим может включать и низкие дозы (менее 0,1 мкг), и высокие дозы (1 мг) в расчете на 1 кг веса тела в день. К тому же могут существовать специфические условия, при которых дозы многофункциональных химерных агонистов гемопоэтических рецепторов могут быть выведены за пределы рекомендуемого диапазона в 0,2-150 мкг на 1 кг веса тела. Это может быть связано с одновременным введением других колониестимулирующих факторов или вариантов интерлейкина-3, или факторов роста, с одновременным введением хемотерапевтических лекарственных средств и (или) облучением, использованием гликозилированного белка многофункционального химерного агониста гемопоэтических рецепторов и различными связанными с отдельными пациентами факторами, отмечавшимися выше. Как было отмечено выше, терапевтические методы и рецептуры могут включать одновременное введение и других белковых факторов человека. Неисчерпывающий перечень других подходящих колониестимулирующих факторов (CSFs), цитокинов, лимфокинов, гемопоэтических факторов роста и интерлейкинов для одновременного или последовательного (сериального) введения с полипептидами по настоящему изобретению таков: GM-CSF, G-CSF, лиганд с-mpl (также известный как ТРО или MGDF), M-CSF, эритропоэтин (ЕРО), IL-1, IL-4, IL-2, IL-3, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-13, IL-15, LIF лиганд flt3/flt2, гормон роста человека, фактор роста В-клеток, фактор дифференциации В-клеток, фактор дифференциации эозинофилов и фактор стволовых клеток (SCF), также известный как "жесткий" фактор или лиганд c-kit или их сочетания. Обозначенные выше дозы должны модифицироваться с целью компенсации присутствия таких дополнительных компонентов в составе терапевтических процедур. Прогресс у пациентов может отслеживаться с помощью периодической оценки гематологических показателей, например, дифференцированного подсчета клеток и подобное.
Материалы и методы
За исключением специально оговоренных случаев, все специальные реактивы получены от Sigma Co. (St.Louis, МО). Рестриктазы и ДНК-лигаза Т4 получены от New England Biolabs (Beverley, MA) или Boehringer Mannheim (Indianapolis, IN).
Трансформация штаммов E.coli
Штаммы E.coli, такие как DH5а™ (Life Technologies, Gaithersburg, MD) и TG1 (Amersham Corp., Arlington Heights, IL), использовали для трансформации продуктов лигирования и были использованы в качестве источника плазмидной ДНК для трансфицирования клеток млекопитающих. Штаммы E.coli, такие как JM101 (Yanish-Perron et al., 1985, Gene, 33, 103-119) и MON105 (Obukowicz et al., 1992, Appl. Environ. Microbiol., 58, 1511-1523), могут быть использованы для экспрессии многофункционального химерного агониста гемопоэтических рецепторов по настоящему изобретению в цитоплазме или периплазматическом пространстве.
MON105 ATCC#55204: F-, λ-, IN (rrnD, rrE)l, rpoD+, rpoH358
DH5а™: F-, φ80dlacZΔM15, Δ(lacZYA-argF) U169, deoR, recAl, endA1, hsdR17 (rk-, mk+), phoA, supE44λ-, thi-1, gyrA96, relA1
TG1: Δ(lас-рrо), supE, thi-1, hsd05/F’(traD36, proA+B+, lacIq, lacZΔM15)
JM101 ATCC#33876; Δ(lac-pro), supE, thi, F’(traD36, proA+B+, lacIq, lacZΔM15)
Эффективные субклоны РН5а™ выбирают в качестве компетентных клеток, готовых к трансформации с использованием предлагаемой производителем процедуры, в то время как оба штамма E.coli - TG1 и MON105 - становятся компетентными по включению ДНК с применением метода с хлоридом кальция. В принципе, 20-50 мл клеток растят в среде LB (1% бактотриптона, 0,5% бактериально-дрожжевого экстракта, 150 мМ Nacl) до достижения плотности, оцениваемой приблизительно в 1,0 единицу оптической плотности на 600 нм (OD600) при измерении на спектрофотометре Baush & Lomb Spectronic (Rochester, NY). Клетки собирают с помощью центрифугирования и ресуспензируют в одной пятой культурального объема в растворе CaCl2 (50 мМ CaCl2, 10 мМ Трис-HCl, рН=7,4) и выдерживают при 4°С в течение 30 минут. Клетки вновь собирают с применением центрифугирования и ресуспензируют в растворе СаСl2 из расчета 1/3 от объема культуральной среды. Лигированную ДНК добавляют к этим клеткам в объеме 0,2 мл: полученный препарат выдерживают при 4°С в течение 30-60 минут. Образцы на 2 минуты помещают на 42°С и 1,0 мл среды LB добавляют до встряхивания образцов при 37°С, проводящегося затем в течение 1 часа. Клетки этих образцов размещают в чашках Петри (среда LB + 1,5% бактериоагара), содержащих либо ампициллин (100 мкг/мл) в случае проведения отбора резистентных к ампициллину трансформантов, либо спектиномицин (75 мкг/мл) при отборе резистентных к спектиномицину трансформантов. Чашки Петри инкубируют в течение ночи при 37°С. Колонии отбирают и инокулируют в среду LB с добавлением необходимого антибиотика (100 мкг/мл ампициллина или 75 мкг/мл спектиномицина) и выращивают при 37°С.
Способы создания генов с новыми N-концами/С-концами
Способ I. Создание генов с новыми N-концами/С-концами, содержащих линкерный сегмент (L2)
Гены с новыми N-концами/С-концами, содержащие линкерный сегмент (L2), который разделяет исходные С- и N-концы, могут быть сформированы в принципе согласно методу, описанному Маллинзом с соавт. (Mullins et al., 1994, J. Amer. Chem. Soc., 116, 5529-5533). Последовательные этапы амплификации с помощью полимеразной цепной реакции (ПЦР) используют для перестройки последовательностей ДНК, кодирующих первичную аминокислотную последовательность полипептида. Эти этапы показаны на фиг.2.
На первом этапе первый набор заправок ("новая стартовая" и "линкерная стартовая") используют для создания и амплификации на основе исходной генной последовательности фрагмента ДНК ("стартовый фрагмент"), который содержит последовательность, кодирующую новый N-концевой участок нового полипептида, переходящий в линкер (L2), контактирующий с С- и N-концами исходного полипептида. На втором этапе второй набор затравок ("новая концевая" и "линкерная концевая") используют для создания и амплификации на основе исходной генной последовательности фрагмента ДНК ("концевой фрагмент"), который кодирует тот же линкер, что указан выше, продолжаемый С-концевой частью нового полипептида. В составе затравок "новая стартовая" и "новая концевая" обеспечивают наличие подходящих сайтов рестрикции, что позволяет клонировать новый ген в состав экспрессирующих плазмид. Типовые параметры ПЦР таковы: 1 цикл при 95°С в течение 2 минут (плавление), 25 циклов при 94°С в течение 1 минуты (денатурация), 50°С в течение 1 минуты (отжиг) и 72°С в течение 1 минуты (достройка); плюс 1 цикл при 72°С в течение 7 минут (дополнительная достройка). Для ПЦР используют реактивы GeneAmp PCR Core Reagents Kit (Perkin-Elmer). Реакционная смесь в объеме 100 мкл содержит 100 пкМ каждой затравки и 1 мкг матричной ДНК, а также 1х ПЦР-буфер, 200 мкМ дГТФ, 200 мкМ дАТФ, 200 мкМ дТТФ, 200 мкМ дЦТФ, 2,5 ед. ДНК-полимеразы ApliTaq и 2 мМ хлорида магния. Реакцию ПЦР осуществляют с использованием прибора Model 480 DNA Thermal cycler (Perkin-Elmer Corp., Norwalk, CT).
"Стартовый фрагмент" и "концевой фрагмент", включающие последовательности, комплементарные линкерному сегменту, и сегмент, кодирующий две аминокислоты с обоих концов линкера, соединяют вместе на третьем этапе ПЦР с целью формирования полноразмерного гена, кодирующего новый полипептид. Фрагменты ДНК "стартовый фрагмент" и "концевой фрагмент" выявляют в 1%-ном ТАЕ-геле, окрашивают бромистым этидием и выделяют с использованием реактивов Qiaex Gel Extraction Kit (Qiagen). Эти фрагменты объединяют в эквимолярных количествах, нагревают до 70°С в течение 10 минут и медленно охлаждают до достижения отжига по их общим последовательностям в "линкерном стартовом" и "линкерном концевом" фрагментах. На третьем этапе ПЦР "новую стартовую" и "новую концевую" затравки добавляют к прошедшим отжиг фрагментам с целью создания и амплификации полноразмерного гена с новыми N- и С-концами. Типовые параметры ПЦР: 1 цикл при 95°С в течение 2 минут (плавление), 25 циклов при 94°С в течение 1 минуты (денатурация), 60°С в течение 1 минуты (отжиг) и 72°С в течение 1 минуты (достройка); плюс 1 цикл при 72°С в течение 7 минут (дополнительная достройка).
Для ПЦР используют реактивы GeneAmp PCR Core Reagents Kit (Perkin-Elmer). Реакционная смесь в объеме 100 мкл содержит по 100 пкМ каждой затравки и 1 мкг матричной ДНК, а также 1х ПЦР-буфер, 200 мкМ дГГФ, 200 мкМ дАТФ, 200 мкМ дТТФ, 200 мкМ дЦТФ, 2,5 ед. ДНК-полимеразы ApliTaq и 2 мМ хлорида магния. Продукты ПЦР очищают с использованием реактивов Wizard PCR Preps Kit (Promega).
Способ II. Создание генов с новыми N-концами/С-концами,не содержащих линкерного сегмента
Новые С-концевые/N-концевые гены без линкерного соединения исходных N-конца и С-конца могут быть сформированы с помощью двух этапов ПЦР-амплификации и лигирования "тупых концов". Эти этапы показаны на фиг.3. На первом этапе набор затравок ("новая стартовая" и "стартовая Р-bl") используют для создания и амплификации по исходной генной последовательности фрагмента ДНК ("стартовый фрагмент"), который содержит последовательность, кодирующую новый N-концевой сегмент нового белка. На втором этапе набор затравок ("новая концевая" и "концевая Р-bl") используют для создания и амплификации по генной последовательности фрагмента ДНК ("концевого фрагмента"), который содержит последовательность, кодирующую новый С-концевой участок нового белка. Затравки "стартовый фрагмент" и "концевой фрагмент" включают подходящие сайты рестрикции, которые позволяют клонировать новый ген в состав экспрессирующего вектора. Типичные условия ПЦР: 1 цикл при 95°С в течение 2 минут (плавление), 25 циклов при 94°С в течение 1 минуты (денатурация), 50°С в течение 45 секунд (отжиг) и 72°С в течение 45 секунд (достройка). Полимеразу Deep-Vent (New England Biolabs) используют для снижения встречаемости выступов в условиях, рекомендуемых производителем. Затравки "стартовая Р-bl" и "концевая Р-bl" фосфорилируют по 5’-концу с целью последующего лигирования "тупых концов" "стартового фрагмента" и "концевого фрагмента" друг на друга. 100 мкл реактива содержат по 150 пкМ каждой из затравок и 1 мкг матричной ДНК, буфер 1 х Vent (New England Biolabs), 300 мкМ дРТФ, 300 мкМ дАТФ, 300 мкМ дТТФ, 300 мкМ дЦТФ и 1 ед. полимеразы Deep-Vent. Реакцию ПЦР проводят на приборе Model 480 Dna Thermal cycler (Perkin-Elmer Corp., Norwalk, CT). Продукт ПЦР очищают с использованием набора реактивов Wizard PCR Preps Kit (Promega).
Затравки созданы таким образом, что включают подходящие сайты рестрикции, которые позволяют клонировать новый ген в состав экспрессирующих векторов. Обычно "стартовый фрагмент" включает сайт узнавания рестриктазой NcoI, а "концевой фрагмент" включает сайт узнавания рестриктазой HindIII. Реакции расщепления рестриктазами проводят с использованием набора реактивов Марс DNA Clean-up System Kit (Promega). Стартовый и концевой фрагменты разделяют в 1%-ном геле ТАЕ, окрашивают бромистым этидием и выделяют с использованием набора Qiaex Gel Extraction Kit (Qiagen). Эти фрагменты смешивают и проводят отжиг на концы NcoI/HindIII-фрагмента длиной примерно 3800 нуклеотидов в составе вектора pMON3934 с помощью нагревания до 50°С в течение 10 минут с последующим медленным охлаждением. Три фрагмента лигируют вместе с использованием ДНК-лигазы Т4 (Boehringer Mannheim). В результате получают плазмиду, включающую полноразмерный ген с новыми N-концом/С-концом. Часть реакционной смеси лигирования используют для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Плазмидную ДНК очищают и последовательность подтверждают, как было описано выше.
Способ III. Создание генов с новыми N-концом/С-концом методом тандемной дупликации
Гены с новыми N-концом и С-концом могут быть сформированы на основе метода, описанного Горликом с соавт. (Horlick et al., 1992, Protein Eng., 5, 427-431). Амплификацию генов с новыми N-концом и С-концом методом полимеразной цепной реакции (ПЦР) проводят с использованием тандемно дуплицированной ДНК-матрицы. Этапы метода показаны на фиг.3.
Тандемно дуплицированную ДНК-матрицу создают путем клонирования: она содержит две копии гена, разделенные последовательностью ДНК, кодирующей линкер, соединяющий исходные С- и N-концы двух копий гена. Набор специфичных затравок используют для создания и амплификации полноразмерного гена с новыми N-концом и С-концом на тандемно дуплицированной ДНК-матрице. Эти затравки созданы таким образом, что включают подходящие сайты рестрикции, которые позволяют клонировать новый ген в состав экспрессирующего вектора. Типичные условия ПЦР таковы: 1 цикл при 95°С в течение 2 минут (плавление), 25 циклов при 94°С в течение 1 минуты (денатурация), 50°С в течение 1 минуты (отжиг) и 72°С в течение 1 минуты (достройка); плюс 1 цикл при 72°С в течение 7 минут (дополнительная достройка). Для ПЦР используют реактивы GeneAmp PCR Core Reagents Kit (Perkin-Elmer). Реакционная смесь в объеме 100 мкл содержит по 100 пкМ каждой затравки и 1 мкг матричной ДНК, а также 1х ПЦР-буфер, 200 мкМ дГГФ, 200 мкМ дАТФ, 200 мкМ дТТФ, 200 мкМ дЦТФ, 2,5 ед. ДНК-полимеразы ApliTaq и 2 мМ хлорида магния. Реакцию ПЦР осуществляют с использованием прибора Model 480 DNA Thermal cycler (Perkin-Elmer Corp., Norwalk, CT). Продукты ПЦР очищают с использованием реактивов Wizard PCR Preps Kit (Promega).
Клонирование генов с новыми N-концом и С-концом в векторы, экспрессирующие многофункциональные агонисты рецепторов
Ген с новыми С-концом и N-концом расщепляют рестриктазами с целью создания концов, необходимых для встраивания в экспрессирующий вектор, содержащий другой ген колониестимулирующего фактора. Этот экспрессирующий вектор подобным образом расщепляют рестриктазами с образованием сочетающихся концов. После очистки генную и векторную ДНК комбинируют и лигируют с использованием ДНК-лигазы Т4. Часть реакционной смеси лигирования используют для трансформации E.coli. Плазмидную ДНК очищают и секвенируют для коррекции вставки. Корректные клоны выращивают для экспрессии кодируемого ими белка.
Выделение и характеристика ДНК
Плазмидная ДНК может быть выделена с помощью ряда различных методов и с использованием стандартных наборов, известных специалистам данной области техники. Некоторые из этих методов описаны здесь. Плазмидную ДНК выделяют с использованием наборов Promega Wizard™ Miniprep Kit (Madison, WI), Qiagen QIAwell Plasmid Isolation Kit (Chatsworth, CA) или Qiagen Plasmid Midi Kit. Эти наборы соответствуют сходной процедуре выделения плазмидной ДНК. Вкратце, клетки гранулируют центрифугированием (5000 g), плазмидную ДНК освобождают последовательной обработкой едким натром и кислотой, а клеточные скопления удаляют центрифугированием (при 10000 g). Надосадочную фракцию (содержащую плазмидную ДНК) загружают в колонку, содержащую ДНК-связывающую смолу, колонку промывают, а плазмидную ДНК элюируют с Трис-НСl. После скрининга колоний на присутствие интересующих плазмид клетки E.coli инокулируют в 50-100 мл объемы среду LB с добавлением антибиотиков с выращиванием в течение ночи при 37°С в воздушном инкубаторе с последующим встряхиванием. Очищенную плазмидную ДНК используют для секвенирования ДНК, последующего расщепления рестриктазами, дополнительного субклонирования фрагментов ДНК и трансфекции в клетки млекопитающих, E.coli и другие клетки.
Подтверждение последовательности
Очищенную плазмидную ДНК ресуспензируют в дистиллированной воде и проводят количественный анализ путем измерения поглощения в диапазоне 260-280 нм на спектрометре Bausch/Lomb Spectronic 601 UV. Образцы ДНК секвенируют с использованием секвенсорного набора ABI PRISM™ DyeDepxy™ (Appl. Biosystems Div., Perkin-Elmer Corp., Lincoln City, CA; части №№ 401388 и 402078) в соответствии с рекомендуемой производителями процедурой, обычно модифицируемой путем добавления 5% ДМСО в секвенируемую смесь. Реакции секвенирования проводят на приборе Model 480 DNA Thermal cycler (Perkin-Elmer Corp., Norwalk, CT) в соответствии с рекомендуемыми условиями амплификации. Образцы очищают с целью удаления избытка окрасочных терминаторов на колонках Centri-Sep™ (Princeton Separations, Adelphia, NJ) и лиофилизируют. Помеченные флуоресцентными красителями секвенационные препараты ресуспензируют в дистиллированном формамиде и секвенируют в денатурирующем 4,75%-ном полиакриламид-8М-мочевинном геле с применением автоматического ДНК- секвенсора ABI Model 373. Перекрывающиеся фрагменты последовательностей ДНК анализируют и собирают в главный ДНК-контиг с применением программного продукта Sequencher-DNA-analysis (Gene Codes Corp., Ann Arbor, MI).
Экспрессия многофункциональных агонистов рецепторов в клетках млекопитающих
Трансфекция и продуцирование клеток млекопитающих в кондиционных средах
Линия клеток ВНК-21 может быть получена в АТСС (Rockville, MD). Клетки культивируют в модифицированной по Дальбекко среде Игла (DMEM с высоким содержанием глюкозы) с добавлением 2 мМ L-глутамина и 10% плодной бычьей сывороткой (FBS). Такой состав обозначен как культуральная среда ВНК. Селективная среда - это ростовая среда ВНК с добавлением 453 ед./мл гигромицина-В (Calbiochem, San Diego, CA). Линия клеток ВНК-21 ранее была стабильно трансфицирована HSV-производным трансактивирующим белком VP16, которые трансактивирует промотор UE110, обнаруженный в плазмиде pMON3359 (см. Hippenmeyer et al., 1993, Bio/Technol., 1037-1041). Белок VP16 контролирует экспрессию генов, встроенных после промотора 1Е110. Клетки ВНК-21, экспрессирующие трансактивирующий белок VP16, обозначают BHK-VP16. Плазмида pMON1118 (см. Highkin et al., 1991, Poultry Sci., 70, 970-981) экспрессирует ген резистентности к гигромицину с промотора SV40. Сходная плазмида, pSV2-hph, может быть получена в АТСС.
Клетки BHK-VP16 высевают на 60-миллиметровые чашки для культуральных тканей из расчета 3·105 клеток на 1 чашку за 24 часа до трансфекции. Клетки трансфицируют в течение 16 часов в 3 мл "OPTIMEM"™ (Gibco-BRL, Gaithersburg, MD), содержащем 10 мкг плазмидной ДНК, включающей интересующий ген, 3 мкг плазмиды с гигромициновой резистентностью (pMON1118) и 80 мкг "LIPOFECTAMINE"™ (Gibco-BRL) на чашку. Далее среду удаляют и замещают 3 мл ростовой среды. Через 48 часов после трансфекции среду из каждой чашки собирают и тестируют на активность (перемежающая кондиционная среда). Клетки удаляют из чашки с помощью трипсина-ЭДТА, разбавляют 1:10 и переносят на 100-мм чашки для культивирования ткани, содержащие 10 мл селективной среды. После приблизительно 7 дней нахождения в селективной среде резистентные клетки вырастают в колонии, имеющей несколько миллиметров в диаметре. Колонии удаляют с чашек с помощью фильтровальной бумаги (разрезаемые на кусочки размером, приблизительно таким же, как и размеры колоний, и пропитанные трипсином-ЭДТА) и переносят на отдельные лунки на 24-луночном планшете, содержащие 1 мл селективной среды. После того, как колонии вырастают до стадии слияния, кондиционную среду тестируют повторно, а позитивные клоны переносят на ростовую среду.
Экспрессия многофункциональных агонистов рецепторов в E.coli
Штаммы E.coli MON105 и JM101, несущие интересующие плазмиды, выращивают при 37°С в среде М9 с добавлением свободных аминокислот при встряхивании в воздушном инкубаторе Model G25 из New Brunswick Scientific (Edison, NJ). Рост контролируют по OD600 (оптическая плотность) до момента достижения величины 1,0: в этот момент добавляют налидиксовую кислоту (10 мг/мл) в 0,1-нормальной NaOH до конечной концентрации 50 мкг/мл. Культуры затем встряхивают при 37°С в течение 3-4 дополнительных часов. Высокую степень аэрации обеспечивают во время периода культивирования с целью достижения максимального выхода желательного генного продукта. Клетки анализируют под световым микроскопом на присутствие включенных телец (IB). 1-миллилитровые аликвоты культуры отбирают для анализа содержания белка путем кипячения гранулированных клеток с последующей обработкой редуцированным буфером и анализом в SDS-PAGE электрофорезе (см. Maniatis et al., 1982, Molecular Cloning: A Laboratory Manual). Культуру центрифугируют (5000g) до момента гранулирования клеток.
Препарирование включенных телец, экстракция, рефолдинг, диализ, хроматография DEAE и характеристика многофункциональных химерных агонистов гемопоэтических рецепторов, которые накапливаются во включенных тельцах в E.coli
Выделение включенных телец:
Клеточные гранулы из культуры E.coii в объеме 330 мл ресуспензируют в 15 мл соникационного буфера (10 мМ 2-амино-(гидроксиметил)-1,3-пропандио-гидрохлорид (Трис-НСl), рН=8,0+1 мМ этилендиаминтетрауксусной кислоты (ЭДТА). Эти ресуспензированные клетки обрабатывают ультразвуком с использованием зонда в Sonicator Cell Disrupotor Model W-375 (Heat Systems Ultrasonics Inc., Farmingdale, NY). Три круга воздействия ультразвука в соникационном буфере с последующим центрифугированием применяют для разрушения клеток и вымывания включенных телец (IB). Первый круг воздействия ультразвука представлен 3-минутным разрывом с последующим 1-минутным разрывом, а заключительные два круга воздействия ультразвука продолжаются по 1 минуте каждый.
Экстракция и рефолдинг белков из гранул включенных телец;
После заключительного этапа центрифугирования гранулы IB ресуспензируют в 10 мл 50 мМ Трис-HCl, рН=9,5, 8 М мочевины и 5мМ дитиотреитола (DTT) и взбалтывают при комнатной температуре в течение приблизительно 45 минут с целью обеспечения денатурации экспрессируемого белка.
Экстрактный раствор переносят в мензурку, содержащую 70 мл с 5М Трис-HCl, рН=9,5 и 2,3 М мочевины и тщательно взбалтывают, а затем оставляют на воздухе при 4°С в течение 18-48 часов для обеспечения рефолдинга белка. Рефолдинг контролируют с помощью анализа на колонках 0,46×25 см жидкостной хроматографии в обратной фазе высокого давления Vydac (Hesperia, СА) С18. Линейный градиент ацетонитрила от 40% до 65%, содержащего 0,1% трифторуксусной кислоты (ТФК), используют для контроля рефолдинга. Этот градиент формируют в течение 30 минут при скорости потока 1,5 мл в минуту. Денатурированные белки в целом элюируются позже в этом градиенте в сравнении с белками, имеющими восстановленную пространственную структуру.
Очистка
Вслед за рефолдингом загрязненные белки Е.coli удаляют с помощью кислотной преципитации. С использованием 15%-ной (объемная концентрация) уксусной кислоты титруют рН рефолдного раствора в диапазоне от рН=5,0 до рН=5,2. Этот раствор взбалтывают при 4°С в течение 2 часов и затем центрифугируют в течение 20 минут при 12000 g до образования гранул всеми нерастворимыми белками.
Надосадочную фракцию, полученную на этапе кислотной преципитации, диализуют с использованием мембраны Spectra/Por-3 с отсечением по величине молекулярной массы 3500 дальтон. Диализ проводят против 2 смен по 4 литра (с 50-кратным избытком) 10 мМ Трис-HCl, рН=8,0 в течение 18 часов в целом. Диализ снижает электропроводимость образца и удаляет мочевину перед проведением хроматографии DEAE. Затем образцы центрифугируют (20 минут при 12000 g) до образования гранул всех нерастворимых белков после диализа.
Колонки Bio-Rad Bio-Scale DEAE2 (7×52 мм) используют для ионобменной хроматографии. Колонки уравновешивают в буфере, содержащем 10 мМ Трис-HCl, рН=8,0 и градиент хлорида натрия в диапазоне 0-500 мМ в равновесном буфере, более 45 объемов колонок используют для элюции белка. Применяют темп потока 1,0 мл в минуту. Колоночные фракции (2,0 мл в расчете на фракцию) собирают по градиенту и анализируют на колонках 0,46×25 см жидкостной хроматографии в обратной фазе высокого давления Vydac (Hesperia, CA) C18. Используют линейный градиент ацетонитрила в диапазоне от 40% до 65%, содержащего 0,1% трифторуксусной кислоты. Этот градиент формируют за 30 минут при скорости потока 1,5 мл в минуту. Объединенные фракции затем подвергают диализу против 2 смен по 4 литра (избыток в 50-500 крат) 10 мМ ацетата аммония, рН=4,0 в течение 18 часов в целом. Диализ проводят с использованием мембраны Spectra/Por-3 с отсечением по величине молекулярной массы 3500 дальтон. В конце образец фильтруют в стерильных условиях с использованием шприц-фильтра 0,22 мкм (шприц-фильтр μStar LB, Costar, Cambridge, МА) и сохраняют при 4°С.
В некоторых случаях белки с нормальной пространственной структурой могут быть аффинно очищены с использованием соответствующих реагентов, таких как моноклональные антитела или рецепторные субъединицы, прикрепленные к подходящему матриксу. С другой стороны (или в дополнение), очистка может быть проведена с использованием любой из группы хроматографических процедур, как-то: ион-обменной, гель-фильтрационной или гидрофобной хроматографии, или ВЭЖХ в обратной фазе.
Эти и другие методы очистки белков подробно описаны в "Methods in Enzymology", Vol. 182 "Guide to Protein Purification", ed. M.Deutscher, Academic Press, San Diego, CA (1990).
Характеристика белка:
Очищенный белок анализируют с помощью ВЭЖХ в обратной фазе, электроспрэй-масс-спектрометрии и SDS-PAGE. Количественный анализ белка проводят на аминокислотный состав с помощью ВЭЖХ в обратной фазе и тестирования белка по Брэдфорду. В некоторых случаях проводят триптическое картирование белков совместно с электроспрэй-масс-спектрометрией с целью подтверждения идентичности белков.
Тест на пролиферацию AML для биологически активного интерлейкина-3 человека
Фактор-зависимую клеточную линию AML-193 получили в Американской коллекции типовых культур (АТСС, Rockville, MD). Эта клеточная линия, сформированная от клеток пациента с острым миелогенным лейкозом, является клеточной линией, зависимой от факторов роста, проявляющей усиленный рост в присутствии в культуральной среде GM-CSF (Lange et al., 1987, Blood, 70, 192; Valtieri et al., 1987, J. Immunol., 138, 4042). Была подтверждена способность клеток AML-193 пролиферировать в присутствии IL-23 человека (Santoli et al., 1987, J. Immunol., 139, 348). Был использован вариант клеточной линии - AML-193/1.3, который был адаптирован для долговременного роста в присутствии IL-3 путем вымывания факторов роста и голодания цитокин-зависимых клеток AML-193 по факторам роста в течение 24 часов. Затем клетки пересевают из расчета 105 клеток на лунку на 24-луночных планшетах в среде, содержащей 100 ед./мл IL-3. Их выдерживают в течение примерно 2 месяцев для обеспечения быстрого роста клеток в присутствии IL-3. Эти клетки поддерживают как AML-193/1.3 путем добавления среды для культивирования тканей (см. описанное выше) в присутствии IL-3 человека.
Клетки AML-193/1.3 промывают 6 раз в холодном сбалансированном солевом растворе Хэнкса (HBSS, Gibco, Grand Island, NY) с помощью центрифугирования клеточных суспензий при 250 g в течение 10 минут с последующим отмучиванием надосадочной фракции. Гранулированные клетки ресуспензируют в HBSS и процедуру повторяют до тех пор, пока шесть промывочных циклов не будут завершены. Клетки, промытые 6 раз согласно описанной процедуре, ресуспензируют в среде для культирования тканей в плотности в диапазоне от 2·105 до 5·105 живых клеток на 1 мл. Эту среду приготавливают путем добавления к модифицированной по Айскоуву среде Дальбекко (IMDM, Hazelton, Lenexa, KS) альбумина, трансферрина, липидов и 2-меркаптоэтанола. Бычий альбумин (Boehringer-Mannheim, Indianapolis, IN) добавляют в дозе 500 мкг/мл; трансферрин человека (Boehringer-Mannheim, Indianapolis, IN) добавляют в дозе 100 мкг/мл; соевый жир (Boehringer-Mannheim, Indianapolis, IN) добавляют в дозе 50 мкг/мл; и 2-меркаптоэтанол (Sigma, St.Louis, МО) добавляют в дозе 5·10-5 М.
Серийные разбавления интерлейкина-3 человека или белков многофункциональных химерных агонистов гемопоэтических рецепторов проводят в сериях с тройными повторами в среде для культивирования тканей с добавлением названных выше компонентов на 96-луночных планшетах Costar-3596 для культивирования тканей. Каждая лунка по завершении серийных разбавлений содержит по 50 мкл среды, включающей интерлейкин-3 или белки многофункциональных химерных агонистов гемопоэтических рецепторов. Контрольные лунки содержат только среду для культивирования тканей (негативный контроль). Суспензии клеток AML-193/1.3, приготовленные, как это было описано выше, добавляют в каждую лунку с помощью пипетки 50-мкл (по 2,5·104 клеток на каждую лунку). Культуральные пластины инкубируют при 37°С с 5% СО2 во влажном воздухе в течение 3 дней. На 3-й день меченый 3H-тимидин с активностью 0,5 микрокюри (2 Кю/мМ, New England Nuclear, Boston, MA), добавляют в 50 мкл среды для культивирования тканей. Культуры инкубируют при 37°С с 5% СО2 во влажном воздухе в течение 18-24 часов. Клеточную ДНК собирают на стеклянную фильтровальную пленку (Pharmacia LKB, Gaithersberg, MD) с использованием прибора ТОМТЕС (ТОМТЕС, Orange, CT), действие которого основано на цикличной промывке водой с последующей цикличной промывкой 70%-ным этанолом. Фильтровальные пленки оставляют до высыхания на воздухе и затем помещают в коробку для образцов, в которую добавляют сцинтилляционную жидкость (Scintiverse II, Fisher Scientific, St.Louis, МО или BetaPlate Scintillation Fluid, Pharmacia LKB, Gaithersburg, MD). Эмиссию β-частиц образцами отдельных лунок для культур тканей считают с помощью сцинтилляционного счетчика LKB BetaPlate 1205 (Pharmacia LKB, Gaithersburg, MD), а полученные данные выражают в виде подсчетов в минуту числа инкорпорированных остатков 3H-тимидина в клетках в каждой из лунок для культивирования тканей. Активность каждого препарата интерлейкина-3 или белка многофункционального химерного агониста гемопоэтических рецепторов анализируют количественно с помощью измерения интенсивности клеточной пролиферации (инкорпорации 3H-тимидина), индуцированной последовательными концентрациями интерлейкина-3 или многофункционального химерного агониста гемопоэтических рецепторов. Типичный диапазон концентраций, анализируемых в данных тестах, находится в пределах от 0,05 пкМ до 105 пкМ. Активность определяют с помощью измерения дозы интерлейкина-3 или белка многофункционального химерного агониста гемопоэтических рецепторов, которая обеспечивает 50% от максимальной пролиферации (EC50=0,5×(максимальный средний подсчет в минуту уровня 3H-тимидина, инкорпорированного в 1 лунку, среди трижды повторенных культур при всех тестированных концентрациях интерлейкина-3 - исходный уровень пролиферации измеряется уровнем инкорпорации 3H-тимидина в трижды повторенных культурах в отсутствие интерлейкина-3). Эта величина ЕС50 также эквивалентна 1 единице биоактивности. Каждый тест проводят с нативным интерлейкином-3, как калибровочным стандартом, что позволяет оценивать относительную активность.
В целом белки многофункциональных химерных агонистов гемопоэтических рецепторов были тестированы в диапазоне концентраций от 2000 пкМ до 0,06 пкМ, титруемых в серии двукратных разведений.
Активность каждого образца определяли по концентрации, которая обеспечивала 50% от максимального ответа при привлечении 4-параметрической документальной модели полученных данных. Было установлено, что верхнее плато (максимальный ответ) образца и стандарт, с которым его сравнивали, не различались. Соответственно, оценка относительного потенциала каждого образца была определена по оценкам EC50 для образца и стандарта, как это было описано выше. Клетки AML-193/1.3 пролиферируют в ответ на hIL-3, hGM-CSF и hG-CSF соответственно, следующие дополнительные тесты были проведены для некоторых образцов с целью демонстрации того, что агонист рецептора G-CSF, являющийся частью белков многофункциональных химерных агонистов гемопоэтических рецепторов, активен. Тест на пролиферацию был проведен с многофункциональным химерным агонистом гемопоэтического рецептора, плюс-минус нейтрализующие моноклональные антитела к части, являющейся агонистом рецептора hIL-3. В дополнение химерную молекулу с сайтом расщепления по фактору Ха расщепляли, затем очищали и полученные половины молекул тестировали на пролиферационную активность. Эти эксперименты показали, что оба компонента белков многофункционального химерного агониста гемопоэтических рецепторов активны.
Тест на пролиферацию, зависимую от лиганда TF1 с-mpl
Пролиферационная активность лиганда с-mpl может быть проверена с использованием субклона плюрипотентной линии клеток TF1 человека (Kitamura et al., 1989, J. Cell Physiol., 140, 323-334). Клетки TF1 поддерживают в присутствии hIL-3 (100 ед./мл). Для формирования субклона, реагирующего на лиганд с-mpl, клетки поддерживали в пересевной среде, содержащей 10% надосадочной фракции клеток ВНК, трансфицированных геном, экспрессирующим форму 1-153 лиганда c-mpl (pMON26448). Большинство клеток погибает, однако некоторые сохраняют жизнеспособность. После дилюционного клонирования отбирают реагирующий на лиганд c-mpl клон, и эти клетки помещают в пересевную среду из расчета 0,3·106 клеток на 1 мл за день до проведения теста.
Пересевная среда для этих клеток следующая: RPMI-1640 (gibco), 10% плодной бычьей сыворотки (Harlan, Lot #91206), 10% надосадочной фракции ВНК-клеток, трансфицированных лигандом c-mp1, 1 мМ пирувата натрия (Gibco), 2 мМ глутамина (Gibco) и 100 мкг/мл пенициллина-стрептомицина (Gibco). На следующий день клетки собирают и промывают дважды в средах RPMI или IMDM с окончательной промывкой в ATL или в тестовой среде. Среда ATL включает следующее: IMDM (Gibco), 500 мкг/мл бычьего сывороточного альбумина, 100 мкг/мл трансферрина человека, 50 мкг/мл соевого жира, 4·10-8 М β-меркаптоэтанола и 2 мл А9909 (Sigma, раствор антибиотиков) на 1000 мл ATL. Клетки разбавляют в тестовой среде с окончательной плотностью 0,25-10 клеток на 1 мл в 96-луночных низкоиспаряющих пластинах (Costar) до окончательного объема в 50 мкл. Временную надосадочную фракцию (кондиционную среду) из трансфицированных клонов добавляют в объеме 50 мкл, как дуплицированного образца, с окончательной концентрацией 50%, и разбавляют втрое до окончательного разбавления на уровне 1,8%. Трипликатные образцы кривой доз варианта IL-3 в составе pMON13288, начиная от 1 нг/мл и, путем трехкратного разбавления, до 0,0014 нг/мл, включают как позитивный контроль. Пластины инкубируют при 5% CO2 и 37°С. На 6-й день культивирования пластины облучают пульсами трития с активностью 0,5 Кю на 1 лунку при объеме 20 мкл на 1 лунку и инкубируют при 5% CO2 и 37°С в течение 4 часов. Пластины собирают и анализируют с помощью счетчика Betaplate.
Тест на пролиферацию клеток MUTZ-2
Клеточная линия, такая как MUTZ-2, которая является производной от линии клеток миелоидного лейкоза (Германская коллекция микроорганизмов и клеточных культур, DSM АСС 271), может быть использована для определения пролиферационной активности агонистов рецепторов flt3. Культуры MUTZ-2 поддерживают с рекомбинантным нативным лигандом flt3 (20-100 нг/мл) в ростовой среде. За 18 часов до проведения теста клетки MUTZ-2 промывают в среде IMDM (Gibco) трижды, ресуспензируют в среде IMDM в концентрации 0,5-0,7·106 клеток на 1 мл и инкубируют при 5% СO2 и 37°С в условиях голодания клеток по лиганду flt3. В день самого теста стандарты и агонисты рецептора flt3 разбавляют в 2 раза свыше желательной итоговой концентрации в тестовой среде в стерильных 96-луночных планшетах для культивирования тканей. Агонисты рецептора flt3 и стандарты тестируют в трех повторностях. Во все лунки, кроме ряда А, заливают 50 мкл тестовой среды. 75 мкл агонистов рецепторов flt3 или стандартов добавляют в ряд А и 25 мкл отбирают обратно из этого ряда путем серийных разбавлений (1:3), перенося на оставшуюся часть планшета (ряды от В до G). Ряд Н сохраняют как контроль, включающий только среду. Голодные клетки MUTZ-2 промывают два раза в среде IMDM и ресуспензируют в 50 мкл тестовой среды. 50 мкл клеток добавляют в каждую лунку, в результате чего итоговая концентрация составляет 0,25·106 клеток на 1 мл. Тестовые планшеты, содержащие клетки, инкубируют при 5% СO2 и 37°С в течение 44 часов. Каждую лунку затем облучают меченным тритием тимидином с активностью 1 Кю на 1 лунку в объеме 20 мкл в течение 4 часов. Планшеты затем собирают и обсчитывают.
Другие тесты на пролиферацию клеток in vitro
Другие клеточные тесты in vitro, известные специалистам в данной области техники, также могут быть использованы для определения активности многофункциональных химерных агонистов гемопоэтических рецепторов, в зависимости от факторов, которые включают молекулу, сходную с таковой, описанной для теста на пролиферацию клеток AML-193/1.3. Приведены следующие примеры применимых тестов.
Тест на пролиферацию TF1: TF1 - плюрипотентная линия клеток человека (Kitamura et al., 1989, J. Cell Physiol., 140, 323-334), реагирующая на hIL-3.
Тест на пролиферацию 32D: 32D - IL3-зависимая линия клеток мыши, которая не реагирует на IL-3 человека, но реагирует на G-CSF человека, для которого отсутствует ограничение по видоспецифичности.
Тест на пролиферацию Baf/3: Baf/3 – IL3-зависимая линия клеток мыши, которая не реагирует на IL-3 человека или лиганд с-mpl человека, но реагирует на G-CSF человека, для которого отсутствует ограничение по видоспецифичности.
Тест на пролиферацию Т1165: Т1165 - IL6-зависимая линия клеток мыши (Nordan et al., 1986), реагирующая на IL-6 и IL-11.
Тест на плазматический Clot meg-CSF человека: применяется для тестирования активности по формированию колоний мегакариоцитов (Mazur et al., 1981).
Линии трансфицированных клеток:
Клеточные линии такие как линия клеток Baf/3 мыши, могут быть трансфицированы рецептором колониестимулирующего фактора, таким как рецептор G-CSF человека или рецептор с-mpl человека, которые в клеточной линии отсутствуют. Эти трансфицированные клеточные линии могут быть использованы для определения активности лиганда, рецептор для которого трансфицирован в клеточную линию.
Одна такая трансфицированная клеточная линия Baf/3 была сформирована из клеточной линии, реагирующей на с-mpl, и клонирована в множественный сайт клонирования плазмиды pcDNA3 (Invitrogen, San Diego, CA). Клетки Baf/3 трансфицировали плазмидой с применением электропорации. Клетки выращивали в условиях отбора G418 в присутствии IL-3 мыши в кондиционной среде Wehi. Клоны были сформированы при ограниченном разбавлении.
По сходной схеме рецептор G-CSF человека может быть трансфицирован в клеточную линию Baf/3 и использован для определения биологической активности многофункциональных химерных агонистов гемопоэтических рецепторов.
Анализ пролиферационной активности лиганда с-mpl
Методы
1, Тест на пролиферацию костного мозга
а. Очистка клеток CD34+:
Аспираты костного мозга (15-20 мл) получали от нормальных аллогенных доноров костного мозга после соответствующего согласия. Клетки разбавляли 1:3 в фосфатном буфере (PBS, Gibco-BRL), 30 мл помещали слоями в 15-мл Histopaque-1077 (Sigma) и центрифугировали в течение 30 минут при 300 RCF. Поверхностный моноядерный слой собирали и промывали в PBS. Клетки CD34+ обогащали из препарата моноядерных клеток с использованием аффинной колонки и инструкций производителей к ней (CellPro Inc., Bothell, WA). После обогащения чистота клеток CD34+ достигала 70% в среднем, что было определено с помощью проточной цитометрии с поликлональным антителом к CD34, соединенным с флуоресцеином, и aHTH-CD38, соединенным с фикоэритрином (Becton Dickinson, San Jose, CA).
Клетки ресуспензировали из расчета 40000 клеток на 1 мл в среде X-Vivo10 (Вю-Whittaker, Walkersville, MD), и 1 мл помещали на 12-луночные планшеты для культивирования тканей (Costar). Фактор роста rhIL-3 добавляли из расчета 100 нг/мл (pMON5873) в некоторые лунки. Варианты hIL-3 использовали в концентрации 10-100 нг/мл. Кондиционную среду клеток ВНК, трансфицированных плазмидой, которая кодирует лиганд с-mpl или многофункциональные химерные агонисты гемопоэтических рецепторов, тестировали путем добавления 100 мкл надосадочной фракции, добавляемой к 1 мл культуры (приблизительно 10%-ное разбавление). Клетки инкубировали при 37°С в течение 8-14 дней при 5% СO2 во влажном инкубаторе с постоянной температурой 37°С.
b. Сбор и анализ клеток:
По окончании периода культивирования общее число клеток подсчитывали в каждом из вариантов. Для флуоресцентного анализа и определения уровня плотности клетки промывали в специальном мегакариоцитном буфере (буфер МК: 13, 6 мМ цитата натрия, 1 мМ теофиллина, 2,2 мкМ PGE1, 11 мМ глюкозы, 3% (весовые проценты) альбумина бычьей сыворотки, растворенные в физиологическом растворе с фосфатным буфером (PBS): рН=7,4 (Tomer et al., 1987, Blood, 70, 1735-1742), ресуспензировали в 500 мкл МК-буфера, содержащего анти-СD41а FITC-антитело (1:200, АМАС, Westbrook, ME), и промывали в МК-буфере. Для анализа ДНК клетки прокачивали буфером МК, содержащим 0,5% Твин-20 (Fisher, Fair Lawn, NJ) в течение 20 минут на льду с последующей фиксацией в 0,5%-ном Твин-20 и 1%-ном параформалине (Fisher Chemical) в течение 30 минут, а затем инкубировали в йодистом пропидие (Calbiochem, La Jolla, CA) (50 мкг/мл) с РНКазой (400 ед./мл) в 55%-ном (объемные проценты) буфере МК (200 m0sm) в течение 1-2 часов на льду. Клетки анализировали на сканере FACScan или проточном цитометре Vantage (Becton Dickinson, San Jose, CA). Сигналы зеленой флуоресценции (CD41a-FITC) отбирали вдоль линейных и логарифмических сигналов красной флуоресценции (йодистый пропидий) с целью определения плоидности ДНК. Все клетки собирали для определения доли клеток, позитивных по CD41. Анализ данных был проведен с использованием компьютерной программы LYSIS (Becton Dickinson, San Jose, CA). Доля клеток, экспрессирующих антиген CD41, была определена с помощью проточной цитометрии (показатель "процент"). Абсолютное число СD41-позитвных клеток в 1 мл определяли по формуле: Абс=(число клеток)×("процент")/100.
Клот-тест с фибрином мегакариоцитов
Обогащенная CD34-позитивными клетками популяция была выделена, как это было описано выше. Клетки суспензировали из расчета на 25000 клеток в 1 мл с или без цитокина (цитокинов) в среде, содержащей основную среду Isco-ves-IMDM, дополненную 0,3% альбумина бычьей сыворотки, 0,4 мг/мл апотрансферрина, 6,67 мкМ FeCl2, 25 мкг/мл CaCl2, 25 мкг/мл L-аспарагина, 500 мкг/мл ε-амино-n-капроевой кислоты и пенициллина/стрептомицина. До посева на 35-мм чашках тромбин добавляли из расчета 0,25 ед./мл с целью индукции "клота" (свертывания). Клетки инкубировали при 37°С в течение 13 дней при 5% СО2 во влажном инкубаторе с постоянной температурой 37°С.
По окончании периода культивирования пластины фиксировали смесью метанола и ацетона в соотношении 1:3, высушивали на воздухе и сохраняли при - 200°С перед окрашиванием. Была использована процедура пероксидазного иммуноцитохимического окрашивания (Zymed, Histostain-SP, San Francisco, CA) с использованием смеси первичных моноклональных антител - к CD41a, CD42 и CD61. Колонии подсчитывали после окрашивания и классифицировали как негативные, CFU-MK (маленькие колонии с 1-2 фокусами и меньше, что соответствует примерно 25 клеткам), BFU-MK (крупные мультифокусные колонии с более чем 25 клетками) или смешанные колонии (смесь и позитивных, и негативных клеток).
Метилцеллюлозный тест
Этот тест отражает способность колониестимулирующих факторов стимулировать нормальные клетки костного мозга к продуцированию различных типов гемопоэтических колоний in vitro (Bradley et al., 1966, Austral. Exper. Biol. Sci., 44, 287-300; Pluznik et al., 1965, J. Cell. Соmр. Physiol., 66, 319-324).
Методы
Приблизительно 30 мл свежего нормального здорового костномозгового аспирата получают от доноров при их согласии. В стерильных условиях образцы разбавляют 1:5 в 1х физиологическом растворе с фосфатным буфером (№ 14040.059 Life Technologies, Gaithersburg, MD) в 50-мл конических пробирках (№25339-50 Corning, Corning, MD). Фиколл (Histopaque 1077 Sigma H-8889) размещают слоями под разбавленным образцом и центрифугируют при 300 g в течение 30 минут. Слой моноядерных клеток выделяют и промывают дважды в 1х физиологическом растворе с фосфатным буфером и один раз в физиологическом растворе с фосфатным буфером, содержащим 1% альбумина бычьей сыворотки (CellPro Col., Bothel, WA). Моноядерные клетки подсчитывают и позитивные по CD34 клетки отбирают с использованием колонки и набора Ceprate LC (CD34) Kit (CellPro Co., Bothel, WA). Такое фракционирование проводят, хотя все стволовые клетки и клетки-предшественники в костном мозге проявляют поверхностный антиген CD34.
Культуры формируют в трех повторностях с конечным объемом 1,0 мл в чашках Петри размером 35×10 мм (Nunc № 174926). Культуральную среду приобретают у Terry Fox Labs. В культуральную среду добавляют среду НСС-4230 (Terry Fox Labs, Vancouver, ВС, Canada) и эритропоэтин (Amgen, Thousand Oaks, CA). На каждую чашку размещают по 3-10 тысяч клеток CD34+. Рекомбинантный IL-3, очищенный из клеток млекопитающих или E.coli, и белки многофункциональных химерных агонистов гемопоэтических рецепторов из кондиционной культуральной среды трансфицированных клеток млекопитающих или очищенные из кондифиционной культуральной среды трансфицированных клеток млекопитающих или E.coli, добавляют с получением окончательной концентрации от 0,001 нМ до 10 нМ. Рекомбинантные hIL-3, GM-CSF, лиганд с-mpl и многофункциональный химерный агонист гемопоэтических рецепторов сохраняют в домашних условиях. G-CSF (Neupogen) получают из Amgen (Thousand Oaks, CA). Культуры ресуспензируют с использованием шприца и 1,0 мл культуры помещают на 1 чашку. В контрольные культуры (с исходным уровнем реакции) не вносят колониестимулирующих факторов. В культуры, являющиеся позитивным контролем, вносят кондиционную среду (стимулированные фитогемагглютинином клетки человека: Теrу Fox Lab. H2400). Культуры инкубируют при 37°С при 5% СO2 во влажном воздухе.
Гемопоэтические колонии, определяемые как колонии с более чем 50 клетками, подсчитывают в день максимального ответа (10-11-й день) с использованием микроскопа с инвертированной световой фазой марки "Никон" с объективом 40х. Группы, содержащие менее 50 клеток, определяют как кластеры. С другой стороны, колонии могут быть идентифицированы путем распределения колоний на предметных стеклах с окрашиванием, или же они могут быть сняты, ресуспензированы и помещены на цитопсиновых препаратах для окрашивания.
Тесты на реакцию пуповинной крови человека на гемопоэтические факторы роста
Клетки костного мозга традиционно используют в тестах in vitro на активность колониестимулирующих факторов (CSF). Однако костный мозг человека не всегда достаточно легко доступен, и при этом существует значительная вариабельность между отдельными донорами. Пуповинная артерия сравнима с костным мозгом в качестве источника стволовых клеток и клеток-предшественников кроветворения (Broxmeyer et al., 1992, Proc. Natl. Acad/ Sci., 89, 4109-4113; Mayani et al., 1993, Blood, 81, 3252-3258). В противопоставлении костному мозгу, пуповинная кровь может быть легко доступна на регулярной основе. Также существует возможность снижения вариабельности результатов тестов за счет взятия достаточного числа клеток сразу от нескольких доноров или благодаря созданию банков криконсервированных клеток для тех же целей. Путем модифицирования условий культивирования и (или) тестирования специфичных для клеточных типов маркеров становится возможным специфически тестировать колонии гранулоцитов/макрофагов (CFU-GM), активности CSF в отношении мегакариоцитов или активности по контролю высокопролиферационного потенциала колоние-формирующих клеток (HPP-CFC).
Методы
Моноядерные клетки (MNC) выделяют из пуповинной крови в пределах суток после отбора образца с использованием стандартного градиента плотности (1,077 г/мл Histopaque). Далее клетки MNC пуповинной крови обогащают по стволовым клеткам и клеткам-предшественникам с помощью ряда процедур, включая магнитоиммунный отбор на клетки CD14-, CD34+, посев фракций CD14-, CD34+ в специальных колбах Appliеd Immune Science (Santa Clara, CA) и отбор клеток CD34+ на авидиновых колонках CellPro (Bothell, WA). Для теста используют как свежевыделенные, так и криоконсервированные фракции, обогащенные по CD34-позитивным клеткам. Культуры в двух повторностях для каждого из серийных разбавлений образца (диапазон концентраций от 1 до 1204 пкМ) готовят из расчета 10000 клеток в 1 мл среды, содержащей 0,9% метилцеллюлозы и лишенной дополнительных факторов роста (Methocult H4230, Stem Cell Technologies, Vancouver, ВС, Canada). В некоторых экспериментах содержащий Methocult-H4330 ритропоэтин был использован вместо Methocult-Н4230, или же добавляли 50 нг/мл фактора стволовых клеток SCF (Biosource Intern., Camarillo, CA). После культивирования в течение 7-9 дней подсчитывали колонии, содержащие более 30 клеток. Подсчеты в тесте проводят "слепым" образом, чтобы избежать субъективных отклонений при подсчете.
Дополнительные подробности по методам рекомбинирования ДНК, которые могут быть использованы для создания вариантов, экспрессии их в бактериях, клетках млекопитающих или клетках насекомых, очистки и рефолдинга желательных белков, и тестам для определения биологической активности белков может быть найдена в описаниях патентов WO 95/00646, WO 94/12639, WO 94/12638, WO 95/20976, WO 95/21197, WO 95/20977, WO 95/21254 и US 08/383035, которые в полном объеме здесь включены в список цитирования.
Дальнейшие подробности, известные специалистам в данной области техники, могут быть найдены у Maniatis et al., 1982, "Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Lab. и ссылках, приведенных в этом издании, а также у Sambroook et al., 1989, "Molecular Cloning, A Laboratory Manual, 2-d ed., Cold Spring Harbor Lab. и в ссылках, приведенных в этом издании.
Табл. 1 - 3 и последовательности приведены после формулы изобретения.
Следующие примеры иллюстрируют изобретение в больших деталях, хотя необходимо понимать, что изобретение не ограничивается этими специфическими примерами.
Пример 1
Конструирование исходного экспрессирующего вектора ВНК
А. Удаление сайта рестрикции AflIII из экспрессирующей плазмиды млекопитающих
Новый экспрессирующий вектор для клеток млекопитающих был сконструирован для соединения генных фрагментов NcoI-HindIII или AflIII-HindIII, находящихся в пределах рамки и 3’-сегмента, к гену агониста рецептора hIL-3 в плазмиде pMON13146 (WO 94/12638) и линкерному фрагменту IgG2b мыши. Во-первых, единственный сайт рестрикции AflIII был удален из плазмиды pMON3934, являющейся дериватом плазмиды pMON359. Плазмида pMON3359 является основанным на плазмиде pUCl8 вектором, содержащим набор элементов для экспрессии в клетках млекопитающих. Этот набор включает герпесвирусный промотор 1Е110 (от -800 до +120) с последующим участком, кодирующим последовательность модифицированного сигнального участка IL-3 человека, и затем поздний полиадениловый сигнал вируса SV40, который субклонируют в полилинкер pUCl8 (см. Hippenmeyer et al., 1993, Bio/Technol., 1037-1041). Модифицированный сигнальный участок IL-3 человека, который обеспечивает секрецию генного продукта за пределами клетки, фланкируют сайтом рестрикции BamHI с 5’-конца и уникальным сайтом рестрикции NcoI с 3’-конца. Уникальный сайт рестрикции HindIII расположен с 3’-стороны от сайта NcoI и с 5’-стороны по отношению к полиадениловому сигналу. Последовательность ДНК, кодирующая сигнальный участок, показана раньше (сайты рестрикции были определены выше). Кодон ATG (метионин) в составе сайта рестрикции NcoI находится в пределах рамки считывания с инициаторным ATG в составе сигнального участка (подчеркнут):
BamHI
5’GGATCCACC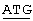 AGCCGCCTGCCCGTCCTGCTCCTGCTCCAACTCCTGGT
AGCCGCCTGCCCGTCCTGCTCCTGCTCCAACTCCTGGT
NcoI
CCGCCCCGCCATGG (SEQ ID NO 857)
Единственный сайт рестрикции AflIII был удален из плазмиды pMON3934 путем расщепления рестриктазой AflIII с последующим заполнением выступов с помощью добавления ДНК-полимеразы и нуклеотидов. Расщепленный фрагмент ДНК очищали с использованием набора Magic PCR Clean-up Kit (Promega) и лигировали с помощью ДНК-лигазы Т4. Продукт лигирования переносили в DH5α™ и клетки высевали на LB-агаре с добавлением ампициллина. Отдельные колонии скринировали на утрату сайта рестрикции AflIII с помощью рестриктазного анализа с AflIII и HindIII, в результате чего был получен единый фрагмент, лишенный сайта AflIII. Полученная в результате плазмида была обозначена pMON30275.
В. Перенос набора элементов pMON13416/IgG2b агониста рецептора hIL-3 в состав pMON30275
Маркируемый рестриктазами NcoI-HindIII фрагмент (длиной примерно 426 пар нуклеотидов) из состава pMON30245 лигировали с фрагментом NcoI-HindIII (длиной примерно 3800 пар нуклеотидов) в составе pMON30275. Плазмида pMON30245 (WO 94/12638) включает ген, кодирующий агонист рецептора hIL-3 pMON13416, соединенный с линкерным фрагментом IgG2b мыши. Сразу с 3’-стороны от IgG2b и 5’-стороны от сайта рестрикции HindIII находится сайт рестрикции AflIII. Гены могут быть клонированы в сайты AflIII-HindIII, как фрагменты NcoI-HindIII или AflIII-HindIII, в пределах рамки кодирования с вариантным линкером pMON13416/IgG2b с целью создания новых химер. Сайты рестрикции NcoI и AflIII характеризуются совместимыми выступами и, соответственно, лигируют друг на друга с потерей сайтов распознавания. Плазмида pMON30304, включающая последовательность ДНК SEQ ID NO 1, кодирующую вариант pMON13416 IL-3, соединенный с линкерным сегментом IgG2c мыши, являлась результатом такого клонирования.
Пример 2
Конструирование промежуточной плазмиды, включающей одну копию гена лиганда с-mpl (1-153) димерной матрицы
С целью создания плазмидной ДНК, включающей кодирующую последовательность лиганда с-mpl (1-153), продолжаемую сайтом рестрикции EcoRI, ген выделяют с применением метода полимеразной цепной реакции с ревертированием (RT-PCR). РНК из плодной (№ 38130) и взрослой (№ 46018) печени человека получают в Clontech (Palo Alto, CA) в качестве источника информационной РНК (мРНК) лиганда c-mpl. Реакции по первой цепи кДНК проводят с использованием набора cDNA Cycle™ Kit (Invitrogen, San Diego, CA). В реакции ревертирования случайно отобранные затравки и олиго-дТ затравка используются для создания кДНК на основе комбинации мРНК плодной и взрослой печени человека. Для амплификации фрагмента гена лиганда с-mpl, кодирующего аминокислоты 1-153, продукт ревертирования используют в качестве матрицы для ПЦР с комбинацией затравок: ведущая затравка - c-mplNcoI (SEQ ID NO 317) и обратная затравка - Ecompl. Затравку c-mplNcoI отжигают на ген лиганда с-mpl (основания 279-311 по последовательности гена лиганда с-mpl из базы данных Genbank за №L33410 или по данным de Sauvage et al., 1994, Nature, 369, 533-538) и помещают сайт рестрикции NcoI сразу с 5’-конца от первого кодона (серин+1) лиганда с-mpl. Сайт рестрикции NcoI включает кодоны метионина и аланина перед Ser+1 и характеризуется вырожденностью по кодону аланина и первых четырех кодонов лиганда с-mpl (Ser, Pro, Ala, Pro). Затравку Ecompl отжигают на основания 720-737 гена лиганда с-mpl: она содержит сайт рестрикции EcoRI (GAATTC) в пределах рамки кодирования, располагающийся за кодирующей частью гена лиганда c-mpl сразу за кодоном аргинина153. Сайт рестрикции EcoRI создает кодоны глутаминовой кислоты и фенилаланина вслед за кодоном Arg153. Продукт ПЦР длиной примерно 480 нуклеотидов очищали, расщепляли рестриктазами NcoI и EcoRI и лигировали во фрагмент NcoI-EcoRI вектора pMON3993 (примерно 4550 нуклеотидов). Плазмида pMON3993 является дериватом плазмиды pMON3359 (описанной в примере 1). Последовательность сигнального участка IL-3 человека, которая была субклонирована в виде BamHI-фрагмента в уникальный сайт рестрикции BamHI между промотором 1Е110 и полиадениловым сигналом, содержит сайт рестрикции NcoI в его 3’-части и завершается уникальным сайтом рестрикции EcoRI. Результатом данного клонирования оказывается плазмида pMON26458, содержащая последовательность ДНК SEQ ID NO 1, кодирующую аминокислоты 1-153 лиганда c-mpl.
Пример 3
Конструирование исходных плазмид, содержащих вторые гены димерных матриц
Для амплификации фрагментов гена лиганда c-mpl, начинающихся с 1-й аминокислоты (серин) со стоп-кодоном вслед за кодоном 153-й аминокислоты (аргинин) реакция ревертирования согласно примеру 2 используется в качестве матрицы для ПЦР с комбинацией следующих затравок: c-mplNcoI (SEQ ID NO 317) (ведущая затравка) и c-mplHindIII (SEQ ID NO 319) (обратная затравка). Затравка c-mplNcoI (SEQ ID NO 317) описана в примере 2. Затравка c-mpl-HindIII (SEQ ID NO 319), которую отжигают на основания 716-737 гена лиганда c-mpl, добавляет и стоп-кодон, и сайт рестрикции HindIII сразу за последним кодоном Arg153.
Создают два типа продуктов ПЦР на образцах кДНК после ревертирования: один с делецией кодонов аминокислот 112-115 и другой без делеции этих кодонов. ПЦР-продукт лиганда c-mpl (длиной примерно 480 нуклеотидов) расщепляют рестриктазами NcoI и HindIII для переноса в экспрессирущий вектор pMON3934 для клеток млекопитающих. Плазмиду pMON3934 расщепляют рестриктазами NcoI и HindIII (длина примерно 3800 нуклеотидов) с целью присоединения продуктов ПЦР.
Плазмида pMON32132 (SEQ ID NO 84), кодирующая аминокислоты 1-153 лиганда с-mpl (SEQ ID NO 546), явилась результатом такого клонирования. Плазмида pMON32134 (SEQ ID NO 86), кодирующая аминокислоты 1-153 лиганда с-mpl (SEQ ID NO 548), явилась результатом такого клонирования. Плазмида pMON32133 (SEQ ID NO 86), кодирующая аминокислоты 1-153 лиганда с-mpl с утратой кодонов 112-115 (Δ112-115) (SEQ ID NO 547), явилась результатом такого клонирования.
Пример 4
Создание димерных ПЦР-матриц 5L с делецией 112-115 во втором гене лиганда с-mpl
ПЦР-матрицу для создания новых форм лиганда с-mpl конструируют путем лигирования фрагмента BstXI-EcoRI длиной 3700 нуклеотидов в pMON-26458 с фрагментом NcoI-BstXI из pMON32133 (содержащим делецию аминокислот 112-115) вместе с синтетическими олигонуклеотидными линкерами EcoRI-AflIII 5L-5’ (SEQ ID NO 322) и 5L-3’ (SEQ ID NO 323).
EcoRI-конец линкера лигируют на EcoRI-конец плазмиды pMON26458. AflIII-конец линкера лигируют на сайт рестрикции NcoI в плазмиде pMON32133: т.е. после лигирования свободных сайтов рестрикции не остается. Также лигируют сайты рестрикции BstXI в составе плазмид pMON26458 и pMON32133. Плазмида pMON28548 является результатом клонирования и содержит последовательность ДНК (SEQ ID NO 3), которая кодирует аминокислоты 1-153 лиганда с-mpl, соединенные через линкер GluPheGlyGlyAsnMet (SEQ ID NO 783) с кодонами аминокислот 1-153 лиганда с-mpl при делеции аминокислот 112-115 (SEQ ID NO 468).
Пример 5
Создание димерной ПЦР-матрицы 4L
ПЦР-матрицу для создания новых форм лиганда с-mpl конструируют путем лигирования фрагмента BstXI-EcoRI длиной 3700 нуклеотидов в pMON26458 с фрагментом NcoI-BstXI из pMON32132 (содержащим делецию аминокислот 112-115) вместе с синтетическими олигонуклеотидными линкерами EcoRI-AflIII 4L-5’ (SEQ ID NO 320) и 4L-3’ (SEQ ID NO 321).
EcoRI-конец линкера лигируют на EcoRI-конец плазмиды pMON26458. AflIII-конец линкера лигируют на сайт рестрикции NcoI в плазмиде pMON32132: т.е. после лигирования свободных сайтов рестрикции не остается. Также лигируют сайты рестрикции BstXI в составе плазмид pMON26458 и pMON32132. Плазмида pMON28500 является результатом клонирования и содержит последовательность ДНК (SEQ ID NO 4), которая кодирует аминокислоты 1-153 лиганда с-mpl, соединенные через линкер 4L GluPheGlyAsnMetAla (SEQ ID NO 223) с кодонами аминокислот 1-153 лиганда с-mpl (SEQ ID NO 469).
Пример 6
Создание димерной ПЦР-матрицы 5L
ПЦР-матрицу для создания новых форм лиганда с-mpl конструируют путем лигирования фрагмента BstXI-EcoRI длиной 3700 нуклеотидов в pMON26458 с фрагментом NcoI-BstXI из pMON32132 (содержащим делецию аминокислот 112-115) вместе с синтетическими олигонуклеотидными линкерами EcoRI-AflIII 5L-5’ (SEQ ID NO 322) и 5L-3’ (SEQ ID NO 323).
EcoRI-конец линкера лигируют на EcoRI-конец плазмиды pMON26458. AflIII-конец линкера лигируют на сайт рестрикции NcoI в плазмиде pMON32132: т.е. после лигирования свободных сайтов рестрикции не остается. Также лигируют сайты рестрикции BstXI в составе плазмид pMON26458 и pMON32132. Плазмида pMON28501 является результатом клонирования и содержит последовательность ДНК (SEQ ID NO 4), которая кодирует аминокислоты 1-153 лиганда с-mpl, соединенные через линкер 4L GluPheGlyGlyAsnMet (SEQ ID NO 783) с кодонами аминокислот 1-153 лиганда c-rnpl (SEQ ID NO 470).
Пример 7
Создание димерной ПЦР-матрицы 8L
ПЦР-матрицу для создания новых форм лиганда с-mpl конструируют путем лигирования фрагмента BstXI-EcoRI длиной 3700 нуклеотидов в pMON26458 с фрагментом NcoI-BstXI из pMON32134 (содержащим делецию аминокислот 112-115) вместе с синтетическими олигонуклеотидными линкерами EcoRI-AflIII 8L-5’ (SEQ ID NO 324) и 8L-3’ (SEQ ID NO 325).
EcoRI-конец линкера лигируют на EcoRI-конец плазмиды pMON26458. AflIII-конец линкера лигируют на сайт рестрикции NcoI в плазмиде pMON32132: т.е. после лигирования свободных сайтов рестрикции не остается. Также лигируют сайты рестрикции BstXI в составе плазмид pMON26458 и pMON32134. Плазмида pMON28502 является результатом клонирования и содержит последовательность ДНК (SEQ ID NO 6), которая кодирует аминокислоты 1-153 лиганда с-mpl, соединенные через линкер 8L GluPheGlyGlyAsnGlyGlyAsnMetAla (SEQ ID NO 224) с кодонами аминокислот 1-153 лиганда c-rnpl (SEQ ID NO 471).
Примеры 8-44
Создание новых генов лигандов с-mpl с новыми N-концом и С-концом
А. ПЦР-генерирование генов, кодирующих новые агонисты рецептора лиганда с-mpl
Гены, кодирующие новые агонисты рецептора лиганда с-mpl, были созданы с использованием способа III (Horlick et al., 1992, Protein Eng., 5, 427-433). Реакции ПЦР проводили с использованием димерных матриц (pMONs 28500, 28501, 28502 или 28548) и одного из наборов синтетических затравок, приведенных ниже. (Первое число обозначает положение первой аминокислоты в исходной последовательности. Например, 31-5’ и 31-3’ обозначает 5’- и 3’-олигозатравки, соответственно, для последовательностей, начинающихся с кодона, кодирующего 31-й аминокислотный остаток в исходной последовательности полипептида).
31-5’ (SEQ ID NO 326) и 31-3’ (SEQ ID NO 327), 35-5’ (SEQ ID NO 328) и 35-3’ (SEQ ID NO 329), 39-5’ (SEQ ID NO 330) и 39-3’ (SEQ ID NO 331), 43-5’ (SEQ ID NO 332) и 43-3’ (SEQ ID NO 333), 45-5’ (SEQ ID NO 334) и 45-3’ (SEQ ID NO 335), 49-5’ (SEQ ID NO 336) и 49-3’ (SEQ ID NO 337), 82-5’ (SEQ ID NO 338) и 82-3’ (SEQ ID NO 339), 109-5’ (SEQ ID NO 340) и 109-3’ (SEQ ID NO 341), 115-5’ (SEQ ID NO 342) и 115-3’ (SEQ ID NO 343), 120-5’ (SEQ ID NO 344) и 120-3’ (SEQ ID NO 345), 123-5’ (SEQ ID NO 346) и 123-3’ (SEQ ID NO 347), 126-5’ (SEQ ID NO 348) и 126-3’ (SEQ ID NO 349).
Матрицы и наборы олигонуклеотидов, использованные для ПЦР, показаны в табл.4. Продукты, которые создаются, характеризуются длиной около 480 нуклеотидов: их очищали с применением наборов Magic PCR Clean-up Kit (Promega).
В. Субклонирование продуктов новых гена агонистов рецепторов с-mpl в экспрессирующий вектор для клеток млекопитающих для создания химер
ПЦР-продукты гена агонистов рецепторов с-mpl расщепляли рестриктазами NcoI и HindIII либо AflIII и HindIII (длиной примерно 470 нуклеотидов) для переноса в экспрессирующий вектор для клеток млекопитающих. Экспрессирующий вектор pMON30304 расщепляли рестриктазами NcoI и HindIII (длиной примерно 4200 нуклеотидов) и присоединяли ПЦР-продукты как фрагменты NcoI-HindIII или AflIII-HindIII. Параметрами рестрикции ПЦР-продуктов и результирующих плазмид показаны в табл. 4.
Пример 45
Конструирование pMON15960
Сконструирована плазмида pMON15960, являющаяся промежуточной плазмидой, используемой в конструировании плазмид, включающих последовательности ДНК, кодирующих G-CSF с серином17 с новыми N-концом и С-концом. ДНК плазмиды рАСУС177 (Chang & Cohen, 1978, J. Bacteriol., 134, 1141-1156) была расщеплена рестриктазами HindIII и BaniHI: в результате образуется HindIII, ВаmНI-фрагмент длиной 3092 нуклеотидов. ДНК плазмиды pMON13037 (WO 95/21254) расщепляли рестриктазами BglII и Fspl: в результате образуется BglII, Fspl-фрагмент длиной 616 нуклеотидов.
ДНК второго образца плазмиды pMONl3037 расщепляли рестриктазами NcoI и HindIII: в результате образуется NcoI,HindIII-фрагмент длиной 556 нуклеотидов. Синтетические ДНК-олигонуклеотиды 1GGGS (SEQ ID NO 380) (прямая затравка) и 1GGGS (SEQ ID NO 381) (обратная затравка) проходили отжиг друг на друга: затем их расщепляли рестриктазами AflIII и Fspl, в результате чего образовывался Af1III, Fspl-фрагмент длиной 21 нуклеотид. Рестрикционные фрагменты лигировали, а реакционную смесь лигирования использовали для трансформации клеток E.coli K-12 штамма JM101. Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и анализировали с использованием рестриктаз для подтверждения точности вставки.
Пример 46
Конструирование pMON15981
Осуществлено конструирование pMON15981 - плазмиды, включающей последовательности ДНК, кодирующие многофункциональный агонист гемопоэтических рецепторов. ДНК плазмиды pMON15960 расщепляли рестриктазой Smal и использовали в качестве матрицы для ПЦР с использованием синтетических ДНК-олигонуклеотидов 38 (SEQ ID NO 369) (концевая затравка) и 39 (SEQ ID NO 368) (стартовая затравка), в результате чего амплифицировался фрагмент ДНК длиной 576 нуклеотидов. Амплифицированный фрагмент расщепляли рестриктазами HindIII и NcoI, в результате чего образовывался HindIII, NcoI-фрагмент длиной 558 нуклеотидов. ДНК плазмиды pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего образовывался HindIII, AflIII-фрагмент длиной 4068 нуклеотидов. Рестрикционные фрагменты лигировали, а реакционную смесь лигирования использовали для трансформации клеток E.coli K-12 штамма JM101. Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли, анализировали с помощью рестриктаз и секвенировали для подтверждения точности вставки. Плазмида pMON15981 включает последовательность ДНК (SEQ ID NO 78), кодирующую следующую аминокислотную последовательность:

Пример 47
Конструирование pMON15982
Осуществлено конструирование pMON15982 - плазмиды, включающей последовательности ДНК, кодирующие многофункциональный агонист гемопоэтических рецепторов. ДНК плазмиды pMON15960 расщепляли рестриктазой Smal и использовали в качестве матрицы для ПЦР с использованием синтетических ДНК-олигонуклеотидов 96 (SEQ ID NO 370) (концевая затравка) и 97 (SEQ ID NO 370) (стартовая затравка), в результате чего амплифицировался фрагмент ДНК длиной 576 нуклеотидов. Амплифицированный фрагмент расщепляли рестриктазами HindIII и NcoI, в результате чего образовывался HindIII, NcoI-фрагмент длиной 558 нуклеотидов. ДНК плазмиды pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего образовывался HindIII, АflIII-фрагмент длиной 4068 нуклеотидов. Рестрикционные фрагменты лигировали, а реакционную смесь лигирования использовали для трансформации клеток E.coli K-12 штамма JM101. Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли, анализировали с помощью рестриктаз и секвенировали для подтверждения точности вставки. Плазмида pMON15982 включает последовательность ДНК (SEQ ID NO 79), кодирующую следующую аминокислотную последовательность:

Пример 48
Конструирование pMON15965
Осуществлено конструирование pMON15965 - плазмиды, включающей последовательности ДНК, кодирующие многофункциональный агонист гемопоэтических рецепторов. ДНК плазмиды pMON15960 расщепляли рестриктазой Smal и использовали в качестве матрицы для ПЦР с использованием синтетических ДНК-олигонуклеотидов 142 (SEQ ID NO 377) (концевая затравка) и 141 (SEQ ID NO 376) (стартовая затравка), в результате чего амплифицировался фрагмент ДНК длиной 576 нуклеотидов. Амплифицированный фрагмент расщепляли рестриктазами HindIII и NcoI, в результате чего образовывался НindIII, NсоI-фрагмент длиной 558 нуклеотидов. ДНК плазмиды pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего образовывался НiildIII, АflIII-фрагмент длиной 4068 нуклеотидов. Рестрикционные фрагменты лигировали, а реакционную смесь лигирования использовали для трансформации клеток E.coli K-12 штамма JM101. Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли, анализировали с помощью рестриктаз и секвенировали для подтверждения точности вставки. Плазмида pMON15965 включает последовательность ДНК (SEQ ID NO 80), кодирующую следующую аминокислотную последовательность:
Пример 49
Конструирование pMON15966
Осуществлено конструирование pMON15966 - плазмиды, включающей последовательности ДНК, кодирующие многофункциональный агонист гемопоэтических рецепторов. ДНК плазмиды pMON15960 расщепляли рестриктазой Smal и использовали в качестве матрицы для ПЦР с использованием синтетических ДНК-олигонуклеотидов 126 (SEQ ID NO 372) (концевая затравка) и 125 (SEQ ID NO 373) (стартовая затравка), в результате чего амплифицировался фрагмент ДНК длиной 576 нуклеотидов. Амплифицированный фрагмент расщепляли рестриктазами HindIII и NcoI, в результате чего образовывался НindIII, NсоI-фрагмент длиной 558 нуклеотидов. ДНК плазмиды pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего образовывался НindIII, АflIII-фрагмент длиной 4068 нуклеотидов. Рестрикционные фрагменты лигировали, а реакционную смесь лигирования использовали для трансформации клеток E.coli K-12 штамма JM101.
Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли, анализировали с помощью рестриктаз и секвенировали для подтверждения точности вставки. Плазмида pMON15966 включает последовательность ДНК (SEQ ID NO 81), кодирующую следующую аминокислотную последовательность:

Пример 50
Конструирование pMON15967
Осуществлено конструирование pMON15967 - плазмиды, включающей последовательности ДНК, кодирующие многофункциональный агонист гемопоэтических рецепторов. ДНК плазмиды pMON15960 расщепляли рестриктазой SmaI и использовали в качестве матрицы для ПЦР с использованием синтетических ДНК-олигонуклеотидов 132 (SEQ ID NO 375) (концевая затравка) и 133 (SEQ ID NO 374) (стартовая затравка), в результате чего амплифицировался фрагмент ДНК длиной 576 нуклеотидов. Амплифицированный фрагмент расщепляли рестриктазами HindIII и NcoI, в результате чего образовывался HindIII, NcoI-фрагмент длиной 558 нуклеотидов. ДНК плазмиды pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего образовывался HindIII, AflIII-фрагмент длиной 4068 нуклеотидов. Рестрикционные фрагменты лигировали, а реакционную смесь лигирования использовали для трансформации клеток E.coli K-12 штамма JM101. Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли, анализировали с помощью рестриктаз и секвенировали для подтверждения точности вставки. Плазмида pMON15967 включает последовательность ДНК (SЕQ ID NO 82), кодирующую следующую аминокислотную последовательность:

Пример 51
Конструирование pMON13180, являющейся промежуточной плазмидой, используемой для конструирования плазмид, которые включают последовательность ДНК, кодирующую многофункциональные агонисты гемопоэтических рецепторов
ДНК плазмиды pMON13046 (WO 95/21254) расщепляли рестриктазами XmaI и SnaBI, в результате чего образовывался векторный фрагмент длиной 4018 нуклеотидов. Векторный XmaI,SnaBI-фрагмент, состоящий из 4018 нуклеотидов, очищали с применением набора Magic DNA Clean-up Kit (Promega), в результате чего 25-нуклеотидный вставочный XmaI, SnaBI-фрагмент не сохранялся. Комплементарную пару олигонуклеотидных затравок glyxal (SEQ ID NO 378) и glyxa2 (SEQ ID NO 379) формировали таким образом, чтобы удалить последовательность, кодирующую сайт расщепления фактора Ха. При условии правильной сборки эти олигонуклеотиды также способствуют появлению концов XmaI и SnaBI. Затравки Glyxal и Glyxa2 отжигали в соответствующем буфере (20 мМ Трис-HCl, рН=7,5, 10 мМ MgCl2, 50 мМ NaCl) путем нагревания до 70°С на 10 минут с последующим медленным охлаждением. XmaI-SnaBI-фрагмент длиной 4018 нуклеотидов из плазмиды pMON13046 лигировали со скомпонованными олигонуклеотидами с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли из трансформантов и анализировали в тесте, основанном на ПЦР. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок олигонуклеотидов. Полученную в результате плазмиду обозначили pMON13180 (SEQ ID NO 88).
Пример 52
Конструирование pMON13181, являющейся промежуточной плазмидой, используемой для конструирования плазмид, которые включают последовательность ДНК, кодирующую многофункциональные агонисты гемопоэтических рецепторов
ДНК плазмиды pMON13047 (WO 95/21254) расщепляли рестриктазами XmaI и SnaBI, в результате чего образовывался векторный фрагмент длиной 4063 нуклеотидов. Векторный XmaI, SnaBI-фрагмент, состоящий из 4063 нуклеотидов, очищали с применением набора Magic DNA Clean-up Kit (Promega), в результате чего 25-нуклеотидный вставочный Xinal, SnaBI-фрагмент не сохранялся. Комплементарную пару олигонуклеотидных затравок glyxal (SEQ ID NO 378) и glyxa2 (SEQ ID NO 379) формировали таким образом, чтобы удалить последовательность, кодирующую сайт расщепления фактора Ха. При условии правильной сборки эти олигонуклеотиды также способствуют появлению концов XmaI и SnaBI. Затравки Glyxal и Glyxa2 отжигали в соответствующем буфере (20 мМ Трис-HCl, рН=7,5, 10 мМ MgCl2, 50 мМ NaCl) путем нагревания до 70°С на 10 минут с последующим медленным охлаждением. XmaI-SnaBI-фрагмент длиной 4063 нуклеотидов из плазмиды pMON13046 лигировали со скомпонованными олигонуклеотидами с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли из трансформантов и анализировали в тесте, основанном на ПЦР. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок олигонуклеотидов. Полученную в результате плазмиду обозначили pMON13181 (SEQ ID NO 87).
Пример 53
Конструирование pMON13182
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13182 был создан с использованием способа I, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 39 (SEQ ID NO 368) и стартовую L-11 (SEQ ID NO 364). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 38 (SEQ ID NO 369) и концевую L-11 (SEQ ID NO 365). Полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом был создан и амплифицирован путем отжига стартовых и концевых фрагментов с использованием затравок стартовой 39 и концевой 38.
Полученный фрагмент ДНК, содержавший новый ген, расщепляли рестриктазами NcoI и HindIII и очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Промежуточную плазмиду pMON13180 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4023 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DHSα (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок. Полученную в результате плазмиду обозначили pMON13182.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13182 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13182 включает последовательность ДНК (SEQ ID NO 17), кодирующую следующую аминокислотную последовательность:

Пример 54
Конструирование pMON13183
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13183 был создан с использованием способа I, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 39 (SEQ ID NO 368) и стартовую L-11 (SEQ ID NO 364). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 38 (SEQ ID NO 369) и концевую L-11 (SEQ ID NO 365). Полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом был создан и амплифицирован путем отжига стартовых и концевых фрагментов с использованием затравок стартовой 39 и концевой 38.
Полученный фрагмент ДНК, содержавший новый ген, расщепляли рестриктазами NcoI и HindIII и очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Промежуточную плазмиду pMONl3181 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4068 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок. Полученную в результате плазмиду обозначили pMON13183.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13183 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMONl3183 включает последовательность ДНК (SEQ ID NO 18), кодирующую следующую аминокислотную последовательность:

Пример 55
Конструирование pMONl3184
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13184 был создан с использованием способа I, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 97 (SEQ ID NO 370) и стартовую L-11 (SEQ ID NO 364). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 96 (SEQ ID NO 371) и концевую L-11 (SEQ ID NO 365). Полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом был создан и амплифицирован путем отжига стартовых и концевых фрагментов с использованием затравок стартовой 97 и концевой 96.
Полученный фрагмент ДНК, содержавший новый ген, расщепляли рестриктазами NcoI и HindIII и очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Промежуточную плазмиду pMON13180 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4023 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок. Полученную в результате плазмиду обозначили pMON13184.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13184 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13184 включает последовательность ДНК (SEQ ID NO 19), кодирующую следующую аминокислотную последовательность:

Пример 56
Конструирование pMON13185
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13185 был создан с использованием способа I, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 97 (SEQ ID NO 370) и стартовую L-11 (SEQ ID NO 364). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 96 (SEQ ID NO 371) и концевую L-11 (SEQ ID NO 365). Полноразмерный ген G-CSF Serl7 с новыми N-концом/С-концом был создан и амплифицирован путем отжига стартовых и концевых фрагментов с использованием затравок стартовой 97 и концевой 96.
Полученный фрагмент ДНК, содержавший новый ген, расщепляли рестриктазами NcoI и HindIII и очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Промежуточную плазмиду pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4068 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок. Полученную в результате плазмиду обозначили pMON13185.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13185 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13185 включает последовательность ДНК (SЕQ ID NO 20), кодирующую следующую аминокислотную последовательность:

Пример 57
Конструирование pMONl3186
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13186 был создан с использованием способа I, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 126 (SEQ ID NO 372) и стартовую L-11 (SEQ ID NO 364). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF SER17 из состава pMON13037 с использованием набора затравок, включающего концевую 125 (SEQ ID NO 373) и концевую L-11 (SEQ ID NO 365). Полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом был создан и амплифицирован путем отжига стартовых и концевых фрагментов с использованием затравок стартовой 126 и концевой 125.
Полученный фрагмент ДНК, содержавший новый ген, расщепляли рестриктазами NcoI и HindIII и очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Промежуточную плазмиду pMON13180 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4023 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок. Полученную в результате плазмиду обозначили pMON13186.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13186 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13186 включает последовательность ДНК (SEQ ID NO 21), кодирующую следующую аминокислотную последовательность:

Пример 58
Конструирование pMON13187
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13187 был создан с использованием способа I, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 126 (SEQ ID NO 372) и стартовую L-11 (SEQ ID NO 364). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 125 (SEQ ID NO 373) и концевую L-11 (SEQ ID NO 365). Полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом был создан и амплифицирован путем отжига стартовых и концевых фрагментов с использованием затравок стартовой 126 и концевой 125.
Полученный фрагмент ДНК, содержавший новый ген, расщепляли рестриктазами NcoI и HindIII и очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Промежуточную плазмиду pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4068 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок. Полученную в результате плазмиду обозначили pMON13187.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13187 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13187 включает последовательность ДНК (SEQ ID NO 22), кодирующую следующую аминокислотную последовательность: 
Пример 59
Конструирование pMON13188
Ген белка с новыми М-концом/С-концом в составе плазмиды pMON13188 был создан с использованием способа I, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 133 (SEQ ID NO 374) и стартовую L-11 (SEQ ID NO 364). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 132 (SEQ ID NO 375) и концевую L-11 (SEQ ID NO 365). Полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом был создан и амплифицирован путем отжига стартовых и концевых фрагментов с использованием затравок стартовой 133 и концевой 132.
Полученный фрагмент ДНК, содержавший новый ген, расщепляли рестриктазами NcoI и HindIII и очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Промежуточную плазмиду pMON13180 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4023 нуклеотидов, который очищали с использованием Маус DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок. Полученную в результате плазмиду обозначили pMON13188.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13188 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13188 включает последовательность ДНК (SEQ ID NO 23), кодирующую следующую аминокислотную последовательность:

Пример 60
Конструирование pMON13189
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13189 был создан с использованием способа I, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 133 (SEQ ID NO 374) и стартовую L-11 (SEQ ID NO 364). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 132 (SEQ ID NO 375) и концевую L-11 (SEQ ID NO 365). Полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом был создан и амплифицирован путем отжига стартовых и концевых фрагментов с использованием затравок стартовой 133 и концевой 132.
Полученный фрагмент ДНК, содержавший новый ген, расщепляли рестриктазами NcoI и HindIII и очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Промежуточную плазмиду pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4068 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок. Полученную в результате плазмиду обозначили pMON13189.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13189 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13189 включает последовательность ДНК (SEQ ID NO 24), кодирующую следующую аминокислотную последовательность:

Пример 61
Конструирование pMON13190
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13190 был создан с использованием способа I, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амллифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 142 (SEQ ID NO 376) и стартовую L-11 (SEQ ID NO 364). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 141 (SEQ ID NO 377) и концевую L-11 (SEQ ID NO 365). Полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом был создан и амплифицирован путем отжига стартовых и концевых фрагментов с использованием затравок стартовой 142 и концевой 141.
Полученный фрагмент ДНК, содержавший новый ген, расщепляли рестриктазами NcoI и HindIII и очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Промежуточную плазмиду pMON13180 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4023 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин.
Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок. Полученную в результате плазмиду обозначили pMON13190.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13190 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13190 включает последовательность ДНК (SEQ ID NO 25), кодирующую следующую аминокислотную последовательность:

Пример 62
Конструирование pMON13191
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13191 был создан с использованием способа I, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 142 (SEQ ID NO 376) и стартовую L-11 (SEQ ID NO 364). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 141 (SEQ ID NO 377) и концевую L-11 (SEQ ID NO 365). Полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом был создан и амплифицирован путем отжига стартовых и концевых фрагментов с использованием затравок стартовой 142 и концевой 141.
Полученный фрагмент ДНК, содержавший новый ген, расщепляли рестриктазами NcoI и HindIII и очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Промежуточную плазмиду pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4068 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставок. Полученную в результате плазмиду обозначили pMON13191.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13191 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13191 включает последовательность ДНК (SEQ ID NO 26), кодирующую следующую аминокислотную последовательность:

Пример 63
Конструирование pMON13192
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13192 был создан с использованием способа II, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 39 (SEQ ID NO 368) и стартовую Р-bl (SEQ ID NO 366). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 38 (SEQ ID NO 369) и концевую Р-bl (SEQ ID NO 367). Стартовый фрагмент расщепляли рестриктазой NcoI, а концевой фрагмент - рестриктазой HindIII. После очистки расщепленные стартовый и концевой фрагменты комбинировали и лигировали в векторный NcoI, HindIII-фрагмент длиной примерно 3800 нуклеотидов в составе плазмиды pMON3934.
Промежуточную плазмиду, описанную выше, содержащую полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом, расщепляли рестриктазами NcoI и HindIII. Расщепленную ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом выделяли с использованием реактивов Geneclean (Bio101, Vista, СА). Промежуточную плазмиду pMON13180 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4023 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставки нового гена. Полученную в результате плазмиду обозначили pMON13192.
Клетки E.coli штамма JM101 трансформировали плазмидой pMONl3192 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMONl3192 включает последовательность ДНК (SEQ ID NO 27), кодирующую следующую аминокислотную последовательность:

Пример 64
Конструирование pMON13193
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13193 был создан с использованием способа II, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 39 (SEQ ID NO 368) и стартовую Р-bl (SEQ ID NO 366). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 38 (SEQ ID NO 369) и концевую P-bl (SEQ ID NO 367). Стартовый фрагмент расщепляли рестриктазой NcoI, а концевой фрагмент - рестриктазой HindIII. После очистки расщепленные стартовый и концевой фрагменты комбинировали и лигировали в векторный NcoI, HindIII-фpaгмeнт длиной примерно 3800 нуклеотидов в составе плазмиды pMON3934.
Промежуточную плазмиду, описанную выше, содержащую полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом, расщепляли рестриктазами NcoI и HindIII. Расщепленную ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом выделяли с использованием реактивов Geneclean (Bio101, Vista, СА). Промежуточную плазмиду pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4068 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставки нового гена. Полученную в результате плазмиду обозначили pMON13193.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13193 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13193 включает последовательность ДНК (SEQ ID NO 28), кодирующую следующую аминокислотную последовательность:

Пример 65
Конструирование pMON25190
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON25190 был создан с использованием способа II, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 97 (SEQ ID NO 370) и стартовую Р-bl (SEQ ID NO 366). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 96 (SEQ ID NO 371) и концевую Р-bl (SEQ ID NO 367). Стартовый фрагмент расщепляли рестриктазой NcoI, а концевой фрагмент - рестриктазой HindIII. После очистки расщепленные стартовый и концевой фрагменты комбинировали и лигировали в векторный NcoI, HindIII-фрагмент длиной примерно 3800 нуклеотидов в составе плазмиды pMON3934.
Промежуточную плазмиду, описанную выше, содержащую полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом, расщепляли рестриктазами NcoI и HindIII. Расщепленную ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом выделяли с использованием реактивов Geneclean (Bio101, Vista, СА). Промежуточную плазмиду pMON13180 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4023 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставки нового гена. Полученную в результате плазмиду обозначили pMON25190.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON25190 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON25190 включает последовательность ДНК (SEQ ID NO 29), кодирующую следующую аминокислотную последовательность:

Пример 66
Конструирование pMON25191
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON25191 был создан с использованием способа II, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 97 (SEQ ID NO 370) и стартовую Р-bl (SEQ ID NO 366). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 96 (SEQ ID NO 371) и концевую Р-bl (SEQ ID NO 367). Стартовый фрагмент расщепляли рестриктазой NcoI, а концевой фрагмент - рестриктазой HindIII. После очистки расщепленные стартовый и концевой фрагменты комбинировали и лигировали в векторный NcoI, HindIII-фрагмент длиной примерно 3800 нуклеотидов в составе плазмиды pMON3934.
Промежуточную плазмиду, описанную выше, содержащую полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом, расщепляли рестриктазами NcoI и HindIII. Расщепленную ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом выделяли с использованием реактивов Geneclean (Bio101, Vista, СА). Промежуточную плазмиду pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4068 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставки нового гена. Полученную в результате плазмиду обозначили pMON25191.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON25191 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON25191 включает последовательность ДНК (SEQ ID NO 30), кодирующую следующую аминокислотную последовательность:

Пример 67
Конструирование pMON13194
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13194 был создан с использованием способа II, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 126 (SEQ ID NO 372) и стартовую Р-Ы (SEQ ID NO 366). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 125 (SEQ ID NO 373) и концевую Р-bl (SEQ ID NO 367). Стартовый фрагмент расщепляли рестриктазой NcoI, а концевой фрагмент - рестриктазой HindIII. После очистки расщепленные стартовый и концевой фрагменты комбинировали и лигировали в векторный NcoI, HindIII-фрагмент длиной примерно 3800 нуклеотидов в составе плазмиды pMON3934.
Промежуточную плазмиду, описанную выше, содержащую полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом, расщепляли рестриктазами NcoI и HindIII. Расщепленную ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом выделяли с использованием реактивов Geneclean (Bio101, Vista, СА). Промежуточную плазмиду pMON13180 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4023 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставки нового гена. Полученную в результате плазмиду обозначили pMON13194.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13194 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13194 включает последовательность ДНК (SEQ ID NO 31), кодирующую следующую аминокислотную последовательность:

Пример 68
Конструирование pMON13195
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13195 был создан с использованием способа II, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 126 (SEQ ID NO 372) и стартовую P-bl (SEQ ID NO 366). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 125 (SEQ ID NO 373) и концевую P-bl (SEQ ID NO 367). Стартовый фрагмент расщепляли рестриктазой NcoI, а концевой фрагмент - рестриктазой HindIII. После очистки расщепленные стартовый и концевой фрагменты комбинировали и лигировали в векторный NcoI, HindIII-фрагмент длиной примерно 3800 нуклеотидов в составе плазмиды pMON3934.
Промежуточную плазмиду, описанную выше, содержащую полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом, расщепляли рестриктазами NcoI и HindIII. Расщепленную ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом выделяли с использованием реактивов Geneclean (Bio101, Vista, СА). Промежуточную плазмиду pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4068 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Manhleim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставки нового гена. Полученную в результате плазмиду обозначили pMON13195.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13195 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13195 включает последовательность ДНК (SEQ ID NO 32), кодирующую следующую аминокислотную последовательность:
Пример 69
Конструирование pMON13196
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13196 был создан с использованием способа II, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 133 (SEQ ID NO 374) и стартовую Р-bl (SEQ ID NO 366). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 132 (SEQ ID NO 375) и концевую Р-bl (SEQ ID NO 367). Стартовый фрагмент расщепляли рестриктазой NcoI, а концевой фрагмент - рестриктазой HindIII. После очистки расщепленные стартовый и концевой фрагменты комбинировали и лигировали в векторный NcoI, HindiII-фрагмент длиной примерно 3800 нуклеотидов в составе плазмиды pMON3934.
Промежуточную плазмиду, описанную выше, содержащую полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом, расщепляли рестриктазами NcoI и HindIII. Расщепленную ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом выделяли с использованием реактивов Geneclean (Bio101, Vista, СА). Промежуточную плазмиду pMON13180 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4023 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставки нового гена. Полученную в результате плазмиду обозначили pMON13196.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13196 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13196 включает последовательность ДНК (SEQ ID NO 33), кодирующую следующую аминокислотную последовательность:

Пример 70
Конструирование pMON13197
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13197 был создан с использованием способа II, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 133 (SEQ ID NO 374) и стартовую P-bl (SEQ ID NO 366). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 132 (SEQ ID NO 375) и концевую Р-bl (SEQ ID NO 367). Стартовый фрагмент расщепляли рестриктазой NcoI, а концевой фрагмент - рестриктазой HindIII. После очистки расщепленные стартовый и концевой фрагменты комбинировали и лигировали в векторный NcoI, HindiII-фрагмент длиной примерно 3800 нуклеотидов в составе плазмиды pMON3934.
Промежуточную плазмиду, описанную выше, содержащую полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом, расщепляли рестриктазами NcoI и HindIII. Расщепленную ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом выделяли с использованием реактивов Geneclean (Bio101, Vista, СА). Промежуточную плазмиду pMON13181 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4068 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставки нового гена. Полученную в результате плазмиду обозначили pMON13197.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13197 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13197 включает последовательность ДНК (SEQ ID NO 34), кодирующую следующую аминокислотную последовательность:

Пример 71
Конструирование pMON13198
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13198 был создан с использованием способа II, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 142 (SEQ ID NO 376) и стартовую Р-bl (SEQ ID NO 366). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 141 (SEQ ID NO 377) и концевую Р-bl (SEQ ID NO 367). Стартовый фрагмент расщепляли рестриктазой NcoI, а концевой фрагмент - рестриктазой HindIII. После очистки расщепленные стартовый и концевой фрагменты комбинировали и лигировали в векторный NсоI, НindiII-фрагмент длиной примерно 3800 нуклеотидов в составе плазмиды pMON3934.
Промежуточную плазмиду, описанную выше, содержащую полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом, расщепляли рестриктазами NcoI и HindIII. Расщепленную ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом выделяли с использованием реактивов Geneclean (Bio101, Vista, СА). Промежуточную плазмиду pMON13180 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4023 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставки нового гена. Полученную в результате плазмиду обозначили pMON13198.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13198 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13198 включает последовательность ДНК (SEQ ID NO 35), кодирующую следующую аминокислотную последовательность:

Пример 72
Конструирование pMON13199
Ген белка с новыми N-концом/С-концом в составе плазмиды pMON13199 был создан с использованием способа II, описанного в разделе "Материалы и методы". Стартовый фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего стартовую 142 (SEQ ID NO 376) и стартовую Р-bl (SEQ ID NO 366). Концевой фрагмент был создан и амплифицирован на основе последовательности G-CSF Ser17 из состава pMON13037 с использованием набора затравок, включающего концевую 141 (SEQ ID NO 377) и концевую P-bl (SEQ ID NO 367). Стартовый фрагмент расщепляли рестриктазой NcoI, а концевой фрагмент - рестриктазой HindIII. После очистки расщепленные стартовый и концевой фрагменты комбинировали и лигировали в векторный NcoI, HindiII-фрагмент длиной примерно 3800 нуклеотидов в составе плазмиды pMON3934.
Промежуточную плазмиду, описанную выше, содержащую полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом, расщепляли рестриктазами NcoI и HindIII. Расщепленную ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и полноразмерный ген G-CSF Ser17 с новыми N-концом/С-концом выделяли с использованием реактивов Geneclean (Bio101, Vista, СА). Промежуточную плазмиду pMONl3181 расщепляли рестриктазами HindIII и AflIII, в результате чего получали векторный фрагмент длиной 4068 нуклеотидов, который очищали с использованием Magic DNA Clean-up Kit (Promega, Madison, WI). Очищенные фрагменты рестрикции комбинировали и лигировали с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК из отобранных трансформантов секвенировали с целью подтверждения правильности вставки нового гена. Полученную в результате плазмиду обозначили pMON13199.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON13199 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON13199 включает последовательность ДНК (SEQ ID NO 36), кодирующую следующую аминокислотную последовательность:

Пример 73
Конструирование тандемно дуплицированной плазмидной матрицы Syntanl
Для создания тандемно дуплицированной матрицы pMON13416 агониста рецептора hIL-3 - Syntanl - три ДНК соединяли друг с другом путем лигирования с использованием ДНК-лигазы Т 4 (Boehringer Mannheim). Эти три ДНК таковы: 1) pMON13046, включающая плазмиду агониста рецептора hIL-3 pMON13416, расщепленную рестриктазами BstEII и SnaBI; 2) прошедшая отжиг пара комплементарных олигонуклеотидов - стартовая затравка L1syn (SEQ ID NO 352) и концевая затравка L1syn (SEQ ID NO 353), которые содержат последовательность, кодирующую линкер, который обеспечивает контакт С-конца и N-конца исходного полипептида и небольшие участки окружающих последовательностей из состава pMON13416, которые при точной сборке обусловливают появление концов BstEII и Clal; и 3) часть плазмиды pMON13416 агониста рецептора hIL-3, отщепленной от pMON13046 с помощью рестриктаз Clal (ДНК растили в dam-клетках: DM1 (Life Technologies)) и SnaBI. Расщепленные ДНК растворяли в 0,9%-ном геле ТАЕ, окрашивали бромистым этидием и выделяли с помощью реактивов Geneclean (Bio101).
Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Минипрепарируемую ДНК выделяли из трансформантов, а трансформанты скринировали в тесте на основе ПЦР. Плазмидную ДНК из отобранных трансформантов секвенировали для получения правильной матрицы. Полученную в результате плазмиду обозначали syntanl: она включала последовательность ДНК по SEQ ID NO 7.
Пример 74
Конструирование тандемно дуплицированной матрицы syntan3
Для создания тандемно дуплицированной матрицы pMON13416 агониста рецептора hIL-3 - Syntan3 - три ДНК соединяли друг с другом путем лигирования с использованием ДНК-лигазы Т4 (Boehringer Mannheim). Эти три ДНК таковы: 1) pMON13046, включающая плазмиду агониста рецептора hIL-3 pMON13416, расщепленную рестриктазами BstEII и SnaBI; 2) прошедшая отжиг пара комплементарных олигонуклеотидов - стартовая затравка L3syn (SEQ ID NO 354) и концевая затравка L3syn (SEQ ID NO 355), которые содержат последовательность, кодирующую линкер, который обеспечивает контакт С-конца и N-конца исходного полипептида и небольшие участки окружающих последовательностей из состава pMON13416, которые при точной сборке обусловливают появление концов BstEII и SnaBI; и 3) часть плазмиды pMON13416 агониста рецептора hIL-3, отщепленной от pMON13046 с помощью рестриктаз Clal (ДНК растили в dam-клетках: DM1 (Life Technologies)) и SnaBI. Расщепленные ДНК растворяли в 0,9%-ном геле ТАЕ, окрашивали бромистым этидием и выделяли с помощью реактивов Geneclean (Bio101).
Часть реакционной смеси лигирования использовали для трансформации клеток E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Минипрепарируемую ДНК выделяли из трансформантов, а трансформанты скринировали в тесте на основе ПЦР. Плазмидную ДНК из отобранных трансформантов секвенировали для получения правильной матрицы. Полученную в результате плазмиду обозначали syntan3: она включала последовательность ДНК по SEQ ID NO 8.
Пример 75
Конструирование pMON31104
Ген белка с новыми N-концом/С-концом в плазмиде pMON31104 создавали с использованием способа III, описанного выше в разделе "Материалы и методы". Полноразмерный ген агониста рецептора hIL-3 с новыми N-концом/С-концом в составе pMONl3416 создавали и амплифицировали из промежуточной плазмиды Syntanl с использованием затравок прямой 35 (SEQ ID NO 356) и обратной 34 (SEQ ID NO 357).
Получаемый в результате фрагмент ДНК, содержащий новый ген, расщепляли рестриктазами NcoI и SnaBI. Расщепленный фрагмент ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и выделяли с помощью реактивов Geneclean (Bio101, Vista, CA). Очищенный расщепленный фрагмент ДНК лигировали в состав экспрессирующего вектора pMON13189 с использованием ДНК-лигазы Т 4 (Boehringer Mannheim, Indianapolis, IN). ДНК плазмиды pMON13189 предварительно расщепляли рестриктазами NcoI и SnaBI с целью удаления кодирующей последовательности агониста рецептора hIL-3 pMON13416: в результате с использованием реактивов Geneclean (Bio101, Vista, CA) выделили векторный фрагмент длиной 4254 нуклеотидов после разгонки в 0,8%-ном геле ТАЕ и окрашивания бромистым этидием. Частью реакционной смеси лигирования трансформировали клетки E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и секвенировали для подтверждения точности вставки. Полученную в результате плазмиду обозначили pMON31104.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON31104 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON31104 содержит последовательность ДНК (SEQ ID NO 9), которая кодирует следующую аминокислотную последовательность:

Пример 76
Конструирование pMON31105
Ген белка с новыми N-концом/С-концом в плазмиде pMON31105 создавали с использованием способа III, описанного выше в разделе "Материалы и методы". Полноразмерный ген агониста рецептора hIL-3 с новыми N-концом/С-концом в составе pMON13416 создавали и амплифицировали из промежуточной плазмиды Syntanl с использованием затравок прямой 70 (SEQ ID NO 358) и обратной 69 (SEQ ID NO 359).
Получаемый в результате фрагмент ДНК, содержащий новый ген, расщепляли рестриктазами NcoI и SnaBI. Расщепленный фрагмент ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и выделяли с помощью реактивов Geneclean (Bio101, Vista, CA). Очищенный расщепленный фрагмент ДНК лигировали в состав экспрессирующего вектора pMON13189 с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). ДНК плазмиды pMON13189 предварительно расщепляли рестриктазами NcoI и SnaBI с целью удаления кодирующей последовательности агониста рецептора hIL-3 pMON13416: в результате с использованием реактивов Geneclean (Bio101, Vista, CA) выделили векторный фрагмент длиной 4254 нуклеотидов после разгонки в 0,8%-ном геле ТАЕ и окрашивания бромистым этидием. Частью реакционной смеси лигирования трансформировали клетки E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и секвенировали для подтверждения точности вставки. Полученную в результате плазмиду обозначили pMON31105.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON31105 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON31105 содержит последовательность ДНК (SEQ ID NO 10), которая кодирует следующую аминокислотную последовательность:

Пример 77
Конструирование pMON31106
Ген белка с новыми N-концом/С-концом в плазмиде pMON31106 создавали с использованием способа III, описанного выше в разделе "Материалы и методы". Полноразмерный ген агониста рецептора hIL-3 с новыми N-концом/С-концом в составе pMON13416 создавали и амплифицировали из промежуточной плазмиды Syntanl с использованием затравок прямой 91 (SEQ ID NO 360) и обратной 90 (SEQ ID NO 361).
Получаемый в результате фрагмент ДНК, содержащий новый ген, расщепляли рестриктазами NcoI и SnaBI. Расщепленный фрагмент ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и выделяли с помощью реактивов Geneclean (Bio101, Vista, CA). Очищенный расщепленный фрагмент ДНК лигировали в состав экспрессирующего вектора pMON13189 с использованием ДНК-лигазы Т 4 (Boehringer Mannheim, Indianapolis, IN). ДНК плазмиды pMONl3189 предварительно расщепляли рестриктазами NcoI и SnaBI с целью удаления кодирующей последовательности агониста рецептора hIL-3 pMON13416: в результате с использованием реактивов Geneclean (Bio101, Vista, CA) выделили векторный фрагмент длиной 4254 нуклеотидов после разгонки в 0,8%-ном геле ТАЕ и окрашивания бромистым этидием. Частью реакционной смеси лигирования трансформировали клетки E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и секвенировали для подтверждения точности вставки. Полученную в результате плазмиду обозначили pMON31106.
Клетки Е.coli штамма JM101 трансформировали плазмидой pMON31106 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON31106 содержит последовательность ДНК (SEQ ID NO 11), которая кодирует следующую аминокислотную последовательность:

Пример 78
Конструирование pMON31107
Ген белка с новыми N-концом/С-концом в плазмиде pMON31107 создавали с использованием способа III, описанного выше в разделе "Материалы и методы". Полноразмерный ген агониста рецептора hIL-3 с новыми N-концом/С-концом в составе pMON13416 создавали и амплифицировали из промежуточной плазмиды Syntanl с использованием затравок прямой 101 (SEQ ID NO 362) и обратной 100 (SEQ ID NO 363).
Получаемый в результате фрагмент ДНК, содержащий новый ген, расщепляли рестриктазами NcoI и SnaBI. Расщепленный фрагмент ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и выделяли с помощью реактивов Geneclean (Bio101, Vista, CA). Очищенный расщепленный фрагмент ДНК лигировали в состав экспрессирующего вектора pMON13189 с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). ДНК плазмиды pMON13189 предварительно расщепляли рестриктазами NcoI и SnaBI с целью удаления кодирующей последовательности агониста рецептора hIL-3 pMON-13416: в результате с использованием реактивов Geneclean (Bio101, Vista, CA) выделили векторный фрагмент длиной 4254 нуклеотидов после разгонки в 0,8%-ном геле ТАЕ и окрашивания бромистым этидием. Частью реакционной смеси лигирования трансформировали клетки E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и секвенировали для подтверждения точности вставки. Полученную в результате плазмиду обозначили pMON31107.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON31107 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON31107 содержит последовательность ДНК (SEQ ID NO 12), которая кодирует следующую аминокислотную последовательность:

Пример 79
Конструирование pMON31108
Ген белка с новыми N-концом/С-концом в плазмиде pMON31108 создавали с использованием способа III, описанного выше в разделе "Материалы и методы". Полноразмерный ген агониста рецептора hIL-3 с новыми N-концом/С-концом в составе pMON13416 создавали и амплифицировали из промежуточной плазмиды Syntan3 с использованием затравок прямой 35 (SEQ ID NO 356) и обратной 34 (SEQ ID NO 357).
Получаемый в результате фрагмент ДНК, содержащий новый ген, расщепляли рестриктазами NcoI и SnaBI. Расщепленный фрагмент ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и выделяли с помощью реактивов Geneclean (Bio101, Vista, CA). Очищенный расщепленный фрагмент ДНК лигировали в состав экспрессирующего вектора pMON13189 с использованием ДНК-лигазы Т 4 (Boehringer Mannheim, Indianapolis, IN). ДНК плазмиды pMON13189 предварительно расщепляли рестриктазами NcoI и SnaBI с целью удаления кодирующей последовательности агониста рецептора hIL-3 pMON13416: в результате с использованием реактивов Geneclean (Bio101, Vista, CA) выделили векторный фрагмент длиной 4254 нуклеотидов после разгонки в 0,8%-ном геле ТАЕ и окрашивания бромистым этидием. Частью реакционной смеси лигирования трансформировали клетки E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и секвенировали для подтверждения точности вставки. Полученную в результате плазмиду обозначили pMON31108.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON31108 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON31108 содержит последовательность ДНК (SEQ ID NO 13), которая кодирует следующую аминокислотную последовательность:

Пример 80
Конструирование pMON31109
Ген белка с новыми N-концом/С-концом в плазмиде pMON31109 создавали с использованием способа III, описанного выше в разделе "Материалы и методы". Полноразмерный ген агониста рецептора hIL-3 с новыми N-концом/С-концом в составе pMON13416 создавали и амплифицировали из промежуточной плазмиды Syntan3 с использованием затравок прямой 70 (SEQ ID NO 358) и обратной 69 (SEQ ID NO 359).
Получаемый в результате фрагмент ДНК, содержащий новый ген, расщепляли рестриктазами NcoI и SnaBI. Расщепленный фрагмент ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и выделяли с помощью реактивов Geneclean (Bio101, Vista, CA). Очищенный расщепленный фрагмент ДНК лигировали в состав экспрессирующего вектора pMON13189 с использованием ДНК-лигазы Т4 (Boehringer Mannрeim, Indianapolis, IN). ДНК плазмиды pMON13189 предварительно расщепляли рестриктазами NcoI и SnaBI с целью удаления кодирующей последовательности агониста рецептора hIL-3 pMON13416: в результате с использованием реактивов Geneclean (Bio101, Vista, CA) выделили векторный фрагмент длиной 4254 нуклеотидов после разгонки в 0,8%-ном геле ТАЕ и окрашивания бромистым этидием. Частью реакционной смеси лигирования трансформировали клетки E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и секвенировали для подтверждения точности вставки. Полученную в результате плазмиду обозначили pMON31109.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON31109 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON31109 содержит последовательность ДНК (SEQ ID NO 14), которая кодирует следующую аминокислотную последовательность:

Пример 81
Конструирование pMON31110
Ген белка с новыми N-концом/С-концом в плазмиде pMON31110 создавали с использованием способа III, описанного выше в разделе "Материалы и методы". Полноразмерный ген агониста рецептора hIL-3 с новыми N-концом/С-концом в составе pMON13416 создавали и амплифицировали из промежуточной плазмиды Syntan3 с использованием затравок прямой 91 (SEQ ID NO 360) и обратной 90 (SEQ ID NO 361).
Получаемый в результате фрагмент ДНК, содержащий новый ген, расщепляли рестриктазами NcoI и SnaBI. Расщепленный фрагмент ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и выделяли с помощью реактивов Geneclean (Bio101, Vista, CA). Очищенный расщепленный фрагмент ДНК лигировали в состав экспрессирующего вектора pMON13189 с использованием ДНК-лигазы Т4 (Boehringer Mannheim, Indianapolis, IN). ДНК плазмиды pMON13189 предварительно расщепляли рестриктазами NcoI и SnaBI с целью удаления кодирующей последовательности агониста рецептора hIL-3 pMON13416: в результате с использованием реактивов Geneclean (Bio101, Vista, CA) выделили векторный фрагмент длиной 4254 нуклеотидов после разгонки в 0,8%-ном геле ТАЕ и окрашивания бромистым этидием. Частью реакционной смеси лигирования трансформировали клетки E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и секвенировали для подтверждения точности вставки. Полученную в результате плазмиду обозначили pMON31110.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON31110 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON31110 содержит последовательность ДНК (SEQ ID NO 15), которая кодирует следующую аминокислотную последовательность:

Пример 82
Конструирование pMON31111
Ген белка с новыми N-концом/С-концом в плазмиде pMON31111 создавали с использованием способа III, описанного выше в разделе "Материалы и методы". Полноразмерный ген агониста рецептора hIL-3 с новыми N-концом/С-концом в составе pMON13416 создавали и амплифицировали из промежуточной плазмиды SyntanS с использованием затравок прямой 101 (SEQ ID NO 362) и обратной 100 (SEQ ID NO 363).
Получаемый в результате фрагмент ДНК, содержащий новый ген, расщепляли рестриктазами NcoI и SnaBI. Расщепленный фрагмент ДНК растворяли в 1%-ном геле ТАЕ, окрашивали бромистым этидием и выделяли с помощью реактивов Geneclean (Bio101, Vista, CA). Очищенный расщепленный фрагмент ДНК лигировали в состав экспрессирующего вектора pMONl3189 с использованием ДНК-лигазы Т 4 (Boehringer Mannheim, Indianapolis, IN). ДНК плазмиды pMONl3189 предварительно расщепляли рестриктазами NcoI и SnaBI с целью удаления кодирующей последовательности агониста рецептора hIL-3 pMON13416: в результате с использованием реактивов Geneclean (Bio101, Vista, CA) выделили векторный фрагмент длиной 4254 нуклеотидов после разгонки в 0,8%-ном геле ТАЕ и окрашивания бромистым этидием. Частью реакционной смеси лигирования трансформировали клетки E.coli штамма DH5α (Life Technologies, Gaithersburg, MD). Трансформированных бактерий отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и секвенировали для подтверждения точности вставки. Полученную в результате плазмиду обозначили pMON31111.
Клетки E.coli штамма JM101 трансформировали плазмидой pMON31111 с целью экспрессии белка и его выделения из включенных телец.
Плазмида pMON31111 содержит последовательность ДНК (SEQ ID NO 16), которая кодирует следующую аминокислотную последовательность:

Пример 83
Конструирование pMON31112
Осуществлено конструирование pMON31112 - плазмиды, содержащей последовательность ДНК, кодирующую многофункциональный агонист гемопоэтических рецепторов, который активирует рецептор hIL-3 и рецептор G-CSF. ДНК плазмиды pMON13189 расщепляли рестриктазами NcoI и XmaI, в результате чего был выделен и очищен в 0,8%-ном агарозном геле векторный NcoI, -XmaI-фрагмент. ДНК второй плазмиды - pMON13222 (WO 94/12639, патент США 08/411796) - расщепляли рестриктазами NcoI и EcoRI, в результате чего образуется NIl,EcoRI-фрагмент длиной 281 нуклеотид. Этот фрагмент выделяли и очищали в 1,0%-ном агарозном геле. Два олигонуклеотида - SYNNOXA1.REQ (SEQ ID NO 350) и SYNNOXA2.REQ (SEQ ID NO 351) - отжигали друг на друга и лигировали с 281-нуклеотидным фрагментом ДНК из плазмиды pMON13222 на векторный фрагмент из плазмиды pMON13189. Часть реакционной смеси лигирования затем использовали для трансформации клеток E.coli K-12 штамма JM101. Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и подвергали рестрикционному анализу с целью установления присутствия EcoRI-фрагмента и секвенировали для подтверждения точности вставки.
Плазмида pMON31112 содержит последовательность ДНК (SEQ ID NO 37), кодирующую следующую аминокислотную последовательность:

Пример 84
Конструирование pMON31113
Осуществлено конструирование pMON31113 - плазмиды, содержащей последовательность ДНК, кодирующую многофункциональный агонист гемопоэтических рецепторов, который активирует рецептор hIL-3 и рецептор С-СSF.ДНК плазмиды pMON13197 расщепляли рестриктазами NcoI и XmaI, в результате чего был выделен и очищен в 0,8%-ном агарозном геле векторный NcoI, -XmaI-фрагмент. ДНК второй плазмиды - pMON13239 (WO 94/12639, патент США 08/411796) - расщепляли рестриктазами NcoI и EcoRI, в результате чего образуется NcoI, EcoRI-фрагмент длиной 281 нуклеотид. Этот фрагмент выделяли и очищали в 1,0%-ном агарозном геле. Два олигонуклеотида - SYNNOXA1.REQ (SEQ ID NO 350) и SYNNOXA2.REQ (SEQ ID NO 351) - отжигали друг на друга и лигировали с 281-нуклеотидным фрагментом ДНК из плазмиды pMON13239 на векторный фрагмент из плазмиды pMON13197. Часть реакционной смеси лигирования затем использовали для трансформации клеток E.coli K-12 штамма JM101. Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и подвергали рестрикционному анализу с целью установления присутствия EcoRI-фрагмента и секвенировали для подтверждения точности вставки.
Плазмида pMON31113 содержит последовательность ДНК (SEQ ID NO 38), кодирующую следующую аминокислотную последовательность:

Пример 85
Конструирование pMON3114
Осуществлено конструирование pMON31114 - плазмиды, содержащей последовательность ДНК, кодирующую многофункциональный агонист гемопоэтических рецепторов, который активирует рецептор hIL-3 и рецептор G-CSF,ДНК плазмиды pMON13189 расщепляли рестриктазами NcoI и XmaI, в результате чего был выделен и очищен в 0,8%-ном агарозном геле векторный NcoI, -XmaI-фрагмент. ДНК второй плазмиды - pMON13239 (WO 94/12639, патент США 08/411796) - расщепляли рестриктазами NcoI и EcoRI, в результате чего образуется NcoI,EcoRI-фрагмент длиной 281 нуклеотид. Этот фрагмент выделяли и очищали в 1,0%-ном агарозном геле. Два олигонуклеотида - SYNNOXA1.REQ (SEQ NO 350) и SYNNOXA2.REQ (SEQ ID NO 351) - отжигали друг на друга и лигировали с 281-нуклеотидным фрагментом ДНК из плазмиды pMON13239 на векторный фрагмент из плазмиды pMON13189. Часть реакционной смеси лигирования затем использовали для трансформации клеток E.coli K-12 штамма JM101. Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли и подвергали рестрикционному анализу с целью установления присутствия EcoRI-фрагмента и секвенировали для подтверждения точности вставки.
Плазмида pMON31114 содержит последовательность ДНК (SEQ ID NO 39), кодирующую следующую аминокислотную последовательность:

Пример 86
Конструирование pMON31115
Осуществлено конструирование pMON31115 - плазмиды, содержащей последовательность ДНК, кодирующую многофункциональный агонист гемопоэтических рецепторов, который активирует рецептор hIL-3 и рецептор G-CSF.ДНК плазмиды pMON13197 расщепляли рестриктазами NcoI и XmaI, в результате чего был выделен и очищен в 0,8%-ном агарозном геле векторный NcoI, -XmaI-фрагмент. ДНК второй плазмиды - pMON13222 - расщепляли рестриктазами NcoI и EcoRI, в результате чего образуется NcoI, EcoRI-фрагмент длиной 281 нуклеотид. Этот фрагмент выделяли и очищали в 1,0%-ном агарозном геле. Два олигонуклеотида - SYNNOXA1.REQ (SEQ ID NO 350) и SYNNOXA2.REQ (SEQ ID NO 351) - отжигали друг на друга и лигировали с 281-нуклеотидным фрагментом ДНК из плазмиды pMONl3222 на векторный фрагмент из плазмиды pMON13197. Часть реакционной смеси лигирования затем использовали для трансформации клеток E.coli K-12 штамма JM101. Трансформированные бактерии отбирали на чашках, содержащих ампициллин. Плазмидную ДНК выделяли подвергали рестрикционному анализу с целью установления присутствия EcoRI-фрагмента и секвенировали для подтверждения точности вставки.
Плазмида pMON31115 содержит последовательность ДНК (SEQ ID NO 40), кодирующую следующую аминокислотную последовательность:

Пример 87
Определение in vitro активности белков многофункциональных агонистов гемопоэтических рецепторов
Концентрация белка многофункционального агониста гемопоэтических рецепторов может быть определена с использованием метода ТИФА-сэндвича по аффинности очищенного поликлонального антитела. С другой стороны, концентрация белка может быть определена с помощью анализа аминокислотного состава. Биологическая активность многофункционального агониста гемопоэтических рецепторов может быть определена в ряде тестов in vitro. Например, многофункциональный агонист гемопоэтических рецепторов, который связывается с рецептором hIL-3 и рецептором G-CSF, может быть протестирован в тестах на пролиферацию клеток с использованием клеточных линий, экспрессирующих рецепторы hIL-3 и (или) G-CSF. Один из таких тестов - тест на пролиферацию клеток AML-193. Клетки AML-193 реагируют на IL-3 и G-CSF, что позволяет определять биологическую активность многофункционального агониста гемопоэтических рецепторов IL-3/G-CSF. Другим таким тестом является тест на пролиферацию клеток TF1.
Дополнительно другие фактор-зависимые клеточные линии, такие как M-NFS-60 (ATCC CRL 1838) или 32D, являющиеся линиями зависимых от IL-3 клетками мыши, могут быть использованы. Активность IL-3 является видоспецифичной, в то время как активность G-CSF таковой не является: соответственно, биологическая активность компонента G-CSF в составе многофункционального агониста гемопоэтических рецепторов IL-3/G-CSF может быть определена независимо. Клеточные линии, такие как ВНК или Baf/3 мыши, которые не экспрессируют рецепторы данных лигандов, могут быть трансфицированы плазмидой, включающей ген желательного рецептора. Примером такой линии является клеточная линия BaF3, трансфицированная рецептором hG-CSF (BaF3/hG-CSF). Активность многофункционального агониста гемопоэтических рецепторов в этих клеточных линиях может быть использована для сравнения с hIL-3 или G-CSF отдельно или вместе. Биологическая активность примеров многофункциональных агонистов гемопоэтических рецепторов по настоящему изобретению, оцениваемая по пролиферации клеток BaF3/hG-CSF и пролиферации клеток TF1, показана в табл. 5 и табл. 6. Биологическая активность многофункционального агониста гемопоэтических рецепторов выражена как относительная активность по отношению со стандартным белком pMON13056 (WO 95/21254), который проявляет связывающую активность рецепторов IL-3 и G-CSF. Биологическая активность примеров многофункциональных агонистов гемопоэтических рецепторов по настоящему изобретению, оцениваемая по пролиферации клеток BaF3/c-mpl и пролиферации клеток TF1, показана в табл.7 и табл.8.
Сходным способом другие фактор-зависимые клеточные линии, известные специалистами в данной области техники, могут быть использованы для измерения биологической активности желательного многофункционального агониста гемопоэтических рецепторов. Метилцеллюлозный тест может быть использован для определения влияния многофункциональных агонистов гемопоэтических рецепторов на экспансию клеток-предшественников кроветворения и параметры различных типов гемопоэтических колоний in vitro. Метилцеллюлозный тест может позволить оценить число предшественников по тому, что можно измерить число долю предшественников в расчете на 100 тысяч тестируемых клеток. Долговременные, зависимые от стромы культуры используют для разграничения примитивных клеток-предшественников кроветворения и стволовых клеток. Этот тест может быть использован для определения того, стимулирует ли многофункциональный агонист гемопоэтических рецепторов экспансию очень примитивных клеток-предшественников и (или) стволовых клеток. В дополнение могут быть поставлены культуры с ограниенным разбавлением, которые позволят определить число примитивных клеток-предшественников, стимулированных многофункциональным агонистом гемопоэтических рецепторов.
Активность дуалистичных агонистов рецепторов IL-3/G-CSF по индукции пролиферации клеток
* - биологическая активность многофункционального агониста гемопоэтических рецепторов выражена как относительная активность по отношению к стандартному белку pMON13056: n=3 или больше
Активность дуалистичных агонистов рецепторов IL-3/G-CSF по индукции пролиферации клеток
* - биологическая активность (n=1 или 2) многофункционального агониста гемопоэтических рецепторов выражена как относительная активность по отношению к стандартному белку pMON 13056
"+" обозначает, что молекула сравнима с pMON13056
Активность пролиферации клеток
** - активность измеряли по отношению к pMON13056
Пример 88
Варианты G-CSF, которые содержат одиночные или множественные аминокислотные замены, были сформированы с использованием методик ПЦР-мутагенеза, как это было описано в патентах WO 94/12639 и WO 94/12638. Эти и другие варианты (т.е. обусловливаемые аминокислотными заменами, вставками или делециями и удлинениями N- или С-конца) также могут быть сформированы, как это известно в данной области техники, с применением ряда других методов, включая сборку синтетических генов или направленный мутагенез (см. Taylor et al., 1985, Nucl. Acids Res., 13, 7684-8785; Kunkel et al., 1985, Proc. Natl. Acad. Sci. USA, 82, 488-492; Sambrook et al., 1989, "Molecular Cloning: A Laboratory Manual, 2d ed., Cold Spring Harbor Lab. Press, Cold Spring Harbor, NY; патенты WO 94/12639 и WO 94/12638). Эти замены могут осуществляться поодиночке или же в комбинации с другими аминокислотными заменами, и (или) с делециями, и (или) со вставками, и (или) с удлинениями концов. После верификации изменения последовательности плазмидная ДНК может быть трансфицирована в подходящие клетки млекопитающих, клетки насекомых или бактериальные штаммы, такие как продукционные штаммы E.coli Известные варианты G-CSF, которые являются активными, включают замены в положениях 1 (треонин на серин, аргинин или глицин), 2 (пролин на лейцин), 3 (лейцин на аргинин или серин) и 17 (цистеин на серин) и делеции аминокислот 1-11 (Kuga et al., 1989, Biochem. Biophys. Res. Commun., 159, 103-111). Эти варианты G-CSF с аминокислотными заменами могут быть использованы в качестве матриц для создания агонистов рецепторов G-CSF, в которых создаются новый N-конец и новый С-конец. Примеры вариантов G-CSF с аминокислотными заменами показаны в табл. 9.
н/о - не определяли
Пример 89
Определение биологической активности вариантов G-CSF с аминокислотными заменами
Варианты G-CSF, имеющие аминокислотные замены, могут быть протестированы на активность по индукции клеточной пролиферации с использованием линии клеток Baf/3, трансфицированной рецептором G-CSF человека. Биологическая активность примеров вариантов G-CSF с аминокислотными заменами показана в табл. 9 в сравнении с активностью нативного G-CSF человека. Знак "+" обозначает активность, сопоставимую с активностью нативного фактора, а знак "-" обозначает существенно сниженную или не поддающуюся измерению активность.
Пример 90
Выделение кДНК, кодирующей лиганд flt3
Три клона лиганда flts были амплифицированы из полиаденилированных РНК клеток костного мозга человека с использованием ПЦР-затравок NCOFLT, HIND160 и HIND165 (в соответствии с условиями, представляемыми производителями), Эти амплифицированные продукты ПЦР очищали в геле и клонировали в ВНК-экспрессирующий вектор pMON5723, создавая тем самым плазмиды pMON30237 (NCOFLT + HIND160), pMON30238 (NCOFLT + HIND165) и делеционный клон pMON30239 (NCOFLT + HIND165). Делеция в составе pMON 30239 соответствует утрате аминокислот 89-106.
Пример 91
Агонисты рецептора flt3 с измененными последовательностями конструировали с использованием ряда методик и линкерных типов. Первый набор конструкций, включающий линкерные пептиды (SerGlyGlyAsnGly)х (где x=1, 2 или 3), с точками разрывов 39/40, 65/66 и 89/90 были созданы с использованием двухэтапной ПЦР, описаной Mullins et al.: в них переднюю половину и заднюю половину каждой итоговой последовательности перестроенной молекулы создавали раздельно на первом этапе ПЦР, а затем попарные продукты первой реакции комбинировали на втором этапе ПЦР и удлиняли в отсутствие экзогенных затравок. Например, шесть исходных ПЦР-продуктов были генерированы с целью создания трех молекул-предшественников с точкой разрыва 89/90 - использовали аминокислотные линкеры SerGlyGlyAsnGly (SEQ ID NO 786), SerGlyGlyAsnGlySerGlyGlyAsnGly (SEQ ID NO 787) и SerGlyGlyAsnGlySerGlyGlyAsnGlySerGlyGlyAsnGly (SEQ ID NO 788) (соответственно, pMON32326, pMON32327 и pMON32328). Следующие пары затравок были использованы на первом этапе реакции ПЦР: a) 89For/L5B; b) 89For/L10B; с) 89For/L15B; d) 89Rev/L5A; e) 89Rev/L10A; f) 89Rev/L15A. Идентичный подход был использован для формирования предшественников pMON32321 (точка разрыва 39/40, пары затравок 39For/L108 и 39Rev/L10A) и pMON32325 (точка разрыва 65/66, пары затравок 65For/LSB и 65Rev/LSA). С учетом исключений, отмеченных выше, во всех последующих реакциях ПЦР использовали компоненты PCR Optimizer Kit (Invitrogen), а условия амплификации принимали с учетом рекомендуемых производителем процедур. Реакции были выстроены следующим образом: комбинировали 50 пкМ каждой затравки, 10 мкл 5х буфера В [300 мМ Трис-НСl (рН=8,5), 10 мМ MgCl2, 75 мМ (NH4)2SO4], 5 ед. Taq-полимеразы и 100 нг нагретой денатурированной ДНК-матрицы (в данном образце pMON30238) и затем доводили до конечного объема в 45 мкл добавлением дистиллированной воды. Реакционные смеси преинкубировали при 80°С в течение 1-5 минут, затем добавляли по 5 мкл 10 мМ дНТФ в каждую реакционную смесь и проводили денатурацию нагреванием до 94°С в течение 2 минут перед последующей амплификацией в приборе Model 480 (Perkin-Elmer). Семь циклов амплификации ДНК осуществляли при следующих условиях: денатурация нагреванием до 94°С в течение 1 минуты, отжиг при 65°С в течение 2 минут, достройка при 72°С в течение 3 минут. Двадцать три дополнительных цикла проводили по схеме - денатурация нагреванием до 94°С в течение 1 минуты, отжиг/достройка при 72°С в течение 4 минут с последующим заключительным удлиняющим циклом при 72°С в течение 7 минут. За исключением плазмиды pMON32328, продукты ПЦР-амплификации разгоняли в 1,2%-ном агарозном геле ТАЕ, а подходящие по размеру зоны амплификации (т.е. основной амплификационный продукт) вырезали и очищали с использованием реактивов Geneclean II (Bio101). Образцы ресуспензировали в 10 мкл дистиллированной воды. Продукты амплификации pMON32328 очищали напрямую с помощью реактивов Wizard PCR Clean-up Kit (Promega), а ДНК элюировали в 50 мкл дистиллированной воды.
Способ конструирования предшественников pMON32322 (точка разрыва 39/40, пары затравок 39For/L5B и 39Rev/L5A) модифицировали путем увеличения количества матрицы до 1 мкг и изменения условий ПЦР-амплификации по следующей схеме: шесть циклов при 94°С в течение 1 минуты, при 65°С в течение 2 минут и при 72°С в течение 2,5 минут с последующими 15 циклами при 94°С в течение 1 минуты, при 70°С в течение 2 минут и при 72°С в течение 2 минут и единственным заключительным циклом достройки при 72°С в течение 7 минут.
На втором этапе ПЦР использовали очищенные в геле предшественники, полученные на первом этапе ПЦР в виде комбинации "затравка/матрица" в следующем сочетании: 5 мкл каждой молекулы-предшественника (т.е. ПЦР-продукты по затравкам 89For/L5B и 89Rev/L5A для pMON32328), 10 мкл 5х буфера В, 5 ед. Taq-полимеразы и 24 мкл дистиллированной воды. Реакционные смеси нагревали до 80°С в течение 5 минут, к ним добавляли 5 мкл 10 мМ дНТФ и реакционные смеси денатурировали нагреванием до 94°С в течение 2 минут.
Условия ДНК-амплификации были следующими: 15 циклов при 94°С в течение 1 минуты, при 69°С в течение 2 минут с последующим достроечным циклом при 72°С в течение 3 минут. С целью достижения полной достройки последний цикл завершают единственным этапом при 72°С в течение 7 минут. Время инкубации при 80°С сокращали до 2 минут и число циклов снижали до 10 для плазмиды pMON32325 (ПЦР-продукты пар затравок 65For/L5B и 65Rev/L5A). Продукты реакции ПЦР, имеющие подходящий размер, очищали в 1,2%-ном агарозном геле ТАЕ с использованием реактивов Geneclean II. В случае с плазмидой pMON32322 (пары затравок 39For/LSB и 39Rev/L5A) температуру отжига снижали до 68°С, а время достройки сокращали до 2 минут. В дополнение продукт ПЦР очищали с использованием реактивов Wizard OCR Clean-up Kit (Promega) в соответствии с предлагаемыми производителем рекомендациями. Второй этап ПЦР в случае с плазмидой pMON32326 (ПЦР-продукты пар затравок 89For/L15B и 89Rev/L15A) модифицировали следующим образом. Три набора реакционных смесей для ПЦР выстраивали также, как это было описано выше, за исключением выбора типа буфера (5х буфер либо В, либо D, либо J: реактивы PCR Optimizer Kit). Состав буферов D и J отличается от буфера В только по величине рН или содержанию MgCl2. Концентрация хлорида магния в буфере D составляет 3,5 мМ, а в буфере J - 9,5 мМ. Процедуру модифицировали путем увеличения числа циклов ПЦР при том, что 20- и 15-мкл аликвоты отбирали в конце 10-го, 15-го и 2-го циклов. По 5 мкл каждого аликвота использовали для анализа присутствия амплифицированного материала в 1,2%-ном агарозном геле ТАЕ. Остатки реакционных смесей ПЦР в буферах В, D и J сохраняли и последовательно очищали с использованием реактивов Wizard ОРСК Clean-up Kit. ДНК элюировали в 50 мкл дистиллированной воды.
Очищенные образцы из реакционной смеси по второму этапу ПЦР расщепляли рестриктазами NcoI/HindIII с использованием одного из двух стандартизованных вариантов расщепления. Для образцов, очищенных в Geneclean II, 10 мкл ДНК расщепляли в 20 мкл реакционной смеси при 7,5 ед. каждой из рестриктаз NcoI/HindIII в течение 2 часов при 37°С, а гель очищали в 1,1%-ном агарозном геле ТАЕ снова в Geneclean II. Готовые к лигированию образцы ресуспензировали в 10 мкл дистиллированной воды. В случае плазмиды pMON32322 20 мкл образца расщепляли в 50 мкл реакционной смеси с 20 ед. каждой из рестриктаз NcoI и HindIII в течение 3 часов при 37°С. После этого добавляли 0,1 объема 3 М NaOAc (рН=5,5) и 2,5 объема этанола, размешивали и сохраняли в течение ночи при - 20°С. ДНК восстанавливали путем пеллетирования в течение 20 минут на микроцентрифуге Sigma Mk-202 при 13000 об/мин при 4°С. Гранулы ДНК промывали в охлажденном 70%-ном этиловом спирте, лиофилизировали и ресуспензировали в 10 мкл дистиллированной воды.
Пример 92
Альтернативный подход был использован для конструирования плазмид pMON32320 (точка разрыва 39/40, линкер из 15 аминокислот), pMON32323 (точка разрыва 65/66, линкер из 15 аминокислот) и pMON32324 (точка разрыва 65/66, линкер из 10 аминокислот). Новые затравки (L15, L15D, L15E) были сформированы таким образом, чтобы включить сайт рестрикции BamHI в затравку, соответствующую участку в пределах рамки кодирования, что позволит клонировать в BamHI-сайт и поддерживать правильную кодирующую рамку. Условия первого этапа ПЦР были выбраны идентичными таковым, которые были описаны для pMON32322, за исключением того, что были использованы следующие пары затравок: 65For/Ll5D и 65Rev/L15E (pMON32324), 39For/L15D и 39Rev/L15C (pMON32320) и 65For/Ll5D и 65Rev/L15C (pMON32323). Продукты ПЦР очищали с использованием реактивов Wizard PCR Clean-up Kit, как это было описано выше, и элюировали в 50 мкл дистиллированной воды. Образцы расщепляли рестриктазами NcoI/BamHI (39For/L15D и 65For/L15D) или BamHI/HindIII (39Rev/L15C, 65Rev/L15C и 65Rev/L15E). Рестриктазное расщепление было осуществлено следующим образом: 10 мкл очищенного продукта ПЦР, 3 мкл 10х универсального рестрикционного буфера, 15 ед. либо NcoI, либо HindIII, 15 ед. BamHI при конечном объеме реакционной смеси 30 мкл. Реакционные смеси инкубировали при 37°С в течение 90 минут, а продукты ПЦР очищали в 1,1%-ном агарозном геле ТАЕ с использованием Geneclean II. Готовую к лигированию ДНК ресуспензировали в 10 мкл дистиллированной воды.
Вставки лигировали в обработанные NcoI/HindIII плазмиды pMON3977 (экспрессирующий вектор для клеток ВНК млекопитающих), которые обрабатывали щелочной фосфатазой креветки (SAP) при трехэтапной (pMON32320, pMON32323 и pMON32324) или двухэтапной (pMON32321, pMON32322, pMON32325, pMON32326, pMON32327 и pMON32328) реакции лигирования следующим образом: 2,5 мкл вставки (2 мкл каждого затравочного амплификона для pMON32320, pMON32323 и pMON32324) добавляли к 50 нг вектора в 10 мкл реакционной смеси при стандартных условиях лигирования. По 2 мл каждой реакционной смеси использовали для трансформации 100 мкл химически компетентных клеток DH5α (Gibco/BRL) в соответствии с рекомендациями производителя. Аликвоты по 25 мкл и 200 мкл высевали на чашках LB, содержащих 50 мкг/мл ампициллина и инкубировали в течение ночи. Выделенные колонии удаляли и ДНК препарировали из 50 мл "переночевавших" культур с использованием наборов Qiagen DNA midiprep. Количесвтенный анализ ДНК проводили по параметрам поглощения при А260/А280 и проверяли точность размера вставки ДНК с помощью электрофореза в агарозном геле с последующим расщеплением 1 мкг матрицы рестриктазами NcoI/HindIII. Образцы, включающие вставки предсказываемого размера, секвенируют в обоих направлениях с использованием вектор-специфичных затравок на автоматическом флуоресцентном секвенсоре ДНК модели 373А (Perkin-Elmer ABI). Реакции секвенирования проводили при 20 мкл объема реакционной смеси с использованием термического циклера модели 480 (Perkin-Elmer) следующим образом: 1 мкг матрицы, 3,2 пкМ затравки, 1 мкл ДМСО, 9,5 мкл Taq-терминаторной дидезокси-смеси (Perkin-Elmer ABI) смешивали и подвергали 25 циклам секвенирующей амплификации по следующей схеме: 30 секунд при 94°С, 15 секунд отжига при 50°С и 4 минуты достроечного цикла при 60°С. Образцы очищали на колонках Centri-Sep spin (Princeton Separations) в соответствии с рекомендуемыми процедурами, затем лиофилизировали и подвергали секвенированию. Образцы, включающие предсказываемые аминокислотные последовательности, отбирали для анализа и обозначения под номерами серии pMON.
Пример 93
Сходный подход в конструировании pMON32320, pMON32323 и pMON32324 использовали для встраивания линкера другого типа (SerGlyGlySerGly)x, где x=2 или 3, в состав двух последовательностей с перестроенной структурой, включающих точку разрыва 39/40 (pMON32348 и pMON32350). Пары затравок были следующими: сочетание 339For2/339Rev3 и 339Rev2/339-10For3 для pMON32348 и 339For2/339Rev3 и 339Rev2/339-15For3 для pMON32350 были использованы для создания трех продуктов ПЦР-амплификации. Каждую реакцию ПЦР-амплификации выстраивали следующим образом: 50 пкМ каждой пары затравок, 10 мкл 5х буфера В, 5 ед. Taq-полимеразы и дистиллированную воду добавляли к 100 нг денатурированной нагреванием pMON32320 до конечного объема 45 мкл. Реакционные смеси проинкубировали, как это было описано выше. Пятнадцать циклов амплификации осуществляли следующим образом: денатурация нагреванием при 94°С в течение 1 минуты, отжиг при 70°С в течение 2 минут и достройка при 72°С в течение 3 минут. По завершении последнего цикла проводили единственный цикл достройки. при 72°С в течение 7 минут. Продукты ПЦР-амплификации по парам затравок 339For/339Rev3, 339Rev2/339-10For3 и 339Rev2/339-15For2 очищали с использованием реактивов Wizard PCR Clean-up Kit (Promega) и элюировали в 50 мкл дистиллированной воды. Расщепление рестриктазами NcoI/BamHI в паре затравок 339For2/339Rev3 проводили следующим образом: 8 мкл ДНК-матрицы смешивали с 2 мл универсального рестрикционного буфера и по 10 ед. NcoI и BamHI в 20 мкл реакционной смеси и инкубировали при 37°С в течение 90 минут. Продукты расщепления очищали с использованием Geneclean II согласно фирменной процедуре, а готовую к лигированию ДНК ресуспензировали в 10 мкл дистиллированной воды. Рестрикционное расщепление и последующую очистку продуктов амплификации по парам затравок 339Rev2/339-10For2 и 339Rev2/339-15For2 осуществляли точно так же, как это было описано для амплификона 339For2/339Rev3, за исключением того, что использовали 10 ед. HindIII вместо NcoI. Стандартный способ лигирования был применен путем добавления к 50 нг обработанной NcoI/HindIII/SAP, очищенной в геле pMON3977 0,5 мкл амплификона 339For2/Rev3, 1 мкл либо амплификона 339Rev2/339-10For3 (pMON32348), либо амплификона 339Rev2/339-15For3 (pMON32350), 5 ед. ДНК-лигазы Т4 и 1 мкл 10х лигазного буфера в 10-мкл реакционном объеме на 60 минут при обычной температуре. Последующие шаги, необходимые для подтверждения полученной в итоге последовательности ДНК, выполняли так же, как это было описано выше.
Пример 94
Линкер третьего типа с варьирующимся повторяющимся мотивом (GlyGlyGlySer)x инкорпорировали в другой набор агонистов рецептора flt3 с перестроенной последовательностью, происходящий от матриц, выстроенных по модульному типу. Длина этих линкеров следующая: линкер из 6 аминокислот (GlyGlyGlySerGlyGly, SEQ ID NO 792), линкер из 7 аминокислот (GlyGlyGlySerGlyGlyGly, SEQ ID NO 793), линкер из 10 аминокислот (GlyGlyGlySerGlyGlyGlySerGlyGly, SEQ ID NO 794), линкер из 13 аминокислот (GlyGlyGlySerGlyGlyGlySerGlyGlyGlySerGly, SEQ ID NO 795), линкер из 15 аминокислот (GlyGlyGlySer GlyGlyGlySerGlyGlyGlySerGlyGlyGly, SEQ ID NO 796) и линкер из 21 аминокислоты (GlyGlyGlySerGlyGlyGlySerGlyGlyGlySerGlyGly GlySerGlyGlyGlySerGly, SEQ ID NO 797). Эти модульные матрицы, каждая из которых включает димер лигандов hflt3, разделенных ВamHI-содержащий линкер уникальной длины, конструировали следующим образом. Шесть промежуточных плазмидных матриц (FL3N, FL7N, FL11N, FL3C, FL4C и FL10C) были сформированы методом ПЦР с использованием пар затравок и pMON30238 в качестве матрицы с использованием параметров циклов ПЦР, какие были применены для pMON32322. Для реакции 50 пкМ каждой затравки добавляли к 100 нг денатурированной нагреванием матрице, и реакционную смесь составляли также, как это было описано выше для pMON32322. Условия ПЦР были следующими: 7 циклов при 94°С в течение 1 минуты, 2 минуты при 65°С и 2,5 минуты при 72°С с последующими 10 циклами по 1 минуте при 94°С, 2 минуты при 70°С и 2,5 минуты при 72°С. Единственный 7-минутный цикл достройки при 72°С завершал ПЦР. Пары затравок, использовавшиеся для создания каждой из промежуточных матриц, были следующими: N-term/FLN3 (FL3N), N-term/FLN7 (FL7N), N-term/FLN11 (FL11N), C-term/FLC3 (FL3C), C-term/FLC4 (FL4C) и С-term/FLC10 (FL10C). Продукты ПЦР-амплификации очищали с использованием реактивов Wizard PCR Clean-up Kit (Promega) и элюировали в 50 мкл дистиллированной воды. Очищенную ДНК для первого набора (FL3N, FL7N и FL11N) расщепляли рестриктазами NcoI/BamHI, очищали в геле, как это было описано выше, и лигировали в обработанный NcoI/BamHI/Sap вектор pSE420 (Invitrogen). Промежуточные матрицы второго набора (FL3C, FL4C и FL10C) создавали идентичным способом, за исключением использования рестриктаэы HindIII вместо NcoI. Последующие шаги по подтверждению конечной последовательности ДНК проводили так же, как это было описано выше.
Пример 95
Для получения шести конечных матриц два набора интермедиатов в рSЕ420 расщепляли либо рестриктазами NcoI/BamHI (FL3N, FL7N, FL11N - набор 1), либо рестриктазами BamHI/HindIII (FL3C, FL4C, FL10C - набор 2) и очищали в геле с использованием Geneclean II, как это было описано выше. По одному промежуточному амплификону из каждого набора лигировали в обработанную NcoI/HindIII/Sap плазмиду pMON3977 для каждой реакции ПЦР и трансформировали в клетках DH5α, как это было описано выше, с использованием следующих сочетаний с целью формирования специфических размеров линкеров: линкер из 6 аминокислот (FL3N и FL3C), линкер из 7 аминокислот (FL3N и FL4C), линкер из 10 аминокислот (FL7N и FL3C), линкер из 13 аминокислот (FL3N и FL10C), линкер из 15 аминокислот (FL11N и Н4С) и линкер из 21 аминокислоты (FL11N и FL10C). ДНК выделяли из единичных колоний 50-мл "переночевавших" культур для каждого из 6 сочетаний так, как это было описано выше, тестировали на правильность размера вставки путем рестрикционного анализа NcoI/HindIII и использовали в качестве матрицы. Пары затравок 39For/39Rev (точка разрыва 39/40, 65For/65Rev (точка разрыва 65/66) и 89For/89Rev (точка разрыва 89/90) использовали для ПЦР-амплификации каждой матрицы, как это было выше описано для pMON32322, за исключением того, что использовали по 75 пкМ каждой затравки. Условия амплификации были модифицированы следующим образом: 6 циклов при 94°С в течение 1 минуты, 2 минуты при 70°С, 2,5 минуты при 72°С с последующими 9 циклами при 94°С в течение 1 минуты и 72°С в течение 3 минут. После последнего цикла заключительную достройку затравок проводили при 72°С в течение 6 минут, что обеспечивало полную полимеризацию получаемых продуктов. Образцы очищали с использованием Wizard PCR Clean-up Kit (Promega), как это было описано выше, и подвергали двойному расщеплению рестриктазами NcoI/HindIII. Полученные продукты амплифицирования вновь очищали с использованием Wizard PCR Clean-up Kit. Дополнительно все шесть молекул с различной длиной линкера для точки разрыва 39/40 были клонированы в обработанную NcoI/HindIII/Sap плазмиду pMON3977 как гены отдельных белков (pMON32365, pMON32366, pMON32367, pMON32368, pMON32369 и pMON32370). Последующие шаги по подтверждению конечной последовательности ДНК проводили так же, как это было описано выше.
Пример 96
Были сконструированы гены, кодирующие молекулы многофункциональных химерных агонистов рецепторов, включающие агонист рецептора IL-3 (по pMON13416; WO 94/12638), соединенные через линкер IgG2b либо с нативным лигандом flt3, либо с агонистами рецептора flt3 с измененной последовательностью (согласно примерам 91-93). Вставки, включающие желательные молекулы агонистов рецептора flt3 с измененной последовательностью, выделяли из исходной плазмиды в виде рестрикционного фрагмента NcoI/Hindiii и лигировали в плазмиду pMON30304, расщепленную AflIII/HindIII/SAP. Последующие шаги по подтверждению конечной последовательности ДНК проводили так же, как это было описано выше. В табл.10 показаны полученные в результате плазмиды, включающие последовательности ДНК, кодирующие молекулы многофункциональных химерных агонистов рецепторов, включая агонист рецептора IL-3 (по pMON13416) и агонист рецептора flt3 с измененной последовательностью (см. табл. 10).
Пример 97
Были сконструированы гены, кодирующие молекулы многофункциональных химерных агонистов рецепторов, включающие агонист рецептора IL-3 (по pMON13288; WO 94/12638), соединенные через линкер IgG2b либо с нативным лигандом flt3, либо с агонистами рецептора flt3 с измененной последовательностью (согласно примерам 91-93). Вставки, включающие желательные молекулы агонистов рецептора flt3 с измененной последовательностью, выделяли из исходной плазмиды в виде рестрикционного фрагмента NcoI/HindIII и лигировали в плазмиду рМО30311, расщепленную AflIII/HindIII/SAP. Последующие шаги по подтверждению конечной последовательности ДНК проводили так же, как это было описано выше. В табл. 11 показаны полученные в результате плазмиды, включающие последовательности ДНК, кодирующие молекулы многофункциональных химерных, включая агонист рецептора IL-3 (по pMON13288) и агонист рецептора flt3 с измененной последовательностью.
Пример 98
Две химерные молекулы с агонистом рецептора hflt3 с перестроенной последовательностью с N-конца химерной молекулы были сконструированы с помощью ПЦР с использованием плазмид pMON32360 и pMON32362 в качестве матриц и пар затравок N-term/134rev и N-term/139rev с целью замены стоп-кодона в С-конце молекулы нативного лиганда flt3 и внесения в кодирующую рамку сайта рестрикции SnaBI. Реакционные смеси формировали также, как это было описано выше для pMON32322. Условия ПЦР были следующими: 7 циклов при 94°С в течение 1 минуты, при 65°С в течение 2 минут и при 72°С в течение 2,5 минут, дополнительно 10 циклов амплификации проводили таким образом, что температура отжига в них была увеличена с 65°С до 70°С. Образцы очищали с использованием Wizard PCR Purification Kit и элюировали в 50 мкл дистиллированой воды: по 20 мкл каждого образца расщепляли рестриктазами NcoI и SnaBI. ДНК плазмиды pMON26431 расщепляли рестриктазами NcoI и SnaBI и лигировали с использованием реакционной смеси ПЦР, обработанной NcoI/SnaBI. Трансформацию компетентных клеток DH5α и последующие шаги по подтверждению конечной последовательности ДНК осуществляли также, как это было описано выше.
Пример 99
Пять дополнительных точек разрывов в лиганде hflt3 были сделаны с использованием определенных затравок 28/29 (28For/28Rev), 34/35 (34For/35Rev), 62/63 (62For/62Rev), 94/95 (94For/94Rev) и 98/99 (98For/98Rev) с амплификацией линкеров (GlyGlyGlySer)х, состоящих из 10 и 15 аминокислот, как это было описано выше. Полученные в результате ПЦР-продукты расщепляли рестриктазами NcoI/HindIII и лигировали в плазмиду pMON30311, обработанную AflII/HindIII/SAP, как было описано выше. Трансформацию компетентных клеток DH5α и последующие шаги по подтверждению конечной последовательности ДНК осуществляли также, как это было описано выше.
Пример 100
Для усиления экспрессии агонистов рецептора hflt3 с перестроенной последовательностью в клетках E.coli были использованы специфичные для N-концов затравки, кодирующие вырожденные кодоны, с целью реорганизации форм 1-134 и 1-139 нативного лиганда hflt3 в составе экспрессирующего в E.coli вектора pMON5723. Пары затравок FH3AFor/SCF.rev (А1а2) и Flt23For/SCF (Gly2) использовали для ПЦР-амплификации N-концевой вырожденной смеси последовательностей, кодирующих нативный лиганд flt3 с использованием условий амплификации, какие были описаны для pMON23222, за исключением того, что число циклов амплификации снижено до 15 циклов при 95°С в течение 1 минуты, при 55°С в течение 2 минут и при 72°С в течение 2,5 минут. Амплификоны очищали с использованием Wizard РСЕ Clean-up Kit (Promega) и элюировали в 50 мкл дистиллированной воды. Расщепление рестриктазами NcoI/-HindIII и последующая гелевая очистка с использованием Geneclean II были проведены также, как это было описано выше. Вставки лигировали в обработанную NcoI/HindIII/SAP, очищенную в геле плазмидную ДНК pMON5723, которой трансформировали компетентные клетки DH5α, как это было описано ранее. Аликвоты трансформированных клеток высевали на чашках с агарозной средой LB, содержащей 75 мкг/мл спектиномицина, и инкубировали при 37°С в течение 14-16 часов, а затем подсчитывали колонии. После подтверждения колоний остатки каждой трансформационной смеси инкубировали в течение ночи (14-16 часов) при 37°С в 2х5 мл среды LB, содержащей 75 мкг/мл спектиномицина. Миниперпарированную ДНК готовили с использованием реактивов Wizard DNA 373 А Miniprep Kit (Promega) в соответствии с рекомендуемой процедурой. Очищенную минипреп-ДНК элюировали в 50 мкл дистиллированной воды и 1-2 мкл использовали для трансформации химически компетентных клеток MON207. Аликвоты объемом 25 и 200 мкл высевали в чашках со средой LB, содержащей 75 мкг/мл спектиномицина, и инкубировали при 37°С в течение 12-15 часов. По 40-50 хорошо отделенных колоний, представляющих каждую исходную пару затравок, отбирали, очерчивали на основных LВ/спектиномициных чашках и инкубировали еще в течение 4-6 часов при 37°С.
Клоны отдельных агонистов рецепторов hflt3 с перестроенными последовательностями скринировали по экспрессии в клетках E.coli в 96-ячейном микротитровальном устройстве с целью выбора улучшенных уровней экспрессии. По 100 мкл минимальной среды М9 (включая 1% свободных аминокислот) вводили в каждую ячейку наряду с единичной колонией (по 40-50 изолятов анализировали для каждой пары затравок в ПЦР hflt3), инкубировали при 37°С при 200 об/мин в течение 3-4 часов (1=0) и индуцировали добавлением в каждую ячейку по 5 мкл свежеприготовленного раствора 1 мг/мл налидиксовой кислоты (в 0,1 N NaOH). После дополнительных 4 часов инкубации при 37°С (1=4) приблизительные аликвоты по 5-10 мкл отбирали из каждой ячейки и анализировали на световом микроскопе на присутствие преломляющих свет телец, а результаты определяли как приблизительный процент клеток, содержащих преломляющие свет тельца по отношению к общему числу клеток. Клоны, характеризующиеся наивысшим уровнем экспрессии, отбирали для анализа шкалы уровней экспрессии, проводившегося по следующей схеме. 5 мл "переночевавшей" культуры растили в среде LB в присутствии 75 мкг/мл спектиномицина при 37°С. Инокулят удовлетворительных культивированных клеток добавляли к 10 мл свежеприготовленной минимальной среды М9 (с добавлением 1% свободных аминокислот) в 125-мл колбах-шейкерах с целью достижения исходного показателя в 20 единиц по Клетту: затем проводили инкубацию при 37°С в течение приблизительно 3-4 часов при взбалтывании до достижения уровня примерно в 110-150 единиц по Клетту (I=0) с последующим индуцированием 50 мкл свежеприготовленным раствором налидиксовой кислоты (10 мг/мл в 0,1 N NaOH). Аликвоты по 1 мл удаляли и клетки пеллетировали в течение 1 минуты на микроцентрифуге. Надосадочные фракции удаляли с помощью аспирации, а гранулы сохраняли при - 20°С до готовности проведения анализа в SDS-PAGE (электрофорезе в полиакриламидном геле с натрийдодецилсульфатом). Остатки индуцированных клеток инкубировали еще в течение 4 часов при 37°С при постоянном взбалтывании, после чего (время отсчета I=4) плотность клеток измеряли в единицах по Клетту. Аликвоты по 1 мл выделяли из каждого образца, пеллетировали центрифугированием и сохраняли также, как описано выше. Другие аликвоты по 5-10 мкл выделяли из каждой колбы и анализировали на световом микроскопе на присутствие преломляющих свет телец. Пеллетированные образцы ресуспензировали в объеме (в мкл) 2х загрузочного буфера (включающего 1% β-меркаптоэтанол), равном показателю по Клетту во время I=4, кипятили в течение 5 минут и 6-7 мкл переносили в 12%- или 14%-ный Трис-глициновый SDS-полиакриламидный гель (Novex) и подвергали электрофорезу в течение 90 минут при напряжении 90 В. Гели фиксировали, окрашивали и препарировали высушиванием в соответствии с рекомендуемой (Novex) процедурой. На данном этапе отбирали клоны для масштабирования с учетом повышения уровня экспрессии отдельного индуцированного белка, полоса которого на фореграмме в момент I=4 соответствовала предсказываемому размеру в сравнении с образцами в момент I=0.
Минипрепарированную ДНК также получали из отобранных клонов, экспрессирующих высокий уровень индуцированного белка, как это было описано ранее, а проверку итоговых последовательностей ДНК осуществляли точно также, как это было описано выше. Эти клоны были обозначены pMON32329, pMON32330, pMON32341 и pMON32342.
Пример 101
Набор химерных молекул многофункциональных агонистов рецепторов, включающий агонист рецептора IL-3 (из pMON13288) и нативный лиганд flt3, также был сконструирован для экспрессии в клетках E.coli. Гены, кодирующие химерные молекулы многофункциональных агонистов рецепторов из pMON32364 и pMON32377, были выделены из исходного вектора расщеплением рестриктазами NcoI/HindIII и лигированы в вектор pMON5677, с помощью которого трансформировали клетки MON207, а отдельные изоляты отбирали так, как это было описано выше. Эти конструкции были обозначены pMON32394 (вставка, происходящая из pMON32364) и pMON32395 (вставка, происходящая из pMON32377).
Пример 102
Укороченный рецептор flt3 был выделен в виде ПЦР-продукта длиной 1400 нуклеотидов с использованием 50 пкМ затравок FLTAFLS1 и FLTR1N и примерно 10 нг плазмидной ДНК pMON27184, используемой в качестве матрицы. Затравки были сформированы таким образом, чтобы образовать сайт рестрикции AflIII сразу с 5’-конца первого аспарагинового кодона последовательности, кодирующей зрелый полипептид, равно как и сайт рестрикции EcoRI сразу с 5’-конца последовательности, кодирующей трансмембранный сегмент. Реакционную смесь обрабатывали рестриктазами AflIII и EcoRI в стандартных реакционных условиях и осуществляли лигирование в расщепленную NcoI/EcoRI плазмиду pMON26458. Эта плазмида включает следующую последовательность ДНК: 5’-GGATCCACCATGAGCCGCCTGCCCGTCCTGCTCCTGCTC CAACTCCTGGTCCGCCCCGCCATGGCTAAAGCTT-3’ (SEQ ID NO 857), - кодирующую сигнальный участок полипептида IL-3. Эта последовательность включает сайт рестрикции BamHI в 5’-конце и включает метиониновый кодон ATG, соответствующий первой аминокислоте сигнального сегмента с 3’-стороны от BanlHI-сайта. Этот сигнальный сегмент отщепляется клеткой, оставляя с 5’-конца Met/Ala, образованный слиянием NcoI-сайта сигнального сегмента и AflIII-сайта собственно рецептора в реакции ПЦР. Полная укороченная форма рецептора вместе с сигнальным сегментом IL-3 может быть снова вырезана из вектора путем расщепления BamHI/EcoRI (hIL3L/hFlt3R).
Каталитический домен, кодируемый pMON30298 (hG-CSF), был реорганизован с целью создания с помощью ПЦР в пределах кодирующей рамки сайта рестрикции EcoRI на месте соединения трансмембранного и цитоплазматического доменов согласно следующей схеме. К 0,5 мкг денатурированной нагреванием pMON30298 добавляли по 100 пкМ каждой из затравок HGCFfor и HGCFrev, 10 мкл 5х буфера J, 5 ед. Taq-полимеразы и дистиллированной воды с достижением конечного объема в 45 мкл также, как это было описано выше. Амплификацию с помощью ПЦР проводили следующим образом: шесть циклов (1 минута при 94°С, 2 минуты при 64°С и 3 минуты при 70°С) с последующими 9 циклами (1 минута при 94°С и 4 минуты при 70°С). Окончательную достройку проводили в течение 7 минут при 70°С. По 10 мкл каждой ПЦР-реакционной смеси очищали в геле с использованием Geneclean II, как это было описано ранее, и элюировали в 10 мкл дистиллированной воды. Образцы расщепляли EcoRI и HindIII (по 10 ед. каждой рестриктазы) в 20 мкл реакционной смеси в течение 90 минут при 37°С. Образцы снова очищали в геле (с использованием Geneclean II), как это было описано выше, и элюировали в 10 мкл дистиллированной воды. 2 мкл вставки лигировали в 50 нг вектора рSЕ420, обработанного NcoI/HindIII/фосфатазой, в 10 мкл реакционной смеси так, как это было описано выше. Трансформацию компетентных клеток DH5α и последующие этапы подтверждения конечной последовательности ДНК проводили точно также, как это было описано выше. Отобранные клоны затем секвенировали с целью подтверждения присутствия в пределах кодирующей рамки EcoRI-сайта, также как и подтверждения последовательности ДНК, кодирующей каталитический домен рецептора G-CSF Клоны, включающие предсказанную последовательность, расщепляли рестриктазами EcoRI/HindIII, как это было описано выше, и очищали в геле. Очищенные вставки фрагментов hG-CSFR (EcoRI/-HindIII) и hIL3L/hFlt3R (BamHI/EcoRI) лигировали в вектор pcDNA-3.1(-) (Invitrogen), обработанный BamHI/HindIII/фосфорилазой. Трансформацию компетентных клеток DH5α и последующие этапы подтверждения конечной последовательности ДНК проводили точно также, как это было описано выше.
Пример 103
Дополнительные гены, кодирующие лиганды flt3 с перестроенными последовательностями, были сконструированы с использованием промежуточных димерных матриц, как это было описано выше. Димерные интермедиаты Flt4C.seq и Flt11N.seq были использованы в качестве матриц для получения агонистов рецепторов flt3 с перестроенными последовательностями, имеющих 15-аминокислотные линкеры (GlyGlyGlySer)3GlyGlyGly (SEQ ID NO 795). Точки разрывов, соответствующие аминокислотным остаткам 28/29, 34/35, 62/63, 94/95 и 98/99 в составе flt3, были сформированы на основе метода ПЦР с использованием реактивов PCR Optimizer Kit (Invitrogen) и следующих пар затравок: FL29For/FL29Rev, FL35For/FL35Rev, FL63For/FL63Rev, FL95For/FL95Rev и FL99For/FL99Rev, как это было описано в примере 94. Условия амплификации были следующими: семь циклов при 94°С в течение 1 минуты, при 62°С в течение 2 минут и при 70°С в течение 2,5 минут, затем двенадцать циклов при 94°С в течение 1 минуты, при 68°С в течение 2 минут и при 70°С в течение 2,5 минут, затем заключительный цикл при 72°С в течение 7 минут. Продукты ПЦР, соответствующие предсказанному размеру вставки, расщепляли рестриктазами NcoI и HindIII и очищали в геле с использованием Geneclean II (Biol91), как это было описано выше, в соответствии с рекомендуемой производителем процедурой. Образцы ресуспензировали в 10 мкл конечного объема дистиллированной воды. Вставки клонировали в виде отдельных генов в экспрессирующий вектор pMON3934 (обработанный NcoI/HindIII/SAP) для клеток млекопитающих и обозначали pMON35712, pMON35713, pMON35714, pMON35715, pMON35716, pMON35717 и pMON35718, соответственно.
Гены, кодирующие химерные белки, включающие агонист рецептора IL-3, кодируемый hMON13288 (WO 94/12638), обозначенный здесь как "агонист-I рецептора IL-3", и лиганд рецептора flt3 с перестроенной последовательностью, были сформированы путем клонирования очищенных, обработанных рестриктазами продуктов ПЦР, полученных при использовании пар затравок для точек разрывов 28/29, 34/35, 62/63, 94/95 и 98/99, в плазмиду pMON30311, обработанную NcoI/HindIII/SAP. Полученные в результате плазмиды были обозначены pMON32398, pMON35700, pMON35702, pMON35704 и pMON35706, соответственно. Дополнительно те же пары затравок были использованы в сочетании с промежуточными димерными матрицами Fl. t7N.seq и Flt3C.seq с целью конструирования форм этих химерных белков, состоящих из агониста-I рецептора IL-3 и Flt3L, с 10-аминокислотным линкером (GlyGlyGlySer)2GlyGly (SEQ ID NO 793): соответствующие плазмиды обозначены pMON32397, pMON32399, pMON35701, pMON35703 и pMON35705.
Пример 104
Гены, кодирующие химерные белки, состоящие из агонист-I рецептора IL-3 и Flt3L, а также 21-аминокислотный линкер (GlyGlyGlySer)5Gly (SEQ ID NO 796), были сконструированы с использованием такого же ПЦВ-метода и с использованием промежуточных димерных матриц FItllN.seq и FIt 10C.seq и следующих пар затравок: Flt36/36Rev, Flt37/37Rev, Flt38/38Rev, Flt39/39Rev, Flt41/41Rev, Flt42/42Rev и Flt43/43Rev. Эти пары затравок соответствуют следующим точкам разрывов в лиганде рецептора flt3 - 35/36, 36/37, 37/38, 38/39, 40/41, 41/42 42/43 (точка разрыва 39/40 была прежде использована при конструировании плазмиды pMON32376): они были использованы для ПЦР-амплификации в следующих условиях: семь циклов при 94°С в течение 1 минуты, при 66°С в течение 2 минут и при 70°С в течение 2,5 минут: затем пятнадцать циклов при 94°С в течение 1 минуты и при 70°С в течение 4 минут, заключительный цикл проводили при 72°С в течение 7 минут; использовали реактивы Invitrogen PCR Optimizer Kit (с буфером В). После подтверждения последовательностей ДНК эти конструкции были обозначены как pMON35733, pMON35734, pMON35735, pMON35736, pMON35738, pMON35739, pMON35740, pMON35741, pMON35742 и pMON35743, соответственно. Как часть этой серии вариантов, были сконструированы и протестированы ошибочные по ПЦР-инкорпорации варианты, в результате которых в составе Flt3 химерной молекулы имеются две точковые аминокислотные замены (pMON35741, точка разрыва 35/36, и pMON-35743, точка разрыва 42/43), а один вариант (pMON35742, точка разрыва 38/39) включал две аминокислотные замены (Q133 → R133, Q100 → R100, L112 → Р112) в составе части Flt3L.
Дополнительно химерные белки, состоящие из Flt3L и агониста-I рецептора IL-3, в которых имелись точки разрывов Flt3L, соответствующие остаткам аминокислот 28/29, 34/35, 62/63, 65/66, 89/90, 94/95 и 98/99 в аминокислотной последовательности лиганда flt3, как это было описано выше, были сконструированы с 15-аминокислотным линкером (GlyGlyGlySer)3GlyGlyGly на матрицах FLt4C и FLtllN. Смеси реакционных ПЦР-смесей были сходными с таковыми, которые были описаны в примере 103, за исключением того, что обратные затравки, соответствующие С-концам Flt3 с перестроенными последовательностями, были модифицированы путем замещения сайта рестрикции HindIII сайтом распознавания рестриктазой SnaBI. Параметры ПЦР-амплификационных циклов были следующими: семь циклов при 94°С в течение 1 минуты, при 66°С в течение 2 минут и при 70°С в течение 2,5 минут, затем четырнадцать циклов при 94°С в течение 1 минуты и при 70°С в течение 4 минут, затем заключительный цикл при 72°С в течение 7 минут. Выделение продуктов ПЦР, обработку рестриктазами и очистку проводили также, как это было описано выше. Вставки лигировали в плазмиду pMON26431 (экспрессирующий вектор для клеток ВНК, включающий агонист-I рецептора IL-3 и линкер IgG2b), обработанный NcoI/-SnaBI/SAP, по следующей схеме: 50 нг обработанного вектора, вставку (при соотношении вставки и вектора 10:1), 1 ед. ДНК-лигазы Т 4 (Gibco BRL) и 1 мкл 10х лигазного буфера смешивали в 10 мкл реакционного объема. Лигирование проводили в течение 1 часа при температуре окружающей среды, затем 2 мкл каждой реакционной смеси отбирали и использовали для трансформации 100 мкл химически компетентных клеток DH10B (Gibco BRL) (как альтернативу клеткам DH5α) в соответствии с рекомендуемой производителем процедурой. Одну пятую и 1/25 объемов каждой трансформационной смеси высевали на чашках со средой LB, в которую добавляли необходимые маркерные антибиотики: инкубацию проводили в течение ночи (14-16 часов) при 37°С. Выделенные колонии сортировали и выделяли ДНК в соответствии с мидипрепарационной процедурой (Oiagen), как это было описано выше.
Секвенирование отобранных клонов подтвердило присутствие точек разрывов 28/29 (pMON35719), 34/35 (pMON35720), 62/63 (pMON35721), 65/66 (pMON35722), 89/90 (pMON35723) и 98/99 (pMON35725). Плазмида pMON35726 содержит точковую аминокислотную замену (лейцина на фенилаланин в 94-м положении) в точке разрыва 94/95. Химерная конструкция, состоящая из Flt3L и агониста-I рецептора IL-3, с точкой разрыва в положении 39/40 в составе Flt3L и варьирующимся числом аминокислот в линкере (10, 15 или 21) представлены плазмидами pMON35707, pMON35708, pMON35709, pMON35710 и pMON35711. Эти конструкции были формированы с помощью ПЦР-амплификации одной из следующих матриц: pMON32373, pMON32375 или pMON32376 и специфичной для Flt3L пары затравок 39N, term-l/SNABIC,term.
Стандартные ПЦР-реакционные смеси компоновали, как это было описано выше, а ДНК амплифицировали по следующей схеме: семь циклов при 94°С в течение 1 минуты, при 62°С в течение 2 минут и при 70°С в течение 2,5 минут, затем двенадцать циклов при 94°С в течение 1 минуты, при 68°С в течение 2 минут и при 70°С в течение 2,5 минут, заключительный цикл проводили при 72°С в течение 7 минут. Продукты ПЦР, соответствующие предсказываемому размеру вставки, расщепляли полностью рестриктазами NcoI и SnaBI, очищали в геле и клонировали в соответствии с вышеописанной процедурой в экспрессирующий вектор pMON26431 (обработанный NcoI/SnaBI/SAP) для клеток млекопитающих, как фактор химерных белков, включающих Flt3L, IgG2b и агонист-I рецептора IL-3. Две из этих конструкций содержали ошибки ПЦР-инкорпорации в составе части химерного белка F113, в результате которых имелись точковые замены аминокислот (F96 → L96 и Е58 → G58) (соответственно, плазмиды pMON35710 и pMON35711).
Пример 105
Была также сконструирована другая серия химерных белков, включающих Flt3L с перестроенной последовательностью и агонист-I рецептора IL-3 с точками разрывов, соответствующими аминокислотным остаткам 35/36, 36/27, 38/39, 40/41, 41/42, 42/43 и 65/66 в полипептиде лиганда Flt3, как это было описано выше: в качестве матриц использовали отобранные конструкции/ кодирующие агонист-I рецептора IL-3 и Flt3L с перестроенной последовательностью (см. табл. 12). Одно исключение было связано с тем, что матрица для 15-аминокислотного линкера (pMON35715) была использована для конструирования варианта с точкой разрыва 65/66 (pMON35771).
Конструкции Flt3L/агонист-I рецептора IL-3
Были использованы пары затравок, соответствующие тем же сайтам рестрикции, которые были использованы при конструировании плазмид pMON35719-35725. Обратные затравки 36Rev’, 37Rev’, 38Rev’, 39Rev’, 41Rev’, 42Rev’ и 43Rev’ были использованы для создания сайта рестрикции SnaBIs в пределах кодирующей рамки. Были использованы такие же прямые затравки Flt36, Flt37, Flt38, Flt39, Flt41, Flt42 и Flt43. ПЦР-реакционные смеси были идентичны с таковыми, которые были описаны выше, однако за исключением pMON35771, а условия амплификации были модифицированы следующим образом: 18 циклов при 94°С в течение 1 минуты, при 68°С в течение 2 минут и при 70°С в течение 2,5 минут, затем единственный цикл достройки при 70°С в течение 7 минут. Для pMON35771 условия амплификации были следующими: шесть циклов при 94°С в течение 1 минуты, при 68°С в течение 2 минут и при 70°С в течение 2,5 минут, затем пятнадцать циклов при 94°С в течение 1 минуты и при 70°С в течение 4 минут, заключительный цикл проводили при 72°С в течение 7 минут. Специфичные для Flt3 продукты ПЦР-амплификации расщепляли рестриктазами, очищали и клонировали в плазмиду pMON26431 (экспрессирующий вектор, кодирующий агонист-I рецептора IL-3 и линкер IgG2b, для клеток ВНК), как это было описано в примере 104. Один из вариантов (pMON-32179) был сконструирован по точке разрыва 34/40 с использованием пары затравок для ПЦР Flt40/34Rev и промежуточных димерных матриц FIt11lN.seq и FIt10C.seq. Условия ПЦР-амплификации и последующего клонирования были идентичными тем, которые были использованы для клона pMON35771. Три дополнительных химерных белка, включающих Flt3L и агонист-I рецептора IL-3 (точка разрыва 38/39), были сформированы с тем, чтобы тестировать влияние различной длины линкера и состава белка. С использованием pMON35709 в качестве матрицы, длина линкерного мотива GlySer была распространена до 29 остатков по формуле (GlyGlyGlySer)7Gly с использованием пар затравок BamForl/38Rev (продукт реакции, обозначенный как ПЦР-А) и Flt38/BamRev1 (продукт реакции, обозначенный как ПЦР-В). Условия амплификации были следующими: шесть циклов при 94°С в течение 1 минуты, при 66°С в течение 2 минут и при 70°С в течение 2,5 минут, затем пятнадцать циклов при 94°С в течение 1 минуты и при 70°С в течение 4 минут, затем заключительный цикл проводили при 72°С в течение 7 минут. Полученные в результаты ПЦР-продукты выделяли с использованием PCR Clean-up Kit (Pronega), расщепляли либо рестриктазами NcoI/BamHI (ПЦР-А), либо рестриктазами BamHI/SnaBI (ПЦР-В), очищали в геле и лигировали в состав pMON26431 (экспрессирующий вектор, кодирующий агонист-I рецептора IL-3 и линкер IgG2b, для клеток В НК), как это было описано выше. Полученную в результате конструкцию подтверждали по включенной последовательности и обозначали pMON35774. В сравнительном аспекте варианты pMON35775 и pMON35776 различались в том, что их линкерный мотив GlySer был замещен аминокислотными фрагментами 140-154 (pMON35775) или 140-160 (pMON35776) нативного Flt3L, содержащими точковую замену аминокислот. Условия ПЦР были идентичными тем, которые были описаны для плазмиды pMON35774, за исключением того, что были использованы следующие пары затравок: 38For/Navfor и 38Rev/NavRevS (для pMON35775) и 38For/Navfor и 38Rev/NavRevL (для pMON35776). Рестриктаза KasI была заменена рестриктазой BamHI при том, что этапы клонирования этих продуктов ПЦР-амплификации были идентичны тем, которые использовались для pMON35774. Секвенирование показало, что ПЦР привела к индукции ошибок во множественных изолятах и в pMON35775, и в pMON35776. Для выявления конечных точных последовательностей необходимо было перерасщепить отобранные субклоны рестриктазами NarI/SnaBI и NcoI/NarI с использованием этих очищенных в геле фрагментов для реклонирования желательных конструкций.
Ряд химерных молекул димерного Flt3L был также сконструирован для тестирования экспрессии в клетках ВНУК. Была собрана плазмида pMON32173, включающая гены двух нативных молекул Flt3L, соединяемых линкером IgG2b, с использованием 2 предварительных молекул следующим образом: содержащая Flt3L NcoI/SnaBI-вставка из pMON32393 была лигирована в очищенную в геле, разрезанную NcoI/SnaBI плазмиду pMON32377, в которую также был внесен ген агониста-L рецептора IL-3, являющегося партнером Flt3L в химерном белке.
Сходным образом pMON35727 (точка разрыва 39/40, линкер из 15 аминокислот) была сконструирована путем внесения вставки Flt3L из pMON35798 (как NcoI/SnaBI-вставки) в очищенную в геле плазмиду pMON32375, из которой ген агониста-I рецептора IL-3, являющегося партнером Flt3L в химерном белке, был вырезан. Третий димер Flt3L - pMON32168 (точка разрыва 39/40, линкер из 21 аминокислоты) был сконструирован следующим образом; NcoI/SnaBI-вставку из состава pMON32165 (включенный в E.coli эквивалент pMON35709, сформированный путем субклонирования NcoI/BamHI-фрагмента из pMON32163 и BamHI/HindIII-фрагмента из pMON35709 в состав pMON5723, обработанной NcoI/HindIII). NcoI/SnaBI-вставку из pMON32165 (Flt3L 1-139 (39/40)L21) и SnaBI/HindIII-вставку из pMON32376 (IgG2b/Flt3L 1-139 (39/40L21) субклонировали в продукционный вектор pMON5723 E.coli, что позволило сформировать pMON32167. Затем NcoI/HindIII-вставку из pMON32167 субклонировали в состав pMON30304 и обозначили как pMON32168.
Пример 106
Серия тримерных молекул, каждая из которых содержит по две части Flt3L и по единственной копии агониста-I рецептора IL-3 или агониста-II рецептора IL-3 (агонист рецептора IL-3, кодируемый плазмидой pMON13416 [WO 94/12638], обозначаемый здесь как "агонист-II рецептора IL-3") также была сконструирована из предшествующих молекул с использованием рестрикционного расщепления с очисткой фрагментов в геле. Плазмида pMON35728 была собрана с использованием NcoI/EcoRI-вставки (Flt3L/IgG2b/агонист-I рецептора IL-3) из состава pMON32375 и EcoRI/HindiII-вставки (агонист-I рецептора IL-3/IgG2b/Flt3L) из состава pMON35708. Эти два фрагмента затем перелигировали в обработанный NcoI/HindIII/SAP вектор pMON3934 для клеток млекопитающих и субклонировали так, как это было описано выше. Плазмиду pMON32205 (агонист-II рецептора IL-3/IgG2b/Flt3 l-139/IgG2b/Flt3 1-139) собирали путем лигирования NcoI/HindIII-фрагмента из состава pMON32173 в AflIII/HindIII-сайт плазмиды pMON30304. Сходный подход был использован для конструирования плазмиды pMON32206 (агонист-II рецептора IL-3/IgG2b/-Flt3L (39/40)L21/IgG2b/Flt3L(39/40)L21). NcoI/HindIII-фрагмент из состава pMON32167 был очищен в геле и субклонирован в состав AflIII/HindIII-расщепленной плазмиды pMON30304 (в состав которой частью входит агонист-II рецептора IL-3/IgG2b). Плазмида pMON32207 (Flt3L (39/40)L21/IgG2b/Flt3L (39/40)L21/G-CSF) была собрана путем субклонирования очищенной в геле NcoI/HindIII-вставки из плазмиды pMON32170 в состав промежуточной плазмиды pMON32198 (AflIII/HindIII).
***Плазмида pMON32208 (Flt3L l-139/IgG2b/G-CSF/IgG2b/Flt3L 1-139) была собрана путем субклонирования очищенной в геле SnaBI-вставки из плазмиды pMON30320 (как IgG2b/G-CSF) в состав расщепленной SnaBI, обработанной SAP плазмиды pMON32173. Плазмида pMON32204 была собрана путем субклонирования NcoI/HindIII-вставки из состава pMON32173 в обработанную рестриктазами AflII/HindIII плазмиду pMON30309 (которая включает G-CSF/IgG2b). Плазмида pMON32195 (Flt3L 1-139(39/40)L21/IgG2b/G-CSF/Flt3L 1-139(39/40) L21) была сконструирована путем субклонирования NcoI/SacI-вставки из состава pMON32190 и SacI/HindIII-фрагмента из состава pMON32171 в расщепленную рестриктазами NcoI/HindIII плазмиду pMON30304. Плазмида pMON32196 (G-CSF/IgG2b/Flt3L 1-139(39/40) L21/IgG2b/Flt3L 1-139) была собрана путем субклонирования NcoI/HindIII-фрагмента из состава pMON30309 (как G-CSF/IgG2b) в обработанную NcoI/SAP плазмиду pMON32168 (Flt3L1-139(39/40)L21/IgG2b/Flt3L 1-139) с подтверждением ориентации с помощью секвенирования ДНК и рестрикционного анализа. Плазмида pMON32197 (G-CSF/IgG2b/Flt3L 1-139(39/40) L21/IgG2b/Flt3L 1-139(39/40)L21) была сконструирована путем субклонирования NcoI/HindIII-вставки pMON32167 (Flt3L 1-139(39/40) L21/IgG2b/Flt3L 1-139(39/40) L21) в AflIII/HindIII-сайт плазмиды pMON30309 (G-CSF/IgG2b).
Пример 108
Серия молекул, включающих Flt3L, была сконструирована в качестве экспрессантов в клетках ВНК путем замещения агониста-I рецептора IL-3 или агониста-II рецептора IL-3 сегментом G-CSF, как химерного партнера. При формировании химерных белков, по перемежающемуся типу экспрессируемых клетками ВНК и созданных с помощью вектора pMON3934, сегмент G-CSF может кодировать в 17-м положении полипептида как серия, так и цистеин. В молекулах, экспрессируемых клетками E.coli или экспрессируемых по неперемежающемуся типу в клетках млекопитающих, положение 17 в составе G-CSF должно быть представлено исключительно серином. Химерный белок, включающий нативные Flt3L и G-CSF, был сконструирован для экспрессии в клетках ВНК при обоих вариантах ориентации: G-CSF/IgG2b/Flt3L (pMON-30329) и Flt3L/IgG2b/G-CSF (pMON32175). Плазмида pMON30329 была собрана путем субклонирования вставки Flt3L 1-139 из состава pMON30238 (как продукт расщепления NcoI/HindIII) в плазмиду pMON30309 (которая содержит G-CSF/IgG2b), расщепленную рестриктазами AflIII/HindIII, в то время как плазмида pMON32175 была сконструирована с использованием очищенной в геле NcoI/SnaBI-вставки из состава pMONM32393 в обработанную рестриктазами NcoI/SnaBI плазмиду pMON26420 (которая содержит ген IgG2b/G-CSF). Третья химерная молекула с нативными G-CSF/Flt3L (pMON32191) отличается от pMON32175 тем, что он включает линкер GlySer на месте химерного линкера IgG2b: она была сформирована для экспрессии в клетках E.coli. Плазмида pMON32191 был собрана с использованием такой же очищенной в геле NcoI/SnaBI-вставки из состава pMON32393 в расщепленную рестриктазами NcoI/SnaBI плазмиду pMON31123 (которая включает ген GlySer/G-CSF). Эквивалент для клеток ВНК (pMON35767) был создан путем субклонирования очищенного в геле химерного NcoI/HindIII-гена из состава pMON32191 в вектор pMON3934, приспособленный для экспрессии в клетках ВНК.
Пример 109
Две серии химер, включающих Flt3L с перестроенной последовательностью, были сконструированы путем замещения агониста-I рецептора IL-3 другим компонентом - G-CSF. Первый набор, характеризующийся ориентацией G-CSF/IgG2b/Flt3L с перестроенной последовательностью, был создан следующим образом; плазмиду pMON30329 (G-CSF/IgG2b/Flt3L 1-139) расщепляли рестриктазами SnaBI/HindIII, а вектор, содержащий ген компонента G-CSF, очищали в геле, как это было описано выше. Обработанные рестриктазами SnaBI/HindIII вставки из подходящих конструкций, включающих агонист-I рецептора IL-3/Flt3L, показанных ниже в табл. 13, были затем субклонированы в плазмиду pMON30329 (SnaBI/HindIII).
Конструкции G-CSF/IgG2b/Flt3L и их аналоги, включающие агонист-I рецептора IL-3
Плазмида pMON32169 (G-CSF/IgG2b/Flt3L 1-139(39/40)L21) была создана с использованием NcoI/BamHI-вставки из состава PMON32163 и ВаmНI/-НindIII-вставки из состава pMON32370, субклонированных в плазмиду pMON30309, расщепленную рестриктазами AflIII/HindIII. Три молекулы в этой серии не включали прямо ориентированных компонентов агониста-I рецептора IL-3. Первая из них (pMON39914) была собрана с использованием экспрессирующего в клетках ВНК вектора pMON30309 (который включает G-CSF/IgG2b), расщепленного рестриктазами AflIII/ HindIII, и вставки Flt3L 1-139(39/40)L29 из состава pMON32243 (как NcoI/HindIII). При создании плазмиды pMON39915 ген Flt3L 1-154 (39/40) из состава pMON32242 (как NcoI/HindII-вставка) был субклонирован в исходный вектор pMON30309. Плазмида pMON39916 была создана точно так же как pMON39915, за исключением того, что была использована вставка Flt3L 1-160 (39/40) из состава pMON32252. Плазмиды pMON32242, pMON32243 и pMON32252 являются конструкциями, экспрессирующимися в E.coli, и включают нехимерный ген Flt3L с перестроенной последовательностью (как NcoI/HindIII). Наконец вставка из состава pMON35799 была субклонирована в плазмиду pMON5723 (как NcoI/HindIII-фрагмент) с целью экспрессии в клетках E.coli. Эта продуктивная в E.coli плазмида была обозначена pMON39904.
Пример 110
Многие химеры G-CSF из второй серии, характеризующиеся ориентацией Flt3L/IgG2b/G-CSF, также были сконструированы на основе их аналогов, включающих агонист-I рецептора IL-3, как это показано в табл.14.
Конструкции Flt3L/IgG2b/G-CSF и их аналоги, включающие агонист-I рецептора IL-3
Эти конструкции были собраны с использованием расщепленной рестриктазами NcoI/SnaBI плазмиды pMON36113 (вектор для клеток ВНК, включающий ген IgG2b/G-CSF) и специфической, расщепленной NcoI/SnaBI вставки Flt3L с перестроенной последовательностью из состава химерных белков, включающей агонист-I рецептора IL-3/Flt3L, показанных в табл.14. Полученные в результате плазмиды были обозначены pMON32170, pMON32871, pMON32271, pMON32172, pMON32174, pMON35751, pMON35752, pMON35753, pMON35754, pMON35755, pMON35756, pMON35757, pMON35758, pMON35759, pMON35760, pMON35761, pMON35762, pMON35763, pMON35764, pMON35765, pMON35766, pMON35767, pMON35768, pMON35770, pMON35772, pMON35773, pMON35777, pMON35778, pMON35779, pMON35780, pMON35782 и pMON39908.
Плазмиды pMON35777 и pMON35778 были сконструированы с помощью ПЦР и выстроены с теми же NcoI/NarI- и NarI/SnaBI-вставками, какие были описаны для pMON35775 и pMON35776, за исключением того, что расщепленная рестриктазами NcoI/SnaBI плазмида pMON35751 была использована в качестве исходного вектора, включающего ген ISG2b/G-CSF. Для конструирования эквивалента pMON35778, несущего точку разрыва 39/40, пара затравок Flt40/SnaBI-Cterm была использована для реамплификации матрицы pMON35778. Применены те же самые условия амплификации, которые были описаны выше для плазмиды pMON35771, за исключением того, что исходная температура отжига была снижена с 66 до 55°С. Полученная в результате конструкция была обозначена pMON35782 (Flt3 1-160 (39/40)/IgG2b/G-CSF).
Плазмида pMON32170 (Flt3L 1-139(39/4)L21/IgG2b/G-CSF) была создана с использованием NcoI/SnaBI-вставки из состава pMON32165, лигированной в расщепленную рестриктазами NcoI/SnaBI плазмиду pMON26430 (которая включает IgG2b/G-CSF). Плазмида pMON35764 (Flt3L (38/39)L21/IgG2b/G-CSF) была клонирована следующим образом: вставка Flt3L с перестроенной последовательностью была амплифицирована с применением ПЦР с использованием pMON35736 в качестве матрицы и пары затравок Flt39/39Rev. Условия амплификации были теми же, которые были использованы для pMON35771, за исключением того, что исходная температура отжига была снижена с 66 до 56°С. Расщепленный рестриктазами NcoI/SnaBI продукт ПЦР-амплификации был субклонирован в расщепленную рестриктазами NcoI/SnaBI плазмиду pMON35754, включающую ген IgG2b/G-CSF. Плазмида pMON35768 (Flt3L (38/39)L21/IgG2b/G-CSF) несет мутацию в 15-м остатке (замена серина на фенилаланин) части Flt3 химерного белка.
Плазмиды pMON35762 (Flt3 с матрицы pMON35739), pMON35763 (Flt3 с матрицы pMON35738), pMON35758 (Flt3 с матрицы pMON35740), pMON35770 (Flt3 с матрицы pMON35743) были сконструированы точно также, как это было описано для pMON35764.
Плазмида pMON35772, представляющая мутацию серин125 → фенилаланин125 в гене Flt3 с перестроенной последовательностью (согласно pMON35760), была клонирована с помощью ПЦР с использованием pMON35715 в качестве матрицы для Flt3 и пары затравок 65For/65SnaBI. Условия циклов ПЦР были идентичны тем, которые были использованы для амплификации генов Flt3 из состава плазмид pMON35733, pMON35734, pMON35735 и pMON35736, описанных выше. Плазмида pMON35761 представляет мутацию глутамин133 → аргинин133 в гене Flt3 с перестроенной последовательностью из состава плазмиды pMON35758. Плазмида pMON35773 (Flt3L 1-139 (38/39)L29/-IgG2b/G-CSF) была клонирована также, как это было описано выше для плазмиды pMON35774, за исключением того, что в качестве исходного вектора использовали плазмиду pMON25430, обработанную NcoI/SnaBI/SAP, включающую ген IgG2b/G-CSF.
Для конструирования эквивалента, характеризующегося точкой разрыва 39/40, плазмида pMON35773 была использована в качестве матрицы в ПЦР-амплификации с парой затравок Flt40/SnaBI-Cterm. Амплификация была проведена точно так же, как это было описано для pMON35771. Расщепленный рестриктазами NcoI/SnaBI продукт амплификации был субклонирован в плазмиду pMON26430 (обработанную NcoI/SnaBI/SAP), в результате чего была создана плазмида pMON35779 (Flt3L 1-139 (39/40)L29/IgG2b/G-CSF). Плазмида pMON35780 является вариантом pMON35779 и кодирует часть Flt3 с перестроенной последовательностью в составе химерного белка, несущую мутацию лейцин60 → пролин60. Плазмида pMON32190 (Flt3L 1-139 (39/40)L21/GS/G-CSF) включает альтернативный химерный линкер GlySer, который замещает линкер IgG2b, имеющийся в плазмиде pMON32170. NcoI/SnaBI-фрагмент гена Flt3L из состава pMON32165 (Flt3L 1-139 (39/40)L21/IgG2b/агонист-I рецептора IL-3 в составе вектора pMON5723, экспрессирующегося в клетках E.coli) был субклонирован в плазмиду pMON31123, расщепленную рестриктазами NcoI/SnaBI. Эквивалентный вариант pMON35766, предназначенный для экспрессии в клетках ВНК, был сконструирован путем субклонирования в плазмиду pMON3934 полной химерной вставки Flt3L/GlySer/G-CSF в виде NcoI/HindIII-фрагмента.
Плазмида pMON39908 сходна с pMON35779, за исключением того, что аминокислотные остатки 133-160 в составе Flt3L замещены аминокислотной последовательностью VETVFHRVSQDGLDLLTS (SEQ ID NO 798), которая гомологична варианту Flt3L, образующемуся в результате альтернативного сплайсинга (№ в базе данных GenBank HSU29874). Плазмида pMON32190 была использована в качестве матрицы для ПЦР с такими использованными парами затравок; Flt40/XbaRev и SnaBICterm/XbaFor. Условия амплификации были такими же, как это было описано для pMON35771, за исключением того, что температура отжига была снижена с 66 до 64°С. Оба очищенных в геле продукта ПЦР-амплификации были обработаны либо рестриктазами NcoI/Xbal (ПЦР-продукт Flt40/XbaRev), либо рестриктазами XbaI/SnaBI (ПЦР-продукт SnaBICterm/XbaFor) и субклонированы в плазмиду pMON26430 (обработанную NcoI/SnaBI/SAP). Плазмида pMON32273 (Flt3L 1-139 (39/40) L21/IgG2b/Cs-CSF) была сконструирована с помощью ПЦР pMON35777 с использованием пар затравок FltConNco/Grev с целью реамплификации варианта Flt31 38/39 в вариант Flt3L с точками разрыва 39/40. Очищенный амплификон обрабатывали рестриктазами NcoI/SnaBI и субклонировали в расщепленную NcoI/SnaBI плазмиду pMON32191, а полученный вариант обозначали pMON32259 (для экспрессии в клетках E.coli). Для экспрессии в клетках ВНК NcoI/HindIII-вставка из состава pMON32259 была субклонирована в плазмиду pMON3934 (NcoI/HindIII).
Пример 103
(на самом деле - Пример 111)
Другая серия химерных белков была сконструирована: в ней часть Flt3L характеризовалась одной или двумя мутациями в остатках цистеина (табл. XIA и XIB). Плазмида pMON35790 (Flt3L 1-139 [цистеин4 → серин4, цистеин85 → серин85]/GS/G-CSF[Ser17]) была сконструирована с помощью ПЦР с использованием pMON32191 в качестве матрицы и пар затравок ClFor/C3Rev и C3For/-139Rev в двух реакциях. Плазмида pMON35791 (Flt3L 1-139 [цистеин93 → серин93, цистеин132 → серин132]/GS/G-CSF[Ser17]) также была сконструирована с помощью ПЦР с использованием pMON32191 в качестве матрицы и пар затравок CSFor/C6Rev и CSRev/N-term. Условия амплификации были теми же, что были описаны выше для pMON35771, за исключением того, что температура отжига была снижена с 66 до 64°С. Второй раунд ПЦР был проведен с использованием амплификонов (10 мкл каждого амплификона) из первого раунда ПЦР; затем продукты ПЦР были очищены, обработаны рестриктазами NcoI/SnaBI и субклонированы в расщепленную рестриктазами NcoI/SnaBI плазмиду pMON-32191. Условия ПЦР-амплификации во втором раунде были модифицированы следующим образом: исходная температура отжига была увеличена до 68°С, а число циклов увеличено с 6 до 15. При этом дополнительная амплификация не требовалась. Эти конструкции - pMON35787 (цистеин4 → серин4, цистеин85 → серин85) и pMON35788 (цистеин93 → серин93, цистеин132 → серин132) - были использованы для экспрессии в клетках E.coli. Эквиваленты для экспрессии клеток ВНК - pMON35790 и pMON35791 - были сконструированы путем субклонирования правильно мутировавших химерных вставок Flt3L/GlySer/G-CSF, взятых в виде NcoI/HindIII-фрагментов в плазмиду pMON3934. Плазмида pMON35792 (Flt3L l-132 [цистеин132 → серин132]/GlySer/G-CSF [Ser17]) была сконструирована с помощью ПЦР с использованием pMON32191 в качестве матрицы и пары затравок FLDlRev/FltNTerm.
Плазмида pMON39905 (Flt3L 1-139 [цистеин132 → серин132]/ GlySer/CSF [Ser17]) была сконструирована с помощью ПЦР с использованием pMON32191 в качестве матрицы и пары затравок FLMlRev/FltNTerm.
Плазмида pMON39906 (Flt3L 1-139 [цистеин127 → серин127, цистеин32 → серин32]/GlySer/G-CSF [Ser17]) содержит единственную аминокислотную замену в 127-м положении части Flt3L с перестроенной последовательностью, являющуюся результатом индуцированной ПЦР ошибки, произошедшей при ПЦР-амплификации pMON39905.
Плазмида pMON32276 (Flt3L 1-139 (39/40)L21 [цистеин4 → серин4, цистеин85 → серин85]/GlySer/G-CSF [Ser17]) была сконструирована в двух раундах ПЦР. Были сформированы три исходных амплификона: ПЦР1 (pMON32190 в качестве матрицы и пары заправок G10L/85N); ПЦР7 (pMON32190 в качестве матрицы и пары затравок 4N/85S) и ПЦР4 (pMON32198 в качестве матрицы и пары затравок 4S/3605Rev). Для второго раунда ПЦР1, ПЦР4 и ПЦР7 были реамплифицированы в комбинированной смеси, в результате чего была получена ПЦР-А. ПЦР-А была очищена, обработана рестриктазами NcoI/SnaBI и субклонирована в плазмиду pMON30277 (GlySer/G-CSF). Следующие три конструкции были сформированы сходным образом. Плазмида pMON32277 (G-CSF [Ser17]/IgG2b/ Flt3L 1-139 (39/40) L21 [цистеин4 → серин4, цистеин85 → серии85]) в первом раунде ПЦР сформировала три исходных амплификона: ПЦР1 (pMON32190 в качестве матрицы и пары затравок G10L/85N); ПЦР7 (pMON32190 в качестве матрицы и пары затравок 4N/85S) и ПЦР6 (pMON32169 в качестве матрицы и пары затравок 4S/3605Rev). Для второго раунда ПЦР1, ПЦР4 и ПЦР6 были реамплифицированы в комбинированной смеси, в результате чего была получена ПЦР-В. ПЦР-В была очищена, обработана рестриктазами NcoI/HindIII и субклонирована в расщепленную рестриктазами NcoI/HindIII плазмиду pMON30309 (G-CSF [Ser17]/IgG2b). Плазмида pMON32278 (Flt3L 1-139 (39/40)L21 [цистеин93 → серин93, цистеин132 → серин132]/GlySer/G-CSF [Ser17]) в первом раунде ПЦР сформировала три исходных амплификона: ПЦР2 (pMON32190 в качестве матрицы и пары затравок G10L/93N); ПЦР8 (pMON32190 в качестве матрицы и пары затравок 132N/93S) и ПЦРЗ (pMON32198 в качестве матрицы и пары затравок 132S/3605Rev). Для второго раунда ПЦР2, ПЦРЗ и ПЦР8 были реамплифицированы в комбинированной смеси, в результате чего была получена ПЦР-С. ПЦР-С была очищена, обработана рестриктазами NcoI/SnaBI и субклонирована в плазмиду pMON30277 (GlySer/G-CSF). Плазмида pMON32279 (G-CSF [Ser17]/IgG2b/Flt3L 1-139 (39/40)L21 [цистеин93 → серин93, цистеин132 → серин132] в первом раунде ПЦР сформировала три исходных амплификона: ПЦР2 (pMON32190 в качестве матрицы и пары затравок G10L/93N); ПЦР8 (pMON32190 в качестве матрицы и пары затравок 132N/93S) и ПЦР5 (pMON32169 в качестве матрицы и пары затравок 132S/3605Rev). Для второго раунда ПЦР2, ПЦР5 и ПЦР8 были реамплифицированы в комбинированной смеси, в результате чего была получена ПЦР-D. ПЦР-D была очищена, обработана рестриктазами NcoI/HindIII и субклонирована в расщепленную рестриктазами NcoI/HindIII плазмиду pMON30309 (G-CSF [Ser17] /IgG2b).
Пример 112
Плазмида pMON39909 (Flt3L 1-139 (39/40)L21/GS/G-CSF [Ser17] (133/132) представляет один из двух химерных белков Flt3/G-CSF, в которых последовательности обоих компонентов перестроены. NcoI/AflIII-фрагмент из состава pMON32198, включающий ген Flt3L 1-139 (39/40)L21/GlySer, был субклонирован в обработанную NcoI/SAP плазмиду pMON25187 (продукционная для клеток E.coli плазмида, несущая единственную копию гена G-CSF [Ser17] (133/132)). После подтверждения последовательности ДНК химерную вставку субклонировали в плазмиду pMON3934 в виде NcoI/HindIII-фрагмента и обозначали pMON39909. Плазмида PMON39910 (G-CSF [Ser17] (133/132)/IgG2b/Flt3L 1-139 (39/40L21) была сконструкирована с помощью ПЦР с использованием pMON25187 в качестве матрицы и пары затравок GPForl/GFRev2. Условия амплификации были идентичны тем, которые использовались для pMON39908. Обработанный NcoI/SnaBI ген G-CSF (Ser17) (133/132) был субклонирован в NcoI/SnaBI-сайт плазмиды PMON32376, включающий ген IgG2b/Flt3L 1-139 (39/40)L21.
Пример 113
Плазмида pMON40000 является продукционной плазмидой, модифицированной на базе плазмиды pClneo (Promega), включающей ген G-CSF (Ser17)/GlySer/Flt3L 1-139 (39/40)L21, предназначенной для экспрессии в клетках NSO (является эквивалентом плазмиды pMON32169, экспрессирующейся в клетках ВНК). Плазмида pMON40000 включает промоторно-энхансерный элемент CMV-IE, лидерный мотив IL-3, расположенный сразу перед геном CSF (Ser17)/GlySer/Flt3L 1-139 (39/40)L21, промотор укороченной тимидинкиназы, полиадениловый сигнал позднего гена SV40 и несколько сайтов гиперчувствительности к ДНКазе (как часть минимального сегмента LCR гена IgH 3).
Пример 114
Биологическая активность многофункциональных химерных агонистов гемопоэтических рецепторов (см. табл. 15).
Биотест на активность многофункциональных химерных агонистов гемопоэтических рецепторов in vitro
+ - снижение потенциала (смещение вправо) в сравнении с контролем
++ - равный потенциал в сравнении с контролем (в пределах двух крат)
+++ - увеличенный потенциал (смещение влево) в сравнении с контролем
1- в сравнении с контролем pMON30247
2- в сравнении с контролем pMON32352
3- в сравнении с подходящим добавочным контролем
4 - тестирована в смешанном пуле pMON40000 и pMON40002
Пример 115
Определение биологической активности (см. табл. 16).
+ - сниженная активность по сравнению с IL-3, IL-6, SCF, G-SCF (контроль по литературным данным)
++ - сходная (в пределах 20%) активность с IL-3, IL-6, SCF, G-CSF (контроль по литературным данным)
+++ - увеличенная активность в сравнении с IL-3, IL-6, SCF, G-CSF (контроль по литературным данным)
Условия культивирования: среда X-Vivo 10, 37°С, 5% CO2, 11 дней инкубации
2Обозначения:
+ - сниженная активность в сравнении с GM-CSF, TNFα, SCF
(контроль по литературным данным)
++ - сходная (в пределах 20%) активность в сравнении с GM-CSF, TNFα, SCF (контроль по литературным данным)
+++ - увеличенная активность в сравнении с GM-CSF, TNFα, SCF (контроль по литературным данным)
Условия культивирования: среда IMDM-20 с добавлением 100 нг/мл GM-CSF, 100 нг/мл TNFα, 20 нг/мл SCF, при 37°С, 5% СO2,
в течение 18-22 дней
агонист-I MFR = pMON31140 (WO 95/21i97)
агонист-II MFR = pMON28571 (WO 97/12985)
Тест на гемопоэтическую экспансию ex vivo
Выделенную обогащенную фракцию СD34-позитивных клеток-предшественников костного мозга человека культивировали из расчета 5·104 клеток на 1 мл в среде ex-vivo 10 с добавлением 1% HSA с тестируемыми цитокинами и с контролем для оценки потенциала цитокиновой экспансии. Клетки экспансировали и пересевали из расчета 5·104 клеток на 1 мл в новую среду с добавлением цитокинов примерно на 5-й день в зависимости от роста культуры. На 10-й день клетки отбирали и характеризовали. Клетки собирали с чашек и доводили их концентрацию до 106 клеток на 1 мл. Определяли общий уровень клеточной экспансии, а клетки тестировали на соответствие клеткам-предшественникам кроветворения в пре- и постэкспанзионном тесте на метилцеллюлозе (Stem Cell Technol., Methocult HCC3534). Экспансированные клетки также тестировали с помощью проточной цитометрии для определения типовой принадлежности: CDllb(РЕ)/CD15/(FITC), CD34 (FITC), CD41a (НТС).
Тест на экспансию дендритных клеток ex vivo
Выделенную обогащенную фракцию CD34-позитивных клеток-предшественников костного мозга человека культивировали из расчета 2·105 клеток на 1 мл в среде IMDM с добавлением 20% плодной телячьей сыворотки с тестируемыми цитокинами и контролем для оценки экспансионного потенциала. Клетки экспансировали и пересевали из расчета 5·104 клеток на 1 мл в новую среду с добавлением цитокинов примерно на 5-й день в зависимости от роста культуры. На 18-22-й день клетки отбирали и характеризовали. Определяли общий уровень экспансии клеток, а экспансированные клетки тестировали с помощью проточной цитометрии с целью определения типовой принадлежности: HLA-DR+ (PE)/CDla+ (FITC), CD86+(РЕ)/CDla+ (FITC), CD19- (FITC). Степень экспансии дендритных клеток определяли как произведение общего уровня клеточной экспансии и доли клеток HLA-DR+/CD1a+. Функциональную активность клеток определяли с использованием монотипной реакции смешанных лимфоцитов. Промытые, облученные культивируемые дендритные клетки добавляли в серии доз во фракции моноядерных клеток периферической крови, являющихся аллогенетическими иммунокомпетентными клетками, помещенными в 96-луночный микротировальный планшет. Способность дендритных клеток функционировать в качестве антиген-презентирующих клеток определяли по степени пролиферации, стимулированной в препаратах иммунокомпетентных клеток, измеряемой по уровню инкорпорации 3H-тимидина.
Пример 116
Связывание рецепторов (см. табл. 17).
Данные приведены в виде среднего ± стандартная ошибка, определенные по крайней мере по трем экспериментам в серии из трех повторностей, за исключением pMON323600, когда только два (2) эксперимента были проведены полностью.
Аффинность агониста Flt-3, включающего химерные молекулы, оценивали в тестах на связывание рецепторами. Анализ BIACORE был проведен путем прямой иммобилизации Flt3-Fc, a величину Rd подсчитывали путем определения констант ассоциации и диссоциации. Сравнительные тесты на связывание были использованы для оценки взаимодействий химерных молекул либо с рецептором G-CSF, трансфицированного в клетки BaF3, либо с γ-субъединицей рецептора IL-3, экспрессируемой клетками ВНК. С использованием этих клеток были проведены тесты на конкурентное связывание агонист-специфичных меченых лигандов: показатели IC50 определяли для конкурентных химер с использованием log-log анализа кривых зависимости от дозы.
Пример 117
Биологическая активность In vivo (см. табл. 18).
Данные по тестированию многофункциональных химерных агонистов гемопоэтических рецепторов у мышей in vivo
Мышей линии C57BL/6 инъецировали подкожно плазмидами рМОХ-30247, pMON32342 или pMON32360 (150 мкг в день) или pMON32191 (200 мкг/день) или мышиным сывороточным альбумином (200 мкг/день) в течение 10 дней. На 11-й день несовместимые с жизнью кровотечения образовывали путем прямой пункции сердца. Подсчет лейкоцитов проводили в цельной крови. Лейкоциты периферической крови выделяли с помощью градиентного центрифугирования (Histopaque) с последующим лизированием хлоридом аммония с целью дальнейшего удаления эритроцитов. Клетки окрашивали для проточной цитометрии с использованием прямых моноклональных антител, соединенных с флуоресцеином или фикоэритрином (Pharmingen). Перед окрашиванием неспецифичное связывание рецептором Fc было заблокировано с использованием реактива FcBlock (Pharmingen). Клетки анализировали на цитометре FacScan (Becton/Dickinson). Долю позитивных клеток определяли интегрально, а распределение фенотипов клеток определяли исходя из подсчета WBC (лейкоцитарная формула). Селезенки у инъецированных животных удаляли в стерильных условиях и препарировали иглами в среде RPMI. Клеточные суспензии получали с использованием плоской поверхности задней части 5-кубового шприца с последующей фильтрацией через хлопковую пробку с целью удаления комков. Эритроциты удаляли с помощью лизиса хлоридом аммония, клетки промывали, ресуспензировали и подсчитывали с помощью счетчика Coulter (Coulter Electronics). Клетки подготавливали для проточной цитометрии также, как это было описано выше. Фенотип маркировали как число клеток в расчете на 1 селезенку, основываясь на доле клеток с позитивным фенотипом и общей лейкоцитарной формуле селезенки. Культуры CFU получали путем посева клеток селезенки из расчета 1,5·105 клеток селезенки на 1 мл в трехраздельных метилцеллюлозных лунках с мышиными цитокинами и эритропоэтином (Stem Cell Technologies). Культуры инкубировали в течение 10 дней при 37°С и обсчитывали под световым микроскопом. Показатель CFU определяли как колонию, включающую более 50 клеток. Степень увеличения числа CFU на 1 селезенку определяли как отношение общего числа CFU в 1 селезенке при тестируемом соединении к общему числу CFU в 1 селезенке в контроле (собственный альбумин).
Контрольные значения (при инъекции собственного альбумина) для периферической крови составили 15 и 347 клеток в 1 мкл для фенотипов I-Ab+/CD11c+ и I-Ab+/-CD8+, соответственно. Контрольные значения (при инъекции собственного альбумина) для селезеночных лейкоцитов составили 2·106 и 1·106 клеток на 12 селезенку для фенотипов I-Ab+/CD11c+ и I-Ab+/-CD8+, соответственно.
В отсутствие дополнительных разработок можно быть уверенным, что специалисты в данной области техники на основе предложенных описаний смогут использовать настоящее изобретение в самой полной степени. Последующее предпочтительное специальное воплощение, такие образом, должно осуществляться как просто иллюстративные и ничем не ограниченные пути применения в любой соответствующей области.
Более подробное описание молекулярно-биологических методов очистки белков и биотестов можно найти в патентах WO 94/12639, WO 94/12638, WO 95/20976, WO 95/21197, WO 95/20977, WO 95/21254 и WO 96/23888, которые полностью включены в настоящий текст в виде ссылок.
Все ссылки, патенты и сообщения, тестированные в настоящем тексте, включены в виде ссылок в полном необходимом объеме.
Различные другие примеры могут стать доступными специалисту в данной области техники, ознакамливающемуся с настоящим изобретением без отклонений от духа и буквы настоящего изобретения. Имеется в виду, что все такие другие примеры будут включены в формулировку цели в формуле изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2019 |
|
RU2822196C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР КЛЕТОК | 2015 |
|
RU2751362C2 |
| БЕЛКОВЫЙ КОМПЛЕКС ИНТЕРЛЕЙКИНА 15 И ЕГО ПРИМЕНЕНИЕ | 2015 |
|
RU2711979C2 |
| МОДИФИЦИРОВАННЫЕ ГЕМОПОЭТИЧЕСКИЕ СТВОЛОВЫЕ КЛЕТКИ/КЛЕТКИ-ПРЕДШЕСТВЕННИКИ И Не-Т ЭФФЕКТОРНЫЕ КЛЕТКИ, И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2733652C2 |
| КОМПОЗИЦИИ Т-КЛЕТОК С НЕДОСТАТОЧНОСТЬЮ РЕЦЕПТОРОВ Т-КЛЕТОК | 2013 |
|
RU2781979C2 |
| Иммунотерапия с применением антител, связывающих лиганд 1 белка программируемой смерти клеток (PD-L1) | 2017 |
|
RU2766582C2 |
| ИММУНОТЕРАПИЯ С ПРИМЕНЕНИЕМ АНТИТЕЛ, СВЯЗЫВАЮЩИХ БЕЛОК 1 ПРОГРАММИРУЕМОЙ СМЕРТИ КЛЕТОК (PD-1) | 2017 |
|
RU2768404C2 |
| АНТИТЕЛА К CD40 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2715597C2 |
| Рекомбинантная плазмидная ДНК pET19b-Surv-OL, обеспечивающая синтез гибридного белка сурвивин-обелин (Surv-OL) и гибридный белок, связываемый анти-сурвивин антителами и обладающий биолюминесцентной активностью | 2021 |
|
RU2770490C1 |
| МНОГОФУНКЦИОНАЛЬНЫЕ СЛИТЫЕ БЕЛКИ И ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2815388C2 |















































































































































































































































































































































































































































































































































































































































































































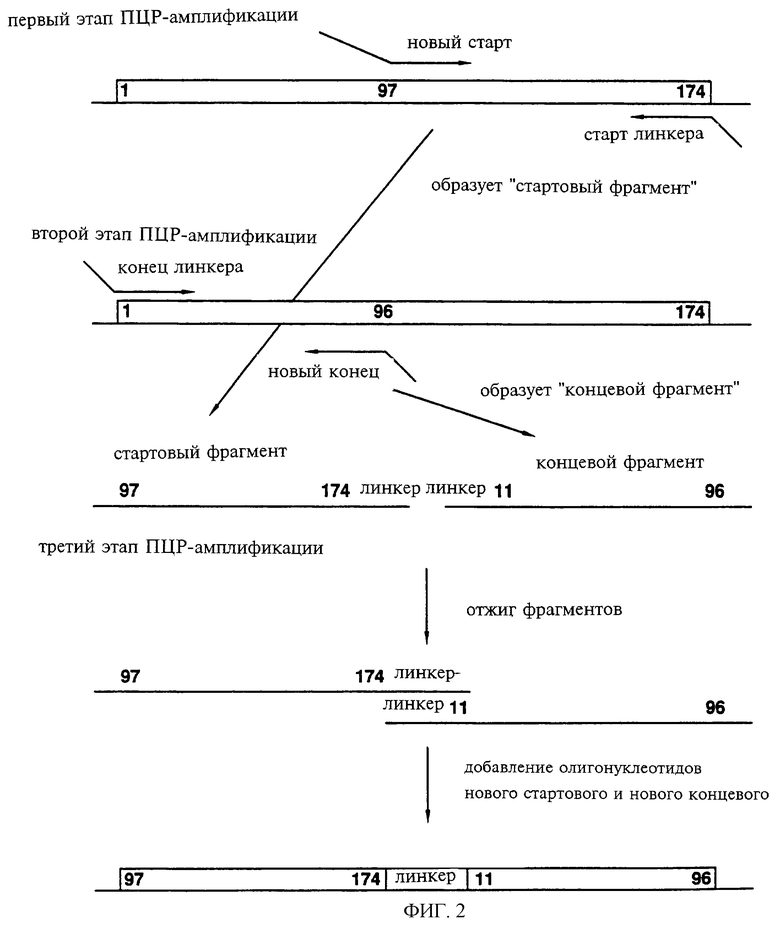
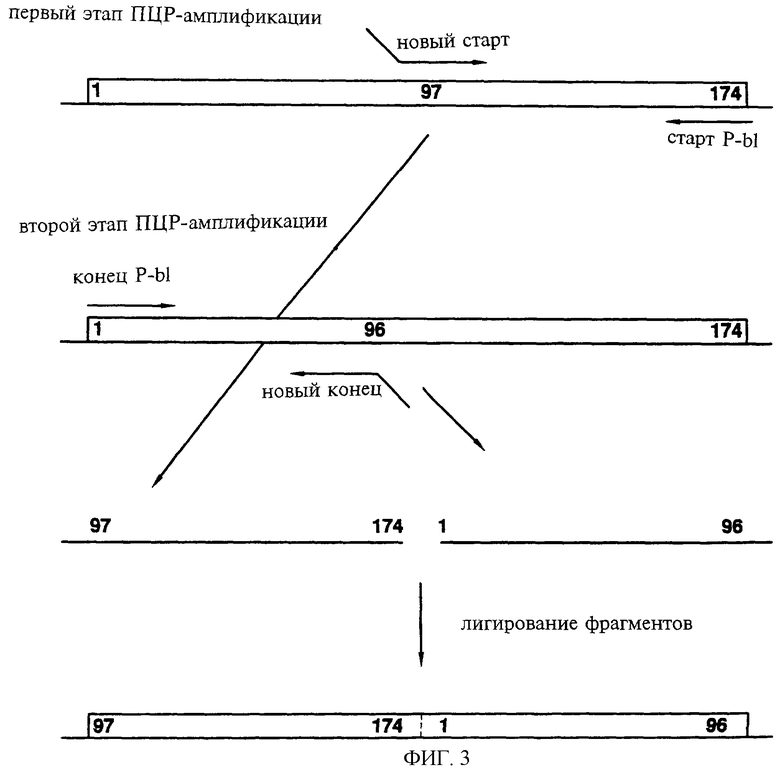
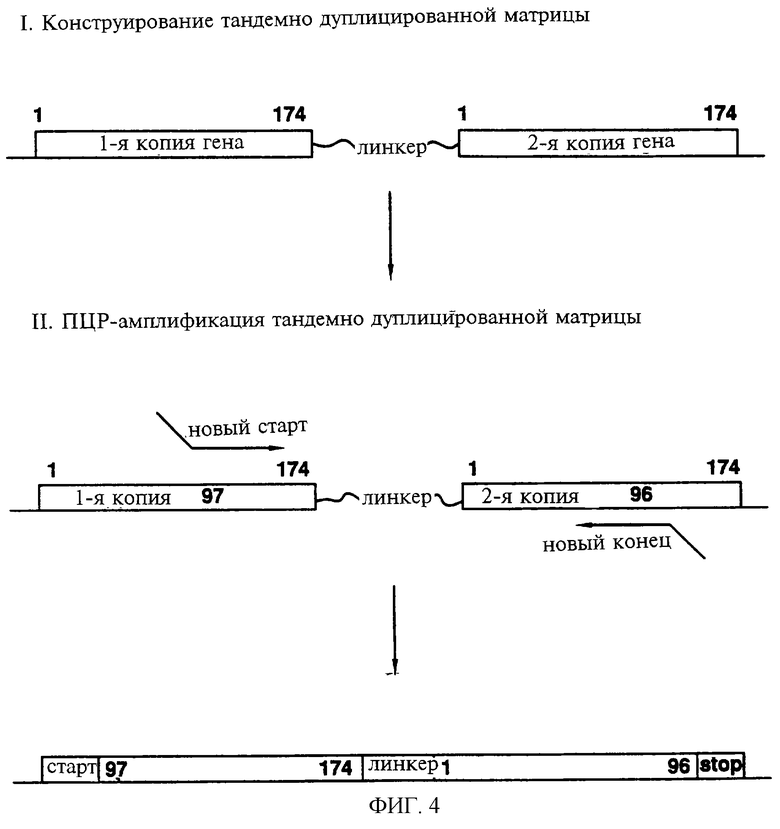
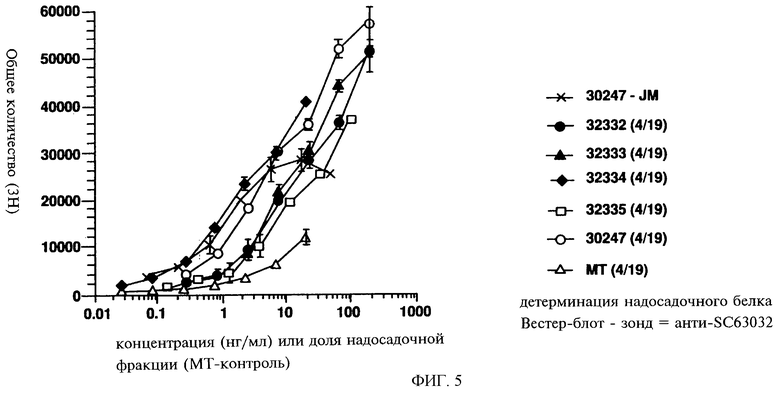














Изобретение относится к биотехнологии, в частности генной инженерии, и может быть использовано для получения химерных многофункциональных гемопоэтических белков. Гемопоэтический белок содержит аминокислотную последовательность формулы R1–L1–R1, R2–L1–R1, R1–R2 или R2–R1, где R1 представляет собой модифицированный лиганд flt–3, R2 представляет собой модифицированный IL-3 человека, модифицированный или немодифицированный колониестимулирующий фактор. Модификацию R1 получают путем присоединении N-конца с С-концом непосредственно или через линкер (L2), способный соединять N-конец с С-концом с образованием новых С- и N-концов. Модифицированный IL-3 человека получают путем замены аминокислот в положении 17-123. G-CSF человека модифицируют путем аминокислотных замен. Гемопоэтический белок получают путем культивирования клеток, трансформированных вектором, содержащим РНК, кодирующую гемопоэтический белок. Гемопоэтический белок стимулирует продуцирование кроветворных клеток. В составе фармацевтической композиции его используют для лечения человека, страдающего опухолью, инфекционным или аутоиммунным заболеванием. Изобретение позволяет получать многофункциональные гемопоэтические белки, проявляющие повышенную активность стимуляции кроветворных клеток и обладающие улучшенными физическими характеристиками. 10 н. и 12 з.п. ф-лы, 10 ил., 18 табл.
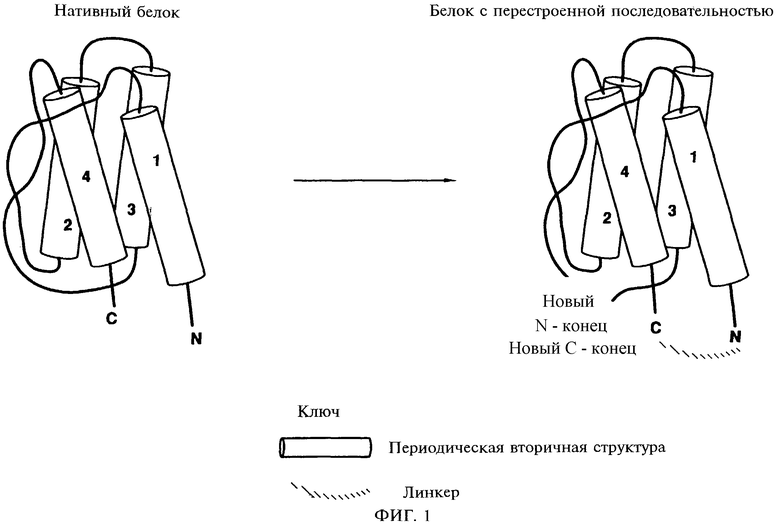
причем R2 является полипептидом, содержащим модифицированную аминокислотную последовательность IL-3 человека SEQ ID NO:859, где
Хаа в положении 17 is Ser, Lys, Gly, Asp, Met, Gln или Arg;
Xaa в положении 18 is Asn, His, Leu, Ile, Phe, Arg или Gln;
Xaa в положении 19 is Met, Phe, Ile, Arg, Gly, Ala или Cys;
Хаа в положении 20 is Ile, Cys, Gln, Glu, Arg, Pro или Ala;
Xaa в положении 21 is Asp, Phe, Lys, Arg, Ala, Gly, Glu, Gln, Asn, Thr, Ser или Val;
Хаа в положении 22 is Glu, Trp, Pro, Ser, Ala, His, Asp, Asn, Gln, Leu, Val или Gly;
Xaa в положении 23 is Ile, Val, Ala, Gly, Trp, Lys, Phe, Leu, Ser или Arg;
Хаа в положении 24 is Ile, Gly, Val, Arg, Ser, Phe или Leu;
Хаа в положении 25 is Thr, His, Gly, Gln, Arg, Pro или Ala;
Хаа в положении 26 is His, Thr, Phe, Gly, Arg, Ala или Trp;
Xaa в положении 27 is Leu, Gly, Arg, Thr, Ser или Ala;
Хаа в положении 28 is Lys, Arg, Leu, Gln, Gly, Pro, Val или Trp;
Xaa в положении 29 is Gln, Asn, Leu, Pro, Arg или Val;
Хаа в положении 30 is Pro, His, Thr, Gly, Asp, Gln, Ser, Leu или Lys;
Хаа в положении 31 is Pro, Asp, Gly, Ala, Arg, Leu или Gln;
Xaa в положении 32 is Leu, Val, Arg, Gln, Asn, Gly, Ala или Glu;
Хаа в положении 33 is Pro, Leu, Gln, Ala, Thr или Glu;
Хаа в положении 34 is Leu, Val, Gly, Ser, Lys, Glu, Gln, Thr, Arg, Ala, Phe, Ile или Met;
Xaa в положении 35 is Leu, Ala, Gly, Asn, Pro, Gln или Val;
Хаа в положении 36 is Asp, Leu или Val;
Xaa в положении 37 is Phe, Ser, Pro, Trp или Ile;
Хаа в положении 38 is Asn или Ala;
Xaa в положении 40 is Leu, Trp или Arg;
Хаа в положении 41 is Asn, Cys, Arg, Leu, His, Met или Pro;
Хаа в положении 42 is Gly, Asp, Ser, Cys, Asn, Lys, Thr, Leu, Val, Glu, Phe, Tyr, Ile, Met или Ala;
Xaa в положении 43 is Glu, Asn, Tyr, Leu, Phe, Asp, Ala, Cys, Gln, Arg, Thr, Gly или Ser;
Хаа в положении 44 is Asp, Ser, Leu, Arg, Lys, Thr, Met, Trp, Glu, Asn, Gln, Ala или Pro;
Хаа в положении 45 is Gln, Pro, Phe, Val, Met, Leu, Thr, Lys, Trp, Asp, Asn, Arg, Ser, Ala, Ile, Glu или His;
Хаа в положении 46 is Asp, Phe, Ser, Thr, Cys, Glu, Asn, Gln, Lys, His, Ala, Tyr, Ile, Val или Gly;
Xaa в положении 47 is Ile, Gly, Val, Ser, Arg, Pro или His;
Хаа в положении 48 is Leu, Ser, Cys, Arg, Ile, His, Phe, Glu, Lys, Thr, Ala, Met, Val или Asn;
Хаа в положении 49 is Met, Arg, Ala, Gly, Pro, Asn, His или Asp;
Xaa в положении 50 is Glu, Leu, Thr, Asp, Tyr, Lys, Asn, Ser, Ala, Ile, Val, His, Phe, Met или Gln;
Xaa в положении 51 is Asn, Arg, Met, Pro, Ser, Thr или His;
Хаа в положении 52 is Asn, His, Arg, Leu, Gly, Ser или Thr;
Хаа в положении 53 is Leu, Thr, Ala, Gly, Glu, Pro, Lys, Ser или Met;
Хаа в положении 54 is Arg, Asp, Ile, Ser, Val, Thr, Gln, Asn, Lys, His, Ala или Leu;
Хаа в положении 55 is Arg, Thr, Val, Ser, Leu или Gly;
Xaa в положении 56 is Pro, Gly, Cys, Ser, Gln, Glu, Arg, His, Thr, Ala, Tyr, Phe, Leu, Val или Lys;
Xaa в положении 57 is Asn или Gly;
Xaa в положении 58 is Leu, Ser, Asp, Arg, Gln, Val или Cys;
Xaa в положении 59 is Glu Tyr, His, Leu, Pro или Arg;
Xaa в положении 60 is Ala, Ser, Pro, Tyr, Asn или Thr;
Хаа в положении 61 is Phe, Asn, Glu, Pro, Lys, Arg или Ser;
Xaa в положении 62 is Asn, His, Val, Arg, Pro, Thr, Asp или Ile;
Xaa в положении 63 is Arg, Tyr, Trp, Lys, Ser, His, Pro или Val;
Xaa в положении 64 is Ala, Asn, Pro, Ser или Lys;
Xaa в положении 65 is Val, Thr, Pro, His, Leu, Phe или Ser;
Хаа в положении 66 is Lys, Ile, Arg, Val, Asn, Glu или Ser;
Хаа в положении 67 is Ser, Ala, Phe, Val, Gly, Asn, Ile, Pro или His;
Хаа в положении 68 is Leu, Val, Trp, Ser, Ile, Phe, Thr или His;
Хаа в положении 69 is Gln, Ala, Pro, Thr, Glu, Arg, Trp, Gly или Leu;
Xaa в положении 70 is Asn, Leu, Val, Trp, Pro или Ala;
Хаа в положении 71 is Ala, Met, Leu, Pro, Arg, Glu, Thr, Gln, Trp или Asn;
Хаа в положении 72 is Ser, Glu, Met, Ala, His, Asn, Arg или Asp;
Хаа в положении 73 is Ala, Glu, Asp, Leu, Ser, Gly, Thr или Arg;
Хаа в положении 74 is Ile, Met, Thr, Pro, Arg, Gly, Ala;
Хаа в положении 75 is Glu, Lys, Gly, Asp, Pro, Trp, Arg, Ser, Gln или Leu;
Хаа в положении 76 is Ser, Val, Ala, Asn, Trp, Glu, Pro, Gly или Asp;
Хаа в положении 77 is Ile, Ser, Arg, Thr или Leu;
Xaa в положении 78 is Leu, Ala, Ser, Glu, Phe, Gly или Arg;
Хаа в положении 79 is Lys, Thr, Asn, Met, Arg, Ile, Gly или Asp;
Xaa в положении 80 is Asn, Trp, Val, Gly, Thr, Leu, Glu или Arg;
Хаа в положении 81 is Leu, Gln, Gly, Ala, Trp, Arg, Val или Lys;
Хаа в положении 82 is Leu, Gln, Lys, Trp, Arg, Asp, Glu, Asn, His, Thr, Ser, Ala, Tyr, Phe, Ile, Met или Val;
Хаа в положении 83 is Pro, Ala, Thr, Trp, Arg или Met;
Хаа в положении 84 is Cys, Glu, Gly, Arg, Met или Val;
Xaa в положении 85 is Leu, Asn, Val или Gln;
Хаа в положении 86 is Pro, Cys, Arg, Ala или Lys;
Хаа в положении 87 is Leu, Ser, Trp или Gly;
Хаа в положении 88 is Ala, Lys, Arg, Val или Trp;
Хаа в положении 89 is Thr, Asp, Cys, Leu, Val, Glu, His, Asn или Ser;
Хаа в положении 90 is Ala, Pro, Ser, Thr, Gly, Asp, Ile или Met;
Хаа в положении 91 is Ala, Pro, Ser, Thr, Phe, Leu, Asp или His;
Хаа в положении 92 is Pro, Phe, Arg, Ser, Lys, His, Ala, Gly, Ile или Leu;
Хаа в положении 93 is Thr, Asp, Ser, Asn, Pro, Ala, Leu или Arg;
Хаа в положении 94 is Arg, Ile, Ser, Glu, Leu, Val, Gln, Lys, His, Ala или Pro;
Xaa в положении 95 is His, Gln, Pro, Arg, Val, Leu, Gly, Thr, Asn, Lys, Ser, Ala, Trp, Phe, Ile или Tyr;
Хаа в положении 96 is Pro, Lys, Tyr, Gly, Ile или Thr;
Хаа в положении 97 is Ile, Val, Lys, Ala или Asn;
Xaa в положении 98 is His, Ile, Asn, Leu, Asp, Ala, Thr, Glu, Gln, Ser, Phe, Met, Val, Lys, Arg, Tyr или Pro;
Хаа в положении 99 is Ile, Leu, Arg, Asp, Val, Pro, Gln, Gly, Ser, Phe или His;
Хаа в положении 100 is Lys, Tyr, Leu, His, Arg, Ile, Ser, Gln или Pro;
Хаа в положении 101 is Asp, Pro, Met, Lys, His, Thr, Val, Tyr, Glu, Asn, Ser, Ala, Gly, Ile, Leu или Gln;
Хаа в положении 102 is Gly, Leu, Glu, Lys, Ser, Tyr или Pro;
Xaa в положении 103 is Asp или Ser;
Xaa в положении 104 is Trp, Val, Cys, Tyr, Thr, Met, Pro, Leu, Gln, Lys, Ala, Phe или Gly;
Хаа в положении 105 is Asn, Pro, Ala, Phe, Ser, Trp, Gln, Tyr, Leu, Lys, Ile, Asp или His;
Xaa в положении 106 is Glu, Ser, Ala, Lys, Thr, Ile, Gly или Pro;
Хаа в положении 108 is Arg, Lys, Asp, Leu, Thr, Ile, Gln, His, Ser, Ala или Pro;
Хаа в положении 109 is Arg, Thr, Pro, Glu, Tyr, Leu, Ser или Gly;
Хаа в положении 110 is Lys, Ala, Asn, Thr, Leu, Arg, Gln, His, Glu, Ser или Trp;
Хаа в положении 111 is Leu, Ile, Arg, Asp или Met;
Хаа в положении 112 is Thr, Val, Gln, Tyr, Glu, His, Ser или Phe;
Хаа в положении 113 is Phe, Ser, Cys, His, Gly, Trp, Tyr, Asp, Lys, Leu, Ile, Val или Asn;
Хаа в положении 114 is Tyr, Cys, His, Ser, Trp, Arg или Leu;
Хаа в положении 115 is Leu, Asn, Val, Pro, Arg, Ala, His, Thr, Trp или Met;
Хаа в положении 116 is Lys, Leu, Pro, Thr, Met, Asp, Val, Glu, Arg, Trp, Ser, Asn, His, Ala, Tyr, Phe, Gln или Ile;
Xaa в положении 117 is Thr, Ser, Asn, Ile, Trp, Lys или Pro;
Хаа в положении 118 is Leu, Ser, Pro, Ala, Glu, Cys, Asp или Tyr;
Хаа в положении 119 is Glu, Ser, Lys, Pro, Leu, Thr, Tyr или Arg;
Хаа в положении 120 is Asn, Ala, Pro, Leu, His, Val или Gln;
Хаа в положении 121 is Ala, Ser, Ile, Asn, Pro, Lys, Asp или Gly;
Хаа в положении 122 is Gln, Ser, Met, Trp, Arg, Phe, Pro, His, Ile, Tyr или Cys;
Хаа в положении 123 is Ala, Met, Glu, His, Ser, Pro, Tyr или Leu;
причем L1 является линкером, способным связывать R1 и R2, и указанному гемопоэтическому белку может непосредственно предшествовать (метионин-1), (аланин-1) или (метионин-2, аланин-1);
причем L1 выбран из группы, состоящей из перестановок аминокислотных последовательностей, содержащих Gly, Asn, Ser, Thr, Ala, шарнирную область тяжелой цепи иммуноглобулина IgG, IgA, IgM, IgD или IgE, шарнирную область гамма-цепи 2b мышиного IgG, SEQ ID NO: 786-788, 793-797, 800-802, 861-865; и указанные линкеры дополнительно необязательно содержат последовательность распознавания эндопептидазы; причем от 1 до 14 аминокислот необязательно делетированы с N-конца и/или от 1 до 15 аминокислот необязательно делетированы с С-конца указанной модифицированной аминокислотной последовательности IL-3 человека, причем аминокислоты 0-44, обозначенные Хаа, отличаются от соответствующих аминокислот нативного (1-133) интерлейкина-3 (IL-3) человека.
причем R2 выбран из группы, состоящей из GM-CSF, G-CSF, G-CSF Ser17, G-CSF Ala17, c-mpl-лиганда (также известного как TPO или MGDF), М-CSF, эритропоэтина (EPO), IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-13, IL-15, LIF, f1t3/f1k2-лиганда, человеческого гормона роста, фактора роста В-клеток, фактора дифференцировки В-клеток, фактора дифференцировки эозинофилов и фактора стволовых клеток (SCF) (также известного как steel-фактор или с-kit-лиганд); причем L1 является линкером, способным связывать R1 и R2, и указанному гемопоэтическому белку может непосредственно предшествовать (метионин-1), (аланин-1) или (метионин-2, аланин-1); причем L1 выбран из группы, состоящей из перестановок аминокислотных последовательностей, содержащих Gly, Asn, Ser, Thr, Ala, SEQ ID NO: 786-788, 793-797, 800-802 и 861-865, шарнирную область тяжелой цепи иммуноглобулина IgG, IgA, IgM, IgD или IgE, шарнирную область гамма-цепи 2b мышиного IgG; и указанные линкеры дополнительно необязательно содержат последовательность распознавания эндопептидазы.
причем R2 является полипептидом, содержащим модифицированную последовательность аминокислот G-CSF человека SEQ ID NO:858, где
Хаа в положении 1 представляет собой Thr, Ser, Arg, Tyr или Gly;
Хаа в положении 2 представляет собой Pro или Leu;
Хаа в положении 3 представляет собой Leu, Arg, Thr или Ser;
Xaa в положении 13 представляет собой Phe, Ser, His, Thr или Pro;
Хаа в положении 16 представляет собой Lys, Pro, Ser, Thr или His;
Хаа в положении 17 представляет собой Cys или Ser;
Хаа в положении 18 представляет собой Leu, Thr, Pro, His, Ile или Cys;
Хаа в положении 22 представляет собой Arg, Tyr, Ser, Thr или Ala;
Xaa в положении 24 представляет собой Ile, Pro, Tyr или Leu;
Хаа в положении 27 представляет собой Asp или Gly;
Хаа в положении 30 представляет собой Ala, Ile или Leu;
Хаа в положении 34 представляет собой Lys или Ser;
Xaa в положении 43 представляет собой His, Thr, Gly, Val, Lys, Trp, Ala, Arg, Cys или Leu;
Хаа в положении 44 представляет собой Pro, Gly, Arg, Asp, Val, Ala, His, Trp, Gln или Thr;
Хаа в положении 46 представляет собой Glu, Arg или Ala;
Хаа в положении 47 представляет собой Leu или Thr;
Хаа в положении 49 представляет собой Leu, Phe, Arg или Ser;
Хаа в положении 50 представляет собой Leu или His;
Хаа в положении 54 представляет собой Leu или His;
Хаа в положении 67 представляет собой Gln, Lys, Leu или Cys;
Хаа в положении 70 представляет собой Gln, Pro, Leu, Arg или Ser;
Хаа в положении 104 представляет собой Asp, Gly или Val;
Xaa в положении 108 представляет собой Leu, Ala, Val, Arg, Trp, Gln или Gly;
Хаа в положении 115 представляет собой Thr, His, Leu или Ala;
Хаа в положении 144 представляет собой Phe, His, Arg, Pro, Leu или Glu;
Хаа в положении 146 представляет собой Arg или Gln;
Xaa в положении 147 представляет собой Arg или Gln;
Хаа в положении 156 представляет собой His, Gly или Ser;
Хаа в положении 159 представляет собой Ser, Arg, Thr, Tyr, Val или Gly;
Хаа в положении 162 представляет собой Glu, Leu, Gly или Trp;
Хаа в положении 163 представляет собой Val, Gly, Arg или Ala;
Хаа в положении 169 представляет собой Ser, Leu, Arg или Cys;
Xaa в положении 170 представляет собой His, Arg или Ser;
причем 1-11 аминокислот с N-конца и/или 1-5 аминокислот с С-конца необязательно делетированы из указанной модифицированной аминокислотной последовательности G-CSF человека и причем N-конец присоединен к С-концу непосредственно или через линкер (L2), определенный выше и способный соединять N-конец с С-концом с образованием новых С- и N-концов при аминокислотах:
соответственно
причем L1 является линкером, способным связывать R1, и R2, и указанному гемопоэтическому белку может непосредственно предшествовать (метионин-1), (аланин-1) или (метионин-2, аланин-1); причем L1 выбран из группы, состоящей из перестановок аминокислотных последовательностей, содержащих Gly, Asn, Ser, Thr, Ala, SEQ ID NO: 786-788, 793-797, 861-865 и 800-802, шарнирную область тяжелой цепи иммуноглобулина IgG, IgA, IgM, IgD или IgE; шарнирную область гамма-цепи 2b мышиного IgG; и указанные линкеры дополнительно необязательно содержат последовательность распознавания эндопептидазы.
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| US 5376367 A, 27.12.1994 | |||
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| HORLICK, R.A | |||
| et al., Permuteins of interleukin 1 beta | |||
| Protein engineering, 1992, v.5, №2, p.427-431. | |||

